Embed Size (px)
Citation preview

http://www.neur-one.fr
[email protected] - 1 - 17/03/2009 1
MUSCLES ET PHYSIOLOGIE MUSCULAIRE (1/3) MUSCLES ET PHYSIOLOGIE MUSCULAIRE (1/3) EN CONSTRUCTION EN CONSTRUCTION
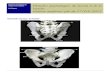
Il existe 3 sortes de muscles (fig. 1): Il existe 3 sortes de muscles (fig. 1):
• Les muscles striés, volontaires (par exemple: le biceps). Innervés par le système nerveux somatique, ils obéissent à la volonté, et peuvent répondre de façon réflexe à une stimulation. Leurs contractions peuvent être lentes (fibres rouges, de type1) ou rapides (fibres blanches, de type 2) .
• Les muscles striés, volontaires (par exemple: le biceps). Innervés par le système nerveux somatique, ils obéissent à la volonté, et peuvent répondre de façon réflexe à une stimulation. Leurs contractions peuvent être lentes (fibres rouges, de type1) ou rapides (fibres blanches, de type 2) .
• Les muscles lisses, involontaires (par exemple les parois de l'intestin). Innervés par le système nerveux autonome, leur action n'est pas liée à la volonté. Leurs contractions, bien que lentes, peuvent être soutenues longtemps.
• Les muscles lisses, involontaires (par exemple les parois de l'intestin). Innervés par le système nerveux autonome, leur action n'est pas liée à la volonté. Leurs contractions, bien que lentes, peuvent être soutenues longtemps.
• Le muscle cardiaque, au fonctionnement très particulier, possède en son sein ses propres ressources à l'origine de son automatisme. L'innervation du système nerveux autonome qu'il reçoit exerce un rôle modulateur de cet automatisme: le système nerveux orthosympathique est cardio-accélérateur, tandis que le système nerveux parasympathique est cardio-modérateur, du rythme et de la force de contraction du cœur.
• Le muscle cardiaque, au fonctionnement très particulier, possède en son sein ses propres ressources à l'origine de son automatisme. L'innervation du système nerveux autonome qu'il reçoit exerce un rôle modulateur de cet automatisme: le système nerveux orthosympathique est cardio-accélérateur, tandis que le système nerveux parasympathique est cardio-modérateur, du rythme et de la force de contraction du cœur.
Un muscle a besoin d'énergie pour produire du travail en se contractant. Il produit de la chaleur en travaillant. Un muscle a besoin d'énergie pour produire du travail en se contractant. Il produit de la chaleur en travaillant.

http://www.neur-one.fr
[email protected] - 2 - 17/03/2009 2 - 17/03/2009 2
Fig. 1-Les trois sortes de muscle Fig. 1-Les trois sortes de muscle
Les muscles somatiques sont normalement au repos; ils sont recrutés pour générer des mouvements (locomotion, mastication, respiration, mouvements oculaires etc…). Les contractions et le tonus du cœur et des muscles lisses sont régulés par les activités antagonistes des systèmes nerveux ortho- et para sympathique (péristaltisme, vasomotricité, miction etc…).
Les muscles somatiques sont normalement au repos; ils sont recrutés pour générer des mouvements (locomotion, mastication, respiration, mouvements oculaires etc…). Les contractions et le tonus du cœur et des muscles lisses sont régulés par les activités antagonistes des systèmes nerveux ortho- et para sympathique (péristaltisme, vasomotricité, miction etc…).
Excepté le coeur, les sphincters, l’oesophage supérieur et certains muscles attachés à la peau (muscles horripilateurs ou arrecteurs), les muscles striés somatiques s’attachent au squelette par des tendons ou aponévroses (collagène). Les muscles lisses et cardiaque (strié) sont des éléments constitutifs des organes "creux".
Excepté le coeur, les sphincters, l’oesophage supérieur et certains muscles attachés à la peau (muscles horripilateurs ou arrecteurs), les muscles striés somatiques s’attachent au squelette par des tendons ou aponévroses (collagène). Les muscles lisses et cardiaque (strié) sont des éléments constitutifs des organes "creux".
Un muscle est excitable (sensible à une stimulation), élastique (déformable si on l'étire), contractile (capable de se raccourcir).
Un muscle est excitable (sensible à une stimulation), élastique (déformable si on l'étire), contractile (capable de se raccourcir).
Le tissu musculaire se définit par une propriété physiologique, la contractilité, aboutissant à la motilité des masses musculaires.
Le tissu musculaire se définit par une propriété physiologique, la contractilité, aboutissant à la motilité des masses musculaires.
Différents caractères fonctionnels et morphologiques font distinguer le muscle strié squelettique (à contraction volontaire), le muscle strié cardiaque (à contraction involontaire) et le muscle lisse (à contraction involontaire).
Différents caractères fonctionnels et morphologiques font distinguer le muscle strié squelettique (à contraction volontaire), le muscle strié cardiaque (à contraction involontaire) et le muscle lisse (à contraction involontaire).
I. MUSCLE STRIE SQUELETTIQUE I. MUSCLE STRIE SQUELETTIQUE
En termes de taille absolue, l'organe le plus développé du corps de l'homme et des autres vertébrés est la musculature, la « chair ». Les muscles constituent 40 à 50% du poids total du corps. Leur fonction principale consiste à produire du travail en se contractant. Parmi les autres fonctions, on peut citer l'important rôle qu'ils jouent dans la régulation thermique corporelle, mais nous ne discuterons pas ici - où le propos est neurophysiologique - de l'intervention des muscles dans la production de chaleur.
En termes de taille absolue, l'organe le plus développé du corps de l'homme et des autres vertébrés est la musculature, la « chair ». Les muscles constituent 40 à 50% du poids total du corps. Leur fonction principale consiste à produire du travail en se contractant. Parmi les autres fonctions, on peut citer l'important rôle qu'ils jouent dans la régulation thermique corporelle, mais nous ne discuterons pas ici - où le propos est neurophysiologique - de l'intervention des muscles dans la production de chaleur.
L'homme travaille et peut modeler son environnement par la seule utilisation de ses muscles. Ceci ne vaut pas seulement pour le travail manuel, mais aussi pour l'activité intellectuelle, car écrire ou parler requiert une coordination musculaire très précise. On peut considérer - peut-être de façon un peu sommaire - le système nerveux comme un organe qui provoque des contractions musculaires appropriées en réponse aux stimuli que lui envoie l'environnement. Ceci veut dire que les muscles constituent un sujet d'études fort important pour le neurophysiologiste. De plus, les modalités de fonctionnement des cellules musculaires sont bien mieux connues que celles de la plupart des autres cellules. La morphologie, les composants chimiques, les réactions et les fonctions physiologiques des cellules musculaires, ont été très étudiés et les diverses approches des dernières années ont permis une synthèse qui se présente sous la forme d'une théorie unifiée de la contraction musculaire. Tout en discutant la fonction des muscles, nous devrons ainsi porter notre attention sur leurs caractéristiques structurales et biochimiques.
L'homme travaille et peut modeler son environnement par la seule utilisation de ses muscles. Ceci ne vaut pas seulement pour le travail manuel, mais aussi pour l'activité intellectuelle, car écrire ou parler requiert une coordination musculaire très précise. On peut considérer - peut-être de façon un peu sommaire - le système nerveux comme un organe qui provoque des contractions musculaires appropriées en réponse aux stimuli que lui envoie l'environnement. Ceci veut dire que les muscles constituent un sujet d'études fort important pour le neurophysiologiste. De plus, les modalités de fonctionnement des cellules musculaires sont bien mieux connues que celles de la plupart des autres cellules. La morphologie, les composants chimiques, les réactions et les fonctions physiologiques des cellules musculaires, ont été très étudiés et les diverses approches des dernières années ont permis une synthèse qui se présente sous la forme d'une théorie unifiée de la contraction musculaire. Tout en discutant la fonction des muscles, nous devrons ainsi porter notre attention sur leurs caractéristiques structurales et biochimiques.
La partie la plus importante de la musculature, la musculature squelettique, est constituée de muscles séparés. Un tel muscle est un « morceau de chair » oblong, terminé à ses deux extrémités par des tendons ressemblant à des cordes. Le muscle est relié aux os, au « squelette », par ces tendons qui lui permettent donc d'agir sur les pièces squelettiques.
La partie la plus importante de la musculature, la musculature squelettique, est constituée de muscles séparés. Un tel muscle est un « morceau de chair » oblong, terminé à ses deux extrémités par des tendons ressemblant à des cordes. Le muscle est relié aux os, au « squelette », par ces tendons qui lui permettent donc d'agir sur les pièces squelettiques.
Le muscle strié squelettique est un tissu musculaire à contraction volontaire, intervenant dans le maintien postural et le mouvement. Il est constitué principalement de fibres multinucléées qui doivent leur nom à une double striation longitudinale et transversale.
Le muscle strié squelettique est un tissu musculaire à contraction volontaire, intervenant dans le maintien postural et le mouvement. Il est constitué principalement de fibres multinucléées qui doivent leur nom à une double striation longitudinale et transversale.
1. Organisation générale : constitution d'un muscle strié squelettique (figure 2)1. Organisation générale : constitution d'un muscle strié squelettique (figure 2)
Les muscles sont des organes propres au sein desquels des fibres musculaires striées sont juxtaposées parallèlement et organisées en faisceaux, tendus entre deux insertions tendino-aponévrotiques.
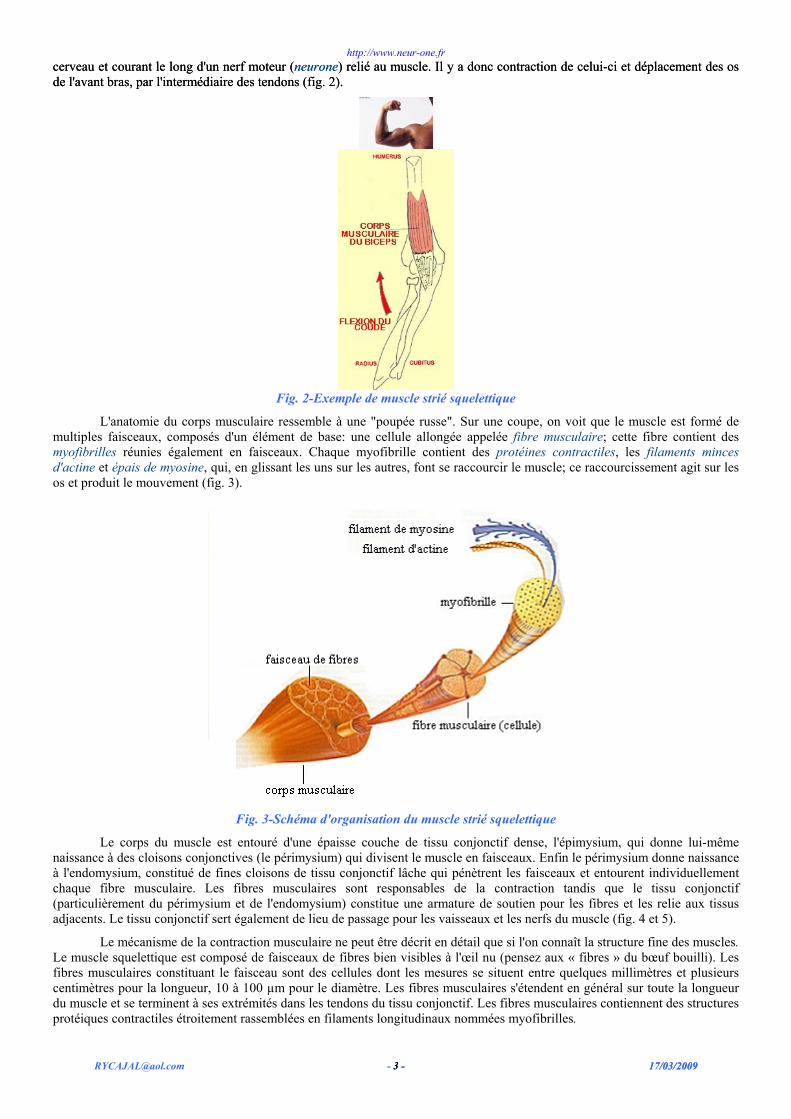
Si l'on prend l'exemple simple et bien connu d'un muscle long tel que le biceps, on voit qu'il est composé d'un corps musculaire rouge qui s'attache à l'os grâce à une structure appelée tendon, de couleur blanc- nacrée. Quand je décide de plier le coude, mon cerveau va commander au biceps de tirer sur mon avant-bras grâce à une onde électrique (influx nerveux) venant du
http://www.neur-one.fr
[email protected] - 3 - 17/03/2009 3 - 17/03/2009 3
cerveau et courant le long d'un nerf moteur (neurone) relié au muscle. Il y a donc contraction de celui-ci et déplacement des os de l'avant bras, par l'intermédiaire des tendons (fig. 2). cerveau et courant le long d'un nerf moteur (neurone) relié au muscle. Il y a donc contraction de celui-ci et déplacement des os de l'avant bras, par l'intermédiaire des tendons (fig. 2).
