Embed Size (px)
Citation preview

L E S R H I N O C E R O T I D A E ( M A M M A L I A , P E R I S S O D A C T Y L A ) D U M I O C I ~ N E
T E R M I N A L A U P L E I S T O C I ~ N E S U P t ~ R I E U R D ' E U R O P E O C C I D E N T A L E
C O M P A R t ~ S A U X ESPI~CES A C T U E L L E S : T E N D A N C E S t ~ V O L U T I V E S E T
R E L A T I O N S P H Y L O G I ~ N I ~ T I Q U E S
par
CLAUDE GUt~RIN *
R~sum~ Abstract
Pour les cinq lign6es constitu6es par les repr6sen- tants des sous-familles Aceratheriinae, Dicerorhini- nae, Dicerotinae, Rhinocerotinae, Teleoceratinae, les tendances 6volutives sont mises en 6vidence par com- paraison statistique d'6chantillons d'~ge diff6rent, permettant de reconna~tre chez certaines esp~ces plu- sieurs stades 6volutifs successifs. Un essai d'interpr6- tation phylog6n6tique est fourni.
In the five lineage constituted by the Aceratheriinae, Dicerorhininae, Dicerotinae, Rhinocerotinae and Teleoceratinae subfamilies, the evolutionary trends are analysed by statistical comparisons of specific samples of different ages ; in several species it leads to the definition of successive evolutionary stages. A tentative phylogenetic interpretation is given.
MOTS-CL~S:MAMMALIA, PERISSODACTYLA, RHINOCEROTIDAE, NI~OGI~NE, PLI~ISTOCI~NE, ACTUEL, t~VOLUTION, PHYLOGENI~SE, EUROPE OCCIDENTALE.
KEY.WORDS:MAMMALIA, PERISSODACTYLA, RHINOCEROTIDAE, NEOGENE, PLEISTOCENE, RECENT, EVOLUTION, PHYLOGENY, WESTERN EUROPE.
L'6tude des Rhinocerotidae du Mioc6ne terminal au P16istoc6ne d'Europe occidentale et de leurs rapports avec les esp6ces actuelles (1) a donn6, outre les r6sul- tats biostratigraphiques d6jh expos6s (2), des r6sultats concernant l'6volution et la phylog6n6se. Toutes les mesures prises oAt 6t6 trait6es statistiquement ; la
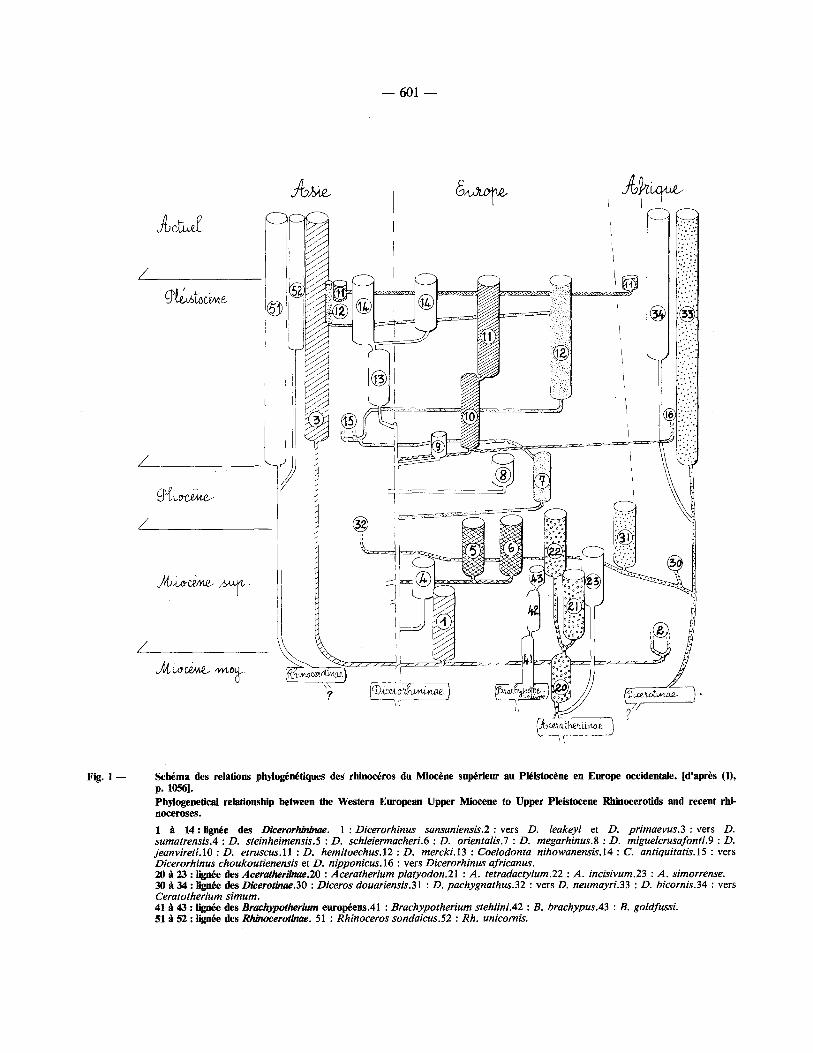
variation intrasp6cifique a 6t6 appr6ci6e ~ l'aide du coefficient de variation, et les diff6rences, en particu- lier entre les stades 6volutifs de la m~me esp6ce, OAt 6t6 analys6es h l'aide du test t de Student. La fig. 1 r6sume les relations phylog6n6tiques et la fig. 2 tente une interpr6tation de celles-ci.
* Centre de Pal6ontologie stratigraphique et Pal6o6cologie, Laboratoire associ6 au CNRS (LA 11), D6partement des Sciences de la Terre, Uni- versit6 Lyon I - Claude Bernard, 69622 Villeurbanne Cedex, France.
