Embed Size (px)
Citation preview

JEMMAL Sarah
BASCUÑANA Céline
Projet de recherche :
Étude de l’activité transcriptionnelle
des gènes var subtélomériques au
niveau des pores nucléaires chez
Plasmodium falciparum
NIETO Laurence
DEMANGE Pascal Année universitaire : 2006-2007

2
Résumé
Notre projet de recherche consiste à étudier l’activité transcriptionnelle des gènes var
subtélomériques au niveau des pores nucléaires chez Plasmodium falciparum. Ces gènes
codent pour des protéines de surface responsables des variations antigéniques du parasite lui
permettant d’échapper au système immunitaire de l’hôte. Plasmodium falciparum est un
organisme eucaryote unicellulaire haploïde qui est un agent étiologique du paludisme qui
cause encore de nos jours plus d’un million de morts par an.
De telles études ont déjà été réalisées chez la levure à l’aide de techniques permettant
de mettre en évidence in situ la colocalisation d’un locus particulier avec les pores nucléaires.
Plusieurs étapes sont indispensables pour parvenir à ce résultat. Il faut, dans un premier
temps, marquer le locus du gène var avec une répétition d’opérateurs lac qui sera reconnue
par la protéine répresseur LacI fusionnée à la GFP. En parallèle, il faut marquer le pore
nucléaire et plus précisément une nucléoporine en la fusionnant à la BFP. Une fois ces
constructions réalisées, la colocalisation du locus d’intérêt et des pores nucléaires pourra être
visualisée au microscope à épifluorescence par la technique du FRET correspondant à un
transfert de fluorescence entre la BFP et la GFP et donc entre le pore nucléaire et le locus. Des
expériences utilisant différentes variantes du FISH permettront ensuite d’étudier l’état
transcriptionnel du gène var.
Cette étude des mécanismes de régulation des gènes var pourrait contribuer à la
recherche de cibles pour le développement de nouvelles thérapies anti-parasitaires.

3
Tables des abréviations
3C : Capture de la Conformation des Chromosomes
Blast : Basic Local Alignment Search Tool
bsd : blasticidine désaminase
BFP : Blue Fluorescent Protein
CAT : Chloramphénicol AcétylTransférase
CFP : Cyan Fluorescent Protein
CT : Territoire Chromosomique
CTD : Domaine CarboxyTerminal
DHFR : DiHydroFolate Réductase
FISH : Fluorescent In Situ Hybridization
FRET : Fluorescent Resonance Energy Transfer
GFP : Green Fluorescent Protein
HXK1 : hexokinase isoenzyme 1
IC : régions InterChromatiques
IC50 : Concentration Inhibitrice 50
NLS : Nuclear Localization Signal
NPC : Nuclear Pore Complexe
Nup : nucléoporine
PCR : Polymerase Chain Reaction
PfEMP1 : Plasmodium falciparum erythrocyte membrane protein 1
qRT-PCR : quantitative Real-Time Polymerase Chain Reaction
SIR : Silent Information Regulatory
TARE : Telomere Associated Repeat Elements

4
Sommaire
Résumé ....................................................................................................................................... 2 Tables des abréviations .............................................................................................................. 3 Sommaire ................................................................................................................................... 4
I- INTRODUCTION....................................................................................................... 5 I.1- L’organisation nucléaire chez les eucaryotes..................................................... 5
I.1.1- Les territoires chromosomiques et les régions interchromatiques........................... 5 I.1.2- Les machineries de transcription .............................................................................. 6 I.1.3- Les pores nucléaires.................................................................................................. 7
I.2- Plasmodium falciparum et la famille de gènes var ........................................... 8
II- RÉSULTATS............................................................................................................. 11 II.1- Localisation in vivo de séquences d’ADN et visualisation de l’organisation de la chromatine à grande échelle en utilisant une reconnaissance répresseur/opérateur Lac. Robinett et al 1996. .............................................................................................................. 11 II.2- L’association au pore nucléaire confère des niveaux d’expression optimum pour un gène inductible de levure. Taddei et al 2006. ...................................................................... 12 II.3- La répression de l’hétérochromatine et le repositionnement de locus sont liés à la régulation de gènes de virulence chez Plasmodium falciparum. Duraisingh et al 2005. .... 15
III- PROJET DE RECHERCHE:............................................................................ 19 Étude de l’activité transcriptionnelle des gènes var subtélomériques au niveau des pores nucléaires chez Plasmodium falciparum.................................................................................. 19
III.1- Synthèse de la répétition en tandem d’opérateurs lac................................................. 20 III.2- Insertion dans un plasmide de cette répétition en amont d’un gène d’intérêt............. 20 III.3- Transfection stable de la séquence répétée en tandem d’opérateurs lac dans le génome de Plasmodium falciparum par recombinaison homologue................................................. 21 III.4- Expression transitoire de protéines de fusion ............................................................. 21
III.4.1- Répresseur-LacI-NLS-GFP.................................................................................. 21 III.4.2- Nucléoporine de Plasmodium falciparum ........................................................... 22
III.5- Expériences de FRET ................................................................................................. 22 III.6- Expériences de FISH................................................................................................... 23
IV- CONCLUSION ET PERSPECTIVES........................................................ 24
BIBLIOGRAPHIE......................................................................................................... 25

5
INTRODUCTION
I.1- L’organisation nucléaire chez les eucaryotes
Le cytoplasme est compartimenté en organites bien définis tels que l’appareil de golgi,
le réticulum endoplasmique, les mitochondries, les lysosomes, les peroxysomes qui sont
caractérisés par des fonctions bien précises. Contrairement au cytoplasme, le noyau ne
comporte pas de compartiments physiquement individualisés. Cependant, il existe des régions
nucléaires dédiées à certaines fonctions, la plus étudiée étant le nucléole. Le nucléole est un
composé d’une auto-agrégation des ADN codants pour les ARN ribosomiques auxquels
s’ajoutent un certain nombre de protéines qui interviennent dans ce processus.
I.1.1- Les territoires chromosomiques et les régions interchromatiques
Un autre constituant capital du noyau est la chromatine. Elle adopte une conformation
spécifique dans le temps et dans l’espace ce qui confère aux gènes un état transcriptionnel
plus ou moins actif. Des mécanismes épigénétiques tels que la méthylation de l’ADN, la
modification des histones et le remodelage de la chromatine participent à cette régulation des
gènes. Ainsi pour un gène, une colocalisation avec l’hétérochromatine péricentriolaire,
subtélomérique ou dans la périphérie nucléaire est généralement synonyme de répression alors
qu’une localisation plus interne dans le noyau (euchromatine) se traduit par une activité
transcriptionnelle.
L’ADN interphasique a longtemps été considéré comme un amas aléatoirement
disposé dans le noyau mais les techniques d’hybridations in situ par fluorescence (FISH) ont
pu permettre de visualiser une organisation territoriale particulière de l’ADN lors de
l’interphase. Le premier à suggérer une organisation territoriale des chromosomes est Boveri
en 1909. Dans les années 1980, Cremer et al. ont démontré, par des expériences de micro-
irradiations au laser UV, l’existence de structures spécifiques de chromatine : les territoires
chromosomiques (CT). La relation des gènes avec les territoires chromosomiques est
hautement corrélée à leur expression : les gènes actifs sont loin des CT, dans des régions
interchromatiques (IC), alors que les gènes inactifs se retrouvent au sein des CT ou à leur
surface.
En 1999, l’équipe de Croft a prouvé que deux chromosomes de tailles similaires ont
une position différente en fonction de leur contenance en gènes, en prenant l’exemple des
chromosomes 18 et 19. En effet, ils ont pu remarquer que le chromosome 18, pauvre en gènes

