Embed Size (px)
Citation preview

Insectes Sociaux, Paris.
1978. T o m e 25, n ~ 1, p p . 13-30.
�9 Masson, Paris, 1978.
PRODUCTIVITI~ DE LA FOURMI F O R M I C A D A K O T E N S I S
DANS LA PESSIC'-RE TOURBEUSE.
2. VARIATIONS ANNUELLES DE LA DENSITI~ DES COLONIES,
DE L'OCCUPATION DES NIDS
ET DE LA RI~PARTITION SPATIALE
P a r A. F R A N C ( E U R et D. P I P P I N (*)
Universitd da Qadbec, Chicoatimi, Qndbec, Canada G7H 2B1.
Re~u le 20 j u i l l e t 1976. Acceptd le 9 oc tohre 1976.
R~SUME
Apr6s q u a t r e a n s de p r o d u c t i o n , la dens i t6 de Formica dakolensis aff lche u n e m o y e n n e de 172,2 co lon ies p a r hec t a re , d a n s la pess i~re t o u r h e u s e de S a i n t - A m b r o i s e , Qudbec. Avec u n coeff ic ient de v a r i a t i o n de 2 % s e u l e m e n t , la p o p u l a t i o n des co lon ies a p p a r a i t en dqu i l i h r e avee le m i l i eu . Une te l le s t ab i l i td cache t o u t e f o i s des f l u c t u a t i o n s i n t e r n e s qui se t r a d u i s e n t p a r u n t a u x effeet i f de r e n o u v e l l e m e n t a n n u e l des co lon ies de 18,7 %, h la fin d ' u n e s a i s o n de e ro i s sance , a lo r s que le t a u x spdc i f ique de r e n o u v e l l e - m e n t a n n u e l de la p o p u l a t i o n de co lon ie s p e n d a n t le t e m p s d ' o b s e r v a t i o n s 'dl~ve h 23,6 %. Cet te de rn i6 re v a l e u r es t c o m p a r d e h eel les o b t e n u e s p o u r d ' a u t r e s esp~ces de Formica. Le t a u x m o y e n de n a i s s a n c e des co lon ies s 'dl~ve /t 22,9 % et ce lui de m o r t a l i t d h 23,5 %. La longdvi td po t en t i e l l e d ' u n e co lonic es t e s t imde h p a r t i r des d o n n d e s s u r l ' o c e u p a t i o n c o n t i n u e des n i d s r ecensds en 1972, La r d o c e u p a t i o n de n i d s a h a n d o n n d s se rdv61e d ' a u t a n t p l u s g r a n d e que ceux-e i son t j e u n e s , c ' e s t -h -d i r e en h o n dta t . D ' a u t r e s esp6ces de f o u r m i u t i l i s e n t p a r f o l s les n i d s c o n s t r u i t s et a b a n d o n n d s p a r F. dakotensis. I1 ex is te u n lot p e r m a n e n t de n i d s d d f i n i t i v e m e n t a b a n d o n n d s qu i e n t r e n t a lo r s en d d c o m p o s i t l o n , p u i s d i s p a r a i s s e n t . La r d p a r t i t i o n spa t i a l e d e m e u r e a n h a s a r d avec u n e f a ib l e t e n d a n c e ve r s le type c o n t a g i e u x . Les co lon ies se c o n c e n t r e n t d a n s u n h a b i t a t p a r t i c u l i e r du b io tope . L ' d v o l u t i o n de ce d e r n i e r in f luence l ' a v e n i r de la p o p u l a t i o n de co lon ies et sa dens i td . Les d i m e n s i o n s des n o u v e a u x n i d s s o n t a n a l y s d e s en f o n c t i o n de l e u r occupa t ion . Le c o m p o r t e m e n t de c o n s t r u c t i o n de cette f o u r m i i l l n s t r e r a i t u n e d tape in i t i a l e d a n s 1 ' appa r i t i on du po lyca l i s rne chez le gen re Formica.
(*) Adres se ac tue l l e : L a b o r a t o i r e de la F a u n e S a u v a g e et de C y n d g d t i q u e , I.N.R.A., F 78350 J o u y - e n - J o s a s .

14 A. FRANCO?,UR E T D. P E P 1 N
S U M M A R Y
Productivity of the ant Formica dakotensis in a spruce bog.
2. Annual variations in colony density, nest inhabitation and spatial distribution.
After 4 years of product ion the mean densi ty of Formica dakotens is is 172.2 colonies per hectare in the spruce bog of Saint-Ambroise, Quebec. A v a r i a t i o n index of only 2 % means t h a t the colony popula t ion has reached s tab i l i ty w i t h i n the hab i ta t . However at the end of a growing season an effective ra te of annua l r ep lacement of 18.7 % affects the colonies whi le the specific annua l rep lacement ra te of the colony popula t ion reached 23.6 % dur ing the period of census. This las t value is compared to those o b t a i n e d for o ther species of Formica. Mean b i r t h ra te of colonies is 22.9 % and mean mor t a l i t y is 23.5 %. The potent ia l longevity of a colony is e s t imated f rom data on con t inuous ly inhab i t ed nests f rom 1972 to 1975. The percentage of deserted nests t ha t are used aga in is h igher among younger nests, t h a t is those sti l l well preserved. Somet imes o ther an t species use the nests bu i l t and deserted by F. dako- tensis. There is a n u m b e r of mounds definit ively ahandoned which then are progessively decomposed and e l imina ted f rom the surface of the organic soil by the l iv ing mosses and Ericaeeae. Spat ia l d i s t r ibu t ion is basical ly r a n d o m wi th a weak tendency towards aggregation and colonies favour a character is t ic par t of the hab i ta t . The evolu t ion of the hab i t a t towards a m a t u r e tree s tand influences the fu tu re and densi ty of the censused populat ion. New mound size is s tudied in re la t ion to occupation. The nes t -hu i ld ing behav io r of th i s an t shows a possible ini t ial step leading to the appearance of poly- calism in the genus Formica.
I N T R O D U C T I O N
L a d e n s i t 6 des c o l o n i e s de F o r m i c a d a k o t e n s i s d a n s c i n q p e s s i ~ r e s t o u r b e u s e s
de la r 6 g i o n d u S a g u e n a y a 6t6 d 6 t e r m i n 6 e d a n s u n p r e m i e r t e m p s (FRaNr et
P~PIN, 1975). A c a u s e des g r a n d e s v a r i a t i o n s o b s e r v 6 e s d a n s l ' a b o n d a n c e de ce t t e
f o u r m i en f o n c t i o n de l '~ge des b i o t o p e s r e c e n c 6 s , il est a p p a r u n ~ c e s s a i r e de
s u i v r e u n e p o p u l a t i o n p a r t i c u l i 6 r e p e n d a n t q u e l q u e s a n n 6 e s , clans le b u t de
c o n n a l t r e s o n 6 v o l u t i o n u l t 6 r i e u r e et s on n i v e a u d ' 6 q u i l i b r e a v e c le b i o t o p e
occup6 .
N o t r e c h o i x s ' e s t p o r t 6 s u r u n e p o p u l a t i o n a f f i c h a n t u n e f o r t e d e n s i t 6 , h
s a v o i r ce l le de la p e s s i b r e t o u r b c u s e de S a i n t - A m b r o i s e , corot6 de C h i c o u t i m i
(48o32 , N, 71018 , O). D a n s u n e te l le s i t u a t i o n , on p e u t s ' a t t e n d r e 'h o b t e n i r s i m u l -
t a n 6 m e n t des d o n n 6 e s i n t 6 r e s s a n t e s s u r les v a r i a t i o n s a n n u e l l e s de la d e n s i t 6
des c o l o n i e s , s u r les c h a n g e m e n t s d a n s l ' o c c u p a t i o n des n i d s , s u r le c o m p o r t e m c n t
de c o n s t r u c t i o n et s u r la r 6 p a r t i t i o n s p a t i a l e des c o l o n i e s . R a p p e l o n s i c i q u e
les n i d s se c o m p o s e n t cn s u r f a c e de d 6 b r i s v6g6 taux , en p a r t i c u l i e r de f eu i l l e s
m o r t e s d ' E r i c a c 6 e s , ce qu i p c r m e t de les r e p 6 r e r f a c i l e m e n t .

PRODUCTIV1Tff? DE LA FOURMI F O R M I C A D A K O T E N S I S 15
METHODES D'ETUDE
Les d o n n 6 e s f u r e n t r eeue i l l i e s h la fin de la s a i s o n de c r o i s s a n c e des co lon ies , e ' e s t -h -d i r e fin a o f i t - d 6 b u t s e p t e m b r e , de 1972 5. 1975. El les p r o v i e n n e n t d ' u n e s u r f a c e car r6e d ' u n h e c t a r e d41imit6e p a r des p i q n e t s et des cordes . Cet te super f i c ie e s t e l l e - m ~ m e
0 oe }
ffT~ Milieu
i ' ~ Milieu
[ ~ Milieu
Fig,. 1. - - T y p e s de m i l i e u d a n s l ' h ee t a r e reeens6 .