Fig. 2-Exemple de muscle strié squelettique
L'anatomie du corps musculaire ressemble à une "poupée russe". Sur une coupe, on voit que le muscle est formé de multiples faisceaux, composés d'un élément de base: une cellule allongée appelée fibre musculaire; cette fibre contient des myofibrilles réunies également en faisceaux. Chaque myofibrille contient des protéines contractiles, les filaments minces d'actine et épais de myosine, qui, en glissant les uns sur les autres, font se raccourcir le muscle; ce raccourcissement agit sur les os et produit le mouvement (fig. 3).
Fig. 3-Schéma d'organisation du muscle strié squelettique
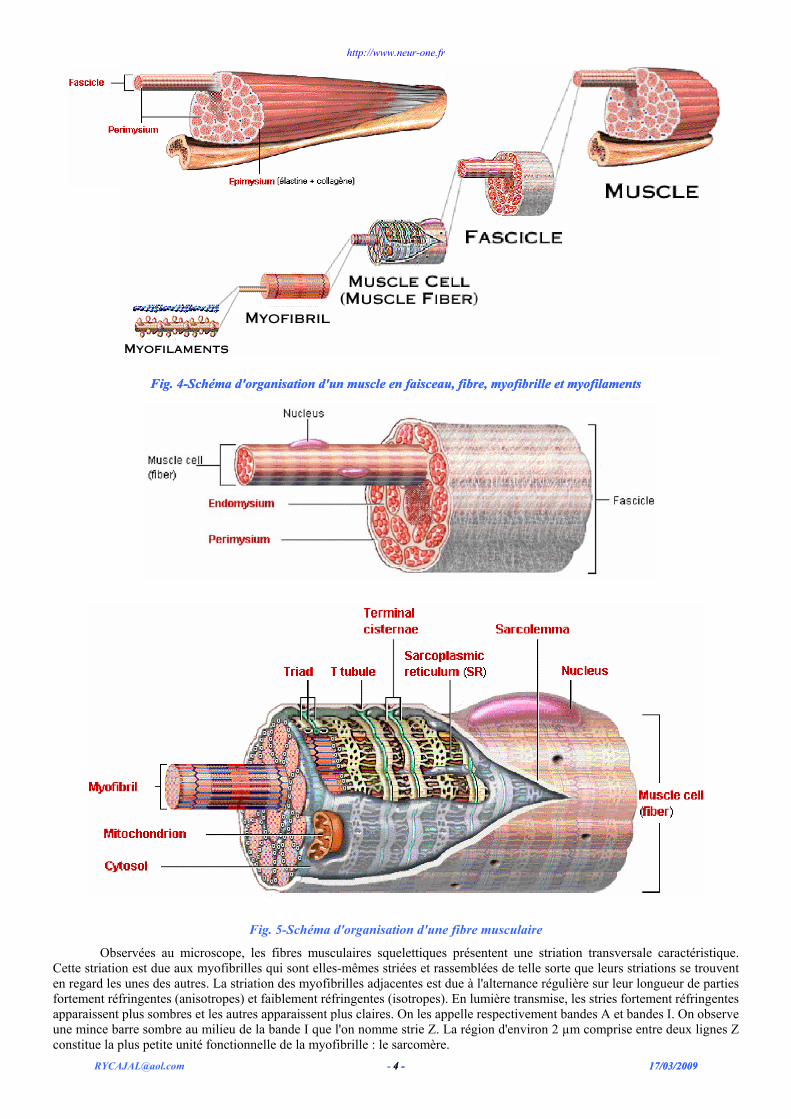
Le corps du muscle est entouré d'une épaisse couche de tissu conjonctif dense, l'épimysium, qui donne lui-même naissance à des cloisons conjonctives (le périmysium) qui divisent le muscle en faisceaux. Enfin le périmysium donne naissance à l'endomysium, constitué de fines cloisons de tissu conjonctif lâche qui pénètrent les faisceaux et entourent individuellement chaque fibre musculaire. Les fibres musculaires sont responsables de la contraction tandis que le tissu conjonctif (particulièrement du périmysium et de l'endomysium) constitue une armature de soutien pour les fibres et les relie aux tissus adjacents. Le tissu conjonctif sert également de lieu de passage pour les vaisseaux et les nerfs du muscle (fig. 4 et 5).
Le mécanisme de la contraction musculaire ne peut être décrit en détail que si l'on connaît la structure fine des muscles. Le muscle squelettique est composé de faisceaux de fibres bien visibles à l'œil nu (pensez aux « fibres » du bœuf bouilli). Les fibres musculaires constituant le faisceau sont des cellules dont les mesures se situent entre quelques millimètres et plusieurs centimètres pour la longueur, 10 à 100 µm pour le diamètre. Les fibres musculaires s'étendent en général sur toute la longueur du muscle et se terminent à ses extrémités dans les tendons du tissu conjonctif. Les fibres musculaires contiennent des structures protéiques contractiles étroitement rassemblées en filaments longitudinaux nommées myofibrilles.
http://www.neur-one.fr
[email protected] - 4 - 17/03/2009 4 - 17/03/2009 4
Fig. 4-Schéma d'organisation d'un muscle en faisceau, fibre, myofibrille et myofilaments Fig. 4-Schéma d'organisation d'un muscle en faisceau, fibre, myofibrille et myofilaments
Fig. 5-Schéma d'organisation d'une fibre musculaire
Observées au microscope, les fibres musculaires squelettiques présentent une striation transversale caractéristique. Cette striation est due aux myofibrilles qui sont elles-mêmes striées et rassemblées de telle sorte que leurs striations se trouvent en regard les unes des autres. La striation des myofibrilles adjacentes est due à l'alternance régulière sur leur longueur de parties fortement réfringentes (anisotropes) et faiblement réfringentes (isotropes). En lumière transmise, les stries fortement réfringentes apparaissent plus sombres et les autres apparaissent plus claires. On les appelle respectivement bandes A et bandes I. On observe une mince barre sombre au milieu de la bande I que l'on nomme strie Z. La région d'environ 2 µm comprise entre deux lignes Z constitue la plus petite unité fonctionnelle de la myofibrille : le sarcomère.
http://www.neur-one.fr
[email protected] - 5 - 17/03/2009 5
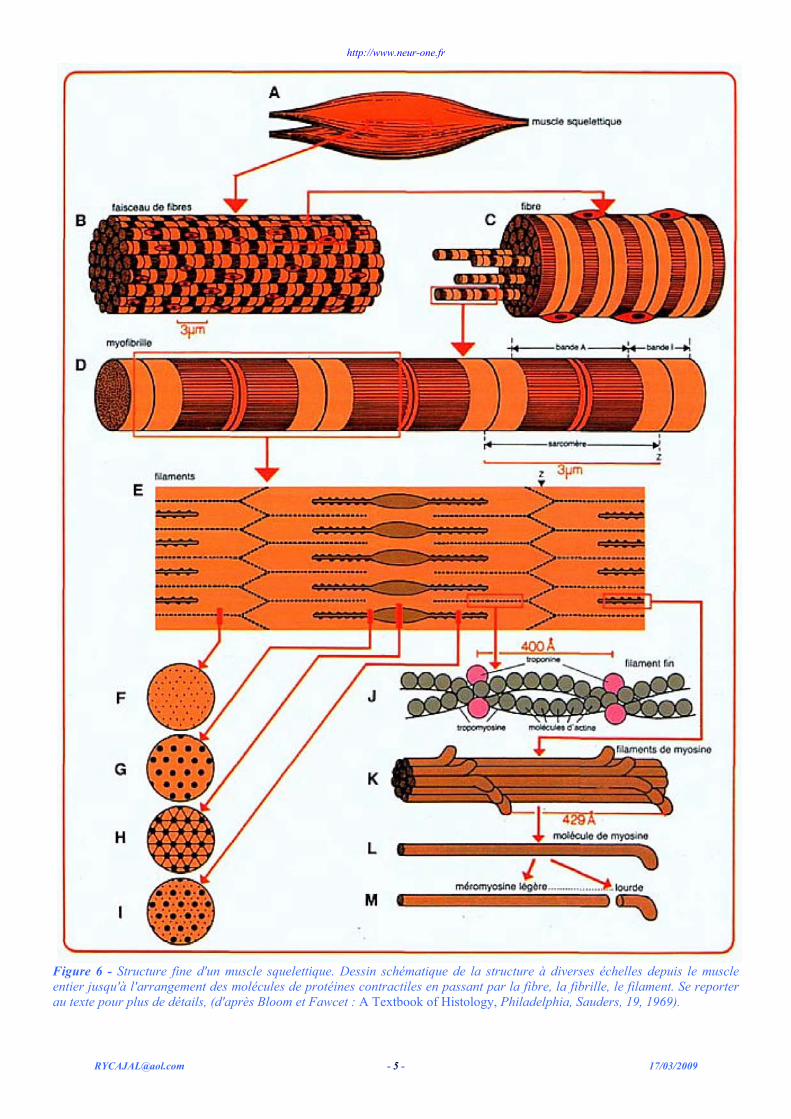
Figure 6 - Structure fine d'un muscle squelettique. Dessin schématique de la structure à diverses échelles depuis le muscle entier jusqu'à l'arrangement des molécules de protéines contractiles en passant par la fibre, la fibrille, le filament. Se reporter au texte pour plus de détails, (d'après Bloom et Fawcet : A Textbook of Histology, Philadelphia, Sauders, 19, 1969).

http://www.neur-one.fr
[email protected] - 6 - 17/03/2009
Fig. 7 – Micrographie optique de fibres musculaires squelettiques
En microscopie optique (fig. 7), les fibres musculaires extra-fusales apparaissent comme des éléments allongés, plurinucléés et présentant une striation transversale régulière. De forme plus ou moins cylindrique, elles mesurent 10 à 100 µm de diamètre avec une longueur variable de quelques mm à plusieurs cm. Plusieurs noyaux, en position périphérique, sont allongés dans le sens de la fibre; en coupe transversale à 5 µm d'épaisseur, chaque fibre contient en moyenne 2 à 5 noyaux.
La majeure partie du cytoplasme est occupée par des myofibrilles, qui se groupent en amas apparaissant en coupe longitudinale sous forme de colonnes (Leydig) et transversalement sous forme de polygones (champs de Cohnheim). Les myofibrilles et le cytoplasme restant constituent par définition le sarcoplasme, lequel est limité en périphérie par la membrane plasmique. Celle-ci est doublée d'une lame basale, l'ensemble constituant le sarcolemme.
Les myofibrilles sont des cylindres d'un diamètre moyen d'un à deux µm, disposés parallèlement avec une striation transversale périodique concordant de l'une à l'autre, ce qui explique l'aspect strié de l'ensemble de la fibre. Cette striation se caractérise par l'alternance régulière de disques sombres A anisotropes (d'une longueur moyenne d'1,5 µm) et de disques clairs I isotropes (d'une longueur moyenne de 0,8 µm). La partie centrale des disques I est marquée une strie régulière, ou strie Z. Une zone plus claire au centre du disque A correspond à la bande H, parfois centrée par la strie M.
