Embed Size (px)
Citation preview
- 1 -
Master « Expression Génique et Protéines Recombinantes »Novembre 2005
« Inhibition par des peptidessynthétiques
de la fibrillogénèse dans la maladied’Alzheimer »
KOKA VondaMONDEIL Sophie
Le peptide β amyloïde1-40 sous sa forme
soluble
Biotechnologie
Master
Protéines
Recombinantes

- 2 -
SOMMAIRE
La Maladie d’Alzheimer
1- Introduction
2- Caractérisation de la maladie
3- Facteurs et causes de la maladie3-1 Facteurs génétiques3-2 Facteurs sporadiques
4- Formation du peptide Aβ 4-1 L’APP
4-2 Le peptide Aβ4-3 L'amyloïdogénèse
5- Inhibition de l’amyloïdogénèse5-1 Clefs thérapeutiques
Inhibition par des peptides synthétiques de la fibrillogénèse
1-Les peptides inhibiteurs1-1 Présentation1-2 Synthèse1-3 Exemple de construction : incorporation de la proline ou d’acides
aminés N- méthylés1-4 Caractérisation des peptides inhibiteurs
2- Etudes réalisées sur l’inhibition des feuillets β2-1 Stratégies de recherche2-2 Etudes
a- Amélioration de la perméabilité cérébraleb- Résistance à la protéolyse plasmatiquec- Caractérisation de l’inhibition de la formation des fibres
amyloïdesd- Dissociation des fibres amyloïdes
3-Résultats3-1 Homologie de séquence3-2 Résultats obtenus avec les peptides inhibiteurs3-3 Hypothèses du mode d’action des peptides inhibiteurs
4- Discussion et perspectives
Références bibliographiques

- 3 -
La Maladie d’Alzheimer
1- INTRODUCTION
La maladie d’Alzheimer est une démence dégénérative. Elle survient en moyenneautour de 65 ans et concerne actuellement 350 000 personnes en France. Sa prévalence (5%de plus de 65 ans, 20 % des plus de 80 ans) et sa charge économique (70% des lits enhôpitaux de long séjour) en fait un problème majeur de santé publique dans tous les paysindustrialisés (1).
2- CARACTERISATION DE LA MALADIE
La maladie est caractérisée par la formation de lésions spécifiques :- L’atrophie corticale :
Chez les patients atteints le cerveau peut perdre 8 à 10 % de son poids tous les dix ans,alors que chez des individus sains, cette perte n’est que de 2%.
- Les plaques sénilesCe sont des plaques amyloïdes contenant des formes agrégées du peptide Aβlocalisées dans le cerveau. Ces amas d’amyloïdes sont entourés de prolongementsneuritiques et de cellules gliales. Une proportion significative de la population âgéeprésente des plaques amyloïdes sans traces de la maladie. Par conséquent, le dépôtd’amyloïde n’est pas un facteur déterminant de la maladie même s’il constitue un desmécanismes lié à la physiopathologie de celle-ci. Mais dans les formes familiales, il ya une corrélation nette entre le dépôt anormal de l’amyloïde et la pathogénicité de lamaladie.
- Les dégénérescences neurofibrillairesElles se situent à l’intérieur des neurones. Ce sont des écheveaux de filamentsanormaux constitués, entre autres, d’une forme hyperphosphorylée de la protéine Tauqui prend alors l’aspect de paires de filaments hélicoïdaux.(1, 2, 3)
3- FACTEURS ET CAUSES DE LA MALADIE
3-1 Facteurs génétiques
La mise en évidence de facteurs génétiques s’est faite par l’étude des personnesaffectées par la forme familiale ou par la trisomie 21. Ils représentent 5 à 10% de l’ensembledes cas, et sont caractérisés par des manifestations précoces de la maladie (parfois avant 40ans, voire 30 ans). La présence de gènes spécifiques de la maladie a contribué à mieuxcomprendre le mécanisme de celle-ci.Les gènes majeurs impliqués sont les suivants :- Amyloid protein precursor (APP)- Présenilines 1 et 2 (PS1 et PS2)- alpha 2 macroglobulines- apoprotéine E
Le gène de l’APP, localisé sur le chromosome 21, présente chez l’homme de raresmutations (moins de 30 familles connues dans le monde). Des souris mutantes présentent une

- 4 -
perte de poids, une mobilité réduite, des altérations du comportement, une mort prématurée etla présence de plaques amyloïdes cérébrales. (10)
PS1 est impliqué dans la formation de l’APP normal. Des mutations altérant PS1 àdifférents endroits semblent être responsables du mauvais clivage de l’APP et de laproduction de peptide Aβ (1-42), qui est le variant le plus virulent dans la formation desplaques (2,4). Ainsi, la mutation A246E va provoquer une formation rapide des plaquesamyloïdes, une dystrophie neuritique et de l’astrogliosis (5). De plus, PS1 interagit avec laglycogène synthase kinase (GSK 3 B) qui est une des kinases impliquées dans laphosphorylation de la protéine Tau. La protéine Tau est une petite protéine qui s'associe auxmicrotubules et fait le lien entre les microtubules et les autres éléments du cytosquelette. Chezl'homme, elle est exprimée dans les neurones et joue un rôle important dans l'assemblage desmonomères en microtubules. Des mutations sur PS1 pourraient conduire àl’hyperphosphorylation de Tau (2).
Dans certains cas de maladie de type « familial » il a été mis en évidence desmutations au niveau du gène PS1 et PS2. Des recherches supplémentaires ont permis demettre en évidence l’implication d’autres gènes :
-dipeptidylcarboxypeptidase (DCP1)-bléomycine hydrolase- α 2 macroglobuline-récepteur pour des lipoprotéines « légères » (LRP) (2)
3-2 Facteurs sporadiques
La maladie peut être due à d’autres facteurs non génétiques. Les paramètresresponsables du passage du peptide d’une structure hélice α (non pathogène) auxfeuillets β (pathogène) sont nombreux et ce mécanisme n’est pas complètement élucidé.Grâce à un peptide modélisé présentant une structure α, on a mis en évidence l’influence deconditions environnementales, telles que le pH ou la force ionique. Cette transformation feraitaussi intervenir les propriétés intrinsèques des différents acides aminés composant le peptide,avec entre autres leur degré d’hydrophobicité. En effet, il a été montré que plus on avait derésidus hydrophobes et plus on avait de chance d’obtenir des structures en feuillets β (6)
D'autres hypothèses impliquent des ions métalliques dans l’agrégation du peptide,comme l’aluminium, le fer, le zinc et le cuivre. Il y aurait un lien entre des taux anormauxdans le cerveau de ces métaux et la progression de la maladie. L’aluminium est peut être unfacteur et les études ont montré que le métabolisme du fer était lui aussi perturbé dans lamaladie. Le métabolisme et la régulation anormale du zinc semblent jouer un rôle net dansl’agrégation de Aβ (30). Enfin, quant au cuivre, il parait avoir une influence protectrice vis àvis de la formation de la plaque amyloïde, mais on a montré aussi qu’il stimulait la voie de laβ sécrétase qui intervient dans la protéolyse de l’APP. Par ailleurs, le cuivre augmente lasécrétion de l’APP, au niveau duquel il se fixe. En présence de concentrations micromolairesde cuivre, on a observé l’agrégation de peptides synthétiques sous forme d’amyloïde (30). Laréoxydation rapide de ce métal pourrait être impliquée dans la fragmentation de l’APP.
Des facteurs comme la concentration en peptide, l’existence d’un stress oxydatif, ou laprésence de radicaux libres sembleraient aussi jouer un rôle dans l’agrégation du peptide Aβ(3)

- 5 -
4- FORMATION DU PEPTIDE Aβ
4-1 L’APP
Le peptide APP fait partie d’une famille de glycoprotéines transmembranaires de type1 que l’on retrouve dans différents types de cellules.
La partie N-term de l’APP se trouve dans le domaine extracellulaire et peut être localiséedans les vésicules du réticulum endoplasmique, dans celles de l’appareil de Golgi ou dans lesendosomes. La partie C-term quant à elle est localisée au niveau du cytoplasme. La protéineAPP est donc une protéine transmembranaire des neurones (2)
Cette protéine aurait des fonctions variées, comme l’inhibition de la sérine protéase,l’adhésion cellulaire et des propriétés de protection des neurones (2). De plus, il a été montréqu’elle présente des domaines de fixation pour des substrats tels que l’héparine, le collagèneet la laminine (7). L’APP intervient aussi comme facteur de croissance des cellules neuronales(8)
Individu normal :L’enzyme α sécrétase coupe une partie du fragment de l’APP, entre la Lys 16 et la Leu 17 quiest située à l’extérieur du neurone. Il s’en suit d’une libération d’un fragment spécifique, lefragment sAPP (secreted AP). Ce peptide est sécrété par les cellules neuronales et se retrouvedans le liquide cérébro-spinal et dans le sang. Il n’est pas neurotoxique et est sensible à laprotéolyse. sAPP commande certaines actions essentielles dans la cellule neuronale :
- il est neuroprotecteur- c’est un récepteur membranaire des kinésines- il inhibe les sérines protéases- il interviendrait dans les phénomènes d’adhésion cellulaire (2)I- il facilite le contact entre les synapses et permet aussi d’augmenter leur nombre- il participe au maintien de la structure des neurones, ainsi qu’à leur croissance- il permet la création de nouvelles connexions- il stabilise la concentration en calcium intracellulaire (8)
Individu malade :Des mutations portant sur la séquence codant pour l’APP modifieraient sa structure. Lesprotéases (β et γ sécrétases) au lieu de découper le fragment sAPP découperaient alors unfragment différent, le peptide Aβ existant sous deux formes :
-Aβ (1-40) : 95% des peptides Aβ-Aβ (1-42) qui s’auto agrège plus facilement, 40 fois plus vite que la forme précédente
(Forme la plus toxique) (9)
4-2 Le peptide Aβ
La séquence du peptide Αβ est la suivante :
NH2 - DAEFRHDSGY10EVHHQKLVFF20AEDVGSNKGA30IIGLMVGGVV40- COOH
Dans la forme pathologique le peptide Aβ (4.3 KDa) s’auto-associe sous forme defeuillets β en conformation antiparallèle (3) Elle est caractérisée par une grande insolubilité,par sa capacité à former de agrégats et par sa toxicité. Elle est résistante à la protéolyse. Si onne connaît pas encore de façon précise les interactions qui existent entre les différents brins,une alternance d’acides aminés hydrophiles et hydrophobes est nécessaire. La stabilité des

