Embed Size (px)
Citation preview

PCEM2 Fiches
Biologie
By Puyraimond-Zemmour Jérémy ©

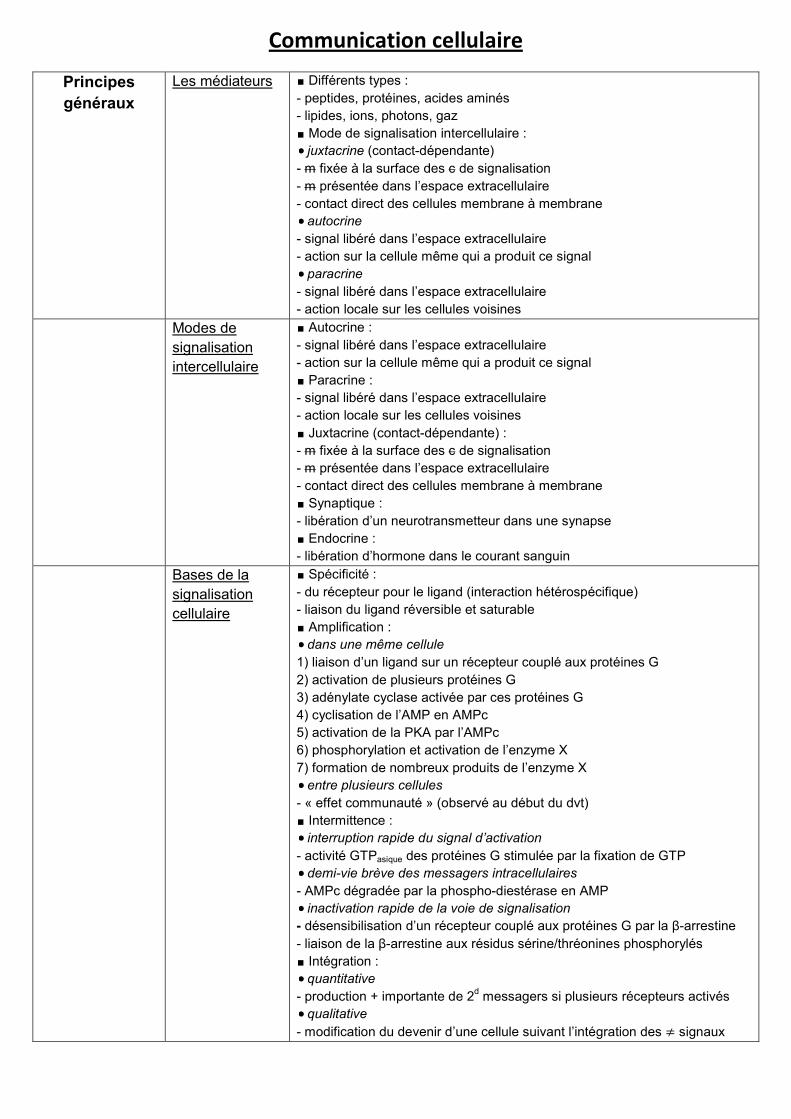
Communication cellulaire
Principes
généraux
Les médiateurs ∎ Différents types :
- peptides, protéines, acides aminés
- lipides, ions, photons, gaz
∎ Mode de signalisation intercellulaire :
juxtacrine (contact-dépendante)
- m fixée à la surface des c de signalisation
- m présentée dans l’espace extracellulaire
- contact direct des cellules membrane à membrane
autocrine
- signal libéré dans l’espace extracellulaire
- action sur la cellule même qui a produit ce signal
paracrine
- signal libéré dans l’espace extracellulaire
- action locale sur les cellules voisines
Modes de
signalisation
intercellulaire
∎ Autocrine :
- signal libéré dans l’espace extracellulaire
- action sur la cellule même qui a produit ce signal
∎ Paracrine :
- signal libéré dans l’espace extracellulaire
- action locale sur les cellules voisines
∎ Juxtacrine (contact-dépendante) :
- m fixée à la surface des c de signalisation
- m présentée dans l’espace extracellulaire
- contact direct des cellules membrane à membrane
∎ Synaptique :
- libération d’un neurotransmetteur dans une synapse
∎ Endocrine :
- libération d’hormone dans le courant sanguin
Bases de la
signalisation
cellulaire
∎ Spécificité :
- du récepteur pour le ligand (interaction hétérospécifique)
- liaison du ligand réversible et saturable
∎ Amplification :
dans une même cellule
1) liaison d’un ligand sur un récepteur couplé aux protéines G
2) activation de plusieurs protéines G
3) adénylate cyclase activée par ces protéines G
4) cyclisation de l’AMP en AMPc
5) activation de la PKA par l’AMPc
6) phosphorylation et activation de l’enzyme X
7) formation de nombreux produits de l’enzyme X
entre plusieurs cellules
- « effet communauté » (observé au début du dvt)
∎ Intermittence :
interruption rapide du signal d’activation
- activité GTPasique des protéines G stimulée par la fixation de GTP
demi-vie brève des messagers intracellulaires
- AMPc dégradée par la phospho-diestérase en AMP
inactivation rapide de la voie de signalisation
- désensibilisation d’un récepteur couplé aux protéines G par la β-arrestine
- liaison de la β-arrestine aux résidus sérine/thréonines phosphorylés
∎ Intégration :
quantitative
- production + importante de 2d messagers si plusieurs récepteurs activés
qualitative
- modification du devenir d’une cellule suivant l’intégration des ≠ signaux
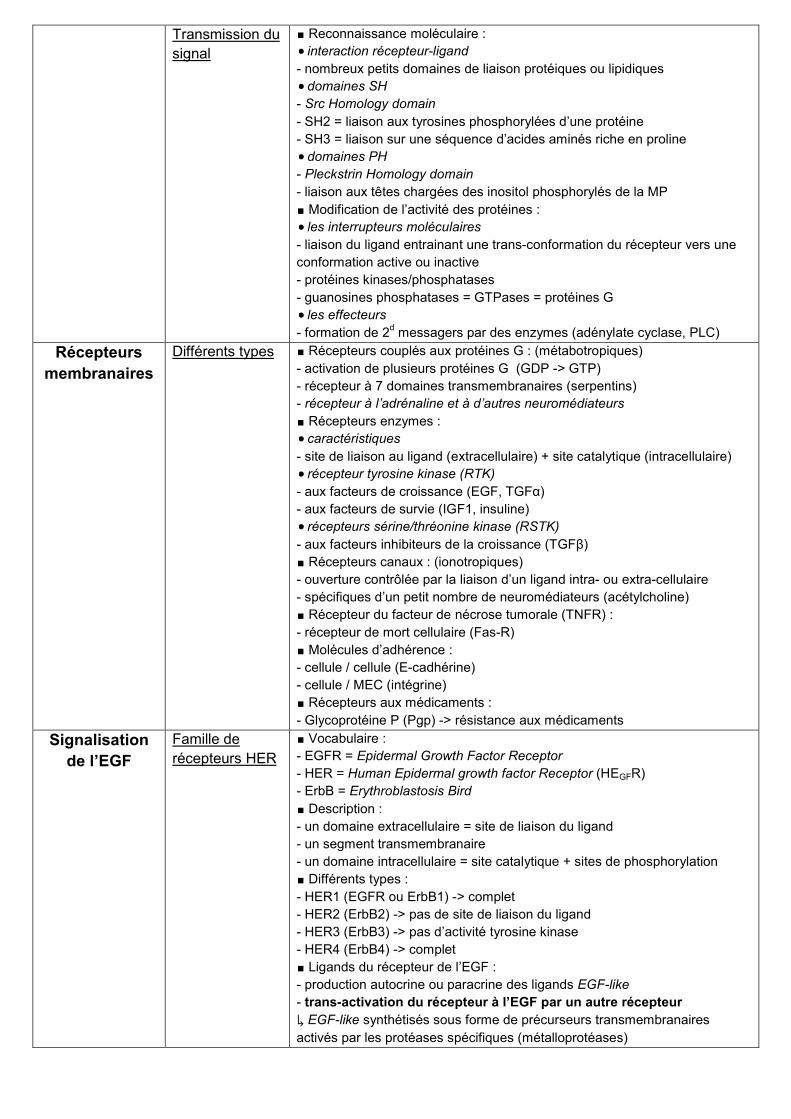
Transmission du
signal
∎ Reconnaissance moléculaire :
interaction récepteur-ligand
- nombreux petits domaines de liaison protéiques ou lipidiques
domaines SH
- Src Homology domain
- SH2 = liaison aux tyrosines phosphorylées d’une protéine
- SH3 = liaison sur une séquence d’acides aminés riche en proline
domaines PH
- Pleckstrin Homology domain
- liaison aux têtes chargées des inositol phosphorylés de la MP
∎ Modification de l’activité des protéines :
les interrupteurs moléculaires
- liaison du ligand entrainant une trans-conformation du récepteur vers une
conformation active ou inactive
- protéines kinases/phosphatases
- guanosines phosphatases = GTPases = protéines G
les effecteurs
- formation de 2d messagers par des enzymes (adénylate cyclase, PLC)
Récepteurs
membranaires
Différents types ∎ Récepteurs couplés aux protéines G : (métabotropiques)
- activation de plusieurs protéines G (GDP -> GTP)
- récepteur à 7 domaines transmembranaires (serpentins)
- récepteur à l’adrénaline et à d’autres neuromédiateurs
∎ Récepteurs enzymes :
caractéristiques
- site de liaison au ligand (extracellulaire) + site catalytique (intracellulaire)
récepteur tyrosine kinase (RTK)
- aux facteurs de croissance (EGF, TGFα)
- aux facteurs de survie (IGF1, insuline)
récepteurs sérine/thréonine kinase (RSTK)
- aux facteurs inhibiteurs de la croissance (TGFβ)
∎ Récepteurs canaux : (ionotropiques)
- ouverture contrôlée par la liaison d’un ligand intra- ou extra-cellulaire
- spécifiques d’un petit nombre de neuromédiateurs (acétylcholine)
∎ Récepteur du facteur de nécrose tumorale (TNFR) :
- récepteur de mort cellulaire (Fas-R)
∎ Molécules d’adhérence :
- cellule / cellule (E-cadhérine)
- cellule / MEC (intégrine)
∎ Récepteurs aux médicaments :
- Glycoprotéine P (Pgp) -> résistance aux médicaments
Signalisation
de l’EGF
Famille de
récepteurs HER
∎ Vocabulaire :
- EGFR = Epidermal Growth Factor Receptor
- HER = Human Epidermal growth factor Receptor (HEGFR)
- ErbB = Erythroblastosis Bird
∎ Description :
- un domaine extracellulaire = site de liaison du ligand
- un segment transmembranaire
- un domaine intracellulaire = site catalytique + sites de phosphorylation
∎ Différents types :
- HER1 (EGFR ou ErbB1) -> complet
- HER2 (ErbB2) -> pas de site de liaison du ligand
- HER3 (ErbB3) -> pas d’activité tyrosine kinase
- HER4 (ErbB4) -> complet
∎ Ligands du récepteur de l’EGF :
- production autocrine ou paracrine des ligands EGF-like
- trans-activation du récepteur à l’EGF par un autre récepteur
↳ EGF-like synthétisés sous forme de précurseurs transmembranaires
activés par les protéases spécifiques (métalloprotéases)
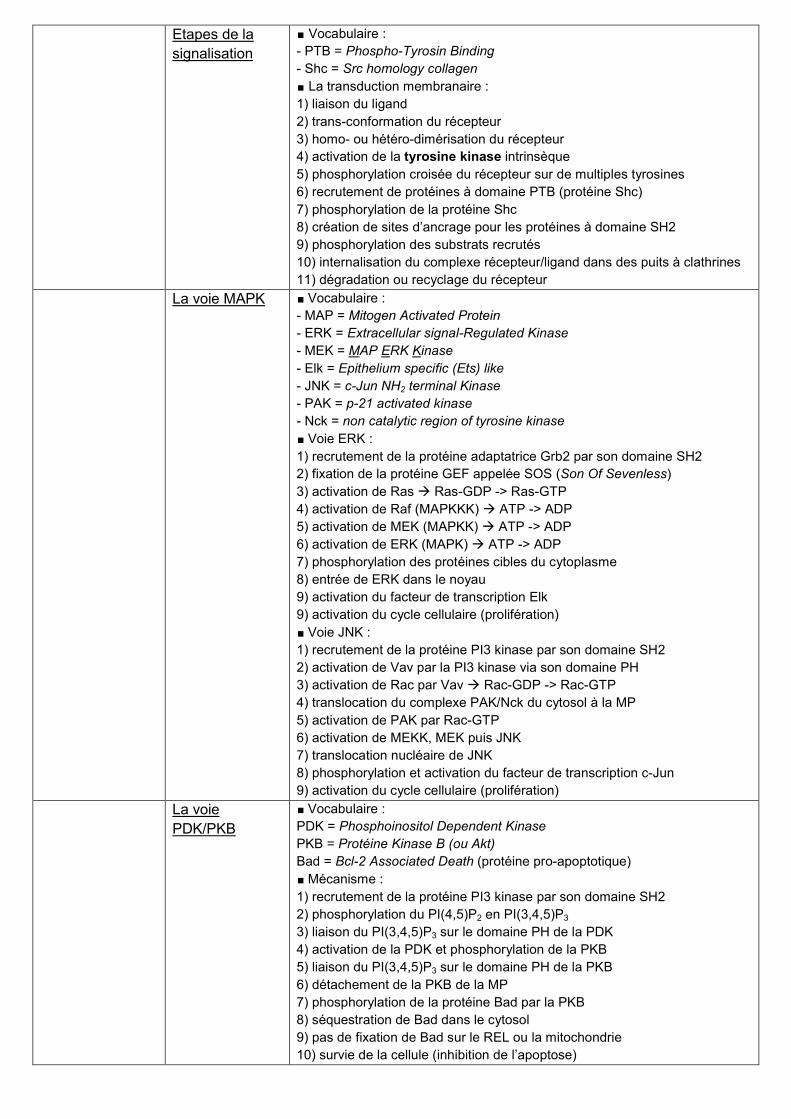
Etapes de la
signalisation
∎ Vocabulaire :
- PTB = Phospho-Tyrosin Binding
- Shc = Src homology collagen
∎ La transduction membranaire :
1) liaison du ligand
2) trans-conformation du récepteur
3) homo- ou hétéro-dimérisation du récepteur
4) activation de la tyrosine kinase intrinsèque
5) phosphorylation croisée du récepteur sur de multiples tyrosines
6) recrutement de protéines à domaine PTB (protéine Shc)
7) phosphorylation de la protéine Shc
8) création de sites d’ancrage pour les protéines à domaine SH2
9) phosphorylation des substrats recrutés
10) internalisation du complexe récepteur/ligand dans des puits à clathrines
11) dégradation ou recyclage du récepteur
La voie MAPK ∎ Vocabulaire :
- MAP = Mitogen Activated Protein
- ERK = Extracellular signal-Regulated Kinase
- MEK = MAP ERK Kinase
- Elk = Epithelium specific (Ets) like
- JNK = c-Jun NH2 terminal Kinase
- PAK = p-21 activated kinase
- Nck = non catalytic region of tyrosine kinase
∎ Voie ERK :
1) recrutement de la protéine adaptatrice Grb2 par son domaine SH2
2) fixation de la protéine GEF appelée SOS (Son Of Sevenless)
3) activation de Ras Ras-GDP -> Ras-GTP
4) activation de Raf (MAPKKK) ATP -> ADP
5) activation de MEK (MAPKK) ATP -> ADP
6) activation de ERK (MAPK) ATP -> ADP
7) phosphorylation des protéines cibles du cytoplasme
8) entrée de ERK dans le noyau
9) activation du facteur de transcription Elk
9) activation du cycle cellulaire (prolifération)
∎ Voie JNK :
1) recrutement de la protéine PI3 kinase par son domaine SH2
2) activation de Vav par la PI3 kinase via son domaine PH
3) activation de Rac par Vav Rac-GDP -> Rac-GTP
4) translocation du complexe PAK/Nck du cytosol à la MP
5) activation de PAK par Rac-GTP
6) activation de MEKK, MEK puis JNK
7) translocation nucléaire de JNK
8) phosphorylation et activation du facteur de transcription c-Jun
9) activation du cycle cellulaire (prolifération)
La voie
PDK/PKB
∎ Vocabulaire :
PDK = Phosphoinositol Dependent Kinase
PKB = Protéine Kinase B (ou Akt)
Bad = Bcl-2 Associated Death (protéine pro-apoptotique)
∎ Mécanisme :
1) recrutement de la protéine PI3 kinase par son domaine SH2
2) phosphorylation du PI(4,5)P2 en PI(3,4,5)P3
3) liaison du PI(3,4,5)P3 sur le domaine PH de la PDK
4) activation de la PDK et phosphorylation de la PKB
5) liaison du PI(3,4,5)P3 sur le domaine PH de la PKB
6) détachement de la PKB de la MP
7) phosphorylation de la protéine Bad par la PKB
8) séquestration de Bad dans le cytosol
9) pas de fixation de Bad sur le REL ou la mitochondrie
10) survie de la cellule (inhibition de l’apoptose)
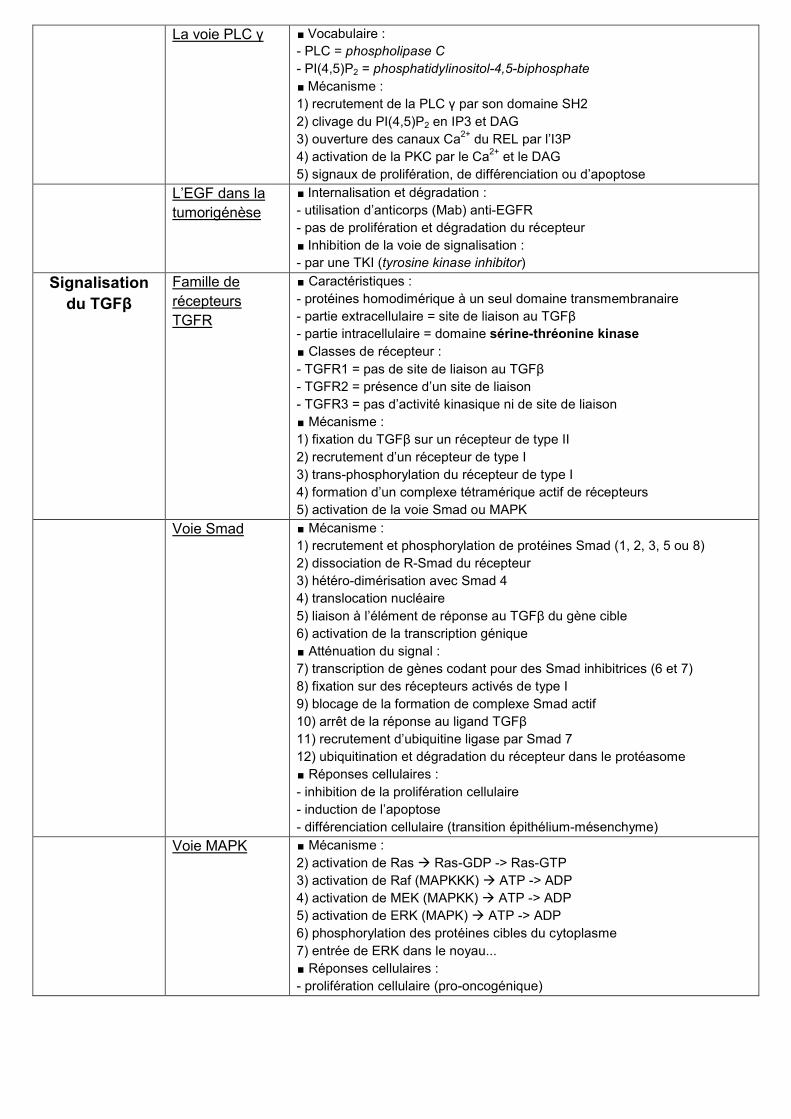
La voie PLC γ ∎ Vocabulaire :
- PLC = phospholipase C
- PI(4,5)P2 = phosphatidylinositol-4,5-biphosphate
∎ Mécanisme :
1) recrutement de la PLC γ par son domaine SH2
2) clivage du PI(4,5)P2 en IP3 et DAG
3) ouverture des canaux Ca2+
du REL par l’I3P
4) activation de la PKC par le Ca2+
et le DAG
5) signaux de prolifération, de différenciation ou d’apoptose
L’EGF dans la
tumorigénèse
∎ Internalisation et dégradation :
- utilisation d’anticorps (Mab) anti-EGFR
- pas de prolifération et dégradation du récepteur
∎ Inhibition de la voie de signalisation :
- par une TKI (tyrosine kinase inhibitor)
Signalisation
du TGFβ
Famille de
récepteurs
TGFR
∎ Caractéristiques :
- protéines homodimérique à un seul domaine transmembranaire
