Embed Size (px)
Citation preview

LES PROTEINESPrincipaux constituants du
vivant

PLAN
I. Spécificité des protéines liées à leur ���������������� ���������
II. Spécificité de fonction des protéines liées àleur ������� �������������������������
III. Economies de moyen des protéines à��������������������������������������

������ !����Quantitativement • Présentes ���"�����
les organismes (même virus)
• Chez l'Homme # $%&des substances organiques
• Dans une cellule animale = '('('('(& du poids frais
• Dans une bactérie> 2000 protéines
• Omniprésence et Abondance justifient leur nom(προτειον = je prends la première place)

������ !����Qualitativement ) *+�������� du
fonctionnement cellulaire ) ���������,������,=
complexe et définie avec précision
qui assure leur bon fonctionnement
� assemblage des constituants cellulaires
� catalyse des transformations chimiques
� production du mouvement

������ !����Historique • Rôle enzymatique
évident dès 1850• expression génétique
1940• rôle des lipides
membranaires 1960• rôle des polysaccharides
pas toujours connu
� L'hémoglobine prise comme exemple de protéine

-�.��*���������� ���� �������
) /.�#�'%%�0��
• synthétisées à partir de petites molécules PM<1200 Da
• 1 Da = 1/12 masse du C• PM = Masse de la
particule / (1/12) masse de C
• 1 aa = 110 Da• donc 1000 aa = 110 kDa
= ����� ��1��� répétées+ liaisons ��2�������

I. Spécificité des protéines liées àleur ���������������
� ���������
• Composés ������������ !343�3�) .��������2������"���
Ex : décomposition à chaud de l'albumine� libération d'ammoniac NH35 HCl ���� NH4Cl (fumée blanche)
• ���������������������� ����

I. Spécificité de �������������
/�������������
� à la �� �������� en acides aminés � à la ������������������ au �� 1�� et à ����� des acides aminés

I. Spécificité de �������������67�/��������������8 ����� ������������������� ����
20 acides aminés standards= groupe � ��� ��� ����+ ��������1�+������sur le même C
sauf la proline = groupe aminésecondaire = αααα-imino acide.
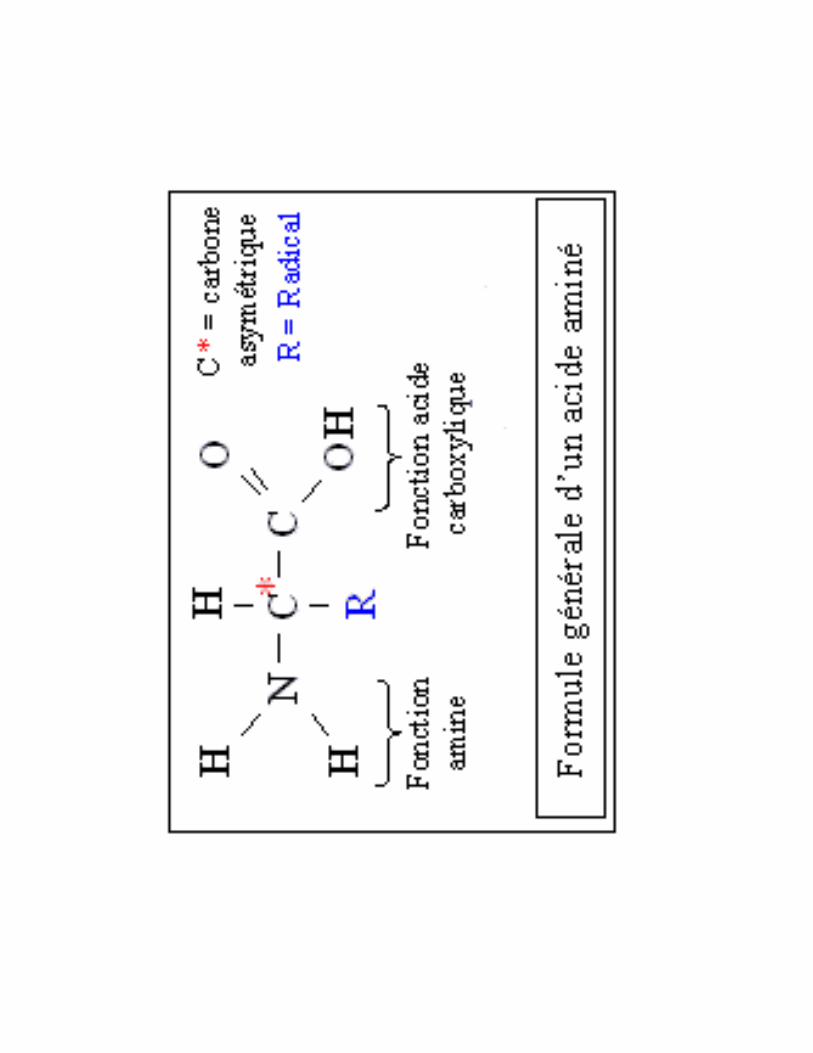

I. Spécificité de �������������67�/��������������8 ����� ������������������� ����
'9 ����1����� ���������� �����:���������������
a) Les groupes acides et amines sont complètement ionisésdans conditions physiologiques à pH=7 : pKCOOH = 2,2 ���� carboxylate COO-pKNH2 = 9,4 ���� ion ammonium NH3+
a.a = ampholytes

'9 ����1����� ���������� �����:���������������
a) Les groupes acides et amines sont complètement ionisésb) Les acides aminés sont des zwitterions
� /�������;����� élevé ~ 300°C au lieu de 100°C
� Très ����1��� dans l'eau,assez solubles dans les solvants polaires maispas du tout dans les solvants organiques apolaires

'9 ����1����� ���������� ����<9 /��������������� ������8 ���������������������
a) Acides aminés à chaînes non polaires
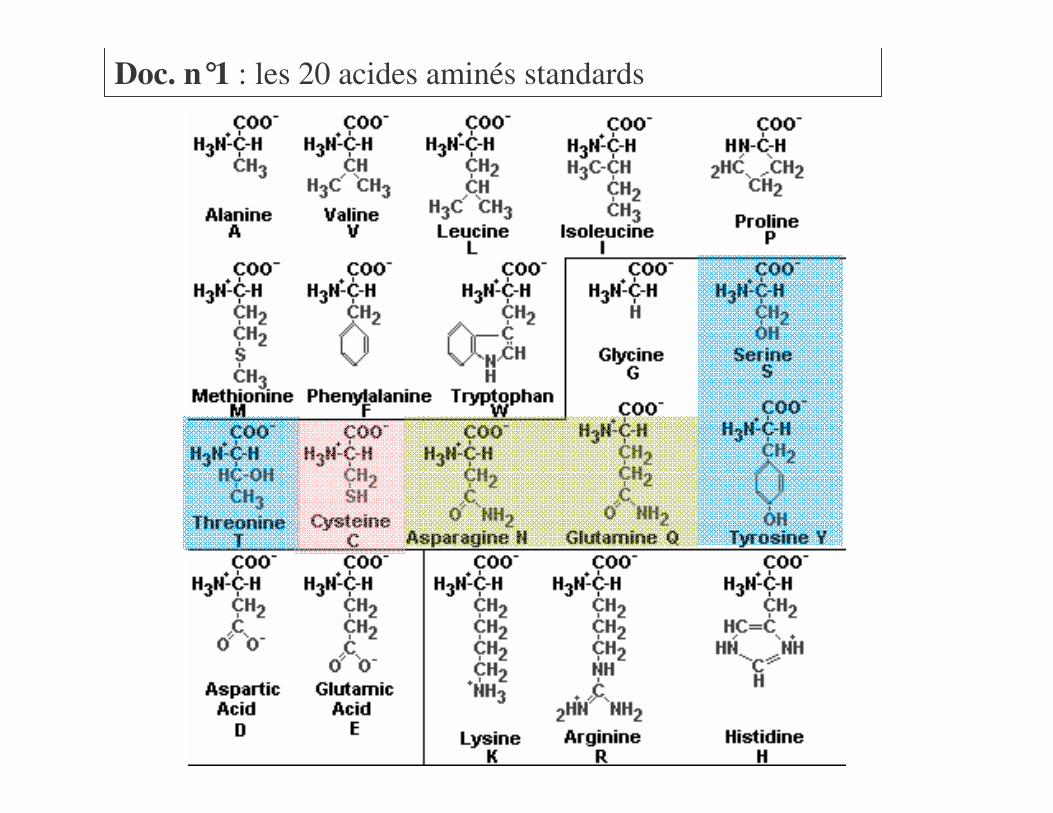
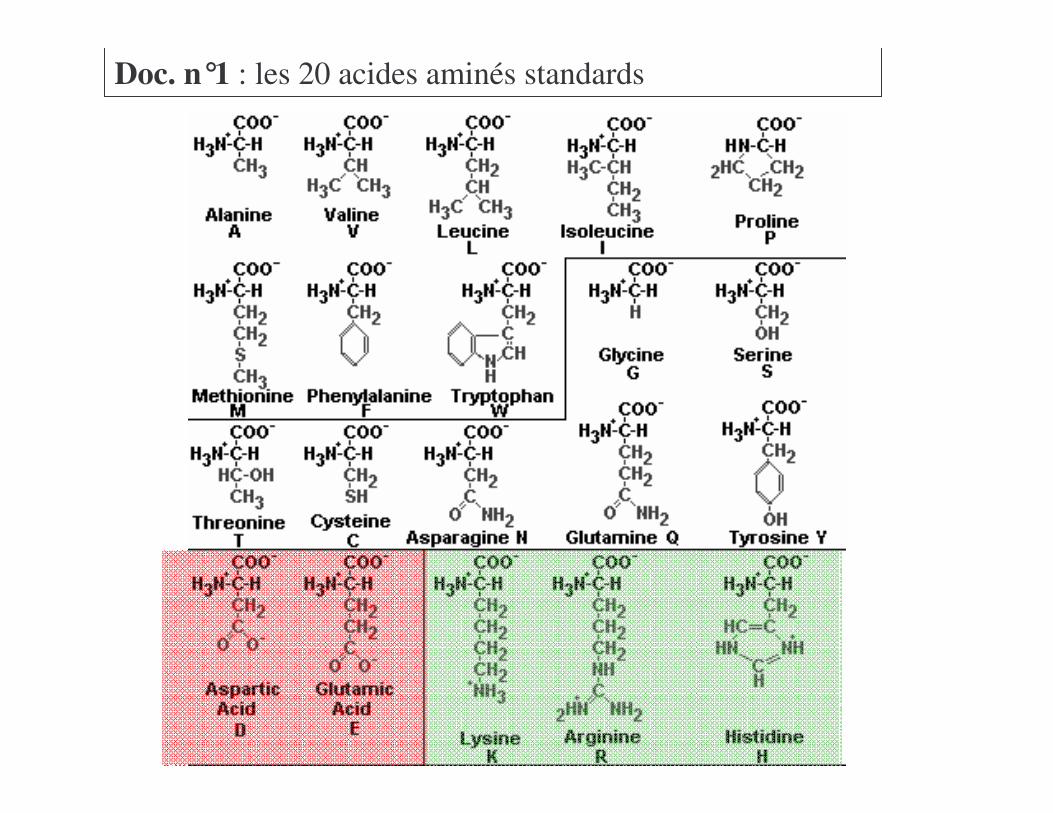
Voir Doc n°1
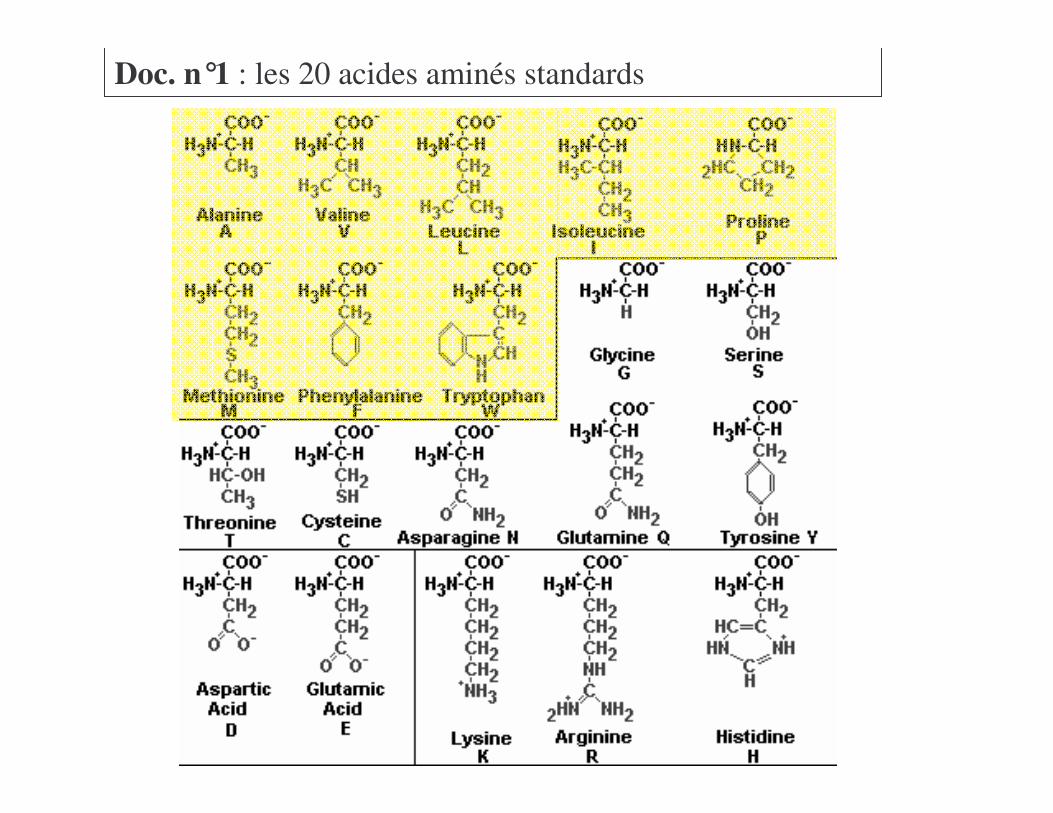
Doc. n°1 : les 20 acides aminés standards

<9 /��������������� ������8 ���������������������
a) Acides aminés à chaînes non polairesb) Acides aminés polaires non chargés
Voir Doc n°1=��!������>� ����+����� � ���� ������
Doc. n°1 : les 20 acides aminés standards

<9 /��������������� ������8 ���������������������
a) Acides aminés à chaînes non polairesb) Acides aminés polaires non chargésc) Acides aminés polaires chargés
Voir Doc n°1 !46�?*��>� ������2��� ��@���2��
Doc. n°1 : les 20 acides aminés standards

