Embed Size (px)
Citation preview

Nicolas MARC Septembre 2012
Adaptation et test d’un nouvel outil de développement postlarvaire Couplage avec la culture en masse de microalgues en Lagunage à Haut Rendement
Algal (LHRA)
Pré-étude de faisabilité d’une micro-nurserie d’huîtres creuses (Crassostrea gigas)

2 Cépralmar / Préfaisabilité micro-nurserie Crassostrea gigas / Septembre 2012
Sommaire
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
2. . . 3 2. Matériels et Méthodes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
3. Résultats et discussion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11
4. Bilan et Perspectives . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15
Bibliographie . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 17
Annexe . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21

3 Cépralmar / Préfaisabilité micro-nurserie Crassostrea gigas / Septembre 2012
lml
L’huître creuse Crassostrea gigas est devenue une espèce d'importance vitale pour la
conchyliculture, et représente plus de 99% de la production d'huîtres en France (FAO, 2010).
Bien que la majorité des huîtres cultivées proviennent du recrutement naturel (5,6 milliards de
naissains sauvages captés: 80% de l'approvisionnement), la demande de juvéniles d’écloserie
ne cesse d'augmenter (Ponis et al., 2006). Cette expansion se produit en raison de la forte
saisonnalité et des variations dans le recrutement de cette espèce (Robert et Gerard, 1999).
Les producteurs ont également montré un intérêt croissant pour de nouveaux produits
conchylicoles, comme les huîtres triploïdes, qui offrent une fertilité réduite, l'augmentation de
la croissance et potentiellement une plus grande résistance à la mortalité estivale (Samain et
McCombie, 2007). De plus la conchyliculture française connaît actuellement une crise sévère,
liée à la mortalité du naissain de C. gigas, quel que soit son origine (captage ou écloserie ;
Pernet et al., 2010). L’approvisionnement est donc devenu une priorité pour la filière et les
écloseries commerciales sont de ce fait fortement sollicitées.
En Languedoc-Roussillon, l’ostréiculture est entièrement dépendante des apports de juvéniles
provenant de la façade atlantique. Les professionnels sont donc confrontés à la fois à une
dépendance mais également à un manque de naissain. La crise de mortalité des juvéniles qui
touche la production d’huîtres creuses a été partiellement amortie par l’augmentation des prix
de vente en gros, mais est amplifiée par les difficultés d’approvisionnement en naissain. En
effet, moins de la moitié des besoins en naissain ont été satisfaits en 2011 (le besoin pour
soutenir la production ostréicole est estimé à environ 300 millions de naissains).
Dans un objectif de limitation des transferts de naissains afin d’éviter les vectorisations de
pathologies, et surtout, pour parvenir à terme à une indépendance de la filière ostréicole en
Languedoc-Roussillon, un test de faisabilité d’une micro-nurserie est lancée.
Le premier volet de ce projet consiste en l’étude de pré-faisabilité de la production de
postlarves d’huîtres creuses.
La reproduction contrôlée des bivalves est relativement récente (Loosanoff et Davis, 1963;
Walne, 1970), et malgré les progrès incontestables qui ont été accomplis dans le
conditionnement des géniteurs, la production de phytoplancton et les développements des
élevages larvaires et post-larvaires (Helm et al., 2004), les techniques de production en
écloserie ne sont pas encore pleinement définies pour la plupart des espèces économiquement
importantes. La connaissance ayant été principalement acquise par une approche empirique,
l’optimisation du processus d'écloserie est donc nécessaire et implique une meilleure
compréhension des besoins physiologiques des mollusques à tout stade de développement.
Un objectif majeur de la production en écloserie est d'améliorer la survie des larves et post-
larves tout en maximisant leur croissance. Aussi, l'alimentation a été considérée comme
l'aspect le plus important et de nombreux travaux ont porté principalement sur le choix des
micro-algues fourrage associé à leur composition biochimique (Brown et al, 1998; Knuckey
et al, 2002; Ponis et al, 2006). Cependant, l'amélioration des techniques d'élevage doit aussi
permettre une amélioration des performances du développement en répondant au mieux aux
besoins écophysiologiques des larves et postlarves de mollusques bivalves.
L’amélioration des techniques d’élevage passe par une optimisation de la structure d’élevage.
Ainsi, lors de cette étude nous tenterons d’adapter un outil utilisé dans un autre contexte à la
station Ifremer de Palavas. Il s’agit de colonnes de filtration biologiques en lit fluidisé
1. Introduction

4 Cépralmar / Préfaisabilité micro-nurserie Crassostrea gigas / Septembre 2012
(cylindres PVC transparents utilisées pour épurer les eaux d’effluent piscicole (Interdonato,
2012). Or, leur forme cylindrique combinée à leur système hydraulique en flux ouvert
ascendant pourrait permettre le développement postlarvaire de C. gigas. Cependant, afin
d’être utilisée comme structure d’élevage postlarvaire, une colonne subira des modifications
puis sera testée pour aboutir le cas échéant à la validation de ce nouvel outil d’élevage
postlarvaire.
De plus, la production de mollusques bivalves en écloserie est fortement soumise à la qualité
et à la quantité des microalgues fournies. En effet, les coûts de production phytoplanctoniques
représentent plus de 30 % de la gestion d’une écloserie (Benemann, 1992 ; Coutteau et
Sorgeloos, 1992). Le choix des microalgues cultivées et de leur proportion dans la ration est
donc d’une importance stratégique afin de diminuer au maximum ce coût.
Sur la plateforme d’Ifremer Palavas, des cultures d’algues en LHRA (Lagunage à Haut
Rendement Algal) sont menées pour le traitement des eaux d’effluents d’élevages piscicoles.
Ces cultures sont lancées à partir d’un inoculum naturel (eau brute de l’étang du Prévost)
chargé en phytoplancton. Le milieu de culture est enrichi en P et N pour permettre la
croissance des microalgues i.e. la multiplication cellulaire.
Le lagunage est donc une culture d’algues qui reproduit les phénomènes d’autoépuration qui
réalisés naturellement dans les milieux aquatiques en réponse aux apports de sels nutritifs et
de matières organiques. Ils sont utilisés depuis l’antiquité pour la pisciculture en Asie et en
Grèce (Gloyna, 1972). De nombreux auteurs ont étudié l’utilisation des algues pour le
traitement des effluents marins d’aquaculture (Goldman et al, 1974 a et b; Krom et Van Rijm,
1989 ; Vandermeulen et Gordin, 1990 ; Cohen et Neori, 1991 ; Neori et al, 1991 ; Jimenez
Del Rio et al, 1994; Craggs et al, 1995 ; Schuenhoff et al, 2003). Le LHRA diffère du
lagunage classique par une plus faible profondeur des bassins et un brassage continu de la
masse d’eau qui favorisent la croissance des algues (Deviller, 2003). Cette technique semble
ainsi répondre aux objectifs de diminution du coût de production des microalgues (pas de
salle d’algue et inoculum disponible et gratuit).
Dans cette étude de pré-faisabilité, nous allons donc tenter de vérifier l’appétence des jeunes
postlarves d’huîtres creuses pour ces microalgues ensemencées naturellement et présentes
dans le milieu naturel à l’échelle régionale. Nous pourrons alors vérifier leur valeur
nutritionnelle pour le naissain.
Ainsi les deux objectifs majeurs de cette étude de préfaisabilité sont:
1. Adapter et tester l’outil existant à Ifremer Palavas en proposant des
améliorations afin d’autoriser le développement postlarvaire de C. gigas.
2. Tester la faisabilité du couplage production phytoplanctonique de masse (LHRA)
/ croissance de jeunes postlarves d’huîtres creuses
.

