Embed Size (px)
Citation preview

BIOCHIMIE, 1977, 59, 9'27-932.
Hdmisynth se d'antibiotiques aminoglycosidiques : I . - M i s e a u point d'un r6acteur enzymatique h c o f a c t e u r .
Fran$ois LE GoFFIC <>, Sames SIcsIc et Chris t ian VINCENT. C . E . R . C . O . A . - - C . N . R . S . 2 ~ 8, r u e H e n r y D t m a n t - - 9 ~ 3 2 0 T h i a i s ( F r a n c e ) .
( 1 ~ - I I - 1 9 7 7 ) .
S u m m a r y . - - In order to produce specifically N-monoalkylated derivatives of amino- glycoside antibiotics of potential therapeutic values, ~ve have developed an enzymatic reactor. This system uses the aminoglyeoside acetyltransferase as catalyst and acetyl- coenzyme A as acetyl donor.
The immobilization of one aminoglycoside aeetyltransferase on different resins has been studied. The coretieulation of this enzyme on DEAE cellulose in the presence of glutaraldehyde gives rise to an enzymatic resin of high efficiency. On the other hand, we have also studied the aeetylation of coenzyme A in a simple manner. Acetylation occurs in a quantitative yield vehen the reaetion is performed in the presence of polyvinyl-4 pyridine/divinylbenzene 2 per cent.
These conclusions enabled to develop t~'o types of acetylating reactors which give rise without purification to 3-acetyl gentamicin.
La r6sistance des bact6ries, d 'or ig ine hospi- talibre, aux ant ib iot iques aminoglycos id iques s 'expl ique p r inc ipa l emen t par la modif ica t ion enzymat iqne de ces mo16cules (phosphoryla t ion , nucl6ot idylat ion, ac6tylation), ce qui alt6re consi- d6rablement leurs propri6t6s biologiques.
Les c l in ic iens se t rouvent done confront6s des prob16mes d i f f i c i l e s : t rouver des moyens pour juguler les infec t ions ~ bact6ries r6sistantes. Plusieurs solutions d 'o rd re ch imique ont 6t6 rete- nues pour r6soudre ce problbme :
- - enlever la fonct ion concern6e par la modi- fication enzymat ique. C'est ainsi qu'est n6e, pa r exemple, la dib6kacine ou dideoxy-3 ' ,4 ' kanamy- cine B [1],
- - e f f e c t u e r une modif ica t ion ch imique telle que le complexe E-S, p remi6re 6tape d 'une r6ac- t ion enzymat ique, ne puisse plus se fo rmer : ainsi a 6t6 d6velopp~e l ' amikac ine [2],
- - enfin, cr6er un encombremen t st6rique sur la cible de 1'enzyme.
L 'ac6tylat ion enzymat ique rev6t une impor tance route par t icul i6re puisque trois enzymes sont impliqu6es dans ce processus :
- - une ac6tyl transf6rase AAC (3), t rouv6e chez une s~rie de bact6vies gram ( - - ) , ac6tyle l 'azote port6 par le carbone 3 de ces ant ib iot iques [3],
- - une ac6tyl transf6rase AAC (2'), t rouv6e chez des Prov idenc ia , ac6tyle l 'azote 2' [4],
A qui toute correspondance doit gtre adress6e.
- - une ac6tyl transf6rase AAC (6'), t rouv6e chez les bact6ries gram (_+) ac6tyle l 'azote port6 p a r le carbone 6' [5], dans la mesure off celui-ci n 'est pas substitu6.
Nous nous sommes done propos6s de cr6er un encombremen t st6rique sur ces diff6rentes fonc- t ions azot6es. Leur aIkylat ion nous paralt , en effet, une solution ra isonnable puisque l ' an t ib io t ique gardera i t ainsi le m6me nombre de sites basiques.
L 'a lkytat ion ch imique n'6tant pas s61ective, i l 6tait tentant de r ecou r r i r h la sp6cificit6 des enzy- mes d ' inac t iva t ion . Le sch6ma suivant r6sume la s6quence r6act ionnel le pe rmet tan t d 'ob ten i r un ant ib io t ique aminoglycos id ique N-6thyl6 d 'une mani6re hautement sp6cifique :
R NH 2 AAC R-NH~CO-CH 3 ( I )
A~ CoA CoA SH
R-NH-CO-CH 3 B2H6 .> R-NH- CH 2-CH 3 ( ] I )
Si l '6tape (II) ne posait , a p r i o r i , pas de pro- bl6me impor t an t puisqu 'on conna i s sa i t un cer ta in nombre de r6actifs ch imiques capables de r6duire une amide en amine, l '6tape (I), malgr6 l 'avantage de sa sp6cificit6, soulevait de nombreuses ques- tions :
a) les enzymes capables d 'ac6ty!er les amino- glycosides sont instables, pa r cons6quent leur uti- l isation est unique,
66

928 F. L e G o f f i c e t co l l .
b) le p r o d u i t de la rdac t ion est diff lci le h pu- r if ier ,
c) l ' ac6ty l coenzyme A est d 'un p r ix tel que la m6thode devient i m p r a t i c a b l e pou r p r o d u i r e des quant i t6s notables d ' an t ib io t iques N-ac6tyl6s.