<9 /��������������� ������8 ���������������������
a) Acides aminés à chaînes non polairesb) Acides aminés polaires non chargésc) Acides aminés polaires chargésd) Identification des acides aminés
� 61���1��� à <A%�� � �����+���������� absorbent aussi dans les <$%9A%%��
� /��2������������� >��énantiomères) en général ;�� ���----

<9 /��������������� ������8 ���������������������
a) Acides aminés à chaînes non polairesb) Acides aminés polaires non chargésc) Acides aminés polaires chargésd) Identification des acides aminése) Autres acides aminés non standards

� !��=��.6������ (Ex antibiotiques actinomycine D)
� 6767�.���=�*� >
e) Autres acides aminés non standards
� 4���+��������Hydroxyproline (15 à 30%par prolylhydroxylase qui nécessite acideascorbique)Hydroxylysinedu �����@B�� (le stabilisent par liaison H)
� .����������, acétylation ou phosphorylationex. : �������� ou protéinesribosomiales

� �*���*��>
• ���������� �������Ex1. ?6C6 (γ-aminobutyric acid, produit de décarboxylation du glutamate),Ex2. ��� ��� (produit de la tyrosine)
• ��� ���Ex1. �����+��� (produit par tyrosine)médiateurs de réactions allergiquesEx2 4���� ���
)����� �������� ���1������� importants
Ex !���������, Ornithine (de la biosynthèse de l'urée)

!��!- ���� �����������6�>
Propriétés physico-chimiques très variées des 20 acides aminés =� polarité� acidité/basicité� aromaticité� encombrement� flexibilité conformationnelle� capacité à former des liaisons covalentes/hydrogène
Un grand nombre de ces propriétés sont corrélées
Responsables de la grande diversité de propriétés des protéines

I. Spécificité de �������������67�/��������������8 ����� ������������������� ����
C7�/��������������8 ��������������������

C7�/��������������8 ��������������������
'9 !��������������������������������������
• Formée par �����������• Liaison covalente ;����• Caractère à D%&���1��• C-N plus ������ (1,33 Å) que C-Cα (1,46 Å) et C=O plus courte également de 0,02 Å• Conformation ��6�� plus stable (de 8kJ/mol) sauf avec Pro (coude!)
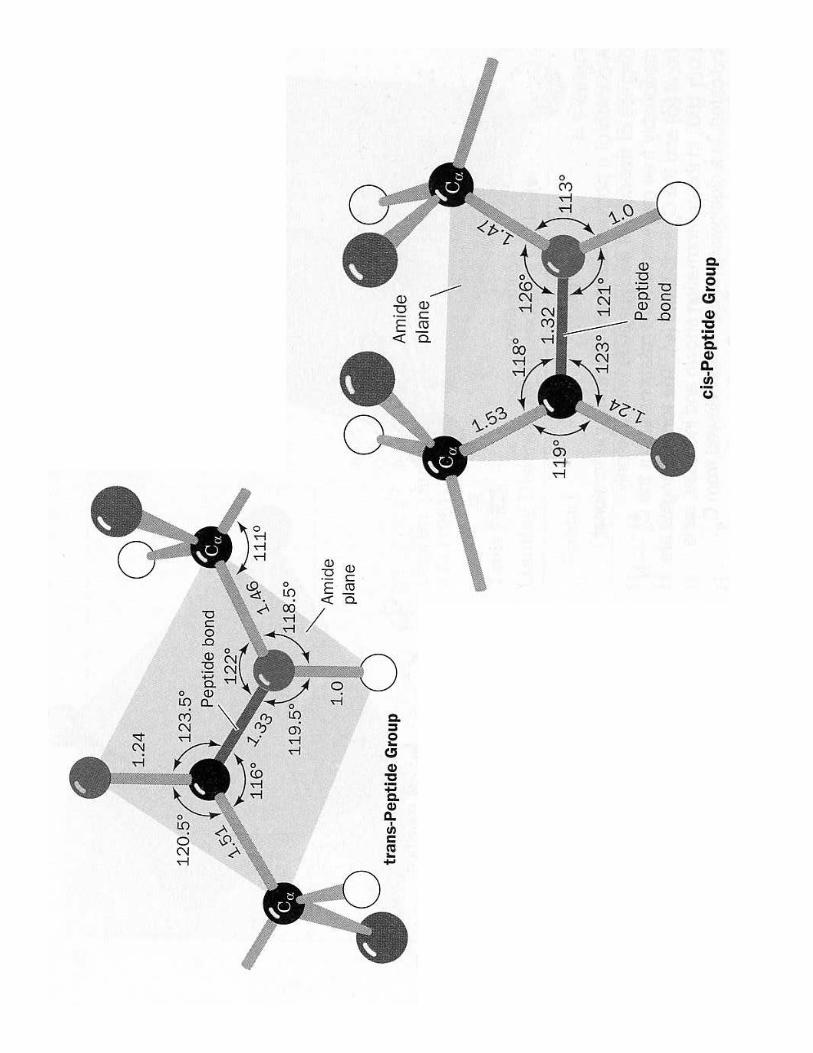

C7�/��������������8 ��������������������'9 !��������������������������������������<9 -�������� ������� B���- ���@�������� < 10 aa- /����� s.s < 50 aa- /���/���/���/������������������� = une ou plusieurs �������peptidiques- Pas d'embranchement (cf code génétique)- Combinaisons pas toutes réalisées
20100=1,27.10130 possibilités(nombre d'atomes dans l'univers ≅ 9.1078 le)
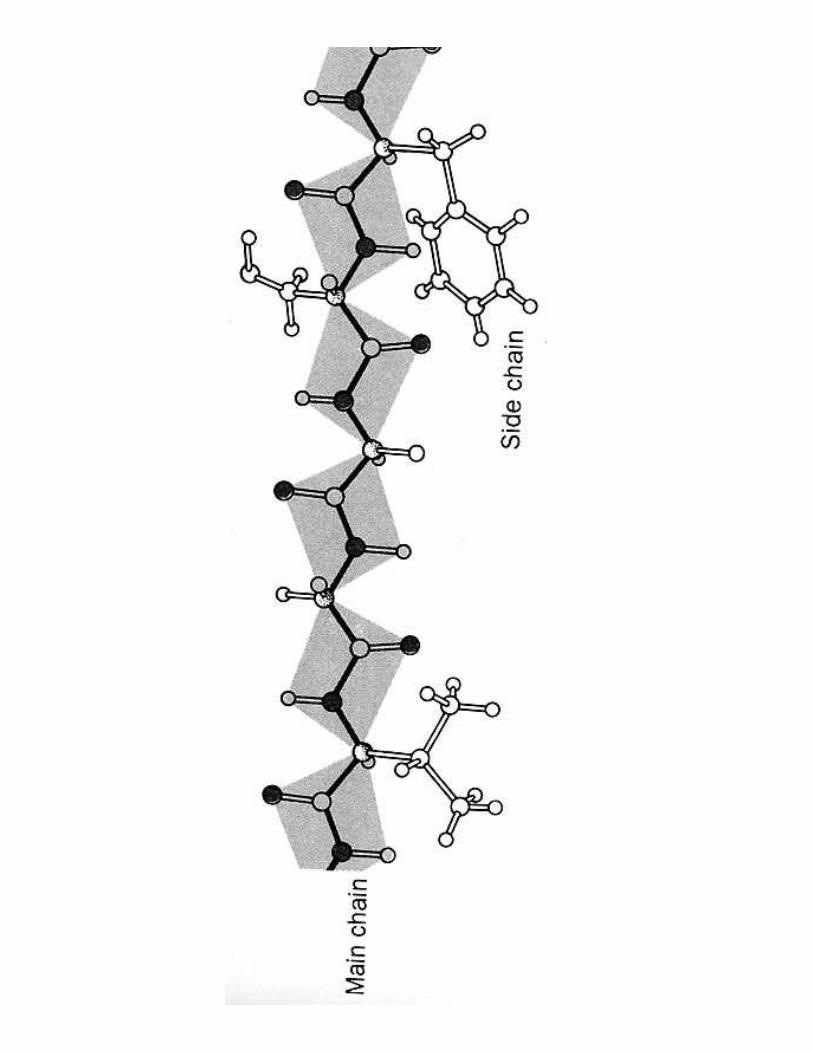

I. Spécificité de �������������67�/��������������8 ����� ������������������� ����
C7�/��������������8 ��������������������
!7�/������������������� 1������8���������������� ����

!7�/������������������� 1������8����������7��7
'9 /�������������� 1�������
- !���1������������� complexe(dépend de la concentration saline du milieu)
- Utilisé pour �����������B��
repérées par la ninhydrine(rouge violacée ou jaune pour proline)

!7�/������������������� 1������8����������7��7
'9 .��;��������������������������� 1�������<9 .��;�����������������������@�������� FONCTION DE :
� �� 1��� ��������� ��������������� ���
Ex. 41� ;����;�� � : 141aa sur chaîne α et 146 sur chaîne βVal 6 de chaine β (hydrophobe) au lieu de Glu (hydrophile)N.B : certaines modifications silencieuses

6�����������>�6�����������>�6�����������>�6�����������>�������� �������������������������� ������� �������������������������� ������� �������������������������� ������� �������������������������� ����
� Préparation = séparer groupements prosthétiques / sous unités / ponts S-S
� Séquençage : fragmentation ( 20 à 60aa) par trypsine puis recoupement des séquences
� Méthode de Sanger automatisée = enlèvement séquentiel des aa terminaux + identifiication avec fluorodinitrobenzènecoloré
� Identification des aa
� absorption des UV� dialyse fonction de PM� point isoélectrique fonction de pKi,)
� Reconnaissance des protéines (par chromatographie 1, 2 ou 3D ou d'affinité, colonne, gel filtration ou ultracentrifugation)

Est-ce que la �2������ ����������������� ����� suffit pour
déterminer la �2������ ���;�������� protéiques
?

I. Spécificité de �������������II. Spécificité de fonction des protéines liées à leur ������� �������������������������

���.�����������
• Spectrométrie (analyse du spectre de dispersion rotatoire)
• Analyse de viscosité• Analyse par diffraction aux rayons X
(l=0,1 à 0,2 nm � diagramme de diffraction) sur cristaux
• RMN jusqu'à 20.000 PM
A. Conformation native des protéinesII. Spécificité de fonction et S3D
'9 .��������2��������������;�� ������������������

