Embed Size (px)
Citation preview

This article was downloaded by: [Le Moyne College]On: 17 October 2014, At: 10:53Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH,UK
Acta Botanica Gallica: BotanyLettersPublication details, including instructions for authorsand subscription information:http://www.tandfonline.com/loi/tabg20
Phylogénie moléculaire dugenre Moraea (Iridaceae:Irideae): apports duséquençage d'une région d'ADNchloroplastiqueBenoît Loeuille a , Peter Goldblatt b , John C.Manning c & Vincent Savolainen da Département de Botanique, Faculté des SciencesPharmaceutiques et Biologiques , BP 83, F-59006 ,Lilleb Missouri Botanical Garden , PO. Box 299, St. Louis ,Missouri , 63166 , USAc National Botanical Institute, Kirstenbosch , PrivateBag X7, Cape Town , South Africad Molecular Svstematics Section, Jodrell Laboratory ,Royal Botanic Gardens, Kew , Richmond , TW9 3DS ,UKPublished online: 26 Apr 2013.
To cite this article: Benoît Loeuille , Peter Goldblatt , John C. Manning & VincentSavolainen (2003) Phylogénie moléculaire du genre Moraea (Iridaceae: Irideae):apports du séquençage d'une région d'ADN chloroplastique, Acta Botanica Gallica:Botany Letters, 150:3, 345-353, DOI: 10.1080/12538078.2003.10516001
To link to this article: http://dx.doi.org/10.1080/12538078.2003.10516001
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all theinformation (the “Content”) contained in the publications on our platform.However, Taylor & Francis, our agents, and our licensors make no

representations or warranties whatsoever as to the accuracy, completeness,or suitability for any purpose of the Content. Any opinions and viewsexpressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of theContent should not be relied upon and should be independently verified withprimary sources of information. Taylor and Francis shall not be liable for anylosses, actions, claims, proceedings, demands, costs, expenses, damages,and other liabilities whatsoever or howsoever caused arising directly orindirectly in connection with, in relation to or arising out of the use of theContent.
This article may be used for research, teaching, and private study purposes.Any substantial or systematic reproduction, redistribution, reselling, loan,sub-licensing, systematic supply, or distribution in any form to anyone isexpressly forbidden. Terms & Conditions of access and use can be found athttp://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
Le
Moy
ne C
olle
ge]
at 1
0:53
17
Oct
ober
201
4

Acta Bot. Gallica, 2003, 150 (3), 345-353.
Phylogenie moleculaire du genre Moraea (Iridaceae: Irideae): apports du sequen~age d'une region d' ADN chloroplastique
par Benoit Loeuille( 1 ), Peter Goldblatte), John C. Manninge) et Vincent Savolainen(4)
(/) Departement de Botanique, Faculte des Sciences Pharmaceutiques et Biologiques, BP 83, F-
59006 Lille
(2)Missouri Botanical Garden, P.O. Box 299, St. Louis, Missouri 63166, USA
(3) National Botanical institute, Kirstenbosch, Private Bag X7, Cape Town, South Africa
(4) Molecular Systematics Section, Jodrell Laboratory, Royal Botanic Gardens, Kew, Richmond TW9
3DS. UK
arrive le 15 mars 2002, accepte le 7 jevrier 2003
Resume.- L'analyse de Ia sequence de l'intron rps 16de I'ADN chloroplastique pour chacune des 66 especes du genre africain Moraea (lridaceae : lrideae), y compris des exemples des groupes majeurs d'especes du genre, apporte de nouvelles donnees pour en dresser l'arbre phylogenetique. Les resultats mantrent que Ia classification actuelle du genre reflete mal les relations phylogenetiques des especes au sein de Moraea. Ce travail confirme les resultats d'une etude phylogenetique qui avail utilise des caracteres morphologiques. Moraea est paraphyletique si Bamardie/la, Galaxia, Hexaglottis, Homeria et Gynandriris son! consideres comme des genres distincts.
Mots-cles : Moraea- lridaceae- phylogenetique- Afrique australe.
Abstract.- DNA sequence analysis of the plastid intron rps 16 from each of 66 species in the African genus Moraea (lridaceae: lrideae), including examples of all the major species clusters in the genus, brings new data for the phylogenetic tree. The results show that the current subgeneric classification poorly reflects the phylogenetic relationships of species within Moraea. This work confirms the results of a phylogenetic study using morphological characters. Moraea is paraphyletic if Bamardie/la, Galaxia, Hexaglottis, Homeria and Gynandriris are recognized as separate genera.
