Embed Size (px)
Citation preview

Primer-mediated Enzymatic Amplification of Cytomegalovirus (CMV) DNAApplication to the Early Diagnosis of CMVInfection in Marrow Transplant Recipients
Sharon A. Cassol, Man-Chiu Poon, Rajinderpal Pal, Margaret J. Naylor, Jeremy Culver-James, Thomas J. Bowen,James A. Russell, Stephen A. Krawetz, Richard T. Pon, and David 1. HoarThe Canadian Red Cross, Blood Transfusion Service, Calgary, Alberta, Canada T2R IJI; and the Departments of MedicalBiochemistry, Pediatrics, and Medicine, the University of Calgary, Calgary, Alberta, Canada T2N4NI
Abstract
A nucleic acid amplification procedure, the polymerase chainreaction (PCR), has been used to establish a diagnostic assayfor the identification of cytomegalovirus (CMV) immediate-early sequences in clinical specimens. Preliminary testingagainst virus-infected cell cultures indicated that the PCRassay was highly CMV-specific, recognizing both wild-typeand laboratory strains of CMV. There was no cross-reactivitywith human DNAor with DNAfrom other herpes viruses. Thesensitivity of the assay, using cloned CMVAD169 Eco RIfragment-J as template, was 1 viral genome per 40,000 cells.In a prospective study of CMVinfection in bone marrowtransplant recipients, the PCRassay correctly identified fourpatients with confirmed CMVinfection. In three of these pa-tients who were followed longitudinally, correlation of DNAreactivity with CMVculture and CMVantibody status overtime indicated that DNAwas the most sensitive marker for thediagnosis of CMVinfection.
Introduction
Cytomegalovirus (CMV)' infections are a major cause of ill-ness and death in immunocompromised patients (1, 2). Asmany as 25% of allogeneic bone marrow transplant (BMT)recipients die of disseminated CMVdisease with interstitialpneumonitis (IP) (3). Rapid diagnosis and early treatmentform the basis for the effective clinical management of thesepatients.
Currently most diagnoses depend on a rise in anti-CMVantibody titer or on the direct isolation of CMVfrom infectedtissues. Neither procedure is satisfactory. Antibody responsesare often delayed or completely absent in immunosuppressedmarrow recipients (4) and recovery of CMVin culture cantake up to 6 wk to obtain a definitive result (5).
Address reprint requests to Dr. S. Cassol, The Canadian Red Cross,Blood Transfusion Service, 737 13th Avenue, S.W., Calgary, Alberta,Canada T2R 1J 1.
Receivedfor publication 12 February 1988 and in revisedform 23September 1988.
1. Abbreviations used in this paper: BMT, bone marrow transplant;CMV, cytomegalovirus; HSV, herpes simplex virus; IE, immediate-early; IP, interstitial pneumonitis; PCR, polymerase chain reaction;VZV, varicella zoster virus.
Recently it has been shown that the rapidity of CMVdiag-nosis can be increased using either MAbto demonstrate thepresence of viral antigens (6) or DNAprobes to detect viralnucleic acid (7-9). The major drawbacks of these technologieshave been their lack of sensitivity when compared with con-ventional culture methods (10, 1 1) and, in the case of DNA,the frequent hybridization of probes with bacterial sequencesfound in nonsterile specimens (12, 13).
To circumvent these problems and facilitate the diagnosisof CMVdisease, we have used an in vitro gene amplificationtechnique to increase both the sensitivity and specificity ofCMVDNAdetection. The technique, known as the polymer-ase chain reaction (PCR), uses synthetic oligonucleotideprimers and DNApolymerase to enzymatically amplify se-lected DNAtarget sequences, up to several million-fold in 2 h(14). PCRamplification has been successfully applied to thediagnosis of sickle cell anemia (15), hemophilia (16), humanpapilloma virus (17), HIV-1 (18, 19), and the human T celllymphoma/leukemia virus (20). Wereport the adaptation ofPCRtechnology to the identification of CMVDNAin blood,urine, saliva, and lung tissue and we describe our preliminaryexperience using this technology to longitudinally monitorCMVinfections in BMTrecipients. To evaluate the efficacy ofPCRtesting we have correlated CMVDNApatterns over timewith clinical outcome and with CMVserology and the abilityto isolate CMVin culture.
Methods
Patients and specimen collection. Seven allogeneic BMT recipientsreceiving their transplants at the Foothills Hospital, Calgary, AB, werelongitudinally sampled pretransplant and on several occasions (up to10 times) posttransplant. An eighth BMTrecipient, a patient at theMontreal General Hospital, was sampled on two occasions only.Serum was collected for antibody studies; urine, saliva, and blood forCMVDNA studies. An aliquot of each specimen was sent to theProvincial Laboratory at Foothills Hospital for viral culture. Allograftdonors were similarly sampled at the time of transplantation. Controlsconsisted of 10 healthy apheresis donors sampled serially (every 3 mo)over an 18-mo period.
Viral and control DNA. Cloned Eco RI fragment-J (10.6 kb) con-taining the complete DNAsequence coding for the major immediate-early (IE) gene of CMVstrain AD169 was provided by Dr. B. Flecken-stein, Universitat Erlangen-Niirenberg, Erlangen, FRG(21). CMVAD169 (VR-538) and Davis (VR-807) were obtained from the Ameri-can Type Culture Collection in Rockville, MD, as were herpes simplexvirus type I (HSV-I) strain F (VR-733), HSV-II strains G(VR-734) andMS(VR-540), and varicella zoster virus (VZV) strains Oka (VR-795),Webster (VR-916), and Ellen (VR-586). EBV was the gift of Dr. J.Roder, Mount Sinai Medical Research Institute, Toronto, Ont., andpurified HSV-I total virion DNAwas obtained from Dr. R. Chan,University of Toronto, Toronto, Ont. Wild-type strains of herpes vi-ruses including eight isolates of CMV, six of VZV, and three each of
Improved Detection of Cytomegalovirus DNA in Clinical Specimens 1109
J. Clin. Invest.© The American Society for Clinical Investigation, Inc.0021-9738/89/04/1 109/07 $2.00Volume 83, April 1989, 1109-1115

Figure 1. Design ofPCRfor detection ofCMVIE sequences.(Top) The structure ofthe major IE gene ofCMVAD169 as deter-mined by Akrigg et al.(24). The bars num-bered 1-4 denote thefour exon sequences.(Middle) The region ofthe fourth exon whichis amplified in the PCRreaction. The two syn-thetic oligonucleotide
primers, SC-6 and SC-8, are 33 and 30 bases long and are comple-mentary to the (-) and (+) strands, respectively. The arrows indicatethe direction of primer extension. The primers are positioned so thatthe enzyme-catalyzed, template-directed extension product of one
primer serves as the template for the other. Repeated cycles of dena-turation, primer annealing, and extension result in the accumulationof a 368-bp CMV-specific amplification product that is defined bythe distance between the 5' ends of the two primers. (Bottom) Theamplification product and its relationship to the probe, SC-2. Theprobe is 30 bases long and is complementary to the (-) strand of theamplified product, but not to the primers.