5 Cépralmar / Préfaisabilité micro-nurserie Crassostrea gigas / Septembre 2012
2.1 Matériel biologique
Origine des postlarves
Les postlarves sont issues de la ponte réalisée le
11/06/2012 à l’écloserie commerciale NOVOSTREA, à
partir d’un pool de géniteurs diploïdes provenant de
Méditerranée (Etang de Thau, 34) et de l’Atlantique
(Abers, 29). La phase larvaire s’est déroulée sur 25 jours
et a été suivie de 10 jours de fixation.
Afin d’obtenir une population homogène pour l’étude,
les postlarves ont subi un tamisage sélectif entre 500 et
750 µm.
A leur réception les 500 000 postlarves représentaient un
poids total de 106,9 g soit un poids individuel moyen de
0,21 mg pour une taille individuelle moyenne de 0,63
mm (figure 1).
Microalgues
LHRA
Les microalgues testées sont issues de la production en lagunage à haut rendement algal
(LHRA) de la station Ifremer de Palavas les Flots.
Le milieu de culture est ensemencé avec un inoculum provenant directement du milieu
naturel. Les espèces microalgales se développant ultérieurement sont donc les mêmes que
celles rencontrées localement, dans l’étang du Prévost (34).
Microalgues fourrages de références
Les deux espèces de microalgues qui servent de référence à la production de postlarves
d’huitres creuses ont été utilisées comme alimentation témoin.
- Isochrysis galbana (Iso, souche CCAP 927/14 ; Parke, 1949)
Classe des Prymnesiophyceae (Hibberd, 1976)
Ordre des Isochrysidacaea
Il s’agit d’une espèce unicellulaire de petite taille (2 à 5 µm), de couleur brune. Elle est
caractérisée par sa forme ovoïde, ses deux flagelles lisses et un haptomène très court. Son
volume cellulaire moyen est d’environ 40 µm3.
Elle est considérée comme une bonne algue fourrage et sert de référence dans la plupart des
travaux touchant à la nutrition des mollusques bien que la confusion avec Isochrysis affinis
galbana (T. iso) ne facilite pas l’interprétation dans la littérature.
- Chaetoceros calcitrans (Cc, souche UTEX LB 2375 ; Ehrenberg, 1844)
Classe des Bacillarriophyceae (Haeckel, 1878 emend. Round et al., 1990)
Ordre des Chaetocerotales
Cette diatomée unicellulaire possède un corps rectangulaire ou carré de 3-4 µm et des soies
d’environ 10 µm. Son volume cellulaire moyen est d’environ 80 µm3.
2. Matériels et Méthodes
Figure 1 : Photographie de
l’ensemble des postlarves
réceptionnées

6 Cépralmar / Préfaisabilité micro-nurserie Crassostrea gigas / Septembre 2012
2.2 Système d’élevage postlarvaire en flux ouvert vertical
Adaptation de l’outil existant
A l’origine, la colonne d’élevage était utilisée à la
station d’Ifremer Palavas comme filtre biologique
(Interdonato, 2012) en lit fluidisé (média) afin de
traiter les effluents d’élevage piscicole (figure 2).
Dans le cadre de notre étude, la colonne a dû être
adaptée pour autoriser la production de postlarves
d’huîtres creuses.
Afin d’empêcher les postlarves de s’échapper par
le bas de la colonne en cas d’arrêt de la pompe
d’arrivée d’eau, un tamis d’une maille de 400 µm
a été placé au dessus de la réduction en PVC. Cette
segmentation du tube permet de plus de se trouver
dans la partie en PVC transparent, ce qui rend
possible l’observation des animaux et le réglage
visuel du débit nécessaire autorisant un bon
développement postlarvaire.
Les 500 000 naissains seront répartis sur ce tamis à
plat d’un diamètre de 15 cm (voir figure 3), et
pourront ensuite se développer dans toute la
hauteur de la colonne d’eau jusqu’à atteindre
l’objectif de taille T2 (naissain restant au dessus
d’une maille de 2 mm).
Figure 3 : Photographies de la colonne d’élevage adaptée pour l’expérimentation, et zoom sur
les postlarves au sein de cette structure.
Figure 2 : Filtre biologique développé
à la station Ifremer de Palavas

7 Cépralmar / Préfaisabilité micro-nurserie Crassostrea gigas / Septembre 2012
Afin de limiter le coût de production, les postlarves seront élevées dans un système recirculé.
La recirculation permet de récupérer les microalgues non consommées par les postlarves lors
de leur passage dans la colonne. Pour que l’ensemble des postlarves aient accès à la nourriture
et que l’environnement trophique soit propice à de bonnes croissances (linéaires et
pondérales), il est nécessaire que la concentration algale en sortie
d’élevage soit d’environ 1,5 x 106 µm
3/µL. Cette concentration
algale de l’effluent de sortie constituerait donc une biomasse
inutilisée et perdue dans le cas d’un élevage conduit en circuit
ouvert. La recirculation permet ainsi à ces microalgues non
consommées de retourner dans le système d’élevage et de
diminuer significativement le besoin en microalgues qui
constitue le coût principal dans la production de naissain de C.
gigas en micronurserie.
Pour compenser la perte d’algues consommées par les postlarves,
une pompe doseuse délivrera en continu un débit d’algues
fraîches provenant d’un bassin de réserve de 200 L avec bullage
situé en extérieur (figure 4).
Enfin, une surverse reliée à l’égout permettra d’éliminer le volume d’eau d’élevage équivalent
au taux de renouvellement adopté.
Le schéma simplifié de ce système d’élevage est reporté en figure 5.
Figure 5 : Schéma simplifié du système de production postlarvaire, avec recirculation de l’eau
d’élevage.
Tamis 400 µm
Figure 4 : Réserve d’algues
Tube d’élevage
Postlarves C. gigas
Bac réserve 500 L

8 Cépralmar / Préfaisabilité micro-nurserie Crassostrea gigas / Septembre 2012
L’outil de référence
En parallèle, afin de tester l’influence du nouveau système d’élevage sur les performances de
développement postlarvaire, un élevage témoin a été lancé dans l’outil de référence utilisé à
l’écloserie expérimentale d’Ifremer Argenton (29).
Il s’agit de tubes cylindro-coniques de 500 mL en plexiglas transparent Artemio® JBL
(Figure 6). Ces tubes ont été conçus à l’origine pour la production d’artemias en
aquariophilie, mais leur forme en entonnoir associée au brassage énergique de l’eau permet de
les utiliser pour la phase de nurserie de C. gigas (Marc, 2011).
Les postlarves ont été maintenues en flux ouvert, i.e., avec un apport d’eau enrichi en
phytoplancton en continu adapté à la biomasse en élevage, afin de limiter les contaminations
bactériennes et la pollution des tubes induite par l’alimentation. Le procédé d’élevage en flux
continu facilite la gestion des structures d’élevage en minimisant les dépôts organiques dans
les enceintes d’élevage.
Figure 6: Elevage postlarvaire d’huîtres creuses en flux ouvert vertical, en réacteurs de
500mL (photographie Nicolas MARC, Ifremer Argenton)
Eau neuve
L’eau de mer provient d’un pompage en mer, situé à 4,5 m de profondeur au large de la station
Ifremer de Palavas les Flots. Elle est filtrée sur filtre à sable (40 µm) puis stérilisée à travers une
unité de rayons ultraviolets (UV). Une fois traitée, l’eau est stockée dans des réserves cylindro-
coniques de 500L (une réserve par système d’élevage). Ces dernières sont équipées d’un bullage
permanent permettant un équilibre des gaz dissous afin d’éviter d’éventuels problèmes de
saturation dans les enceintes d’élevage.
Enfin, une pompe de reprise permet d’envoyer cette eau dans les structures d’élevage par le bas.
Un contrôle bijournalier de ce flux est indispensable.
Lumière
L’éclairage de la salle est permanent (24 h lumière / 0 h obscurité), et ceci pour toute la durée
de l’élevage postlarvaire. Les néons diffusent une lumière d’un spectre dit « lumière du jour ».
L’intensité lumineuse relevée à proximité des tubes d’élevage est de 14,5 µmole m-2
s-1
.