La case musculaire, ou sarcomère, a été décrite en 1868 par Krause comme un prisme de substance anisotrope (disque A) entouré de part et d'autre de substance isotrope (deux demi-disques I). Cet ensemble, limité latéralement par deux stries Z, correspond à l'unité élémentaire de contraction du tissu musculaire.
Le microscope électronique (figure 7) permet d'aller plus avant dans l'étude de la structure fine du sarcomère. Comme on l'a vu, la strie Z relie entre eux des myofilaments fins adjacents. Dans la région centrale se trouvent des myofilaments épais, encadrés par les myofilaments fins. Les coupes transversales montrent que les myofilaments fins et épais sont organisés d'une manière stricte qui fait penser à un arrangement cristallin. L'attaque par voie chimique montre que les filaments fins sont surtout constitués d'actine, alors que les filaments épais sont constitués par les longues molécules d'une autre protéine, la myosine. Ainsi, les bandes I des myofïbrilles sont surtout constituées par de l'actine, et la partie centrale des bandes A est exclusivement constituée de myosine ; les parties extrêmes des bandes A sont constituées d'actine et de myosine. C'est la myosine qui confère aux bandes A leur forte réfringence.
Figure 7 – Micrographie électronique montrant l'organisation des sarcomères
Le sarcomère contient deux types de myofilaments,, myofilaments de myosine et d'actine superposés les uns aux autres et orientés dans l'axe des myofibrilles. Les filaments épais de myosine, de 12 à 14 nm d'épaisseur et d'1,6 µm de long, sont situés au milieu du sarcomère au niveau du disque A. Les filaments fins d'actine, de 5 à 7 nm d'épaisseur et de 0,98 µm de long, s'insèrent sur les stries Z. Ils se situent dans les disques I et les parties latérales du disque A (en dehors de la bande H).
6
http://www.neur-one.fr
[email protected] - 7 - 17/03/2009
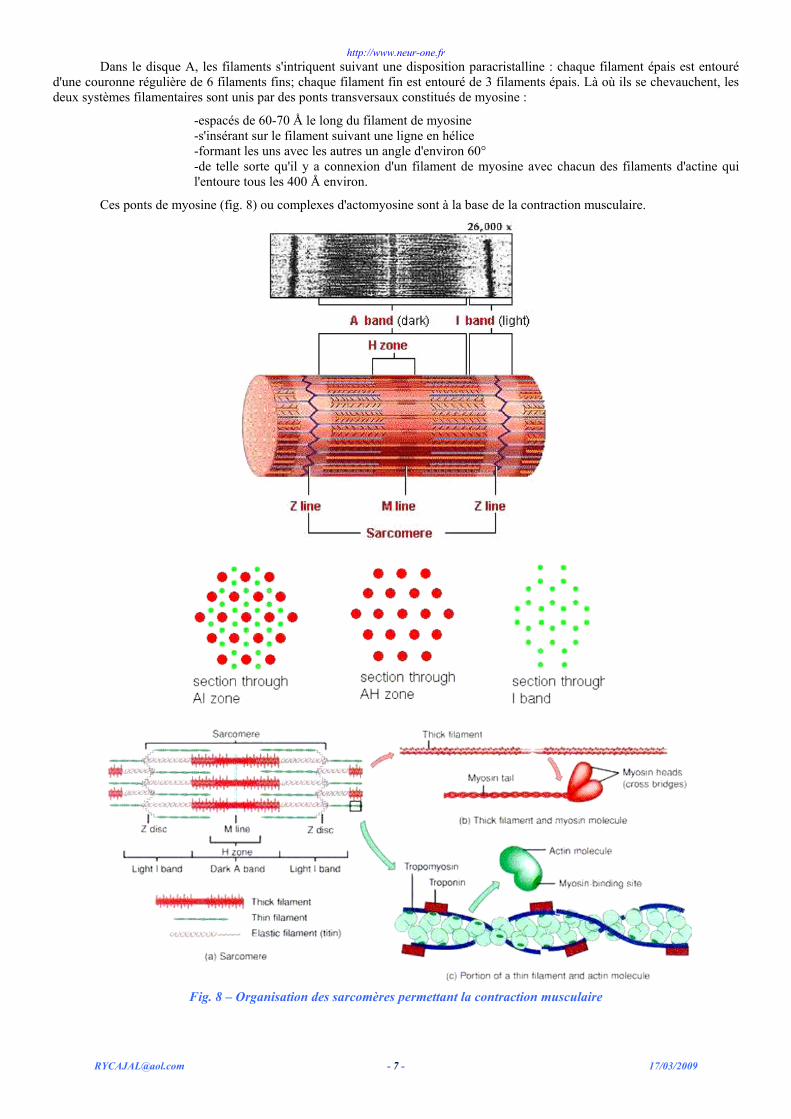
Dans le disque A, les filaments s'intriquent suivant une disposition paracristalline : chaque filament épais est entouré d'une couronne régulière de 6 filaments fins; chaque filament fin est entouré de 3 filaments épais. Là où ils se chevauchent, les deux systèmes filamentaires sont unis par des ponts transversaux constitués de myosine :
-espacés de 60-70 Å le long du filament de myosine -s'insérant sur le filament suivant une ligne en hélice -formant les uns avec les autres un angle d'environ 60° -de telle sorte qu'il y a connexion d'un filament de myosine avec chacun des filaments d'actine qui l'entoure tous les 400 Å environ.
Ces ponts de myosine (fig. 8) ou complexes d'actomyosine sont à la base de la contraction musculaire.
Fig. 8 – Organisation des sarcomères permettant la contraction musculaire
7
http://www.neur-one.fr
[email protected] - 8 - 17/03/2009 8
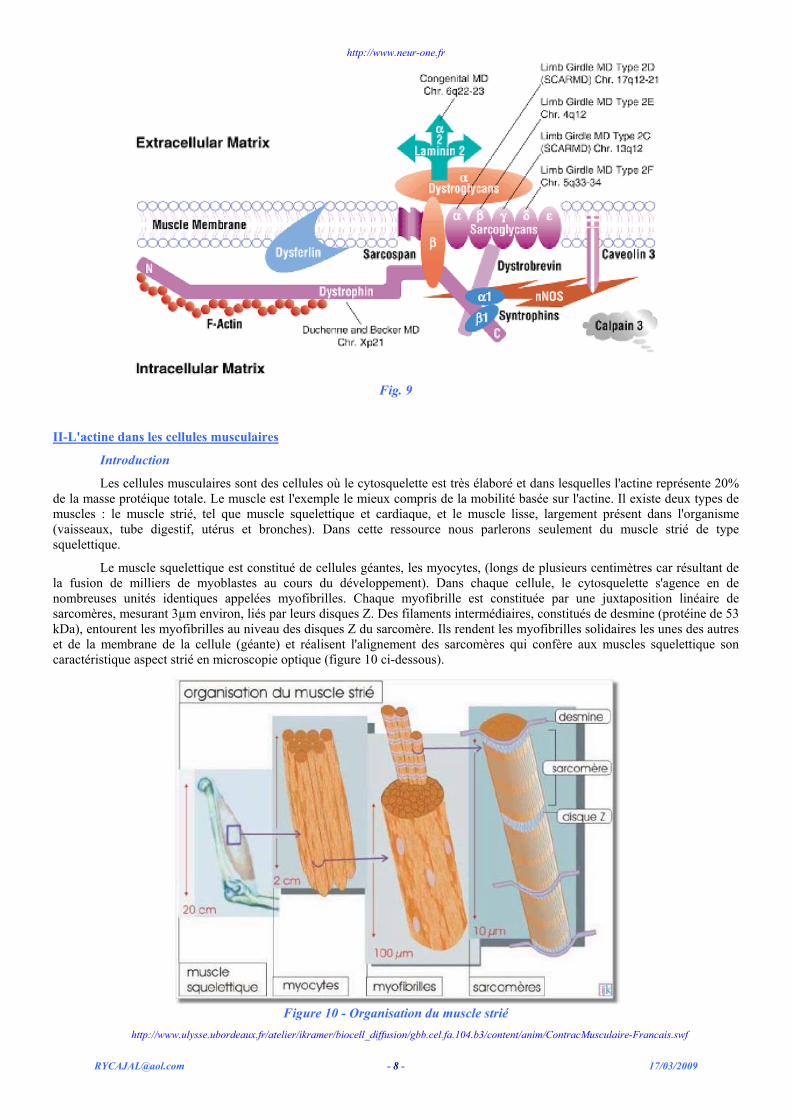
Fig. 9
II-L'actine dans les cellules musculaires
Introduction
Les cellules musculaires sont des cellules où le cytosquelette est très élaboré et dans lesquelles l'actine représente 20% de la masse protéique totale. Le muscle est l'exemple le mieux compris de la mobilité basée sur l'actine. Il existe deux types de muscles : le muscle strié, tel que muscle squelettique et cardiaque, et le muscle lisse, largement présent dans l'organisme (vaisseaux, tube digestif, utérus et bronches). Dans cette ressource nous parlerons seulement du muscle strié de type squelettique.
Le muscle squelettique est constitué de cellules géantes, les myocytes, (longs de plusieurs centimètres car résultant de la fusion de milliers de myoblastes au cours du développement). Dans chaque cellule, le cytosquelette s'agence en de nombreuses unités identiques appelées myofibrilles. Chaque myofibrille est constituée par une juxtaposition linéaire de sarcomères, mesurant 3µm environ, liés par leurs disques Z. Des filaments intermédiaires, constitués de desmine (protéine de 53 kDa), entourent les myofibrilles au niveau des disques Z du sarcomère. Ils rendent les myofibrilles solidaires les unes des autres et de la membrane de la cellule (géante) et réalisent l'alignement des sarcomères qui confère aux muscles squelettique son caractéristique aspect strié en microscopie optique (figure 10 ci-dessous).
Figure 10 - Organisation du muscle strié
http://www.ulysse.ubordeaux.fr/atelier/ikramer/biocell_diffusion/gbb.cel.fa.104.b3/content/anim/ContracMusculaire-Francais.swf

http://www.neur-one.fr
[email protected] - 9 - 17/03/2009 9
Le sarcomère comme unité de contraction
L'actine et la myosine sont à la base de la contractilité des sarcomères qui sont constitués par un assemblage de filaments parallèles d'actine (filaments minces) et de myosine-II (filaments épais) (figure 11 ci-dessous). Les filaments d'actine, longs d'environ 1µm, sont attachés aux disques Z par l'intermédiaire de capZ (protéine de coiffage qui se fixe à l'extrémité plus) et de l'α-actinine. L'extrémité moins (libre) est stabilisée par la tropomoduline. Sur sa longueur, le filament d'actine est associé à d'autres protéines qui interviennent dans la contraction musculaire (voir ci-dessous).
Figure 11 - Vue de détail du sarcomère
Les filaments de myosine-II, structures bipolaires résultant de l'association de nombreuses molécules de myosine-II, alternent régulièrement avec les filaments actine (figure 11 ci-dessus). La myosine-II est une protéine motrice formée d'une tête et d'une queue. La queue sert à insérer la protéine dans le filament et la tête, responsable d'une activité ATPase, interagit avec les filaments d'actine. Deux petites chaînes protéiques légères (17 kDa) entourent la myosine-II au niveau de la transition tête-queue.
Le filament épais de myosine-II est maintenu en place par un troisième filament, constitué de titine. C'est une protéine élastique de 3300 kDa (sa taille lui a valu son nom qui fait, semble-t-il, référence à un géant mythologique du nom de Titin (les étudiants susceptibles d'élucider l'origine étymologique et mythologique du mot titine sont priés de nous en informer). C'est une des plus grandes protéines codées par le génome humain. La titine fait la liaison entre le disque Z et le filament épais de myosine-II. Par sa forme, la titine est une molécule élastique qui permet d'entretenir dans le muscle le phénomène de tension passive. De plus elle permet de centrer parfaitement le filament épais de myosine-II entre les filaments d'actine (figure 11 ci-dessus).
