Embed Size (px)
Citation preview

IÉCANISMES DE COLONISATION PAR LA VÉGÉTATION D'UN INSELBERG GRANITIQUE EN GUYANE FRANçAISE
Corinne SARTHOU" et Catherine GRIMALDI**
La zone humide intertropicale est le domaine des sols ferr4llitiques recouverts par la forêt dense sempervirente. Sous les mêmes latitukles, les inselbergs représentent un milieu original avec des sols peu épais et une vagétation peu dense.
Sur les inselbergs, différentes formations végétales coexistent : (( savane- roche D, forêt basse et claire, et forêt dense à la base. En Gupane, elles ont fait l'objet d'études botaniques (de Granville, 1978 ; Sabatier et! Prévost, 1992) et d'inventaires floristiques, répertoriés dans la Banque de donhées (( Aublet )) de l'herbier du Centre ORSTOM de Cayenne (Hoff et al., 1989).
étudié par Sarthou (1992) sur l'inselberg granitique des Nouragues, représente une végétation pionnikre sur le rocher. Il était donc intéressant de connaître la nature des sols associés à cette végétation. De plus, l'analyse des eaux qui s'écoulent ou stagnent sur le granite permet de discuter les différents processus d'altération, d'échanges avec la végétation ou le sol participant au cycle des Cléments nutritifs sur l'inselberg. Une attention particulière a été apportée au rôle joué dans ce cycle par les Cyanophycées qui recouvrent le rocher.
L'écosystème (( savane-roche
1. - PRÉSENTATION DU MILIEU
La station des Nouragues est située dans le massif des Montagnes Balenfois, constitué de petites collines et d'un inselberg granitique, en bordure du bassin de l'Arataye, par 52"42' de longitude Ouest et 4"3' de latitude Nord.
1.1. Géologie
En Guyane française, le domaine géosynclinal est composé d'une série volcano-sédimentaire métamorphique appelée série de Paramaca (2,l à 2,O GA) (Gruau et al., 1985), recoupée par deux générations de granites, granites guyanais syntectoniques et granites carai%es post-tectoniques (1,9 GA) (Choubert, 1974 ;
* URA 1183, Laboratoire de Botanique tropicale, 12 rue Cuvier, 75005 Paris. ** Pédologie, INRA, mise à disposition de I'ORSTOM, B.P. 165, 97323 Cayenne Cedex.
?lB§TOm Fonds Documentaire Rev. Ecol. (Terre Vie), vol. 47, 1992.

Gruau et al., 1985 ; Teixeira et al., 1989). Ces derniers prédominent dans la moitié orientale du Massif Central de la Guyane et forment le plus souvent des reliefs en demi-orange. L'inselberg étudié constitue un affleurement tabulaire de ce granite caraïbe. I1 culmine à 411 mètres.
I1 s'agit d'un granite de type monzonitique, porphyro'ide, de couleur rosée, qui contient du quartz, de l'oligoclase, du microcline perthitique en proportions égales, et très peu de minéraux ferromagnésiens, biotite, muscovite et accessoire-
p ment hornblende (Depagne et Lelong, 1965).
1.
1.2. Climat
Le climat guyanais est du type tropical humide avec environ 8 mois de saison pluvieuse et une saison sèche diversement marquée. La hauteur des précipitations annuelles varie entre 2 500 et 4 O00 mm et l'intensité des averses est souvent forte. La température moyenne journalière oscille entre 25 et 27 "C.
Des données complémentaires ont été recueillies sur le lieu même de l'étude. Les précipitations mesurées sur deux années (1 988-1989) totalisent environ 2 300 et 3 200 mm, avec respectivement 75 et 110 jours présentant une pluviométrie comprise entre 10 et 100 mm. La température de l'air mesurée au-dessus de la surface du rocher se caractérise par des amplitudes journalières importantes (entre 20 et 38 "C et jusqu'à 45 "C en saison sèche). Dans la journée, la température de l'air 6 la surface du rocher peut atteindre 65 "C. L'humidité relative au-dessus du rocher varie entre 18 et 100 % (Sarthou, 1992).
1.3. Végétation
La végétation étudiée constitue un écosystème original, typique des inselbergs et connu sous le terme local de ¿( savane-roche D. C'est une couverture végétale discontinue qui peut être définie comme une mosaïque composée du rocher et de sa strate algale épilithe, de groupements herbacés et de groupements arbustifs (Fig. 1).
Les algues appartiennent presque toutes à la classe des Cyanophycées. Les espèces récoltées et déterminées (Sarthou et al., 1992) sont pour la majorité des formes. hétérocystées : Scytonema guyanense, S. multiramosum, StigonemaJEexuo- sum, S. hormoïdes, S. ocellatum. S. panniforme. Hapalosiphon fontinalis, H. luteolus ... Leur thalle filamenteux présente des hétérocystes, cellules spécialisées dans la fixation de l'azote atmosphérique (Lewin, 1962 ; Fogg et al., 1973).
Parmi les groupements herbacés, nous avons distingué des mares temporai- res, des prairies et des (( coussins D de Pitcairnia geyskesii (Broméliacée).
Les mares gravillonnaires temporaires (Fig. 2) sont caractérisées par une végétation subaquatique, très discontinue, composée de nombreuses espèces de Lentibulariacées (Utricularia hispida, U. nana, U. amethystina ...), de Xyridacées (Abolboda americana, Xyris savanensis), de Cypéracées (Rhynchospora barbuta, R. tenuis), d'Eriocaulacées (Paepalanthus lamarckii), de Rubiacées (Perama hirsuta)
Les prairies sont aussi temporairement inondées. Elles sont constituées d'un tapis de Graminées et de Cypéracées, dominé par Axonopus ramosus et Rhynchos- pora subdicephala.
-
u et de Poacées (Panicum sp.).
Figure 1. - Zone de savane-roche sur l'inselberg des Nouragues.
Les mares et les prairies se rencontrent dans les petites dépressions au sommet de l'inselberg.
Des (( coussins de Pitcairnia geyskesii caractérisent aussi le paysage des savanes-roche. Ils peuvent former des plages monospécifiques importantes, ou bien se diversifient, dans certaines conditions écologiques, définissant ainsi plusieurs sous-groupements (Sarthou, 1992).
Ils se développent aussi bien sur les pentes très fortes que sur les replats. Ces coussins présentent une accumulation d'humus et de Cyanophycées (Fig. 3 et 4) soit sur leurs bordures amont et latérales en position de pente, soit sur le pourtour des dépressions.
