Embed Size (px)
Citation preview

" REPUBLIQUE DE COTE D'IVOIRE
Union-Discipline-Travail
Ministère de l'Enseignement Supérieur et de la Recherche Scientifique
Année universitaire 2013-2014
UNIVL::HSITE NANGUIABROGOUA
UFR Sciences de la Nature Laboratoire de Biologie et d, Amélioration
des Productions Végétale
MÉMOIRE DE MASTER
Biodiversité et Gestion Durable des Écosystèmes
Thème: , Etude de l'effet de quelques fongicides et nématicides
utilisés en bananeraie sur la croissance et la sporulation de
Metarhizium sp agent de lutte biologique contre le
charançon noir du bananier, Cosmopolites sordidus Gemar
(Coléoptère: Curculiooidae)
Présenté par :
SÉKA Chapaud Landry Vigile
Soutenu publiquement le Vendredi 29 Août 2014, devant le jury composé de
M. TRA BI. Maître de Conférences, Université Nangui Abrogoua. Président
M. KOUAKOU Tanoh Hilaire, Maître de Conférences, Université Nangui Abrogoua Codirecteur
M. KOBENAN Kouman, Directeur de Recherche, CNRA
M. SIÉ Raoul. Maître de Conférences, Université Nangui Abrogoua
M. DOUMBIA Mamadou, Maître de Conférences, Université Nangui Abrogoua
Codirecteur
Examinateur
Examinateur

DÉDICACE
À mon Dieu et Seigneur Jésus-Christ pour m'avoir
donné la santé, le courage et mis sur mon chemin des
personnes disposées à m'aider, me secourir ...
À mon père OSSIN Séka Joseph-Désiré et à ma mère
DAGOU Opri Anne Marie. Je vous dédie ce mémoire comme
le fruit de trente-deux ans d'amour et de sacrifices
à mon égard.
À ma grande sœur BOSSEY Djoman Antoinette,
À mes grands frères SÉKA Alain, SÉKA Pacôme, SÉKA
Guy, SÉKA Thierry et SÉKA Hermann,
À mes petits frères SÉKA Stéphane et SÉKA Olivier,
À mes oncles OSSIN Yapi, Ossin Adjamey, ABÉ Basile,
DAGOU Rodrigue, DAGOU Zéphirin, DAGOU Théophile,
DAGOU Moïse, DAGOU Alain et DAGOU Guy.
À ma bien aimée ABIO Clémentine.
Pour votre AMOUR et votre SOUTIEN.

REMERCIEMENTS
Cette étude a été menée au laboratoire de phytopathologie du Centre National de
Recherche Agronomique (CNRA), à la Station de Recherche de Bimbresso. J'aimerais
remercier les personnes morales et physiques dont le concours a été fructueux pour accomplir
ce travail.
Permettez moi de remerciez d'abord le CNRA et particulièrement toute l'équipe de la
Station de Recherche de Bimbresso.
Je remercie aussi l'Université Nangui Abrogoua et surtout l'UFR des Sciences de la
Nature pour la formation académique que j'y ai reçue depuis ma première année universitaire.
Mes sincères remerciements vont aussi à l'endroit de:
)- Monsieur TIDO Seydou, Doyen de l'UFR des Sciences de la Nature de l'Université
Nangui Abrogoua.
)- Monsieur KOU A.KOU Tanoh Hilaire, Maître de Conférences à l'UFR des Sciences de
la Nature de l'Université Nangui Abrogoua qui a accepté d'être le codirecteur
scientifique de ce travail.
)- Monsieur GNONHOURI Philippe, Directeur de la station CNRA de Bimbresso.
)- Monsieur KOBENAN Kouman, Directeur de Recherche au CNRA pour son
encadrement et pour avoir accepté que je fasse mon stage pratique à ladite station. U
m'a donné le goût de la recherche et je lui suis reconnaissant pour les suggestions
qu'Il a faites dans la conception de ce mémoire.
)- Monsieur TRAORÉ Siaka, Chercheur au CNRA, qui a suivi de bout en bout toutes les
activités ici présentées.
)- Monsieur ABY N'goran, Checheur au CNRA, qui a été au four et au moulin dans
l'exécution de toutes les manipulations et la rédaction de ce travail. Oui. ces
enseignements et ces conseils m'ont été bénéfiques. Il s'est montré disponible à tout
moment bien qu'il ne soit pas mon encadreur. Vraiment, je lui suis reconnaissant.
Je remercie particulièrement les membres du jury dont :
)- Monsieur TRA BI, Maître de Conférences à l'UFR des Sciences de la Nature de
l'Université Nangui Abrogoua, qui a accepté de présider le jury et aussi pour la
formation en biologie végétale que j'ai reçue de lui depuis ma deuxième année
universitaire à Daloa.
)- Monsieur SIÉ Raoul, Maître de Conférences à J 'UFR des Sciences de la Nature de
l'Université Nangui Abrogoua, qui m'a enseigné la génétique formelle.
ii

)"' Monsieur DOUMBIA Mamadou, Maître de Conférences à l 'UFR des Sciences de la
Nature l'Université Nangui Abrogoua, auprès de qui j'ai reçu les bases fondamentales
en entomologie.
Je dois reconnaissance à Mesdames KRAMON Pierrette et ZAKO Jorgina, les assistantes
techniques qui ont été les chevilles ouvrières dans l'exécution de toutes les activités
rapportées dans ce document.
Mes remerciements vont aussi. à l'endroit de mes aînés KONAN Yao François, ESSI Brice
et mes amis KIMOU Serge, KOUADIO Samuel, DIDI Roland, ASTIN Olivier, Y AO Alban,
SEKA Hervé qui m'ont épaulé à travers leurs conseils et leurs aides matériels.
iii

SOMMAIRE ,
DEDICACE I
REM:ERCIEMENTS II
RÉSUMÉ VI
LISTE DES ABRÉVIATIONS ET ACRONYMES IX
INTRODUCTION l
PREMIÈRE PARTIE: REVUE BIBLIOGRAPHIQUE 4
1.1. Généralités sur les pesticides 5
1.1.1. Définition d'un pesticide 5
1.1.2.
1.1.3.
Classification des pesticides 5
Mode d'action et cibles biochimiques des fongicides 7
1.2. Généralités sur le bananier 12
1.2.l.
1.2.2.
1.2.3.
1.2.4.
1.2.5.
1.2.6.
Origine et Systématique 12
Description du bananier 12
Ecologie du bananier 12
Productions de la banane en Côte d'Ivoire 14
Principales contraintes biotiques à la culture du bananier 14
Méthodes de lutte contre Cosmopolites sordidus 17
DEUXIÈME PARTIE : MATÉRIEL ET MÉTHODES 26
2.1. Site d'étude 27
2.2. Matériel 27
2.2.1.
2.2.2.
2.2.3.
2.2.4.
Matériel de laboratoire 27
Champignon entomopathogène 27
Insecte ravageur 27
Pesticides utilisés 27
2.3. Méthodes 27
2.3 .1. Préparation des milieux de culture 28
2.3.2.
2.3.3.
2.3.4.
Effet des pesticides sur la germination des conidies 28
Effet des pesticides sur la croissance mycélienne 30
Effet des pesticides sur la sporulation 30
iv

2.3.5. Pouvoir germinatif des conidies produites après tratement de
fongicide 30
2.3.6. Pouvoir infectieux des conidies produites après traitement de
fongicide 30
2.3.7. Evaluation de l'effet des pesticides 32
2.3.8. Analyse statistique 34
TROISIÈME PARTIE: RÉSULTATS 35
3.1. Effet des fongicides sur l'isolat de Metarhizium sp 36
_3. 1. 1. Effet des fongicides sur la germination des conidies 36
3.1.2.
3.1.3.
Effet des fongicides sur la croissance mycélienne 38
Effet des fongicides sur la sporulation 40
3.1.4. Pouvoir germinatif des conidies produites après traitement de
fongicide 42
3.1.5. Pouvoir infectieux des conidies produites après traitement de
fongicide 44
3.2. Effets des nématicides sur l'isolat de Metarhizium sp 49
3 .2.1. Effet des nématicides sur la germination des conidies 49
3 .2.2. Effet des nématicides sur la croissance du mycélium 49
3 .2.3. Effet des nématicides sur la sporulation 51
QUATRIEME PARTIE: DISCUSSION 54
CONCLUSION ET PERSPECTIVES 61
RÉFÉRENCES BIBLIOGRAPHIQUES 63
V

RÉSUMÉ Pour utiliser Metarhizium sp comme agent de lutte biologique contre
Cosmopolites sordidus dans les plantations où sont appliqués des pesticides afin de
protéger Je bananier, l'effet de trois fongicides et deux nématicides sur ce champignon
a été évalué. La germination conidienne, la croissance mycélienne et la sporulation ont
été d'abord évaluées sur des milieux de culture amendés de différentes doses de
chaque pesticide ou pas pour le témoin. Ensuite, le taux de germination des conidies
produites après traitement de fongicide a été déterminé sur milieu gélosé. Puis ces
mêmes conidies produites après traitement de fongicide ont été servi à inoculer des
adultes de Cosmopolites sordidus pour évaluer leur pouvoir infectieux.
Les taux d'inhibition de la germination conidienne provoqués par les
fongicides méthyl-thiophanate, tritloxystrobine et propiconazole ont été relativement
faibles, inférieurs à 50 %. Par contre, la croissance mycélienne et la sporulation ont été
fortement inhibées par la plupart des doses du trifloxystrobine et du propiconazole, 64
à 99 % de taux d'inhibition. Alors que l'incidence des doses du rnéthyl-thiophanate sur
la croissance mycélienne et la sporulation ont été relativement faibles, 13 à 56 % de
taux de réduction. Les conidies produites après traitement de fongicide ont présenté
des pouvoirs germinatifs variables et inférieurs à celui des conidies produites sur le
milieu témoin. Ces mêmes conidies produites après traitement de fongicide ont été
effectivement pathogènes des charançons adultes de Cosmopolites sordidus avec des
taux de mortalité de 16 à 67 % et inférieurs au taux de mortalité (75 %) provoqué par
les conidies produites sur le milieu témoin.
Contrairement aux fongicides, les doses utilisées d'oxamyl et d'éthoprophos
ont stimulé la germination conidienne, - 20 à - 11 % de taux d'inhibition. La
croissance mycélienne a été aussi stimulée par la plupart des doses de ces nématicides.
De même, la sporulation a été stimulée par les doses de 3 et 5 ppm de ces nématicides.
Alors que les doses de 10 et 15 ppm des deux nématicides ont provoqué des taux de
réduction relativement élevés et compris entre 58 et 64 %.
Mots clés : charançon, champignon, entornopathogène, fongicide, nématicide,
inhibition, germination, conidie, croissance, mycélium, sporulation, pathogène,
mortalité.
vi

LISTE DES FIGURES Figure 1: Schéma d'un bananier en phase de fructification 13
Figure 2: Zones de production des bananes et bananes plantain en
Côte d'Ivoire 15
Figure 3: Dessin de l'adulte du charançon noir du bananier
Cosmopolites sordidus 18
Figure 4 : Stades de la métamorphose chez Cosmopolites sordidus 18
Figure 5 : Décorticage tangentiel d'une souche de bananier
montrant les galeries larvaires 18
Figure 6 : Piège de Vilardebo utilisé pour la collecte des
charançons 18
Figure 7: Culture pure de Metarhizium sp sur milieu PDA et
Conidies de Metarhizium sp vues au microscope photonique Gx400 23
Figure 8: Charançon noir du bananier présentant la muscardine
verte causé par Met arhizium sp 25
Figure 9: Piège à contre plaqué muni d'inoculum de l'isolat de
Met arhizium sp. BME2 25
Figure 10: Situation géographique de la sous-préfecture de Songon 29
Figure 11 Evolution du taux de mortalité des adultes de
Cosmopolites sordidus inoculés avec des suspensions de conidies
produites après traitement de Méthyl-thiophanate et de
Propiconazole 45
Figure 12 : Evolution du taux de mortalité des adultes de
Cosmopolites sordidus inoculés avec des suspensions de conidies
produites après traitement de Trifloxystrobine .48
vii

LISTE DES TABLEAUX
Tableau 1: Différentes doses des fongicides et nématicides utilisés 29
Tableau 2 : Coefficients des périmètres de comptage de
l'hématimètre de Malassez 31
Tableau 3 : Taux d'inhibition de la germination des conidies de
l 'isoJat BME 2 de Metarhizium sp sur milieu géJosé amendé des
fongicides Methyl-thiophanate, Trifloxystrobine et Propiconazole 37
Tableau 4 : Taux d'inhibition de la croissance mycélienne de
l'isolat BME 2 de Met arhizium sp sur milieu PDA amendé des
fongicides Metbyl-thiophanate, Trifloxystrobine et Propiconazole 39
Tableau 5 : Taux d'inhibition de la sporulation de l'isolat BME 2
de Metarhizium sp sur milieu PDA amendé des fongicides Methyl-
thiophanate, Trifloxystrobine et Propiconazole .41
Tableau 6 : Taux de germination des conidies de l'isolat BME 2 de
Metarhizium sp produites après traitement de Methyl-thiophanate,
Trifloxystrobine et Propiconazole .43
Tableau 7 : Taux de mortalité des charançons inoculés avec les
conidies produites après traitement de Méthyl-thiophanate,
Trifloxystrobine et Propiconazole .46
Tableau 8: Taux d'inhibition de la germination des conidies de
l'isolat BME 2 de Metarhizium sp sur milieu gélosé amendé des nématicides Oxamyl et Ethoprophos 50
Tableau 9 : Taux d'inhibition de la croissance mycélienne de
l'isolat BME 2 de Metarhizium sp sur milieu PDA amendé des
nématicides Oxamyl et Ethoprophos 50
Tableau 10 : Taux d'inhibition de la sporulation de l'isolat BME 2
de Metarhizium sp sur milieu PDA amendé des nématicides Oxamyl
et Ethoprophos 53
viii

LISTE DES ABRÉVIATIONS ET ACRONYMES
ANOVA : Analyse des Variances
BME2: Batia Mort Ensemencé 2
BME5: Batia Mort Ensemencé 5
CNRA : Centre National de Recherche Agronomique
FAO : Organisation Mondiale pour I' Alimentation et l' Agriculture
FIRCA : Fond Interprofessionnel pour la Recherche et le Conseil Agricole
IBS : Inhibiteurs de la Biosynthèse des Stérols
LUBILOSA : Lutte Biologique contre les Locustes et Sautereaux
OCAB : Organisation Centrale des Exportateurs d' Ananas-Bananes
ODEADOM: Office de Développement de l'Economie Agricole d'Outre Mer
PDA : Potato Dextrose Agar
PIB : Produit Intérieur Brut
SAKJ : Société Agricole Kablan Joubin
SBMK : Société Bananière Maubert Kabla
SCB : Société pour le développement de la Culture de la Banane
TC : Talt Conidia
TL50: Temps Létal pour 50 % d'individus
TL90: Temps Létal pour 90 % d'individus
TLlOO : Temps Létal pour 100 % d'individus
ix