a) Méthodes d'étude1�����������
�Conformation = S3D ± compactée stable dans des conditions physiologiques
�Structure native (reployée dans conditions physiologiques) présente caractéristiques uniques.
�Propriétés "moyennes" communes à toutes protéines dénaturées (déroulées)
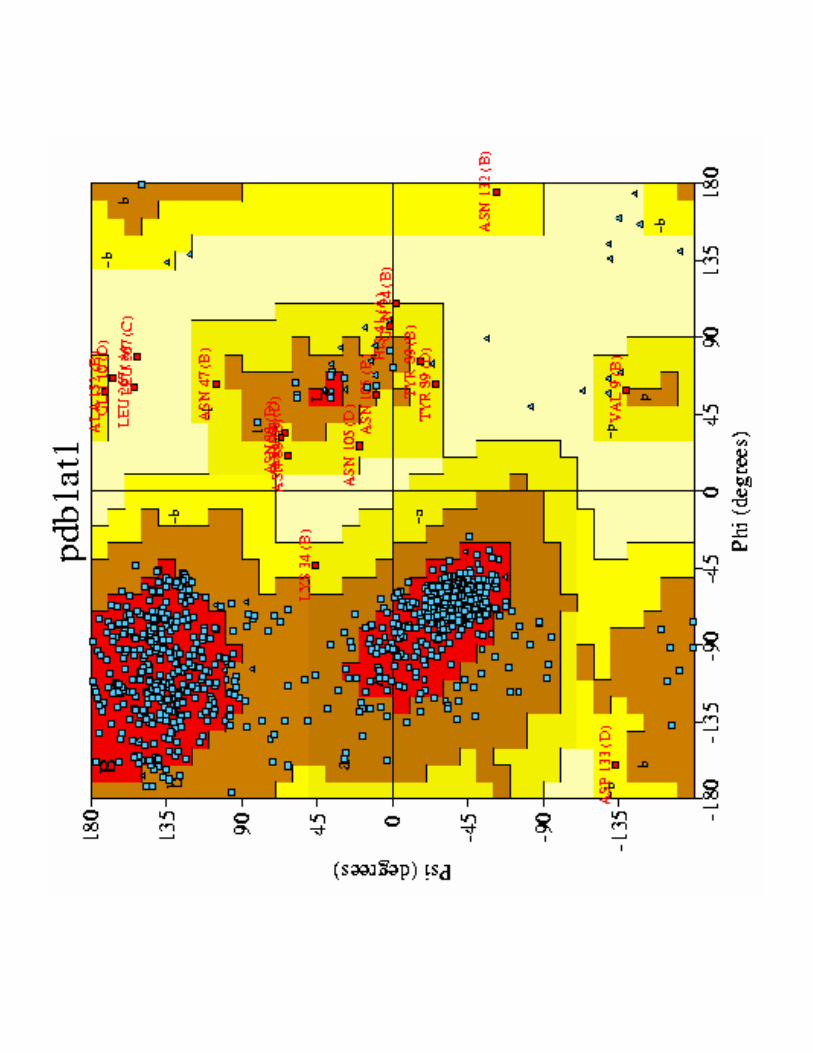
� Diagramme de Ramachandran donne angles stériquement possibles
ΦΦΦΦ= Cαααα - NΨΨΨΨ = Cαααα - C=O
1- Mise en évidence de la conformation
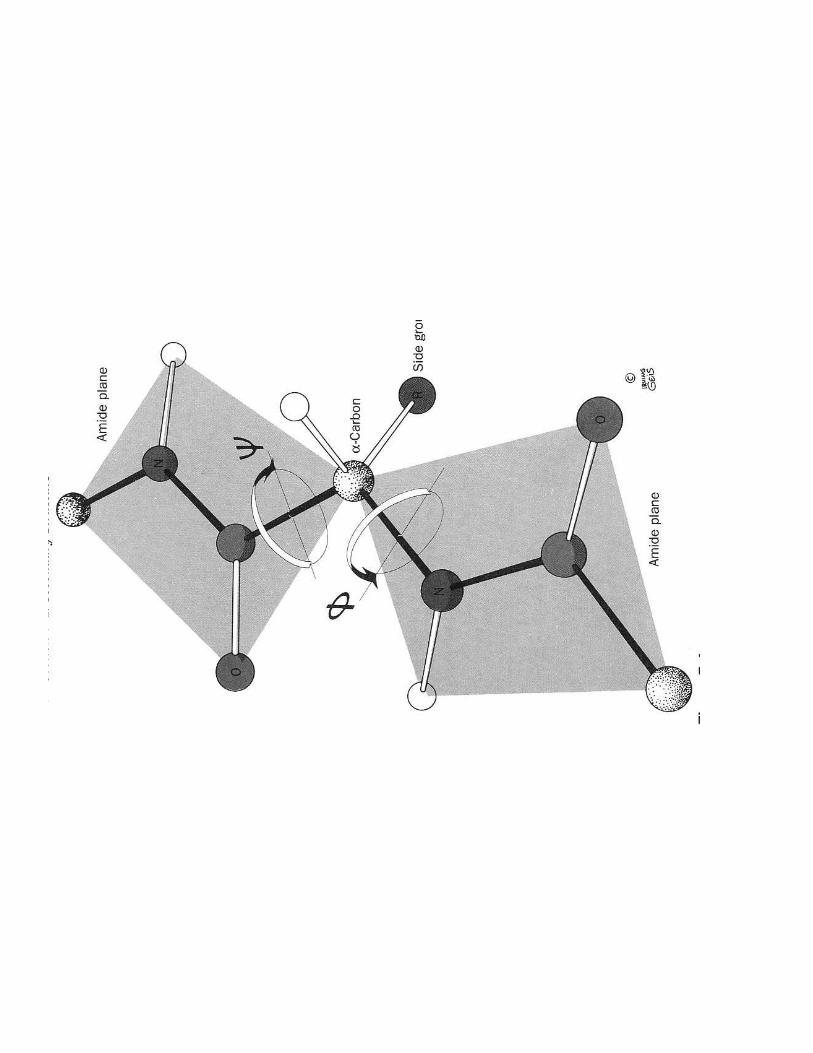
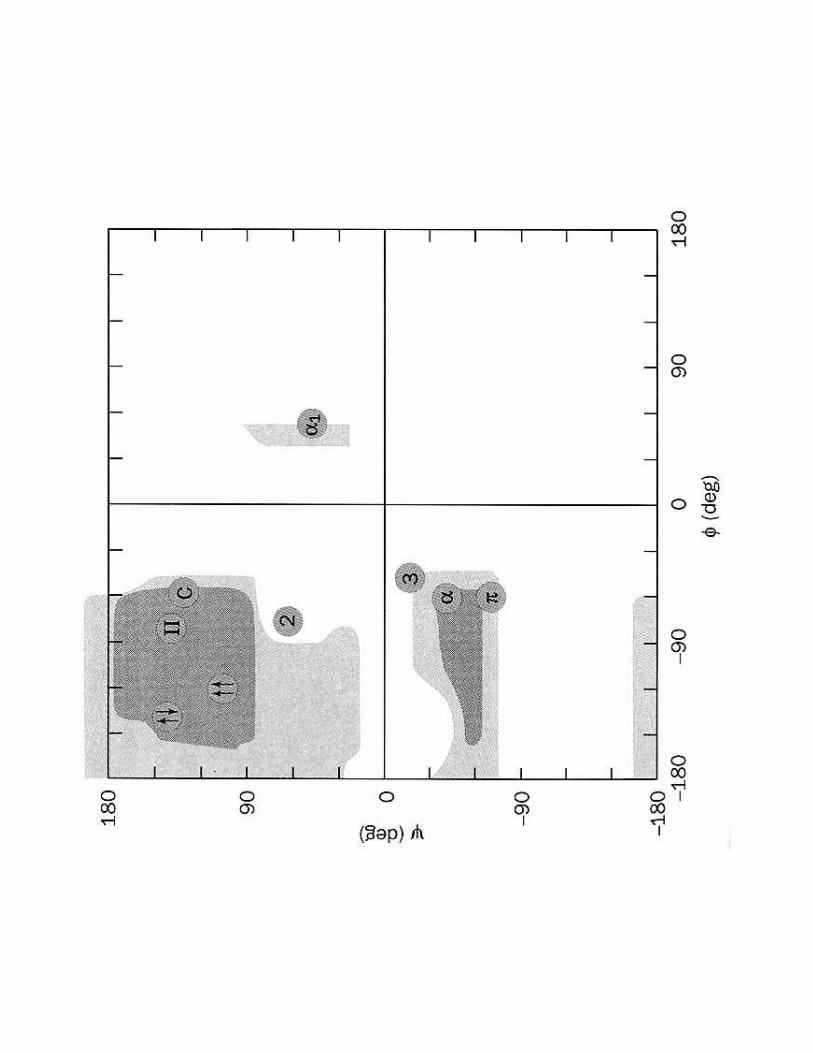

���.��������2����� >������'E$F�6�;�����
1- Mise en évidence de la conformation<9 .������ ������������ ����������������
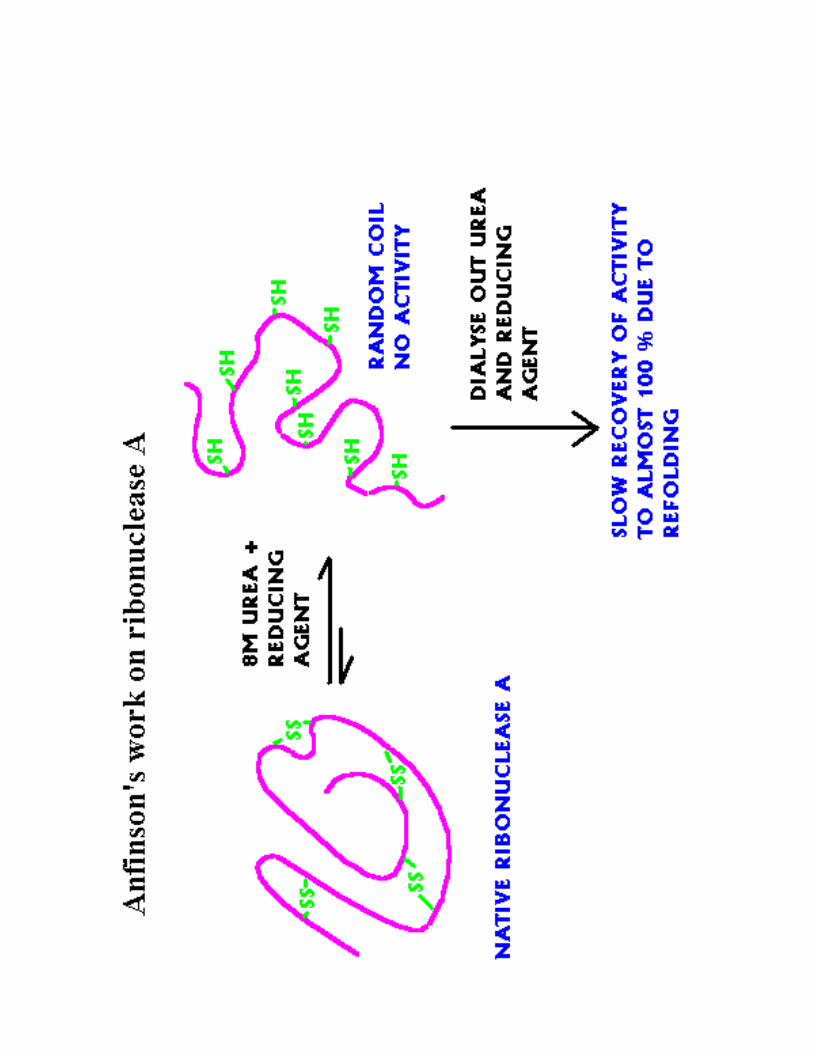

Chances que le reploiement se fasse correctement :
(1/7)*(1/5)*(1/3)*(1/1)='G'%$
Reploiement = 10 h in vitro= 3 minutes in vivo
avec disulfure protéine-disulfide isomérase

a) Mise en évidence : 1957 Anfinsen1��������� ����@��� ����;�������������1��
<9 .������ ������������ ����������������

-������������������H��-������������������H��-������������������H��-������������������H�� (permanents ou induits)-��H����H����H����H�� (forces de Van der Waals) = faibles mais nombreuses �stabilisent fortement les protéines.
� !��������H������� ������- ex : entre COOH de 4,3. 10-30C.m disposés en série séparés de 5 Å dans un milieu D=4 � 9,3 kJ.mol-1
;������:�;��'G�A �
- Rôle d'autant plus important que le cœur de la protéine possède un D faible
- Forces s'additionnent dans structures où les dipôles des groupe amide et carbonyl pointent tous dans la même direction : ex feuillets β et hélices α

� !������������ ������forces résultantes beaucoup plus faibles que dipolespemanents
� !������;��������������������-����- Changement très rapides de répartition des électrons pour des molécules non polaires presque électriquement neutres- De l'ordre de 9%3A�0I7 ��9'(ex. CH4 vaporisé avec 8,2 kJ.mol-1, qui établi des contacts H...H avec 12 "voisins")- Diminue très rapidement avec la distance :�;�'G�J�- Très nombreuses (par reploiement protéique)�
���1�����K
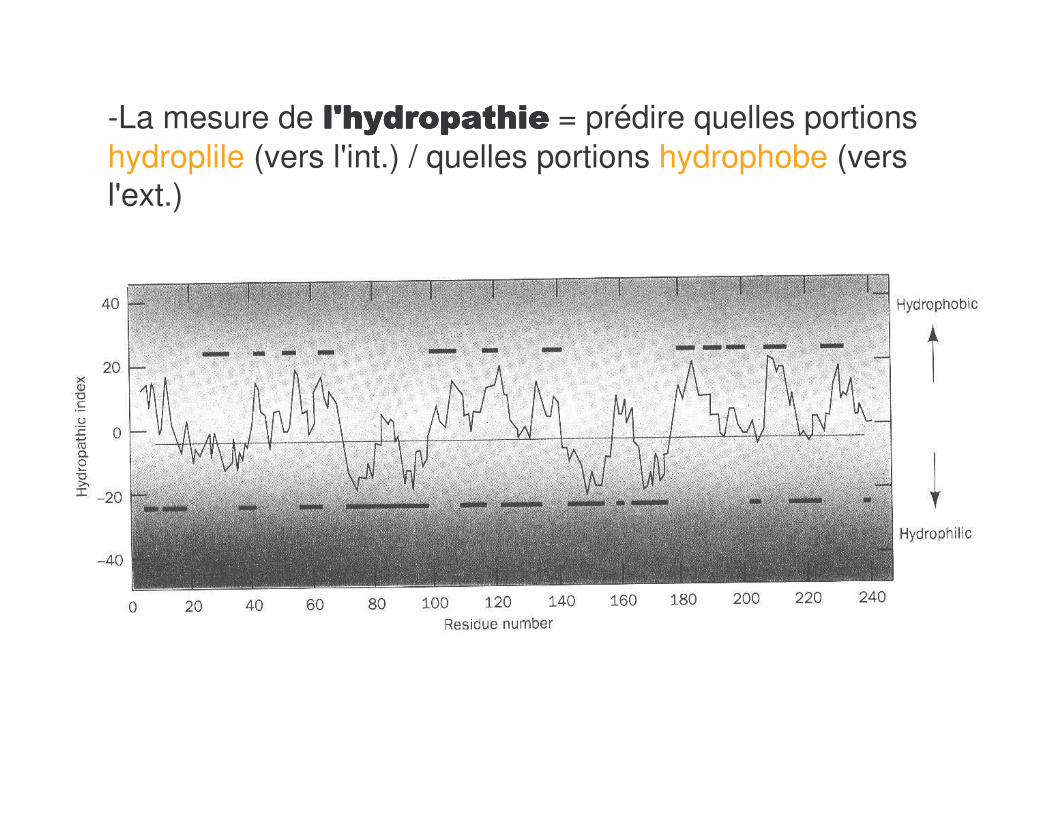
-La mesure de �������������������������������������������� = prédire quelles portions hydroplile (vers l'int.) / quelles portions hydrophobe (vers l'ext.)