30-base long synthetic probe (Fig. 1). Aliquots of each amplificationreaction (usually one-tenth of the final volume) were electrophoreti-cally separated in 2.0% agarose gels and transferred by alkaline blotmethod (29) onto Zeta-Probe (Bio-Rad Laboratories, Mississauga,Ont.) nylon membranes. After neutralization the filters were individu-ally prehybridized at 420C for 2 h in 10 ml of 0.5 Msodium phosphatebuffer (355 mgNaHPO4, 0.2 ml 85% H3PO4/l0 ml, pH 7.2) contain-ing 0.1 MNaCl, 1% BSA, 2% SDS, 30% formamide, 1.0 AsM EDTA,and 0.4 mg/ml yeast RNA. Hybridization with 8 ng/ml of [32P]SC-2probe in 10 ml of the same solution was carried out for 3 h at 420C.The probe was phosphorylated at its 5' terminus to a specific activity ofbetween 6 and 8 X 108 cpm/,ug using [gamma-32P]dATP and T4 poly-nucleotide kinase (30). Filters were washed three times at room tem-perature and once at 420C for 15 min each in 0.75 MNaCl, 0.075 Msodium citrate, and 0.1% SDS, pH 7.2. Autoradiography was routinelyperformed for 18 h at -70°C using X-Omat ARfilm (Eastman KodakCo., Rochester, NY) and a single intensifying screen (Lightning Plus;Dupont Instruments, Mississauga, Ont.).
Antibody studies. IgG and IgM antibodies to CMVwere deter-mined separately using a solid-phase enzyme immunoassay, Enzyg-nost anti-CMV (Behring Diagnostic, Montreal, Que.). For IgM anal-ysis, test samples were pretreated with RFAbsorbent (Behring Diag-nostic) to remove rheumatoid factor. Serum samples from individualpatients and donors were batched and analyzed simultaneously in asingle assay.
Results
HSV-I and HSV-II were provided by the Alberta Provincial Labora-tory. The presence of CMVin wild-type strains was confirmed byindirect immunofluorescence using an MAbagainst the 72,000-Dearly nuclear protein (1 1).
Isolation of DNA. The methods used for DNAextraction fromvirus-infected cell cultures, whole blood, PMN- and mononuclear-enriched blood fractions have been described previously (22, 23). Forstudies of CMVexcretion, 12 ml of urine or saliva (collected in saline)were centrifuged for 1 h at 38,000 rpm (180,000 g) in an SW-41 rotorin a centrifuge (Beckman Instruments, Inc., Palo Alto, CA). The con-
centrated viral pellets were then lysed and DNAwas extracted (22).10-,sl aliquots of each DNAextract (equivalent to 3.0 ml of the originalfluid) were amplified as described below.
Selection and preparation of DNAprimers and probe. For PCRstudies, we chose to amplify a 368-bp segment of the fourth exon of themajor CMVIE gene. This region of the genome was selected since it ishighly conserved between CMVstrains (24, 25) and it has no reportedhomology with human DNA(26, 27). Synthesis of the primers andprobe were performed on a DNAsynthesizer (model 380 A; AppliedBiosystems, Foster City, CA) using published sequence data (24) andthe methoxyphosphoramidite method (28). The design of the PCRprocedure for detection of CMVIE sequences is outlined in Fig. 1.
Sequence amplifpication with DNApolymerase. Amplification reac-
tions were performed according to the procedure of Kogan et al. ( 16). 1
Mg samples of DNA(or 10 Ml of urine/saliva) were routinely amplifiedfor 40 cycles using 1.0 U of Taq DNApolymerase (Perkin Elmer-Cetus, Rexdale, Ont.) and reaction times of 2 min at 50°C for anneal-ing, 3 min at 70°C for extension, and 1 min at 85°C for denaturation.In some experiments, crude cell lysates (representing 0.2 X 106 cells)were amplified directly without the use of DNAextraction. Lysateswere prepared by heating cell suspensions to 95°C for 5 min. DNAspecimens from individual patients (or donors) were analyzed ingroups, each of which represented a serial collection obtained over a
period of months. As a control on internal consistency, four referencestandards were included in each PCRreaction. These standards were
prepared by seeding l-,ug quantities of uninfected human Wi 38 fibro-blast DNAwith known amounts of CMVAD169 Eco RI fiagment-Jranging from 2.86 X 10-10 to 1.43 X I0- pmol.
Southern analysis of amplification products. Amplified sequenceswere detected by Southern transfer and hybridization with [32P]SC-2, a
Sensitivity and specificity of CMVPCRtesting. In preliminaryamplification studies, purified CMVAD169 Eco RI frag-ment-J was used as the IE gene template. The product of am-
plification was an SC-2 hybridizing fragment that migrated atthe position expected of an amplified 368-bp target sequence
(Fig. 2 B). When 1.43 X l0-7 pmol of Eco RI fragment-J was
amplified in the presence of 1 Atg of Wi 38 fibroblast DNA, theefficiency of the reaction, calculated by the method of Saiki etal. (14), was 75%. No hybridization signal was detected withunamplified DNAor with uninfected Wi 38 fibroblast DNA.
1 2 3 4 5 6 7 Figure2. Autoradio-graphs showing South-ern analysis of viral andcontrol DNAamplifiedby the CMVPCRpro-cedure (18-h exposure).(A) Amplification (40
1 2 3 1 2 3 4 5 cycles) of whole cell ly-sates infected withHSV-I strain F (lane 1),HSV-II strain MS(lane2), VZV strains Oka,
'_ = Webster, and Ellen* _ t (lanes 3, 4, and 5, re-
spectively), and CMVwild-type strains iso-
lated from clinical specimens (lanes 6 and 7). Only CMV-infectedsamples yielded a positive hybridization signal. (B) Amplification ofthe cloned IE sequence, CMVAD169 Eco RI fragment-J (400 pg).The intensity of the hybrization signal increased with increasingnumbers of PCRcycles. Lane 1, 15 cycles; lane 2, 20 cycles; lane 3,25 cycles. (C) Amplification (40 cycles) of DNAextracted from urine(lane 1), saliva (lane 2), whole blood (lane 3), and PMN-enriched pe-ripheral blood cells (lane 4) of an asymptomatic CMVseronegativeblood donor (negative control). Lane 5 contains 1 pg of Eco RI frag-ment-J (positive control).
