Embed Size (px)
Citation preview

Eur. J . Biochem. 73, 607-615 (1977)
Methionyl-tRNA synthetase des embryons de ble : dissociation en sous-unites Philippe CHAZAL, Jean-Claude THOMES et Raymond JULIEN
Laboratoire de Biochimie, Unit6 d'Enseignement et de Recherches des Sciences, Universite de Limoges
(Requ le 17 mai/22 septembre, 1976)
Methionyl-tRNA Synthetase from Wheat Embryo : Dissociation into Subunits
Wheat-embryo methionyl-tRNA synthetase is a dimeric protein of PZ structure. When highly diluted, it loses the capacity to catalyze ATP-[32P]PPi exchange and to aminoacylate tRNAM": at low enzymatic concentrations the rates of formation of r2P]ATP and [14C]methionyl-tRNAMet are lower than those predicted by extrapolating the rates determined at higher enzyme concentrations. The difference between observed and expected rates becomes greater with decreasing enzyme concentration.
Filtration of purified, dilute enzyme preparations on Sephadex G-200 results in the separation of dimer and monomer fractions. The proportion of monomer present increases with increasing pre-incubation times before the assay and demonstrates an equilibrium between active dimers and inactive monomers. The above-mentioned loss of enzymatic activity is explained by the equilibrium being shifted towards the production of monomers. Data previously gathered for Escherichia coli prolyl-tRNA synthetase and for bovine-pancreatic tryptophanyl-tRNA synthetase, coupled with the present results, suggests that the dissociation of dimeric synthetases may be a general phenomenon in eukaryotes as well as in prokaryotes.
The number of sub-units and the dissociation constant were obtained at equilibrium, according to relations adapted to the case of oligomeric enzymes (KD = 13 nM at 25 "C and pH 7.5).
Rate constants were determined by kinetic studies of the attainment of equilibrium. The rate con- stant kl for monomolecular dissociation was determined to be 1.85 . s-' and k~ for the bimolecular association to be 0.145 . lo6 M-' s-' . The KD calculated from kl and kz was coherent with the experimentally determined value, at equilibrium.
The sub-unit interactions, which involve only a small quantity of energy (AGO = + 11 kcal mol-'; +45 kJ mol-') at 25 "C, depend on the ionic environment of the medium and the presence of substrates. Alkaline pH favors monomer production, while the presence of methionine, (Mg-ATP)2 - and tRNAMe' protect the synthetase from dissociation. 2-mercaptoethanol and dithio- erythritol prevent only slightly the loss of activity. Bovine serum albumin, however, protects the enzyme from dissociation under dilute conditions.
Dans les reactions catalyskes par de nombreuses enzymes, les vitesses initiales de formation des pro- duits sont proportionnelles aux concentrations en- zymatiques utilisees. Ce n'est pas le cas de la methionyl- tRNA synthetase des embryons de blk, proteine de poids molkculaire 165000 et de structure dimere
Abreviations. Unite ,4260, quantite de materiel contenue dans 1 ml d'une solution qui a une absorbance de 1 a 260 nm, pour un trajet optique de 1 cm.
Enzyme. Methionyl-tRNA synthetase ou L-mtthionine : tRNA ligase (EC 6.1.1.10).
P2 [l]. Cette derniere perd, aux faibles concentra- tions enzymatiques, son aptitude a catalyser l'echange ATP-[32P]PPi ainsi que l'aminoacylation des tRNAMe'. Dans ce present travail, nous montrons que l'equilibre entre dimeres actifs et monomeres inactifs, mis en evidence pour la prolyl-tRNA synthetase de E. coli B [2] et la tryptophanyl-tRNA synthetase du pancreas de boeuf [3], existe pour la methionyl-tRNA syn- thetase des embryons de ble et que le deplacement de cet equilibre vers la production de monomeres inactifs, aux faibles concentrations, est responsable de la perte d'activite observee.

608 Methionyl-tRNA synthetase: dissociation en sous-unitts
MATkRIEL ET MkTHODES
Enzyme
La methionyl-tRNA synthetase et les tRNAMe‘ des embryons de ble sont purifies comme precedem- ment decrit [l 1. Une modification est apportee cepen- dant au processus de purification de I’enzyme: un agent inhibiteur des proteases a skrine, le fluorure de phenyl-methyl-sulfonyle est introduit dans tous les tampons d’extraction et de purification. L’activite specifique, au plus haut degre de purete est, en amino- acylation: 117 nmol de tRNAMe‘ aminoacylees x mg proteine-’ x min-’ a 25 “C, et en &change ATP-[32P]- PPi : 621 nmol de [32P]ATP neosynthetisees x mg pro- teine-’ x min-’ a 25 “C.
Produits radioactifs
La [‘4C]methionine (activite specifique, 53 mCi/ mmol) provient du Commissariat a 1’Energie Atomi- que (Saclay) et le [32P]pyrophosphate de sodium anhydre (activite specifique, 25 a 40 mCi/mmol) du Radiochemical Centre (Amersham).
Rkactijs chimiques
L’albumine de serum bovin (fraction V), l’oval- bumine, l’aldolase, la catalase et 1’ATP proviennent de Boehringer, le tampon Tris de Sigma, le fluorure de phenyl-methyl-sulfonyle de Calbiochem, la Sepha- dex G-200 et les cdextranes bleusi) de Pharmacia et tous les autres reactifs chimiques sont de Merck.