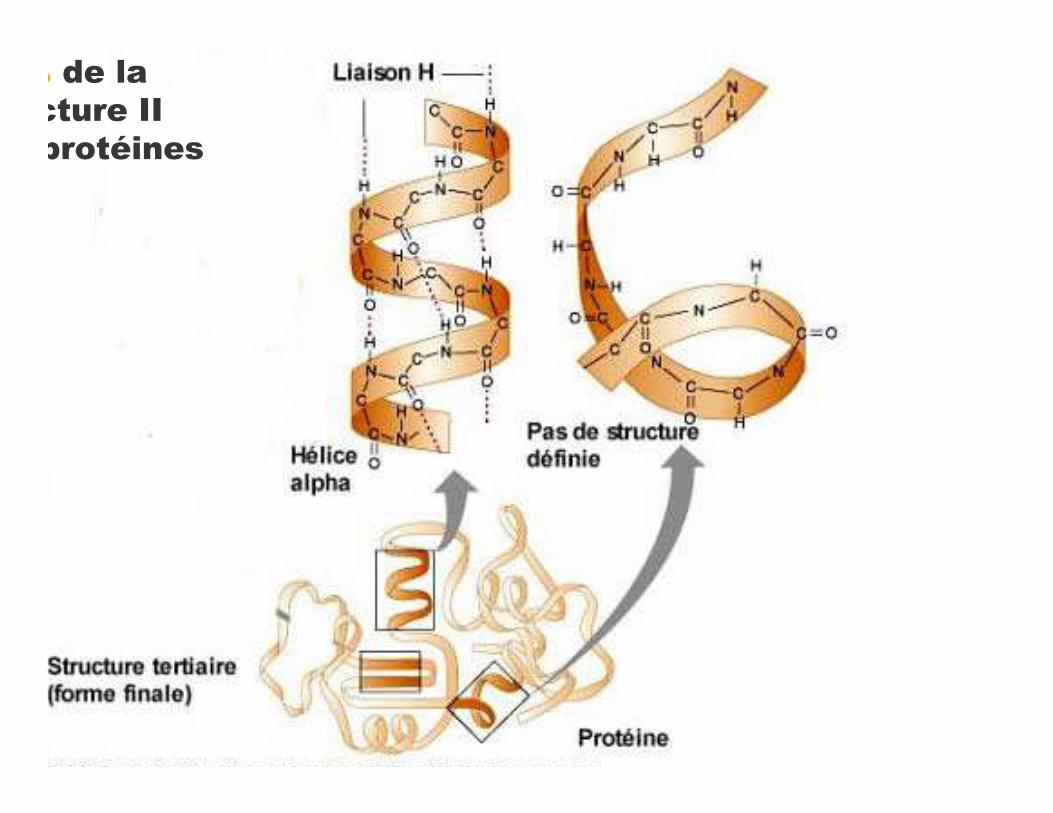
• Pauling et Corey (1951)
A. Conformation native des protéinesB. Feuillets/hélices/Tours = motifs communs
II. Spécificité de fonction et S3D
'9 =���������β
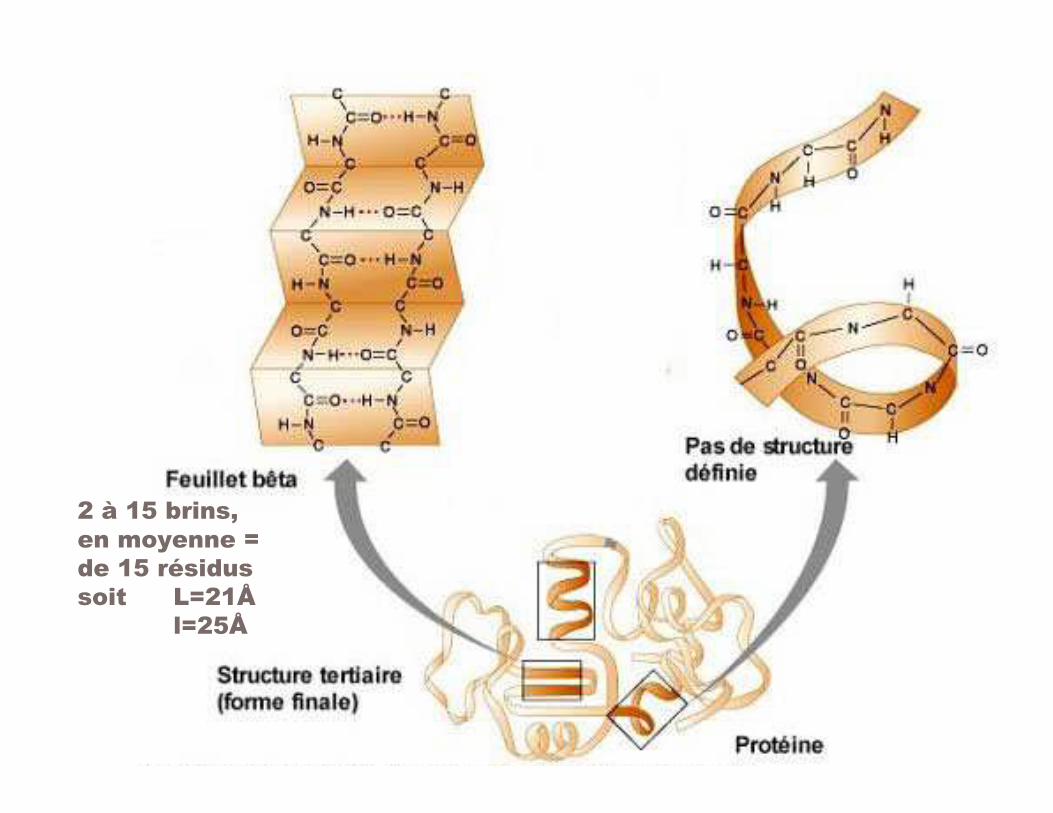
<�8 '$�1����3���� �������:���'$������������ -:<'L
�:<$L
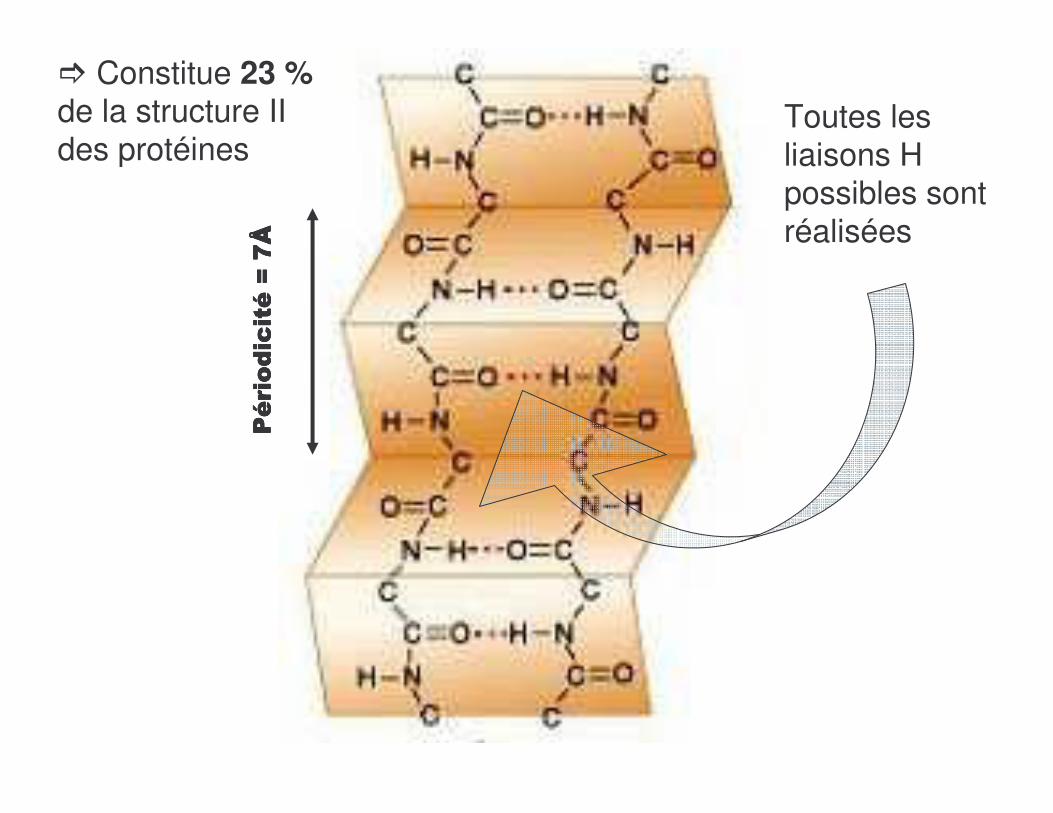
� Constitue 23 %de la structure II des protéines
Toutes les liaisons H possibles sont réalisées
// //�� ���������
�������
�������
��������� ��:�F:�F:�F:�FLL LL
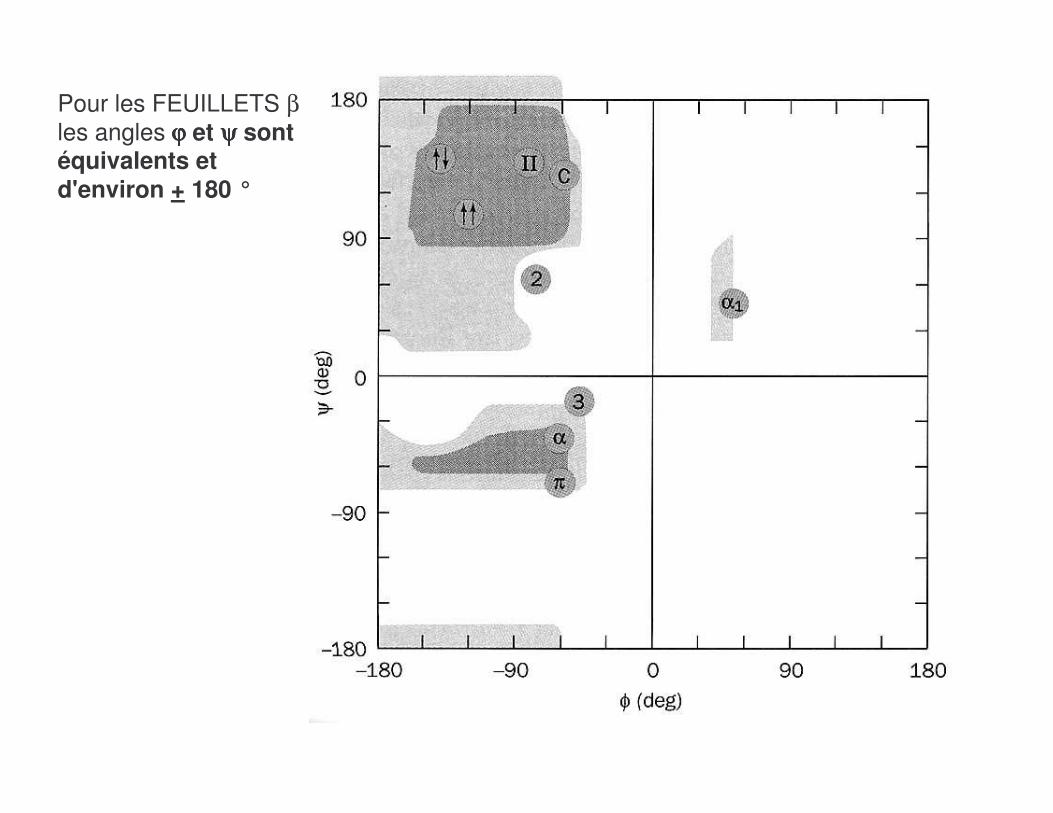
Pour les FEUILLETS βles angles ϕϕϕϕ et ψψψψ sont équivalents et d'environ + 180 °
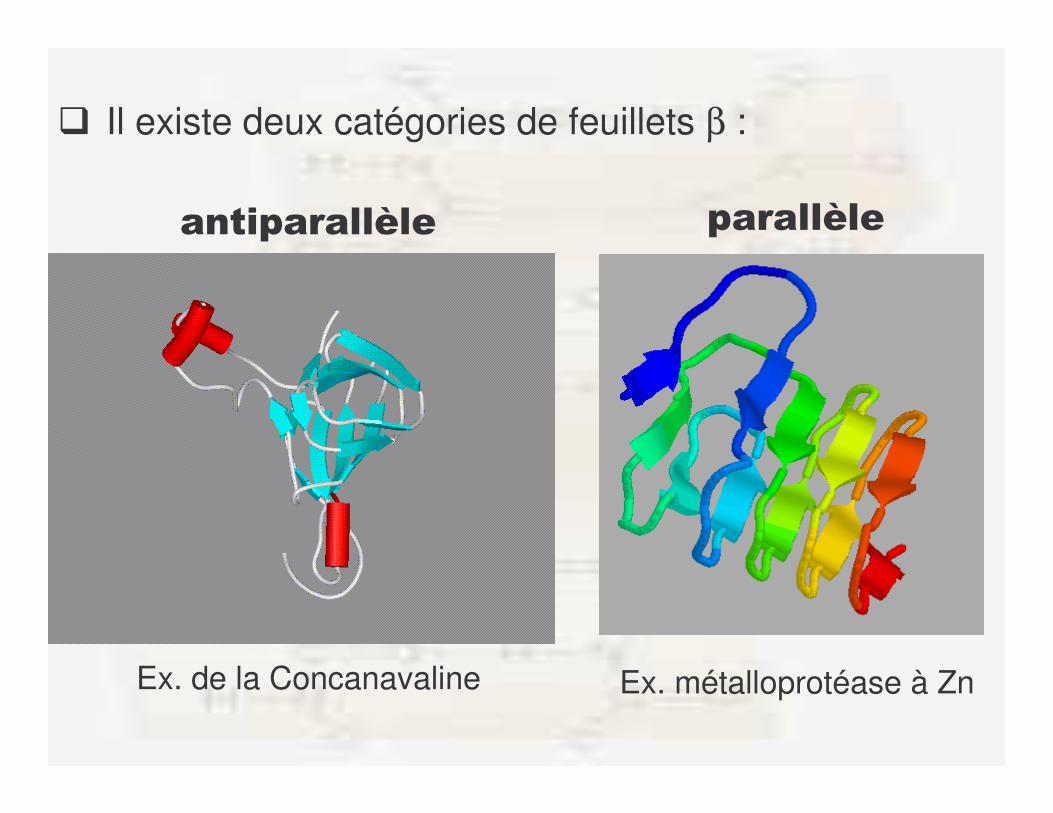
� Il existe deux catégories de feuillets β :
����������B�� ������B��
Ex. de la Concanavaline Ex. métalloprotéase à Zn

