Embed Size (px)
Citation preview

Proies captur6es et strat6gies pr6datrices chez deux esp~ces d'araign6es orbit~ies: A rgiope bruennichi et A raneus marmoreus
[Prey and predatory strategies of two orb-weaving spiders: Argiope bruennichi and Araneus marmoreus]
Alain Pasquet Laboratoire de Biologie du Comportement, Universitb de Nancy I. B.P. 239, 54506 Vandoeuvre-les-Nancy Cedex, France
Mots cl6s: Araneae, Argiopidae, rbgime alimentaire, s61ection des proies [Keywords: Orb-weaving spiders, diet, prey-selection]
Abstract
L'analyse des r6gimes alimentaires de deux espbces d'Araign6es b, toile g6ombtrique, Argiope bruennichi (Scopoli) et Araneus marmoreus (Clerk), montre qu'A. marmoreus capture 14,1 proies/jour surtout des Diptbres (75%) et des proies de petite taille (0 4 mm) (86%) et qu'A. bruennichi capture 4,3 proies/jour. Les proies de cette dernibre espbce sont plus diversifibes (Dipt~res, Hym6noptbres, Orthoptbres et H6mipt~res) et de plus grande taille que pour A. marmoreus. L'analyse des proies potentielles disponibles montre que ces deux esp~ces occupent des strates diff6rentes faunistiquement, dans un m~me milieu.
Introduction
Pour appr6cier rimpact des Araign6es sur les peuplements d'Insectes, il est n6cessaire de conna~- tre les m6canismes qui r6gissent les relations prb- dateur-proies dans un espace donn6. La s61ection des proies est un de ces m6canismes; elle se traduit par une diffbrence qualitative ou quantitative entre proies disponibles et r6ellement captur6es.
Des 6tudes quantifient le r6gime alimentaire des Araign6es tissant une toile (Turnbull, 1960; Kajak, 1965a, b; Robinson & Robinson, 1970; Schaefer, 1974; Turner, 1979; Nyffeler, 1982; Nyffeler & Benz, 1978, 1979; Nentwig, 1980, 1982, 1983 et Olive, 1980). Mais il n'existe pratiquement pas d'6- tudes qui comparent le r6gime alimentaire aux pro- ies potentielles qui circulent dans l'habitat des Araign6es (Kajak, 1965a, b).
Notre 6rude porte sur le rbgime alimentaire de deux esp~ces d'Araign6es orbitbles qui exploitent le
Entomol. exp. appl. 36, 177-184 (1984). © Dr W. Junk Publishers, Dordrecht. Printed in the Netherlands.
m~me habitat: Argiope bruennichi (Scopoli) et Araneus marmoreus (Clerk).
Pour aborder la sblection des proies pour ces deux espbces nous avons compar+ d'une part les proies potentielles des diff6rentes zones occup6es par les Araignbes et d'autre part les proies poten- tielles et r6ellement captur6es par les Araign6es.
Mat6riel et m6thodes
Le terrain d'6tude (3300 m 2) est une ancienne prairie, en friches depuis quelques ann6es, bord6e sur trois c6t6s par de la for~t (dont il est s6par6 par une lisibre d'une dizaine de m~tres) et par un champ cultiv+ sur le 4 ~me c6t6. Les gramin6es et les 16gu- mineuses, esp~ces v6g6tales dominantes, fournis- sent les supports n6cessaires aux Araign6es pour la construction de leur toile; leur hauteur varie de 40 cm ~ 60 cm (gramin6es) et de 80 cm ~ 100 cm (16gumineuses).

178
A. marmoreus et A. bruennichi construisent 1cur toile le matin et effectuent le maximum des captures durant la journbe; les toiles souvent tr~s abim6es le soir ne sont pas reconstruites / tce moment. Ceci justifie nos observations durant la pbriode diurne. Nous n'avons pris en consid6ration que des toiles de femelles adultes. La p6riode d'6tude s'6tend du 15-08 au 15-09-1981.
Les toiles sont choisies au hasard et les pr6Dve- ments s'effectuent routes les heures de 7 h h 19 h selon un circuit d6termin6. Les proies r6ellement captur6es par les Araign6es sont prises directement dans la toile ou dans les ch6licbres des Araignbe fl l'aide de pinces et conserv6es dans l'alcool 5. 70 ° C. Pour ne pas entrainer de perturbations trop impor- tantes dans l'6tat psycho-physiologique des Araig- n6es, ces pr61bvements ont lieu tousles deux jours.
Les proies circulant dans l 'habitat sont pr61ev6es aux m~mes dates 5. l'aide d'assiettes colorbes (Pas- quet, 1982; Pasquet & Krafft, 1980). Nous avons choisi des assiettes de couleur jaune et de couleur rouge de fagon h couvrir au max imum le spectre d 'at traction des Insectes (Roth & Couturier, 1966). Ces assiettes, plac6es par paire de couleurs diff6- rentes 5- 4 hauteurs (0 cm, 25 cm, 50 cm, et 75 cm) de fagon 5- couvrir toute la v6g6tation, d61imitent ainsi dans le milieu 4 strates appel6es respectivement: strate 0, (de 0 cm 5- 25 cm), strate 25 (de 25 cm h 50 cm), strate 50 (de 50 cm fl 75 cm) et strate 75 (au-dessus de 75 cm). Les captures sont conserv6es dans l 'alcool 5- 70 ° C. Les assiettes colo- r6es sont des pibges attractifs (Roth & Couturier, 1966) qui peuvent conduire 5- une surestimation de certains ordres (Dipt~res); nous ne les avons utilis6s qu'en rant qu'indice d 'abondance relative entre les diffbrentes zones exploit6es par les deux espbces d'Araign6es.
