Embed Size (px)
Citation preview

OSTRACODES DU PLATEAU D OUTH (OCeaN. INDIEN) : REMARQUES SYSTEMATIQUES ET EVOLUTION
DES ENVIRONNEMENTS OCEANIQUES?ROFONDS AU COURS DU CENOZO QUE
CLAUDE G U E R N E T Uniuersitd P. et M. Curie, Laboratoire de Micropaldontologie, Ddpartement de Gdologie s~dimentaire et URA 1315, F-75252 Paris cedex 05.
GUERNET C. 1993. Ostracodes du plateau d'Exmouth (Ocdan Indien) : remarques syst~matiques et dvolution des environnements oc~aniques profonds au cours du Cdnozolque. [Ostracodes of the Exmouth Plateau (Indian Ocean) : systematic observations and deep sea environmental evolution during the Cenozoic]. Ostracoden aus dem Exmouth Plateau (Indischer Ozean) : Bemerkungen tiber die Systematic und pal~loSkological Entwicklung der Ozeantiefe w~lhrend des K~ozoikums. GEOBIOS, '26, 3 : 345-360. 80 .06 .1998.
Manuscrit d~pos~ le 22.04.1992 ; accept~ ddfinitivement le 22.06.1992.
R~]SUM~
Les son dages des sites O.D.P. 762 et 763 (Plateau d~Exmouth, Ocean Indien) permettent de suivre, de FEoc~ne au Pl~istec~ne, l'~volution des associations d'Ostracodes. Celles-ci sont relativement diversifi~es jusqu'au Miocene inf~rieur, elles sont pauvres, en g~n~ral, ~ partir du Miocene moyen-sul~rieur. Une trentaine d'esl~ces sont reconnues, leur r~partition stratigraphique pr~cis~e et certaines d'entres elles, Trachyleberis orientalis, T. sp. 2, Bradleya thomasi, Henryhowella reticulata, etc., qui semblent avoir une vaste extension g~ographique, sont l'objet d'observations syst~matiques. D'autre part, la persistance, au cours du C~nozo'fque, du m&me type d'association g~n~rique et le renouvellement ~chelonn~ des esp~ces indique que l'~volution des conditions physiques bathyales a ~t~, au moins ~ partir de l'Eoc~ne, elle-m~me progressive.
Mots-cl~s : Ostracodes, C~nozoique, Ocean indien, Pal~oenvironnement, Pal~og~ographie, Stratigraphie.
ABSTRACT
Sites O.D.P. 762 and 763 (Exmouth Plateau, Indian Ocean) allow the opportunities to study, from Eocene to Pleistocene, the evolution of the ostracode associations which are relatively diverse up to the early Miocene and generally poor in the middle-late Miocene to Pleistocene interval. More than thirty species are present ; their stratigraphical distribution is summarized and some species, such as, Trachyleberis orientalis, T. sp. 2, Bradleya thomasi, Henryhowella reticulata, etc., which seem to have been widely distributed are described or commented upon. Persistence of the same type of generic association and progressivity of the specific renewal demonstrate that evolution of bathyal environmental conditions were progressive through the Cainozoic.
Key-words : Ostracodes, Cenozoic, Indian Ocean, Paleoenvironnement, Paleogeography, Stratigraphy.
ZUSAMMENFASSUNG
Dutch probebohrungen von site O.D.P. 762 und 763 ("Exmouth Plateau", Indischer Ozean) konnte die Entwicl~lung der Ostrakodenassoziationen von Eoz~n bis Pleistoz~in verfolgt werden. Sie sind relatif abwechslungsreich bis zum unteren Mioz~in und sie sind general arm ab mittlerem-oberem Mioz~in bis zum Pleistoz~in. Mehr als dreissig Arten wurden erkannt, ihre stratigraphische Verfeilung bestimmt und einige unter ihnen, Trachyleberis orientalis, T. sp. 2, Bradleya thomasi, HenryhoweUa reticulata.., die eine breite geographische Verbreitung zu haben scheinen, wurden beschrieben oder diskutiert. Andererseits, w~.rend des IZdinozoikums, Durchl~iufer vom selben Typus der Artenvergeselschaftung und die fortschreitende Ernenerung der Arten zeigt, dass die bathyale, physikalische Verh~iltnisse shad gleich progressiv mindestens seit dem EozBn.
Schlfisselw6rter : Ostrakoden, K~nozoikum, Indscher Oz~an, Pal~ioSkologie, Pal~iogeographie, Stratigraphie.

346
INTRODUCTION
Les sondages ODP du leg 122 prdsentent des caract6ristiques particuli~rement intdressantes, a priori, pour r6tude des microfossiles d'origine benthique parmi lesquels les ostracodes, t6moins privi l6gi6s de l '6volut ion des envi . r o n n e m e n t s profonds . D'une part, ces sondages furent implant6s en domaine bathyal (- 1360 m et - 1367 m, respectivement, pour les deux sites s61ectionn6s, les sites 762 et 763) mais aussi en bordure oc6anique d'un large plateau probable- ment tr~s ancien - le plateau Exmouth - donc l 'abri des appo r t s n6r i t iques (ceux-ci, par la pollution biologique qu'ils entra/nent, rendent plus d~licates les interpr6tations pal6odco- logiques). D'autre part, les sondages dtudi6s ici t r a v e r s e n t la quas i total i t~ (forages 762B et 762C) ou une p a t t i e i m p o r t a n t e (forages 763A et B) de la s6rie c6nozoique. Les forage 762C, 763B et 763C atteignent le Cr6tacd infdrieur mais des lacunes ou la raretd des benthiques, ne nous ont pas permis, ~ Damotte et ~ moi-m6me, de suivre de fa~on continue l'6volution des ostracodes du Cr6tacd ~ l'Eoc~ne. Cependant, les esp~ces maestrichtiennes ou campaniennes obser- vdes dans les sondages ou d6crites ~ terre dans le bassin de Carnarvon (Bate 1972) et les associa- tions g6ndriques elles-m6mes sont diff6rentes de celles du C6nozo~'que.
Les ostracodes du plateau d'Exmouth ont dtd robjet des publications de Crasquin-Soleau, Ddp~- che& Galbrun (1990) et de Ddp~che & Crasquin- Soleau (1992) pour le Trias, de Damotte (1992) pour le Cr6tac6 et de Guernet & Galbrun (1992)
pour le Cdnozoique, les dchantillons ayant dtd pr61evds par Galbrun ~ bord du Joides Resolu- tion. Darts la note consacrde au Cdnozo~que, une r~partition stratigraphique des principales esp~- ce~ est dtablie, notre conclusion dtant que "there is, at least on the Exmouth Plateau, a remar- kable permanence throughout the Cenozoic of the bathyal physical conditions, particularly bathy- metric and thermic". Cependant, bien que nous ayons figurd certaines esp~ces du leg 122, aucune 6tude syst6matique ne rut alors entreprise, dtude qui ici doit nous permettre de proposer une 6bauche d'6chelle stratigraphique bas~e sur rutilisation des ostracodes.
C A D R E G I ~ O G R A P H I Q U E E T GI~OLOGI- QUE (d'apr~s Haq, yon R a d et al. 1990)
Le plateau sous-marin d'Exmouth est situd en bordure de l'Oc~an Indien, ~ 250 km environ de la cSte NW de l'Australie (Fig. 1). Sa surface culmine aux alentours de - 1000 m. Les sites 762 et 763 sont eux-m~mes sur la marge oc6anique (NW) de ce plateau, loin des influences cSti~res.
Le plateau d'Exmouth et son prolongement vers le NE, le plateau de Wombat, constituent aujour- d'nui deux horst s~pards par le graben de Wombat. L'existence de ces structures est li6e aux Amlncissements crustaux et aux mouvements verticaux qui affectent la marge continentale australienne au cours du M~sozoique. Le Trias et le Jurassique sont reprdsent6s par une sdrie s6dimentaire paralique (fluviodelta/que, ddtriti- que ou houill~re, ou bien ndritique, principa- lement carbonat~e). Au Jurassique sup~rieur et
Figure 1 - Carte mon t r an t la position des sondages O.D.P. (Leg 122) sur le p la teau d 'Exmouth (sites 762 et 763). Location map showing O.D.P. 122 drill sites on the Exmouth Plateau (sites 762 and 763).
347
que ou houill~re, ou bien n~ritique, principa- lement carbonat~e). Au Jurassique sup~rieur et ,~ la partie inf~rieure du Cr~tac~ (N~ocomien) correspondent principalement des lacunes ou des dSp6ts delta/ques. A partir de rAptien la s~di- mentation est marine, & caract~re p~lagique de plus en plus marqu~ ; cependant une importante laeune peut 6 t r e observ~e & la limite Cr~tac~- Pal~og~ne au site 763. Au cours du C~nozoique et actuellement se d~posent des "boues & Globi- g4rines" ; des lacunes, sans doute dues
l'~rosion, s'observent localement, notamment au site 763, en position topographique relativement ~lev~e.
Au site 762, & rouest du plateau d'Exmouth (Fig. 1), & 19"53.24'S et 112°15.24'E, correspondent trois forages r~alis~s sous une tranche d'eau ~paisse de 1360 m. Le 762A, cit4 pour m~moire, n'a permis de recueillir qu'une earotte de s~diments quaternaires. Le 762B a dfi 8tre stopl~ dans rOligoc~ne inf~rieur. Le 762C a atteint le
~= ~ ~.~ ~b
~ , ~ " . ~ ~ ~ . ~ ~ ~ ~ ~ •
~ ~ .