B. Feuillets/hélices/Tours = motifs communs
1- Feuillets β<9 4������
!���������������������α : Pauling et Corey (1951)
<A�& ������������������������������
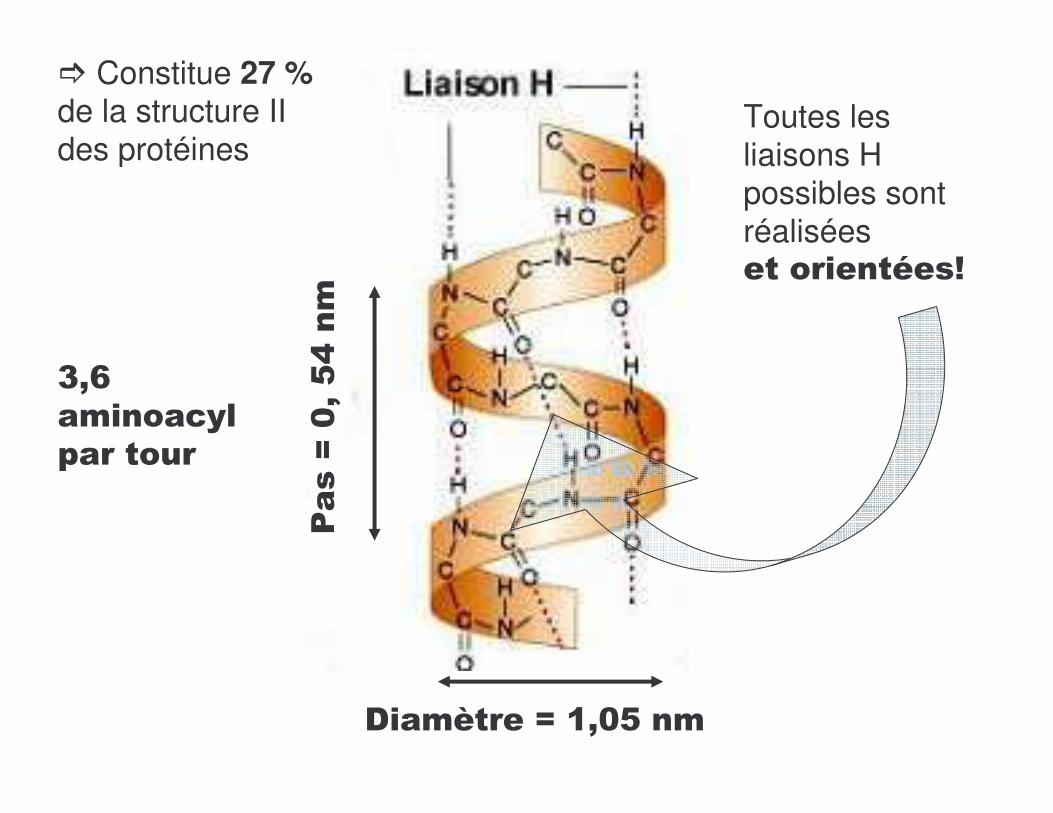
Toutes les liaisons H possibles sont réalisées������������K
/���:�%3�$D��
��� B����:�'3%$��
A3J�� ���������������
� Constitue 27 %de la structure II des protéines
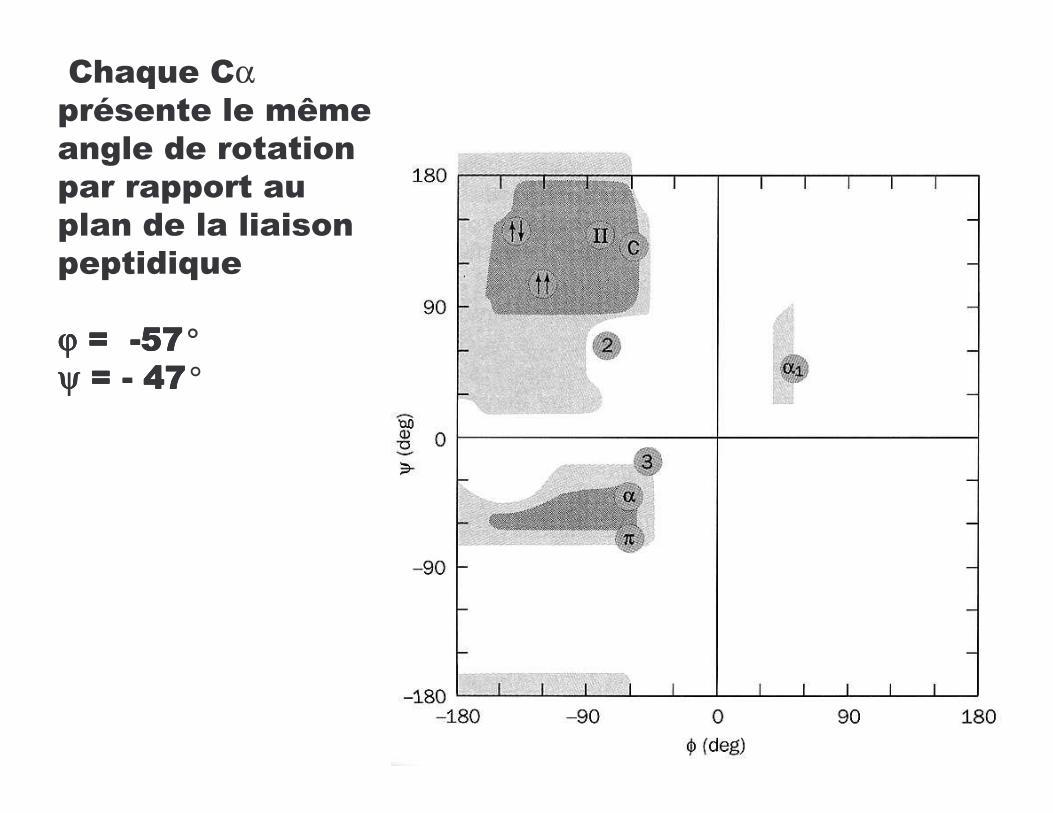
!������!����������� M ����@��������������������������������������������������������
ϕϕϕϕ :��:��:��:��9999$F$F$F$F°ψψψψ :�:�:�:�9999 DFDFDFDF°
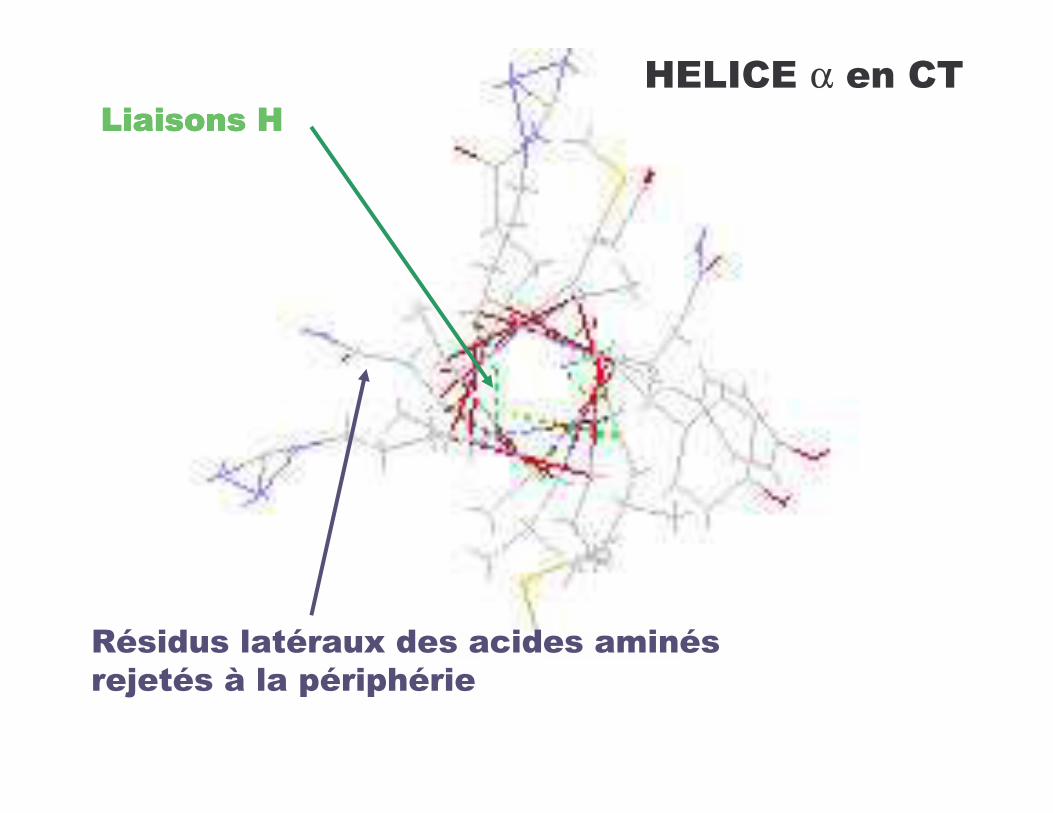
-��������4-��������4-��������4-��������4
��������������+����������� �������N�����8 �������������
4*-�!*�α ���!�

6������������������� : • hélices gauche type collagène• hélice π

B. Feuillets/hélices/Tours = motifs communs
1- Feuillets β2- HélicesA9 6�����������������������������2��
• Environ 50% des protéines
• Dès 60 a.a, ∃ boucle et/ou tour
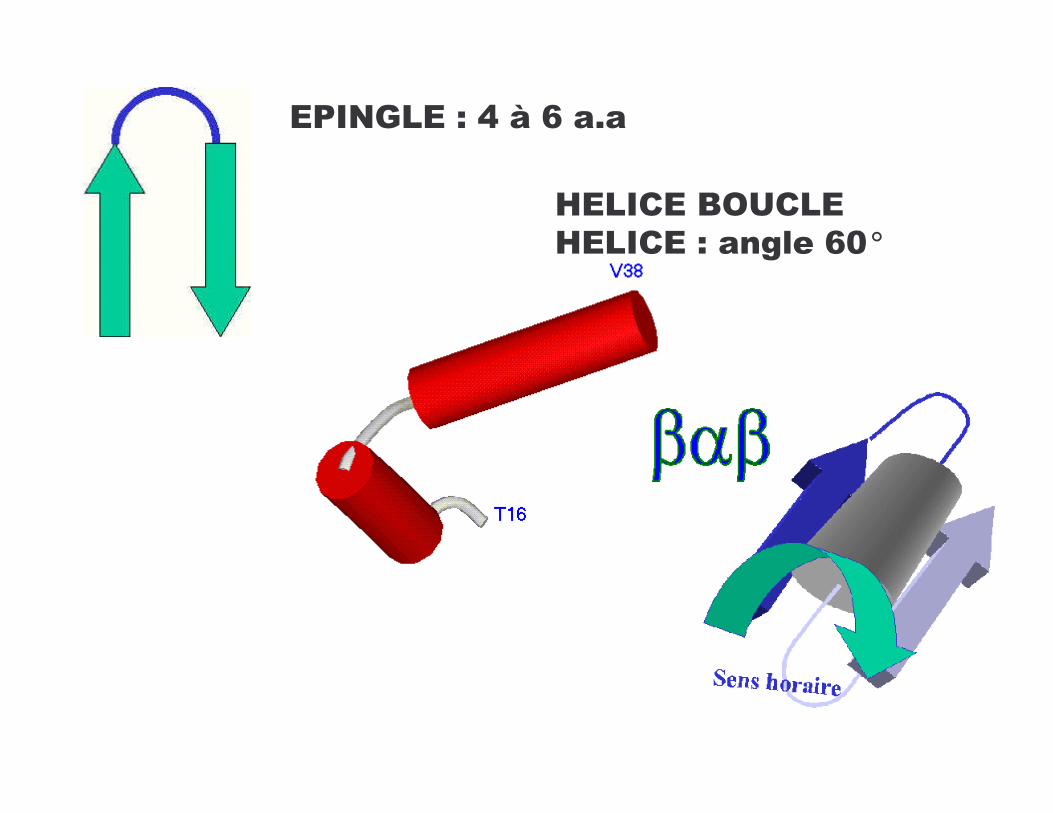
*/��?-*�>�D�8 J��7�
4*-�!*�C� !-*�4*-�!*�>���@���J%°
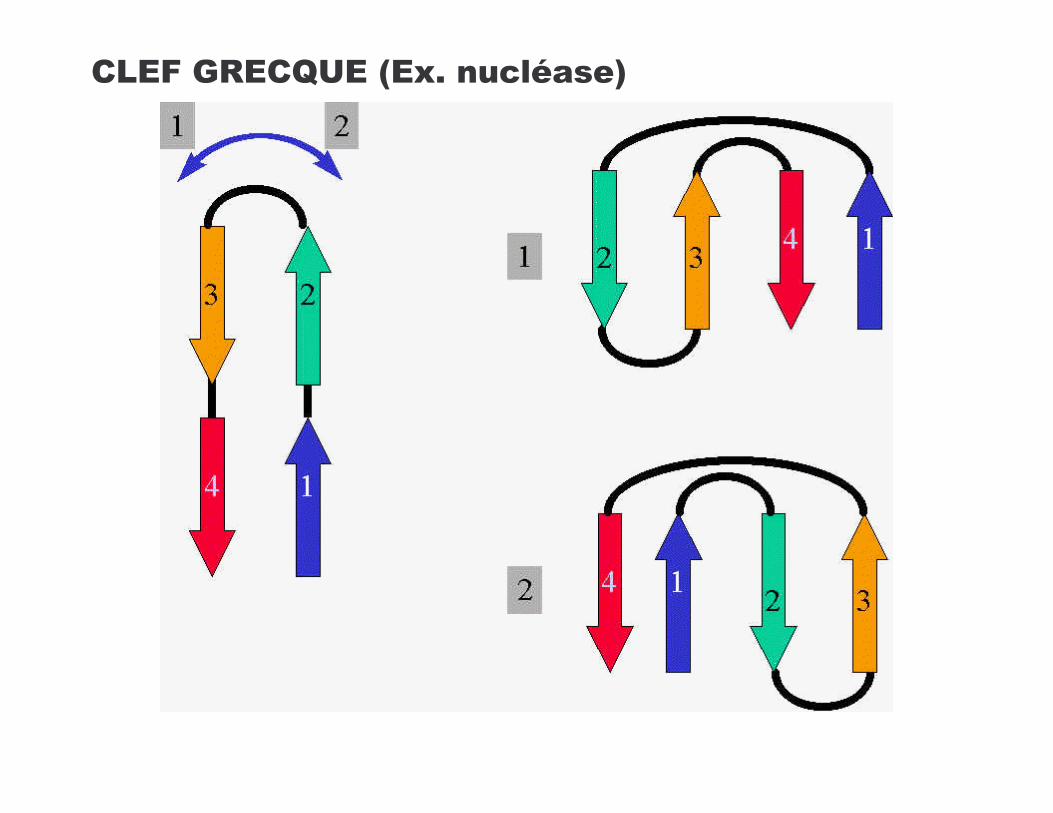
!-*=�?�*!O *��*+7����������

A. Conformation native des protéinesB. Feuillets/hélices/Tours = motifs communsC. Protéines fibrillaires (de structure) ou globulaires (à activité métabolique)
II. Spécificité de fonction et S3D
'9 /���������;�1���������
) �A��:����
) ��-*�:������ !� �*
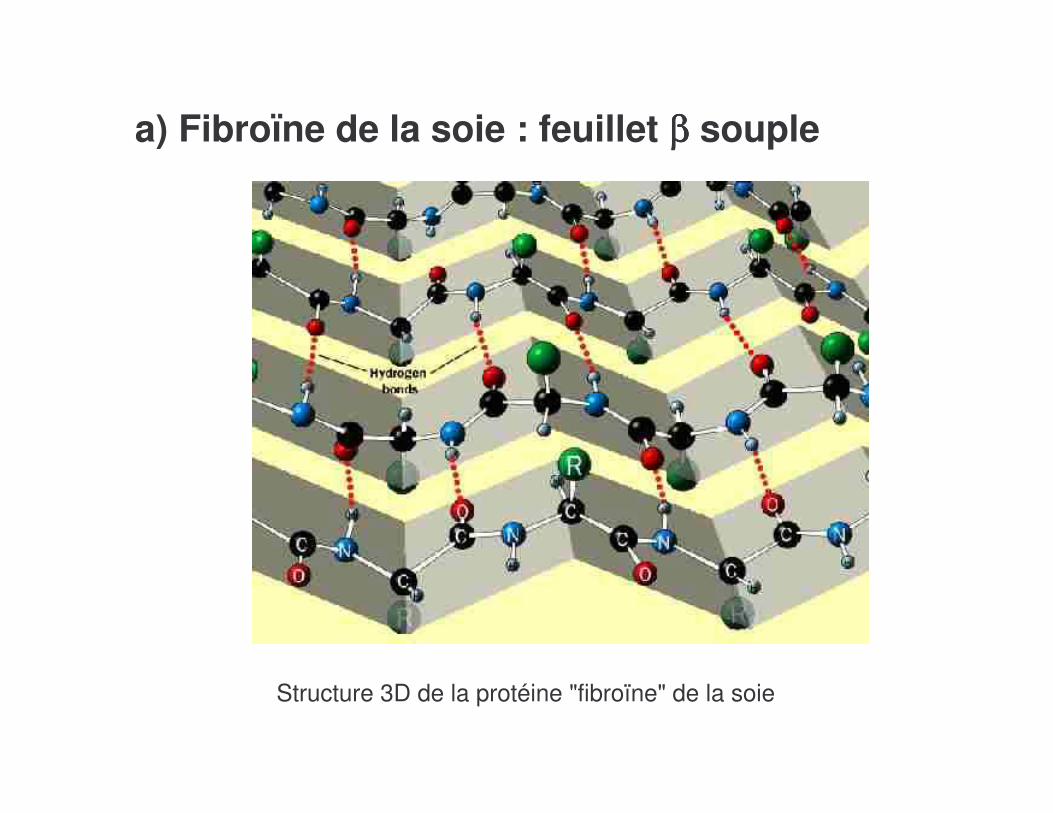
a) Fibroïne de la soie : feuillet ββββ souple
Structure 3D de la protéine "fibroïne" de la soie