9 Cépralmar / Préfaisabilité micro-nurserie Crassostrea gigas / Septembre 2012
Alimentation
L’alimentation des postlarves est automatisée et continue (24/24h) pendant toute la durée de
l’élevage. L’utilisation de pompes doseuses équipées de variateurs de vitesses permet
d’ajuster et de maintenir un environnement nutritionnel optimal. La valeur de la biomasse
disponible doit être proche de 1 500 µm3.µl
-1 soit environ 40 000 cellules algales par mL en
sortie d’élevage (Rico-Villa et al., 2008).
Un contrôle (matin et soir) dans l’effluent de sortie à l’aide d’un comptage à la cellule de
Malassez permet d’ajuster au plus juste cet environnement phytoplanctonique.
2.3 Protocole expérimental
Test de l’outil expérimental et comparaison avec le système de référence
Tout d’abord, une colonne a été testée pour l’élevage de 500 000 postlarves de C. gigas, ce
qui représente une biomasse totale de 106.5g. Ces postlarves ont été nourries avec du
phytoplancton extérieur produit en LHRA, et élevées à une température constante de 25°C.
Cette expérimentation s’est déroulée sur 10 jours. Cependant, pour des raisons de refus
alimentaire (cf partie résultats), nous avons décidé de tester l’incidence de l’outil d’élevage
sur le développement postlarvaire de C. gigas.
Pour ce faire, un tube d’élevage Artemio a été utilisé et a reçu une biomasse initiale de 7 g de
postlarves (poids total égoutté) soit environ 32 000 postlarves. Les animaux ont donc été
élevés dans les mêmes conditions de température et d’alimentation que la colonne d’élevage
mise en place préalablement. Ainsi, nous tenterons de vérifier si le refus alimentaire est
engendré par un problème zootechnique.
Incidence des espèces phytoplanctoniques utilisées
Trois régimes alimentaires ont été testés, dans trois tubes cylindro-coniques Artémio
(référence):
- Condition 1 : Microalgues issues du LHRA
- Condition 2 : Référence : Isochrysis galbana/Chaetoceros calcitrans (1 : 1)
- Condition 3 : A jeun
Chaque tube a reçu aléatoirement une biomasse initiale de 7 g de postlarves (poids total
égoutté), ce qui correspondait à environ 32 000 huîtres par structure.
Le renouvellement d’eau a été ajusté à la biomasse en élevage, i.e., mise en suspension
suffisante des postlarves mais sans choc entre les animaux. Il a donc varié de 2 L h-1
à 6 L h-1
au cours de cette expérimentation.
La température d’élevage était constante et égale pour toutes les conditions tout au long de
l’élevage (25 °C ± 0,5).

