Embed Size (px)
Citation preview

Original article
Myosin expression in semitendinosus muscleduring fetal development of cattle:
immunocytochemical and electrophoretic analyses
J Robelin B Picard A Listrat C JurieC Barboiron F Pons Y Geay
1 INRA-Theix, laboratoire Croissance et Métabolismes des Herbivores,63122 Saint-Genès-Champanelle;
2 INSERM-U300, Unité de Recherche de Physiopathologie Cellulaire et Moléculaire,Av Charles Flahaut, 34100 Montpellier, France
(Received 12 March 1992; accepted 29 October 1992)
Summary ― The pattern of expression of different types of myosin and the development of differentmuscle cell populations were studied in the semitendinosus muscle of cattle from 39 d of gestationto 30 d of post-natal life. Monoclonal antibodies specific to different myosin heavy chains were used.Two cell generations were identified during myogenesis. They appeared successively and werecharacterized by different patterns of expression of myosins. The first population, which was presentfrom the first stage studied (39 d of gestation), gave rise to type I fibers, which, in the mature animal,express only slow myosin. A second generation became differentiated at about 120 d of fetal life andthen developed into type II fibers (Ila, lib or llc). The beginning of differentiation was characterized inall the cell populations by the expression of specific types of embryonic or fetal myosins.A comparison of these results with findings from previous works shows a marked similarity betweenspecies in the pattern of myogenesis but great differences in the length of the different stages of de-velopment. In this respect, myogenesis in cattle closely resembles that in man.
bovine I fetus I muscle fiber I differentiation I myosin
Résumé ― Expression de différentes formes de myosine dans le muscle semitendinosus aucours du développement foetal chez le bovin : étude immunocytochimique et électrophoréti-que. Le profil d’expression des différents types de myosines, et le développement de différentes gé-nérations de cellules musculaires ont été étudiés au cours de la vie fcstale chez le bovin, à l’aide
d’anticorps monoclonaux dirigés contre les chaînes lourdes de myosines. On a identifié 2 popula-tions cellulaires apparaissant successivement au cours du développement, et caractérisées par desprofils d’expression des myosines différents. La première population, déjà pésente lors du premierstade fcetal étgudié (39 j de gestation), est à l’origine des fibres de type 1 qui expriment chez l’adulteune myosine de type lent. Une seconde population est reconnaissable dès l’âge fcetal de 120 j, etconduit ultérieurement aux fibres de type Il (Ila, Ilb et Ilc). Enfin, le début de la différenciation desfibres musculaires est caractérisé dans les 2 générations de cellules par l’expression de myosinesspécifiques de type fcetal ou embryonnaire que l’on ne retrouve pas chez l’adulte. La comparaisonde ces résultats avec ceux obtenus chez d’autres mammifères fait apparaître une grande similarité

entre espèces dans le profil général de la myogenèse, mais aussi des différences importantes dansla durée des différentes étapes du développement musculaire. À cet égard, la myogenèse chez lebovin ressemble beaucoup à ce que l’on observe chez l’homme.
bovin / fcetus / fibre musculaire / differenciation / myosine
INTRODUCTION
In mammals and birds, most muscles aremade up of a heterogeneous population ofmuscle fibers. In the adult animal, these fi-bers are classified into 3 types (I, Ila and
lib) according to their rate of contractionand energy metabolism (for a review, seeGauthier, 1986). The classification of fibertypes in the mature animal was first estab-lished by histochemical and histoenzymat-ic methods (Brooke and Kaiser, 1970).The characteristics of muscle fibers are re-lated to the expression of different types ofcontractile proteins, particularly myosins(Lowey, 1986).
The development of the different fibertypes occurs in the process of differentia-tion and growth of the muscle cells duringembryonic and fetal life (for a review, seeMuntz, 1990). In adult animals, the pheno-type of a muscle fiber results from a com-bination of genetic and environmental ef-fects, in which innervation is thought toplay a role (Pette and Vrbova, 1985). How-ever, it is not sufficient to account for fiber
type diversity, as shown by Hughes andBlau (1992) on the basis of earlier work(Butler et al, 1982; Harris et al, 1989; Con-don et al, 1990)..