1110 Cassol et al.
4 3 2 1OI.i ,,,,,,^t,X-7y-r 77 1r en
A
0I.74 0
to0.742 0.75U
, s
fI
MuWWIp cyclesof ampification
3 5'(OP) - probe (sC-2)
L........38 base pair-product
To estimate the sensitivity attainable with CMVPCRam-plification, Wi 38 fibroblast DNAwas spiked with serial dilu-tions of Eco RI fragments and amplified for 40 cycles. One-tenth of each amplification reaction was then analyzed bySouthern transfer hybridization with [32P]SC-2. Using this re-construction system, as little as 0.0001 pg (0.143 X 10-10 pmol,8.6 molecules) of the CMVIE gene was readily detectable in 2Asg of cellular DNA(320,000 cells) to give a sensitivity (after 18h of autoradiographic exposure) of 1 viral genome per 40,500cells. As outlined in Methods, a selection of these reconstitutedspecimens was included in each PCRreaction as a measure ofintertest variability. Our results showed little or no variationbetween runs. Weconsistently detected at least one viral ge-nome per 20,000 cells.
Specificity studies were performed on established cell lines(ATCC) harboring known strains of CMV(AD 169, Davis),HSV-I (F), HSV-II (G and MS), and VZV(Oka, Webster, andEllen). Additional controls included DNAextracted fromEBV-infected B95-8 marmoset cells, as well as 100 pg each ofpurified HSVvirion DNA, pBR322 DNAand Escherichia coliDNA. The high degree of specificity contributed by the PCRamplification procedure was shown by the complete lack ofany hybridization signal with amplified HSV, EBV, VZV,pBR322, or E. coli DNAs. Representative results are shown inFig. 2 A, lanes 1-5. In contrast, strong positive hybridizationsignals were obtained by the amplification of both Davis andAD169 strains of CMV(Fig. 3 B, lanes 8 and 9). Retesting ofthe same samples gave identical results; negatives were consis-tently negative and positives were consistently positive.
CMVPCRtesting of wild-type strains. Before using PCRfor diagnosis, it was necessary to establish whether the assaycould identify wild-type strains of CMVfound in clinical spec-imens. Eight clinical strains, grown from the urines of differentpatients, were passaged in human fibroblast cell lines. DNAwas extracted and amplified as described. In some studies theinfected cells were lysed and analyzed by direct amplificationwithout previous DNAextraction. All eight clinical strains ofCMVgave reproducible hybridization signals correspondingto the CMV-specific IE gene amplification product. Fig. 2 Ashows representative results for two of these isolates (lanes 6and 7). Wild-type HSVand VZV, on the other hand, failed togive any detectable hybridization signals in three differentPCRruns.
Direct CMVPCRtesting of clinical specimens: correlationswith clinical disease. A total of 504 specimens were analyzedby CMVPCR: 154 from BMTpatients and 350 from healthyasymptomatic blood donors. The results of PCR testing areshown in Table I. Again, many of these samples (both positiveand negative) were retested and identical results were ob-tained.
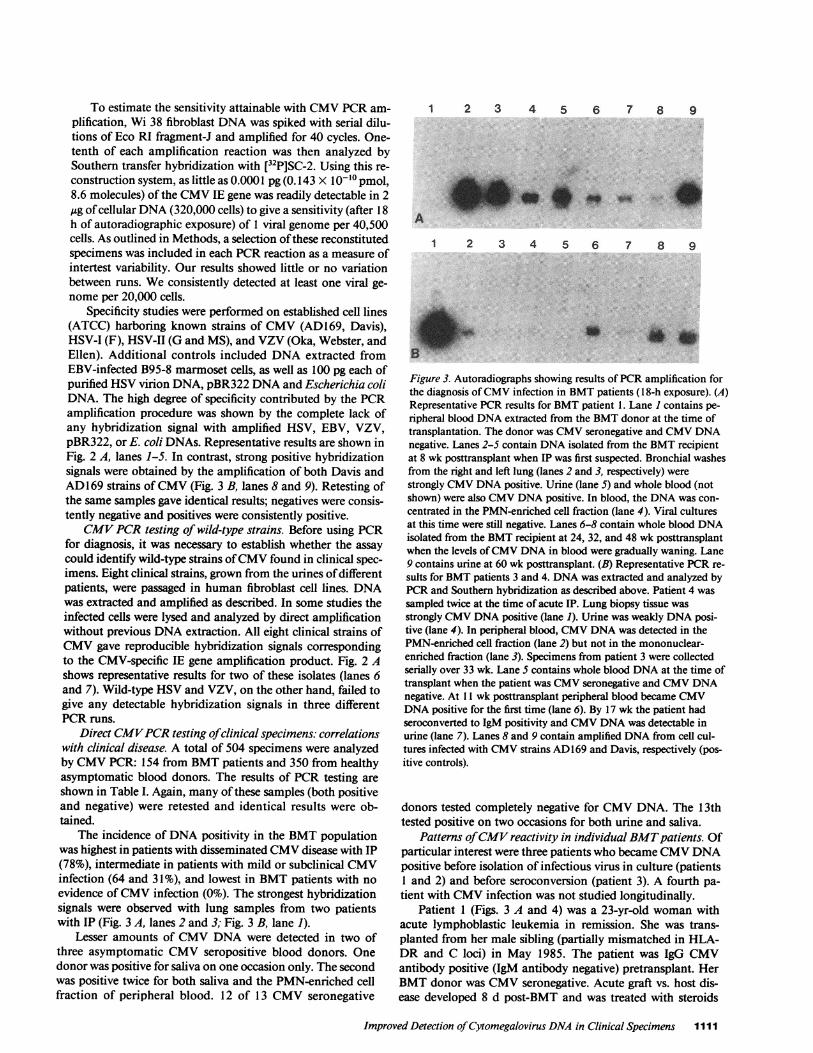
The incidence of DNApositivity in the BMTpopulationwas highest in patients with disseminated CMVdisease with IP(78%), intermediate in patients with mild or subclinical CMVinfection (64 and 31%), and lowest in BMTpatients with noevidence of CMVinfection (0%). The strongest hybridizationsignals were observed with lung samples from two patientswith IP (Fig. 3 A, lanes 2 and 3; Fig. 3 B, lane 1).
Lesser amounts of CMVDNAwere detected in two ofthree asymptomatic CMVseropositive blood donors. Onedonor was positive for saliva on one occasion only. The secondwas positive twice for both saliva and the PMN-enriched cellfraction of peripheral blood. 12 of 13 CMVseronegative
1 2 3 4 5 6 7 8 9
.M.&.