- 6 -
feuillets β serait due à des interactions électrostatiques, ainsi qu’à la formation de liaisonshydrogène entre les brins peptidiques (10)
Des nombreuses études ont montré que la région hydrophobe comprise entre les acidesaminés 17 à 21 (en gras et en rouge dans la séquence ci-dessus) était impliquée dans latoxicité et dans l'auto-agrégation de Aβ (11,12). La substitution des acides aminés Lysine16, Leucine 17 et Phénylalanine 20 par une Alanine a provoqué des changementsconformationnels du peptide et a inhibé la formation des fibres d’amyloïde (13).
4-3 L’amyloïdogenèse
L’agrégation des peptides Aβ conduit à la formation de fibres d’amyloïde. Des étudespar diffraction aux rayons X ont montré que les fibres d’amyloïde étaient composées defeuillets β en configuration antiparallèles (3).
La formation et l’élongation des fibres d’amyloïde se fait en plusieurs étapes.Le processus de formation des fibres est initié par une augmentation de la concentration enpeptide Aβ . Le peptide Aβ, sous forme de monomères va s’assembler en micelles. Au départ,il y a un équilibre réversible entre les formes monomériques et les micelles (diamètre 14 nm).Ces micelles fusionnent ensuite entre elles et forment alors un microagrégat de petite taille(diamètre 8nm). Cette étape pourrait faire intervenir l’apolipoprotéine E4. Une fois ce petitnoyau formé, on va avoir une élongation à une extrémité, par fixation de monomères de Aβ etde petits précurseurs protofibrillaires.Ensuite, ces structures se fixent face à face pour former des protofilaments hélicoïdaux qui ontentre 2,5 et 3,5 nm de diamètre. Chaque protofilament est composé de résidus polypeptidiquesorganisés en feuillets β, qui sont alignés perpendiculairement par rapport à l’axe de la fibre.Les protofilaments s’associent ensuite par 5, 6 ou 8 pour former des fibres d’amyloïde.Celles-ci ont un diamètre moyen compris entre 7,5 et 8 nm (14, 15,16).
La figure ci-dessous présente un diagramme schématique des différents niveaux destructure en partant des feuillets β, (liés par des liaisons hydrogène et en conformationantiparallèle) dans un protofilament jusqu’à l’organisation de ceux-ci dans une fibre amyloïde(Fig.1 extraite de la publication 15)
5- INHIBITION DE L’AMYLOIDOGENESE
Feuillets β
Fig. 1 Schéma del’amyloïdogénèse

- 7 -
5-1 Clefs thérapeutiques
Actuellement il n’existe aucun traitement efficace pour la maladie d’Alzheimer. Laprévention de l’agrégation du peptide Aβ en plaques ou en fibres amyloïdes ainsi quel’inhibition de leur formation peut être un des objectifs de la thérapie et de la prévention de lamaladie d’Alzheimer.
Toutefois, le peptide Aβ est extrêmement insoluble. Sa dissolution demande desagents chaotropiques ou des solvants organiques forts, ce qui fait que la destruction desplaques ne peut être envisagée in situ.Différentes stratégies peuvent être développées pour diminuer l’amyloïdogénèse :- bloquer l’expression de l’APP- inhiber l’action des sécrétases- inhiber la formation des fibres amyloïdes- tenter d’éliminer les plaques grâce à des enzymes, telles que la néprilysine (17, 18) ou lagelsoline (3, 18). La néprilysine est une métalloprotéase à zinc de 97 KDa. Elle est connuepour dégrader les enképhalines, les endothélines et le peptide amyloïde Aβ (35). Quand à lagelsoline, elle inactive le complexe formé par la DNase I avec l’actine. Par conséquent, ellerégule l’activité de la DNase I, qui est impliquée dans l’apoptose des cellules (34).Il a étémontré récemment qu’elle se fixait sur le peptide Aβ, dissociait les fibres amyloïdespréformées et inhibait la cytotoxicité du peptide β amyloide (36).- réaliser une immunisation avec le peptide Aβ 1-42. Cependant, les essais en phase 2 chezl’homme ont dus être arrêtés à la suite d’encéphalopathies d’origine inconnue.- agir au niveau du peptide :
• en inhibant l’activité neurotoxique de l’amyloïde• en limitant ses interactions avec des protéines associées• en provoquant son élimination du cerveau• en empêchant la formation des plaques amyloïdes.
Les molécules individuelles de peptide Aβ ne sont pas liées par des liaisons covalenteset la formation de plaques peut être un processus réversible. La conformation pathologique dupeptide amyloïde sous forme de feuillets β confère à cette protéine un potentiel d’agrégationélevé.
Un mauvais repliement d’une protéine est le point commun de maladiesneurodégénératives comme la maladie d’Alzheimer, la maladie de Creutzfeldt-Jakob ou lamaladie d’Huntington. Dans celles ci, la protéine obtenue présente une structure riche enfeuillets β.
Des composés capables de déstabiliser les feuillets β ou d’empêcher leur formationpourraient constituer des inhibiteurs efficaces, et donc des médicaments potentiels pour letraitement de la maladie d’Alzheimer (10)
Dans les publications A et B, les chercheurs ont synthétisé des peptides capables de sefixer sur le peptide β amyloïde et d’en déstabiliser la structure. Ces peptides ont été choisispour interagir avec la région hydrophobe centrale du peptide β amyloïde, entre les acidesaminés 17 à 21 (19).
- 8 -
Inhibition par des peptides synthétiques de lafibrillogénèse
1- LES PEPTIDES INHIBITEURS
1-1 Présentation
Les peptides inhibiteurs sont des petits peptides synthétiques homologues à différentesrégions de Aβ. Dans un premier temps les chercheurs ont étudié les effets de ces derniers surl’agrégation de Aβ. Des études ont montré que le remplacement des acides aminés hydrophiles par desacides aminés hydrophobes au niveau de la région centrale de Aβ inhibait la formation desfibres amyloïdes. Par conséquent l’assemblage de Aβ est en partie dû à des interactionshydrophobes. La conformation adoptée par le peptide Aβ semble être un facteur importantdans la formation de l’amyloïde. En effet, le peptide Aβ présentant une structure sous formed’hélice α, ou random coil va s’agréger très doucement. Par contre, le même peptide avec uneconformation en feuillets β présente une vitesse d’agrégation rapide. (12).
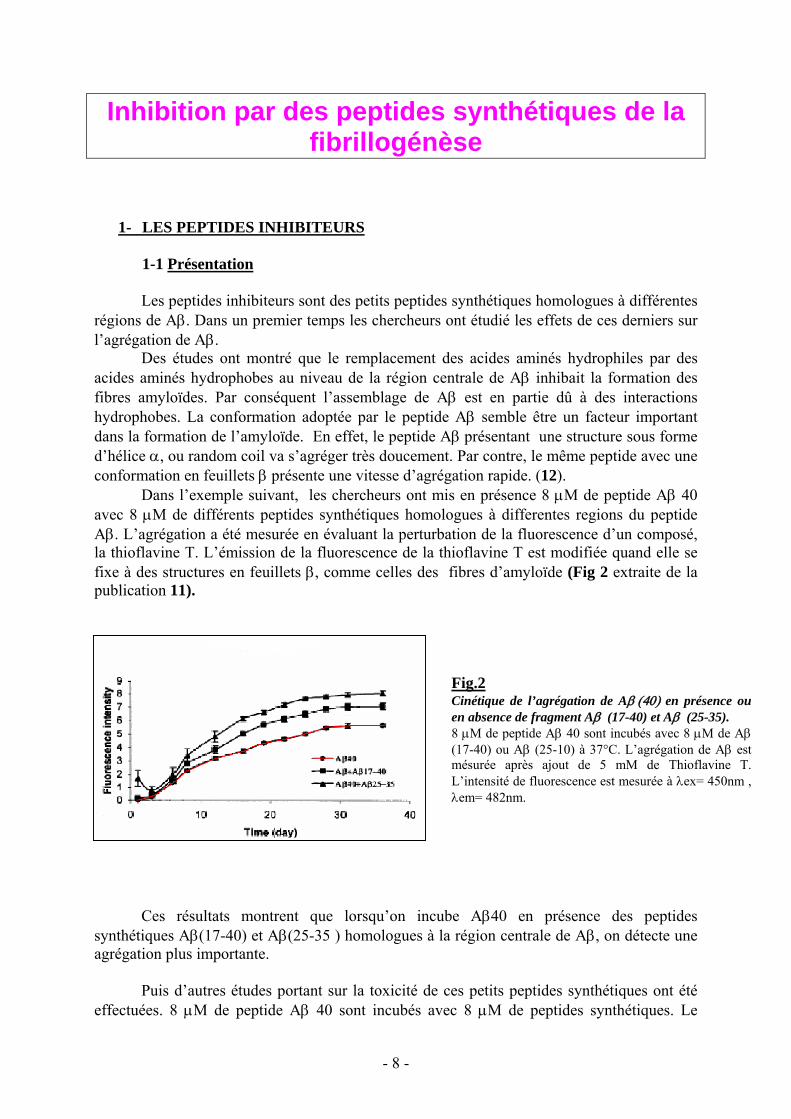
Dans l’exemple suivant, les chercheurs ont mis en présence 8 μM de peptide Aβ 40avec 8 μM de différents peptides synthétiques homologues à differentes regions du peptideAβ. L’agrégation a été mesurée en évaluant la perturbation de la fluorescence d’un composé,la thioflavine T. L’émission de la fluorescence de la thioflavine T est modifiée quand elle sefixe à des structures en feuillets β, comme celles des fibres d’amyloïde (Fig 2 extraite de lapublication 11).
Ces résultats montrent que lorsqu’on incube Aβ40 en présence des peptidessynthétiques Aβ(17-40) et Aβ(25-35 ) homologues à la région centrale de Aβ, on détecte uneagrégation plus importante.
Puis d’autres études portant sur la toxicité de ces petits peptides synthétiques ont étéeffectuées. 8 μM de peptide Aβ 40 sont incubés avec 8 μM de peptides synthétiques. Le
Fig.2Cinétique de l’agrégation de Aβ (40) en présence ouen absence de fragment Aβ (17-40) et Aβ (25-35).8 μM de peptide Aβ 40 sont incubés avec 8 μM de Aβ(17-40) ou Aβ (25-10) à 37°C. L’agrégation de Aβ estmésurée après ajout de 5 mM de Thioflavine T.L’intensité de fluorescence est mesurée à λex= 450nm ,λem= 482nm.