Concentration protkique
La concentration proteique est estimee par la methode de Lowry et al. [4] et par le quotient d’ab- sorption 280/260 decrit par Warburg et al. [5].
Activite‘ enzymatique
L’attachement de la L-mtthionine au tRNAMe‘ est mesure comme precedemment dkcrit [I], excepte que le pH de test est 7,5 et que l’incubation est effec- tuke durant 5 min a 25 “C.
L’activite d’echange ATP-[32P]PPi de la methionyl- tRNA synthetase est determinee selon une methode inspiree de Lemoine et al. [6]. Le milieu reactionnel (0,l ml) contient du tampon cacodylate de sodium 100 mM a pH 7,5, MgClz 5 mM, ATP(Na) 2 mM, L-methionine 2 mM, 10 pg d’albumine de serum bovin, [32P]PPi 0,l mM (500000 coups. min-’, par essai) et des quantites variables de preparations enzymatiques. L’incubation s’effectue durant 5 min a 25 “C.
Filtration molkculaire
La filtration moleculaire sur SCphadex G-200 de la methionyl-tRNA synthetase des embryons de blC
est realisee selon une technique inspiree de Andrews [7]. Le volume mort de la colonne, VO, est mesure en eluant des ((dextranes blew)), et le calibrage est effectue a l’aide d’albumine de serum bovin, oval- bumine. catalase et aldolase.
R~SULTATS
Variation de l’activitt; en ,fonction de la concentration enzymatique
Les vitesses de formation de [‘4C]methionyl- tRNAMe‘ ou de [32P]ATP restent inferieures aux valeurs prevues lorsqu’on extrapole au domaine des faibles concentrations, les resultats obtenus pour les fortes concentrations (Fig. 1 A et 3 B).
Trois hypotheses peuvent @tre envisagees afin d’expliquer la perte d’activite observee.
(a) Une adsorption de I’enzyme sur les parois du recipient entourant le milieu de dilution, ce qui diminuerait, aux faibles concentrations, la quantite de methionyl-tRNA synthetase libre et fonctionnelle.
(b) Une inactivation de l’enzyme resultant d’une modification structurale. Dans ce cas, deux possibi- lit& peuvent Etre retenues: soit une isomerisation de la methionyl-tRNA synthetase en une forme inactive, soit une dissociation de l’enzyme dimere active en sous-unites non fonctionnelles.
(c) Enfin, une inactivation de I’enzyme i la suite d’alterations provoquees soit par la presence de metaux lourds apportks par les reactifs chimiques meme a un haut degre de purete, soit par proteolyse. Puisque l’adjonction d’acide ethylene-diamine tetra- cetique (sel de sodium) lo-’ M a toutes les solutions tamponnees utilisees (resultats non presentes) ne modifie en rien les courbes u = f(concentration en- zymatique), l’action des metaux lourds sur l’activite enzymatique peut @tre negligee. De plus, l’incubation a 25 “C pendant 30 min, en presence de fluorure de phknyl-methyl-sulfonyle, de preparations enzymati- ques hautement purifiees et concentrees, ne permet pas de mettre en evidence une perte d’activite enzyma- tique. L’hypothese d’une proteolyse doit donc Etre elle aussi exclue.
Itre hypothtse: adsorption
tion, I’equation de l’isotherme de Langmuir [8] : A l’equilibre, dans des conditions de non satura-
ou a: fraction de sites occupes au niveau de la paroi, [ P z ] ~ : concentration en enzyme dimi.re libre, K : con- stante d’equilibre de desorption, peut Ctre modifiee en

P. Chazal, J.-C. Thomes et R . Julien 609
5 10 Concentration enrymatique (nM)
Fi:. I . i:vo/ution di, iu vitessc rt~uctionnclle enfbnction dc la conccntra- ti011 en mitliionyl-tRNA synthirase. La solution enzymatique mere (1820 nM conserveei pH = 7.2) est dilueedans le tampon phosphate de potassium 10 mM, pH = 7 3 , fluorure de phenyl-methyl-sul- fonyle 0,l mM et glycerol 10 ”/,. Apres 30 min d’incubation a 25 T, I’enzyme est introduite d a m le milieu reactionnel ou elle subit une dilution supplementaire d’un facteur 5. (A): aminoacylation des tRNAM“; (O), vitesse reactionnelle mesuree en fonction de la concentration analytique [P,]o. Chaque point est la moyenne de trois determinations; (- -), droite prevue, par extrapolation, aux faibles concentrations, des resultats obtenus aux fortes concen- trations: elle permet, connaissant la vitesse de reaction et la concen- tration analytique en enzyme [P&, de determiner pour la suite des calculs, la concentration de dimkres actifs et fonctionnels: [P.], [P2], ou [P2IL selon I’hypothese envisagee. (B): Echange ATP-[32P]PPi; (O), vitesse mesuree; (-), droite obtenue d e la mime faGon que celle de (A)
tenant compte de l’equation de conservation de l’en- zyme :
dans laquelle
avec [Pz]F : concentration de dimeres correspondant A la quantite d’enzyme adsorbee, [Pz]o : concentra- tion analytique, S: surface de la paroi au contact de la solution, Q: facteur de proportionnalite; ce qui conduit a :
Les courbes des Fig.1A et 1 B fournissent les valeurs des concentrations en dimeres actifs, inter- pretees dans l’hypothese d’adsorption comme [Pz]L.
