Embed Size (px)
Citation preview

A
P
C
MS
i
HAD2
MCPVRPF
KCPVCPF
1h
Revue du rhumatisme monographies 79 (2012) 203–209
Disponible en ligne sur
www.sciencedirect.com
rticle original
hysiologie du calcium et des phosphates
alcium and phosphate physiology
arion Vallet , Ivan Tack ∗
ervice des explorations fonctionnelles physiologiques, laboratoire de physiologie, hôpital de Rangueil, CHU de Toulouse, 1, avenue J.-Poulhes, 31059 Toulouse cedex, France
n f o a r t i c l e
istorique de l’article :ccepté le 16 juillet 2012isponible sur Internet le 14 septembre012
ots clés :alciumhosphatesitamine Décepteur sensible au calciumTHGF-23
r é s u m é
L’homéostasie phosphocalcique est soumise à trois challenges. Le premier consiste à accumuler et main-tenir un important stock osseux de phosphates de calcium qui leur confère leur résistance mécanique. Cetobjectif, indépendant des apports alimentaires, repose sur : (1) l’augmentation de l’absorption intesti-nale phosphocalcique par la vitamine D (calcitriol) ; (2) le verrouillage de la calciurie par la parathormone(PTH) ; (3) l’effet coordonné de ces deux hormones qui stimulent le turn-over osseux avec, en conditionsphysiologiques, un effet anabolisant et d’accrétion prédominant. Le second challenge consiste à main-tenir constants le calcium extracellulaire et le stock cellulaire de phosphates. La calcémie ionisée estun déterminant vital de l’excitabilité neuromusculaire et de la coagulation. Sa stabilité repose sur ladétection de variations minimes par le récepteur sensible au calcium (CaSR) qui module la sécrétion dePTH et l’excrétion rénale du calcium. La PTH agit également sur la mobilisation osseuse du calcium etla synthèse de calcitriol. Le maintien du stock cellulaire de phosphates est crucial pour les phénomènesde phosphorylation et la synthèse d’ATP. Le calcitriol rend le bilan des phosphates positif. Inversement,l’excès de phosphates est prévenu par l’action phosphaturiante de la PTH et du fibroblast growth factor-23 (FGF-23), ce dernier inhibant aussi la synthèse du calcitriol. Le dernier challenge homéostatique est depermettre l’accrétion osseuse tout en prévenant la formation de dépôts phosphocalciques dans les tissusmous. Cette capacité, rattachée à la sénescence et à la longévité, repose sur le FGF-23 et son interactionavec le co-récepteur Khloto.
© 2012 Publié par Elsevier Masson SAS. pour la Société française de rhumatologie.
eywords:alciumhosphatesitamin Dalcium-sensing receptorTHGF-23
a b s t r a c t
Calcium and phosphate homeostasis is subjected to three challenges. The first one consists in accu-mulating and maintaining an important stock of calcium and phosphates which confers bones theirmechanical resistance. Independently of nutritional constraints, this objective is achieved by: (1) thevitamin D (calcitriol)-dependent increase of calcium and phosphates intestinal absorption; (2) the capa-bility of parathormone (PTH) to reduce calciuria; (3) the coordinated effect of both hormones to stimulatebone turnover and to exert, in physiological conditions, a predominant anabolic effect and mineral accre-tion. The second challenge consists in maintaining stable both extracellular calcium and the cellular stockof phosphates. Ionized calcium is a vital determinant of neuromuscular excitability and coagulation. A cal-cium sensing receptor (CaSR) detects even small variations of extracellular calcium and modulates bothrenal calcium excretion and PTH secretion. In turn, PTH mobilizes calcium from bones and stimulatesthe synthesis of calcitriol. The preservation of the cellular stock of phosphates is crucial for phospho-rylation and the synthesis of ATP. Global effect of calcitriol results in a positive phosphates balance,
while an excess of phosphates stimulates both PTH and Fibroblast Growth Factor-23 (FGF-23) secretion.Both hormones induce a marked phosphaturia whereas FGF-23 only inhibits the synthesis of calcitriol.The last homeostatic challenge is to produce mineral accretion in bones while simultaneously preven-ting calcium-phosphates complexes deposits in soft tissues. This capability, based on FGF-23 activityand interaction with the co-redetermine life longevity.© 2012 Published by
∗ Auteur correspondant.Adresse e-mail : [email protected] (I. Tack).
878-6227/$ – see front matter © 2012 Publié par Elsevier Masson SAS. pour la Société frttp://dx.doi.org/10.1016/j.monrhu.2012.07.006
ceptor Khloto, appears to be connected with senescence and could partly
Elsevier Masson SAS. on behalf of the Société française de rhumatologie.
ançaise de rhumatologie.

204 M. Vallet, I. Tack / Revue du rhumatisme monographies 79 (2012) 203–209
Fs
1
embdptltucbldl
2
2
22d2t[mrmddCduddn
2sem
Fig. 2. Régulation de l’homéostasie des phosphates. Les flèches vertes indiquentun effet stimulant, les flèches rouges un effet inhibiteur. RT phosphates : réabsorp-
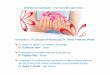
ig. 1. Régulation de l’homéostasie calcique. Les flèches vertes indiquent un effettimulant.
. Introduction
Résumer le métabolisme phosphocalcique en quelques pagesst une véritable gageure. Ses implications pathologiques sontultiples (ostéoporose, ostéomalacie, hyperparathyroïdies, dia-
ètes phosphatés, ostéodystrophie rénale. . .) et la compréhensione ses mécanismes régulateurs évolue sans cesse. Bien-sûr, lesrincipaux acteurs de ce métabolisme : calcium, phosphates, para-hormone et vitamine D, tube digestif, os et rein, sont connus depuisongtemps. Toutefois, de nouveaux éléments, tels que le récep-eur sensible au calcium ou le fibroblast growth factor-23 (FGF-23),ne hormone osseuse, s’ajoutent sans cesse, éclairant mais aussiomplexifiant notre compréhension physiopathologique. Cetterève revue a choisi de proposer un regard intégratif sur le métabo-
isme phosphocalcique et ses régulations (Fig. 1 et 2) plutôt que deétailler ses aspects classiques que l’on retrouvera aisément dans
es ouvrages et revues générales [1,2].
