Embed Size (px)
Citation preview

Netrin-1 is a survival factor during commissuralneuron navigationCeline Furne*†, Nicolas Rama*†, Veronique Corset†, Alain Chedotal‡§, and Patrick Mehlen†¶
†Apoptosis, Cancer, and Development Laboratory, Equipe labellisee ‘‘La Ligue,’’ Centre Leon Berard and Unite Mixte de Recherche 5238, Centre National dela Recherche Scientifique, Universite de Lyon, 69008 Lyon, France; ‡Unite Mixte de Recherche S592, Institut National de la Sante et de la RechercheMedicale, Institut de la Vision, Universite Pierre et Marie Curie, 17 rue Moreau, 75012 Paris, France; and §Assistance Publique - Hopitaux de Paris, GroupeHospitalier Pitie-Salpetriere, Federation de Neurologie, Paris, F-75013 France
Edited by N. M. Le Douarin, College de France, Paris, France, and approved June 26, 2008 (received for review April 15, 2008)
DCC (Deleted in Colorectal Cancer) is a putative tumor suppressorwhose expression is lost in numerous cancers and whose tumorsuppressor activity appears to be dependent on its ability to triggerapoptosis when disengaged by its ligand netrin-1. In this sense,netrin-1 is a survival factor that controls tumorigenesis. However,netrin-1 is also the prototypical axon guidance cue and has beenshown to orient many neurons or axons, especially commissuralaxons, during spinal cord development. Here we show that ne-trin-1 is not only an attractive cue for developing commissuralaxons but also promotes their survival. In primary neuronal culture,in mice or in chick embryos, netrin-1 inhibits the proapoptoticactivity of DCC in developing commissural neurons. Thus, adequatecommissural neurons navigation requires both the attractive ac-tivity of netrin-1 and the anti-apoptotic function of this cue.
apoptosis � deleted in colorectal cancer � nervous system development �dependence receptor � axon guidance
The development of colonic carcinoma from normal colonicepithelium is associated with the mutation of a specific set of
genes (1). Allelic deletions [loss of heterozygosity (LOH)] onchromosome 18q in 70% of primary colorectal tumors promptedthe search for a tumor suppressor gene at that locus, leading tothe cloning of a putative cell-surface receptor, DCC (Deleted inColorectal Cancer) (2). DCC expression was then shown to bemarkedly reduced in 50% of colorectal tumors. Moreover, theloss of DCC expression is not restricted to colon carcinoma, buthas been observed in many other tumors, suggesting that DCCis a tumor suppressor (3).
However, DCC is also the main component of a receptorcomplex that mediates the activity of the secreted axon-guidancemolecule netrin-1 (4, 5). The key role of DCC and netrin-1 inmediating growth cone extension is supported by a large numberof studies, such as the analysis of DCC and netrin-1 knock-outmice, which display the disorganization of many axonal tracts inthe central nervous system (6).
The link between a tumor suppressor in colorectal cancer anda receptor mediating axon outgrowth and turning remainedunclear until recently. We proposed that the tumor suppressorfunction could be explained by the fact that DCC is a depen-dence receptor (7). DCC is functionally related to other depen-dence receptors such as p75NTR, the common neurotrophinreceptor, RET, Patched, UNC5H, neogenin, ALK, or TrkC(8–13). Such receptors create cellular states of dependence totheir respective ligands by inducing apoptosis when unoccupiedbut inhibiting apoptosis in the presence of their ligands (12). Weand others have shown that the in vitro expression of DCCinduces apoptosis in the absence of netrin-1 (7, 14), while thepresence of netrin-1 blocks this proapoptotic activity (15).
The fact that DCC displays such proapoptotic activity whenunbound to its ligand has led to the hypothesis that DCCnormally functions to kill tumor cells that grow in an inappro-priate context (e.g., local growth in a setting of constant andlimited netrin-1 concentration or growth at a secondary sitewhere there is no netrin-1 expression). Accordingly, mice over-
expressing netrin-1 in the gut show a marked decrease of celldeath in the intestinal epithelium associated with an increasedpredisposition to develop colorectal tumors (16). Thus, DCC maybe viewed as a tumor suppressor that controls tumorigenesis byregulating apoptosis (17, 18) and netrin-1 expression as a selectivesurvival advantage for tumor growth and metastasis (19).
However, it is clear that the main role of netrin-1 in thedeveloping CNS is not survival but axon guidance. The maindefects of netrin-1 and DCC knock-out mice may, to a largeextent, be attributed to a loss of the DCC/netrin-1 axon-guidancesystem. Yet, netrin-1 knock-out mice also display increased celldeath in the developing brainstem (20). Thus, one may wonderwhether netrin-1 in the developing nervous system also shows asurvival activity that may participate to the neuronal navigationactivity of this cue. Here we addressed this question in commis-sural neurons that are the prototype neurons guided by thenetrin-1/DCC pair. We show that in primary cultures, commis-sural neurons die except if netrin-1 is provided and that apoptoticactivity depends on DCC, as primary commissural neurons fromDCC knock-out embryos survive in the absence of netrin-1. Wealso show enhanced apoptosis in developing mouse spinal cordfrom netrin-1 knock-out embryos. Moreover, we show that indeveloping chick neural tube, the DCC/netrin-1 pair acts as apair dependence receptor/ligand as electroporation of DCCtriggers cell death that is blocked by coelectroporation ofnetrin-1 while down-regulation of netrin-1 is associated withincreased cell death.
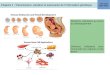
ResultsNetrin-1 Acts as a Survival Factor for Commissural Neurons. In thedorsal spinal cord, commissural axons that express DCC areattracted toward the ventral midline by floor-plate-derivednetrin-1. To assess whether netrin-1 may also display survivalactivity toward these neurons, we cultured commissural neuronsfrom E13 rat or E11.5 mouse embryos. Dorsal spinal cords weredissected out and dissociated and primary cells were then grownin neurobasal medium (Fig. 1A). The cells grown in theseconditions were commissural neurons as they expressed theneuronal marker class III �-tubulin and DCC as shown byimmunohistochemistry [Fig. 1B, specificity of the DCC antibodyshown in supporting information (SI) Fig. S1]. When the ratcommissural neuron cultures were analyzed 16 h after initialplating, about all of the cells were dead (data not shown and Fig.