Toutes les captures sont ramenbes au laboratoire et y sont class6es par ordre syst6matique en 7 groupes: Orthopt~res (ORT), L6pidoptbres (LEP), H6mipt~res (H6tbroptbres plus Homopt~res) (HEM), Col6optbres (COL), Hym6nopt~res (HYM), Dipt~res (DIP) et Divers (DIV) et par taille en trois classes:.Petites (0 ~ 4 mm), Moyennes (4 5. 10 mm) et Grandes (Plus de 10 ram).
Enfin, nous avons relev6 certaines caract6ris- tiques des toiles: le diambtre oblique, interm6diaire entre un diambtre horizontal et un diam~tre verti- cal, le nombre de rayons, le nombre de tours de spires. La localisation de la toile est caract6ris6e par la hauteur de son centre par rapport au sol.
R~sultats
Les proies r6ellement capturbes par les Araign6es. Nous avons obtenu 958 proies dans 68 toiles d'A. marmoreus et 233 proies dans 57 toiles d'A. bruen- nichi, soit 14,1 proies (o = 8,2)/toile/p6riode de 12 h pour la premiere et 4,3 proies (o = 4,7)/toile/ p6riode de 12 h pour la seconde.
A. marmoreus capture un nombre de proies sig- nificativement plus grand par toile et par pbriode de 12 h qu'A. bruennichi (test Mann et Whitney, z = 7,9 p < 0.001).
Les proies d'A. marmoreus sont plus de 75% de Dipt~res, 16% d'Hym6nopt~res; les autres ordres sont plus faiblement repr6sent6s. Les Orthopt~res et les L6pidopt~res ne sont pas repr6sent6s (Fig. 1A, a).
Le proies de petite taille sont les plus nombreuses (86%) et celles de grande taille sont faiblement repr6sent6es (4,5%) (Fig. 1A, c). Cette espbce cap- ture 0,63 proies de grande taille par toile par p6ri- ode de 12 h.
Les proies d'A. bruennichi sont 60% de Dipt~res, 14,1% d'Hym6nopt~res, 11,7% d'Orthopt~res et 10,6% d'H6mipt~res. Les autres ordres sont trbs faiblement repr6sent6s (Fig. 1 B, a).
Les proies de petite taille sont les plus nom- breuses (55%). Les proies de grande taille sont les moins nombreuses (17,6%) soit 0,72 proie de grande taille par toile par p6riode de 12 h (Fig. 1B, c).
Comparaison entre les deux gammes de proies des deux espbces. Les captures pr6lev6es directement dans les toiles d'A. marmoreus et d'A. bruennichi different par leur composition syst6matique (X 2 = 112,17, ddl = 18, p < 0.001) et par leur taille (X 2 = 86,5, ddl = 6, p < 0.001) (Tableau 1).
En ce qui concerne la composition syst6matique, il y a plus d'H6miptbres (p < 0.001) et d'Orthoptbres (p < 0.001) dans le r6gime alimentaire d'A. bruenni- chi que dans celui d'A. marmoreus. I1 n'y a pas de diff6rences significatives en ce qui concerne les au- tres ordres (Tableau 1).
Pour la taille, il y a moins de proies de petite taille (p < 0.001) mais plus de proies de taille moyenne (p < 0.001) et de grande taille (p < 0.001) dans le r6gime alimentaire d'A. bruennichi que dans celui d'A. marmoreus (Tableau 1).
Les diff6rences entre les gammes de proies captu- r6es par chacune des espbces reflbtent une diff6rence de composition. En effet, pourA, bruennichi, par-
179
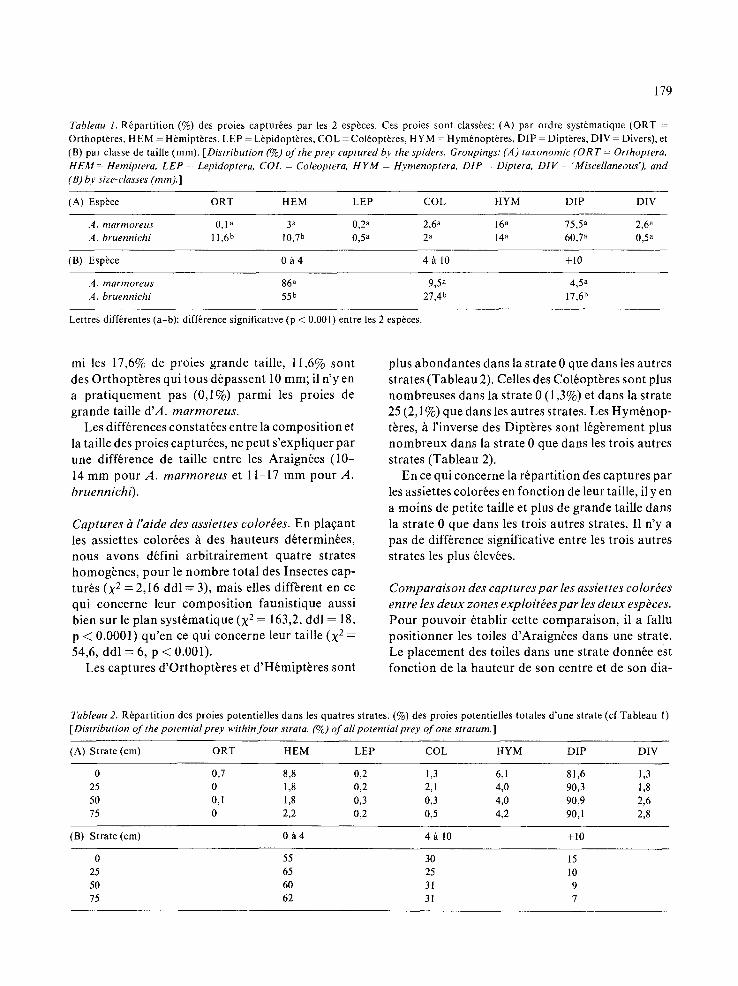
Tableau l. R6partition (%) des proies captur6es par les 2 esp6ces. Ces proies sont class6es: (A) par ordre syst6matique (ORT = Orthoptbres, HEM = H6mipt~res, LEP = L6pidoptbres, COL = Col6opt~res, HYM = Hym6noptbres, DIP = Dipt~res, DIV = Divers), et (B) par classe de taille (mm). [Distribution (%) of the prey captured by the spiders. Groupings: (A) taxonomic (ORT = Orthoptera, HEM = Hemiptera, LEP = Lepidoptera, COL = Coleoptera, HYM = Hymenoptera, DIP = Diptera. DIV = 'Miscellaneous'), and (B) by size-classes (mm).]