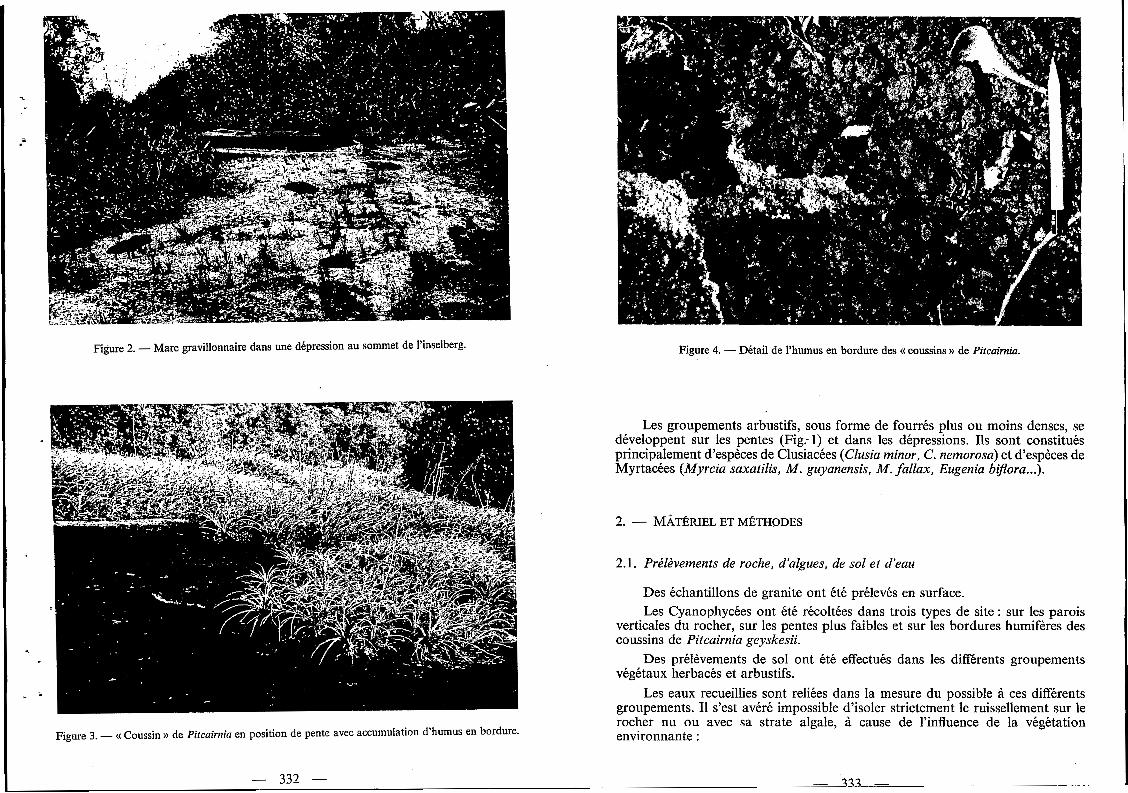
Figure 2. - Mare gravillonnaire dans une dépression au sommet de l’inselberg. Figure 4. - Détail de l’humus en bordure des (( coussins N de Pitcuirnia.
Figure 3. - (( Coussin )) de Pitcuirniu en position de pente avec accumulation d’humus en bordure.
Les groupements arbustifs, sous forme de fourrés plus ou moins denses, se développent sur les pentes (Figyl) et dans les dépressions. Ils sont constitués principalement d’espèces de Clusiacées (Clusia minor, C . nemorosa) et d’espèces de Myrtacées (Myrcia saxatilis, M. guyanensis, M. fallax, Eugenia bgora ...).
2. - MÄTÉRIEL ET MÉTHODES
2.1. Prtlèvements de roche, d’algues, de sol et d’eau
Des échantillons de granite ont été prélevés en surface. Les Cyanophycées ont été récoltées dans trois types de site : sur les parois
verticales du rocher, sur les pentes plus faibles et sur les bordures humifères des coussins de Pitcairnia geyskesii.
Des prélèvements de sol ont été effectués dans les différents groupements végétaux herbacés et arbustifs.
Les eaux recueillies sont reliées dans la mesure du possible à ces différents groupements. I1 s’est avéré impossible d’isoler strictement le ruissellement sur le rocher nu ou avec sa strate algale, à cause de l’influence de la végétation environnante :

- une partie des eaux collectées ont ruisselé sur la roche au cours des pluies et sont retenues dans les dépressions, où elles stagnent et sont donc soumises à I’évaporation ; ces dépressions correspondent aux mares et aux prairie? tempo- raires ; - les eaux traversant les coussins de Pitcairnia sont des eaux de ruisselle-
ment sur pente plutôt faible, à débit modéré, rapidement taries après la pluie ; elles
- d’autres eaux de ruissellement sont associées directement aux fourrés .* qu’elles traversent (Fig. 5). S’y ajoutent des eaux prélevées dans des rigoles,
véritables canalisations qui collectent l’eau après passage sous les fourrés de la falaise bordant le sommet tabulaire de l’inselberg. Cet écoulement persiste plusieurs jours après une précipitation et devient permanent en saison des pluies ; - les eaux des sources situées plus bas dans le paysage se sont sans doute
infiltrées à la faveur de fissures, et constituent un écoulement permanent en contact prolongé avec la roche.
4, j .- ont généralement circulé au préalable sous des fourrés ; c
Figure 5. - Fourré de CIusiu sur pente ; à l’aval, zone d’écoulement dépourvue de Cyanophycées.
2.2. Mesures et analyses chimiques i L‘analyse chimique totale des échantillons de granite a été complétée par la reconnaissance des principaux minéraux sur lames minces. La porosité d’échan-
- btillons de surface a été estimée par double pesée après dégazage et immersion dans l’eau (Costet et Sanglerat, 1983).
Avant analyse, les Cyanophycées ont été séparées des grains de sable restés agrégés lors du prélèvement. Elles sont ensuite attaquées par l’acide nitrique A
chaud, puis par l’eau oxygénée, avant le dosage des cations basiques totaux par absorption atomique. Leur teneur en carbone est analysée par la méthode Walkley et Black, leur teneur en azote par la méthode Kjedhal.
Les sols, dont on analyse la fraction inférieure à 2 mm, sont caractérisés au laboratoire par leur granulométrie, leurs pH,,, et pHKCI, leur teneur en matière organique (carbone total par la méthode Walkley et Black), leur teneur en azote (méthode Kjedhal) et en bases totales (attaque triacide puis analyse des ions en solution par absorption atomique). La capacité d’échange et les bases échangea- bles sont déterminées après saturation à l’acétate d’ammonium. La couleur des sols est définie à partir de la charte Munsell.
Sur les eaux filtrées à 0,45 pm sont réalisées les analyses suivantes: pH, conductivité, concentrations de Na+, K+, Caf +, Mg’ par absorption atomi- que, concentrations de CI-, NO,-, SO4’, PO4’ - et H4Si04 par colorimétrie, alcalinité par titration potentiométrique.