INTRODUCTION
1

Le charançon noir du bananier, Cosmopolites sordidus Gemar (Coléoptère :
Curculionidae), responsable d'au moins 40 % des pertes dans les bananeraies, est l'une des
contraintes majeures de la culture de banane (Gold et al., 2001). L'incidence du parasitisme
est telle que la lutte chimique représente la stratégie de lutte dominante à cause de son
efficacité. Protéger le bananier contre ce ravageur est devenu une priorité pour assurer
l'autosuffisance alimentaire. Malheureusement, les pesticides sont de plus "en plus sur la
sellette pour des questions de toxicité, de pollution de l'environnement. de santé et même
de biodiversité (Faurie el al., 2009). Il se pose également le problème de l'efficacité des
produits phytosanitaires qui, comme les antibiotiques utilisés en médecine humaine
provoquent l'apparition de résistance qui les rendent inefficaces (He et al., 2005). Face à
ces problèmes, l'utilisation de plantes transgéniques a été préconisée, mais elle rencontre
actuellement de nombreuses oppositions au niveau éthique (Kouser et Qaim, 2011). Dans
ce contexte, il apparait nécessaire de rechercher des alternatives plus efficaces pour le
développement d'une agriculture durable. Ainsi, la lutte biologique a été proposée comme
stratégie de lutte par plusieurs auteurs (Schiffers, 2011), parce qu'elle est plus respectueuse
de l'environnement. De plus, la lutte biologique au moyen d'organismes antagonistes tels
que les champignons Beauveria et Metarhizium a été réalisée avec succès par Gold et al.
(2003). En Côte d'Ivoire, des tests au champ ont montré une meilleure performance des
souches locales de Metarhizium sp par rapport au biopesticide à base de Beauveria bassiana, une souche exogène. Les mortalités du charançon noir du bananier provoquées
par des isolats locaux de Metarhizium sp (60 à 70%) ont été supérieures à celles induites
(10 à 21 %) par ce biopesticide (Aby, 2013). D'où l'intérêt porté aux isolats locaux de
Metarhizium sp.
Compte tenu de l'efficacité de cet agent de lutte biologique sur Cosmopolites
sordidus. il est important d'identifier les conditions propices à son application au champ. Il
s'agit de déterminer une synergie entre l'emploi de ce champignon entomopathogène et
celui des pesticides (insecticides, fongicides, nématicides, herbicides) et des engrais déjà
utilisés pour protéger les bananiers et optimiser leur rendement. Pour ce faire, Aby (2013)
a testé sur des milieux amendés avec la dose appliquée au champ de certains pesticides
l'efficacité de 11 isolats de Metarhizium sp dont l'isolat BME2 a été le plus performant.
Ainsi, dans cette étude, le comportement de l'isolat BME2 de Metarhizium sp dans des
milieux amendés d'une part de fongicides utilisés contre la cercosporiose noire et d'autre
2

part de nématicides utilisés contre le nématode migrateur Radopholus similis a été
évalué.
L'objectif général de ce travail est de déterminer les conditions propices à
l'application au champ de Metarhizium sp afin de réduire la quantité d'insecticide utilisée
pour lutter contre le charançon noir du bananier, Cosmopolites sordidus.
De façon spécifique, il s'est agi de:
).:> déterminer l'effet de quelques fongicides et de nématicides sur la germination, la
croissance mycélienne et la sporulation de l'isolat BME2 de Metarhizium sp ;
).:> évaluer le pouvoir germinatif et le pouvoir infectieux des conidies produites après
traitement de fongicides.
Le manuscrit s'organise en quatre parties. La première partie est consacrée à la
revue bibliographique. La deuxième partie décrit le matériel et la méthode d'étude utilisés
et la troisième partie expose les résultats qui sont ensuite discutés dans la quatrième partie.
Enfin, le travail se termine par une conclusion, les perspectives et des références
bibliographiques.
3

' PREMIERE PARTIE:
REVUE BIBLIOGRAPHIQUE
4

1.1. Généralités sur les pesticides
1.1.1. Définition d'un pesticide
Un pesticide est toute substance ou mélange de substances qui est destinée à
repousser, détruire ou combattre les ravageurs (y compris les vecteurs de maladies
humaines ou animales) et les espèces indésirables de plantes ou d'animaux causant des
dommages ou se montrant autrement nuisibles durant la production. la transformation. Je
stockage, le transport ou la commercialisation des denrées alimentaires, des produits
agricoles, du bois et des produits ligneux, ou des aliments pour animaux, ou qui peut être
administrée aux animaux pour combattre les insectes, les arachnides et les autres endo- ou
ectoparasites. Le terme inclut les substances destinées à être utilisées comme régulateur de
croissance des plantes, comme défoliant, comme agent de dessiccation. comme agent
d'éclaircissage des fruits ou pour empêcher la chute prématurée de ceux-ci, ainsi que les
substances appliquées sur les cultures, avant ou après la récolte, pour protéger les produits
contre la détérioration durant l'entreposage et le transport (FAO, 2003).
1.1 .2. Classification des pesticides
Plusieurs critères permettent de classer les pesticides. Ces critères sont entre autres
le domaine d'utilisation, les propriétés biologiques, les propriétés chimiques ou physico
chimiques.
1.1.2.1. Classification selon le domaine d'utilisation
Le mot pesticide recouvre en réalité deux catégories de produits distincts qui sont
les produits phytosanitaires et les produits biocides. Les produits phytosanitaires ou
phytopharmaceutiques sont utilisés le plus souvent en milieu végétal, agricole et ont pour
objet de prévenir, contrôler ou éliminer des organismes vivants jugés indésirables. Tandis
que les produits biocides sont utilisés en milieu non agricole pour détruire ou repousser les
nuisibles, notamment dans le domaine de la conservation du bois, la désinfection d'objets
en milieu hospitalier ainsi que dans certains usages domestiques.
1 1 1 5

1.1.2.2. Classification selon les propriétés biologiques
En fonction des organismes nuisibles qui sont éliminés par les pesticides, ceux-ci
sont repartis en plusieurs groupes tels que :
)"' les insecticides destinés à éliminer les insectes qui se nourrissent ou pondent sur les
cultures;
)"' les rodenticides qui tuent les rongeurs ;
)"' les moUuscicides qui ont la propriété de tuer les mollusques (limace, escargot, etc);
)"' les nématicides destinés à élimer les nématodes considérés nuisibles ;
)"' les fongicides utilisés pour lutter contre les mycoses, maladies provoquées par les
champignons ;
)"' les herbicides destinés à tuer les mauvaises herbes ou adventices qui étouffent les
végétaux d'intérêt.
1.1.2.3. Classification selon les propriétés chimiques
Selon les propriétés chimiques ou physico-chimiques, on distingue quatre grandes
familles de pesticides.
a. Famille des organochlorés
Les composés organiques du chlore appelés organochlorés sont les pesticides dits
hydrocarbures ou halogénés. Ils sont caractérisés par la présence de plusieurs atomes de
chlore autour du noyau phénolique ou benzénique. Ils sont d'une très grande rémanence
dans les milieux naturels, doués de propriétés neurotoxiques et utilisés comme insecticides.
Il s'agit de la famille du DDT (dichloro-diphényl-trichloroéthane) et ses dérivés, Je lindane,
interdits du fait de leur persistance et des risques d'accumulation dans les sols, les tissus
végétaux et les graisses animales. Seuls restent autorisés: le diénochlore et l'endosulfan
(Couteux et Lejeune, 2005).
b. Famille des organophosphorés
Le nom du chimiste allemand Schrader est lié à cette classe de composés
organiques du phosphore qui ont été développés à partir de 1944 pour remplacer les
organochlorés désormais interdits. Ils se sont rapidement imposés du fait de leur
rémanence plus faible, et de leur grande efficacité. Ces produits sont des esters de l'acide
orthophosphorique (P04H3) et sont doués de propriétés insecticides, acaricides et
nématicides, Il s'agit de la famille des phosphates (le dichlorvos), des thiophosphates (le
6

parathion), des dithiophospahtes (le dimethoate), des phosphonates (le cruformate)
halogénés ou non, fluorés, alkylés, hétérocycliques (Cluzeau, 1995).
c. Famille des carbamates
La synthèse chimique des carbamate a été effectuée à partir des composés
anticholinestérasiques d'origine végétale. Les carbamates sont des dérivés (esters et
amides) de l'acide carbamique (HOOC-NH2) et sont doués de propriétés insecticide
fongicide, nématicide, herbicide et molluscide. Cette famille est subdivisée en trois
groupes dont les naphthylcarbamates tel que le carbaryl, les phénylcarbamates dont le
propoxur, les carbamates hétérocycliques tel que l'aldicarbe (Cluzeau, 1995; Couteux et
Lejeune, 2005).
d. Famille des pyrétroïdes
Le groupe des pyréthroïdes au sens large, englobe à la fois les pyréthrines dérivés
naturels des capitules de la pyrèthre ( Chrysanthemum cinerariaefoliumï et les pyréthrines
de synthèse photostables dits encore pyréthrinoïdes. Les pyréthrines naturels sont des
dérivés de l'acide chrysanthémique et de l'acide pyréthrique. Ces deux acides donnent
respectivement avec trois alcools différents : pyrétrolone, jasmolone, cinérolone ; les
pyréthrines I et II, jasmolines I et 11, cinérines I et II. Les pyréthrines naturels sont
photolabiles et de ce fait, sont utilisés comme des insecticides domestiques, vétérinaires et
hygiéniques. Les pyréthrines de synthèse tels que le Decis ou la deltaméthrine, la
permétrhine et l'alphamétrine sont des esters de l'acide cyclopropanecarboxilique
(Cluzeau, 1995). Du fait de leur très bonne biodégradabilité, les pyréthrines de synthèse
sont largement utilisés contre les arthropodes des cultures maraîchères et en santé publique
contre les diptères, vecteurs de nombreuses maladies.
1.1.3. Mode d'action et cibles biochimiques des fongicides
Les fongicides sont des substances destinées à combattre les champignons
responsables de maladies chez les plantes. Un fongicide est donc un produit phytosanitaire
conçu pour tuer ou limiter le développement des champignons parasites des végétaux. Pour
ce faire, certains fongicides agissent à la fois sur plusieurs cibles biochimiques et sont
appelés fongicides multi-sites. Tandis que d'autres fongicides affectent une réaction
biochimique bien précise et sont appelés fongicides uni-sites.
7

1.1.3.1. Fongicides multi-sites
Les fongicides multi-sites sont des produits qui agissent sur plusieurs mécanismes
impliqués dans la production d'énergie cellulaire. Ces mécanismes de base existent chez
tous les êtres vivants y compris les champignons, ce qui donne à ces produits un large
spectre d'action c'est-à-dire Jeur polyvalence. Ces fongicides agissent par contact en
inhibant la germination des spores avant que celles-ci n'aient pu émettre des :filaments
pénétrant dans les tissus de l'hôte et ne peuvent donc être utilisés qu'à titre préventif. Ces
fongicides multi-sites sont entre autres des produits à base de métaux et des
dithiocarbamates (Cluzeau, 1995).
a. Les produits à base de métaux
Ces produits sont le cuivre sous forme de sel (forme minérale) ou combiné à une
molécule organique et le soufre sous forme de poudre ou en mélange avec des tensions
actiifs sous forme de suspension dans l'eau. Les métaux sont des agents thiolprives c'est-à
dire qu'ils se fixent sur le groupement SH ou thiol (R-SH) constituant la partie active de
nombreuses enzymes ayant un rôle dans les phénomènes d'oxydoréduction assurant le
transport d'énergie cellulaire. Les métaux ont l'inconvénient de posséder une phytotoxicité
à l'égard de la culture. Il a donc été nécessaire de les préparer sous une forme qui réduit
cette phytotoxicité (forme organique). Les formes organiques ont de plus l'avantage d'avoir
une meilleure rémanence, ce qui évite leur lessivage à la première pluie.
b. Les dithiocarbamates
Les ditbiocarbamates agissent en libérant des isocyanates ou du thirame, molécules
actives qui bloquent les groupements SH des enzymes, perturbant ainsi le métabolisme des
champignons à 3 niveaux : inhibition de l'oxydation du glucose, inhibition de la synthèse
d'acide nucléique, inhibition de la dégradation des acides gras. Les dithiocarbamates sont
entre autres le mancozèbe. le manèbe, le métam-sodiurn, le ferbame, le nabame, le
cuprobane, le carbatène, le métirame-zinc, Je thirame. Ces fongicides ont en commun leur
absence totale de phytotoxicité sauf pour le nabame et ont même une certaine influence
favorisante sur la végétation, une polyvalence assez grande et une très faible toxicité
(Cluzeau, 1995 ; Couteux et Lejeune, 2005)
1.1.3.2. Fongicides uni-sites
Les fongicides uni-sites sont des produits qui affectent une réaction biochimique bien
précise et sont spécifiques à une classe de champignon ou polyvalents. La plupart sont
8

systémiques c'est-à-dire sont absorbés par la plante, se déplacent à l'intérieur de celle-ci
pour inhiber des phénomènes de biosynthèses précis et présentent donc l'avantage d'être à
la fois préventifs et curatifs. Couteux et Lejeune (2005) ont classé certains fongicides uni
sites en fonction des processus biochimiques que ces fongicides affectent.
a. Fongicides affectant les processus respiratoires et
la production d'énergie cellulaire
Les processus respiratoires inhibés par ces fongicides sont multiples.
~ L'inhibition du complexe mitochondrial Il (succinate-ubiquinone
réductase) est provoquée par le carboxine et Je flutolanil qui appartiennent à
la famille des amides ou amines et à la sous-famille des anilides ou
phénylamides. Ces fongicides sont spécifiques des Basidiomycètes.
~ Le complexe mitochondrial III (face externe du cytochrome B « Qol »)
est inhibé par le fenamidone et le famoxadone qui sont des carbarnates
appartenant à la famille des hétérocycles azotés et aux sous-familles des
imidazolinones et des oxazolidinedones, respectivement. Ce complexe
mitochondrial est aussi affecté l'azoxysrobine, le krésoxim-méthyl, le
picoxystrobine, le pyraclostrobine et le trifloxystrobine qui sont aussi des
carbamates hétérocycliques azotés et appartenant à la sous-famille des
strobilurines.
~ La phosphorilation oxydative est découplée par le fluazinarn et le dinocap
qui appartiennent à la famille des amides ou amines et aux sous-familles des
pyridinamioes et des dérivés du phénol, respectivement.
~ La libération de l'ATP (adénosine triphosphate) est inhibée par le
sithiofam de la famille des silylamides (Couteux et Lejeune, 2005).
b. Fongicides agissant sur le métabolisme des
glucides et des polyols
Ces fongicides qui appartiennent aux familles des dicarboximides ou imides
cycliques, des carbamates hétérocycliques azotés et des organophosphorés agissent sur
l'histidine et la protéine kinase pour inhiber la biosynthèse des polyols et l'osmorégulation.
Les dicarboximides concernés sont l'iprodione, le procymidone et le vinchlozoline.
9

L'hétérocylce azoté et l'organophosphoré concernés sont respectivement le fludioxonil et
le tochlofos-méthyl appartenant aux sous-familles des pyrolles et des phosphates
respectivement (Couteux et Lejeune, 2005).
c. Fongicides affectant la biosynthèse des lipides
C'est un groupe important regroupant en plusieurs familles la moitié des matières
actives. La plupart provoquent l'inhibition d'enzymes impliquées dans la synthèse des
stérols, entraînant une perturbation du fonctionnement et de la formation des membranes
cellulaires des champignons. Ces inhibiteurs de la biosynthèse des stérols (IBS) ont
généralement une action systémique, mais certains sont translaminaires (prochlorase). Les
IBS sont classés en deux groupes. Les IBS du groupe I inhibent la 14-a déméthylase et
provoquent la déméthylation des stérols. Ces IBS sont des carbamates hétérocycliques
azotés appartenant aux sous-familles des imidazoles, des pyrimidines et des triazoles. Les
imidazoles concernés sont l'imazalil, le prochloraze et le prochloraze-manganèse tandis
que les pyrimidines sont le fénarimol et le nuarimol. Les triazoles concernés sont entre
autres le metconazole, le flutriafol, le penconazole, le triadiméfon et le propiconazole. Les
IBS du groupe Il inhibent sur la ôl4 réductase et sont entre autres le fenpropimorphe, le
dodémorphe et le tridémorphe appartenant à la famille des hétérocycles azotés et à la sous
famille des morpholines (Couteux et Lejeune, 2005).
d. Fongicides affectant la biosynthèse des acides
aminés ou des protéines Ces fongicides agissent sur la B-lyase et perturbent la synthèse de la méthionine qui
est le premier acide aminé produit lors de la traduction de l'acide ribonucléique (ARN)
messager. Les matières actives des fongicides inhibant la synthèse des acides aminés sont
le cyprodinil, le mepanipyrin et le pyriméthanil. Elles sont des carbamates hétérocycliques
azotés et appartenant à la sous-famille des anilinopyrimidines (Couteux et Lejeune, 2005).
e. Fongicides affectant la biosynthèse des acides
nucléiques et de leurs précurseurs
La biosynthèse de I' ARN est affectée à travers l'inhibition de l'adénosine
désaminase par le buprimate ou à travers l'inhibtion de l' ARN-polymérase 1 par le
bénalaxy et le méfenoxarn. Le bupirimate est un hétérocycle azoté appartenant à la sous
famille des pyrimidines tandis que le bénalaxy et le méfenoxam sont des amides
10

appartenant à la sous-famille des phénylamides. Le bénalaxy et le méfénoxam sont
spécifiques des Oomycètes (Couteux et Lejeune, 2005).
La biosynthèse de l'acide désoxyribonucléique (ADN) est inhibée par l'hymexazol
qui appartient à la famille des carbamates hétérocycliques azotés et à la sous-famille des
isoxazoles (Couteux et Lejeune, 2005).
f. Fongicides agissant sur la division cellulaire et les
microtubules
Une fois absorbés, ces produits se transforment en carbendazime qui est un
antimitotique. Ces produits bloquent la division cellulaire et nucléaire (mitose) en
perturbant la formation et le fonctionnement du fuseau achromatique. La carbendazime a
de plus une action au niveau de J'ADN: elle se substitue aux bases puriques (adénine et
guanine) des acides nucléiques et provoquent des erreurs dans la transcription du génome.
Ces fongicides sont entre autres la carbendazime, le thiabendazole et le méthyl-thiophanate
qui sont des carbamates hétérocycliques azotés appartenant à la sous-famille des
benzimidazoles (Couteux et Lejeune, 2005).
11