Les deux p r e m i e r s probl~mes, couran ts dans les r6ac t ions enzymat iques monomol6cula i res , ont d6jh t rouv6 une solut ion sa t i s fa isante [fi, 7] p a r r i m m o b i l i s a t i o n de l ' enzyme sur suppor t polym6- r ique pe rme t t an t h la fois la s tabi l i sa t ion , l 'u t i l i - sa t ion mul t ip le de l ' enzyme et l ' i so lement ais6 des p r o d u i t s de r6ac t ion p a r s imple f i l t rat ion.
Si le p rob l6me de la reg6n6rat ion du cofac teur a t rouv6 des solu t ions plus ou moins sa t i s fa isantes dans le cas de cer ta ins coenzymes d 'oxydo- r6duc- t ion tels que le couple NAD ÷, NADH ou les com- p lexes FMN, FMNH2 ; FAD, FADH 2 [8], le p ro- blbme de la reg6n6rat ion r a p i d e et quant i ta t ive de l ' ac6 ty lcoenzyme A h p a r t i r du coenzyme A n 'a j amais 0 6 abord6.
Dans le p r6sen t ar t ic le , nous exposerons succes- s ivement :
1) les 6tudes que nous avons r6alis6es sur la s tab i l i sa t ion d 'un aminog lycos ide ac6tyl t ransf6- rase (AAC(3)), don t la cible est l 'azote 3 de ces mol6cules, p a r immobi l i s a t ion sur d ivers suppor ts ,
2) les 6tudes qui ont about i ~ la reg6n6rat ion de l ' ac6ty l coenzyme A /i p a r t i r du coenzyme A avec un r endemen t quasi quant i la t i f ,
R'\CH/R2
0
H ~ / C H 3
NH
Fro. 1. - - S tructure de la gentamieine C, C~ RI = H 1% = NH~ C~ R1 ---- CH8 1% ---- NH-CH~ C~ R~ ---- CH8 R~ ---- NH~
3) la r6a l i sa t ion d 'un r6ac teur enzymat ique pro- p r e m e n t di t qui p e r m e t la synth6se de la N-3 ac6- tyl gentamic ine .
BIOCHIMIE, 1977, 59, n ° 11-12.
MATERIEL ET METHODES.
L ' an t ib io t ique uti l is6 dans ce t r ava i l est la gen tamic ine C (fig. 1) et p rov i en t des Labora to i r e s Unilabo.
La source d ' enzyme est un co l ibac i l l e [3] r6sis- tant h la gentamic ine . Ce m i c r o o r g a n i s m e cst cul- tiv6 sur mi l ieu t ryp t i ca se soy bro th (30 g/ l) pen- dant 24 h h 37 °. Apr6s les t r a i t ements c lass iques (r6colte, lyse, cent r i fuga t ion) , ce qui condui t h u n << ex t ra i t bac t6r ien b ru t >>, l ' enzyme est purif i6e p a r c h r o m a t o g r a p h i e d 'aff ini t6 [9]. L 'ac t iv i t6 ac6- ty lan te est d6termin6e su ivant la m6thode de Da- vies [10J h l ' a ide de p a p i e r de phosphoce l lu lose P 81.
I - - I m m o b i l i s a t i o n de l ' e n z y m e .
a) I m m o b i l i s a t i o n s u r p o l g s a c c h a r i d e r~ t icu l~ : l ' i ndub iose A4 nous a 6t6 fourni p a r l ' Ins t i tu t Biologique F ran~a i s et a 6t6 fonct ionnal is6 p a r l ' i n t roduc t i on d 'un b ras t r ig lyc ine apr6s act iva- t ion du suppor t p a r le b romure de cyanogbne [113. Le couplage de l ' enzyme h cette r6sine est a lors r6alis6 h l ' a ide de c h l o r h y d r a t e de N 6thyl N ' (3-d im6thylamino- lubne sulfonate.
b) I m m o b i l i s a t i o n Whatman) : 2,5 g r inc6 p a r 1OO ml de
p ropy l ) c a r b o d i i m i d e p. to-
s u r D E A E c e l h d o s e (DE 52 de cel lulose p r 6 a l a b l ~ e n t t ampon B : Tr is HC1 10 -2 M
MgC12 10 -2 M NH4C1 2.10 -2 M, ~ mercap to6 thano l 6.10 -3 M (pH 7) sont ajout6s ~ 3 ml de solut ion enzymat ique . 70 td de g lu ta ra ld6hyde h 25 p. cent sont ajout6s au mil ieu. Le m61ange est a lors con- gel6 p e n d a n t 2: h puis laiss6 fi 4 ° p e n d a n t 24 h. La r6sine est a lors r inc6e pa r le t ampon pr6c6dem- ment d6cri t , jusqu 'h ce que les eaux de r incage n ' a ien t plus d 'ac t iv i t6 enzymat ique .
c) I n c l u s i o n d o n s u n ge l de f i b r i n e : cette inclu- s ion est effectu6e p a r add i t i on de 10 UI de th rom- b ine (Sigma) h 120. mg de f lbr inog6ne (Sigma) p r6a l ab l emen t d issous darts 3 ml d ' ex t r a i t enzy- mat ique. Le ca i l lo t obtenu apr6s 15 mn d ' a g i t a - t ion ~ 37°C est r inc6 p a r 20 ml de t ampon B.