In
A,.-*_.41 t
A
B
1 2 3 4 5 6 7 8 9
h_.IS
Figure 3. Autoradiographs showing results of PCRamplification forthe diagnosis of CMVinfection in BMTpatients (1 8-h exposure). (A)Representative PCRresults for BMTpatient 1. Lane I contains pe-ripheral blood DNAextracted from the BMTdonor at the time oftransplantation. The donor was CMVseronegative and CMVDNAnegative. Lanes 2-5 contain DNAisolated from the BMTrecipientat 8 wk posttransplant when IP was first suspected. Bronchial washesfrom the right and left lung (lanes 2 and 3, respectively) werestrongly CMVDNApositive. Urine (lane 5) and whole blood (notshown) were also CMVDNApositive. In blood, the DNAwas con-centrated in the PMN-enriched cell fraction (lane 4). Viral culturesat this time were still negative. Lanes 6-8 contain whole blood DNAisolated from the BMTrecipient at 24, 32, and 48 wk posttransplantwhen the levels of CMVDNAin blood were gradually waning. Lane9 contains urine at 60 wk posttransplant. (B) Representative PCRre-sults for BMTpatients 3 and 4. DNAwas extracted and analyzed byPCRand Southern hybridization as described above. Patient 4 wassampled twice at the time of acute IP. Lung biopsy tissue wasstrongly CMVDNApositive (lane 1). Urine was weakly DNAposi-tive (lane 4). In peripheral blood, CMVDNAwas detected in thePMN-enriched cell fraction (lane 2) but not in the mononuclear-enriched fraction (lane 3). Specimens from patient 3 were collectedserially over 33 wk. Lane S contains whole blood DNAat the time oftransplant when the patient was CMVseronegative and CMVDNAnegative. At 11 wk posttransplant peripheral blood became CMVDNApositive for the first time (lane 6). By 17 wk the patient hadseroconverted to IgM positivity and CMVDNAwas detectable inurine (lane 7). Lanes 8 and 9 contain amplified DNAfrom cell cul-tures infected with CMVstrains AD169 and Davis, respectively (pos-itive controls).
donors tested completely negative for CMVDNA. The 13thtested positive on two occasions for both urine and saliva.
Patterns of CMVreactivity in individual BMTpatients. Ofparticular interest were three patients who became CMVDNApositive before isolation of infectious virus in culture (patients1 and 2) and before seroconversion (patient 3). A fourth pa-tient with CMVinfection was not studied longitudinally.
Patient 1 (Figs. 3 A and 4) was a 23-yr-old woman withacute lymphoblastic leukemia in remission. She was trans-planted from her male sibling (partially mismatched in HLA-DR and C loci) in May 1985. The patient was IgG CMVantibody positive (IgM antibody negative) pretransplant. HerBMTdonor was CMVseronegative. Acute graft vs. host dis-ease developed 8 d post-BMT and was treated with steroids
Improved Detection of Cytomegalovirus DNAin Clinical Specimens 1111
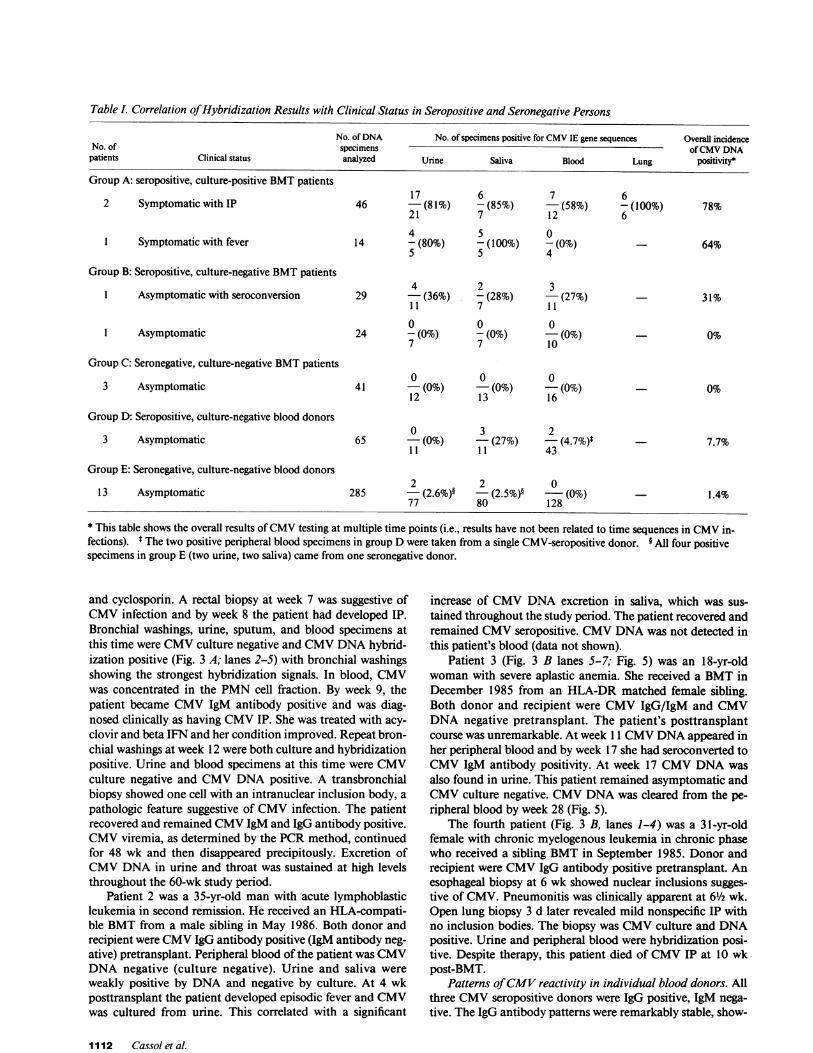
Table L Correlation of Hybridization Results with Clinical Status in Seropositive and Seronegative Persons
No. of DNA No. of specimens positive for CMVIE gene sequences Overall incidenceNo. of specimens of CMVDNA
patients Clinical status analyzed Urine Saliva Blood Lung positivity*
Group A: seropositive, culture-positive BMTpatients17 6 7 6
2 Symptomatic with IP 46 -(81%) - (85%) - (58%) - (100%) 78%2 1 7 12 64 5 0
1 Symptomatic with fever 14 - (80%) - (100%) - (0%) _ 64%5 5 4
Group B: Seropositive, culture-negative BMTpatients4 2 3
1 Asymptomatic with seroconversion 29 - (36%) - (28%) - (27%) - 31%1 1 7 1 1
0 0 01 Asymptomatic 24 - (0%) - (0%) - (0%) - 0%
7 7 10
Group C: Seronegative, culture-negative BMTpatients0 0 0
3 Asymptomatic 41 - (0%) - (0%) - (0%) - 0%1 2 1 3 1 6
Group D: Seropositive, culture-negative blood donors0 3 2
3 Asymptomatic 65 - (0%) - (27%) - (4.7%)* - 7.7%1 1 1 1 43
Group E: Seronegative, culture-negative blood donors2 2 0
13 Asymptomatic 285 -(2.6%)§ -(2.5%)§ - (0%) - 1.4%77 80 128
* This table shows the overall results of CMVtesting at multiple time points (i.e., results have not been related to time sequences in CMVin-fections). t The two positive peripheral blood specimens in group Dwere taken from a single CMV-seropositive donor. § All four positivespecimens in group E (two urine, two saliva) came from one seronegative donor.