Key-words: Moraea -lridaceae- phylogenetic- Southern Africa.
Dow
nloa
ded
by [
Le
Moy
ne C
olle
ge]
at 1
0:53
17
Oct
ober
201
4

346
I. INTRODUCTION
La region tloristique du Cap (CFR, Cape Floristic Region) en Afrique du Sud sous l'intluence de conditions climatiques mediterraneennes, fait )'objet de nombreuses etudes scientifiques sur Ia diversite des plantes. En effet, elle possede une grande richesse en especes (9 000 especes de plantes vasculaires) ainsi que des niveaux d'endemisme et de densite Ies plus eleves au monde (67% des especes sont confinees a Ia CFR). La zone sudouest du Cap pn!sente le plus haut degre de diversite, deux fois celui de Ia zone sud-est. Cette tlore est connue pour sa diversite en succulentes et geophytes, dont le genre Moraea est un remarquable exemple. Environ 150 especes de Moraea (plus de 75% du total) se rencontrent dans cette region. Une etude phylogenetique de Moraea apportera les informations cles pour expliquer les raisons d'un tel degre de diversite dans cette zone du monde.
La circonscription du genre africain Moraea (lridaceae : lrideae), pres de deux cents especes, a longtemps ete confuse. Les premieres definitions incluaient des especes ayant un bulbe a entre-nc:eud unique, des feuilles bifaciales et des tleurs de type Iris avec des lobes du style petaloldes, chacun portant une crete proeminante. Des especes a bulbes et feuilles similaires mais a tleurs etoilees ayant des lobes du style etroits et filiformes avec des cretes minuscules ou absentes etaient incluses dans les genres Galaxia ( 15 especes), Hexaglottis (6 especes), Homeria (32 especes) et Roggeveldia (2 especes). Plusieurs especes a extension tubulaire de l'ovaire ont ete incluses dans Iris (250 especes) ou Gynandriris (9 especes), alors que Barnardiella (I espece) possede des tleurs etoilees sans cretes sur les styles et une extension tubulaire de I'ovaire. Des etudes cytologiques et des experiences de croisements (Goldblatt, 1980) ont montre que Ia distinction traditionnelle entre Moraea et Homeria n'etait plus de mise. Certaines especes a tleurs etoilees sans style petalo"ide, ni cretes partageaient un caryotype unique et une morphologie vegetative avec des especes particulieres de Mora ea. Des etudes anatomiques (Rudall & Goldblatt, 1993) de Moraea et de ses genres allies ont aussi montre que Moraea, comme il est circonscrit actuellement, etait paraphyletique. Le degre de relation entre les especes variees de Moraea et de ses genres allies reste incertain a cause des convergences morphologiques. En 1993, cependant, il etait evident que Barnardiella, Galaxia, Gynandriris, Hexaglottis, Homeria et Roggeveldia etaient combines dans Moraea. En 1998, Goldblatt transferra les especes de ces genres dans Moraea (Goldblatt, 1998).
Ce transfert porta le nombre total d'especes chez Moraea, deja un genre important avec plus de 150 especes, a 195. Une etude moleculaire fut entreprise pour fournir un support a l'hypothese que Moraea etait paraphyletique et pour determiner les relations entre Ies especes des genres precedents maintenant inclus dans Moraea. L:etude consiste en l'analyse des sequences d'ADN de quatre regions: l'exon rbcL, l'intron rps/6, l'intron trnL et l'espaceur intergenique trnL-trnF, pour 66 especes de Moraea. Les genres utilises dans les groupes externes etaient des membres de Ia tribu des lrideae. Nous presentons ici les resultats obtenus avec Ia sequence intron du gene codant Ia proteine ribosomale rps/6.