6
actifs, se retrouvait à la périphérie du noyau alors que le chromosome 19, riche en gènes
actifs, avait une position plus centrale.
I.1.2- Les machineries de transcription
Les ARN polymérases II peuvent être détectées par immunoFISH à l’aide d’anticorps
couplés à des fluorophores dirigés contre les ARN polymérases II en cours de transcription
c’est-à-dire hyperphosphorylées sur leur domaine carboxyterminal (CTD). Ainsi, il a pu être
mis en évidence que la transcription des gènes s’effectuait dans certaines zones nucléaires
restreintes où tous les éléments nécessaires (facteurs de transcription,…) sont réunis : il s’agit
des machineries de transcription.
L’équipe d’Osborne en 2004 a voulu mettre en évidence la colocalisation d’allèles
transcrits chez la souris en utilisant la technique des 3C ou Capture de la Conformation des
Chromosomes. Cette technique consiste à fixer des cellules dans le formaldéhyde ce qui
entraîne la formation de pontages, à digérer par une enzyme de restriction, à favoriser les
ligations intramoléculaires, à enlever les pontages précédemment formés et enfin à purifier
l’ADN avant de le détecter par PCR.
Territoires chromosomiques(CT)
Régions Interchromatiques
Figure n°1 : Représentation schématique de l’organisation territoriale nucléaire des chromosomes
Fraser et al. 2006.
Machinerie de transcription (regroupement des composants nécessaires
à la transcription)
Interactions régulatrices
Figure n°2 : Représentation de la méthodologie 3C : Étapes : Pontage au formaldéhyde ; digestion enzymatique ; ligation intramoléculaire ; réversion du pontage suivi détection des produis de ligation par PCR. NB : les astérisques montrent les sites de restriction nouvellement formés
Dekker et al,2002

7
Le principe de l’étude d’Osborne chez la souris était de détecter par PCR les produits
de ligation entre Hbb-b1, gène de la globine, et différents fragments de restriction dans deux
types cellulaires (cellules érythroïdes et cellules du cerveau). Le temps de fixation nécessaire
au pontage entre Hbb-b1 et le gène étudié corrélait en fait avec la fréquence d’expression de
ce gène sachant que Hbb-b1 est un gène hautement exprimé. Ces résultats appuient le modèle
d’association dynamique de gènes avec les machineries transcriptionnelles illustré par la
figure n°1.
I.1.3- Les pores nucléaires
Les pores nucléaires sont des structures macromoléculaires complexes ancrées dans
l’enveloppe nucléaire. C’est au niveau de ces structures, représentées sur la figure 3, que se
fait la totalité des échanges bidirectionnels entre le cytoplasme et le noyau dans les cellules
eucaryotes. Les études réalisées par l’équipe de Belgareh en 2001 ont permis de caractériser,
dans les cellules humaines, les différents constituants (nucléoporines) du complexe Nup107,
présent au niveau des pores nucléaires, déjà identifiés chez la levure S. pombe. Au cours de la
mitose, il y a désorganisation de ces
composants sous forme de sous-complexes
dispersés dans le nucléoplasme. Certaines
nucléoporines (Nup107 et Nup133) vont
alors s’associer aux kinétochores qui sont des
structures assemblées au niveau des
centromères permettant la fixation des
chromosomes aux microtubules. Ceci
pourrait permettre la régulation coordonnée
de deux processus essentiels du cycle
cellulaire : le désassemblage et la
restructuration de l’enveloppe et des pores
nucléaires en mitose et la ségrégation
correcte des chromosomes. (Belgareh et al,
2001).Drummond et al . 2006
Figure n°3 : Représentation schématique de l’organisation d’un pore nucléaire de vertébré Légende : schéma de dessus : filament interne , filament cytoplasmique, particule cytoplasmique, anneaux « thin », « star », « spoke » ,canal transporteur central ; schéma de dessous : anneau nucléoplasmique, filament interne, filament du panier, anneau distal.

8
Il a également été montré que l’association de gènes à ces complexes de pores
nucléaires entraîne leur transcription (Casolari et al. 2004).
I.2- Plasmodium falciparum et la famille de gènes var
Plasmodium falciparum est l’agent étiologique du paludisme. Son invasion est initiée
par une interaction avec la cellule hôte par l’intermédiaire de protéines de surface. Il se
développe de façon indépendante dans le globule rouge et modifie cette cellule de sorte à en
tirer ses nutriments. C’est un parasite unicellulaire et haploïde. Sa caractéristique majeure est
d’être constamment capable de varier son expression génique pour produire des formes
phénotypiques différentes et ainsi contourner les réponses immunitaires de l’hôte. Le
séquençage du génome de Plasmodium falciparum achevé en 2002 (Gardner et al, 2002).
constitue un intérêt majeur de l’utilisation de cet organisme dans les études. Le génome du
clone 3D7 de Plasmodium falciparum est composé de 22,8 mégabases (Mb) réparties en 14
chromosomes dont la taille varie de 0,643 à 3,29Mb. La composition en nucléotides présente
une forte richesse en adénines (A) et en thymines (T), en moyenne de 80,6% du génome voire
plus de 90% pour les régions intergéniques et les introns. La taille moyenne des gènes de
Plasmodium falciparum est de 2300 pb sans les introns présents à 54%. Plasmodium
falciparum possède une machinerie typique des eucaryotes capable d’engendrer les
modifications et le remodelage de la chromatine pour contrôler la régulation
transcriptionnelle.
Des aperçus de la fonction des regroupements de télomères en « clusters » sont fournis
par des pathogènes tels que Plasmodium falciparum, Trypanosoma brucei, ou Candida
glabrata. Chez Plasmodium falciparum notamment, les télomères sont groupés au niveau de
l’enveloppe nucléaire, comme chez la levure de bière. De plus, les protéines de la levure SIR
(Silent Information Regulatory) orthologues ont été identifiées chez Plasmodium falciparum
augmentant la probabilité d’avoir des voies conservées de regroupements et d’ancrage de
télomères. Dans ce pathogène, le regroupement des télomères est clairement lié à sa virulence.
Cela implique des mécanismes épigénétiques d’exclusion allélique pour permettre une
variation antigénique des protéines de surface. Ces protéines de surface, Plasmodium
falciparum erythrocyte membrane protein 1, PfEMP1, sont codées par une famille de gènes,
appelés var, et jouent un rôle dans l’adhésion aux cellules hôtes. Il existe cinquante neuf
copies de gènes var dans le génome mais seule une copie est active au niveau transcriptionnel,
toutes les autres étant réprimées.

