Embed Size (px)
Citation preview

Morphologie, structure et ontogenie des nectaires floraux de 19Ailanthus glandulosa
G ~ R A R D BORY ET DANIELLE CLAIR-MACZULAJTYS Universite' Paris 7, Laboratoire de Botanique, 2 Place Jussieu, 75005 Paris, France
R e p le 19 mai 1981
BORY, G., et D. CLAIR-MACZULAJTYS. 1982. Morphologie, structure et ontoghie des nectaires floraux de 1'Ailanthus glandulosa. Can. J. Bot. 60: 818-824.
Les fleurs miles et hermaphrodites de 1'Ailante possedent un disque hypogyne composC d'une partie basale et d'une partie apicale nettement sCparCes. Ces territoires histologiques distincts doivent &re considCrCs comme des nectaires. 11s diffkrent par leur position, leur forme, le contenu et 1'Ctat de diffkrenciation de leurs cellules. Les nectaires basaux sont dCpourvus de vascularisation, alors que l'apex du disque re~oi t des ClCments de phloeme. L'initiation des masses nectarifkres se fait aprks la mise en place des pikces florales et leur origine reste superficielle. De nombreux stomates, ronds, anomocytiques et a ostiole ouvert sont presents a la surface de ces formations glandulaires, surtout au sommet du disque. 11s sont beaucoup plus rares sur les nectaires basaux.
BORY, G. , and D. CLAIR-MACZULAJTYS. 1982. Morphologie, structure et ontoghie des nectaires floraux de 1'Ailanthus glandulosa. Can. J. Bot. 60: 818-824.
Staminate and hermaphroditic flowers show an hypogynous disc which is composed of two separate parts, an apical and a basal part. These histologic zones should be considered as nectaries. They differ by their position, their shape, and the content and state of differentiation of their cells. Basal nectaries are not vascularized but phloem bundles reach the tip of the disc. Initiation of nectariferous tissue occurs after that of the floral parts. The basal nectaries appear first but the number of cells still increases during the formation of the apical part of the disc. Both basal and apical nectaries have a superficial origin. Numerous anomocytic stomata are present especially on the surface of the apical nectaries; they are rounded and keep their stomata1 pores open. On basal nectaries, however, they occur much less frequently.
Introduction Les fleurs des Simarubactes sont dtcrites comme
prtsentant souvent un disque hypogyne plus ou moins dtveloppt . Toutefois, c e disque peut manquer complk- tement chez les Eurycoma. I1 peut &tre trks volumineux (Brucea), allongt en colonne (Quassia), Ccailleux et port t par un gynophore (Samandura), entier (Picrasma) ou fragment6 (Brucea). Dans la plupart des genres, il correspond ?i un simple bourrelet annulaire. Au con- traire,- chez les ~ i c r a m n i a , il apparait trks festonnt. Enfin, le disque hypogyne est gtntralement glabre (Ailanthus, Brucea . . .) ou exceptionnellement velu (Simaruba). Souvent signalts comme nectarifkres (Jadin 1901; Brizicky 1962), ces disques floraux n'ont jamais fait l'objet d't tudes cytologiques prtcises.
L'Ailante glanduleux, Ailanthus glandulosa Desf. encore appelt "Faux-vernis du Japon" est un arbre dioi'que dont les nombreuses petites fleurs verdltres sont grouptes e n panicules terminales. I1 existe des individus dont les fleurs sont uniquement staminkes et des indivi- dus ?i fleurs uniquement hermaphrodites. Pour les fleurs mlles cornrne pour les fleurs hermaphrodites l'ensemble des pi2ces florales est port t par un disque tpais , charnu, chlorophyllien, multilobt e t que l'on peut diviser en surface distinctes. A notre connaissance, c'est B Davies (1943) que l'on doit l a seule et brkve analyse morpholo- gique de cette partie de la fleur.
C e prtsent article a pour but d e prtciser les caracttris- tiques morphologiques, cytologiques et ontogtniques d e
ces glandes nectarifkres. I1 s'inskre dans l'ensemble des recherches mentes actuellement sur le systkme secreteur de 1'Ailante et vient complt ter des notions rtcemment acquises sur les nectaire; extra-floraux ou le trichome glanduleux (Bory et Clair-Maczulajtys 1978, 1980).
Materiel et methodes Les fleurs ont CtC rCcoltCes au mois de juin, sur de vieux
arbres du Jardin des plantes (Museum National d'Histoire naturelle de Paris).