3. - RBSULTATS
3.1. Le granite
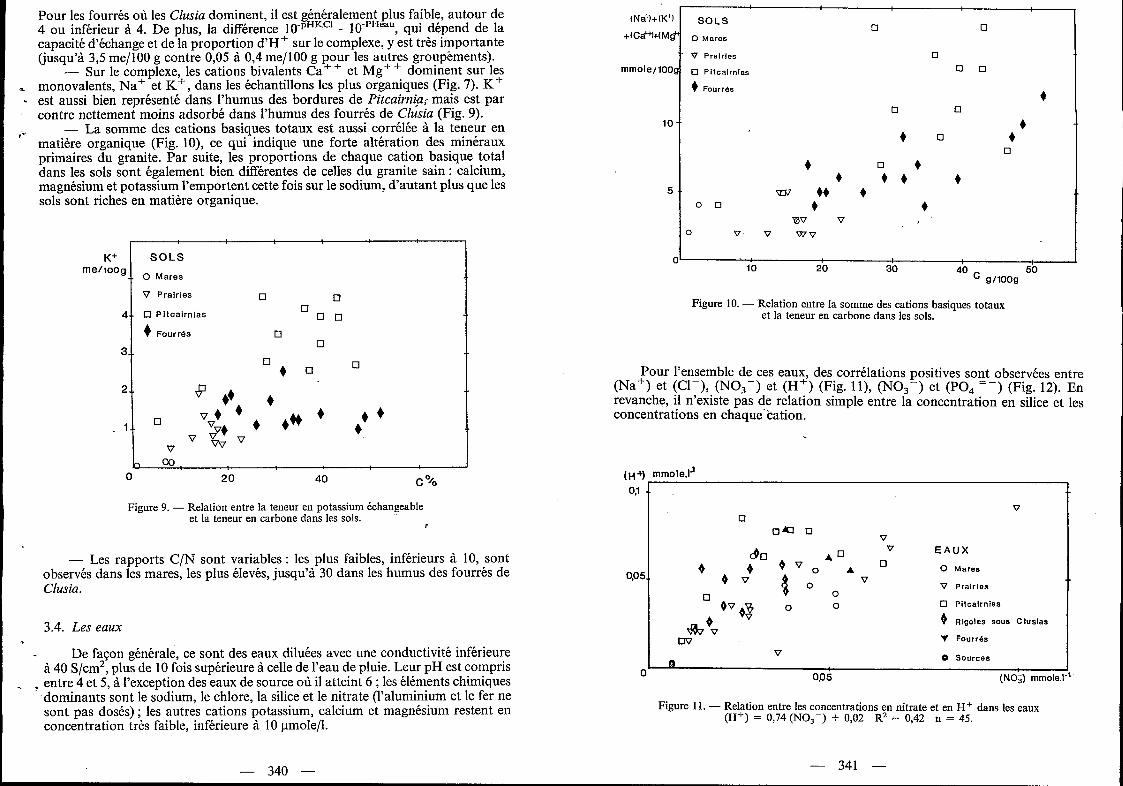
L‘analyse chimique totale de ce granite (Tab. I) met en évidence son caractère très siliceux (plus de 76 % de SO,) et alcalin (importance de K 2 0 et Na20). Ce granite renferme un peu plus de 30 % de quartz sous forme de cristaux grossiers, des plagioclases et feldspaths potassiques en proportion à peu près égale, et 2 YO de minéraux accessoires (biotite, amphibole verte, épidote et alanite) (Tab. II). 11 s’agit d’un leucogranite.
La mesure de l’absorption d’eau de deux échantillons, l’un peu altéré retenant 1,2g d’eau pour 100 g, l’ailtre plus altéré en retenant jusqu’à 3,2 g, révèle l’importante porosité superficielleLdu granite, évaluée respectivement pour les deux échantillons à 3,l YO et 7,8 ‘3’0 (porosité volumique).
TABLEAU I
Analyse chimique totale du granite.
Eléments majeurs % Eléments traces ppm
76,43 12,91 0,70 0,14 0,06 0,45 4,19 457 0,05 0.10 0,25
99,85
Ba Be Co Cr Cu Ga Nb Ni
71 Rb 20 1 2,7 sc 3,9
t 5 Sr 27 11 Th 14 15 V t 5 20 Y 12 16 Zn 61 13 Zr 55
- 114 - - 335 -
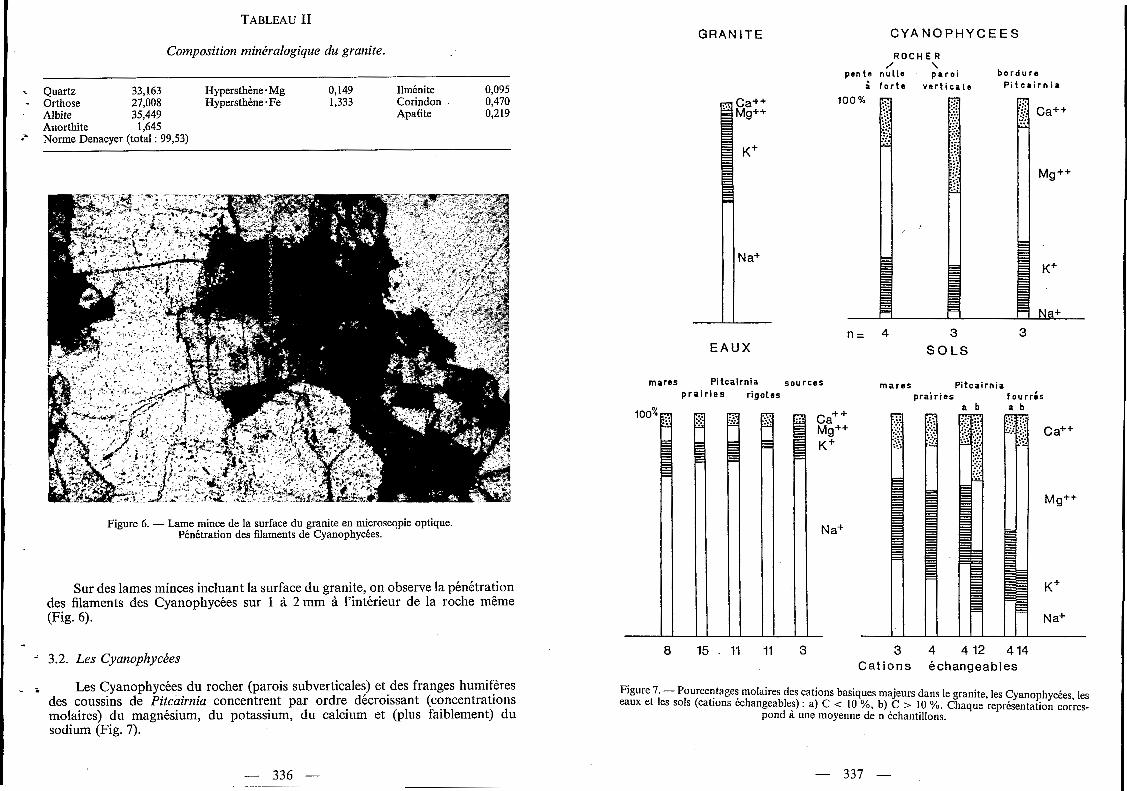
TABLEAU II
c
.-
Composition minéralogique du granite.
Quartz 33,163 Hypersthene. Mg 0,149 Ilmenite 0,095 Orthose 27,008 Hypersthène.Fe 1,333 Corindon . 0,470 Albite 35,449 Apafite 0,219 Anorthite 1,645 Norme Denaeyer (total : 99,53)
Figure 6. - Lame mince de la surface du granite en microscopie optique. Pénétration des filaments de Cyanophycées.
Sur des lames minces incluant la surface du granite, on observe la pénétration des filaments des Cyanophycées sur 1 à 2mm à l'intérieur de la roche même (Fig. 6).