9
Ces gènes var représentent une grande partie des gènes situés à proximité des
télomères dans les régions subtélomériques des 14 chromosomes de Plasmodium falciparum
plus précisément localisés en aval de la séquence TARE6 (Telomere Associated Repeat
Elements). Ces séquences sont voisines des gènes rifin et stevor et aussi des gènes var
supplémentaires (cf. Figure n°4).
Figure n°4 :Modèle d’organisation des chromosomes chez Plasmodium falciparum
Légende : (a) Génome haploïde organisé en 14 chromosomes linéaires. Chaque chromosome est composé d’une région interne dans laquelle les gènes de ménage sont localisés et d’une extrémité chromosomique montrant une haute organisation de l’ADN commune à tous les chromosomes. (b) L’extrémité du bras gauche du chromosome 3 est schématiquement représenté comme un exemple d’organisation d’extrémité chromosomique.
Le regroupement des différents gènes var au niveau de foci, correspondant aux foci de
télomères permet d’accroître l’efficacité de recombinaison entre de tels gènes, augmentant
ainsi la virulence du parasite.
Figure n°5 : Modèle schématique du cluster de télomères de Plasmodium falciparum
Ce modèle est déterminé par l’alignement physique de régions subtélomériques hétérologues dans lesquelles le
taux de recombinaison augmente.
Scherf et al 2001
Scherf et al 2001

10
En 2002, Figueiredo et son équipe ont montré que le regroupement des télomères de
Plasmodium falciparum dépendait d’éléments subtélomériques répétés.
Les gènes var sont contrôlés par trois principaux types de promoteur définis comme
upsA, B et C (Gardner et al, 2002). Les gènes régulés par les promoteurs upsA et upsB sont
généralement localisés dans les régions subtélomériques mais parfois certains sont présents
dans les régions centrales. Les promoteurs des gènes var type upsC sont exclusivement
retrouvés dans les clusters des chromosomes centraux.

11
II- RÉSULTATS
II.1 - Localisation in vivo de séquences d’ADN et visualisation de l’organisation de la
chromatine à grande échelle en utilisant une reconnaissance répresseur/opérateur Lac.
Robinett et al 1996.
Robinett et son équipe en 1996 ont développé une nouvelle méthode de localisation in
situ de séquences d’ADN qui permet d’effectuer des observations directes in vivo tout en
préservant l’ultrastructure chromosomique et nucléaire. Cette méthode est basée sur la
reconnaissance d’une séquence répétée en tandem de 256 opérateurs lac qui est ajoutée au
vecteur utilisé pour la transfection comme le montre la figure n°6. Cette reconnaissance se fait
par le répresseur Lac I étiqueté à la GFP à l’aide d’un microscope à épifluorescence.
Figure n°6 : Construction d’une répétition d’opérateurs lac dans un vecteur
Légende : E1, E2 et E3 sont trois enzymes de restriction différentes. Les extrémités générées par E1 et E3 sont compatibles.
Dans son étude, Robinett a utilisé comme enzyme Sal I pour E1, Bam HI pour E2 et Xho I pour E3 ; le plasmide utilisé est
dérivé de pUC18. Plasmide Lacop*1= plasmide avec une seule copie de l’opérateur lac ; Lacop*2 = plasmide avec 2 copies de
l’opérateur lac. Au bout de n cycles on obtient 2n copies d’opérateurs lac dans le plasmide.
Dans cette méthode, une unité de base est construite et contient l’opérateur lac ainsi
que trois sites de restriction correspondant à trois enzymes de restriction différentes. Cette
unité va être insérée dans un plasmide préalablement digéré par Sal I et Bam HI. Ce plasmide
nouvellement formé va subir de nouvelles digestions : celle par Sal I et Bam HI va permettre
Plasmidelacop
*1Ligation
E1 E2E3
Digestion par E1 et E2Digestion par E2 et E3
Plasmide
lacop*1
Plasmidelacop
*2
Ligation
*
Opérateur lac = unité de base
E1 E2E3
Plasmide digéré
par E1 et E2
n fois
���� 2n copies de lacop
Plasmidelacop
*1Ligation Plasmide
lacop*1
Ligation Plasmidelacop
*1Ligation
E1 E2E3
Digestion par E1 et E2Digestion par E2 et E3
Plasmide
lacop*1
Plasmidelacop
*2
Ligation
*
Opérateur lac = unité de base
E1 E2E3
Plasmide digéré
par E1 et E2
n fois
���� 2n copies de lacop
E1 E2E3
Digestion par E1 et E2Digestion par E2 et E3
Plasmide
lacop*1
E1 E2E3
Digestion par E1 et E2Digestion par E2 et E3
Plasmide
lacop*1
Plasmide
lacop*1
Plasmidelacop
*2
Ligation
*Plasmidelacop
*2
Ligation
Plasmidelacop
*2
Ligation
*
Opérateur lac = unité de base
E1 E2E3
Plasmide digéré
par E1 et E2
n fois
���� 2n copies de lacop
Opérateur lac = unité de base
E1 E2E3
Plasmide digéré
par E1 et E2
Opérateur lac = unité de base
E1 E2E3
Plasmide digéré
par E1 et E2
Plasmide digéré
par E1 et E2
n fois
���� 2n copies de lacop
n fois
���� 2n copies de lacop