10 Cépralmar / Préfaisabilité micro-nurserie Crassostrea gigas / Septembre 2012
Les mélanges nutritionnels des deux premières conditions étaient stockés dans des bacs
réserves cylindro-coniques de 20 L. Ces réserves étaient vidées et rincées avant le remplissage
avec la nouvelle ration de microalgues fraîches.
Les postlarves élevées à jeun étaient élevées dans de l’eau sans enrichissement
phytoplanctonique. Cette eau était directement injectée dans le tube d’élevage après filtration
mécanique et passage à la lampe U.V.
2.4 Mesure des performances de développement postlarvaire
Poids
La biomasse totale de chaque structure d’élevage était obtenue deux fois par semaine à l’aide
d’une balance électronique de précision 10-3
g (XS 6035 DeltaRange® ; Mettler Toledo) pour
déterminer le poids total par tube et le poids individuel égoutté (mg). Un échantillon de
chacune de ces pesées était alors récupéré pour l’estimation du nombre de postlarves et de la
survie. Ces échantillons servaient également à la mesure de la taille des postlarves.
Taille
Deux fois par semaine, 200 postlarves était échantillonnées par tube d’élevage suite à la
pesée, pour déterminer la longueur maximale de la coquille (en mm).
Ces données ont été obtenues par analyse d’images numériques acquises avec un scanner à
plat (Scanner photo HPTM
Officejet 6210®).
Chaque échantillon était disposé puis étalé sur un récipient en verre, lui même complété d’eau
de mer. En effet, la prise de vue d’objets immergés évite la formation de nombreux reflets et
irisations qui perturbent l’analyse d’image (Moriceau, 2008). Les numérisations étaient
effectuées à une résolution de 600 ppp (nombre de pixels par pouce ; ici 236 pixels par cm).
Le centrage du récipient en verre s’avère important afin de limiter au mieux les déformations
dues aux erreurs de parallaxe créées par l’objectif du scanner associé à une profondeur de
champ importante (Moriceau, 2010).
Une fois la numérisation réalisée, l’image est alors « épurée » grâce au logiciel de retouche
d’image Photofiltre 6.5.2 ®. Seules les postlarves utilisables pour le traitement numérique de
l’image sont conservées, tous les « bruits » de l’image étant supprimés.
Cette nouvelle image est alors analysée à l’aide du logiciel ImageJ ®. Une fois cette image
reconstituée en niveaux de gris (8-bits), elle est « seuillée » afin d’obtenir un ensemble de
pixels soit blancs, soit noirs. Le même niveau de seuillage (0 à 220) a été utilisé tout au long
de l’élevage postlarvaire.
Il est alors possible de mesurer le paramètre choisi qui est appelé « Fit ellipse » dont le grand
axe (Major) représente la longueur maximale théorique d’une postlarve (Marc, 2011). Les
données obtenues sont alors enregistrées sous forme de tableau avec le logiciel Microsoft
Office Excel ® pour l’analyse future des résultats.
11 Cépralmar / Préfaisabilité micro-nurserie Crassostrea gigas / Septembre 2012
3.1 Test de l’enceinte d’élevage
Le premier essai a été conduit dans la colonne d’élevage avec l’intégralité des postlarves.
Elles étaient nourries en continu à partir du phytoplancton cultivé en LHRA. Nous n’avons
pas observé de gain pondéral au cours des 19 jours d’expérimentation.
L’évolution de la taille des animaux confirme cette inhibition de croissance. En effet, nous
n’avons pas enregistré de croissance linéaire significative au cours de l’expérimentation
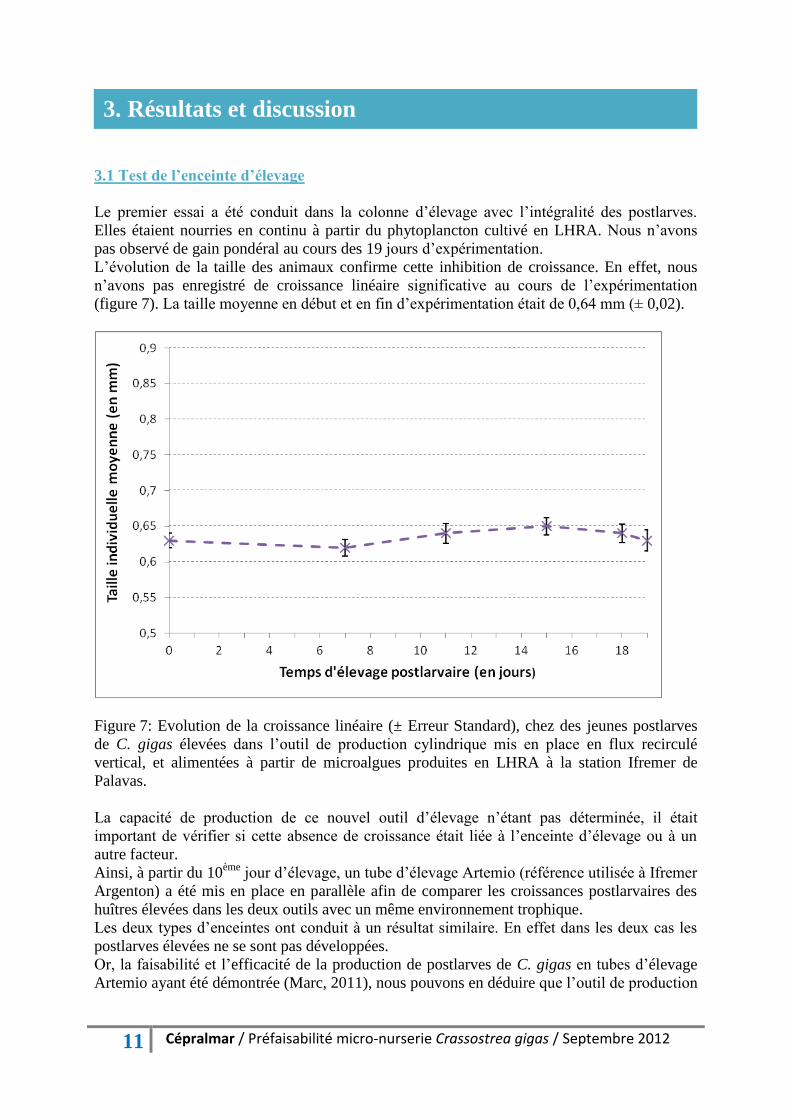
(figure 7). La taille moyenne en début et en fin d’expérimentation était de 0,64 mm (± 0,02).
Figure 7: Evolution de la croissance linéaire (± Erreur Standard), chez des jeunes postlarves
de C. gigas élevées dans l’outil de production cylindrique mis en place en flux recirculé
vertical, et alimentées à partir de microalgues produites en LHRA à la station Ifremer de
Palavas.
La capacité de production de ce nouvel outil d’élevage n’étant pas déterminée, il était
important de vérifier si cette absence de croissance était liée à l’enceinte d’élevage ou à un
autre facteur.
Ainsi, à partir du 10ème
jour d’élevage, un tube d’élevage Artemio (référence utilisée à Ifremer
Argenton) a été mis en place en parallèle afin de comparer les croissances postlarvaires des
huîtres élevées dans les deux outils avec un même environnement trophique.
Les deux types d’enceintes ont conduit à un résultat similaire. En effet dans les deux cas les
postlarves élevées ne se sont pas développées.
Or, la faisabilité et l’efficacité de la production de postlarves de C. gigas en tubes d’élevage
Artemio ayant été démontrée (Marc, 2011), nous pouvons en déduire que l’outil de production
3. Résultats et discussion
12 Cépralmar / Préfaisabilité micro-nurserie Crassostrea gigas / Septembre 2012
utilisé n’est pas à l’origine de cette inhibition de croissance. Un outil de production inadapté
aurait pu freiner le développement postlarvaire mais pas l’inhiber de façon totale.
Nous pouvons émettre l’hypothèse d’un refus alimentaire lié à la qualité spécifique ou
nutritionnelle du phytoplancton, ou bien encore à l’état de santé et à la condition intrinsèque
des postlarves. Ces deux hypothèses ont donc été vérifiées en testant l’incidence de différents
régimes alimentaires sur la croissance postlarvaire.
3.2 Incidence de l’alimentation sur le développement postlarvaire
Poids
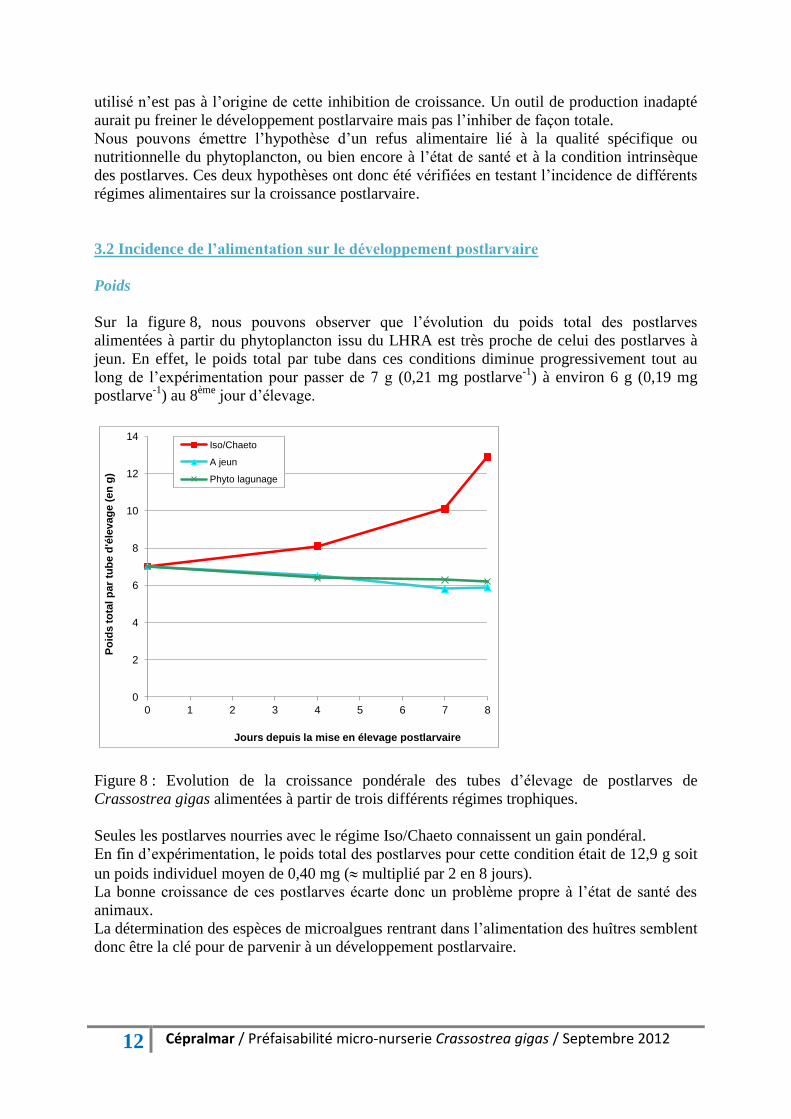
Sur la figure 8, nous pouvons observer que l’évolution du poids total des postlarves
alimentées à partir du phytoplancton issu du LHRA est très proche de celui des postlarves à
jeun. En effet, le poids total par tube dans ces conditions diminue progressivement tout au
long de l’expérimentation pour passer de 7 g (0,21 mg postlarve-1
) à environ 6 g (0,19 mg
postlarve-1
) au 8ème
jour d’élevage.
Figure 8 : Evolution de la croissance pondérale des tubes d’élevage de postlarves de
Crassostrea gigas alimentées à partir de trois différents régimes trophiques.
Seules les postlarves nourries avec le régime Iso/Chaeto connaissent un gain pondéral.
En fin d’expérimentation, le poids total des postlarves pour cette condition était de 12,9 g soit
un poids individuel moyen de 0,40 mg ( multiplié par 2 en 8 jours).
La bonne croissance de ces postlarves écarte donc un problème propre à l’état de santé des
animaux.
La détermination des espèces de microalgues rentrant dans l’alimentation des huîtres semblent
donc être la clé pour de parvenir à un développement postlarvaire.
0
2
4
6
8
10
12
14
0 1 2 3 4 5 6 7 8
Po
ids t
ota
l p
ar
tub
e d
'éle
vag
e (
en
g)
Jours depuis la mise en élevage postlarvaire
Iso/Chaeto
A jeun
Phyto lagunage
13 Cépralmar / Préfaisabilité micro-nurserie Crassostrea gigas / Septembre 2012
Taille
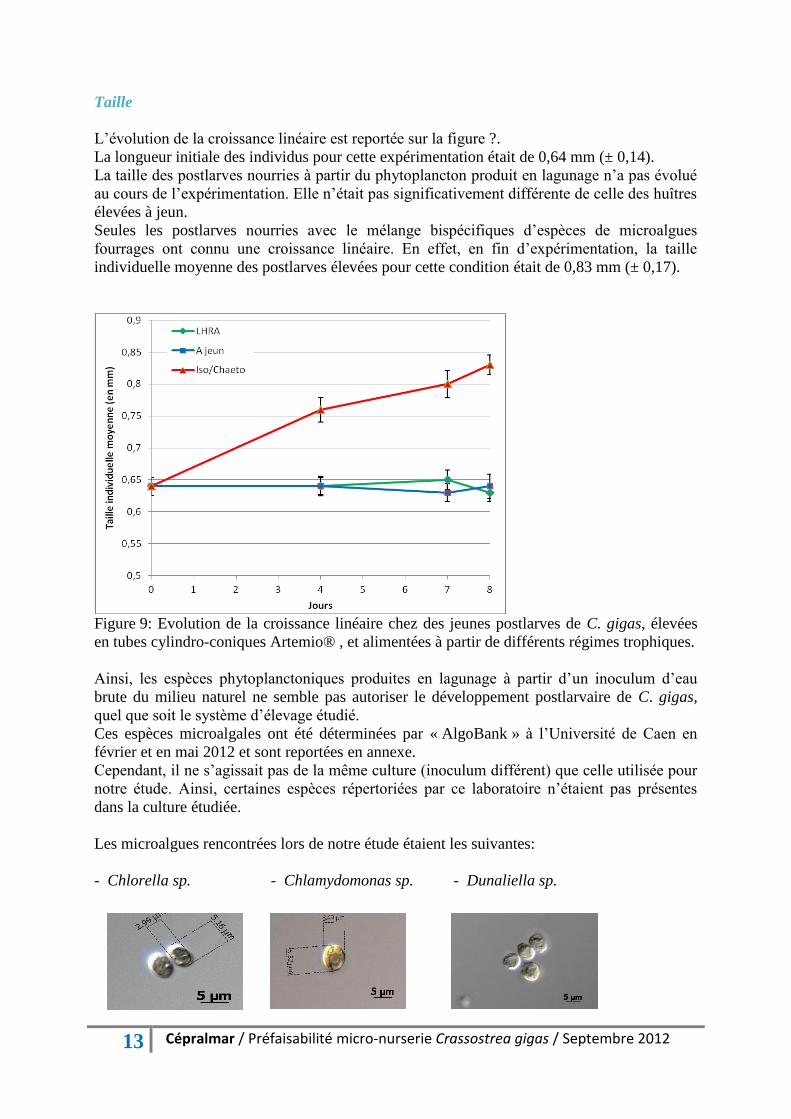
L’évolution de la croissance linéaire est reportée sur la figure ?.
La longueur initiale des individus pour cette expérimentation était de 0,64 mm (± 0,14).
La taille des postlarves nourries à partir du phytoplancton produit en lagunage n’a pas évolué
au cours de l’expérimentation. Elle n’était pas significativement différente de celle des huîtres
élevées à jeun.
Seules les postlarves nourries avec le mélange bispécifiques d’espèces de microalgues
fourrages ont connu une croissance linéaire. En effet, en fin d’expérimentation, la taille
individuelle moyenne des postlarves élevées pour cette condition était de 0,83 mm (± 0,17).
Figure 9: Evolution de la croissance linéaire chez des jeunes postlarves de C. gigas, élevées
en tubes cylindro-coniques Artemio® , et alimentées à partir de différents régimes trophiques.
Ainsi, les espèces phytoplanctoniques produites en lagunage à partir d’un inoculum d’eau
brute du milieu naturel ne semble pas autoriser le développement postlarvaire de C. gigas,
quel que soit le système d’élevage étudié.
Ces espèces microalgales ont été déterminées par « AlgoBank » à l’Université de Caen en
février et en mai 2012 et sont reportées en annexe.
Cependant, il ne s’agissait pas de la même culture (inoculum différent) que celle utilisée pour
notre étude. Ainsi, certaines espèces répertoriées par ce laboratoire n’étaient pas présentes
dans la culture étudiée.
Les microalgues rencontrées lors de notre étude étaient les suivantes:
- Chlorella sp. - Chlamydomonas sp. - Dunaliella sp.