Author contributions: C.F., N.R., and P.M. designed the research; C.F., N.R., and V.C.performed the research; A.C. contributed new reagents/analytic tools; C.F., N.R., V.C., A.C.,and P.M. analyzed the data; and P.M. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
*C.F. and N.R. contributed equally to this work.
¶To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/cgi/content/full/0803645105/DCSupplemental.
© 2008 by The National Academy of Sciences of the USA
www.pnas.org�cgi�doi�10.1073�pnas.0803645105 PNAS � September 23, 2008 � vol. 105 � no. 38 � 14465–14470
DEV
ELO
PMEN
TAL
BIO
LOG
Y
Dow
nloa
ded
by g
uest
on
Janu
ary
6, 2
020
1C). To confirm this observation, apoptosis was measured bycounting TUNEL positive cells or by quantification of caspaseactivity. As shown in Fig. 1 C–E, commissural neurons grown inneurobasal medium died by apoptosis within the first 16 h.Interestingly, when the culture was performed in the presence ofz-VAD-fmk, a general caspase and cell death inhibitor, 50% ofthe neurons survive after 16 h of culture and this effect wasassociated with a marked decrease in TUNEL- (Fig. 1 C and D)and active caspase-3 (Fig. 1E) -positive neurons. Netrin-1 wasthen added, instead of z-VAD-fmk, to the cultures and as shownin Fig. 1 C–E, netrin-1 presence increased cell survival and was
associated with decreased number of TUNEL- and activecaspase-3-positive cells to a similar extent than z-VAD-fmk (Fig.1 C and E). Similar results were observed when mice commis-sural neurons instead of rat were cultured (data not shown andFig. 1F). Thus, netrin-1 similarly to z-VAD-fmk enhances sur-vival of commissural neurons.
Netrin-1 Acts as a Survival Factor by Inhibiting DCC ProapoptoticActivity. In tumor cells, netrin-1 has been shown to act as asurvival factor inhibiting the proapoptotic activity of DCC orUNC5H receptors (7, 16, 19–21). To determine whether this
control
netrin-1
z-VAD-fmk
C
A
B
hoechst
DCC
-tubulin III
merge
DCC +/+ DCC -/-
net-1
z-VA
D
net-1
z-VA
D
Fco
ntro
l
cont
rol
Inde
x of
cas
pase
-3po
sitiv
e ne
uron
s
00.10.20.30.40.50.60.70.80.9
1*
*
**
**
***
******
: p<0.05: p<0.01: p<0.001
00.10.20.30.40.50.60.70.80.91
Inde
x of
cas
pase
-3po
sitiv
e ne
uron
s
cont
rol
netri
n-1
z-VAD-fm
k
E
00.10.20.30.40.50.60.70.80.9
1
Inde
x of
T
UN
EL
posi
tive
neur
ons
cont
rol
netri
n-1
z-VAD-fm
k
D ** : p < 0.05
Fig. 1. Netrin-1 is a survival factor for commissural neurons. (A) The dorsal spinal cord that contains the commissural neurons is represented. (B) The culturedcells are commissural neurons. Immunostainings using an anti-DCC antibody (A20) (central panel) and an anti-beta-tubulin III antibody (below panel) ofcommissural neurons from E13 rat embryos dorsal spinal cord culture. Merge signal between DCC and beta-tubulin signals is shown (Right) together with ahoechst. Insets, control staining without primary antibodies. (C) Representative photographs of commissural neurons cultured 16 h in the absence (control) orin the presence of netrin-1 (300 ng/ml) or of z-VAD-fmk (20 �M) and stained by TUNEL assay. Right panels are a magnification of left panels. (Scale bars, 50 �M.)(D and E) Histograms representing cell death quantified by numbering cells stained with TUNEL assay (D) or with anti-active caspase-3 (E). Index of cell stainedwith TUNEL or active caspase-3 is shown as the ratio between the number of cells stained in treated conditions (netrin-1 or z-VAD-fmk) to the number of cellstained in control condition. Standard deviations are indicated (n � 3). (F) Commissural neurons from E11.5 mice embryos were cultured 16 h in the presenceor absence of netrin-1 (300 ng/ml) or z-VAD-fmk (20 �M). Cultures were from DCC�/� or DCC-/- embryos as indicated. Histogram representing cell deathmeasured by the number of cells stained with anti-active caspase-3. Index of cell stained with active caspase-3 is shown as the ratio between the number of cellsstained in treated conditions (netrin-1 or z-VAD-fmk) to the number of cell stained in control condition. Standard deviations are indicated (n � 3). *, **, and
***, respectively, indicates a P � 0.05, P � 0.01, and P � 0.001, calculated using a two-sided Mann–Whitney test.
14466 � www.pnas.org�cgi�doi�10.1073�pnas.0803645105 Furne et al.
