Embed Size (px)
Citation preview

Biochimie 70 (1988) 145-165 © Soci6t6 de Chimie biologique/Elsevier, Paris 145
Review
Hydration of transfer RNA molecules" a crystallographic study
Eric WESTHOF, Philippe DUMAS and Dino MORAS
Laboratoire de Cristailographie Biologique, lnstitut de Biologie Mol~cu/aire et Cellulaire, Centre National de la Recherche Scient(fique, 15, rue R.-Descartes, F-67084 Strasbourg Cedex, France
(Received 6-5-1987, accepted after revision 2-9-1987)
Summary - Four crystal structures of transfer RNA molecules were refined at 3 ,A resolution with the inclusion of the solvent molecules found in the difference maps: yeast tRNA-phe in the ortho- rhombic form, yeast tRNA-phe in the monoclinic form and yeast tRNA-asp in the A and B forms. Over 100 solvent molecules were located in each tRNA crystal. Several hydration schemes are found repeatedly in the 4 crystals. The tertiary interactions in the corner of the L-shaped molecule attract numerous solvent molecules which bridge the ribose hydroxyl 0(2') atoms, base exocyclic atoms and phosphate anionic oxygen atoms. Conservation of bases leads to conservative localized hydra- tion patterns. Several solvent molecules are found stabilizing unusual base pairs like the G-U pairs and those involving the pseudouridine base. Water bridges between the 0(2') and the exocyclic atom 02 of pyrimidines or the N3 atom of purines are common. Water bridges occur frequently between successive anionic oxygen atoms of each strand as well as between N7 or other exocyclic atoms of successive bases in the major groove. Magnesium ions or spermine molecules are found to bind in the major groove of tRNA helices without specific interactions.
hydration / solvent / tRNA / X-ray crystallography
Introduction
Knowledge of the aqueous hydration of biolo- gical macromolecules is essential for the understanding of the tertiary structures of biomolecules, the interactions between them and their functions. Usually, tRNA crystals have a high solvent content, between 60 and 75% [1]. Most of the solvent molecules occupy the solvent channels between the loosely packed tRNA molecules, exchange dynami- cally with the mother liquor surrounding the crystal and can be considered in a state identi- cal to bulk phase solution. These solvent mo le - cules lead to featureless electron density and contribute predominantly to the low-angle region of the diffraction pattern [2, 3]. That region is usually excluded from the data used in the refinement [4]. A small percentage of the
crystal solvent is crystallographically ordered and forms the hydration shell or the bound water of the macromolecule. The definition of bound water is necessarily operational and depends upon the technique used to determine it [5]. With X-ray crystallography, one can iden- tify localized hydration sites, that is sites on the macromolecule surface which are locally stable so that these positions are, on an average, frequently occupied by solvent molecules. Depending upon the temperature factor and the occupancy of the site, one speaks of a tightly or weakly bound solvent molecule, although these two parameters reflect the attraction potential of the site for solvent mole- cules.
The amount of information about the solvent structure around a macromolecule that one can extract from the diffraction data

146 E. Westhof et al.
depends strongly upon the resolution of the data and on the state qf the refinement of crystal structure. At 2-3 A resolution, only the strongly bound solvent molecules are expected to be extracted [6, 7]. In the present analysis, data between 10 and 3 A resolution were used. Therefore, one aim of the study was to compensate for the relatively low resolution by refining and analyzing data from 4 different tRNA crystals and from 3 different laborato- ries. The same refinement program was used, but 2 methods for extracting the solvent peaks were employed. This was done, since it is known that the interpretation of solvent density involves some degree of subjective judgment (e.g. [8]). With these approaches, it was hoped that the analysis and the compari- sons of the solvent structures in the 4 tRNA crystals would ),ield a meaningful and consis- tent picture of the organization of solvent molecules around tRNA molecules.
The 4 crystals of yeast tRNA molecules analyzed were: yeast tRNA-phe in the ortho- rhombic form, yeast tRNA-phe in the mono- clinic form and yeast tRNA-asp in the A and B forms. The diffraction data for the ortho- rhombic form were from S.H. Kim's laboratory and the starting coordinates as well [9, 10]. The monoclinic form of yeast tRNA-phe [11, 12] was refined using the data from M. Sundaralin- gam's laboratory [13]. For the B-form of yeast tRNA-asp, the already published refinement [14] was pursued in order to extract the solvent
molecules. The details of the ref inement of the A-form of yeast tRNA-asp has recently been described (P. Dumas, E. Wes thof and D. Moras, submitted). Several results of these comparative refinements have already been addressed and discussed [15, 16]. Here, the solvation of these 4 tRNA molecules are described, compared and discussed.
Crystallographic refinement
The refinement techniques used for these 4 tRNA crystals have been described ([13, 14, 17], P. Dumas et al., submitted). For the refinement of the 4 crystals, about the same number of data (see Table I) between 10 and 3 A, were used [16]. ,In principle [18], with interplanar spacings of 3 A, details separated by more than 2.1 A can be seen. Other factors will, however, increase this value so that it may be difficult to resolve peaks involved in H-bonds, which are typically separated by 2.6-3.1 ~,. Usually, the assign- ment of solvent peaks to particular chemical entities is made on the basis of the geometric parameters associated with the peak. However, at 3 A resolution, water molecules cannot be distinguished from, say, an ammon ium ion. Sodium or magnesium ions can be assigned in terms of distances and coordination. At best, we might thus expect to observe the more ordered solvent sites and some local water networks in crevices or channels - " " - O l t i l e i i l i : l . l , , ; iu-
molecular surfaces without any specific details
Table I. Total and average numbers of contacts made by the water molecules with other water molecules, with the nucleic acid, the phosphates, the sugars, and the bases.
Crystal To water To nucleic acid Phosphates Sugars Bases
tRNA-phe 40 198 85 47 66 ortho 0.4 1.9 0.8 0.5 0.6 tRN A-phe 60 181 79 31 71 mono 0.7 2.0 0.9 0.3 0.8 t RNA-asp 71 216 89 41 86 B-form 0.6 1.9 0.8 0.4 0.7 tRNA-asp 32 97 55 13 29 A-form 0.4 1.1 0.6 0.2 0.3 mean 0.5 1.7 0.8 0.35 0.6 A-DNA 176 110 57 10 43 octamer 2.0 i.3 0.7 0.1 0.5
The values for the A-DNA octamer are based on a structure from [23]. The total number ofwater molecules determined in each structure, in the order ofthe table, are : 104, 9 I, ! 16, 85 and 86. The total number of reflections used and the R-factor, in the same order are : 4585, 4335, 4508, 40 ! 9 and 18.8%, ! 9.4%, 1.7.2% and ! 7,1% at 3 A, resolution for the 4 tRN A structures, For the, A-ONA octamer, 4428 reflections between 10 and !.7 A reflections were used and the final R-factor was 15% [231.
Hydration of tRNA 147
on the water structure itself. In addition, Savage [19] has remarked that the use of O...O distance criteria alone for H-bonding may be misleading, since he observed water molecules or hydroxyl groups partially surrounded by water molecules that do not form H-bonds to the enclosed species despite intermolecular distances between 3.1 and 3.5 ,~.
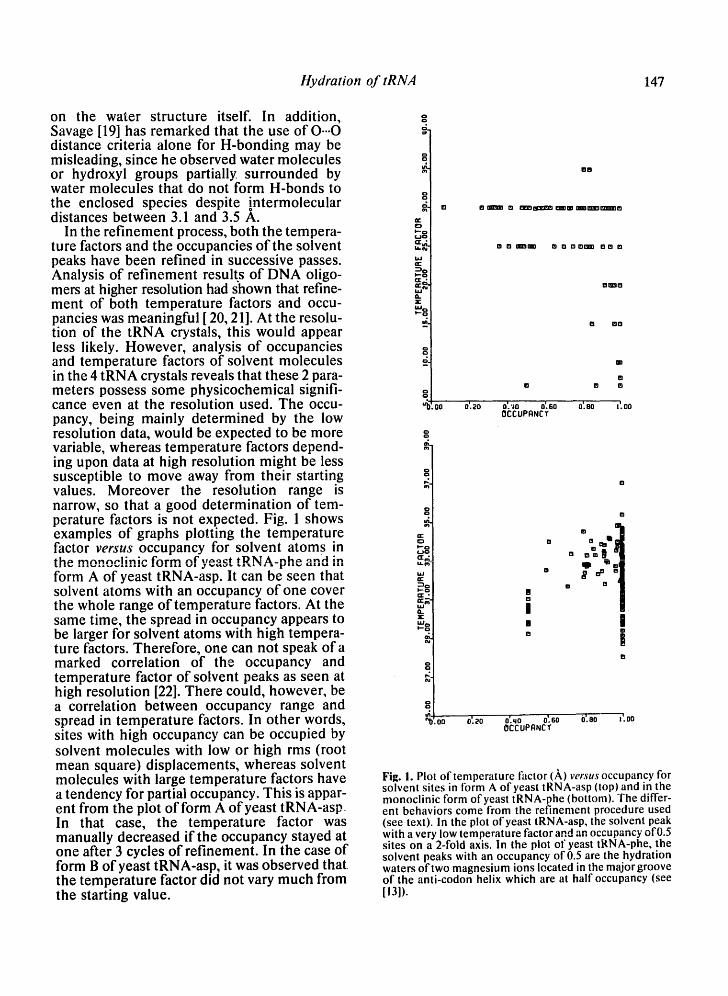
In the refinement process, both the tempera- ture factors and the occupancies of the solvent peaks have been refined in successive passes. Analysis of refinement results of DNA oligo- mers at higher resolution had shown that refine- ment of both temperature factors and occu- pancies was meaningful [ 20, 21]. At the resolu- tion of the tRNA crystals, this would appear less likely. However, analysis of occupancies and temperature factors of solvent molecules in the 4 tRNA crystals reveals that these 2 para- meters possess some physicochemical signifi- cance even at the resolution used. The occu- pancy, being mainly determined by the low resolution data, would be expected to be more variable, whereas temperature factors depend- ing upon data at high resolution might be less susceptible to move away from their starting values. Moreover the resolution range is narrow, so that a good determination of tem- perature factors is not expected. Fig. 1 shows examples of graphs plotting the temperature factor versus occupancy for solvent atoms in t h ~ m n n n c J J n i c f n r m n f ' w ~ ¢ t tDl~l A_nh~- on~1 ~n i l i i . Z l i % J ,17 W I ~ , I J I ~ L I • • I [~11~11 i l l
form A of yeast tRNA-asp. It can be seen that solvent atoms with an occupancy of one cover the whole range of temperature factors. At the same time, the spread in occupancy appears to be larger for solvent atoms with high tempera- ture factors. Therefore, one can not speak of a marked correlation of the occupancy and temperature factor of solvent peaks as seen at high resolution [22]. There could, however, be a correlation between occupancy range and spread in temperature factors. In other words, sites with high occupancy can be occupied by solvent molecules with low or high rms (root mean square) displacements, whereas solvent molecules with large temperature factors have a tendency for partial occupancy. This is appar- ent from the plot of form A of yeast tRNA-asp. In that case, the temperature factor was manually decreased if the occupancy stayed at one after 3 cycles of refinement. In the case of form B of yeast tRNA-asp, it was observed that the temperature factor did not vary much from the starting value.