I I - - R d a c t e u r s d ' a c d t y l a t i o n .
a ) R6acteur tubu la i re : 10 g de r6sine enzyma- tiqu'e h u m i d e Pr6a lab lement gonfl6e dans l0 ml de t ampon B sont in t rodu i t s dans une colonne (•,6 × 20 cm) thermosta t~e h 40 ,o . Une so lu t ion de 22,4 mg de sulfate de gen tamic ine C et 30 mg d 'ac6ty l [14C3 Coenzyme A ~ 0,4 ~Ci /mole dans 5 ml de t ampon B, est i n t rodu i t e au sommet de la colonne. La colonne est r inc6e p a r le t ampon. Le d6bi t d e 1 a co lonne est r6g16 h 0,2 m l / m n . L'~vo-
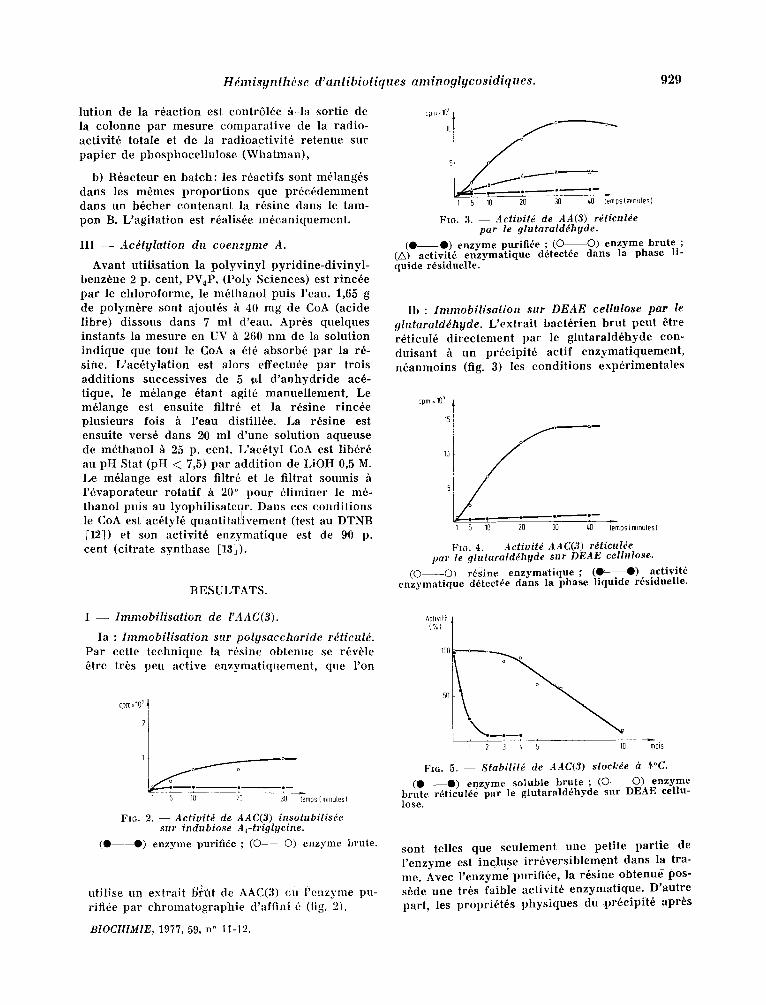
H d m i s y n l h d s e d ' a n t i b i o t i q u e s a m i n o g l y c o s i d i q u e s . 929
lut ion de la r6act ion est contr616e ~ la sortie de la colonne par mesure comparat ive de la radio- activit6 totale et de la radioact ivi t6 retenue sur papier de phosphocellulose (WhatmanL
b) R6acteur en ba tch: les r6actifs sont m61ang6s dans les m6mes propor t ions que pr6c6demment dans un b6cher con tenan t la r6sine dans le tam- pon B. L'agitat ion est r6alis6e m6eaniquement .
III - - Ac~ ty la t i on du c o e n z y m e A.