Le raccourcissement du sarcomère est provoqué par le glissement des filaments d'actine sur les filaments de myosine-II (force motrice), déclenché par l'hydrolyse de l'ATP (figure 12 ci-dessous).
Remarques étymologiques:
Sarco, du grec muscle, équivaut au latin myo. Le vocabulaire utilise indifféremment la racine grecque ou la racine latine. Par exemple on parle de myofibrilles, assemblage de sarcomères, contenant des filaments de myosine-II. Les myofibrilles sont entourées par du sarcoplasme (cytoplasme), contenant du réticulum sarcoplasmique (réticulum endoplasmique lisse), le tout emballé dans un sarcolemme (plasmolemme ou membrane plasmique).
Le déplacement de l'actine induit par la myosine-II
Ce déplacement s'effectue selon un cycle de modifications successives. Au début du cycle, la tête de myosine-II est attachée à l'actine. Cette interaction est de très courte durée car une molécule d'ATP se lie à la tête et provoque une réduction d'affinité pour l'actine. La tête de myosine-II s'éloigne. L'hydrolyse de l'ATP s'ensuit (étape limitante) et induit un changement de la position de la tête de myosine-II (ADP et Pi restent associés à la myosine-II). Dans cet état, la tête s'attache de nouveau à l'actine. La perte subséquente de phosphate (Pi) remet la tête de myosine-II à la position de départ, ce qui déplace le filament d'actine d'environ 10 nm. La perte de Pi est le moment où l'énergie libérée par l'hydrolyse de l'ATP est convertie en mouvement. L'ADP se détache et est remplacé en moins d'une milliseconde par une nouvelle molécule d'ATP et un nouveau cycle peut commencer. La répétition de ce cycle engendre une contraction dynamique (figure 13 ci-dessous).
Le Ca2+ et la troponine/tropomyosine-II interviennent dans la régulation de la contraction du muscle
L'interaction actine/myosine est hautement régulée pour prévenir les contractions musculaires indésirables (par exemple, imaginez les conséquences désastreuses sur la respiration d'une contraction des muscles intercostaux maintenue pendant quelques minutes). La contraction du muscle squelettique est déclenchée par des motoneurones qui forment des

http://www.neur-one.fr
[email protected] - 10 - 17/03/2009 10
synapses spécialisées, les jonctions neuro-musculaires (ou plaques motrices) (figure 14 ci-dessous). L'ensemble constitué par un motoneurone et une ou quelques cellules musculaires est appelé « unité motrice ». Le système nerveux influence la force de contraction d'un muscle :
en mobilisant plus au moins d'unités motrices et
en réglant la fréquence d'activation de chacune de ces unités motrices (avec un maximum de 200 potentiels d'action car chaque cycle dure 50 millisecondes) (revoir aussi la figure 17 .
Le Ca2+ et la troponine/tropomyosine-II interviennent dans la régulation de la contraction du muscle
L'interaction actine/myosine est hautement régulée pour prévenir les contractions musculaires indésirables (par exemple, imaginez les conséquences désastreuses sur la respiration d'une contraction des muscles intercostaux maintenue pendant quelques minutes). La contraction du muscle squelettique est déclenchée par des motoneurones qui forment des synapses spécialisées, les jonctions neuro-musculaires (ou plaques motrices) (figure 14 ci-dessous). L'ensemble constitué par un motoneurone et une ou quelques cellules musculaires est appelé « unité motrice ». Le système nerveux influence la force de contraction d'un muscle :
en mobilisant plus au moins d'unités motrices et
en réglant la fréquence d'activation de chacune de ces unités motrices (avec un maximum de 200 potentiels d'action car chaque cycle dure 50 millisecondes.
Figure 12 - Le sarcomère comme unité de contraction
Figure 13 - Déplacement de l'actine (contraction du sarcomère) dû au changement de la position de la tête de myosine
http://www.neur-one.fr
[email protected] - 11 - 17/03/2009 11
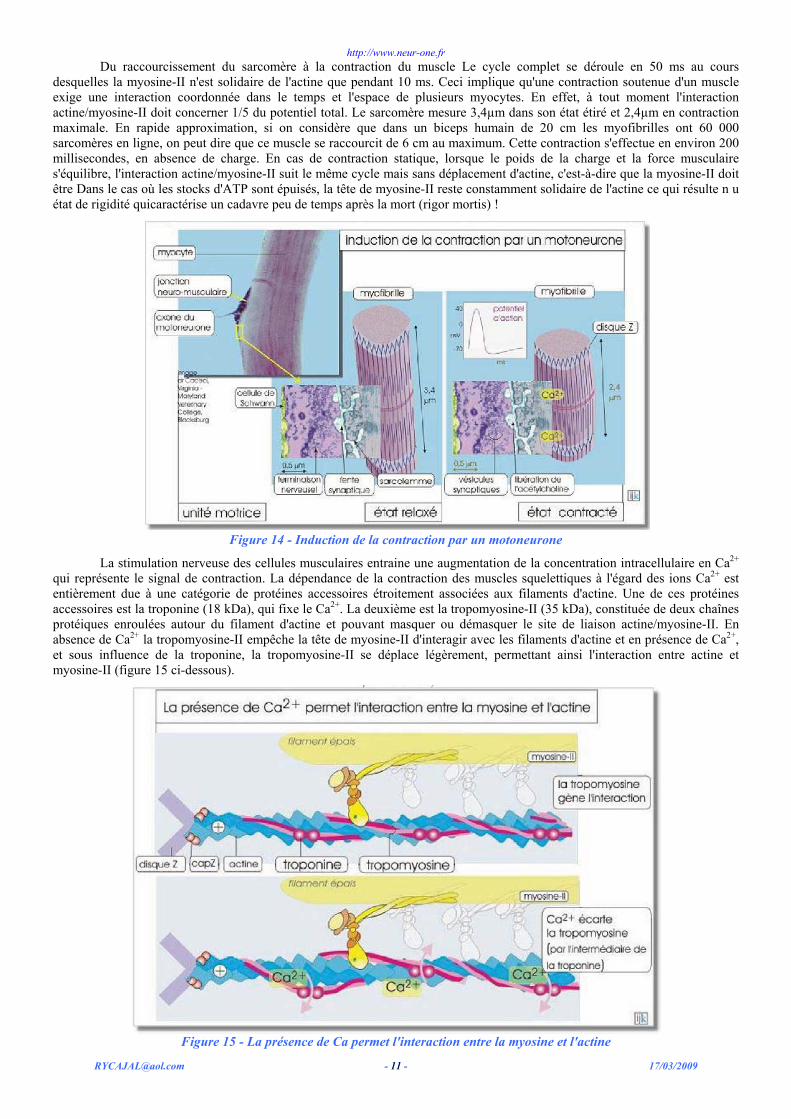
Du raccourcissement du sarcomère à la contraction du muscle Le cycle complet se déroule en 50 ms au cours desquelles la myosine-II n'est solidaire de l'actine que pendant 10 ms. Ceci implique qu'une contraction soutenue d'un muscle exige une interaction coordonnée dans le temps et l'espace de plusieurs myocytes. En effet, à tout moment l'interaction actine/myosine-II doit concerner 1/5 du potentiel total. Le sarcomère mesure 3,4µm dans son état étiré et 2,4µm en contraction maximale. En rapide approximation, si on considère que dans un biceps humain de 20 cm les myofibrilles ont 60 000 sarcomères en ligne, on peut dire que ce muscle se raccourcit de 6 cm au maximum. Cette contraction s'effectue en environ 200 millisecondes, en absence de charge. En cas de contraction statique, lorsque le poids de la charge et la force musculaire s'équilibre, l'interaction actine/myosine-II suit le même cycle mais sans déplacement d'actine, c'est-à-dire que la myosine-II doit être Dans le cas où les stocks d'ATP sont épuisés, la tête de myosine-II reste constamment solidaire de l'actine ce qui résulte n u état de rigidité quicaractérise un cadavre peu de temps après la mort (rigor mortis) !
Figure 14 - Induction de la contraction par un motoneurone
La stimulation nerveuse des cellules musculaires entraine une augmentation de la concentration intracellulaire en Ca2+ qui représente le signal de contraction. La dépendance de la contraction des muscles squelettiques à l'égard des ions Ca2+ est entièrement due à une catégorie de protéines accessoires étroitement associées aux filaments d'actine. Une de ces protéines accessoires est la troponine (18 kDa), qui fixe le Ca2+. La deuxième est la tropomyosine-II (35 kDa), constituée de deux chaînes protéiques enroulées autour du filament d'actine et pouvant masquer ou démasquer le site de liaison actine/myosine-II. En absence de Ca2+ la tropomyosine-II empêche la tête de myosine-II d'interagir avec les filaments d'actine et en présence de Ca2+, et sous influence de la troponine, la tropomyosine-II se déplace légèrement, permettant ainsi l'interaction entre actine et myosine-II (figure 15 ci-dessous).
Figure 15 - La présence de Ca permet l'interaction entre la myosine et l'actine

http://www.neur-one.fr
[email protected] - 12 - 17/03/2009 12
Le cycle de modification de la myosine en absence de Ca2+ (état de repos):
En absence de Ca2+, le cycle d'hydrolyse de l'ATP se déroule mais de façon plus lente (environ 1000 fois). En effet, les têtes de myosine-II restent plus longtemps dans l'état de liaison ADP+Pi avant que le Pi et l'ADP soient libérés et remplacés par une nouvelle molécule d'ATP. C'est le contact actine-myosine-II qui accélère la perte de Pi et ADP et donc le cycle d'hydrolyse. On peut ainsi prendre conscience de l'énorme demande d'ATP (1000 fois) lorsque le muscle se contracte, une demande qui doit être compensée par une production mitochondriale équivalente, de façon à éventuellement soutenir une contraction prolongée.
III-Déplacement des filaments d'actine et de myosine pendant la contraction
Les processus chimiques et physiques à l'origine de la contraction s'éclairent si on observe le comportement des éléments constituant le muscle pendant cette contraction. Lorsque le muscle est raccourci - par exemple dans le cas d'une contraction isotonique - la longueur des bandes A reste constante alors que les bandes I se raccourcissent. Si les bandes A ne changent pas de longueur, les filaments de myosine doivent également conserver leur longueur. Le microscope électronique confirme ce fait, mais il révèle aussi que les filaments d'actine, seuls constituants de la bande I, conservent eux aussi leur longueur pendant la contraction. Puisque les longueurs des filaments d'actine et de myosine ne changent pas, la longueur du sarcomère ne peut diminuer pendant la contraction que si les filaments glissent les uns par rapport aux autres, augmentant ainsi leur degré de recouvrement. Cette interprétation, généralement admise à l'heure actuelle, est connue sous le nom de théorie du glissement des filaments. Pendant la contraction le sarcomère se raccourcit et la distance entre les stries Z devient moindre car les filaments de myosine s'insinuent entre les filaments d'actine. Les extrémités des filaments de myosine s'approchent des stries Z et les bandes I se réduisent en conséquence (fig.16).
Fig. 16 - Mécanisme de la contraction. Diagramme de la disposition des filaments d'actine et de myosine dans un sarcomère (entre deux lignes Z) et des ponts entre filaments. On pense que les modifications de configuration des ponts engendrent une force de traction qui déplace les filaments d'actine entre ceux de myosine. A - disposition au début de la. contraction, B - état du muscle lorsque la contraction est maximale : le sarcomère est raccourci.