g
g g m
g i - - o 1 . ,~o
h i a " "-~¢
I---¢
O .
-2.
d~
,00
O i . - o ¢.)L~ r r , , i u - t ~
W a--
~ O I - - O
a . x
g
g
0'.ao
B 8
• B DI B ~ ~ Im~'zm'm'mO
• o rmp'zmm ~ zg 2 I O ~ D m 0 In
mmngl
i~ om
0 m
o'. '~o o'. so o'. eo 0 C C U P f l N C g
B
t'.oo
0
0 m 8
B B
! |
o'.ao o'.~o o'.so O'.eo :'.on OCCUPANCY
Fig. I. Plot of temperature factor (,~,) v e r s u s occupancy for solvent sites in form A ofyeast tRNA-asp (top) and in the monoclinic form ofyeast tRNA-phe (bottom). The differ- ent behaviors come from the refinement procedure used (see text). In the plot of yeast tRNA-asp, the solvent peak with a very low temperature factor and an occupancy of 0.5 sites on a 2-fold axis. In the plot of yeast tRNA-phe, the solvent peaks with an occupancy of 0.5 are the hydration waters of two magnesium ions located in the major groove of the anti-codon helix which are at half occupancy (see [13]).
148 E. Westho. f et al.
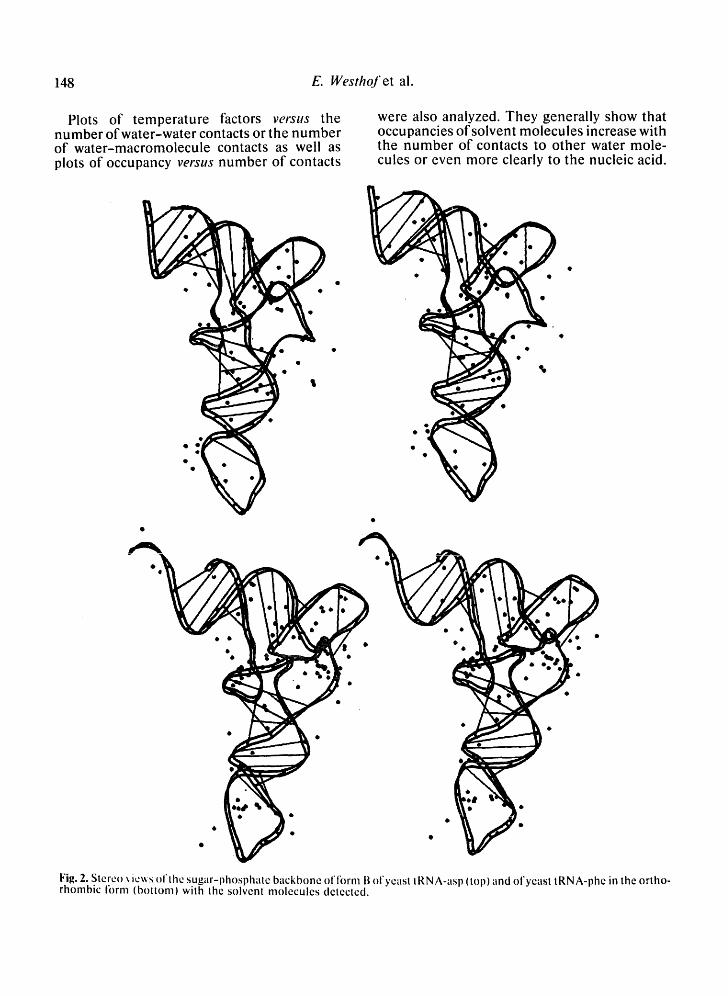
Plots of tempera ture factors versus the number of water -water contacts or the number of water-macromolecule contacts as well as plots of occupancy verstts number of contacts
were also analyzed. They generally show that occupancies of solvent molecules increase with the number of contacts to other water mole- cules or even more clearly to the nucleic acid.
• Qt
Fig. 2. Stereo ~ ie~s ol'lhe sugar-phospllate backbone of fornl B ol'yeasl tRNA-asl~ (top) and ol'yeast tRNA-phe in the ortho- rhonlbic form (bottom) wilh the solvent molecules detected.
H y d r a t i o n o f t R N A 149
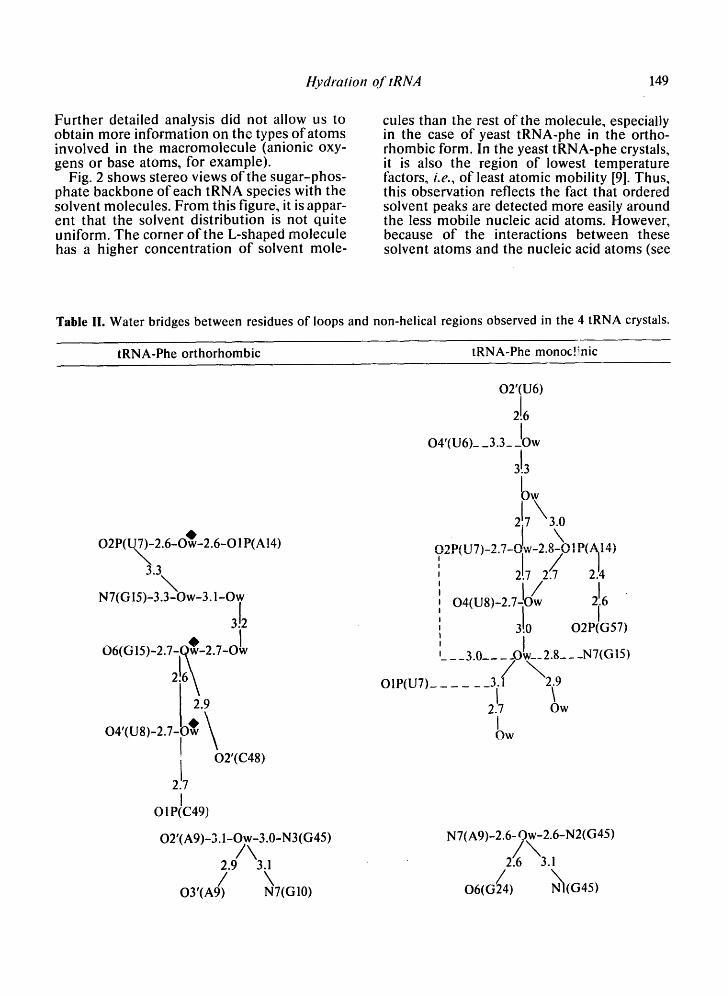
Further detailed analysis did not allow us to obtain more information on the types of atoms involved in the macromolecule (anionic oxy- gens or base atoms, for example).