"-~ c3 ~ ~ *~ ~- ~ "~ ¢~ ~ 3 . ' " ~ *"~'~ ~-~ "'~
J 20 i i i i i i i i i i i i i i i i i i i i i " } " ' Slcll./Ca .I ........... ]~ si .......... i'"'":'"""""'"'""='"'a'"''""'""'"~'l ~ i i i i i ! .......... l"'"~'"''"'"s""'=i ! i II-'"'! .......... a! ......... I i i i , i""~"s'""' .......... ,.....I.....I~ i .... 18 i ! ~ . . . . . i i ~ i i I i i i I i i i I
' f 5 i { z a o o , e ~ ~"~ 4 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . " ' i ; . . . . . . . . . . . . . i ; . . . . . . . . i : i : i ; . . . . . . . . . i:i : : 2:: iiill :ii 1121111111 iiiii iiiii21 . . . . . . . . 11 ....
"f2' Mess./Tort. 1 1 i i ! ~ . . . . i i I i ! ! I ~ ! i I '~1"0' '" ' ! ' l ! t"" i i ..r--..~.----e.---~.-..-.r....~ ........... ~ ~ ..... , F"T~""T""F'"~'"~' , .... ~"~"'""{ ............ i ......... i II ...................................... I i.....?.....i......t.....t.....,..... } .......... i .......... 'i.iH
S e r r a v a l l . ' 7 ' i 6 ........... i ' '~ [ i i i i i i i ................. i i I . . . . . . . . . i ! i ~ ! ! i I . . . . . i i ! 71' . . . . . .
Langhien .... 5 J ..... ...... }'""-"'"I'"" I "'""I'"'" ÷ "'"'l"-"'l'"'-'e'"" ÷ '-'"!"-'"!'"'":-"-.-+-" -.e .... ~" i i ....................... -
,-' ................. ..................... '--'-" .......... i ......................... ::::::::::::::::::::::::::::::::::::::::::::::::::::: Aquitanien ..... 2" . . . . . . . . . . I ! i
Chattien NPii2ili5iI" I'2t.t, ililill)=ii)~LiiiiIiiiilir ii ' I I Iiili '] '1 ! I I t i i ~ ~ - ~ ~ ~ = ~ = : : = : " - . ' i i i 'i i i i t
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . lt iL__ '2"~ """ ................................ ~ '"", .......................................................................... T'"'i"""!'"'., """i .....
Rup~lien 2~ ! i i i '" "~'2 ....................................................................................................... : "'":'"'"~'""r "'"r "'"r"".~'""~ ..... Priabonien 2(~ ! { ] i i i i i i i i i i
~8 ............ ~ .......................... ' ............................................................... i i ~ . ~ i i i i i ..... B a r t o n i e n ~1"7 '": . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . i i i i i i " " i t i .....
Lutetien i ~[" 'I .......... I ............. [ ................................ .T. .......... T- .......... - T i i - - i - Y t i I ~~ ............ T I ............................... T T ! ........... T - - ~ F - ...................... i ................. i i .....
" ' i " '~ . . . . . . . . . . . . . . . . . . . . . . I : " ' " ' r " " ~ . " ' " ' ~ ' " " ! ' " ' " ~ . " " ' ~ . ' " " " ' " " r ' " " ' " " ? . . . . . . . . . . . . . . . . . . . . . . ~: . . . . . . . . . . . . . . . . . . . . . . ~: . . . . . . . . . . . . . . . . . . . . . . ~:.....:-. Ypresien
'T~ '~ .... J ......... ,--i-~.--.~-i-...-.H-.i----.~.--..!...-.i-..--.!.....~ ................. !.-...~ ............... ,.i..-..~ ................. i.......: ......
T n a n ~ t , e ~ .... ~ .. . . . . . . . i ' " " i " ' " i " " i ' " i i ~ ' ~ e P ~ i cer!aineldu ~.. 'xon i""'i""~"'"i'""i'"'"i'"'"i'""~'""! ..... 7
..... "51 ........ T .......... ~ " i " ' " i ' " i T [ [ ~ p r ~ s e n c e i~ossi6ie: au taxon i i ~ T i i " i ' i | ..... 4t .......... "1 .......... "'""t'~'"'!t'"'".:: . . . . ~ ~ ~ ~ ~ , #"i"!t"tg".. ' i t~ l ' i { i i ! { i l i { i } i ! { i i i i l ! I Danien ................... i .................................................................................................................................................... ...... '1" ............ ..-" . . . . ~ '" 'i ~ 3 .... i 't ~" ! i .t' i i !' i .... ~ i" . . . . . . . : : • . : . : . : : :
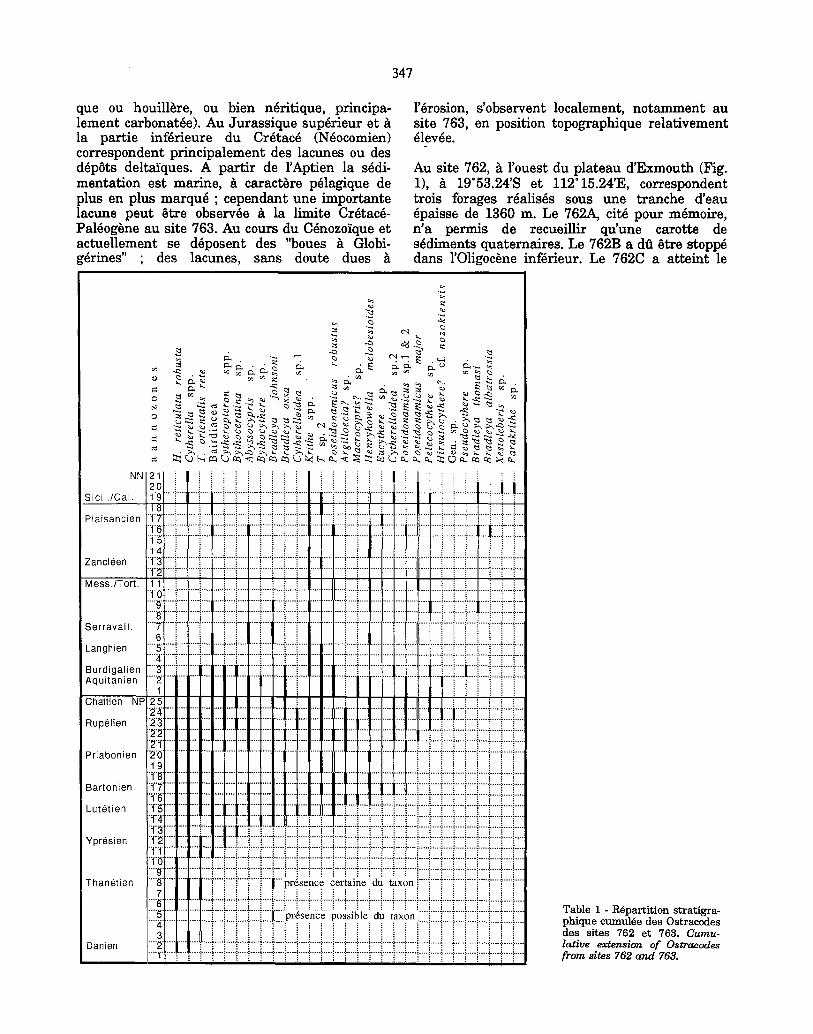
Table 1 - R6par t i t i on s t ra t ig ra - ph ique cumul6e des Os t raeodes des si tes 762 et 763. Cumu- lative extension of Ostracodes from sites 762 and 763.

348
Berriasien ~ environ - 850 m sous le fond marin e t a travers~ la limite Cr~tac~- Pal~og~ne A - 555 m ; cependant les carottes pr~lev~es dans le Pal~og~ne et particuli~rement dans le Pal~oc~ne et dans l'Eoc~ne inf~rieur sont souvent incom- plates. L'ensemble du C~nozoique est repr~sent~ par d'anciens s~diments p~lagiques A nanofossiles (craies du Pal~oc~ne A l'Eoc~ne moyen, boues au-dessus)
Au site 763 (20"35.19'S, 112°12.52'E), A l'ouest du plateau d'Exmouth, A 85 km environ au sud du site 762, correspondent aussi trois forages r~alis~s sensiblement sous la m~me tranche d'eau, A - 1367 m. Le 763A n'a pas d~pass~ la limite Eoc~ne-Oligoc~ne (A - 195 m sous le fond matin) et le 763B, prolong~ du 762C, a atteint le Berriasien A - 670 m (base du forage, dans cet ~tage, A - 1036 m). Le site 763 est remarquable par rimportance de ses lacunes (lacunes d'~rosion ?). Ainsi l'Eoc~ne moyen repose directement sur le Campanien et le Miocene manque partielle- ment. La s~rie s~dimentaire c~nozo'que pal~o- g~ne du site 763 est repr~sent~e par des craies, celle du N~og~ne et du Pleistocene par des boues.
Au total, 122 ~chantillons ont ~t~ pr~lev~s par Galbrun, 97 au site 762, 39 au site 763. Au point de vue stratigraphique, il est important de noter que les 17 ~chantillons pal~oc~nes (dont 10 st~riles) n'ont fourni, au total, que 20 valves ou carapaces en m~diocre ~tat de conservation. Avec 15 ~chantillons (dont 4 d~pourvus d'ostracodes identifiables) et 43 valves et carapaces l'Eoc~ne inf~rieur est A peine mieux loti. Plus haut dans les s~ries, les hombres d'~chantillons et de valves et carapaces sont respectivement, darts l'Eoc~ne moyen s.1. 16 et plus de 150, l'Eoc~ne sup~rieur 9
et plus de 100, l'Oligoc~ne 19 et environ 300, le Miocene inf~rleur 4 et plus de 100, le Miocene moyen 5 et environ 40, le Miocene sup~riettr 5 et 20, le Plioc~ne 19 et 30 et enfin le Pleistocene 12 et ~nviron 35. Ainsi, pour des raisons sans doute diverses (en partie trophiques comme le sugg~re ailleurs, dans le Pacifique Central, Steineck et al. 1988 ou bien li~e ~ la vitesse de la s~dimentation p~lagique), les ~chantillons provenant des parties inf~rieure (Pal~oc~ne et Eocene inf~rieur) et sup~rieure (Miocene ~ Pleistocene) des deux sites sont tr~s pauvres en ostracodes, avec en moyenne 1 ~ 4 valves ou carapaces (souvent de CythereUa ou de Bairdiacea) par ~chantillons, alors que dans les parties moyenne et sup~rieure de rEo- c~ne ainsi que dans l'Oligoc~ne cette moyenne est de rordre de 10. Naturellement, il rdsulte de cette plus ou moins grande pauvretd des dchantil- lons en individus que l'extension stratigraphique rdelle des esp~ces, en parfiiculier des plus rares, est certainement souvent supdrieure ~ celle constatde ici.