- 9 -
mélange a été ajouté à des cellules de la lignée SH-SY5Y, issues de neuroblastome humain.Après 18 jours d’incubation, un test au MTT a été réalisé. Celui ci permet de déterminer laviabilité des cellules. On ajoute le MTT (sel de tétrazolium jaune) à des cellules. Si celles-cisont vivantes, le MTT est métabolisé au niveau des mitochondries en formazan de couleurbleu. L’absorbance est ensuite déterminée. Une absorbance élevée correspond à une bonneviabilité. (11)La Fig.3 (extraite de la publication 11) indique que les peptides synthétiques Aβ (17-40) etAβ (25-35) présentent une toxicité aussi importante que le peptide Aβ 40.
Par ailleurs, d’autres expériences complémentaires basées sur le même principe ontmis en évidence une forte implication de la région centrale de Aβ (17-20) dans l’agrégation etla toxicité du peptide Aβ40.
Ces études ont permis de choisir la zone d’homologie 17-20 qui est la plus adéquatepour la synthèse d'un peptide inhibiteur.
1-2 Synthèse des peptides inhibiteurs
Les peptides inhibiteurs doivent être capables de se lier à Aβ afin de déstabiliser etd’empêcher la formation d’amyloïde. Ces peptides sont issus d’une synthèse chimique etpeuvent être éventuellement marqués à l’iode 125 I suivant les études effectuées (B). Lapurification a été faite par chromatographie haute pression (HPLC) avec une colonne C18 enphase inverse. Puis la séquence peptidique synthétisée est contrôlée par spectrométrie demasse (ESI-MS/MS ou MALDI-TOF/MS).
Des peptides de petite taille ont déjà été largement utilisés comme médicaments.Toutefois, il y a plusieurs problèmes associés à leur utilisation dans les maladies du systèmenerveux central. En effet, ils sont rapidement éliminés du plasma par protéolyse. Environ 50%des peptides inhibiteurs sont dégradés par des protéases au bout de 5 minutes (B).De plus, ces peptides présentent une mauvaise perméabilité au niveau des membranes desméninges. Celles ci sont connues pour empêcher le passage d’environ 95% des médicaments.Enfin, leur nature hydrophobe fait qu’ils sont peu solubles (B).
Fig.3Test de toxicité des différentes régions de Aβ.8 μM de peptide Aβ 40 sont incubés avec 8 μMde peptides synthétiques à 37°C pendant 18jours . Le mélange est ajouté à des cellules dela lignée SH-SY5Y, issues de neuroblastomehumain. Puis un test de viabilité cellulaire auMTT est réalisé.