' 3.2. Les Cyanophycées
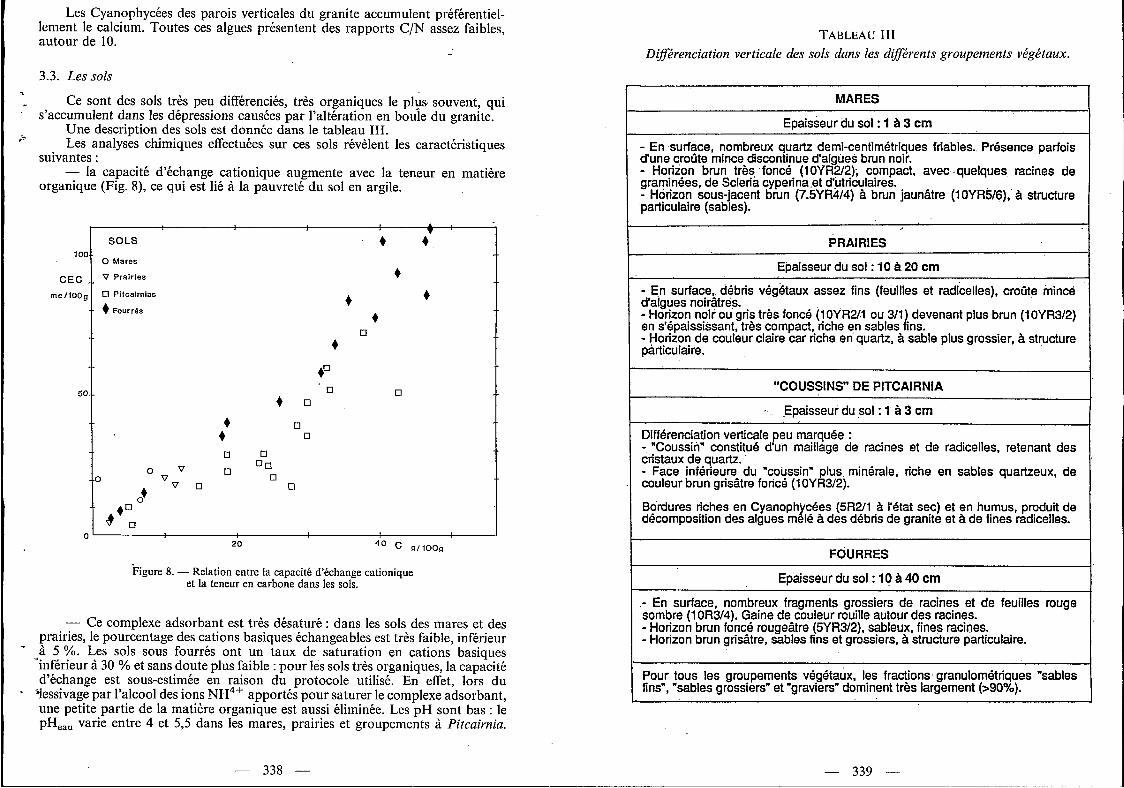
Les Cyanophycées du rocher (parois subverticales) et des franges humifères des coussins de Pitcairizia concentrent par ordre décroissant (concentrations molaires) du magnésium, du potassium, du calcium et (plus faiblement) du sodium (Fig. 7).
GRANITE
E A U X
mares Pitcairnia sources p r a i r i e s rigoles
8 15 . 11 11
C Y A N O P H Y C E E S
R O C H E R / \
pente n u l l e p a r o i b o r d u r e i f o r t e v e r t i c a l s P i t c a i r n i t
n = 4 3 3 S O L S
Pitca i rn i a m a r e s f o u rrós p r a i r i e s
a b a b Ca+ +
Mg++ K+
N a+
- 3 3 4 412 414
Cat ions echangeables
Ca++
Mg++
K+
N a+
Figure 7. - Pourcentages molaires des cations basiques majeurs dans le granite, les cyanophycées, les eaux et les sols (cations échangeables) : a) C < I O YO, b) C > I O YO. Chaque représentation corres-
pond à une moyenne de n échantillons.
- 336 - - 331 -
Les Cyanophycées des parois verticales du granite accumulent préférentiel- lement le calcium. Toutes ces algues présentent des rapports C/N assez faibles, autour de 10.
3.3. Les sols
s'accumulent dans les dépressions causées par l'altération en boule du granite.
suivantes :
organique (Fig. 8), ce qui est lié à la pauvreté du sol en argile.
1.
Ce sont des sols très peu différenciés, très organiques le plus souvent, qui
Une description des sols est donnée dans le tableau III. Les analyses chimiques effectuées sur ces sols révèlent les caractéristiques
- la capacité d'échange cationique augmente avec la teneur en matière
;7-
SOLS 100.-
O Mares
V Prairies CEG mel lOog 13 Pitcairnias
-- Fourres
V V v o
+ + o o
4
o o
o 0 0
O o
O ' 40 c glloog 20
Figure 8. - Relation entre la capacité d'échange cationique et la teneur en carbone dans les sols.
- Ce complexe adsorbant est très désaturé : dans les sols des mares et des prairies, le pourcentage des cations basiques échangeables est très faible, inférieur à 5 %. Les sols sous fourrés ont un taux de saturation en cations basiques
*inférieur à 30 % et sans doute plus faible : pour les sols très organiques, la capacité d'échange est sous-estimée en raison du protocole utilisé. En effet, lors du
*lessivage par l'alcool des ions NH4+ apportés pour saturer le complexe adsorbant, une petite partie de la matière organique est aussi éliminée. Les pH sont bas : le pH,,, varie entre 4 et 5'5 dans les mares, prairies et groupements à Pitcairnia.
-
-
TABLEAU III Différenciation verticale des sols dans les diffirents groupements végétaux.
MARES I Epaisseur du sol : 1 B 3 cm
- En surface, nombreux quartz demi-centimétriques friables. Présence parfois d'une croûte mince discontinue d'algues brun noir. - Horizon brun très foncé (lOYR2/2), compact. avec.quelques racines de graminées, de Scleria cyperina et &utriculaires. - Horizon sous-jacent brun (7.5YR4/4) à brun jaunâtre (lOYR5/6), à structure particulaire (sables).
PRAIRIES
Epaisseur du sol : 10 B 20 cm -~
- En surface, débris véghtaux assez fins (feuilles et radicelles), croûte mince d'algues noirâtres. - Horizon noir ou gris tres foncé (10YR2/1 ou 3/1 devenant plus brun (10YR3/2) en s'épaississant, tres compact, riche en sables lins. - Horizon de couleur claire car riche en quartz, à sable plus grossier, à structure particu taire.
"COUSSINS DE PlTCAlRNlA
Epaisseur du sol : 1 ir 3 cm
Différenciation verticale peu marquée : - "Coussin" constitué d'un maillage de racines et de radicelles, retenant des cristaux de quartz. - Face inférieure du "coussin" plus minérale, riche en sables quartzeux, de couleur brun grisâtre foncé (1 OYR3/2).
Bordures riches en Cyanophycées (5R2/1 à I'état sec) et en humus, produit de décomposition des algues mel6 à des débris de granite et à de fines radicelles.
FOURRES
.