12
de fournir des unités de base tandis que celle par Bam HI et Xho I va permettre une nouvelle
insertion d’une unité de base. Les extrémités générées par Sal I et Xho I étant compatibles, on
va avoir insertion de l’unité de base sans formation d’un site de restriction. Grâce à cela, on va
pouvoir renouveler huit fois les étapes de digestion et insertion et ainsi obtenir une séquence
répétée en tandem de 256 (28) opérateurs lac sur notre vecteur de départ. Cette séquence va
être reconnue par le répresseur lac qui est fusionné à la protéine GFP ou Green Fluorescent
Protein pour pouvoir être détecté par microscopie à épifluorescence comme l’illustre la figure
n°7. Ainsi on va localiser notre gène d’intérêt puisque celui-ci est positionné en aval de la
répétition.
Figure n°7 : Localisation d’un gène d’intérêt par la protéine de fusion répresseur LacI/GFP
Plusieurs applications de cette technique ont été testées par Robinett. Ainsi des copies
simples et multiples du vecteur intégré ont pu être détectées dans les cellules d’ovaire de
hamster (CHO) vivantes avant amplification du gène d’intérêt. De plus, la détection d’une
seule répétition de 256 opérateurs Lac et sa stabilité pendant la mitose ont été démontrées par
son insertion ciblée dans le génome de levures de bière par recombinaison homologue.
II.2- L’association au pore nucléaire confère des niveaux d’expression optimum pour un
gène inductible de levure. Taddei et al 2006.
Cette technique basée sur la reconnaissance répresseur/opérateur lac a également été
utilisée en 2006 par l’équipe de Taddei pour localiser un gène subtélomérique de levure de
bière. En effet, plusieurs études ont montré que les changements dans l’organisation
tridimensionnelle du génome s’accompagne de changements dans l’expression des gènes.
Chez la levure, la chromatine transcriptionnellement silencieuse, incluant notamment les
Gène d’intérêtRépétition de 256
opérateurs lac
exon exonexon
Protéine de fusion LacI répresseur/ GFP
Gène d’intérêtRépétition de 256
opérateurs lac
exon exonexon
Protéine de fusion LacI répresseur/ GFP
13
télomères, se lie à l’enveloppe nucléaire alors que la transcription de certains gènes a lieu à
cette même périphérie nucléaire. Selon une hypothèse, la « gene gating hypothesis » de
Blobel , le gène serait capturé par le pore nucléaire où il se ferait transcrire permettant ainsi un
export rapide vers le cytoplasme de ces transcrits. L’équipe de Taddei s’est donc intéressé au
positionnement périnucléaire des gènes et plus particulièrement aux niveaux d’expression
qu’il entraînait. Leur étude a consisté à tester si l’activation transcriptionnelle d’un gène
subtélomérique de levure de bière, HXK1 ou hexokinase isoenzyme 1, conduisait à sa
relocalisation aux pores nucléaires. Pour cela, ils ont utilisé la technique de Robinett en
insérant en amont du promoteur de HXK1 une répétition d’opérateurs lac. Parallèlement, ils
ont exprimé des protéines de fusion dans les levures de bière : une protéine répresseur lacI-
GFP afin de suivre le positionnement du gène HXK1 comme expliqué précédemment et une
nucléoporine, la Nup49, étiquetté avec la GFP afin de localiser les pores nucléaires et donc
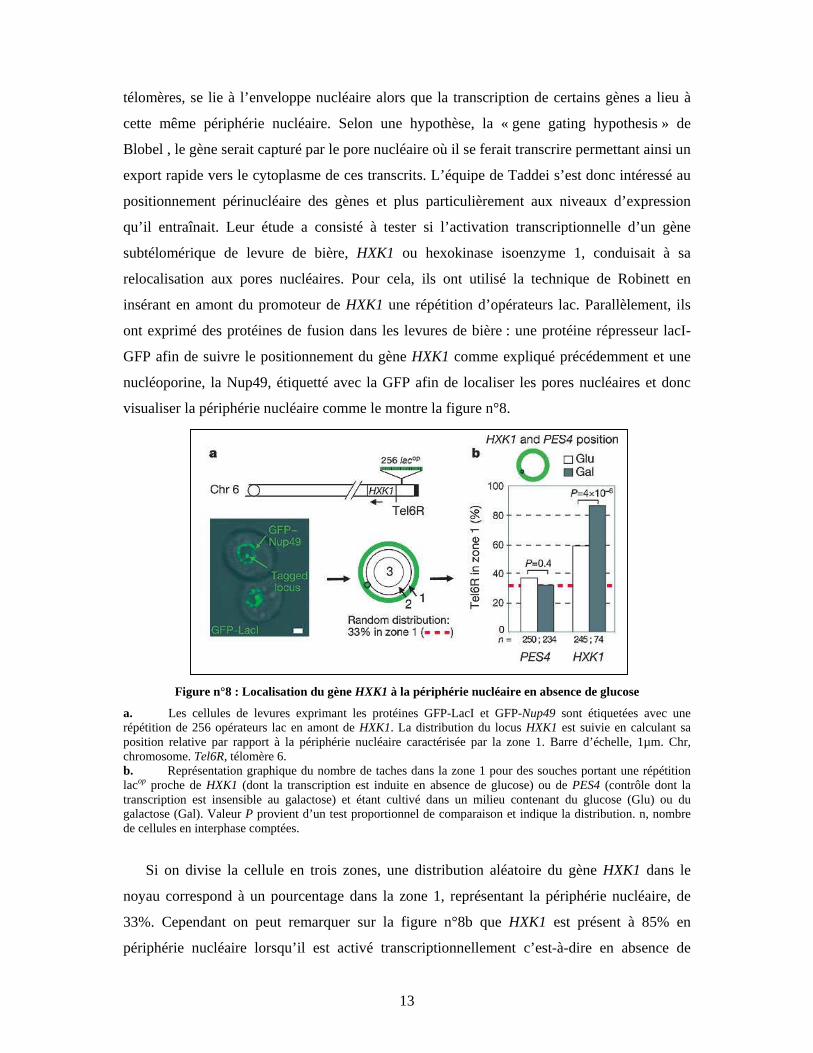
visualiser la périphérie nucléaire comme le montre la figure n°8.
Figure n°8 : Localisation du gène HXK1 à la périphérie nucléaire en absence de glucose
a. Les cellules de levures exprimant les protéines GFP-LacI et GFP-Nup49 sont étiquetées avec une répétition de 256 opérateurs lac en amont de HXK1. La distribution du locus HXK1 est suivie en calculant sa position relative par rapport à la périphérie nucléaire caractérisée par la zone 1. Barre d’échelle, 1µm. Chr, chromosome. Tel6R, télomère 6. b. Représentation graphique du nombre de taches dans la zone 1 pour des souches portant une répétition lacop proche de HXK1 (dont la transcription est induite en absence de glucose) ou de PES4 (contrôle dont la transcription est insensible au galactose) et étant cultivé dans un milieu contenant du glucose (Glu) ou du galactose (Gal). Valeur P provient d’un test proportionnel de comparaison et indique la distribution. n, nombre de cellules en interphase comptées.
Si on divise la cellule en trois zones, une distribution aléatoire du gène HXK1 dans le
noyau correspond à un pourcentage dans la zone 1, représentant la périphérie nucléaire, de
33%. Cependant on peut remarquer sur la figure n°8b que HXK1 est présent à 85% en
périphérie nucléaire lorsqu’il est activé transcriptionnellement c’est-à-dire en absence de
14
glucose. Des expériences complémentaires mesurant le niveau d’expression du gène par
analyse de ces transcrits par qRT-PCR confirment cette observation et amènent donc à
affirmer que l’ancrage du gène HXK1 à la périphérie nucléaire est induite par l’activation du
gène.
A partir de ces premiers résultats Taddei a cherché à savoir si l’ancrage du gène à la
périphérie nucléaire se faisait au niveau de complexes de pores nucléaires (NPC). Elle a pour
cela utilisé une souche délétée de Nup133 dans laquelle le regroupement des pores nucléaires
est étiqueté avec la CFP (Cyan Fluorescent Protein). Les résultats de son expérience sont
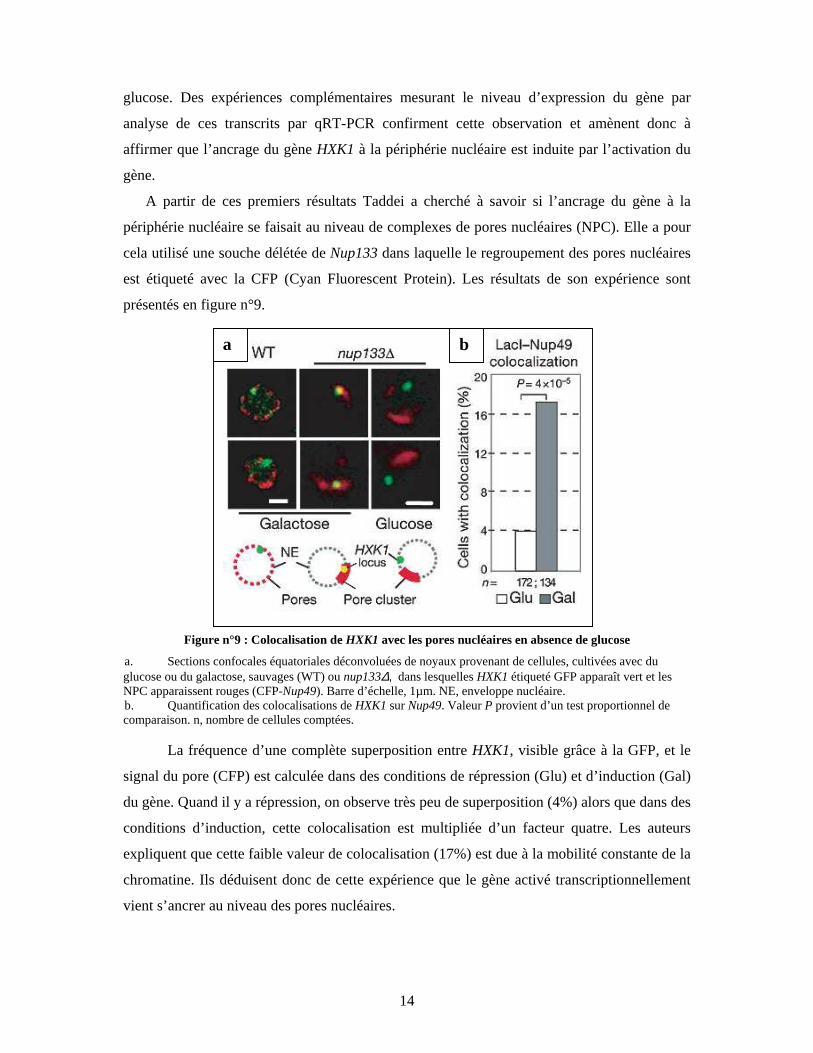
présentés en figure n°9.
Figure n°9 : Colocalisation de HXK1 avec les pores nucléaires en absence de glucose
a. Sections confocales équatoriales déconvoluées de noyaux provenant de cellules, cultivées avec du glucose ou du galactose, sauvages (WT) ou nup133∆, dans lesquelles HXK1 étiqueté GFP apparaît vert et les NPC apparaissent rouges (CFP-Nup49). Barre d’échelle, 1µm. NE, enveloppe nucléaire. b. Quantification des colocalisations de HXK1 sur Nup49. Valeur P provient d’un test proportionnel de comparaison. n, nombre de cellules comptées.
La fréquence d’une complète superposition entre HXK1, visible grâce à la GFP, et le
signal du pore (CFP) est calculée dans des conditions de répression (Glu) et d’induction (Gal)
du gène. Quand il y a répression, on observe très peu de superposition (4%) alors que dans des
conditions d’induction, cette colocalisation est multipliée d’un facteur quatre. Les auteurs
expliquent que cette faible valeur de colocalisation (17%) est due à la mobilité constante de la
chromatine. Ils déduisent donc de cette expérience que le gène activé transcriptionnellement
vient s’ancrer au niveau des pores nucléaires.
a b