Dow
nloa
ded
by g
uest
on
Janu
ary
6, 2
020
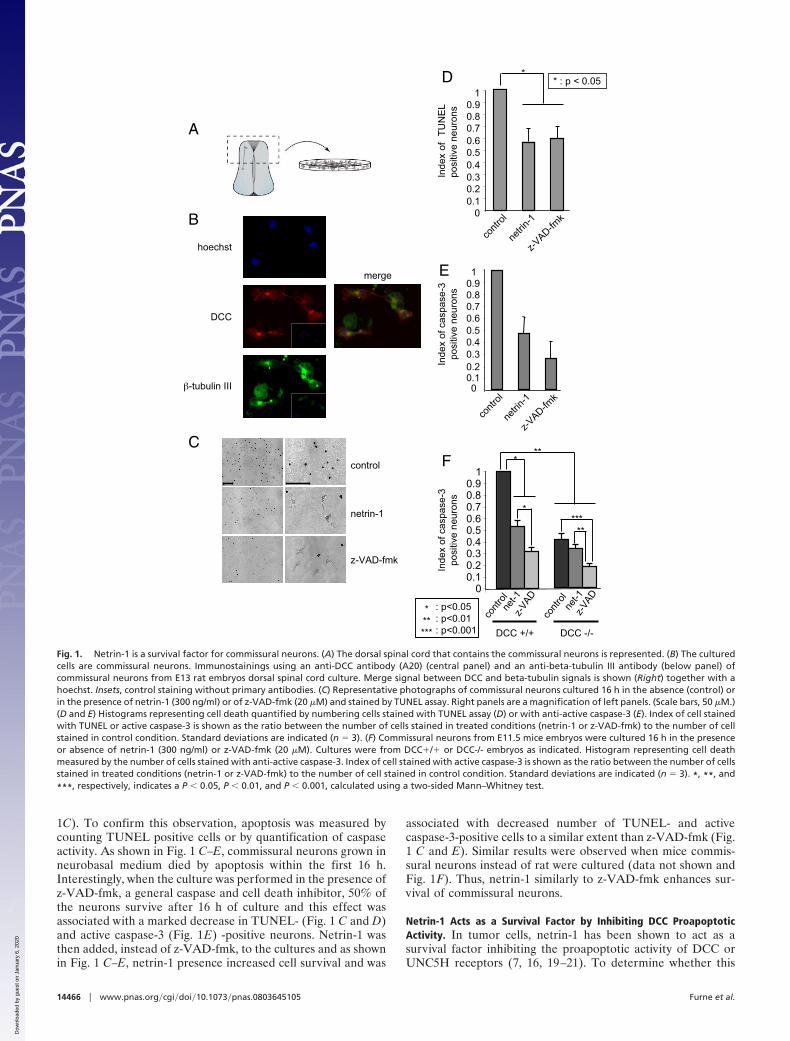
involves a neurotrophic factor-like effect of netrin-1 (i.e., acti-vation upon ligand binding of survival signaling pathways likePI3K or MAPK pathways) or occurs through the inhibition ofDCC proapoptotic activity, we then compared commissuralneuron survival in primary cultures from DCC mice mutantembryos. If netrin-1 behaved as a classic neurotrophic factor, oneshould expect the loss of DCC to be associated with a similareffect on survival instead of a loss of the ligand (i.e., the DCC-/-commissural neurons should die even in the presence of netrin-1at a same level than DCC�/� commissural neurons cultured inthe absence of netrin-1). Alternatively, if netrin-1 were to act byinhibiting DCC-induced apoptosis, the survival of DCC-/- neu-rons should be higher than DCC�/� ones in the absence ofnetrin-1. As shown in Fig. 1F, in the absence of netrin-1treatment, commissural neurons from DCC�/� embryos diedand showed caspase activation, whereas DCC-/- commissuralneurons showed increased survival and decreased caspase acti-vation. Moreover, adding netrin-1 to DCC�/� culture increasedneuronal survival, while it failed to increase the survival ofDCC-/- neurons. Taken together, these data demonstrate thatnetrin-1 is a survival factor that inhibits the proapoptotic activityof the DCC dependence receptor in spinal cord commissuralneurons.
Netrin-1 Controls Cell Survival in the Developing Neural Tube. Thephenotypic analysis of netrin-1 knockout mice revealed pro-found defects in the organization of many axonal tracts, but noincrease in cell death was noted (5). Yet, because the numerationof commissural neurons is not easy and was not done precisely,and because these mutants were hypomorphic mice rather thannull mice, we then decided to assess whether netrin-1 inactivationis associated with increased cell death in the developing spinalcord. An increased caspase-3 activity was observed in spinal cordextracts from netrin-1-/- embryos compared with wild-typeextracts (Fig. 2A). Interestingly, this increased caspase-3 activityis specific for the spinal cord as when a similar caspase activitymeasurement was performed in brain rather than spinal cord, nosimilar increase of caspase activity was observed in netrin-1mutant mice. Moreover, this increased caspase activity is asso-ciated with an increased number of apoptotic (i.e., TUNEL-positive cells) in the spinal cord of E13.5 of netrin-1 mutantembryos (Fig. 2 B and C). These TUNEL-positive cells moreovercolocalize with DCC staining (Fig. 2D), further supporting theview of commissural neurons death. Thus, netrin-1 appears toinhibit apoptotic cell death in the developing spinal cord andmore specifically in commissural neurons.
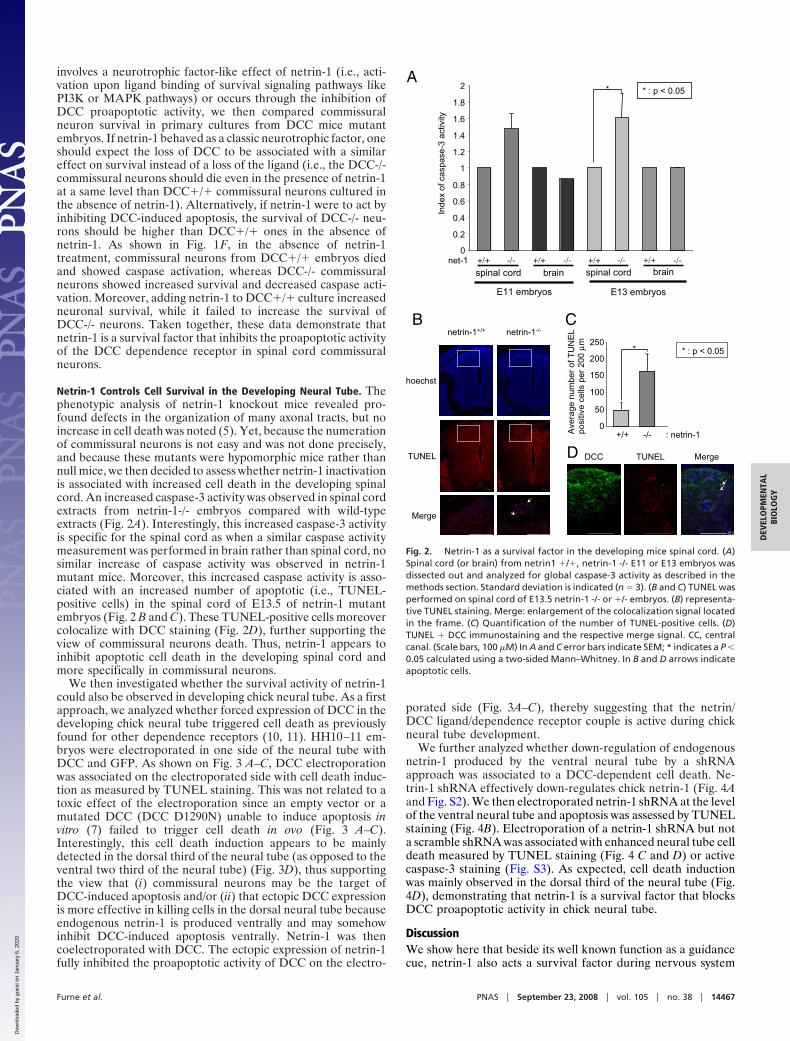