- partie extracellulaire = site de liaison au TGFβ
- partie intracellulaire = domaine sérine-thréonine kinase
∎ Classes de récepteur :
- TGFR1 = pas de site de liaison au TGFβ
- TGFR2 = présence d’un site de liaison
- TGFR3 = pas d’activité kinasique ni de site de liaison
∎ Mécanisme :
1) fixation du TGFβ sur un récepteur de type II
2) recrutement d’un récepteur de type I
3) trans-phosphorylation du récepteur de type I
4) formation d’un complexe tétramérique actif de récepteurs
5) activation de la voie Smad ou MAPK
Voie Smad ∎ Mécanisme :
1) recrutement et phosphorylation de protéines Smad (1, 2, 3, 5 ou 8)
2) dissociation de R-Smad du récepteur
3) hétéro-dimérisation avec Smad 4
4) translocation nucléaire
5) liaison à l’élément de réponse au TGFβ du gène cible
6) activation de la transcription génique
∎ Atténuation du signal :
7) transcription de gènes codant pour des Smad inhibitrices (6 et 7)
8) fixation sur des récepteurs activés de type I
9) blocage de la formation de complexe Smad actif
10) arrêt de la réponse au ligand TGFβ
11) recrutement d’ubiquitine ligase par Smad 7
12) ubiquitination et dégradation du récepteur dans le protéasome
∎ Réponses cellulaires :
- inhibition de la prolifération cellulaire
- induction de l’apoptose
- différenciation cellulaire (transition épithélium-mésenchyme)
Voie MAPK ∎ Mécanisme :
2) activation de Ras Ras-GDP -> Ras-GTP
3) activation de Raf (MAPKKK) ATP -> ADP
4) activation de MEK (MAPKK) ATP -> ADP
5) activation de ERK (MAPK) ATP -> ADP
6) phosphorylation des protéines cibles du cytoplasme
7) entrée de ERK dans le noyau...
∎ Réponses cellulaires :
- prolifération cellulaire (pro-oncogénique)
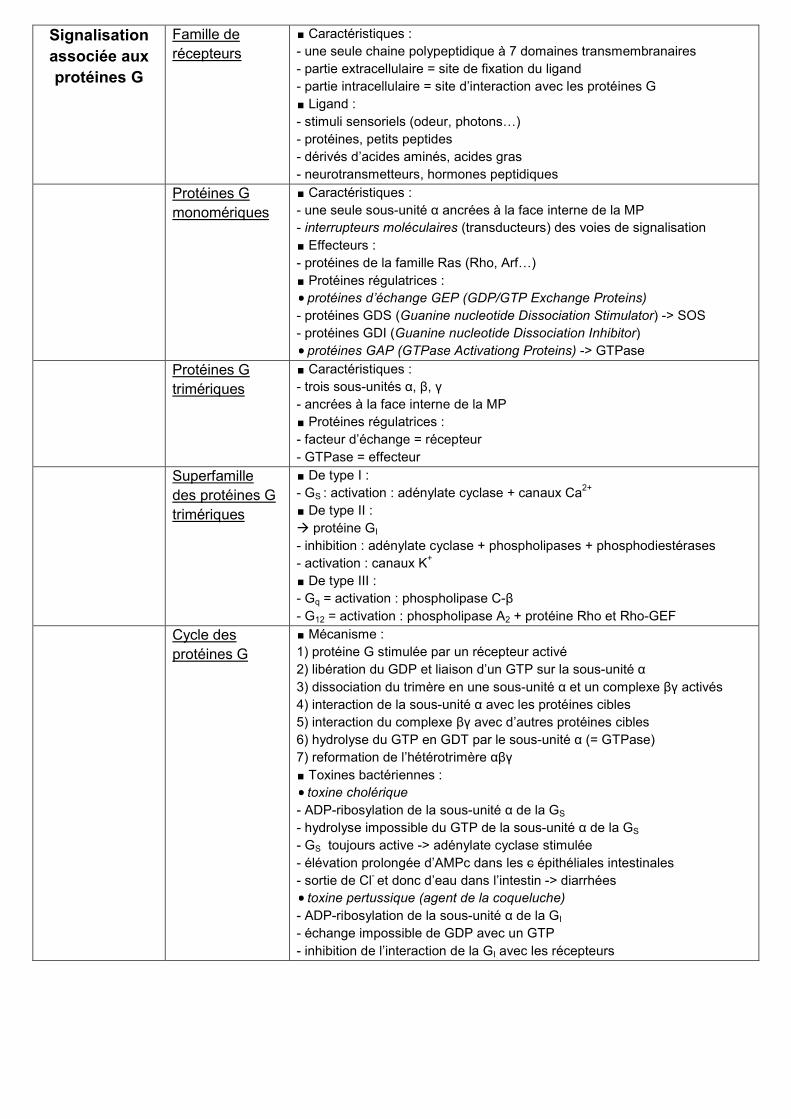
Signalisation
associée aux
protéines G
Famille de
récepteurs
∎ Caractéristiques :
- une seule chaine polypeptidique à 7 domaines transmembranaires
- partie extracellulaire = site de fixation du ligand
- partie intracellulaire = site d’interaction avec les protéines G
∎ Ligand :
- stimuli sensoriels (odeur, photons…)
- protéines, petits peptides
- dérivés d’acides aminés, acides gras
- neurotransmetteurs, hormones peptidiques
Protéines G
monomériques
∎ Caractéristiques :
- une seule sous-unité α ancrées à la face interne de la MP
- interrupteurs moléculaires (transducteurs) des voies de signalisation
∎ Effecteurs :
- protéines de la famille Ras (Rho, Arf…)
∎ Protéines régulatrices :
protéines d’échange GEP (GDP/GTP Exchange Proteins)
- protéines GDS (Guanine nucleotide Dissociation Stimulator) -> SOS
- protéines GDI (Guanine nucleotide Dissociation Inhibitor)
protéines GAP (GTPase Activationg Proteins) -> GTPase
Protéines G
trimériques
∎ Caractéristiques :
- trois sous-unités α, β, γ
- ancrées à la face interne de la MP
∎ Protéines régulatrices :
- facteur d’échange = récepteur
- GTPase = effecteur
Superfamille
des protéines G
trimériques
∎ De type I :
- GS : activation : adénylate cyclase + canaux Ca2+
∎ De type II :
protéine GI
- inhibition : adénylate cyclase + phospholipases + phosphodiestérases
- activation : canaux K+
∎ De type III :
- Gq = activation : phospholipase C-β
- G12 = activation : phospholipase A2 + protéine Rho et Rho-GEF
Cycle des
protéines G
∎ Mécanisme :
1) protéine G stimulée par un récepteur activé
2) libération du GDP et liaison d’un GTP sur la sous-unité α
3) dissociation du trimère en une sous-unité α et un complexe βγ activés
4) interaction de la sous-unité α avec les protéines cibles
5) interaction du complexe βγ avec d’autres protéines cibles
6) hydrolyse du GTP en GDT par le sous-unité α (= GTPase)
7) reformation de l’hétérotrimère αβγ
∎ Toxines bactériennes :
toxine cholérique
- ADP-ribosylation de la sous-unité α de la GS
- hydrolyse impossible du GTP de la sous-unité α de la GS
- GS toujours active -> adénylate cyclase stimulée
- élévation prolongée d’AMPc dans les c épithéliales intestinales
- sortie de Cl- et donc d’eau dans l’intestin -> diarrhées
toxine pertussique (agent de la coqueluche)
- ADP-ribosylation de la sous-unité α de la GI
- échange impossible de GDP avec un GTP
- inhibition de l’interaction de la GI avec les récepteurs
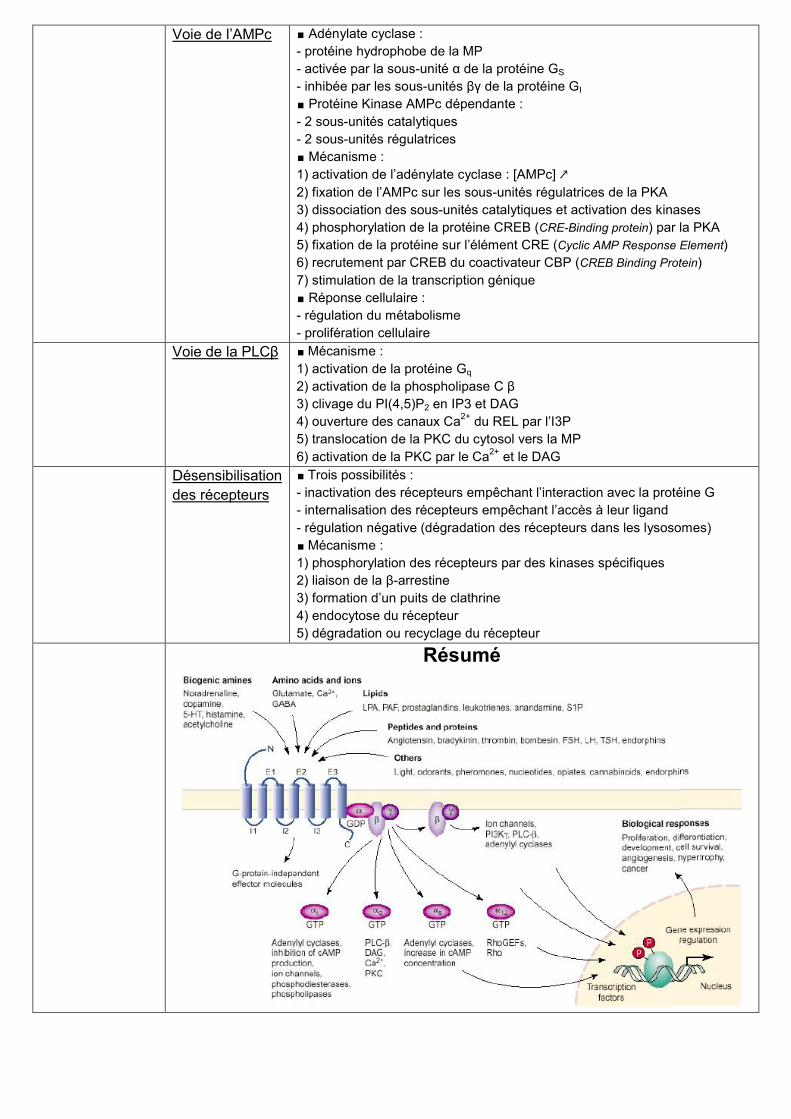
Voie de l’AMPc ∎ Adénylate cyclase :
- protéine hydrophobe de la MP
- activée par la sous-unité α de la protéine GS
- inhibée par les sous-unités βγ de la protéine GI
∎ Protéine Kinase AMPc dépendante :
- 2 sous-unités catalytiques
- 2 sous-unités régulatrices
∎ Mécanisme :
1) activation de l’adénylate cyclase : [AMPc] ↗
2) fixation de l’AMPc sur les sous-unités régulatrices de la PKA
3) dissociation des sous-unités catalytiques et activation des kinases
4) phosphorylation de la protéine CREB (CRE-Binding protein) par la PKA
5) fixation de la protéine sur l’élément CRE (Cyclic AMP Response Element)
6) recrutement par CREB du coactivateur CBP (CREB Binding Protein)
7) stimulation de la transcription génique
∎ Réponse cellulaire :
- régulation du métabolisme
- prolifération cellulaire
Voie de la PLCβ ∎ Mécanisme :
1) activation de la protéine Gq
2) activation de la phospholipase C β
3) clivage du PI(4,5)P2 en IP3 et DAG
4) ouverture des canaux Ca2+
du REL par l’I3P
5) translocation de la PKC du cytosol vers la MP
6) activation de la PKC par le Ca2+
et le DAG
Désensibilisation
des récepteurs
∎ Trois possibilités :
- inactivation des récepteurs empêchant l’interaction avec la protéine G
- internalisation des récepteurs empêchant l’accès à leur ligand
- régulation négative (dégradation des récepteurs dans les lysosomes)
∎ Mécanisme :
1) phosphorylation des récepteurs par des kinases spécifiques
2) liaison de la β-arrestine
3) formation d’un puits de clathrine
4) endocytose du récepteur
5) dégradation ou recyclage du récepteur
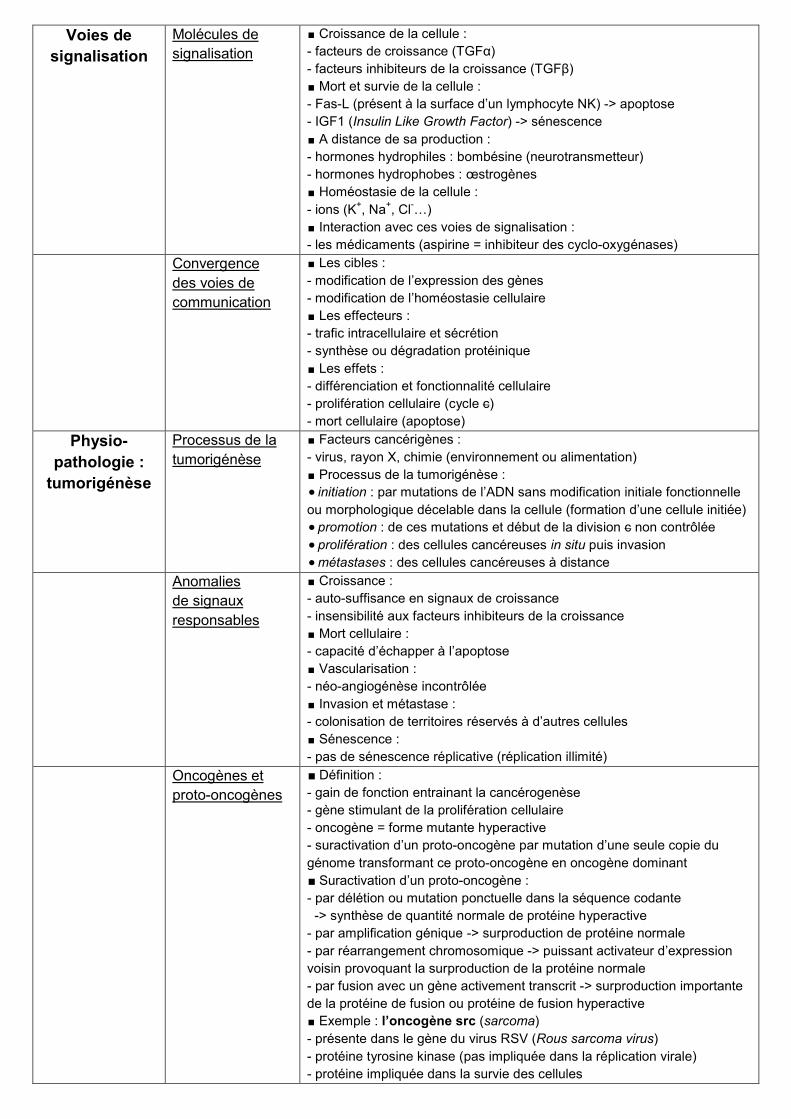
Résumé
Voies de
signalisation
Molécules de
signalisation
∎ Croissance de la cellule :
- facteurs de croissance (TGFα)
- facteurs inhibiteurs de la croissance (TGFβ)
∎ Mort et survie de la cellule :
- Fas-L (présent à la surface d’un lymphocyte NK) -> apoptose
- IGF1 (Insulin Like Growth Factor) -> sénescence
∎ A distance de sa production :
- hormones hydrophiles : bombésine (neurotransmetteur)
- hormones hydrophobes : œstrogènes
∎ Homéostasie de la cellule :
- ions (K+, Na
+, Cl
-…)
∎ Interaction avec ces voies de signalisation :
- les médicaments (aspirine = inhibiteur des cyclo-oxygénases)
Convergence
des voies de
communication
∎ Les cibles :
- modification de l’expression des gènes
- modification de l’homéostasie cellulaire
∎ Les effecteurs :
- trafic intracellulaire et sécrétion
- synthèse ou dégradation protéinique
∎ Les effets :
- différenciation et fonctionnalité cellulaire
- prolifération cellulaire (cycle c)
- mort cellulaire (apoptose)
Physio-
pathologie :
tumorigénèse
Processus de la
tumorigénèse
∎ Facteurs cancérigènes :
- virus, rayon X, chimie (environnement ou alimentation)
∎ Processus de la tumorigénèse :
initiation : par mutations de l’ADN sans modification initiale fonctionnelle
ou morphologique décelable dans la cellule (formation d’une cellule initiée)
promotion : de ces mutations et début de la division c non contrôlée
prolifération : des cellules cancéreuses in situ puis invasion
métastases : des cellules cancéreuses à distance
Anomalies
de signaux
responsables
∎ Croissance :
- auto-suffisance en signaux de croissance
- insensibilité aux facteurs inhibiteurs de la croissance
∎ Mort cellulaire :
- capacité d’échapper à l’apoptose
∎ Vascularisation :
- néo-angiogénèse incontrôlée
∎ Invasion et métastase :
- colonisation de territoires réservés à d’autres cellules
∎ Sénescence :
- pas de sénescence réplicative (réplication illimité)
Oncogènes et
proto-oncogènes
∎ Définition :
- gain de fonction entrainant la cancérogenèse
- gène stimulant de la prolifération cellulaire
- oncogène = forme mutante hyperactive
- suractivation d’un proto-oncogène par mutation d’une seule copie du
génome transformant ce proto-oncogène en oncogène dominant
∎ Suractivation d’un proto-oncogène :
- par délétion ou mutation ponctuelle dans la séquence codante
-> synthèse de quantité normale de protéine hyperactive
- par amplification génique -> surproduction de protéine normale
- par réarrangement chromosomique -> puissant activateur d’expression
voisin provoquant la surproduction de la protéine normale
- par fusion avec un gène activement transcrit -> surproduction importante
de la protéine de fusion ou protéine de fusion hyperactive
∎ Exemple : l’oncogène src (sarcoma)
- présente dans le gène du virus RSV (Rous sarcoma virus)