Mécanisme moléculaire de la contraction. -
Quelle peut être la cause de ce déplacement de myofibrilles pendant la contraction ? Des traits rouges en oblique relient, sur la figure 16 , les filaments d'actine et de myosine. Ils symbolisent les groupes de molécules-ponts reliant ces filaments entre eux. Lorsqu'un tel lien est constitué entre les filaments, il se raccourcit, ce qui a pour effet de tirer un filament d'actine entre deux filaments de myosine. Après raccourcissement, le lien se brise et le cycle peut recommencer : un nouveau lien se forme, qui se raccourcit, se brise, etc.
Les « liens » ou « ponts » dont le raccourcissement produit le déplacement des filaments de myosine, et donc la contraction, ont leur origine dans l'extrémité épaisse de la molécule de myosine. La figure 6 montre que le filament de myosine est composé de molécules (figure 6K, L) étroitement juxtaposées en quinconce. Les têtes épaissies des molécules de myosine sont régulièrement réparties dans ce filament. Cette partie du filament de myosine porte le nom de « méromyosine lourde (figure 6 M). La méromyosine lourde peut, d'une part entrer en combinaison chimique avec l'actine, d'autre part agir comme enzyme et hydrolyser l'adénosine triphosphate (ATP). Ce phénomène s'accompagne de libération d'énergie, laquelle est en partie utilisée pour assurer le raccourcissement des ponts jetés entre myosine et actine, et en partie transformée en chaleur. La cause de la contraction musculaire est donc la formation de complexes de myosine, d'actine et d'ATP, ce dernier se trouvant amputé d'un groupe phosphate et les ponts entre myosine et actine se trouvant raccourcis. Beaucoup d'observations conduisent à penser que les deux segments rigides qui constituent les ponts font un angle l'un par rapport à l'autre. Pendant le raccourcissement l'angle entre les deux segments se trouve réduit.
La réaction entre la myosine, l'actine et l'ATP qui constitue la base moléculaire de la contraction ne peut se manifester que lorsque la concentration des ions Ca++ dans la cellule est d'environ 10-5 mol /1. La contraction requiert également la présence d'ions Mg++. Les ions Ca++ ne se trouvent à la concentration de 10-5 mol /1 dans le milieu intracellulaire que pendant la

http://www.neur-one.fr
[email protected] - 13 - 17/03/2009 13
contraction. Dans le muscle au repos, la concentration intracellulaire des ions Ca++ est d'environ 10-8 mol /1, en conséquence de quoi, la réaction myosine, actine, ATP ne peut avoir lieu. Les cellules possèdent un mécanisme d'accroissement de leur concentration en ions Ca++ et c'est ce qui permet la mise en jeu des réactions moléculaires.
Non seulement l'ATP fournit l'énergie nécessaire à la contraction musculaire, mais il joue un autre rôle vis-à-vis du système de contraction. Après que le muscle se soit contracté, et même lorsque la concentration des ions Ca++ est tombée à 10-8 mol /1, les liens entre myosine et actine ne se brisent qu'en présence d'ATP. Cette action de l'ATP a été désignée du nom d'effet plastifiant. Lorsque la concentration en ATP d'un muscle tombe du fait d'une réduction de la fourniture d'énergie, le muscle ne peut se relâcher : il reste « dur » et rigide : on le dit en état de « rigidité ». Après la mort, également, la concentration en ATP dans le muscle tombe, son effet plastifiant sur les complexes myosine-actine n'existe plus et la rigidité cadavérique s'installe.
Caractéristiques de la contraction musculaire en fonction de la longueur de la fibre et de la vitesse de contraction.
Les filaments d'actine sont tirés entre les filaments de myosine pendant la contraction. Toutefois, la force développée sur les tendons par ce processus, et le raccourcissement même de la fibre, dépendent des circonstances qui accompagnent la contraction. Expérimentalement on démontre que la tension développée par un muscle dépend beaucoup de la longueur des fibres au moment de l'activation - c 'est-à-dire du degré de « tension préalable » - ainsi que de la vitesse avec laquelle s'opère le raccourcissement. Ces paramètres étant également de grande importance in vivo, nous les discuterons en détails maintenant.
La courbe d'équilibre tension-longueur au repos. - Les muscles ou les fibres musculaires isolées maintenus à leur « longueur de repos » n'exercent aucune force sur leurs points d'attache. Si on tire sur une extrémité du muscle, celui-ci s'étend. Si on note la force nécessaire pour produire une extension dans ces conditions, et qu'on construise ses variations en fonction de la longueur du muscle, on obtient une courbe tension-longueur du type de celle représentée par la figure 17. La force croît exponentiellement avec la longueur à partir de la valeur 0, correspondant à la longueur de repos 10. Le muscle peut supporter sans dommage d'être étiré 1,8 lois sa longueur de repos, mais tout étirement supplémentaire lèse les libres musculaires.
Figure 17 - Courbe tension-longueur et courbe de maximum isométrique. Les relations observées entre la force développée par le muscle et sa longueur au repos sont représentées sur la courbe en noir ; le maximum de tension engendrée activement en conditions isométriques est représenté sur la courbe en rouge. 10 est la longueur du muscle au repos ; l'axe des abscisses est gradué en multiples de 10. P0 est la tension maximale additionnelle qu'on peut engendrer isométriquement.
Lorsqu'on étire le muscle, l'élongation se manifeste d'abord dans le sarcomère où les filaments d'actine et de myosine glissent les uns par rapport aux autres (cf. figure 16). Les bandes I s'allongent et les bandes A sont inchangées. Outre l'élongation des éléments contractiles (les sarcomères). on observe également une élongation passive des éléments élastiques pendant l'étirement du muscle. C'es éléments passifs sont principalement les tendons par lesquels se terminent les muscles. Comme on le montrera plus loin de façon détaillée, l'étirement passif des éléments élastiques est un facteur important de la dynamique de la contraction musculaire.
Tension isométrique et longueur. - Le degré d'étirement préalable d'un muscle n'a pas seulement une importance en regard de la tension de repos de ce muscle, mais aussi vis-à-vis de la force qu'il développe pendant sa contraction. Par exemple, si on mesure la tension isométrique, il est possible de modifier la longueur de ce muscle avant chaque excitation en déplaçant ses points d'attache. Lorsqu'on provoque la contraction d'un muscle, après lui avoir fait prendre diverses longueurs, on constate que la force qu'il développe croît avec la longueur, jusqu'à un « maximum isométrique ». La courbe du maximum isométrique est tracée en rouge sur le graphique 17. Pour toute longueur, la force développée par un muscle, comparée à la force qu'il développe au repos, est la différence entre les ordonnées de la courbe de maximum isométrique et de la courbe tension-longueur relativement à la longueur donnée.
http://www.neur-one.fr
[email protected] - 14 - 17/03/2009 14
Le muscle développe sa force maximum, P0, au voisinage de sa longueur de repos 10. La force développée se réduit très peu si le muscle est plus court que 10 quand la contraction commence. Le point d'intersection de la courbe rouge avec l'axe des abscisses sur la figure 17 indique la plus courte longueur à laquelle un muscle peut encore développer une force. Ainsi, dans le cas d'une contraction isotonique, le muscle ne peut pas supporter d'être raccourci à plus de 70% environ de sa longueur de repos. La force développée pendant une contraction isométrique diminue également lorsque le muscle est étiré au-delà de sa longueur de repos. Les courbes de la figure 17 se rapprochent lorsque le muscle atteint environ 1,8 fois sa longueur de repos ; au point où elles se rencontrent, le muscle ne peut développer aucune force supplémentaire par rapport à la force développée au repos. En résumé, le muscle ne peut développer une force nouvelle que lorsque sa longueur varie entre 70 et 180% de sa longueur au repos.
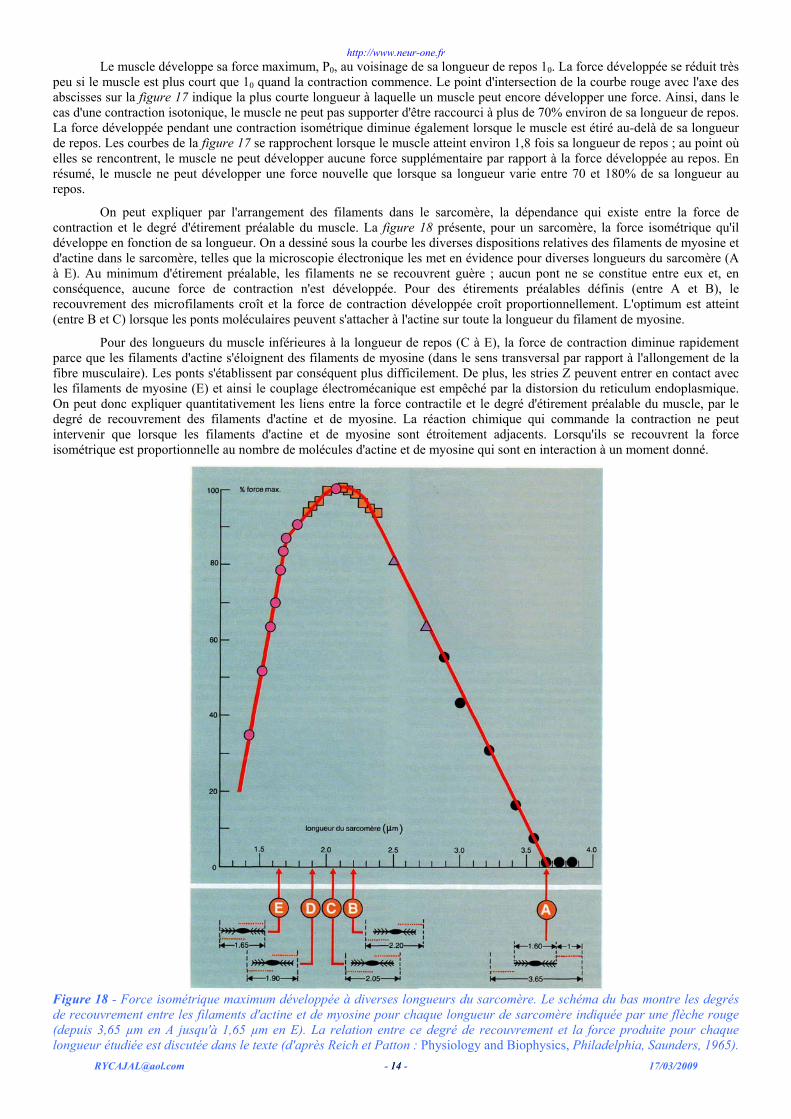
On peut expliquer par l'arrangement des filaments dans le sarcomère, la dépendance qui existe entre la force de contraction et le degré d'étirement préalable du muscle. La figure 18 présente, pour un sarcomère, la force isométrique qu'il développe en fonction de sa longueur. On a dessiné sous la courbe les diverses dispositions relatives des filaments de myosine et d'actine dans le sarcomère, telles que la microscopie électronique les met en évidence pour diverses longueurs du sarcomère (A à E). Au minimum d'étirement préalable, les filaments ne se recouvrent guère ; aucun pont ne se constitue entre eux et, en conséquence, aucune force de contraction n'est développée. Pour des étirements préalables définis (entre A et B), le recouvrement des microfilaments croît et la force de contraction développée croît proportionnellement. L'optimum est atteint (entre B et C) lorsque les ponts moléculaires peuvent s'attacher à l'actine sur toute la longueur du filament de myosine.
Pour des longueurs du muscle inférieures à la longueur de repos (C à E), la force de contraction diminue rapidement parce que les filaments d'actine s'éloignent des filaments de myosine (dans le sens transversal par rapport à l'allongement de la fibre musculaire). Les ponts s'établissent par conséquent plus difficilement. De plus, les stries Z peuvent entrer en contact avec les filaments de myosine (E) et ainsi le couplage électromécanique est empêché par la distorsion du reticulum endoplasmique. On peut donc expliquer quantitativement les liens entre la force contractile et le degré d'étirement préalable du muscle, par le degré de recouvrement des filaments d'actine et de myosine. La réaction chimique qui commande la contraction ne peut intervenir que lorsque les filaments d'actine et de myosine sont étroitement adjacents. Lorsqu'ils se recouvrent la force isométrique est proportionnelle au nombre de molécules d'actine et de myosine qui sont en interaction à un moment donné.