Les noms des principaux genres et esp~ces reconnus (une trentaine) sont port, s sur le tableau 1 de r~partition stratigraphique.
I~TUDE SYSTI~MATIQUE
L'~mde syst~matique porte principalement sur les esp~ces les plus caract~ristiques, appartenant aux Trachyleberididae et pourvues de valves orn~es. Les autres esp~ces ne sont le plus souvent que figur~es.
Quoiqu'il en soit, les esp~ces sont consid~r~es dans une acceptation relativement large en ce
PLANCHE 1
Fig. 1 - Cytherella cf. lata BRADY, 1880, 122-762C-13X-02, 88-89 cm., V.D. ~, Eoc. moyen. Fig. 2 - Cytherella cf. Iata, 122-762B-16H-04, 74-76, V. G. 9, Miocene. Fig. 3, 5 - "Bairdia" c£ minima BRADY, 1880, 122-762C-9X, CC, V.G. & V.D., Eoc. sup~rieur. Fig. 4 - Cytherella cf. serratula (BRADY, 1880), 122-762C-10X-02, 50-52 cm., V.D. (larv.?), Eoc. moyen. Fig. 6 - Cytherella cf. serratula (BRADY, 1880), 122-763A-19H-03, 60-62 cm., V.G., Oligoc. Fig. 7 - Cytherelloidea sp.1, 122-763A-19H-03, 60-62 era., V.D., Oligoc. Fig. 8 - Neonesidea? sp. aft. rara TITTERTON & WHATLEY, 1988, 122-763A-17H-02, 66-68 cm., V.D.,
Mioc. inf. Fig. 9 - "Bairdia" sp., 122-762B-16H-02, 67-69 cm., V.G., Mioc. in£ Fig. 10 - Krithe sp., 122-763A-16H-04, 74-76 cm., V.D., Mioc. Fig. 11 - "Bairdia" (Neonesidea?) sp. aft. rara TITTERTON & WHATLEY, 1988, 122-762B-16H-02, 74-76
cm., V.G., Mioc. in£ Fig. 12 - Gen. sp., 122-763A-19H-03, 60-62 cm., V.D., Oligoc. Fig. 13, 16 Krithe capensis DINGLE, LORD & BOOMER, 1990 (=? K. producta BRADY, 1880),
122-762B-2H-05, 67-69 cm., V.D.& V.G.v. int., Quatern. Fig. 14 - Krithe capensis, 122-762B-2H-02, 67-69 cm., V.G., Quat. Fig. 15 - Parakrithe sp. 122-762B-2H-02, 67-69 cm., V.G., Quat. Echelles graphiques, graphic scales : 0,100 ram. V.D. : valve droite - left valve ; V.G. : valve gauche - left valve.
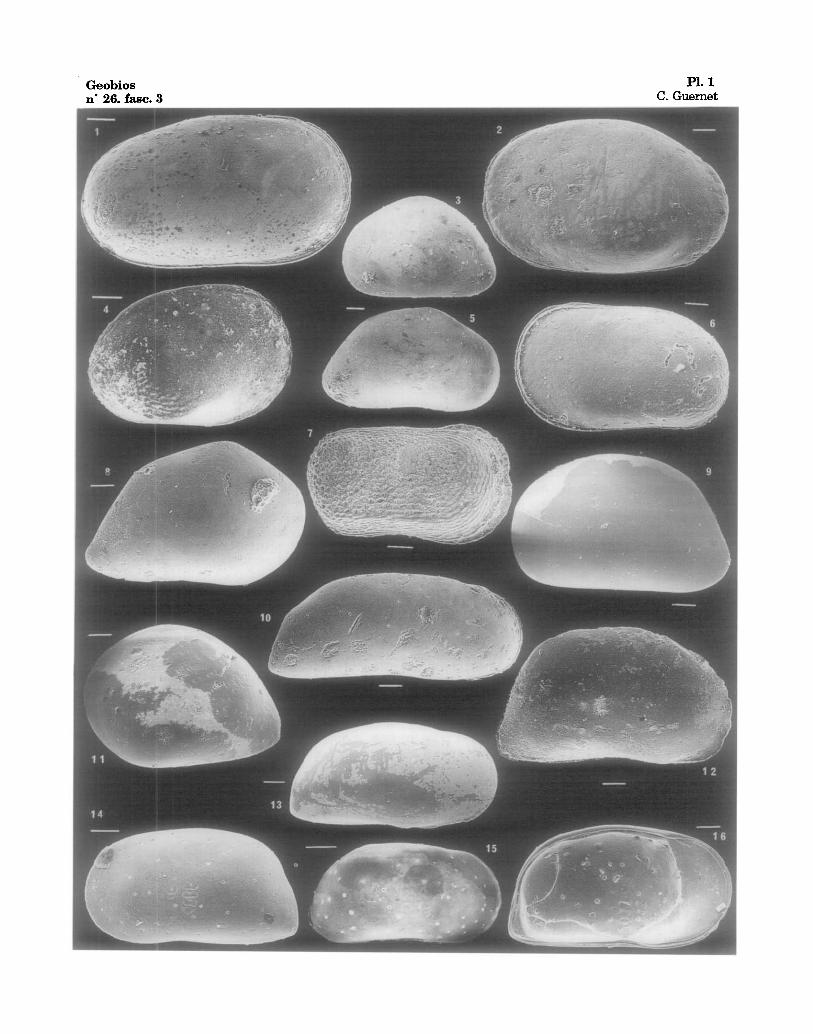
Geobios PI. 1 n" 26, fasc. 3 C. Guernet

350
sens que pour l'attribution des ostracodes recueil- lis dans le C~nozo~'que du plateau d'Exmouth, des differences morphologiques jugdes mineures (mais constantes) avec des types ou typoides d'esp~ces d~crites en d'autres lieux sont consi- ddrdes comme rexpression d'un polytypisme (c'est-~-dire de variations gdnotypique intraspd- cifiques). Ce choix est arbitraire, mais il n'existe pas la possibilitd, pour des organismes fossiles, d'appliquer les crit~res biologiques qui seuls per- mettent la distinction entre esp~ces et sous- esp~ces. Le meilleur choix, le plus "dconome", est donc celui qui dvite la creation d'esp~ces insuffi- sament caractdrisde ou sans signification strati- graphique ou dcologique ddfinie. Des recherches ultdrieures conduiront peut-~tre ~ dlever au rang spdcifique les deux seules sous-esp~ces propos~es dans cet article - Trachyleberis orientalis rete, Henryhowella reticulata robusta - et dont les types sont ddposds darts la collection du Labora- toire de Micropal~ontologie de l~Jniversitd P. & M. Cur ie .
Ordre PODOCOPIDA Mtiller, 1894 Sous-ordre PLATYCOPA Sars, 1866 Famille CYTHEI~ET,LTI]AE Sars, 1866 Genre Cytherella JONES, 1849
nales de Cythere? serratula (reals correspondent- etles bien ~ une Cytherella?, apparernment ce n'~tait pas l'id~e de Brady lui-m~me e t les figurations ne sugg~re pas ce genre) ; cependant elles sont proches, ou pour certaines d'entres elles peut-~tre consp~cifiques, du lectotype d~sign~ par Puri & Hulings (1976) et des valves figur4es par Dingle et al. (1990, fig. 5, A-C) ; elles sont identiques ~ une pattie au moins des Cytherella sp. de l'Eoc~ne moyen de la ride 90 Est (Guernet 1985, voir aussi Dingle et al. 1990, p. 254). Une des caractdristiques des valves du plateau d'Exmouth et de la ride 90 E est l'exis- tence de courtes rides dispos~es en plusieurs lignes parall~les au bord postdrovental. Au plateau d'Exmouth, elles sont pr~sentes de l'Eoc~ne infdrieur ou moyen au Quaternaire.
Les C. sp. gr. lata (pl. 1, fig. 1, 2) semblent tr~s proches de ceux figures par Brady et du lectotype ddsignd par Purl & Hulings (1976, pl. 24, fig. 17-18). Au plateau d~xmouth , je les ai observ~es de rEoc~ne moyen ~ l'Oligoc~ne.
Sous-ordre PODOCOPA Sars, 1866 Super-famille BAIRDIACEA Sars,1888 Famille BAIRDIIDAE Sars,1888
Les Cytherella sent presents dans la plupart des dchantiUons pal~og~nes ; ils sont g~n~ralement absents dans ceux d'~ge miocene moyen quaternaire, tr~s pauvres pour la plupart en ostrac0des. Beaucoup de ces CythereUa semblent p0uvoir ~tre rattach~es aux groupes serratula (BRADY, 1880) et lata BRADY, 1880. Les attribu- tions spdcifiques sont n~anmoins d~licates, compte tenu du tr~s grand hombre d'esl~ces d~crites, souvent de faqon insuffis8mment precise, compte tenu aussi de rimportance du dimorphisme bilatdral et/ou sexuel et de l'imrnaturit6 de nombreuses valves.
: !