Enormous advances in the study of em-bryonic origin of the muscle fiber typeshave been made by the development ofmonoclonal antibodies raised againstdifferent types of myosins (Gauthier et al,1978) and by the use of molecular probescorresponding to these proteins (Lyons et
al, 1990; Miller, 1990). The most detailedresults have been obtained in birds (Band-man et al, 1982; Crow and Stockdale,1986; Miller and Stockdale, 1986; for
reviews, see Stockdale and Miller, 1987,and Stockdale, 1990), in small mammals,such as mice (Rubinstein and Kelly, 1981;Whalen et al, 1981; Lyons et al. 1983;Vivarelli et al, 1988) and cats (Hoh andHughes, 1989). Studies have also beenmade on the human fetus (Pons et al,1986; Draeger et al, 1987). There are stilllarge gaps in our knowledge of how themuscle fibers develop into different types,in species whose advanced stage of ma-turity at birth (cattle, sheep, humans) sug-gests that their myogenesis is character-ized by specific features (Robelin et al,1991 ).
For this reason, we studied the expres-sion of different types of myosins duringmuscle differentiation in the fetal calf. Two
techniques were used, immunocytochem-istry, to identify the different types of mus-cle cells, and electrophoresis, followed byimmunoblotting to analyse the expressionof the different types of myosin in the mus-cle tissue.
This study shows that 2 distinct cell pop-ulations develop in succession during myo-genesis in the fetal calf, and describes thepattern of expression of the different typesof myosins in these populations. It alsoshows that there are many similarities be-tween cattle and humans in muscle diffe-
rentiation, especially in the temporal se-quence of events.

MATERIALS AND METHODS
Semitendinosus muscle was sampled from 10bovine fetuses, obtained from cows who hadbeen artificially inseminated and killed after ges-tation periods of 39, 53, 64, 84, 98, 122, 143,180, 224 and 270 d, and from 1 calf aged 30 dafter birth. Samples of the median part of themuscle were prepared and frozen in liquid nitro-gen as described previously (Robelin et al,1991 ).
lmmunocytochemistry
The myosins present at the different stages offetal life were evidenced by immunofluores-cence on serial frozen sections, 10 micronsthick. The procedure used was that describedby Pons et al (1986). We used 4 monoclonal an-tibodies (MAb) raised against different types ofhuman myosin heavy chains: a p type (slow)myosin from the heart ventricle of an adult (anti-body A), a fast myosin immunopurified from thequadriceps muscle of an adult (antibody B), amyosin from a 22-wk-old fetus (antibody C) anda myosin from adult atrium (antibody D). Thespecificity of these antibodies has been de-scribed in earlier works (Pons et al, 1986; Mariniet al, 1990). However, in preliminary experi-ments, we analysed their reaction with myosinfrom fetal and adult cattle. Immunofluorescenceand classical histochemistry (ATPase activity)analyses were carried out on frozen cut sectionsof cutaneus trunci muscle (entirely fast), masset-er muscle (entirely slow) and semitendinosusmuscle (predominantly fast) from an 8-yr-oldcow. They showed (data not presented) that an-tibodies A and B reacted with type I and type 11fibers respectively. Immunoblotting confirmedthat antibody A reacted with type I myosin andantibody B with the myosins of type Ila and Ilb.Antibody C did not react with adult muscle fibers(data not shown), but during fetal life reactedwith all muscle cells up to = 140 d of gestation,and with some until 220 d (results presented inthis article). Immunocytochemical analysis andimmunoblotting showed that antibody D raisedin human cardiac muscle reacted strongly withadult bovine cardiac muscle (data not shown). Italso reacted with fetal muscle cells at the begin-ning of differentiation (fig 1). However, it did notreact with any of the myosin heavy chain in im-
munoblotting analysis. Tests made with 2 otherantibodies raised against human cardiac muscleyielded similar results.