Epaisseur du sol : 10 A 40 cm
- En surface, nombreux fragments grossiers de racines et de feuilles rouge sombre (1 OR3/4). Gaine de couleur rouille autour des racines. - Horizon brun foncé rougeâtre (5YR3/2), sableux, fines racines. - Horizon brun grisâtre, sables fins et grossiers, à structure particulaire.
Pour tous les groupements végétaux, les fractions granulométriques "sables fins", "sables grossiers" et "graviers" dominent très largement (>go%).
- 338 - - 339 -
Pour les fourrés où les Clusia dominent, il est généralement plus faible, autour de 4 ou inférieur à 4. De plus, la différence 10-pHKC’ - qui dépend de la capacité d’échange et de la proportion d’H’ sur le complexe, y est très importante Cjusqu’à 3,5 me/100 g contre 0,05 à 0,4 me/100 g pour les autres groupements).
et Mg’ + dominent sur les monovalents, Na+ et K”, dans les échantillons les plus organiques (Fig. 7). K’ est aussi bien représenté dans l’humus des bordures de Pitcairn@;- mais est par contre nettement moins adsorbé dans l’humus des fourrés de CIusia (Fig. 9). - La somme des cations basiques totaux est aussi corrélée à la teneur en
matière organique (Fig. lo), ce qui indique une forte altération des minéraux primaires du granite. Par suite, les proportions de chaque cation basique total dans les sols sont également bien différentes de celles du granite sain : calcium, magnésium et potassium l’emportent cette fois sur le sodium, d’autant plus que les sols sont riches en matière organique.
- Sur le complexe, les cations bivalents Ca+
S O L S
O Mares melioog
o o V Prairies
O 4 I O Pitcairnias O 0
Fourres
3 I -1
O 0
o o + o
S O L S o o O Mares
V Prairies o O Pitcairnias O 0
+ Fourres + O o + + o +
o + o + + + + + ++ + O 0 + +
@V v 3 v v w v
10 20 30 50 40 c g/1oog
Figure 10. - Relation entre la somme des cations basiques totaux et la teneur en carbone dans les sols.
Pour l’ensemble de ces eaux des corrélations positives sont observées entre (Na+) et (Cl-), (NO,-) et (H+’) (Fig. l l ) , (NO,-) et (PO, =-) (Fig. 12). En revanche, il n’existe pas de relation simple entre la concentration en silice et les concentrations en chaque.cation.
20 40 c OJO 0
Figure 9. - Relation entre la teneur en potassium échangeable et la teneur en carbone dans les sols.
?
- Les rapports C/N sont variables : les plus faibles, inférieurs à 10, sont observés dans les mares, les plus élevés, jusqu’à 30 dans les humus des fourrés de Clusia.
3.4. Les eaux
- De façon générale, ce sont des eaux diluées avec une conductivité inférieure à 40 S/cm2, plus de 10 fois supérieure à celle de l’eau de pluie. Leur pH est compris
E entre 4 et 5 , à l’exception des eaux de source où il atteint 6 ; les éléments chimiques dominants sont le sodium, le chlore, la silice et le nitrate (l’aluminium et le fer ne sont pas dosés) ; les autres cations potassium, calcium et magnésium restent en concentration très faible, inférieure à 10 pmole/l.
C
nmote.1”
t o
V o b o
I V E A U X
O Marea
V Prairias
o B O A O
V 6 6 O V O A
4 v I7 U
O O Pitcairnias
6 Rigoles sous Glusias
‘I Fourres du7 *v
uv V
1 O Sources I
Figure 11. - Relation entre les concentrations en nitrate et en H + dans les eaux (H+) = 0,74(NO,-) + 0,02 R2 = 0,42 n = 45.
- 340 - - 341 -
linOl*.lJ
EAUX V
Figure 12. - Relation entre les concentrations en phosphate et en nitrate dans les eaux (NO,-) = 17,9 (PO,’-) + 0,001 R2 = 0,52 n = 44.
(H,SiO,)mmole.l~’
EAUX o
OJ\ .
O
r
0.01 0.0 2 (K*) mmole.1-l O’
Figure 13. - Concentrations en silice et potassium dans les eaux.
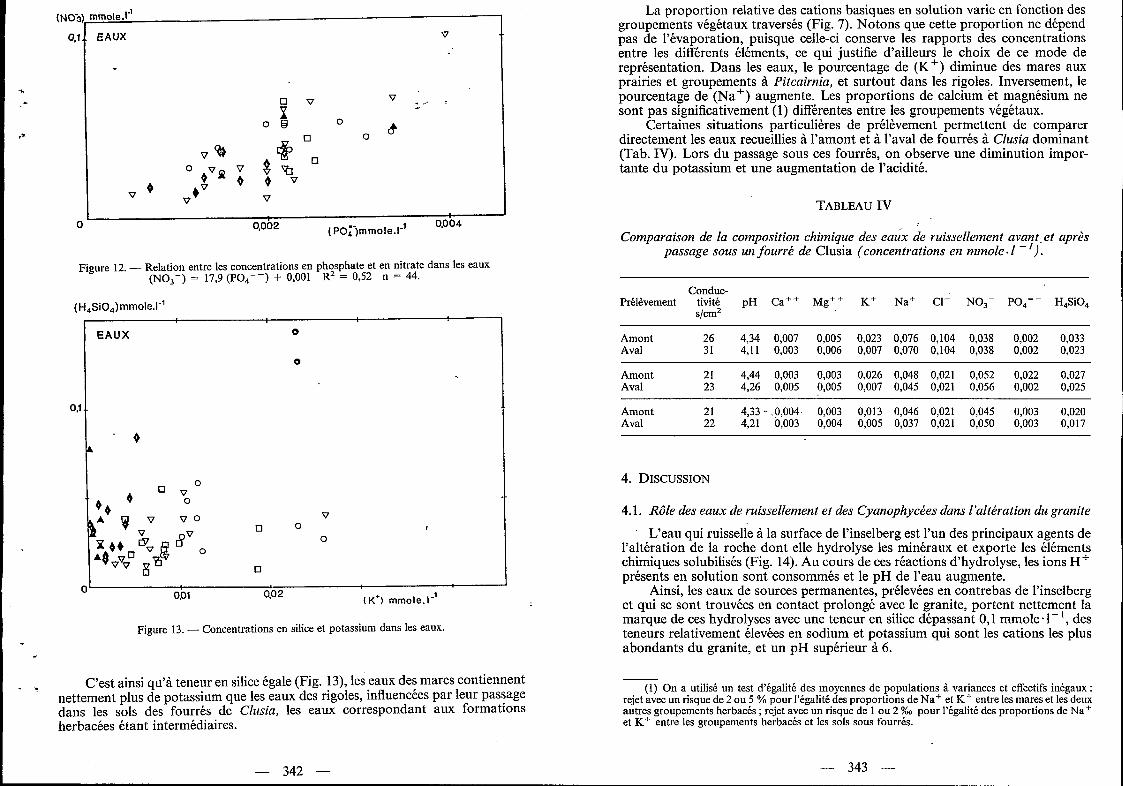
C’est ainsi qu’à teneur en silice égale (Fig. 13), les eaux des mares contiennent nettement plus de potassium que les eaux des rigoles, influencées par leur passage dans les sols des fourrés de Clusia, les eaux correspondant aux formations herbacées étant intermédiaires.