. Métabolisme minéral
.1. Calcium [3–9]
.1.1. Distribution et fonctions dans l’organisme
.1.1.1. Le calcium osseux. Le calcium osseux représente prèse 99,9 % du calcium total du corps humain adulte (1 kg soit5 moles). Environ 99 % de ce calcium est déposé autour de larame osseuse protéique sous forme de cristaux d’hydroxyapatiteCa10(PO4)6(OH)2] qui confèrent à l’os ses propriétés de résistance
écanique. Cette masse cristalline représente aussi une abondanteéserve, lentement mobilisable, dans les situations où la balanceétabolique du calcium demeure durablement négative. Le reste
u calcium osseux (≈ 5–10 g) est déposé sous forme de phosphatee calcium complexé et constitue le « pool calcique échangeable ».e pool est capable de compenser rapidement (quelques dizainese minutes) une variation brutale du bilan calcique, soit en stockantn excès soit en libérant du calcium en cas de déficit. La mesure de laensité minérale osseuse par Dual X-ray Absorption (DXA) permete connaître la masse calcique osseuse mais ni sur sa distributioni la qualité de sa cristallisation.
.1.1.2. Le calcium extracellulaire. Le calcium extracellulaire repré-ente seulement 0,1 % du calcium de l’organisme. Ce calciumst réparti en une phase ionisée (45 %), la seule physiologique-ent active, une phase liée aux protéines du plasma (45 %),
tion tubulaire fractionnelle des phosphates ; Pi : phosphates inorganiques ; NPT2a/c :symporteurs sodium-phosphates.
essentiellement à l’albumine, et une phase liée à de petits anions(10 %) tels que phosphates, citrates et sulfates. Le maintien à l’étatstable du calcium ionisé correspond à la fonction d’homéostasierapide du calcium, cruciale pour la survie car le calcium ionisécontribue à la régulation de l’excitabilité neuromusculaire età la fonction de coagulation. Le dosage de la calcémie ioni-sée par méthode électrochimique est le plus pertinent (valeurusuelle 1,16–1,30 mmol/L à pH 7,40). Toutefois, sa complexité pré-analytique lui fait préférer en pratique le dosage de la calcémietotale (valeur usuelle : 2,20–2,60 mmol/L). Son interprétation estdélicate dans deux situation cliniques communes :
• lorsque la protidémie varie (en particulier l’albumine et lesimmunoglobulines), la calcémie totale varie alors dans le mêmesens que la protidémie, mais sans lien avec le calcium ionisée,donc sans effet physiologique. Pour limiter cette interférence,une formule de correction de la calcémie totale en fonction del’albuminémie peut être utilisée : Ca total – 0,02 × [40–albumine]avec Ca total en mmol/L et albuminémie en g/L ;
• les variations du pH plasmatique modifient la liaison du cal-cium aux protéines. Une acidose aiguë diminue la liaison auxprotéines et augmente proportionnellement la calcémie ioniséesans modifier la calcémie totale. Inversement, l’alcalose aiguëd’hyperventilation au cours des attaques de panique (connue enFrance sous le nom de « spasmophilie ») augmente la liaison auxprotéines et entraîne une crise tonico-clonique sans variationnotable de la calcémie totale.
2.1.1.3. Le calcium intracellulaire. Le calcium intracellulaire repré-sente environ un millionième du calcium total. De fait, le fortgradient de concentration transmembranaire (1/1000) favorisel’entrée cellulaire de calcium. Le calcium intracytosolique se
comporte comme un puissant second messager. Le secteur intra-cellulaire est cliniquement inabordable et il n’existe pas deproportionnalité entre la calcémie et le calcium intracellulaire.
tisme monographies 79 (2012) 203–209 205
2
d2nd1à
•
•
hten
2
ldlmaddpismCpitc
Tableau 1Principaux aliments riches/pauvres en calcium.
Aliments riches en calciumProduits laitiers Lait : 100–120 mg/100 mL
Yaourt : 150 mg/yaourt ou/100 gfromage blancFromage pâte dure : 1000 mg/100 gFromage pâte molle : 400 mg/100 g
Fruits secs, légumesa Noisettes, amandes, figues sèches :200 mg/100 gCresson, choux, persil :150–180 mg/100 gHaricots secs : 100 mg/100 g
Eaux, boissons Talians : 600 mg/LVittel Hépar : 590 mg/LContrexéville : 510 mg/LPerrier : 140 mg/LCidre : 120 mg/LEau du robinetb : 40–120 mg/L
Aliments pauvres en calciumProtéines animales Viande < 40 mg/100 g
Poisson < 60 mg/100 g
Végétaux Blé, riz, pomme de terreépluchée < 40 mg/100 gLaitue, carottes, tomates,avocats < 40 mg/100 gBananes, raisin, agrumes < 50 mg/100 g
Eaux Volvic, Mont Roucous : 10 mg/L
a A teneur identique, les végétaux fournissent moins de calcium absorbable queles produits laitiers car leur contenu en fibres non digestibles réduit la biodisponi-bilité du calcium d’origine végétal.