Avant u t i l i sa t ion la po lyv iny l py r id ine -d iv iny l - benz6ne 2 p. cent, PV4P, (Poly Sciences) est r inc6e par le chloroforme, le m6thanol puts l 'eau. 1,65 g de polym6re sont ajout6s h 40 mg de CoA (acide libre) dissous dans 7 ml d'eau. Apr6s quelques ins tants la mesure en UV h 260 n m de la solution ind ique que tout le CoA a 6t6 absorb6 par la r6- sine. L 'ac6tylat ion est alors effectu6e par trois addi t ions successives de 5 ~I d ' anhydr ide ac6- tique, le m61ange 6tant agit6 manuel lement . Le m6lange est ensuite filtr6 et ]a r6sine r inc6e plusieurs fois h l 'eau distill6e. La r6sine est ensuite vers6 dans 20 ml d 'une solution aqueuse de m6thanol h 25 p. cent. L'ac6tyl CoA est lib6r6 au pH Star (pH < 7,5) par addi t ion de LiOH 0,5 M. Le m61ange est alors filtr6 et le filtrat SOulnis "~ l '6vaporateur rotat if /t 20 ° pour 61trainer le m6- thanol puts au lyophil isateur . Dans ces condi t ions I le CoA est ac6tyl6 quant i ta f ivement (test au DTNB [12]) et son activit6 enzymat ique est de 90 p. cent (citrate synthase [13]).
RESULTATS.
zPm ~IB~ A
s ~ / ~ . . ~ _ _ _ . _ _ ~ _
I 5 I0 ZO 30 40 temps(minutes)
F1G. 3. - - Activit~ de AA(3) r~ticul~e par le 9lutaralddhyde.
(0 0) enzyme purifide ; (O O) enzyme brute ; (A) aetivit6 enzymatique d6tect6e dans la phase li- quide r6siduelle.
Ib : I m m o b i l i s a l i o n sur D E A E cel lulose par le 91utaratd~hyde. L'extrai t bact6rien brut peut 6tre r6ticul6 d i rec tement par le glutarald6hyde con- duisant h un pr6cipit6 actif enzymat iquement , n6anmoins (fig. 3) les condi t ions exp6rimentales
cpm ~ I0 ~ ,~
15 f ~
5
5 I0 ZO 30 40 temps(minutes)
FI6. 4. - - Activitd AAC(3) rdticul~e par le 9lutaralddhyde stzr DEAE cellulose.
(O O) r6sine enzymatique ; (0 O) aetivit6 enzymatique d6tectde dans la phase liquide r6siduelle.
I - - I m m o b i l i s a t i o n de I 'AAC(3).
Ia : I m m o b i l i s a t i o n sur p o l y s a c c h a r i d e r~ t icu l& Par cette technique la r6sine obtenue se r6vble 6tre tr6s peu active enzymat iquement , que l 'on
cprn~103
Z
" ~ 5 10 20 30 temps (minutes)
FIG. 2. - - Activitd de AAC(3) insohlbilis~e sur indubiose A~-triglyeine.
( 0 - - - - 0 ) enzyme purifi6e ; (O O) enzyme brute.
ntil ise tin extrait b~ri~it de AAC(3) (;u Fenzyme pu- riti6e par chromatographie d'affini_6 (fig. 2).
50 ~
1 Z 3 ~ 5 10 mois
Fro. 5, - - Stabilit~ de AAC(3) stock~e & .~°C.
(0 0) enzyme soluble brute ; (0 O) enzyme brute rdticul6e par le glutarald6hyde sur DEAE cellu- lose.
sont relies que seulement une petite part ie de l 'enzyme est incluse i r r6vers ib lement dans la tra- me. Avec l ' enzyme purifi6e, la r6sine obtenue pos- s6de une tr6s faible activit6 enzymatique. D'autre part, les propri6t6s physiques du pr6cipi t6 apr6s
BIOCHIMIE, 1977, 59, n ° 11-12.