Electrophoresis
Samples of semitendinosus muscle were frozenin liquid nitrogen, stored at -80 °C and thenminced in a solution containing 30% (v/v) glyce-rol, 5% (w/v) sodium dodecyl sulfate (SDS), 1 MTris-HCI (pH 6.8), 7.5% (v/v) p-mercaptoethanofand 0.05% (w/v) pyronin Y. After 10 min at roomtemperature, the samples were heated for 10 minat 90 °C. An amount of solution corresponding to4 pg of proteins was placed in each well. Themyosin heavy chains (MHC) were separated on apolyacrylamide gradient gel 5-8% (Laemmli,1970; Bar and Pette, 1988) on plates measuring160 x 160 x 15 mm. As in the study of Sugiuraand Murakami (1990), a gradient of glycerol wasalso used, 30% (v/v) in the solution at 5%, and40% (v/v) in the solution at 8%, to improve theseparation of the different MHCs. Migration wascarried out at 50 constant volts for half an hour,then at 110 constant volts for = 18 h at 4 °C. The
temperature was an important factor in the suc-cessful resolution of the different MHCs. After mi-
gration, the gels were stained with Coomassieblue or used for immunoblotting.
ImmunoblottingThe gels for immunoblotting were obtained bythe previously described technique, with a small-er amount of protein (0.4 pg) per well. To deter-mine the nature of the different isoforms of the
myosin heavy chains, the proteins were trans-ferred from the polyacrylamide gel on to a mem-brane of polyvinyl (Millipore, lmmobilon P), in asemi-dry system, at 24 constant volts for 1 h
(Towbin et al, 1979). The blot was treated withone of the anti-myosin antibodies mentionedabove (A, B, C or D), and then by a second anti-body raised against the first, and labelled withalkaline phosphatase (mouse IgG; Jackson lm-munotech) according to the method of Matus etal (1980). In all the buffers used in this proce-dure, bovine serum albumin was replaced by3% (w/v) skimmed milk.
Image-analysisAn image-analysis system was used to make aquantitative analysis of the muscle slices. After


digitizing the images of the muscle cut sectionsunder the microscope, we carried out a thresh-old to isolate the cells that emitted fluorescence,and measured their total surface (Si). With an-other threshold, we measured the total surfaceof all the muscle fibers visible in the field of the
microscope (ST). This surface corresponds tothe total surface visible in the field minus the in-terfascicular tissue. For each group of cells thatreacted with an antibody, we calculated the pro-portion of surface they occupied: 100* Si/ST.This measurement gives only an indicative val-ue, since it was made in just 1 animal per stageof development, and has no decisive incidenceon the conclusions of this study, which are
drawn from the presence or absence of expres-sion of various types of myosins in the musclecells.
RESULTS
Expression of the different typesof myosins in the muscle fibers
Age 39-64 days
The muscles were not yet well organizedat this age, and it was not possible to
clearly distinguish muscle masses in theback or limbs. However, numerous fibersalready reacted with antibodies A, C andD, which shows that muscle differentiationand the synthesis of myosin had actuallybegun.
Age 84 days
The muscle masses were now better orga-nized and most fibers had a myotube-likeappearance (fig 2). All the fibers reacted
uniformly with the 3 antibodies A, C and Das before, but also with antibody B. Thepopulation of myotubes therefore seemedperfectly homogeneous in terms of myosinsynthesis.
Age 122 days (fig 3)
Two muscle fiber populations were ob-served at this age. The first, which repre-sented = one-third of the total population,reacted strongly with antibody A and weak-ly with antibody B. These slow fibers, re-
sembling large myotubes, corresponded tothe primary generation. The second popu-lation of fibers that appeared at this agewere smaller, and all reacted very weaklywith antibody A, but strongly with antibod-ies C and D. Some of them also reactedwith antibody B.
Age 143 days (fig 3)
The large primary fibers reacted with anti-body A but not at all with B. Their propor-tion was now close to 10% only, probablybecause of the increase in the number of
secondary myotubes. These smaller sec-ondary fibers (30% of the total population)did not react with antibody A. Their differ-entiation program is therefore unlike that of
the primary generation, and they constitutea real second population and not merely acontinuation of the proliferation of the pri-mary one. All the secondary fibers reactedwith antibodies C and D, but only some ofthem reacted with antibody B. Thus, at thisage we distinguished 3 fiber populations,which differed in size and in the type ofmyosin they synthesized at this time, andwhich, to make designation simpler, canbe called primary-large-slow (10%), secon-dary-small-fast (30%) and secondary-small-embryonic (60%).