- protéine tyrosine kinase (pas impliquée dans la réplication virale)
- protéine impliquée dans la survie des cellules

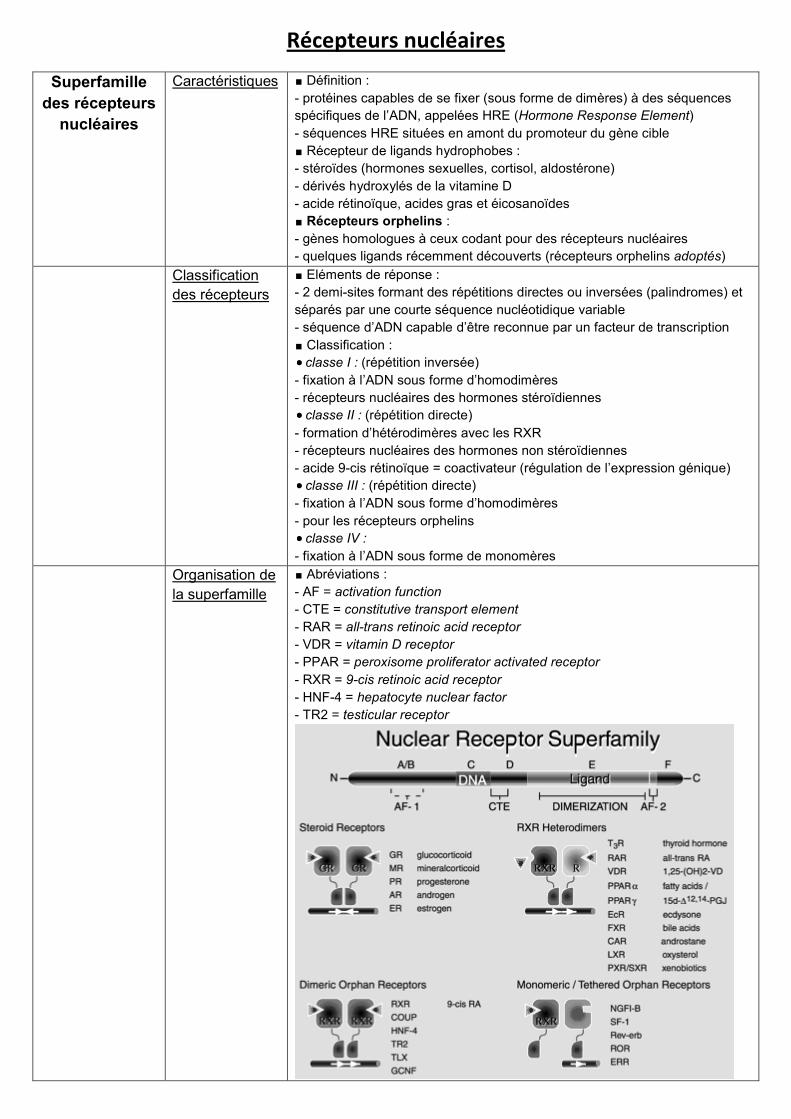
Récepteurs nucléaires
Superfamille
des récepteurs
nucléaires
Caractéristiques ∎ Définition :
- protéines capables de se fixer (sous forme de dimères) à des séquences
spécifiques de l’ADN, appelées HRE (Hormone Response Element)
- séquences HRE situées en amont du promoteur du gène cible
∎ Récepteur de ligands hydrophobes :
- stéroïdes (hormones sexuelles, cortisol, aldostérone)
- dérivés hydroxylés de la vitamine D
- acide rétinoïque, acides gras et éicosanoïdes
∎ Récepteurs orphelins :
- gènes homologues à ceux codant pour des récepteurs nucléaires
- quelques ligands récemment découverts (récepteurs orphelins adoptés)
Classification
des récepteurs
∎ Eléments de réponse :
- 2 demi-sites formant des répétitions directes ou inversées (palindromes) et
séparés par une courte séquence nucléotidique variable
- séquence d’ADN capable d’être reconnue par un facteur de transcription
∎ Classification :
classe I : (répétition inversée)
- fixation à l’ADN sous forme d’homodimères
- récepteurs nucléaires des hormones stéroïdiennes
classe II : (répétition directe)
- formation d’hétérodimères avec les RXR
- récepteurs nucléaires des hormones non stéroïdiennes
- acide 9-cis rétinoïque = coactivateur (régulation de l’expression génique)
classe III : (répétition directe)
- fixation à l’ADN sous forme d’homodimères
- pour les récepteurs orphelins
classe IV :
- fixation à l’ADN sous forme de monomères
Organisation de
la superfamille
∎ Abréviations :
- AF = activation function
- CTE = constitutive transport element
- RAR = all-trans retinoic acid receptor
- VDR = vitamin D receptor
- PPAR = peroxisome proliferator activated receptor
- RXR = 9-cis retinoic acid receptor
- HNF-4 = hepatocyte nuclear factor
- TR2 = testicular receptor
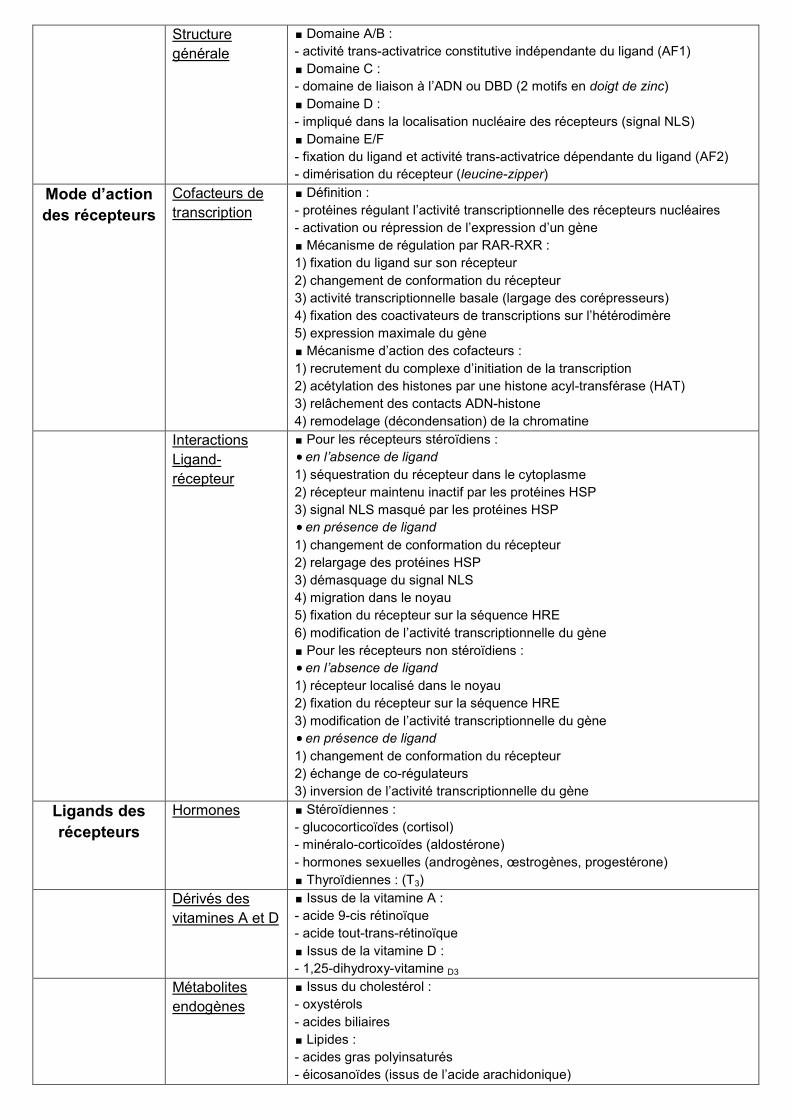
Structure
générale
∎ Domaine A/B :
- activité trans-activatrice constitutive indépendante du ligand (AF1)
∎ Domaine C :
- domaine de liaison à l’ADN ou DBD (2 motifs en doigt de zinc)
∎ Domaine D :
- impliqué dans la localisation nucléaire des récepteurs (signal NLS)
∎ Domaine E/F
- fixation du ligand et activité trans-activatrice dépendante du ligand (AF2)
- dimérisation du récepteur (leucine-zipper)
Mode d’action
des récepteurs
Cofacteurs de
transcription
∎ Définition :
- protéines régulant l’activité transcriptionnelle des récepteurs nucléaires
- activation ou répression de l’expression d’un gène
∎ Mécanisme de régulation par RAR-RXR :
1) fixation du ligand sur son récepteur
2) changement de conformation du récepteur
3) activité transcriptionnelle basale (largage des corépresseurs)
4) fixation des coactivateurs de transcriptions sur l’hétérodimère
5) expression maximale du gène
∎ Mécanisme d’action des cofacteurs :
1) recrutement du complexe d’initiation de la transcription
2) acétylation des histones par une histone acyl-transférase (HAT)
3) relâchement des contacts ADN-histone
4) remodelage (décondensation) de la chromatine
Interactions
Ligand-
récepteur
∎ Pour les récepteurs stéroïdiens :
en l’absence de ligand
1) séquestration du récepteur dans le cytoplasme
2) récepteur maintenu inactif par les protéines HSP
3) signal NLS masqué par les protéines HSP
en présence de ligand
1) changement de conformation du récepteur
2) relargage des protéines HSP
3) démasquage du signal NLS
4) migration dans le noyau
5) fixation du récepteur sur la séquence HRE
6) modification de l’activité transcriptionnelle du gène
∎ Pour les récepteurs non stéroïdiens :
en l’absence de ligand
1) récepteur localisé dans le noyau
2) fixation du récepteur sur la séquence HRE
3) modification de l’activité transcriptionnelle du gène
en présence de ligand
1) changement de conformation du récepteur
2) échange de co-régulateurs
3) inversion de l’activité transcriptionnelle du gène
Ligands des
récepteurs
Hormones ∎ Stéroïdiennes :
- glucocorticoïdes (cortisol)
- minéralo-corticoïdes (aldostérone)
- hormones sexuelles (androgènes, œstrogènes, progestérone)
∎ Thyroïdiennes : (T3)
Dérivés des
vitamines A et D
∎ Issus de la vitamine A :
- acide 9-cis rétinoïque
- acide tout-trans-rétinoïque
∎ Issus de la vitamine D :
- 1,25-dihydroxy-vitamine D3
Métabolites
endogènes
∎ Issus du cholestérol :
- oxystérols
- acides biliaires
∎ Lipides :
- acides gras polyinsaturés
- éicosanoïdes (issus de l’acide arachidonique)
Le cortisol ∎ Régulation du jeûne :
1) hypoglycémie
2) sécrétion de corticotrophine (ACTH) par l’hypophyse
-> ACTH = adreno-corticotropic hormon
3) sécrétion de cortisol par les surrénales
4) liaison du cortisol à son récepteur au niveau du foie
5) entrée de ce facteur trans-régulateur dans le noyau des hépatocytes
6) liaison du trans-régulateur sur l’élément GRE
-> GRE = glucocorticoid responsive element (cis-régulateur)
7) activation du gène en aval et synthèse accrue de messagers
8) synthèse d’enzymes de la gluconéogenèse (F16BPase, G6Pase, PEPCK)
9) transformation des aa en glucose -> augmentation glycémie
∎ Régulation de l’effort:
1) stress
2) activation des médullo-surrénales par le cerveau -> sécrétion adrénaline
3) sécrétion de glucagon par pancréas (si hypoglycémie)
4) synthèse d’AMPc par les cellules musculaires et hépatiques
5) activation de la PKA (protéine kinase A)
6) phosphorylation de la CREB (facteur trans-régulateur)
-> CREB = cAMP Responsive Element Binding
7) liaison du CREB-P sur le CRE (cis-régulateur)
8) activation du gène en aval et synthèse accrue de messagers
9) synthèse d’enzymes de la glycogénolyse (glycogène phosphorylase)
10) glycogène -> glucose (énergie)
La vitamine D ∎ Au niveau de la peau :
1) action des UV sur le 7-déhydrocholestérol
2) synthèse de la vitamine D3 (cholécalciférol)
∎ Dans le foie :
3) hydroxylation par la 25-hydroxylase
4) formation du 25-OH-vitamine D3
∎ Dans le rein : (action de la PTH)
5) hydroxylation par la 1-hydroxylase
6) formation du 1,25-dihydroxyvitamine D3 (calcitriol)
∎ Dans l’intestin :
7) absorption du calcium et du phosphore
∎ Dans les chondrocytes :
8) hydroxylation par la 24-hydroxylase
9) formation du 1,24,25-trihydroxyvitamine D3
10) fixation du calcium sur l’os
Les acides
biliaires
∎ Formation des acides biliaires :
- à partir du cholestérol
∎ Cycle entéro-hépatique des sels biliaires :
acides biliaires primaires
- acide cholique et acide chénodésoxycholique
- excrétion de ces sels dans la bile puis dans l’intestin
- indispensables à l’action de la lipase pancréatique
acides biliaires secondaires
- acide cholique -> acide désoxycholique
- acide chénodésoxycholique -> acide lithocholique
- réabsorption dans l’iléon puis transport par la veine porte vers le foie
- recapture par le foie ou excrétion dans les urines
acides biliaires tertiaires
- acide lithocholique -> acide sulfolithocholique
- élimination dans les fèces (selles) d’une partie des acides biliaires
Pathologie et
pharmacologie
Cicatrisation
cutanée
∎ Phase vasculaire et inflammatoire :
étape vasculaire
1) mise à nu de l’endothélium vasculaire
2) adhésion et activation des plaquettes
3) formation d’un caillot pour arrêter le saignement et constituer une matrice
provisoire servant à la migration des cellules
4) libération de facteurs de croissance (PDGF, FGF, TGF) par les plaquettes
étape inflammatoire
1) polynucléaires attirés au niveau de la zone cicatricielle
2) libération par les neutrophiles d’enzymes protéolytiques favorisant la
pénétration des cellules dans la plaie
3) libération par les neutrophiles de cytokines pro-inflammatoires participant à
la migration et à la prolifération des fibroblastes et kératinocytes
4) différenciation des monocytes de la plaie en macrophage
5) libération de facteurs de croissance par les monocytes (TGF, TNF, VGF)
6) amplification de la réponse inflammatoire
7) stimulation de la formation du tissu de granulation
∎ Phase de réparation tissulaire :
formation du tissu de granulation
1) migration précoce des fibroblastes
2) synthèse et remodelage de la MEC composé de collagène III puis I
3) migration des cellules endothéliales à partir des vaisseaux
4) développement d’un néo-réseau vasculaire dans la zone cicatricielle
5) transformation progressive de certains fibroblastes en myofibroblastes
6) contraction de la plaie permettant le rapprochement des berges
épithélialisation
1) migration des cellules épithéliales à partir des berges de la plaie
2) plaie fermée par une monocouche de kératinocytes