and cyclosporin. A rectal biopsy at week 7 was suggestive ofCMVinfection and by week 8 the patient had developed IP.Bronchial washings, urine, sputum, and blood specimens atthis time were CMVculture negative and CMVDNAhybrid-ization positive (Fig. 3 A; lanes 2-5) with bronchial washingsshowing the strongest hybridization signals. In blood, CMVwas concentrated in the PMNcell fraction. By week 9, thepatient became CMVIgM antibody positive and was diag-nosed clinically as having CMVIP. She was treated with acy-clovir and beta IFN and her condition improved. Repeat bron-chial washings at week 12 were both culture and hybridizationpositive. Urine and blood specimens at this time were CMVculture negative and CMVDNApositive. A transbronchialbiopsy showed one cell with an intranuclear inclusion body, apathologic feature suggestive of CMVinfection. The patientrecovered and remained CMVIgM and IgG antibody positive.CMVviremia, as determined by the PCRmethod, continuedfor 48 wk and then disappeared precipitously. Excretion ofCMVDNA in urine and throat was sustained at high levelsthroughout the 60-wk study period.
Patient 2 was a 35-yr-old man with acute lymphoblasticleukemia in second remission. He received an HLA-compati-ble BMTfrom a male sibling in May 1986. Both donor andrecipient were CMVIgG antibody positive (IgM antibody neg-ative) pretransplant. Peripheral blood of the patient was CMVDNA negative (culture negative). Urine and saliva wereweakly positive by DNAand negative by culture. At 4 wkposttransplant the patient developed episodic fever and CMVwas cultured from urine. This correlated with a significant
increase of CMVDNAexcretion in saliva, which was sus-tained throughout the study period. The patient recovered andremained CMVseropositive. CMVDNAwas not detected inthis patient's blood (data not shown).
Patient 3 (Fig. 3 B lanes 5-7; Fig. 5) was an 18-yr-oldwoman with severe aplastic anemia. She received a BMTinDecember 1985 from an HLA-DR matched female sibling.Both donor and recipient were CMVIgG/IgM and CMVDNAnegative pretransplant. The patient's posttransplantcourse was unremarkable. At week 11 CMVDNAappeared inher peripheral blood and by week 17 she had seroconverted toCMVIgM antibody positivity. At week 17 CMVDNAwasalso found in urine. This patient remained asymptomatic andCMVculture negative. CMVDNAwas cleared from the pe-ripheral blood by week 28 (Fig. 5).
The fourth patient (Fig. 3 B, lanes 1-4) was a 31-yr-oldfemale with chronic myelogenous leukemia in chronic phasewho received a sibling BMTin September 1985. Donor andrecipient were CMVIgG antibody positive pretransplant. Anesophageal biopsy at 6 wk showed nuclear inclusions sugges-tive of CMV. Pneumonitis was clinically apparent at 61/2 wk.Open lung biopsy 3 d later revealed mild nonspecific IP withno inclusion bodies. The biopsy was CMVculture and DNApositive. Urine and peripheral blood were hybridization posi-tive. Despite therapy, this patient died of CMVIP at 10 wkpost-BMT.
Patterns of CMVreactivity in individual blood donors. Allthree CMVseropositive donors were IgG positive, IgM nega-tive. The IgG antibody patterns were remarkably stable, show-
1112 Cassol et al.
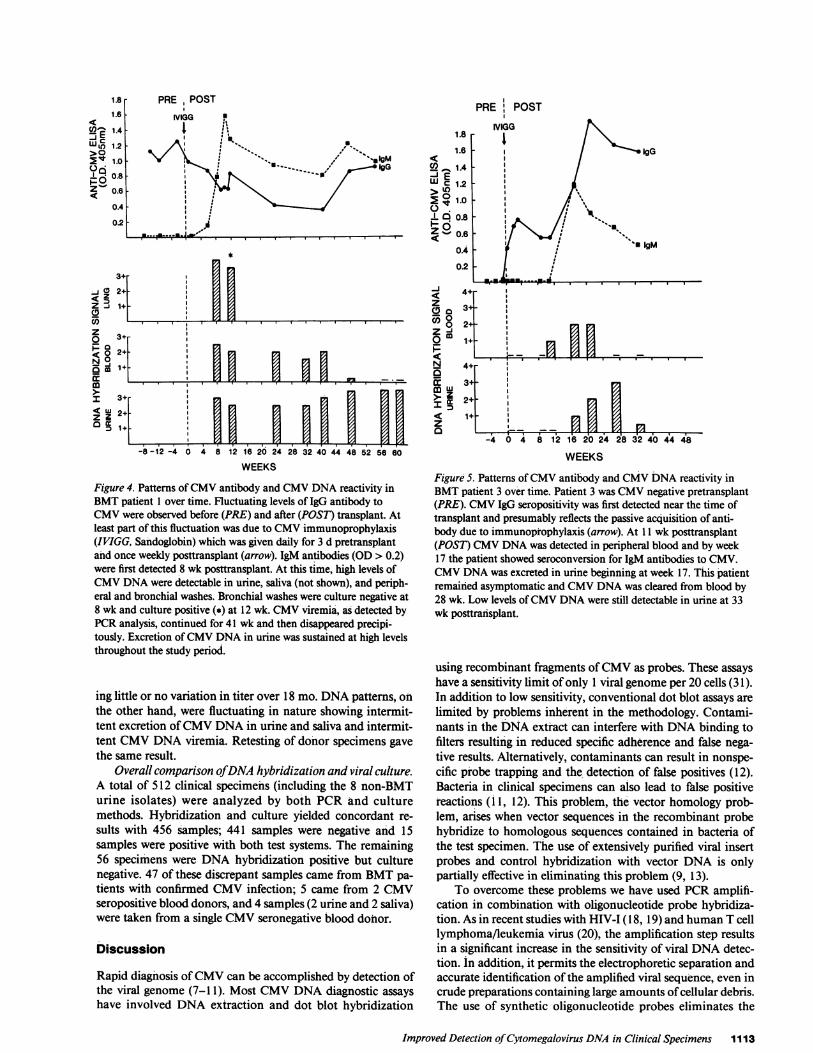
Figure 4. Patterns of CMVantibody and CMVDNAreactivity inBMTpatient 1 over time. Fluctuating levels of IgG antibody toCMVwere observed before (PRE) and after (POST) transplant. Atleast part of this fluctuation was due to CMVimmunoprophylaxis(IVIGG, Sandoglobin) which was given daily for 3 d pretransplantand once weekly posttransplant (arrow). IgM antibodies (OD > 0.2)were first detected 8 wk posttransplant. At this time, high levels ofCMVDNAwere detectable in urine, saliva (not shown), and periph-eral and bronchial washes. Bronchial washes were culture negative at8 wk and culture positive (*) at 12 wk. CMVviremia, as detected byPCRanalysis, continued for 41 wk and then disappeared precipi-tously. Excretion of CMVDNAin urine was sustained at high levelsthroughout the study period.
