Embed Size (px)
Citation preview

S655L’Encéphale, 33 : 2007, Septembre, Cahier2
Hypothèses neuro-développementales
de la dépression
B. MilletCHU, Service Universitaire de Psychiatrie, 35703 Rennes Cedex 7
L’hypothèse neurodéveloppementale a permis d’accumuler beaucoup de connaissances à propos des schizophrénies.B. Millet examine cette hypothèse pour les troubles de l’humeur. Il montre que l’hypothèse ne manque pas d’arguments.
Un modèle neurodévelop-pemental pour une patholo-gie sous entend l’existence d’une vulnérabilité génétique et d’une interaction gène/envi-ronnement. Le phénomène pathologique retentit sur le plan neuro-anatomique par des anomalies structurales et fonc-tionnelles.
QU’EST CE QU’UN MODELE NEURODEVELOPPEMENTAL ?
Le modèle neuro-développemen-tal postule que la pathologie observée est la résultante d’un processus du développement neurologique, débutant bien avant l’apparition des symp-tômes cliniques et déterminée par la combinaison de facteurs environnementaux et généti-ques (23). Ce type de modèle, généralement appliqué à la schi-zophrénie, prend en compte des : - facteurs génétiques ;- stresseurs environnementaux au cours de l’enfance en parti-culier autour de la naissance ;
- stresseurs environnementaux à l’âge adulte (consommation de cannabis…);- anomalies cérébrales morpho-logiques et histologiques.
ASPECTS GENETIQUES DE LA DEPRESSION
Etudes de risques relatifs
La prévalence de l’épisode dépressif majeur en population générale est de 8%. Son risque de survenue est deux fois plus important chez la femme que chez l’homme (28). Les études de jumeaux retrou-vent un taux de concordance de 40 à 50 % chez les jumeaux
homozygotes (16). Les quel-q u e s é t u d e s d ’ a d o p t i o n concluent à un l ien généti-que. Selon les études familia-les, le risque de survenue de dépression chez des apparen-tés de sujets souffrant d’épi-sode dépressif est deux fois supérieur au risque de surve-nue de dépression en popula-tion générale (27). Certains facteurs environnementaux ont un rôle important dans la survenue d’une dépression tels que l’abus ou les négligences au cours de l’enfance ainsi que les événements de vie stres-sants (14,15).Le poids génétique de la dépres-sion apparaît d’autant plus fort que :
La d
épre
ssion
: de
s pra
tique
s aux
théo
ries 9

S656
La d
épre
ssion
: de
s pra
tique
s aux
théo
ries 9
- l’âge de début est précoce (âge inférieur à 30 ans);- les épisodes sont récurrents;- la précocité de survenue et le nom-bre de récurrences sont associés;- l’épisode dépressif s’inscrit dans un trouble bipolaire plutôt qu’uni-polaire.
Approche dimensionnelle ou facteur de risque ?
Doit-on considérer le trouble anxieux généralisé et le neuroti-cisme comme des facteurs de ris-que ou comme des dimensions de l’épisode dépressif majeur ?Le neuroticisme est un facteur unitaire de traits (dysphorie, anxiété, tension interne, réac-tivité émotionnelle) identifi é au cours de l’analyse factorielle de la personnalité. (11). Un score haut de neuroticisme est un facteur robuste de risque de survenue d’un épisode dépressif majeur. Certains facteurs génétiques prédisposant à la fois à l’épisode dépressif majeur, au neuroticisme et au trouble anxieux généralisé pourraient sous tendre le phéno-type anxiodépressif.
Etudes d’association
Les études d’association ont per-mis de mettre en évidence un lien entre des polymorphismes géné-tiques et la dépression :
- Le polymorphisme du gène du transporteur de la sérotonine 5HTTLPR (SLC6A4)
Beaucoup d’études ont montré l’existence d’une association entre la dépression et le gène du trans-
porteur de la sérotonine 5HTT-LPR. La fonctionnalité de la protéine dérivée du gène 5HTT-LPR est partiellement connue. La protéine issue de l’allèle court (s) du gène diminuerait la quantité de transporteur de la sérotonine et augmenterait la réactivité de l’amygdale aux images de peur et d’agressivité (comme chez les sujets déprimés) (13).
- Le polymorphisme du gène BDNF
Le Brain Derived Neurotrophic Factor (BDNF) est un facteur neurotrophique diminué par le stress et augmenté par la prise d’antidépresseurs. Le rôle du polymorphisme fonctionnel Val-66Met apparaît dans les varia-tions du volume hippocampique et de la mémoire épisodique : ceci n’est pas sans évoquer ce qui est observé chez les déprimés.