Age 180 days (fig 4)
The group of large primary fibers that re-acted very strongly with antibody A, no
longer reacted with antibody C or antibodyD. In this respect, these fibers were exactlythe same as type I fibers in the adult. The


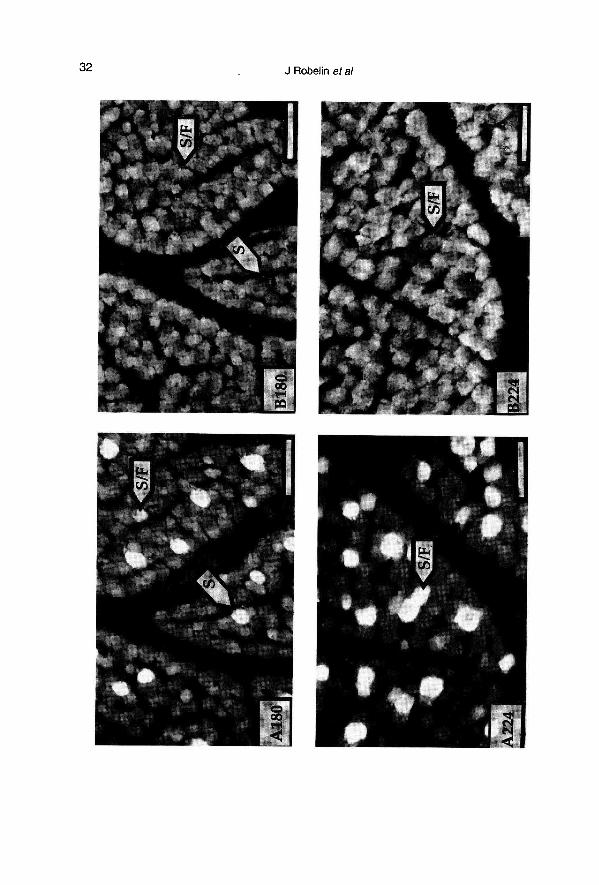

population of smaller fibers reacted uniform-ly with antibody B, while a small proportion(5% of the total) also began to react with an-tibody A. The small fibers that reacted withantibodies A and B also reacted with C andD. The fibers of the second generationtherefore seemed to begin differentiation byexpressing only myosins that react with Cand D, and then to continue their develop-ment by expressing either fast myosins or,simultaneously, fast and slow myosins.
Age 224 days (fig 4)
The proportion of type I fibers, which ex-press only slow myosins, remained practi-cally stable at around 10%. The group of fi-bers that had previously expressed slowand fast myosins simultaneously was nowclearly distinctive, as the reaction with anti-body B was stronger. They also represent-ed about 10% of the total but were largerand close in size to those of the type I fi-bers previously described. These fibers,which expressed simultaneously fast andslow myosins, correspond to type Ilc fi-
bers. The third group of fibers, which wasby far the most numerous (about 80% ofthe total population), were fibers which
now expressed very clearly a fast myosinbut no slow myosin. The reaction with anti-bodies C and D became increasingly vari-able and yielded no additional information.
Age 270 days (fig 5)
The main observation was an increase inthe size of the secondary fast fibers, whichwere now as large as types I and lic. The
reaction to antibodies C and D had be-
come very weak.
Age 30 days after birth (fig 5)
The fibers were all larger at this age. A sig-nificant proportion of them were still syn-thetizing simultaneously fast and slow
myosins, but none reacted to antibodies Cand D.
Expression of different typesof myosins
The results of electrophoresis and immu-noblotting shown in figure 6 are presentedin diagrammatic form in figure 7. The mus-



cle tissue expressed a slow myosin fromthe earliest ages onwards (64 d). The sig-nal grew in intensity at about 180 days offetal life and decreased after birth. The fe-tal myosin (recognized by antibody C),which migrated just above the slow myo-sin, was expressed throughout fetal life butwas no longer detectable on the gels afterbirth. Fast myosins (types Ila and lib) rec-ognized by antibody B were only ex-
pressed from 143 days of gestation. Its
signal increased at around 210 days andremained high thereafter.