We then investigated whether the survival activity of netrin-1could also be observed in developing chick neural tube. As a firstapproach, we analyzed whether forced expression of DCC in thedeveloping chick neural tube triggered cell death as previouslyfound for other dependence receptors (10, 11). HH10–11 em-bryos were electroporated in one side of the neural tube withDCC and GFP. As shown on Fig. 3 A–C, DCC electroporationwas associated on the electroporated side with cell death induc-tion as measured by TUNEL staining. This was not related to atoxic effect of the electroporation since an empty vector or amutated DCC (DCC D1290N) unable to induce apoptosis invitro (7) failed to trigger cell death in ovo (Fig. 3 A–C).Interestingly, this cell death induction appears to be mainlydetected in the dorsal third of the neural tube (as opposed to theventral two third of the neural tube) (Fig. 3D), thus supportingthe view that (i) commissural neurons may be the target ofDCC-induced apoptosis and/or (ii) that ectopic DCC expressionis more effective in killing cells in the dorsal neural tube becauseendogenous netrin-1 is produced ventrally and may somehowinhibit DCC-induced apoptosis ventrally. Netrin-1 was thencoelectroporated with DCC. The ectopic expression of netrin-1fully inhibited the proapoptotic activity of DCC on the electro-
porated side (Fig. 3A–C), thereby suggesting that the netrin/DCC ligand/dependence receptor couple is active during chickneural tube development.
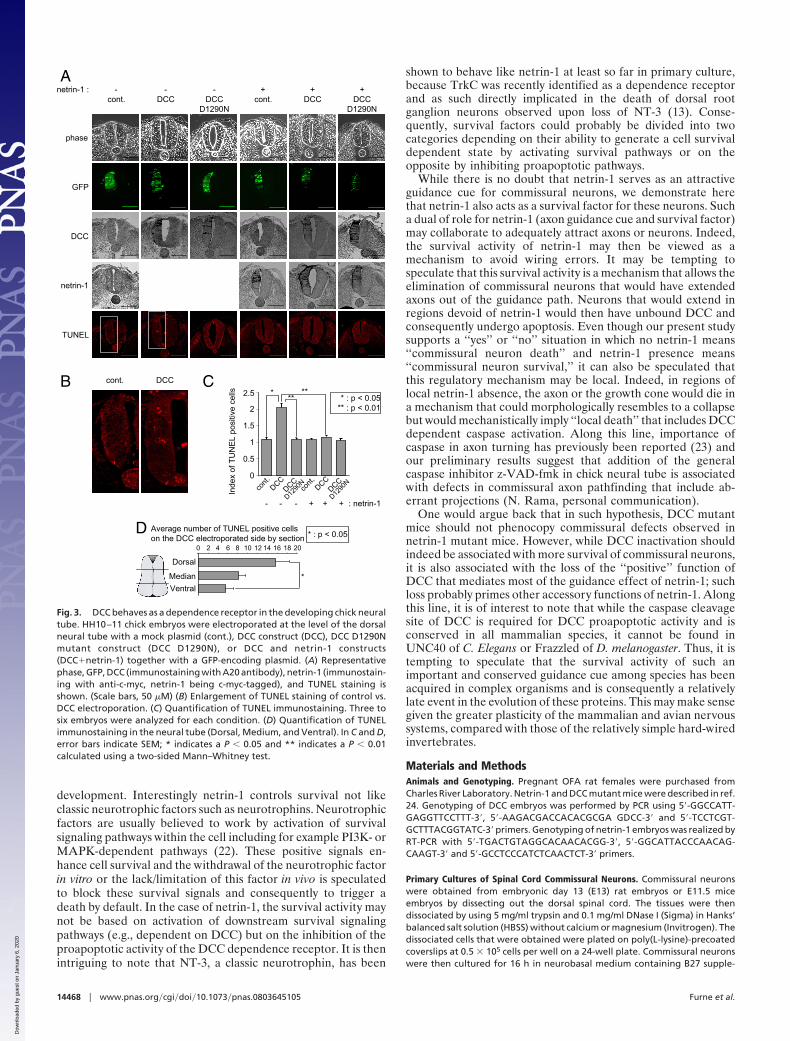
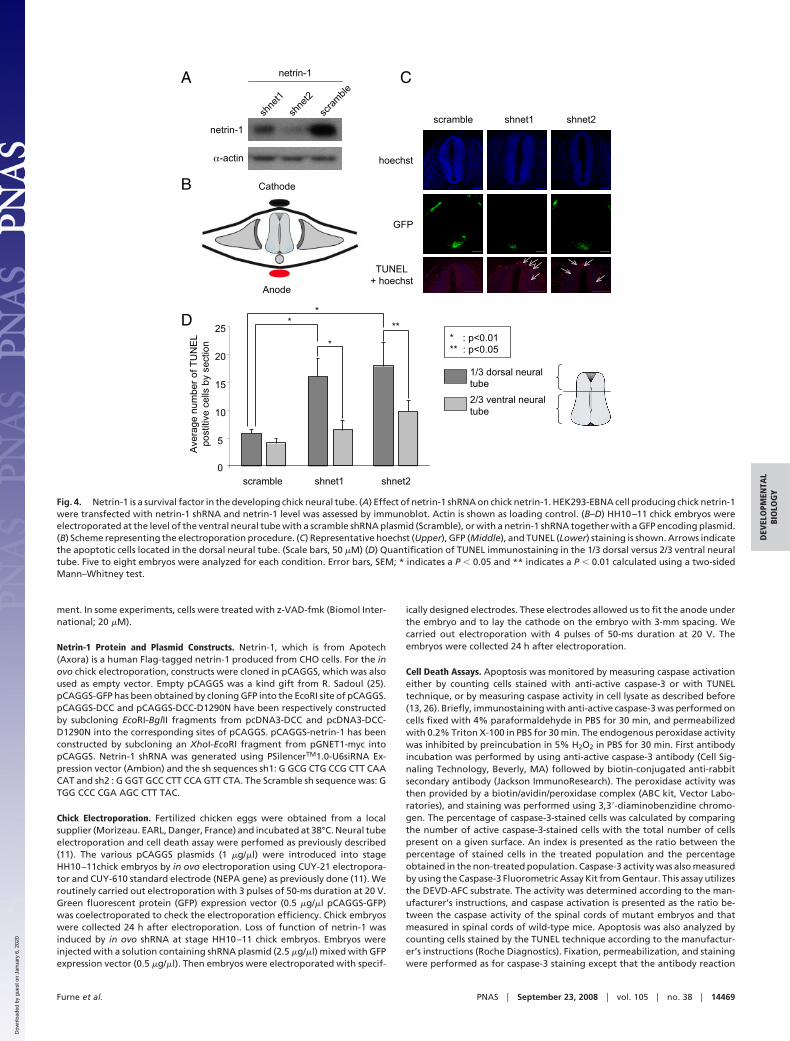
We further analyzed whether down-regulation of endogenousnetrin-1 produced by the ventral neural tube by a shRNAapproach was associated to a DCC-dependent cell death. Ne-trin-1 shRNA effectively down-regulates chick netrin-1 (Fig. 4Aand Fig. S2). We then electroporated netrin-1 shRNA at the levelof the ventral neural tube and apoptosis was assessed by TUNELstaining (Fig. 4B). Electroporation of a netrin-1 shRNA but nota scramble shRNA was associated with enhanced neural tube celldeath measured by TUNEL staining (Fig. 4 C and D) or activecaspase-3 staining (Fig. S3). As expected, cell death inductionwas mainly observed in the dorsal third of the neural tube (Fig.4D), demonstrating that netrin-1 is a survival factor that blocksDCC proapoptotic activity in chick neural tube.
DiscussionWe show here that beside its well known function as a guidancecue, netrin-1 also acts a survival factor during nervous system
Bnetrin-1+/+ netrin-1-/-
hoechst
TUNEL
Merge
C
Ave
rage
num
ber
of T
UN
EL
posi
tive
cells
per
200
µm
+/+ -/- : netrin-10
50
100
150
200
250* * : p < 0.05
A
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
E11 embryos
spinal cord brain
E13 embryos
spinal cord brain
Inde
x of
cas
pase
-3 a
ctiv
ity
+/+ -/- +/+ -/- +/+ -/- +/+ -/-net-1
*
0
* : p < 0.05
DCC TUNEL MergeD
CC
Fig. 2. Netrin-1 as a survival factor in the developing mice spinal cord. (A)Spinal cord (or brain) from netrin1 �/�, netrin-1 -/- E11 or E13 embryos wasdissected out and analyzed for global caspase-3 activity as described in themethods section. Standard deviation is indicated (n � 3). (B and C) TUNEL wasperformed on spinal cord of E13.5 netrin-1 -/- or �/- embryos. (B) representa-tive TUNEL staining. Merge: enlargement of the colocalization signal locatedin the frame. (C) Quantification of the number of TUNEL-positive cells. (D)TUNEL � DCC immunostaining and the respective merge signal. CC, centralcanal. (Scale bars, 100 �M) In A and C error bars indicate SEM; * indicates a P �0.05 calculated using a two-sided Mann–Whitney. In B and D arrows indicateapoptotic cells.