Fig. 2 shows stereo views of the sugar-phos- phate backbone of each tRNA species with the solvent molecules. From this figure, it is appar- ent that the solvent distribution is not quite uniform. The corner of the L-shaped molecule has a higher concentration of solvent mole-
cules than the rest of the molecule, especially in the case of yeast tRNA-phe in the ortho- rhombic form. In the yeast tRNA-phe crystals, it is also the region of lowest temperature factors, i . e . , of least atomic mobility [9]. Thus, this observation reflects the fact that ordered solvent peaks are detected more easily around the less mobile nucleic acid atoms. However, because of the interactions between these solvent atoms and the nucleic acid atoms (see
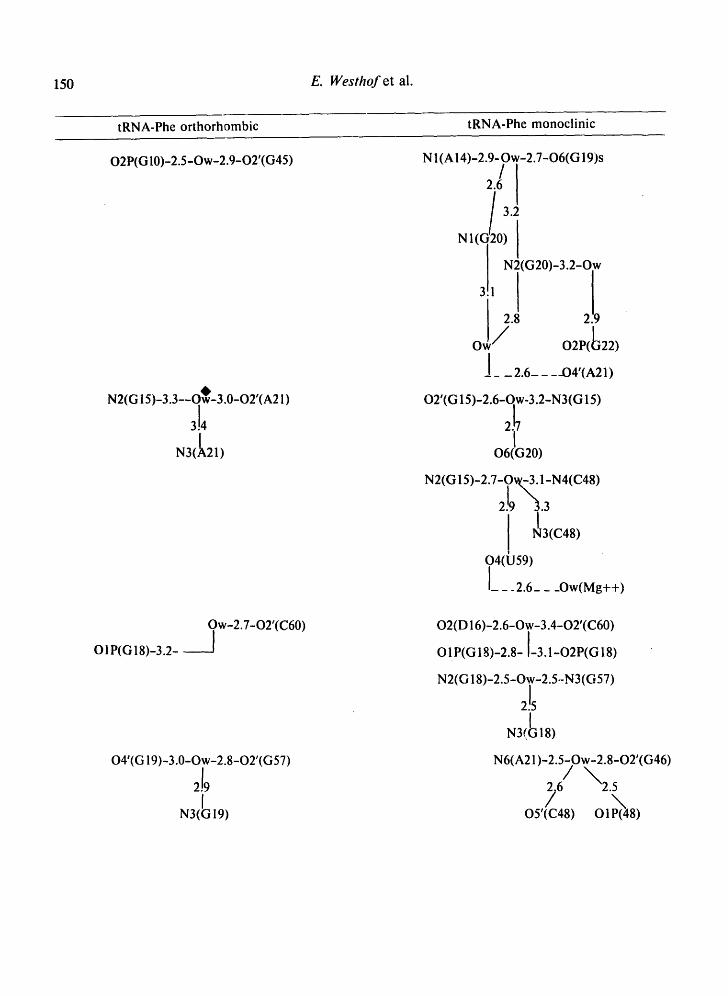
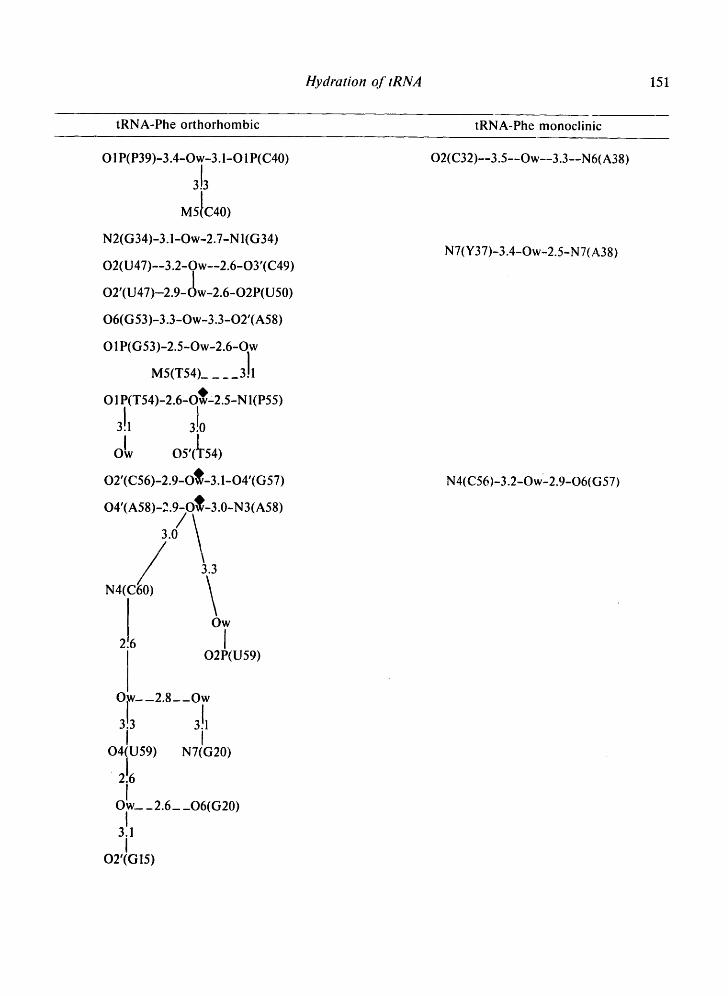
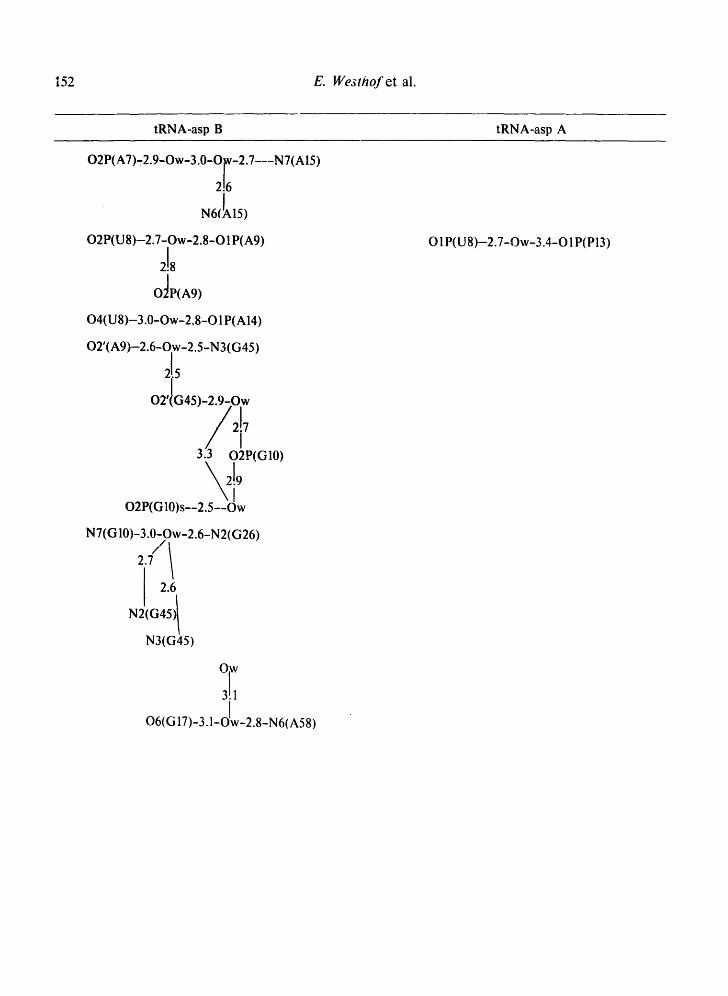
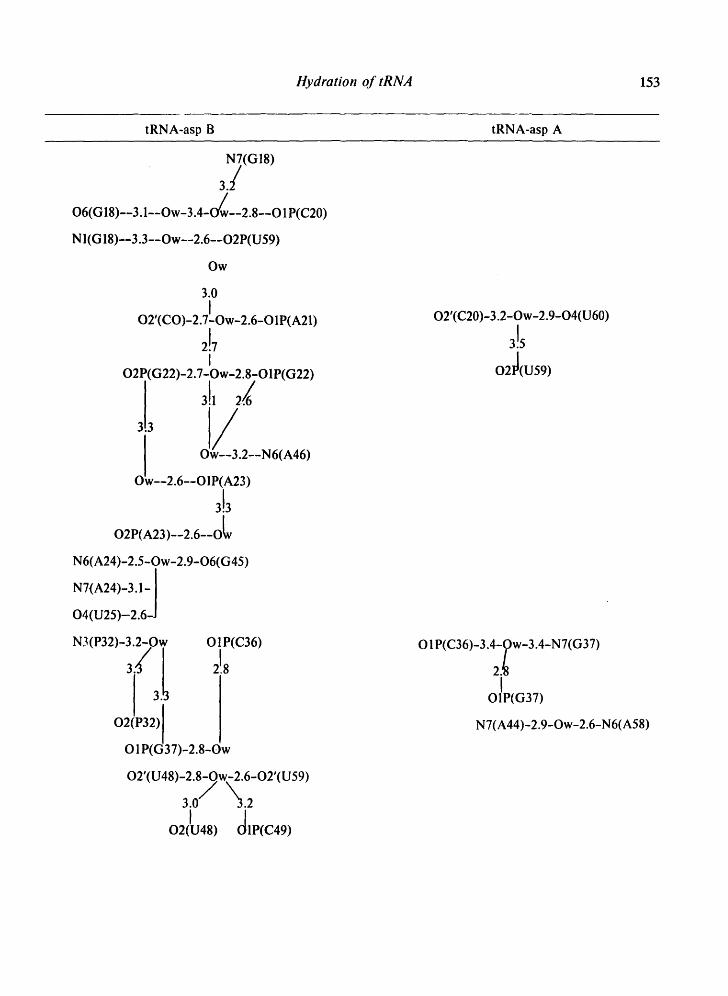
Table II. Water bridges between residues of loops and non-helical regions observed in the 4 tRNA crystals.