3) formation progressive d’une membrane basale
4) colonisation secondaire de l’épiderme par les mélanocytes
∎ Phase de maturation et de remodelage :
- remodelage de la MEC dans les 2 mois suivant la fermeture de la plaie
- diminution progressive du tissu de granulation
- élaboration d’une structure collagénique plus dense
Polycystines ∎ Structure des polycystines :
- codées par les gènes PKD1 et 2 (Polycystin Kidney Disease)
polycystine 1
- protéine de la famille des mécano-récepteurs
- longue portion N-terminale extracellulaire
- 11 domaines transmembranaires
- courte portion C-terminale intracellulaire avec des domaines SH3 et des
sites de phosphorylation par Src, FAK, PKA…
polycystine 2
- protéine de la famille des canaux ioniques voltage dépendant
- 6 domaines transmembranaires
- 2 domaines intracellulaires N- et C-terminaux
- un domaine de liaison à l’actine
∎ Fonction des polycystines :
dans la cellule tubulaire
- complexe polycystine 1/2 = canal ionique mécano-sensible
- localisé au niveau de la membrane plasmique
au pôle apical
- complexe localisé au niveau de la surface du cil primaire
- transmission à la cellule les signaux de la lumière
au pôle basal
- polycystine 1 localisés au niveau des points d’adhérence focale
- transmission des signaux de la MEC aux points d’adhérence focale
dans le domaine latéral
- polycystine 1 localisée au niveau des jonctions adhérentes
- transmission à la cellule les signaux des cellules adjacentes
Cil primaire ∎ Structure :
- constitué de 9 doublets de μtubules (sans paire centrale)
- structure différente des cils motiles (9 doublets + 1 paire centrale)
∎ Fonction sensorielle :
1) contrainte mécanique
2) capture du signal par la polycystine 1
3) transmission de ce signal à la polycystine 2
4) influx de Ca2+
induit par la polycystine 2
5) activation des récepteurs de type ryanodine
6) libération du Ca2+
du réticulum endoplasmique
7) rôle dans la prolifération et la différenciation
Polykystose
hépatique et
rénale
∎ Polykystose rénale : (1/800)
- maladie autosomique dominante
- mutation des gènes codant pour les polycystines (PDK 1 et 2)
- souvent associée à une polykystose hépatique
∎ Polykystose hépatique isolée : (1/3000)
- maladie autosomique dominante
- mutation du gène codant pour l’hépatocystine

Les cellules et leur environnement
La matrice extracellulaire
Généralité Caractéristiques ∎ Définition :
- réseau tridimensionnel de macromolécules sécrétées par les cellules
formant le lien entre les différents groupes de cellules ou tissus
∎ Fonction :
- charpente des tissus
- support pour l’adhésion et la migration cellulaire
- libération de signaux relatifs à l’environnement des cellules
Localisation présente dans tous les tissus de l’organisme
- abondante dans les tissus conjonctifs lâches
- pauvre des les épithéliums
- rigide dans les tissus cartilagineux
- minéralisée dans les tissus osseux
Tissu
conjonctif
Composants ∎ Composants :
- fibroblastes sécrétant la MEC
- cellules du système immunitaire (macrophages et mastocytes)
- protéines fibreuses : collagène, élastine
- protéines d’adhérence : fibronectine, laminine
- GAG, protéoglycanes et glycoprotéines
Propriétés ∎ Propriétés :
- MEC plus volumineuse que les cellules l’entourant
- gel hydratée -> GAG, protéoglycanes et glycoprotéines
- résistance aux fortes pressions -> GAG
- résistance aux forces de tension -> collagène et élastine
Composants
de la MEC
Les GAG ∎ Caractéristiques :
- grandes chaînes polysaccharidiques non ramifiée faites de la répétition d’un
motif disaccharidique {acide uronique + osamine}
- fortement hydrophiles car forte charge négative (gpt sulfate ou carbonyle)
∎ GAG non liés à une protéine : -> pas de formation de protéoglycanes
- acide hyaluronique (acide glucuronique + N-acétyl-glucosamine)
-> le seul non sulfaté
∎ GAG liés à une protéine :
- chondroïtine, kératane, dermatane et héparane sulfate, héparine
Les
protéoglycanes
∎ Structure :
- liaison covalente de GAG à un noyau protéique
- décorine = 1 noyau protéique + 1 GAG (chondroïtine/dermatane sulfate)
-> dans tous les tissus conjonctifs
- perlécan = 1 noyau protéique + 1 GAG (héparane sulfate)
-> dans les membranes basales
- aggrécan = 1 noyau protéique + plusieurs GAG -> dans le cartilage
∎ Organisation en agrégat :
- au centre = acide hyaluronique
- protéoglycanes liés à l’acide hyaluronique par l’intermédiaire de protéines
de liaisons (liaisons non covalentes)
Le collagène ∎ Caractéristiques :
- glycoprotéine fibreuse principale de la MEC
- protéine la plus abondante de l’organisme
- dégradation par des collagénases (métalloprotéinase matricielle)
- 22 collagènes différents identifiés
- tropocollagène formés de 3 chaines α enroulées en hélice
- 42 chaines α différentes : motif COL (glycine, proline et hydroxyproline)
∎ Différents types :
20 collagènes différents identifiés
- collagène fibreux ou fibrillaire (I, II, III)
- collagène non fibreux, en réseau (IV)
- collagène associé aux membranes cellulaires

Composants
de la MEC
L’élastine ∎ Caractéristiques :
- sécrétée sous forme de tropoélastine
- protéine très élastique car fortement hydrophobe
- composant amorphe, hydrophobe riche en proline et glycine
∎ Fibres élastiques :
- composé d’élastine principalement mais aussi de fibrilline
- présente dans les tissus les plus élastiques (vaisseaux…)
∎ Syndrome de Marfan :
- mutation du gène de la fibrilline 1
- risques de rupture de l’aorte
La fibronectine ∎ Caractéristiques :
- grosse glycoprotéine de 2500 acides aminés - hétérodimère constitué de 2 grosses sous-unités réunies par 2 ponts S-S à l’extrémité C-term ∎ Domaine protéique particulier :
- la répétition de type III de la fibronectine -> liaison à l’intégrine
↳ la séquence RGD (Arg-Glu-Asp) -> séquence de liaison aux intégrines
∎ Sites d’interaction pour : - le collagène - les GAG (héparine) et protéoglycanes (perlécan) - l’intégrine ∎ Rôles : - adhérence cellulaire avec le tissu conjonctif - communication cellulaire
Membrane
basale
Caractéristiques ∎ Définition : - tapis souple et fin formé par une MEC spécifique ∎ Rôles : - structural en séparant les cellules épithéliales du tissus conj. sous jacent ou en entourant les cellules musculaires, adipeuses et de Schwann - de filtre sélectif en séparant 2 feuillets cellulaires distincts (glomérule rénal) - pour déterminer la polarité cellulaire - pour la migration cellulaire
Composants ∎ Composants : - collagène IV - laminine - nidogène (= entactine) - protéoglycane (perlécan) ∎ Laminine : - 3 chaînes polypeptidiques (α, β1, β2) très longues disposées en forme de croix asymétrique et maintenues ensemble par des ponts S-S - chaîne A : 3 domaines globulaires + une longue queue - chaînes B : 2 domaines globulaires + un domaine α-hélicoïdal super-enroulé autour de la queue de la chaîne A - sites d’interaction avec le perlécan, le nidogène et les récepteurs protéiques des laminines à la surface des cellules (dystroglycane) ∎ Nidogène : - appelé aussi entactine - sites d’interaction avec la laminine, le collagène IV et le perlécan - rôle dans le maintien de la structure et dans l’assemblage de la lame basale ∎ Interaction entre les ≠ composants :
Les interactions cellule-cellule
Jonctions
serrées Jonction étanche
Protéines
d’adhésion
membranaires
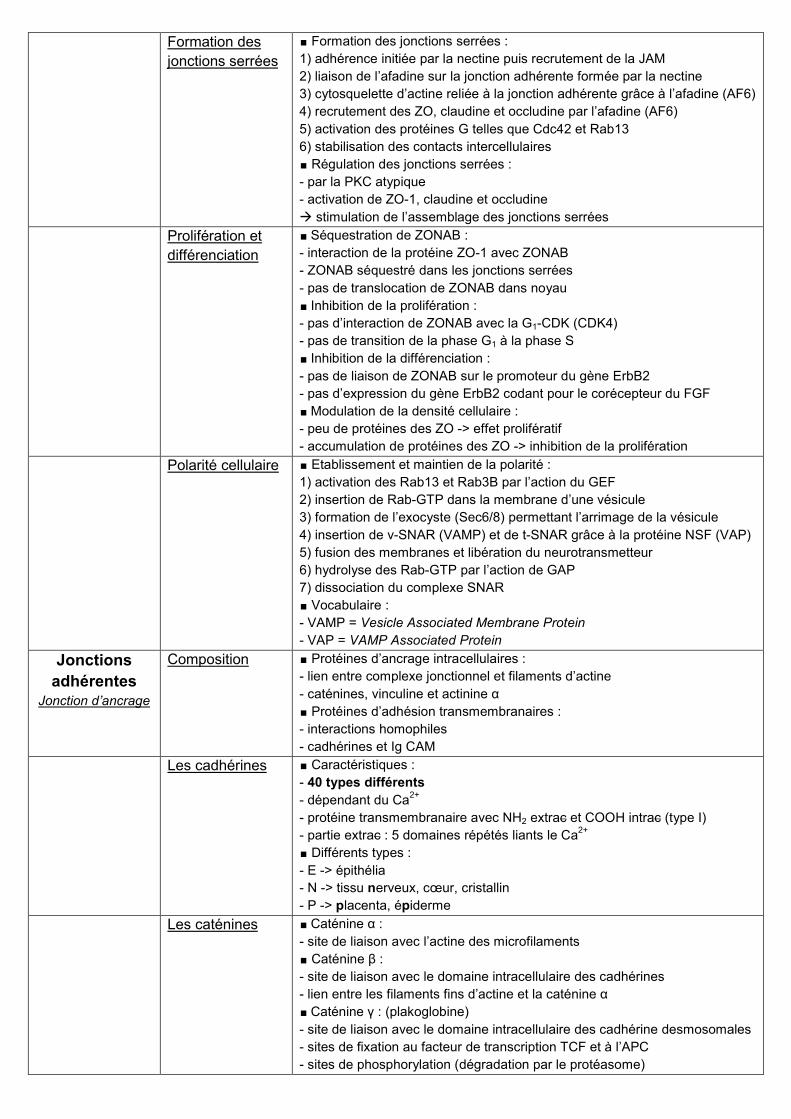
∎ Les claudines et occludines :
- protéines à 4 domaines transmembranaires (= tétraspanines)
- COOH et NH2 intracellulaires, interactions homophiles
- 2 boucles extracellulaires hydrophobes (occludine) ou chargées (claudine)
∎ La protéine JAM et la nectine :
- JAM = Junctional Adhesion molecule
- m d’adhérences de la famille des Ig permettant d’initier l’adhésion c / c
Protéines
de la plaque
cytoplasmique
∎ Protéines adaptatrices :
caractéristiques
- domaine PDZ (post-synaptic disc large zona) permettant la reconnaissance
d’un motif peptidique (Ser/Thr-X-Val/Iso) des protéines membranaires
- rôle dans la liaison des protéines membranaires avec le cytosquelette
d’actine ou avec des protéines régulatrices
protéines ZO (zonula occludens)
- ZO-1 : interaction avec les claudines et l’afadine
- ZO-2 et ZO-3 : interaction avec les occludines et l’afadine
autres protéines adaptatrices
- ASIP = Atypical isotype-Specific Interacting Protein
- MAGUK = Membrane Associated Guanylate Kinase
- AF-6 = Acute lymphoblastic leukemia Fusion (from chromosome 6)
- cinguline = domaines coiled-coil interagissant avec les ZO et la myosine
∎ Protéines régulatrices :
- Rab13 et Rab3B
- cdc42 = cell division control protein