Geobios, n ° 15, fasc. 4 p. 599-605, 2 fig. Lyon, Aofit 1982

- - 600
1 - - Sous-famille
a. S o u s - g e n r e Aceratherium
Aceratherium tetradactylum et A. incisivum d6ri- vent tous deux de A. platyodon de l 'Orl6anien d 'Europe.
A. tetradactylum existe pendant les zones 6 ~t 9. Le mat6riel provenant de la zone 9 a 6t6 compar6 au mat6riel astaracien : la branche horizontale de la mandibule devient plus puissante, les dents sup6rieu- res montrent une r6duction de largeur des pr6molaires et un 61argissement des molaires, la rang6e dentaire inf6rieure r6duit son segment pr6molaire, l 'hypsodon- tie des M 3 s'accroit, les diaphyses de certains os deviennent plus massives (radius, Mc III).
A. incisivum existe pendant les zones 9 A 13. J 'a i compar6 le mat6riel r6parti en trois 6chantillons : zone 9, zones 10 et 11, zones 12 et 13. L'esp~ce est remarquablement stable ; il semble qu'i l y ait ten- dance d 'abord ~ l 'affinement, puis ~ l'616vation de la branche horizontale de la mandibule, 61argissement des jugales inf6rieures et accroissement de la massivit6 de certains m&apodes et tarsiens ; la forme ultime de l'esp~ce (zones 12 et 13) ne parak pas confirmer les tendances montr6es par l '6chantillon interm6diaire (zones 10 et 11), au contraire : elle serait p lu t f t plus petite et moins massive. La discontinuit6 des tendan- ces 6volutives et les diff6rences biom6triques qui sont
Aceratheriinae
souvent significatives permettent de distinguer trois stades 6volutifs, le stade I pour le Vall~sien inf6rieur de la zone 9, le stade II pour les zones 10 et 11, le stade III pour les zones 12 et 13.
I1 serait tentant de faire descendre A. incisivum de A. tetradactylum du fait de la ressemblance morpho- logique des deux esp~ces, qui de plus occupent un bio- tope analogue, mais les deux esp~ces coexistent pen- dant la zone 9, et il est logique de leur assigner une origine commune par cladogen~se A partir de A. platyodon (fig. 1 et 2).
b. S o u s - g e n r e Alicornops
Aceratherium simorrense existe de la zone 6 ~ la zone 10. J 'ai compar6 le mat6riel vall6sien des zones 9 et 10 au mat6riel des zones 7 et 8. Les tendances 6volu- tives sont l 'augmentation de taille des dents jugales, l 'accroissement de l 'hypsodontie des jugales sup6rieu- res, l 'augmentation de la taille et de la robustesse des os des membres.