tRNA-Phe orthorhombic tRNA-Phe monoc!inic
O2 P(UN7)-2.6- O~-2.6-O 1P(A14)
"~.3
N7(G 15)-3.3"~-Ow-3.1-Ow
3!2 o6(o15)-2.7-o* -2.7-o
2!6 2.9
O4'(U8)-2.7- 9~ ~
02'(C48)
O1P(C49)
O2'(A9)-3.1-Ow-3.0-N3(G45) / \
2.9 3.1 / \
OY(A9) NT(GI0)
OIP(U7)_
O2'(U6)
2!6 /
O4'(U6)_ _3.3_ _lOw
3!3 bw
• I \ n ~ D / I T ' l ~ ", "I r~. . , '~ 8 n ~ p ~ 14' v _ , , , - , , , - ,- . , - , -T, , - ,~ . ? , - , , , ~ ,
2!7 2".7 2.'4
O4(U8) 2 7 IO/w 216
_ __ 3.0__ __Olw__ 2.8__ -N7(GIS)
.3.1 / \ 2 . 9 2.17 Ow
t OW
N7( A9 )-2.6-Ow-2.6-N2(G45)
/6\3, o6,o2/4,
150 E. Westhof et al.
tRNA-Phe orthorhombic tRNA-Phe monoclinic
02 P(G 10)-2.5-Ow-2.9-O2'(G45)
N2(G 15)-3.3--OwO-3.0-O2'(A21)
3!4 N3(IA21)
N 1 (A 14)-2.9- Ow-2.7-O6(G 19)s
~l 3"2
NI( I 0)N!(G20)_3.2_O. w
o// o~(b,~) J_ _ 2.6-- --O4'(A21)
O2'(G 15)-2.6-qw-3.2-N3 (G 15)
I O6(G20)
N2(G 15)-2.7-0~-3.1-N4(C48)
~!9~
O4(U59) I__ _2.6__ _Ow(Mg++)
O 1P(G 18)-3.2- ~ w-2.7-02'(C60)
O4'(G 19)-3.0-Ow-2.8-O2'(G57)
2!9 N3(~G 19)
O2(D 16)-2.6-Ow-3.4-O2'(C60)
OIP(G18)-2.8-I-3.1-O2P(G18)
N2(G 18)-2.5-Ow-2.5.-N3(G57) 2!s
N3(~18)
N6(A21)-2.5-Ow-2.8-O2'(G46) / ,~6 ~ 2 5
O5'(C48) O1P~((48)
Hydration 03" tRNA 151
tRNA-Phe orthorhombic tRNA-Phe monoclinic
O 1P(P39)-3.4-Ow-3.1-O 1P(C40)
3!3
M5tC40)
N2(G34)-3.1-Ow-2.7-Nl(G34)
O2(U47)--3.2-Ow--2.6-03'(C49)
O2'(U47)-2.9- (~w-2.6-O2P(U50)
O6(G53)-3.3-Ow-3.3-O2'(A58)
O 1P(G53)-2.5-Ow-2.6-Ow
M5(T54)_ ___ 311.
O 1P(T 54 )-2.6-OOw z2.5-N I(P55) 3!1 3!o OIw O5',~54)
O2'(C56)-2.9-O~-3.1-O4'(G57)
O4'(A58)-2.9-O~-3.0-N3(A58)
3.0 /
I ow 2.6 I
02P(U59)
O,w-_2.8__Ow
3!3 3!1 I I
O4(U59) N7(G20)
2!6 JW__ __2.6__ _O6(G20)
I 3.1 I
O2'(G15)
O2(C32)--3.5--Ow--3.3--N6(A38)
N7(Y37)-3.4-Ow=2.5-N7(A38)
N4(C56)-3.2-Ow-2.9-O6(G57)
152 E. Westhof et al.
tRNA-asp B tRNA-asp A
O2P(A7)-2.9-Ow-3.0-O~-2.7---N7(AI5)
216
N6(IAIS)
O2P(U8)-2.7-Ow-2.8-01 P(A9)
2!8 O~P(A9)
O4(U 8)-3.0-Ow-2.8-O 1 P(A 14)
O2'(A9)-2.6-Ow-2.5-N3(G45)
O2P(G10)s--2.5--Ow
N7(GI0)-3.0-Ow-2.6-N2(G26) / t 27" / 2.6
N2(G45) /
N3(G45)
O,w
3!1 !
O6(G 17)-3.1-OIw-2.8-N 6(A58)
O 1P(U8)-2.7-Ow-3.4-O 1 P(P13)
Hydration of tRNA 153
tRNA-asp B tRNA-asp A
N7(G18)
3.:/ O6(1318)--3. l--Ow-3.4-O/w--2.8--O 1 P(C20)
NI(G 18)--3.3--Ow--2.6--O2P(U59)
Ow
3.0 , I
02 (CO)-2.7-Ow-2.6-O1P(A21)
2!7 I
O2P(G22)-2.7-Ow-2.8-O1P(G22)
oL--~.6--o1~;21;- 3!3
O2P(A23)--2.6--OL
N6(A24)-2.5-Ow-2.9-O6(G45)
N3(P32)-3.2-Ow O1P(C36)
O 28
(G37)-2.8-Ow
O2'(U48)-2.8-Qw-2.6-O2'(U59)
3.0 / "3.2 I (~1P(C49) 02(U48)
O2'(C20)-3.2-Ow-2.9-O4(U60)
3!s O2~(U59)
O 1P(C36)-3.4-Ow-3.4-N 7(G37)
I O1P(G37)
N7(A44)-2.9-Ow-2.6-N6(A58)

154 E. Westhof et al.
tRNA-asp B tRNA-asp A
O21P55 )-3.2-Ow-2.7-O 1 P(A58) /
3.2 I
Ow
O2(C56)-3.3-Ow-2.6-N3(C56)
3!3 I
N3(A57)
O2'(A58)-2.8-Ow-3.1-O1P(U59) I I
2.9 2.6 I I
Ow Ow
O2P(U59)-3.2-Ow-2.9-04(U60)
O2(U60)-3.4-Ow-2.6-O5'(DI6)
3Jl I
O2P(G 18)-2.7-Ow-3.2-O2'(D 16)
For the yeast tRNA-phe crystal in the orthorhombic form, a dark diamond indicates those bridges within !.7 ,~, in the present re-refinement and in the previous refinement [9].
Table II), some of these peaks must contribute also to the stabilization of that region rich in tertiary interactions and base-pairing.
The interactions between the solvent mole- cules and the nucleic acid atoms will now be described in more detail. In Table I, the average numbers of contacts made by the water mole- cules with other water molecules and with the nucleic acid are given for the 4 crystal forms and for an A-DNA octamer [23]. The low number ofcontacts with other water molecules reflects the low resolution of the tRNA data. The similarity between the average numbers of contacts to nucleic acid atoms, not only in the four crystal forms but also in the A-DNA crystal (1.7 ,~ resolution), gives some confi- dence in the analysis. The average number of contacts made by the water molecules to the sugar atoms is 3 times larger in the tRNAs than in the A-DNA octamer, naturally because of the presence of the ribose hydroxyl 0(2'). For
the A-form of yeast tRNA-asp the average values are less than those of the other tRNAs. This reflects the method employed for picking up the solvent peaks (P. Dumas et ai., sub- mitted), which puts more emphasis on contacts between the macromolecule and the solvent than on contacts between the solvent peaks themselves. With the experience gained in the refinement process and ana:ysis of the results, the following directions for searching solvent peaks at such resolution can be given. First, it is worthwhile to refine alternatively occupancy and temperature factor of solvent peaks. Secondly, the temperature factor has to be manually decreased either if the occupancy stays at 1.0 after cycles of refinement or accord- ing to the average temperature factor of the atoms with which the solvent peaks interact. And thirdly, the search for new solvent peaks has to be made on the basis of contacts not only with the macromolecule but also with the

Hydration of tRNA 155
already determined solvent peaks. The differ- ence maps should also be frequently searched and viewed for large or longish peaks, some- times without direct contact with the macro- molecule, to pick up molecules like spermine.
Tables II-IV present the bridges and i:ontacts between the solvent molecules and the 4 tRNA molecules in the helical stems (Table III), in the loop regions (Table II), and between symmetrically-related molecules (Table IV). In Tables II and IV, the water bridges are arranged with increasing residue number and so as to be on the same level in different structures.
It is interesting to compare the positions of the solvent molecules determined in the pres- ent refinement of the orthorhombic form of yeast tRNA-phe and in the previous one, for which coordinates of 83 solvent positions have been deposited at the Brookhaven Data Bank [9]. Of those solvent peaks, 50 are within 1.7 A in the two refinements with the 4 magnesium sites within 0.6 ,A. Besides the hydration waters
Table II1. Some observed one-water bridges it: tRNA crystals.
of the magnesium ions, 15 of the peaks within 1.7 ,A are bound to anionic phosphate oxygen atoms and 10 to hydroxyl 0(2') atoms. Thus, excepting the magnesium ions and their waters of hydration, about 40% of the solvent peaks are common to both refinements. The water bridges found in the two refinements are marked by a dark diamond in Tables II and IV.