Les exons rps/6 sont separes par un intron du groupe II (Oxelman eta/., 1997). La longueur de l'intron rps/6 varie de 790 bp a 887 bp; les scores de similarite varient de 95% (entre deux Solanaceae : Nicotiana tabacum et Solanum tuberosum) a 85-86% (entre trois Poaceae: Zea mays, Hordeum vulgare et Oryza sativa) (Oxelman eta/., 1997). Ces scores indiquent que cette region peut etre utile dans les etudes phylogenetiques de famille ou de genre. La possibilite de construire des sondes universelles dans les exons tlanquants ou dans les extremites conserves 5' et 3' de l'intron, Ia possibilite de couvrir )'ensemble de l'intron avec une seule reaction d'amplification par polymerisation en chaine (PCR) et
Dow
nloa
ded
by [
Le
Moy
ne C
olle
ge]
at 1
0:53
17
Oct
ober
201
4

347
deux reactions de sequen~;age nous laissent penser que l'intron rpsl6 est un candidat ideal aux etudes phylogenetiques de Moraea (Oxelman eta/., 1997).
II. MATERIELS ET METHODES
A. Materiel vegetal, extraction d' ADN, PCR et sequen~age L'extraction d'ADN a ete realisee a partir de feuilles fraiches dessechees par du gel de
silice, Ia methode 2x CTAB a ete uti Iisee (Doyle & Doyle, 1987), I' ADN total obtenu fut purifie par un gradient de chlorure de cesium (1,55 g/ml) ou avec des colonnes de silice QIAquick™ (Qiagen, Crawley, West Sussex, UK) selon le protocole pour produits de PCR du fabricant. L'intron rpsl6 a ete amplifie par PCR (28-30 cycles, I minute de denaturation a 95 °C, 30 secondes d'hybridation a 50 °C, I minute d'extension a 72 °C, 7 minutes d'extension finale) en utilisant Master Mix™ (2,5 mM MgC12 ; AB gene Ltd., Epsom, Surrey, UK) et les sondes d'Oxelman eta/. ( 1997)
rpsF : GTGGTAGAAAGCAACGTGCGACTT et rpsR2 : TCGGGATCGAACATCAATTGCAAC.
L'extremite 3' de rpsF est localisee a II positions a l'interieur de l'intron, alors que l'extremite 3' de rpsR2 est localisee a 18 positions en aval vers l'extremite 5' de l'exon 2. La presence de fragments a ete verifiee par gel d'agarose a I %.Avant Ia reaction de sequencage, les amplifiats ont ete purifies en utilisant les colonnes QIAquick™ (Qiagen, Crawley, West Sussex, UK) ou Concert™ (Life Technologies, Paisley, Scotland, UK). La reaction de sequencage (26 cycles, 10 secondes de denaturation a 96 °C, 5 secondes d'hybridation a 50 °C, 4 minutes d'extension a 60 oq avec BigDye™ Terminators (v2.0 ; Applied Biosystems, Inc., Warrington, Cheshire, UK) a ete realisee dans un volume de 5 J..Ll de produits de PCR purifies et ensuite precipites avec de !'ethanol. Les deux brins ont ete sequences en utilisant les sondes d'amplification qui ont donne un recouvrement de 80-90%. Les echantillons remis en suspension ont passe sur un sequenceur automatique d' ADN Applied Biosystems Inc. 377 suivant le protocole du fabricant. Les sequences obtenues ont ete editees avec Sequence Navigator (Applied Biosystems, Inc., Warrington, Cheshire, UK). Pour chaque nucleotide Ia position a ete verifiee sur les deux brins.
B. Alignement et reconstruction phylogenetique Les sequences d' ADN ont ete alignees manuellement car aucun programme d'aligne
ment ne prend en consideration les dift'erentes categories de changement dans les regions non codantes d' ADN chloroplastique (regions homopolymeres, repetitions indirectes et directes, repetition unique ... ), qui ont besoin d'etre traitees differement (Kelchner, 2000). Les insertions/deletions (indels) ont ete codees comme caractere present/absent en utilisant PaupGap 1.12. Les indels ont ete considerees comme homologues quand elles partageaient exactement Ia meme postion sur Ia matrice. Les 66 sequences ont ete ajoutees a Ia matrice de Goldblatt et Manning qui en contenait deja 75, apportant ainsi le nombre total a 141 sequences.