L'esp~ce constitue la fin d 'un phylum bien distinct de celui constitu6 par le sous-genre pr6c~dent ; le sous-genre Alicornops, immigr6 en Europe au d6but de l'Orl6anien, existe ~ Wintershof-West (MN3)d~s la base du Miocene moyen.
2 - - S o u s - f a m i l l e Dicerorhininae
a. S o u s - g e n r e Dicerorhinus
Je classe dans ce sous-genre des formes primitives sans cloison nasale ossifi~e et pourvues d 'une denture ant6rieure fonctionnelle. Ces formes comprennent l 'actuel D. sumatrensis, qui n 'a gu~re vari~ depuis le P16istoc~ne moyen. D. sansaniensis est connu de la zone 4 b ~ la zone 9 . En d6pit d 'une vaste extension dans le temps l'esp~ce est rare et je n 'ai pas r6ussi ~t mettre en 6vidence ses tendances 6volutives. Morpho- logiquement D. sansaniensis est tr6s voisin du D. lea- keyi qui est son contemporain en Afrique, et proche de l 'actuel D. sumatrensis ; il a dfi ~tre tr~s semblable /l la forme souche du phylum (fig. 1 et 2).
D. steinheimensis est une esp6ce plus r6cente que la pr6c6dente (zones 7 ~ 9). Une comparaison du mat6- riel vall6sien au mat6riel astaracien montre, malgr6 la pauvret6 des 6ehantillons que les pr6molaires sup6- rieures tendent ~ accroitre leur taille et leur hypsodon- tie. Le crane de l'esp~ce n'est pas connu, et il est bien
difficile de distinguer en son absence ~ quelle lign6e appartient l'esp~ce. Toutefois les caract~res dentaires sont ceux du sous-genre nominal, et ceux du squelette des membres ne s'en distinguent que par la taille tr~s petite et l 'allure trapue ; il est donc logique de faire d6river D. steinheimensis de la lign6e des Dicerorhi- nus comme indiqu6 fig. 2.
Dicerorhinus schleiermacheri est cantonn6 ~t l 'Europe occidentale et centrale mais existe pendant tout le Mioc6ne terminal (zones 9/L 13). La comparai- son d 'un 6chantillon vall6sien et d 'un 6chantiUon p i k e r m i e n m o n t r e les t e n d a n c e s 6volu t ives suivantes • r6duction de la taille de la branche hori- zontale de la mandibule, diminution du volume des jugales inf6rieures, augmentation de la puissance des carpiens et tarsiens, accroissement de la massivit6 des m6tapodes m6dians ; il y a deux stades 6volutifs, le stade I pour le VaU6sien et le stade II pour les zones 12 et 13. I1 est possible que la d6couverte de mat6riel nouveau amine a reconnaitre un troisi~me stade car
- - 601 - -
/
i®
i
'I
Z
/
/
,! ®
I@
Fig. 1 - Scbema des relations phylog~n~tiques des rhinoceros du Miocene sup~rieur au Pleistocene en Europe occidentale. [d'apr~s (D, p. 10561. Phylogenelical relationship between the Western European Upper Miocene to Upper Pleistocene Rhinocerofids and recent rhi- noceroses.