Similar values were reached after other comparative refinement studies. Thus, the re- refinement of the B-dodecamer d ( C - G - C - G - A - A - T - T - C - G - C - G ) indicated that at most, half of the assigned solvent peaks were common to both refinements, with the hydra- tion spine down the minor groove particularly well conserved [24]. The latter point was also noted after the refinement of the same dodeca- mer with a G - A pair [25]. Similarly, a compar- ison of two independently refined models of ribonuclease A at 2.0 A resolution showed that just over half of the solvents were in similar positions (within 1.9 ,A) with almost all of the best determined waters in the first shell of hydration [26]. That small percentage of the crystal solvent, which is crystallographically ordered and forms the first hydration shell or the bound water of the macromolecule, is espe- cially crucial for understanding the role of water in the structure and stability ofbiomole- cules.
O2'(i)---Ow---O2(i) ( 11 ) O2'(i)---Ow---N3(i) (16) O2'(i)---Ow---O4'(i + I) (20)
N2(i)---Ow---N3(i) (8)
O 1P(i)---Ow---N 7(i) (9) OlP(i)---Ow---OIP(i + 1) (10) OY(i)---Ow---O2P(i + 1) (8) OIP(i)---Ow---O2P(i + 1) (7)
O 1P(i)---Ow---O2 P(i) (30) O 1 P( i)---Ow---05'( i ) (16) O2P(i)---Ow---O5'(i) (13)
N7(i)---Ow---N6(i) N7(i)---Ow---O6(i) O4,N4(i)---Ow---O6,N6(i + 1) N7(i)---Ow---N7(i + 1)
The total number of occurences is given between paren- theses, except for those involving exocyclic polar atoms of the bases due to their great variety and, consequently, weak statistical weight. Those shown on the last 4 lines are most probably, due to the resolution, two-water bridges instead of one-water bridges.
Contacts within helical stems
Some of the water contacts in the helical Stems (Table III) are found repeatedly. They occur in water bridges involving either the ribose hydroxyl 0(2') and 02 of pyrimidine bases (N3 of purine bases) or the 0(2') atom and the ribose endocyclic oxygen 0(4') of the next residue. They also occur in water bridges involving the phosphate oxygens and will be described below. It has already been noticed that the hydroxyl oxygen atom 0(2') of a sugar residue often forms a hydrogen-bond to the sugar ring oxygen atom 0(4') of the next resi- dues when the two residues belong to a helical stem [10, 14, 27]. In Table III, it can be seen that, not unfrequently, a water molecule links these two atoms. Often, the same bridging water molecule also links the hydroxyl 0(2') to the N3 (purine) or the 02 (pyrimidine) of the attached base (Fig. 3, top). This pattern was often seen in smaller nucleic acid fragments [28-30]. Also, in the structure of r(ApU) [31], a

156 E. Westhof et al.
water molecule links the hydroxyl 0(2') to the ring 0(4') of the next residue. Water molecules binding in that small cavity of the minor groove certainly contribute to the stabilization of the A-type helix typical of RNA molecules. Para- doxically, such water molecules might play a crucial role in the non-enzymatic hydrolysis of RNA molecules by participating, under appro- priate conditions, in the removal of a proton from 0(2') which can then attack the phospho- diester linkage.
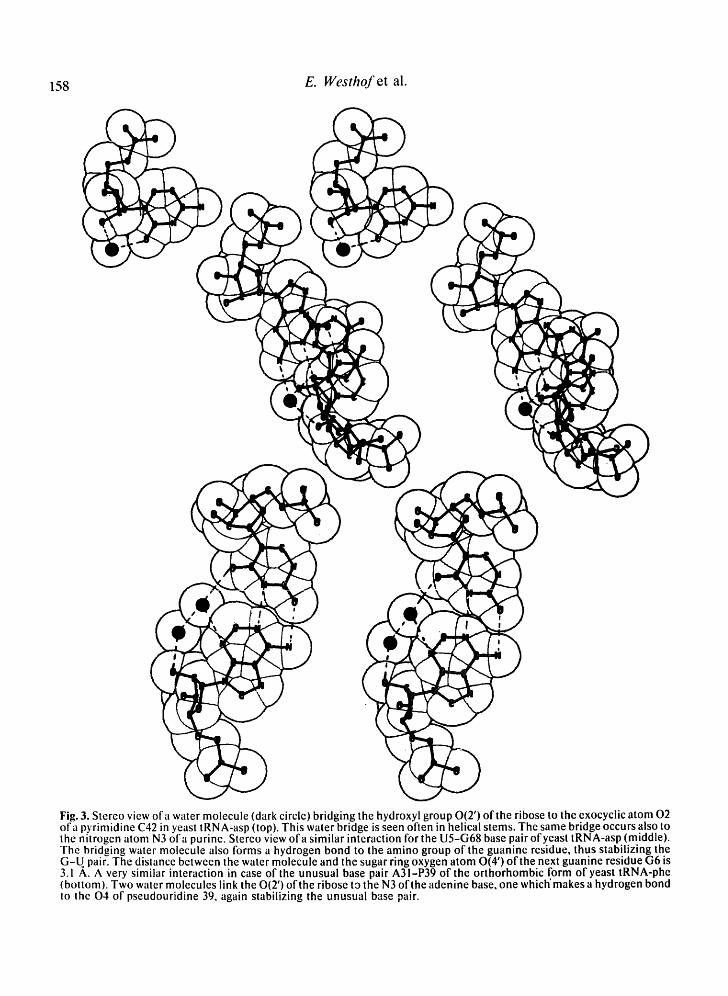
It is interesting that most of the G-U pairs of the 4 structures have these water bridges (Fig. 3, middle). With a G-U pair, the minor groove is slightly deeper and water molecules tend to be stabilized in such a cavity (for a similar effect, see the case of the Z-DNA oligomers [20]). Also, in the cases of two G-U pairs, the water molecule hydrogen-bonded to the 0(2') and 02 of the uracil makes an additional hydro- gen-bond to the amino group N2 of the gua- nine (Fig. 3, middle). Similarly, the unusual base
Table IV. Water bridges between symmetrically-related tRNA molecules found in the 4 crystal structures.
tRNA-Phe orthorhombic tRNA-Phe monoclinic
N2(G4)-3.0-O~'-2.7-O2'(U52) (1,-100)
O2'(U6)-3.0-Ow-3.3-Ow-3.3-O2P(U52) (1,-100)
O2P(G 15)-2.5-OOw-2.7-O3'(C56)
314" (1,-100)
I O2P(G57) (1,-100)
O4'(G20)-2.8-Ow-2.9-O4(D 16)
O2'(G20)
O1P(C56) (3,-100)
O2'(C56)-3.0-Ow-2.6-Ow(Mg++) (3,000)
O2(C 13)-2.9-Ow-3.3-N3 (D 17)
.Jlt (2,2-11 ) 3
I t " ~ , " ~ l l ' ~ , " I x l , , ' ,b ~ '11 ! x
u ~ t u l !1 tz ,z- [ l)
O2(C27)-2.5-Ow-2.6-N3(C75)
(2,1-11) 3 . / ~ 4 ' 2:8
/ \ O2'(C27) N4(C75) (2,1-11)

H y d r a t i o n o f t R N A 157
Table IV continued.
tRNA-asp B
O2'(G4)-2.7-Ow-2.8-O2P(G41) (6,000)
O2P(G 10)-2.9-Ow-2.4-O2P(G 10) (2,000)
O2(C49)-2.9-Ow-3.0-O4'(U 11 ) (2,000)
3.3
N2(G 10) (2,000)
O2'(C56)-2.7-Ow-2.9-O3P(U 1 ) (5,- 100)
O2'(C67)-2.9-Ow-2.9-O2P(U 12) (2,000)
2.8
O2(C67)
O 1P(G68)-2.9-Ow-3.2-O4'(D 19) (5,0-10)
tRNA-aspA
O2P(G 10)-2.5-Ow-2.5-O2P(G 1 O)
(2,000)
O4'(C29)-3.2-Ow-3.2-N3(D16) (6,-100)
For the yeast tRNA-phe crystal in the orthorhombic form, a dark diamond indicates those bridges within 1.7 ,A in the present re-refinement and in the previous refinement [9]. In yeast tRNA-asp, the solvent peak linking two symmetrically-related O2P atoms of residue G10 was assigned an occupancy of one-half. That peak could well be an ammonium ion. Notice how the solvent-mediated links between symmetry-related molecules building up the crystal are well preserved in the two refinements of the orthorhombic form yeast tRNA-phe and in the two forms ofyeast tRNA-asp. For the symmetry operation to be applied on the second residue, the first number refers to one of the transformations below and the three subsequent numbers to the translation along x, y, z. For the orthorhombic form: (i) x, y, z; (2) -x, y, -z: (3) 0.5 + x. -y. 11.5 --: (4~ 0.5 -x. -.v 0.5 + z. For the monoclinic form: (1) x, y, z; (2) -x, 0.5 + y, -z. For tRNA -asp: (1) x, y, z; (2) x, -y, -z; (3) -x. -y, 0.5 + z; (4) -x , y, 0 .5-z; (5) 0.5 + x, 0.5 + y, z; (6) x + 0.5, 0.5 -y, -z; (7) 0.5 -x, 0.5 -y, 0.5 + x; (8) 0.5 -x, 0.5 + y, 0.5 -z.
pair be tween aden ine 31 and pseudour id ine 39 of yeast t R N A - p h e presents , on the adenine site, a bridging water molecule which is linked to an addit ional water molecu le hydrogen- bonded to the exocyclic oxygen a tom 0 4 of the pseudour id ine in the a n t i conformat ion (Fig. 3, bot tom). Such a water molecule might stabilize the a n t i conformat ion of the pseudour id ine base. N M R results have been interpreted as indicating al ternat ively a s y n conformat ion [32-34] or an a n t i c o n f o r m a t i o n [35]. Close association o f water molecules with the func- tional groups of misma tched bases was also observed in D N A ol igomers [36, 37]. Because of the absence of the hydroxyl 0(2 ' ) a tom, hydrat ion is extensive on the major groove side in D N A with a water molecule br idging the N2 of guanine and the 0 2 of thymine . It has been
sugges ted that free energy differences between matched and mismatched nucleot ides are magnif ied at the D N A polymerase active site by exclusion of water a round the base pairs, thereby encouraging fidelity [38].