1.2. Généralités sur le bananier
1.2.1. Origine et Systématique
Le bananier serait originaire d'Asie du Sud-est. Il est une Monocotylédone
(Deysson, 1967) qui appartient à la famille des Musaceae qui comprend deux
genres: Musa et Ensete. Le genre Musa est constitué d'environ 40 espèces dont celles des
bananiers à fruits comestibles actuellement cultivés dans la zone tropicale. Les bananiers
desserts sucrés et les bananiers plantains cultivés en Afrique de l'Ouest appartiennent
respectivement aux groupes génomiques AA, AAB (Bakry et al., 1997).
1.2.2. Description du bananier
Le bananier est une herbe géante (Lassoudière, 2007). À maturité, le plant de
bananier se compose d'une tige souterraine appelée rhizome portant un système radiculaire
fasciculé dont 80% des racines ne dépassent pas 1 m (Lavagine, 1997). Les feuilles dont
les gaines, imbriquées les unes dans les autres, forment le pseudo-tronc qui porte une
inflorescence dont la partie proximale est occupée par les fleurs femelles et la partie distale
par les fleurs mâles (Figure l ). Les ovaires des fleurs femelles donnent des fruits
parthénocarpiques appelés bananes ou doigts (Lassoudière, 1978). Un régime de fruits peut
peser entre 5 et 70 Kg.
1.2.3. Ecologie du bananier
Le bananier est une plante adaptée aux zones des régions équatoriales et sub
tropicales. Les besoins en eau sont estimés à 125-150 mm /mois. La température optimale
de développement est de 28 °C. Un éclairement d'une durée de 2200 heures par an est utile
au bon développement du bananier. Le vent peut causer la chute des bananiers. De ce fait,
l'utilisation des zones abritées du vent ou des haies brise-vent, ou encore de tuteurs s'avère
nécessaire (Tezenas, 1985). La plante préfère un sol bien structuré, léger, aéré
(Lassoudière, 1978) avec un pH compris entre 6 et 7,5. Les besoins sont plus importants en
azote et en potassium. La culture du bananier nécessite seulement des faibles quantités de
phosphore, de calcium et de magnésium (Dessaw, 1988). La culture de la banane permet
de lutter contre la pauvreté en participant à la diversification des revenus dans les zones
rurales et urbaines, au Produit Intérieur Brut (PIB) en Afrique centrale et occidentale
(Temple et Tentchou, 2000). De plus, les bananes sont aussi utilisées dans les domaines de
la médecine, de la cosmétique et de l'industrie de fabrication des fibres et de l'alcool
(Dadzie, I 995). 12

Hampe ou axe de l'inflorescence
Mains de fleurs femelles ou bractée de bananes (doigts)
Pseudo tronc
Rejet fils
Tige ou souche ou bulbe ou rhizome Racine
Figure 1 : Schéma d'un bananier en phase de fructification (Aby, 2013)
13

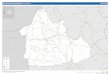
1.2.4. Productions de la banane en Côte d'Ivoire
1.2.4.1. Productions de la banane dessert en Côte d'Ivoire
La zone de production de la banane dessert est principalement située dans un
rectangle délimité, d'une part, par les 5e et 6e degrés de latitude Nord, et d'autre part, par
les 3e et 5e degrés de longitude Ouest (Figure 2) ; dans un rayon de 200 Km autour
d'Abidjan (Lassoudière, 1978). La production en 2009 a été de 399.000 tonnes
(ODEADOM, 2010) et la filière emploie environ 22 000 personnes. Le secteur de la
banane a réalisé 7 à 8 % de la valeur ajoutée agricole, il a représenté 2 à 3 % du PIB
national et 1,3 % de la valeur des exportations en 2007 (OCAB, 2012). La production de la
banane dessert est du ressort de grandes compagnies multinationales telles que la SCB, la
SBM et la SAKJ auxquelles sont associés quelques rares petits producteurs. Les surfaces
exploitées sont estimées à plus de 5000 ha de culture pure (Kobenan et al., 2006, Aby et
al., 2008).
1.2.4.2. Productions de la banane plantain en Côte d'Ivoire
La zone de production de la banane plantain inclut les limites de production de la
banane dessert et d'autres zones (Figure 2). Elle correspond au secteur préforestier du
domaine guinéen (Koulndoh, 1986) et est limitée dans sa partie Nord par une ligne passant
par Bondoukou, Sud de Bouaké, Oumé, Bouaflé, Daloa, Man et Danané. La banane
plantain est en général associée aux cultures industrielles (hévéa, café, cacao ... ) et
vivrières. Cependant, certaines plantations sont en culture pure. La production ivoirienne a
été de 1.700.000 tonnes en 2010 (Ministère de l'environnement et du développement
durable, 2013).
1.2.5. Principales contraintes biotiques à la culture du bananier
Le bananier subit l'action de nombreux bio-agresseurs qui se rencontrent dans
plusieurs groupes : les virus, les bactéries, les nématodes, les champignons et les
arthropodes.
14

M~Ll I N
t
A
• • •.. •
~ SC8T1ssal6 ·~ /
COIICITissai.
J ::. SPDCle T1ssal6
SCB Banacomoé
r.•
Ealln Aabovllle
11 &tin Azamllé
N
t
Diby
B
SPDBde Dabou :-,
coaa Az.aaulé ~ • - Rouchard 'I ~ ,-..
• • ( sbmk
Echelle : l/1000 000
Figure 2: Zones de production des bananes et bananes plantains en Côte d'Ivoire. A : limites septentrionales, B : position des plantations de banane dessert dans les régions productrices. - - Limite septentrionale de la zone de production de banane plantain - Limite septentrionale d'exploitation de la banane dessert C:=J Agnéby C:=J Lagune C:=J Sud Comoé
i.:_ i:&·; !Positions des plantations de banane dessert (Gnonhouri el al. 2008, adapté Aby)
15

1.2.5.1. Viroses et bactérioses du bananier
Les maladies virales sont souvent liées aux infections généralisées de la plante,
excepté le méristème (Lassouclière, 2007). Les plus répandues sont la chlorose infectieuse
causée par le Cucumber Mosaic Virus, et la mosaïque en tirets du bananier causée par le
Banana Bract Mosaic Virus. qui occasionnent d'importantes pertes (Nazaire et al., 2008;
Aka et al., 2009).
Les maladies bactériennes entravent aussi la production de cette culture. Ce sont
entre autres le flétrissement dû à Xanthomonas campestris pv. Musacearum.
(Tusbemereirwe et al., 2003) et la maladie de Moko, causée par Ralstonia solanacearum
Smith qui entraîne le nanisme et la malformation d'organes (Smith, 2006).
1.2.5.2. Mycoses du bananier
Les principales contraintes parasitaires du bananier sont des attaques fongiques
aériennes liées à trois espèces du genre Mycosphaere/la: M musicola, l'agent causal de la
maladie de Sigatoka, encore appelée cercosporiose jaune, M. fijiensis, l'agent causal de la
maladie des raies noires ou cercosporiose noire et M eumusae, l'agent causal de la maladie
des taches foliaires (Mulder et Stover 1976 ; Fouré et Mouliom, 1988).
Les attaques fongiques souterraines liées au champignon tellurique Zythia sp.,
causent des nécroses qui, par coalescence, engainent la racine du bananier et entrainent la
perte des fonctions d'absorption (Kobenan et al., 1997).
1.2.5.3. Ravageurs du bananier
a. Nématodes phytophages
Les phytonématodes les plus dévastateurs sont entre autres les endoparasites
migrateurs Radopholus similis et Pratylenchus cojfeae (Lassoudière, 2007). Ils
endommagent les racines du bananier, seuls ou associés à des champignons des genres
Cylindroc/adium, Fusarium (Loridat, 1989) en formant un complexe parasitaire racinaire
(Risède, 2002). En Côte d'Ivoire, 19 espèces de nématodes appartenant à 15 genres ont été
décrites sur les bananiers plantains (Adiko, 1988). La paire P. cojfeae-R. similis a été
associée au raccourcissement du cycle de culture des bananeraies du Sud comoé
(Gnonhouri et al., 2009).
16

b. Charançon noir du bananier, Cosmopolites sordidus
Le charançon noir, Cosmopolites sordidus, est le ravageur d'importance mondiale
du bananier (Lassoudière, 2007). Il provoque des pertes de rendement de l'ordre de 70% en
absence de contrôle chimique (Ganry, 2004).
Originaire de l'Asie du Sud-est, Le charançon noir du bananier (Figure 3)
Cosmopolites sordidus GERMAR (Coleoptère: Curculionidae) s'est diffusé dans toutes les
régions tropicales et subtropicales productrices de bananes et de bananes plantains.
Ce ravageur a une longue durée de vie (2 ans et plus) mais une faible fécondité
(Treverrow et al., 1992). C. sordidus est un insecte nocturne caractérisé par un fort
hygrotropisme et un grégarisme. Les adultes se nourrissent des tissus de bananier en
décomposition (Budenberg et al., 1993). Les femelles pondent préférentiellement dans les
gaines foliaires et à la partie supérieure du bulbe des plants ayant atteint le stade de la
floraison. A l'émergence, la larve qui passe par 5 à 7 stades (Figure 4) se nourrit du
rhizome, des tissus foliaires du pseudo tronc et cause ainsi d'énormes dégâts en creusant
des galeries (Figure 5). Ces dommages sont entre autres la section des vaisseaux
conducteurs, des racines émises et la destruction des assises racinaires (Gold et al., 1994;
Cerda et al., 1995).
1.2.6. Méthodes de lutte contre Cosmopolites sordidus
1.2.6.1. Pratiques culturales
Certaines pratiques culturales permettent de créer un environnement défavorable à
la multiplication de cet insecte et stoppent sa migration. Il s'agit entre autres:
• de l'usage de matériels sains tels que les vitro plants;
• de la capture systématique des adultes à travers des pièges à pseudo tronc (Figure
6) et des pièges à phéromone seule ou le plus souvent en association avec les
champignons entomopathogène (Tinzaara et al., 2002) ;
• du maintien de la propreté de la plantation car les résidus de récoltes et
l'enherbement constituent une source d'alimentation, un site de reproduction
et de refuge des charançons (Treverrow et Bedding, 1993)
• de l'association de culture qui maintient la pullulation de l'insecte à un niveau bas par manque d'habitats propices et surtout par l'accroissement d'ennemis naturels;
• de l'exploitation des variétés résistantes telles que les cultivars plantains Kedong
et Kekang utilisés au Cameroun (Fogain et Price, 1994).
17

Soie
Antenne
Patte postérieure
Ponctuation sur l'élytre
Strie sur l'élytre
Ponctuation sur le thorax
Patte médiane
Patte antérieure
Rostre Thorax Abdomen
Figure 3: Dessin de l'adulte du charançon noir du bananier C. sordidus (Aby, 2013).
Figure 4 : Stades de la métamorphose chez C. sordidus (Hoarn, 2005). A : œufs déposés à la base de la souche de bananier B : Stades larve-adulte, LJ : larve au stade 1, L3 : stade 3 ; Ij : jeune imago, Ny: Nymphe, lm : Imago mature
Pseudo tronc de bananier
Fragment de pseudo tronc
Figure 5: Décorticage tangentiel d'une souche Figure 6 : Piège de Vilardebo utilisé pour la de bananier montrant les galeries larvaires (Aby, collecte des charançons (Aby, 2013) 2013)
18

1.2.6.2. Lutte chimique contre le charançon noir du bananier
La lutte contre le charançon noir du bananier par Ja voie chimique s'opère
actuellement sous deux formes : la lutte chimique classique et la lutte chimique sur
avertissement biologique.
c. Lutte chimique classique
C'est la première méthode de lutte chimique. Elle repose sur l'emploi systématique
d'insecticides spécifiques ou de nématicides à effets insecticides pour protéger le matériel
de plantation (immersion des rejets dans la solution d'insecticide) et les bananiers par des
applications périodiques à la base de ces derniers. Elle est d'une efficacité immédiate. Elle
a commencé par l'usage des pesticides non synthétiques tels que l'arsenate de calcium et
de barax (W eddel, 1945) qui ont été suivis à partir de 1951 par les insecticides synthétiques
(Vilardebo, 1984; Simon, 1994). Plusieurs classes de composés chimiques ont été testées
et recommmandées, entre autres 1 'aldicarb, l' aldrin, le chlordane, le
dichlorodiphényltrichloroéthane (DDT), l'hexachlorocyclohexane (HCH) etc (Sponagel et
al., 1995 ; Seshu et al., 1998).
Les insecticides à base de cyclodiènes (dieldrin et aldrin) ont été pendant longtemps
recommandés et utilisés contre le charançon noir du bananier. Ces molécules ont été
abandonnées face au développement de popuJatioos de charançons résistants signalés dans
toutes Jes aires de production et du fait de la pollution de l'environnement. Le pesticide
efficace le plus utilisé fut le chlordécone. Cependant, son utilisation abusive a causé des
foyers de résistance. TI en a résulté une augmentation des doses d'utilisation. Aujourd'hui,
les résidus du chlordécone sont souvent rencontrés dans les vivriers qui sont rejetés des
marchés d'exportation (Luc et al., 2010).
Des organophosphorés moins rémanents existent, mais sont très toxiques et plus
coûteux. Aussi, il est assuré que le charançon peut développer une résistance à la plupart
des familles de pesticides (Gold et al., 1999 ; Gold et al., 2003).
d. Lutte chimique par avertissement biologique
Dans le principe de la lutte chimique par avertissement biologique, la décision de
traitement ou d'intervention chimique est liée au niveau réel et actueJ des dégâts sur le bananier spécifiquement, à l'importance et à la nature des galeries larvaires. Elle consiste à
n'intervenir que là où cela est réellement nécessaire. Cette méthode repose sur l'estimation
19

actuelle du taux et de l'intensité des attaques pour situer un seuil d'intervention qui
lorsqu'il est atteint, impose de faire des applications d'insecticides.
En Côte d'Ivoire, depuis l'initiation de l'avertissement biologique, les traitements
systématiques réalisés 4 à 6 fois par an, sont passés en moyenne entre 1 et 2 traitement(s)
par an (K.éhé, 2006), ce qui constitue des économies énormes et une voie de préservation
de l'environnement.
1.2.6.3. Lutte biologique contre le charançon noir du bananier
Cette lutte est l'usage d'un ou de combiné d'ennemis naturels (parasitoïdes,
prédateurs, agents pathogènes), d'antagonistes pour maintenir à un seuil tolérable la
population d'un ennemi de la culture (Debach, 1964; Van Driesche et Bellows, 1996).
a. Lutte biologique contre le charançon par utilisation de
prédateurs
La lutte biologique contre le charançon par les ennemis naturels a
commencé dans son aire d'origine en Asie du Sud Est, par la recherche de tous les
prédateurs opportunistes. Le coléoptère, Plaesius javanus Erickson (Coléoptère :
Histeridae) s'est montré le plus efficace. Les adultes et les larves attaquent les
larves du charançon (Gold et al., 2003 ).
Néanmoins. des résultats satisfaisants ont été obtenus avec ! 'usage des
fourmis myrimicines Pheidole megacephala (Hymenoptère : Formicidae) à Cuba.
Les potentialités d'utilisation de ces fourmis font objet d'études courantes (Gold
et al., 2003).
b. Lutte biologique contre le charançon par utilisation de
champignons entomopatbogènes
Le plus grand nombre de champignons pathogènes se trouvent dans la classe des
Zygomycètes, mais les plus utilisés en lutte biologique proviennent des Deuteromycètes
(champignon imparfait). Les espèces des genres, Beauveria, Metharizium, Verticillium,
Erynia, Hirsute/la, Entomophtora et Entomophaga sont les plus utilisées en lutte biologique (Wraight et Roberts, 1987; Goettel, 1992). Beauveria bassiana et Metharizium anisoplae
ont été éprouvés comme agents de lutte biologique contre le charançon noir du
bananier (Nank.inga, 1994 et 1997).
20

bl. Champignon Beauveria bassiana
Ce champignon imparfait est pathogène pour de nombreux insectes. B. bassiana
est un agent de lutte biologique très intéressant du fait qu'il peut infecter l'hôte par
ingestion ou par simple contact contrairement aux autres agents microbiologiques. Tous les
stades de l'insecte (oeuf, larve, adulte) sont sensibles. Son large spectre d'action et sa
virulence ont permis de l'utiliser avec succès en milieu agricole en Ex-URSS et en Europe
de l'Est (Lipa, 1985). Les premiers résultats positifs dans la répression des ravageurs avec
B. bassiana en Amérique ont été obtenus aux USA (Roberts; 1981; Carbonell et al.,
1985). En Côte d'Ivoire, un test de l'échantillon commercial de B. bassiana sur deux
sites expérimentaux (domaine ELIMA et plantation SBMK) a permis d'obtenir un
taux moyen de charançon mort de 33 %.
b2. Champignon Methuriziun anisoplae
- Taxonomie de Metarhizium anisopliae Metarhizium anisopliae est un champignon mitosporique (conidie, Figure 7) avec
une reproduction asexuée. Il est classé parmi les Hyphomycètes (Cloyd, 1999), du phylum
des Deutéromycètes appelés aussi champignon imparfait. La position systématique selon
Bischoff el al. (2009) est :
Règne: Sous règne: Phylum: Classe: Ordre: Famille: Genre: Espèce:
Champignon Dicaryon Deutéromycètes Sordariomycètes Hypocreales Clavicipitaceae Metarhizium Metarhizium anisopliae
- Biologie de M. anisopliae
M. anisopliae est un champignon tellurique naturellement présent dans tous
les écosystèmes à travers le monde. Il cause diverses parasitoses chez les insectes.
Il est reconnu parasite de plus de 200 espèces (Cloyd, 1999). Il est couramment
utilisé comme insecticide biologique contre divers ravageurs de cultures et
vecteurs de maladies. La distribution de M. anisopliae dans les écosystèmes des
pays tropicaux n'a pas encore été étudiée.
21