Une partie du matCriel rCcoltC, destinC a l'examen au microscope Clectronique a balayage et a l'obtention de coupes semi-fines a CtC fixCe avec du glutaraldkhyde a 4,5% dans du tampon cacodylate de sodium (pH 6,8) pendant 4 h a la temperature ambiante, puis pendant 12h a 4°C. Les Cchantillons subissent ensuite des rin~ages successifs pendant plusieurs heures dans le tampon cacodylate. 11s sont ensuite post-fixCs dans du tCtroxyde d'osmium a 2% dans le tampon cacodylate pendant 1 h, rincCs puis dCshydratCs par des bains dlCthanol. Pour les observations en microscopic Clectronique a balayage, 1'Cthanol est substituC progressivement par l'acttate d'amyle. Les Cchantillons sont soumis au point critique selon la technique utilisant le COz. Le materiel est recouvert d'une fine pellicule d'or avant d'&tre examink au microscope JEOL JS M 35. Pour l'obtention des coupes semi-fines, les Cchantil- lons aprks dkshydratation dans des bains d'Cthanol croissant sont inclus dans une rCsine Cpoxy (Spurr 1969). Les sections sont rCalisCes au couteau de verre a l'ultramicrotome LKB Huxley et colorCes au bleu de toluidine boratC.
L'ontogenkse des glandes florales a CtC suivie a paratir de boutons floraux prealablement fixCs au mClange alcool-
0008-4026/82/0608 18-07$0 1 .OO/O 01982 National Research Council of Canada/Conseil national de recherches du Canada
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
WIN
DSO
R o
n 11
/18/
14Fo
r pe
rson
al u
se o
nly.

BORY ET CLAIR
formol-acide acCtique (AFA), puis dCshydratCs sous pression rCduite par des bains d'Cthano1 et inclus dans la paraffine 58°C. Les coupes sCriCes, (5 a 6 pm) sont colorCes par le vert de mCthyle-pyronine.
Un certain nombre de recherches histochimiques classiques dCcrites par Jensen (1962) ont CtC Cgalement rCalisCes. Ainsi les polysaccharides sont mis en Cvidence par la mCthode i I'acide pCriodique - riactif de Schiff, (APS), la lignine par la phloroglucine chlorhydrique, les lipides par le rouge Soudan 111, les tannins et tannoydes par le rCactif de Braemer et leper- chlorure de fer i 1 %.
Resultats Aspects morphologiques
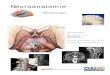
Toutes les parties du disque sur lequel sont insCrees les pikces florales laissent suinter un liquide limpide, visqueux et sucrC. L'ensemble de ce systkme nectari- gkne peut Ctre divisC en deux parties distinctes: nectaires apicaux et basaux (fig. 1).
Cinq volumineuses glandes chlorophylliennes occu- pent la base de la fleur. SituCes chacune en face des sCpales, elles sont sCparCes les unes des autres par des sillons profonds (fig. 1). Le point d'insertion des pCtales se trouve au milieu de la zone de jonction de ces masses nectarifkres. Entre les sCpales il existe frkquemment des glandes surnumQaires d'importance variable, mais tou- jours plus petites (fig. 2). A proximitC du point d'inser- tion des pCtales ou des Ctamines, le disque nectarifkre peut aussi se fractionner. Des petits massifs glandulaires bien individualisCs sont alors visibles aussi bien chez les fleurs staminCes que chez les fleurs hermaphrodites (fig. 1).
La partie apicale du disque, Cgalement chlorophyl- lienne est nettement isolCe des nectaires basaux: les deux cercles d7Ctamines sont insCrCs au niveau 'de cette jonction (fig. 1 et 3). Chez les fleurs hermaphrodites, le disque apical rCduit en raison mCme de la prCsence des samares forme une sorte de couronne plus ou moins lobCe (fig. 3,5). Chez les fleurs staminCes, il occupe tout le sommet de la fleur et prCsente un aspect partiellement fragment6 par des sillons longitudinaux (fig. 4).
La rkpartition des stomates, ronds, anomocytiques et a ostiole ouvert (fig. 6) est trks inkgale suivant les deux types de nectaires. 11s sont rarissimes sur les nectaires basaux (en moyenne trois ou quatre par masse nectari- fkre). Par contre sur le disque apical leur frCquence est bien plus ClevCe (100 a 120 par disque entier). 11s sont en principe rCpartis trks rkgulikrement a la surface de ce disque, toutefois chez les fleurs hermaphrodites, ils sont beaucoup plus nombreux dans 1'Cchancrure mCnagCe dans le disque par la presence des samares (fig. 3).