a) Fibroïne de la soie : feuillet ββββ souple
b) Kératine : association résistantes de 2 hélices αααα
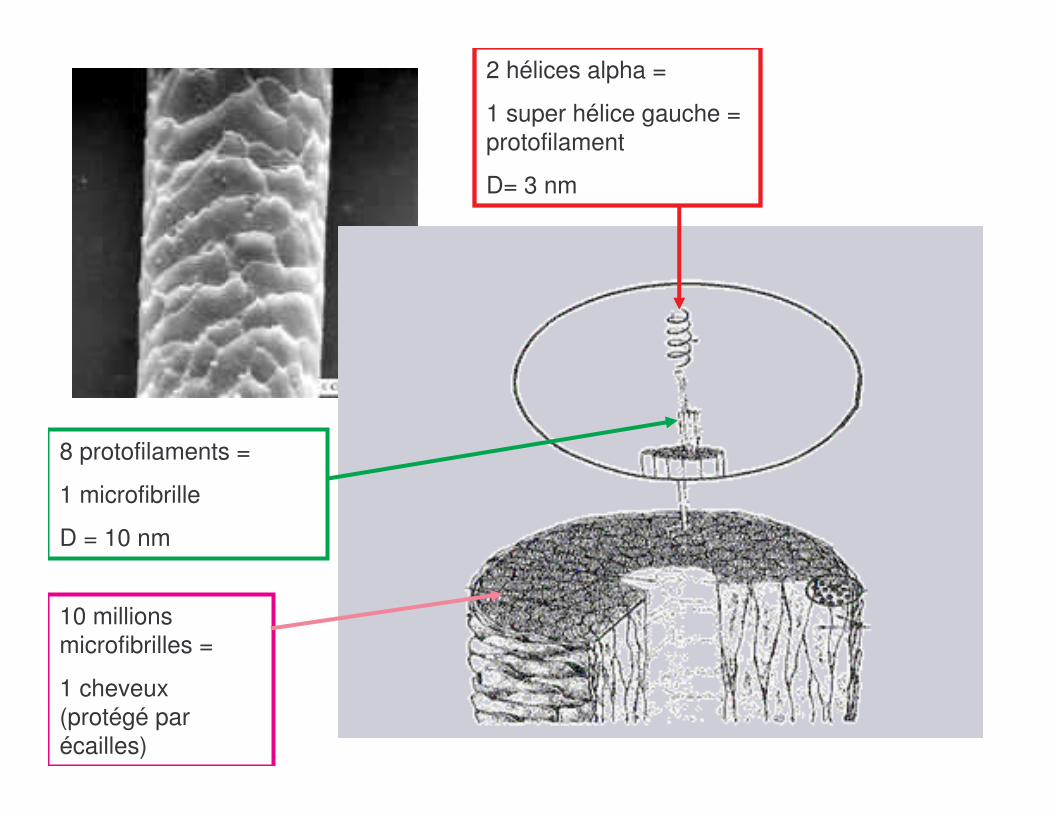
2 hélices alpha =
1 super hélice gauche = protofilament
D= 3 nm
8 protofilaments =
1 microfibrille
D = 10 nm
10 millions microfibrilles =
1 cheveux (protégé par écailles)

a) Fibroïne de la soie : feuillet ββββ souple
b) Kératine : association résistantes de 2 hélices αααα
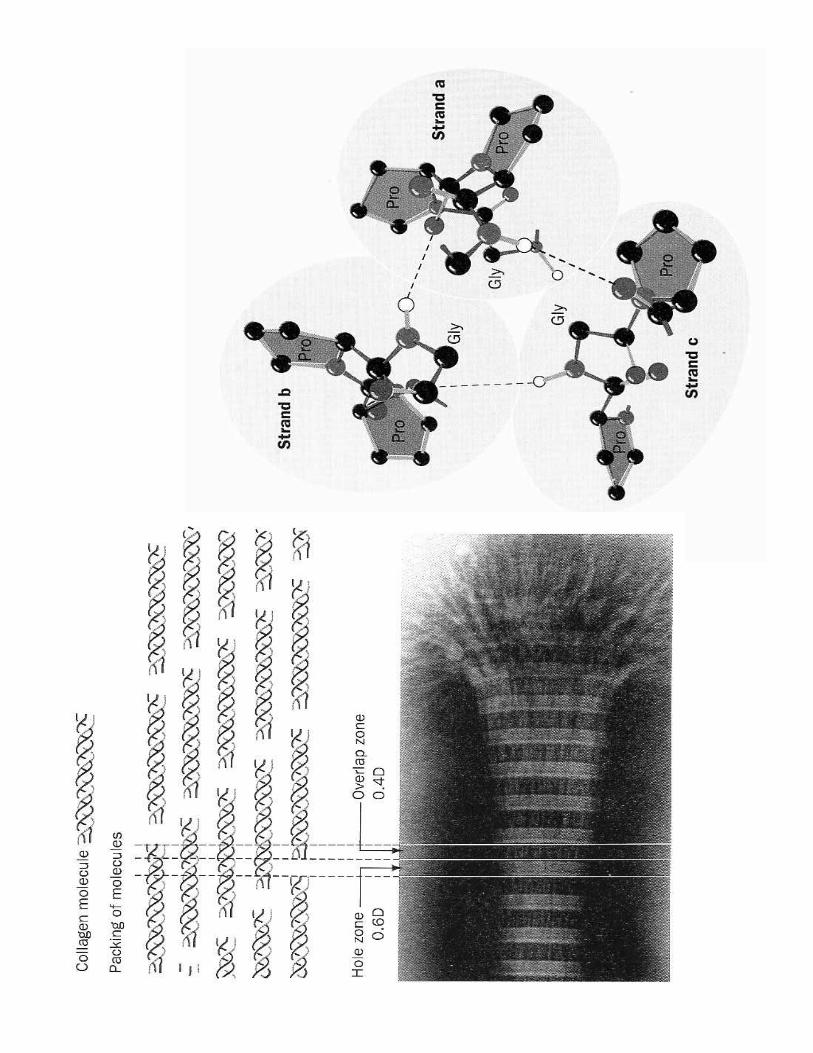
c) Collagène : un câble hélicoïdal dextre de trois hélices gauches très résistant
(cf. cours matrices extracellulaires )

C. Protéines fibrillaires ou globulaires
1- Protéines fibrillaires<9 /���������@��1��������8 ����2���
) *+7���4� �@��1�����41�
) ����������������;;���������P

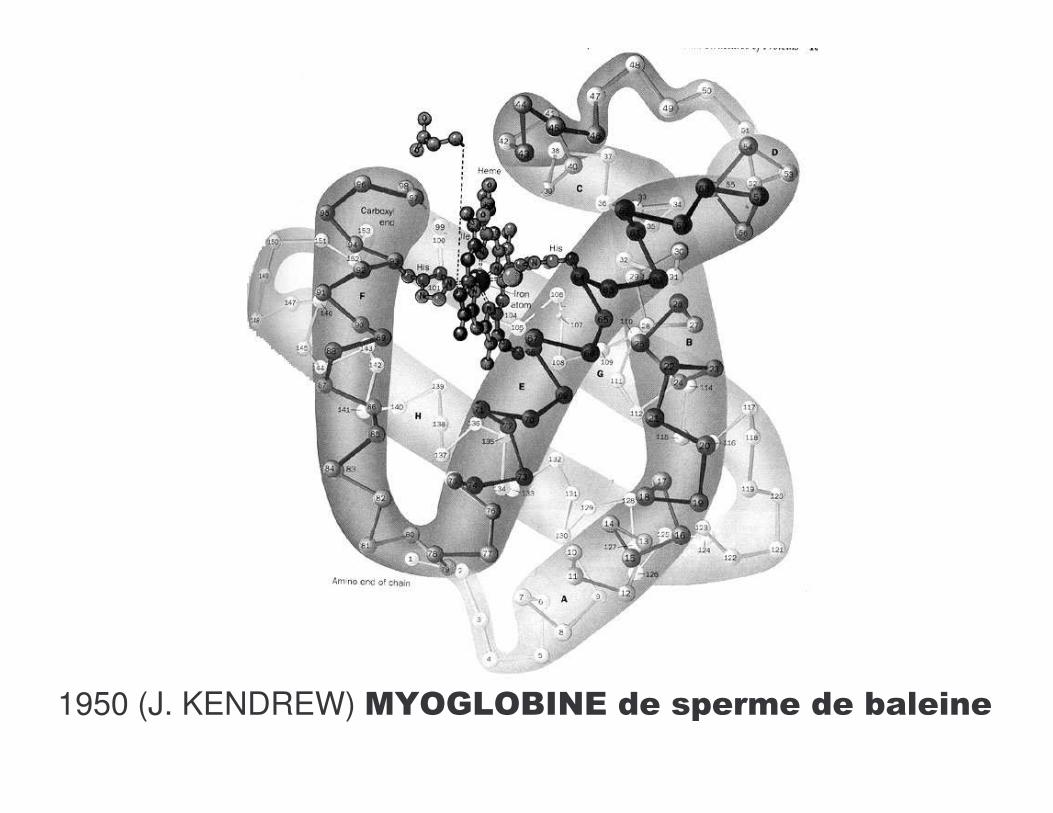
1950 (J. KENDREW) .Q�?-�C��*������� ����1������
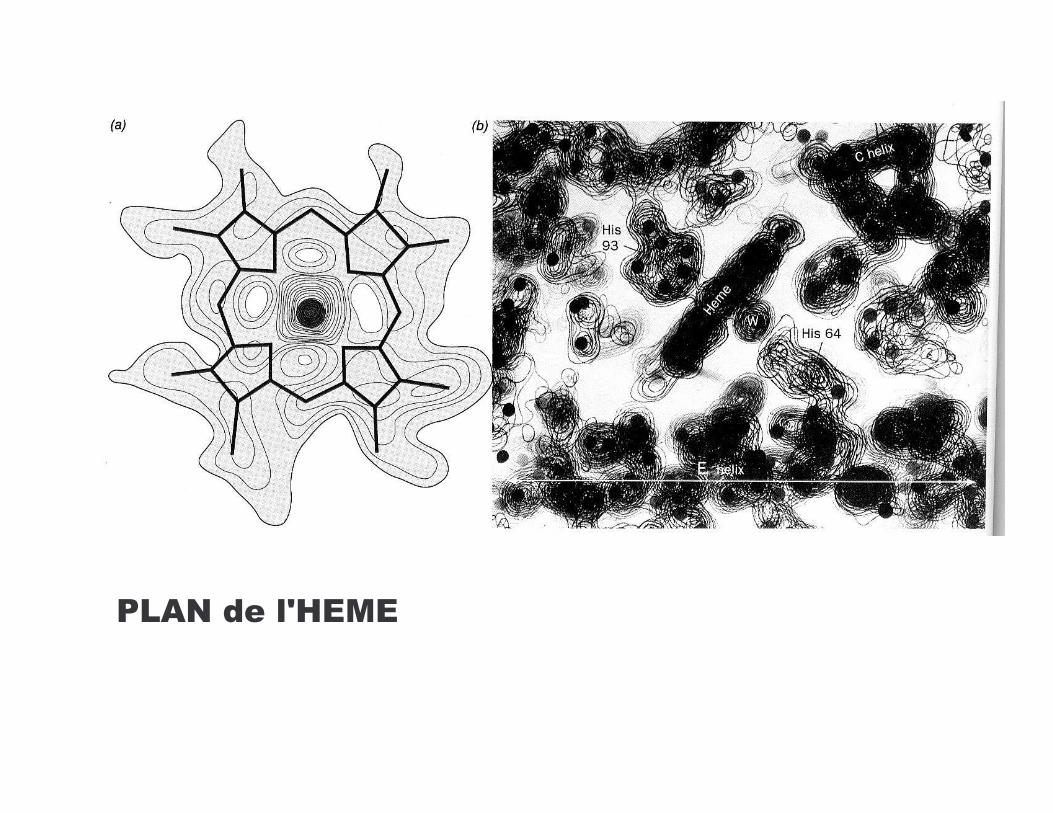
/-6�����4*.*

I. Spécificité de �������������II. Spécificité de fonction liées S3D���7�/���������8 ����������������� �����
Nota : Organisation symétrique des protomères et chiralité

���7�/���������8 ����������������� �����69 � �������������������2�����������: Cas des très grands assemblages
- Synthétiser petites sous unités (cf.préfabriqués)- Réparation aisée des défauts- Lieu de fabrication différent du lieu d'assemblage- IG nécessaire au codage des protéines plus courte

���7�/���������8 ����������������� �����69 � �������������������2�����������: Cas des très grands assemblages
C9 � ������������������2�����������>
Cas des protéines à activité spécifique
Cas de l'Hb

- 1849 : observation cristalisée- 1909 : atlas de 109 espèces- (1926 : première enzyme : l'uréase de pois)- Masse moléculaire- ultracentrifugation et rôle physiologique- mutation liée au changement d'un seul acide aminé - Formulation de théories expliquant le contrôle de l'activitéenzymatique- Structure déterminée aux RX
41 :�/�� �B���8 ������������������2���>