L'analyse phylogenetique a ete realisee avec PAUP*4.0b2a qui utilise le principe de parcimonie maximum (algorithme de Wagner). Cette methode donne l'arbre le plus court (le plus parcimonieux). Le principe de parcimonie maximum signifie : minimiser Ia quantite de changements evolutifs necessaires pour expliquer les donnees disponibles et attribuer le maximum de quantite de similarite parmi les sequences a l'ancetre commun (Darlu &
Dow
nloa
ded
by [
Le
Moy
ne C
olle
ge]
at 1
0:53
17
Oct
ober
201
4

348
Tassy, 1993). PAUP peut utiliser deux principales methodes pour rechercher l'arbre optimum : une methode exacte et une methode heuristique. La methode exacte garantit Ia solution optimale mais requiert un temps de calcul important qui devient incompatible avec une matrice des donnees trop importante (nombre eleve de taxons et caracteres) ; l'algorithme heuristique permet d'obtenir un resultat en un temps de calcul raisonnable, mais Ia decouverte de l'arbre optimal n'est pas toujours garantie, le resultat etant sensible a l'ordre d'introduction des taxons terminaux. Pour pallier cette difficulte, il existe une strategie heuristique dite de ((rearrangement des branches)) (branch swapping). L'amelioration d'un arbre initial est operee par deplacement des branches. Si un rearrangement donne un arbre plus court, ce demier devient le sujet d'un nouveau rearrangement. A force de rearrangements, l'arbre minimal peut etre trouve. Pour le « rearrangement des branches », l'algorithme TBR (Tree Bisection Reconnection) a ete utilise.
Iris a ete specifie comme l'outgroup ultime et les sequences de Bobartia, Dietes, Belamcanda et Pardanthopsis ont ete incluses en plus comme groupe exteme.
III. RESULTATS
Nous avons amplifie Ia region rpsl6 de 66 especes de Moraea. L'amplification a ete couronnee de succes pour !'ensemble des especes. Les premiers resultats de Ia reaction de sequen9age ont ete mauvais mais, apres avoir reajuste le rapport quantite de sonde-quantite d' ADN, les resultats ont ete meilleurs et le signal (produit par le fluorochrome) dans EditView a ete dans Ia plupart des cas superieur a 100 (unite intrinseque au logiciel). Les deux brins ont ete obtenus pour ces 66 especes ; parfois il a ete necessaire de recommencer une seconde fois le sequen9age pour l'un des deux brins car, le signal etant trop faible il devenait hasardeux de lire l'electrophoregramme ; !'autre raison ayant amene a refaire un sequen9age a ete Ia presence d'un bruit de fond trop important qui apportait une ambigui"te sur !'interpretation de l'electrophoregramme. L'assemblage des deux brins a ete fait avec AutoAssembler; !'assemblage etait de 80 a 90% entre les deux brins, ce qui a permis d'avoir des sequences de bonne qualite a ajouter a Ia matrice.
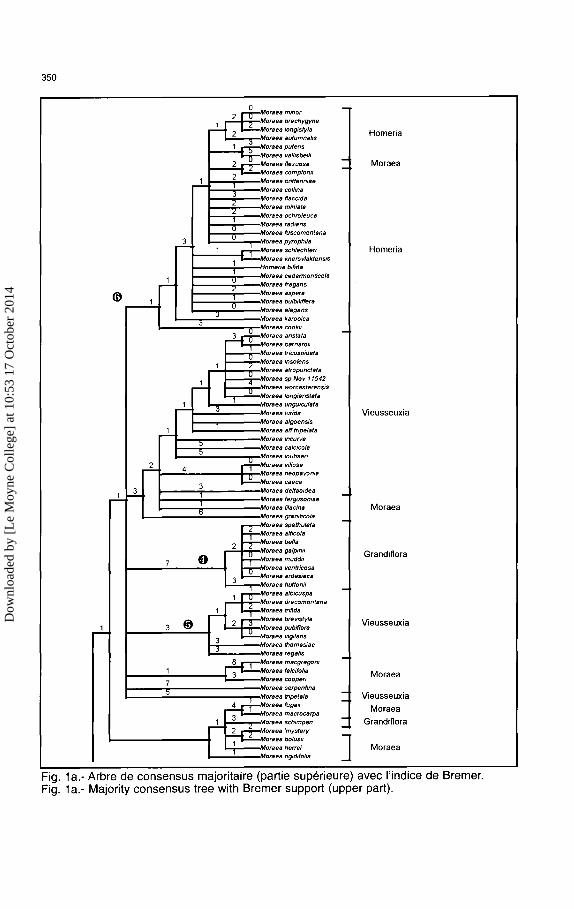
La taille de Ia matrice a impose !'utilisation d'une methode heuristique qui est une methode approximative. Apres Ia recherche heuristique, nous avons obtenu 13 000 arbres avec un score minimal (arbre le plus court) de 586 pas. L'arbre de consensus compile !'information presente dans !'ensemble de ces arbres. Un arbre de strict consensus est construit en ne retenant des arbres compares que les groupements de taxons qui sont identiques dans to us les arb res (I' in dice de consensus est 100% ), mais cet arbre est faiblement resolu et presente de nombreuses multifurcations (polytomies). L'indice de consensus peut etre modifie afin d'obtenir un arbre plus resolu (moins et/ou de plus petites multifurcations) ; ainsi nous avons abaisse cet indice a 50% et le resultat est represente par l'arbre des figures Ia et 1 b. Cet arbre est dit de consensus majoritaire (majority rule consensus tree).