I1 faut encore souligner que toutes les parties du nectaire floral sont glabres. Les poils glanduleux frequents en particulier a la base des carpelles ou sur les sCpales manquent complktement au niveau des nec-
taires. Des poils hyalins occupent la base des Ctamines et des pCtales. 11s manquent sur les nectaires (fig. 1 a 4).
Histologie et cytologie des nectaires fonctionnels Chaque masse nectarifkre est constituCe par un tissu
homogkne bien distinct du reste des tissus rCceptacu- laires sous-jacents (fig. 7 et 8). En plus des caractkristi- ques gCnCrales des cellules sCcrCtrices de nectar, ces zones montrent une forte reaction a 1'APS qui permet Cgalement de les distinguer du reste des tissus de la fleur (fig. 8). Ce tissu nectarifkre trks Cpais dans le nectaire basal (15 a 20 couches cellulaires) est beaucoup plus mince au niveau du disque apical (4 B 10 couches cellulaires). L'opposition entre les nectaires de la base de la fleur et le sommet du disque est surtout Cvidente chez les fleurs staminCes. D'abondants prCcipitCs chro- mophiles granuleux ou en plaque sont observables a l'apex du disque (fig. 7 et 9) tandis que les nectaires basaux en sont dCpourvus. Ceux-~i , identifiables par ailleurs a des tannins, apparaissent tout d'abord en soulignant les contours de la vacuole qu'ils remplissent progressivement (fig. 10). Dans les deux types de fleurs, les nectaires basaux sont toujours dCpourvus de vacula- risation, l'irrigation des sCpales et des pitales voisins passant 21 proximitC. Par contre, l'apex du disque re~o i t une vascularisation constituCe par des ClCments de phlokme (fig. 9). Elle remonte parfois trks haut jusqu'au niveau des assises sous-Cpidermiques. L'ensemble des nectaires est recouvert par un Cpidenne dont les cellules renferment Cgalement des pr6cipitC.s assimilables ti des tannins (fig. 10 et 1 1). Ace niveau des stomates de petite taille (30 p,m) et dCpourvus de chambre sous-stomatique sont visibles (fig. 11). Les cellules du tissu nectarifkre montrent souventunsystkme vacuolaireparticulikrement dCveloppC, constituC par une vacuole centrale et de nombreuses petites vCsicules plus ou moins coalescentes (fig. 10 et 12). Certaines cellules Cparses dans le tissu nectarifkre contiennent de volumineuses mlcles d'oxa- late de calcium. Le noyau des cellules nectarifkres, toujours trks important, montre un nuclCole volumineux (fig. 12). Le cytoplasme des cellules du nectaire basal ou apical des fleurs staminCes renferme d'abondantes gra- nulations osmiophiles Cgalement colorables par le rouge Soudan 111. Dans les cellules des nectaires apicaux, la phase fonctionnelle se marque par l'apparition de nombreux amyloplastes (fig. 13).
Ontogenbse des nectaires$oraux La mise en place des diffkrentes masses nectarifkres
est retardCe par rapport B celle des pikces florales qu'il s'agisse des fleurs mlles ou hermaphrodites, des nectaires apicaux ou basaux. L'ontogCnkse de ces deux types de nectaires suit le mCme schCma. Les cellules Cpidermiques et sous-Cpidermiques situCes entre les initiums des pikces florales (fig. 14) conservent plus
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
WIN
DSO
R o
n 11
/18/
14Fo
r pe
rson
al u
se o
nly.

820 CAN. J . BOT. VOL. 60, 1982
longtemps leurs caractkres mCristCmatiques. Lorsque les pCtales, les Ctamines ou les carpelles commencent leur diffkrenciation basipkte, les six 2 sept cellules Cpidermiques constituant l'initium glandulaire se divi- sent d'une mani6re anticline (fig. 15). La formation des nectaires basaux dCbute avant celle des nectaires api- caux. L'assise sous-Cpidermique subit les mCmes types de divisions que l'assise Cpidermique (fig. 15) et tr6s rapidement elle est le sikge de recloisonnements pCri- clines et obliques (fig. 16). Progressivement, il se met en place un "massif prC-glandulaire" (fig. 17) dont l'tlargissement va &tre assurC par le jeux de divisions anticlines supeficielles et le soulkvement par des divisions pCriclines et obliques en profondeur. Ceci va concourir a Ccarter les diffkrentes pikces florales les unes des autres. Pour les nectaires de la base de la fleur, les activitCs de mCrkse se poursuivent pendant le stade fonctionnel: ainsi, des figures de division sont encore visibles au sein du tissu nectarifkre alors que la fleur est Cpanouie et que les nectaires apicaux prCsentent dCji des prCcipitCs chromophiles denses.