Cependant, les études menées dans les pays tempérés et froids. montrent
que M. anisopliae est une espèce associée aux sols cultivés (Meyling et Eilenberg,
2006, Quesada-Moraga et al., 2007). Ce champignon est également associé à la
rhizosphère (Hu et St. Leger, 2002) où il semble être activé par les exudats des
racines. En dépit de ces observations, 1' insecte hôte demeure le principal
constituant de la source organique pour le développement du champignon
(Meyling et Eilenberg, 2006). La dissémination de M anisop/iae se fait par les
insectes hôtes, les arthropodes, les collemboles, le vent et les eaux de
ruissellement.
- Mode d'infection de M. anisopliae
La maladie causée par le champignon est dénommée la muscardine verte, en
raison de la couleur de ses spores (Figure 8). Lorsque la conidie entre en contact
avec la cuticule d'un hôte compatible, elle germe et produit l'appressorium.
L'appressorium produit un tube germinatif qui pénètre dans la cuticule de l'hôte
par la libération d'enzymes hydrolysantes telles que les protéases. Par la suite. le
champignon se développe à l'intérieur de l'hôte, surtout dans l'bémolymphe avant
sa mort. Une toxine peptidique cyclique (destruixine) semble aider le champignon
à surmonter les défenses de l'hôte. Après la mort de l'insecte, le champignon se
développe d'abord dans les espaces inter-segmentaires et ensuite sur tout l'hôte
avec un mycélium blanc. L'insecte mort est ensuite enveloppé d'un feutrage vert
de conidies (Freimoser et al., 2003).
- Applications pratiques de M. anisopliae comme agent de lutte
biologique et de bioinsecticide
Les applications pratiques en Afrique utilisant le champignon M
anisopliae sont développées dans les pays sahéliens et au Bénin contre les locustes et les
sauteriaux à travers un vaste projet dénommé LUBILOSA : Lutte Biologique Contre
Locustes et Sauteriaux. Le projet LUBILOSA a mis sur le marché un produit à base du
champignon appelé "Green Muscle".
22

Figure 7: Culture pure de Metarhizium sp sur milieu PDA (Photo A) et Conidies de Metarhizium sp vues au microscope photonique Gx400 (Photo B).
23

Ce biopesticide consiste en des spores du champignon Metarhizium anisoplae var. acridum
Ga.ms & Rozsypal (Deuteromycotina : Hyphomycetes) sous forme de poudre sèche
(formulation TC) ou de suspension huileuse : Formulation OF (Mohamed et Amadou,
2001).
En Côte d'Ivoire, l'usage de souches locales de Metarhizium sp dans des pièges
(Figure 9) a donné des résultats intéressants soit 70 à 86% de taux de mortalité enregistré
au niveau des charançons noirs du bananier capturés (Aby, 2013).
- Effets de quelques pesticides sur des souches locales de
Metarhlzium sp.
Ce champignon est utilisé dans un environnement où des fongicides, des
nématicides et d'autres pesticides sont fréquemment appliqués pour protéger la culture
principale. li est donc indispensable de déterminer les effets de ces pesticides sur cet agent
de lutte biologique afin d'éviter d'éventuelles inhibitions de ce champignon par ces intrants
agricoles. En effet, Striel et al. (2002) ont montré que des analogues de Strobilurine ont
inhibé la germination de spores et la croissance mycélienne de divers champignons
dont Altenaria alternata (Moshe, 2005). Les travaux de Wyss et al. (2004) ont
aussi indiqué que la plupart des fongicides inhibait la germination de Phomopsis
amaranthicola, un bioherbicide utilisé contre Amaranthus spp. De même que les
travaux de Grant et al. (1999) ont montré que la germination des conidies de
Colletotrichum gloeosporioides, un bioherbicide utilisé contre Malva pusilla a été inhibée
par la majorité des fongicides testés. Concernant les souches locaJes de Metarhizium sp
les travaux d' Aby (2013) ont montré que les fongicides méthyl-thiophanate et
propiconazole ont induit de faibles taux d'inhibition (inférieurs à 10%) sur leur
germination contrairement au trifloxystrobine qui a provoqué des taux d'inhibition alJant
de 70 à 90%. Ces mêmes fongicides ont été plus ou moins compatibles avec la croissance
mycélienne et la sporulation des isolats de Metarhizium sp. Ce champignon serait aussi
compatible avec le nématicide oxamyl et l'herbicide glyphosate (Aby, 2013).
24

Duvet mycélien
Amas de conidies
Figure 8: Charançon noir du bananier présentant la muscardine verte causée par Metarhizium sp.
Morceau de pseudo tronc
Contre plaqué
Figure 9: Piège à contre-plaqué muni d'inoculum de l'isolat de Metarhlzium sp BME2 (Aby, 2013) A: Piège stabilisé par un morceau de pseudo tronc. Les flèches rouges indiquent l'espace d'entrée des charançons. 8: Piège recouvert par un film de polystyrène.
25

' DEUXIEME PARTIE : MATÉRIEL ET MÉTHODES
26

2.1. Site d'étude
Nos travaux ont été effectués au laboratoire de phytopathologie de la Station de
Recherche du CNRA (Centre National de Recherche Agronomique) à Bimbresso. Cette
station est située dans la sous-préfecture de Songon (Figure 10), au Sud-ouest d'Abidjan,
sur l'axe routier Abidjan-Dabou. Ses coordonnées géographiques sont les suivantes: 5°22
latitude Nord, 4 °08 longitude Ouest.
2.2. Matériel
2.2.1. Matériel de laboratoire
Le matériel de laboratoire a été essentiellement composé d'un autoclave, un
agitateur aimenté, un microscope optique, des lames et lamelles, un compteur mécanique
l 'hématimètre de Malassez, de pipettes et micropipettes, de ruban adhésif. de la
mousseline, de l'eau distillée un appareil photo numérique et de la verrerie.
2.2.2. Champignon entomopathogène
Le champignon entomopathogène faisant 1 'objet de cette étude est une souche
locale de Métarhizium sp isolée sur des charançons morts récoltés dans la bananeraie
BA TIA (Tiassalé ; Aby et al., 2008).
2.2.3. Insecte ravageur
L'insecte ravageur qui est l'objet de cette étude est le charançon noir du bananier
Cosmopolites sordidus. Les charançons ont été collectés dans les bananeraies ivoiriennes
puis élevés à la Station de Recherche du CNRA à Bimbresso.
2.2.4. Pesticides utilisés
Différentes concentrations de trois matières actives de fongicides et de deux matières actives de nématicides ont été testées (Tableau 1 ).
2.3. Méthodes Cette étude a été réalisée pour déterminer l'effet de quelques fongicides et
nématicides sur la germination, la croissance mycélienne, la sporulation et le pouvoir
infectieux des conidies de l'isolat BME 2 de Metarhizium sp. Tous les tests ont été
réalisés deux fois.
27

2.3.1. Préparation des milieux de culture
2.3.1.l. Préparation des solutions mères de 1000 ppm
La solution mère a été obtenue en dissolvant le pesticide commercial dans l'eau
distillée stérile. Ainsi, 0,1 ml de méthyl-thiophanate (400 g/1), 1,6 ml de
propiconazole (250 g/l) ont été dissouts chacun dans 40 ml d'eau distillée stérile
tandis que 0,4 ml de trifloxystrobine (75 g/l) a été dissout dans 30 ml d'eau
distillée stérile pour obtenir une concentration de 1000 ppm. Concernant les autres
pesticides, 0,2 ml d'oxamyl (240 g/1) et 0,4 g d'ethoprophos () 00 g/kg) ont été
respectivement dissouts dans 48 ml et 40 ml d'eau distillée stérile pour obtenir
aussi une concentration de 1000 ppm.
2.3.1.2. Amendement des milieux de culture avec les pesticides
Deux milieux de culture ont été utilisés: le milieu gélosé (20 g d'agar pour 1 litre
d'eau distillée) et le milieu PDA; c'est-à-dire Potato Dextrose Agar (20 g de pomme de
terre, 20 g de dextrose et 20 g d'agar pour 1 litre d'eau distillée). Les milieux ont été
préparés dans des erlenmeyers et autoclavés pendant 30 min à 121 °C sous une pression de
1 bar.
A partir des solutions mères, différentes quantités d'un pesticide (Tableau 1) ont été
incorporées dans les milieux de culture contenus dans les erlenmeyers autoclavés. Le
mélange a été ensuite bien homogénéisé à l'aide d'un agitateur à raison de 150 tr/min
pendant 30 secondes. Puis, ce mélange a été distribué dans des boîtes de Petri à raison de
10 à 15 ml par boîte.
2.3.2. Effet des pesticides sur la germination des conidies
Les spores ont été obtenues après culture des isolats sur milieu PDA
pendant 21 jours. Une suspension de conidies a été faite dans de l'eau distillée
stérile. Cinq gouttes de suspension de conidies ont été déposées sur milieu gélosé
amendé du pesticide étudié et sur milieu gélosé sans pesticide pour le témoin.
Les conidies ont été étalées par légère inclinaison des boîtes créant ainsi une trainée
qui a été suivie lors du comptage des conidies germées. Trois répétitions ont été réalisées
pour chaque concentration du pesticide testé. Les boîtes de Petri ont été disposées de
manière aléatoire sur un plancher, à l'obscurité, dans une chambre d'incubation pendant 48
h à la température ambiante.
28

 N
Région de rAgnéby
'· ' Any;ama • Département de Dabou
Océan Atlantique
Limite, de Région [j] Capitale économique Limite de département • Sous-préfecture Limite di, Sous-préfecture Cours d'eau
0 S lOKm
Source : Ministère de fintérieur et de la décentralisation, 2009.
Figure 10: Situation géographique de la sous-préfecture de Songon (Ministère de l'intérieur et de la décentrai isation, 2009)
Tableau 1: Différentes doses des fongicides et nématicides utilisées Pesticides Matière active Doses (ppm)
Trifloxystrobine 0 0,01 0,03 0, 1 0,5 1 (Strobilurines)
Fongicides Propiconazole 0 0,01 0,03 0,1 (Triazoles)
Méthyl-tbiophanate 0 0,5 1 3 5 (Benzimidazo les)
Nérnaticides Oxarnyl (Carbamates) 0 3 5 10 15 Insecticide- Ethoprophos 0 3 5 10 15 Nérnaticide (Organophosphorés)
29

2.3.3. Effet des pesticides sur la croissance mycélienne
L'effet d'un pesticide sur la croissance mycélienne a été étudié sur milieu PDA
amendé par ce pesticide et sur un milieu PDA sans pesticide servant de témoin. Une
rondelle de 5 mm de diamètre de culture pure de 15 jours d'âge a été déposée au centre de
la boîte de Petri contenant le milieu de culture. Les fronts de croissance du mycélium sur
deux axes perpendiculaires dont le point séquent est le centre de la rondelle. ont été
mesurés tous les deux jours jusqu'à la confluence de la boîte de Petri.
Trois répétitions ont été effectuées pour chaque concentration de chaque pesticide.
Les boîtes de Petri ont été mises dans une chambre d'incubation avec une photopériode de
12 b à la température ambiante.
2.3.4. Effet des pesticides sur la sporulation
Après 21 jours de culture de l'isolat sur le milieu PDA amendé du pesticide étudié
ou sur milieu PDA sans pesticide. la colonie formée dans une boîte a été raclée avec une
spatule stérile et agitée à 150 tours/min pendant 30 secondes. Dans 100 ml d'eau distillée
stérile, la solution obtenue a été filtrée deux fois sur de la mousseline et sa concentration
déterminée à l'hématimètre de Malassez dont les coefficients des périmètres de comptages
sont mentionnés dans le tableau 2. Pour chaque dose de pesticide, trois répétitions ont été
effectuées.
2.3.5. Pouvoir germinatif des conidies produites après tratement de fongicide
Après 21 jours de culture sur milieu PDA amendé du fongicide étudié, une
suspension de spores a été faite. Cinq gouttes de suspension de conidies de l'isolat ont été
déposées sur milieu gélosé sans pesticide. Les conidies ont été étalées par légère
inclinaison des boîtes créant ainsi une trainée qui a été suivie lors du comptage des
conidies germées. Trois répétitions ont été réalisées pour chaque traitement. Les boîtes de
Petri ont été disposées dans une chambre d'incubation pendant 48 h à la température
ambiante avec une photopériode de 12 heures.
2.3.6. Pouvoir infectieux des conidies produites après traitement de fongicide
Les suspensions de conidies obtenues après traitement de fongicide ont été utilisées
pour inoculer les charançons. Ces supensions ont d'abord été diluées pour obtenir une
concentration identique. Deux gouttes de Tween-80 ont été ajoutées à chaque
suspension de conidies. Pour chaque traitement, cinq charançons ont été immergés pendant
30

Tableau 2 : Coefficients des périmètres de comptage de l'hématimètre de Malassez
Périmètres de Dimensions (mm) Volumes Volumes (ml) Coefficients comptage (mm3) (ml")
Petit carré 0,05 X 0,05 X 0,2 0,5.10-3 0,5.10-6 20.105
Rectangle 0,05 X 0,25 X 0,2 2,5.10-3 2,5.10-6 4.105 horizontal
Rectangle vertical 0,05 X 0,2 X 0,2 2.10-3 2.10-6 5.105
Grand rectangle 0,2 X 0,25 X 0,2 10.10-3 10.10-6 1.105
31

30 secondes dans la suspension de conidies correspondante. A cet effet ; trois répétitions
ont été réalisées pour chaque traitement. Les cinq charançons ont été déposés sur du
papier filtre imbibé d'eau distillée dans une boîte de Petri stérile. Toutes les boîtes
ont été placées à l'obscurité dans une chambre d'incubation à la température ambiante.
Les charançons inoculés ont été observés au quotidien pendant 34 jours pour
compter le nombre de charançons morts. Chaque charançon mort a été désinfecté à
l'hypochlorite de sodium (1 %) pendant 30 secondes et rincés 3 fois avec de J'eau
distillée stérile. Il a été ensuite déposé sur du papier filtre imbibé d'eau distillée
dans des boîtes de Petri stériles et mis en incubation à l'obscurité. Le délai
d'apparition du duvet mycélien a été noté. Le champignon a été identifié au
microscope photonique.
2.3.7. Evaluation de l'effet des pesticides
2.3.7.1. Effet des pesticides sur la germination des conidies
Cent spores ont été observées par boîte de Pétri. Une conidie a germé
lorsque la longueur du tube germinatif est supérieure à sa demi-longueur. Le
rapport du nombre de conidies ayant germé sur le nombre de conidies observées a
permis de déterminer le taux de germination (Tg).
Tg Nombre de conidies ayant germé
X 100 Nombre de conidies observées au microscope
Trois taux de germination ont été obtenus pour chaque traitement et pour le
témoin. Ensuite, Ja moyenne des taux de germination du témoin a été calculée.
Cette moyenne a été utilisée pour déterminer trois taux d'inhibition de la
germination (Tig) pour chaque traitement, selon la formule suivante :
T. Tg du témoin - Tg d'un traitement 1g _'----------------
Tg du témoin X 100
32