/��1�B ������ >���� �@��1����:
) ���,���0,���������������<
��
) �������B ������������� �����������+�1��������������������������������������2������
R
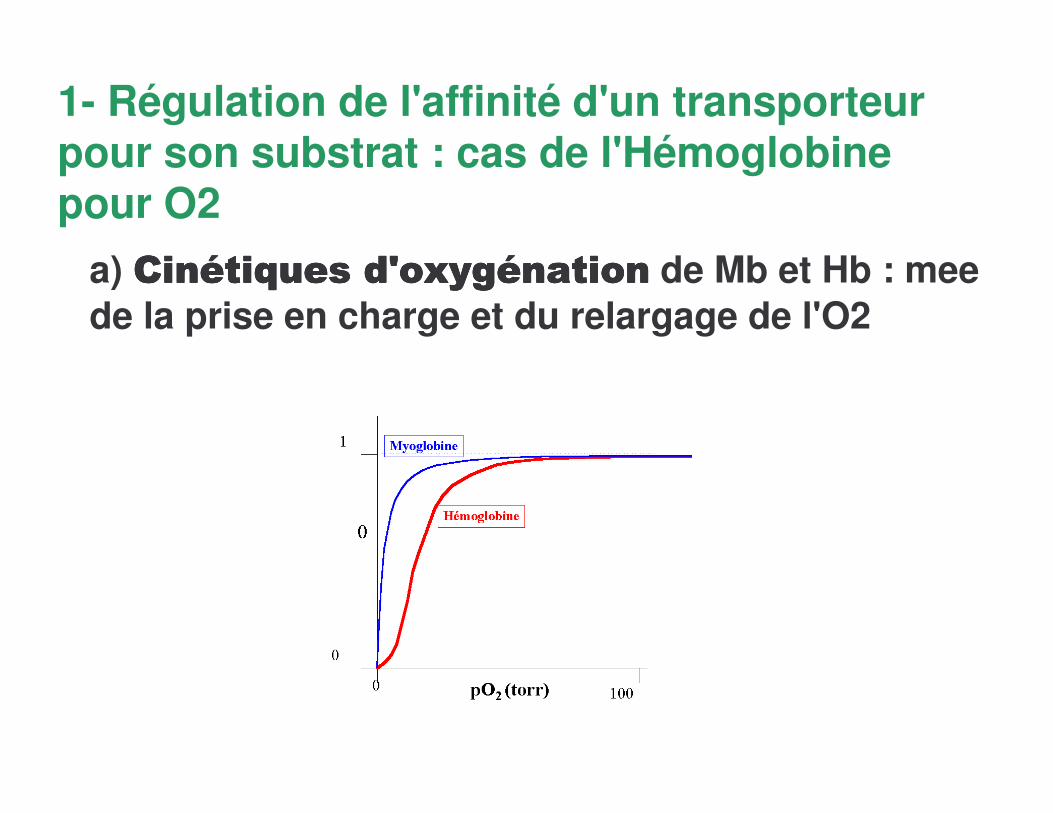
1- Régulation de l'affinité d'un transporteur pour son substrat : cas de l'Hémoglobine pour O2
a) !��!��!��!��������������+�@��������+�@��������+�@��������+�@���������������������������� de Mb et Hb : meede la prise en charge et du relargage de l'O2
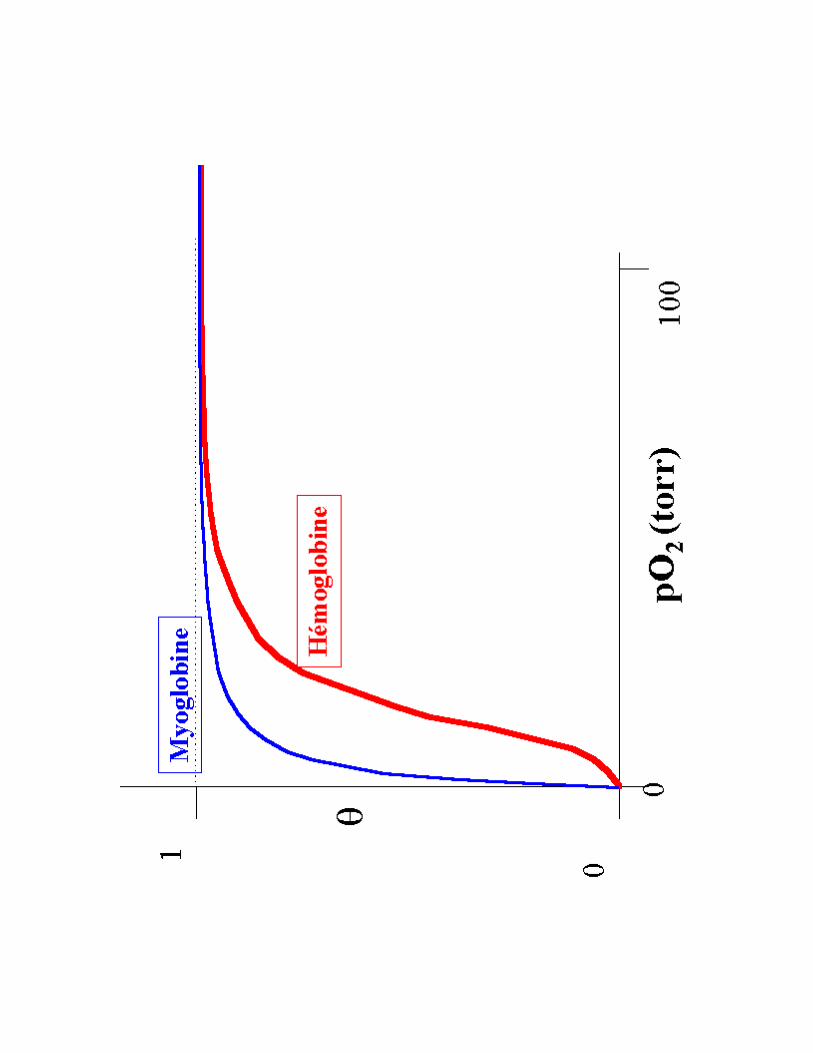

1- Régulation de l'affinité d'un transporteur pour son substrat : cas de l'Hémoglobine pour O2
a) !��!��!��!��������������+�@��������+�@��������+�@��������+�@���������������������������� de Mb et Hb : meede la prise en charge et du relargage de l'O2
b) Mécanismes moléculaires de la fixation de O2
c) Régulation allostérique de la fixation de 02

� Interprétation de l'allure sigmoïde des courbes d'activité
� Interprétation comparée de la structure 3D de Mb et Hb
� Notion d'allostérie
� Notion de transition forme T/R (Tendue / Relâchée) et Régulation allostérique

!������<3A�C�/������?��������
- S<3A�C/?T�:�$� .����1������;�� ���(liaisons supplémentaires entre les protomères : se fixe dans une cavité centrale grâce à a.a chargé +) �favorise le relarguage de O2 dans les tissus
- 6�;��1������������������3�;�2������;�� ���
- �����<3A�C/?3�41 ������1����������@����< dans les tissus = plus d'effet allostérique (Hb se comporte comme Mb!)
- /�� ������������ dans conditions particulières ex. en altitude ( [2,3 BPG] augmente jusqu'à 8 mM)

Bilan :� ������������������������ ��������>
- PO2 forte- CO2 faible- pH faible- Hb saturée : forme R (Hb(O2)4)
� ��������������������������@���������� �������>
- PO2 faible- CO2 forte- pH élevé- Hb relargue tout son O2 : forme T
(désoxyHb)