Les valves attribudes au groupe serratula (pl. 1, fig. 4, 6) sent peu conformes aux figurations origi-
Les Bairdiidae sont relativement frequents de l'Eoc~ne moyen au Miocene inf~rieur aux sites 762 et 763. Ils appart iennent ~ au moins trois esp~ces diff~rentes mais leur ddtermination off-re des difficult~s comparables ~ celle des Cytherella, en rapport avec le nombre trds ~levd d'esp~ces de description et figuration souvent anciennes. "Bairdia" cf. minima BRADY, 1880 (pl. 1, fig. 3, 5) dent le type est actuel (lectotype dans Purl & Hulings 1976) est, de l'Eoc~ne moyen au Miocene infdrieur, la plus abondante de ces esp~ces. "Bair- dia" sp. (pl. 1, fig. 9), du Miocene inf~rieur du plateau d'Exmouth est caract~ris4e par son bord
ventral subrectiligne et son bord postdro-dorsal tr~s inclind vers l'arri~re et ~galement subrec-
P L A N C H E 2
Fig. 1-2 - Trachyleberis orientalis fete nov. sbsp., holotype, 122-763A-19H-03, 60-62 cm., V.G.,v. ext. & int., Oligoc.
Fig. 3 - Trachyleberis orientalis rete nov. subsp., 122-762C-8X-05, 25-27 cm., V.D., Eoc.sup. Fig, 4 - Bradleya ossa, 122-762B-18H-05, 10-12 cm., V.D., Oligoc. Fig. 5 - Trachyleberis sp. 2, 122-762B-16H-02, 74-76 cm., V.D., Mioc. Fig. 6 - Bradleya albatrossia, 122-762B-6H-03, 89-91 cm., V.D., Plioc. sup. Fig. 7 - Hirsutocythere? cf. nozokiensis, 122-762B-16H-02, 74-76 cm., V.D., Mioc. inf. Fig. 8 - Trachyleberis orientalis orientalis, (repris de - from Guernet, 1985) holotype, 22-214-34-6,
15-19, V.G., Eoc. Fig. 9 - Trachyleberis sp., 122-762B-17H-01, 20-22 cm., V.G., Oligoc. Fig. 10- Bradleyajohnsoni, 122-762C-03X-05, 10-12 cm., V.D., Oligoc.
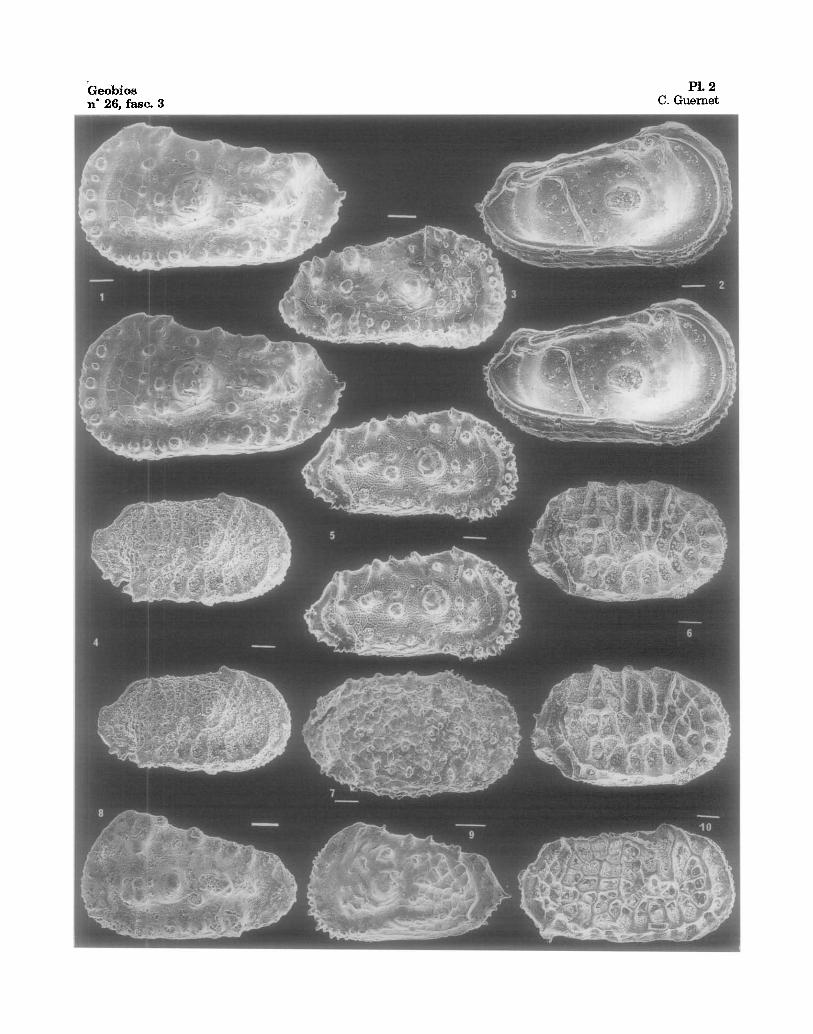
Geobios n" 26, fasc. 3
PI. 2 C. Guernet

352
tiligne. Enfin, Neonesidea? sp. aft. rara TITTER- TON & WHATLEY est pr~sente dans~ l'Eoe~ne sup~rieur et l'Oligoe6ne des deux sites ~tudi~s.
Super-famille CYTHERACEA Baird, 1853 F-mille TRACHYLEBERIDAE sensu Sylvester- Bradley, 1948 Genre Trachyleberis BRADY, 1898 Esp~ce-type Cythere scabrocuneata BRADY, 1880
TRACY~EBERIS ORIENTALIS RETE N. S. SP.: pl. 2, fig. 1-2.
1992 Actinocythereis orientalis, Guernet, p. 835-838, tabl. 1-2 et pl.1, fig.l-2.
E t y m o l o g i e , Du latin rete, allusion au earact~re de l 'ornementation.
H o l o t y p e - Une valve gauche, n ° P6M 1223 (pl. 2, fig. 1), ~chantillon O.D.P. 762B-19H-03 ; 60-62 cm. (Oligoc~ne).
Dimensions de l ' ho lo type - L = 1, 1 mm., h= 0,65 ram.
Para typoYdes - Cinq valves immatures, m~me ~chantillon.
D iagnose T. orientalis rete est une esp~ce probablement eonsp~eifique de T. orientalis orientalis GUERNET, 1985 (pl. 2, fig. 8 dans cet article). Elle s'en distingue par sa grande taille, par la presence d'une r~ticulation ~tendue ~ la plus grande pattie des valves, par ses tubercules moins ~¢ol-mlneux et par le moindre d~veloppe- ment du bourrelet subvertical situ~ ~ l'emplace- ment habituel du tubercule oculaire (une fossette oculaire sans doute vestigiale s'observe en vue interne des valves de T. orientalis).
R e m a r q u e - C'est au genre Actinocythereis que j'ai attribu~ ant~rieurement T. orientalis orien- talis et T. orientalis rete (GUERNET, 1985, 1992). Cependant, par leur forme g~n~rale et notam- ment leur extr~mit~ post~rieure ne t tement trian- gulaire y compris fi la valve droite et par leur ornementation (absence de cr~te h~micirculaire la marge ant~rieure ici enti~rement bord~e de tubercules arrondis, diff~renciation d'un tuber- cule subcentral ~ l 'emplacement des empreintes des muscles adducteurs), elles s'~loignent beaucoup, morphologiquement de A. exanthe- mata, esp~ce-type de Acanthocythereis (lectotypes et figurations dans Forester 1980) ; par leur contour elles sont sont proehes de Cythere scabrocuneata, esp~ce-type de Trachyleberis, mais s'en s'~loignent par leur ornementation. Leur attribution g~n~rique est donc provisoire.
R6partition stratigraphique - T orientalis rete constitue un des ~l~ments les plus caract~ris- tiques de la mierofaune c~nozoique des sites 762 et 763. Elle semble apparaitre d~s le Pal~oc~ne, est repr~sent~e dans la plupart des ~chantillons ~oc~nes et oligoc~nes et disparait au Miocene inf~rieur. Trachyleberis sp. BENSON & PEYPOU- QUET, 1983 (pl. 2, fig. 2, 4, 6, 8), de l'Oligoe~ne e t du Miocene du Sud de l'Atlantique, s'identifie peut-~tre fi cette esp~ce. T. orientalis est ~gale- ment proche de Trachyleberis anteplana BATE, 1972 dont elle se distingue par la forme et la disposition des tubercules ; elle a ~t~ d~crite dans le Campauien du bassin de Carnarvon e t elle est sigual~e au m~me niveau au plateau d'Exmouth par Damotte (1992).
TRACHYLEBERIS SP. 2 : pl. 2 , fig. 5.
1985 -A. sp. 2 Guernet, p. 282 et pl. II, fig. 11.
PLANCHE 3
Fig. 1 Fig. 2 Fig. 3 Fig. 4 Fig. 5
- Poseidonamicus - Poseidonamicus - Poseidonamicus - Poseidonamicus - Poseidonamicus
major, 122-763A-1H-04, 01-03 cm., V.G., Quatern. robustus, 122-763B-2X-01, 77-79 cm., V.D., Eoc. sup. major, 122-763A-5H-03, 81-83 cm., V.G., Plioc. sup. cf. robustus, 122-762B-7H-03, 19-21 cm., V.D., Eoc. sup. robustus, 122-762C-14X-03, 80-88 era., V.G., Eoc. moy.