1 Ces valeurs appliquees a la relation ~ ~ ~ ~
/ I \ [PZIO - [PZII.
ne conduisent pas a une representation
lineaire et de ce fait, l’hypothese envisagee ne parait donc pas rendre compte du phenomene observe, quelle que soit d’ailleurs la nature chimique de la paroi au contact de la solution.
2ime hypothese: modification structurale
Isomkrisation. Dans le cas d’une isomerisation :
PZ I (active) PZ I1 (inactive);
avec :
[Pzlo = [PZII + [PZIII.
[PZ lo Le rapport ~- ‘pz I] est constant quel que soit le fac-
teur de dilution impose. Une telle modification, monomoleculaire dans les deux sens, ne peut pas rendre compte de la non proportionnalite entre la vitesse reactionnelle observee et la concentration enzymatique.
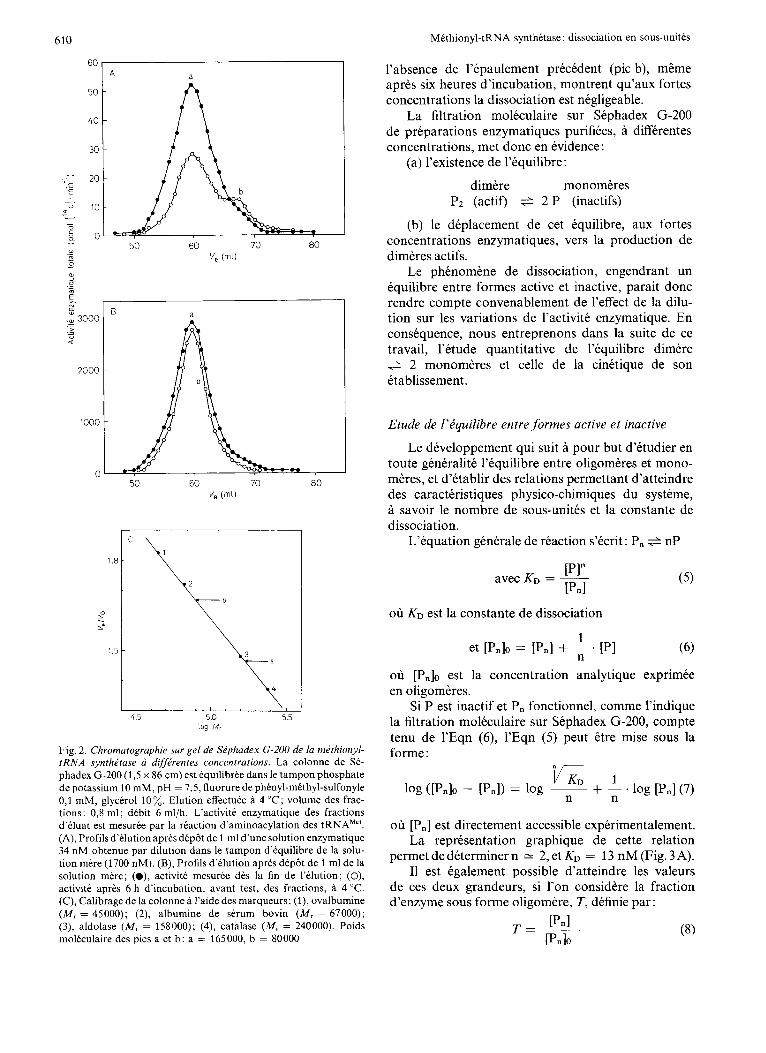
Dissociation. La filtration moleculaire sur Se- phadex G-200 de preparations enzymatiques purifiees et peu concentrees permet de separer une fraction ((dimere)) ( M , = 165000) et une fraction ((mono- mere)) ( M , = 80000) dont l’activite diminue et aug- mente respectivement en fonction de la duree d’incuba- tion avant test (Fig.2A). Dans le cas des solutions enzymatiques peu concentrees, dimeres et mono- meres sont en equilibre avant le dep6t sur la colonne de Sephadex G-200. Etant donnee leur difference de masse molaire, la filtration moleculaire permet leur separation. Lorsque l’activite enzymatique est mesuree dans les differentes fractions d’kluat, juste apres la fin de l’elution, le profil d’activite a la forme d’un pic symetrique a dont le maximum correspond au volume d’elution des dimeres. Apres six heures d’incubation avant le test des fractions d’eluat, les dimeres actifs se sont dissocies en monomeres inactifs, ce qui ex- plique la perte d’activite dans les fractions du pic a. Par contre, dans les fractions d’eluat contenant les monomeres inactifs, ces derniers se sont reassocies en dimeres actifs, de faCon a satisfaire a l’equilibre dimere e 2 monomeres; cette reassociation conduit a l’apparition d’un epaulement b dans le profil d’activite.
Dans le cas de solutions enzymatiques concen- trees (Fig. 2 B), l’existence et le maintien a leur niveau initial d’activite des fractions du pic a ainsi que
610 Methionyl-tRNA synthetase: dissociation en sous-unites
60 A a
B a
1.8
P 1- .
1 5
l’absence de l’kpaulement precedent (pic b), mtme apres six heures d’incubation, montrent qu’aux fortes concentrations la dissociation est negligeable.
La filtration moleculaire sur Sephadex G-200 de preparations enzymatiques purifiees, a differentes concentrations, met donc en evidence :
(a) l’existence de l’equilibre:
dimere monomeres Pz (actif) f 2 P (inactifs)
(b) le deplacement de cet equilibre, aux fortes concentrations enzymatiques, vers la production de dimeres actifs.