Electrophoretic analysis showed a pro-tein which migrates at the same molecular
weight as type Ila myosin from 56 d to theend of the period analysed. However, thisprotein did not react with antibody B in im-munoblotting analysis (fig 6).
DISCUSSION
The aim of this work was to analyse thetemporal sequence of myogenesis duringthe development of fetal calves, and in par-ticular to determine the various myosinheavy chain isoforms expressed in the dif-ferent populations of muscle fibers, no
published information being available in

this species. The main points of discussionare: the number and nature of the myosinisoforms detected, the characterization ofthe muscle fiber populations observed, thechronology of their development during fe-tal growth, and the specificity of myogene-sis in cattle in comparison with that of oth-er species.
Expression of the different typesof myosin
Our study shows the presence of 4 typesof MHC during myogenesis identified byelectrophoresis and immunoblotting, in thefollowing increasing order of migrationrate: 2 fast types which react with antibodyB, a fetal type recognized by antibody Cand a slow type recognized by antibody A.We also evidenced, by immunocytochemi-cal analysis alone, a myosin that reactswith antibody D. It is likely that the myosincharacterized by this antibody is an em-
bryonic isoform, since it is only expressedin cells as they begin their differentiation.This embryonic myosin may share a com-mon epitope with alpha cardiac myosin, asevidenced by the reaction in immunocyto-chemistry. This protein did not react in im-munoblotting, possibly because of its lowabundance. It was therefore not possibleto situate it on the electrophoretic gel.
Electrophoretic analysis also showedanother type of myosin that is expressed atthe beginning of fetal life, and which mi-grates with the same molecular weight astype Ila myosin but without reacting withantibody B or any other antibodies. Thisfast-like myosin may also be an embryonicisoform of the fast myosin.
Adult slow myosin is expressed from thefirst stages of muscle development in cat-tle, as in the human fetus (Draeger et al,1987), in cats (Hoh et al, 1988), in rats (La-Framboise et al, 1991 in mice (Lyons et
al, 1990) and in chickens (Stockdale andMiller, 1987). All the above studies con-firmed that this expression is not necessar-ily dependent on innervation, which is notfully developed at this stage of growth.However, it is now widely recognized thatthe last stage of myogenesis is dependenton environmental influence (see reviews ofStockdale, 1990; Hughes and Blau, 1992).
Adult fast myosin is expressed at about180 d of gestation in cattle, thus, some-what later than slow myosin. A similar se-quence was observed in the human fetus
by Pons et al (1986) and Draeger et al(1987), who reported that the fast myosinwas not expressed until the appearance ofthe secondary generation, at = 100 d of
gestation. In rats, the adult fast myosin isonly expressed after birth (LaFramboise etal, 1991 at an even later stage than in hu-mans.
Characterization of the fiber
generations in developing muscle
Two cell populations, with very differentcell sizes, called primary and secondary fi-bers, were first observed in mammal fetus-es, in particular in the fetus of sheep, al-most 20 years ago (Ashmore et al, 1972).In a previous study on cattle (Robelin et al,1991 ), we obtained results with histochem-istry that showed the differentiation of 2
populations of fibers = 140 d of fetal life. Inthe present study, by characterizing thetypes of myosin synthetized by the cells,we have added to these results and shownthat these fibers expressed different kindsof myosin during their differentiation pro-grams.
During its development, the primarygeneration began by expressing slow myo-sins, then 2 fetal or embryonic myosins,and afterwards another embryonic fast-likemyosin. This finding closely agrees withthe results of Vivarelli et al (1988) in mice

and those of Draeger et al (1987) in hu-
mans. In cattle, as in humans (Draeger etal, 1987), this primary population evolvesduring growth and expresses only a slowmyosin at maturity.
The second generation, which ap-peared around the age of 120 d, differsfrom the primary in initially expressing onlyfetal and embryonic myosins. Later, it ex-
presses a fast myosin and in some cells afast and a slow myosin simultaneously.This second population is similar to thatfound in the 100-d-old human fetus (Ponset al, 1986; Draeger et al, 1987) or to thatobserved in 15-17 d-old rat fetuses (Ru-binstein and Kelly, 1981 ). Its change intoslow or fast types is probably dependenton innervation, as in other species.