Fig. 6 - Poseidonamicus robustus, 122-762B-16H-02, 74-78 cm., V.D., Mioe. Fig. 7 - Henryhowella reticulata robusta nov. subsp., holotype, 122-762C-26X-04, 98-99 era., V.D., Eoc.
inf. Fig. 8 - Henryhowella melobesioides, 122-763B-06X-05, 78-80 cm., V.D., Eoc. moy. Fig. 9 - Henryhowella reticulata robusta nov. subsp, 122-762C-21X-01, 20-22 cm., V.G., Eoc. inf. Fig. 10 - Poseidonamicus sp. 1, 122-762B-16H-04, 74-78 cm., V.D., Mioc. Fig. 11, 12 - Henryhowella melobesioides, 122-762B-13H-06, 72-74 cm., V.G., v. ext. & int., Mioc. moy. Fig. 13 - Poseidonamicus sp. 2, 122-762B-17H-01, 20-22 cm., VoD., Oligoc. sup. Fig. 14 - Henryhowella melobesioides, 122-762C-13X-01, 87-89 cm., V.D., Eoc. moy. Fig. 15 - Henryhowella reticulata robusta nov. subsp., 122-762C-31X-01, 132-134 cm., V.G., Pal~oc. sup.
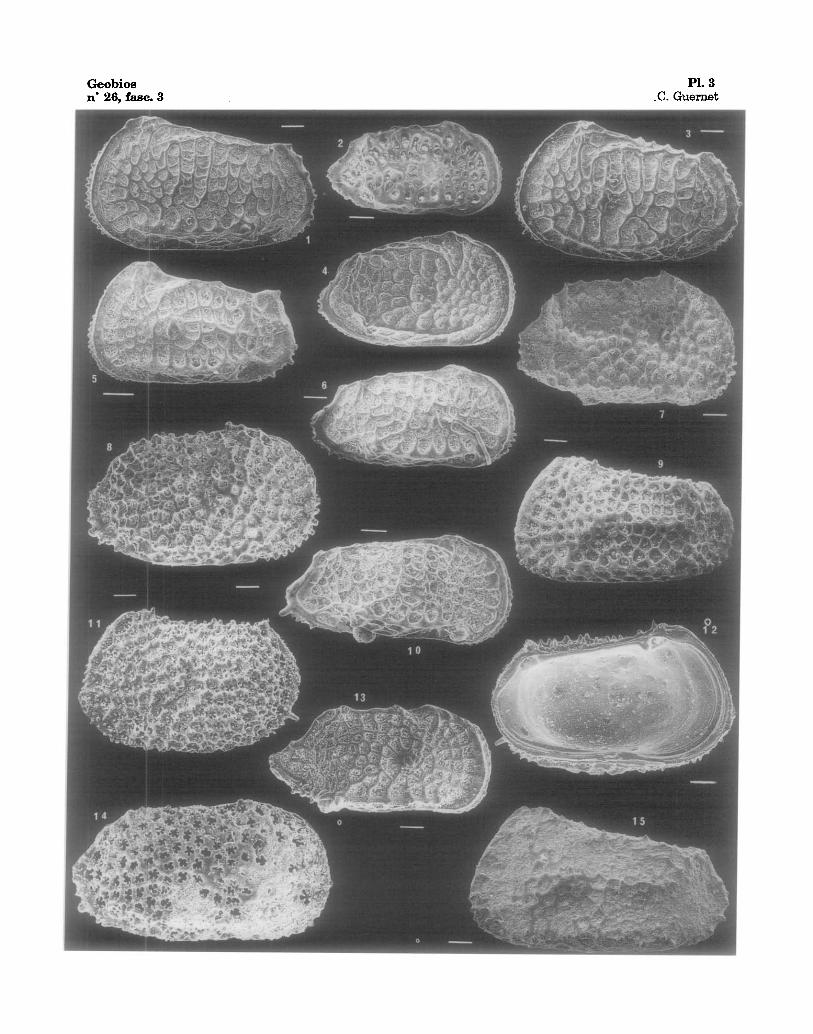
Geobios PI. 3 n" 26, fasc. 3 .C. Guernet

354
1992- A. orientalis n . s. sp, e t A. n. s, sp., Guer- net, p. 835-838, tabl. 1 et 2 et pl. 1, fig. 3 et
.
T. sp. 2 se distingue de l'esp~ce pr~c~dente par sa r~tieulation de deuxi~me ordre d~limitant de tr~s petites fossettes ; elle semble, en outre, ~tre ne t tement plus allong~e. La r~ticulation h~che de T. orientalis rete est visible, au moins sur certaines valves de T. sp. 2.
R6partition - T. sp. 2 coexiste avec T. orientalis rete dont elle semble d~river, de l'Eoc~ne moyen au Miocene inf~rieur ; ~ moins qu'elle n'occupe une niche ~cologique distinete, son existence t~moigne alors du polymorphisme de resp~ce. Par contre, ~ partir du Miocene sup~rieur, elle est seule pr~sente, darts les pr~16vements ~tudi~s, et peut alors ~tre consid~r~e comme une esp~ce nouvelle. Au site 214 (ride 90-Est), quelques valves repr~sentent cette esp~ce au voisinage de la limite Eoc~ne-Oligoc~ne (Guernet 1985) : son extension g~ographique est donc importante
TRACHYLEBERIS SP. : pl. 2, fig. 9.
Une valve provenant de l'Oligoc~ne du site 762 (762B-17H-01 ; 20-22) plus triangulaire, que eelle de T. rete se singularise en outre par son orne- mentat ion (disposition et forme des tubercules, r~ticulation en partie effac~e ant~rieurement au tubercule musculaire et au contraire mieux marquee post~rieurement, etc.).
Genre Bradleya HORNIBROOK, 1952 Esp~ce-type : Cythere arata BRADY, 1880
Les s~diments c~nozoiques des sites 762 et 763 eontiennent plusieurs esp~ces de Bradleya, genre eonnu comme l'un des plus caract~ristiques des milieux matins profonds, m~me si certains de ses repr~sentants - dont l'esp~ce-type - sont n~riti- ques.
BRADLEYA JOHNSONI BENSON, in BENSON & PEYPOUQUET, 1983 : pl. 2, fig. 10.
1992 - B. gr. johnsoni, pro parte, Guemet, p. 835-838, tableau 1, pl. 1, fig. 4.
B. johnsoni est la plus commune des esp~ces de Bradleya du site 762. Ses valves ne different de l'holotype figur~ par Benson que par leur forme g~n~rale apparemment plus quadrangulaire et par des d~tails de rornementat ion (en partieulier la cSte dorsale plus anguleuse)
Sur le plateau d'Exmouth, B. johnsoni est repr~- sent~e de l'Eoc~ne moyen au Miocene moyen, les valves plioe~nes et quaternaires, bris~es ou immatures que nous avions at tr ibutes ~ B. gr. johnsoni (Guernet & Galbrun 1992) appartenant probablement ~ P. thomasi.
B. johnsoni est une espSce cosmopolite connue darts l'Oligo-Mioc~ne de l'Atlantique nord (in Steineck et al. 1988) et du Pacifique central et dans le Miocene inf~rieur de l 'Atlantique sud (Benson & Peypouquet 1983). B. caeca GUERNET, 1985, de l'EocSne moyen de la ride 90-Est, se distingue de B. johnsoni par sa petite taille et sa r~ticulation plus grossi~re. B. caeca est peut-Stre synonyme de B. tongaensis HAZEL & HOLDEN, 1971, de l'EocSne sup~rieur du Pacifique, ainsi que Steineck me ravait sugg~r~ (communic. personn.) et elle est tr~s proche de B. solida WHATLEYet al. 1984. Ces esp~ces appart iennent
la souche des Bradleya du groupe dictyon auquel se rattache B. johnsoni.
BRADLEYA THOMASI STEINECK & YOZZO, 1988.
1988 - B. thomasi STEINECK & YOZZO, p. 187-189 et pl. 3, fig. 1-11.
1992 -B. ef. thomasi, Guemet, p. 835-838, tabl. 1-2 et pl. 1, fig. 4.