(A) Espbce ORT HEM LEP COL HYM DIP DIV
A. marmoreus 0,1 a 3 a 0,2 a 2,6 a 16 a 75,5 a 2,6 a A. bruennichi 11,6 b 10,7 b 0,5 a 2 a 14 a 60,7 a 0,5 a
(B) Espbce 0/l 4 4 ~ 10 +10
A. marmoreus 86 a 9,5 a 4,5 a A. bruennichi 55 b 27,4 b 17,6 b
Lettres diff6rentes (a-b): difference signifieative (p < 0.001) entre les 2 esp~ces.
m i l e s 17,6% de p ro ies g r a n d e tail le, 11,6% son t
des O r t h o p t b r e s qui t ous d6passen t l0 ram; il n 'y en
a p r a t i q u e m e n t pas (0 ,1%) p a r m i les p ro ies de
g r a n d e tai l le d 'A. m a r m o r e u s .
Les d i f f6rences cons ta t6es en t re la c o m p o s i t i o n et
la ta i l le des pro ies captur6es , ne peu t s ' e x p l i q u e r pa r
une d i f f6rence de tai l le en t r e les Ara ign6es ( 1 0 -
14 m m p o u r A. m a r m o r e u s et 11-17 m m p o u r A.
bruennichi ) .
Cap tures h l 'aide des assiet tes colorkes. En p l agan t
les ass ie t tes co lor6es h des h a u t e u r s d6termin6es ,
nous a v o n s d6fini a r b i t r a i r e m e n t q u a t r e s t ra tes
h o m o g ~ n e s , p o u r le n o m b r e to t a l des Insec tes cap -
tur6s (X 2 = 2,16 dd l = 3), mais elles d i f fbrent en ce
qu i c o n c e r n e leur c o m p o s i t i o n f aun i s t i que aussi
b ien sur le p l an sy s t6ma t ique (X 2 = 163,2, dd l = 18,
p < 0.0001) q u ' e n ce qu i c o n c e r n e leur ta i l le (X 2 =
54,6, dd l = 6, p < 0.001).
Les cap tu re s d ' O r t h o p t b r e s et d ' H 6 m i p t ~ r e s son t
plus a b o n d a n t e s dans la s t ra te 0 que dans les au t re s
s t ra tes ( T a b l e a u 2). Cel les des Co l6op t~res son t plus
n o m b r e u s e s dans la s t ra te 0 (1,3%) et dans la s t ra te
25 (2,1%) que dans les au t res strates. Les H y m 6 n o p -
t~res, h r i n v e r s e des Dip t~res son t 16g~rement plus
n o m b r e u x dans la s t ra te 0 que dans les t ro is au t res
s t ra tes ( T a b l e a u 2).
En ce qui c o n c e r n e la r6pa r t i t i on des cap tu res pa r
les ass ie t tes co lor6es en f o n c t i o n de leur tai l le , il y e n
a m o i n s de pe t i te tai l le et plus de g r a n d e tai l le dans
la s t ra te 0 que dans les t ro is au t res s trates. I1 n 'y a
pas de d i f f6rence s ign i f ica t ive en t re les t ro is au t res
s t ra tes les plus 61ev~es.
C o m p a r a i s o n des c a p t u r e s p a r les assiet tes colorbes
entre les d e u x zones explo i tbes p a r les d e u x espbces.
P o u r p o u v o i r 6 tabl i r ce t te c o m p a r a i s o n , il a fa l lu
p o s i t i o n n e r les to i les d ' A r a i g n 6 e s dans une strate .