INTERACTION GENE ET ENVIRONNEMENT DANS LA DEPRESSION
Interaction entre gène 5HTTLPR et environnement
L’interaction gène et environne-ment a été étudiée par Caspi et al. en 2003 (6). Huit cent quarante sept sujets souffrant de dépression ont été suivis de 3 à 26 ans et évalués à différents temps de leur vie (3,5,7,11,13,15,18,21 ans). Les données recueillies pour chaque sujet comprenaient : - le type d’allèle du gène 5HTT-
LPR porté afi n de classer les indi-vidus en 3 groupes : s/s, s/l, ll ;- 14 événements de vie recensés concernant les domaines de l’em-ploi, des fi nances, de la famille, de la santé, des stresseurs relation-nels entre 21 et 26 ans ;- l’évaluation du diagnostic de dépression à 26 ans à l’aide du DIS (Diagnostic Interview Schedule).Trois sous groupes ont été exa-minés : - sujets homozygotes pour l’allèle S (s/s) soit 37% de la cohorte,- sujets hétérozygotes (s/l) soit 51% de la cohorte,- sujets homozygotes pour l’allèle L (l/l) soit 31% de la cohorte. Cette étude conclut que : - le nombre d’événements de vie stressants entre 21 et 25 ans prédit la survenue d’un épisode dépressif majeur (EDM) lorsque le sujet est porteur de l’allèle « s » et non lorsqu’il est porteur de l’al-lèle « l » du gène ;- la maltraitance de l’enfant entre 3 et 11 ans prédit la survenue de l’EDM lorsque le sujet est porteur de l’allèle « s ».Les événements de vie sont donc stressants en eux-mêmes, mais d’autant plus chez les sujets porteurs de l’allèle « s » du gène 5HTTLPR. Ces résultats ont été répliqués 5 fois de manière positive alors que deux études se sont révélées être négatives.
Rôle du gène 5HTTLPR sur les conséquences des événements de vieLe rôle du gène 5HTTLPR sur les conséquences des événements de vie a été évalué chez 118 sujets
L’Encéphale, 2007 ; 33 : 655-660, Cahier 2B. Millet
S657
La d
épre
ssion
: de
s pra
tique
s aux
théo
ries 9
âgés de 18 à 29 ans. Cette étude (26) s’appuie sur : - une évaluation psychologique et de la dépression grâce à l’inven-taire de Beck (BDI);- un recensement des événe-ments de vie précoces positifs et négatifs : amour de la famille, mal-traitance, abus verbal ou sexuel, violence intrafamiliale, etc ... ;- un recensement des événe-ments de vie actuels durant les 6 derniers mois à la fois positifs et négatifs. Les auteurs ne retrouvent pas d’effet direct du génotype en lui même sur le risque de survenue de la dépression. Mais les sujets homozygotes « s/s » sont plus à risque que les autres participants de présenter des symptômes dépressifs lorsque l’environne-ment précoce est défavorable. Inversement, les sujets « s/s » ont un risque moins important de présenter des symptômes dépressifs lorsque l’environne-ment précoce est favorable. Le risque de survenue de dépression apparaît donc corrélé à la pola-rité des événements de vie chez les sujets homozygotes « s/s ». Le génotype « s/s » ne serait pas un facteur de risque de la dépres-sion mais un facteur de risque de la réactivité émotionnelle.
Facteurs génétiques de vulnérabilité et liens neuroanatomiques
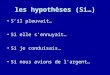
L’équipe de Pezawas s’est inté-ressée aux aspects structuraux cérébraux en fonction de la pré-sence ou non de l’allèle « s » du gène 5HTTLPR chez le sujet sain. La réalisation d’une IRM de haute résolution chez plus de
100 sujets sains, sans histoire psy-chiatrique a permis de mettre en évidence une hypotrophie cingu-laire chez les sujets porteurs de l’allèle « s » du gène 5HTTLPR (21) (fi gure 1).