La proportion relative des cations basiques en solution varie en fonction des groupements végétaux traversés (Fig. 7). Notons que cette proportion ne dépend pas de l’évaporation, puisque celle-ci conserve les rapports des concentrations entre les différents éléments, ce qui justifie d’ailleurs le choix de ce mode de représentation. Dans les eaux, le pourcentage de (K+) diminue des mares aux prairies et groupements à Pitcairnia, et surtout dans les rigoles. Inversement, le pourcentage de (Na+) augmente. Les proportions de calcium et magnésium ne sont pas significativement (1) différentes entre les groupements végétaux.
Certaines situations particulières de prélèvement permettent de comparer directement les eaux recueillies à l’amont et à l’aval de fourrés à C h i a dominant (Tab. IV). Lors du passage sous ces fourrés, on observe une diminution impor- tante du potassium et une augmentation de l’acidité.
TABLEAU IV
Comparaison de la composition chimique des eau, de ruissellement avant, et après passage sous un fourrk de Clusia (concentrations en mmole. I - I ) .
Conduc-
s/cm2 Prélèvement tivité pH Ca++ Mg++ K’ Na+ CI- NO,- PO,=- H,SiO,
Amont 26 4,34 0,007 0,005 0,023 0,076 0,104 0,038 0,002 0,033 Aval 31 4,11 0,003 0,006 0,007 0,070 0,104 0,038 0,002 0,023
Amont 21 4,44 0,003 0,003 0,026 0,048 0,021 0,052 0,022 0,027 Aval 23 4,26 0,005 0,005 0,007 0,045 0,021 0,056 0,002 0,025
Amont 21 4,33 .. .0,004 0,003 0,013 0,046 0,021 0,045 0,003 0,020 Aval 22 4,21 0,003 0,004 0,005 0,037 0,021 0,050 0,003 0,017
4. DISCUSSION
4.1. RÓ1e des eaux de ruissellement et des Cyanophycées dans l’altération du granite
L‘eau qui ruisselle à la surface de l’inselberg est l’un des principaux agents de l’altération de la roche dont elle hydrolyse les minéraux et exporte les éléments chimiques solubilisés (Fig. 14). Au cours de ces réactions d’hydrolyse, les ions H+ présents en solution sont consommés et le pH de l’eau augmente.
Ainsi, les eaux de sources permanentes, prélevées en contrebas de l’inselberg et qui se sont trouvées en contact prolongé avec le granite, portent nettement la marque de ces hydrolyses avec une teneur en silice dépassant 0,l mmole.1- I , des teneurs relativement élevées en sodium et potassium qui sont les cations les plus abondants du granite, et un pH supérieur à 6.
(1) On a utilisé un test d’égalité des moyennes de populations à variances et effectifs inégaux : rejet avec un risque de 2 ou 5 % pour I’égalité des proportions de Na+ et K + entre les mares et les deux autres groupements herbacés ; rejet avec un risque de 1 ou 2 % pour l’égalité des proportions de Na+ et K+ entre les groupements herbacés et les sols sous fourrés.
- 342 - - 343 -

PLUIES CONDITIONS MICROCLIMATIQUES EXTREMES
quartz residue1
I I I
Nai, Ct
ì EAUX DE RUISSELLEMENT
I A I I I I
I I I I I Si, Na+,
I P, Ca++. I Mg+* ... I I
1 I
hydrolyses I
I I
I I I
atm. I I
I ' I
mati8re organique '
I I I I r I 1 I I
I I I I I 1 I
adaptations morphologiques el biologiques
Figure 14. - Schéma d'altération physique et biogéochimique du granite et cycle des éléments nutritifs majeurs.
Au contraire, dans les eaux superficielles recueillies dans la savane-roche pendant ou peu de temps après les pluies, la concentration en silice est plus faible, inférieure à 0,05 mmole-I-' et le pH, inférieur à 5. La dilution des eaux de ruissellement s'explique par leur faible temps de résidence au contact du granite et la forte intensité des averses. Leurs faibles DH confirment que la consommation : des H+ par les hydrolyses est moins rapide que les prodûctions d'acides. Ces productions sont d'origine diverse : acides organiques issus de la décomposition de la matière organique, absorption des cations par les Cyanophycées et par la végétation libérant en échange des H', réactions d'oxydation comme la nitrifi- cation et la phosphorylation mises en évidence dans les eaux superficielles. L'importance de cette dernière origine est illustrée par la corrélation présentée plus haut entre (H+) et (NO,-) (Fig. 1 l), corrélation faible du fait de l'existence des autres sources d'H+ et du devenir différent des deux ions.
. i
- 344 -
Dilution et acidité confèrent à ces eaux une agressivité importante puisqu'el- les restent sous-saturées par rapport aux minéraux du granite, et même par rapport au quartz, minéral le plus résistant qui s'altère aussi à leur contact. De même, les faibles concentrations en silice et en cations ne permettent pas la néoformation d'argile à partir de ces solutions.
C'est pourquoi l'altération du granite, dès lors qu'elle est amorcée, est rapide et complète: il n'existe pas d'arène granitique où les plagioclases les plus altérables (albite ou anorthite) seraient hydrolysés, tandis que les minéraux plus résistants (feldspath potassique, muscovite, quartz) seraient relativement conser- vés ; ici, seul le quartz s'accumule en graviers généralement friables, visibles dans les mares gravillonnaires, ou bien en sables grossiers et fins qui représentent la fraction granulométrique largement dominante dans tous les sols développés sur la savane-roche.
Les Cyanophycées participent aussi à l'altération de la roche et accélèrent sa dégradation (Fig. 14). En effet, ces organismes adhèrent énergiquement au granite et provoquent une désagrégation et une microdivision des minéraux par la pénétration de leur thalle filamenteux à l'intérieur de la roche (Fig. 6). Les conditions microclimatiques très contrastées, avec l'alternance d'humectation et de dessiccation (Fig. 14) préparent l'arrachement des minéraux adhérents au thalle. Les synthèses bibliographiques de Krumbein (1972), de Robert et Berthelin (1986), insistent également sur l'action à la fois mécanique et chimique des êtres vivants (bactéries, algues, champignons, lichens) dans les premiers stades d'alté- ration des roches. Plus particulièrement, l'action corrosive des Cyanophycées a été reconnue par Marathe et Chaudari (1975) et Danin et al. (1983) ...
Cependant, les Cyanophycées ne se développent pas sur certaines zones du granite situées à l'aval des fourrés de Clusia et des << coussins )) de Pitcairnia, en position de pente. Ces zones correspondent à l'écoulement des eaux les plus acides (pH entre 4,l et 4,3) parmi l'ensemble des prélèvements effectués. Cette observa- tion est à mettre en relation'avec divers travaux montrant l'effet négatif de l'acidité sur la croissance des Cyanophycées (Fogg, 1962) ; plus précisément, d'après Brock (1973), les Cyanophycées sont absentes dans les sols de pH inférieur à 5 et dans les eaux de pH 4 ou parfois même de pH inférieur à 4,8. Par contre, une strate algale constituée d'espèces de Chlorophycées se développe dans les eaux des mares, des prairies et de certains << coussins )) de Pitcaìrnia, oÙ le pH varie entre 4 et 5.