Sur cet arbre de consensus majoritaire nous avons ajoute l'indice de Bremer qui reflete Ia robustesse de 1 'hypothese phylogenetique. Cet indice correspond au nombre d'etapes supplementaires avant de perdre un noeud sur l'arbre de strict consensus des arbres les plus courts (ou les plus proches de l'arbre le plus court) (Bremer, 1988). Quand tousles caracteres sont parfaitement congruents, Ia solution Ia plus parcimonieuse est unique, alors !'indice de Bremer correspond a Ia longueur des branches de l'arbre (Darlu & Tassy, 1993). La quantite d'information phylogenetique a ete evaluee par l'indice de coherence et l'indice de retention (Tableau 1).
Dow
nloa
ded
by [
Le
Moy
ne C
olle
ge]
at 1
0:53
17
Oct
ober
201
4

A partir de cet arbre nous avons pu comparer Ia classification anatomomorphologique de Goldblatt aux donnees phylogenetiques apportees par
349
Tableau 1.- Caracteristiques de Ia matrice d'ADN (intron rps16).
Table 1.- Statistics of the DNA matrix (rps16 intron).
l'intron rpsl6, ainsi que Ia pertinence de Nombre total de caracteres inclus 882 Ia reduction des genres Barnardiella, Nombre de caracteres variables 214 (24,2%) Galaxia, Gynandriris, Hexaglottis, Nombre de caracteres porteurs Homeria et Roggeveldia au sein du d'une information phylogenetique
Indice de coherence genre Moraea. Indice de retention
La longueur de l'intron rpsl6 inclus dans l'analyse est de 882 nucleotides,
Ill (12,6%) 0,54 0,76
dont 214 (24,2%) bases sont variables et Ill ( 12,6%) sont potentiellement porteurs d'une information. La longueur des arbres obtenus apn!s une recherche heuristique est de 586 pas avec un indice de coherence (CI) de 0,54 et un indice de retention (RI) de 0,76. l?arbre de consensus majoritaire est relativement faiblement resolu.
IV. DISCUSSION
A un niveau de divergence comparable a celui de Moraea, l'intron rpsl6 possede plusieurs proprietes qui le rendent interessant dans les etudes phylogenetiques des angiosperrnes incluant plusieurs sequences. II est facilement amplifiable par rapport a I' ADN ribosomal qui comprend souvent des sequences paralogues au sein d'un meme individu. l?alignement de multiples sequences est Ia plupart du temps simple et Ia variabilite est distribuee uniforrnement le long de l'intron (Oxelman eta/., 1997).
l?aspect le plus frappant de ces resultats est l'inclusion dans Moraea d'especes qui,jusqu'a recemment, etaient reportees dans Barnardiella, Galaxia, Gynandriris, Hexaglottis et Homeria. Chacun de ces genres est evidemment monophyletique ( ou pratiquement a cause du manque de resolution de Ia partie <I>(Fig. la et lb) et se retrouve dans des positions differentes au sein de l'arbre Moraea. Ces genres ont ete consideres comme differents du genre Moraea sur Ia base de leurs morphologies florales divergentes, mais its partagent avec Moraea un ensemble constant de caracteres vegetatifs, dont les bulbes a entre-mx:ud unique, Ia derivation du bulbe a partir d'un bourgeon lateral, les tuniques fibreuses du bulbe bien developpees et des feuilles bifaciales, canaliculees sans cotes.