Fla. 1. - - H a b i t a t t y p e s in the e e n s u s e d hec ta re .
ouvert: sans epinettes noires
intermediaire=epinettes noires eparses
ferme= epinettes noires en touffes
subd iv i s6e en 16 s e c t e u r s 6gaux de 25 m de e6t6 c h a c u n (fig. 1), d a n s le b u t de f ac i l i t e r la r eche rche s y s t 6 m a t i q u e des n i d s s u r le t e r r a i n , l e a r l o c a l i s a t i o n s u r u n p l a n q u a d r i l l ~ e t l ' a n a l y s e de l ' o e e u p a t i o n des d i f f6 ren t s h a b i t a t s ident i f i6s .
Une d e s c r i p t i o n g6n6ra le de la v6g6 t a t i on du b io tope es t d o n n d e d e n s un a r t i c l e p r6c6dent (FRaNcceva et P~.PIN, 1975). L ' h e c t a r e i nven to r i 6 rec61e t r o i s t y p e s d i f f6 ren t s

16 A . F R A N C ( E U R E T D . P E P I N
d ' h a b i t a t d o n t les l i m i t e s g6ndra les a p p a r a i s s e n t s u r la f igure 1. Ce son t , en f o n c t i o n de l '~ge : a) u n m i l i e u ouve r t , e o n s t i t u 6 e s s e n t J e l l e m e n t de S p h a i g n e s et d ' E r i c a c 6 e s ; l)) u n m i l i e u i n t e r m ~ d i a i r e , carac t~r i s~ p a r la p r6 sence e n v a h i s s a u t e de t iges gdn6 ra l e - mer i t i sol6es d ' E p i n e t t e no i r e ( P i c e a m a r i a n a ) ; c) u n m i l i e u fe rmd, d o m i n 6 p a r des touf fes p l u s ou m o i n s d e n s e s d ' E p i n e t t e no i re , h l ' i n t ~ r i e u r d e s q u e l l e s les Er icac~es s o n t s o u v e n t a b s e n t e s . Le d e u x i ~ m e t ype oceupe la p l u s g r a n d e superf ic ie .
La p r e m i 6 r e a n n 6 e de l '6 tude , t o u s l e s n i d s de F . d a k o t e n s i s e x i s t a n t h l ' i n t 6 r i e u r du p 6 r i m ~ t r e de l ' h e e t a r e s o n t iden t i f ids p a r u n p i q u e t n u m 6 r o t 6 . P o u r c h a q u e h id d~couver t , on 6 t ab l i t u n e fiche i n d i q u a n t le n u m S r o d ' i den t i f i ca t ion , les e o o r d o n n 6 e s p o u r la c a r t o g r a p h i e , la p o s i t i o n t o p o g r a p h i q u e ( su r t e r t r e de s p h a i g n e , d u n s d 6 p r e s s i o n , etc.), l ' o r i e n t a t i o n , les d i m e n s i o n s a p p a r e n t e s ( l o n g u e u r , l a r g e u r , h a u t e u r ) , la n a t u r e des d6br i s v6g6 taux . On v6rifle e n s u i t e si le n i d es t oeeupd. D u n s l ' a f f i r m a t i v e , d e u x ou t ro i s i n d i v i d u s s o n t pr61ev6s et d6pos6s avee le n u m S r o du h id d u n s u n e pe t i t e b o u t e i l l e e o n t e n a n t de l ' a leool , af in de les i den t i f i e r au l a b o r a t o i r e . Le n o m de l ' espbce p o u r r a a lo r s 8tre i n sc r i t s u r l a fiche.
Les a r m i e s s u b s ~ q u e n t e s , on v i s i t e d ' a b o r d les n i d s d6jh r e p o r t s et ident i f i~s p o u r vdrif ier s ' i l s s o u l t o u j o u r s h a b i t , s p a r ]a m 6 m e esp~ee ou s ' i l s on t 6t6 a b a n d o n n 6 s . D u n s le eas de n i d s p r 6 c 6 d e m m e n t i noeeup6s qu i s o n t h n o u v e a u u t i l i s~s p a r les f o u r m i s , on pr61~ve d e u x i n d i v i d u s p o u r i den t i f i ca t ion . La fagon de t r a i l e r les n id s dt~couverts la p r e m i d r e a n n i e s ' a p p l i q u e a u x n o u v e a u x n ids .
L ' u t i l i s a t i o n de la m ~ t h o d e de CLARK et EVANS (1954), bustle s u r la d i s t a n c e m o y e n n c e x i s t a n t e n t r e les p l u s p roehes vo i s ins , p e r m e t de d 6 t e r m i n e r le t ype de r~pa r t i t i on s p a t i a l e des eoIonies .
R I ~ ' S U L T A T S E T D I S C U S S I O N
L e r c c e n s e m e n t g 6 n 6 r a l d e s n i d s c o n s t r u i t s p a r F . d a k o t e n s i s a p p a r a i t a u
t a b l e a u I a v e c l a l i s t e d e s e s p ~ c e s q u i l e s o c c u p a i e n t a u c o u r s d e 1972 .it 1975. L e
t a b l e a u I I c o n s t r u e u n e r e p r 6 s e n t a t i o n d e s d o n n 6 e s s u r l ' u s a g e d e s n i d s a n c i e n s
et n o u v e a u x d u r a n t c e t t e p 6 r i o d e . T o u t e s l e s p o s s i b i l i t 6 s d e c h a n g e m e n t d u n s
l ' o c c u p a t i o n d e s n i d s s o n t c o n s i g n 6 e s , s a n s 6 t r e o b l i g a t o i r e m c n t r e p r d s e n t 6 c s
( v o i r e n p a r t i c u l i e r l e s d o n n 6 e s r e e u e i l l i e s e n 1975 ) . L e s n i d s c o m p t a n t p l u s
TABLEAU I. - - R e e e n s e m e n t a n n u e l des n i d s c o n s t r u i t s p a r F o r m i c a d a k o l e n s i s d u n s u n h e c t a r e de pes s ib re t o u r b e u s e , h S a i n t - A m b r o i s e , et i den t i f i ca t ion des o e v u p a n t s .
TABLE I. - - A n n u a l c e n s u s of the n e s t s b u i l t by F o r m i c a d a k o l e n s i s in one h e c t a r e o f s p r u c e bog, a t S a i n t - A m b r o i s e , a n d l i s t of t he i n h a b i t i n g an t s .
Ann6e 1972 1973 197.1 1975
Nids oceup6s . . . . . . . . . . . . . . . . . . . 183 Nids a b a n d o n n ~ s . . . . . . . . . . . . . . . . 71 Nids reeens~s . . . . . . . . . . . . . . . . . . . 254
Nids oeenp~s p a r : F o r m i c a d a k o t e n s i s . . . . . . . . . . . . . 172 F. n e o r u f i b a r b i s . . . . . . . . . . . . . . . . . 9 L e p t o l h o r a x m u s c o r u m . . . . . . . . . . 1 F . s u b n u d a . . . . . . . . . . . . . . . . . . . . . 1 F . m h q m p e r i . . . . . . . . . . . . . . . . . . . . 0 C a m p o n o l u s h e r c u l e a n u s . . . . . . . . 0 M . q r m i c a d i s e o n t i n a a . . . . . . . . . . . . 0
178 184 192 117 138 155 295 322 347
171 177 1[;9 7 5 19 0 1 _'2 0 0 0 0 1 0 0 0 1 0 0 l

P R O D U C T I V I T E DE L A FOURM1 F O R M I C A D A K O T E N S I S 17
TABLEAU I I . - Sta t is t iques annuel les sur l 'usage des aneiens et des nouveaux nids pa r F. dakotensis de 1972 h 1975.
TABLE II. - - Annua l s tat is t ics On the use of old and new nests by F. dakotensis f rom 1972 to 1975.
N I D S F L 1 9 7 2 I f 1 9 7 3 f I 1 9 7 4 f
R E C E N S E S [_ ~ 17=1,~1 17~17317,]
o c c u .p
E S
(+)
( - )
+ : 1 3 4
Lz(+) 1 7 2 1 L~(+) 1 7 1
8 2 1 I:~ (-) 12 4 [ i ! 7 4 3 8
1 2
+ + + 1 1 2 - 4- 4- " 7
+ -- + 5
o + + 1 9 o - + 5
o o + 2 6 5 - ( + ) 1 7 7
[Z (-)
o = i n e x i s t a n t
E(-)
i 0 + -
io o
I I 1 9 7 5
1,21,~f ,,t ,51
0 - - +
iO "1" §
! ~ ' I ~ I 2
o -; ~_~j_ o 9
~:(+) 1 6
1 4 5 !z(- ) 1 7 8 7 1 . . . . 6 9 3 3 * - - - 3 1 2 2 § - - 2 2
1 + + § 1 5 7 - + - - 1
1 0 + - + - 0 1 - - + - 0
- * * - 3
o + + - 5
0 + - - 9
o - - - 7
o - + - 0
, o o - - 1
~ o + - 6
o o 9
d ' u n e s a i s o n de c r o i s s a n c e s e r o n t qua l i f i6s d 'anc iens et c e u x c r66s l ' a n n 6 e d ' u n
r e c e n s e m e n t , de nouveaux .