15
Pour vérifier que l’association à l’enveloppe nucléaire du gène HXK1 activé est bien
caractéristique d’une transcription active, ils induisent HXK1 d’une autre façon. Les
expériences qu’ils mènent montrent que l’association stable aux NPC n’est pas absolument
nécessaire pour la transcription chez la levure. Ils cherchent alors à déterminer si l’association
aux pores qu’ils observent est nécessaire pour des niveaux élevés de transcription en analysant
les niveaux de transcrits et la position du gène pour les deux méthodes d’induction du gène
HXK1. Ils montrent alors que la deuxième technique d’induction présentée annule le niveau
normal d’expression de HXK1 induite par la première méthode, l’induction au galactose. Ils
proposent donc que l’association de HXK1 aux pores nucléaires est nécessaire pour une
expression maximale. En améliorant l’ancrage du gène aux pores nucléaires, les auteurs
montrent en effet une augmentation de l’activation de HXK1 par le galactose ce qui confirme
la proposition formulée précédemment.
Cette technique a permis à Taddei et al d’affirmer que les différents modes
d’activation d’un gène peuvent conduire à des positions nucléaires différentes et de proposer
que la position nucléaire a un rôle actif dans la détermination du niveau d’expression optimal
d’un gène chez la levure.
II.3- La répression de l’hétérochromatine et le repositionnement de locus sont liés à la
régulation de gènes de virulence chez Plasmodium falciparum. Duraisingh et al 2005.
Pour aborder la régulation périnucléaire de gènes subtélomériques de virulence chez
Plasmodium falciparum, Duraisingh et al insèrent le gène de la dihydrofolate réductase
humaine (hDHFR) dans la région TARE6 du chromosome 3 (cf. Figure n°4). Ils démontrent
que l’expression de ce gène marqueur aussi bien que celles des gènes var endogènes sont
épigénétiquement contrôlés et que le repositionnement de locus et la structure de la
chromatine jouent un rôle dans la répression et l’activation de ces gènes chez Plasmodium
falciparum.
Pour insérer un marqueur de transcription dans le subtélomère Rep20 du chromosome
3, ils transfectent les parasites Plasmodium falciparum (3D7) avec un plasmide pHdhfr, sous
contrôle d’un promoteur calmoduline, contenant le gène hDHFR humain codant pour la
résistance à une drogue, l’antifolate (WR99210). Le plasmide pHdhfr inclut 506 pb de la
région Rep20, pour permettre l’intégration homologue dans le subtélomère du chromosome 3,
et la séquence CAT (Chloramphénicol AcétylTransférase) comme cible pour l’hybridation in
situ par fluorescence :

16
Figure n°10 : Structure du subtélomère du chromosome3 de Plasmodium falciparum et intégration de
hDHFR.
Légende : La structure conservée du subtélomère du chromosome de Plasmodium falciparum est montrée. La région noire à gauche est le télomère. TARE6 correspond à la région Rep20. Le gène var sur le chromosome 3 (PF0005w) est représenté par la flèche noire. Le plasmide pHdhfr est présenté et contient Rep20 comme cible pour l’intégration. Les sites d’enzymes de restriction sont : H, HindIII ; M, MseI ; B, BsaAI ; S, StuI ; A, AatI ; X, XbaI . La structure d’intégration est montrée (2 copies du plasmide sont insérées 3.3kb en amont de l’ATG du gène var.
Les parasites transfectés sont sélectionnés sur WR pour obtenir des lignées 3D7/H
avec l’intégration de pHdhfr proche du gène var. Des expériences d’électrophorèse en champ
pulsé et de Southern blot permettent de confirmer l’intégration de la structure.
Pour déterminer si le gène hDHFR maintient une activité transcriptionnelle sans la
sélection (drogue WR), les lignées de parasites transfectées sont clonées et poussent sur 50
générations en absence de WR pour obtenir des lignées 3D7/Hc1. Après détermination des
IC50 pour la drogue sur les populations, ils considèrent qu’approximativement 40% de la
population ont un gène hDHFR silencieux ou inactivé et que la localisation subtélomérique
peut être importante pour cet état transcriptionnel.
Ils ont pu démontrer que le transgène subtélomérique hDHFR est réversiblement
silencieux. En analysant la sensibilité d’une dizaine de clones à la drogue par RT-PCR, ils ont
montré que hDHFR est transcrit dans les lignées résistantes au WR mais pas dans les
parasites sensibles. De plus, ils ont montré que les lignées sensibles pouvaient intégrer le gène
hDHFR et le transcrire après sur une nouvelle exposition à la drogue.
Dans cette étude, ils ont mis en évidence par des expériences de digestion de la
chromatine native avec une nucléase micrococcale (MNAse) que la répression (tout comme
l’activation) est associée à des altérations de la structure de la chromatine « locale » et à la
fonction de PfSir2.
Pour tester si hDHFR occupe une position spatiale différente au sein du noyau de
Plasmodium falciparum en fonction de son état (actif ou silencieux), ils emploient la