4.2. Importance des Cyanophycées comme producteurs primaires
Les Cyanophycées, procaryotes autotrophes, s'inscrivent parmi les végétaux qualifiés de reviviscents, qui prisentent une très grande adaptabilité et tolérance aux conditions les plus rudes. Leur pouvoir de colonisation d'un milieu a priorì hostile a, depuis longtemps, été observé par Treub (1888) sur les laves et les cendres du volcan Krakatau, par Fritsch (1922) sur des surfaces rocheuses arides, et par Flechter et Martin (1948) sur des sables désertiques.
L'installation pionnière des Cyanophycées sur le granite de l'inselberg est rendue possible par leur aptitude à métaboliser directement le carbone et aussi l'azote atmosphérique grâce à leur thalle hétérocysté. Leur analyse chimique montre qu'elles accumulent de manière sélective le calcium et le magnésium (ces deux cations sont peu abondants dans le granite) et le potassium. Rappelons que le magnésium est en effet un élément essentiel de la chlorophylle.
- 345 -

Les Cyanophycées jouent un rôle déterminant pour la colonisation végétale de l’inselberg en permettant un premier stockage d’éléments nutritifs, disponibles pour les autres végétaux lorsque les produits de leur décomposition sont entraînés dans les dépressions avec les débris de la roche. Vivantes, elles libèrent aussi par exsudation une fraction de l’azote fixé, sous forme d’ammoniac, d’amides, de peptides et d’acides aminés (Fogg, 1962, Dommergues et Mangenot, 1970) qui grâce aux réactions de nitrification produisent des ions $$rate, directement assimilables par les végétaux supérieurs (Fig. 14).
Enfin, les Cyanophycées forment des (( croûtes )) sur les bordures des coussins de Pitcairnia (Fig. 3 et 4) et des fourrés de Clusia. Elles constituent alors un apport important en matière organique et en éléments nutritifs, La formation et la cohésion de ces croûtes sont rendues possibles grâce aux gaines mucilagineuses qui entourent le trichome des Cyanophycées. L’importance de ces gaines a déjà été mentionnée dans le travail de Durrel et Shields (1961), sur une espèce de Microcoleus participant à la constitution de croûtes.
Fritsch (1922) et Booth (1941), ont reconnu depuis longtemps l’importance des Cyanophycées dans la formation des sols. Certains auteurs insistent sur leur pouvoir de limiter leur activité métabolique aux périodes oÙ l’eau est disponible. D’autres, observant les croûtes qu’elles forment sur des sables de milieu désertique soulignent leur effet protecteur contre l’érosion et leur rétention d’humidité.
4.3. Nature des sols sur l’inselberg
Le squelette sableux et la pauvreté en argile des sols (Tab. III) entraînent une faible capacité de rétention d‘eau, donc des déficits hydriques importants en période sèche pour la végétation ou la microfaune. En période pluvieuse, au contraire, des engorgements se produisent du fait de la faible profondeur des sols sur le soubassement granitique. La présence de gaines de couleur rouille, ferrugineuses, autour des racines de Clusia, est le signe d’une hydromorphie temporaire. Par ailleurs, l’épaisseur importante de la litière, la couleur noire ou gris foncé du sol, associée à des teneurs élevées en carbone, donnant des rapports C/N de 30 sous fourré, sont liées à une décomposition lente de la matière organique, due sans doute à ces conditions hydriques contrastées.
Ces contraintes hydriques sont à mettre en relation avec certains caractères adaptatifs de la végétation à la sécheresse : succulence des feuilles, épaisseur des cuticules, petite taille des feuilles (Sarthou, 1992).
Parallèlement à l’augmentation de la biomasse végétale des différents grou- pements, la profondeur des sols augmente, accroissant leur capacité de rétention en eau et permettant ainsi le développement d’une végétation plus abondante et ligneuse.
Au niveau chimique, la fraction minérale des sols est très pauvre, puisqu’elle est constituée essentiellement de quartz, sans argile dont les propriétés adsorban- tes permettraient la fixation de cations échangeables. Ce complexe adsorbant existe par contre sur la matière- organique. I1 est cependant très désaturé et acide en raison de la mauvaise décomposition de cette matière organique et du lessivage intense du sol en période pluvieuse.
4.4. Echanges avec la végétation
Du fait de la faible capacité de rétention d‘eau des sols, c’est au cours des épisodes pluvieux que les échanges chimiques entre les eaux de ruissellement et la
végétation sont importants. La composition chimique des eaux superficielles reflète moins les hydrolyses du granite que les échanges avec les Cyanophycées, les autres végétaux ou le complexe adsorbant du sol. Ceci est confirmé par l’absence de corrélation entre la concentration en silice et les concentrations des cations.
La rapidité du recyclage des Cléments nutritifs à partir des eaux de ruisselle- ment est particulièrement nette en ce qui concerne le potassium: lorsque la biomasse végétale augmente, des mares aux prairies et (( coussins D de Pitcairnia et surtout aux fourrés de Clusia, les eaux s’appauvrissent en potassium relative- ment aux autres cations (Fig. 7) ou relativement à la concentration en silice (Fig. 13) ; l’absorption du potassium dans les eaux circulant à travers des fourrés est mise en évidence dans le tableau IV. De même, la matière organique est relativement pauvre en potassium sous ces fourrés de Clusia (Fig. 9), ce qui indique un recyclage rapide du potassium.
II est cependant possible que différentes étapes se succèdent dans les échanges chimiques entre la végétation et les eaux de ruissellement : comme le montrent les expériences d’Abrahamsen et al. (1979), dans le cas’d’une simulation de pluies sur un rocher de granite recouvert de lichens, les cations sont absorbés surtout au début du ruissellement, en échange d’une libération d’H+. De plus, l’importance relative de l’absorption ou du lessivage des cations dépend certainement de l’intensité de la pluie.
D’autres auteurs (Isichei et al., 1990) montrent les variations du flux des Cléments minéraux sur un inselberg du Nigeria au cours d’un cycle annuel : une absorption importante du potassium et du nitrate par la végétation et les lichens du rocher a été observée en début de saison des pluies, et inversement une libération de ces ions par les lichens eut lieu en milieu de saison des pluies.
t CONCLUSION
Les Cyanophycées, grâce à leur propriété d’assimiler le carbone et l’azote atmosphériques, jouent un rôle essentiel dans la colonisation végétale de l’insel- berg étudié, en produisant les premiers apports organiques nécessaires aux végétaux supérieurs. Elles participent également à l’altération de la roche et accumulent les cations nutritifs. Par contre, les sols issus de l’altération du granite ne constituent pas un support très favorable pour la végétation : d’une part, peu épais et essentiellement sableux, ils entraînent des conditions hydriques très contrastées entre des dessiccations extrêmes ou des excès d’eau ; d’autre part, ils accumulent une matière organique qui se révèle très acide. Ce sont en fait les eaux de ruissellement qui jouent un rôle fondamental dans le cycle des déments nutritifs.