1- Régulation de l'affinité d'un transporteur
2- Régulation de l'activité d'une enzyme non michaélienne : cas de l'ATCase
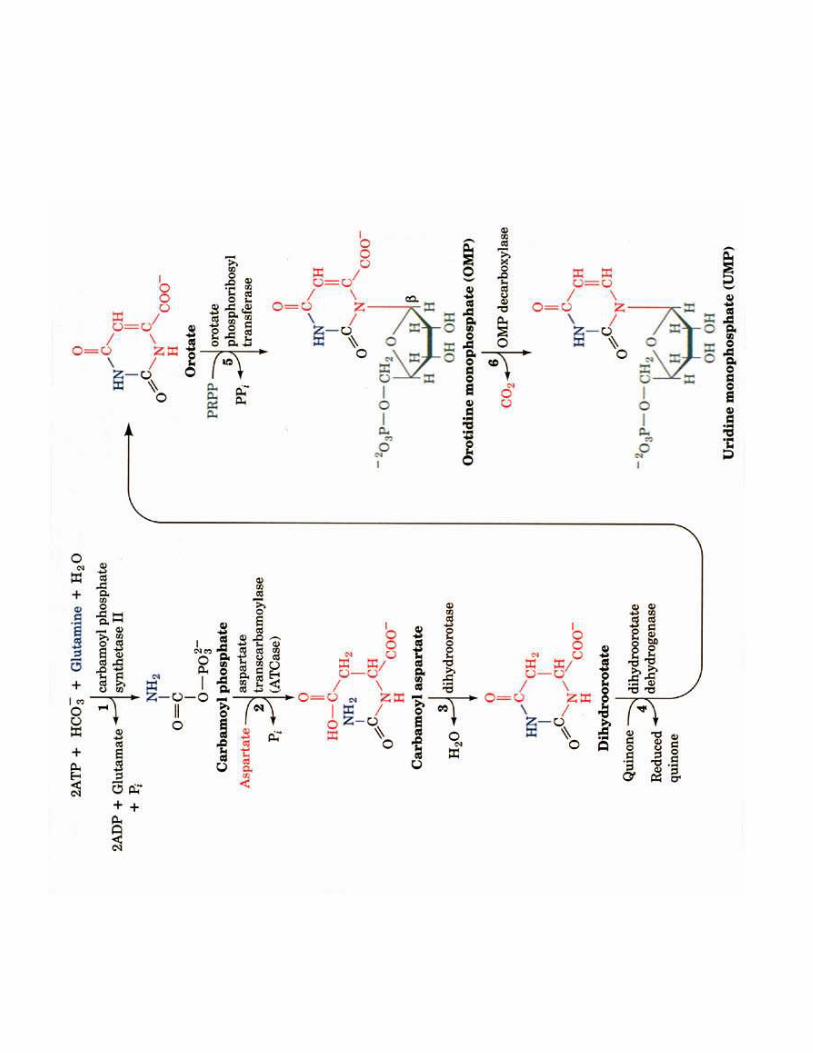
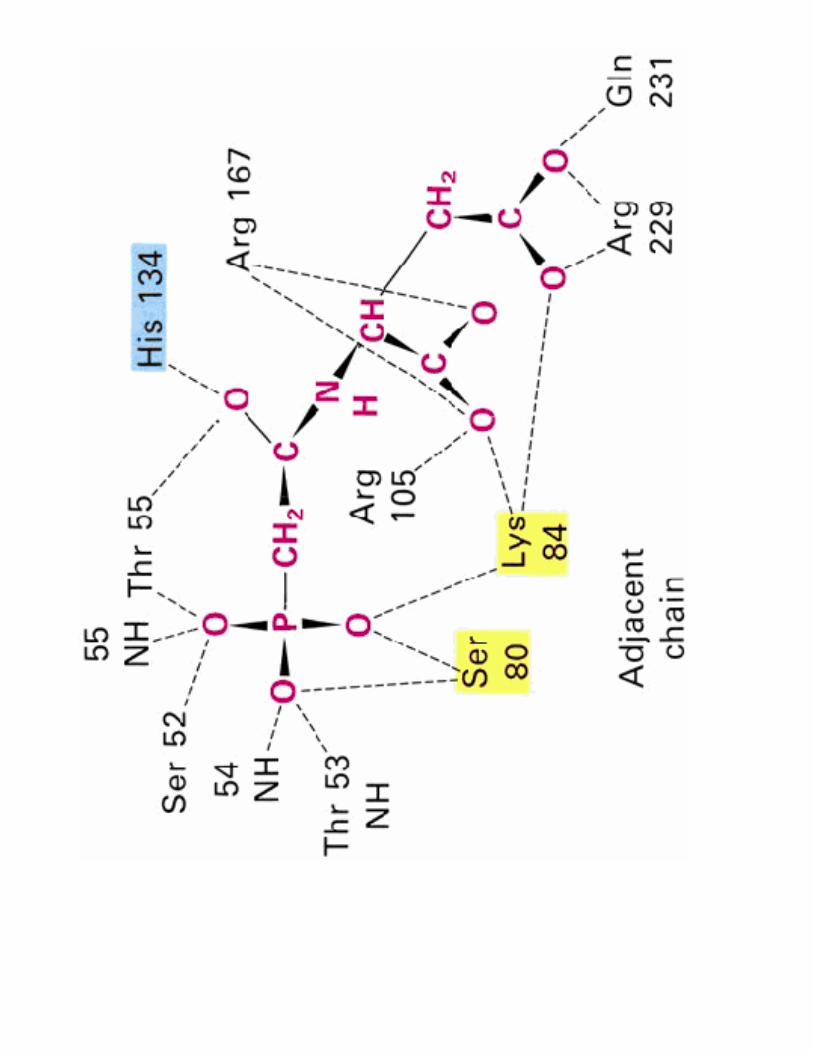
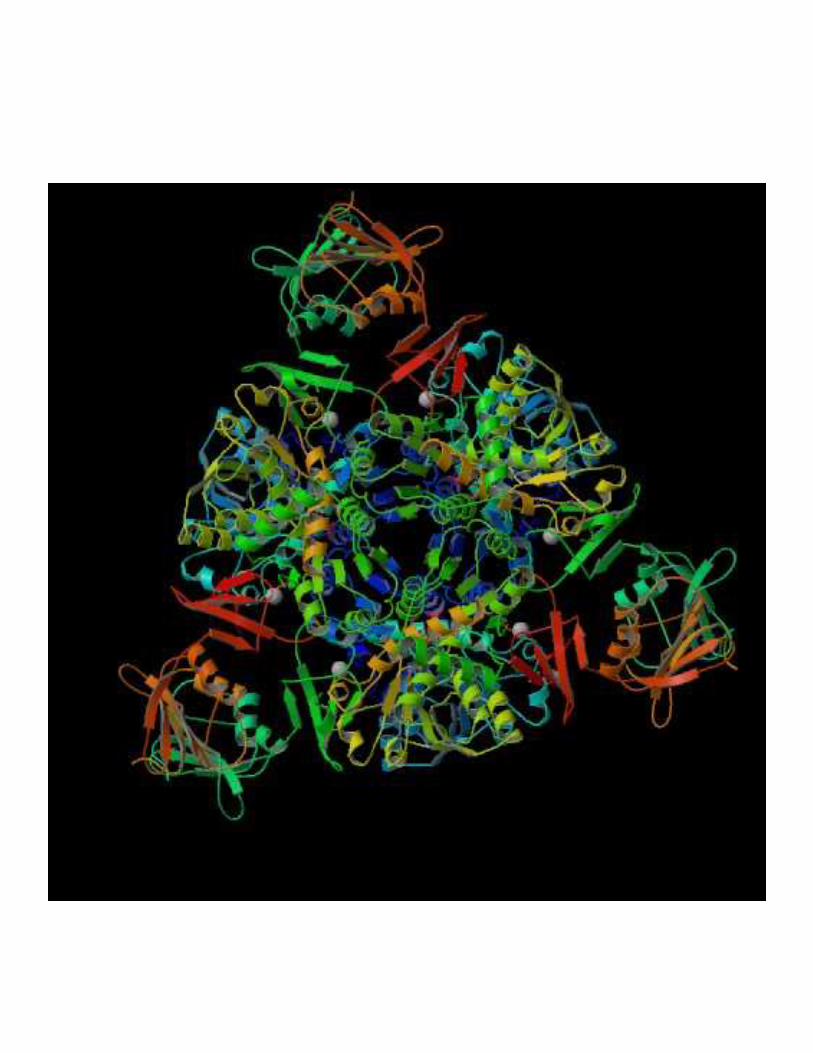
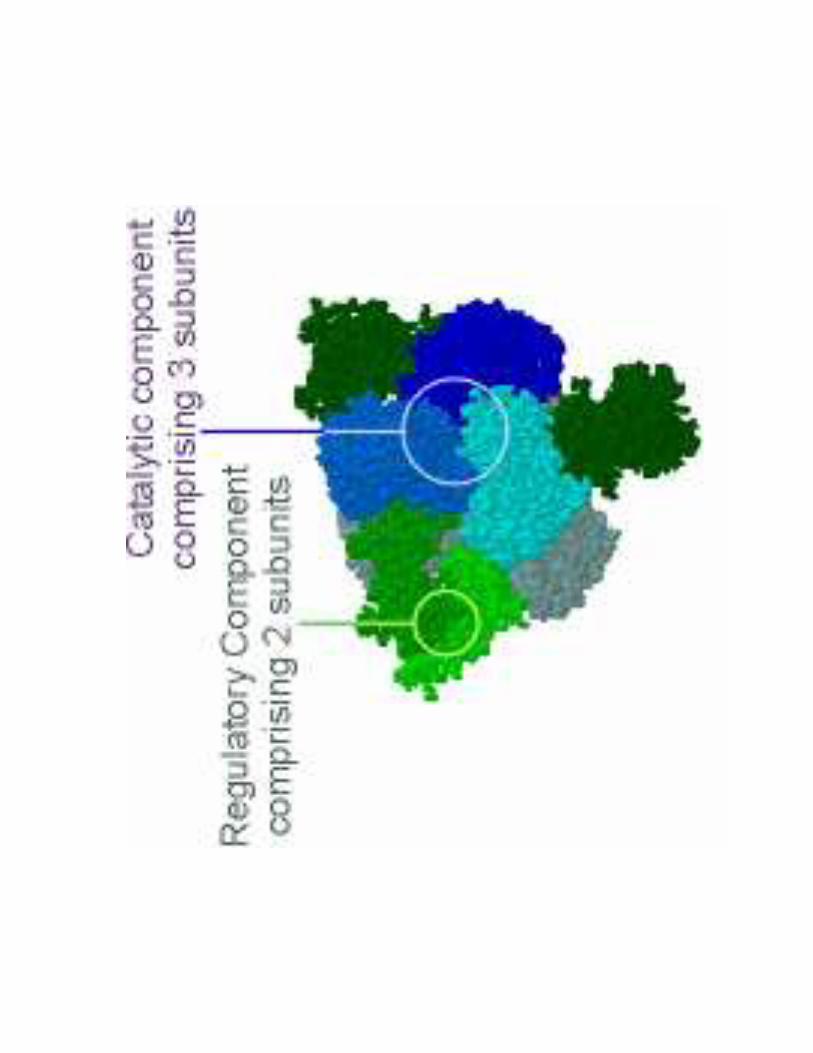
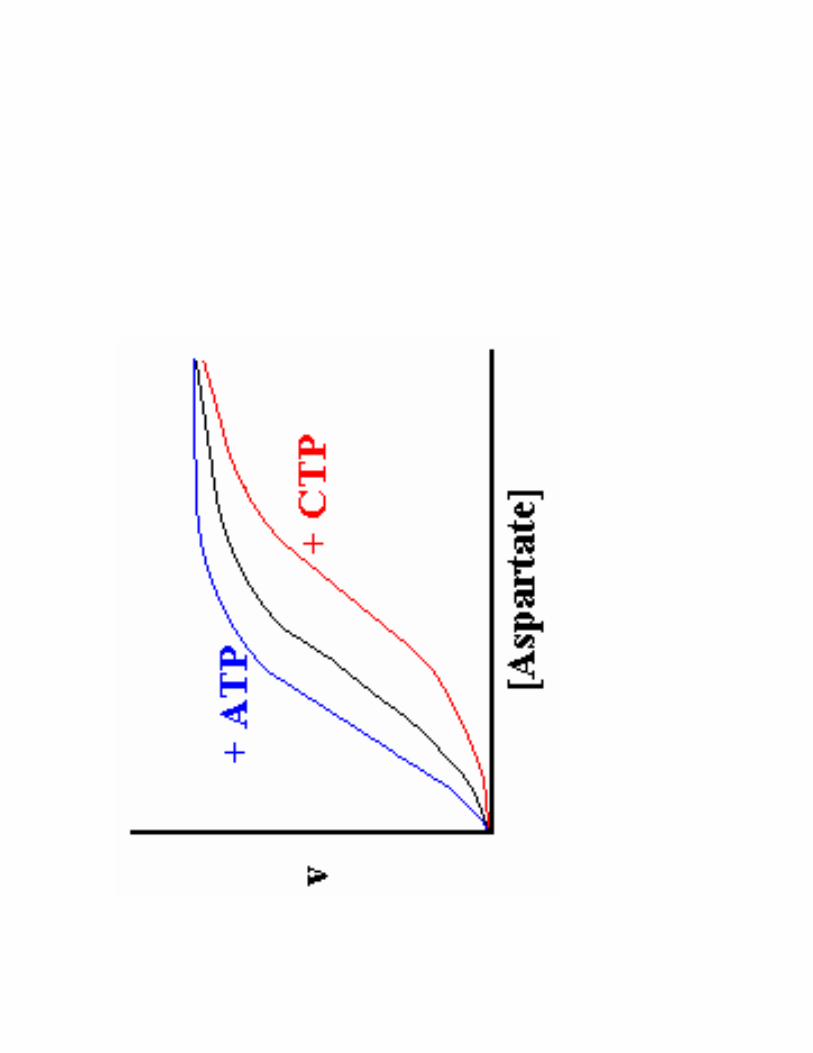
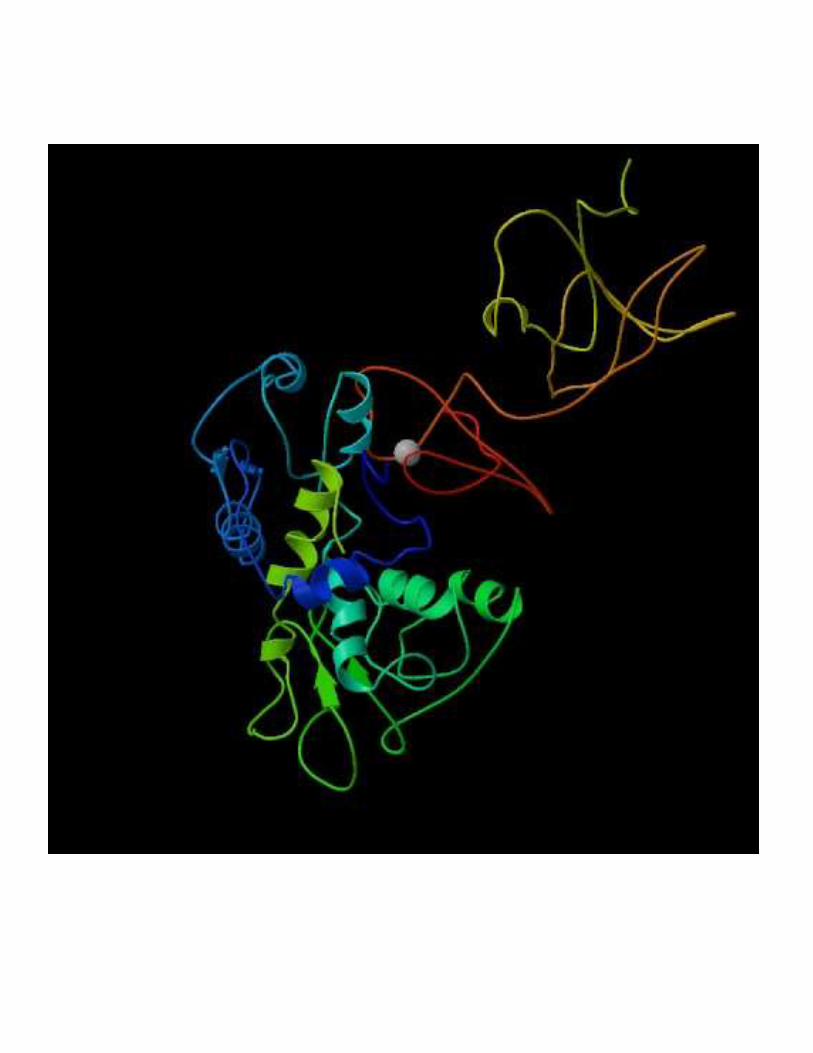
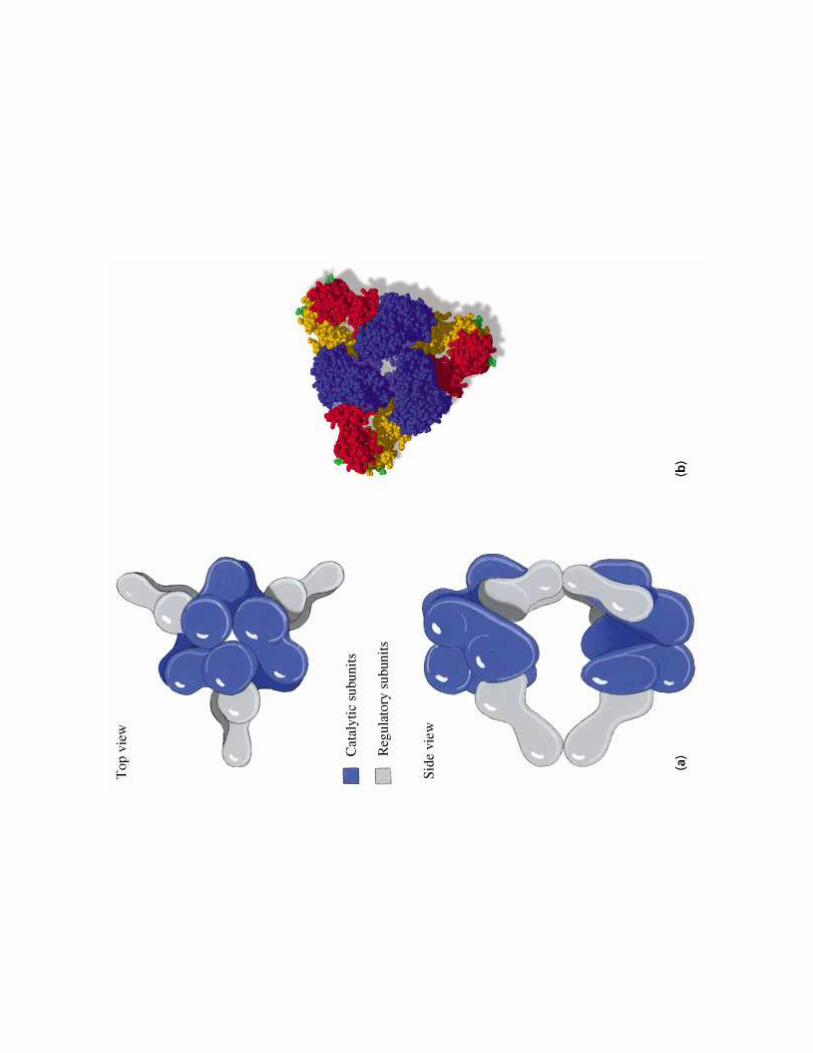
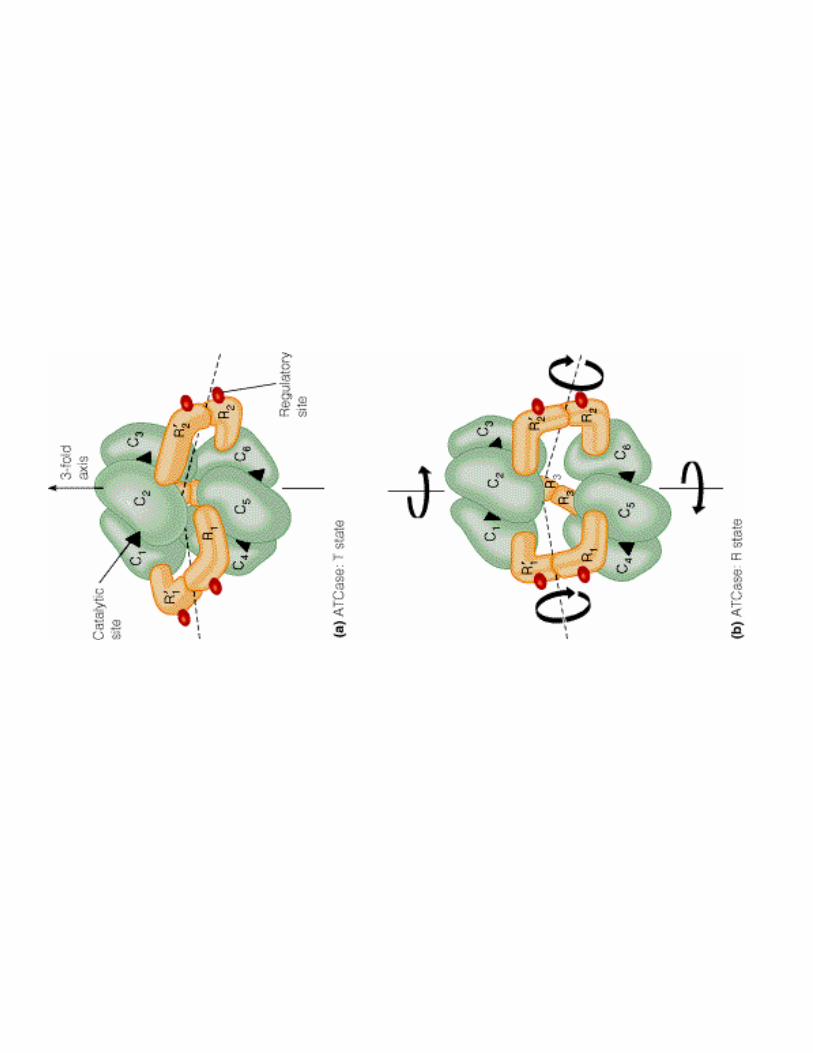

!��!- �����> La stabilité des conformations permet d'établir des interactions moléculaires
'9 ������������� ��������������� �������a) Autoassemblages structuraux fonctionnelsb) Insertion de protéines dans la membrane
<9 ������������� ������������������������a) Récepteur/ligandb) Enzyme/substrat
A9 -�������@� ����������;�� ������������������ �1����� ����������a) Exemple de la contraction de la fibre musculaireb) Généralisation à la mobilité cellulaire