ing little or no variation in titer over 18 mo. DNApatterns, onthe other hand, were fluctuating in nature showing intermit-tent excretion of CMVDNAin urine and saliva and intermit-tent CMVDNAviremia. Retesting of donor specimens gavethe same result.
Overall comparison ofDNA hybridization and viral culture.A total of 512 clinical specimens (including the 8 non-BMTurine isolates) were analyzed by both PCRand culturemethods. Hybridization and culture yielded concordant re-sults with 456 samples; 441 samples were negative and 15samples were positive with both test systems. The remaining56 specimens were DNAhybridization positive but culturenegative. 47 of these discrepant samples came from BMTpa-tients with confirmed CMVinfection; 5 came from 2 CMVseropositive blood donors, and 4 samples (2 urine and 2 saliva)were taken from a single CMVseronegative blood donor.
Discussion
Rapid diagnosis of CMVcan be accomplished by detection ofthe viral genome (7-1 1). Most CMVDNAdiagnostic assayshave involved DNAextraction and dot blot hybridization
z
V50
I
CoLUza
WEEKS
FigureS5. Patterns of CMVantibody and CMVDNAreactivity inBMTpatient 3 over time. Patient 3 was CMVnegative pretransplant(PRE). CMVIgG seropositivity was first detected near the time oftransplant and presumably reflects the passive acquisition of anti-body due to immunoprophylaxis (arrow). At 11 wk posttransplant(POST) CMVDNAwas detected in peripheral blood and by week17 the patient showed seroconversion for IgM antibodies to CMV.CMVDNAwas excreted in urine beginning at week 17. This patientremained asymptomatic and CMVDNAwas cleared from blood by28 wk. Low levels of CMVDNAwere still detectable in urine at 33wk posttransplant.
using recombinant fragments of CMVas probes. These assayshave a sensitivity limit of only 1 viral genome per 20 cells (31).In addition to low sensitivity, conventional dot blot assays arelimited by problems inherent in the methodology. Contami-nants in the DNAextract can interfere with DNAbinding tofilters resulting in reduced specific adherence and false nega-tive results. Alternatively, contaminants can result in nonspe-cific probe trapping and the detection of false positives (12).Bacteria in clinical specimens can also lead to false positivereactions (i 1, 12). This problem, the vector homology prob-lem, arises when vector sequences in the recombinant probehybridize to homologous sequences contained in bacteria ofthe test specimen. The use of extensively purified viral insertprobes and control hybridization with vector DNA is onlypartially effective in eliminating this problem (9, 13).
To overcome these problems we have used PCRamplifi-cation in combination with oligonucleotide probe hybridiza-tion. As in recent studies with HIV-I (18, 19) and human T celllymphoma/leukemia virus (20), the amplification step resultsin a significant increase in the sensitivity of viral DNAdetec-tion. In addition, it permits the electrophoretic separation andaccurate identification of the amplified viral sequence, even incrude preparations containing large amounts of cellular debris.The use of synthetic oligonucleotide probes eliminates the
Improved Detection of Cytomegalovirus DNAin Clinical Specimens 1113
WE 1.4
t 1.2>0
t 1.0oci
6 0.8
Z 060.6
OA
0.2
3+
J 0 2+Ad ZZ J 1+0C)z 3+o= a
8 2+N0
1+
m<U 2
o 31
PRE POST
IVIGG1.8
1.6
U>_1A~I
> U1.0 1.0
~0in 0.8 -;
02_.
0.4
0.2flIII77Kinv nP nI.IB.I;E |B l
0|
-8-12 -4 0 4 8 12 16 20 24
WEEKS28 32 40 44 48 52 56 60

vector homology problem associated with recombinant DNAtechnology.
In preliminary testing we have shown that PCRcan repro-ducibily detect one copy of the CMVIE target sequence in40,000 cells. This corresponds to a 2,000-fold increase in sensi-tivity over unamplified DNAassays for the detection of CMVIE sequences (1 1, 31). The successful amplification of Davisand AD169 strains, as well as eight of eight wild-type strains ofCMV, indicates that the PCRassay recognizes most, if not all,CMVstrains. The ability to identify a wide range of strains wasexpected since amplification sequences were selected from ahighly invariant region of the CMVgenome. These sequencesare known to be identical in both Towne and AD169 strains(24, 25). The high specificity of the PCRassay was shown bythe complete lack of hybridization with human cellular DNA,E. coli DNA, or DNAfrom other herpes viruses.
The clinical applicability of the CMVPCR test has beenshown in a longitudinal surveillance study of CMVinfectionin BMTpatients. The PCRassay correctly identified four pa-tients with documented CMVinfection. Three of these pa-tients were followed longitudinally. In these patients, the ap-pearance of CMVDNApreceded clinical and/or serologicalevidence of CMVinfection. These results suggest that PCRis avaluable marker for the early stages of CMVinfection. Sincethis work was completed, Loche and Mach (32) have reportedsimilar results for HIV. Using PCR methods, they detectedHIV DNAin 5 of 16 high-risk seronegative sexual partners ofHIV seropositive persons. They concluded that PCRtechnol-ogy was capable of identifying infected individuals before thedevelopment of antibody. Clinical and serological follow-up ofthese patients, however, has not yet been completed.
The ability of PCRto identify CMVin the peripheral bloodof BMTpatients is particularly important since CMVviremiais frequently associated with serious disease (33). In this study,CMVDNAwas identified in peripheral blood of both patientswho developed IP. Although the levels of CMVin blood werelower than in lung and urine, we believe this reflects differ-ences in viral titer rather than a lack of sensitivity with bloodspecimens. This is based on reconstruction experiments andon studies in AIDS patients where high levels of CMVarefrequently detected in peripheral blood (unpublished results).The concentration of CMVin the PMNcell fraction suggeststhat blood is functioning primarily to clear CMVinfections.