14 Cépralmar / Préfaisabilité micro-nurserie Crassostrea gigas / Septembre 2012
Or, dans la littérature nous avons pu vérifier que Chlorella sp. (Walne, 1966 ; Babinchak et
Ukeles, 1979), Chlamidomonas sp. (His et Robert, 1986), et Dunaliella sp. étaient qualifiées
de mauvaises algues fourrages par rapport à leur faible qualité nutritionnelle pour les
mollusques. De plus, la valeur nutritionnelle des microalgues pour les bivalves dépend avant
tout de leur taille, autorisant leur ingestion, et leur digestibilité (Robert et Trintignac, 1997).
En effet, certaines espèces peuvent être ingérées mais non digérées (Robert, 1998). La
digestibilité d’une microalgue dépend à la fois de sa nature et de l’épaisseur de sa paroi. Ces
différentes études confortent ainsi les résultats que nous avons acquis.
L’observation lors de l’expérimentation de pseudo fèces en grande quantité était liée à cette
activité de filtration qui n’était pas suivie par l’étape de digestion. La production de pseudo-
fécès constitue une perte d'énergie non négligeable dans le bilan énergétique de l'animal et
l'optimisation des élevages (Soletchnik et al., 1996). La production de pseudo-fécès est une
caractéristique des mollusques filtreurs et constitue un moyen de contrôler l'ingestion de
matière en suspension (Newell et Jordan, 1983). Des études de physiologie ont démontré la
capacité des bivalves à exercer une sélection au sein du phytoplancton. Les mécanismes de tri
sont décrits en détail par Barillé et al. (1994). Ils peuvent s’exercer sur des caractéristiques
biométriques de la matière particulaire (e.g., taille, densité) (Atkins, 1937 ; Bernard,
1974) ; sur des critères de densité (Bernard, 1974); sur le type d'espèces
phytoplanctoniques (Loosanoff, 1949; Menzel, 1955; Kiorboe et Mohlenberg, 1981;
Newell et Jordan, 1983; Bricelj et Malouf, 1984, Iglésias et al., 1992; Navarro et al.,
1992); ou encore sur des critères organiques ou de nature biochimique (Foster-Smith, 1975 ;
Razet et al., 1990 ; Bayne et al., 1993).
Les bivalves filtreurs s'alimentent à partir de la matière en suspension dans l'eau de mer dont
la valeur nutritionnelle présente de fortes variations (Widdows et al., 1979 ; Héral et al.,
1980). En effet, dans le milieu naturel, les blooms phytoplanctoniques sont cycliques et
dépendent des conditions climatiques d’irradiance et de température et de la présence de
nutriments. Ainsi, en fonction de la saison, la diversité phytoplanctonique est modifiée. Dans
les conditions estivales de la station Ifremer de Palavas, les espèces produites en LHRA
(présentes dans l’étang du Prévost) ne convenaient pas aux postlarves.
Afin de vérifier si le LHRA modifiait la qualité nutritionnelle des microalgues et la diversité
spécifique, un test ad hoc à partir de l’eau brute concentrée du milieu naturel a été conduit.
Ainsi, nous avons constaté que les postlarves n’ont pas connu de croissance. Ceci nous permet
donc d’affirmer qu’à cette période de l’année (dans les conditions de l’étude) les espèces
microalgales présentes dans le milieu naturel ne permettaient pas le développement
postlarvaire de C. gigas.