b La teneur en calcium de l’eau de ville (généralement entre 1 et 5 mmol/L, soit40–200 mg/L) n’est pas réglementée en France ce qui explique sa forte variabilité
M. Vallet, I. Tack / Revue du rhuma
.1.2. Absorption intestinale et apports alimentairesL’absorption intestinale du calcium est incomplète. La plupart
es ouvrages de référence indiquent une absorption moyenne de0 % de l’apport alimentaire de calcium. . . valeur théorique quie reflète ni la réalité pratique ni la complexité du phénomène’absorption intestinale du calcium, en fait fortement variable (de0 à 50 % de l’apport). L’absorption digestive du calcium est limitée
l’intestin grêle et comporte deux mécanismes intriqués :
une absorption paracellulaire passive tout au long du grêle, peuefficiente mais non saturable ;une absorption transcellulaire active essentiellement duodé-nale mais aussi jéjunale, qui est particulièrement efficace maissaturable. Cette absorption active est dépendante du calcitriol (1-25(OH)2 vitamine D). Pour une concentration infraphysiologiquede calcitriol, l’absorption intestinale totale du calcium représenteà peine 10 %, ce qui correspond essentiellement à l’absorptionparacellulaire. Ensuite, plus la concentration de calcitriol croît,plus l’absorption intestinale du calcium augmente. Ainsi, avecune concentration physiologique maximale de calcitriol et unapport calcique faible, l’absorption intestinale nette du calciumpeut atteindre 50 %. Si l’on tient compte de la perte de calcium parles sécrétions digestives, un apport de 400 mg/j (10 mmol/j) suffitthéoriquement, chez un adulte dont la capacité de synthèse calci-triol est optimale, pour maintenir la balance calcique nulle. En fait,la large variabilité interindividuelle de l’absorption intestinalede calcium explique que la valeur recommandée soit d’environle double (800 mg/j, soit 20 mmol/j). Cela évite le risque decarence chez les moins bons absorbeurs. Lorsque l’apport calciquedépasse 800 à 1000 mg/j (20–25 mmol/j) et que la concentra-tion sanguine de calcitriol est normale, l’absorption intestinalen’augmente pratiquement plus. C’est la raison pour laquelle lessupplémentations calciques en excès souvent utilisés en rhuma-tologie (1–2 g/j) n’entraînent généralement pas d’hypercalciurie,limitant le risque de lithiase rénale et/ou de néphrocalcinose. Parailleurs, l’absorption intestinale de calcium diminue avec l’âge, cequi réduit d’autant l’éventuel impact d’une telle supplémentationchez les personnes âgées.
L’apport alimentaire de calcium varie largement en fonction desabitudes alimentaires et des régions du monde. Les produits lai-iers sont la source principale de calcium en Europe, mais certainesaux (embouteillées ou de réseau) peuvent apporter une fractionotable de la ration quotidienne de calcium (Tableau 1).
.1.3. Turn-over osseuxLe remodelage osseux joue aussi rôle permanent dans
’homéostasie minérale. Chez le sujet adulte sain, environ 500 mge calcium est mobilisé chaque jour via la solubilisation de
’hydroxyapatite dans les zones de résorption osseuse. Parallèle-ent, chez l’adulte jeune, la même quantité de calcium se dépose
u sein de la phase minérale des zones d’accrétion. La rupturee cet équilibre survient physiologiquement aux âges extrêmese la vie. En phase de croissance, le bilan minéral osseux estositif sous réserve d’un apport alimentaire suffisant et d’une
mprégnation adaptée en vitamine D. Inversement, lors de laénescence, l’établissement d’un bilan phosphocalcique légère-ent négatif contribue à la déminéralisation osseuse progressive.
e bilan négatif est déterminé par la raréfaction de la matrice
rotéique extracellulaire mais aussi par la moindre absorptionntestinale du calcium. De fait, les carences d’apport calcique reten-issent d’autant sur la minéralisation que le sujet est en phase deroissance ou de sénescence.
intercommunale. Dans les sites où l’eau potable est dure (calcaire), l’apport calciquerésultant de la boisson d’eau du robinet doit être pris en compte.
2.1.4. Excrétion rénaleLors de la filtration glomérulaire, seul le calcium ionisé ou
complexé est filtré (soit 55 à 60 % de la calcémie totale). Pour unefiltration glomérulaire normale de 180 litres par jour, la quan-tité de calcium filtrée quotidiennement représente environ 10 g(240 mmol). La totalité du calcium ionisé extracellulaire est doncfiltrée plusieurs fois par jour. La réabsorption tubulaire rénalereprésente plus de 98 % du calcium filtré et est finement régulée. Encas d’augmentation du calcium extracellulaire, le rein est capabled’excréter une importante quantité de calcium, tant que la filtrationglomérulaire est préservée.
La réabsorption au niveau tubulaire proximal représente 70 % ducalcium filtré et est passive, par voie paracellulaire, à la faveur de laréabsorption de sodium et d’eau. Plus loin, au niveau de la brancheascendante large de Henle (BALH), 20 % du calcium est réabsorbépar voie paracellulaire de fac on dépendante de celle du chlorure desodium par le cotransporteur NaK2Cl. Ce transporteur est bloquépar les diurétiques de l’anse (furosémide), ce qui induit une aug-mentation de la calciurie. Finalement, le tubule distal ne réabsorbeque 8 % de la charge filtrée, par voie transcellulaire active sous ladépendance de la parathormone (qui réduit physiologiquement lacalciurie). Les diurétiques thiazidiques (indapamide, hydrochloro-thiazide), en bloquant la réabsorption du NaCl dans ce segment,favorisent la réabsorption du calcium, diminuent la calciurie etaugmentent la calcémie.
L’excrétion urinaire quotidienne de calcium dépend, chezl’adulte sain, de l’absorption intestinale nette de calcium. La cal-ciurie est en général de l’ordre de 3 à 5 mmol/24 h. On parle
d’hypercalciurie pour des valeurs supérieures à 7,0 mmol/24 h chezl’homme et supérieures à 6,0 mmol/24 h chez la femme, soit un ratiocalcium/créatinine urinaires supérieur à 0,60 mmol/mmol sur unrecueil urinaire de 24 heures.
2 tisme monographies 79 (2012) 203–209
2
2
drldos
2cbrde
2ldldédpn
2lpe1ptf(lclrcplmépacc
2
rrCqdedcdcdpm
Tableau 2Principaux aliments riches/pauvres en phosphore.