- ZONAB = ZO-1 associated nucleic acid protein
∎ Protéines du cytosquelette :
actine, myosine
- filaments fins d’actine (microfilaments)
- filaments épais de myosine
cinguline
- domaines coiled-coil interagissant avec les ZO et la myosine
- rôle dans la contraction actine-myosine
Protein
interactions at the
tight junction
Formation des
jonctions serrées
∎ Formation des jonctions serrées :
1) adhérence initiée par la nectine puis recrutement de la JAM
2) liaison de l’afadine sur la jonction adhérente formée par la nectine
3) cytosquelette d’actine reliée à la jonction adhérente grâce à l’afadine (AF6)
4) recrutement des ZO, claudine et occludine par l’afadine (AF6)
5) activation des protéines G telles que Cdc42 et Rab13
6) stabilisation des contacts intercellulaires
∎ Régulation des jonctions serrées :
- par la PKC atypique
- activation de ZO-1, claudine et occludine
stimulation de l’assemblage des jonctions serrées
Prolifération et
différenciation
∎ Séquestration de ZONAB :
- interaction de la protéine ZO-1 avec ZONAB
- ZONAB séquestré dans les jonctions serrées
- pas de translocation de ZONAB dans noyau
∎ Inhibition de la prolifération :
- pas d’interaction de ZONAB avec la G1-CDK (CDK4)
- pas de transition de la phase G1 à la phase S
∎ Inhibition de la différenciation :
- pas de liaison de ZONAB sur le promoteur du gène ErbB2
- pas d’expression du gène ErbB2 codant pour le corécepteur du FGF
∎ Modulation de la densité cellulaire :
- peu de protéines des ZO -> effet prolifératif
- accumulation de protéines des ZO -> inhibition de la prolifération
Polarité cellulaire ∎ Etablissement et maintien de la polarité :
1) activation des Rab13 et Rab3B par l’action du GEF
2) insertion de Rab-GTP dans la membrane d’une vésicule
3) formation de l’exocyste (Sec6/8) permettant l’arrimage de la vésicule
4) insertion de v-SNAR (VAMP) et de t-SNAR grâce à la protéine NSF (VAP)
5) fusion des membranes et libération du neurotransmetteur
6) hydrolyse des Rab-GTP par l’action de GAP
7) dissociation du complexe SNAR
∎ Vocabulaire :
- VAMP = Vesicle Associated Membrane Protein
- VAP = VAMP Associated Protein
Jonctions
adhérentes Jonction d’ancrage
Composition ∎ Protéines d’ancrage intracellulaires :
- lien entre complexe jonctionnel et filaments d’actine
- caténines, vinculine et actinine α
∎ Protéines d’adhésion transmembranaires :
- interactions homophiles
- cadhérines et Ig CAM
Les cadhérines ∎ Caractéristiques :
- 40 types différents
- dépendant du Ca2+
- protéine transmembranaire avec NH2 extrac et COOH intrac (type I)
- partie extrac : 5 domaines répétés liants le Ca2+
∎ Différents types :
- E -> épithélia
- N -> tissu nerveux, cœur, cristallin
- P -> placenta, épiderme
Les caténines ∎ Caténine α :
- site de liaison avec l’actine des microfilaments
∎ Caténine β :
- site de liaison avec le domaine intracellulaire des cadhérines
- lien entre les filaments fins d’actine et la caténine α
∎ Caténine γ : (plakoglobine)
- site de liaison avec le domaine intracellulaire des cadhérine desmosomales
- sites de fixation au facteur de transcription TCF et à l’APC
- sites de phosphorylation (dégradation par le protéasome)
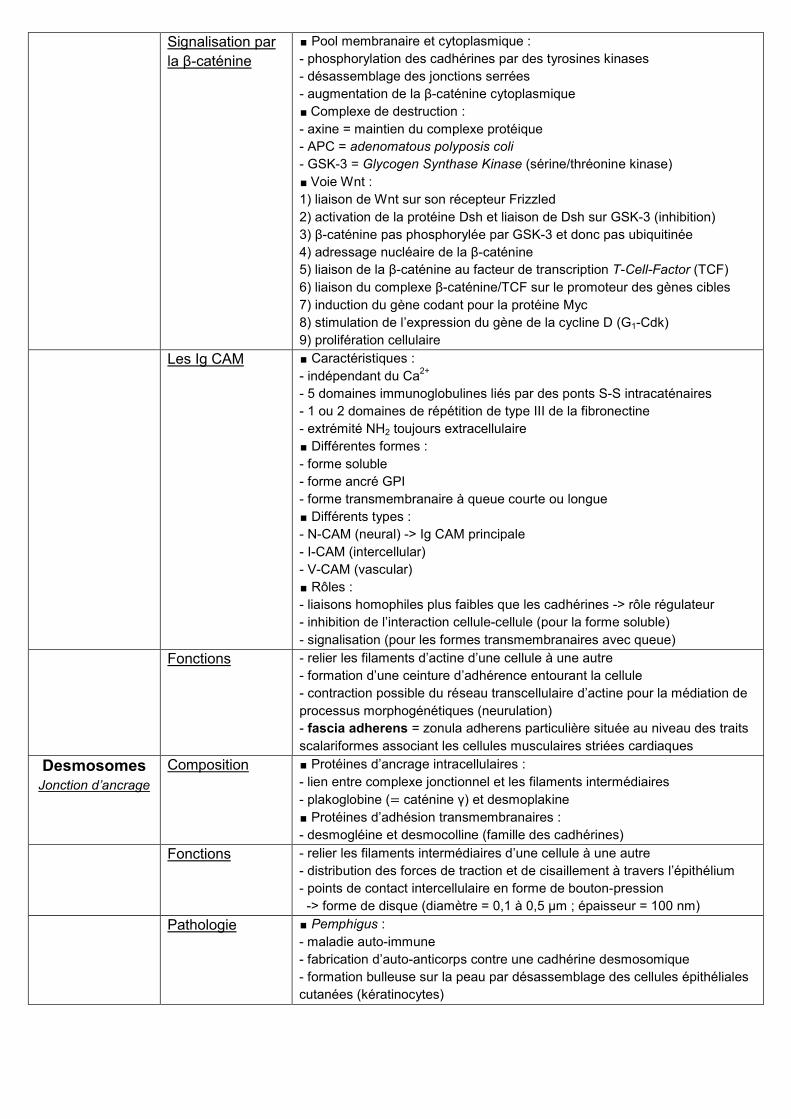
Desmosomes Jonction d’ancrage
Composition ∎ Protéines d’ancrage intracellulaires :
- lien entre complexe jonctionnel et les filaments intermédiaires
- plakoglobine (= caténine γ) et desmoplakine
∎ Protéines d’adhésion transmembranaires :
- desmogléine et desmocolline (famille des cadhérines)
Fonctions - relier les filaments intermédiaires d’une cellule à une autre
- distribution des forces de traction et de cisaillement à travers l’épithélium
- points de contact intercellulaire en forme de bouton-pression
-> forme de disque (diamètre = 0,1 à 0,5 μm ; épaisseur = 100 nm)
Pathologie ∎ Pemphigus :
- maladie auto-immune
- fabrication d’auto-anticorps contre une cadhérine desmosomique
- formation bulleuse sur la peau par désassemblage des cellules épithéliales
cutanées (kératinocytes)
Signalisation par
la β-caténine
∎ Pool membranaire et cytoplasmique :
- phosphorylation des cadhérines par des tyrosines kinases
- désassemblage des jonctions serrées
- augmentation de la β-caténine cytoplasmique
∎ Complexe de destruction :
- axine = maintien du complexe protéique
- APC = adenomatous polyposis coli
- GSK-3 = Glycogen Synthase Kinase (sérine/thréonine kinase)
∎ Voie Wnt :
1) liaison de Wnt sur son récepteur Frizzled
2) activation de la protéine Dsh et liaison de Dsh sur GSK-3 (inhibition)
3) β-caténine pas phosphorylée par GSK-3 et donc pas ubiquitinée
4) adressage nucléaire de la β-caténine
5) liaison de la β-caténine au facteur de transcription T-Cell-Factor (TCF)
6) liaison du complexe β-caténine/TCF sur le promoteur des gènes cibles
7) induction du gène codant pour la protéine Myc
8) stimulation de l’expression du gène de la cycline D (G1-Cdk)
9) prolifération cellulaire
Les Ig CAM ∎ Caractéristiques :
- indépendant du Ca2+
- 5 domaines immunoglobulines liés par des ponts S-S intracaténaires
- 1 ou 2 domaines de répétition de type III de la fibronectine
- extrémité NH2 toujours extracellulaire
∎ Différentes formes :
- forme soluble
- forme ancré GPI
- forme transmembranaire à queue courte ou longue
∎ Différents types :
- N-CAM (neural) -> Ig CAM principale
- I-CAM (intercellular)
- V-CAM (vascular)
∎ Rôles :
- liaisons homophiles plus faibles que les cadhérines -> rôle régulateur
- inhibition de l’interaction cellule-cellule (pour la forme soluble)
- signalisation (pour les formes transmembranaires avec queue)
Fonctions - relier les filaments d’actine d’une cellule à une autre
- formation d’une ceinture d’adhérence entourant la cellule
- contraction possible du réseau transcellulaire d’actine pour la médiation de
processus morphogénétiques (neurulation)
- fascia adherens = zonula adherens particulière située au niveau des traits
scalariformes associant les cellules musculaires striées cardiaques
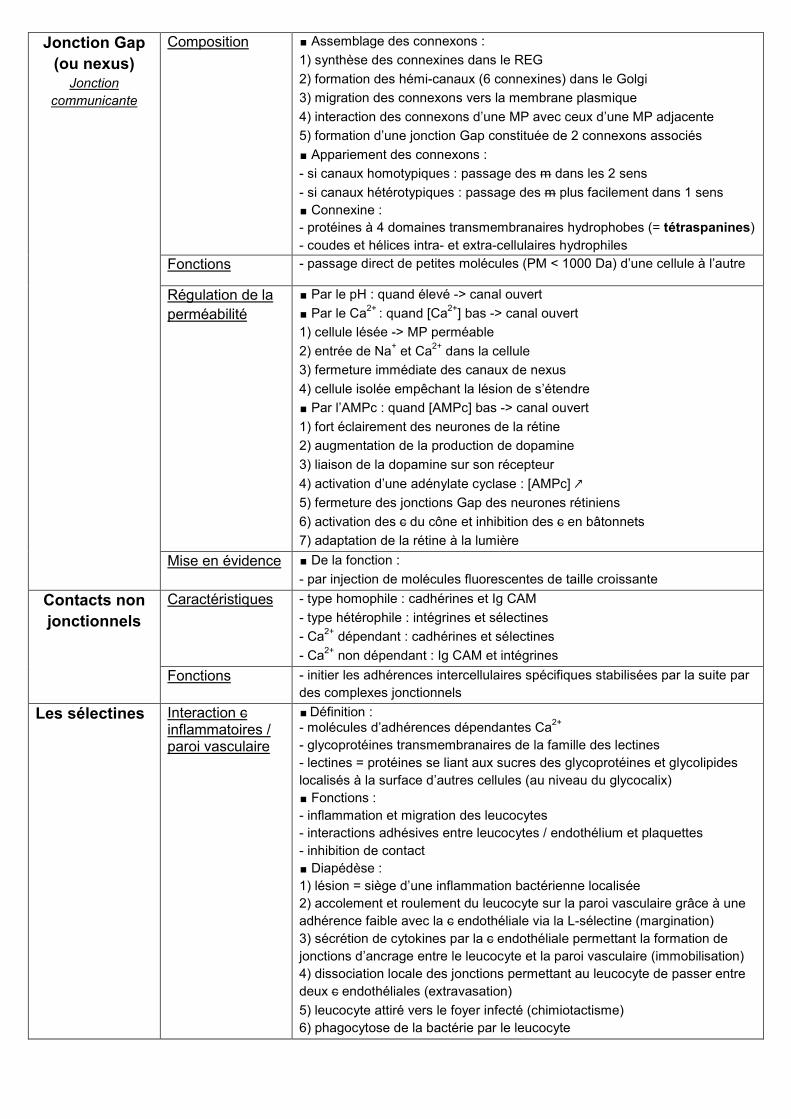
Jonction Gap
(ou nexus) Jonction
communicante
Composition ∎ Assemblage des connexons :
1) synthèse des connexines dans le REG
2) formation des hémi-canaux (6 connexines) dans le Golgi
3) migration des connexons vers la membrane plasmique
4) interaction des connexons d’une MP avec ceux d’une MP adjacente
5) formation d’une jonction Gap constituée de 2 connexons associés
∎ Appariement des connexons :
- si canaux homotypiques : passage des m dans les 2 sens
- si canaux hétérotypiques : passage des m plus facilement dans 1 sens
∎ Connexine :
- protéines à 4 domaines transmembranaires hydrophobes (= tétraspanines)
- coudes et hélices intra- et extra-cellulaires hydrophiles
Fonctions - passage direct de petites molécules (PM < 1000 Da) d’une cellule à l’autre
Régulation de la
perméabilité
∎ Par le pH : quand élevé -> canal ouvert
∎ Par le Ca2+
: quand [Ca2+
] bas -> canal ouvert
1) cellule lésée -> MP perméable
2) entrée de Na+ et Ca
2+ dans la cellule
3) fermeture immédiate des canaux de nexus
4) cellule isolée empêchant la lésion de s’étendre
∎ Par l’AMPc : quand [AMPc] bas -> canal ouvert
1) fort éclairement des neurones de la rétine
2) augmentation de la production de dopamine
3) liaison de la dopamine sur son récepteur
4) activation d’une adénylate cyclase : [AMPc] ↗
5) fermeture des jonctions Gap des neurones rétiniens
6) activation des c du cône et inhibition des c en bâtonnets
7) adaptation de la rétine à la lumière
Mise en évidence ∎ De la fonction :
- par injection de molécules fluorescentes de taille croissante
Contacts non
jonctionnels
Caractéristiques - type homophile : cadhérines et Ig CAM
- type hétérophile : intégrines et sélectines
- Ca2+
dépendant : cadhérines et sélectines
- Ca2+
non dépendant : Ig CAM et intégrines
Fonctions - initier les adhérences intercellulaires spécifiques stabilisées par la suite par
des complexes jonctionnels
Les sélectines Interaction c inflammatoires / paroi vasculaire
∎ Définition : - molécules d’adhérences dépendantes Ca
2+
- glycoprotéines transmembranaires de la famille des lectines
- lectines = protéines se liant aux sucres des glycoprotéines et glycolipides
localisés à la surface d’autres cellules (au niveau du glycocalix)
∎ Fonctions :
- inflammation et migration des leucocytes
- interactions adhésives entre leucocytes / endothélium et plaquettes
- inhibition de contact
∎ Diapédèse :
1) lésion = siège d’une inflammation bactérienne localisée
2) accolement et roulement du leucocyte sur la paroi vasculaire grâce à une
adhérence faible avec la c endothéliale via la L-sélectine (margination)