17
Figure n°12 :Position nucléaire de hDHFR à l’extrémité du chromosome 3 par rapport à
l’épisome pBsd/Rep20
Analyse FISH de 3D7/Hc1.3+/pB et 3D7/Hc1.3/pB respectivement Chr3Lactif et Chr3L silencieux. Images d’épifluorescence de noyaux colorés au DAPI et hybridé avec pGem(Rouge) pour identifier à la fis l’ »pisome pBsd/Rep20 et le hDHFR et CAT (vert) pour identifier la position de hDHFR intégré dans le chromosome 3 seulement.
technique du FISH. Pour fournir un marqueur de l’activation de cluster de chromosome
périphérique, ils génèrent un plasmide pBsd/Rep20(cf. Figure n°11) qui porte le gène de la
blasticidine désaminase (bsd) conférant une résistance à la blasticidine S, Rep20 pour assurer
l’association de l’épisome au cluster de chromosome à la périphérie nucléaire, et le gène de la
luciférase (lux) comme sonde.
Figure n°11 : Schéma du plasmide pBsd/Rep20
Légende : Le gène bsd (rouge) est sous le contrôle du promoteur hsp86 (bleu) et flanqué d’un terminateur dhfr de P. bergei (PbDt3’ ; vert). Le gène de la luciférase (jaune) est intégrée comme cible d’hybridation.
L’hybridation avec pGem détecte le gène hDHFR intégré dans le chromosome 3 dans
tous les clones transfectés. Chez Plasmodium falciparum, les épisomes contenant Rep20
s’associent physiquement avec les clusters de chromosomes localisés en périphérie, un
procédé qui permet la répartition des plasmides durant la mitose. Comme les parasites
transfectés avec le plasmide pBsd/Rep20 ont poussé sur blasticidine S le gène bsd doit être
exprimé et le plasmide contenu au sein de la région transcriptionnellement active. De plus, ce
plasmide sert de marqueur de domaines transcriptionnellement actifs à la périphérie nucléaire.
Pour déterminer si la transcription périnucléaire chez Plasmodium falciparum est restreinte à
des zones, les auteurs réalisent un FISH sur les lignées blasticidine S résistantes hDHFR actif
et hDHFR silencieux respectivement 3D7/Hc1.3+/pB et 3D7/Hc1.3/pB.

18
Ils emploient le FISH à deux couleurs utilisant le gène cat (vert) qui détecte
spécifiquement le hDHFR subtélomérique sur le chromosome 3 et le plasmide (rouge) pour
détecter les deux c’est-à-dire le locus du transgène hDHFR et l’épisome (cf. Figure n°12).
Si la présence d’une sous population parasite dans 3D7/Hc1.3/pB avec un hDHFR
activé est pris en compte (50% approximativement), on pourrait s’attendre à ce que ce gène
puisse colocaliser avec le plasmide pBsd/Rep20 soit en raison de l’intégration, soit en raison
d’une association physique avec la région transcriptionnellement active, ces résultats
deviennent hautement significatifs. Cela suggèrent que chez Plasmodium falciparum, les loci
subtélomériques transcriptionnellement actifs et silencieux occupent différentes localisations
périnucléaires. Finalement , ils proposent le modèle de régulation suivant :
Légende :
À gauche : Dans les noyaux avec hDHFR, Chr3L est séparé de l’épisome périnucléaire transcriptionnellement actif dans les clusters attachés à la membrane nucléaire (MN). La formation de cluster, médiée par les TAREs apparaît impliquée dans le pontage de Rep20. Étant donné que dans la chromatine native le promoteur devant le gène hDHFR est moins accessible à la nucléase miccrococale, il est probable que l’extrémité Chr3L silencieux soit empaquetée dans l’hétérochromatine (repésentée par les chromosomes « ondulés »). Ici, le plasmide BSD actif transcriptionnellement s’attache à la séquence Rep20 d’un autre chromosome qui se situe dans un sous compartiment périphérique transcriptionnellement actif. À droite : Suivant l’activation de la transcription du gène hDHFR par la sélection WR, le Chr3L transcriptionnellement actif et le plasmide épisomique colocalisent. Chr3L se relocaliserait dans un sous compartiment transcriptionnellement actif, occupé par le plasmide épisomique. De plus, l’accessibilité augmentée aux endonucléases suggèrent qu e le promoteur est dans une conformation plus ouverte. Il est important de remarquer que le gène var voisin n’est pas activé à ce moment, en accord avec les niveaux supplémentaires de régulation transcriptionnelle ( i.e. la répression coopérative intron-promoteur).
Figure n°13 : Modèle de repositionnement de locus et de répression de gène pour la région subtélomérique
des chromosomes chez P. falciparum.

19
III- PROJET DE RECHERCHE:
Étude de l’activité transcriptionnelle des gènes var subtélomériques au
niveau des pores nucléaires chez Plasmodium falciparum
Les "clusters" de télomères de Plasmodium falciparum sont à l'origine des variations
antigéniques des protéines de surface qui permettent l'échappement aux réponses
immunitaires de l'hôte. En effet, ces clusters favorisent les événements de recombinaison
entre les gènes qui codent pour ces protéines de surface. Les télomères de Plasmodium
falciparum peuvent être considérés comme le talon d'achille du pathogène car leur
perturbation empêcherait les mécanismes de variations antigéniques et donc le pathogène
serait plus vulnérable. C'est pourquoi il est intéressant de se pencher sur les mécanismes de
régulations des gènes var qui peuvent constituer une cible pour le développement de
nouvelles thérapies anti-parasitaires.
Au moyen des techniques utilisées par Taddei, nous allons étudié l’activité
transcriptionnelle des gènes var en fonction de leur localisation dans le noyau. Pour
approfondir les résultats obtenus par Duraisingh, nous avons voulu étudier l’implication des
pores nucléaires dans l’activité transcriptionnelle d’un gène var.
Dans un premier temps, nous allons devoir intégrer une répétition en tandem
d’opérateurs lac en amont d’un gène var par transfection stable au sein du génome de
Plasmodium falciparum. Pour suivre la localisation nucléaire de cette répétition et donc du
locus d’intérêt, nous allons produire la protéine de fusion répresseur LacI/GFP. Parallèlement,
nous allons produire une nucléoporine de Plasmodium falciparum fusionnée avec la BFP ou
Blue Fluorescent Protein afin de détecter la localisation des pores nucléaires. Ainsi, cette
technique pourra mettre en évidence une éventuelle colocalisation entre notre locus d’intérêt
et les pores nucléaires. Cette colocalisation pourrait être confirmée par des expériences de
FRET (Fluorescent Resonance Energy Transfer). Pour élargir cette étude, différentes
variantes de la technique du FISH pourront être utilisées dans le but de contrôler l’état
transcriptionnel du gène d’intérêt.