Aliments riches en phosphoreProduits laitiers Lait : 90 mg/100 mL
Yaourt : 120–160 mg/100 gFromage (gruyère) : 500 mg/100 g
Végétaux et dérivés Poudre cacao : 40 mg/6 gPomme de terre : 120 mg/100 gPâtes : 80 mg/100 gPain blanc : 100 mg/100 g
Protéines animales Viandes (muscle) : 250 mg/100 gFoie : 400 mg/100 gPoisson : 150–250 mg/100 gŒuf : 100 mg/unité
Aliments pauvres en phosphoreBoissons Thé : 0 mg/100 mL
« Lait » de soja : 20 mg/100 mLCoca-Cola : 17 mg/100 mL
Végétaux Concombre : 15 mg/100 gRiz : 20 mg/100 gFruits < 30 mg/100 g
06 M. Vallet, I. Tack / Revue du rhuma
.2. Phosphates [10,11]
.2.1. Distribution et fonction dans l’organismeLe phosphore est un élément important mais peu abondant
e notre organisme dont il représente 1 % de la masse, soit envi-on 700 g. Il est présent sous forme complexée, principalement à’oxygène et au calcium, aussi parle-t-on de phosphates plutôt quee phosphore. Sa répartition est inhomogène avec 85,5 % dans less et les dents, 14 % dans les tissus mous, 0,4 % dans les hématies eteulement 0,1 % dans le secteur extracellulaire.
.2.1.1. Les phosphates osseux. Les phosphates osseux sont desomposants essentiels des cristaux d’hydroxyapatite. De fait, unealance négative prolongée en phosphates impacte la miné-alisation osseuse provoquant des tableaux d’ostéoporose et’ostéomalacie, parfois rapidement progressive (formes oncogènest syndromes de Fanconi).
.2.1.2. Le phosphate des tissus mous. Il est essentiellement cellu-aire mais on le retrouve aussi, complexé au calcium, au niveaues matrices extracellulaires où ses dépôts s’accroissent avec
’âge. Le phosphore est présent dans les membranes cellulaires etans le cytosol où il forme les liaisons pyrophosphates de hautenergie de l’ATP et contribue à la signalisation par phénomènee phosphorylation des protéines. Au sein du noyau, les phos-hates participent à la structuration et à la stabilisation des acidesucléiques.
.2.1.3. Les phosphates extracellulaires. Les phosphates extracel-ulaires sont présents sous forme organique, au sein deshospholipides circulants, et sous forme inorganique, la seule doséen routine. La concentration sérique des phosphates varie de 0,80 à,40 mmol/L (soit 25 à 43 mg/L, exprimé sous forme pondérale dehosphore). Les phosphates se comportent comme un systèmeampon présent sous deux formes à pH physiologique : 20 % sousorme acide, monovalente (H2PO4
−), et 80 % sous forme bivalenteHPO4
2−), accepteuse de protons si le pH diminue. Le dosage dea phosphatémie est simple et robuste à condition de prendreertaines précautions préanalytiques : patient à jeun (ni produitsaitiers, ni sucres) et prélèvement de préférence sur tube hépa-iné centrifugé rapidement afin d’éviter l’hémolyse ou le relargageellulaire de phosphates. Le rythme nycthéméral marqué des phos-hates justifie d’effectuer la recherche d’hypophosphatémie plutôt
e matin (moment du nadir). La phosphatémie varie avec l’âge :aximale chez le nourrisson, elle demeure encore 1,5 fois plus
levée chez l’adolescent que chez l’adulte. Après la puberté, la phos-hatémie décroît rapidement pour atteindre les valeurs de l’âgedulte. Durant la sénescence, elle diminue lentement et de fac onontinue chez l’homme alors qu’elle se stabilise après la ménopausehez la femme.
.2.2. Absorption intestinale et apports alimentairesÀ l’instar du calcium, l’absorption intestinale des phosphates
ésulte d’un double mécanisme : passif et prédominent lors desepas mais aussi actif via le symport sodium-phosphates NPT2b.e transport principalement duodénal et jéjunal n’est critiqueue dans les rares cas de carence d’apport et est sous la dépen-ance du calcitriol. Pour une alimentation équilibrée (Tableau 2),nviron 60 à 75 % des phosphates sont absorbés. À la différenceu calcium, ce phénomène n’est pratiquement pas saturable,e qui favorise la survenue des hyperphosphatémies en cas deéfaut d’excrétion rénale (hypoparathyroïdie, insuffisance rénale
hronique). En revanche, la présence d’un excès de calcium ou’aluminium dans la lumière intestinale réduit l’absorption deshosphates via la formation de complexes insolubles. Ce phéno-ène est utilisé pour lutter contre l’hyperphosphatémie au coursL’absorption intestinale des phosphates est réduite en présence de calcium,d’aluminium, de copolymères d’oxirane (ex. : Sevelamer®).
de l’insuffisance rénale mais peut, exceptionnellement, induire unehypophosphatémie chez les forts consommateurs d’anti-acides àbase de sels d’aluminium.
2.2.3. Excrétion rénaleEnviron 85 % des phosphates inorganiques circulants sont ultra-
filtrables. Sur environ 180 mmol de phosphates filtrés par jour, 85 à90 % sont réabsorbés de fac on active, essentiellement au niveau dutubule proximal. La réabsorption fractionnelle des phosphates (RTphosphates) est facilement déterminée à partir d’un prélèvementsanguin et urinaire : RT phosphates = [1 – (phosphaturie × créati-ninémie)/(phosphatémie × créatininurie)] × 100. L’unité (mg/L oummol/L) doit être identique pour le sang et les urines. En casd’hypophosphatémie, une RT phosphates supérieure à 85 % signeune réponse rénale d’épargne adaptée. Inversement, une valeurinférieure à 85 % signe une fuite tubulaire de phosphates. Laréabsorption tubulaire proximale repose sur l’expression api-cale de symporteurs sodium-phosphates (NPT2a et NPT2c) dontl’expression est fortement augmentée par le calcitriol et réduitepar la parathormone et le FGF-23.