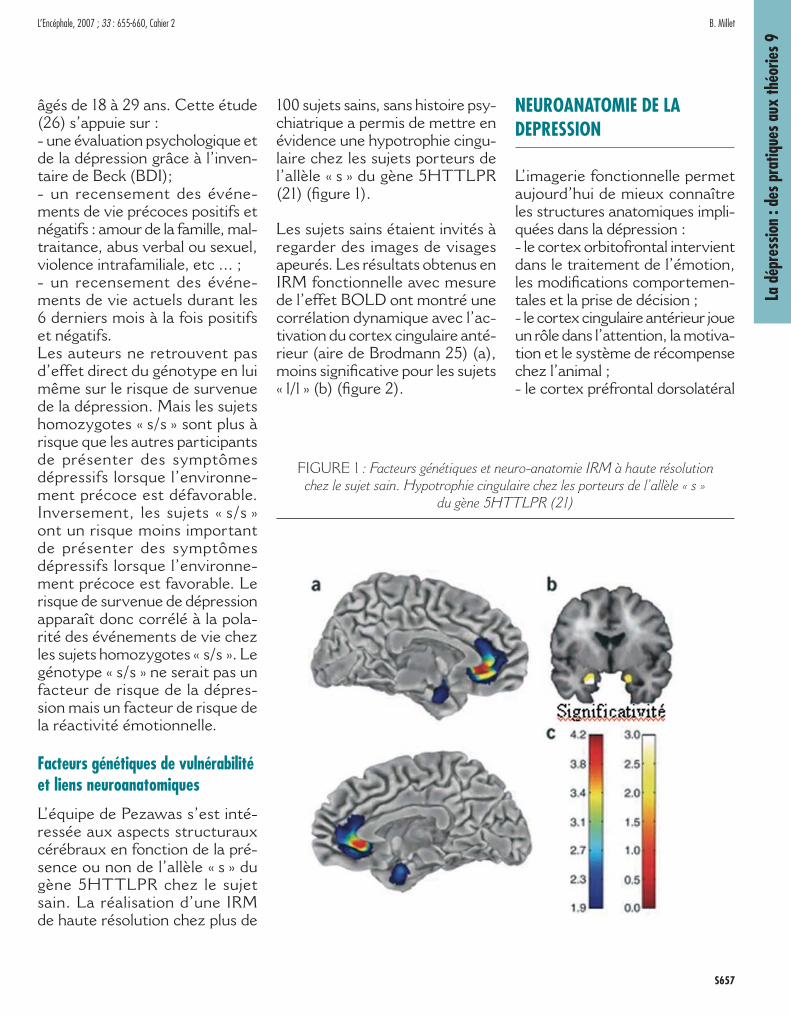
Les sujets sains étaient invités à regarder des images de visages apeurés. Les résultats obtenus en IRM fonctionnelle avec mesure de l’effet BOLD ont montré une corrélation dynamique avec l’ac-tivation du cortex cingulaire anté-rieur (aire de Brodmann 25) (a), moins signifi cative pour les sujets « l/l » (b) (fi gure 2).
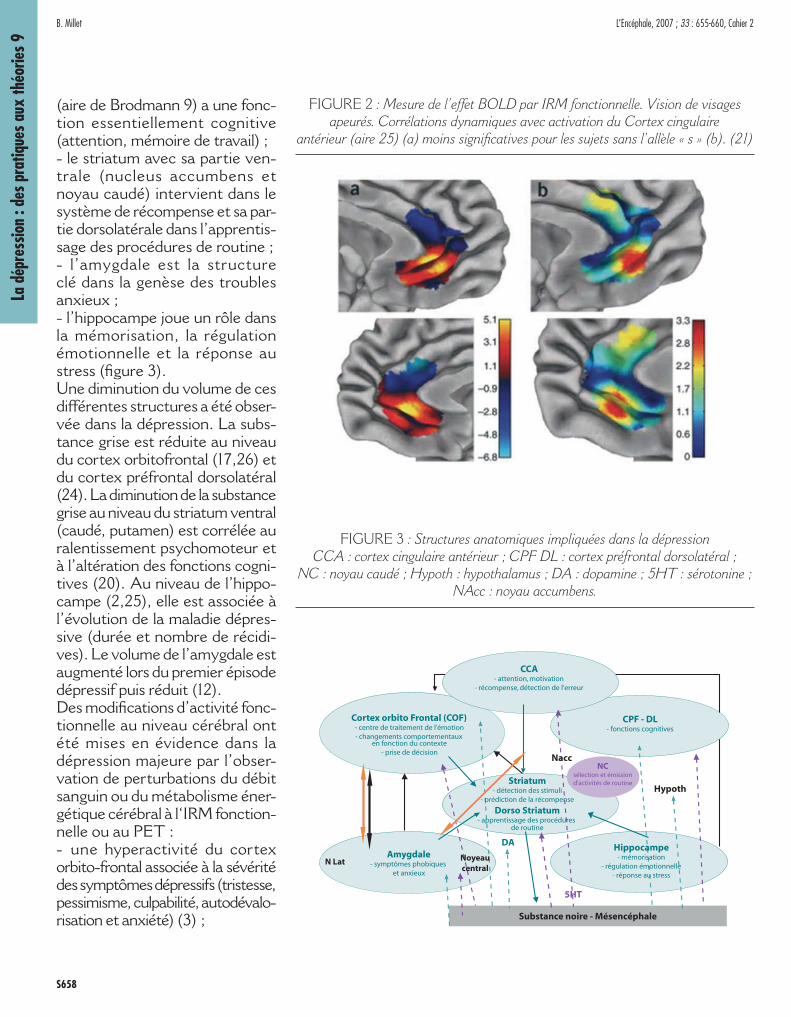
NEUROANATOMIE DE LA DEPRESSION
L’imagerie fonctionnelle permet aujourd’hui de mieux connaître les structures anatomiques impli-quées dans la dépression : - le cortex orbitofrontal intervient dans le traitement de l’émotion, les modifi cations comportemen-tales et la prise de décision ;- le cortex cingulaire antérieur joue un rôle dans l’attention, la motiva-tion et le système de récompense chez l’animal ;- le cortex préfrontal dorsolatéral
FIGURE 1 : Facteurs génétiques et neuro-anatomie IRM à haute résolutionchez le sujet sain. Hypotrophie cingulaire chez les porteurs de l’allèle « s »
du gène 5HTTLPR (21)
B. MilletL’Encéphale, 2007 ; 33 : 655-660, Cahier 2
S658
La d
épre
ssion
: de
s pra
tique
s aux
théo
ries 9
(aire de Brodmann 9) a une fonc-tion essentiellement cognitive (attention, mémoire de travail) ;- le striatum avec sa partie ven-trale (nucleus accumbens et noyau caudé) intervient dans le système de récompense et sa par-tie dorsolatérale dans l’apprentis-sage des procédures de routine ;- l’amygdale est la structure clé dans la genèse des troubles anxieux ;- l’hippocampe joue un rôle dansla mémorisation, la régulation émotionnelle et la réponse au stress (fi gure 3).Une diminution du volume de ces différentes structures a été obser-vée dans la dépression. La subs-tance grise est réduite au niveau du cortex orbitofrontal (17,26) et du cortex préfrontal dorsolatéral (24). La diminution de la substance grise au niveau du striatum ventral (caudé, putamen) est corrélée au ralentissement psychomoteur et à l’altération des