Furne et al. PNAS � September 23, 2008 � vol. 105 � no. 38 � 14467
DEV
ELO
PMEN
TAL
BIO
LOG
Y
Dow
nloa
ded
by g
uest
on
Janu
ary
6, 2
020
development. Interestingly netrin-1 controls survival not likeclassic neurotrophic factors such as neurotrophins. Neurotrophicfactors are usually believed to work by activation of survivalsignaling pathways within the cell including for example PI3K- orMAPK-dependent pathways (22). These positive signals en-hance cell survival and the withdrawal of the neurotrophic factorin vitro or the lack/limitation of this factor in vivo is speculatedto block these survival signals and consequently to trigger adeath by default. In the case of netrin-1, the survival activity maynot be based on activation of downstream survival signalingpathways (e.g., dependent on DCC) but on the inhibition of theproapoptotic activity of the DCC dependence receptor. It is thenintriguing to note that NT-3, a classic neurotrophin, has been
shown to behave like netrin-1 at least so far in primary culture,because TrkC was recently identified as a dependence receptorand as such directly implicated in the death of dorsal rootganglion neurons observed upon loss of NT-3 (13). Conse-quently, survival factors could probably be divided into twocategories depending on their ability to generate a cell survivaldependent state by activating survival pathways or on theopposite by inhibiting proapoptotic pathways.
While there is no doubt that netrin-1 serves as an attractiveguidance cue for commissural neurons, we demonstrate herethat netrin-1 also acts as a survival factor for these neurons. Sucha dual of role for netrin-1 (axon guidance cue and survival factor)may collaborate to adequately attract axons or neurons. Indeed,the survival activity of netrin-1 may then be viewed as amechanism to avoid wiring errors. It may be tempting tospeculate that this survival activity is a mechanism that allows theelimination of commissural neurons that would have extendedaxons out of the guidance path. Neurons that would extend inregions devoid of netrin-1 would then have unbound DCC andconsequently undergo apoptosis. Even though our present studysupports a ‘‘yes’’ or ‘‘no’’ situation in which no netrin-1 means‘‘commissural neuron death’’ and netrin-1 presence means‘‘commissural neuron survival,’’ it can also be speculated thatthis regulatory mechanism may be local. Indeed, in regions oflocal netrin-1 absence, the axon or the growth cone would die ina mechanism that could morphologically resembles to a collapsebut would mechanistically imply ‘‘local death’’ that includes DCCdependent caspase activation. Along this line, importance ofcaspase in axon turning has previously been reported (23) andour preliminary results suggest that addition of the generalcaspase inhibitor z-VAD-fmk in chick neural tube is associatedwith defects in commissural axon pathfinding that include ab-errant projections (N. Rama, personal communication).