Le phenomene de dissociation, engendrant un kquilibre entre formes active et inactive, parait donc rendre compte convenablement de l’effect de la dilu- tion sur les variations de l’activite enzymatique. En consequence, nous entreprenons dans la suite de ce travail, l’etude quantitative de l’equilibre dimere e 2 monomeres et celle de la cinetique de son Ctablissement.
Etude de I’kquilibre entre formes active et inactive
Le developpement qui suit a pour but d’ktudier en toute generalit6 l’equilibre entre oligomeres et mono- meres, et d’etablir des relations permettant d’atteindre des caracteristiques physico-chimiques du systeme, a savoir le nombre de sous-unites et la constante de dissociation.
L’equation gentrale de reaction s’ecrit : P, e nP
Fig. 2. Chromatographie sur gel de Siphadex G-200 de la mithionyl- tRNA synthdtase a dffirentes concentrations. La colonne de Se- phadex G-200 (1,5 x 86 cm) est equilibree dans le tampon phosphate de potassium 10 mM, pH = 7,5, fluorure de phenyl-methyl-sulfonyle 0,l mM, glycerol 10%. Elution effectuee a 4 “C; volume des frac- tions: 0,8 ml; debit 6 ml/h. L’activitt enzymatique des fractions d’tluat est mesuree par la reaction d’aminoacylation des tRNAMe’. (A), Profils d’elution apres dep8t de 1 ml d’une solution enzymatique 34 nM obtenue par dilution dans le tampon d’equilibre de la solu- tion mere (1700 nM). (B), Profils d’elution apres dep8t de 1 ml de la solution mere; (O), activite mesuree des la fin de I’elution; (O), activite aprts 6 h d’incubation, avant test, des fractions, a 4 “C. (C), Calibrage de la colonne a l’aide des marqueurs: (I), ovalbumine ( M , = 45000); (2), albumine de serum bovin ( M , = 67000); (3), aldolase (M, = 158000); (4), catalase ( M , = 240000). Poids moleculaire des pics a et b: a = 165000, b = 80000
ou KD est la constante de dissociation 1
ou [P,]o est la concentration analytique exprimee en oligomeres.
Si P est inactif et P, fonctionnel, comme l’indique la filtration moleculaire sur Sephadex G-200, compte tenu de 1’Eqn (6), 1’Eqn (5 ) peut &tre mise sous la forme :
K 1 log ([pn]~ - [Pn]) = log ~ + ~ . log [P”] (7) n n
oh [P,] est directement accessible experimentalement. La representation graphique de cette relation
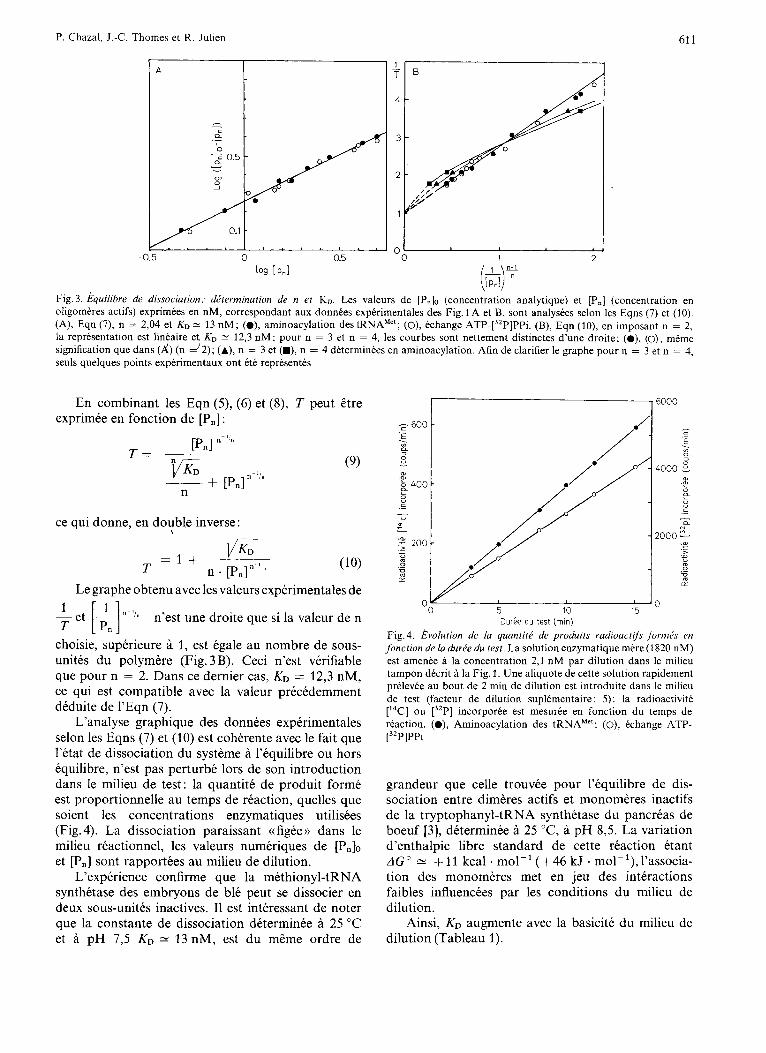
permet de determiner n E 2, et KD = 13 nM (Fig. 3A). I1 est egalement possible d’atteindre les valeurs
de ces deux grandeurs, si l’on considere la fraction d’enzyme sous forme oligomere, T, definie par:
P. Chazal, J.-C. Thomes et R. Julien 61 1
1 T a
4 -
- A
- -
Q
-0.5 0 0.5 0 1 2 (&IY log IP"1
Fig. 3. hquilibre de dissociation: dPterminution de n et KD. Les valeurs de [P& (concentration analytique) et [Pn] (concentration en oligomeres actifs) exprimees en nM, correspondant aux donnees experimentales des Fig. I A et B, sont analysees selon les Eqns (7) et (10). (A), Eqn (7), n = 2,04 et KO = 13 nM; (o), aminoacylation des tRNA'"; (O), echange ATP-[32P]PPi. (B), Eqn (lo), en imposant n = 2, la representation est lin'aire e KO 12,3 nM; pour n = 3 et n = 4, les courbes sont nettement distinctes d'une droite; (a), (o), mCme
seuls quelques points experimentaux ont ete representes signification que dans ( / J 5 = ) (n - 2); (A), n = 3 et (M), n = 4 determinees en aminoacylation. Afin de clarifier le graphe pour n = 3 et n = 4,
En combinant les Eqn (9, (6) et (8), T peut etre exprimee en fonction de [P,] :