2.3.7.2. Effet des pesticides sur la croissance mycélienne
La croissance journalière du mycélium (Cm) a été déterminée selon la formule
suivante:
Cm -Nombre de mesures
mj : mesure du front de croissance du mycélium du jour j
mj-Z : mesure du front de croissance du mycélium du jour j-2
n : nombre de jours au cours desquels les mesures ont été
effectuées iusou'à la confluence de la boîte de Petri
Pour le traitement témoin, la moyenne des trois croissances journalières a été
calculée. Cette moyenne a permis de déterminer le taux d'inhibition de la
croissance du mycélium (Ticm) selon la formule suivante :
T. Cm du témoin - Cm d'un traitement 1cm _---------------
Cm du témoin X 100
2.3. 7.3. Effet des pesticides sur la sporulation
Pour chaque dose de chaque pesticide ainsi que pour le témoin, les nombres de
conidies obtenus ont permis de déterminer les concentrations de conidies en tenant compte
des périmètres de comptage de l'hématimètre de Malassez (Tableau 2) utilisés. Puis les
taux d'inhibition de la sporulation (Tisp) ont été déterminés selon la formule suivante :
Tisp Sporulation du témoin - Sporulation d'un traitement
Sporulation du témoin X 100
2.3. 7.4. Pouvoir germinatif des conidies produites après traitement de
fongicide. Le nombre de conidies ayant germées parmi cent conidies a permis de
calculer le taux de germination des conidies produites sur le milieu témoin et celui
des conidies produites après traitement de fongicide.
2.3. 7.5. Pouvoir infectieux des conidies produites après traitement de
pesticide
Le pouvoir infectieux des conidies produites après traitement de fongicide a été
évalué à travers Je temps de latence, les temps létaux pour 50 ; 90 et 100 % de charançons
inoculés et les taux de mortalité. Pour chaque traitement, le rapport du nombre de
charançons morts présentant le duvet mycélien de Metarhizium sp sur le nombre de
charançons inoculés a permis de calculer l.es taux de mortalité provoqués par les conidies
33

produites après traitement. Les taux de mortalité cumulés ont permis de déterminer le
temps létal pour 50 % (TLSO), le temps létal pour 90 % (TL90) et le temps létal pour 100
% (TL 100) de charançons inoculés.
2.3.8. Analyse statistique
Pour chaque traitement ainsi que pour le témoin, La courbe de Gauss a d'abord été
construite pour éliminer les valeurs aberrantes contenues dans les taux de germination,
croissance mycélienne et sporulation avant de calculer les taux d'inhibition
correspondants. Ensuite, ces taux dinhibtion ont été soumis à une analyse de variance à un
facteur grâce au logiciel XL STAT version 7.5.3. Puis le test de Duncan au seuil de 5 % a
été utilisé pour regrouper et classer les moyennes qui ont été significativement différentes.
Concernant les taux de germination des conidies produites après traitement, une
analyse de variance à un facteur a aussi été réalisée avec le même logiciel et les moyennes
ont été séparées et classées avec le test de Duncan. Par contre, le temp de latence, les
TLSO, TL90, TLIOO et les taux de mortalité n'ont pas été soumis à une analyse de
variance.
34

TROISIÈME PARTIE: RÉSULTATS
35

3.1. Effet des fongicides sur l'isolat de Metarhizium sp
3.1.1. Effet des fongicides sur la germination des conidies
Sur le milieu sans fongicide, le taux de germination des conidies a été de 77,59 %.
Toutefois, aucun taux d'inhibition de la germination des conidies n'a atteint 50 %.
3.1.1.1. Effet du métbyl-tbiopbaoate sur la germination des conidies
Le méthyl-thiophanate à 3 et 5 ppm (dose homologuée) a inhibé de façon
significative la germination des conidies avec respectivement 43,93 et 46.88 % de taux
d'inhibition (F = 8,885 ; P = 0,001 < 0,05). Par contre, les doses de 0,5 et 1 ppm ont
provoqué les inhibitions les plus faibles, 18,11 et 19,44 %, respectivement (Tableau 3).
L'inhibition de la germination a donc diminué proportionnellement avec la quantité du
fongicide apportée au milieu de culture.
3.1.1.2. Effet du trifloxystrobine sur la germination des conidies
L'inhibition de la germination a augmenté proportionnellement avec la quantité du
fongicide utilisée. En effet, l'utilisation du tri:floxytrobine à 1 ppm ( dose appliquée au
champ) a provoqué une inhibition de la germination des conidies de 39,94 % qui a été
significativement la plus élevée (F = 7,852; P = 0,001 < 0,05). Ensuite, suivent les doses
de OJ et de 0.5 ppm, avec respectivement 17,53 et 21.88 % de taux d'inhibition de la
germination des conidies. Il n'y a pas eu de différence statistique entre les taux d'inhibition
provoqués par les doses de 0,1 et 0,5 ppm. Puis, les doses de 0,01 et 0,03 ppm ont induit
les inhibitions les plus faibles, respectivement 2,08 et 7,75 % (Tableau 3).
3.1.1.3. Effet du propiconazole sur la germination des conidies
Concernant le propiconazole, la dose homologuée (O,lppm) a permis d'obtenir
42,54 % de taux d'inhibition de la germination des conidies. Les doses de 0,03 et 0,01
ppm de propiconazole ont provoqué respectivement 34.38 et 29.51 % de taux d'inhibition
de la germination des conidies (Tableau 3). Cependant, aucune différence significative n'a
été observée entre ces taux d'inhibition induits par les différentes doses de ce fongicide (F
= 0,940 ; P = 0,426 > 0,05). L'inhibition de la germination a donc diminué
proportionnellement avec la quantité du fongicide apportée au milieu de culture.
36

Tableau 3: Taux d'inhibition de la germination des conidies de l'isolat BME 2 de
Metarhizium sp sur milieu gélosé amendé des fongicides Methyl-thiophanate,
Tritloxystrobine et Propiconazole
Fongicides Doses (ppm)
Méthyl-thiophanate Tryfloxystrobine Propiconazole
O,Ol n.e 2,08 ± 4,06 C 29,51 ± 6,79 a
0,03 n.e 7,75 ± 6,42 C 34,38 ± 9,60 a
0,1 n.e 17,53 ± 6,42 be 42,54 ± 9,60 a
0.5 18,11 ± 5,32 b 21,88 ± 6,02 b n.e
1 19,44 ± 7,53 b 39,94 ± 6,42 a n.e
3 43,93 ± 7,53 a n.e n.e
5 46,88 ± 7,21 a n.e n.e
n.e: non effectué Les taux d'inhibition surmontés de la même lettre ne sont pas significativement différents (Duncan à 5 %)
37

3.1.2. Effet des fongicides sur la croissance mycélienne
Sur le milieu témoin, la croissance du mycélium a été en moyenne de 2,15 mm/j.
3.1.2.1. Effet du méthyl-thiophanate sur la croissance mycélienne
L'inhibition de la croissance mycélienne a été plus faible avec les doses de 0,5 et 1
ppm. Cette inhibition a augmenté proportionnellement à la quantité de fongicide apportée
au milieu de culture. L'analyse de variance a séparé les concentrations en trois groupes
différents (F = 65,730; P < 0,0001 < 0,05). Le premier groupe constitué de la dose de 5
ppm a provoqué un taux d'inhibition relativement élevé (56,52 %). Il est suivi par la dose
de 3 ppm avec un taux d'inhibition de 43,47 %. Puis le troisième groupe comprend les
doses de 1 et 0,5 ppm avec les taux d'inhibition les plus faibles, respectivement 17,87 et
13,76 % (Tableau 4).
3.1.2.2. Effet du trifloxystrobine sur la croissance mycélienne
Le taux d'inhibition de la croissance mycélienne dû au trifloxystrobine a varié avec
la concentration en matière active. Les différences significatives ont mis en évidence 3
groupes (F = 145,767; P < 0,0001 < 0,05). Le premier groupe constitué des doses de 0,5 et
1 ppm a provoqué les taux d'inhibition les plus élevés; à savoir 69,35 et 71,80 %. Le
deuxième groupe comprend la dose de 0, 1 ppm qui a provoqué 39,42 % de taux
d'inhibition. Le troisième groupe est constitué des doses de 0,01 et 0,03 ppm avec les taux
d'inhibition les plus faibles, respectivement 6,83 et 13,04 % (Tableau 4). L'inhibtion de la
croissance mycélienne a diminué proportionnellement avec la dose du fongicide apportée
au milieu de culture.
3.1.2.3. Effet du propiconazole sur la croissance mycélienne
L'inhibition de la croissance mycélienne a diminué proportionnellement à la
quantité de ce fongicide apportée au milieu de culture. Le propiconazole à 0,1 ppm a
provoqué une inhibition de 81,96 % qui est significativement supérieure au taux
d'inhibition (68, 11 %) induit par la dose de 0,03 ppm (F = 186,626 ; P < 0,0001 < 0,05). Puis la dose de 0,01 ppm a provoqué le taux d'inhibition le plus faible, 23,55 % (Tableau
4).
38

Tableau 4: Taux d'inhibition de la croissance mycélienne de l'isolat BME 2 de
Metarhizium sp sur milieu PDA amendé des fongicides Methyl-thiophaoate,
Trifloxystrobine et Propiconazole
Fongicides Doses (ppm)
Méthyl-thiopbanate Tryfloxystrobine Propiconazole
0,01 n.e 6,83 ± 4,03 C 23,55 ± 2,62 C
0,03 n.e 13,04 ± 3,08 C 68,11 ± 3,45 b
0,1 n.e 39,42 ± 3,77 b 81,96±3,lOa
0,5 )3,76±2,73 C 69,35 ± 4,03 a n.e
1 17,87 ± 4,17 C 71,80 ± 3,71 a n.e
3 43,47 ± 3,86 b n.e n.e
5 56,52 ± 3,47 a n.e n.e
n.e : non effectué Les taux d'inhibition surmontés de la même lettre ne sont pas significativement différents (Duncan à 5 %)
39

3.1.3. Effet des fongicides sur la sporulation
La sporulation obtenue sur le milieu témoin a été de 44,88. 105 conidies/ml.
3.1.3.1. Effet du méthyl-thiophanate sur la sporulation
Les taux d'inhibition de la sporulation provoqués par les doses de ce fongicide ont
été relativement faibles, inférieurs à 50 %. Ces inhibtions ont été plus faibles sur les
milieux amendés avec les plus petites doses (Tableau 5). En effet, les taux d'inhibition,
20,21 ; 22,25 et 24,71, statistiquement identiques et respectivement provoqués par les
doses de 0,5 ; 1 et 3 ppm ont été les plus faibles. Par contre, la dose de 5 ppm a provoqué
48,57 % de taux d'inhibition qui a été significativement le plus élevé (F = 5,67 ; P = 0,04 < 0,05).
3.1.3.2. Effet du trifloxystrobine sur la sporulation
Les doses de 0,1 ; 0,5 et 1 ppm ont provoqué des inhibitions respectives de 89,30;
98,93 et 99,07 % qui ont été significativement les plus élevées (F = 66,89 ; P < 0,0001 < 0,05). Ensuite, suit la dose de 0,03 ppm avec 17,57 % de taux d'inhibition. Puis la dose de
0,03 ppm a provoqué - 11,21 % de taux d'inhibition (Tableau 5). La sporulation a donc
été, fortement inhibée par les doses de 0,1 ; 0,5 et 1 ppm, faiblement inhibée par la dose de
0,03 ppm, mais stimulée par la plus petite dose (0,01 ppm) utilisée de ce fongicide.
3.1.3.3. Effet du propiconazole sur la sporulation
Les taux d'inhibition provoqués par les doses utilisées de ce fongicide ont été
relativement élevés. Cette inhibtion de sporulation a diminué proportionnellement avec la
dose utilisée pour amender le milieu de culture. En effet, les doses de 0,03 et 0, 1 ppm,
classées dans le même groupe statistique, ont provoqué les inhibitions les plus élevées ; à
savoir 84,26 et 90,27 %. Ensuite, suit la dose de 0,01 ppm avec 64, 10 % de taux de
réduction de la sporulation (F = 4,07 ; P = 0,03 < 0,05).
40

Tableau 5: Taux d'inhibition de la sporulation de l'isolat BME 2 de Metarhizium sp sur
milieu PDA amendé des fongicides Methyl-thiophanate, Trifloxystrobine et Propiconazole
Fongicides Doses (ppm)
Métbyl-tbiophanate Trytloxystrobine Propiconazole
0,01 n.e - 11,21 ± 6,94 C 64,10 ± 6,79 b
0,03 n.e 17,57 ± 9,82 b 84,26 ± 9,60 a
0,1 n.e 89,30 ± 8,35 a 90,27 ± 9,60 a
0,5 20,21 ± 9,32 b 98,93 ± 8,96 a n.e
I 22,25 ± 7,12 b 99,07 ± 8,70 a n.e
3 24,71 ± 8,88 b n.e n.e
5 48,57 ± 8,88 a n.e n.e
n.e : non effectué Les taux d'inhibition surmontés de la même lettre ne sont pas significativement différents (Duncan à~%)
41

3.1.4. Pouvoir germinatif des conidies produites après traitement de fongicide
3.1.4.1. Pouvoir germinatü des conidies produites après traitement de
méthyl-thiophante
Les conidies produites après traitement de 0,5, 1 et 3 ppm du methyl-thiophanate,
classées dans le même groupe statistique, ont permis d'enregistrer des taux de germination
respectifs de 66,98, 66,53 et 61,81 % qui ont été significativement inférieurs au taux de
germination (82,75 %) des conidies produites sur le milieu témoin (F = 11,33; P = 0.0001
< 0,05). Le plus faible taux de germination (49,72 %) a été obtenu avec les conidies
produites après traitement de 5 ppm (Tableau 6). Les taux de germination les plus élevés
ont donc été obtenus avec les conidies produites après les traitements qui ont utilisé les
plus petites doses de ce fongicide.
3.1.4.2. Pouvoir germinatif des conidies produites après traitement de
trifloxystrobine
Les conidies produites sur le milieu témoin et celles produites après traitement de
0,01 ppm du trifloxystrobine ont eu des taux de germination respectifs de 82,75 et 78,40
%, qui ont été statistiquement identiques et les plus élevés (F = 16,832; P < 0,0001 <
0,05). Ensuite, les conidies produites après traitement de 0,03 et 0, 1 ppm ont présenté 65
et 56.33 % de taux de germination respectifs et ont été classées dans le même groupe
statistique. Puis, les conidies produites après traitement de 0,5 et 1 ppm, classées dans le
même groupe statistique, ont permis d'enregistrer les taux de germination les plus faibles:
45 et 40,66 % (Tableau 6). Les conidies produites après les traitements ayant utilisé les
plus petites doses ont présenté les taux de germination les plus élevés.
3.1.4.3. Pouvoir germinatif des conidies produites après traitement de
propiconazole
Le taux de germination des conidies produites après traitement de 0,01 ppm a été de
69,19 % et significativement inféreieur à celui des conidies issues du milieu témoin (F = 21,894; P < 0,0001 < 0,05). Les conidies produites après traitement de 0,03 et 0,1 ppm ont
permis d'enregistrer les taux de germination les plus faibles, respectivement 50,59 et 43,25
% (Tableau 6). Les taux de germination les plus élevés ont donc été obtenus avec les
conidies produites après les traitements qui ont utilisé les plus petites doses de ce
fongicide.
42

Tableau 6: Taux de germination des conidies de l'isolat BME 2 de Metarhizium sp
produites après traitement de Methyl-thiophanate, Trifloxystrobine et Propiconazole
Fongicides Doses (ppm)
Méthyl-thiophanate Tryfloxystrobine Propiconazole
0 82,75 ± 3,52 a 82,75 ± 2,79 a 82, 75 ± 3,82 a
0,01 n.e 78,40 ± 4,26 a 69,19 ± 5,41 b
0,03 n.e 65,00 ± 4,26 b 50,59 ± 5,41 C
0,1 n.e 56,33 ± 4,26 b 43,25 ± 5,41 C
0,5 66,98 ± 4,98 b 45,00 ± 4,83 C n.e
1 66,53 ± 5,38 b 40,66 ± 4,26 C n.e
3 61,81 ± 4,98 b n.e n.e
5 49,72 ± 4,98 C n.e n.e
n.e : non effectué. La dose de O ppm représente le témoin. Les taux d'inhibition surmontés de la même lettre ne sont pas significativement différents (Duncan à 5 %)
43