fonctions cogni-tives (20). Au niveau de l’hippo-campe (2,25), elle est associée à l’évolution de la maladie dépres-sive (durée et nombre de récidi-ves). Le volume de l’amygdale est augmenté lors du premier épisode dépressif puis réduit (12). Des modifi cations d’activité fonc-tionnelle au niveau cérébral ont été mises en évidence dans la dépression majeure par l’obser-vation de perturbations du débit sanguin ou du métabolisme éner-gétique cérébral à l‘IRM fonction-nelle ou au PET : - une hyperactivité du cortex orbito-frontal associée à la sévérité des symptômes dépressifs (tristesse, pessimisme, culpabilité, autodévalo-risation et anxiété) (3) ;
FIGURE 3 : Structures anatomiques impliquées dans la dépression CCA : cortex cingulaire antérieur ; CPF DL : cortex préfrontal dorsolatéral ;
NC : noyau caudé ; Hypoth : hypothalamus ; DA : dopamine ; 5HT : sérotonine ; NAcc : noyau accumbens.
FIGURE 2 : Mesure de l’effet BOLD par IRM fonctionnelle. Vision de visages apeurés. Corrélations dynamiques avec activation du Cortex cingulaire
antérieur (aire 25) (a) moins signifi catives pour les sujets sans l’allèle « s » (b). (21)
L’Encéphale, 2007 ; 33 : 655-660, Cahier 2B. Millet

S659
La d
épre
ssion
: de
s pra
tique
s aux
théo
ries 9
- une hyperactivité de la région ventrale du cortex cingulaire anté-rieur et du striatum corrélée à la sévérité de l’épisode dépressif et son caractère résistant (7,19) ; - une hypoactivité des régions dorsolatérales (aire de Brodmann 9 et 46) et dorsomédianes du cor-tex préfrontal (aire de Brodmann 10, 24 et 32) (4,8) corrélée à une dégradation des fonctions cogni-tives (1,9) (fi gure 4).La réponse aux différents traite-ments antidépresseurs a aussi été étudiée en neuroimagerie fonc-tionnelle. Après 6 à 10 semaines de traitement par fl uoxétine, paroxé-tine, sertraline, venlafaxine, imi-pramine ou bupropion, le pattern d’activité cérébrale présent avant la mise en place du traitement se normalise (3,4,5).