Chronology of myogenesis in cattle
The synthesis of myosin in the bovine fetusbegins very soon, before 39 d of age. Up to122 d of gestation, we observed only 1 pop-ulation of large myotubes, which succes-sively expressed several types of myosins.These myotubes later transformed into typeI fibers. This is practically the same devel-opment as that observed in humans (Ponset al, 1986; Draeger et al, 1987).A secondary generation of smaller cells
appeared at 122 d of gestation. Its patternof expression of myosins was differentfrom that of the primary population and it
was made up of smaller cells, whose pro-portion increased rapidly until the end of
fetal life. It is likely therefore that this sec-ondary population is more directly involvedin the increase in muscle fiber number ob-served during this period in cattle (Robelinet al, 1991 At a later stage of develop-ment, this second generation gave rise to2 types of fibers, one of which expressedfast myosin and the other fast and slowmyosins simultaneously.
At birth, we found the 4 muscle cell
types commonly observed in other spe-
cies: type I fibers (slow), types Ila and lib(fast), and type llc fibers (fast/slow). Thedistribution of the fiber types observed in
this study was closely related to the kind ofmuscle we chose, semitendinosus, whichis made up of = 10, 30 and 60% type I,type Ila and type Ilb fibers respectively inadult cattle (Holmes and Ashmore, 1972;Johnston et al, 1975; Robelin, unpublisheddata).
Comparison of the temporal sequenceof myogenesis in different speciesFor the sake of simplicity, we comparedonly 4 animal species: chickens, rats, cat-tle and humans. There are considerabledifferences between these species in theiraduld size (a ratio of 1:1 000), and thelength of their gestation period (from = 20-280 d), and also variations in the extent towhich weight at birth compares with that ofthe mature animal (< 1% in rats; = 2% in
chicks; and between 4 and 6% in cattle
and humans).We used the results of various studies
to compare myogenesis in these species:those of Crow and Stockdale (1984) onchickens, of Rubinstein and Kelly (1981), ),Lyons et al (1983) and LaFramboise et al(1991) on rats, and the findings of Pons etal (1986) and Draeger et al (1987) in hu-
mans. While the chronological periods var-ied greatly in length, there was, neverthe-less, an overall similarity in the sequenceof events. The first recognizable stage in
myogenesis is signalled by the presence oftissue containing cells that synthesizemyosin. It occurs at = 4-6 d of gestation inchickens, 10―12 d in rats, and a little be-fore 40 d in cattle. The following stage ismarked in all the species by the appear-ance of a second cell population: at 8-12 d dof gestation in chicks, 17 d in rats, and= 100 d in humans and cattle. The third
stage culminates in the maturation of themuscle fibers, by which time the fetal andembryonic myosins are no longer synthe-

sized or only in a very small number ofcells. This takes place relatively late in
post-natal life in chickens, at = 20 d in rats,and shortly after birth in cattle and hu-
mans. The muscle tissue of the last 2 spe-cies is thus more fully developed at birththan in chickens or rats. There is probablya link between this more advanced stageof development and the fact that the
weight of newborn calves or humans is
comparatively high in relation to that of theadult.
CONCLUSION
These results show that during myogene-sis in cattle, and in mammals in general, 2successive cell generations develop, whichhave different patterns of expression of
myosin types. The first generation givesrise to type I fibers, while the second gen-eration transforms into type II fibers (Ila, libor lic) in rapid muscles. Adult slow myosinis expressed early on in fetal life, whereasfast type is synthesized at a later time. Theperiod of muscle differentiation in both cat-tle and humans is characterized by the ex-pression of specific myosins, which are
classicaly designated as embryonic or fe-tal. There are many similarities in myogen-esis between cattle and humans, both in
the sequence of events, which is practical-ly the same, as in the number and charac-teristics of the cell populations involved,classically termed primary and secondarymuscle fibers.