3.1.5. Pouvoir infectieux des conidies produites après traitement de fongicide
3.1.5.1. Pouvoir infectieux des conidies produites après traitement de
méthyl-thiophante
Les conidies produites après traitement de 0,5 ; 1 et 5 ppm ont montré des temps de
latence respectifs de 8 ; 3 et 3 jours qui ont été plus courts (10 jours) que celui des conidies
issues du milieu témoin (Figure 11 ). Par contre, le temps de latence de 12 jours présenté
par les conidies produites après traitement de 3 ppm a été plus long que celui du témoin. Le
temps de latence n'a donc pas varié en fonction du traitement.
Seules les conidies obtenues après traitement de 0,5 et 1 ppm ont provoqué
respectivement des TL50 de 14 et 21 jours. Ces TL50 ont été plus longs (12 jours) que
celui obtenu avec les conidies produites sur le milieu sans fongicide (Figure 11 ).
Les taux de mortalité provoqués par les conidies produites après traitement de
méthyJ-thiophanate ont varié de 33,33 à 50 %. Ces taux de mortalité ont été inférieurs à
celui (75 %) provoqué par les conidies produites sur le milieu témoin (Tableau 7). Le
délai d'apparition du duvet mycélien a varié entre 2 et 6 jours contre 3 à 6 jours pour le
témoin.
3.1.5.2. Pouvoir infectieux des conidies produites après traitement de
propinaconazole Les temps de latence de 3 et 6 jours ont été enregistrés chez les conidies produites
après traitement de 0,01 et 0,1 ppm de propiconazole. Ces temps de latence ont été plus
courts que celui présenté par les conidies produites sur le milieu témoin. Par contre, les
conidies produites après traitement de 0,03 ppm ont présenté le même temps de latence que
celui des conidies obtenues sur le milieu témoin (Figure 11 ).
Seules les conidies produites après traitement de 0,01 ppm ont présenté un TL50
de 10 jours qui a été plus court (12 jours) que le TL50 des conidies produites sur le milieu
témoin (Figure 11 ).
Les taux de mortalité provoqués par les conidies produites après traitement de ce fongicide
ont varié de 33,33 à 50 %. Ces taux de mortalité ont été inférieurs à celui provoqué par les
conidies obtenues sur le milieu témoin. Le délai d'apparition du duvet mycélien a varié
entre 3 et 6 jours de même que celui enregistré avec les conidies du milieu témoin (Tableau
7).
44

Témoin
1: l /"""""""""" 0 •••••••
1 5 9 13 17 21 25 29 33
Jours après Inoculation
100
•QI~ ~ - 50 - QI Ill •QI t: :i 0 E o ::E a
Methyl-thlophanate 0,5 ppm
1 5 9 13 17 21 25 29 33 Jours après Inoculation
Methyl-thlophanate 1 ppm 60
~ g 40 ·- CU i] 20 ~ ::, 0 u
1 5 9 13 17 21 25 29 33 Jours après Inoculation
40
Methyl-thlophanate 3 ppm
1 5 9 13 17 21 25 29 33 Jours après inoculation
Methyl-thiophanate 5 ppm
0 1 5 9 13 17 21 25 29 33
Jours après Inoculation
- 60 •CU *' ;!: ';' 40 g ~ 20 ~ E :::, u 0
Proplconazole 0,01 ppm
1 5 9 13 17 21 25 29 33 Jours après Inoculation
Propiconazole 0,03 ppm
40
20 1 r1"""'"""""' 0 ••••••••
1 5 9 13 17 21 25 29 33
Jours après Inoculation
Proplconazole 0,1 ppm
40
1:! ~ /""'"""" """ - QI 20 ! 1 0 J'""""
1 5 9 13 17 2125 29 33 Jours après Inoculation
Figure 11 : Evolution du taux de mortalité des adultes de C. sordidus inoculés avec des suspensions de conidies produites après traitement de Méthyl-thiophanate et de Propiconazole.
45

Tableau 7 : Taux de mortalité des charançons inoculés avec les conidies produites après
traitement de Méthyl-thiophanate, Trifloxystrobine et Propiconazole
Fongicides Doses (ppm)
Méthyl-thiophanate Tryfloxystrobine Propiconazole
0 75 75 75
0,01 n.e 25 50
0,03 n.e 66,66 33,33
0,1 n.e 50 33,33
0,5 50 50 n.e
1 50 16,66 n.e
3 33,33 n.e n.e
5 33,33 n.e n.e
n.e : non effectué. La dose de O ppm représente le témoin.
46

3.1.5.3. Pouvoir infectieux des conidies produites après traitement de
trifloxystrobine
Les conidies produites après traitement de 0,01 ; 0,03 et 0,5 ppm de trifloxystrobine
ont présenté des temps de latence respectifs de 3 ; 8 et 8 jours qui ont été plus courts (10
jours) que celui du témoin. Le temps de latence des conidies produites après traitement de
1 ppm a été identique à celui du témoin. Cependant, les conidies produites après traitement
de 0, 1 ppm ont présenté 15 jours de temps de latence qui a été plus long que celui des
conidies produites sur le milieu témoin (Figue 12). Le temps de latence n'a donc pas varié
en fonction du traitement.
Les conidies produites après traitement de 0,03 ppm ont provoqué un TL50 de 14
jours alors que les conidies obtenues après traitement de 0, 1 ppm et 0,5 ppm de ce
fongicide ont présenté le même TL50 de 21 jours. Ces TL50 ont été supérieurs au TL50
(10 jours) induit par les conidies produites sur le milieu témoin (Figure 12). Toutefois, les
conidies obtenues après traitement de 0,01 et 1 ppm n'ont pas présenté de TL50 au cours
des 34 jours d'observation après inoculation des charançons.
Les conidies produites après traitement de ce fongicide ont provoqué des taux de
mortalité compris entre 16,66 et 66,66 % qui ont été inférieurs au taux de mortalité (75 %)
provoqué par les conidies produites sur le milieu témoin. Le délai d'apparition du duvet
mycélien a varié entre 1 et 10 jours tandis que celui du témoin a varié entre 3 et 6 jours
(Tableau 7).
La germination des conidies, la croissance du mycélium et la sporulation ont été
observées sur les milieux amendés de fongicide. Cependant, ces trois paramètres ont été
inhibées par les fongicides utilisés. Concernant la germination, le metylthiophanate a été le
plus inhibiteur des fongicides. La croissance mycélienne quant à elle, a été plus réduite par
le propiconazole. La sporulatioo a été plus inhibée par le trifloxystrobine. Toutefois, à très
faibles concentrations (0,01 ppm), le trifloxystrobine a stimulé la sporulation de l'isolat
BME 2 de Metarhizium sp.
Les conidies produites après traitement de fongicide ont présenté des pouvoirs
germinatifs variables. Ces mêmes conidies ont été effectivement pathogènes des
charançons inoculés.
47

Trifloxystroblne 0,01 ppm 30
-«1 * 20 •.. - = CU g -~ 10
~ § 0 u 1 5 9 13 17 21 25 29 33
Jours après Inoculation
100 Trifloxystroblne 0,03 ppm
. ., ~ ~ "; 50 t:!! :::, 0 E o ~ :::,
u 1 5 9 13 17 21 25 29 33
Jours après Inoculation
Trifloxystrobine 0,1 ppm _ 60
1 ["'"""""
•CU .f. •.. - :: CU 40 "' -QI ~ "3 20 '1 § u 0 •••••••••••••
1 5 9 13 17 21 25 29 33 Jours après Inoculation
Trifloxystroblne 1 ppm 20
1 l """""""""""' - . ., ~ ~ "; 10 ~ :!! .. :::, 0 E ~ 0 •••••••• a 1 5 9 13 17 21 25 29 33
Jours après Inoculation
- 60 -QI *- •.. - :: CU 40 g ~ 20 '1 E a a
Trifloxystroblne 0,5 ppm
T
' 1 1 5 9 13 17 21 25 29 33
Jours après Inoculation
Figure 12 : Evolution du taux de mortalité des adultes de C. sordidus inoculés avec des suspensions de conidies produites après traitement de Triflox:ystrobine.
48

3.2. Effets des nématicides sur l'isolat de Metarhizium sp
3.2.1. Effet des nêmaticides sur la germination des conidies
Le taux de la germination des conidies sur Je milieu témoin a été de 77,59 %.
3.2.1.1. Effet de l'oxamyl sur la germination
Les doses de 3; 5 ; 10 et 15 ppm d'oxamyl ont respectivement provoqué des taux
d'inhibition de - 19,79; - 16,97; - 14,42 et- 11,16 % (Tableau 8). Les valeurs négatives
de ces taux d'inhibition ont montré une stimulation de la germination conidienne par ce
nématicide. Cette stimulation a été plus élevée sur les milieux amendés avec les plus
petites quantités du nématicide bien qu'il n'y ait pas eu de différence statistiques entre les
taux d'inhibition (F = 0,64 ; P = 0,604 > 0,05).
3.2.1.2. Effet de l'ethoprophos sur la germination des conidies
Les taux d'inhibition provoqués par les doses de 3; 5; 10 et 15 ppm d'ethopropbos
ont été respectivement de - 20,50 ; - 13,84 ; - 13,54 et - 13,49 % (Tableau 8). Les
valeurs négatives de ces taux d'inhibition ont montré une stimulation de la germination
conidienne par ce nématicide. Cette stimulation a été plus élevée sur les milieux amendés
avec les plus petites quantités du nématicide bien qu'il n'y ait pas eu de différence
statistiques entre les taux d'inhibition (F = 0,249 ; P = 0,859 > 0,05).
3.2.2. Effet des nématicides sur la croissance du mycélium
Sur le milieu témoin, la croissance du mycélium a été de 2,02 mm/j.
3.2.2.1. Effet de l'oxamyl sur la croissance du mycélium
L'oxamyl à 15 ppm a inhibé la croissance mycélienne de façon significative avec
33,92 % de taux d'inhibition (F = 20,39 ; P < 0,0001 < 0,05). Cependant, les taux d'inhibition de - 6,34; - 2,98 et - 0,44 % statistiquement identiques et respectivement
provoqués par les doses de 3 ; 5 et 10 ppm de ce nématicide ont montré une stimulation de
cette croissance mycélienne {Tableau 9). La stimulation de la croissance du mycélium a
donc été plus élevée sur les milieux de culture amendés avec les plus faibles doses du
nématicide.
49

Tableau 8: Taux d'inhibition de la germination des conidies de l'isolat BME 2 de
Metarhizium sp sur milieu gélosé amendé des nématicides Oxamyl et Ethoprophos
Nématicides Doses (ppm)
Oxamyl Ethoprophos
3 -19,79 ± 4,59 a -20,50 ± 9,02 a
5 - 16,97 ± 6,50 a -13,84±5,71 a
10 -14,42 ± 6,50 a - 13,54 ± 9,02 a
15 -11,16 ± 6,50 a - 13,49 ± 9,02 a
n.e : non effectué. La dose de O ppm représente le témoin. Les taux d'inhibition surmontés de la même lettre ne sont pas significativement différents (Duncan à 5 %)
Tableau 9 : Taux d'inhibition de la croissance mycélienne de l'isolat BME 2 de
Metarhizium sp sur milieu PDA amendé des nématicides Oxamyl et Ethoprophos
Nématicides Doses (ppm)
Oxamyl Ethoprophos
3 -6,34 ± 3,98 b -16,81 ± 5,26 b
5 -2,98 ± 5,09 b -13,27 ± 6,15 b
10 - 0,44 ± 4,44 b -11,75±4,83 b
15 33,92 ± 5,54 a 20,00 ± 3,55 a
n.e : non effectué. La dose de O ppm représente le témoin. Les taux d'inhibition surmontés de la même lettre ne sont pas significativement différents (Duncan à 5 %)
50

3.2.2.2. Effet de l'ethoprophos sur la croissance du mycélium
L'incidence de l'ethoprophos sur la croissance mycélienne a été une inhibition sur
les milieux de culture amendés de 1 5 ppm. Par contre, une stimulation de cette croissance
mycélienne a été provoquée par les doses de 3 ; 5 et 10 ppm. En effet, l'analyse de
variance a présenté deux groupes significativement différents (F = 19,70; P < 0,0001). Le premier groupe a été constitué de la dose de 15 ppm avec 20 % de taux d'inhibition. Le
second groupe a été composé des doses de 3 ; 5 et 10 ppm qui ont respectivement
provoqué des taux d'inhibition de- 16,81 ; - 13,27 et- 11,75 % (Tableau 9).
3.2.3. Effet des nématicides sur la sporulation
Sur le milieu témoin. la sporulation a été de 37,2.105 spores/ml d'eau.
3.2.3.1. Effet de l'oxamyl sur la sporuJation
L'incidence de l'oxamyl sur la sporulation a varié en fonction de la quantité utilisée
pour amender les milieux de culture. Des inhibitions de cette sporulation ont été
provoquées par les doses de 10 et 15 ppm tandis que des stimulations ont été induites par
les plus faibles doses, 3 et 5 ppm. En effet, quatre groupes ont été mis en évidence par les
différences significatives (F = 23,884; P < 0,0001 < 0,05). Le premier a été constitué de la
dose de 15 ppm tandis que la dose de l O ppm a été classée dans le deuxième groupe. Ces
doses de 15 et l O ppm ont respectivement provoqué des taux d'inhibition de 58.39 et
11,25 % (Tableau 9). Les doses de 5 et 3 ppm qui ont induit des taux d'inhibition respectifs
de - 18,65 et - 56,87 % ont respectivement été classées dans le troisième et le quatrième
groupe.
3.2.3.2. Effet de l'ethoprophos sur la sporuJation
Les doses utilisées de l'ethoprophos ont provoqué des effets différents sur la
sporulation. Les doses de 10 et 15 ppm inhibé la sporulation du champignon tandis que les
plus faibles doses, 3 et 5 ppm, ont stimulé cette sporulation. L'analyse de variance a en
effet mis en évidence trois groupes (F = 28,822; P < 0,0001 < 0,05). Le premier groupe a été composé des doses de 10 et 15 ppm qui a provoqué respectivement les taux d'inhibition
de 63,22 et 63,95 %. Ensuite, le deuxième groupe a été constitué de la dose de 3 ppm avec
- 2,92 % de taux d'inhibition. Puis, la dose de 5 ppm qui a provoqué - 78,69 % de taux
d'inhibtion a été classée dans le troisième groupe (Tableau 9).
51

Contrairement aux fongicides, toutes les doses utilisées des nématicides ont stimulé
la germination des conidies. Par contre, l'incidence de ces nématicides sur la croissance
mycélienne et la sporulation a varié en fonction des doses utilisées pour amneder les
milieux de culture. La croissance mycélienne et la sporulation ont été inhibées par les
concentrations les plus élevées tandis que les plus faibles doses ont induit la stimulation de
ces deux paramètres.
52

Tableau 9: Taux d'inhibition de la sporulation de l'isolat BME 2 de Metarhizium sp sur
milieu PDA amendé des nématicides Oxamyl et Ethoprophos
Nématicides Doses (ppm)
Oxamyl Ethoprophos
3 -56,87 ± 9,62 d -2,92 ± 12,02 b
5 -18,65 ± 13,61 C - 78,69 ± 18,03 C
10 11,25 ± 12,83 b 63,22 ± 17,00 a
15 58,39 ± 14,16 a 63,95 ± 16,25 a
n.e : non effectué. La dose de O ppm représente le témoin. Les taux d'inhibition surmontés de la même lettre ne sont pas significativement différents (Duncan à 5 %)
53