On observe :- une augmentation de l’activité dans les régions corticales (en particulier au niveau des cortex préfrontal dorsolatéral, cingulaire antérieur dorsal et postérieur),
- une réduction de l’activité des structures sous-cortico-limbi-ques (cortex cingulaire anté-rieur subgénual, noyau caudé, amygdale, hippocampe) en cas de réponse favorable au traite-ment. L’interaction entre structures cor-ticales et sous-corticales joue-rait un rôle dans la réponse aux traitements antidépresseurs en permettant soit la correction du dysfonctionnement du circuit cortico-striato-thalamique soit en rétablissant l’équilibre entre l’ac-tivité corticale et limbique.
CONCLUSION
L’hypothèse neurodéveloppemen-tale de la dépression est évolutive puisque la vulnérabilité pour les troubles thymiques à la naissance est modulée par des facteurs envi-ronnementaux, notamment lors de la période postnatale. Par ailleurs, certaines anomalies mor-phologiques cérébrales pourraient
précèder la survenue de la dépres-sion, les séquelles cérébrales des épisodes dépressifs récurrents semblant accentuer l’altération initiale de ces fonctions cérébra-les. La dépression répondrait donc plus à un modèle de stress / vul-nérabilité. La superposition d’une prédisposition génétique avec des événements de vie et des stress responsables d’une dysfonc-tion cérébrale (boucles cortico-striato-thalamiques) aboutirait ainsi à la survenue d’un épisode dépressif majeur évoluant vers la chronicité ou la rémission.
Références
1. Bench CJ, Friston KJ, Brown RG, Frackowiak RS, Dolan RJ. Regional cerebral blood fl ow in depres-sion measured by positron emission tomography: the relationship with clinical dimensions. Psychol Med 1993;23:579-590.
2. Bremner JD, Narayan M, Anderson ER, Staib LH, Miller HL, Charney DS. Hippocampal volume reduction in major depression. Am J Psychiatry 2000;157:115-118.
3. Brody AL, Saxena S, Mandelkern MA, Fairbanks LA, Ho ML, Baxter LR. Brain metabolic changes associated with symptom factor improvement in major depressive disorder. Biol Psychiatry 2001;50:171-178.
4. Buchsbaum MS, Wu J, Siegel BV, Hackett E, Tre-nary M, Abel L, Reynolds C. Effect of sertraline on regional metabolic rate in patients with affective disorder. Biol Psychiatry 1997;41:15-22.
5. Canli T, Cooney RE, Goldin P, Shah M, Sivers H, Tho-mason ME, Whitfi eld-Gabrieli S, Gabrieli JD, Gotlib IH. Amygdala reactivity to emotional faces predicts improvement in major depression. Neuroreport 2005;16:1267-1270.
6. Caspi A, Sugden K, Moffi tt TE, Taylor A, Craig IW, Harrington H, McClay J, Mill J, Martin J, Braithwaite A, Poulton R. Infl uence of life stress on depression: moderation by a polymorphism in the 5-HTT gene. Science 2003;301:386-389.
FIGURE 4 : Structures cérébrales et changements fonctionnels dans la dépression
B. MilletL’Encéphale, 2007 ; 33 : 655-660, Cahier 2

S660
La d
épre
ssion
: de
s pra
tique
s aux
théo
ries 9
7. Drevets WC. Neuroimaging and neuropathological studies of depression: implications for the cognitive-emotional features of mood disorders. Curr Opin Neurobiol 2001;11:240-249.
8. Drevets WC, Bogers W, Raichle ME. Functional anatomical correlates of antidepressant drug treat-ment assessed using PET measures of regional glucose metabolism. Eur Neuropsychopharmacol 2002;12:527-544.
9. Drevets WC, Price JL, Bardgett ME, Reich T, Todd RD, Raichle ME. Glucose metabolism in the amyg-dala in depression: relationship to diagnostic sub-type and plasma cortisol levels. Pharmacol Biochem Behav 2002;71:431-447.