20
III.1- Synthèse de la répétition en tandem d’opérateurs lac
La technique utilisée pour construire la répétition en tandem des 256 opérateurs lac est
celle décrite par Robinett et illustrée par la figure n°6. Il faudra donc se procurer le plasmide
utilisé par Robinett qui contient cette séquence.
III.2- Insertion dans un plasmide de cette répétition en amont d’un gène d’intérêt
L’insertion de la répétition en tandem des 256 opérateurs lac dans le chromosome 3 de
Plasmodium falciparum va se faire par recombinaison homologue en amont du gène var
d’intérêt. Pour cela, il faut avoir une région d’homologie entre le plasmide et la région du
chromosome 3 afin de cibler l’intégration : il s’agit de la séquence Rep20. Le plasmide utilisé
par Duraisingh et al en 2005 comporte ces caractéristiques (cf. Figure n°14). Il suffirait donc
de demander à leur équipe de nous fournir le plasmide et de procéder à quelques
modifications pour effectuer nos expériences. En effet, la séquence lacop*256 présente sur le
plasmide de Robinett doit être intégrée dans le plasmide pHdhfr pour obtenir le plasmide
pHdhfr- lacop*256. Cela peut se faire par digestion enzymatique en utilisant l’enzyme MseI,
ainsi la séquence lacop*256 se retrouvera en amont de la séquence CAT. Si l’intégration au
génome ne se fait pas correctement (en raison d’une taille de l’insert trop importante), on
pourra envisager de retirer le gène CAT.
Une fois cette intégration réalisée, la reconnaissance de la séquence lacop*256 et donc
du locus d’intérêt se fera par une protéine de fusion répresseur LacI-NLS-GFP comme
l’illustre la figure n°7.
Figure 14 : Plasmide pHdhfr Légende : Le plasmide pHdhfr est présenté et contient Rep20 comme cible pour l’intégration, le gène hDHFR humain codant pour la résistance à une drogue (WR99210), la séquence CAT chloramphénicol acétyltransférase comme cible pour l’hybridation in situ par fluorescence Les sites d’enzymes de restriction sont : H, HindIII ; M, MseI ; B, BsaAI ; S, StuI ; A, AatI ; X, XbaI .

21
III.3- Transfection stable de la séquence répétée en tandem d’opérateurs lac dans le
génome de Plasmodium falciparum par recombinaison homologue
Le fait que le génome de Plasmodium falciparum soit haploïde facilite cette
manipulation. En effet, avec un génome diploïde on peut augmenter l’efficacité de
recombinaison par augmentation de la pression de sélection ce qui favorise les événements de
crossing-over inégaux et peut ainsi contribuer au fait que plusieurs copies s’intègrent dans le
génome. Ici, on souhaite intégrer au génome une seule copie de la répétition d’opérateurs lac
pour localiser le locus d’intérêt. Cependant les expériences de Duraisingh montrent qu’ils ont
obtenus plusieurs insertions de leur plasmide. Afin d’éviter cela, on pourra procéder à une
numération cellulaire dans le but d’effectuer une dilution clonale, c’est à dire dans des
conditions telles qu’on se retrouve avec zéro ou une cellule après dilution. Cette dilution sera
réalisée après la transfection du plasmide pHdhfr-lacop*256 juste avant d’appliquer la
sélection. Ainsi, la population obtenue ne proviendra que d’une seule cellule et donc toutes les
cellules de la lignées seront identiques.
La lignée de Plasmodium falciparum employée sera la lignée 3D7 qui a été utilisée par
l’équipe de Duraisingh en 2005. Les transfections de plasmide dans les cellules 3D7 se feront
par électroporation (Wu et al. 1995) comme pour l’équipe de Duraisingh. Des expériences
d’électrophorèse en champ pulsé et de Southern blot permettront de confirmer l’intégration de
la structure(Duraisingh et al, 2005).
III.4- Expression transitoire de protéines de fusion
III.4.1- Répresseur-LacI-NLS-GFP
La protéine nécessaire à la détection de la répétition en tandem d’opérateurs lac est la
protéine de fusion GFP-répresseur LacI. Robinett et son équipe en 1996 ont déjà produit une
telle protéine qui comporte un signal de localisation nucléaire de SV40. Nous pourrions nous
procurer le vecteur qu’ils ont utilisés. Le point de essentiel de la production de cette protéine
de fusion répresseur LacI-NLS-GFP pour une détection convenable de la répétition en tandem
d’opérateurs lac est la quantité de protéines de fusion au sein du noyau. En effet, il faut se
mettre dans des conditions où tous les répresseurs LacI-NLS-GFP se fixent uniquement au
niveau de la répétition d’opérateurs lac pour éviter qu’il y ait émission de fluorescence non

22
spécifique du locus. Le promoteur en amont du gène ne doit donc pas être un promoteur très
fort.
III.4.2- Nucléoporine de Plasmodium falciparum
La production d’une nucléoporine permettra de localiser les pores nucléaires de
Plasmodium falciparum et ainsi de voir s’il y a une colocalisation entre ces pores et notre
gène d’intérêt var. Afin de trouver des protéines de pores nucléaires accessibles de
Plasmodium falciparum, nous allons effectuer un BLASTP (Basic Local Alignment Search
Tool Proteic) de la séquence protéique de Nup49, la nucléoporine de levure utilisée dans les
expériences de Taddei, contre le protéome de Plasmodium falciparum. Une autre alternative
serait de déterminer une séquence consensus de nucléoporine de levure et une séquence
consensus de nucléoporine de mammifères dans le but de procéder à un BLASTP de cette
séquence contre le protéome de Plasmodium falciparum. Cette deuxième possibilité semble
plus plausible étant donné qu’elle tient compte d’un plus grand nombre de protéines.
De plus, le séquençage du génome de cet organisme achevé en 2002 se révèle être un
outil intéressant. Effectivement, si une nucléoporine a été annotée, on pourrait l’utiliser mais
il faudrait auparavant vérifier que celle-ci soit bien accessible au niveau du pore nucléaire.
Une fois la séquence de la nucléoporine d’intérêt déterminée, il faudra la cloner dans
un vecteur qui permettra de produire la protéine en fusion à la BFP. Qbiogen commercialise
ce type de vecteur contenant des promoteurs plus ou moins forts. Un paramètre essentiel est à
prendre en considération pour la suite de la démarche expérimentale. Il faut avoir une idée du
nombre de nucléoporines présentes dans le complexe du pore nucléaire. En effet, plus le
nombre sera important, plus le signal de fluorescence générée par la BFP sera amplifié.
III.5- Expériences de FRET
Pour mettre en évidence la colocalisation entre le locus d’intérêt et les nucléoporines,
la technique de choix est le FRET. En effet, cela permet de mettre en évidence la proximité de
deux molécules. Dans notre projet, le gène d’intérêt est détecté grâce à la GFP et les
nucléoporine grâce à la BFP. On peut donc utiliser les propriétés (longueur d’onde
d’excitation et longueur d’onde d’émission) de ces deux protéines fluorescentes pour mettre
en évidence un transfert d’énergie révélant la proximité des deux protéines.