RÉSUMÉ
Sur l’inselberg granitique des Nouragues, en forêt tropicale guyanaise, l’écosystème (( savane-roche )) est défini comme une mosaïque végétale constituée de Cyanophycées recouvrant le rocher, et de plages herbacées et arbustives discontinues. Les sols développés dans ces plages de végétation montrent un squelette quartzeux prédominant, issu de l’altération du granite, associé à une
- 346 - - 347 -

matière organique d'autant plus abondante que la biomasse végétale augmente. Le complexe adsorbant est acide et désaturé malgré l'apport de cations par les eaux de ruissellement. Dans la mise en place du cycle des Cléments nutritifs, les Cyanophycées jouent un rôle essentiel en fkant l'azote atmosphérique. Elles produisent de plus une matière organique riche en calcium, magnésium et potassium. L'analyse des eaux superficielles, acides et diluées, permet d'expliquer l'altération intense du granite et de mettre en évidence l'attivi$,é-nitrificatrice, ainsi que le recyclage rapide du potassium, par la végétation.
SUMMARY
The soils and vegetation of a granitic inselberg of French Guiana are described. This (< rock-savanna )) ecosystem is a mosaic composed of Cyanophytes growing on the rock surface, and of herbaceous and shrubby patches. The soils in these vegetation patches mainly have a quartz-grained skeleton, which originates from the weathering of the granite. The organic matter content of the soils increases with plant biomass. The adsorbing complex is acid and desaturated in spite of cation enrichment by the runoff waters. The Cyanophytes are of primary importance in the establishment of the nutrient cycles on the rock surface, as they are able to fìx atmospheric nitrogen. They also produce organic matter with high calcium, magnesium and potassium contents. The analysis of the runoff waters, which are acid and dilute, helps to explain the intense weathering of the granite, and brings to the fore the existence of nitrification processes, and the rapid turnover of potassium.
REMERCIEMENTS
Nos remerciements s'adressent plus particulièrement à Louis Latouche du Laboratoire de Minéralogie du Museum National d'Histoire Naturelle, pour son aide dans la détermination et les analyses du granite, ainsi qu'à Denis Larpin du Laboratoire de Botanique tropicale de l'université Paris VI, sollicité pour quelques récoltes complémentaires d'échantillons sur le terrain.
REFERENCES i-
ABRAHAMSEN, G., STUANES, A. & BIOR, K. (1979). - Interaction between simulated rain and barren
BOOTH, W.E. (1941). -Algae as pioneers in plant succession and their importance in erosion control.
BROCK, T.D. (1973). - Lower pH limit for the existence of blue-green algae: evolutionary and
CHOUBERT, B. (1974). - Les Précambriens des Guyanes. Mémoires, B.R.G.M., 81. COSTET, J. & SANGLERAT, G. (1983). - Cours pratique de mécanique des sols. Dunod. DANIN, A., GERSON, R. & GARTY, J. (1983). -Weathering patterns on hard limestone and dolomite
by endolithic lichens and cyanobacteria : supporting evidence for eolian contribution to Terra Rossa.soi1. Soil Science, 136 : 213-217.
DEPAGNE, J. & LELONG, F. (1965). -Feuille de Haute Comté et notice explicative. Carte géologique à l'échelle du 1/100 000". Carte géologique détaillée de la France, Département de la Guyane, Paris.
rock surface. Water, Air, and Soil Pollution, 11 : 191-200.
Ecology, 22 : 38-46.
ecological implications. Science, 179 : 480-483.
DOMMERGUES, Y. & MANGENOT, F. (1970). - Ecologie microbienne du sol. Masson, Paris. DURREL, L.W. & SHIELDS, L.M. (1961). - Characteristics of soil algae relating to crust formation.
Trans. Amer. Microscop. Soc., 80 : 73-79. FLECHTER, J.E. & MARTIN, W.P. (1948). -Some effects of algae and molds in the rain-crust of desert
soils. Ecology, 29 : 95-100. FOGG, G.E. (1962). - Nitrogen Fixation. In Lewin R.A. (ed.). Physiology and Biochemistry of Algae.
Academic Press, New York and London. FOGG, G.E., STEWART, W.D.P., FAY, P. & WALSBY A.E. (1973). - The Blue-green Algae. Academic
Press, London & New York. FRITSCH, F.E. (1922). - The moisture relation of terrestrial algae, Some general observations and
experiments. Ann. Bot., 36 : 1-20. GRANVILLE, J.J. de (1978). - Contributions à IPtude de lafrore et de la végétation des inselbergs des
Tumuc Humac (Guyane française). In Recherches sur lafrore et la végétation guyanaise. Thèse de Doctorat d'Etat, USTL, Montpellier.
GRUAU, G., MARTIN, H., LEVEQUE, B., CAPDEVILA, R. & MAROT, A. (1985). - Rb-Sr and Sm-Nd geochronology of lower Proterozoic granite-greenstone terrains in French Guiana, South America. Precamb. Research, 30 : 63-80.
HOFF, M., CREMERS, G., FEUILLET, C. & GRANVILLE, J.J. de (1989). - La Banque de.données Aublet P de l'herbier du centre ORSTOM de Cayenne (CAY). Bull. Jard. Bor. Nat. Belg.,
ISICHEI, A.O., MORTON, A.J. & EKELEME, F. (1990). - Mineral nutrient flow from an inselberg in
KRUMBEIN, W.E. (1972). - Rôle des microorganismes dans la genèse, la diagenèse et la dégradation
LEWIN, R.A. (1962). -Physiology and Biochemistry of Algae. Academic Press, New York & London. MARATHE, K.V. and CHAUDHARI, P.R. (1975). -An example of algae as pioneers in the lithosphere
ROBERT, M. et BERTHELIN, N.J. (1986). - Role of biological and biochemical factors in soil mineral
59 : 171-178.
south-western Nigeria. J. Trop. Ecol.. 6 : 479-492.
des roches en place. Rev. Ecol. Biol. Sol, 9 : 283-319.
and their role in rock corrosion. J. Ecol., 63 : 65-69.
weathering : 453-496. I n Interactions of Soil MLerals with Natural Organics and Microbes, SSSA Spec. Pub., 17.
SABATIER, D. & PREVOST, F. (1992). - Variations du peuplement forestier à l'échelle stationnelle ; le cas de la station des Nouragues en Guyane française. Atelier MABIUNESCO, IUFRO. FAO, 12-16 mars 1990, Cayenne. -
SARTHOU, C. (1992). - Dynamique de la végétation pionnière sur un inselberg en Guyane française. Thèse de Doctorat de l'université de Paris VI.
SARTHOU, C., THEREZIEN, Y. & COUTE, A. (à paraître). - Cyanophycées de l'inselberg des Nouragues (Guyane française). Nova Hedwigia.
TEIXEIRA, W., TASSINARI, C.C.G., CORDANI, U.G. & KAWASHITA, K. (1989). - A review of the geochronology of the amazonian craton : tectonic implications. Precamb. Research, 42 :
TREUB, M. (1888). -Notice sur la nouvelle flore de Krakatau. Ann. Jard. Bot. Buitenzorg. 7 : 221-223. 213-227.
- 349 - - 348 -





