C h a q u e n i d o c c u p 6 r e p r 6 s e n t e n o r m a l e m e n t u n e c o l o n i c a v e c u n e seu le
r e i n e d ' a p r b s les i n v e n t a i r e s e f fec tu6s p o u r e s t i m e r le n o m b r e d ' i n d i v i d u s p a r
c o l o n i c et l e u r b i o m a s s e (ces r ~ s u l t a t s s e r o n t d o n n 6 s dar ts le t r o i s i ~ m e a r t i c l e
de ce t t e s6r ie ) . Des t e s t s p r 6 l i m i n a i r e s d ' h o s t i l i t 6 se s o n t r6v616s p o s i t i f s & 80 % .
I1 s e m b l e que ce t t e f o u r m i ne so i t p a s v 6 r i t a b l e m e n t p o l y c a l i q u e , b i e n que le
c o m p o r t e m e n t de c o n s t r u c t i o n l a i s se e n t r e v o i r p a r f o i s u n d 6 b u t de p o l y c a l i s m e
p r i m i t i f su r l e q u e l n o u s r e v i e n d r o n s p l u s lo in . L ' a n a l y s e q u i s u i t f a i t u n e d is-
t i n c t i o n e n t r e la p o p u l a t i o n des c o l o n i e s et la p o p u l a t i o n des n i d s .
I N S E C T E S S O C I A U X , 1978, T . 2 5 , N ~ 1 . 2
18 A. F R A N C ( E U R E T D. PIs
V a r i a t i o n s d e l a d e n s i t 4 d e s c o l o n i e s .
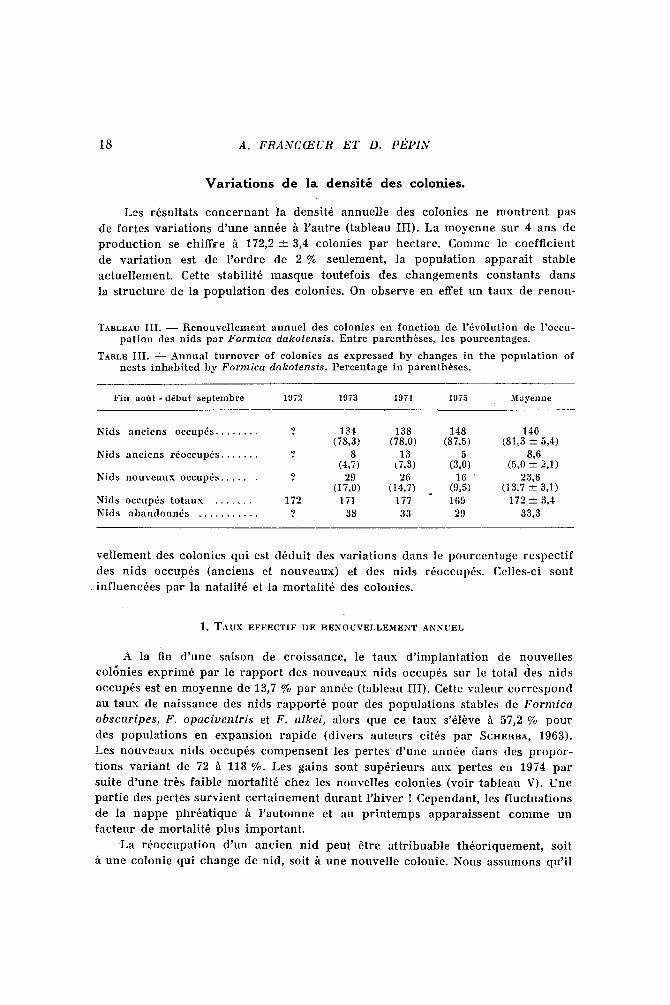
Les r6sultats conce rnan t la densit6 annuel le des colonies ne m o n t r e n t pas
de fortes var ia t ions d 'une ann6e /t l ' au t re (tableau III). La moyenne sur 4 ans de
p roduc t ion se chiffre h 1 7 2 , 2 _ 3,4 colonies par hectare. Comme le coeff ic ient
de var ia t ion est de l ' o rd re de 2 % seulement, la popula t ion appara i t stable
actuel lement . Cette stabilit6 masque toutefois des changements constants dans
la s t ruc ture de la popula t ion des colonies. On observe en effet un taux de renou-
TABLEAU III. - - Renouvellement annuel des colonies en fonction de l'dvolution de l'occu- pation des nids par Formica dakolensis. Entre parenth6ses, les pourcentages.
TABLE I I I . - - Annual turnover of colonies as expressed by changes in the population of nests inhabited by Formica dakotensis. Percentage in parenthbses.
F i n a o f t t - d6but s e p t e m b r e 1972 1973 1974 1975 Moyenne
Nids aneiens occnp6s . . . . . . . . ? 13'1~ 138 148 140 (78,3) (78,0) (87,5) (81~3 ---+ 5,4)
Nids anciens r6occup~s . . . . . . . ? 8 13 5 8,6 (4,7) (7,3) (3,0) (5,0 ~ 2,1)
Nids nouveaux oecnp~s . . . . . . ? 29 26 16 23,6 (17,0) (14,7) (9,5) (13,7 --+ 3,1)
Nids occuptis totaux . . . . . . . 172 171 177 169 172 _ 3,4 Nids abandonn~s . . . . . . . . . . . ? 38 33 29 33,3
vel lement des colonies qui est d6duit des var ia t ions dans le pourcentage respec t i f
des nids occup6s (anciens et nouveaux) et des nids r6occup6s. Celles-ci sont
inf luenc6es par la natalit6 et la mortal i t6 des colonies.
1. TAUX E F F E C T I F DE RENOUVELLEMENT ANNUEL
A la fin d 'une saison de croissance, le taux d ' implanta t ion de nouvel les
col(~nies expr im6 par le r appor t des nouveaux nids occup6s sur le total des nids
occup6s est en moyenne de 13,7 % par ann6e (tableau III). Cette va leur co r r e spond
au taux de naissance des nids rappor t6 pour des popula t ions stables de Formica obscuripes, F. opaciventris et F. ulkei, alors que ce taux s'61~ve h 57,2 % pour
des popula t ions en expansion rap ide (divers auteurs cit6s par SCHERBA, 1963).
Les nouveaux nids occup6s compensent les per tes d 'une ann6e dans des p ropor -
t ions var ian t de 72 h 118 %. Les gains sont sup6r ieurs aux per tes en 1974 par
suite d 'une trbs faible mortal i t6 chez les nouvelles colonies (voir tableau V). Une
par t ie des pertes surv ient ce r t a inement durant l ' h ive r ! Cependant , les f luc tuat ions
de la nappe phr6at ique h l ' au tomne et au p r in t emps appara issent comme un fac teur de mortal i t6 plus impor tant .
La r6occupat ion d 'un ancien nid peut 6ire a t t r ibuable th6or iquement , soit
'h une colonic qui change de nid, soit h une nouvel le colonic. Nous assumons qu ' i l

PRODUCTIVITE DE LA FOURMI FORMICA DAKOTENSIS 19
s'agit de nouvelles colonies d 'apr~s cer ta ines 6vidences expliqu6es plus loin. Le taux moyen de r6cup6rat ion est de 5 % sur trois ans (tableau III).
Le renouvel lement de la popula t ion de colonies est donc 6gal au taux d ' im- p lanta t ion des nouvelles colonies plus le taux de r6occupat ion des anciens nids. Selon les valeurs du tableau III, le renouvel lement annuel varie de 12,5 h 22,0 % et la moyenne s'61~ve h 18,7 • 5,4 %. Au cours des trois ann,~es d 'observat ion, Ie renouvel lement total s'61~ve donc h 56 ,1%.
Ce pourcentage peut 6galement 6tre calcul6 selon les formules d6crites par PETRUSEWICZ et MACFADYEN (1970), en assumant qu 'une colonie 6quivaut h u n individu. Par combinaison, on obt ient le taux de renouvel lement total :
O~ - - _
NT
off Y est le nombre total de colonies ayant exist6 h n ' impor te quel moment duran t la p6riode T d 'observat ion et N le nombre moyen de colonies pe nda n t ce temps. D'ofi :
0N _ 291 X 100 -~ 56,3% 172,2 • 3
Le taux de renouvel lement total des colonies 07 mult ipl i6 par N donne le hombre total de nouvelles colonies qui furent r6ellement pr6sentes dans la popu- lat ion durant la p6riode d 'observat ion, h savoir 97. E n fait, un total de 119 colonies nouvelles furent recens6es, mais 22 d 'entre elles sont apparues et dis- parues durant la m6me saison de croissance. C'est pourquoi u s e chiffre "h 291, soit 172 colonies relev6es lors du p remier recensement plus les 119 colonies nouvelles recens6es par la suite.