Les ClCments de vascularisation des disques apicaux apparaissent lorsque le massif prC-glandulaire est dCja bien individualis6 (fig. 18). Ces premiers ClCments de procambium naissent en position mCdiane sous chacun des cinq lobes du disque apical. Ce procambium va
prolifkrer et se raccorder au systkme vasculaire de la fleur.
Chez les fleurs hermaphrodites, la structure des disques nectarifkres se maintient jusqu'a la maturation complkte des samares. De plus, on assiste ?I un proces- sus de lignification centripkte des tissus nectarifkres (fig. 19). Cette lignification est toutefois retardee au niveau des sillons sCparant les masses nectarifkres (fig. 19). Les assises profondes montrent d'abondantes m2cles d'oxa- late de calcium. Dans les nectaires floraux vieillissant, la prCsence de tannins est rCvClCe par le rCactif de Braemer. Jusqu'a la maturation complkte des samares, soit 5 mois aprks 1'Cpanouissement du bouton floral, la structure des cellules nectarif6res parait assez bien consemCe, le noyau est encore parfaitement visible.
Discussion Les nectaires floraux de 1'Ailanthus glandulosa ap-
partiennent aux "types discoi'des" dCfinis par Fahn (1953). En effet, ils apparaissent ici comme des dif- fkrenciations locales du disque hypogyne des Simaruba- cCes. Constituts d'un massif plus ou moins homogkne de cellules, ils s'opposent en cela, chez cette plante, aux nectaires extra-floraux dont la structure tissulaire com- plexe et 1'Cvolution schizo-lysigkne a CtC rCcemment CtudiCe (Bory et Clair-Maczulajtys 1978). Deux types
ABREVIATIONS: a , grains d'amidon; c, carpelles ou emplacement des carpelles; cs, cellules stomatiques; cu, cuticule; da, disque apical; e, tpiderme; et, Ctamines ou emplacement des Ctamines; go, granulations osmiophiles intracytoplasmiques; gs, glandes surnumtraires; i , initium glandulaire; I, zone de lignification; rn, massif prt-glandulaire; n, noyau; nb, nectaire basal; nu, nucltole; o, ostiole; p , pttales ou emplacement des pttales; pf, ptdoncule floral; pg, prtcipitts vacuolaires globuleux; ph, phlotme; pp, prtcipitts vacuolaires en plaque; pr, procambium; s , stpales ou emplacement des sCpales; sc, canaux sCcrCteurs; si, sillon stparant les masses nectarifiires; st, stomates; t, trichome; v, vascularisation des piiices florales; va, vacuole; ve, vtsicules.
FIG. 1-6. Observations en microscopie tlectronique h balayage des nectaires floraux de 1'Ailanthus glandulosa. (Les difftrentes pitces florales ont t t t Gttes.) Fig. 1. Le disque floral peut Ctre divist en deux ensembles glandulaires: le nectaire basal et le disque apical; des glandes surnumtraires sont visibles. X40. Fig. 2. Nectaire basal surnumtraire chez une fleur hermaphrodite. x40. Fig. 3. Aspect du disque apical chez une fleur hermaphrodite. x40. Fig. 4. Chez les fleurs stamintes, le disque apical montre cinq lobes stparks par de profonds sillons. x40. Fig. 5. Formation du disque nectarifere apical dans un bouton floral hermaphrodite. X 100. Fig. 6. Les stomates des nectaires floraux sont arrondis et prtsentent un ostiole ouvert. X 1500.
FIG. 7-13. Aspects histologiques et cytologiques des nectaires floraux de 1'Ailanthus glandulosa. Fig. 7. Coupe longitudinale dans la fleur staminte (AFA, bleu de mCthyliine). X35. Fig. 8. Section transversale au niveau des nectaires basaux d'une fleur staminte (AFA, rCaction h l'APS). X40. Fig. 9. Coupe semi-fine dans le disque apical d'une fleur staminte. Des Cltments de phloiime ptnttrent h la base de la zone nectarifere. Prtsence de prtcipitts en plaque assimilables h des tannins. X250. Fig. 10. Les prCcipitts osmiophiles (tannins) soulignent les contours de la vacuole qu'ils remplissent progressivement. x 1000. Fig. 11. Stomate de petite taille dtpourvu de chambre sous-stomatique, au niveau du disque apical (fleur staminte). x700. Fig. 12. Aspect des cellules nectarifiires au dtbut de la phase fonctionnelle. X 1000. Fig. 13. Nombreux amyloplastes dans les cellules des nectaires apicaux en phase fonctionnelle. x 1000.