One would argue back that in such hypothesis, DCC mutantmice should not phenocopy commissural defects observed innetrin-1 mutant mice. However, while DCC inactivation shouldindeed be associated with more survival of commissural neurons,it is also associated with the loss of the ‘‘positive’’ function ofDCC that mediates most of the guidance effect of netrin-1; suchloss probably primes other accessory functions of netrin-1. Alongthis line, it is of interest to note that while the caspase cleavagesite of DCC is required for DCC proapoptotic activity and isconserved in all mammalian species, it cannot be found inUNC40 of C. Elegans or Frazzled of D. melanogaster. Thus, it istempting to speculate that the survival activity of such animportant and conserved guidance cue among species has beenacquired in complex organisms and is consequently a relativelylate event in the evolution of these proteins. This may make sensegiven the greater plasticity of the mammalian and avian nervoussystems, compared with those of the relatively simple hard-wiredinvertebrates.
Materials and MethodsAnimals and Genotyping. Pregnant OFA rat females were purchased fromCharles River Laboratory. Netrin-1 and DCC mutant mice were described in ref.24. Genotyping of DCC embryos was performed by PCR using 5�-GGCCATT-GAGGTTCCTTT-3�, 5�-AAGACGACCACACGCGA GDCC-3� and 5�-TCCTCGT-GCTTTACGGTATC-3� primers. Genotyping of netrin-1 embryos was realized byRT-PCR with 5�-TGACTGTAGGCACAACACGG-3�, 5�-GGCATTACCCAACAG-CAAGT-3� and 5�-GCCTCCCATCTCAACTCT-3� primers.
Primary Cultures of Spinal Cord Commissural Neurons. Commissural neuronswere obtained from embryonic day 13 (E13) rat embryos or E11.5 miceembryos by dissecting out the dorsal spinal cord. The tissues were thendissociated by using 5 mg/ml trypsin and 0.1 mg/ml DNase I (Sigma) in Hanks’balanced salt solution (HBSS) without calcium or magnesium (Invitrogen). Thedissociated cells that were obtained were plated on poly(L-lysine)-precoatedcoverslips at 0.5 � 105 cells per well on a 24-well plate. Commissural neuronswere then cultured for 16 h in neurobasal medium containing B27 supple-
A
B cont. DCC
cont. DCC DCCD1290N
cont. DCC DCCD1290N
netrin-1 : - + + +- -
phase
GFP
TUNEL
DCC
netrin-1
C
Inde
x of
TU
NE
L po
sitiv
e ce
lls ** * : p < 0.05** : p < 0.01
- - - + + + : netrin-1
cont
.DCC
DCC
D1290
Nco
nt.
DCC0
0.5
1
1.5
2
2.5 ***
DCC
D1290
N
Average number of TUNEL positive cellson the DCC electroporated side by section
Median
Dorsal
Ventral
0 2 4 6 8 10 12 14 16 18 20
*
* : p < 0.05D
Fig. 3. DCC behaves as a dependence receptor in the developing chick neuraltube. HH10–11 chick embryos were electroporated at the level of the dorsalneural tube with a mock plasmid (cont.), DCC construct (DCC), DCC D1290Nmutant construct (DCC D1290N), or DCC and netrin-1 constructs(DCC�netrin-1) together with a GFP-encoding plasmid. (A) Representativephase, GFP, DCC (immunostaining with A20 antibody), netrin-1 (immunostain-ing with anti-c-myc, netrin-1 being c-myc-tagged), and TUNEL staining isshown. (Scale bars, 50 �M) (B) Enlargement of TUNEL staining of control vs.DCC electroporation. (C) Quantification of TUNEL immunostaining. Three tosix embryos were analyzed for each condition. (D) Quantification of TUNELimmunostaining in the neural tube (Dorsal, Medium, and Ventral). In C and D,error bars indicate SEM; * indicates a P � 0.05 and ** indicates a P � 0.01calculated using a two-sided Mann–Whitney test.
14468 � www.pnas.org�cgi�doi�10.1073�pnas.0803645105 Furne et al.
Dow
nloa
ded
by g
uest
on
Janu
ary
6, 2
020
ment. In some experiments, cells were treated with z-VAD-fmk (Biomol Inter-national; 20 �M).
Netrin-1 Protein and Plasmid Constructs. Netrin-1, which is from Apotech(Axora) is a human Flag-tagged netrin-1 produced from CHO cells. For the inovo chick electroporation, constructs were cloned in pCAGGS, which was alsoused as empty vector. Empty pCAGGS was a kind gift from R. Sadoul (25).pCAGGS-GFP has been obtained by cloning GFP into the EcoRI site of pCAGGS.pCAGGS-DCC and pCAGGS-DCC-D1290N have been respectively constructedby subcloning EcoRI-BglII fragments from pcDNA3-DCC and pcDNA3-DCC-D1290N into the corresponding sites of pCAGGS. pCAGGS-netrin-1 has beenconstructed by subcloning an XhoI-EcoRI fragment from pGNET1-myc intopCAGGS. Netrin-1 shRNA was generated using PSilencerTM1.0-U6siRNA Ex-pression vector (Ambion) and the sh sequences sh1: G GCG CTG CCG CTT CAACAT and sh2 : G GGT GCC CTT CCA GTT CTA. The Scramble sh sequence was: GTGG CCC CGA AGC CTT TAC.