Overall, CMVDNAwas identified in seven of seven cul-ture-positive BMTspecimens. In addition, CMVDNAwasidentified in 47 of 147 CMVculture-negative specimens. All ofthese specimens came from the four patients with confirmedinfection. Since we have probed with synthetic oligonucleo-tides (devoid of vector contamination), it is unlikely that theseresults are due to nonspecific probe binding to bacteria. Analternative explanation is that some specimens may containdefective virions that do not grow in culture. However, theearly detection of CMVDNAin patients who later becameculture positive suggests that PCR technology can identifyCMVwith higher sensitivity than viral culture. Similar resultshave been reported for HIV. Ou and colleagues (19), usingPCR methods, were able to demonstrate HIV in 11 of 11(100%) DNAsamples from HIV culture-positive, seropositivehomosexual men. They also detected HIV in 7 of 11 (64%)DNAsamples from high-risk seropositive, but culture-nega-tive homosexual men, suggesting that PCRwas the more sen-sitive technology.
Validation of our donor data is more difficult since donorinfections are usually asymptomatic and CMVcannot be cul-tured from donor units. Previous infection can be inferred bythe presence of IgG antibodies to CMV. However, it appearsthat only a small percentage of previously infected peopleenter a latent phase that remains infectious in a blood donor'sproduct. Schrier and colleagues (34), using in situ hybridiza-tion, were able to detect CMVRNAtranscripts in 0.03-2.0%of PBMCin eight of eight asymptomatic seropositive donors.Spector and associates (35), using a dot blot assay, detectedCMVin the peripheral blood of 3 of 41 seropositive donors.However, we and others (31, 36), using similar technology,have been repeatedly unable to identify CMVDNAin blooddonors. In the present study, using sensitive PCRmethods, wewere able to detect CMVin two of three seropositive donorsand in 1 of 13 seronegative donors. Patterns of CMVDNAreactivity in individual donors over time showed only inter-mittent CMVviremia and excretion. This is in contrast to theprotracted DNA patterns observed in the BMTpopulationand suggests a low level of viral activity that is only periodicallydetectable by hybridization. The detection of CMVDNAinonly 2 of 43 (4.7%) of peripheral blood specimens from sero-positive donors is consistent with what is known about CMVcarrier rates in the donor population. These rates have beenestimated to range between 1 and 13% (37).
There are several possibilities to explain the lack of CMVDNAdetection in two seropositive, culture-negative individ-uals (one donor and one BMTpatient). One possibility is thatthe PCRassay is not sufficiently sensitive to detect the verylow levels of virus present in these persons. This possibilitycould, in theory, be resolved by increasing the number of PCRamplification cycles. Alternatively, it could indicate that theseindividuals have been exposed to CMVin the past but thattheir infection has been successfully resolved. A third possibil-ity is that these individuals harbor strains of CMVthat are notdetectable by the present PCRassay or that they carry viralgenomes that are defective in the IE region. A fourth possibil-ity is that these individuals are temporarily in an inactive (la-tent) phase and that continued monitoring will eventually re-sult in the detection of positives. The detection of CMVDNAin one CMVseronegative donor is more difficult to explain.We are currently sequencing this amplification product toconfirm its identity.
To summarize, our findings suggest that PCRamplifica-tion technology is more sensitive than conventional culturemethods for the diagnosis of CMV. One of the major advan-tages of the technology is its applicability to a wide variety ofbiological specimens. In longitudinal surveillance studies wehave shown that DNAtesting of multiple tissues can provideadditional information on the overall CMVstatus of BMTpatients. It is anticipated that ongoing serial study will eventu-ally lead to the identification of CMVreactivity patterns thatare of diagnostic importance. The potential of the PCRassayto detect early CMV, especially CMVviremia, has importantimplications since early diagnosis may allow physicians tomodify antiviral or immunosuppressive regimens in time toalter the course of serious CMVdisease.
AcknowledgmentsThe authors would like to thank Dr. K. Buchan and Mr. T. Stowe ofthe clinical virology laboratory, Foothills Hospital, for providing viralculture studies.
1114 Cassol et al.

These studies were supported by the Canadian Red Cross, BloodTransfusion Service, grants CA-02-84 and CA-02-87.
References
1. Hirsch, M. S. 1981. Herpes group virus infections in the com-promised host. In Clinical Approach to Infection in the CompromisedHost. R. H. Rubin and L. S. Young, editors. Plenum Medical BookCo., NewYork. 389-415.
2. Drew, W. L., E. S. Mocarski, E. Sweet, and R. C. Miner. 1984.Multiple infections with CMVin AIDS patients: documentation bySouthern blot hybridization. J. Infect. Dis. 150:954-960.
3. Meyers, J. D., N. Fluornoy, and E. D. Thomas. 1982. Nonbac-terial pneumonia after allogeneic marrow transplantation: a review of10 years' experience. Rev. Infect. Dis. 4:1119-1132.
4. Rasmussen, L., D. Kelsall, and R. Nelson. 1982. Virus-specificIgG and IgM antibodies in normal and immunocompromised subjectsinfected with cytomegalovirus. J. Infect. Dis. 145:191-199.
5. Starr, S. E., and H. M. Friedman. 1980. Humancytomegalovi-rus. In E. H. Lenette, A. Balows, W. J. Hausler, and J. P. Truant,editors. Manual of Clinical Microbiology. 3rd ed. American Society forMicrobiology, Washington, DC. 790-797.
6. Emanuel, D., J. Peppard, D. Stover, J. Gold, D. Armstrong, andU. Hammerling. 1986. Rapid immunodiagnosis of cytomegaloviruspneumonia by bronchoalveolar lavage using human and murinemonoclonal antibodies. Ann. Intern. Med. 104:476-481.
7. Chou, S., and T. C. Merigan. 1983. Rapid detection and quanti-tation of human cytomegalovirus in urine through DNAhybridiza-tion. N. Engl. J. Med. 308:921-925.
8. Spector, S. A., and D. H. Spector. 1985. The use of DNAprobesin studies of human cytomegalovirus. Clin. Chem. 31:1514-1520.
9. Kahan, B. D., and T. A. Landers. 1985. Rapid detection ofcytomegalovirus infection using a DNAprobe. Transplant. Proc.17:989-993.
10. Virtanen, M., A.-C. Syvanen, J. Oram, H. Soderlund, and M.Ranki. 1984. Cytomegalovirus in urine: detection of viral DNAbysandwich hybridization. J. Clin. Microbiol. 20:1083-1088.
11. Augustin, S., T. Popow-Kraupp, F. X. Heinz, and C. Kunz.1987. Problems in detection of cytomegalovirus in urine samples bydot blot hybridization. J. Clin. Microbiol. 25:1973-1977.