2. TAUX SPI~CIFIQUE DE RENOUVELLEMENT ANNUEL
Tel que present6 ci-dessus, le renouvel lement annuel ind ique le pourcentage de nouvelles colonies int6gr6es, h la fin d 'une saison de croissance, "h une populat ion de colonies survivantes . I1 ne rend pas compte de celles qui appa- raissent et meurent durant cette p6riode. Pour pouvoir comi~arer diff~rentes populat ions en consid6rant toutes les colonies effectivement produi tes (ici r6v616es par les nids), nous uti l isons le taux de renouvel lement par unit6 de temps d~fini par les derniers auteurs cit6s et que nous qualifions de sp6cifique :
0s ~ __1 e t ' t - NT t v E
od t -es t la long6vit~ ~cologique moyenne et "r le nombre de colonies 61imin~es, pendan t le temps T. D'oh :
0s _ v~, = 122 • 100 = 23,6 % N--T 172,2 • 3
Le hombre 122 des pertes inc lu t les 100 colonies des nids abandonn6s du tableau III plus les 22 colonies d6jh identifi6es dans la subdivis ion pr6c6dente.

20 A. F R A N C t : E U R E T D. P E P I N
Si l 'on soustrait du taux sp6cifique le pourcentage que ces dernibres repr6sentent , soit 4,3 %, on obt ient un taux moyen de renouvel lement de 19,3 %. Ce chiffre peut se d6duire des donn6es du tableau III en faisant le rappor t des nouvel les colonies sur la p o p u l a t i o n de l ' ann6e pr6c6dente, plut6t que sur le total de l 'ann6e qui se t e rmine (fin de la saison de croissance), pour t en i r compte des fuc tua t i ons d 'une ann6e h l 'autre. E n effet, la diff6rence de 0,6 Vo par r appor t au taux effectif de renouvel lement de 18,7 % repr,6sente l ' impor tance de la fluc- tuat ion du niveau de la popula t ion de colonies pe nda n t la dur6e de l '6tude.
La diff6rence entre le taux sp6cifique de renouvel lement et le taux effectif de renouvel lement de la populat ion, pendan t un interval le de temps, s ' expl ique par l ' appar i t ion et la d ispar i t ion d 'un cer ta in nombre de colonies p e n d a n t ce m6me temps. Cette valeur (tant6t gain, tant6t perte) exerce une in f luence qui sera p ropor t ionne l le h sa grandeur . En d6 te rminan t 0 s pour une unit6 de temps donn6e, il devient poss ib le de. comparer , selon le point de rue de la product ivi t6 , le renouvel lement de diff6rentes popula t ions appar tenan t h u n e ou p lus ieur s esp~ces, v ivant dans des biotopes diff6rents. Le tableau IV pr6sente des valeurs
TABLEAU IV. - - Taux sp~cifique (0s) de renouvellement annuel pour quelques populations de colonies appartenant h des esp~ees diff~rentes de Formica.
TABLE IV. - - Specific annual replacement rate (0s) for some colony populations of different ants species of Formica.
Esp~ces Interval le ~ de t emlas v Os Source
1. F. dakotens is . . . . . . . . 3 ans 172,2 122 23,6 Texte 2. F. obscuripes . . . . . . . . 9 ans 119,6 149 13,8 KING et 8ALLEE (1956) 3. F. opacioentris . . . . . . 3 ans 415 81 6,5 SCHERBA (1963)
�9 4. F. ulkei . . . . . . . . . . . . . 10 ans 430,5 131 3,0 DnEYEn (1942) 5. F. exsectoides . . . . . . . . 10 ans 79,5 27 3,4 HAVILAND (1948)
de 0 s pour quelques esp~ces de F o r m i c a . I1 appara l t c la i rement que la vitesse de changement sur une base annuel le diff~re pour chaque cas. En par, t icul ier , les popula t ions du groupe F o r m i c a r u f a (1 et 2) se renouvel lent beaucoup plus vite que celles du groupe e x s e c t o i d e s (3 h 5). Remarque taxonornique : contra i - rement h la posi t ion de CREmHTON (1950), il faut consid6rer le groupe n6arct ique e x s e c t o i d e s comme diff6rent du groupe pa16arctique exsec ta . L'analyse de cette diff6rence, qui n 'a pas" sa place ici, devrai t signifier beaucoup de choses, par exemple, en rappor t avec le processus de fondat ion des colonies.
3. TAUX DE NATALITE ET DE MORTALITE
La somme des nouveaux nids cr66s (qu'i ls soient ou non occup6s au moment de leur d6couverte), compar6e au nombre de n ids occup6s h la fin de la pr6c6dente saison de croissance, donne des taux de natalit6 de 23,8, 15,8 et 14,1% de 1973 h 1975, pour une moyenne de 17,9 _ 4,5 %, selon les chiffres du tableau II. Dans
P R O D U C T I V I T b 7 D E L A F O U R M I FORMICA D A K O T E N S I S 21
l ' h y p o t h ~ s e od la r 6 o c c u p a t i o n d ' a n c i e n s n i d s est le fa i l de n o u v e l l e s c o l o n i e s
s eu l emen t , le t aux m o y e n a n n u e l de na ta l i t6 des c o l o n i e s p a s s e h 22,9-+-5,8 %.
De la m ~ m e m a n i ~ r e , le h o m b r e to ta l des n i d s a b a n d o n n 6 s ( a n c i e n s q-
n o u v e a u x ) au c o u r s d ' u n e ann6e d o n n e des t a u x de mor t a l i t 6 de 29,1, 19,9 et 21,5 %
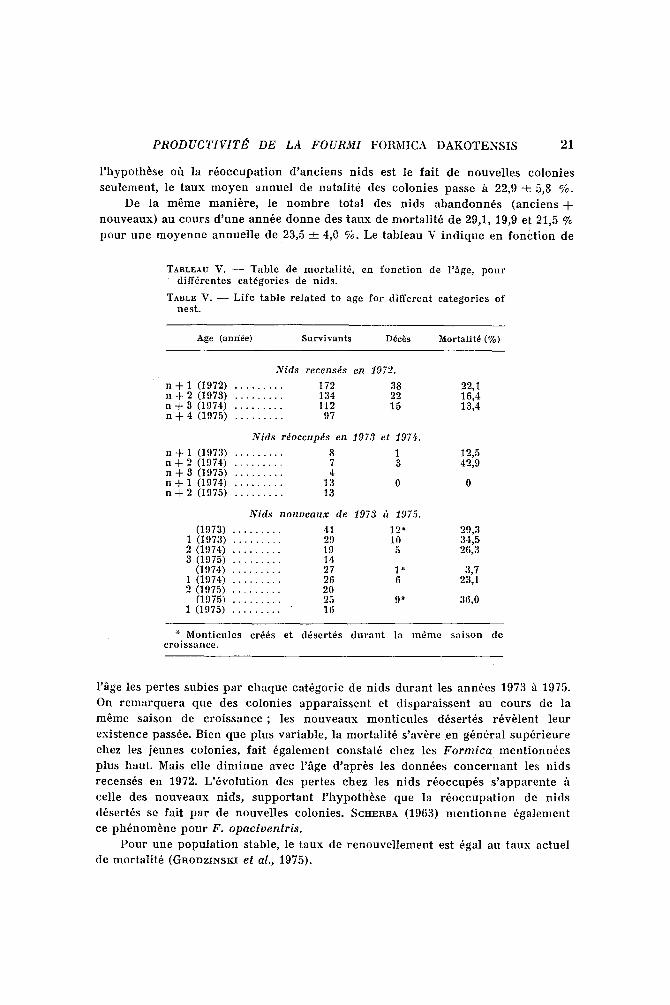
p o u r u n e I n o y e n n e a n i m e l l e de 23,5 _ 4,0 %. Le tab leau V i n d i q u e en f o n c t i o n de
TABLEAU V. - - Table de mortalitY, en fonction de i'~ge, pour diff~rentes cat6gories de nids.
TABLE V. - - Life table related to age for different categories of nest.
Age (anr/6e) Survivants D6cds Mortalit6 (%)
Nids recensds en 1972.
n q- 1 (1972) . . . . . . . . . 172 38 22,1 n -}- 2 (1973) . . . . . . . . . 134 22 16,4 n q- 3 (1974) . . . . . . . . . 112 15 13,4 n -}- 4 (1975) . . . . . . . . . 97
N i ds rdoccupds en 1973 et 197!~.
n q- 1 (1973) . . . . . . . . . 8 1 12,5 n + 2 (1974) . . . . . . . . . 7 3 42,9 n q- 3 (1975) . . . . . . . . . 4 n -I- 1 (1974) . . . . . . . . . 13 0 0 n d- 2 (1975) . . . . . . . . . 13
N i ds n o a o e a u x de 1973 ~'~ 1975.
(1973) . . . . . . . . . 41 12" 29,3 1 (1973) . . . . . . . . . 29 10 34,5 2 (1974) . . . . . . . . . 19 5 26,3 3 (1975) . . . . . . . . . 14
(1974) . . . . . . . . . 27 1" 3,7 1 (1974) . . . . . . . . . 26 6 23,1 2 (1975) . . . . . . . . . 20
(1975) . . . . . . . . . 25 9* 36,0 1 (1975) . . . . . . . . . " 16
"* Monticules crffis et d~sert6s durant la m~me saison de croissanee.
l"~ge les p e r t e s sub ie s p a r c h a q u e ca t~gor i e de n ids d u r a n t les an n 6 es 1973 "5 1975.