10. Duman RS, Monteggia LM. A neurotrophic model for stress-related mood disorders. Biol Psychiatry 2006;59:1116-1127.
11. Eysenck HJ. Neurosis, constitution and personalityZ Psychol Z Angew Psychol 1966;172:145-181.
12. Frodl T, Meisenzahl E, Zetzsche T, Bottlender R, Born C, Groll C, Jager M, Leinsinger G, Hahn K, Moller HJ. Enlargement of the amygdala in patients with a fi rst episode of major depression. Biol Psy-chiatry 2002;51:708-714.
13. Hariri AR, Drabant EM, Munoz KE, Kolachana BS, Mattay VS, Egan MF, Weinberger DR. A suscepti-bility gene for affective disorders and the response of the human amygdala. Arch Gen Psychiatry 2005;62:146-152.
14. Kendler KS, Sheth K, Gardner CO, Prescott CA. Childhood parental loss and risk for fi rst-onset of major depression and alcohol dependence: the
time-decay of risk and sex differences. Psychol Med 2002;32:1187-1194.
15. Kendler KS, Kuhn J, Prescott CA. The interrela-tionship of neuroticism, sex, and stressful life events in the prediction of episodes of major de-pression. Am J Psychiatry 2004;161:631-636.
16. Kendler KS. Twin studies of psychiatric illness: an update. Arch Gen Psychiatry 2001;58:1005-1014.
17. Lacerda AL, Keshavan MS, Hardan AY, Yorbik O, Brambilla P, Sassi RB, Nicoletti M, Mallinger AG, Frank E, Kupfer DJ, Soares JC. Anatomic evaluation of the orbitofrontal cortex in major depressive disor-der. Biol Psychiatry 2004;55:353-358.
18. Levinson DF. The genetics of depression: a review. Biol Psychiatry 2006;60:84-92.
19. Mayberg HS, Lozano AM, Voon V, McNeely HE, Seminowicz D, Hamani C, Schwalb JM, Kennedy SH. Deep brain stimulation for treatment-resistant depression. Neuron 2005;45:651-660.
20. Naismith S, Hickie I, Ward PB, Turner K, Scott E, Little C, Mitchell P, Wilhelm K, Parker G. Caudate nucleus volumes and genetic determinants of ho-mocysteine metabolism in the prediction of psycho-motor speed in older persons with depression. Am J Psychiatry 2002;159:2096-2098.
21. Pezawas L, Meyer-Lindenberg A, Drabant EM, Ver-chinski BA, Munoz KE, Kolachana BS, Egan MF, Mattay VS, Hariri AR, Weinberger DR. 5-HTTLPR polymorphism impacts human cingulate-amyg-dala interactions: a genetic susceptibility me-chanism for depression. Nature Neuroscience
2005;8:828-834. 22. Posener JA, Wang L, Price JL, Gado MH, Province
MA, Miller MI, Babb CM, Csernansky JG. High-di-mensional mapping of the hippocampus in depres-sion. Am J Psychiatry 2003;160:83-89.
23. Rapoport JL, Addington AM, Frangou S, Psych MR. The neurodevelopmental model of schizophrenia: update 2005. Mol Psychiatry 2005;10:434-449.
24. Rajkowska G, Miguel-Hidalgo JJ, Wei J, Dilley G, Pittman SD, Meltzer HY, Overholser JC, Roth BL, Stockmeier CA. Morphometric evidence for neu-ronal and glial prefrontal cell pathology in major depression. Biol Psychiatry 1999;45:1085-1098.
25. Sheline YI, Gado MH, Kraemer HC. Untreated de-pression and hippocampal volume loss. Am J Psy-chiatry 2003;160:1516-1518.
26. Taylor SE, Way BM, Welch WT, Hilmert CJ, Lehman BJ, Eisenberger NI. Early family environment, cur-rent adversity, the serotonin transporter promoter polymorphism, and depressive symptomatology. Biol Psychiatry 2006;60:671-676.
27. Weissman MM, Kidd KK, Prusoff BA. Variability in rates of affective disorders in relatives of de-pressed and normal probands. Arch Gen Psychiatry 1982;39:1397-1403.
28. Weissman MM, Bland RC, Canino GJ, Faravelli C, Greenwald S, Hwu HG, Joyce PR, Karam EG, Lee CK, Lellouch J, Lepine JP, Newman SC, Rubio-Stipec M, Wells JE, Wickramaratne PJ, Wittchen H, Yeh EK. Cross-national epidemiology of major depres-sion and bipolar disorder. JAMA 1996;276:293-299.
L’Encéphale, 2007 ; 33 : 655-660, Cahier 2B. Millet