1 i 14 : f ign6e des Dicerorh'minae. l : D i c e r o r h i n u s sansaniens is .2:vers D. leakeyi et D. p r i m a e v u s . 3 : v e r s D. sumatrensis.4 : D. steinheimensis.5 : D. schleiermacheri.6 : D. orientalis.7 : D. megarhinus.8 : D. miguelcrusafonti.9 : D. jeanvireti . lO : D. e truscus . l l : D. hemitoechus.12 : D. mercki.13 : Coelodonta nihowanensis.14 : C. antiquitatis.15 : vers Dicerorhinus choukoutienensis et D. nipponicus.16 : vers Dicerorhinus africanus. 20 ~ 23 : lign6e des Aceratheriinae.20 : Aceratherium platyodon.21 : A . tetradactylum.22 : A . incisivum.23 : A . simorrense. 30/ t 34 : lign~e des Dicerot~uTe.30 : Diceros douariensis.31 : D. pachygnathus.32 : vers D. neumayri.33 : D. bicornis.34 : vers Ceratotherium s imum. 41 ~t 43 : lign~e des B r a c h y p o t h e ~ m eurol~ens .41 : Brachypotherium stehlini.42 : B. brachypus.43 : B. goldfussL 51 ~ 52 : lign~e des Rhinoceroanae. 51 : Rhinoceros sondaicus.52 : Rh. unicornis.

- - 602
les restes de la zone 9 semblent particuli~rement volu- mineux. D. schleiermacheri se distingue par l'acquisi- tion d 'une grande taille et d 'un squelette des membres de type semi-cursorial, deux caract~res synapomor- phes qui apparaissent ~t plusieurs reprises dans la famille (fig. 2).
D. megarhinus n'existe qu 'au Ruscinien (zones 14 et 15) et constitue l '6volution ultime du sous-genre • la denture ant6rieure existe encore mais n'est plus fonc- tionelle. La comparaison de deux 6chantillons des zones 14 et 15 montre comme tendances 6volutives une for te r6duct ion de volume des jugales sup6rieures,une faible r6duction des jugales inf6rieu- res, un net accroissement de l 'hypsodontie, un rac- courcissement et un affinement des os longs, une r6duction de taille des carpiens et tarsiens, un amincis- sement des m6tacarpiens et un raccourcissement des m6tatarsiens qui deviennent plus graciles. Les diff6- rences sont souvent significatives ou hautement signi- ficatives et deux stades 6volutifs, un pour la zone 14 et l 'autre pour la zone 15, se distinguent ais6ment. Les fig. 1 et 2 montrent deux interpr6tations possibles des relations phylog6n6tiques de l'esp6ce.
b. Sous-gem'e ind6termin6
D. miguelcrusafonti n'est connu que de deux gise- ments de la zone 15. Les dents sont caract6ristiques du genre, les os des membres sont trapus, mais l 'absence de crfme et de mandibulene permet pas de savoir dans
- quelle lign6e se situe l'esp6ce ; la r6partition tempo- relle trop br6ve emp~che pour l ' instant de conna~tre les tendances 6volutives.
c. Sous-genre Brandtorhinus
Ce sous-genre d6finit une lign~e qui se caract6rise par l 'acquisition d 'une cloison nasale ossifi6e et la perte de la denture ant6rieure.
Dicerorhinus jeanvireti est le plus ancien repr6sen- tant connu du sous-genre. L'esp~ce n'existe que pen- dant la zone 16 (Villafranchien inf6rieur) mais cette zone dure suffisamment longtemps pour que la com- paraison de l '6chantillon le plus ancien (mat6riel de Vialette, ~ l'extr~me d6but de la zone) avec le reste du mat6riel fasse appara~tre quelques tendances 6voluti- ves : raccourc issement du f6mur, allongement du radius et du tibia, allongement et affinement des m6tapodes ; les fig. 1 et 2 montrent deux interpr&a- tions possibles des relations phylog6n6tiques de l'esp~ce.