On r e m a r q u e r a que des c o l o n i e s a p p a r a i s s e n t et d i s p a r a i s s e n t au c o u r s de la
m 6 m e sa i son de c r o i s s a n c e ; les n o u v e a u x Inon t i cu l e s d6ser t6s r~v~lent l eu r
e x i s t e n c e passde . Bien que p lus va r i ab l e , la mor t a l i t 6 s ' av~re en g~n6ra l s u p 6 r i e u r e
chez les j eunes co lon ie s , fa i l 6ga l emen t cons ta t~ chez les F o r m i c a m e n t i o n n ~ e s
p lus haut . Mais elle d i m i n u e avec l '~ge d ' a p r ~ s les d o n n ~ e s c o n c e r n a n t les n i d s
r e c e n s 6 s en 1972. L ' 6 v o l u t i o n des p e r t e s chez les n i d s r 6 o c c u p 6 s s ' a p p a r e n t e h
cel le des n o u v e a u x n ids , s u p p o r t a n t l ' h y p o t h ~ s e que la r 6 o c c u p a t i o n de n i d s
d~ser t6s se fai t p a r de n o u v e l l e s co lon ies . SCHERUA (1963) m e n t i o n n e dga le inen t
ce p h ~ n o m ~ n e p o u r F . o p a c i v e n t r i s .
P o u r u n e p o p u l a t i o n s table , le t aux de r e n o u v e l l e m e n t est ~gal au t a u x ac tue l
de mor ta l i t6 ( G R o D Z I N S K I e t al., 1975).
22 A. FRANC(EUR E T D. PEPIN
Dans le cas de la popula t ion de colonies ~tudi~es ici, la mortal i t~ m o y e n n e
annuelle est tout h fair comparab le fi la natalit~ m oyenne annuelle. Les faibles
var ia t ions enregistr~es dans le nombre de colonies t6moignent donc de la re-
cherche d 'un ~quil ibre annuel entre ces d e u x param~tres. De plus, les taux de
natalit~ et de mortal i t~ tendent ~t d iminuer actuel lement , ce qui la issera i t c ro i re
que ]a popula t ion est en voie d 'a t te indre la charge b io t ique maximale du b io tope
ou que le mi l ieu se t r ans fo rme et devient moins favorable.
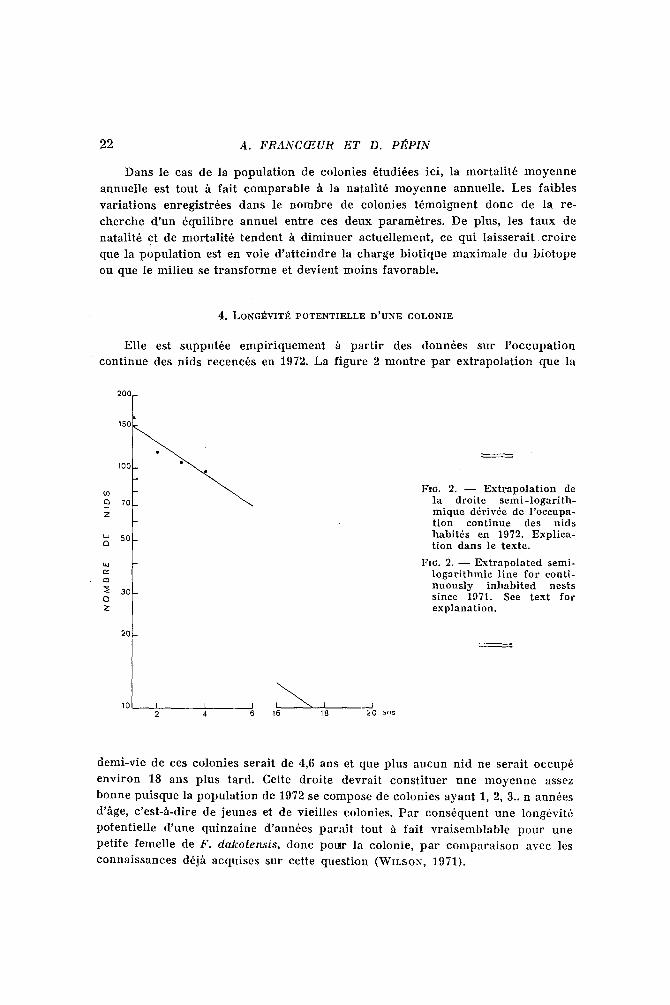
4. LONG~VIT]E POTENTIELLE D'UNE COLONIE
Elle est supput~e empi r iquemen t fi pa r t i r des donn~es sur l ' occupa t ion
cont inue des nids recenc~s en 1972. La f igure 2 mout re pa r ex t rapola t ion que la
20c
15r
100
70
50
30
2O
10 I I I I ' ~ I I 2 4 6 16 18 20 ans
I:m. 2. - - Extrapolation de la droite semi-logarith- mique d~riv6e de l'oceupa- tion continue des nids habit,s en 1972. Explica- tion darts le texte.
Fx6. 2. - - Extrapolated semi- logarithmic line for conti- nuously inhabited nests since 1971. See text for explanation.
demi-vie de ces colonies serait de 4,6 ans et que plus aucun nid ne serai t occup6
envi ron 18 ans plus tard. Cette droi te devrai t cons t i tuer une m oyenne assez
bonne puisque la popula t ion de 1972 se compose de colonies ayant 1, 2, 3.. n ann6es
d'~ge, c 'est-h-dire de jeunes et de viei l les colonies. Par cons6quent une long6vit6
potent ie l le d 'une quinzaine d 'ann6es para~t tout h fai t v ra i semblable pour une
peti te femelle de F. d a k o t e ~ i s , donc pour la colonic, pa r compara i son avec les
connaissances d6jh acquises sur cette quest ion (WILSON, 1971).

PRODUCTIVITE DE LA FOURMI FORMICA DAKOTENSIS 23
Variations dans l'occupation des nids.
Le tableau II fait ne t tement ressort i r l 'existence d 'une occupat ion cont inue, d 'une occupat ion d iscont inue r6sul tant du ph6nom6ne d ' a ba ndon et de r6oc- cupation, enfin d 'un apport constant de nouveaux nids qui s ' in t6grent d~s l ' ann6e de leur appar i t ion h Fun des deux types d 'occupation.
1. OCCUPATION CONTINUE ET DISCONTINUE
Des 172 nids habit6s en 1972, seulement 97 l '6taient encore en 1975. I1 s 'agit d 'une d iminu t ion totale de 43,6 % sur 3 ans r6sultant de pertes annuelles . L'oc- cupat ion d i s c o n t i n u e pour ce m6me groupe se t radui t en 1975 par un taux de r6occupat ion de 3,1% soit 7 nids d6sert6s en 1973, dont 5 r6cup6r6s en 1974 et 2 en 1975 (tableau II). La baisse r6elle en nids abandonn6s d iminue ainsi 40,5 % pour une moyenne annuel le de 13,5 %.
2. OCCUPATION SECONDAIRE ET MIXTE
Quand un n id est abandonn~ d6finit ivement par F. dakotensis h la suite de la mort de la colonie, il reste ut i l isable par d 'autres esp~ces du ran t une p~riode de 3 h 5 ans selon sa taille. Le tableau I pr~sente la liste des diff~rentes fourmis retrouv~es dans ces nids. Formica neorufibarbis semble marque r un certain int~rf~t pour ee type de micromil ieux, tandis que la presence des autres esp~ces s'av~re plut6t rare et temporaire .
A trois occasions, un nid habi ts par F. clakotensis abri ta i t 6galement des ind iv idus de F. neorufibarbis. Nods ne savons pas s'il existait, ou non, des relat ions entre les deux esp~ces.
3. REOCCUPATION DE NIDS ABANDONNI~S
Bien que ce ph~nom~ne ne cont r ibue pas aussi eff icacement h ma i n t e n i r la popula t ion en ~quil ibre que la creat ion de nouveaux nids, il repr~sente tout de m(~me de 21,6 "~ 33,3 % (moyenne : 26,3 • 2,7 %) des changements annuels dans l 'occupat ion.
Le tableau VI aide h d~terminer l ' in l luence de l 'usage ant~r ieur et de l'hge des nids sur ce ph6nom~ne. Des changements annuels f requents ne favorisent pas la r6occupat ion et i nd iquen t que le taux de d6m~nagement, s ' i l existe, reste tr~s faible. Les nids sont d 'au tant plus r~occup~s qu'i ls n 'on t pas ~t~ abandonn~s depuis tr~s l on~emps . La re la t ion est inversement propor t ionnel le . Ainsi, l'~ge ou l'~tat de conservat ion des nids abandonn~s apparai t comme un facteur impor- tant dans le ph~nom~ne de la r~occupation.