Figure 18 - Force isométrique maximum développée à diverses longueurs du sarcomère. Le schéma du bas montre les degrés de recouvrement entre les filaments d'actine et de myosine pour chaque longueur de sarcomère indiquée par une flèche rouge (depuis 3,65 µm en A jusqu'à 1,65 µm en E). La relation entre ce degré de recouvrement et la force produite pour chaque longueur étudiée est discutée dans le texte (d'après Reich et Patton : Physiology and Biophysics, Philadelphia, Saunders, 1965).
http://www.neur-one.fr
[email protected] - 15 - 17/03/2009 15
Force et vitesse de contraction. - On peut expliquer la dépendance de la force de contraction vis-à-vis du degré d'étirement préalable du muscle en termes de structure de l'appareil contractile. De même, il est possible d'attribuer aux caractéristiques de cet appareil, l'influence exercée par la vitesse de contraction sur l'importance de la force développée.
Pensons à quelques exemples familiers du fonctionnement de notre propre musculature. Nous ne pouvons tirer de nos muscles un maximum de force que si le mouvement en cours ne leur fait pas subir de compression ou seulement une compression faible : par exemple, quand nous poussons ou que nous pressons quelque chose. A l'opposé, nous ne pouvons effectuer des mouvements rapides que lorsque nos muscles ne sont que légèrement sollicités : par exemple, lorsque nous jetons une pierre ou que nous jouons du piano.
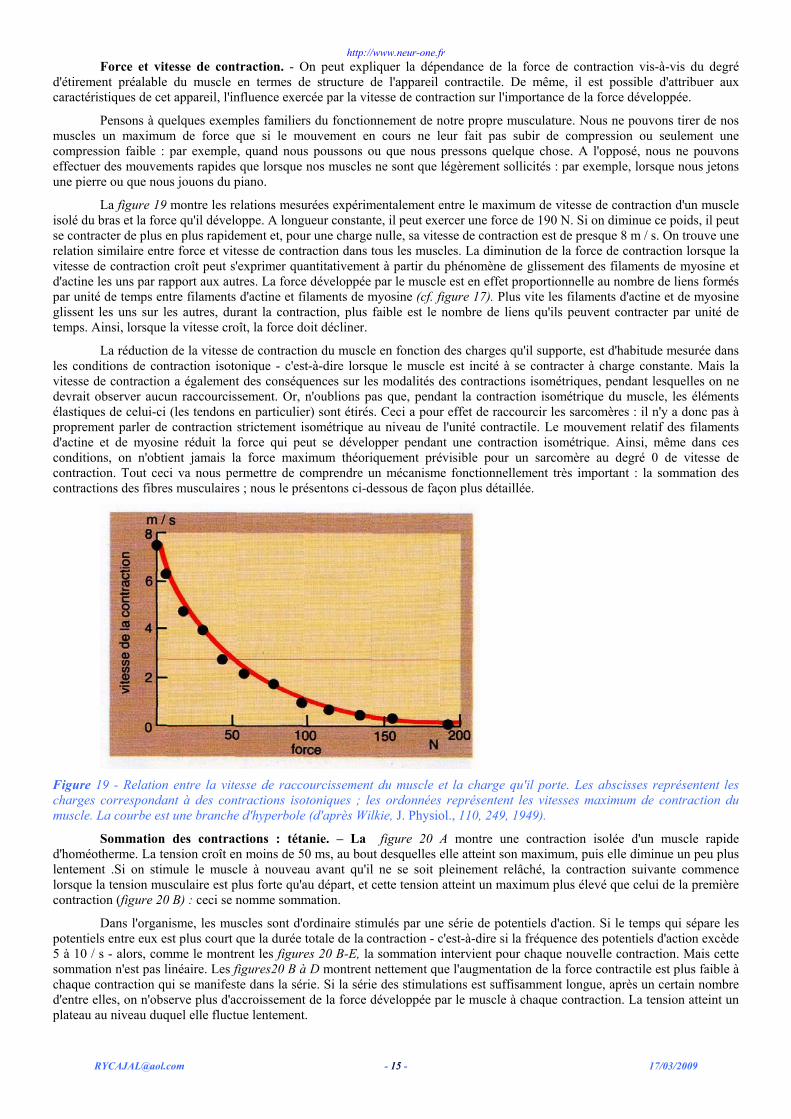
La figure 19 montre les relations mesurées expérimentalement entre le maximum de vitesse de contraction d'un muscle isolé du bras et la force qu'il développe. A longueur constante, il peut exercer une force de 190 N. Si on diminue ce poids, il peut se contracter de plus en plus rapidement et, pour une charge nulle, sa vitesse de contraction est de presque 8 m / s. On trouve une relation similaire entre force et vitesse de contraction dans tous les muscles. La diminution de la force de contraction lorsque la vitesse de contraction croît peut s'exprimer quantitativement à partir du phénomène de glissement des filaments de myosine et d'actine les uns par rapport aux autres. La force développée par le muscle est en effet proportionnelle au nombre de liens formés par unité de temps entre filaments d'actine et filaments de myosine (cf. figure 17). Plus vite les filaments d'actine et de myosine glissent les uns sur les autres, durant la contraction, plus faible est le nombre de liens qu'ils peuvent contracter par unité de temps. Ainsi, lorsque la vitesse croît, la force doit décliner.
La réduction de la vitesse de contraction du muscle en fonction des charges qu'il supporte, est d'habitude mesurée dans les conditions de contraction isotonique - c'est-à-dire lorsque le muscle est incité à se contracter à charge constante. Mais la vitesse de contraction a également des conséquences sur les modalités des contractions isométriques, pendant lesquelles on ne devrait observer aucun raccourcissement. Or, n'oublions pas que, pendant la contraction isométrique du muscle, les éléments élastiques de celui-ci (les tendons en particulier) sont étirés. Ceci a pour effet de raccourcir les sarcomères : il n'y a donc pas à proprement parler de contraction strictement isométrique au niveau de l'unité contractile. Le mouvement relatif des filaments d'actine et de myosine réduit la force qui peut se développer pendant une contraction isométrique. Ainsi, même dans ces conditions, on n'obtient jamais la force maximum théoriquement prévisible pour un sarcomère au degré 0 de vitesse de contraction. Tout ceci va nous permettre de comprendre un mécanisme fonctionnellement très important : la sommation des contractions des fibres musculaires ; nous le présentons ci-dessous de façon plus détaillée.
Figure 19 - Relation entre la vitesse de raccourcissement du muscle et la charge qu'il porte. Les abscisses représentent les charges correspondant à des contractions isotoniques ; les ordonnées représentent les vitesses maximum de contraction du muscle. La courbe est une branche d'hyperbole (d'après Wilkie, J. Physiol., 110, 249, 1949).
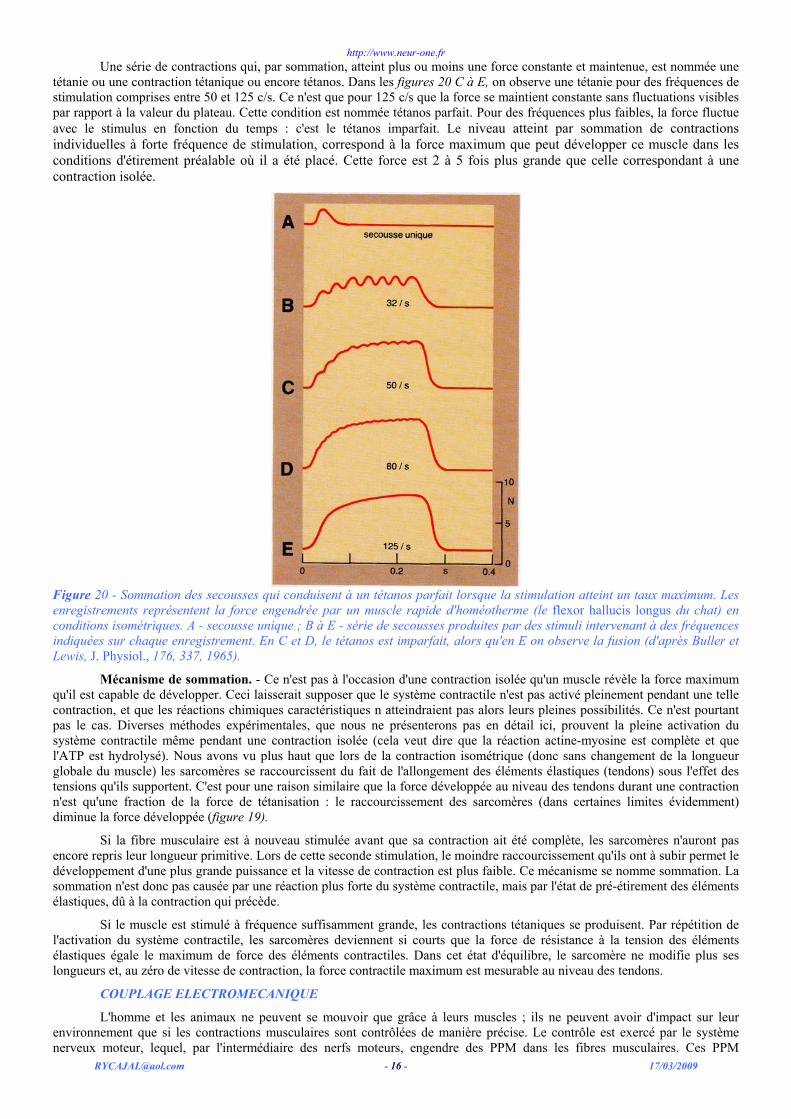
Sommation des contractions : tétanie. – La figure 20 A montre une contraction isolée d'un muscle rapide d'homéotherme. La tension croît en moins de 50 ms, au bout desquelles elle atteint son maximum, puis elle diminue un peu plus lentement .Si on stimule le muscle à nouveau avant qu'il ne se soit pleinement relâché, la contraction suivante commence lorsque la tension musculaire est plus forte qu'au départ, et cette tension atteint un maximum plus élevé que celui de la première contraction (figure 20 B) : ceci se nomme sommation.
Dans l'organisme, les muscles sont d'ordinaire stimulés par une série de potentiels d'action. Si le temps qui sépare les potentiels entre eux est plus court que la durée totale de la contraction - c'est-à-dire si la fréquence des potentiels d'action excède 5 à 10 / s - alors, comme le montrent les figures 20 B-E, la sommation intervient pour chaque nouvelle contraction. Mais cette sommation n'est pas linéaire. Les figures20 B à D montrent nettement que l'augmentation de la force contractile est plus faible à chaque contraction qui se manifeste dans la série. Si la série des stimulations est suffisamment longue, après un certain nombre d'entre elles, on n'observe plus d'accroissement de la force développée par le muscle à chaque contraction. La tension atteint un plateau au niveau duquel elle fluctue lentement.
http://www.neur-one.fr
[email protected] - 16 - 17/03/2009 1
Une série de contractions qui, par sommation, atteint plus ou moins une force constante et maintenue, est nommée une tétanie ou une contraction tétanique ou encore tétanos. Dans les figures 20 C à E, on observe une tétanie pour des fréquences de stimulation comprises entre 50 et 125 c/s. Ce n'est que pour 125 c/s que la force se maintient constante sans fluctuations visibles par rapport à la valeur du plateau. Cette condition est nommée tétanos parfait. Pour des fréquences plus faibles, la force fluctue avec le stimulus en fonction du temps : c'est le tétanos imparfait. Le niveau atteint par sommation de contractions individuelles à forte fréquence de stimulation, correspond à la force maximum que peut développer ce muscle dans les conditions d'étirement préalable où il a été placé. Cette force est 2 à 5 fois plus grande que celle correspondant à une contraction isolée.