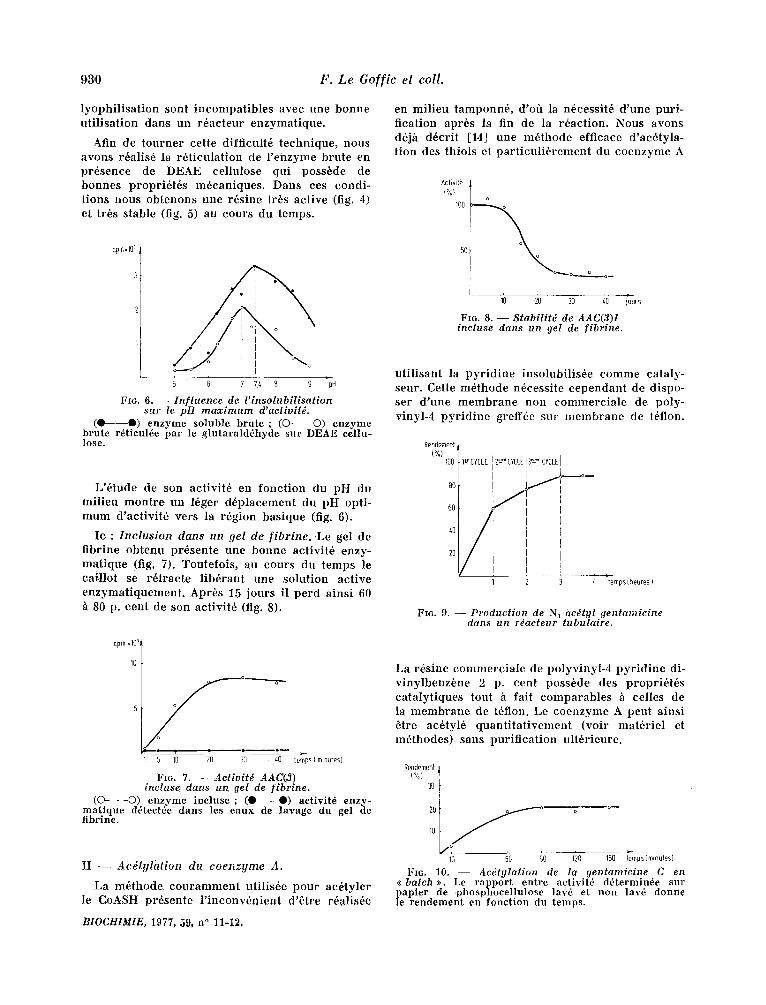
930 F . L e G o f f i c e l co l l .
lyophi l i sa t ion sont incompat ib les avec une bonne ut i l isat ion dans un r6acteur enzymatique.
Afin de tourner cette difficult6 technique, nous avons r6alis6 la r6t iculat ion de l ' enzyme brute en pr6sence de DEAE cellulose qui poss6de de bonnes propri6t6s m6caniques. Dans ces condi- t ions nous obtenons une r6sine tr6s act ive (fig. 4) et trbs stable (fig. 5) au cours du temps.
cpm,103
[ ° o
5 5 7 ~z, 8 g pH
Fro. 6 . - In f luence de l ' insolubi l i sat ion sur le pH m a x i m u m d'acfivi t&
(O O) enzyme soluble bru te ; (O O) enzyme brute r6ticul6e par le glutarald~hyde sur DEAE cellu- lose.
L '6tude de son activit6 en fonct ion du pH du mi l ieu mon t r e un 16ger d6placement du pH opti- mum d 'act ivi t6 vers la r6gion basique (fig. 6).
Ic : I n c l u s i o n clans u n ge l de [ i b r i n e . Le gel de f ibrine obtenu pr6sente une bonne activit6 enzy- mat ique (fig. 7). Toutefois , au cours du temps le cail lot se r6tracte l ib6rant une solution act ive enzymat iquement . Apr6s 15 jours il perd ainsi 60 h 80 p. cent de son activit6 (fig. 8).
en mi l ieu tamponn6, d'ofl la n6cessit6 d 'une puri- fication apr6s la fin de la r6action. Nous avons d6j/i d6cri t [14] une m6thode efficace d 'ac6tyla- t ion des thiols et pa r t i cu l i6 rement du coenzyme A
Activile
IO0
I0 20 30 LO
FIG. 8 . - Stabi l i t6 de AAC(3)I incluse dans un gel de f ibrine.
Jours
uti l isant la py r id ine insolubil is6e comme cataly- seur. Cette m6thode n6cessite cependant de dispo- ser d 'une membrane non eommerc ia l e de poly- vinyl-4 py r id ine greff6e sur membrane de t6flon.
Rendemenl (%)
ioo
'BO
60
@
2[I
WCYCL[ IZt~CY[LE]3~'CYCLEI
I I
1 ¢ temps (heures)
FIG. 9. - - Product ion de N3 ac6tgl gentamicine dans un rdacteur tubulaire.
~prn .10 ~,
10
5 10 70 ?0 . 40 temps (minufes)
Fro. 7. - - Act ivi t6 AAC(3) incluse dans un gel de [ibrine.
(O O) enzyme incluse ; (O O) activit6 enzy- matique d~tectde dans les eaux de lavage du gel de fibrine.
II A c 6 t g l a t i o n d u c o e n z g m e A.
La m6thode couramment utilis6e pour ac6tyler le CoASH pr6sente l ' inconv6nien t d '6tre r6a!is6e
La r6sine commerc ia le de polyvinyl-4 py r id ine di- v inylbenz6ne 2 p. cent poss6de des propri6t6s cata lyt iques tout ~ fait comparables ~ celles de la membrane de t6flon. Le coenzyme A peut ainsi 6tre ae6tyl6 quan t i t a t ivement (voir mat6riel et m6thodes) sans pur i f icat ion ult6rieure.
Rendemenl I
(%) 30 [
10
10 60 ~0 I)0 150 bmps (minutes)
FIG. 10. - - Ac6tylat ion de la gentamieine C en << batch >>. Le rapport entre activitg d6termin6e sur papier de phosphocellulose law" et non lav6 donne le rendement ea fonction du temps.
BIOCHIM1E, 1977, 59, n ° 11-12.