ce qui donne, en double inverse: \
Le graphe obtenu avec les valeurs experimentales de
- et -~ n'est une droite que si la valeur de n
choisie, superieure a 1, est egale au nombre de sous- unites du polymere (Fig. 3 B). Ceci n'est verifiable que pour n = 2. Dans ce dernier cas, KD = 12,3 nM, ce qui est compatible avec la valeur precedemment deduite de 1'Eqn (7).
L'analyse graphique des donnees experimentales selon les Eqns (7) et (10) est coherente avec le fait que l'etat de dissociation du systeme a l'equilibre ou hors equilibre, n'est pas perturbe lors de son introduction dans le milieu de test: la quantite de produit forme est proportionnelle au temps de reaction, quelles que soient les concentrations enzymatiques utiliskes (Fig. 4). La dissociation paraissant ((figee)) dans le milieu reactionnel, les valeurs numeriques de [P,]o et [P,] sont rapportees au milieu de dilution.
L'experience confirme que la methionyl-tRNA synthetase des embryons de ble peut se dissocier en deux sous-unites inactives. I1 est interessant de noter que la constante de dissociation determinee a 25 "C et a pH 7,5 KD = 13 nM, est du mEme ordre de
[i"l
I I 6000
Durbe du test (rnin)
Fig.4. Evolution de iu quantite de produits radioactrfs JormGs en fonction de la durie du test. La solution enzymatique mere (1820 nM) est amenee a la concentration 2,l nM par dilution dans le milieu tampon decrit a la Fig. 1. Une aliquote de cette solution rapidement prelevee au bout de 2 min de dilution est introduite dans le milieu de test (facteur de dilution suplementaire: 5 ) : la radiodctivitt ['"C] ou [3zP] incorporee est mesuree en fonction du temps de reaction. (O), Aminoacylation des tRNAM"; (O), echange ATP- [3zP]PPi
grandeur que celle trouvte pour l'equilibre de dis- sociation entre dimeres actifs et monomeres inactifs de la tryptophanyl-tRNA synthetase du pancreas de boeuf [3], determinee a 25 "C, a pH 8,5. La variation d'enthalpie libre standard de cette reaction etant AGO = + 11 kcal . mol-' ( f46 kJ . mol-'), l'associa- tion des monomeres met en jeu des interactions faibles influencees par les conditions du milieu de dilution.
Ainsi, KD augmente avec la basicit6 du milieu de dilution (Tableau 1).

612 Methionyl-tRNA synthttase: dissociation en sous-unites
Tableau 1. Variations de K D en,fonction du p H Determinations faites a 25 ”C. Milieux tampons utilises : phosphate de potassium 10 mM, fluorure de phenyl-methyl-sulfonyle 0,l mM, glycerol 10% pour les pH = 6,8-7,5; Tris-HCI 10 mM, fluorure de phinyl-methyl-sulfonyle O, i mM, et glycerol 10% pour les pH = 8,2-9
nM
6 3 9,2- 9,9 7.5 12,3-13 8,2 21 -22,4 9 31,2-33,l
6 ‘%3 1 m
?o{ , , , , 1 0 1 2 3 4 Concentration en alburnine de serum de bceuf (rng/rnl)
Fig. 5. Influencr dr l‘cllhuminc dr ,sc;rum hovin rt du 2-mrrcapto- 6thanol sur la dissociation. La solution enzymatique est amenee a une concentration de 4,5 nM par dilution directe de la solution mere (1820 nM) dans le milieu tampon decrit a la Fig. 1 contenant des quantites croissantes d’albumine de serum bovin et de 2-mer- captoethanol. L’activite enzymatique est mesuree apres 30 min d’incubation avant test a 25 ”C. Concentration en 2-mercapto- ethanol: (O), 0 mM; (m), 5 m M ; (A), 10 mM
Des groupes acido-basiques jouent donc un r81e important dans l’association des monomeres.