3) sécrétion de cytokines par la c endothéliale permettant la formation de
jonctions d’ancrage entre le leucocyte et la paroi vasculaire (immobilisation)
4) dissociation locale des jonctions permettant au leucocyte de passer entre
deux c endothéliales (extravasation)
5) leucocyte attiré vers le foyer infecté (chimiotactisme)
6) phagocytose de la bactérie par le leucocyte
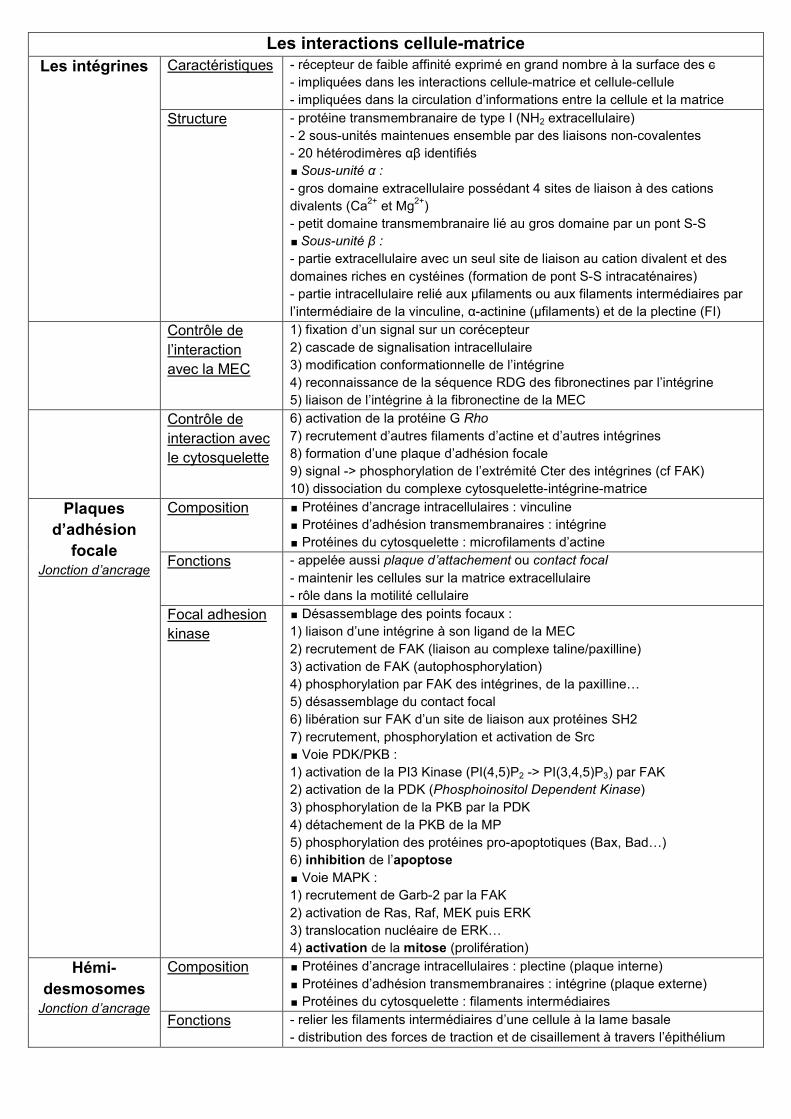
Les interactions cellule-matrice
Les intégrines Caractéristiques - récepteur de faible affinité exprimé en grand nombre à la surface des c
- impliquées dans les interactions cellule-matrice et cellule-cellule
- impliquées dans la circulation d’informations entre la cellule et la matrice
Structure - protéine transmembranaire de type I (NH2 extracellulaire)
- 2 sous-unités maintenues ensemble par des liaisons non-covalentes
- 20 hétérodimères αβ identifiés
∎ Sous-unité α :
- gros domaine extracellulaire possédant 4 sites de liaison à des cations
divalents (Ca2+
et Mg2+
)
- petit domaine transmembranaire lié au gros domaine par un pont S-S
∎ Sous-unité β :
- partie extracellulaire avec un seul site de liaison au cation divalent et des
domaines riches en cystéines (formation de pont S-S intracaténaires)
- partie intracellulaire relié aux μfilaments ou aux filaments intermédiaires par
l’intermédiaire de la vinculine, α-actinine (μfilaments) et de la plectine (FI)
Contrôle de
l’interaction
avec la MEC
1) fixation d’un signal sur un corécepteur
2) cascade de signalisation intracellulaire
3) modification conformationnelle de l’intégrine
4) reconnaissance de la séquence RDG des fibronectines par l’intégrine
5) liaison de l’intégrine à la fibronectine de la MEC
Contrôle de
interaction avec
le cytosquelette
6) activation de la protéine G Rho
7) recrutement d’autres filaments d’actine et d’autres intégrines
8) formation d’une plaque d’adhésion focale
9) signal -> phosphorylation de l’extrémité Cter des intégrines (cf FAK)
10) dissociation du complexe cytosquelette-intégrine-matrice
Plaques
d’adhésion
focale Jonction d’ancrage
Composition ∎ Protéines d’ancrage intracellulaires : vinculine
∎ Protéines d’adhésion transmembranaires : intégrine
∎ Protéines du cytosquelette : microfilaments d’actine
Fonctions - appelée aussi plaque d’attachement ou contact focal
- maintenir les cellules sur la matrice extracellulaire
- rôle dans la motilité cellulaire
Focal adhesion
kinase
∎ Désassemblage des points focaux :
1) liaison d’une intégrine à son ligand de la MEC
2) recrutement de FAK (liaison au complexe taline/paxilline)
3) activation de FAK (autophosphorylation)
4) phosphorylation par FAK des intégrines, de la paxilline…
5) désassemblage du contact focal
6) libération sur FAK d’un site de liaison aux protéines SH2
7) recrutement, phosphorylation et activation de Src
∎ Voie PDK/PKB :
1) activation de la PI3 Kinase (PI(4,5)P2 -> PI(3,4,5)P3) par FAK
2) activation de la PDK (Phosphoinositol Dependent Kinase)
3) phosphorylation de la PKB par la PDK
4) détachement de la PKB de la MP
5) phosphorylation des protéines pro-apoptotiques (Bax, Bad…)
6) inhibition de l’apoptose
∎ Voie MAPK :
1) recrutement de Garb-2 par la FAK
2) activation de Ras, Raf, MEK puis ERK
3) translocation nucléaire de ERK…
4) activation de la mitose (prolifération)
Hémi-
desmosomes Jonction d’ancrage
Composition ∎ Protéines d’ancrage intracellulaires : plectine (plaque interne)
∎ Protéines d’adhésion transmembranaires : intégrine (plaque externe)
∎ Protéines du cytosquelette : filaments intermédiaires
Fonctions - relier les filaments intermédiaires d’une cellule à la lame basale
- distribution des forces de traction et de cisaillement à travers l’épithélium

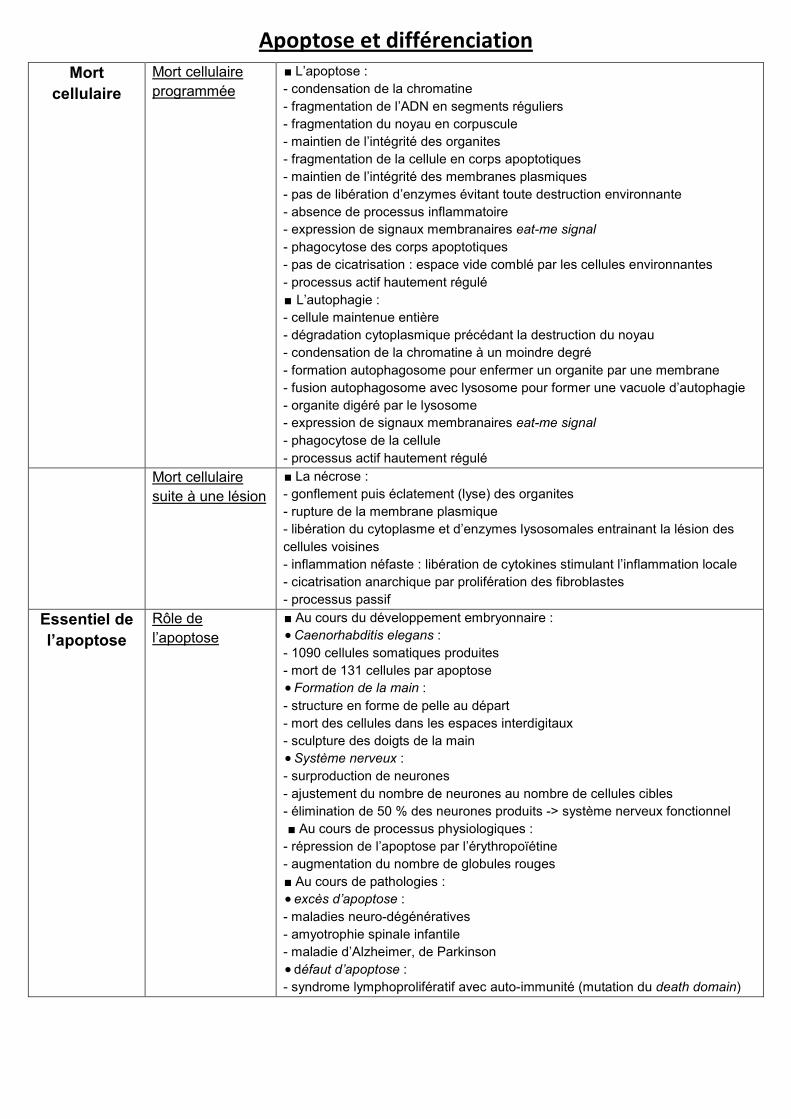
Apoptose et différenciation
Mort
cellulaire
Mort cellulaire
programmée
∎ L’apoptose :
- condensation de la chromatine
- fragmentation de l’ADN en segments réguliers
- fragmentation du noyau en corpuscule
- maintien de l’intégrité des organites
- fragmentation de la cellule en corps apoptotiques
- maintien de l’intégrité des membranes plasmiques
- pas de libération d’enzymes évitant toute destruction environnante
- absence de processus inflammatoire
- expression de signaux membranaires eat-me signal
- phagocytose des corps apoptotiques
- pas de cicatrisation : espace vide comblé par les cellules environnantes
- processus actif hautement régulé
∎ L’autophagie :
- cellule maintenue entière
- dégradation cytoplasmique précédant la destruction du noyau
- condensation de la chromatine à un moindre degré
- formation autophagosome pour enfermer un organite par une membrane
- fusion autophagosome avec lysosome pour former une vacuole d’autophagie
- organite digéré par le lysosome
- expression de signaux membranaires eat-me signal
- phagocytose de la cellule
- processus actif hautement régulé
Mort cellulaire
suite à une lésion
∎ La nécrose :
- gonflement puis éclatement (lyse) des organites
- rupture de la membrane plasmique
- libération du cytoplasme et d’enzymes lysosomales entrainant la lésion des
cellules voisines
- inflammation néfaste : libération de cytokines stimulant l’inflammation locale
- cicatrisation anarchique par prolifération des fibroblastes
- processus passif
Essentiel de
l’apoptose
Rôle de
l’apoptose
∎ Au cours du développement embryonnaire :
Caenorhabditis elegans :
- 1090 cellules somatiques produites
- mort de 131 cellules par apoptose
Formation de la main :
- structure en forme de pelle au départ
- mort des cellules dans les espaces interdigitaux
- sculpture des doigts de la main
Système nerveux :
- surproduction de neurones
- ajustement du nombre de neurones au nombre de cellules cibles
- élimination de 50 % des neurones produits -> système nerveux fonctionnel
∎ Au cours de processus physiologiques :
- répression de l’apoptose par l’érythropoïétine
- augmentation du nombre de globules rouges
∎ Au cours de pathologies :
excès d’apoptose :
- maladies neuro-dégénératives
- amyotrophie spinale infantile
- maladie d’Alzheimer, de Parkinson
défaut d’apoptose :
- syndrome lymphoprolifératif avec auto-immunité (mutation du death domain)
Voie des
récepteurs
de mort
Machinerie
intracellulaire
∎ Mécanisme (irréversible) :
1) signal inducteur apoptotique
2) auto-activation des procaspases initiatrices en caspases initiatrices
-> coupure au niveau du site aspartique
3) activation d’autres procaspases effectrices par les caspases initiatrices
-> cascade protéique amplificatrice
4) protéolyse des protéines clefs de la cellule par les caspases activées
∎ Protéines cibles de caspases :
- les lamines nucléaires -> désassemblage irréversible de la lamina nucléaire
- enzyme dégradant l’ADNase -> coupure en fragments de l’ADN nucléaire
Le récepteur Fas
(récepteur de
mort cellulaire)
∎ Caractéristiques :
- récepteur de la famille des TNF-R (= tumor necrosis factor receptor)
- protéine transmembranaire de type I (NH2 extracellulaire)
- trimérisation du récepteur après liaison du ligand Fas-L (Fas-Ligand)
∎ Structure :
partie extracellulaire :
- 3 domaines riches en cystéines impliqués dans la liaison du ligand
- CRD = cystein rich domain
partie intracellulaire :
- 1 domaine de mort de 80 acides aminés (DD = death domain)
∎ Protéine adaptatrice de Fas = FADD
- FADD = Fas associated death domain
- 1 domaine de mort (DD)
- 2 domaines effecteurs de la mort (DED = death effector domain)
Le complexe de
signalisation
induisant la mort
∎ Définition :
- DISC = death inducing signaling complex
∎ Mise en place :
1) ligand de Fas (Fas-L) présenté à la surface d’un lymphocyte tueur (killer)
2) liaison du Fas-L sur le récepteur Fas-R (domaine extracellulaire)
3) trimérisation du récepteur Fas-R
4) association homophile de FADD et de Fas-R par leur domaine de mort (DD)
5) association homophile de procaspases 8 et de FADD par leur domaines DED
6) agrégation de procaspases 8
Les caspases ∎ Famille des caspases (15 caspases différentes) :
- protéases particulières au cœur de la machinerie moléculaire de l’apoptose
- coupure des protéines cibles sur des acides aspartiques
- une cystéine sur leur site actif
∎ La caspase 8 :
- 2 domaines effecteurs de la mort en N-term (DED)
- une grande sous-unité
- une petite sous-unité en C-term
- cFLIP = cellular FLICE (FADD like interleukin converting enzyme) inhibitory protein
- cFLIP = caspase 8 sans site protéasique actif (inhibition de l’apoptose)
∎ La caspase 3 :
- une grande sous-unité en N-term
- une petite sous-unité en C-term
∎ La caspases 9 :