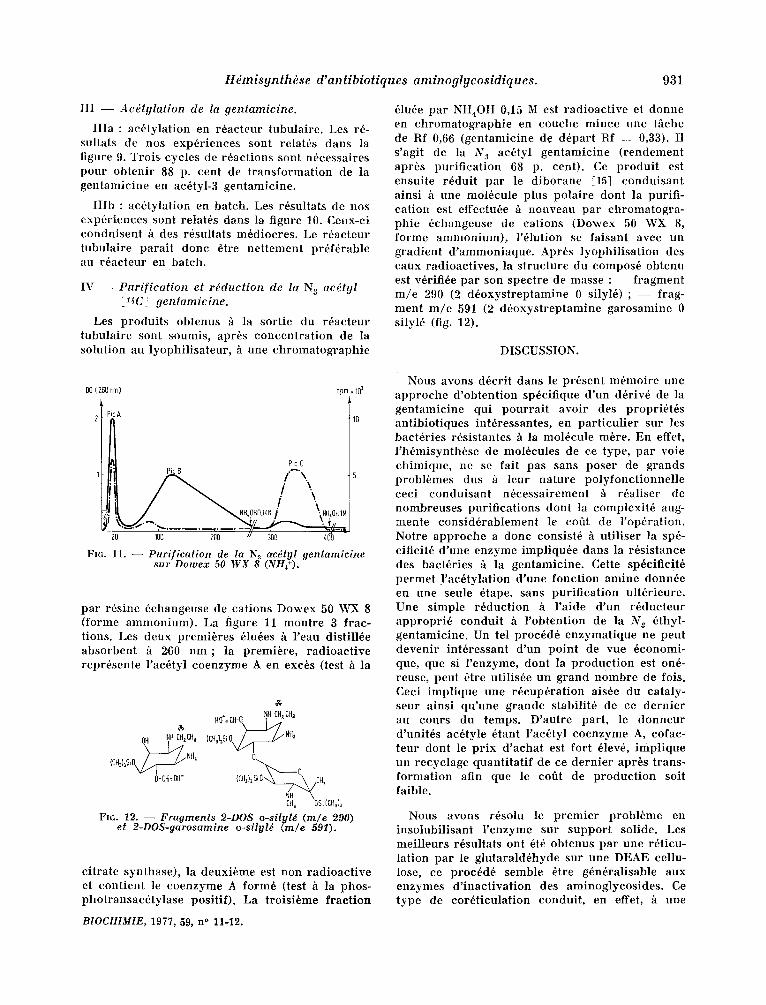
Hdmisynth~se d'antibiotiques aminoglycosidiques. 931
III -- Ac~tglation de la gentamicine.
IIIa : ac6tylat ion en r6acteur tubulaire . Les r6- sultats de nos exp6riences sont relat6s dans la figure 9. Trois cycles de r6actions sont n6cessaires pour obteni r 88 p. cent de t ransformat ion de la gentamicine en ac6tyl-3 gentamicine.
I I Ib : ac6tylal ion en batch. Les r6sultats de nos exp6riences sont retat6s darts la figure 10. Ceux-ci conduisent fi des r6sultats m6diocres. Le r6acteur tubula i re parai t donc Ore ne t tement pr6f6rable au r6acteur en batch.
IV - - Puri f icat ion et r~duction de la N a acOtyl [u,C] gentamieine.
Les produi ts obtenus ~ la sortie du r~acteur tubula i re sont soumis, apr~s concen t ra t ion de la solut ion au lyophil isateur , "~ une chromatographie
61u6e par NH4OH 0,15 M est radioact ive et donne en chromatographie en couche mince une t~che de Rf 0,66 (gentamicine de d6part Rf : 0,33). Il s 'agit de la N s ac6tyl gentamicine ( rendement apr~s puri f icat ion 68 p. cent). Ce produi t est ensuite r~duit par le d iborane [15] conduisan t ainsi h une mol6cule plus polaire dont la purifi- cat ion est effectu6e ~ nouveau par chromatogra- phie 6changeuse de cations (Do~'ex 50 WX 8, forme ammonium) , l'61ution se faisant avec un gradient d ' ammoniaque . Apr6s lyophi l i sa t ion des eaux radioactives, la s t ructure du compos6 obtenu est v6rifi6e par son spectre de masse : - - f ragment m / e 290 (2 d6oxyst reptamine 0 silyl6) ; - - frag- ment m / e 591 (2 d6oxyst reptamine garosamine 0 silyl6 (fig. 12).
DISCUSSION.
O0 (26Ohm)
L
Pica
cpm .I0 3
l0
Pic C Pic B {--'N. 5
/ : , ,! . . . .