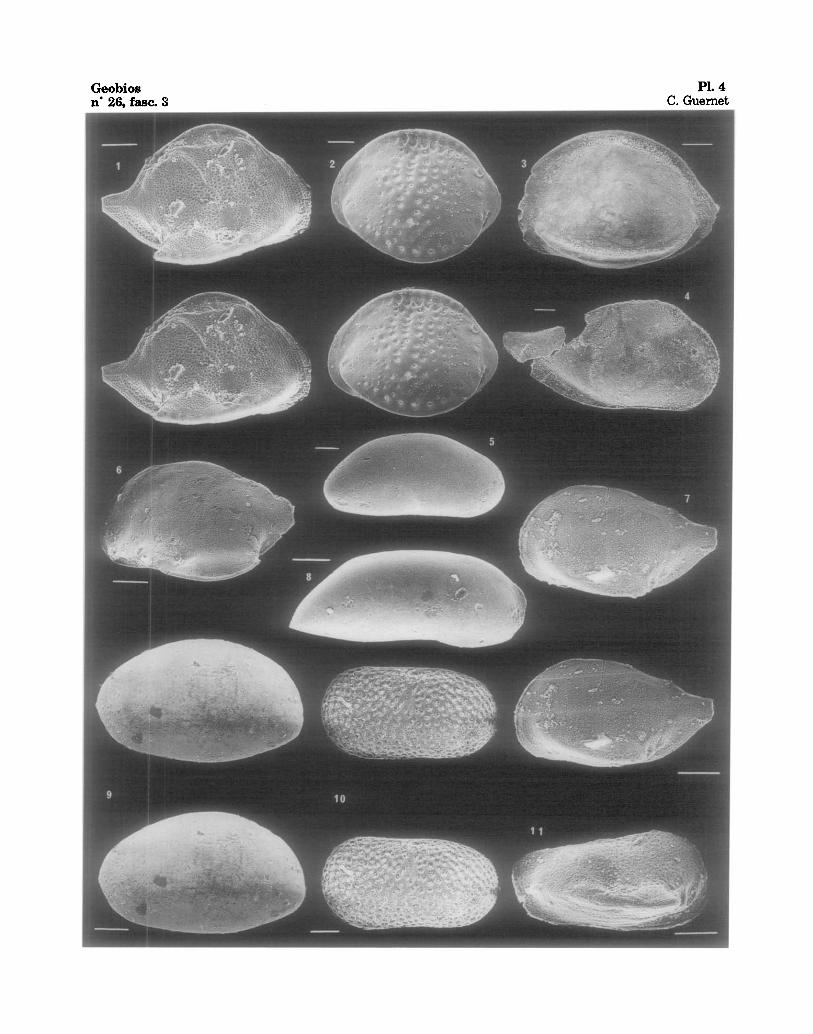
PLANCHE 4
Fig. 1 - Cytheropteron sp., 122-762B-17H-03, 20-22 cm., V.D., Oligoc. sup. Fig. 2 - Pelecocythere sp. aft. foramena WHATLEY • COLES, 1987, 122-762B-13H-06, 72-74 cm., V.D.,
Mioc. moy. Fig. 3 - Bosquetina sp. aft. mucronalatum WHATLEY & COLES, 1987, 122-762B-2H-05, 67-69 cm., V.D.,
Quatern. Fig. 4 - Phlyctocythere? sp . , 122-762B-16H-02, 74-76 cm., V.D., Mioe. Fig. 5 -Abyssocypris? sp. 122-763A-20H-01, 80-82 cm., V.D., Oligoc. inf. Fig. 6 - Cytheropteron sp., 122-763A-17H-02, 66-68 cm., V.G., Mioc. Fig. 7 - Cytheropteron sp., 122-762C-21X-01, 20-22 cm., V.G,, Eoc. inf. Fig. 8 -Argilloecia? sp. gr. acuminata MUELLER, 1894, 122-762B-18H-03, 10-12 cm., V.D., Oligoc. Fig. 9 - Abyssocypris? sp., 122-762C-17X-01, 49-52 cm., V.G., Eoc. moy. Fig. 10 - Cytherelloidea? (ou Platella?) sp. 2, 122-762C-11X-05, 60-62 cm., V.D., Eoc. moy. Fig. 11 -Bythocythere sp., 122-762C-18X-02, 22-23 cm., V.D., Eoc. inf.
Geobios n" 26, fasc. 3
PI. 4 C. Guernet

356
Les valves attribudes ~ B. thomasi - du groupe de B. dictyon (Brady 1880, voir aussi Benson 1972 et Malz 1990) - sont reconnaissables ~ leur grande taille (longueur de valves prdsumdes adultes de l'ordre de 1,1 ram), ~ la relative ddlicatesse de leur ornementation, ~ la position tr~s en retrait de la ride ant~rieure par rapport ~ la marge du test, et, enfin, ~ la disposition des fossettes au niveau et en arri~re de la structure centrale. Celle-ci ("bridge") est tr~s peu marquee et ~voque, par ce caract~re, celle de B. lordhowensis (WHATLEY et al. 1984).
A "Exmouth Plateau", B. thomasi est prdsente du Miocene supdrieur au Quaternaire. Ailleurs, l'esp~ee est cit~e par ses auteurs dans le Paei- flque (d'ofl provient le type), le canal de Mozam- bique et rAtlantique nord, avec une extension stratigraphique semblable, du Miocene supdrieur
l'Actuel. B. thomasi est tr~s proche de B. dictyon ; elle ne semble gu~re en diff~rer que par des ddtails de l 'ornementation et sa rdpartition tant stratigraphique que g~ographique est sensiblement identique.
B R A D L E Y A A L B A T R O S S I A BENSON, 1972 : pl. 2, fig. 6.
1972 - B. albatrossia BENSON, p. 39-40 et pl. 7, fig. 1-2.
?1984 - B. sp. 3, Whatley et al. p. 288 et pl. 3, fig. 9.
?1988 - B. sp. YAJ~MA, p. 1080, pl.2, fig. 15.
Une valve et un d~bris de valve provenant de deux ~chantillons du plioc~ne sup~rieur du site 763 sont attribuables ~ cette esp~ce dont le type, actuel, provient de la Mer de Chine. Elles en poss~dent la forme g~n~rale et l 'ornementation. L'absence de tubercule oculaire, de car~ne dorsale et de ride m~diane la distingue de B. andamanae BENSON, 1972 (voir aussi, pour cette esp~ce, Whatley & Quanhong 1988). La confluence d'al- v~oles m~diodorsales, par quasi disparition de murs horizontaux, la rapproche de B. sp. 3 WHATLEY et al., 1984, peut-~tre synonyme de B. sp. YAJIMA, 1988, l'une et Fautre en provenance du Miocene du Pacifique.
B R A D L E Y A O S S A WHATLEY et al., 1984 : pl. 2, fig. 4.
1984 - B. ossa, Whatley et al., p. 285, pl. 2, fig. 20-21, pl. 3, fig. 1-2.
1992 - B. ossa, Guernet, p. 835-838, tabl. 1 et 2, pl. 1, fig. 9
Des valves d'~ge oligoc~ne ~ miocene inf~rieur en provenance ~ la fois des sites 762 et 763
pr~sentent les principales caract6ristiques de B. ossa : forte car~ne ventro-lat~rale, cote dorsale rdduite ~ une tang le de 2-3 grosses ~pines, bridge ~ ~l~ments post~rieurs et dorsaux ~paissis en un grossier tubercule, r~ticulation secondaire ; cependant, ici, l'extrdmit~ post~roventrale des valves semble ~tre plus d~velopp~e et les fossettes ant~roventrales ne sont pas combines. D'autres valves, d'~ge ~oc~ne, se distinguent des pr~e~dentes par une r~ticulation plus l~che, apparemment simple et par l'~bauche d'une cr~te lat~rale m~diane.
Bradleya ossa dont le type provient du Miocene du site 207, ~ r e s t de la Nouvelle-Z~lande est cit~e jusque dans le Pleistocene (Whatley et al. 1984). Les individus recueillis sur le plateau d~lxmouth sont donc g~n~ralement plus anciens et en constituent peut-~tre les formes ancestrales
Genre Poseidonamicus BENSON, 1972 (Esp~ce-type : P. major BENSON, 1972). Diagnose ~mend~e in Whatley et al., 1986.
Le genre Poseidonamicus est repr~sent~ dans le C~nozo~'que du plateau d'Exmouth par au moins deux esp~ces : P. major et P. robustus WHATLEY et al., 1986. Ces deux esp~ces ou groupe d'esp~ces se relaient dans le temps, au cours de l'Oligoc~ne.
P O S E I D O N A M I C U S M A J O R BENSON, 1972 : pl. 3, fig. 1,3.
71972 - P. major nov. sp., Benson, p. 52-53, pl. 8, fig 5 et pl. 10, fig. 1-6.
1992 - P. sp., Guernet, p. 835-838, tabl. 1 et 2. , pro parte et pl. 1, fig. 7.
Les esp~ces connues du groupe major - P. major BENSON, 1972, P. minor BENSON, 1972, P. mioce- nica BENSON, 1983 in BENSON & PEYPOUQUET, 1983, P. riograndensis BENSON, 1982, P. ocularis WHATLEY et al., 1986, etc. - different entres elles principalement par la taille plus ou moins grande et par des d~tails de l 'ornementation. Les valves n~og~nes du plateau d'Exmouth attribuables ~ ce groupe dvoquent tant6t P. major par l 'alignement vertical des fossettes m~dio-dorsales et par l'esquisse d'une boucle dorsal, tant6t P. minor par la dominance des tours "verticaux" et l'excavation des alv~oles. Etant donn~ le petit nombre de valves recueillies, il n'est pas possible de savoir si ces variations rel~vent ou non d'un polymor- phisme intrasp~cifique.
Les esp~ces du groupe major sont cities par les auteurs de l'Oligoc~ne ou du Miocene ~ l'Actuel. Au plateau d'Exmouth la plus ancienne valve se

357
ra t tachant ~ ce groupe provient de l'intervalle 762B-18H-5 ; 10-12 cm, attribug ~ la biozone N-P 22, et l'esp~ee est repr~sent~e jusque dans le Pl$istoc~ne. En outre, de l'Oligoc~ne ~ l'Actuel, les 6chantillons livrent des valves plus grandes que celles de P. robustus (voir ci-apr~s), avec des alvgoles qui tendent ~ confluer (donc ~ 6tre plus grandes et moins nombreuses) et qui sent peut-~tre des valves larvaires de P. major (pl. 3, fig. 4)
POSEIDONAMICUS ROBUSTUS WHATLEY et al., 1986 : pl. 3, fig. 2 et 5-6.
1986 - P. robustus nov. sp., Whatley eta/., p.393 - 394, pl. 2, fig. 8-11.
1992 - P. sp., Guernet, pro parte, p .835-838 et tabl. 1 et 2.
L'absence de vestibule, la petitesse des pores normaux et la simplicitd des canaux de pores marginaux (non ramifids) plaident en faveur du ra t tachement des esp6ces qui suivent au genre HenryhoweUa plutSt qu'au genre hom6omorphe Wichmanella BERTELS, 1969.
HENRYHOWELLA RETICULATA ROBUSTA NOV. SUBSP. : pl. 3, fig. 9.
1992 - H.? reticulata, Guernet et Galbrun, p. 835-838, tabl. 1 et 2, pl. 1, fig. 8.