These findings should now be comple-mented by a study in vitro of the differenti-ation of myoblasts sampled at different
stages in fetal life. A more detailed studyshould also be made of the precise role ofthese 2 generations in muscles with ex-
treme characteristics, such as the masset-er or cutaneus trunci, which in ruminantsare composed of a single type of fiber,
slow in the former and fast in the latter. Fi-
nally, cattle represent an interesting modelfor the study of the early stages of myo-genesis because of the similarities in the
temporal sequence with the fetal develop-ment of humans.
ACKNOWLEDGMENTS
The authors thank R Jailler and G Cuylle for ani-mal management and slaughtering, and R Jaill-er for assistance in the dissection of muscles.
REFERENCES
Ashmore CR, Robinson DW, Rattray P, Doerr L(1972) Biphasic development of muscle fi-bers in the fetal lamb. Exp Neurol 37, 241-255
Bandman E, Matsuda R, Strohman RC (1982)Developmental appearance of myosin heavyand light chain isoforms in vivo and in vitro inchicken skeletal muscle. Dev Biol93, 508-518 8
Brooke MH, Kaiser KK (1970) Muscle fiber
types: how many and what kind? Arch Neurol23,369-379
Butler J, Cosmos E, Brierley J (1982) Differenti-ation of muscle fiber types in aneurogenicbrachial muscles of chick embryo. J Exp Zool224, 65-80
Bar A, Pette D (1988) Three fast myosin heavychains in adult rat skeletal muscle. FEBS Lett
235, 153-155Condon K, Silberstein L, Blau HM, Thompson
WJ (1990) Differentiation of fiber types inaneural musculature of prenatal rat hindlimb.Dev Biol 138, 275-295
Crow MT (1987) The determinants of muscle fi-ber type during embryonic development. AmZoo/27,1043-1053
Crow MT, Stockdale FE (1984) Myosin isoformsand the cellular basis of skeletal muscle de-
velopment. Exp Biol Med 9, 165-174Crow MT, Stockdale FE (1986) Myosin expres-
sion and specialization of earliest muscle fi-bers in developing avian limb. Dev Biol 113,238-254

Draeger A, Weeds AG, Fitzsimmons RB (1987)Primary, secondary and tertiary myotubes indeveloping skeletal muscle: a new approachto the analysis of the human myogenesis.J Neurol Sci 81, 19-43
Gauthier GF (1986) Skeletal muscle fiber types.In: Myology (Engel AG, Banker BQ, eds)Mac Graw-Hill, New York, vol I, 255-283
Gauthier SF, Lowey S, Hobbs AW (1978) Fastand slow myosin in developing muscle fiber.Nature 274, 25-29
Harris AJ, Fitzsimons RB, McEvan JC (1989)Neural control of the sequence of expresionof myosin heavy chain isoforms in foetalmammalian muscles. Development 107,751-769
Hoh JFY, Hughes S (1989) Immunocytochemi-cal analysis of the perinatal developmentof cat masseter muscle using anti-myosinantibodies. J Muscle Res Cell Motil 10, 312-325
Hoh JFY, Hughes S, Hale PT, Ftizsimons RB(1988) lmmunocytochemical and electropho-retic analyses of changes in myosin gene ex-pression in cat limb fast and slow musclesduring postnatal development. J Muscle ResCell Motif 9, 30-47
Holmes DM, Ashmore CR (1972) A histochemi-cal study of development of muscle fiber
type and size in normal and &dquo;double mus-cled&dquo; cattle. Growth 36, 351-372
Hughes SM, Blau HM (1992) Muscle fiber pat-tem is independent of cell lineage in postna-tal rodent development. Cell 68, 659-671
Johnston DM, Stewart DF, Moody WG, BolingJ, Kemp J (1975) Effect of breed and time onfeed on the size and distribution of beef mus-cle fiber types. J Anim Sci 40, 613-620
Laemmli UK (1970) Cleavage of structural pro-teins during the assembly of the head of bac-teriophage T4. Nature 227, 680-685