3. Régulation de l’homéostasie phosphocalcique
3.1. Récepteur sensible au calcium (CaSR) [12,13]
La concentration du calcium ionisé extracellulaire est détectéedans différents tissus par un récepteur sensible au calcium (CaSR,pour « Ca + + sensing receptor »). Ce récepteur appartient à la super-famille des récepteurs à sept domaines transmembranaires couplésaux protéines G. CaSR est principalement exprimé à la membranedes cellules principales des parathyroïdes, des cellules C de la thy-roïde (où il stimule la sécrétion de calcitonine), dans le rein maisaussi dans d’autres tissus ou organes tels le tube digestif et l’os. Auniveau des parathyroïdes, CaSR permet une régulation fine et rapidede la sécrétion de PTH en réponse à des variations du calcium ioniséextracellulaire. Le calcium extracellulaire se lie au CaSR, qui estcapable de détecter des variations, même modestes (0,1 mmol/l), dela calcémie ionisée. Lorsque la calcémie diminue, la moindre liaison
du calcium avec la CaSR stimule la sécrétion de PTH et semble aug-menter sa synthèse. Inversement, l’augmentation de la liaison ducalcium avec le CaSR diminue la sécrétion de PTH et l’expression du
tisme
gp
rmslcah
3
3
appdddtu
3
qmpdqemimbltlo
3
ldpsll
3
hpcc
3
t13rh
M. Vallet, I. Tack / Revue du rhuma
ène de la prépro-PTH mais inhibe aussi la prolifération des cellulesarathyroïdiennes.
Le CaSR est aussi exprimé dans différents segments du tubuleénal où il module la perméabilité de l’épithélium au calcium et auagnésium. Ainsi, lors d’une augmentation de calcémie, la réab-
orption tubulaire de calcium diminue, ce qui favorise l’excrétion de’excédent de calcium. L’activation du CaSR réduirait aussi les capa-ités de concentration des urines en inhibant l’action de l’hormonentidiurétique et expliquant le diabète insipide rénal associé auxypercalcémies sévères.
.2. Parathormone (PTH) [13,14]
.2.1. Synthèse et sécrétion de PTHLa PTH est une hormone polypeptidique constituée de 84 acides
minés. Elle est synthétisée dans les cellules principales des glandesarathyroïdiennes sous forme d’une prépro-hormone, clivée enro-PTH au niveau du réticulum endoplasmique puis en PTH lorsu stockage dans des vésicules de sécrétion. La sécrétion de PTHépend directement de la concentration du calcium extracellulaireétectée par le CaSR. Son dosage est rendu délicat par la circula-ion de plusieurs formes dégradées (problématique détaillée dansn autre article de ce numéro).
.2.2. Principales ciblesLa PTH agit sur le rein et l’os, en se liant à son récepteur (PTH1R),
ui appartient à la famille des récepteurs à sept domaines trans-embranaires couplés aux protéines G. Lorsqu’elle est sécrétée,
ar exemple en cas d’hypocalcémie, la PTH augmente la libératione calcium osseux et stimule la réabsorption rénale du calcium, ceui tend à normaliser la calcémie. La PTH stimule aussi l’expressiont l’activité de la 1-alpha hydroxylase dans le tubule proximal, aug-entant la synthèse de calcitriol et donc indirectement l’absorption
ntestinale de calcium. Au-delà de son action sur le métabolismeinéral, la PTH stimule, via son récepteur exprimé dans les ostéo-
lastes, la synthèse protéique osseuse et, dans certaines conditions,a différenciation et l’activité ostéoclastiques contribuant ainsi for-ement à la régulation du turn-over osseux. Cette action expliquee bénéfice osseux de la PTH 1-34 humanisée dans le traitement desstéoporoses fracturaires.
.3. PTH-related protein [14–16]
L’existence d’un facteur capable de mimer l’action de la PTH sure métabolisme minéral a été évoquée il y a 60 ans dans le cadre’hypercalcémies malignes. La structure protéique de la PTH relatedrotein (PTH-rp) a depuis été identifiée et ses multiples actions phy-iologiques, majoritairement paracrines, ont été décrite au cours de’embryogenèse, de la croissance mais aussi de la grossesse et de laactation.
.3.1. Origine cellulaireLa PTH-rp est issue de la transcription d’un gène partiellement
omologue à celui de la PTH. Ses trois isoformes sont expriméesar de nombreux types cellulaires, plus particulièrement par lesellules épithéliales ce qui rend compte des principaux types deancers responsables de sa sécrétion paranéoplasique.
.3.2. Actions hormonalesLa PTH-rp est une poly-hormone dont la maturation abou-
it à trois fragments clivés. Seul le fragment N-terminal
-36 présente une homologie notable avec le fragment 1-4 de la PTH, ce qui lui confère la capacité de se lier auécepteur de la PTH (PTH1R) et de mimer l’action de cetteormone. Le syndrome paranéoplasique d’hypersécrétion demonographies 79 (2012) 203–209 207
PTHrp produit un tableau d’hyperparathyroïdie : hypercalcé-miev – hypercalciurie – hypophosphorémie – hyperphosphaturieassocié à une PTH freinée (généralement ≤ 15 pg/mL). Physiolo-giquement, la concentration circulante de PTH-rp est trop faiblepour mimer l’action de la PTH. Toutefois, à la fin de la grossesse, saforte sécrétion par les cellules épithéliales des glandes mammaires(régulée par le CaSR) permet à la PTH-rp d’atteindre une concen-tration circulante suffisante pour agir comme une hormone. LaPTH-rp permet alors la mobilisation du calcium osseux nécessaireà l’allaitement, parallèlement au verrouillage de la calciurie.
3.4. Vitamine D et calcitriol [11,17–20]
3.4.1. Production et métabolismeLa vitamine D3 (cholécalciférol) provient de la transformation,
sous l’action des rayons ultraviolets B au niveau de la peau, du 7-déhydrocholestérol en vitamine D3. La vitamine D2 (ergocalciférol)est la forme produite par les plantes. Chez l’humain, la vitamineD est en partie apportée par l’alimentation, en particulier par laconsommation de poissons gras (vitamine D3) ou de certains végé-taux comme les champignons ou les céréales (vitamine D2).