Chick Electroporation. Fertilized chicken eggs were obtained from a localsupplier (Morizeau. EARL, Danger, France) and incubated at 38°C. Neural tubeelectroporation and cell death assay were perfomed as previously described(11). The various pCAGGS plasmids (1 �g/�l) were introduced into stageHH10–11chick embryos by in ovo electroporation using CUY-21 electropora-tor and CUY-610 standard electrode (NEPA gene) as previously done (11). Weroutinely carried out electroporation with 3 pulses of 50-ms duration at 20 V.Green fluorescent protein (GFP) expression vector (0.5 �g/�l pCAGGS-GFP)was coelectroporated to check the electroporation efficiency. Chick embryoswere collected 24 h after electroporation. Loss of function of netrin-1 wasinduced by in ovo shRNA at stage HH10–11 chick embryos. Embryos wereinjected with a solution containing shRNA plasmid (2.5 �g/�l) mixed with GFPexpression vector (0.5 �g/�l). Then embryos were electroporated with specif-
ically designed electrodes. These electrodes allowed us to fit the anode underthe embryo and to lay the cathode on the embryo with 3-mm spacing. Wecarried out electroporation with 4 pulses of 50-ms duration at 20 V. Theembryos were collected 24 h after electroporation.
Cell Death Assays. Apoptosis was monitored by measuring caspase activationeither by counting cells stained with anti-active caspase-3 or with TUNELtechnique, or by measuring caspase activity in cell lysate as described before(13, 26). Briefly, immunostaining with anti-active caspase-3 was performed oncells fixed with 4% paraformaldehyde in PBS for 30 min, and permeabilizedwith 0.2% Triton X-100 in PBS for 30 min. The endogenous peroxidase activitywas inhibited by preincubation in 5% H2O2 in PBS for 30 min. First antibodyincubation was performed by using anti-active caspase-3 antibody (Cell Sig-naling Technology, Beverly, MA) followed by biotin-conjugated anti-rabbitsecondary antibody (Jackson ImmunoResearch). The peroxidase activity wasthen provided by a biotin/avidin/peroxidase complex (ABC kit, Vector Labo-ratories), and staining was performed using 3,3�-diaminobenzidine chromo-gen. The percentage of caspase-3-stained cells was calculated by comparingthe number of active caspase-3-stained cells with the total number of cellspresent on a given surface. An index is presented as the ratio between thepercentage of stained cells in the treated population and the percentageobtained in the non-treated population. Caspase-3 activity was also measuredby using the Caspase-3 Fluorometric Assay Kit from Gentaur. This assay utilizesthe DEVD-AFC substrate. The activity was determined according to the man-ufacturer’s instructions, and caspase activation is presented as the ratio be-tween the caspase activity of the spinal cords of mutant embryos and thatmeasured in spinal cords of wild-type mice. Apoptosis was also analyzed bycounting cells stained by the TUNEL technique according to the manufactur-er’s instructions (Roche Diagnostics). Fixation, permeabilization, and stainingwere performed as for caspase-3 staining except that the antibody reaction
Cathode
Anode
C
B
scramble shnet1 shnet2
hoechst
GFP
TUNEL+ hoechst
A
scra
mble
shne
t1
shne
t2
netrin-1
netrin-1
-actin
D
scramble shnet1 shnet2
Ave
rage
num
ber
of T
UN
EL
post
itive
cel
ls b
y se
ctio
n
* : p<0.01** : p<0.05
1/3 dorsal neural tube
2/3 ventral neural tube
0
5
10
15
20
25
*
****
Fig. 4. Netrin-1 is a survival factor in the developing chick neural tube. (A) Effect of netrin-1 shRNA on chick netrin-1. HEK293-EBNA cell producing chick netrin-1were transfected with netrin-1 shRNA and netrin-1 level was assessed by immunoblot. Actin is shown as loading control. (B–D) HH10–11 chick embryos wereelectroporated at the level of the ventral neural tube with a scramble shRNA plasmid (Scramble), or with a netrin-1 shRNA together with a GFP encoding plasmid.(B) Scheme representing the electroporation procedure. (C) Representative hoechst (Upper), GFP (Middle), and TUNEL (Lower) staining is shown. Arrows indicatethe apoptotic cells located in the dorsal neural tube. (Scale bars, 50 �M) (D) Quantification of TUNEL immunostaining in the 1/3 dorsal versus 2/3 ventral neuraltube. Five to eight embryos were analyzed for each condition. Error bars, SEM; * indicates a P � 0.05 and ** indicates a P � 0.01 calculated using a two-sidedMann–Whitney test.
Furne et al. PNAS � September 23, 2008 � vol. 105 � no. 38 � 14469
DEV
ELO
PMEN
TAL
BIO
LOG
Y
Dow
nloa
ded
by g
uest
on
Janu
ary
6, 2
020

was replaced by cell incubation for 1 h at 37°C with terminal deoxynucleoti-dyltransferase and biotin-dUTP. For Figs. 2, 3, and 4, TUNEL on sections wasperformed as above except that it was revealed with streptavidine coupledCy-3 (Jackson ImmunoResearch).
Cell Death Quantification. For themousespinal cord, serial section20-�msectionswere performed and TUNEL-positive cells located in the spinal cord were countedblinded on 200 �m. For the electroporation of DCC and the coelectroporation ofDCC�netrin-1, the index corresponds to the number of TUNEL-positive cells onthe electroporated side versus the non-electroporated side. For the DCC electro-poration experiment, on each slice the spinal cord was divided into three parts:dorsal, median, and ventral via the use of computer analysis (Axovision software,Zeiss). The number of TUNEL-positive cells was counted blinded and an averagecalculated for each part. For the electroporation of shRNA, chick spinal cord wasdivided in two parts: 1/3 dorsal and 2/3 ventral using the Axovision software. Oneach section, the number of TUNEL-positive cells was counted blinded on eachpart and an average was calculated. In each case, the counting was done in blindby two independent experimenters.