Wate r molecules interacting with 0(2 ' ) and 0(4 ' ) o f the sugar, N3 of purines or 02 of pyri- midines form the main bridges in the minor and shallow groove of the tRNA molecules. Several times, the guanine amino group is also involved in water bridges, especially when the guan ine is at the end of s tems or in single- s t randed regions.
In the major groove, several exocyclic amino or carbonyl groups hydrogen bond to water molecules , but do not frequently form an extens ive solvation network. Most o f those water molecules l inked to the major groove
158 E. Westhof et al.
J
( (
Fig. 3. Stereo view of a water molecule (dark circle) bridging the hydroxyl group 0(2') of the ribose to the exocyclic atom 02 ofa pyrimidine C42 in yeast tRNA-asp (top). This water bridge is seen often in helical stems. The same bridge occurs also to the nitrogen atom N3 ofa purine. Stereo view of a similar interaction for the U5-G68 base pair ofyeast tRNA-asp (middle). The bridging water molecule also forms a hydrogen bond to the amino group of the guanine residue, thus stabilizing the G-U pair. The distance between the water molecule and the sugar ring oxygen atom 0(4') of the next guanine residue G6 is 3.1 A. A very similar interaction in case of the unusual base pair A31-P39 of the orthorhombic form of yeast tRNA-phe (bottom). Two water molecules link the 0(2') ofthe ribose ta the N3 ofthe adenine base, one which'makes a hydrogen bond to the 04 of pseudouridine 39, again stabilizing the unusual base pair.
Hydration o./" tRN,4 159
sites form intra- or inter-strand bridges (purine N7/O6/N6 to either purine N7/O6/N6 or pyri- midine N4/O4). These water bridges can involve up to 4 bases belonging either to 2 consecutive base pairs or to each strand but staggered, tRNA-asp, which has a sequence of four G-C base pairs in the thymine stem, presents (especially in form A) the interesting situation of 2 separate water networks without much interaction between them, one bonding
to the guanine strand and the other to the cytosine strand. In A-DNA oligomers, one- water bridges are generally not frequent, espe- cially in the major grove; instead two-water bridges are observed [21]. In this study, mainly one-water bridges have been observed. We suspect that these one-water bridges are an arti- fact due to the low resolution, which tends to smear the peaks so that two-water bridges may appear as one-water bridges.
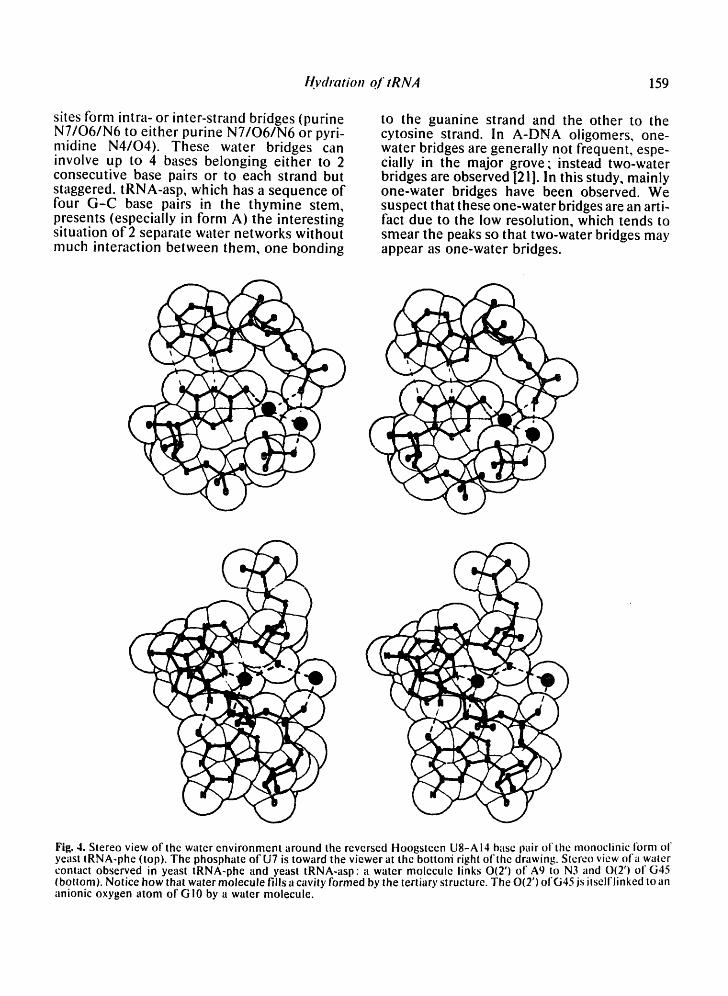
Fig. 4. Stereo view of the water environment around the reversed Hoogsteen U8-A 14 base pair of the monoclinic i'orm of yeast tRNA-phe (top). The phosphate of U7 is toward the viewer at the bottoni right ofthe drawing. Stereo view of a water contact observed in yeast tRNA-phe and yeast tRNA-asp: a water molecule links O(2') of A9 to N3 and 0(2') of G45 (bottom). Notice how that water molecule fills a cavity formed by the tertiary structure. The O(2') ofG45 js itselflinked to an anionic oxygen atom ot'G 10 by a water molecule.

160 E. Westhof et al.
Solvent contacts in the loops and tertiary interactions
The solvent contacts in the non-helical regions are quite interesting, on the one hand, by their variety, and on the other hand, by their posi- tions. Some bridging water molecules are found in the two types of tRNA molecules studied. In the comparison of crystallographic structures, one should keep in mind that the hydration networks of nucleic acids are very dynamic. Further, variations or rearrange- ments in local geometry, as well as changes in crystallization conditions, can lead to changes in the water structure. The occurrence of two identical hydration schemes in different crys- tals is therefore of significance; for example, the water molecule linking 04 of U8 to the anionic oxygen O1P of AI4 in the reversed- Hoogsteen U8-A14 base pair (Fig. 4, top). This water molecule helps stabilize the reversed- Hoogsteen pair at this tight corner of the mole- cule. It would be interesting to analyze the effect of a 4-thiouridine at position 8 with the presence of a bridging water molecule. In the monoclinic form of yeast tRNA-phe, this bridg- ing water molecule is linked to two other water molecules, one of which is hydrogen bonded to O2P of U7 and the other one to N7 of G15.
Three other water bridges are found in yeast tRNA-phe and in yeast tRNA-asp. One links 0(2') of A9 with N3 of G45; the same water
molecule or another one links N7 of G10 to N3 of G45. And the third one links O2P of G10 with 0(2') of G45. Thus, the base triple between base pairs 10-25 and base 45, which is held only through the interactions between the amino group of G45 and the O6/N7 of G !9, is further stabilized by at least 2 water molecules (see Fig. 4, bottom). This base triple is impor- tant for the overall stability of the tRNA mole- cule, since it is at the interface between the dihydrouridine stem and the anti-codon stem. In the two types of tRNA molecules, there is also a water molecule bridging the exocyclic group at position 6 of the conserved purine 24 with guanine 45, which is also a conserved purine.
Other water bridges are found in similar posi- tions in the 4 structures; for example, between pseudouridine 55 (N1 or 02) and an anionic oxygen atom of either thymine 54 or the base 58 (Fig. 5). Notice in Fig. 5 how water mole- cules, anionic oxygen atoms, and carbonyl oxygen atoms surround the methyl group of thymine T54. As already suggested [39], the presence of a water molecule on the pseudou- ridine 55 might explain the peculiar NMR effects [39, 40] observed on the N1 proton of tile pseudouridine 55 upon removal of magnesi- um ions or of spermine. Most of these water molecules clearly help stabilize tight turns or loops. Such is the case of the water molecule(s) bridging O2P of U59 and the exocyclic atom at
Fil~. 5. Stereo.view of the envirc, nment of the methyl group (dark triangle) of thymine 54 in the orthorhombic form ofyeast tRNA-phe. A water molecule b~ idges N! ofpseudouridine 55 to an anionic oxygen atom ofT54 and is at a distance of 3.4 A from tin anionic oxygen of P55. Also, two water molecules help form a ring oofoxygen atoms around ihe methyl group. The distance between the second water molecule and the methyl group is 3.1 A.