24 A. F R A N C t E U R E T D. P E P I N
TABLEAU VI. - - Pourcentage de r6occupation en fonetion de l'hge et de l'usage ant~rieur des nids. Signification des sym- boles au tableau II.
TABLE VI. - - Percentage of utilisation of empty nests in relatiol~ to age and preceding yearly use of ne~ts. Meaning of symbols as in table II.
Usage en 1972-1973 Abandon Rfioeeupalion Poureentage
1972 1973 82 8 9,8
1973 197~ 74 3 4,1
+ 38 5 13,2 0 12 5 41,7
197~ 1975 71 2 2,8
+ - - R3 2 6,1 + + 22 0 0 - - + 1 0 0
0 - - 7 0 0 0 --- 10 1 10,0 00 1 0 0
4. ABANDON DES NIDS
A la fin d 'une saison de croissance, le nombre total d 'abandons ine lu t des nouveaux et des anciens nids. Le tableau II mont re que l ' appor t respec t i f de
chaque cat6gorie var ie d 'ann6e en ann6e et en fonct ion de l'~ge. Ces f luctuat ions
appara issent toutefois plus impor tan tes chez les nouveaux aids. Les a ids const ru i t s
dans des posi t ions d~favorables (pente nord d 'un t e r t re ou base du nid t rop pr6s
de la nappe phr6at ique) sont rapid.ement 61imin6s. Expr im6 par le r appor t des abandons sur les a ids occup~s, le taux annuel
moyen d ' abandon s'61bve ~ 23,6 ___ 5 , 1 % et co r respond dans ce eas-ci au taux de mortal i t6 des colonies, puisque la migra t ion est r6put6e nulle. I1 est sup~rieur
de 4,9 % au taux effectif de r enouve l l ement annuel. Cet 6cart comprend , d 'une
part, le d6ficit moyen (0,6 %) entre les per tes et le renouvel lement duran t la
p~r iode d 'observa t ion et, d 'aut re part , l ' im por t ance re la t ive (4,3 %) des nouveaux mont icules abandonn~s l 'ann~e de leur const ruct ion . La difference entre les deux
taux moyens s 'av~re hautement s ignif icat ive (P < 0,01). Bien qne peu blev~,
l ' appor t de la derni~re cat6gorie de n id n 'est done pas n~gligeable, d ' au tan t
plus qu 'e l le pe rmet essent iel lement le ph6nom~ne de la r6occupat ion.
Des 82 nids trouv6s abandonn6s en 1972, 13 furent r6occup6s par la suite
dont 9 l '6 ta ient en 1975, landis que 69 ou 84,1% demeurbrent toujours d6serts
(tableau II). Bien que l '6tude de la d6gradat ion des nids abandonn6s n ' en t ra i t
pas dans les objectifs in i t iaux du projet , des notes accumul6es lors des recen-
sements annuels et un relev6 per t inen t en 1975 permet ten t de faire quelques
observat ions. Le nombre de mont icu les r i de s e o m p r e n d des a ids encore en boa
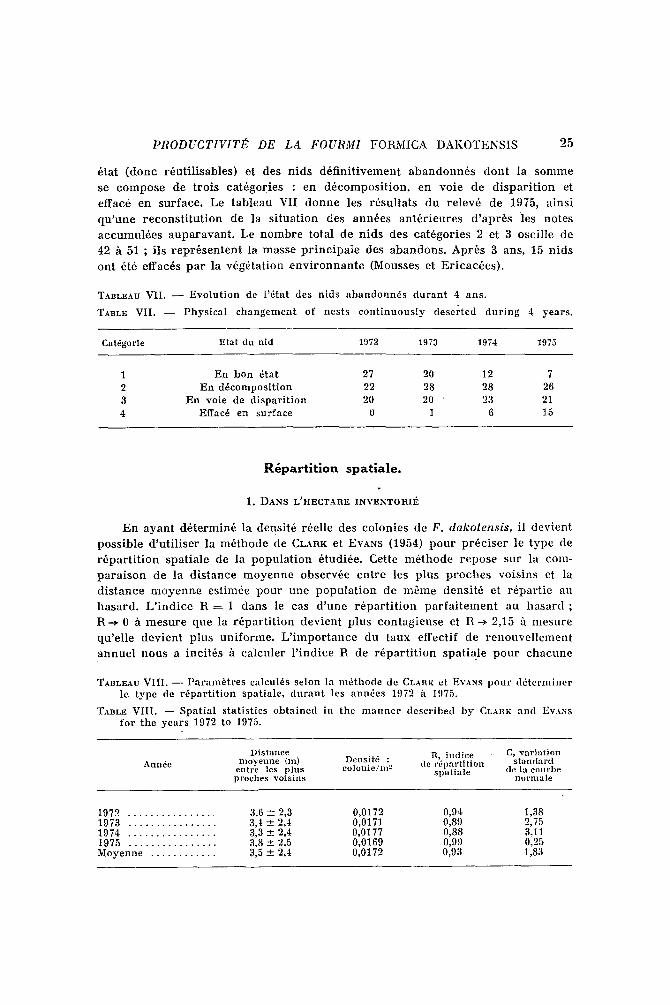
PRODUCTIVITE DE LA FOURMI F O R M I C A D A K O T E N S I S 25
~ta t ( d o n c r 6 u t i l i s a b l e s ) e t des n i d s d 6 f i n i t i v e m e n t a b a n d o n n 6 s d o n t la s o n m l e
se c o m p o s e de t r o i s c a t 6 g o r i e s : en d 6 c o m p o s i t i o n , e n vo l e de d i s p a r i t i o n et
e f fac6 e n s u r f a c e . Le t a b l e a u VI I d o n n e les r 6 s u l t a t s d u r e l e v 6 de 1975, a i n s i
q u ' u n e r e c o n s t i t u t i o n de la s i t u a t i o n des a n n ~ e s a n t 6 r i e u r e s d ' a p r ~ s les n o t e s
a c c u m u l 6 e s a u p a r a v a n t . Le n o m b r e t o t a l de n i d s de s c a t 6 g o r i e s 2 et 3 o sc i l l e de
42 h 51 ; i l s r e p r 6 s e n t e n t la m a s s e p r i n c i p a i e des a b a n d o n s . A p r ~ s 3 ans , 15 n i d s
o n t 6t6 e f fac6s p a r l a v 6 g 6 t a t i o n e n v i r o n n a n t e (Mousses et E r i c a c ~ e s ) .
TABLEAU VII. - - Evolu t ion de l '6tat des nids abandonn6s du ran t 4 ans.
TABLE VII. - - Physical chaugement of nests con t inuous ly deserted dur ing 4 years.
Cat~gorie Etat du nid 1972 1973 1974 1975
1 En boa 6tat 27 20 12 7 2 En d6composit ion 22 28 28 26 3 En vole de d ispar i t ion 20 20 " 23 21 4 Effac6 en surface 0 1 6 15
R 6 p a r t i t i o n s p a t i a l e .
1. DANS L'HECTARE INVENTORIE
E n a y a n t d 6 t e r m i n 6 la dens i t 6 rde l le des c o l o n i e s de F. dakotensis, i l d e v i e n t
p o s s i b l e d ' u t i l i s e r la m 6 t h o d e de CLARK et EVANS (1954) p o u r pr(~ciser le t y p e de
r 6 p a r t i t i o n s p a t i a l e de la p o p u l a t i o n 6 tudi~e . Cet te m 6 t h o d e r e p o s e s u r la com-
p a r a i s o n d e la d i s t a n c e m o y e n n e o b s e r v ~ e e n t r e les p l u s p r o c h e s v o i s i n s et la
d i s t a n c e m o y e n n , e e s t i m 6e p o u r u n e p o p u l a t i o n d e m S m e d e n s i t 6 et r ~ p a r t i e au
h a s a r d . L ' i n d i c e R = 1 d a n s le cas d ' u n e r 6 p a r t i t i o n p a r f a i t e m e n t au h a s a r d ;
R - > 0 h m e s u r e que la r 6 p a r t i t i o n d e v i e n t p l u s c o n t a g i e u s e et R - > 2,15 'h m e s u r e
q u ' e l l e d e v i e n t p l u s u n i f o r m e . L ' i m p o r t a n c e du t a u x e f fec t i f de r e n o u v e l l e m e n t
a n n u e l n o u s a i n c i t e s '/1 e a l c u l e r l ' i n d i c e R de r 6 p a r t i t i o n spat ia , le p o u r c h a c u n e
TABLEAU VIII. - - Param~tres calcul6s selon la m~thode de CLANK ct EVANS pour ddtcrminer le type de r~par t i t ion spatiale, du ran t les ann~es 1972 h 1975.
TABLE VIII. - - Spat ial s tat ist ics obta ined in the m a n n e r described by CLANK and EVANS for the years 1972 to 1975.
Dis tnnce R, i n d i t e C, v a r i a t i o n A n n i e m o y e n n e (m) Den s i t6 : s t a n d a r d
en t re les p l u s c o l o n i c / i n ~ (le r d p a r t i t i o n p roches v o i s i n s s p a t i a l e denormalela co t t rbe
1972 . . . . . . . . . . . . . . . . 1973 . . . . . . . . . . . . . . . . 1974 . . . . . . . . . . . . . . . . 1975 . . . . . . . . . . . . . . . . Moyenne . . . . . . . . . . . .