/
QUATRIÈME PARTIE: DISCUSSION
54

L'intérêt de ce travail est de déterminer la capacité de Metarhizium sp à germer. croître
sporuler et à infecter le charançon noir du bananier, Cosmopolites sordidus, dans un milieu
où des fongicides et des nématicides sont appliqués pour protéger Je bananier.
4. Effet des fongicides sur l'isolat de Metarhizium sp
4.1. Effet des fongicides sur la germination des conidies
La germination conidienne de cet agent de lutte biologique a été inhibée par les
trois fongicides testés, le methyl-thiophanate (Benzirnidazole), le trifloxystrobine
(Strobilurine) et le propiconazole (Triazole). Des études similaires avec Phomopsis
amaranthicola, un bioherbicide contre Amaranthus spp ont montré que la plupart des
fongicides inhibait la germination du bioherbicide (Wyss et al., 2004). De même que les
travaux de Grant et al. (1990) ont montré que la germination des conidies de
Colletotrichum gloeosporioides, un bioherbicide utilisé contre Malva pusilla a été inhibée
par la majorité des fongicides testés. Cependant les taux d'inhibition provoqués par ces
fongicides ont été relativement faibles. inférieurs à 50 % tout comme au cours des travaux
d'Aby (2013).
Les taux d'inhibition (18, 1 l - 46,88 %) relativement faibles ne seraient pas
révélateurs du mode d'action du methyl-thiophante, un carbamate hétérocyclique azoté
appartenant à la sous-famille des benzimidazoles. En effet, une fois absorbés, les
benzimidazoles se transforment en carbendazime qui est un antimitotique. Ces produits
affectent les microtubules et les divisions cellulaires (Couteux et Lejeune, 2005). Ils
inhibent la mitose du parasite en se liant à une protéine des microtubules (la tubuline)
empêchant leur réunion correcte dans le fuseau achromatique (Davidse et al., 1978). Les
travaux de Kobenan et al. (2011 et 2012) ont montré que Mycosphaerella fijiensis
responsable de la cercosporiose noire du bananier, est très sensible au methyl-thiophanante
à 5 ppm. Par contre, Je bénomyl et le thiabendazole, deux benzimidazoles, ont faiblement
inhibé la germination de plusieurs champignons tels que Fusarium oxysporum, Rhizopus
stolonifer, Saccharomyces cerevisiae, Alternaria alternata (Atrassi et al., 2007). Le
benzimidazole utilisé serait donc inefficace sur la germination de cet agent de lutte
biologique.
Les inhibitions (2 - 39,94 %) provoquées par les doses du trifloxystrobine, un
strobilurine, ont été les plus faibles. Ces résultats ne semblent pas s'accorder à ceux de
Kobenan et al. (2011 et 2012) qui ont montré que les strobilurines inhibent fortement la
55

germination de Mycosphaere/la fijiensis, le champignon responsable de la cercosporiose
noire. Des analogues de strobilurine ont aussi inhibé la germination de spores et la
croissance mycélienne de divers champignons (Stierl et al., 2002) dont Altenaria alternata
(Mosbe, 2005). Une éventuelle résistance de Metarhizium sp pourrait expliquer nos
résultats car les mêmes travaux de Ko ben an et al., (2011 et 2012) effectués sur d'autres
plantations de bananiers ont décelé des problèmes de résistance de Mycosphaerella
fijiensis à la famille des strobilurines.
Le propiconazole, un triazole, qui n'a pas pour vocation principale d'inhiber la
germination des spores a provoqué des taux d'inhibtions (29,51 - 42,54 %) semblables aux
inhibitions induits par le benzimidazole et le strobilurine testés. Ces taux d'inhibitions de la
germination conidienne provoqués par ce triazole ont été largement supérieurs à ceux
obtenus au cours des travaux d'Aby (2013).
Ces trois fongicides seraient partiellement compatibles avec la germination
conidienne de Metarhizium sp. En effet, pour Wiss et al. (2004), lorsque la germination des
conidies est partiellement réduite par un fongicide, celui-ci est classé comme étant peu
compatible avec le champignon.
4.2. Effet des fongicides sur la croissance mycélienne
La croissance mycélienne a été plus affectée que la germination des conidies et les
taux d'inhibition les plus élevés ont été aussi enregistrés par les doses homologuées des
fongicides. Ces taux d'inhibition de 56,52, 7 l ,80 et 81,96 % causés respectivement par
les doses appliquées au champ du méthyl-thiophanate, du trifloxystrobine et du
propiconazole sont conformes aux travaux d' Aby (2013).
Ces inhibitions relativement élevées confirmeraient le mode d'action du
trifloxystrobine et du propiconazole. En effet, Je trifloxystrobine, un carbamate
hétérocyclique azoté appartenant à la sous-famille des strobilurines. est reconnu comme un
inhibiteur du complexe mitochondrial li de la chaîne respiratoire et donc de la production
d'énergie cellulaire (Couteux et Lejeune, 2005). Hors, l'élongation du tube germinatif et la
croissance du mycélium sont des phénomènes qui utilisent l'énergie cellulaire. Ce
strobilurine inhiberait la croissance mycélienne en empêchant la production d'énergie
cellulaire. Concernant le propiconazole, ce carbamate hétérocyclique azoté appartenant à la
sous-famille des triazoles affecte la biosynthèse des stérols en provoquant la déméthylation
des stérols à travers l'inhibition de la 14-a déméthylase (Couteux et Lejeune, 2005). Ce
56

fongicide est un inhibiteur de la biosynthèse des stérols (IBS) qui empêche la conversion
du lanostérol en ergostérol, produit final de la constitution de la paroi cellulaire fongique
(Koller et Scheinpflug, 1987). Ainsi, les IBS inhibent la croissance du tube germinatif et la
croissance du mycélium de Metarhizium sp (Aby, 2013) et de Mycosphaerella fijiensis
(Kobenan et al., 2011 et 2012). Ce strobilurine et ce triazole testés seraient très peu
compatibles avec la croissance mycélienne de Metarhizium sp qui a été inhibée, mais
observée sur les milieux de culture amendés de ces fongicides. Pour Wiss et al. (2004)
lorsque la croissance mycélienne n'est pas totalement inhibée par un fongicide. celui-ci est
classé comme étant peu compatible avec le champignon. Les mêmes travaux de Wiss et
al. (2004) ont montré une incompatibilité entre la croissance mycélienne du bioherbicide
Phomopsis amaranthicola et certains fongicides.
Le benzimidazole testé a été le moins inhibiteur des fongicides étudiés. Les taux
d'inhibition (13,73 - 56,52 %) provoqués par les différentes concentrations de ce fongicide
pourraient indiquer que le champignon entomopathogène étudié est plus compatible au
rnethyl-thiophanate que les autres fongicides testés. Ces inhibitions relativement faibles
montreraient aussi que le benzimidazole étudié n'est pas efficace sur la croissance
mycélienne de Metarhizium sp. Des études similaires ont montré que le bénomyl et le
thiabendazole, deux benzimidazoles, inhibent la croissance mycélienne d'Aspergillus niger
et Aspergillus fumigatus mais ne sont pas efficaces sur la croissance du mycélium
d'Alternaria alternata, Saccharomyces cerevisiae et Cryptosporiopsis malicorticis (Atrassi
et al., 2007)
4.3. Effet des fongicides sur la sporuJation
La sporulation a été plus affectée que la germination des conidies. Les taux
d'inhibition de la sporulation de 90,27 et 99,07 % relativement élevés et provoqués
respectivement par les doses appliquées au champ du propiconazole et du trifloxytrobine
confirmeraient aussi leur mode d'action. Le trifloxystrobine inhiberait fortement la
sporulation en empêchant la production d'énergie cellulaire indispensable à la réalisation
de la mitose, processus par lequel ce champignon imparfait produit ses conidies. Ce
strobilurine et ce triazole testés pourraient entraver fortement la reproduction de ce
champignon entomopatbogène et donc l'infection du ravageur Cosmopolites sordidus par
Metharhizium sp. Ces taux d'inhibition très élevés (supérieurs à 90 %) indiqueraient que la
sporulation sur les milieux amendés avec les doses homologuées de ces deux fongicides a
57

été presque nulle. Cela signifierait, selon les classes de compatibilité de Wiss et al. (2004),
que les doses homologuées de propiconazole et de trifloxystrobine seraient incompatibles
avec la sporulation de Metarhizium sp. De plus les taux d'inhibition (64,10 - 98,93 %)
relativement élevés et induits par les quantités infimes du propiconazole et par trois doses
infimes de trifloxystrobine confirmeraient encore leur mode d'action. Par contre, la plus
faible dose (0,01 ppm) de trifloxystrobine pourrait stimuler la sporulation car elle a
provoqué un taux d'inhibition inférieur à zéro. À certaines concentrations, ce strobilurine
aurait donc des effets inhibiteurs ou stimulateurs sur Metarhizium sp, l'agent de lutte
biologique.
Concernant le benzirnidazole testé (méthyl-thiophanate), la dose appliquée au champ et
les quantités infimes ont induit des taux d'inhibition relativement faibles (inférieurs à 50
%). Ce fongicide qui a faiblement inhibé la germination conidienne, serait plus compatible
avec la sporulation que les deux autres fongicides testés. Le méthyl-thiophanate ne serait
donc pas un obstacle à la reproduction du champignon entomopathogène et à l'infection du
ravageur. L'effet de résistance ou de tolérance pourrait aussi expliquer les faibles
taux d'inhibition induits par le benzimidazole étudié. En effet, les fongicides
benzimidazolés sont des fongicides systémiques unisites. Contrairement aux fongicides
multisites, les fongicides unisites, possédant un seul site d'action, favorisent le
développement de résistance (Bacon, 2002). Cette résistance peut être présente
naturellement à l'intérieur de la population d'un champignon ou plus rarement résulter
d'une mutation génétique (Gilbert, 1999; Bacon. 2002 ; Sholberg et al., 2005).
La germination a été le paramètre le moins affecté par les trois fongicides testés tout
comme au cours des travaux d'Aby (2013). Ces fongicides n'empêcheraient donc pas
l'infection du ravageur par Metarhizium sp. Car selon Wyss et al. (2004) l'effet des
pesticides sur la germination conidienne est plus important que sur la croissance
mycélienne et la sporulation, car la spore est la source d'initiation de l'infection et de
dispersion de l'inoculum. De plus, les taux d'inhibition de la germination conidienne, la
croissance mycélienne et la sporulation induits par les quantités infimes des fongicides ont
été inférieurs à ceux provoqués par leurs doses appliquées au champ. Ces résultats
indiqueraient que le mode d'application aérien et la systémie de ces fongicides pourraient
réduire l'inhibition de la germination des conidies et faciliter l'infection du ravageur. En
effet, selon Aby (2013), le mode d'application aérien de ces fongicides rend difficile leur
58

contact avec les inocula protégés en premier lieu par le feuillage des bananiers et ces
fongicides étant systémiques, une infime quantité parvient au sol où se trouvent les
champignons.
4.4. Pouvoir germinatif et pouvoir infectieux des conidies produites après
traitement de fongicide
Les conidies produites après traitement de fongicide ont présenté des taux de
germination compris entre 40 et 79 % mais inférieurs à celui (82,75 %) des conidies
produites sur le milieu témoin. Les fongicides utilisés auraient donc diminué le pouvoir
germinatif des conidies produites après traitement. Cette diminution du pouvoir germinatif
s'expliquerait par la présence de résidus des fongicides à l'intérieur des conidies produites
après traitement. En effet, l'utilisation de pesticides pour améliorer la production agricole
résulte par la présence de résidus de ces pesticides dans les aliments (Chubilleau et al.,
2011 ). Le pouvoir germinatif observé chez les conidies produites après traitement a été
corroboré par les paramètres pathogéniques de ces mêmes conidies. Ces conidies produites
après traitement de fongicide ont été effectivement pathogènes des charançons inoculés. En
effet, les conidies produites après traitement de fongicide ont présenté des TL50 compris
entre 10 et 21 jours contre 12 jours pour les conidies produites sur le milieu témoin. Tandis
que les taux de mortalité (16 - 67 %) provoqués par les conidies produites après traitement
de fongicide ont été inférieurs à celui (75 %) induit par les conidies produites sur le milieu
témoin. L'observation du pouvoir infectieux des conidies produites après test s'accorderait
avec les travaux d' Aby (2013) qui ont présenté que ce même isolat de Metarhizium sp a
provoqué un taux élevé (93 % ) de mortalité des charançons. Ce champignon pourrait donc
sporuler et infecter le charançon noir du bananier dans une plantation où les fongicides
sont appliqués pour protéger le bananier.
5. Effet des nématicides sur l'isolat de Metarhizium sp
5.1 Effet des nématicides sur la germination
Les deux nématicides testés que sont l'Oxamyl et l'Ethoprophos ont stimulé la
germination des conidies. Nos résultats se rapprochent des travaux d'Aby (2013) qui ont
indiqué que l'Oxamyl a faiblement inhibé la germination des conidies du même
champignon. Ces deux nématicides seraient donc compatibles avec la germination des
conidies de Metharhizium sp. L'application de ces nématicides dans le sol pour protéger le
59

· bananier pourrait donc augmenter le pouvoir germinatif et le pouvoir pathogène de
Metarhizium sp, qui est un champignon tellurique.
5.2 Effet des nématicides sur la croissance du mycélium
Les faibles inhibitions et les stimulations de la croissance mycélienne enregistrées
indiqueraient une compatibilité entre ces nématicides et la croissance du mycélium cet
agent de lutte biologique. Les travaux d' Aby (2013) ont également présenté une absence de
différence significative entre la croissance mycélienne en présence de 5 ppm d'oxamyl et
la croissance mycélienne de Metarhizium sp sur le milieu témoin. Ces nématicides
n'empêcheraient donc pas le développement de ce champignon entomopathogène.
5.3 Effet des nématicides sur la sporulation
Les doses élevées (10 et 15 ppm) d'oxamyl et d'éthoprophos qui ont inhibé la
sporulation pourraient entraver la reproduction de cet agent de lutte biologique. Par contre,
ces nématicides à faibles doses (3 et 5 ppm) stimuleraient la sporulation et la reproduction
du champignon entomopathogène. La stimulation d'un champignon par un nématicide a
été aussi observée par Culbreath et al. (1986). Ceux-ci ont montré que la chitine, un
nématicide, stimule l'activité de Paecilomyces lilacinus, un champignon utilisé comme
agent de lutte biologique contre le nématode Meloidogyne sp. Nos résultats s'accordent
avec ceux d'Aby (2013) qui ont indiqué une faible inhibition de la sporulation induite par
la dose 5 ppm d'oxamyl. Une compatibilité existerait aussi entre certaines concentrations
de ces nématicides et la sporulation de cet agent de lutte biologique.
60

CONCLUSION ET PERSPECTIVES
61

La germination des conidies, la croissance mycélienne et la sporulation de Metarhizium
sp ont été inhibées par le méthyl-thiophante, le trifloxystrobine et le propinaconazole qui
sont des fongicides utilisés pour protéger le bananier contre les mycoses telles que la
cercosporiose noire. L'utilisation de Metarhizium sp comme agent de lutte biologique
contre Comopolites sordidus devrait en tenir compte. Cette précaution est utile même si le
feuillage des bananiers et la systémie de ces fongicides testés constituent de sérieux
obstacles avant que les fongicides atteignent le sol où se trouvent les inocula. Toutefois
l'élément infectieux qui est la conidie a germé, a présenté une croissance mycélienne et a
sporulé en présence ces fongicides. Et, les conidies produites après traitement de fongicide
ont germé et ont causé la mort du charançon noir du bananier, Comopolites sordidus.
De plus, la plupart des doses utilisées des deux nématicides ont été compatibles avec la
germination des conidies, la croissance mycélienne et la sporulation. L'application de ces
nématicides pour protéger le bananier favoriserait l'infection de Cosmopolites sordidus par
Metarhizium :,p. Les doses de 3 et 5 ppm de ces nématicides qui ont stimulé la sporulation
pourraient être utilisées pour la multiplication des inocula.
Ce champignon entomopathogène pourrait donc être utilisé comme agent de lutte
biologique en plantation où sont appliqués ces fongicides et nématicides pour protéger la
culture principale.
Nos travaux ont montré que Metarhizium sp peut être utilisé comme agent de lutte
biologique contre le charançon noir dans un environnement où plusieurs pesticides sont
appliqués pour protéger la culture principale. Cependant, il est indispensable d'effectuer
des travaux complémentaires pour une large exploitation du champignon dans toutes les
plantations de Côte d'Ivoire. Ces travaux pourraient s'articuler autour de:
};:> l'effet des herbicides, des rodonticides, des engrais, des insecticides et d'autres
fongicides et nématicides sur Metarhizium sp ;
};:> la préparation de substrats organiques non alimentaires favorisant la sporulation de
Metarhizium :,p et l'infection du charançon noir du bananier, Cosmopolites
sordidus;
};:> l'étude approfondie des conditions de sporulation de Metarhizium sp ;
};:> la mise en place d'une unité de formulation et de production de l'inoculum;
};:> la lutte génétique à travers ! 'utilisation de bananiers résistants ou tolérants ;
62