FIG. 14-19. Ontoghie des nectaires floraux de 1'Ailanthus glandulosa. (Fig. 14 21 18: fixation AFA, coloration vert de mCthyle-pyronine. Fig. 19: rtaction h la phloroglucine chlorhydrique.) Fig. 14. Section transversale h la base du bouton floral montrant la zone d'initiation du nectaire. X 150. Fig. 15. De nombreuses divisions anticlines contribuent h tlargir l'initium glandulaire (section longitudinale). x 150. Fig. 16. Section longitudinale dans le jeune bouton floral montrant au niveau de l'initium glandulaire des divisions ptriclines et obliques qui contribuent h sa surrection. x 150. Fig. 17. Dans le bouton floral, le dtveloppement des massifs prt-glandulaires contribue h tcarter les difftrentes piiices florales les une des autres (section transversale). x70. Fig. 18. Coupe longitudinale dans un jeune bouton floral montrant, h la base du disque apical, la mise en place d'tltments de procambium. X 150. Fig. 19. Aspect du nectaire basal d'une fleur hermaphrodite deux mois apriis la floraison (section transversale). x 80.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
WIN
DSO
R o
n 11
/18/
14Fo
r pe
rson
al u
se o
nly.

BORY ET CLP
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
WIN
DSO
R o
n 11
/18/
14Fo
r pe
rson
al u
se o
nly.

822 CAN. J. BOT. VOL. 60, 1982
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
WIN
DSO
R o
n 11
/18/
14Fo
r pe
rson
al u
se o
nly.

BORY ET CLAIR-I
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
WIN
DSO
R o
n 11
/18/
14Fo
r pe
rson
al u
se o
nly.

824 CAN. J. BOT. VOL. 60, 1982
de nectaires floraux peuvent Ctre distinguks (nectaires situCs ii la base de la fleur et apex du disque floral). Les nectaires basaux ne sont pas vascularisCs, alors que l'apex du disque r e ~ o i t une irrigation constituCe princi- palement de phlokme. Nair et Joshi (1958) relevent la prCsence de nombreuses traces de faisceaux conducteurs dans le disque d'Ailanthus malabarica (D.C.) et d'Ai- lanthus glandulosa Desf. Mais ces auteurs ne dCtaillent pas la structure du disque et ne font pas mention des diffkrentes parties irrigukes. 11s soulignent aussi l'absence de vascularisation du disque nectarifkre chez d'autre Ailantes comme A. grandis Prain et A. excelsa Roxb. Enfin, ils assimilent les nectaires floraux du Picrasma quassioides L. ii un verticille modifiC d'Cta- mines. Enfin, plusieurs auteurs (Nair et Joshi 1958; Brizicky 1962) pensent que dans le genre Ailanthus, le disque doit Ctre de nature rkceptaculaire. Les nectaires floraux posskdent une origine superficielle et sont initiCs aprks la mise en place des organes de la fleur. Toutefois, la prCsence de vascularisation propre au nectaire apical pose le problkme de I'identitC morphologique de cette partie de la fleur.
Au cours de la phase de sCcrCtion, le vacuome apparait constituk de nombreuses vCsicules de toutes tailles plus ou moins confluentes en une grande vacuole. Une telle Cvolution a CtC signalCe 2 propos des nectaires floraux des CaryophyllacCes (Zandonella 1972), du Cynoglossum oficinale (Tacina 1973) et du Trifolium pratense (Eriksson 1977). Cependant, Rachmilevitz et Fahn (1975) observent le contraire chez Trapaeolum majus. Egalement en phase sCcrCtoire et au cours du vieillissement de la cellule nectarifkre, on observe aussi une augmentation du nombre des amyloplastes. Eriks- son (1977) observe chez le nectaire floral du Trifolium pratense en phase sCcrCtoire un accroissement consi- dQable des enclaves amylifkres. D'autres auteurs men- tionnent au contraire une diminution progressive de la quantitC d'amidon plastidial pendant la phase de matura- tion de certains nectaires floraux (Fahn et Rachmilevitz 1970; Tacina 1973).