Douze des treize especes de Galaxia forrnent un clade (0) assez divergent du reste des Moraea ; il possede un groupe frere constitue par M. angusta et M. neglecta (M. sous-genre Monocephalae). La morphologie de ce dernier groupe ne montre pas de synapomorphies evidentes avec Galaxia. On ne retrouve pas les deux principales lignees de Galaxia, certainement par manque de resolution car elles se retrouvent sur les arbres obtenus a partir de rbcL. Apparamment le clade Galaxia represente une specialisation precoce de Ia lignee Moraea, qui en derive par son port acaulescent, ses unites d'inflorescence a fleur solitaire et ses fleurs avec un tube perianthaire, des tepales de taille identique et les filets soudes en une colonne. A part le tube perianthaire, Ia fleur est de type Homeria, avec des tepales larges et de dimensions egales, une colonne staminate proeminente et les cretes du style reduites. De telles fleurs se sont developpees a maintes reprises chez Moraea, le plus notable etant dans le clade Homeria mais aussi en d'autres endroits dans le genre (Goldblatt, 1986).
Dow
nloa
ded
by [
Le
Moy
ne C
olle
ge]
at 1
0:53
17
Oct
ober
201
4
350
oraea m1nor ...,:;.4...;;.-Moraea brachygyne
oraea longJstyla
1--!'-....,.-Moraea autumns/IS
1--..!...f"o"'::oraea putens
....:~u-:::;::: ::;~:~~~ 1--....;..-Moraea tncusp1data
.-!..4--r-Moraea atropunctata Moraea sp Nov 11542
1--...;;--k oraea worcesterensts Moraea fongiBflstata
1-..-~--...11 oraea ungU/culata
L--====:t=====:z:oraea 11/acma oraea gramhcola
0
oraea spathulata oraea altlcofa oraea bella
Moraea V1g1fans oraea thomaSJae
oraea regs/is oraea macgregom oraea falclfol1a oraea coopen oraea serpentma oraea tr1petala oraea fugax oraea macrocarpa oraea schtmpen oraea 'mystery' oraea bolusu oraea herre1
oraea ngtdlfolla
Homeria
Morae a
Homeria
Vieusseuxia
Moraea
Grandiflora
Vieusseuxia
Moraea
Vieusseuxia
Morae a
Grandiftora
J Morae a
Fig. 1 a.- Arbre de consensus majoritaire (partie superieure) avec l'indice de Bremer. Fig. 1 a.- Majority consensus tree with Bremer support (upper part).
Dow
nloa
ded
by [
Le
Moy
ne C
olle
ge]
at 1
0:53
17
Oct
ober
201
4
Moraea tulbaghens1s
Moraea hesperantha
0 oraea afbtffora
Moraea namaquamontana
1etes
obartla 15 elamcanda
12 ardanthops1s
351
Morae a
Galaxia Home ria
Grandiflora Vieusseuxia
Moraea Grandi flora
Moraea
Gynandriris
Vieusseuxia Gynandriris
Moraea
Hexaglottis
Moraea
Galaxia
Monocephalae
Moraea
Visciramosa
Moraea
Visciramosa
Outgroup
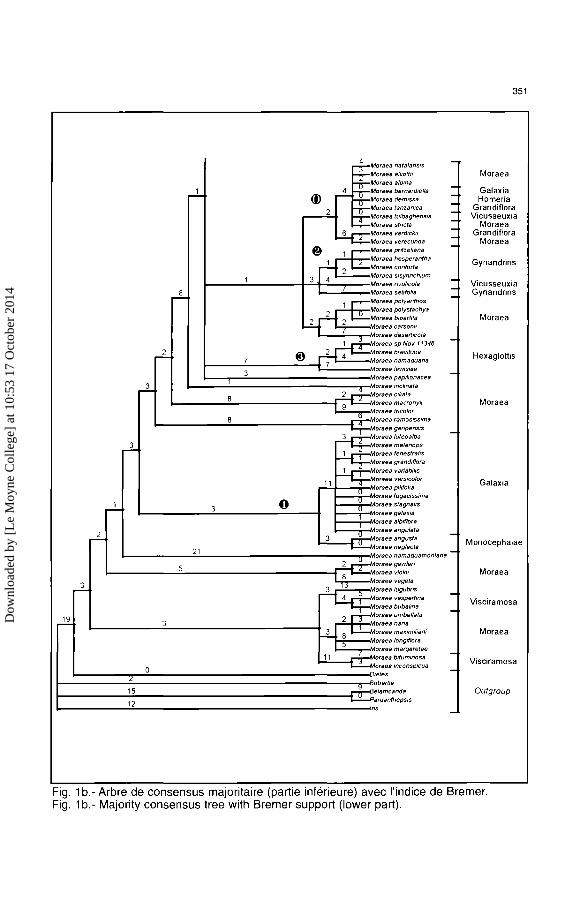
Fig. 1 b.- Arbre de consensus majoritaire (partie inferieure) avec l'indice de Bremer. Fig. 1 b.- Majority consensus tree with Bremer support (lower part).