Fla. 11. - - Purificalion de la N.~ ae~ly! .qenlamieine sur Dolwex 50 WX 8 (NHF).
par r6sine 6changeuse de cations Do'wex 50 ~VX 8 (forme ammonium) . La figure 11 mont re 3 frac- tions. Les deux premi6res 61u6es h l 'eau distill6e absorbent h 260 n m ; la premibre, radioact ive repr6sente l 'ac6tyl coenzyme A en exc6s (test h la
OH NH CH~CH 3 ~ (CH3laSIOx~ NH2)----V /
(CHASl
0- CH = OH ~ {CHela CH~
~, \OSi(CH,),
Fro. 12. -- Fraymenls 2-DOS o-siIyl~ (m/e 290) et 2-DOS-(larosamine o-silyl6 (m/e 591).
citrate synthase), la deuxibme est non radioact ive et cont ient le coenzyme A form6 (test h la phos- photransac6tylase positif). La troisi6me f rac t ion
Nous avons d6crit dans le pr6sent m6moire une approche d 'obtent ion sp6cifique d 'un d6riv6 de la gentamicine qui pour ra i t avoir des propri6t6s ant ibiot iques int6ressantes, en par t icu l ie r sur les bact6ries r6sistantes h la mol6cule m6re. En effet, l 'h6misynthbse de mol6cules de ce type, par vote chimiquc, ne se fait pas sans poser de grands probl6mes dus h lcur nature polyfonct ionnel le ceci conduisan t n6cessairement h r6aliser de nombreuses purif icat ions dont la complexit6 aug- mente cons id6rablement le coflt de l 'op6ration. Notre approche a donc consist6 ~ ut i l iser la sp6- cificit6 d 'une enzyme impliqu6e dans ]a r6sistance des bact6ries h la gentamicine. Cette sp6cificit6 permct l 'ac6tylat ion d 'une fonct ion amine donn6e en une seule 6tape, sans pur i f icat ion ult6rieure. Une simple r6duct ion fi l 'a ide d 'un r6ducteur appropr i6 condui t h l 'ob tent ion de la N3 6thyl- gentamicine. Un tel proc6d6 enzymat ique ne peut deveni r in t6ressant d 'un po in t de vue 6conomi- que, que si l 'enzyme, dont la p roduc t ion est on6- reuse, peut 6tre utilis6e un grand hombre de lois. Ceci impl ique une r6cup6rat ion ais6e du cataly- seur ainsi qu 'une grande stabilit6 de ce dern ie r au cours du temps. D'autre part , le donneur d 'unit6s ac6tyle 6tant l 'ac6tyl coenzyme A, cofac- teur dont le pr ix d 'achat est fort 61ev6, impl ique un recyclage quant i ta t i f de ce dern ie r aprbs trans- format ion afin que le cofit de product ion soit faible.
Nous avons r6solu le p remier probl6me en insolubi l i sant l 'enzyme sur suppor t solide. Les meil leurs r6sultats ont 06 obtenus par une r6ticu- lat ion par le glutarald6hyde sur une DEAE cellu- lose, ce proc6d6 semble 6tre g6n6ralisable aux enzymes d ' inac t iva t ion des aminoglycosides. Ce type de cor6t iculat ion conduit , en effet, h une
BIOCHIMIE, 1977, 59, no 11-12.

932 F. Le Gof f ic et coll.
r 6 s i n e b i o a c t i v e (fig. 4) p o s s 6 d a n t des p r o p r i d t 6 s m 6 c a n i q u e s c o m p a t i b l e s a v e c s o n u t i l i s a t i o n d a n s u n r 6 a c t e u r e n z y m a t i q u e . D ' a u t r e p a r t , l ' e n z y m e i n s o l u b i l i s 6 e p r 6 s e n t e u n e t r b s b o n n e s t a b i l i t 6 d a n s le t e m p s ( s t o c k a g e h 4°C) p u i s q u ' o n o b s e r v e s e u l e m e n t u n e p e r t e d ' a c t i v i t 6 de 48 p. c e n t au b o u t d u c i n q u i ~ m e m o i s a l o r s que l ' e n z y m e so lu- b le d e v i e n t i n a c t i v e a p r b s q u i n z e j o u r s d a n s les m 6 m e s c o n d i t i o n s (fig. 5).
U n r 6 a c t e u r f o n c t i o n n a n t en f lux s e m i - c o n t i n u a d o n e p u O r e m i s au p o i n t , les p r o d u i t s r6ac - t i o n n e l s 6 t a n t s 6 p a r 6 s p a r r 6 s i n e s 6 c h a n g e u s e s d ' i o n s . D e u x t y p e s de r 6 a c t e u r s o n t 6t6 e n v i s a g 6 s : u n r 6 a c t e u r f e r m 6 , sous a g i t a t i o n m 6 c a n i q u e ou < (ba t ch ~, e t u n r 6 a c t e u r t u b u l a i r e . Ce d e r n i e r s e m b l e 6 t re le p l u s a p p r o p r i 6 p o u r la s y n t h b s e d e n o s p r o d u i t s , le r e n d e m e n t d ' a c 6 t y l a t i o n 6 ran t de 88 p. c e n t a p r b s t r o i s r e c y c l a g e s a u l i eu de 20 p. c e n t a v e c le r 6 a c t e u r en ~ b a t c h ~ (fig. 9 et 10). A p r ~ s s 6 p a r a t i o n de la Ns a c 6 t y l g e n t a m i c i n e , le c o e n z y l n e A p e u t 6 t re ac6 ty l6 q u a n t i t a t i v e m e n t p a r la m 6 t h o d e d 6 c r i t e d a n s ce m 6 m o i r e , p r 6 s e n . r a n t l ' a v a n t a g e de n ' a v o i r a u c u n e p u r i f i c a t i o n u l t 6 r i e u r e h e f f ec tue r .