Figure 20 - Sommation des secousses qui conduisent à un tétanos parfait lorsque la stimulation atteint un taux maximum. Les enregistrements représentent la force engendrée par un muscle rapide d'homéotherme (le flexor hallucis longus du chat) en conditions isométriques. A - secousse unique ; B à E - série de secousses produites par des stimuli intervenant à des fréquences indiquées sur chaque enregistrement. En C et D, le tétanos est imparfait, alors qu'en E on observe la fusion (d'après Buller et Lewis, J. Physiol., 176, 337, 1965).
Mécanisme de sommation. - Ce n'est pas à l'occasion d'une contraction isolée qu'un muscle révèle la force maximum qu'il est capable de développer. Ceci laisserait supposer que le système contractile n'est pas activé pleinement pendant une telle contraction, et que les réactions chimiques caractéristiques n atteindraient pas alors leurs pleines possibilités. Ce n'est pourtant pas le cas. Diverses méthodes expérimentales, que nous ne présenterons pas en détail ici, prouvent la pleine activation du système contractile même pendant une contraction isolée (cela veut dire que la réaction actine-myosine est complète et que l'ATP est hydrolysé). Nous avons vu plus haut que lors de la contraction isométrique (donc sans changement de la longueur globale du muscle) les sarcomères se raccourcissent du fait de l'allongement des éléments élastiques (tendons) sous l'effet des tensions qu'ils supportent. C'est pour une raison similaire que la force développée au niveau des tendons durant une contraction n'est qu'une fraction de la force de tétanisation : le raccourcissement des sarcomères (dans certaines limites évidemment) diminue la force développée (figure 19).
Si la fibre musculaire est à nouveau stimulée avant que sa contraction ait été complète, les sarcomères n'auront pas encore repris leur longueur primitive. Lors de cette seconde stimulation, le moindre raccourcissement qu'ils ont à subir permet le développement d'une plus grande puissance et la vitesse de contraction est plus faible. Ce mécanisme se nomme sommation. La sommation n'est donc pas causée par une réaction plus forte du système contractile, mais par l'état de pré-étirement des éléments élastiques, dû à la contraction qui précède.
Si le muscle est stimulé à fréquence suffisamment grande, les contractions tétaniques se produisent. Par répétition de l'activation du système contractile, les sarcomères deviennent si courts que la force de résistance à la tension des éléments élastiques égale le maximum de force des éléments contractiles. Dans cet état d'équilibre, le sarcomère ne modifie plus ses longueurs et, au zéro de vitesse de contraction, la force contractile maximum est mesurable au niveau des tendons.
COUPLAGE ELECTROMECANIQUE
L'homme et les animaux ne peuvent se mouvoir que grâce à leurs muscles ; ils ne peuvent avoir d'impact sur leur environnement que si les contractions musculaires sont contrôlées de manière précise. Le contrôle est exercé par le système nerveux moteur, lequel, par l'intermédiaire des nerfs moteurs, engendre des PPM dans les fibres musculaires. Ces PPM
6
http://www.neur-one.fr
[email protected] - 17 - 17/03/2009 1
engendrent des potentiels d'action qui sont propagés le long des fibres musculaires. L'excitation de la membrane est suivie d'une contraction de la fibre. Une modification du potentiel de membrane contrôle ainsi la réaction des protéines contractiles du muscle ; ce phénomène se nomme couplage électromécanique ; nous allons le décrire en détails.
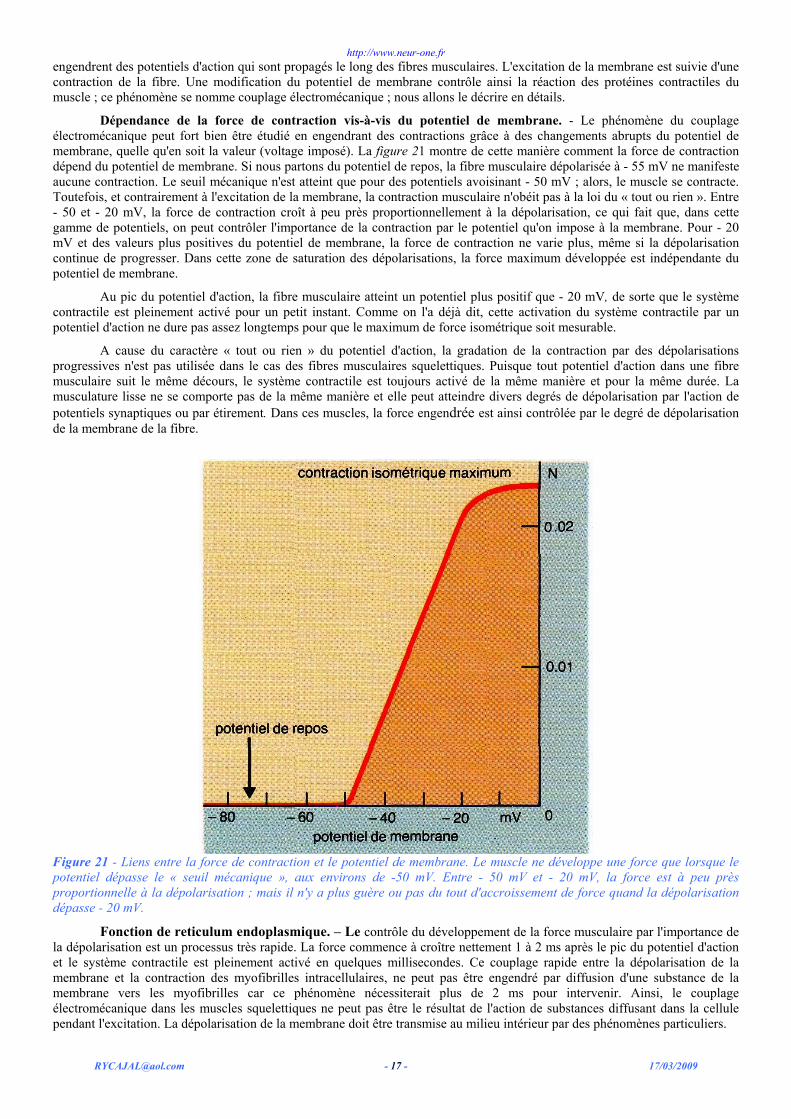
Dépendance de la force de contraction vis-à-vis du potentiel de membrane. - Le phénomène du couplage électromécanique peut fort bien être étudié en engendrant des contractions grâce à des changements abrupts du potentiel de membrane, quelle qu'en soit la valeur (voltage imposé). La figure 21 montre de cette manière comment la force de contraction dépend du potentiel de membrane. Si nous partons du potentiel de repos, la fibre musculaire dépolarisée à - 55 mV ne manifeste aucune contraction. Le seuil mécanique n'est atteint que pour des potentiels avoisinant - 50 mV ; alors, le muscle se contracte. Toutefois, et contrairement à l'excitation de la membrane, la contraction musculaire n'obéit pas à la loi du « tout ou rien ». Entre - 50 et - 20 mV, la force de contraction croît à peu près proportionnellement à la dépolarisation, ce qui fait que, dans cette gamme de potentiels, on peut contrôler l'importance de la contraction par le potentiel qu'on impose à la membrane. Pour - 20 mV et des valeurs plus positives du potentiel de membrane, la force de contraction ne varie plus, même si la dépolarisation continue de progresser. Dans cette zone de saturation des dépolarisations, la force maximum développée est indépendante du potentiel de membrane.
Au pic du potentiel d'action, la fibre musculaire atteint un potentiel plus positif que - 20 mV, de sorte que le système contractile est pleinement activé pour un petit instant. Comme on l'a déjà dit, cette activation du système contractile par un potentiel d'action ne dure pas assez longtemps pour que le maximum de force isométrique soit mesurable.
A cause du caractère « tout ou rien » du potentiel d'action, la gradation de la contraction par des dépolarisations progressives n'est pas utilisée dans le cas des fibres musculaires squelettiques. Puisque tout potentiel d'action dans une fibre musculaire suit le même décours, le système contractile est toujours activé de la même manière et pour la même durée. La musculature lisse ne se comporte pas de la même manière et elle peut atteindre divers degrés de dépolarisation par l'action de potentiels synaptiques ou par étirement. Dans ces muscles, la force engendrée est ainsi contrôlée par le degré de dépolarisation de la membrane de la fibre.
Figure 21 - Liens entre la force de contraction et le potentiel de membrane. Le muscle ne développe une force que lorsque le potentiel dépasse le « seuil mécanique », aux environs de -50 mV. Entre - 50 mV et - 20 mV, la force est à peu près proportionnelle à la dépolarisation ; mais il n'y a plus guère ou pas du tout d'accroissement de force quand la dépolarisation dépasse - 20 mV.
Fonction de reticulum endoplasmique. – Le contrôle du développement de la force musculaire par l'importance de la dépolarisation est un processus très rapide. La force commence à croître nettement 1 à 2 ms après le pic du potentiel d'action et le système contractile est pleinement activé en quelques millisecondes. Ce couplage rapide entre la dépolarisation de la membrane et la contraction des myofibrilles intracellulaires, ne peut pas être engendré par diffusion d'une substance de la membrane vers les myofibrilles car ce phénomène nécessiterait plus de 2 ms pour intervenir. Ainsi, le couplage électromécanique dans les muscles squelettiques ne peut pas être le résultat de l'action de substances diffusant dans la cellule pendant l'excitation. La dépolarisation de la membrane doit être transmise au milieu intérieur par des phénomènes particuliers.
7

http://www.neur-one.fr
[email protected] - 18 - 17/03/2009 18
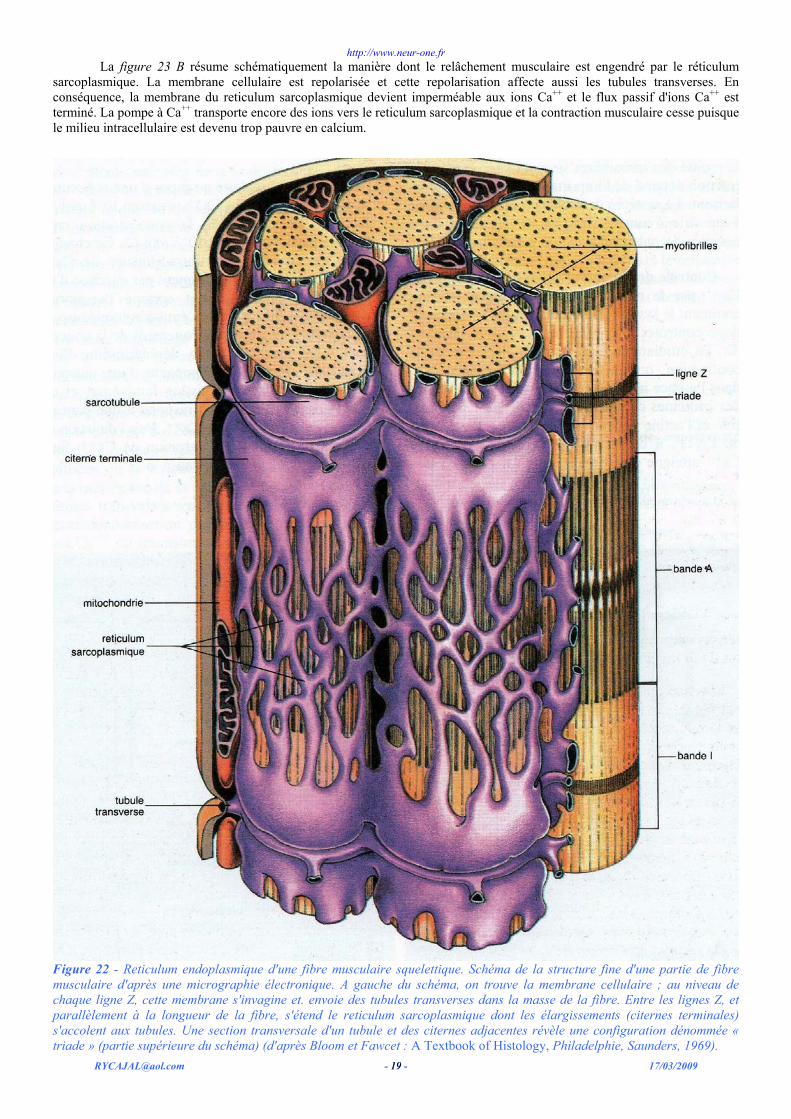
Une structure complexe spéciale se développe dans le cas de fibres assez épaisses comme les fibres musculaires squelettiques. Cette structure assure le couplage rapide de la dépolarisation membranaire et de la contraction ; elle provient d'une différenciation du reticulum endoplasmique. Celui-ci comporte deux systèmes de cavités ou « tubes » occupant la fibre musculaire. Sur la droite de cette figure, on voit la membrane cellulaire. A intervalles réguliers, de fins tubes, les tubules transverses, s'invaginent dans la cellule à partir de la membrane cellulaire externe. Ces tubules s'enfoncent profondément au niveau des stries Z. Le fait que les tubules soient des invaginations de la paroi cellulaire fait qu'ils sont en connexion directe avec l'espace extracellulaire.