- 10 -
Les nouvelles améliorations de ces peptides portent sur la synthèse de nouveauxpeptides inhibiteurs modifiés ayant intégré différents paramètres tels que :
- l’ajout de résidus chargés aux extrémités induit une augmentation de la solubilité - l’ajout de polyamines naturelles (putrescine, spermidine et spermine) augmente laperméabilité cérébrale et la protection contre la protéolyse plasmatique. Le choix a été faitd’ajouter une putrescine, fixée au niveau du COOH du peptide inhibiteur. La formule de laputrescine est la suivante : NH2 - (CH2)4 - NH2.Il a été mis en évidence que cette amidation augmente la perméabilité de l’inhibiteur d’unfacteur 2. (B, 20)
- une acétylation à l’extrémité N terminale, une amidation à l’extrémité C terminale ouune N-méthylation (A) permettent une protection contre la protéolyse. (B)- utilisation des peptides sous une forme énantiomérique D. Cette forme augmentel’efficacité de l’inhibiteur. (B, 21)- ajout de résidus proline, qui sont des acides aminés rares dans les feuillets β. Leur
introduction devrait permettre une amélioration de l'inhibition de la formation des feuilletsβ (13)
1-3 Exemple de construction: incorporation de la proline ou d’acides aminés N-méthylés
Dans la publication B, les chercheurs ont synthétisé des peptides inhibiteurshomologues à la région centrale de Aβ (LVFFA), portant un résidu putrescine à l’extrémité Cterminale, 3 résidus proline et un résidu tyrosine. Par ailleurs, ces peptides ont été synthétiséssous forme D et sous forme L. (Exemple de séquence : YiAβ11 : RDLPFYPVPID).
Dans la publication A, le choix a été fait de travailler avec des peptides inhibiteursméthylés. La méthylation des acides aminés peut être réalisée de plusieurs façons. On peutméthyler des acides aminés qui se suivent, ou alors réaliser une méthylation alternée, enintercalant un acide aminé normal non méthylé et un acide aminé méthylé. (Fig.4 publicationA)
1-Peptide Aβ 16-22 méthylé (méthylation alternée)
NH2-K(MeL)V(MeF)F(MeA)E-CO-NH2
2-Peptide Aβ16-22 méthylé (méthylation consecutive)
NH2-KL(MeV)(MeF)(MeF)(MeA)E-CO-NH2
Fig.4Peptides inhibiteurs Aβ (16-22) méthylés.Représentation schématique des peptides inhibiteurs présentant une méthylationalternée (1) et une méthylation consécutive (2). Dans cette figure les atomes decarbone sont en noir, les azotes sont en bleu, les atomes d’oxygène sont en rouge, leshydrogènes liés aux atomes d’azote sont en jaune, et les autres atomes d’hydrogène nesont pas représentés.
1
2
- 11 -
1-4 Caractérisation des peptides inhibiteurs
Les peptides synthétiques ont été caractérisés par différentes techniques biophysiques.Des expériences d’ultracentrifugation ont montré que les peptides Aβ (16-22) méthylésétaient sous une forme monomérique en solution.
Puis des études en dichroïsme circulaire ont été effectuées sur le peptide Aβ (16-22)méthylé ou non. Les résultats mettent en évidence une structure en random coil pour lepeptide non méthylé. Quand on méthyle ce même peptide, on observe une structure enfeuillets β. Les auteurs font varier la concentration en peptide méthylé et n’observent pas devariation au niveau du minimum obtenu sur le spectre de dichroïsme circulaire. Parconséquent, le signal en dichroïsme circulaire correspondant à des feuillets β serait dûuniquement à la présence de la méthylation. Celle ci aurait une influence au niveau de larépartition des charges sur le peptide inhibiteur (A).
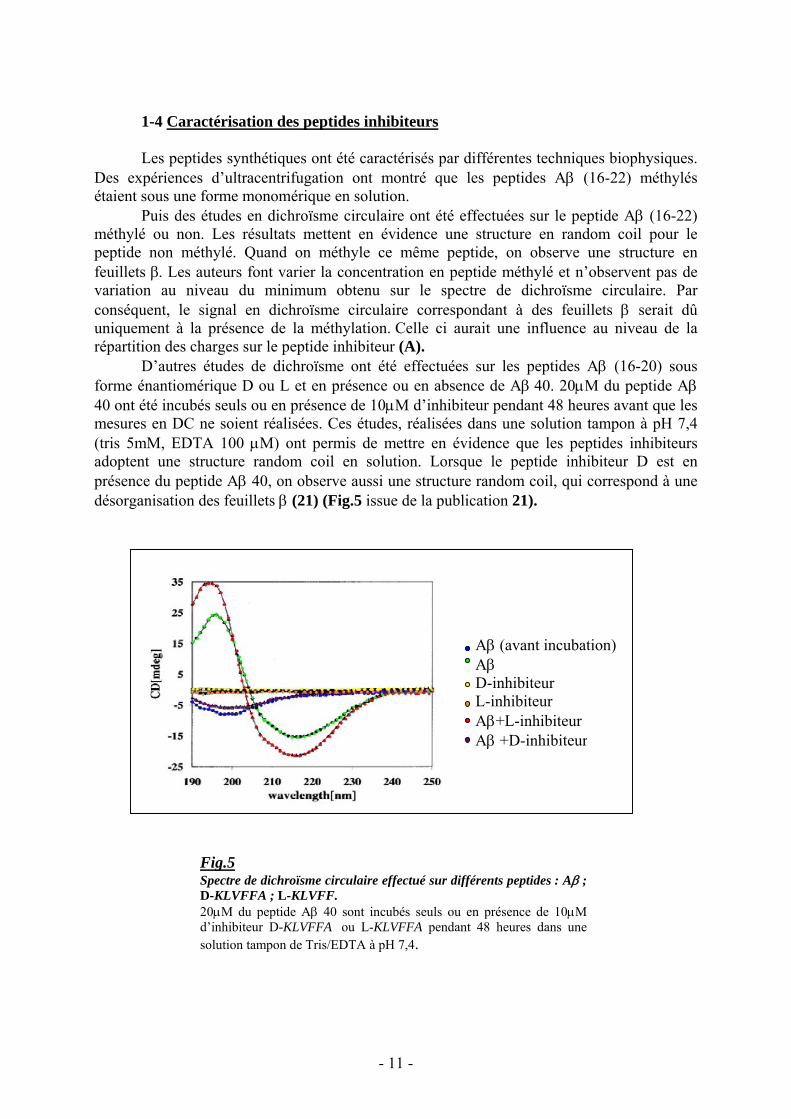
D’autres études de dichroïsme ont été effectuées sur les peptides Aβ (16-20) sousforme énantiomérique D ou L et en présence ou en absence de Aβ 40. 20μM du peptide Aβ40 ont été incubés seuls ou en présence de 10μM d’inhibiteur pendant 48 heures avant que lesmesures en DC ne soient réalisées. Ces études, réalisées dans une solution tampon à pH 7,4(tris 5mM, EDTA 100 μM) ont permis de mettre en évidence que les peptides inhibiteursadoptent une structure random coil en solution. Lorsque le peptide inhibiteur D est enprésence du peptide Aβ 40, on observe aussi une structure random coil, qui correspond à unedésorganisation des feuillets β (21) (Fig.5 issue de la publication 21).
Aβ (avant incubation) Aβ D-inhibiteur L-inhibiteur Aβ+L-inhibiteur Aβ +D-inhibiteur
Fig.5Spectre de dichroïsme circulaire effectué sur différents peptides : Aβ ;D-KLVFFA ; L-KLVFF.20μM du peptide Aβ 40 sont incubés seuls ou en présence de 10μMd’inhibiteur D-KLVFFA ou L-KLVFFA pendant 48 heures dans unesolution tampon de Tris/EDTA à pH 7,4.