Dicerorhinus etruscus existe de la zone 16 ~ la zone 22, donc pendant tout le Villafranchien et une bonne part du P16istoc~ne moyen ancien. La sous-esp~ce
nominale compte deux stades 6volutifs, le stade I pour le Villafranchien inf6rieuret moyen (zones 16 et 17) et le stade II pour le Villafranchien sup6rieur (zones 18 et 19) ; du stade I au stade II les tendances 6volutives sont les suivantes : modification des proportions du cr~me avec recul de l 'orbite, ~largissement de l'arri~re- cr,~ne et du toit cr~nien, abaissement de la face occipi- tale ; amincissement et abaissement de la branche horizontale de la mandibule ; diminution de taille des den t s et r 6 d u c t i o n r e l a t i v e des s egmen t s pr6molaires ; r6duction de taille du radius et modifi- cation de taille des 6piphyses des os longs, raccourcis- sement et modification des proportions des m6tapo- des. La sous-esp~ce brachycephalus se reconna~t h son crane plus allong6, plus large et plus haut, ~ ses dents plus volumineuses et pour certaines plus hypsodontes, ~t ses os longs plus puissants et ses m6tapodes plus allong6s. Cette sous-esp~ce d6rive directement de la sous-esp6ce nominale.
Dicerorhinus mercki apparait en Europe occiden- tale au d6but du P16istoc6ne moyen (zone 20) et y sub- siste jusqu'au P16istoc6ne sup6rieur terminal (zone 26). J 'ai r6parti le mat6riel dont je disposais en trois 6chantillons, l 'un correspondant aux zones 20 et 21, le second aux zones 22 et 23, le troisi~me aux zones 24, 25 et 26. De l'6chantillon le plus ancien h l'6chantillon le plus r6cent les tendances 6volutives sont les suivan- tes • raccourcissement et affinement de la mandibule, modification des dimensions relatives des dents (en g6n6ral r6duction de taille mais augmentation de lon- gueur pour p4 et M3 et augmentation de largeur pour p3 et p4), r6duction de taille des os longs, carpiens, tarsiens et m6tapodes. I1 y a deux stades 6volutifs dis- tincts, un pour les zones 20 et 21, un pour les zones 24 ~t 26 ; l '~chantillon interm6diaire sugg~re qu'il pour- rait lui aussi correspondre ~ un stade, mais il n'est pas suffisant pour qu'il soit possible d' en d6cider actuelle- ment.
L'origine de D. mercki peut ~tre recherch6e du c6t6 de D. megarhinus (fig. 1), ou, si l 'on adopte le point de vue cladistique, et en vertu du principe de parcimo- nie, dans le rameau qui m~ne d 'une part ~ D. mega- rhinus, d'autre part aux Brandtorhinus plus 6volu6s et