Les nectaires apicaux s'opposent aux nectaires ba- saux non seulement par la vascularisation ou par le prCsence de prCcipitCs chromophiles chez les fleurs staminCes mais aussi par la rCpartition des stomates. Ces derniers ont toutes les caractCristiques des stomates des nectaires: ostiole arrondi, ouvert, parois Cpaissies, chambre sous-stomatique rCduite ou nulle (Frey- Wissling et Hausermann 1960; Ancibor 1969). 11s sont souvent interprCtCs comme des orifices d'exudation du nectar, mais les observations directes sont rares. Zando- nella (1972), souligne l'inexistence de donnCes sur la physiologie de ces stomates.
Pour les nectaires basaux, la raretC ou l'absence des stomates permet difficilement de les envisager comme le lieu de sortie exclusif du nectar. Leur abondance au niveau du disque apical peut laisser supposer un r61e
diffkrent dans le fonctionnement de ce nectaire. Toute- fois, la participation Cventuelle des stomates dans la sortie du nectar reste ii prouver. Une Ctude de l'ultras- tructure des cellules des diffkrents types de nectaires, actuellement en cours en relation avec l'analyse des produits sCcrCtCs permettra une meilleure comprihen- sion du fonctionnement de ces organes.
Remerciements Les observations en microscopic electronique a ba-
layage ont CtC effectuCes g r k e a l'assistance technique de Mme Meury, Laboratoire d'Anatomie comparke, UniversitC Paris 7 , France.
ANCIBOR, E. 1969. Los nectarios florales en leguminosas- Mimosoideas. Darwinians, 15: 128- 142.
BORY, G., et D. CLAIR-MACZULAJTYS. 1978. Morphologie, anatomie et dCveloppement des glandes foliaires chez 1'Ailanthus glandulosa Desf. (SimarubacCes). Rev. Gen. Bot. 85: 137-157.
1980. Morphology, ontogeny and cytology of tri- chomes of Ailanthus altissima. Phytomorphology, 30: 67-78.
BRIZICKY, G. K. 1962. The genera of Simarubaceae and Burseraceae in the Southeastern United States. J. Arnold Arbor. Harv. Univ. 43: 173-183.
DAVIES, P. A. 1943. Floral glands in Ailanthus altissi~na. Trans. Ky. Acad. Sci. 11: 12-16.
ERIKSON, M. 1977. Ultrastructure of the nectary of red clover (Trifolium pratense). J. Apic. Res. 16: 184- 193.
FAHN, A. 1953. The topography of the nectary in the flower and its phylogenetical trend. Phytomorphology, 3: 424- 426.
FAHN, A., et T. RACHMILEVITZ. 1970. Ultrastructure and nectar secretion in Lonicera japonica. Duns New research in plant anatomy. Biol. Chem. Umbelliferae, Pap. Int. Symp. 1970, Bot. J. Linn. Soc. Suppl. No. 1, 1977. pp. 51-56.
FREY-WISSLING, A., et E. HAUSERMANN. 1960. Deutung der gestaltlosen Nektarien. Ber. Schweiz. Bot. Ges. 70: 150- 162.
JADIN, F. 1901. Contribution a 1'Ctude des SimarubacCes. Ann. Sci. Nat. Bot. Biol. Veg. 13: 201-304.
JENSEN, W. A. 1962. Botanical histochemistry. W. H. Freeman and Co., San Fransisco.
NAIR, N. C., et R. K. JOSHI. 1958. Floral morphology of some members of the Simaroubaceae. Bot. Gaz. (Chicago), 119: 88-99.
RACHMILEVITZ, T., et A. FAHN. 1975. The floral nectary of Tropaeolum majus L. The nature of the secretory cells and the manner of nectar secretion. Ann. Bot. 39: 721-728.
SPURR, A. R. 1969. A low-viscosity epoxy resin embedding medium for electron microscopy. J. Ultrastruct. Res. 26: 31-43.
TACINA, F. L. 1973. La structure optique et electronomicr~s- copique de la glande nectarifkre chez Cynoglossum oflcina- lis L. Rev. Roum. Biol. Ser. Bot. 18: 201-209
ZANDONELLA, P. 1972. Le nectaire floral des Centrospema- les. Localisation, morphologie, anatomie, histologie, cyto- logie. Thkse de doctorat d'Ctat, UniversitC de Lyon, Lyon.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
WIN
DSO
R o
n 11
/18/
14Fo
r pe
rson
al u
se o
nly.