Le p l a c e m e n t des to i les dans une s t ra te donn6e est
f o n c t i o n de la h a u t e u r de son cent re et de son dia-
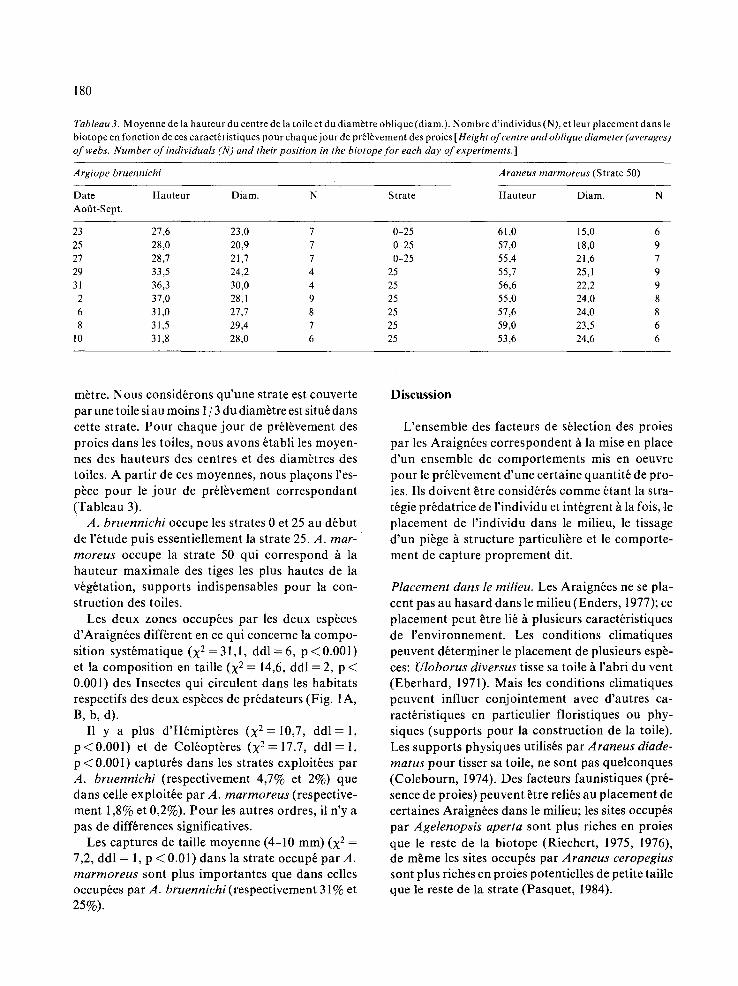
Tableau 2. R6partition des proies potentielles dans les quatres strates. (%) des proies potentieltes totales d'une strate (cf Tableau 1) [Distribution of the potential prey within four strata. (%) of all potential prey of one stratum.]
(A) Strate(cm) ORT HEM LEP COL HYM DIP DIV
0 0,7 8,8 0,2 1,3 6,1 81,6 1,3 25 0 1,8 0,2 2,1 4,0 90,3 1,8 50 0,1 1,8 0,3 0,3 4,0 90,9 2,6 75 0 2,2 0,2 0,5 4,2 90,1 2,8
(B) Strate(cm) 0 h 4 4 h i 0 +10
0 55 30 15 25 65 25 10 50 60 31 9 75 62 31 7
180
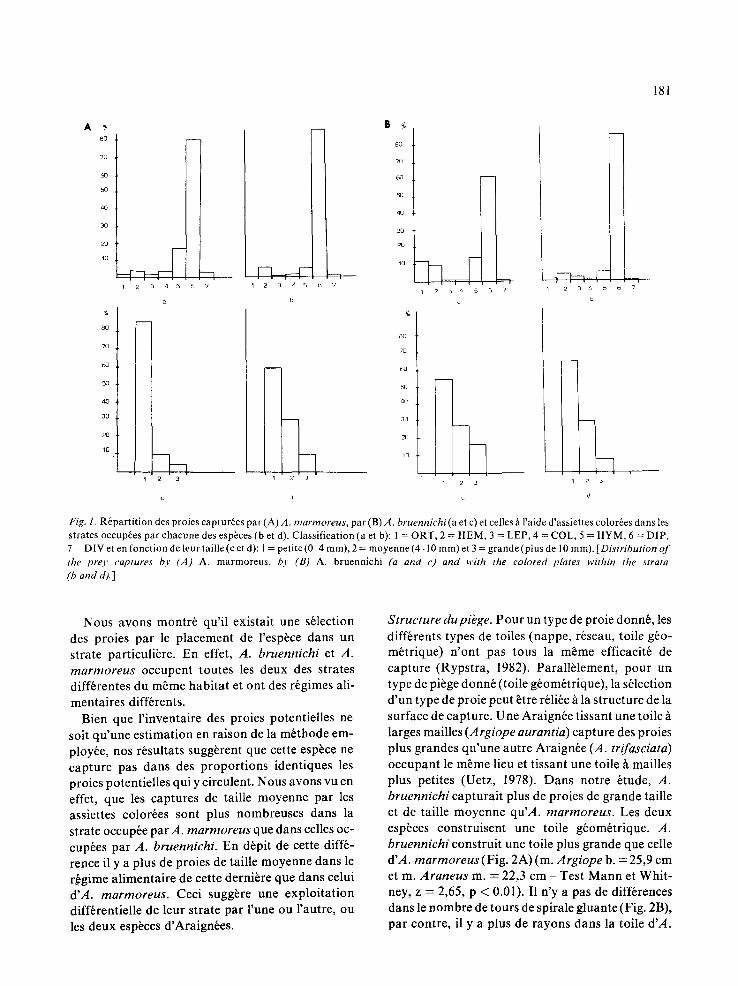
Tableau 3. M oyenne de la hauteur du centre de la toile et du diambtre oblique (diam.). Nombre d'individus (N), et leur placement dans le biotope en fonction de ces caract6ristiques pour chaque j our de pr61bvement des proies [Height of centre and oblique diameter (averages) of webs. Number of individuals (N) and their position in the biotope for each day of experiments.]