- 12 -
2- ETUDES REALISEES SUR L’INHIBITION DES FEUILLETS β
2-1 Stratégies de recherche
Les études effectuées sur l’inhibition des feuillets β par des peptides synthétiques sontgénéralement basées sur les thèmes suivants :
- l’amélioration de la perméabilité cérébrale- la résistance à la protéolyse plasmatique- la caractérisation de l’inhibition de la formation des fibres amyloïdes- la dissociation des fibres d’amyloïde
2-2 Etudes
a-Amélioration de la perméabilité cérébrale
Ces études ont été réalisées in vivo chez des souris. Des peptides inhibiteurs marqués àl’ 125I ont été injectés au niveau de la veine brachiale. Après 15 minutes, l’animal est sacrifié,et les différentes parties du cerveau sont disséquées. Une mesure quantitative de laradioactivité dans celles ci est ensuite faite avec un compteur pour les rayons gamma. Lespeptides inhibiteurs portant un résidu putrescine présentent une meilleure perméabilitécérébrale (B).
b-Résistance à la protéolyse plasmatique
La résistance des peptides inhibiteurs à la protéolyse a été testée in vitro et in vivochez des rats. Dans les études in vitro, les peptides inhibiteurs, méthylés ou non, ont étéincubés en présence de chimiotrypsine (1%, w/v) pendant 24 H à 37°C. La quantité de peptidenon digérée, évaluée par HPLC en phase inverse, a mis en évidence que les peptidesinhibiteurs, lorsqu’ils étaient méthylés, ne subissaient pas de dégradation enzymatique. Lespeptides non méthylés présentent des sites de coupures et sont donc digérés (A).
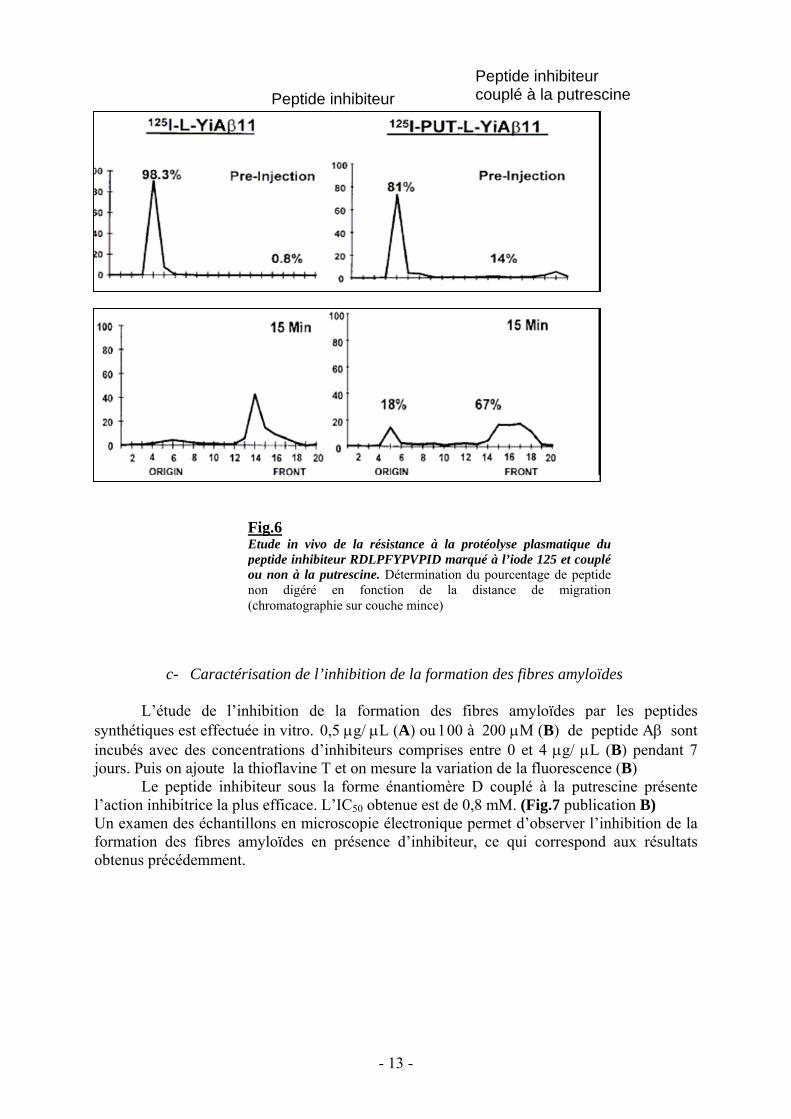
Les études in vivo ont été faites quand à elles sur des peptides inhibiteurs portant ounon un résidu putrescine, marqué à l’iode 125 I. Après injection dans la veine brachiale de rat,des cinétiques de prélèvements sanguins ont été réalisées, entre le moment de l’injection et20 minutes plus tard. Après centrifugation du sang, le plasma des différents prélèvements estanalysé en chromatographie sur couche mince. Les peptides intacts marqués ne migrentpratiquement pas et se retrouvent sur la ligne de dépôt, alors que les peptides dégradés, pluspetits, vont migrer avec le front du solvant. (Fig.6 publication B).
Au bout de 15 minutes, 18% de peptides couplés à la putrescine sont intacts alors quetous les peptides non couplés à la putrescine ont été dégradés (B).
- 13 -
c- Caractérisation de l’inhibition de la formation des fibres amyloïdes
L’étude de l’inhibition de la formation des fibres amyloïdes par les peptidessynthétiques est effectuée in vitro. 0,5 μg/ μL (A) ou 100 à 200 μM (B) de peptide Aβ sontincubés avec des concentrations d’inhibiteurs comprises entre 0 et 4 μg/ μL (B) pendant 7jours. Puis on ajoute la thioflavine T et on mesure la variation de la fluorescence (B)
Le peptide inhibiteur sous la forme énantiomère D couplé à la putrescine présentel’action inhibitrice la plus efficace. L’IC50 obtenue est de 0,8 mM. (Fig.7 publication B)Un examen des échantillons en microscopie électronique permet d’observer l’inhibition de laformation des fibres amyloïdes en présence d’inhibiteur, ce qui correspond aux résultatsobtenus précédemment.
Peptide inhibiteurPeptide inhibiteurcouplé à la putrescine
Fig.6Etude in vivo de la résistance à la protéolyse plasmatique dupeptide inhibiteur RDLPFYPVPID marqué à l’iode 125 et coupléou non à la putrescine. Détermination du pourcentage de peptidenon digéré en fonction de la distance de migration(chromatographie sur couche mince)
- 14 -
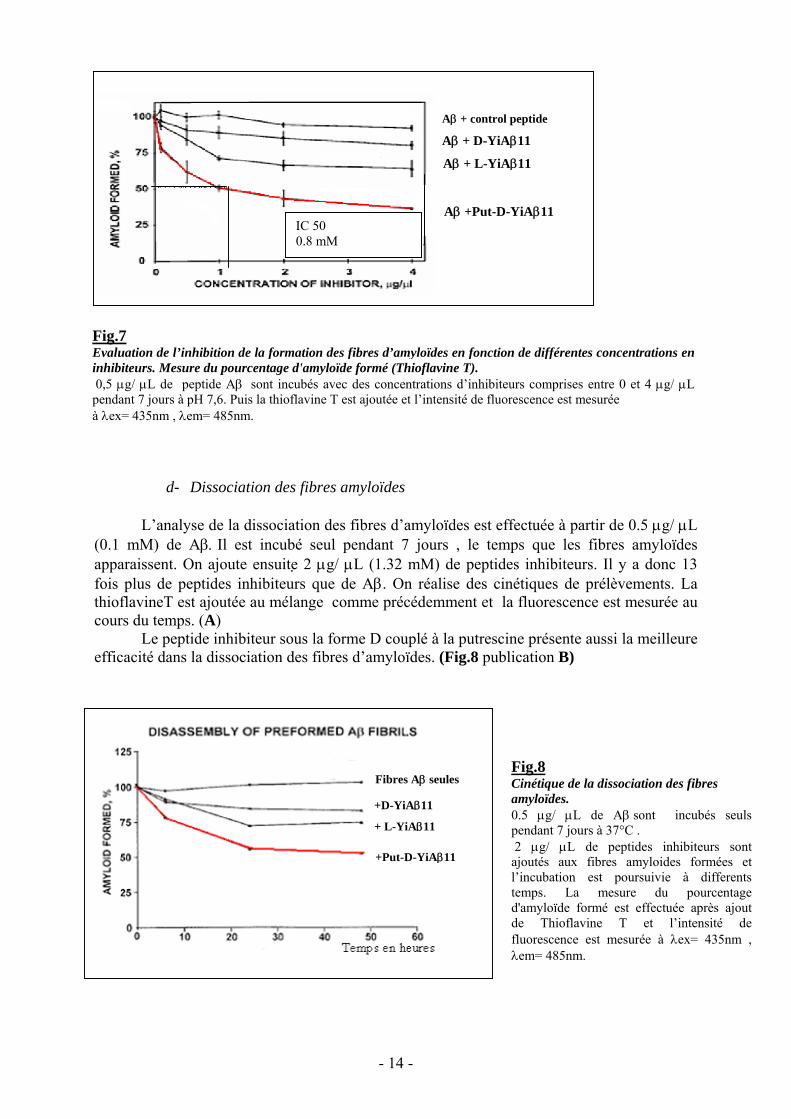
d- Dissociation des fibres amyloïdes
L’analyse de la dissociation des fibres d’amyloïdes est effectuée à partir de 0.5 μg/ μL(0.1 mM) de Aβ. Il est incubé seul pendant 7 jours , le temps que les fibres amyloïdesapparaissent. On ajoute ensuite 2 μg/ μL (1.32 mM) de peptides inhibiteurs. Il y a donc 13fois plus de peptides inhibiteurs que de Aβ. On réalise des cinétiques de prélèvements. LathioflavineT est ajoutée au mélange comme précédemment et la fluorescence est mesurée aucours du temps. (A)
Le peptide inhibiteur sous la forme D couplé à la putrescine présente aussi la meilleureefficacité dans la dissociation des fibres d’amyloïdes. (Fig.8 publication B)
Fig.8Cinétique de la dissociation des fibresamyloïdes.0.5 μg/ μL de Aβ sont incubés seulspendant 7 jours à 37°C . 2 μg/ μL de peptides inhibiteurs sontajoutés aux fibres amyloides formées etl’incubation est poursuivie à differentstemps. La mesure du pourcentaged'amyloïde formé est effectuée après ajoutde Thioflavine T et l’intensité defluorescence est mesurée à λex= 435nm ,λem= 485nm.
IC 500.8 mM
Aβ + control peptide
Aβ + D-YiAβ11
Aβ + L-YiAβ11
Aβ +Put-D-YiAβ11
Fig.7Evaluation de l’inhibition de la formation des fibres d’amyloïdes en fonction de différentes concentrations eninhibiteurs. Mesure du pourcentage d'amyloïde formé (Thioflavine T). 0,5 μg/ μL de peptide Aβ sont incubés avec des concentrations d’inhibiteurs comprises entre 0 et 4 μg/ μLpendant 7 jours à pH 7,6. Puis la thioflavine T est ajoutée et l’intensité de fluorescence est mesuréeà λex= 435nm , λem= 485nm.
Fibres Aβ seules
+D-YiAβ11
+ L-YiAβ11
+Put-D-YiAβ11