B i e n q u e la r 6 d u c t i o n a i t l i eu en p h a s e h 6 t 6 r o - g~ne (N~ a c 6 t y l g e n t a m i c i n e i n s o l u b l e d a n s T H F ) ce l le -c i es t q u a n t i t a t i v e et la s t r u c t u r e d u c o m p o s 6 a 6t6 v6r i f i6e p a r s on s p e c l r e de m a s s e (fig. 12).
O u t r e le p o u v o i r a n t i b i o t i q u e p o t e n t i e l d e ce c o m p o s 6 , ce d e r n i e r p o u r r a n o u s r e n s e i g n e r s u r l ' i m p o r t a n c e de la f o n c t i o n a m i n e en 3 d u c y c l e 2,-DOS p o u r l ' a c t i v i t 6 b i o l o g i q u e des a m i n o g l y c o - s ides .
D ' u n p o i n t de v u e g6n6ra l , ce t y p e de r 6 a c t e u r p e u t 6 t r e p r a t i q u 6 a v e c les d i f f 6 r e n t e s e n z y m e s d ' i n a c t i v a f i o n a c 6 t y l a n t les a m i n o g l y c o s i d e s en des p o s i t i o n s d i f f 6 r e n t e s te l q u e AAC(2 ' ) et AAC(6 ' ) . L ' o b t e n t i o n de s d 6 r i v 6 s N 6thy16s c o r r e s - p o n d a n t s p e r m e t t r a d ' 6 t u d i e r la r e l a t i o n s t r u c - t u r e - a c t i v i t 6 de ces m o l 6 c u l e s .
R~suM~.
Afin d 'ob ten i r des d~riv~s N-monoalkylSs d ' amino- glycosides, h bu t th~rapeut ique , nous u t i l i sons les propri~t~s sp6cifiques des enzymes d ' inac t iva t ion de ces ant ib io t iques . Une ~tude d ' immobi l i s a t i on d 'nne de ces enzymes snr suppor t polym~rique est r6aIis~e, d'oit il r~sulte que la r~t iculat ion sur DEAE cellulose par le g lu tara ld~hyde est le mei l leur proc~dC La r~ac- t ion enzymat ique a y a n t l ieu en prSsence d'AcCoA, comme agent d 'ac~tylat ion, le probl~me de la r~g~n~- r a t i on de ce cofacteur a ~t4 abord4 et une m~thode simple d 'acfi tylation du CoA u t i l i s an t un ca ta lyseur polym~rique insoluble , le po lyv iny l -4-pyr id ine-d iv iny l benz6ne 2 p. cent a ~t~ raise au point. Enfin, une ~tude compara t ive mon t r e que le r~acteur tubu la i r e donne les mei l leurs r endemen t s d 'ac~tylat ion.
BIBLIOGRAPHIE.
1. Umez~wa, S. (1974) Adv. in carbohydrate chemistry and biochemistry, 30, 111-164, Tipson, R. S. and Horton, D. Editor, Academic Press.
2. Kawagochi, H., Naito, T., N~kawa. G. ~ Fujisa~va, K. (1972) J. Antibiot., ~), 695-708.
3. Le Goffic, Martel, A. & Witchi tz , J. (1974) Antimi- crob. Agent. Chemoth., 6, 680-684.
4. Chevereau, M., Daniels, P. J. L., Davies, J. & Le Goffic, F. (1974) Biochemistry, 13, 598-602.
5. Le Goffic, F. & Martel, A. (1974) Biochimie, 56, 893-897.
6. Zaborsky, O. R. (1973) Immobilized enzymes, ~Verst, R. C. Editor, C.R.C. Press.
7. Mosbach, K. (1976) Methods in enzymology, vol. XLIV, Colo'wigl~, S. P., Kaplan, N. O. Editor , Academic Press Ne'w Yor~.
8. Bright , H. ~ Pora th , J. (1974) Enzyme engineering, 2, 183-237.
9. Le Goffic, F., Moreau, N. ~ Cheverean, M. (1973) Biochimie, 55, 1183-1186.
10. Benveniste , R. & Davies, J. (1971) Biochemistry, 10, 1787.
11. Marca, G. S., Pari'k, H. J. & Cuatrecasas, P. (1974) Anal. Biochem., 60, 149-152.
12. Gruner , T. R. P. ,& Phi l ips , P. H. (1951) Arch. Bio- chem., 30, 217.
13. Iau, P. T., Hylary, R. T., David, A. P. S. ~ Bot- teomley, R. G. (1974) Biochem. J., 139, 609-623.
14. Le Goffic, F., Sicsic, S. & Vincent, C. (1976) Tetra- hedron Letters, 33, 2845-2847.
15. Brown, H. C . .a Zweifel, G. (1963) Organic Reac- tions, 13, 32, J. Wiley, Editor, Academic Press New York.
BIOCHIMIE, 1977, 59, n ° 11-12.