3,6 ----- 2,3 0,0172 0,94 1,38 3,4 • 2,4 0,0171 0,89 2,75 3,3 • 2,4 0,0177 0,88 3,11 3,8 • 2,5 0,0169 0,99 0,25 3,5 • 2,4 0,0172 0,93 1,83

2s A. FRANC(EUR E T D. PEPIN
des 4 ann6es d 'observat ions , ainsi que la va leur C, var ia t ion s tandard de la
courbe normale, qui 6value la s ignif icat ion s tat is t ique de R. On consul tera les
auteurs de la m6thode pour conna l t re le d6tail des calculs ou encore ELMES (1974).
Les r6sultats sont pr6sent6s au tableau VIII. Les valeurs de C ind iquen t que
la r6par t i t ion spatiale des nids ne s '6car tent pas de fas s ignif icat ive de la
i il i11 I!ili!ii i!ii!!!ii!i!iiiii!i!iii ::;i::i::iii::;i;::ii ' ,111 I;iiiiii!!iiii::i:=::iiii::~ii::iiiiii!iii=:iiii::::iiii::iJ
. 'i:::~i::::iJ:i:i::::il' I ':i::ii:iiiiiiiiiill i!iiiii!iiiiiiiiiiiii!ii!i!i ~ i ! i ~ i ;~ iiiiiiiiiiliiii ~i:iiii~iiii:!:::iii~i~::iiiiii'iiiiii:i:ii iiiiiiiiiiiiiiii
;ii;ii', iiii ',ii i iiii',ii!i',i',i',iiiii;',i',i;',iii iiii!ii " i~iiill I I I I I 11 t. l=;)iisi:i:!:iii?!:i:i:i:i:ili:i:isi:i:i:i:iii:iii:i:!:i:):i:~i!!l
FIG. 3. - - Densit~ des colo- nies pour i972. L'hectare reeens~ est subdivis6 en unit~ de 100 m e .
Fro. 3. - - Colony density for the year 1972 by units of 100 m-* in the censured hectare.
2 nids et plus 1 nid 0 hid
r6par t i t ion au hasard en 1972 et 1975 et qu 'e l le est fa iblement eontagieuse en
1973 et 1974 (P < 0,01). Pour la p6r iode d 'observat ion , la r6par t i t ion reste au
hasard dans l 'hec tare inventor i6 avec une t endance vers le type contagieux lorsque
la densit6 augmente. La figure 3 i l lustre cette r6par t i t ion pour l 'ann6e 1972.
La moyenne des distances moyennes entre les plus proches voisins 6valu6es
durant quatre ann6es cons6cutives se chiffre h 3,53 m. Si l 'on consid~re que cette valeur co r respond au rayon du t e r r i to i re exploit6 par une colonie, celui-c i est
alors estim6 h une surface moyenne de 39,15 m e.
2. EN FONCTION DES HABITATS
La figure 1 mont re que l ' hec ta re prospect6 r6c61e trois types d 'habi ta t s
diff6rents par r appor t h la pr6sence et au d6veloppement de la strate arborescente .
Cette surface 6chanti l lon se t rouve sur un f ront d ' envahissement de la tourb i6re par l '6pinet te noire.

PRODUCTIVITI~ DE LA FOURMI FORMICA DAKOTENSIS 27
L'analyse de la densit~ des n ids occup~s pour chaque recensement annuel apparai t au tableau IX, en fonct ion des trois habitats identifi~s (ouvert, interm~- diaire et fermi) dont les pourcentages respectifs de recouvrement ont ~t~ calculus par planim~tr ie . On d~termine ensuite le hombre moyen de nids occup~s par habitat , ce qui permet d 'est imer une densit~ moyenne par hectare.
T A B L E A U I X . - - Densit~ des colonies de F. dakotensis en fonetion de trois types de milieu : 1, ouvert; 2, interm~diaire; 3, fermi.
T A B L E I X . - - Colony density of F. dakotensis in three different habi ta ts : 1, open; 2, intermediate; 3, bushy trees.
Milieu Pourcen tage N o m b r e de n ids N o m b r e Densit~
de m o y e n m o y e n n e r e e o u v r e m e n t 1972 1973 197,1 1975 de n ids p a r hec ta re
1 14 11 10 12 9 10,5 75,0 2 67 148 150 150 147 148,7 222,0 3 19 13 11 15 13 13,0 68,4
Total 100 172 171 177 169 172,2
I1 ressort tr~s net tement que plus de 86 % des nids ou colonies se re t rouvent dans le mil ieu interm6diaire , expr imant la pr6f6renee de F. dakotensis. Elle peut 6galement s ' instal ler et survivre dans les deux autres. Dans le cas du mil ieu ferm6, les nids sont situ6s h In p6riph6rie (jamais h l ' in t6r ieur) des bosquets d'6pinettes, plus pr6cis6ment sous les b ranches les plus longues et basses. A l ' in t6r ieur du mil ieu interm6diaire , la dis tance n loyenne du plus proclle voisin s'61~ve h 3,2 • 2,2 m d'apr~s les donn6es de 1974. La valeur de R e s t 0,96 et celle de C 0,98. La r~part i t ion spatiale h l ' in t~r ieur de ce mil ieu par t icu l ie r reste au hasard, alors qu'elle cst fa iblement coutagieuse pour l ' ensemble de l 'hectare, cette annie- lb . Ces rSsultals peuvent s 'expl iquer par l ' inf luence de la t ransfor- 'nat ion du mil ieu sur la fourmi. En outre, selon ELMES (1974), les esp~ces cons- t rn isant des monticules pour ra ien t jouir d 'une plus grande libert~ dans le choix des micromi l ieux de nidification. Dans ce cas-ci, le d~veloppement des ter tres el les f luctuations de la nappe phr~at ique semblent inf luencer ce choix. Cet auteur souligne que les popula t ions de colonies d ' insectes sociaux tenden t vers une r6par t i t ion spatiale uni forme h cause de la competi t ion. Cela ddpend du niveau de la d e n s i t & ' P e u d 'auteurs ont rapport~ jusqu ' ic i une r~par t i t ion au hasard pour les esp~ces de fournfis (BRIAN, 1956; ELMES, 1974; LEWIS, 1975; TaLnOV, 1954 ; WALOFF et BLACKITH, 1962).
3. INFLUENCE DE LA SUCCESSION
La colonisat ion de la tourbi~re ouverte par F. dakotensis peut commencer en bordure. Au stade suivant, les ~pinettes s ' implan ten t progress ivement selon tin front plus ou moins d iscont inu (fig. 1). Cela favorise le d~veloppement de la

28 A. FRANC(EUR ET D. PEPIN
populat ion de cette fourmi, qui passe par un max imum qui ne corr ,espondrai t
pas h la charge b io t ique-poten t ie l le de cet habi ta t in term6diaire , d'ofl une r6par- l i t ton qui reste au hasard avec une faible tendance vers le type contagieux. En effet, le mi l ieu se t r ans fo rme ensui te rap idement en bosquets de plus en plus
denses et vastes que d6serte la fourmi. BOLAS (1956) a d6jh soulign6 que le manque de site ad~quat de n id i f ica t ion peut
l imiter une popula t ion de colonies de fourmis. Pour une superficie donn6e, on peut ainsi vist~aliser le passage du mil ieu in term6diai re et sa popu la t ion de F. dakotensis. Cette derni~re peut 6tre qualifi~e d'esp~ce de t rans i t ion , tout comme le mil ieu qu'elle pr~f~re, en fonct ion de l 'espace et du temps.
La t rans format ion progressive de la pessi~re tourbeuse vers la matur i t6 (arboraie dense sur sol organique h surface s~che) influence l '6volut ion de la populat ion de cette fourmi, darts un espace donn~, en t r ans fo rman t l 'habi ta t , c'est-h-dire la composi t ion des sites favorables de nidif icat ion et les condi t ions microcl imatiques. La densit6 des colonies (mats non le type de r6par t i t i0n spa- tiale, ou alors faiblement) s 'en t rouve affect6e dans ies types d 'habi ta ts consid6r6s. On peut comparer les figures 1 et 3 pour s 'en convaincre. Dans le p remie r article (FBANE(EUR et P~.PIN, 1975), nous avions ddjh percu l ' inf luence de la succession v6g6tale sur les var ia t ions de la densit6 des colonies dans des pessibres h des stades diff6rents d '6volution.
D i m e n s i o n s e t f o r m e s d e s m o n t i c u l e s .