Dow
nloa
ded
by [
Le
Moy
ne C
olle
ge]
at 1
0:53
17
Oct
ober
201
4

352
Le clade Gynandriris (8) regroupe les cinq especes du genre qui ont ete incluses dans I' etude (sur les neuf que compte le genre). On remarquera que les quatre especes venant de Ia Mediterranee (M. sisyrinchium) et de Ia winter-rainfall area d'Afrique du Sud (M. pritzeliana, M. hesperantha et M. contorta) sont regroupees ensemble, avec pour espece sreur M. setifolia. L'ovaire tubulaire en forme de bee, qui etait considere comme une supposee synapomorphie pour Gynandriris et Barnardiella, a clairement evolue de maniere independante dans ces deux lignees (Goldblatt, 1980). II est curieux que Moraea rivulico/a soit un groupe frere de Gynandriris, cette espece du sous-genre Vieusseuxia ayant une morphologie qui lui assure une place sans ambigu"ite dans ce sous-genre, qui est caracterisee par des fleurs a perianthe marcescent, a tepales internes tricuspides, et une feuille solitaire. Les membres du groupe Gynandriris ont habituellement deux feuilles (ou potentiellement deux feuilles), un perianthe fugace (deux caracteristiques plesiomorphes mais considerees ici comme des reversions), des spathes membraneuses et un ovaire sessile avec un bee tubulaire sterile. Cette position de M. rivulicola se retrouve egalement sur I'arbre obtenu par Ie sequencage de rbcL, done une erreur humaine ne peut expliquer ces resultats; une captured' ADN plastidique apparait comme possible, car aucun hybride nature) n'est connu entre ces taxons.
Le clade Hexaglottis ($) apparait ici eloigne du clade Homeria, ce qui est en desaccord avec !'hypothese de Goldblatt qui suggerait une possible relation entre les deux clades sur Ia base d'un caryotype similaire (x = 6). Les fleurs d'Hexaglottis ont des tepales etendus sub-egaux comme Homeria, mais ne possedent pas de colonne staminale. A Ia place, les filets sont courts et fusionnes ensemble sur Ia moitie de leur longueur ; le style est divise en six bras minces qui sortent horizontalement entre Ies bases des antheres.
Ces donnees moleculaires montrent de maniere convaincante Ia paraphylie de Moraea comme circonscrite traditionnellement. Barnardiella, Galaxia, Gynandriris, Homeria et Hexag/ottis ne devraient pas etre consideres comme des genres distincts. En se basant sur des caracteres morphologiques, Goldblatt transfera en 1998 toutes ces especes dans Moraea, un mouvement corrobore par )'analyse presentee ici. En plus de Ia paraphylie de Moraea et des grandes relations de Barnardiel/a, Galaxia, Gynandriris, Homeria et Hexaglottis, cette etude apporte egalement une avancee dans Ia comprehension de Ia phylogenie de Moraea. Le sous-genre Grandiflora {0) est monophyletique (si on exclut Ia position, certainement erronee, de M. verdickii). Le sous-genre Vieusseuxia forme deux clades separes. Le second clade (8) regroupe toutes les especes de Ia partie orientale de I' Afrique australe, alors que Ie premier clade regroupe les especes de Ia winter rainfall area, ce qui constitue un signe clair de I' origine unique des membres du sous-genre Vieusseuxia extemes a Ia region du Cap. Homeria {<D) est monophyletique et semble etre proche des sous-genres Vieusseuxia et Grandiflora.