Argiope bruennichi Araneus marmoreus (Strate 50)
Date Hauteur Diam. N Strate Hauteur Diam. N Aofit-Sept.
23 27,6 23,0 7 0 25 61,0 15,0 6 25 28,0 20,9 7 0-25 57,0 18,0 9 27 28,7 21,7 7 0-25 55,4 21,6 7 29 33,5 24,2 4 25 55,7 25,1 9 31 36,3 30,0 4 25 56,6 22,2 9 2 37,0 28,1 9 25 55,0 24,0 8 6 31,0 27,7 8 25 57,6 24,0 8 8 31,5 29,4 7 25 59,0 23,5 6
10 31,8 28,0 6 25 53,6 24,6 6
mbtre. Nous consid6rons qu 'une strate est couverte par une toile si au moins 1 / 3 du diam~tre est situ6 darts cette strate. Pour chaque jour de pr6l~vement des proies dans les toiles, nous avons 6tabli les moyen- nes des hauteurs des centres et des diam~tres des toiles. A partir de ces moyennes, nous plagons l'es- p~ce pour le jour de pr6l~vement correspondant (Tableau 3).
A. bruennichi occupe les strates 0 et 25 au d6but de l '6tude puis essentiellement la strate 25. A. mar-
moreus occupe la strate 50 qui correspond h la hauteur maximale des tiges les plus hautes de la v6g6tation, supports indispensables pour la con- struction des toiles.
Les deux zones occup6es par les deux esp~ces d 'Araign6es different en ce qui concerne la compo- sition syst6matique (X 2 = 31,1, ddl = 6, p <0 .001) et la composi t ion en taille (X 2 = 14,6, ddl = 2, p < 0.001) des Insectes qui circulent dans les habitats respectifs des deux esp~ces de pr6dateurs (Fig. 1A, B, b, d).
I1 y a plus d 'H6mipt~res (X 2 = 10,7, d d l = 1, p < 0 . 0 0 1 ) et de ColOoptOres (•2= 17.7, d d l = 1, p <0 .001) capturOs dans les strates exploitOes par A. bruennichi (respectivement 4,7% et 2%) que dans celle exploitOe par A. marmoreus (respective- ment 1,8% et 0,2%). P o u r les autres ordres, il n 'y a pas de diff6rences significatives.
Les captures de taille moyenne (4-10 mm) (X 2 = 7,2, ddl = 1, p < 0.01) dans la strate occup~ par A. marmoreus sont plus importantes que dans celles occup6es par A. bruenniehi (respectivement 31% et 25%).
D i s c u s s i o n
L'ensemble des facteurs de s61ection des proies par les Araign6es correspondent h la mise en place d 'un ensemble de compor tements mis en oeuvre pour le pr6lbvement d 'une certaine quantit6 de pro- ies. Ils doivent ~tre consid6r6s comme 6tant la stra- t6gie pr6datrice de r individu et int6grent h la fois, le placement de l ' individu dans le milieu, le tissage d 'un pi~ge ~. structure particulibre et le comporte- ment de capture proprement dit.
Placement dans le milieu. Les Araign6es ne se pla- cent pas au hasard dans le milieu (Enders, 1977); ce placement peut ~tre li6 ~ plusieurs caract6ristiques de renvironnement . Les conditions climatiques peuvent d6terminer le placement de plusieurs esp~- ces: Uloborus diversus tisse sa toile h l 'abri du vent (Eberhard, 1971). Mais les conditions climatiques peuvent influer conjointement avec d 'autres ca- ract~ristiques en particulier floristiques ou phy- siques (supports pour la construct ion de la toile). Les supports physiques utilis6s par Araneus diade-
matus pour tisser sa toile, ne sont pas quelconques (Colebourn, 1974). Des facteurs faunistiques (pr6- sence de proies) peuvent ~tre reli6s au placement de certaines Araign6es dans le milieu; les sites occup~s par Agelenopsis aperta sont plus riches en proies que le reste de la biotope (Riechert, 1975, 1976), de m~me les sites occup6s par Araneus ceropegius
sont plus riches en proies potentielles de petite taille que le reste de la strate (Pasquet, 1984).
181
80
70
6O
5O
40
3O
20
10
I
80
70
60
5G
40
30
20
10
.__q
1 2 3 a n 7
-h , -!l 1 2 3 1 2 .
c ,J
B
8O
90
60
5O
40
3 0
20
10
80
70
6U
50
40
30
20
10
2 3 4 5 ")
a
1 2 3
2 3 4 5 5
a
1 2 3 ~ 5
b
L I 2 3
I
?