Des extraits enzymatiques partiellement purifies, de concentration protkique globale Clevee ne presen- tent pas aux fortes dilutions, la perte d’activite pre- entre formes dimere et monomere peut ttre affecte tres purifiees. Pour ces dernieres solutions, lorsque l’enzyme est diluee dans le milieu tampon habituel, mais contenant initialement de l’albumine de serum bovin en concentration suffisamment forte, la dis- sociation est largement attenuee (Fig. 5) . L’effect protecteur de l’albumine de serum bovin ne peut, a priori, resulter d’interactions specifiques entre cette proteine et l’enzyme vegetale, conduisant a la forma- tion d’une nouvelle espece chimique et deplagant de ce fait l’equilibre etudie. Par contre, l’equilibre entre formes dimere et monomere peut ttre affecte par une variation de l’intensitk des interactions elec- trostatiques dans le milieu de dilution. Bien que l’influence exacte de l’albumine de serum bovin sur de telles interactions reste a demontrer, il est possible
d’envisager a titre d’hypothese, que l’effect protecteur de cette proteine s’exerce dans ce domaine d’inter- actions.
Iborra et al. [3] ont montrk que les groupes -SH sont impliques dans l’association des monomeres de la tryptophanyl-tRNA synthetase du pancreas de boeuf. Dans le cas de la methionyl-tRNA syn- thetase des embryons de ble, l’introduction de 2-mer- captoethanol en quantites croissantes n’a qu’un effet protecteur limite passant par un maximum peu pro- nonce a 5 mM (Fig.5). I1 en est de mtme avec le dithioerythritol. I1 est donc possible que les groupes - SH n’interviennent pas de fagon aussi preponderante que dans le cas precedent.
Lorsque le milieu de dilution contient du 2-mer- captoethanol ( 5 mM) et de l’albumine de serum bovin en concentration saturante (4 mg/ml) la dissociation devient negligeable pour des preparations enzyma- tiques tres purifiees. Dans ces conditions, la vitesse de formation des produits augmente lineairement avec la concentration enzymatique tant pour l’echange ATP-[32P]PPi que pour l’aminoacylation des tRNAMe’, et les droites obtenues sont alors superposables ri celles des Fig. 1 A et B.
Nous avons precedemment montrk que l’adsorp- tion de l’enzyme sur les parois du recipient entourant le milieu de dilution n’etait pas responsable de la perte d’activite observee. Le bon accord des donnees experimentales (Fig. 3) avec les Eqns (7) et (10) montre que le phenomene d’adsorption, s’il existe, doit ttre negligeable devant la dissociation.
Etablissement de l’kquilibve
Les etudes precedentes effectuees a l’equilibre de dissociation ont confirme la valeur du nombre de sous-unites et permis de determiner KD. Cette dernike peut egalement ttre obtenue en combinant kl et k2, respectivement dkfinies comme les constantes de vitesse des reactions de dissociation et d’association :
A , ki P 2 T 2 P a v e c K D =--.
k2
D’apres ce modele, la concentration en dimere actif [P2] evolue en fonction de la duree de dilution suivant la relation :
La combinaison des Eqns conduit ri :
+ 4 . k2 . [P2I2
12) et (6) avec n = 2,
P. Chazal, J.-C. Thomes et R. Julien 613
10 I I
I *
I I I 1 10 20 30
Durke de dilution (min)
Temps ( s ) 180 100
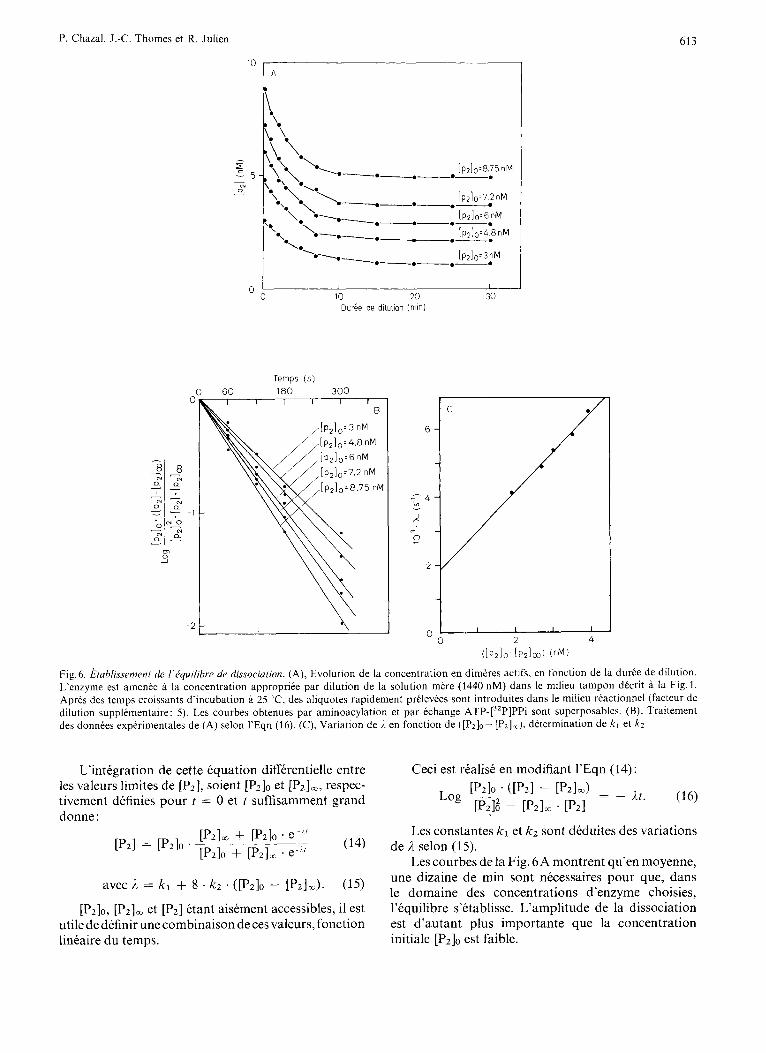
I 2 1 Fig.6. hzh/issernmt cle I’equilihre de dissociation. (A), Evolution de la concentration en dimeres actifs, en fonction de la duree de dilution. L’enzyme est amenee B la concentration appropriee par dilution de la solution mere (1440 nM) dans le milieu tampon decrit a la Fig. 1. Apres des temps croissants d’incubation B 25 ‘C, des aliquotes rapidement prtlevees sont introduites dans le milieu reactionnel ( fx teur de dilution supplementaire: 5). Les courbes obtenues par aminoacylation et par echange ATP-[32P]PPi sont superposables. (B), Traitement des donnees experimentales de (A) selon 1’Eqn (16). (C), Variation de i. en fonction de ([Pzl0- [P,],), determination de kl et k z
L’integration de cette equation differentielle entre les valeurs limites de [Pz], soient [Pz]o et [P2Im, respec- tivement definies pour t = 0 et t suffisamment grand donne :
avec A. = k l + 8 . kz . ([P2l0 - [PzIm). (15)
[P~]o, [PzIm et [Pz] etant aisement accessibles, il est utile dedefinir une combinaison de ces valeurs, fonction lineaire du temps.