- 15 -
3-RESULTATS
3-1 Homologie de séquence
Dans les nombreuses études réalisées sur le peptide β amyloïde, il a été mis enévidence que la région centrale hydrophobe du peptide a une grande importance, tant auniveau de la formation des fibres amyloïdes que de la toxicité (11).
Par ailleurs, une homologie de séquence est nécessaire pour que les peptidesinhibiteurs puissent interagir avec le peptide Aβ, mais elle ne doit pas être totale. En effet,dans ce cas, on obtient alors des peptides inhibiteurs qui présentent une forte affinité pour Aβ,mais qui se trouvent incapables de déstabiliser les feuillets β et donc d’empêcher leurincorporation ultérieure. Enfin, il a été mis en évidence que les peptides inhibiteurs portant denombreux groupements bloquants n’avaient qu’une faible affinité pour Aβ (17, 19)
3-2 Résultats obtenus avec les peptides inhibiteurs
Les peptides inhibiteurs testés dans ces deux publications présentent une capacitéd’inhibition de la formation des fibres amyloïdes. Ces observations ont été mises en évidencepar deux techniques, la mesure de la fluorescence et la microscopie électronique.
Dans la publication B, les peptides ayant une modification putrescine en C terminalepermettent une amélioration de la pénétration de la barrière méningée (taux de pénétrationmultiplié par 5 à 7 fois). Il a eu aussi un effet positif vis à vis de la protéolyse plasmatique, enprotégeant les peptides inhibiteurs. Ainsi, on retrouve environ 18 % de peptide intact au boutde 15 minutes, alors que les peptides non protégés sont dégradés après une incubation de 1minute(B). Ce résultat confirme que l’addition de polyamines naturelles, telles que laputrescine, la spermine ou la spermidine influence de façon intéressante la perméabilitécérébrale. Initialement, leur pénétration semblait être liée à des interactions électrostatiquesentre les polyamines, chargées positivement et les charges négatives portées par les cellulesendothéliales (20).Toutefois, l’existence d’un transporteur spécifique des polyamines a étémis en évidence dans les mitochondries de certains tissus chez la souris (20).
Par ailleurs, il a été montré que des peptides inhibiteurs contenant des résidus prolinesprésentaient aussi une forte capacité d’inhibition (17,19,22).
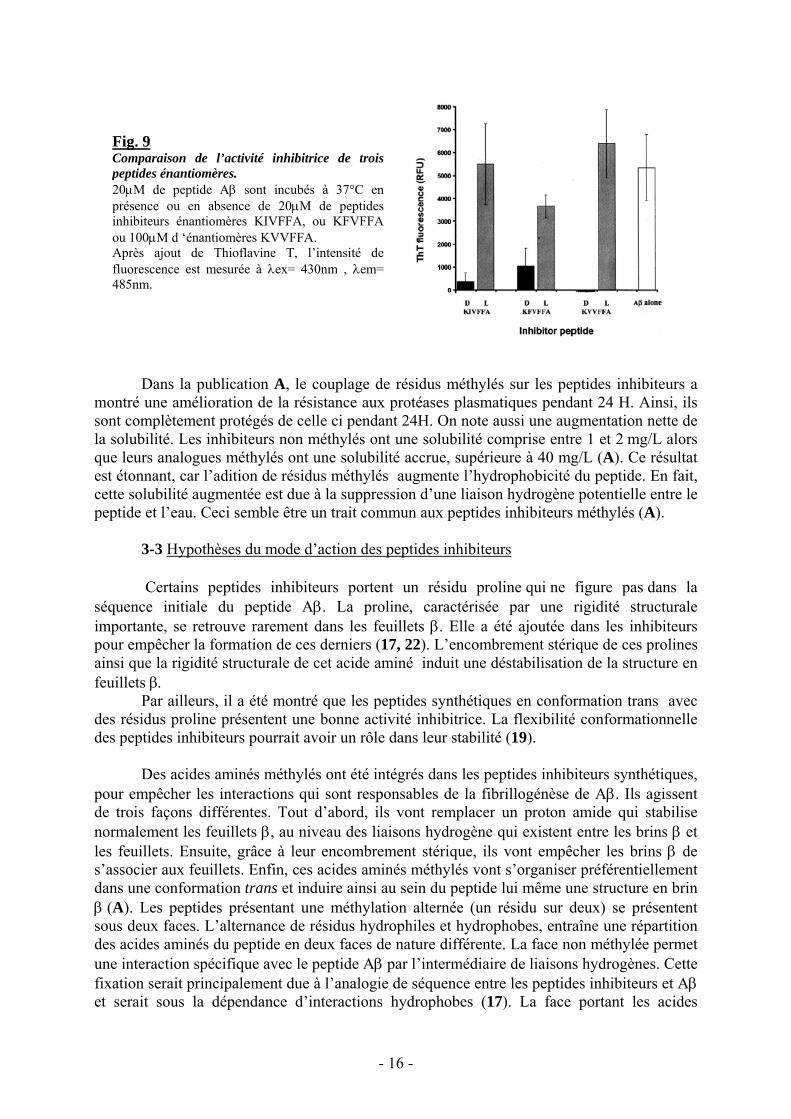
Les chercheurs ont aussi mis en évidence que l’utilisation d’un peptide inhibiteur sousla forme D donnait de meilleurs résultats que sous la forme L. Ce phénomène a été confirmédans une étude récente (21) qui a porté sur la comparaison de l’activité inhibitrice de 3peptides sous les formes D et L. Pour chacun de ces peptides, la forme L montre unesimilitude de structure en feuillets β avec le peptide β amyloïde, alors que les formes Dprésentent une inhibition nette (Fig. 9 extraite de la publication 21).
- 16 -
Dans la publication A, le couplage de résidus méthylés sur les peptides inhibiteurs amontré une amélioration de la résistance aux protéases plasmatiques pendant 24 H. Ainsi, ilssont complètement protégés de celle ci pendant 24H. On note aussi une augmentation nette dela solubilité. Les inhibiteurs non méthylés ont une solubilité comprise entre 1 et 2 mg/L alorsque leurs analogues méthylés ont une solubilité accrue, supérieure à 40 mg/L (A). Ce résultatest étonnant, car l’adition de résidus méthylés augmente l’hydrophobicité du peptide. En fait,cette solubilité augmentée est due à la suppression d’une liaison hydrogène potentielle entre lepeptide et l’eau. Ceci semble être un trait commun aux peptides inhibiteurs méthylés (A).
3-3 Hypothèses du mode d’action des peptides inhibiteurs
Certains peptides inhibiteurs portent un résidu proline qui ne figure pas dans laséquence initiale du peptide Aβ. La proline, caractérisée par une rigidité structuraleimportante, se retrouve rarement dans les feuillets β. Elle a été ajoutée dans les inhibiteurspour empêcher la formation de ces derniers (17, 22). L’encombrement stérique de ces prolinesainsi que la rigidité structurale de cet acide aminé induit une déstabilisation de la structure enfeuillets β.
Par ailleurs, il a été montré que les peptides synthétiques en conformation trans avecdes résidus proline présentent une bonne activité inhibitrice. La flexibilité conformationnelledes peptides inhibiteurs pourrait avoir un rôle dans leur stabilité (19).
Des acides aminés méthylés ont été intégrés dans les peptides inhibiteurs synthétiques,pour empêcher les interactions qui sont responsables de la fibrillogénèse de Aβ. Ils agissentde trois façons différentes. Tout d’abord, ils vont remplacer un proton amide qui stabilisenormalement les feuillets β, au niveau des liaisons hydrogène qui existent entre les brins β etles feuillets. Ensuite, grâce à leur encombrement stérique, ils vont empêcher les brins β des’associer aux feuillets. Enfin, ces acides aminés méthylés vont s’organiser préférentiellementdans une conformation trans et induire ainsi au sein du peptide lui même une structure en brinβ (A). Les peptides présentant une méthylation alternée (un résidu sur deux) se présententsous deux faces. L’alternance de résidus hydrophiles et hydrophobes, entraîne une répartitiondes acides aminés du peptide en deux faces de nature différente. La face non méthylée permetune interaction spécifique avec le peptide Aβ par l’intermédiaire de liaisons hydrogènes. Cettefixation serait principalement due à l’analogie de séquence entre les peptides inhibiteurs et Aβet serait sous la dépendance d’interactions hydrophobes (17). La face portant les acides
Fig. 9Comparaison de l’activité inhibitrice de troispeptides énantiomères.20μM de peptide Aβ sont incubés à 37°C enprésence ou en absence de 20μM de peptidesinhibiteurs énantiomères KIVFFA, ou KFVFFAou 100μM d ‘énantiomères KVVFFA.Après ajout de Thioflavine T, l’intensité defluorescence est mesurée à λex= 430nm , λem=485nm.

- 17 -
aminés méthylés inhibe la formation de liaisons hydrogène entre deux feuillets voisins. Parailleurs, il y a une modification de l’encombrement stérique à ce niveau, ce qui va induire unedéstructuration des empilements de feuillets β.
Il semblerait que la conformation antiparallèle entre les peptides inhibiteurs et lesfeuillets β soit la meilleure hypothèse possible. En effet, cette conformation serait plus stableet favoriserait les interactions entre les résidus lysine et glycine de l’inhibiteur et du peptideAβ.
Une étude récente portant sur un peptide inhibiteur de petite taille contenant uneproline (séquence KPLVFF) a permis d’éclaircir les mécanismes de l’interaction entre lespeptides inhibiteurs et le peptide Aβ 1-40 (23). Ce peptide porte un résidu proline, et unrésidu chargé a été ajouté à l’extrémité C terminale pour augmenter la solubilité de cetinhibiteur. L’addition du peptide inhibiteur en excès (20 fois) a permis de dissocier les fibresde Aβ.
Les fibres de Aβ 1-40 sont composées de deux brins β séparées par un coude de 180°.Les brins β forment des feuillets qui interagissent essentiellement par le biais de liaisonshydrogènes intramoléculaires. A la surface d’un feuillet β, les résidus chargés positivement(K16) et négativement (D22) du premier brin sont disposés en ligne le long de l’axe de lafibre. Les deux lignes chargées sont séparées par une bande hydrophobe et aromatiqueconstituée par les acides aminés V 18 et F 20. Le côté hydrophobe du deuxième brin formeune face hydrophobe de l’autre côté du feuillet (Fig. 10 extraite de la publication 23)
Deux positions de ce petit peptide inhibiteur sont possibles : il peut y avoir uneorientation dans laquelle les deux charges négatives du ligand sont orientées du côté de labande portant les résidus chargés positivement (K16) ou alors ces deux charges ont uneorientation opposée, dans laquelle la charge positive de l’extrémité N terminale va avec
Inhibiteur LFFPDinverse de la séquence homologue
Fig.10The model of iAβ5inv bound to Aβ 1-40 fibrilsusing the orientational restraints from trRDC(transferred residual dipolar coupling) data.