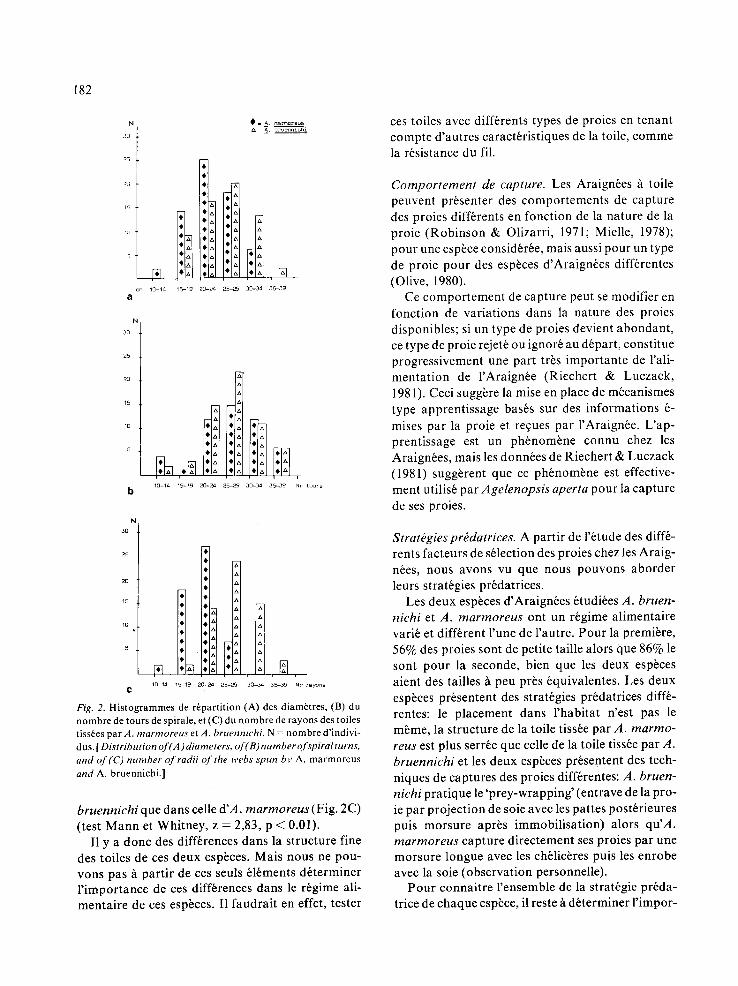
Fig. 1. R6part i t ion des proies capturbes par (A) A. marmoreus, par (B) A. bruennichi (a et c) et celles ~t l'aide d'assiettes color6es dans les strates occup6es par c h a c u n e des espbces (b et d). Class i f icat ion (a et b): 1 = O R T , 2 = H E M , 3 = LEP, 4 = COL, 5 = H Y M , 6 = D I P , 7 = D I V et en fonc t ion de leur tail le (c et d): 1 = petite ( 0 - 4 mm), 2 = m o y e n n e (4 -10 mm) et 3 = grande (plus de I 0 mm). [ Distribution of the prey captures by (A) A. marmoreus , by (B) A. bruennichi (a and e) and with the colored plates within the strata (b and d).]
Nous avons montr6 qu'il existait une s61ect{on des proies par le placement de l'esp~ce dans un strate particuli~re. En effet, A. bruennichi et A. marmoreus occupent toutes les deux des strates diff6rentes du m~me habitat et ont des r6gimes ali- mentaires diff~rents.
Bien que l'inventaire des proies potentielles ne soit qu'une estimation en raison de la m6thode em- ploy6e, nos r6sultats suggbrent que cette esp~ce ne capture pas dans des proportions identiques les proies potentielles qui y circulent. Nous avons vu en effet, que les captures de taille moyenne par les assiettes colorbes sont plus nombreuses dans la strate occup~e par A. marmoreus que dans celles oc- cup6es par A. bruennichi. En d6pit de cette diff6- rence il y a plus de proies de taille moyenne dans le r~gime alimentaire de cette derni~re que dans celui d'A. marmoreus. Ceci sugg~re une exploitation diff6rentielle de leur strate par l'une ou l'autre, ou les deux esp~ces d'Araign6es.
Structure dupibge. Pour un type de proie donn6, les diff6rents types de toiles (nappe, r6seau, toile g6o- m6trique) n'ont pas tous la m~me efficacit~ de capture (Rypstra, 1982). ParallNement, pour un type de pi~ge donn6 (toile g6om6trique), la s6lection d'un type de proie peut btre r61i6e h la structure de la surface de capture. Une Araign6e tissant une toile larges mailles (Argiope aurantia) capture des proies plus grandes qu'une autre Araign6e (A. trifasciata) occupant le m~me lieu et tissant une toile & mailles plus petites (Uetz, 1978). Dans notre 6tude, A. bruennichi capturait plus de proies de grande taille et de taille moyenne qu'A. marmoreus. Les deux esp~ces construisent une toile g6om6trique. A. bruennichi construit une toite plus grande que celle d'A. marmoreus (Fig. 2A) (m. Argiope b. = 25,9 cm et m. Araneus m. = 22,3 cm - Test Mann et Whit- ney, z = 2,65, p < 0.01). I1 n'y a pas de differences dans le nombre de tours de spirale gluante (Fig. 2B), par contre, il y a plus de rayons dans la toile d'A.
182
N
3O
25
2L)
15
lU
5
1 5 - 1 £
= A. bruennichi
.q $1
2 0 - 2 4 2 5 - 2 £ 30-.34 3 5 - 3 9
10 -14 15 -19 20-24 2 5 - 2 9 3 0 - 3 4 3 5 - 3 9 N r [ o u r s
N
3[3
25 ~4
G
15 i 0 ~,
lu l ~
ilD i ~ 5 I'@ a,
10-14 15 -19 2 0 - 2 4 2 5 - 2 9 3 0 - 3 4 3 5 - 3 9 N r r a y o n ~ e
Fig. 2. Histogrammes de r~partition (A) des diam~tres, (B) du nombre de tours de spirale, et (C) du nombre de rayons des toiles tiss6es par A. marmoreus et A. bruenniehi. N = n o m b r e d'indivi- dus. [ Distribution o f (A) diameters, o/'(B) number o f spiral turns, and o f (C) number o f radii o f the webs spun by A. marmoreus and A. bruennichi.]
bruennichi que dans celle d'A. marmoreus (Fig. 2C) (test Mann et Whitney, z = 2,83, p < 0.01).