Hydration of tRN,4 161
position 4 of base 60 (see Table II). The hydroxyl atom 0(2') makes a large
number of direct intramolecular contacts, thereby playing a crucial role in the stabiliza- tion of the secondary and tertiary structures [10, 14, 27]. Above, it was already noted that water molecules might participate in these intramolecular contacts. Many water bridges involving the hydroxyl 0(2') atom in Table II reinforce the role of the 0(2') atom in the stabi- lization of the tertiary folding. Another example is shown in Fig. 6 where a water mole- cule which fits snugly into a cavity of the struc- ture hydrogen bonds to three residues: the amino group of A21, the hydroxyl group of G46, and an anionic oxygen atom of C48.
In the structure of form B of yeast tRNA-asp, the most complex solvent network involves residues 20-21-22-23 and 46 and 6 solvent molecules. Such a network is not observed in the yeast tRNA-phe structures around that region. It has been observed that phosphate 22 is more accessible to ethylnitrosourea in tRNA-phe than in tRNA-asp [41]. The water network observed in the crystal might, contrib- ute to the protection of phosphate 22. In the monoclinic form of yeast tRNA-phe, the most complex network involves nine water mole- cules and residues 6, 7, 8, 14 and 15. In the orthorhombic form, there are 2 networks, each comprising 5 water molecules; one links 15, 20, 58, 59 and 60 and the other 7, 8, 15, 48 and 49. These networks are detailed in Table !!.
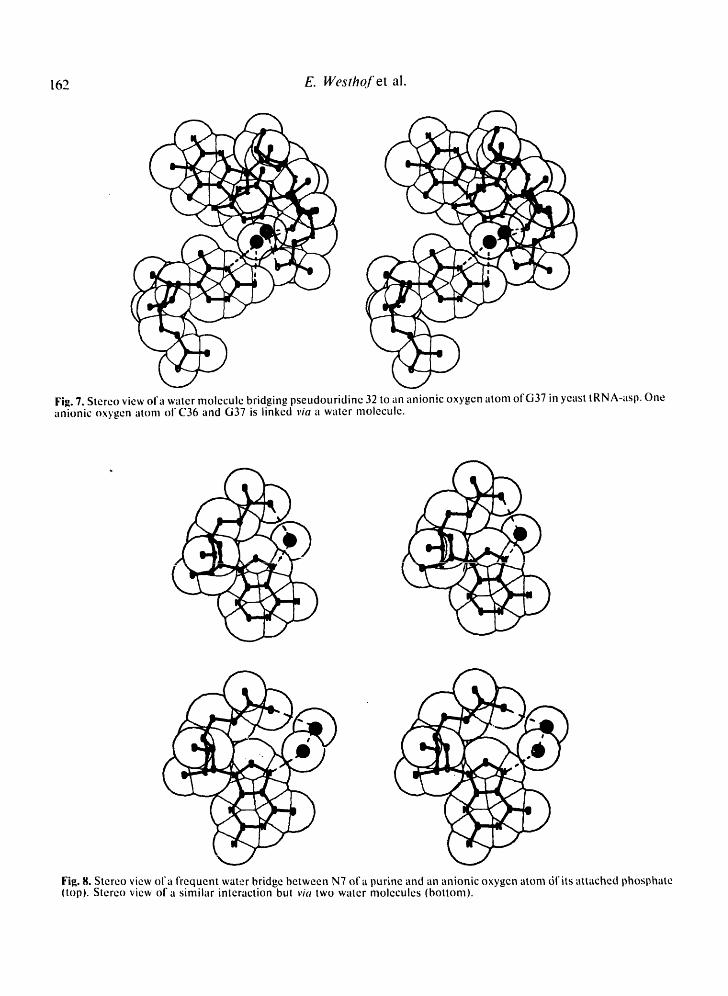
The special hydration of the pseudouridine base is again exemplified in yeast tRNA-asp for residue 32, where a water molecule links residue 32 to an anionic oxygen of base 37 (Fig. 7). This site is a gold site which was used
in the structure solution stage. Interestingly, in the orthorhombic form of yeast tRNA-phe, the same site is occupied by a water molecule of the coordination sphere of the magnesium ion bound to the anti-codon loop. Another such example is described below. In the monoclinic form of yeast tRNA-phe, there is also such an example of a water site which is occupied by a lanthanide ion in crystals soaked with lantha- nide salts. This water molecule links N 1 of G20 to 0(4') of A21.
Contacts involving the phosphate group anionic oxygen atoms
In Tables I1 and III, the participation of the anionic oxygens of the phosphate grou0 is apparent. First, in helical stems, the anionic oxygen O1P interacts frequently with the N7 of punne residues through one (Fig. 8, top) or two water molecules (Fig. 8, bottom). Such a scheme has been observed in nucleotide struc- tures (e.q., [42]). It should be noted that no water bridge between the ribose hydr,q×y! 0(2') and an anionic oxygen of the 3'-phosphate was found. Such bridges havebeen suggested on the basis of NMR experiments [43]. In the case of pyrimidines, instances have been found of a water bridging the anionic oxygen O1P to the C5 or C6, or the methyl group on C5. In the B-dodecamer fAA] .~,o, . . . . . . . . . ,- t-'-'l, a . a t e , I l l U i I ~ I , , U I C iS ---t~4"---- O l t e l l
lodged between the methyl groups of thymine residues and their attached phosphates: There is also the interesting example of the methyl group of thymine 54 linked to an anionic oxygen of the preceding residue by two water
(
Fig. 6. Stereo view of the stack G46-A21-C48 in the monoclinic form of yeast tRNA-phe. Notice how a water molecule, filling a cavity, forms hydrogen bonds to N6 of A21, 0(2') of G46, and O I P of C48.
162 E. Westho.f et al.
( (
Fig. 7. Stereo view of a water molecule bridging pseudouridine 32 to an anionic oxygen atom of G37 in yeast tRNA-asp. One anionic oxygen atom of C36 and G37 is linked via a water molecule.
Fig. 8. Stereo view of a frequent water bridge between N7 ofa purine and an anionic oxygen atom dfits attached phosphate (top). Stereo view of a similar interaction but via two water molecules (bottom).

Hydration of tRNA 163
molecules (see Fig. 5). A characteristic of the contacts in the non-helical regions is that both the hydroxyl atom 0(2') and the anionic oxygen atoms are frequently involved in water bridges. In yeast tRNA-asp, the water molecule bridging the anionic oxygen atoms of residues 8 and 9 is replaced by a gadolinium ion after soaking the crystal in gadolinium sulfate. As above for the water molecule linking 32 and 37, this water molecule belongs to the coordina- tion sphere of the magnesium ion bound to the P10-1oop in the orthorhombic crystal of yeast tRNA-phe.
Water molecules very often bridge phos- phate atoms of the same ~-esidue or of adjacent residues on the same strand (Table III). The latter bridges are frequent in crystals of DNA oligomers in the A forms [23, 45]. Water bridges to phosphates are also involved in strings of water molecules which can span 4 or 5 residues across the major groove from one strand to the other in the 5'-direction. Some examples of these strings can be seen in A form tetramer structure [46]. The water molecules take the shortest path from one strand to the other, as is observed in loop formation by unpaired residues [47].
Intermolecular contacts mediated by water mo|ecnh~
In the orthorhombic form of yeast tRNA-phe and in yeast tRNA-asp, several intermolecular contacts are mediated by water molecules (Table IV). Again, several of these contacts involve either the 0(2') atom or an anionic oxygen atom. The least amount of intermole- cular water bridges is found in the monoclinic form of yeast tRNA-phe. It is also the form with the smallest number of direct intermolecular contacts.
The magnesium ions and spermine molecules
Four magnesium ions were localized in yeast tRNA-phe [10, 11, 48, 49]: one ion binds fully hydrated to phosphates 8, 9, 11 and 12; one binds directly to O2P of G19 with its water molecules linking residues 15, 20, 56 (symmet- rically-related), 59 and 60, thus stabilizing the
interactions between the thymine and the dihydrouridine loops ; one ion binds directly to the anionic oxygen atoms of residues 20 and 21; and finally, there is a magnesium ion interact- ing through its water molecules with residues 31,32, 37, 38 and 39 of the anti~codon loop. The same locations were found in the monoclinic form of yeast tRNA-phe with variants in the anti-codon loop [10, 13]. In the structure of yeast tRNA-asp, no magnesium ion sites could be detected with certainty, although there is a possible site in the anti-codon loop. In this regard, one should mention that these crystals were grown in high ionic strength and, thus, it is not unexpected that the magnesium ions should be quite mobile and not visible in differ- ence maps. As mentioned above, two water sites in yeast tRNA-asp are identical with a water site of two hydrated magnesium ions. In tRNA-phe crystals, spermine molecules could not be identified, although close to a site in the major groove of the anti-codon stem previously assigned to a spermine molecule [10, 48], two magnesium ions that occupy one-half were identified in both forms. In tRNA-asp, a sper- mine molecule was positively identified in the major groove of the amino acid stem (P. Dumas et aL, submitted). Both results indicate that large counter-ions bind in the major groove of RNA helices and that specific interactions are not necessary for binding. Such conclusions were also reached on the basis of theoretical calculations [50, 51].