- 18 -
l’extrémité des charges négatives de E 22 à la surface de la fibre amyloïde. Toutefois, lapremière hypothèse semble la meilleure pour des raisons énergétiques.
Cette étude a permis de mettre en évidence que les résidus K 16 et F 20 de la fibre βamyloïde servaient de point de fixation pour le peptide inhibiteur. D’autres peptidesinhibiteurs sont associés de façon analogue au niveau des fibres. La compétition entre lesinteractions hydrophobes et électrostatiques en même temps que l’incapacité à former desliaisons hydrogène dans les feuillets β provoque un affaiblissement des liaisons hydrogènedans les feuillets des fibres d’amyloïde. Ce dernier phénomène serait responsable de ladissolution des fibres amyloïdes.
4- DISCUSSION ET PERSPECTIVES
Dans les publications A et B, l’accent est mis majoritairement sur la capacité qu’ontles peptides inhibiteurs à empêcher la formation des fibres amyloïdes ou à permettre leurdissociation. Les peptides inhibiteurs testés sont relativement grands, autour de 11 résidus, etsont donc potentiellement immunogènes du fait de leur taille. Actuellement, les études portentsur des peptides bien plus petits, constitués d’environ 6 résidus (23).
La fibrillogénèse étant un phénomène cinétique, on ne sait pas si les peptidesinhibiteurs ont une action temporaire ou définitive sur l’inhibition de celle ci in vivo.
Les mécanismes de l’interaction entre les peptides inhibiteurs et le peptide Aβ ne sontpas encore totalement élucidés aujourd’hui, malgré de nombreuses études portant sur desessais de modélisation de celle-ci ou utilisant la RMN. Toutefois, des analyses de dichroïsmecirculaire mettant en présence les peptides inhibiteurs et le peptide Aβ ont permis deconfirmer la désorganisation des feuillets β (18).
Par ailleurs, il a été montré que les peptides inhibiteurs méthylés ou portant des résidusproline avaient une activité similaire. Bien que le nombre de dérivés méthylés impliqués dansl’inhibition n’aie pas été étudié et optimisé, il pourrait être judicieux de construire un peptideinhibiteur combinant toutes ces possibilités et donc constitué de la façon suivante
- taille la plus petite possible- résidu chargé à l’extrémité C terminale- sous forme énantiomèrique D- portant des prolines- portant des résidus méthylés alternés.
Cependant, il est difficile d’analyser les résultats obtenus avec d’autres résultats publiés.En effet, chaque groupe étudie l’action des inhibiteurs avec des moyens et des techniques quilui sont propres, ce qui rend quasiment impossible la comparaison entre les différentsinhibiteurs en l’absence d’évaluation de leurs constantes d’affinité respectives (Kd)
Par ailleurs, lorsque l’on se réfère aux différentes publications portant sur l’utilisation depeptides inhibiteurs, on se rend compte qu’ils ont été surtout utilisés pour tenter d’élucider lesmécanismes de l’interaction avec le peptide Aβ, mais qu’ils ne constituent pas de réelle pistequand à un usage thérapeutique. De plus, l’interaction des peptides inhibiteurs étant« séquence dépendante », ils ne pourraient être utilisés dans d’autres maladies. Ainsi, unpeptide inhibiteur méthylé contre une autre protéine fibrillaire, la protéine du prion, n’amontré aucune efficacité quant à l’inhibition de la fibrillogénèse due au peptide Aβ (A).
Dans une publication récente, grâce à la résonance plasmonique de surface, deschercheurs ont montré que la constante de dissociation de certains peptides inhibiteurs était

- 19 -
voisine de 30 μM (24). Par conséquent, il semblerait que les peptides inhibiteurs testés n’aientpas une très grande affinité vis à vis du peptide Aβ. Ces tests ayant été réalisés in vitro, onpeut se poser la question quand à l’efficacité de tels inhibiteurs in situ.
Nouvelles pistes
Les dénominateurs communs des maladies neurodégénératives sont des réactionsmétallochimiques. Il a été montré que la concentration cérébrale en métaux lourds étaitélevée dans ces derniers cas et pouvait créer un environnement pro-oxydatif. Le peptide Aβforme des agrégats lorsque l’on baisse le pH, ou lorsque l’on ajoute des ions métalliques. Desétudes ont mis en évidence le rôle du cuivre dans les maladies neurodégénératives comme lamaladie d’Alzheimer (25, 26, 27, 28). A l’aide de la spectroscopie Raman, il a été montré quele cuivre se fixait au niveau de l’atome d’azote porté par les trois résidus histidine du peptideAβ. Par conséquent, le cuivre semble être impliqué au niveau de l’agrégation des feuillets βpar l’intermédiaire des résidus histidine. Un chélateur spécifique du fer, mais ayant aussi unetrès grande affinité pour l’aluminium, le zinc et le cuivre, la desferioxamine (DFO), a montréson potentiel dans la dissolution des plaques. Malheureusement, ce produit est neurotoxique etprovoque des graves effets secondaires. Il est faiblement absorbé au niveau intestinal, etrapidement dégradé après administration. Enfin, sa nature hydrophobe fait qu’il ne pénètreque très difficilement au niveau de la barrière des méninges (28).
D’autres expériences ont utilisé des chélateurs du cuivre. Post mortem, on a pu mettreen évidence qu’ils augmentaient la solubilité de Aβ dans le tissu cérébral de patients ou desouris trangéniques (30). Par conséquent, la recherche de chélateurs dépourvus de toxicitépourrait s’avérer être une piste prometteuse.
Des études faites sur des nanoparticules (moins de 300 nm de diamètre) recouvertes desurfactants ont montré leur potentiel vis à vis du transport de différents produits. Ellespourraient être utilisées pour faciliter la pénétration des substances d’intérêt dans le cerveau, àtravers la barrière cérébrale. Des groupements amines ou carboxyles fixés à la surface de cesnanoparticules pourraient se lier de façon covalente avec des chélateurs de métaux. Enmodifiant des paramètres de surface (par exemple avec des surfactants), on a montré que lecomplexe « nanoparticule-chélateur » était absorbé par l’Apo A1 ou par l’Apo E. En fait, ilmimerait les apolipoprotéines, et serait donc capable de traverser les méninges avec lemécanisme de transport des LDL. On pourrait alors envisager de recouvrir des nanoparticulesavec des apolipoprotéines, puis de les coupler avec des chélateurs de métaux. Ainsi, ellespourraient jouer un rôle très actif dans l’élimination des ions métalliques en excès : arrivéesjusqu’au cerveau, elles fixeraient alors les métaux présents et retraverseraient les barrièrescérébrales pour rejoindre la circulation sanguine (28).
Des substances empêchant l’auto-agrégation de Aβ, des mimétiques de glycosaminoglycanes ont passé avec succès les essais cliniques de phase II. Ils sont actuellement testés enphase III, mais les résultats concernant ces essais ne sont pas publiés à ce jour (33).
Une nouvelle stratégie pour prévenir la maladie d’Alzheimer porte sur les sécrétases etl’inhibition de la γ et de la β sécrétases. La structure de la γ sécrétase vient d’être élucidée(29). Il s’agit en fait d’un complexe formé de quatre protéines transmembranaires, dont lesprésénilines 1 et 2. Des inhibiteurs de ce complexe ont été dessinés, et présentent unehomologie de séquence avec le site de clivage de l’APP. L’utilisation du programmeinformatique ROCS a permis de trouver des molécules ayant une structure tridimensionnellesimilaire. Ces molécules ont été testées sur des cellules CHO, et leurs IC50, déterminés avec

- 20 -
des tests Elisa, donnent des valeurs comprises entre 0,53 μM et 0,64 μM (32). L’inhibitiondes γ sécrétases a bien une action inhibitrice sur la production de peptide Aβ.Malheureusement, des études ont montré qu’il y avait des effets négatifs : on assiste enparallèle à une inhibition de la maturation ou de la différenciation de certaines lignéescellulaires, ainsi qu’à des modifications graves du tractus gastro-intestinal.
La structure de la β sécrétase, quand à elle, est connue depuis 1999. On a montré quecelle-ci était une enzyme transmembranaire, de type aspartic protéase, appelée encore BACE(pour « Beta site APP Cliving Enzyme »). Des substances peptidomimétiques inhibantl’activité de la β sécrétase ont été testées, et ont montré des IC 50 ≈ 4 μM (31). Par ailleurs,des souris délétées pour BACE n’ont présenté aucun problème (au niveau de la croissance,des résultats biochimiques, hématologiques, morphologiques ou comportementaux), ce qui estencourageant. Enfin, la structure tridimensionnelle de la β sécrétase est disponible, et on a pumettre en évidence la présence d’un large site actif (33). Les tests sur l’inhibition de la βsécrétase sont actuellement en phase I.
Au vu des problèmes qui se posent avec l’inhibition totale d’une de ces enzymes, onpourrait mettre en œuvre une inhibition partielle, pour tenter de diminuer les effetsindésirables cités ci-dessus, tout en permettant une réduction de la production de peptide Aβ.On empêcherait ainsi la survenue de la maladie en traitant les gens de façon préventivependant les 10 ou 30 dernières années de la vie.