15 Cépralmar / Préfaisabilité micro-nurserie Crassostrea gigas / Septembre 2012
A travers cette étude de préfaisabilité, nous voyons émerger l’importance de l’alimentation
pour le développement postlarvaire de C. gigas. La maitrise de la production de
phytoplancton en termes de qualité et de quantité est incontournable.
En effet, il semble judicieux d’être capable de produire du phytoplancton adapté à
l’alimentation du naissain d’huîtres avant de réfléchir à l’optimisation des méthodes de
production postlarvaire.
Les conditions du milieu naturel évoluent au cours de l’année et entrainent une modification
de la composition des communautés phytoplanctoniques. Aussi, ces variations spécifiques, ne
peuvent pas permettre d’utiliser cette eau de mer chargée naturellement en microalgues,
comme inoculum pour des cultures de masse en LHRA tout au long de l’année.
Deux axes de réflexion peuvent être proposés :
Ecologie : composition des communautés phytoplanctoniques lagunaires
Durant l’étude, aucune espèce microalgale autorisant le développement postlarvaire de
l’huître creuse n’a été observée. Or, la présence de mollusques bivalves captés naturellement
sur les rochers (moules, huîtres) dans les lagunes à proximité du site d’expérimentation nous
laisse supposer qu’à certaines périodes de l’année et de façon régulière, des blooms
phytoplanctoniques permettent leur développement.
Un suivi régulier des communautés phytoplanctoniques permettrait de vérifier l’éventuelle
présence de certaines espèces microalgales propices à l’alimentation des huîtres.
Des cycles phytoplanctoniques pourrait ainsi être établi, permettant d’appliquer un calendrier
de production de postlarves d’huîtres creuses adapté à la disponibilité du milieu naturel.
Pendant les périodes de blooms adaptés, l’eau brute du milieu naturel pourrait alors servir
d’inoculum au LHRA. Par exemple, l’étude réalisée par Algobank en février 2012 (voir
annexe) a permis d’identifier la microalgue Isochrysis sp. qui se révèle être une algue de
référence pour l’alimentation des mollusques bivalves. Ainsi, à cette période hivernale la
production en LHRA serait potentiellement utilisable pour le développement postlarvaire de
C. gigas.
Au-delà de l’aspect technique, il est intéressant de voir que cette étude ouvre des perspectives
et des questions très larges, aussi bien écologiques qu’économiques.
L’absence de microalgues favorables au développement postlarvaire de l’huître creuse au
cours d’une période pendant laquelle des juvéniles apparaissent spontanément dans le milieu
après les pontes naturelles, suscite un certain nombre de questions. Nous pouvons nous
demander si les blooms phytoplanctoniques qui sont sensés se produire conjointement à la
ponte des huîtres ont bien lieu, ou bien s’ils sont décalés dans le temps. Le cas échéant, les
larves et postlarves de mollusques, qui nécessitent une alimentation régulière durant ces
phases critiques du développement, ne pourraient pas se développer.
De plus, la région dispose de nombreux étang dont celui de Thau, qui est le principal bassin
de production ostréicole de méditerranée. Les compositions phytoplanctoniques étant
différentes dans ce milieu, l’eau de cette lagune pourrait être utilisée pour réitérer
4. Bilan et Perspectives

16 Cépralmar / Préfaisabilité micro-nurserie Crassostrea gigas / Septembre 2012
l’expérimentation et vérifier si des espèces favorables sont présentes à cette période,
contrairement à l’étang du Prévost.
Production contrôlée de microalgues fourrages
Un bassin de culture en LHRA (tel que celui utilisé à Ifremer Palavas) permet une production
de 6 m3 de phytoplancton. Un tel volume de production nécessite un inoculum initial
suffisamment concentré pour permettre le développement microalgal et éviter la
contamination par des espèces pionnières non désirées.
Deux types d’inoculums pourraient être étudiés :
- Eau brute d’un milieu naturel avec la présence d’espèces considérées comme
favorables au développement postlarvaire de l’huître creuse
- Culture de microalgues fourrages de référence pour l’élevage de mollusques bivalves
La faisabilité de la culture d’espèces fourrages telles que Isochrisis galbana ou Chaetoceros
gracilis, en LHRA doit être vérifiée. En effet, les conditions climatiques méditerranéennes
estivales (températures élevées et fort ensoleillement) pourraient jouer un rôle prépondérant
dans le choix des espèces en fonction de leurs preferendum thermiques.
Il est possible que certaines espèces utilisées en routine dans des nurseries sur la côte
atlantique (Skeletonema marinoï, Chaetoceros gracilis…) ne puissent pas se développer dans
ces conditions climatiques.
Enfin, cette étude de pré-faisabilité aura permis de vérifier l’importance de l’alimentation
pour le développement postlarvaire de l’huître creuse, et donc a fortiori la nécessité de
maitriser ce paramètre pour développer à terme une micronurserie en méditerranée.