Un second système tubulaire, le reticulum sar-coplasmique, s'accole aux tubules transverses (figure 22). Ce second système se place perpendiculairement aux tubules et parallèlement aux filaments du sarcomère. Il est ainsi longitudinalement organisé dans la cellule. Le reticulum sarcoplasmique forme un réseau de cavités allant d'une strie Z à l'autre, le long des myofibrilles de chaque sarcomère. Lorsque le reticulum sarcoplasmique s'accole aux tubules transverses, il se dilate en une citerne terminale. Une section transversale au travers d'une telle région révèle une configuration caractéristique dans laquelle on reconnaît un tube mince accompagné de part et d'autre par de gros tubes. Cette configuration constitue une triade (cf. figure 22 à gauche). Il apparaît clairement sur la figure que les tubules transverses et le reticulum sarcoplasmique ne constituent pas un système commun de canaux mais une étroite juxtaposition de canaux.
Fonction des tubules transverses. - Les tubules transverses sont des invaginations de la membrane cellulaire externe et, à ce titre, on peut les considérer comme des extensions de la membrane dans l'espace intracellulaire ainsi que des enclaves d'espace extracellulaire. Il en résulte que toute excitation de la membrane se propage par l'intermédiaire des tubules dans l'intérieur de la cellule. Ainsi, les tubules transverses peuvent conduire rapidement les dépolarisations de la membrane dans l'intimité de la cellule. Ceci permet l'activation rapide du système contractile, même dans les fibres de grand diamètre. Les tubules transverses sont particulièrement développés dans les fibres musculaires épaisses.
Le rôle des tubules transverses dans le couplage électromécanique peut être démontré par une expérience élégante. Si on applique une pipette fine contre la membrane cellulaire, de telle sorte qu'on ne dépolarise que l'orifice d'invagination d'un seul tubule, alors on observe que, seule la moitié des sarcomères se contracte : cette contraction dépend de l'importance du courant dépolarisant. Le système de tubules transverses associé à une strie Z contrôle ainsi les moitiés de sarcomères sises de part et d'autre de cette strie Z.
Contrôle de la concentration intracellulaire de Ca++ par le reticulum sarcoplasmique. - Mais comment le système de tubules transverses peut-il donc contrôler la contraction ? Nous avons déjà dit, en étudiant le mécanisme moléculaire de la contraction, que la concentration de l'ion Ca++ dans l'espace intracellulaire contrôle les réactions des protéines contractiles. La myosine et l'actine peuvent se lier grâce à l'hydrolyse de l'ATP, à condition que la concentration de Ca++ atteigne environ 10-5 mol /1. Le reticulum endoplasmique exerce un contrôle de la contraction en réglant les changements rapides de la concentration intracellulaire en Ca++.
La fibre musculaire au repos a une concentration très faible en ions Ca2+ : environ 10-8 mol /1. Pendant la contraction, la concentration croît jusqu'à une valeur de 2.10-5 mol /1. Ce changement de concentration intracellulaire de Ca++ peut être démontré élégamment par injection d'un colorant vital luminescent comme l'aequorine dans la fibre musculaire. L'émission lumineuse de cette substance dépend étroitement de la concentration en ions Ça2+. La dépolarisation d'une fibre ainsi préparée s'accompagne d'une augmentation rapide de son émission lumineuse, et ce, jusqu'à une valeur qui correspond à une concentration de 10-5 mol /1 de Ca++. Puis l'émission de lumière et donc la concentration de Ca++ diminuent rapidement à nouveau.
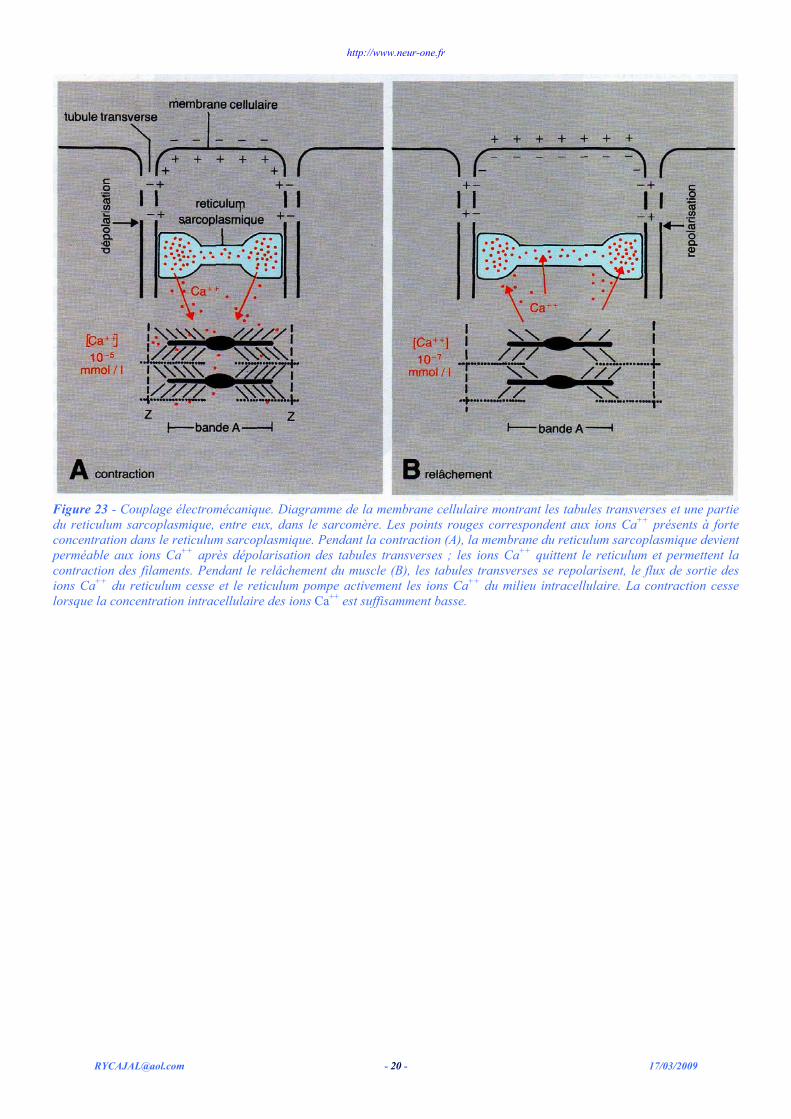
Quelle est la source d'ions Ca++ qui permet cette multiplication rapide de la concentration du milieu intracellulaire par un coefficient de 1 000 ? On a décelé une très forte concentration d'ions Ca++ dans les citernes terminales du reticulum sarcoplasmique du muscle au repos, ces citernes qui avoisinent les tubules transverses figure 22). On pense que, lorsque les tubules sont dépolarisés, ils agissent par un moyen encore inconnu sur le reticulum sarcoplasmique en accroissant transitoirement la conductance de ses membranes à l'ion Ca++. Le très important gradient cause un passage des ions Ca++ du reticulum vers le milieu intracellulaire. Ceci accroît la concentration de ce milieu en Ca++ et permet l'induction de la réaction myosine, actine, ATP. Le couplage électromécanique est ainsi effectué sous le contrôle des concentrations des ions Ca++. Les diverses étapes de ce phénomène sont schématisées sur la figure 23 A. La dépolarisation de la membrane de la cellule est transmise par les tubules transverses vers l'intérieur de la cellule. Cette dépolarisation provoque la libération des ions Ca++ du reticulum sarcoplasmique. Les ions Ca++ diffusent vers les myofilaments adjacents où ils rendent possibles les réactions qui engendrent la contraction.
Lorsque la membrane se repolarise, la concentration du Ca++ intracellulaire chute brutalement et le relâchement commence. La chute rapide de la concentration intracellulaire de Ca++ n'est pas due principalement au fait que le reticulum sarcoplasmique cesse de libérer du calcium. Au contraire, le reticulum reprend activement des ions Ca++ à l'intérieur de la cellule. Une pompe ionique occupe la membrane du reticulum sarcoplasmique. En utilisant l'énergie métabolique, cette pompe transporte à nouveau le Ca++ vers les citernes du reticulum à rencontre du gradient membranaire. La pompe Ca++ du reticulum sarcoplasmique opère à peu près comme la pompe Na+ dans les membranes du neurone, des fibres musculaires et d'autres cellules. C'est grâce à cette pompe que la concentration du Ca++ libre dans le milieu intracellulaire se maintient à un faible niveau (10-8 mol /1) au repos, et que le reticulum peut stocker de fortes concentrations de cet ion. La pompe joue un rôle dans la chute rapide de la concentration en Ca++ à la fin de la phase de dépolarisation. La plupart des ions Ca++ présents alors dans le milieu intracellulaire sont rapidement repris et rejetés dans le reticulum sarcoplasmique par cette pompe.
http://www.neur-one.fr
[email protected] - 19 - 17/03/2009 19
La figure 23 B résume schématiquement la manière dont le relâchement musculaire est engendré par le réticulum sarcoplasmique. La membrane cellulaire est repolarisée et cette repolarisation affecte aussi les tubules transverses. En conséquence, la membrane du reticulum sarcoplasmique devient imperméable aux ions Ca++ et le flux passif d'ions Ca++ est terminé. La pompe à Ca++ transporte encore des ions vers le reticulum sarcoplasmique et la contraction musculaire cesse puisque le milieu intracellulaire est devenu trop pauvre en calcium.
Figure 22 - Reticulum endoplasmique d'une fibre musculaire squelettique. Schéma de la structure fine d'une partie de fibre musculaire d'après une micrographie électronique. A gauche du schéma, on trouve la membrane cellulaire ; au niveau de chaque ligne Z, cette membrane s'invagine et. envoie des tubules transverses dans la masse de la fibre. Entre les lignes Z, et parallèlement à la longueur de la fibre, s'étend le reticulum sarcoplasmique dont les élargissements (citernes terminales) s'accolent aux tubules. Une section transversale d'un tubule et des citernes adjacentes révèle une configuration dénommée « triade » (partie supérieure du schéma) (d'après Bloom et Fawcet : A Textbook of Histology, Philadelphie, Saunders, 1969).
http://www.neur-one.fr
[email protected] - 20 - 17/03/2009 20
Figure 23 - Couplage électromécanique. Diagramme de la membrane cellulaire montrant les tabules transverses et une partie du reticulum sarcoplasmique, entre eux, dans le sarcomère. Les points rouges correspondent aux ions Ca++ présents à forte concentration dans le reticulum sarcoplasmique. Pendant la contraction (A), la membrane du reticulum sarcoplasmique devient perméable aux ions Ca++ après dépolarisation des tabules transverses ; les ions Ca++ quittent le reticulum et permettent la contraction des filaments. Pendant le relâchement du muscle (B), les tabules transverses se repolarisent, le flux de sortie des ions Ca++ du reticulum cesse et le reticulum pompe activement les ions Ca++ du milieu intracellulaire. La contraction cesse lorsque la concentration intracellulaire des ions Ca++ est suffisamment basse.