- un pro-domaine CARD en N-term (caspase-associated recruitment domain)
- une grande sous-unité
- une petite sous-unité en C-term
Cascade des
caspases
∎ Mécanisme :
1) agrégation des procaspases 8 dans le complexe de signalisation DISC
2) auto-activation des procaspases par :
-> faible activité kinasique fortement augmentée par activation mutuelle
-> changement de conformation activant les procaspases
3) clivage au niveau de 2 acides aspartiques
4) séparation des 2 sous-unités et libération de la chaîne avec les 2 DED
5) clivage au niveau d’un acide aspartique de la caspase 3 (par la caspase 8)
6) association des 2 petites sous-unités ensemble liées chacune à une grande
sous-unité pour former un tétramère actif
Clivages des
substrats
∎ Substrats des caspases :
- caspases elles-mêmes
- inhibiteurs d’endonucléases (ICAD = inhibitor of caspase-activated DNAase)
- protéines de régulation de l’apoptose (Bid, Bcl-2)
- protéines du cytosquelette cytoplasmique/nucléaire (gelsoline, kératine, LMN)
- protéines impliquées dans la régulation du cycle cellulaire (P21, Rb)
- enzymes de réparation de l’ADN
∎ Conséquences des clivages des substrats :
activation de l’apoptose :
- procaspases activées en caspases
- inactivation de l’inhibiteur de la caspase activatrice de l’ADNase (ICAD)
-> fragmentation de l’ADN par coupure entre les nucléosomes
- Bid activé par la caspase 8 -> déclenchement de la voie mitochondriale
inactivation de protéines de structures :
- lamines -> désassemblage irréversible de la lamina
- gelsoline/actine -> dissociation du cytosquelette à la membrane
inactivation de protéines du cycle cellulaire :
- Poly ADN ribose polymerase (PARP) inactivé
-> diminution de la synthèse de poly-ADP-ribose
- protéine Rb inactivée -> pas de passage en phase S
Contrôle de la
voie des death
receptors
∎ Compétition:
- ligand Fas-L capté par un récepteur soluble
∎ Séquestration :
- dimérisation de la caspase 8 avec l’usurpine
∎ Infection virale :
- activation des caspases par les protéines virales
Voie mito-
chondriale
Découvertes des
gènes impliqués
∎ Caenorhabditis elegans :
- petit ver transparent de 1 mm de long vivant dans le sable
- vie brève de 2 semaines
- multiplication rapide -> nombreux descendants (300)
- génome beaucoup plus petit que chez l’homme
- cellule œuf de 1090 cellules -> mort de 131 c par apoptose (au cours du dvt)
∎ Mutations des cell-elegans death gene (ced) :
- ced-3 ou ced-4 : pas de mort cellulaire et survie des 131 cellules
- ced-9 : mort de toutes les cellules (1090)
- ced-3, ced-4, ced-9 : survie de toutes les cellules (1090)
∎ Rôle des gènes ced :
- ced-3 et ced-4 : inducteurs de l’apoptose
- ced-9 : inhibiteur de l’apoptose en inhibant ced-3 et ced-4
Gènes impliqués
chez l’homme
∎ Gènes pro-apoptotiques:
- ced 3 = caspase 9
- ced 4 = Apaf-1
- egl-1 = Bid
∎ Gène anti-apoptotique :
- ced 9 = Bcl-2
∎ Vocabulaire :
- ced = caenorhabditis elegans cell-death
- Apaf = apoptosis protease activating factor
- Bcl-2 = B cell lymphoma 2 (identifié dans les lymphomes folliculaires)
- egl-1 = egg laying defective 1 (ponte d’oeuf défectueuse)
La protéine Apaf ∎ Description :
- protéine adaptatrice de la voie mitochondriale
- un domaine CARD permettant l’association avec la caspase 9
- un domaine de liaison avec le cytochrome c
- un domaine de liaison de l’ATP (ATPase domain)
- un domaine de liaison avec Bcl-2 empêchant la liaison de la caspase 9
- plusieurs segments répétitifs
∎ Fonction :
- transmission du signal de mort entre les mitochondries et les caspases
Formation de
l’apoptosome
∎ Formation de l’apoptosome :
1) libération d’Apaf-1, de Cyt c et d’ATP de la mitochondrie
2) association d’Apaf-1 avec de l’ATP
3) hydrolyse de l’ATP par Apaf-1 -> liaison du Cyt c
4) fixation de la procaspase 9 sur Apaf-1 par son domaine CARD
5) apoptosome formé
Famille des Bcl-2 ∎ Famille de Bcl-2 :
- 1 domaine d’ancrage à la membrane mitochondriale (externe) en C-term
- 1 à 4 domaines BH (Bcl-2 homology)
∎ Protéines Bcl-2 et Bcl-xL :
- Bcl-2 = B-cell lymphoma 2
- Bcl-xL = B-cell lymphoma extra large
- protéines anti-apoptotiques
- un domaine hydrophobe d’ancrage à la membrane mitochondriale (en C-term)
- domaine BH4 permettant l’interaction avec Apaf-1 (en N-term)
- domaines BH1/2 impliqués dans la formation du canal
- domaines BH1/2/3 permettant homo/hétéro-dimérisation entre Bcl2 et Bax
-> homo-dimérisation Bcl-2/Bcl-2 = survie cellulaire
-> hétéro-dimérisation Bcl-2/Bax = neutralité
- synthèse de Bcl-2 inhibée par p53
∎ Protéine Bax :
- Bax = Bcl-2 associated X protein
- protéine pro-apoptotique
- un domaine hydrophobe d’ancrage à la membrane mitochondriale (en C-term)
- domaines BH1/2 impliqués dans la formation du canal
- domaines BH1/2/3 permettant homo/hétéro-dimérisation entre Bcl2 et Bax
-> homo-dimérisation Bax/Bax = mort cellulaire
-> hétéro-dimérisation Bcl-2/Bax = neutralité
- synthèse de Bax stimulée par p53
∎ Protéine Bid :
- protéine pro-apoptotique
- un domaine BH3 -> hétéro-dimérisation entre Bid et Bax facilitant le relargage
de cytochrome c dans le cytoplasme
- activation de Bid possible via la voie des récepteurs de mort cellulaire
Pore transitoire
de perméabilité
∎ Description :
membrane externe :
- VDAC = voltage dependant anion chanel
membrane interne :
- ANT (ATP translocase) = adenine nucleotide translocator
∎ Modèle de lyse membranaire :
1) Bax ancré dans la membrane externe mitochondriale
2) stress apoptotique
3) ouverture du pore transitoire de perméabilité induit par Bax
4) dissipation des ions entre le cytosol et la matrice
5) diminution du potentiel transmembranaire mitochondrial (ΔΨm)
6) entrée d’eau et de soluté dans la matrice
7) gonflement de la matrice et éclatement partiel de la membrane externe
8) libération aspécifique de cytochrome c dans le cytosol
∎ Modèle de méga-canal :
1) Bax ancré dans la membrane externe mitochondriale
2) stress apoptotique
3) interaction de Bax avec VDAC pour former un méga-canal
ou association de Bax avec Bid pour former un méga-pore
4) sortie du cytochrome c de l’espace intermembranaire vers le cytosol
∎ Modèle incluant le Ca2+
:
1) Bax ou Bid insérés dans la membrane du réticulum (séquestration Ca2+
)
2) stress apoptotique
3) sortie massive du Ca2+
dans le cytosol
4) ouverture du canal VDAC par le Ca2+
5) sortie du cytochrome c de l’espace intermembranaire vers le cytosol
Mécanisme voie
mitochondriale
∎ Activation exogène :
1) signal inducteur apoptotique exogène
2) ligand de Fas (Fas-L) présenté à la surface d’un lymphocyte tueur (killer)
3) liaison du Fas-L sur le récepteur Fas-R (domaine extracellulaire)
4) trimérisation du récepteur Fas-R
5) association homophile de FADD et de Fas-R par leur domaine de mort (DD)
6) association homophile de procaspases 8 et de FADD par leur domaines DED
7) agrégation de procaspases 8
8) auto-activation de la caspase 8
9) Bid activé par la caspase 8
10) formation d’un hétéro-dimère Bid/Bax facilitant le relargage de cytochrome c
∎ Activation endogène :
1) lésion de l’ADN nucléaire
2) p53 plus détruit dans le protéasome -> accumulation de p53
3) synthèse de Bax stimulée par p53
4) relargage du cytochrome c
∎ Voie mitochondriale :
1) libération Cyt c, Apaf-1, ATP et AIF de la mitochondrie
2) fragmentation de l’ADN induit par AIF (indépendamment des caspases)
3) p53 plus détruit par le protéasome -> accumulation de p53
4) synthèse de Bax stimulée par p53
5) formation d’un méga-canal ou méga-pore libérant encore plus de Cyt c…
6) liaison d’Apaf-1 avec de l’ATP
7) hydrolyse de l’ATP permettant la liaison du Cyt c avec Apaf-1
8) changement de conformation d’Apaf-1
9) agrégation d’Apaf-1 et démasquage du site de liaison CARD
10) fixation de la procaspase 9 sur Apaf-1 par son domaine CARD
11) auto-activation de la caspase 9 (restant liée à Apaf-1)
12) activation des caspases en aval (caspases effectrices)
13) clivage des cibles par les caspases
14) mort cellulaire
∎ Vocabulaire :
- AIF = apoptosis inducting factor
- CARD = caspase-associated recruitment domain
Les IAP et
Smac/DIABLO
∎ Vocabulaire :
- IAP = Inhibitor of Apoptosis Protein
- Smac = Second mitochondria-derived activator of caspases
- DIABLO = direct IPA-binding protein with low pI
∎ Protéines IAP :
- protéines se liant aux caspases empêchant leur clivage et leur activité
- inhibiteurs de la mort cellulaire programmée (apoptose)
∎ Protéines Smac/DIABLO :
- neutralisation des activités anti-apoptotiques des IAP
- répresseur des inhibiteurs de caspases
- inhibiteurs de la mort cellulaire programmée (apoptose)