17 Cépralmar / Préfaisabilité micro-nurserie Crassostrea gigas / Septembre 2012
Atkins O., 1937. On the ciliary mechanisms and interrelationships of the lamellibranchs. Part
II: Sorting devices on the gills. Journal of Microscopic Sciences, 79: 339-373.
Babinchak J. et Ukeles R., 1979. Epifluorescence microscopy, a technique for the study of
feeding in Crassostrea virginica veliger larvae. Marine Biology, Volume 51, 1, 69-76.
Barillé L., Prou J., Héral M., Bougrier S., 1993. No influence of food quality, but ration-
dependent retention efficiencies in the Japanese oyster Crassostrea gigas. J. Exp. Marine
Biology, 171, 91-106.
Bayne B.L., Iglesias J.P., Hawkins A.J.S, Navarro E., Héral M., Deslous-Paoli J.M., 1993.
Feeding behaviour of the mussel, Mytilus edulis: responses to variations in quantity and
organic content. J. Mar. Biol. Assoc. U.K 73, 813-R29.
Benemann J.R., 1992. Microalgae aquaculture feeds. J. Appl. Phycol., 4, 233-245.
Bernard, F.R., 1974. Particle sorting and labial palp function in the pacific oyster Crassostrea
gigas (Thunberg, 1975). Biol. Bull. mar. biol. Lab., Woods Hole 146: 1.
Bricelj V.M., Malouf R.E., 1984. Influence of algal and suspended sediment
concentrations on the feeding physiology of the hard clam Merpenaria mercenaria.-
Marine Biology, 84: 155-165.
Brown, B.L., Butt A.J., Shelton S.W., Paynter K.T., 1998. Growth and mortality of North
Carolina-heritage oysters, Crassostrea virginica, in North Carolina and in Chesapeake Bay. J.
Appl. Aquacult. 8: 25–39.
Cohen, I., Neori, A., 1991. Ulva lactuca biofilters for marine fishpond effluents. I. Ammonia
uptake kinetics and nitrogen content. Botanica Marina, 34: 475-482.
Coutteau P., Sorgeloos P., 1992. The use of algal substitutes and their requirement for live
algae in the hatchery and nursery rearing of bivalve molluscs: an international survey.
J.Shellfish Res., 11, 467-476.
Craggs, R.J., Smith, V.J., McAuley, P.J., 1995. Wastewater nutrient removal by marine
microalgae cultured under ambient conditions in mini-pond. Water Science and
Technicology, 31: 151-160.
Deviller, 2003. Traitement par Lagunage à Haut Rendement Algal (LHRA) des effluents
piscicoles marins recyclés : évaluation chimique et écotoxicologique. Thèse, Université de
Montpellier I. 172 pp.
Foster-Smith R.L., 1975a. The effect of concentration of suspension and inert material on
the assimilation of algae by three bivalves. J. mar. biol. Ass. U.K. 55: 411.
Bibliographie

18 Cépralmar / Préfaisabilité micro-nurserie Crassostrea gigas / Septembre 2012
Gloyna, E.F., 1972. Bassins de stabilisation des eaux usées. OMS, Genève.
Goldman, J.C., Tenore K.R., Stanley H.I., 1974a. Inorganic nitrogen removal in a combined
tertiary treatment-marine aquaculture system - II. Algal bioassays. Water Research, 8: 55-59.
Goldman J.C., Tenore K.R., Ryther J.H., Norwin N., 1974b. Inorganic nitrogen removal in a
combined tertiary treatment-marine aquaculture system-I. Removal efficiencies. Water
Research, 8: 45-54.
Helm M.M., Bourne N., 2004. The hatchery culture of bivalves: a practical manual. FAO
Fish, 1-184.
Héral M., Razet D., Maestrini S., Garnier J., 1980. Composition de la matière organique
particulaire dans les eaux du bassin de Marennes-Oléron. Apport énergétique pour la
nutrition des huîtres. Cons. Inter. Exp. Mer C.N .lL : pp 44.
His E., Robert R., 1986. Croissance des larves de Crassostrea gigas et de Mytilus
galloprovincialis en présence d'algues monocellulaires isolées du tractus digestif des véligères
du milieu naturel. Société Française de Malacologie, Symposium de Rochefort Ecologie,
Ecophysiologie, Energétique des Mollusques Marins et Continentaux.
Iglesias J.I.P., Navarro E., Alvarez Jorna P., Armentia I., 1992. Feeding, particle selection and
absorption in cockles Cerastoderma edule (L.) exposed to variable conditions of food
concentration and quality. J. Exp. Mar. Biol. Ecol. 162, 177 – 198.
Interdonato F., 2012. Recirculating aquaculture system (RAS) biofilters: focusing on bacterial
communities complexity and activity. Ifremer dept. BOME. Università degli studi di Messina.
Dottorato di Ricerca Scienze Ambientali: Ambiente Marino e Risorse. 124 pp.
Jiménez del Rio M., Ramazanov Z., Garcia-Reina G., 1994. Optimisation of yield and
biofiltering efficiencies of Ulva rigida C. Ag. cultivated with Sparus aurata L. wastewaters.
Scientia Marina, 58: 329-335.
Kiarboe T., Mshlenberg G., 1981. Particle selection in suspension-feeding bivalves. Mar.
Ecol. Prog. Ser. 5: 291-296.
Knuckey R.M., Brown M.R., Barret S.M., Hallegraeff G.M., 2002. Isolation of new
nanoplanktonic diatom strains and their evaluation as diets for juvenile Pacific oyster
(Crassostrea gigas). Aquaculture 211, 253-274.
Krom M.D., Van Rijn J., 1989. Water quality processes in fish culture systems: processes,
problems, and possible solutions. In N. De Pauw, E. Jaspers, H. Ackefors & N. Wilkins (eds),
Aquaculture, A Biotechnology in Progress, European Aquaculture Society, Berdene : 1091-
1111.
Loosanoff V.L., Davis H.C., 1963. Rearing of bivalve molluscs. In: Russel, F.S. (Ed.),
Advances in marine biology, vol. 1. Academic Press Inc., London. 136 pp.

19 Cépralmar / Préfaisabilité micro-nurserie Crassostrea gigas / Septembre 2012
Loosanoff V.L., 1949. On the food selectivity of oysters. Science. 110: 122.
Marc N., 2011. Influence de l’alimentation et de la température sur le développement
postlarvaire de l’huître creuse Crassostrea gigas. Mémoire Master 2 BAEMT, Université
Montpellier 2, 38 pp.
Menzel RW., 1955. Sorne phases of the biology of Ostrea equestris (Say) and a
comparison with Crassostrea virginica (Gmelin). Publications of the Institute of marine
Science, University of Texas, 4: 73-148.
Moriceau J., 2008. Les outils permettant l’étude de l’indice gonadique numérique de la nacre
Pinctada margaritifera (Photoshop, ImageJ). Ouvrage de soutien technique PJ0707
Développement durable de l’huître perlière. Laboratoire Ifremer Domestication Huître
Perlière, 17 pp.
Moriceau J., 2010. Biométries larvaires par analyse d’images numériques obtenues avec un
scanner à plat. Ouvrage de soutien technique. PJ0707 Développement durable de l’huître
perlière. Laboratoire Ifremer Domestication Huître Perlière, juin 2010, 16 pp.
Navarro E., Iglesias J.I.P., Ortega M.M., 1992. Natural sediment as a food source for the
cockle Cerastoderma edule (L.): effect of variable particle concentration on feeding, digestion
and the scope for growth. Journal of Experimental Marine Biology and Ecology, 156: 69-87.
Neori, A. Cohen, I., Gordin, H., 1991. Ulva lactuca biofilters for marine fishpond effluents,
II. Growth rate, yield and C:N ratio. Botanica Marina, 34: 483-489.
Newell R.l., S.l. Jordan 1983. Preferential ingestion of organic matter by the American
oyster Crassostrea virginica. Mar. Ecol. Prog. Ser. 13, 47-53.
Pernet F., Barret J., Marty C., Moal J., Le Gall P., Boudry P., 2010. Environmental anomalies,
energetic reserves and fatty acid modifications in oysters coincide with an exceptional
mortality event. Marine Ecology-Progress series, volume 401, 129-146.
Ponis E., Parisi G., Le Coz J.R., Robert R., Zittelli G.C., Tredici M.R., 2006. Effect of the
culture system and culture technique on biochemical characteristics of Pavlova lutheri and its
nutritional value for Crassostrea gigas larvae. Aquaculture Nutr. 12,322-329.
Razet D., M. Héral, J. Prou, J. M. Sornin 1990. Variations des productions de biodépôts
(fécès et pseudofécès) de l'huître Crassostrea gigas dans un estuaire macrotidal: baie de
Marennes-Oléron. Haliotis 10, 143-161.
Rico-Villa B., Whoerther P., Mingant C., Hamont M., Pouvreau S., Lepiver D., Robert R.,
2008. A flow-through rearing system device for Pacific oyster Crassostrea gigas larvae
ecophysiological studies. Aquaculture 282: 54-60.
Robert R., Gérard A., 1999. Bivalve hatchery technology: The current situation for
Crassostrea gigas and Pecten maximus in France. Aquaculture Living Resources 12 (2), 121-
130.