Immunohistolocalisation. DCC and �-tubulin III immunostaining was realizedon mouse commissural neurons fixed and permeabilized as described. Anti-
DCC antibody (DCC-A20) (Santa Cruz) and anti-�-tubulin III (Tuj-1) (AbCam)were detected with Alexa488-Donkey anti-goat antibody (Invitrogen) andcyanin 3 donkey anti-mouse antibody (Jackson ImmunoResearch), respec-tively. The coverslips were mounted in fluoromount (EMS) and sections pho-tographed with a Zeiss Axiovert 200 microscope. To perform DCC immunos-tainings on mouse spinal cord, sections were fixed and permeabilized asdescribed above for TUNEL assay. Anti-DCC antibody (DCC-A20; Santa Cruz)was detected with Alexa488-Donkey anti-Goat (Invitrogen). Sections weretreated with Hoechst (Sigma) and were mounted as described above. DCC andnetrin-1 immunostaining on chick spinal cords was performed with anti-DCCantibody (DCC-A20; Santa Cruz) and anti-c-myc antibody (Sigma), respectively,which were detected with biotinylated-Donkey anti-Goat (Jackson Immu-noResearch) and biotinated-Donkey-anti-mouse (Jackson ImmunoResearch),respectively. The peroxidase activity was revealed as described.
ACKNOWLEDGMENTS. We thank C.Guix for her excellent technical help, H.Bilak for text correction, and L. Pays for helpful discussion. This work wassupported by institutional funds from the Centre National de la RechercheScientifique and by grants from the Ligue Nationale Contre le Cancer, theNational Institute of Health (NIH) and the ANR. CF was supported by a doctoralfellowship from the ARC.
1. Fearon ER, Vogelstein B (1990) A genetic model for colorectal tumorigenesis. Cell61:759–767.
2. Fearon ER, et al. (1990) Identification of a chromosome 18q gene that is altered incolorectal cancers. Science 247:49–56.
3. Mehlen P, Fearon (2004) ER Role of the dependence receptor DCC in colorectal cancerpathogenesis. J Clin Oncol 22:3420–3428.
4. Keino-Masu K, et al. (1996) Deleted in Colorectal Cancer (DCC) encodes a netrinreceptor. Cell 87:175–185.
5. Serafini T, et al. (1996) Netrin-1 is required for commissural axon guidance in thedeveloping vertebrate nervous system. Cell 87:1001–1014.
6. Fazeli A, et al. (1997) Phenotype of mice lacking functional Deleted in colorectal cancer(Dcc) gene. Nature 386:796–804.
7. Mehlen P, Rabizadeh S, Snipas SJ, Assa-Munt N, Salvesen GS, Bredesen DE (1998) TheDCC gene product induces apoptosis by a mechanism requiring receptor proteolysis.Nature 395:801–804.
8. Rabizadeh S, et al. (1993) Induction of apoptosis by the low-affinity NGF receptor.Science 261:345–348.
9. Bordeaux MC, et al. (2000) The RET proto-oncogene induces apoptosis: a novelmechanism for Hirschsprung disease. EMBO J 19:4056–4063.
10. Thibert C, Teillet MA, Lapointe F, Mazelin L, Le Douarin NM, Mehlen P (2003) Inhibitionof neuroepithelial patched-induced apoptosis by sonic hedgehog. Science 301:843–846.
11. Matsunaga E, et al. (2004) RGM and its receptor neogenin regulate neuronal survival.Nat Cell Biol 6:749–755.
12. Mehlen P, Thibert C (2004) Dependence receptors: between life and death. Cell MolLife Sci 61:1854–1866.
13. Tauszig-Delamasure S, et al. (2007) The TrkC receptor induces apoptosis when thedependence receptor notion meets the neurotrophin paradigm. Proc Natl Acad SciUSA 104:13361–13366.
14. Chen YQ, et al. (1999) Induction of apoptosis and G2/M cell cycle arrest by DCC.Oncogene 18:2747–2754.
15. Shin SK, et al. (2007) Epigenetic and genetic alterations in Netrin-1 receptors UNC5Cand DCC in human colon cancer. Gastroenterology 133:1849–1857.
16. Mazelin L, et al. (2004) Netrin-1 controls colorectal tumorigenesis by regulatingapoptosis. Nature 431:80–84.
17. Mehlen P, Llambi F (2005) Role of netrin-1 and netrin-1 dependence receptors incolorectal cancers. Br J Cancer 93:1–6.
18. Grady WM (2007) Making the case for DCC and UNC5C as tumor-suppressor genes inthe colon. Gastroenterology 133:2045–2049.
19. Fitamant J, et al. (2008) Netrin-1 expression confers a selective advantage for tumor cellsurvival in metastatic breast cancer. Proc Natl Acad Sci USA 105:4850–4855.
20. Llambi F, Causeret F, Bloch-Gallego E, Mehlen P (2001) Netrin-1 acts as a survival factorvia its receptors UNC5H and DCC. EMBO J 20:2715–2722.
21. Bernet A, et al. (2007) Inactivation of the UNC5C Netrin-1 receptor is associated withtumor progression in colorectal malignancies. Gastroenterology 133:1840–1848.
22. Yao R, Cooper GM (1995) Requirement for phosphatidylinositol-3 kinase in the pre-vention of apoptosis by nerve growth factor. Science 267:2003–2006.
23. Campbell DS, Holt CE (2003) Apoptotic Pathway and MAPKs Differentially RegulateChemotropic Responses of Retinal Growth Cones. Neuron 37:939–952.
24. Forcet C, et al. (2002) Netrin-1-mediated axon outgrowth requires deleted in colorectalcancer-dependent MAPK activation. Nature 417:443–447.
25. Mahul-Mellier AL, Hemming FJ, Blot B, Fraboulet S, Sadoul R (2006) Alix, making a linkbetween apoptosis-linked gene-2, the endosomal sorting complexes required fortransport, and neuronal death in vivo. J Neurosci 26:542–549.
26. Furne C, Corset V, Herincs Z, Cahuzac N, Hueber AO, Mehlen P (2006) The dependencereceptor DCC requires lipid raft localization for cell death signaling. Proc Natl Acad SciUSA 103:4128–4133.
14470 � www.pnas.org�cgi�doi�10.1073�pnas.0803645105 Furne et al.
Dow
nloa
ded
by g
uest
on
Janu
ary
6, 2
020












![Optical mesoscopy without the scatter: broadband multispectral … · single GFP-labeled neurons within dendritic trees in isolated hippocampi [3]. SPIM has also been able to offer](https://img.pdfslide.fr/doc/110x75/60b4cc937ba1593eee0be699/optical-mesoscopy-without-the-scatter-broadband-multispectral-single-gfp-labeled.jpg)