Coelodonta (fig. 2).
D. hemitoechus appara~t en Europe occidentale vers la fin de la zone 22, au milieu du Pl6istoc~ne moyen, et perdure jusqu'~t la zone 26. J 'a i r6parti le mat6riel en 3 6chantillons et l '6tude statistique montre qu'il y a deux stades 6volutifs, l 'un pour la fin de la zone 22 et la zone 23, l 'autre pour les zones 25 et 16 , ce dernier
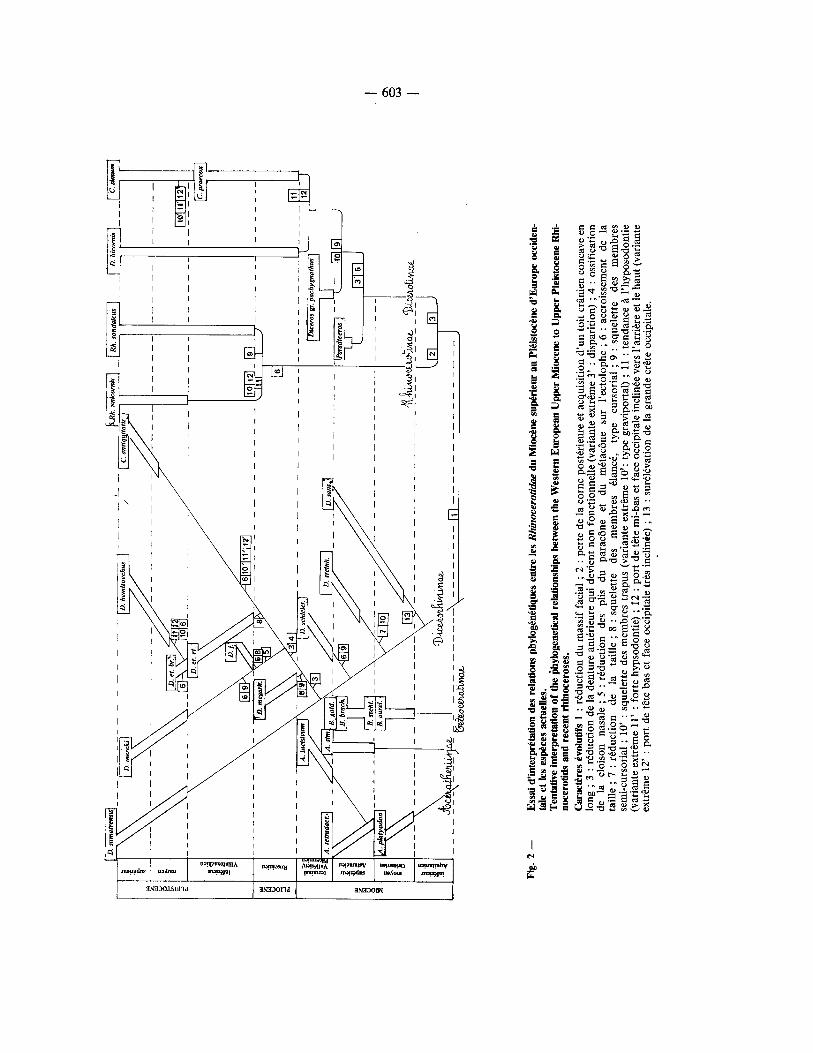
-- 603 --
i
\
I
U 1
i
zn~u?dns uo,(otu ualq~u~elSA
may~tu]
I
I I
T
I I
:IN~30]l~[
i
I I I
l I
/~!~yll~A u~Jrr~sv u"~p~lz 0 ua~u~nbv I~mBu2~l a ~ l m ua~om
"" = ' o "~ .~
• ~ ~ o.. ~_,,~_~ ~ ~ ~ ~.~
~ " ~
o o "° ~ ~ 7.
.! g
@
.g~.
0~
==-~
0 ~ 0 ~ ~
~ rj ~ .~ ~:~
I

604 --
stade correspond peut-~tre ~t la sous-esp~ce aretinus AZZAROLI ; les tendances 6volutives concernent la mandibule (accroissement de hauteur de la branche horizontale), les dents (61argissement des molaires sup6rieures et des jugales inf6rieures, r6duction du segment pr6molaire, 16ger accroissement de l'hypso- dontie) et le squelette des membres (allongement de l'hum6rus, du f6mur et du tibia, raccourcissement du radius, augmentation de volume de l'astragale et du calcan6um, raccourcissement des Mc III et Mc IV, allongement des Mt III et IV).
D. hemitoechus d6rive par cladog6n~se de D. etrus- cus etruscus (fig. 2).
d. G e n r e C o e l o d o n t a
Ce genre correspond ~t l'6volution ultime des Dice- rorhininae. C. antiquitatis d'origine asiatique appa- rait en Europe occidentale au d6but de la zone 24 avec la sous-esp~ce C. a. praecursor ; pendant les zones 25 et 26 c'est la sous-esp~ce nominale qui est pr6sente. D'une sous-esp~ce ~t l'autre les tendances 6volutives concernent surtout la stature : les os longs s'allongent 16g~rement et les m6tapodes se raccourcissent faible- ment, tous devenant plus trapus.
3 - - S o u s - f a m i l l e Dicerotinae
a. G e n r e Diceros
Les premiers repr6sentants de ce genre datent de la fin du Miocene ; il s'agit d'un phylum circum-m6di- terran6en represent6 par les esp~ces pachygnathus d'Europe orientale, neumayri de Turquie et douarien- sis d'Afrique du Nord. Ces trois formes sont regrou- p6es sous le nom de Diceros gr. pachygnathus dans la fig. 2, et ont un repr6sentant en Espagne pendant la zone 13. Les rapports phylog6n6tiques avec les autres membres de la sous-famille sont donn6s fig. 1 et 2.