20 Cépralmar / Préfaisabilité micro-nurserie Crassostrea gigas / Septembre 2012
Robert R., Trintignac P., 1997. Microalgues et nutrition larvaire en écloserie de mollusques.
Société française de malacologie. Haliotis, 26 : 1-13.
Robert R., 1998. Nutritional inadequacy of Nannochloris atomus and Stichoccocus bacilaris
for the oyster Crassostrea gigas (Thunberg) larvae. Haliotis 27, 121-130.
Round F.E, Crowford R.M., Mann D.G., 1990. The Diatoms: biology & morphology of the
genera. Cambridge University Press, 732 pp.
Samain J.F., McCombie H., 2007. Mortalités estivales de l’huître creuse Crassostrea gigas.
Défi Morest. Edition Quae, 319 pp.
Schuenhoff, A., Shpigel, M., Lupatsch, I., Ashkenazi, A., Msuya, F.E., Neori, A., 2003.
Asemi-recirculating, integrated system for the culture of fish and seaweed. Aquaculture,
221:167-181.
Soletchnik P., Geairon P., Razet D., Goulletquer P., 1996. Physiologie de la maturation
et de la ponte de l'huître creuse Crassostrea gigas. RIDRV-96-05- RA/LA TREMBLADE,
27 pp.
Vandermeulen H., Gordin H., 1990. Ammonium uptake using Ulva (Chlorophyta) in
intensive fishpond systems: mass culture and treatment of effluent.J. Appl. Phycol., 2: 363-
374.
Walne P.R, 1966. Large scale culture of larvae Ostrea edulis. L., Fish. Invest., II 25, 1–52.
Walne P.R., 1970. Studies on the food value of nineteen genera of algae to juvenile bivalves
of the genera Ostrea, Crassostrea, Mercenaria and Mytilus, Fisheries Investigation vol. 26,
Ministry of Agriculture Fisheries and Food, U.K. 62 pp.
Widdows J., Fieth P., Worrall M., 1979. Relationship betwecn seston, available food and
feeding activity in the common mussel Mytilus edulis. Mar. Biol. 51, 195-207.

21 Cépralmar / Préfaisabilité micro-nurserie Crassostrea gigas / Septembre 2012
Annexe 1 : Détermination phytoplanctonique (Algobank Caen)
1- Méthode de dénombrement des microalgues en LHRA réalisée par
Algobank Caen
Les comptages ont été effectués sur cellule de comptage Mallassez au grossissement x200.
Un dénombrement par genres et espèces n’a pas été réalisable au vu du mauvais état de
conservation des cellules et de leur petite taille. Nous avons donc procédé à un dénombrement par
compartiment phytoplanctonique. Ainsi, les tableaux suivants présentent les résultats pour 3
différentes classes de tailles écologiques du phytoplancton : le picoplancton pour les cellules
comprises entre 0.2 et 2 µm, le nannoplancton pour les cellules comprises entre 2 et 20 µm et enfin
le microplancton pour les cellules comprises entre 20 et 200 µm.
2- Concentration cellulaire par compartiment phytoplanctonique
Les valeurs présentées sont exprimées en 106 de cellules par mL.
Lag C picoplancton (0.2-2µm) nannoplancton (2-20µm) microplancton(20-200 µm) population totale
16/02/2012 3.86 0.1 - 3.96
06/04/2012 13.9 11.7 - 25.61
04/05/2012 4.76 4.74 0.12 9.62
Lag D picoplancton (0.2-2µm) nannoplancton (2-20µm) microplancton(20-200 µm) population totale
16/02/2012 3.33 0.86 - 4.19
04/05/2012 2.97 3.85 0.15 6.97
3- Floristique
Chaque échantillon a été observé pour la détermination qualitative de la flore phytoplanctonique en
microscopie optique (Zeiss Axio Scope. A1). Pour ce faire, un aliquot de l’échantillon est observé sur
une lame aux grossissements x100, x200, x400 et x1000. Nous précisons que cette floristique
présente les microalgues dominantes de la communauté phytoplanctonique observée pour chaque
échantillon.
Annexes

22 Cépralmar / Préfaisabilité micro-nurserie Crassostrea gigas / Septembre 2012
- Février 2012
Chlorella sp (salina ?) Phylum : Chlorophyta Classe : Chlorophyceae Ordre : Chloroccocales Famille : Oocystaceae
Chlamydomonas sp.
ou Dunaliella
sp.(primolecta)
Phylum : Chlorophyta
Classe : Chlorophyceae
Ordre : Volvocales
Isochrysis sp.
Phylum : Haptophyta
Classe : Coccolithophyceae
Ordre : Isochrysidales
Famille : Isochrysidaceae
- Mai 2012
Chlorella stigmatophora
Phylum : Chlorophyta
Classe : Chlorophyceae
Ordre : Chloroccocales
Famille : Oocystaceae
Cylindrotheca closterium
Phylum : Ochrophyta
Classe : Bacillariophyceae
Ordre : Bacillariales
Famille : Bacillariaceae

23 Cépralmar / Préfaisabilité micro-nurserie Crassostrea gigas / Septembre 2012
Espèces présentes dans les LHRA en plus faible proportion (en février et mai 2012)
Amphora sp.
Phylum : Ochrophyta
Classe : Bacillariophyceae
Ordre : Thalassiophysales
Famille : Catenulaceae
Pinnularia sp.
Phylum : Ochrophyta
Classe : Bacillariophyceae
Ordre : Naviculales
Famille : Pinnulariaceae
Nitzchia sp.
Phylum : Ochrophyta
Classe : Bacillariophyceae
Ordre : Bacillariales
Famille : Bacillariaceae
Cylindrotheca closterium
Phylum : Ochrophyta
Classe : Bacillariophyceae
Ordre : Bacillariales
Famille : Bacillariaceae
Synedra sp.
Phylum : Ochrophyta
Classe : Bacillariophyceae
Ordre : Fragilariales
Famille : Fragilariaceae
























![Nurserie ambiance [Mode de compatibilité] - fidocl.fr« le risque de diarrhée, ... 1 Régulation MVC 10 368 1 Sonde température 12 K 15 m 44 1 Coffret électrique complet 420 12](https://img.pdfslide.fr/doc/110x75/5ad8e2f47f8b9a865b8e045f/nurserie-ambiance-mode-de-compatibilit-le-risque-de-diarrhe-1-rgulation.jpg)