- 21 -
Références bibliographiques
A- « Inhibition of β amyloid (40) fibrillogenesis and disassembly of β amyloid (40)fibrils by short β amyloid congeners containing N-methyl amino acids at alternateresidues »David J. Gordon, Kimberly L. Wsciarretta, and Stephen C. MeredithBiochemistry 2001, 40, 8237-8245
B- “β sheet breaker peptide inhibitor of Alzheimer’s amyloidogenesis with increasedblood-brain barrier permeability and resistance to proteolytic degradation in plasma”Joseph F. Poduslo, Geoffry L. Curran, Asok Kumar, Blas Frangione, and Claudio Soto.Inc. J. neurobiol 39: 371-382 1999
1- O. Zékri « Pharmacologie de la maladie d’Alzheimer »www.med.univ-rennes1.fr/etud/pharmaco/alzheimer.htm
2- R. B. Maccioni, J. P. Munoz and L. Barbeito“The molecular bases of Alzheimer’s disease and other neurodegenerative disorders”ReviewMedical Research, 2001 Vol 32 pp. 367-381
3- B. Clippingdale, J. D. Wade and C. J. Barrow “The amyloid β peptide and its role in Alzheimer’s disease”ReviewJournal of Peptide Science, 2001, Vol 7, pp. 227-249
4- W. Xia, J. Zhang, R. Perez, E. H. Koo and D. J. Selkoe“Interaction between amyloid precursors protein and presenilins in mammalian cells:Implications for the pathogenesis of Alzheimer disease”Proc. Natl. Acad. Sci. USA, 1997, Vol 94, pp.8208-8213
5- B. Permanne, C. Adessi, G. P. Saborio, S. Fraga, M. J. Frossard, J. Van Dorpe, I.dewacgter, W. A. Banks, F. Van Leuven and C. Soto« Reduction of amyloid load and cerebral damage in a trangenic mouse model ofAlzheimer’s disease by treatment with a β sheet breaker peptide »The FASEB Journal, 2002, Vol 16, pp 860-862
6- K.Pafel, T. Vagt and K. Koksch“Directing the secondary structure of polypeptides at will: from helices to amyloïd andback again?”Org. Biomol. Chem, 2005 Vol 3, pp. 3843-3850

- 22 -
7- P.R Turner, O. Connor, W. P. Tate and W. C. Abraham“Role of amyloid precursor protein and its fragment in regulating neural activity,plasticity and memory »ReviewProgress in Neurobiology 70, 2003, pp. 1-32
8- L. Conti and E. Cattaneo“Controlling neural stem cell division within the adult subventricular zone: anAPPealing job”Trends in neurosciences, 2005, Vol 28 N° 2, pp.57-59
9- S. Gandy“The role of cerebral amyloid β accumulation in common forms of Alzheimer’sdisease”Journal of Clinical Investigation, 2005, Vol 115 N° 5, pp. 1121-1129
10- C. Soto“Alzeimer’s and prion disease as disorders of protein conformation: implications forthe design of novel therapeutic approaches”ReviewJ.Mol.Med, 1999, 77: pp. 412-418
11- R.Liu, C. MacAllister, Y. Lyubchenko, and M. R. Sierks“Residues 17-20 and 25-35 of beta amyloid plays critical roles in aggregation”Journal of Neurosciences Research, 2004, Vol 75, pp. 162-171
12- C. Soto, M. S. Kindy, M. Baumann and B. Frangione“ Inhibition of Azheimer’s amyloidosis by peptides that prevent β sheet conformation”Biomedical and Biophysical Research Communication 1996, Vol 226, pp. 672-680
13- L. O. Tjernberg, J. Näslund, F. Lindqvst, J. Johansson, A. R. Karlström,J. Thyberg, L.Terenius and C. Nordstedt“Arrest of β amyloid fibril formation by a pentapeptide ligand”The Journal of Biological Chemistry1996, Vol 271 N° 15, pp. 8545-8548
14- A. Lomakin, D.S. Chung, G.B. Benedek, D. A. Kirschner and D. B. Teplow“On the nucleation and growth of amyloid b protein fibrils: Detection of nuclei andquantification of rate constants”.Proc. Natl. Acad. Sci. USA, 1996, Vol 93, pp. 1125-1129
15- L. C. Serpell, C. C. F. Blake and P. E. Fraser“Molecular structure of a fibrillar Alzheimer’s Aβ fragment”Biochemistry, 2000, Vol 39, pp. 13269-13275
16- E. Zerovnik“Amyloid fibril formation”ReviewEur . J. Biochem, 2002, Vol 269, pp. 3362- 3371

- 23 -
17- C. Adessi and C. Soto“Beta sheet breaker strategy for the treatment of Alzheimer’s disease”Drug Development Research, 2002, Vol 56, pp. 184-193
18- E. R. L. C. Vardy, A. J. Catto and N. M. Hooper“Proeolytic mechanisms in amyloid β metabolism: therapeutic implications forAlzheimer ‘s disease”
19- C Adessi, M. J. Frossard, C. Boissard, S. Fraga, S. Bieler, T. Ruckle, F. Vilbois, S. M.Robinson, M. Mutter, W. A. Bank and C. Soto“Pharmalogical profiles of peptide drug candidates for the treatment of Alzheimer’sdisease”The Jounal of Biological Chemistry, 2003, Vol 278 N° 16, pp. 13905-13911
20- J. F. Poduslo and G. L. Curran“Polyamine modification increase the permeability of proteins at the blood –nerve andblood-brain barriers”Journal of Neurochemistry, 1996, Vol 66 N° 4, pp. 1599-1609
21- R. J. Chalifour, R. W. McLaughin, L. Lavoie, C. Morisette, N. Tremblay, M. Boulé, P.Sarrazin, D. Stéa, D. Lacombe, P. Tremblay and F. Gervais“Stéreoselective interactions of peptide inhibitors with the β amyloid peptide”The Journal of Biological Chemistry, 2003, Vol 278 N° 37; pp. 34874-34881
22- A. Morimoto, K. Irie, K. Murakami, Y. Masuda, H. Ohigashi, M. Nagao, H. Fukuda, T.Shimizu and T. Shirasawa“Analysis of the secondary structure of b amyloid Aβ42 fibrils by systematic prolinereplacement”Journal of Biochemistry, 2004, Vol 279 N° 50, pp. 52781-52788
23- Z. Chen, G. Krause and B. Reif“Structure and orientation of peptide inhibitors bound to beta amyloid fibrils”J. Mol. Biol, 2005, Vol 20, pp. 1-17
24- C. W. Cairo, A. Strzelec, R. M. Murphy and L. L. Kiessling“Affinity based inhibition of b amyloid toxicity”Biochemistry, 2002, Vol 41, pp. 8620-8629
25- C. C. Curtain, F. E. Ali, D. G. Smith, A. I. Bush, C. L. Masters and K. J. Barnham“Metal ions, pH, and cholesterol regulate the interactions of Alzheimer’s diseaseamyloid b peptide with membrane lipids”The Journal of Biological Chemistry, 2003, Vol 278 N° 5; pp. 2977-2982
26- D. J. Waggoner, T. B. Bartnikas and J. D. Gitlin“The role of copper in neurodegenerative disease”ReviewNeurobiology of Disease, 1999, Vol 6, pp. 221-230

- 24 -
27- K. Pagel, T. Vagt, T. Kohajda and B. Koksch“From α helix to β sheet: a reversible metal ion induced peptide secondary structureswitch”Org. Biomol.Chem, 2005, Vol 3, pp.2500-2502
28- G. Liu, M. R. Garett, P. Men, X. Zhu, G. Perry and M. A. Smith“Nanoparticle anf other metal chelation therapeutics in Alzheimer disease”Biochemica et Biophysica Acta, 2005, Vol 1741, pp. 246-252
29- W. T. Kimberly and M. S. Wolfe“Identity and function of γ secretase”Journal of Neuroscience Research, 2003, Vol 74, pp. 353-360
30- D.Strausak, J. F. B. Mercer, H. H. Dieter, W. Stremmel and G. Multhaup“Coper in disorderswith neurological symptoms: Alzheimer’s disease, Menkes andWilson diseases”Brain Reseach Bulletin, 2001, Vol 55 N° 2, pp. 175-185
31- M. Citron“β secretase inhibition for the treatment of Alzheimer’s disease: promise andchallenge”ReviewTrens in Pharmalogical Sciences, 2004, Vol 25 N° 2, pp.92-97
32- E. Gundersen, K. Fan, K. Haas, D. Huryn, J. S. Jacobsen, A. Kreft, R. Martone, J.Sonnenberg-Reines, S. C. Sun and H. Zhou“Molecular-modeling based design, synthetis and activity of substituted piperidines asγ secretase inhibitors”Bio-organic and Medicinal Chemistry Letters, 2005, Vol 15, pp. 1892-1894
33- M. Citron“Strategies for disease, modification in Alzheimer’s diseasse”ReviewNature review Neurosciences, 2004, Vol 5, pp.677-685
34- D. Chhabra, N. J. Nosworthy and C. G. dos Remedios“The N-terminal fragment of gelsolin inhibits the interactionof DNaseI with isolatedactin, but not with the cofilin-actin complex”Proteomics, 2005, Vol 2, pp. 3131-3136
35- T. L. Carter, S. Pedrini, J. Ghiso, M. E. Ehrlich and S. Gandy“Brain neprilysin activity and susceptibility to transgene induced Alzheimeramyloidosis”Neurosciences Letters XXX, 2005, XXX-XXX, pp. 1-5
36- H. Qiao, R. C. Koya, K. Nakagawa, H. Tanaka,H. Fujita, M. Takimoto, N. Kuzumaki« Inhibition of Alzheimer’s amyloid-peptide-induced reduction of mitochondrialmembrane potential and neurotoxicity by gelsolin”Neurobiology of Aging, 2005, Vol 26, pp. 849–855





