Diceros bicornis est une esp~ce actuelle connue depuis le Plioc~ne ; cette esp~ce particuli~rement sta- ble a si peu vari6 depuis son apparition que ses
repr6sentants fossiles n'ont pu pour l'instant ~tre dis- tingu6s taxonomiquement des formes actuelles.
b. G e n r e C e r a t o t h e r i u m
L'actuel Ceratotherium simum est directement issu au P16istoc~ne moyen du Ceratotherium praecox apparu au Plioc~ne. C. simum pr6sente de nombreu- ses convergences (synapomorphies) avec Coelodonta antiquitatis. De C. praecox ~t C. simum l'6volution a port6 sur un accroissement de l'hypsodontie et surtout sur une modification des proportions des os longs et des m&apodes qui deviennent beaucoup plus trapus, les membres prenant un type franchement gravipor- tal.
4 - - S o u s - f a m f l l e Rhinocerotinae
A 1'exception d'une incursion du genre Gandaithe- rium dans le mioc6ne ancien d'Europe occidentale il s'agit l~t d'un phylum exclusivement asiatique. Rhino- ceros sondaicus, connu depuis le d6but du Villafran- chien, est la moins 6volu6e des deux esp~ces actuelles ; deux sous-esp6ces pl6istoc~nes pr6c6dent
les sous-esp~ces actuelles.
Rhinoceros unicornis est une forme bien 6volu6e (denture hypsodonte, port de t~te mi-bas) apparue au P16istoc~ne moyen ; une sous-esp~ce pl6istoc~ne a 6t6 d~cdte ~t Java.
5 - - S o u s - f a m i l l e Teleoceratinae
Le genre Brachypotherium est connu au N6og~ne en Asie, Afrique, Europe. Les formes du Miocene moyen au Miocene terminal d'Europe me semblent constituer une anagen~se avec 4 esp~ces successives, que l'on rencontre de rOrl6anien au Vall6sien inf6- rieur ; chacune des esp~ces exprime les m~mes ten- dances 6volutives (accroissement de taille, accroisse- ment de l'hypsodontie).
Je d6signe cf. Prosantorhinus sp: D quelques restes trouv6s dans le gisement Vall6sien sup6rieur de Mon- tredon (H6rault, France) ; ce materiel est trop pauvre pour pouvoir ~tre compar6 avec les Prosantorhinus de l'Astaracien d'Europe ; si ma d6termination g6n6ri- que s'av6rait, nous aurions l~ les tous derniers repr6- sentants du genre.

- - 605 - -
6 - - S o u s - f a m i l l e Elasmotheriinae
Les Elasmotheriinae sont un phylum d'origine asia- tique aboutissant au genre Elasmotherium du P16isto- c~ne d 'Europe orientale et d 'Asie occidentale. Ce genre extr~mement 6volu6 (par sa denture hyper- hypsodonte en particulier c 'est le plus 6volu6 de tous les Rhinocerotidae) semble avoir v6cu sporadique-
ment en Europe occidentale, mais les restes recueillis ont soit disparu (dent d 'Abbevil le d6truite par fait de guerre) soit sont mal localis6s (arri~re-cr~ne des <~ bords du Rhin >> de la collection Gall au Mus6um national d 'Histoire naturelle A Paris).
7 - - S o u s - f a m i l l e i n d ~ t e r m i n ~ e
Les deux pi~ces du gisement ruscinien ancien de Pont-de-Gaff (Cantal, France) ne sont pas d6termina- bles g6n6riquement, mais pourrai t &re rapproch6es
des genres Chilotherium ou Aceratherium. Dans l 'un ou l 'autre cas on aurait affaire au dernier repr6sentant europ6en du genre.
R 6 f 6 r e n c e s b i b l i o g r a p h i q u e s
(1) GUt~RIN C. - Docum. lab. GdoL Lyon, 79, 1-3, 1980, 1185 p. et Thdse ScL, Universitd Claude Bernard-Lyon/,81-31 (1981).
(2) GUI~RIN C. - Geobios, Lyon, 15,4, 1982, p. 593-598.
Manuscrit d6finitif regu le 14.06.1982





