I1 y a done des diff6rences dans la structure fine des toiles de ces deux esp~ces. Mais nous ne pou- vons pas & partir de ces seuls 616ments d6terminer l ' importance de ces diff6rences dans le r6gime ali- mentaire de ces espbces. I1 faudrait en effet, tester
ces toiles avec diff6rents types de proies en tenant compte d'autres caract6ristiques de la toile, comme la r6sistance du ill.
C o m p o r t e m e n t de capture. Les Araign6es ~ toile peuvent prbsenter des comportements de capture des proies diff6rents en fonction de la nature de la proie (Robinson & Olizarri, 1971; Mielle, 1978); pour une espbce consid6r6e, mais aussi pour un type de proie pour des espbces d'Araign6es diff6rentes (Olive, 1980).
Ce comportement de capture peut se modifier en fonction de variations dans la nature des proies disponibles; si un type de proies devient abondant, ce type de proie rejet6 ou ignor6 au d6part, constitue progressivement une part trbs importante de l'ali- mentation de l'Araign6e (Riechert & Luczack, 1981). Ceci suggbre la mise en place de m6canismes type apprentissage bas6s sur des informations 6- mises par la proie et regues par l'Araign6e. L'ap- prentissage est un ph6nombne connu chez les Araign6es, mais les donn6es de Riechert & Luczack (1981) sugg~rent que ce ph6nom~ne est effective- ment utilis6 par Agelenops is aperta pour la capture de ses proies.
Stratkgiesprkdatrices . A partir de l'6tude des diffb- rents facteurs de s61ection des proies chez les Araig- nbes, nous avons vu que nous pouvons aborder leurs strat6gies pr6datrices.
Les deux espbces d'Araign6es 6tudi6es A. bruen- nichi et A. marmoreus ont un r6gime alimentaire vari6 et diff6rent l'une de l'autre. Pour la premibre, 56% des proies sont de petite taille alors que 86% le sont pour la seconde, bien que les deux esp~ces aient des tailles & peu pros 6quivalentes. Les deux espbces pr6sentent des strat6gies pr6datrices diff6- rentes: le placement dans l 'habitat n'est pas le m~me, la structure de la toile tiss6e par A. marmo- reus est plus serr6e que celle de la toile tiss6e par A. bruennichi et les deux esp~ces pr6sentent des tech- niques de captures des proies diff6rentes: A. bruen- nichi pratique le 'prey-wrapping' (entrave de la pro- ie par projection de soie avec les pattes post6rieures puis morsure apr~s immobilisation) alors qu'A. marmoreus capture directement ses proies par une morsure longue avec les ch61ic~res puis les enrobe avec la soie (observation personnelle).
Pour connaitre l 'ensemble de la strat6gie pr6da- trice de chaque esp~ce, il reste & d6terminer l ' impor-

t ance de c h a c u n de ces p h 6 n o m b n e s dans la s61ec-
t ion des proies pa r c h a c u n e des d e u x espbces.
Remerciements
Je r emerc i e les P ro fe s seu r s B. Kraf f t et V. L a b e y r i e
p o u r leurs c r i t iques cons t ruc t ives .
Summary
Prey and preda tory strategies o f two orb-weaving spiders: A r g i o p e b r u e n n i c h i (Scop. ) and A r a n e u s
m a r m o r e u s (Clark), Araneae, Argiopidae
T h e i m p a c t o f spiders on insect prey p o p u l a t i o n s
d e p e n d s on the n u m b e r o f prey c a u g h t and on p rey
se lect ion. Se l ec t i on depends on m a n y fac tors , bu t
p a r t i c u l a r l y on the p o s i t i o n o f the web in the hab i -
tat.
T h e die t o f t w o species o f sp iders was s tud ied ,
t o g e t h e r wi th the p o t e n t i a l p rey f r o m the s a m e hab-
itat, d e t e r m i n e d by red and ye l low c o l o u r e d t raps .
A. marmoreus c a u g h t an ave r age o f 14.1 p r ey /
day , whi le A. bruennichi c a u g h t an a v e r a g e o f 4.3
p r e y / d a y . T h e p rey o f A. marmoreus were m o s t l y
(86%) smal l insects (0 -4 ram) cons i s t ing m a i n l y o f
D i p t e r a (75%) and H y m e n o p t e r a (16%). A bruen-
nichi c a u g h t D i p t e r a (60%) and H y m e n o p t e r a
(14.1%) but also O r t h o p t e r a (11.7%) and H e m i p t e -
ra (10.6%) wi th fewer smal l insects (55%) and m o r e
m e d i u m - s i z e d (4 -10 m m ) (28.4%) and large p rey
( > 10 m m ) (17.6%).