LaFramboise WA, Daood MJ, Guthrie RD, Schi-affino S, Moretti P, Brozanski B, Ontel MP,Buttler-Browne GS, Whalen RG, Ontell M
(1991) Emergence of the mature myosinephenotype in the rat diaphragm muscle. DevBiol 114,1-15 5
Lowey S (1986) The structure of vertebratemuscle myosin. In: Myology (Engel AG,Banker BQ, eds) Mac Graw-Hill, New York,vol I, 563-587
Lyons GE, Haselgrove J, Kelly AM, RubinsteinNA (1983) Myosin transitions in developingfast and slow muscles of the rat hindlimb. Differentiation 25, 168-175
Lyons GE, Ontell M, Cox R, Sasoon D, Bucking-ham M (1990) The expression of myosingenes in developing skeletal muscle in themouse embryo. J Cell Biol 111, 1465-1476
Marini JF, Pons F, Anoal F, Fardeau M, Leger J(1990) Immunocytochemical study of myosinchanges of muscle fibers of DMD boys andsymptomatic carriers of DMD identified byantidistrophic MAbs. Abstracts of the VII In-temational Congress on neuromuscular dis-ease, Munich. J Neurol Sci (suppl) 98
Matus A, Pehling G, Ackermann M, Maeder J(1980) Brain postsynaptic densities: their re-lationship to glial and neuronal filaments.J Cell Biol 87, 346-359
Miller JB (1990) Myogenic programs of mousecell lines: expression of myosin heavy chainisoforms, MyoD1 and myogenin. J Cell Biol111,1149-1159
Miller JB, Stockdale FE (1986) Developmentalregulation of the multiple cell lineages of theavian embryo. J Cell Biol 103, 2197-2208
Motus A, Pehling G, Ackermann M, Maeder J(1980) Brain postsynaptic densities: their re-lationship to glial and neuronal filaments.
J Cell Biol 87, 346-359
Muntz L (1990) Cellular and biochemical as-
pects of muscle differentiation. Comp Bio-chem Physiol 978, 215-225
Pette D, Vrbova G (1985) Neural control of phe-notypic expression in mammalian muscle fi-bers. Muscle Nerve 8, 676-689
Pons F, L6ger JOC, Chevallay M, Tome FMS,Fardeau M, Leger JJ (1986) Immunocyto-chemical analysis of myosin heavy chains inhuman fetal skeletal muscles. J Neurol Sci76, 151-163
Robelin J, Lacourt A, Béchet D, Ferrara M, Bri-and Y, Geay Y (1991) Muscle differentiationin the bovine fetus: a histological and histo-chemical approach. Growth Dev Aging 55,151-160
Rubinstein NA, Kelly AM (1981) Development ofmuscle fiber specialization in the rat hindlimb.J Cell Biol 90, 128-144
Stockdale FE (1990) The myogenic lineage: evi-dence for multiple cellular precursors during

avian limb development. Proc Soc Exp BiolMed 194, 71-75
Stockdale FE, Miller JB (1987) Review the cellu-lar basis of myosin heavy chain isoform ex-pression during development of avian skele-tal musculature. Dev Biol 123, 1-9
Sugiura T, Murakami N (1990) Separation ofmyosin heavy chain isoforms in rat skeletalmuscles by gradient sodium dodecyl sulfatepolyacrylamide gel electrophoresis. BiomedRes 11, 87-91
Towbin H, Stachelin T, Gordon J (1979) Electro-phoretic transfer of proteins from polyacryla-
mide gels to nitrocellulose sheets: procedureand some applications. Proc Natl Acad SciUSA 76, 4350-4354
Vivarelli E, Brown WE, Whalen RG, Cossu G(1988) The expression of slow myosinduring mammalian somatogenesis and limbbud differentiation. J Cell Biol 107, 2191-2197
Whalen RG, Sell SM, Butler-Browne GS,Schwartz K, Bouveret P, Pinset-Harstrom I(1981) Three myosin heavy chain isozymesappear sequentially in rat muscle develop-ment. Nature 292, 805-809
![Gamme de produits - bonfiglioli.fr · 9 Type Couple [Nm] C 05 C 12 C 22 C 32 C 36 C 41 C 51 C 61 C 70 C 80 C 90 C 100 7200 4000 2300 1600 1000 600 450 300 200 100 45 12000 Lorsque](https://img.pdfslide.fr/doc/110x75/5c644d3b09d3f26b1f8c80a5/gamme-de-produits-9-type-couple-nm-c-05-c-12-c-22-c-32-c-36-c-41-c-51-c.jpg)