V. CONCLUSION
Situee sous un climat tempere, Ia region du Cap en Afrique du Sud est remarquable par sa flore qui est tres differente de celle de I' Afrique tropicale et possede de tres nombreux groupes endemiques et, pour d'autres taxons, une tres riche diversification. Les raisons de cet impressionnant niveau de diversite ne sont pas encore totalement comprises et les etudes phylogenetiques en cours vont apporter des elements de reponse.
Dow
nloa
ded
by [
Le
Moy
ne C
olle
ge]
at 1
0:53
17
Oct
ober
201
4

353
Les resultats obtenus avec I' intron rps 16 sont en concordance avec ceux obtenus par I' analyse de rbcL, meme si certaines regions de l'arbre de consensus majoritaire manquent un peu de resolution (Goldblatt et al., 2002). lis permettent d'affirmer Ia coherence de Ia reduction des genres Barnardiella, Galaxia, Gynandriris, Homeria et Hexaglottis au sein de Moraea qui, sans I' integration de ces genres, apparaitrait paraphyletique. La classification anatomo-morphologique de Goldblatt est confirmee pour le sous-genre Grandiflora, alors que le so us-genre Vieusseuxia est scinde en deux clades distincts selon I' origine geographique des especes. L'analyse phylogenetique souligne le caractere artificiel du sousgenre Moraea qui est eparpille le long de l'arbre etjette les premieres bases d'une analyse plus complete incluant le sequem;:age de quatre loci pour )'ensemble des especes du genre.
Remerciements - Nous remercions Mark W. Chase (Royal Botanic Gardens. Kew. UK) pour l'accueil dans son laboratoire de Benoit Loeuille. Nos remerciements vont, bien sur, a tout le personnel de ce laboratoire. Martyn Powell. Jim Clarkson aussi qu'a Francisco J. Morrel Soler. Une subvention ayant couvert une partie des frais a ete fournie par l'universite de Lille II. Nous exprimons notre sincere gratitude a Bruno de Foucault (departement de botanique, faculte des sciences pharmaceutiques et biologiques de Lille) pour sa lecture attentive du manuscrit et pour ses nombreux conseils.
BIBLIOGRAPHIE
Bremer K., 1988.- The limits of amino acid sequence data in Angiosperm phylogenetic reconstruction. Evolution. 42 (4). 795-803.
Darlu P. & P. Tassy, 1993.- Reconstruction phylogenetique : concepts et methodes. Masson, Paris, Collection biologie theorique, 7, 1-245.
Doyle J.J. & J.L. Doyle, 1987.-A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochemistry Bulletin, 19, 11-15.
Goldblatt P., 1980.- Systematics of Gynandriris (lridaceae), a Mediterranean- southern Africa disjunct. Botaniska Notiser, 133, 239-260.
Goldblatt P., 1986.- Convergent evolution of the 'Homeria' flower type in six new species of Moraea (lridaceae-lrideae) in Southern Africa. Annals of the Missouri Botanical Garden, 73, 102-116.
Goldblatt P., 1998.- Reduction of Barnardiella, Galaxia, Gynandriris, Hexaglottis, Homeria, and Roggeveldia in Moraea (lridaceae: lrideae). Novon, 8, 371-377.
Goldblatt P., V. Savolainen, 0. Porteous, I. Sostaric, M.
Powell, G. Reeves, J.C. Manning, T.G. Barraclough &
M.W. Chase, 2002.- Radiation in the Cape flora and
the phylogeny of peacock irises Moraea (lridaceae) based on four plastid DNA regions. Malec. Phylog. Eva/., 25 (2), 241-360.
Kelchner S.A., 2000.- The evolution of non coding chlo
roplast DNA and its application in plant systematics.
Annals of the Missouri Botanical Garden, 87, 482-
498.
Oxelman B., M. Liden & D. Berglund, 1997.- Chloroplast
rps16 intron phylogeny of the tribe Sileneae
(Caryophyllaceae). Plant Systematics and Evolution, 203, 393-410.
Rudall P.J. & P. Goldblatt, 1993.- Leaf anatomy and sys
tematics of Homeriinae (lridaceae). Botanical Journal of the Linnean Society, 111, 379-397.
Dow
nloa
ded
by [
Le
Moy
ne C
olle
ge]
at 1
0:53
17
Oct
ober
201
4





