E t y m o l o g i e - Du latin robusta, allusion ~ la grossi~ret6 de l 'ornementation et de la costulation
H o l o t y p e - Une valve gauche, n ° P6M 1225, 6chantillon O.D.P. 762C-21X-01 ; 49-52 cm, Eoebne in£
L'Eoe~ne (moyen et sup6rieur) et l'Oligoe~ne du plateau d'Exmouth livrent eommun~ment des valves de P. robustus, esp~ce d6crite dans le Pacifique, ~ lEs t de l'Australie, o6 elle pr~sente la m~me r6partition stratigraphique. Ces valves sent de petite taille (lenr longueur est de l'ordre de 0,7 ram.), quadrangulaires du c6t6 gauche ; le r6ticule est 6pals, les c6tes dorsales et ventrales sent tr~s marquees et il y a souvent 6bauche d'une c6te m6diane ; un bourrelet "vertical" s'inscrit au niveau probable des empreintes des muscles adducteurs et tend parfois ~ s'~largir en un tubercule subcentral ; au-dessus de ce bourrelet ou tubercule ; le reticulum des valves des sites 762 et 763 est normalement d6velopp~, comme chez P. rudis WHATLEY et al., 1986. Cette derni~re esp~ce, plus grande que P. robustus, a elle aussi 6t~ d~crite au SW du Pacifique et son extension stratigraphique est aussi tr~s importante (Eocene - Quaternaire).
Dimens ions de l ' h o l o t y p e - L = 0,89 ; h = 0,53.
P a r a t y p o i d e - Deux valves gauches immatures.
D iagnose Sous-esp~ce de H. reticulata que caract~rise le fort dgveloppement des c6tes longitudinales (par renforcement des muri et des pores-eonuli) et de la structure eentrale (par 6paississement et coalescence des pores-conuli). Les autres caract~ristiques morphologiques sent celles de H. reticulata (GUERNET, 1985).
R 6 p a r t i t i o n s t r a t i g r a p h i q u e - Pal~oc~ne sup~- rieur? - Eocene inf~rieur - Miocene inf6rieur des sites 762 et 763 CExmouth Plateau"). Les deux indications de pr6sence darts la Miocene moyen et sup6rieur in Guernet & Galbrun (1991, tableau 2) correspondent ~ des valves immatures, plus vraisemblablement attribuables ~ H. cf. melobe- sioides (voir ci-dessous).
POSEIDONAMICUS SP. : pl. 3, fig. 10 & 13.
1992 - P. sp., Guernet, pro parte, p. 835-838 et tabl. 1 et 2.
L'Oligoc~ne et le Miocene du site 762 ont livr~ quelques valves de Poseidonamicus plus allon- g6es que celles des esp~ces pr~c~dentes ; en outre les murs du reticulum sent moins ~pais tandis que les alv6oles sent de forme irr~guli~res (P. sp. 1, pl. 3, fig. 10) ou bien plus ou moins quadran- gulaires et dispos6es de faqon concentrique (P. sp. 2, pl. 3, fig. 13).
Genre Henryhowella PURI, 1957 (Esp~ce-type : Cythere evax ULRICH & BASSLER, 1904) Esp~ce Henryhowella reticulata (GUERNET, 1985).
HENRYHOWELLA MELOBESOIDES (BRADY, 1869) : pl. 3, fig. 8 & 11-12.
1869 - Cythere melobesioides, BRADY, 1869, pl. 19, fig. 10-11.
1976 - Cythere melobesioides BRADY, Purl & Hulings, pl. 25, fig. 1-2.
1990 - H. ? melobesioides (BRADY), Dingle, Lord & Boomer, p 311-315, fig. 42 c-f et 43 A-F.
1992 - Gen 2 sp. et H. sp. GUERNET & GALBRUN, p. 835-838, tabl. 1 et tabl. 2.
H. melobesioides appart ient ~ un groupe d'esp~ees qui semblent se distinguer de celles du groupe de H. evax ULRICH • BASSLER, 1904 (=? H. asperrima REUSS, 1850) par l'absence de trois c6tes ou rang~es d'6pines ~ l'arri~re des valves (selon Dingle et al. 1990). H. melobesioides a

358
originellement ~t~ d~crite au large de rile Maurice. Puri & Hulings (1976) en ont figur~ (pl. 25, fig. 1-2) une valve recueillie au large du Cap de Bonne Esp~rance, ~ -50 m de profondeur tandis que Dingle et al. (1990) at tr ibuent ~ cette esp~ce plusieurs valves provenant de la marge sud-orientale de l'Afrique. Ces valves pr~sentent une variabilit~ telle (fig. 42 C-F et 43 A-F) que ces derniers auteurs n'excluent pas "that we have more than one species in our populations". En particulier, les pores-conuli peuvent ~tre massifs, d'aspect noduleux ou bien ~lanc~s, d'aspect ~pineux ; dernier le r~ticule, dont les pores-conuli Qccuperit les noeuds, est lui-m~me plus ou moins d~velopp~. Enfin l 'allongement variable des valves n'est probablement pas li~ uniquement au dimorphisme sexuel. Steineck et al. (1988) ont aussi observ~ de telles differences d'orne- mentat ion et de taille parmi les Henryhowella du Pacifique ~quatorial
C'est ~ ce .type d'ornementation - ~pineux et r~ticul~ - que se rat tachent les valves des sites 762 et 763 at tr ibutes ~ cette esp~ce. Elles se singularisent, par rapport ~ celles de l'Atlantique notamment, par l'existence de minuseules ~pines d~velopp~es radialement dans les alv~oles partir du r~ticule (cellation ~toil~e), caract~re sp~cifique ou infrasp~cifique ou bien caract~re ph~notypique peut-~tre li~ ~ l 'environnement. H. melobesioides est pr~sente, ~ Exmouth Plateau, de l'Eoc~ne moyen au Pleistocene (et ~ l'Actuel ?). Jusqu'au Miocene inf~rieur elle coexiste avec H. reticulata robusta dont elle d~rive peut-~tre et dont elle n'est pas toujours facilement distinguable.
Genre Hirsutocythere HowE, 1951 Esp~ce-type : H. hornotina HOWE, 1951
HIRSUTOCYTHERE CF. NOZOI~ENSIS (ISHI- ZAKI, 1963) : pl. 2, fig. 7.
1963 - Carinocythereis nozokiensis nov. sp., Ishizaki, 1963, p. 28, fig. 10.
1988 - H. nozokiensis (ISHIZAKI), Yajima, 1988, p. 1078-1079, fig. 16.
Les deux valves recueillies dans l'Oligoc~ne et dans le Miocene inf~rieur des sites 762 et 763 semblent plus proches de celle figur~e par Yajima que des types figures par Ishizaki, les uns et les autres en provenance du Miocene inf~rieur et moyen du Japon. Les attributions g~n~riques propos~es par les auteurs pour cette esp~ce ne semblent n~anmoins pas convenir ; les contours (en vue lat~rale, valves arrondies aux deux extr~mit~s et bord ventral convexe ~ la valve droite) et/ou l 'ornementation (absence de costula-
tion, pores-conulis plus ou moins grossiers situ~s aux noeud d'un r~ticule plus ou moins d~velopp~) ne correspondent pas ~ ceux des esp~ces-types soit de Carinocythereis soit de Hirsutocythere.
Quoiqu'il en soit de rappartenance g~n~rique de nozokiensis, sa presence (ou celle d'une esp~ce tr~s voisine) au plateau Exraouth t~moignerait de son extension g~ographique considerable, peut- ~tre restreinte au Miocene, ce qui en ferait un bon marqueur stratigraphique.
S Y N T H E S E E T C O N C L U S I O N
Le nombre d'esp~ces recueillies dans les 122 ~chantillons ~tudi~s des sites 762 et 763 est certainement sup~rieur ~ 30. I1 ne peut ~tre ~tabli avec precision, les dissemblances observ~es entre certaines valves, no tamment au sein des genres Cytherella, "Bairdia", Krithe, etc., pou- vant, d priori, ~tre li~es au dimorphisme sexuel,
des differences de stade de d~veloppement ou de lat~ralit~ ou ~ un polymorphisme intrasp~ci- fique aussi bien qu'~ la diversit~ sp~cifique. Les probl~mes de polymorphisme sont d 'autant plus difficiles ~ r~soudre que, souvent, le nombre de valves susceptibles d'appartenir ~ une m~me esp~ce est g~n~ralement de l'ordre de l'unit~ par ~chantillon et qu'il est au total peu ~lev~ dans
l 'ensemble de la s~rie ; l 'importance de la varia- bilit~ intrasp~cifique ne peut alors ~tre ~tablie. L'attribution des valves ~ des esp~ces d~crites en des lieux plus ou moins ~loign~s se heurtes ~ un probl~me du m~me type, celui d'un ~ventuel polytypisme. I1 est par ailleurs possible que les faibles differences observ~es entre les esp~ces des groupes Bradleya johnsoni, B. dictyon, P. major, etc. rel~vent en pattie d u n e telle variabilit& Enfin les differences observ~es d'une valve ~ une autre peuvent avoir une signification purement ph~notypique. Aucune ~tude statistique suscep- tible d'aider ~ r~soudre partiellement les probl~- rues ainsi pos4s ne peut ~tre envisag~e compte- tenu du tr~s petit nombre de valves eomparables recueillies. Ces i n c e r t i t u d e s ob l i gen t ~ u n e c e r t a i n e p r u d e n c e auss i b ien darts les attri- bu t i ons sp~acifiques q u e lors des crea t ions de n o u v e a u x taxons . Ceux-ci se limitent ainsi deux sous-esp~ces Trachyleberis orientalis rete et Henryhowella reticulata robusta, l'une et l 'autre relativement bien caract~ris~es morphologique- ment et repr~sent~es dans les sondages des deux sites.
D'une fa~on g~n~rale (tableau 1) les esp~ces du plateau d'Exmouth ont une longue r~partition stratigraphique cornme nous l'avons d~j~ soulign~ (Guernet & Galbrun 1991) et comme c'est la r~gle

359
habituelle pour les organismes mat ins des grandes profondeurs. La diversitd sp~cifique est maximum dans l'Eoc~ne et l'Oligoc~ne (consta- tation semblable faite par Steineck et al. 1988, dans le Pacifique dquatorial), mais en ce qui concerne les sites 762 et 763 cette diversitd est au moins partiel lement lide A l'abondance des ostracodes, ceux-ci dtant rares dans les prdleve- ments paldoc~nes et dans ceux du Ndog~ne et du Quaternaire.