La vitamine D est ensuite hydroxylée de fac on non régulée en25-hydroxyvitamine D (25OHD ou calcifédiol) par un cytochromeP450 du foie. Sa demi-vie longue, environ deux semaines, en fait lemeilleur moyen d’évaluer le stock en vitamine D.
La 25OHD est finalement convertie dans les cellules du tubuleproximal du rein dans sa forme active, la 1,25-dihydroxyvitamineD (1,25(OH)2D ou calcitriol), par l’enzyme mitochondriale 1-alphahydroxylase. Une hydroxylation sur le carbone 24 est égalementpossible, avant ou après la 1-alpha hydroxylation, aboutissant àl’inactivation fonctionnelle de la vitamine D.
La vitamine D et ses métabolites circulent dans le sang principa-lement liés à une protéine de transport, la vitamin D-binding protein(VDBP).
3.4.2. Mode d’action cellulaireLa 1,25-dihydroxyvitamine D exerce ses actions principalement
par liaison à son récepteur nucléaire VDR (Vitamin D receptor). LeVDR est un facteur transcriptionnel qui, une fois activé, se lie à l’ADNpour moduler la transcription de ses gènes cibles. Le calcitriol régulel’expression de nombreux gènes dans le tube digestif, l’os, le reinet les parathyroïdes.
3.4.3. Régulation du métabolisme minéralComme détaillé précédemment, le calcitriol augmente
l’absorption intestinale active du calcium et du phosphore. Lecalcitriol augmente également le turnover osseux : il stimule, viales ostéoblastes et en interaction avec la PTH, la maturation etl’activation des ostéoclastes avec pour effet une résorption et lamobilisation du calcium osseux. Parallèlement, l’activation desostéoblastes par la vitamine D exerce un puissant effet anabolisantsur les protéines de l’os et favorise l’accrétion minérale osseuse.En l’absence d’hyperparathyroïdie (primitive ou secondaire) et/oud’hypocalcémie, l’effet anabolisant osseux du calcitriol l’emportece qui en fait l’hormone essentielle du trophisme osseux. Au niveaurénal, le calcitriol stimule la réabsorption des phosphates mais n’aque peu d’effet sur le calcium. Enfin, il inhibe l’expression du gènede la PTH dans les cellules parathyroïdiennes.
3.4.4. Actions « non minérales »Une activité 1-alpha hydroxylase a aussi été décrite dans la
peau, le trophoblaste, les parathyroïdes, le pancréas, la médullo-
surrénale, le côlon et le cervelet. La régulation et le rôle de cettesynthèse extrarénale sont mal connus. Des effets bénéfiques dela vitamine D ont été rapportés dans d’autres circonstances quela régulation du métabolisme minéral, par exemple au cours des
2 tisme
mpLc
3
eamgtPd3Lnàrlélc
3
1leoptràdeldiscmcrd
3
3
etlts
•
08 M. Vallet, I. Tack / Revue du rhuma
aladies cardiovasculaires, du diabète de type 2 et de certainesathologies néoplasiques mais aussi dans la réponse immunitaire.es mécanismes d’action de la vitamine D dans ce cadre sont malonnus.
.4.5. Besoins physiologiquesLes besoins et les cibles thérapeutiques de la vitamine D font
ncore actuellement l’objet de débats d’écoles. La majorité desuteurs s’accorde toutefois pour définir l’insuffisance en vita-ine D comme le seuil en dessous duquel, dans la population
énérale, la PTH augmente. Cela correspond à une concentra-ion de 25OHD inférieure à 20 ng/mL (50 nmol/L). Par ailleurs, laTH est optimalement freinée quand la vitamine D est au-dessuse 30 ng/ml (75 nmol/L). Cela conduit actuellement à proposer0 ng/mL comme seuil inférieur de sa concentration « optimale ».’intoxication à la vitamine D, responsable d’une hypercalcémie,e se voit pas pour des concentrations de vitamine D inférieures
115 ng/mL (375 nmol/L), ce qui permet de définir le seuil supé-ieur acceptable de 25OHD. Concernant les apports en vitamine D,es dernières recommandations de l’IOM (Institute Of Medicine),tablissent le besoin quotidien en vitamine D à 600 UI partir de’âge d’un an, et à 800 UI après 70 ans, l’objectif étant d’obtenir uneoncentration de 25OHD d’au moins 20 ng/mL.
.5. Fibroblast growth factor-23 [11,21–24]
Facteur tumoral phosphaturiant identifié à la fin des années990, le FGF-23 a été considéré comme une hormone clé de
’homéostasie du phosphore dix ans plus tard. Cette protéinest principalement sécrétée par l’os (ostéocytes, ostéoblastes etstéoprogéniteurs) et agit sur les transferts rénaux de phos-hates et la synthèse du calcitriol. Son action phosphaturianteubulaire proximale est synergique avec celle de la PTH. Enevanche, le FGF-23 inhibe la synthèse du calcitriol contrairement
la PTH qui la stimule. En conditions normales, la sécrétione FGF-23 est stimulée par les phosphates (après un repas, parxemple) et l’administration de calcitriol. Elle est inhibée par’hypophosphatémie, comme celle induite par la prise de chélateures phosphates. Le FGF-23 se lie avec le co-récepteur Klotho pour
nteragir avec une série de récepteurs transmembranaires. La pré-ence de ces récepteurs dans le rein ailleurs qu’au niveau du tubuleontourné proximal indique que d’autres propriétés de cette hor-one restent à découvrir. Une sécrétion excédentaire de FGF-23 au
ours des ostéomalacies oncogènes et des rachitismes vitamino-ésistants, lié à l’X ou autosomique dominant, entraîne un tableau’ostéomalacie sévère avec déficit en calcitriol.