Conclusions
A large number of solvent molecules have been detected in each tRNA crystal. Despite the low resolution, several hydration schemes were found repeatedly in all 4 crystals or in the 2 types of transfer RNA molecules. Thus, the conservation or semi-conservation of bases in transfer RNA molecules leads to conservative localized hydration patterns. The role of some water molecules is clearly related to a stabiliza- tion of the secondary and tertiary interactions, since these water molecules bridge ribose hydroxyl 0(2') atoms, base exocyclic atoms and phosphate anionic oxygen atoms, which other- wisewould not be hydrogen bonded. This is quite apparent in the case ofunusual base pairs,

164 E. Westhof et al.
where bonds to water molecules tend to make for the loss of hydrogen bonding. The neglect of some of these localized water molecules can lead to distortion of the local environment during restrained least-squares refinement. A recent study showed that the extent and the location of the solvent shell around a transfer RNA molecule depends upon the solvent envi- ronment [52]. In 0.1 M NaCl, an excess mass of about 85 water molecules was determined [52]. Between 71 and 108 water molecules were local- ized in the present study and one can wonder whether some of these water molecules do not belong to the solvent shell of the polyion. The presence of numerous water molecules in the core of the elbow of the tRNA molecules should be taken into account when considering hinge flexibility in transfer RNA molecules
"[53]. Also, the frequent occurence of some hydration sites and water bridges may prevent straightforward interpretations of NMR results.
It is known that RNA structure is quite insensitive to variations in the degree of hydra- tion but that magnesium ions are crucial for the maintenance of the tertiary structures of the complex structures of tRNAs and ribosomal RNAs. However, as described above, water molecules bound inside pockets formed by loops and stretches of the polynucleotide back- bone contribute further to the stabilization of the sh'ucture. A theoretical study has also emphasized the stabilizing effect of bound
~1.1 **at~ to loop ut;tuI C:~ [2qJ. I nese water moie- cules localized in notches and caviti s of the tertiary structure formed by the polynucleotide folding can be regarded as an integral part of the tRNA structure itself.
Acknowledgments
We are grateful to Professors S.H. Kim (Berkeley University) and M. Sundaralingam (University of Wisconsin) for giving us their X-ray data on, respec- tively, the orthorhombic and monoclinic forms of yeast tRNA-phe. This work would not have been possible without their collaboration. We thank R.E. Dickerson (UCLA), W.A. Hendrickson (Columbia University), W.K. Olson (Rutgers University), G.J. Quigley (MIT), and S.T. Rao (University of Wisconsin) for the use of their programs and for discussions.
References
1 Dock A.C., Lorber B., Moras D., P. ixa G., Thierry J.C. & Giege R. (1984) Biochimie 66, 179-201
2 Moews P.C. & Kretsinger R.H. (1975), J. Mol. BioL 91,201-228
3 Phillips S.E.V. (1980), J. Mol. Biol. 142, 531-554 4 Hendrickson W.A. (1981) #1 : Refinement of Pro-
tein Structures (Machin P.A., Campbell J.W. & Elder M., eds), S.E.R.C. Daresbury Laboratory, Warrington, pp. 1-8
5 Texter J. (1978) Prog. Biophys. Mol. Biol. 33, 83-97
6 Finney J.L. (1979) in: Water: A Comprehensive Treatise (Franks F., ed.), vol. 6, Plenum Press, New York, pp. 47-122
7 Blake C.C.F., Pulford W.C.A. & Artymiuk P.J. (1983) J. MoL Biol. 167, 693-723
8 North A.C.T. & Smith J.C. (1985) Int. J. Biol. Macromol. 7, 223-225
9 Sussman J.L., Holbrook S.R., Wade-Warrant R., Church G.M. & Kim S.H. (1978) J. Mol. Biol. 123, 607-630
10 Holbrook S.R., Sussman J.L., Wade-Warrant R. & Kim S.H. (1978) J. Mol. Biol. 123, 631-660
11 Hingerty B., Brown R.S. & Jack A. (1978) J. Mol. Biol. 124, 523-534
12 Stout C.D., Mizuno H., Rao S.T., Swaminathan P., Rubin J., Brennan T. & Sundaralingam M. (1978) Acta Crystallogr. Sect. B 34, 1529-1544
13 WesthofE. & Sundaralingam M. (1986) Biochem- isuy 25, 4868-4878
14 WesthofE., Dumas P. & Moras D. (1985) J. Mol. Biol. 184, 119-145
15 Dumas P., Ebel J.-P.~ Giege R., Moras D., Thierry J.C. & Westhof E., (1985) Biochimie 67, 597-606
16 Westhof E., Dumas P. & Moras D. (1988) Acta Crystallogr. Sect. A (in press)
17 Hendrickson W.A. & Konnert J.H. (1980) in: Biomolecular Structure, Function, ConJormation and Evolution (Srinivasan R., ed.), Vol. 1, Perga- mon Press, Oxford, pp. 43-57
18 Blundell T.L. & Johnson L.N. (1976) in: Protein Crystallography Academic Press, New York, pp. 117
19 Savage H.F. (1986) l,l, ater Sci. Rev. 2, 1-82 20 Westhof E., Prange T., Chevrier B. & Moras D.,
(1985) Biochimie 67, 811-817 21 WesthofE., (1987) Int. J. Biol. Macromol. 9, 186-
192 22 Watenpaugh K.D., Margulis T.N., Sieker L.C. &
Jensen L.H. (1978) J. Mol. Biol. 122, 175-180 23 Kennard O., Cruse W.B.T., Nachman J., Prange
T., Shakked Z. & Rabinovich D. (1986)J. Biomol. Struct. Dyn. 3, 623-647
24 Westhof E. (1987) J. Biomol. Struct. Dyn. 5, 581- 600

Hydration o f tRN~4 165
25 Hunter W.N., Brown T. & Kennard O. (1986) J. Biomol. Struct. Dvn. 4, 173-191
26 Wlodawer A., ,Borkakoti N., Moss D.S. & Howlin B., (1986) Acta Crystallogr. Sect. B 42, 379-387
27 Jack A., Ladner J.E., Rhodes D., Brown R.S. & Klug A. (1977) J. MoL Biol. 11 !, 315-328
28 Rosenberg J.M., Seeman N.C., Day R.O. & Rich A. (1976) J. Mol. Biol. 104, 145-167
29 WesthofE., Rao S.T. & Sundaralingam M. (1980) J. MoL Biol. 142, 331-370
30 Wang A.H.J., Fujii S., van Boom J.H., van der Marel G.A., van Boeckei S.A.A. & Rich A. (1982) Nature 299, 601-605
31 Seeman N.C., Rosenberg J.M., Suddath F.L., Kim J.J.P. & Rich A. (1976) J. Mol. Bio-I. 104, 109- 144
32 Hurd R.E. & Reid B.R. (1977) Nucleic Acids Res. 4, 2747-2755
33 Roy S., Pe.pastavros M.Z. & Redfield A.G. (1982) Nucleic Acids Res. 10, 8341-8349
34 Heerschap A., Haasnoot C.A.G. & Hilbers C.W. (1983) Nucleic Acids Res. 11, 4483-4499
35 Heerschap A. (1984) Thesis (University of Nijme- gen)
36 Kneale G., Brown T., Kennard O. & Rabinovich D. (1985) Jr. Mol. Biol. 186, 805-814
37 Wang A.H.J., Gessner R.V., van der Marel G.A., van Boom J.H. & Rich A. (1985) Proc. Natl. Acad. Sci. USA 82, 3611-3615
38 Petruska J., Sowers L.C. & Goodman M.F. (1986) Proc. Natl. Acad. Sci. USA 83, 1559-1562
39 Tropp J. & Redfield A.G. (1981) Biochemistry 20, 2133-2140
40 Heerschap A., Haasnoot C.A.G. & Hilbers C.W.
(1983) Nucleic Acids Res. 11, 4501-4520 41 Romby P., Moras D., Bergdoll M., Dumas P.,
Vlassov V.V., Wes.thof E., Ebel J.P. & Giege R. (1985) J. MoL BioL 184, 455-471 "
42 Rao S.T. & Sundaralingam M. (1969) J. Am. Chem. Soc. 91, 1210-1217
43 Bolton P.H. & Kearns D.R. (1979) J. Am. Chem. Soc. 101,479-484
44 Drew I-!.R. & Dickerson R.E. (1981) J~ MoL BioL 151,335-556
45 Saenger W., Hunter W.N. & Kennard O. (1986) Nature 324, 385-388
46 Conner B.N., Yoon C., Dickerson J.L. & Dicker- son R.E. (1984) J. MoL Biol. 174, 663-695
47 Haasnoot C.A.G., Hilbers C.W., van der Marel G.A., van Boom J.H., Singh U.C., Pattabiraman N. & Kollman P.A. (1986) J. BiomoL Struct. Dyn. 3, 843-857
~8 Quigley G.J., Teeter M.M. & Rich A. (1978) Proc. Natl. Acad. Sci. USA 75, 64-68
49 Teeter M.M., Quigley G.J. & Rich A. (1980) in: Nucleic Acid-Metal Ion Interactions (Spiro T.G., ed.), Wiley-lnterscience, New York, pp. 145-177
50 Lavery R., Pullman A., Pullman B. & De Oiiviera M. (1980) Nucleic Acids Res. 8, 5095-5111
51 Lavery R. & Pullman B. (1981) Nucleic Acids Res. 9, 4677-4688
52 Li Z.Q., Giege R., Jacrot B., Oberthur R., Thierry J.C. & Zaccai G. (1983) Biochemistry 22, 4380- 4388
53 Harvey S.C. & McCammon J.A. (1981) Nature 294, 286-287
54 Kim K. & Jhon M.S. (1979) Biochim. Biophys. Acta 565, 131-147