12. Ambinder, R. F., P. Charache, S. Staal, P. Wright, M. Forman,S. D. Hayward, and G. S. Hayward. 1986. The vector homology prob-lem in diagnostic nucleic acid hybridization of clinical specimens. J.Clin. Microbiol. 24:15-20.
13. Schister, V., B. Matz, H. Wiegand, B. Traub, D. Kampa, and D.Neumann-Hafelin. 1986. Detection of human cytomegalovirus inurine specimens by DNA-DNAand RNA-DNAhybridization. J. In-
fect. Dis. 154:309-314.14. Saiki, R. K., S. Shark, F. Faloona, K. B. Mullis, G. T. Horn,
H. A. Erlich, and N. Arnheim. 1985. Enzymatic amplification of beta-globin genomic sequences and restriction site analysis for diagnosis ofsickle cell anemia. Science (Wash. DC). 230:1350-1354.
15. Embury, S. H., S. J. Scharf, R. K. Saiki, M. A. Gholson, M.Golbus, N. Arnheim, and H. A. Erlich. 1987. Rapid prenatal diagnosisof sickle cell anemia by a new method of DNAanalysis. N. Engl. J.Med. 316:656-661.
16. Kogan, S. C., M. Doherty, and J. Gitschier. 1987. An improvedmethod for prenatal diagnosis of genetic diseases by analysis of ampli-fied DNAsequences. N. Engl. J. Med. 317:985-990.
17. Shibata, D. K., N. Arnheim, and W. J. Martin. 1988. Detectionof human papilloma virus in paraffin-embedded tissue using the poly-merase chain reaction. J. Exp. Med. 167:225-230.
18. Kwok, S., D. H. Mack, K. B. Mullis, B. Poiesz, G. Ehrlich, D.Blair, A. Friedman-Kien, and J. J. Sninsky. 1987. Identification of
human immunodeficiency virus sequences by using in vitro enzymaticamplification and oligomer cleavage detection. J. Virol. 61:1690-1694.
19. Ou, C.-Y., S. Kwok, S. W. Mitchell, D. H. Mack, J. J. Sninsky,J. W. Krebs, P. Feorino, D. Warfield, and G. Schochetman. 1988.DNAamplification for direct detection of HIV- I in DNAof peripheralblood mononuclear cells. Science (Wash. DC). 239:295-297.
20. Duggan, D. B., G. D. Ehrlich, F. P. Davey, S. Kwok, J. Sninsky,J. Goldberg, L. Baltrucki, and B. J. Poiesz. 1988. HTLV-1 inducedlymphoma mimicking Hodgkin's disease, diagnosis by polymerasechain reaction amplification of specific HTLV-1 sequences in tumorDNA. Blood. 71:1027-1032.
21. Fleckenstein, B., I. Muller, and J. Collins. 1982. Cloning thecomplete human cytomegalovirus genome in cosmids. Gene. 38:39-46.
22. Hoar, D. I., D. B. Haslam, and D. M. Starozik. 1984. Improveddirect molecular diagnosis and rapid fetal sexing. Prenatal Diagn.2:241-247.
23. Hoar, D. I., T. Bowen, D. Matheson, and M. C. Poon. 1985.Hepatitis B virus DNAis enriched in polymorphonuclear leukocytes.Blood. 66:1251-1253.
24. Akrigg, A., G. W. G. Wilkinson, and J. D. Oram. 1985. Thestructure of the major immediate early gene of human cytomegalovi-rus strain AD169. Virus Res. 2:107-121.
25. Pritchett, R. F. 1980. DNAnucleotide sequence heterogeneitybetween the Towne and AD169 strains of cytomegalovirus. J. Virol.36:152-161.
26. Rudiger, R., G. W. Bornkamm, and B. Fleckenstein. 1984.Humancytomegalovirus DNAsequences with homologies to the cel-lular genome. J. Gen. Virol. 65:1351-1364.
27. Peden, K., P. Mounts, and G. S. Hayward. 1982. Homologybetween mammalian cell DNAsequences and human herpesvirus ge-nomes detected by a hybridization procedure with high complexityprobe. Cell. 31:71-80.
28. Beaucage, S. L., and M. H. Caruthers. 1981. Deoxynucleosidephosphoramidites: a new class of key intermediates for deoxypolynu-cleotide synthesis. Tetrahedron Lett. 22:1859-1862.
29. Southern, E. M. 1975. Detection of specific sequences amongDNA fragments separated by gel electrophoresis. J. Mol. Biol.98:503-517.
30. Maniatis, T., Fritsch, E. F., and J. Sambrook. 1982. MolecularCloning Manual. Cold Spring Harbor Laboratory, Cold Spring Har-bor, NewYork. 122-123.
31. Jackson, J. B., H. T. Orr, J. J. McCullough, and M. C. Jordan.1987. Failure to detect human cytomegalovirus DNAin IgM-seropos-itive blood donors by spot hybridization. J. Infect. Dis. 156:1013-1016.
32. Loche, M., and B. Mach. 1988. Identification of HIV-infectedseronegative individuals by a direct diagnostic test based on hybridisa-tion to amplified viral DNA. Lancet. ii:418-421.
33. Saltzman, R. L., M. R. Quirk, and M. C. Jordan. 1988. Dissem-inated cytomegalovirus infection. Molecular analysis of virus and leu-kocyte interaction in viremia. J. Clin. Invest. 81:75-81.
34. Schrier, R. D., J. A. Nelson, and M. B. A. Oldstone. 1985.Detection of human cytomegalovirus in peripheral blood lymphocytesin a natural infection. Science (Wash. DC). 230:1048-1051.
35. Spector, S. A., J. A. Rua, D. H. Spector, and R. McMillan.1984. Detection of human cytomegalovirus in clinical specimens byDNA-DNAhybridization. J. Infect. Dis. 150:121-126.
36. Cassol, S., D. Hoar, E. Wong, K. Buchan, and C. Snider. 1985.Detection of cytomegalovirus DNAin clinical specimens using DNAdot hybridization. Blood. 66:258a. (Abstr.)
37. Tegtmeier, G. E. 1985. Cytomegalovirus and blood transfusion.In Infection, Immunity and Blood Transfusion. Alan R. Liss, Inc.,NewYork. 175-199.
Improved Detection of Cytomegalovirus DNA in Clinical Specimens 1115




![BCL9PromotesTumorProgressionbyConferringEnhanced ...Res2009;69(19):7577–86] Introduction The Wnt pathway consists of a highly conserved and tightly regulated receptor-mediated signal](https://img.pdfslide.fr/doc/110x75/612571378eb4a3086b4f647d/bcl9promotestumorprogressionbyconferringenhanced-res200969197577a86-introduction.jpg)
