.6. Autres hormones [25,26]
.6.1. Stéroïdes sexuelsSi le rôle critique des stéroïdes sexuels, estradiol chez la femme
t testostérone chez l’homme, sur le statut minéral osseux et leurn-over osseux est aujourd’hui bien connu, leur impact physio-ogique sur le métabolisme phosphocalcique reste imprécis. Desendances émergent toutefois pour les deux principaux stéroïdesexuels :
les estrogènes exercent un effet globalement phosphaturiant quidisparaît à la ménopause et qui est restauré par une substitu-tion en estradiol ; par ailleurs, si les estrogènes endogènes ouexogènes à dose physiologique ne modifient pas la calcémie,leur action calciotrope est démontré. Les estrogènes stimulent
l’absorption intestinale du calcium de fac on indépendante du cal-citriol mais également sa réabsorption tubulaire distale. L’effetdes estrogènes sur son excrétion urinaire est paradoxal : unecarence en estradiol (au cours de la ménopause, par exemple)monographies 79 (2012) 203–209
augmente la calciurie via une diminution de l’anabolisme osseuxet de la réabsorption rénale, toutefois, un apport pharmacolo-gique d’estrogènes majore également la calciurie en stimulantl’absorption intestinale du calcium et en augmentant le turn-overosseux ;
• les androgènes, et plus particulièrement la testostérone, exercentun effet modulateur hormonal (augmentation parallèle de la cal-citonine et de la PTH) modéré, sans impact apparent sur lesconcentrations circulantes de calcium et de phosphates.
3.6.2. Hormones somatotropes : GH et Insulin-like GrowthFactor-1
L’axe somatotrope joue un rôle clé dans la croissance et lamaturation osseuse. L’essentiel des actions osseuses, en particulierdurant la phase de croissance, est rattaché à l’IGF-1 (insulin-like growth factor-1). L’IGF-1 stimule l’absorption intestinale desphosphates et du calcium de fac on calcitriol-dépendante. Parallèle-ment, l’IGF-1 stimule nettement et de fac on durable la réabsorptiontubulaire proximale des phosphates filtrés ce qui augmente laphosphatémie, comme au cours de la puberté mais aussi des acro-mégalies. En revanche, l’IGF-1 n’exerce pas d’effet notable sur laréabsorption tubulaire du calcium. Il en résulte que le surcroîtde calcium absorbé est en partie capté, particulièrement lors del’accrétion osseuse de la croissance, mais aussi en partie éliminépar excrétion rénale, particulièrement chez l’adulte acromégale.
3.6.3. Hormones thyroïdiennesLes hormones thyroïdiennes jouent un rôle dans la crois-
sance et la maturation du squelette mais aussi sur le remodelageosseux, chez l’adulte. Il existe des récepteurs à ces hormonesdans les chondrocytes des cartilages de croissance, les ostéo-blastes et les ostéoclastes. Si une concentration physiologiqued’hormones thyroïdiennes ne modifie pas l’équilibre phosphocal-cique, les hyperthyroïdies augmentent la résorption osseuse nette,libérant du calcium et des phosphates osseux ce qui génère unehypercalciurie modeste et détermine un bilan calcique négatif.Les hyperthyroïdies sévères entraînent une hypercalcémie modé-rée à PTH freinée et une hypercalciurie abondante. Enfin, la priseen charge chirurgicale des thyrotoxicoses sévères s’accompagneparfois d’un impressionnant tableau de « hungry bone syndrome »,qui associe hypocalcémie et hypocalciurie, que l’on distingue aisé-ment d’une simple hypoparathyroïdie postchirurgicale où la PTHest effondrée et la calciurie non verrouillée.
4. Synthèse : les trois challenges du métabolismephosphocalcique
L’homéostasie phosphocalcique répond à trois contraintes :
• accumuler et maintenir un stock suffisant de phosphate de cal-cium (essentiellement sous forme stable d’hydroxyapatite) ausein du squelette osseux afin de lui conférer une résistance méca-nique optimale ;
• maintenir stable, à court terme, la concentration extracellulairede calcium ionisé mais aussi le stock cellulaire de phosphatesqui permet les réactions de phosphorylation indispensables à lasignalisation cellulaire ainsi qu’aux transferts d’énergie ;
• protéger les tissus mous, en particulier les parois vasculaires, dedépôts intempestifs de phosphate de calcium alors que les condi-tions d’une accrétion osseuse sont obtenues.
L’établissement d’un bilan positif ou nul de calcium et dephosphates repose sur un apport alimentaire suffisant et uneabsorption intestinale fonctionnelle. Si tel n’est pas le cas, le défautd’apport/d’absorption est compensé par la synthèse de calcitriol

tisme
qcddsProsndssnp1bdd
circrdommltpsdsfié
D
t
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[[
[
M. Vallet, I. Tack / Revue du rhuma
ui augmente l’absorption intestinale de ces deux ions. Si laarence concerne particulièrement le calcium, la sécrétion CaSR-épendante de PTH permet le verrouillage de l’excrétion urinaireu calcium et la synthèse d’un surcroît de calcitriol. Si ce n’est pasuffisant pour restaurer la calcémie ionisée, la sécrétion continue deTH permet la mobilisation osseuse du calcium. Cette ultime bar-ière homéostatique repose sur le renforcement électif de l’activitéstéoclastique en réponse à une augmentation prolongée de PTH etur l’induction d’une phosphaturie PTH-dépendante qui, en dimi-uant le produit phosphocalcique, permet le maintien en solutionu calcium mobilisé. En cas d’une carence en phosphates, rare car ilsont abondants dans notre alimentation, la vitamine D via son effetur l’absorption intestinale maintient le stock cellulaire. Le méca-isme régulateur est moins clairement défini, mais la détection deshosphates par le tubule proximal et le recrutement de son activité-alpha hydroxylase jouent un rôle important. Lorsque la disponi-ilité phosphocalcique est finalement suffisante, l’action concertéeu calcitriol et de la concentration physiologiquement fluctuantee PTH stimule l’anabolisme osseux et le phénomène d’accrétion.