A. m a r m o r e u s spins its web at the t o p o f the
v e g e t a t i o n (50 to 75 cm) and A. bruennichi spins
l ower (0 -50 cm), a m o n g s t the grasses. Ana lys i s o f
the c o l o u r e d t rap ca tches f r o m these d i f fe ren t s t ra ta
shows tha t p o s i t i o n o f the web in the hab i t a t ac-
c o u n t s pa r t ly fo r the d i f f e r en t diets o f the two spe-
cies. S t r u c t u r a l charac te r i s t i c s o f the webs and the
p r e d a t o r y b e h a v i o u r o f the two species are also
cons ide red .
References
Colebourn, P. H., 1974. The influence of habitat structure on the distribution of Araneus diadematus (Clerk). J. Anim. Ecol. 43:401 410.
183
Eberhard, W. G., 1971. The ecology of the web of Uloborus diversus (Araneae, Uloboridae). Oecologia 6:328 342.
Enders, F., 1977. Web-site selection by orb-web spiders particu- larly Argiope aurantia (Lucas). Anim. Behav. 25: 694-712.
Kajak, A., 1965a. An analysis of food relations between spiders Araneus cornutus (Clerk) and Araneus quadratus (Clerk) and their prey in a meadow. Ekol. Polska A, 13:717 764.
Kajak, A., 1965b. Quantitative analysis of relations between spi- ders (Araneus cornutus (Clerk) and Araneus quadratus (Clerk)), and their prey in meadow. Bull. Acad. Sci. Pol. II, 13: 515-522.
Mielle, D., 1978. Contribution ~ l'6tude du comportement pr~- dateur et des mbcanismes de tol6rance dans le genre Tege- naria (Araneae, Agelenidae). Th~se de 3~me Cycle. Univer- sit6 de Nancy I.
Nentwig, W., 1980. The selective prey of linyphiid-like spiders and their spacewebs. Oecologia 45: 236-243.
Nentwig, W., 1982. Why do only certain insects escape from a spider's web. Oecologia 53: 412-417.
Nentwig, W., 1983. The prey of web-building spiders compared with feeding experiments (Araneae, Araneidae, Linyphiidae, Pholcidae, Agelenidae). Oecologia 56: 132-139.
Nyffeler, M., 1982. Field studies on the ecological role of the spiders as Insect predators in agrosystems (Abandonned grassland, meadows and cereal fields). Th~se Doet. Z~irich 1982:1 174.
Nyffeler, M. & G. Benz, 1978. Die Beutespektren des Netz- spinnen Argiope bruennichi (Scopoli), Araneus quadratus (Clerk), und Agelena labyrinthica (Clerk) in Odlandwiesen bei Zfirich. Rev. Suisse Zool. 85 (4): 747 757.
Nyffeler, M. & G. Benz, 1979. Studies on the ecological impor- tance of spiders population for the vegetation of cereal and rape fields. J. Ang. Entomol. 87: 348-376.
Olive, C. W., 1980. Foraging specializations in orb-weaving spiders. Ecology61:1133 1144.
Pasquet, A., 1982. Placement et s+lection des proies chez les Araign6es d'une prairie. Rev. Arach. 4: 157-165.
Pasquet, A., 1984. Predatory site selection and adaptation of the trap in four orb-weaving spiders. Biol. Bebav. 9:3 19.
Pasquet, A. & B. Krafft, 1980. Relations entre la v6g6tation, la r6partition des proies potentielles et les sites de construction des toiles par les Araign6es. Proc. Int. Aracb. Congress 8th: 145-150.
Riechert, S. E., 1975. The pattern of local distribution in a desert spider: mechanisms and seasonal variation. J. Anim. Ecol. 43: 733-746.
Riechert, S. E., 1976. Web-site selection in the desert spider Agelenopsis aperta. Oikos 27:311 315.
Riechert, S. E. & J. Luczack, 1981. Spider foraging: behavioral responses to prey. In: P. N. Witt & J. S. Rovner (eds.), Spiders Communication. Princeton University Press, Prince- ton, NJ.
Robinson, M. H. & B. Robinson, 1970. Prey caught by a popula- tion of the spider Argiope argentata (Araneae, Araneidae) in Panama. A year's census data. Zool. J. Linn. Soc. 49: 345-357.
Robinson, M. H. & J. Olazarri, 1971. Units of behavior and complex sequences in the predatory behavior of Argiope argentata (Fabricius) (Araneae, Araneidae). Smithsonian Cont. Zool. 65:1 36.

184
Roth, M. & G. Couturier, 1966. Les plateaux color6s en 6cologie entomologique. Ann. Soc. Entomo!. (N.S.) 2 (2): 361 370.
Rypstra, A. L., 1982. Building a better Insect trap; an experi- mental investigation of prey capture in a variety of spider webs. Oecologia 52:31 36.
Schaefer, M., 1974. Experimentelle Untersuchungen zur Be- deutung der interspezifischen Konkurrenz bei 3 Wolspinnen- Arten (Araneida, Lycosidae) einer Salzwiese. Zool. Jb. Syst. 101:213 235.
Turnbull, A. L., 1960. The prey of the spider Linyphia triangula-
ris (Clerk) Araneae, Linyphiidae. Can. J. Zool. 38: 859-873. Turner, M., 1979. Diet and feeding phenology of the green Lynx
spider Peucetia viridens (Araneae, Oxyopidae). J. Arach. 7: 149-154.
Uetz, W. G., 1978. Web placement, web structure and prey capture in orb-weaving spiders. Bull. Brit. Arach. Soc. 4 (4): 141-148.
Accept6 le 17 Mai, 1984.


