R e m a r q u e . Ce travail a ~td rdalisd dans le cadre de I'U.A. 1315 et du GDR 88 du CNRS. I1 fait suite au Leg 122 dont je remercie t o u s l e s participants e t no tamment Galbrun qui, A bord du Joides Resolution, a collectd les dchantillons. J~exprime ~galement ma grati tude ~ Whatley pour ses remarques constructives, no tamment propos des esp~ces provisiorement attribudes au genre Trachyleberis.
Certaines esp~ces sont suffisamment caractd- ristiques et bien repr~sentdes pour que leur signification stratigraphique puisse ~tre envisa- gde. H. reticulata robusta apparait, semble-t-il, d~s la base du Pal~oc~ne et s'~teindrait au cours du Miocene. Trachyleberis orientalis rete semble avoir sensiblement la m~me rdpartition et T. sp. 2 apparait plus tardivement, ~ l'Eoc~ne moyen : la coexistence des deux sous-esp~ces correspond donc dans l'dtat actuel de nos connaissances l'intervalle Eocene moyen-Mioc~ne infdrieur. Bradleya johnsoni, B. ossa, Poseidonamicus robustus et H. melobesioides apparaissent aussi au Lutdtien et P. robustus semble un bon mar- queur de l'intervalle Eocene moyen-OligocAne. Les autres esp~ces ont dtd recueillies en trop petit nombre pour que leur rdpartition aux sites 762 et 763 puisse ~tre considdrde comme signifi- cative. Cependant, compte-tenu des donn~es publides dans d'autres parties du domaine ocdani- que, il est probable que B. fir. thomasi-dictyon qui prend le relais de B. johnsoni au cours du Mio- cene supdrieur (comme ddj~ constatd pax Steineck et al. 1988) et B. albatrossia caractdrisent l'inter- valle Miocene sup~rieur-Actuel. I1 existe donc une dvolution lente mais significative de nature des associations sp~cifiques, peut-~tre plus rapide au d~but de l'Eoc~ne et au Miocene sup~rieur.
La composition g d n d r i q u e des associations est relativement stable, au moins ~ partir de l'Eoc~ne, avec dominance des Cytherella, "Bair- dia", Krithe, Henryhowella, Bradleya, Poseidona- micus, etc. Cette constance tdmoigne de la per- manence des conditions bathymdtriques et, soit d'une permanence, soit d'une ~volution tr~s progressive de la tempdrature (avec peut-~tre des refroidissements un peu plus rapides ~ l'Eoc~ne moyen et au Miocene supdrieur, contemporains de l'apparition de quelques espbces nouvelles). Par contre, un changement important des condi- tions du milieu (refroidissement) se produit probablement ~ la fin du Crdtacd ou au d~but du Paldoc~ne (ou ~ la limite entre les deux), les associations gdndriques du Crdtacd supdrieur (voir Damotte 1992) et du Cdnozo~que infdrieur dtant tr~s diffdrentes.
R E F E R E N C E S B I B L I O G R A P H I Q U E S
BATE R. 1972 - Upper Cretaceous Ostracoda from the Carnavon Basin, Western Australia. Special Papers in Paleontology, 10 : 1-85.
BENSON R. 1972 - The Bradleya problem, with description of two new psychrospheric Ostra- code genera Agrenocythere and Poseidonamicus (Ostracode : Crustacea). Smithsonian contribu- tions to Paleobiology, 12 : 1-138.
BENSON R. & PEYPOUQUET J.-P. 1983 - The upper and mid-bathyal cenozoic ostracode faunas of the Rio Grande Rise found on Leg 72 of the Deep Sea Drilling Project. Init. Repts DSDP, 72 : Washington (U.S. Govt. Printing Office) : 808-818.
BERTELS A. 1969 - Rocaleberidinae, nueva sub- familia (Ostracoda, Crustacea) del l~mite Cretacico-Tertiario de Patagonia septentrional Argentina). Ameghiniana, 6/2 : 146-171.
BRADY 1880 - Report on the Ostracoda dredged by H.M.S. Challenger during the years 1873-1876. Rep. sci. Results Voyage Challenger, Zool., 1/3 : 1-184.
CRASQUIN-SOLEAU S., D]~PI~CHE F. & GALBRUN B. 1990 - Premiers ostracodes triasiques ddeou- verts darts un forage D.S.D.PJO.D.P. : Le Leg 122 (NW de l'Australie). Comptes rendus de l'Acaddmie des Sciences de Paris, 311, 2 : 1081- 1087.
DAMOTTE R. 1992 - Cretaceous ostracodes from Holes 761B and 764A (Wombat Plateau) and Holes 762C, 763B, and763C (Exmouth Plateau). In VON RAD U., HAQ B. U. et al. : Proceeding ODP, Sciences Results ODP, College Station, 122 : 819-834.
D]~P~.CHE F. & CRASQUIN-SOLEAU S. 1992 Triassic marines Ostracodes of the Australian margin (Holes 759B, 760B, 761C, 764A, and 764]3). In VON RAD V., HAQ B.V. et al. : Proceeding ODP, Sciences Results ODP, College Station, 122 : 453-462.
DINGLE R., LORD A. & BOOMER I. 1990 - Deep- water Quaternary Ostracoda from the conti- nental margin off South-Western Africa (SE Atlantic Ocean). Annals of the South African Museum, 99/9 : 1-366.

360
FORESTER R. 1980 - A systemacie Revision of the Ostracode Species Described by Ulrich and Basster and by Malkin from the Chesapeake Group in Maryland and Virginia. Geology Survey Profesional Paper, Washington (U.S. Govt. Printing Office), 1128 : 1-22.
GUERNET C1. 1985 - Ostracodes pal~ogSnes de quelques sites "D.S.D.P." de l'Oc~an Indien (legs 22 et 23). Revue de Paldobiologie, 412 : 279-295.
GUERNET C1. & GALBRUN B. 1992 - Preliminary Report on the Ostracodes of Leg 122 (Exmouth Plateau, Indian Ocean). In VON RAD U., HAQ B. U. et al. : Proceeding ODP, Sciences Results, Leg122, College Station : 835-838.
HAQ B.U., VON RAD V., O CONNEL S. et al. 1990 - Proc. of the Ocean Drilling Program, Initial Reports, Washington (U.S. Govt. Printing Office), 122 : 1-826.
HAZEL J. ~z HOLDEN J. 1971 - Ostracoda of Late Eocene Age From Eua, Tonga. Geological Survey Profesional Paper, Washington (U.S. Govt. Printing Office), 640D: 1-11.
ISHIZAKI I~ 1963 - Japanese Miocene ostraeodes from the Sunakosaka Member of the Yatsuo Fomation, east of Kanazawa City, Ishikawa Prefecture. Japanese Journal Geological and Geographical, 34/1 : 19-34.
MALZ H. 1990 - Tiefseearten leben 18nger. Natur und Museum, 120/5 : 139-152.
PURI H. & HULINGS N. 1976 - Designation of lectotypes of some ostracods from the Challen- ger Expedition. Bulletin British Museum natural History (Zoology), 29, 5 : 249-315.
REUSS A~ 1850 - Die fossilien Entomostraceen des ~lsterreichischen Terti~r-beckens. Naturwiss. Abhandl. : 41-92.
STEINECK P., DEHLER D., HOOSE E. & MCCALLA D. (1988) - Oligocene to Quaternary Ostracods of the Central Equatorial Pacific (Leg 85, DSDP-IPOD). In HANAI T., IKEYA N. 8z ISHIZAKI K. (eds.) : Proceeding of the Ninth International Symposium on Ostracoda : 597- 617.
STEINECK L. • YOZZO D. 1988 - The late Eocene- Recent Bradleya johnsoni Benson lineage (Crustacea, Ostracoda) in the Central Equa- torial Pacific. Journal of micropalaeontology., 7, 2 : 187-199.
WHATLEY R. & COLES G. 1987 - The late Miocene to Quaternary Ostracoda of Leg 94 Deep Sea Drilling Project. Revista Espanola de Micropa- leontologia, 19 : 33-97.
WHATLEY R., DOWNING S., KESLER I~ & HARLOW C. 1984 - New species of the Ostracod Genus Bradleya from the Tertiary and Quaternary of D.S.D.P. sites in the Southwest Pacific. Revista Espanola de Micropaleontologia, 16 : 265-298.
WHATLEY R., DOWNING S., KESLER I~ & HARLOW C. 1986 - The Ostracod genus Poseidonamicus from the Cainozoic of D.S.D.P. sites in the S.W. Pacific. Revista Espanola de Micropaleon- tologia, 18 : 387-400.
WHATLEY R. & QUANHONG Z. 1988 Recent Ostracoda of the Malacca Straits. Revista Espanola de Micropaleontologia, 10 : 5-37.
YAJIMA M. 1988 - Preliminary Notes on the Japanese Miocene Ostracoda. in "Evolutionary Biology of Ostracoda". In HANAI I., IKEYA N. & ISHIZAKI IZ~ (eds.) : Proceedings of the Ninth International Symposium on Ostracoda : 1073- 1085.
















![Examens pré interventionnels systématiques · dans les recommandations publiées en 1998 par l’ANAES [1] exclut les examens pré interventionnels spécifiques de l’acte thérapeutique](https://img.pdfslide.fr/doc/110x75/5b95775e09d3f2de4a8c1b83/examens-pre-interventionnels-systematiques-dans-les-recommandations-publiees.jpg)