Cytotoxicité
médiée par les
lymphocytes T
∎ Les perforines :
1) augmentation du Ca2+
intracellulaire du lymphocyte T
2) induction de l’exocytose des granules cytotoxiques
3) libération de perforine dans l’espace intercellulaire
4) insertion dans la membrane de la cellule cible
5) polymérisation des monomères de perforine
6) formation de pores cylindriques
∎ Les granzymes :
7) pénétration des granzymes dans la cellule cible
8) caspases clivées par les granzymes
9) cascade des caspases
10) induction de l’apoptose
Rôle central de
p53
∎ Caractéristiques :
- gène suppresseur de tumeur
- gène de p53 muté dans 1 cancer sur 2
- rôle dans le contrôle du cycle cellulaire, l’apoptose, la sénescence et
l’entretien de la stabilité génique
∎ Entrée dans le cycle cellulaire :
en condition normale
1) lésion de l’ADN et activation de DNA protein kinase (DNA-PK)
2) phosphorylation de p53 par les DNA-PK
3) dissociation du complexe p53/MDM2 (MDM2 = ubiquitine ligase)
4) p53 plus détruit par le protéasome -> accumulation dans la cellule
5) expression du gène p21 -> inhibition des complexes Cdk2-Cycline E A
6) transition G1/S inhibée car Cdk2-Cycline E inactive
si gène p53 inactivé (sur les 2 copies du génome)
1) pas de production des protéines p53 et p21
2) pas d’inhibition des complexes Cdk2-Cycline A E
3) entrée en phase S non contrôlée (même si ADN lésé)
∎ Entrée en mitose :
1) inactivation du gène p53 sur les 2 copies du génome
2) Cdc25 pas inactivé même si ADN lésé
3) déphosphorylation des Cdk4/6 par Cdc25 (sur les Thr 14 et Tyr 15)
4) Cdk4/6 actif
5) entrée en mitose non contrôlée (même si erreurs de réplication de l’ADN)
∎ Apoptose :
1) inactivation du gène p53 sur les 2 copies du génome
2) pas d’inhibition par p53 de la synthèse de Bcl-2
3) pas de stimulation par p53 de la synthèse de Bax
4) formation d’un complexe Bcl-2/Bax neutralisant
5) pas de libération de cytochrome c…
6) apoptose inhibée (survie de la cellule)
∎ Sénescence :
1) raccourcissement des télomères rendant les cellules fragiles et instables
2) perte de l’expression de p53 et du point de contrôle du cycle cellulaire
3) réplication de l’ADN en dépit d’une cassure non réparée d’un brin
4) fusion des extrémités des chromatides sœurs n’ayant pas de télomères
5) séparation des chromatides fusionnées à la mitose
6) cassure sur un nouveau site
7) une cellule fille héritant d’un chromosome avec le gène dupliqué
8) surexpression du gène en question dans cette cellule fille
9) dégâts chromosomiques massifs (perte des télomères, duplication de gènes)
10) formation de cellules avec des anomalies chromosomiques importantes
11) réactivation de l’expression de la télomérase
12) chromosomes partiellement stabilisés et survie des c avec bc de mutations
13) formation de tumeurs cancéreuses
Voie des facteurs
anti-prolifératifs
∎ Voie des facteurs anti-prolifératifs :
1) fixation du TGFβ sur un récepteur de type II
2) recrutement d’un récepteur de type I
3) trans-phosphorylation du récepteur de type I
4) formation d’un complexe tétramérique actif de récepteurs
5) libération des Smad associées au récepteur (R-Smad = Receptor activated
Smad) du complexe avec SARA (Smad Anchor for Receptor activation)
6) phosphorylation de R-Smad par le récepteur TGFβR de type I activé
7) dissociation de R-Smad du récepteur
8) hétéro-dimérisation avec une Co-Smad (Smad 4)
9) entrée de ce complexe dans le noyau
10) liaison à l’élément de réponse au TGFβ du gène cible
11) inhibition de la prolifération et activation de l’apoptose
Voie des facteurs
de survie
∎ Voie médiée par l’IGF1 :
mécanisme
1) liaison de l’IGF1 sur son récepteur
2) dimérisation (oligomérisation) du récepteur
3) autophosphorylation du récepteur sur ses tyrosines
4) PI3 kinase recrutée au niveau du récepteur via ses domaines SH2
5) activation de la sous-unité catalytique de la PI3K
6) phosphorylation du PI(4,5)P2 en PI(3,4,5)P3
7) liaison du PI(3,4,5)P3 sur le domaine PH de la PDK
8) activation de la PDK et phosphorylation de la PKB
9) liaison du PI(3,4,5)P3 sur le domaine PH de la PKB
10) détachement de la PKB de la MP
11) phosphorylation de IkB (inhibitor of nuclear factor kappa B)
12) dégradation de IkB et libération de NFkB
13) migration dans le noyau et activation des gènes de survie
rétrocontrôle de la voie PI3K/PKB
1) activation de PTEN (Phosphatase and TENsin homolog) par la protéine p53
2) déphosphorylation des lipides membranaires par PTEN
3) décrochage de Akt (PKB) de la MP
4) inactivation de la PKB et arrêt du signal
5) inhibition de la survie cellulaire induite par l’IGF
∎ Voie médiée par le proto-oncogène c-Jun :
1) liaison du TGFα sur son récepteur
2) dimérisation du récepteur
3) autophosphorylation du récepteur sur ses tyrosines
4) PI3 kinase recrutée au niveau du récepteur via ses domaines SH2
5) activation de Vav par la PI3 kinase via son domaine PH
6) activation de Rac par Vav Rac-GDP -> Rac-GTP
7) translocation du complexe PAK/Nck du cytosol à la MP
8) activation de PAK par Rac-GTP
9) activation de MEKK, MEK puis JNK
10) translocation nucléaire de JNK
11) phosphorylation et activation du facteur de transcription c-Jun
12) activation du cycle cellulaire (prolifération)
Effets de
l’apoptose
Apoptose dans le
cerveau
∎ Souris sur-exprimant Bcl-2 :
- souris viables avec un cerveau plus grand et un nerf optique plus épais
- 85 % de plus de neurones dans l’hippocampe (gyrus denté)
- 50 % de plus de neurones dans la rétine (couche ganglionaire)
- 40 % de plus de cellules de Purkinje
- 30 % de plus de neurones olivaires
∎ Conséquence d’un défaut d’apoptose = déficit d’apprentissage
- test de l’orientation spatiale = test de la piscine de Morris
- test de la coordination motrice = test du rotarod (barre en rotation)
Apoptose et
cancer
∎ Apoptose et cancer :
1) p53 peu présent dans la cellule
2) synthèse de Bcl-2 pas inhibée par p53 (même quand ADN lésé)
3) cellule cancéreuse sur-exprimant Bcl-2
4) radiothérapie et chimiothérapie induisant l’apoptose des cellules tumorales
pour un système de signalisation inopérante ou altérée !
5) utilisation de drogues stimulant la libération de Cyt c des mitochondries
Différenciation
cellulaire
Contrôle de la
différenciation
∎ Facteurs diffusibles :
- hormones (myogénine)
- facteurs de transcription (Nerve Growth Factor)
∎ Matrice extra-cellulaire :
signalisation par les intégrines
- prolifération et motilité
- différenciation et survie
∎ Contacts cellule-cellule :
signalisation par les cadhérines
Différenciation
intestinale
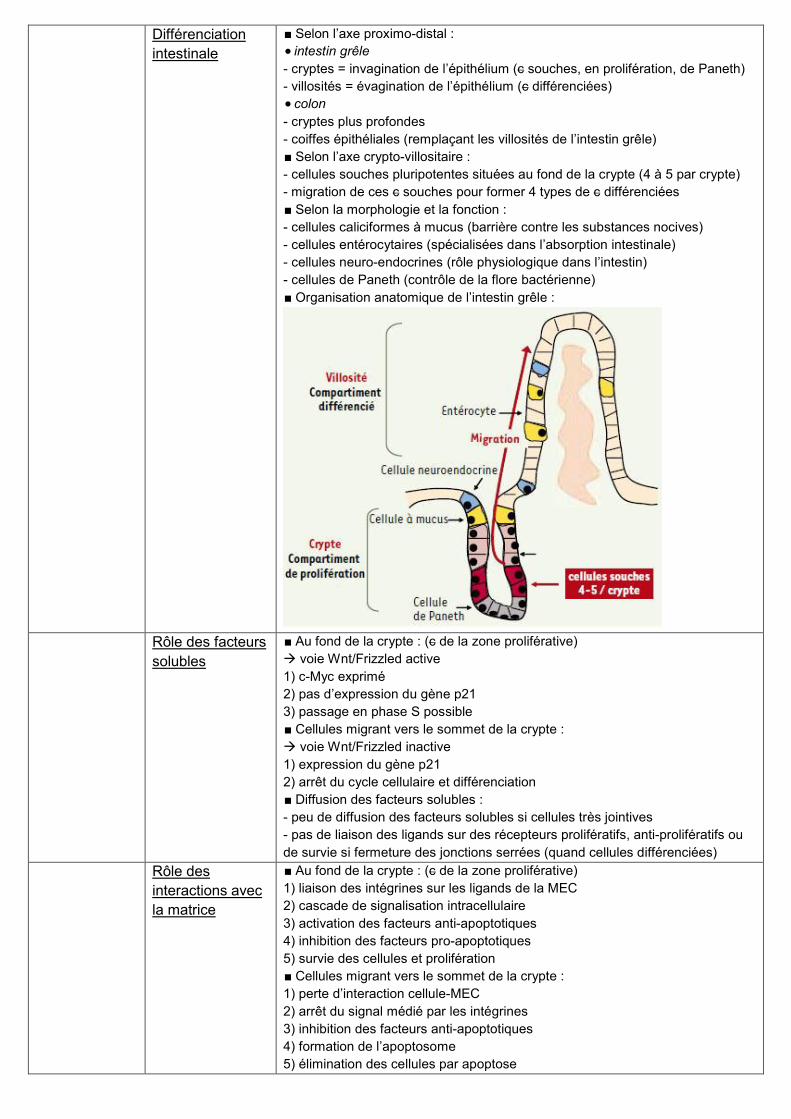
∎ Selon l’axe proximo-distal :
intestin grêle
- cryptes = invagination de l’épithélium (c souches, en prolifération, de Paneth)
- villosités = évagination de l’épithélium (c différenciées)
colon
- cryptes plus profondes
- coiffes épithéliales (remplaçant les villosités de l’intestin grêle)
∎ Selon l’axe crypto-villositaire :
- cellules souches pluripotentes situées au fond de la crypte (4 à 5 par crypte)
- migration de ces c souches pour former 4 types de c différenciées
∎ Selon la morphologie et la fonction :
- cellules caliciformes à mucus (barrière contre les substances nocives)
- cellules entérocytaires (spécialisées dans l’absorption intestinale)
- cellules neuro-endocrines (rôle physiologique dans l’intestin)
- cellules de Paneth (contrôle de la flore bactérienne)
∎ Organisation anatomique de l’intestin grêle :
Rôle des facteurs
solubles
∎ Au fond de la crypte : (c de la zone proliférative)
voie Wnt/Frizzled active
1) c-Myc exprimé
2) pas d’expression du gène p21
3) passage en phase S possible
∎ Cellules migrant vers le sommet de la crypte :
voie Wnt/Frizzled inactive
1) expression du gène p21
2) arrêt du cycle cellulaire et différenciation
∎ Diffusion des facteurs solubles :
- peu de diffusion des facteurs solubles si cellules très jointives
- pas de liaison des ligands sur des récepteurs prolifératifs, anti-prolifératifs ou
de survie si fermeture des jonctions serrées (quand cellules différenciées)
Rôle des
interactions avec
la matrice
∎ Au fond de la crypte : (c de la zone proliférative)
1) liaison des intégrines sur les ligands de la MEC
2) cascade de signalisation intracellulaire
3) activation des facteurs anti-apoptotiques
4) inhibition des facteurs pro-apoptotiques
5) survie des cellules et prolifération
∎ Cellules migrant vers le sommet de la crypte :
1) perte d’interaction cellule-MEC
2) arrêt du signal médié par les intégrines
3) inhibition des facteurs anti-apoptotiques
4) formation de l’apoptosome
5) élimination des cellules par apoptose