23
Ici, on excitera nos préparations cellulaires à la longueur d’onde d’excitation de la
BFP et on mesurera l’intensité de fluorescence émise à la longueur d’onde d’émission de la
GFP comme le montre la figure n°15.
BFP GFP
λ émission GFP≈ 504nmλ excitation BFP≈ 380 nm
λ émission BFP≈450nm
λ excitation GFP≈ 475 nm
FRET(1)
Ro=50-60 Å
(2) Absence de FRET
BFP GFP
λ émission GFP≈ 504nmλ excitation BFP≈ 380 nm
λ émission BFP≈450nm
λ excitation GFP≈ 475 nm
FRET(1)
Ro=50-60 Å
(2) Absence de FRET
Figure n°15 : Principe du FRET Légende : Le Ro ou distance de Förster est la distance entre les chromophores pour laquelle il y a 50 % d’efficacité de transfert. Un contrôle est cependant nécessaire pour valider cette mesure. Il faut exciter la
préparation à la longueur d’onde d’excitation de la GFP pour vérifier la présence du
répresseur LacI/GFP. De plus si on n’observe pas de transfert de fluorescence, il faudra
contrôler la présence de la nucléoporine/BFP en mesurant l’intensité de fluorescence émise à
la longueur d’onde d’émission de la BFP.
III.6- Expériences de FISH
Des expériences de FISH (Du et al 2003) pourront nous donner une information sur
l’activité du gène var étudié. Du RNA-FISH pourra être réalisé en utilisant une sonde
correspondant à une séquence intronique du gène pour pouvoir localiser les ARN pré-
messagers et ainsi pouvoir suivre la transcription du gène. Ces expériences pourront être
combinées à des expériences d’ImmunoFISH. En effet, si l’on détecte à l’aide d’un anticorps
couplé à un fluorophore spécifique, la RNA polymérase II hyperphosphorylée sur son
domaine carboxy-terminale, cela nous permettra de visualiser les machineries de
transcription. Ainsi s’il y a colocalisation des ARN pré-messagers var avec les machineries de
transcription, cela démontrera l’activité transcriptionnelle du gène.

24
Pour compléter cela, on peut envisager de quantifier les transcrits du gène d’intérêt par
qRT-PCR, comme l’a fait Taddei, afin de caractériser le niveau d’expression du gène étudié.
Les télomères pourront également être détectés pour les situer par rapport au pore
nucléaire en utilisant la technique du DNA-FISH. Dans cette expérience, les séquences
TARE, qui détectent tous les télomères, seront utilisées.
IV- CONCLUSION ET PERSPECTIVES
Ce projet de recherche permettra de déterminer si un gène subtélomérique
transcriptionnellement inactif de Plasmodium falciparum, le gène var du chromosome 3, se
retrouve positionné au niveau des pores nucléaires lorsque sa transcription est activée. En
effet, une fois les différentes constructions effectuées, la technique de FRET nous permettra
de voir si le gène d’intérêt colocalise avec les pores nucléaires et le RNA-FISH permettra de
déterminer si notre gène d’intérêt est actif au niveau transcriptionnel. Les résultats obtenus
pourront alors être confrontés à ceux de Taddei chez la levure. Cela nous donnera une vision
plus complète de la régulation de la transcription des gènes positionnés en périphérie
nucléaire chez les eucaryotes.
De plus, étant donné que cette étude traite des mécanismes de régulations des gènes
var responsables des variations antigéniques de Plasmodium falciparum, elle pourrait entrer
dans le cadre d’une recherche de cibles pour le développement de nouvelles thérapies anti-
parasitaires

25
BIBLIOGRAPHIE • Belgareh N, Rabut G, Bai SW, van Overbeek M, Beaudouin J, Daigle N, Zatsepina OV, Pasteau F, Labas V, Fromont-Racine M, Ellenberg J, Doye V. 2001 An evolutionarily conserved NPC subcomplex, which redistributes in part to kinetochores in mammalian cells. The Journal of Cell Biology 154(6):1147-1160. • Blobel G. 1985 Gene gating : a hypothesis. Proc.Natl Acad. Sci. USA, Vol. 82, 8527-8529. • Casolari JM, Brown CR, Komili S, West J, Hieronymus H and Silver PA. 2004 Genome-wide localization of the nuclear transport machinery couples transcriptional status and nuclear organization. Cell, Vol.117, 427-439. • Cremer T, Cremer M, Dietzel S, Müller S, Solovei I, Fakan S. 2006 Chromosomes territories- a functional nuclear landscape. Current Opinion in Cell Biology, Vol.18, 307-316 • Croft J, Bridger J, Boyle S, Perry P, Teague P and Bickmore A. 1999 Differences in the localization and morphology of chromosomes in the human nucleus. The Journal of Cell Biology, Vol. 145, Number 6. • Dekker J, Rippe K, Dekker M, Kleckner N. 2002 Capturing chromosome conformation. Science, 15;295(5558):1306-11. • Drummond SP, Rutherford SA, Sanderson HS, and Allen TD. 2006 High resolution analysis of mammalian nuclear structure throughout the cell cycle: implications for nuclear pore complex assembly during interphase and mitosis. Can. J. Physiol. Pharmacol, 84:423–430 • Du MJ., Liu DP and Liang CC. 2003 The impact of FISH on globin gene regulation research. Experimental Cell Research, Vol. 291, 267-274. • Duraisingh MT, Voss TS, Marty AJ, Duffy MF, Good RT, Thompson JK, Freitas-Junior LH, Scherf A, Crabb BS and Cowman AF. 2005 Heterochromatin Silencing and Locus Repositioning Linked to Regulation of Virulence Genes in Plasmodium falciparum. Cell, Vol. 121 : 13-24. • Figueiredo LM, Freitas-Junior LH, Bottius E, Olivo–Martin JC and Scherf A. 2002 A central role for Plasmodium falciparum subtelomerics regions in spatial positioning and telomere length regulation. The EMBO journal, Vol. 21(4):815-824. • Fraser P. 2006 Transcriptional control thrown for a loop. Current Opinion in Genetics & Development, 16:490–495. • Gardner MJ, Hall N, Fung E, White O, Berriman M, Hyman RW, Carlton JM, Pain A, Nelson KE, Bowman S, et al. 2002 Genome sequence of the human malaria parasite Plasmodium falciparum. Nature, 419 : 498–511.

26
• Osborne CS, Chakalova L, Brown KE, Carter D, Hurton A, Debrand E, Goyenchea B, Mitchell JA, Lopes S, Reik W & Fraser P. 2004 Active genes dynamically colocalize to shared sites of ongoing transcription. Nature genetics, Vol.36/10 1065-1071. • Robinett CC, Straight A, Li G, Willhelm C, Sudlow G, Murray A and Belmont AS. 1996 In Vivo Localization of DNA Sequences and Visualization of Large-Scale Chromatin Organization Using Lac Operator/Repressor Recognition. The Journal of Cell Biology, Vol. 135, Number 6, Part 2, 1685-1700 • Scherf A, Figueiredo LM and Freitas-Junior LH. 2001. Plasmodium telomeres: a pathogen’s perspective. Current Opinion in Microbiology, 4:409–414 • Taddei A, Van Houwe G, Hediger F, Kalck V, Cubizolles F, Schober H & Gasser SM. 2006 Nuclear pore association confers optimal expression levels for an inducible yeast gene. Nature, Vol. 441|8 774-778 • Wu Y, Sifri CD, Lei HH, Su XZ and Wellems TE. 1995 Transfection of Plasmodium falciparum within human red blood cells. Proc. Natl. Acad. Sci. USA, Vol. 92, pp. 973-977