Des apports alimentaires franchement excessifs ou certainesirconstances pathologiques s’accompagnent d’une augmentationmportante du calcium et/ou des phosphates extracellulaires. Leisque immédiat de l’hypercalcémie est lié à son impact neuromus-ulaire, en particulier cardiaque. Il est limité par l’augmentationapide et importante de la calciurie en réponse à l’activationes CaSR rénaux et parathyroïdiens. Une augmentation rapideu prolongée des phosphates extracellulaires expose à la for-ation de dépôts phosphocalciques irréversibles dans les tissusous. La stimulation de la sécrétion de PTH et de FGF-23 par
’hyperphosphatémie inhibe de fac on conjointe la réabsorptionubulaire rénale des phosphates, augmentant fortement la phos-haturie. Parallèlement, le puissant effet inhibiteur du FGF-23 sur laynthèse du calcitriol dépasse l’effet stimulant de la PTH et permete réduire l’absorption intestinale des phosphates. Cette dimen-ion homéostatique est hors du champ de cette revue mais elleait aujourd’hui l’objet d’une attention soutenue en raison de sonmpact sur le vieillissement tissulaire et, plus largement, d’un lientroit avec notre longévité [27,28].
éclaration d’intérêts
Les auteurs déclarent ne pas avoir de conflits d’intérêts en rela-ion avec cet article.
éférences
[1] Coe F, Favus M. Disorders of bone and mineral metabolism. Raven Press; 1992.
[
monographies 79 (2012) 203–209 209
[2] Peacock M. Calcium metabolism in health and disease. Clin J Am Soc Nephrol2010;5(Suppl. 1):S23–30.
[3] Landis WJ. The strength of a calcified tissue depends in part on the molecularstructure and organization of its constituent mineral crystals in their organicmatrix. Bone 1995;16(5):533–44.
[4] Parfitt AM. Letter: correction of plasma calcium measurements. Br J Hosp Med1974;1(5906):520.
[5] Malm OJ. Calcium requirement and adaptation in adult men. Scand J Clin LabInvest 1958;36(10 Supp):1–290.
[6] Hoenderop JG, Nilius B, Bindels RJ. Calcium absorption across epithelia. PhysiolRev 2005;85:373–422.
[7] Favus MJ. Factors that influence absorption and secretion of calcium in thesmall intestine and colon. Am J Physiol Cell Physiol 1985;248:G147–57.
[8] Friedman P. Calcium transport in the kidney. Curr Opin Nephrol Hypertens1999;8:589–95.
[9] Frick KK, Bushinsky DA. Molecular mechanisms of primary hypercalciuria. J AmSoc Nephrol 2003;14:1082–95.
10] Berndt T, Kumar R. Novel mechanisms in the regulation of phosphorus homeo-stasis. Am J Physiol Cell Physiol 2009;24:17–25.
11] Bergwitz C, Jüppner H. Regulation of phosphate homeostasis by PTH, vitaminD, and FGF-23. Annu Rev Med 2010;61:91–104.
12] Egbuna O, Brown E. Hypercalcaemic and hypocalcaemic conditions dueto calcium-sensing receptor mutations. Best Pract Res Clin Rheumatol2008;22(1):129–48.
13] Chen RA, Goodman WG. Role of the calcium-sensing receptor in parathyroidgland physiology. Am J Physiol Renal Physiol 2004;286:F1005–11.
14] Datta NS, Abou-Samra AB. PTH and PTHrP signaling in osteoblasts. Cell Signal2009;21(8):1245–54.
15] Mundy GR, Edwards JR. PTH-related peptide (PTHrP) in hypercalcemia. J AmSoc Nephrol 2008;19(4):672–5.
16] Kronenberg HM. PTHrP and skeletal development. Ann N Y Acad Sci2006;1068:1–13.
17] Monk R, Bushinsky D. Making sense of the latest advice on vitamin D therapy.J Am Soc Nephrol 2011;22:994–8.
18] Hewison M, Zehnder D, Bland R, et al. 1alpha-Hydroxylase and the action ofvitamin D. J Mol Endocrinol 2000;25:141–8.
19] Jones G, Strugnell SA, DeLuca HF. Current understanding of the molecularactions of vitamin D. Physiol Rev 1998;78:1193–231.
20] Sutton AL, MacDonald PN. Vitamin D: more than a “bone-a-fide” hormone. JMol Endocrinol 2003;17:777–91.
21] Shimada T, Mizutani S, Muto T, et al. Cloning and characterization of FGF-23 asa causative factor of tumor-induced osteomalacia. Proc Natl Acad Sci U S A2001;98(11):6500–5.
22] Shimada T, Kakitani M, Yamazaki Y, et al. Targeted ablation of FGF-23 demonstrates an essential physiological role of FGF-23 in phosphate andvitamin D metabolism. J Clin Invest 2004;113(4):561–8.
23] Perwad F, Azam N, Zhang MY, et al. Dietary and serum phosphorus regulatefibroblast growth factor-23 expression and 1,25-dihydroxyvitamin D metabo-lism in mice. J Endocrinol 2005;46(12):5358–64.
24] Gattineni J, Bates C, Twombley K, et al. FGF-23 decreases renal NaPi-2a andNaPi-2c expression and induces hypophosphatemia in vivo predominantly viaFGF receptor 1. Am J Physiol Renal Physiol 2009;297:F282–91.
25] Secreto FJ, Monroe DG, Spelsberg TC. J Bone Miner Metab 2006:84–9.26] Gogakos AI, Duncan Basset JH, Williams GR. Thyroid and bone. Arch Biochem
Biophys 2010;503(1):129–36.27] Memon F, Mohga El-Abbadi M, Nakatani T, et al. Does FGF-23-klotho activity
influence vascular and soft tissue calcification through regulating phosphatehomeostasis? Kidney Int 2008;74(5):566–70.
28] Cannata-Andia JB, Roman-Garcia P, Hruska K. The connections betweenvascular calcification and bone health. Nephrol Dial Transplant 2011;26:3429–36.











![Synthèse et étude structurale des phosphates mixtes des ... · , matériau de choix dans les batteries au lithium [9-10]. 5O 14 De nombreux phosphates constituent des structures](https://img.pdfslide.fr/doc/110x75/5f0228977e708231d402dc8d/synthse-et-tude-structurale-des-phosphates-mixtes-des-matriau-de-choix.jpg)
