Ceci est realise en modifiant 1’Eqn (14) :
Les constantes kl et kz sont deduites des variations de A. selon (1 5).
Les courbes de la Fig. 6A montrent qu’en moyenne, une dizaine de min sont necessaires pour que, dans le domaine des concentrations d’enzyme choisies, I’kquilibre s’etablisse. L’amplitude de la dissociation est d’autant plus importante que la concentration initiale [Pz]O est faible.

614 Methionyl-tRNA synthetase : dissociation en sous-unites
c 0 “ ._ c
m LL
1 10 20 30
Ourek de dilution (rnin) Fig. 7. Ejfet protecteur des suhstruts. it2fluence sur I’itahlissement de I’iquilihre. Le processus de dilution est effectue comme decrit pour la Fig.6A. Concentration enzymatique utilisee: 6 nM. Dilu- tion en absence des substrats: activite enzymatique mesuree par la reaction d’echange ATP-[32P]PPi (W) et par la reaction d’amino- acylation des tRNAM“ (0). Dilution en presence des substrats: (A), methionine 1 mM dans le milieu de dilution; (O), ATP 5 mM, MgClz 5 mM, activite mesuree par la reaction ATP-[32P]PPi; (O), tRNAMe‘ (30 unites Azso/rnl), activite mesuree par la reaction d’arninoacylation des tRNAM“
La representation graphique des donnees experi- mentales de la Fig.6A selon 1’Eqn (16) conduit a l’obtention d’un faisceau de droites passant par l’origine (Fig.6B), ce qui confirme la validite des equations precedentes.
L’experience montre que la variation de /z en fonction de ([P2l0 - [ P z ] ~ ) obeit a la loi linkaire prevue par 1’Eqn (15) (Fig.6C). Les valeurs de kl et kz sont respectivement de 1,85 . lop3 s-l et 0,145
K D , calculte selon 1’Eqn (11) est egale a 12,8 nM, a pH 7,5 et a 25 “C. Ce resultat est compatible avec celui obtenu grBce aux experiences realisees a l’equi- libre ( K D * 13 nM).
Nous avons precedemment montre (Fig. 4) que 1’Ctat de dissociation du systeme, m&me hors-equi- libre thermodynamique, est G bloque )) lors de son introduction dans le milieu de test oh sont presents tous les substrats. La question est donc posee de savoir si ce (( blocage)) resulte d’interactions entre substrats et enzyme. Dans des experiences preli- minaires, la methionine et (Mg-ATP)2- sont intro- duits separement dans les milieux de dilution, l’ac- tivite enzymatique etant mesurke par l’echange ATP- [32P]PPi: le processus cinttique de dissociation, dans les deux cas, n’a alors pas lieu (Fig.7). Le mEme phtnomene est observe en presence de tRNAM“, I’activitk enzymatique etant mesuree par la reaction d’aminoacylation ; le choix de cette derniere technique est motive par le fait que le tRNAMe‘, non indispensable a la reaction d’echange ATP-[32P]PPi, modifie nean-
1 0 + 6 M-1 s - l
moins la vitesse reactionnelle de cette derniere (re- sultats personnels non publies).
Ainsi, chaque substrat pris separement, parait stabiliser la forme dimere de l’enzyme, mais la pre- cision experimentale inherente aux techniques de mesure de l’activite et les conditions experimentales de dissociation utilisees, rendent impossible la distinc- tion entre un blocage reel ou un ralentissement considerable du processus de dissociation.
De plus, lorsque l’enzyme, a l’equilibre de dis- sociation, subit une nouvelle incubation en presence de l’un de ses ligands (methionine ou (ATP-Mg)’- ou tRNA) avant d’&tre introduite dans le milieu reactionnel, l’activite enzymatique avant dissociation est partiellement restaurke. Ce resultat preliminaire indiquerait que les ligands presents dans le milieu reactionnel provoquent un effect different sur l’equi- libre Ctudik de celui exerct par chacun d’entre eux ajoutk separement.