1. DIMENSIONS ~T OCCUPATION
Pour ne pas pe r tu rbe r les colonies, nous ut i l isons les d imens ions ext6r ieures du mont icule qui ne cor responden t pas au volume total du hid, h savoir la largeur, la longueur et la hau teur apparente maximale de la couche de d6bris v6g6taux au-dessus de la sphaigne vivante. Cette est imation par t ie l le util is6e dans le cos des nouveaux mont icu les permet n6anmoins de faire quelques cons- tatal ions int6ressantes.
a) Nouveaux monticules abandonn~s l ' ann6e m6me de leur cons t ruc t ion . Le volume moyen de cette cat6gorie s'dlbve 'h 1 100 cm a. Ceux qui seront r6oc- cup6s par la suite aff ichaient un volume m i n i n m m de 1 000 cmS.
b) Nouveaux monticules occupds l ' ann6e de leur construct ion. P a r mi ceux qui seront abandonn6s l ' ann6e suivante, seulement quatre avaient un volume sup6rieur 'h 1500 cma. Par contre, sur tous les monticules qui demeure ron t oecup6s l ' annde suivante, un seul avail un volume inf6r ieur h cette l imite.
Pour les nouveaux mont icules , il semble donc exister un volume cr i t ique qui s '~tablit :entre 1 000 et 1 500 cma. Les colonies qui at teignent ou d6passent ce seui[ de cons t ruct ion r6uss i ra ient h s ' implan te r de fa~on viable. Celles qui ne peuvent l ' a t te indre avant les mauvaises condi t ions h ivernales aura ien t peu de chance de se main ten i r . I1 para i t 6galement exister un volume m i n i m u m pour

P R O D U C T I V I T ~ DE LA F O U R M I FORMICA DAKOTENSIS 29
que la r6occupat ion des mont icu les d~sert6s l 'ann~e m~me de leur ~dification
soit possible ul t~rieurement. Enfin, l e s d imensions ext~rieures des nids refl~-
tera ient le niveau d6mographique des colonies pendant leur p6r iode de crois-
sance seulement.
2. FORMES ET POLY(2ALISME P R I M I T I F
La figure 4 i l lustre les diff~rentes formes de s t ructures externes d 'apr~s le
profi l . Les formes pet i t d6me et grande plage sont s tandards dans la pessi~re
tourbeuse. Les formes 3 et 4 demeuren t assez rares avec seulement une douzaine
de cas observes dont 4 pour le double d6me. La cons t ruc t ion d 'un d6me simple
sur une viei l le plage a ~t6 observ~e dans la nature et en laboratoire . Le p remie r
Fro. 4. - - Profils sch~matiques des mon- ticules construits par Formica dako- tensis. Sur sol organique (sphaignes) : 1, petit d6me; 2, grande plage; 3-I,, d6me simple et double sur vieille plage; sur sol minSral (parmi les herhes) : 5, nids h trois petits dSmes interreli~s.
Fro. 4. - - Outlined profile of mounds build by Formica dakotensis. On orga-
1 ~ 2 - - - -
nic soil (sphagnum mosses) : 1, small conical mound; 2, large flat mound; 3-/*, single or double conical mound on old fiat base; on mineral soil (among grasses) : 5, small polydomous nest with connections between mounds.
auteur a eu l 'occasion en 1975 de voi r un nid h perils d6mes interrel i~s h la
Reserve E. S. George, au Michigan. TALBOT (1971) a d~jh d~crit ce type de nid et son habitat .
Cette s~quence de formes repr~sente des ~tapes possibles vers l ' appar i t ion
du polyca l i sme typ ique tel qu 'on le connal t chez les F o r m i c a des groupes exsec- toides et ru[a. La phase suivant celle de la forme '~ d6mes reli6s serai t la s~pa-
ra t ion phys ique des monticules, paral l~lement au d~veloppement de nouveaux
modes de fondat ion des colonies (bouturage) et de la polygynie .
C O N C L U S I O N S
Les recensements effectu6s au cours des ann6es 1972 h 1975 ont permis non
seulement de quantif ier certaines var iables n6cessaires pour d6 te rminer la pro-
ductivi t6 d 'une populat ion de colonies de F. dakotens is , mais aussi d 'ob ten i r des donn6es sur l '6cologie de cette esp~ee.
I1 s 'agit d 'une fourmi de t ransi t ion pour la pessi~re tourbeuse. On la re t rouve
durant la phase d 'envahissement de :a tourbi~re par nne strate arbust ive, puis
a rborescente de P i c e a mariana. Pour un espace donn6, c'est le passage dans le

30 A. F R A N C O ~ U R ET D. PEPIN
t e m p s d ' u n h a b i t a t de t r a n s i t i o n qui ab r i t e , e n t r e au t res , une p o p u l a t i o n de
F. dako tens i s . Cet te d e r n i ~ r e a p p a r a i t a c t u e l i e m e n t en 6qu i l i b r e d a n s l ' h e c t a r e
6tudi6 et en vo ie d ' a t t e i n d r e l ' e x p a n s i o n m a x i m a l e que p e r m e t la v i t e s s e de la
s u c c e s s i o n . U n t a u x e f f ec t i f de r e n o u v e l l e m e n t a n n u e l de 18,7 % aft, e r i e la p o p u -
l a t ion de co lon ie s , l eque l d 6 p e n d d ' u n e mor t a l i t 6 m o y e n n e de 23,5 % et d ' u n e
na ta l i t6 m o y e n n e de 22,9 %. La d i f f 6 r e n c e e n t r e le r e n o u v e l l e m e n t e t l a na ta l i t6
s ' e x p l i q u e p a r les p e r t e s s u b i e s d u r a n t la s a i s o n de c r o i s s a n c e et no t6es h la f in
de ce t t e sa i son . La m o r t a l i t 6 61ev6e des j e u n e s co l o n i e s s e m b l e a t t r i b u a b l e p r i n -
c i p a l e m e n t aux f l u c t u a t i o n s de la n a p p e p h r 6 a t i q u e en su r f ace .
REMERCIEMENTS. - - La collaboration et l 'hospitali t6 du D ~ Mary TALBOT ont faeilit~ le s6jour du premier auteur ~t la R~serve E.S. George du Michigan, E.-U. Cette ~tude fut entreprise grhce h une subven t ion du Conseil National de Recherche du Canada.
REFERENCES
BRIAN (M. V.), 1956. - - The natural densi ty of M. rabra and associated ants in Scotland. Ins. sot., 3, 473-487.
CLANK (P. J.) et EVANS IF. C.), 1954. - - Distance to nearest neighbor as a mesure of spatial re la t ionship in populat ions. Ecoloffy, 35, 445-453.
CREIGHTON (W. S.), 1950. - - The ants of North America. Ball. Mils. Comp. Zool. Harvard, 10~, 1-585.
D]~EYEU (W. A.), 1942. - - Fur ther observat ions on the occurrence and size of ant mounds wi th reference to their age. Ecoloyy, 23, 486-490.
ELMES (G. W.), 1974. - - The spatial d is t r ibut ion of a populat ion of two ant species living in l imestone grassland. Pedobioloqia, 1L 412-418.
FRANC(/'~UR (A.) et PIPPIN (D.), 1975. - - Produetivit6 de la fourmi Formica dakotensis dans la pessi6re tourbeuse. 1. Densit6 observ6e et densit6 estim6e des colonies. Ins. soc., 22, 135-150.
GRoazlr~sxi ~V.), KLEKOWSKX (R. Z.) et DUNCAN (A.), 1975. - - Methods for ecological bioener- getics. IPB Handbook No. 24. Blackwell, 6dit., Oxford, 367 p.
I-IAvILAND (E.), 1948. - - Mound changes af ter 10 years in colonies of Formica exsectoides Forel. Ann. Ent. Soc. Amer., M, 438.
KING (R. L.) et SALLEE (R. M.), 1956. - - On the half- l ife of nests of Formica obscaripes Forel. Proc. Iowa Acad. Sci., 63, 721-723.
LEwis IT.), 1975. - - Colony size, densi ty anti dis t r ibut ion of the leaf-cut t ing ant, Aero- myrmex octospinosus (Reich) in cultivated fields. Trans. R. Ent. Soc. Lond., 127, 51-64.
PETRUSEWICZ (K.) et MACFADYEN (A.), 1970. - - Product ivi ty of terrestr ial animals pr incipals and methods. IPB Handbook No. 13, Blackwell, ~dit., Oxford, 190 p.
SCHBnBA (G.), 1963. - - Populat ion characterist ics among colonies of the ant Formica opa- civentris Emery (Hym. : Formieidae). Joar. N. Y. Ent. Soc., 71, 219-232.
TALBOT (M.), 1954. - - Populat ions of the ant Aphaenofasier (Attomyrma) traetae Forel on abandoned f ie lds on the Edwin S. George Reserve. Contrib. Lab. Vertebr. Biol. Univ. Mich., No. 69, 1-9.
TALnOT (M.), 1971. - - Fl ights of the ant Formica dakotensis Emery. Psyche, 78, 169-179. WALOFF IN.) et BLACKITH (R. E.), 1962. - - The growth and dis t r ibut ion of the mounds of
Lasius flaous (Fabricius) (Hym. Formieidae) in Silwood Park, Berkshire. J. Anita. Ecol., 81, 421-437.
WXLSON (E. O.), 1971. - - The insect societies. Belknap Press of Harvard University, 6dit.. Cambridge, 548 p.





