DISCUSSION
La methionyl-tRNA synthetase des embryons de ble perd, aux faibles concentrations enzymatiques, sa capacite a catalyser l’echange ATP-[32P]PPi ainsi que l’aminoacylation des tRNAMet. Cette perte d’ac- tivite resulte de la dissociation de I’enzyme dimere active en monomires inactifs, ces deux formes par- ticipant a l’equilibre :
P2 (actif) 2P (inactifs).
Ceci valide et complite les resultats obtenus dans une etude precedente [l], oh il a ett: montrk que l’enzyme native est dimere et possede une structure p2. Ainsi se trouve confirme pour une enzyme vegetale l’existence de I’equilibre entre formes dimeres actives et monomeres inactives precedemment mis en evidence pour la prolyl-tRNA synthetase de E. coli B [2] et la tryptophanyl-tRNA synthetase du pancreas de boeuf [3]. Des etudes en cours dans ce laboratoire (J. R. Carias, resultats non publies), sur la leucyl et l’arginyl-tRNA synthetase des embryons de ble semblent permettre la genkralisation de ce phenomene aux aminoacyl-tRNA synthetases dimeres de cet organisme.
Comme I’ont souligne Iborra et al. [3], l’activite enzymatique est like aux interactions entre proto- meres et resulterait d’une transconformation de la structure des sous-unites lors de leur association. Une telle modification structurale pourrait &tre sche- matiste de la fagon suivante:
0 - 0 - 6 monomeres inactifs dimere actif
Aucun ecart a la cinetique michaelienne n’ayant ete observe jusqu’a ce jour, pour la methionyl-tRNA

P. Chazal, J.-C. Thomes et R. Julien 615
synthetase des embryons de ble, la structure oligo- mtrique ne parait pas mettre en jeu d'interactions cooperatives.
La presence de tous les substrats dans le milieu de test, ((figen l'etat de dissociation du systeme (Fig. 4), alors que chacun separement, dans le milieu de dilu- tion, stabilise la structure dimerique (Fig. 7) : ceci sousentend la fixation des ligands sur les protomeres, favorisant l'association des sous-unitis. On pourrait envisager que les transconformations des dimeres et des monomeres provoquees par la presence simul- tanee des trois substrats interdisent I'association- dissociation et figent ainsi I'ttat d'tquilibre en main- tenant constantes les concentrations de dimkres et de monomeres.
Les tRNAM"' ne semblent pas indispensables au phenomene de blocage, puisque ce dernier est observe aussi bien dans la reaction d'echange ATP-[3ZP]PPi, ou la presence de tRNAM" n'est pas necessaire au deroulement de la reaction, que dans la reaction d'aminoacylation. Des resultats preliminaires non publies montrent neanmoins que le tRNA'" augmente l'activite d'echange, le systeme etant primitivement ( ( f i g t ~ : on peut alors se poser la question d'un eventuel effet du tRNAM" sur certaines constantes de vitesse de la reaction d'echange ATP-[32P]PPi, modifiant le taux d'incorporation de [3ZP] dans I'ATP.
Le mod& d'association-dissociation que nous avons propose pour la methionyl-tRNA synthetase des embryons de blk, s'integre dans les etudes theori-
ques menkes par Koshland et al. [9], Frieden [lo, 111, Klapper et af. [12] sur les interactions entre oligo- meres, monomeres et ligands. Le modele envisage permet d'etudier les modulations de l'activite enzyma- tique in vitro, en fonction de la concentration de I'enzyme et des substrats. I1 reste pourtant a savoir si la regulation de I'activite catalytique des aminoacyl- tRNA synthetases dimtres utilise, dans les conditions intracellulaires, les deplacements de l'equilibre de dissociation.
Nous remercions Mademoiselle M. Mourkout pour son ex- ceilente collaboration technique.
BIBLIOGRAPHIE 1.
2. 3.
4.
5 . 6.
7. 8. 9.
10. 11. 12.
P. Chazal, J .X . Thomes, and R. Julien, Laboratoire de Biochimie, U.E.R. 123 Rue Albert-Thomas, F-87060 Limoges-Cedex, France
Chazal, P., Thomes, J. C., Julien, R. (1975) FEBS Lett. 56,
Lee, M. L., Muench, K. H. (1969) J . Biol. Chem. 244,223-230. Iborra, F., Dorizzi, M., Labouesse, J. (1973) Eur. J . Biochem.
Lowry, 0. H., Rosebrough, N . J . , Farr, A. L., Randall, R. J.
Warburg, O., Christian, W. (1942) Biochem. J . 310, 384-421. Lemoine, F., Waller, J. P., Van Rapenbusch, R. (1968) Eur. J .
Andrews, P. (1964) Biochem. J . 91, 222-233. Langmuir, I . (1916) J . Am. Chem. Soc. 35, 2221 -2295. Koshland, D. R., Jr, Nemethy, G., Filmer, D. (1966) Biochem-
Frieden, C. (1967) J . Biol. Chem. 242, 4045-4052. Frieden, C. (1970) J . Biol. Chem. 245, 5788-5799. Klapper, M. H., Klotz, I. M. (1968) Biochemistry, 7, 223-231.
268 - 272.
39,275 - 282.
(1951) J. Biol. Chem. 193, 265-275.
Biochem. 4,213 - 221.
istry, 5, 365 - 385.
des Sciences, Universite de Limoges,