RÉFÉRENCES BIBLIOGRAPIDQUES
63

Aby N. (2013). Lutte biologique contre le charançon noir du bananier, Cosmopolites
sordidus Germar (Coleoptere: Curculionidae) en Côte d'Ivoire : Caractéristiques
entomopathologiques d'isolats locaux de Metarhizium sp testés sur les populations au
laboratoire et en bananeraie. Thèse de Doctorat présentée à l 'UFR de Biosciences de
l'Université Félix Houphouët Boigny, Côte d'Ivoire. 181 p.
Aby N., Kéhé M., Kobenan K. & Gnonhouri P. (2008). Recherche de souches
locales de champignons parasites des charançons du bananier dans les principaux
bassins de production de banane dessert en Côte d'Ivoire, Avril-Septembre 2008.
Premier rapport d'étape Projet FIRCA/ CNRA, Décembre 2008. 22p.
Acked J. & Kyamuhangere W. (1996). Postharvest aspects of highland bananas
in Uganda. Tropical Science, 36: 54-64.
Adiko A. (1988). Plant parasitic nematodes associated with plantain, Musa
paradisiaca (AAB), in Ivory Coast. Revue Nematologie, 11 (1): 109-113.
Aka R., Kouassi N., Agneroh T., Amancho N. & Sangare A. (2009).
Distribution et incidence de la mosaïque de concombre (CMV) dans les
bananeraies industrielles au Sud-est de la Côte d'Ivoire. Sciences & Nature, 2:
171-183.
Atrassi K., Benkirane R., Attarassi B., Badoc A. & Douira A. (2007). Efficacité de
deux fongicides benzimidazolés et de l'anilinopyrimidine sur la pourriture des pommes en
conservation. Bull.Soc.Pharm.Bordeaux. 195-21 O. www.socpharmbordeaux.asso.fr/pdf/-
146/J 46-l 95-2 l 0.pdf. Consulté le 12-09-2013.
Bacon R. (2002). Gestion de la résistance aux fongicides. Agriculture et Agroalimentaire
Canada.http://www.agrireseau.qc.ca/petitsfruits/Documents/resistance%20aux%20fongicid
es.PDF. Consulté le 12-09-2013.
Bakry F., Carrreel M., Côte F., Jenny C. & Tézenas Du Montcel H.
(1997). Les bananiers. ln: L'amélioration des plantes tropicales. A
Charrier, M. Jacquot, S. Hamon et D. Nicolas (Eds), CIRAD/ORSTOM,
Montpellier, 109-139.
64

Bischoff J.F., Rehner S.A. & Humber R.A. (2009). A multilocus phylogeny of
the Metarhizium anisopliae lineage". Mycology, 101: 512-530.
Budenberg W.J., Ndiege J.O., Karago F.W. & Hansson B.S. (1993). Behavioral
and electro-physiological responses on the banana weevil Cosmopolites sordidus
to host plant volatiles. Journal of ChemicaJ Ecology, Vol 19: 267-277.
Cluzeau S. (1995). Index phytosanitaire ACTA, 31 e édition, pp 52-244.
Carbonell L. F., Klowden M. J. & Miller K. K. (1985). BacuJovirus-mediated
expression of bacterial genes in dipteran and mammalian cells, J. Virol., 56: 153.
Cerda H., Lopez A., Saooja O., Sancez P. & Jaffe K. (1995). Attraccion olfativa
de Cosmopolites sordidus Germar (Coleoptera: Curculionidae) estimulado por
volatiles originados en Musaceas de distintas edades y variedades genomicas.
Agronomy Tropical, 46 : 413-29.
Chubilleau C., Pubert M., Comte J. & Giraud J. (2011). Pesticides et santé.
Etude écologique du lien entre territoires et mortalité en Poitou-Charentes. Rapport 136.
222 p. http://www.ors-poitou-charentes.org/pdf/k:vUUK4Pesticides 11.pdf. Consulté Je 31-
08-2014.
CIRAD. (2010). (Centre de Coopération Internationale en Recherche Agronomique pour
le Développement). www.cirad.fr/nos-recherches/productions-tropicales/banane-et
plantain/contexte-et-enieux. Consulté le 15 avril 2014
Cloyd R. A. (1999). The Entomopathogenic Fungus Metarhizium anisopliae".
Midwest Biological Control News VI (7). Cosmopolites sordidus (Germar)
damage on highland banana growth, yield and stand duration Cosmopolites
sordidus. Biocontrol Sciences Technology. 3: 177-187.
Culbreath, A.K., R.Rodriguez-Kabao & G.Morgan-Jooes. (1986). Chitin and
Paecilomyces lilacinus for control of Meloidogyne arenaria. Nematropica
16(2):153-166.
Couteux A. & Lejeune V. (2005). Index phytosanitaire ACTA, 41 e edition, 820 p.
Dadzie (1995). Plantain utilization in Ghana. Tropical Science, 35: 405-41 O.
65

Davidse L.C., Flacb W. (1978). Interaction of thiabendazole witb fungal ubulin. -
Biochemistry Biophysique ACTA, 543(1), 82-90.
Debach P. (1964). Biological Control of lnsect Pests and Weeds. London: Chapman and
Hall, 844 p.
Dessaw (1988). Etude des facteurs de la stérilité du bananier (Musa spp.) et des
relations cytotaxonomiqaues entre Musa acumioata COLLA et Musa balbisiana
COLLA. Fruits, 43 (10): 539-558.
Deysson G. (1967). Cours de Botanique Générale. Tome 2. Organisation et
classification des plantes vasculaires. II Systématique. Deysson (Ed.), Paris, 123p.
FAO (2003) (Organisation Mondiale pour l' Alimentation et l' Agriculture). Code
international de conduite pour la distribution et l'utilisation des pesticides, Version révisée
2003. http://www.sustainlabour.org/documentos/fra34 2011.pdf. Consulté le 31-08-2014
Faurie B., Cluzet S., Corio-costct M.F. & Merillon J.M. (2009). Methyl
jasmonate/etephon cotreatment synergically induces stilbène production in vitis vinifera
cell suspension but fails to trigger resistabce to Erysiphe necator. Journal International
Sciences Vigne vin 43 (2): 99-11 O.
Fogain R. & Price N.S. (1994). Varietal screening of some Musa cultivars for
susceptibility to the banana weevil, Cosmopolites sordidus (Coleoptera:
Curculionidae). Fruits, 49: 247-51.
Fouré E. & Mouliom P. A. (1988). Les cercosporioses du bananier et leurs traitements.
Efficacités comparées de différentes molécules Références 211 fongicides sur
Mycosphaerella fijiensis Morelet, agent de la maladie des raies noires des bananiers et
plantains au Cameroun. Fruits 43: 67-72.
Freimoser F. M., Screen S., Bagga S., Hu G. & St. Leger R.J. (2003). EST
analysis of two subspecies of Metarhizium anisopliae reveals a pletbora of
secreted proteins with potential activity in insect hosts". Microbiology, 149: 239-
247.
66

Ganry J., Fouré E., De Lapeyre de Bellaire L. & Lescot T. (2012). An integrated
approach to control the Black leaf streak disease (BLSD) of bananas, while reducing
fungicide use and environmental impact. In, D Dhanasekaran, N Thajuddin, A
Panneerselvam Fungicides for Plant and Animal Diseases, pp 193-226.
Gilbert G. 1999. La résistance des champignons aux fongicides. Semaine Hortic.
http://www.agrireseau.qc.ca/1egumeschamp/documents/R%C3%A9sistance.pdf. Consulté
le 12-09-2013.
Gnonbouri G.P., Adiko A. Kobenan K. & Aké S. (2009). Longévité des
bananeraies industrielles en relation avec le parasitisme des nématodes
Radopholus similis et Pratylenchus coffeae en Côte d'Ivoire. Journal Biosciences:
1100-1 111.
Gnonhouri P., Kobénao K., Kéhé M. & Aby N. (2008). Répartition des
nématodes Radopholus similis et Pratylenchus coffeaea dans les régions de
l' Agnéby, des Lagunes, et du Sud comoé, bassins de production de la banane
dessert en Côte d'Ivoire, Avril-Septembre 2008. Premier rapport d'étape Projet
FIRCA/ CNRA, Décembre 2008. 36p.
Goettel M.S. (1992). Des champignons comme agents de lutte biologique. ln : La
lutte biologique contre les acridiens, sous la direction de C.J. Lomer et C. Prior.
Ibadan, Nigeria: CAB International /UT A 122-131 p.
Gold C.S., Jorge P.E., Karamura E.B. & Rukazambuga N.D. (2003). Biology
and integrated pest management for the banana weevil Cosmopolites sordidus
(Germar). Integrated Pest Management. Revew. 6: 79-155.
Gold C.S., Pena J.E. & Karamura E.B. (2001). Biology and integrated pest
management for the banana weevil, Cosmopolites sordidus (Germar) (Coleoptera:
Curculionidae). Integrated Pest Mangement. Revew. 6: 79-155.
Gold C.S., Bagabe M.1. & Sendege R. (1999). Banana weevil, Cosmopolites
sordidus (Germar) (Coleoptera: Curculionidae): test for suspected resistance to
Carbofuran and Dieldrin in Masaka District, Uganda. African Entomology, 7: 189-
196.
67

Gold C.S., Speijer P.R., Karamura E.B., Tushemereirwe W.K. & Kashaija
I.N. (1994). Survey methodologies for pest and disease assessment in Uganda.
Afr. Crop Sei. J., 2: 309-321.
Grant N.T., Pmsinkiewicz E., Makowski RMD, Holmstrom-Ruddick B. & Mortcnscn
K. (1990) Effect of selected pesticides on survival of Colletotrichum gloeosporioides f. sp.
malvae, a bioherbicide for round-leaved mallow (Malva pusilla). Weed Technol. 4, 701-
715.
Kéhé M. (2006). La méthode de lutte sur avertissement biologique contre le
charançon noir du bananier, Cosmopolites sordidus (Coléoptère, Curculionidae),
document de formation FIRCA/CNRA, Juin 2006. 1 Op.
Kobenan K., Traoré S. & Essi B. (2012). Niveau de sensibilité du champignon
responsable de la cercosporiose noire aux fongicides systémiques appliqués sur les
bananiers en plantations industrielles en Côte d'Ivoire. Rapport CNRA. 31 p.
Kobenan K., Traoré S. & Essi B. (2011). Niveau de sensibilité du champignon
responsable de la cercosporiose noire aux fongicides systémiques appliqués sur les
bananiers en plantations industrielles en Côte d'Ivoire. Rapport CNRA. 27 p.
He Z.L., Yang X.E. & Stofella P.J. (2005). Trace elements in agroecosystems and
impacts on the environment. Journal of Trace Elemenl in Medicine and Biology, 19:125-
140.
Hu G. & St. Leger R.J. (2002). Field studies using a recombinant
mycoinsecticide Metarhizium anisopliae reveal that it is rhizosphere competent.
Appl. Environ. Microbiol., 68: 6383-6387.
Kobenan K., Traoré S. & Gnonhouri Ph. (2006). Situation des populations de
Mycosphaerella spp. Champignons responsables des cercosporioses dans les
bananeraies en Côte d'Ivoire au 30 novembre 2005. Premier rapport d'étape Projet
FIRCA/ CNRA, Février 2006. 35p.
Kobenan K., Ake S. & Koné D. (1997). Etude des réactions des racines de
bananiers Musa (AAA) Cv. "Grande Naine" à des lésions artificielles. Info Musa,
6 (1): 21-23.
68

Koller W. & Scbeinpflug H. (1987). Fungal resistance to sterol biosynthesis
inhibitors: a new challenge. Plant Dis .• 71: 1066-1074.
Koulndob B. (1986). Les circuits de distribution de la banane plantain dans
l'agglomération abidjanaise. Mémoire de maîtrise de Géographie rurale.
Université d'Abidjan, Côte d'Ivoire, 48p.
Kouser S., Qaim M. (2011). Impact of Bt cotton on pesticide poisoning in smallholder
agriculture: A panel data analysis. Ecological Economies, 70: 2105-2113.
Lassoudière A. (2007). Le bananier et sa culture, Edition Quae RD 78026
Versailles Cedex, France, 383 p.
Lassoudière A. (1978). Quelques aspects de la croissance et du développement du
bananier 'Poyo' en Côte d'Ivoire. Le système radical. Fruits 33: 314-338.
Lavigne C. (1987). Contribution à l'étude du système racinaire du bananier. Mise
au point du rhizotron et premiers résultats. Fruits, 42 : 264-271.
Leroux P. (2002). Mode d'action des produits phytosanitaires sur les organismes
pathogènes des plantes. Biologie et pathologie végétale, Académies des
sciences/Editions scientifiques et médicales, 1631-0691 (3) 00005-2.
Lipa J. J. (1985). White muscardine (Beauveria sp.). ln: An Outline of Josect
Pathology. Foreign Sei. Publ. Dept NCSTEI, Warsaw, Poland, 139-142.
Luc M., Jean R. N., Arnaud G., Marc R., Helene D. M., Sylvaine C., Bernard
J., Jean P. T. & Pascal B. (2010). Chlordecone Exposure and Risk of Prostate
Cancer: JCO Jun 21 2010: doi:10.1200/JC0.2009.27.2153], étude française
publiée le 21 juin dans le Journal of Clinicat Oncology dans le cadre d'un
programme de recherche nommé Karuprostate.
Loridat P. (1989). Etude de la mycoflore fongique et des nématodes associés aux
nécroses de l'appareil souterrain du bananier en Martinique. Mise en évidence du
pouvoir pathogène du genre Cylindrocladium. Fruits 44 : 587-598.
69

Meyling N. V. & Einlenberg J. (2006). Occurrence and distribution of soil borne
entomopathogenic fungi within a single organic agroecosystem. Agr. Ecosyst.
Environ. 113: 336-341.
Ministère de l'environnement et du développement durable. (2013). Etude de la
Vulnérabilité du Secteur Agricole face aux Changements Climatiques en Côte d'Ivoire.
PNUD Rapport final. l 05 p.
Mohamed E. H.O.T. & Amadou D. (2001). lmproving pesticide application
techniques for desert locust control, report prepared under GCP/INT/651/NO~
'Improving Pesticide Application Techniques for Desert Locust Control". a
collaborative project between FAO, the Centre National Lutte Antiacridienne
Nouakchott. Mauritania, and otber countries of the EMPRES Western Region.
17p.
Mosbe R. (2005). Inhibition of germination and growth of Alternaria alterna/a
and mouldy-core development in Red Delicious apple fruit by Bromuconazole and
Sygnum. Crop Protection, 25: 253-258.
Mulder JL, Stover RH. (1976). Mycosphaeralla species causing banana leaf spot. Trans.
Brit. Mycol. Soc. 67: 77-82
Nankinga C.M. (1997). Cbaracterisation of Beauveria and Metarhizium, Isolates
and the ilnfluence of delivery systems on their use as biological control agents of
banana weevil, Cosmopolites sordidus, University of Reading upgrading report. Reading, UK.
Nankinga C.M. (1994). Potential of Indigenous FungalPathogens for the
Biological Control of the Banana Weevil, Cosmopolites sordidus (Germar), m
Uganda. Masters thesis, Makerere University, Kampala, Uganda, 95p.
Nazaire K. K., Romain A. & Abdourabamane S. (2008). La chlorose infectieuse
du bananier dans les bananeraies industrielles de Côte d'Ivoire. Points sur les
acquis, CNRA en 2007, Document interne: 14-15.
ODEADOM (2010). Office de développement l'économie agricole <l'outre mer. Recueil
Statistique Banane 2010. 52 p.
70

OCAB. (2012). Organisation Centrale des producteurs-exportateurs del' Ananas et
de la Banane. Stratégie pluriannuelle pour la période 2012-2013. dans le cadre des
mesures d'accompagnement de l'Union européenne pour les pays producteurs de bananes
Côte d'Ivoire. 27 p.
Quesada-moraga E., Navas-cortes J.A., maranhao E.A.A., Ortiz- Urquiza A.
& Santiago-Alvarez C. (2007). Factors affecting the occurrence and distribution
of entomopathogenic fungi in natural and cultivated soils. Mycol. Res. 111: 94 7-
966.
Risède J.M. (2002). Analyse de la diversité des champignons du genre Cylindrocladium
Morgan : Application à la caractérisation phénotypique, moléculaire et du pouvoir
pathogène d'isolats de la rhizosphère du bananier. Thèse de doctorat, ENSAM,
Montpellier, France. 191p.
Roberts D. W. (1981). Toxins of entomopathogenic fungi. ln: Microbial control
of pest and plant diseases 1970-1980. H. D. Burges, Academic Press, NY, 441-
465.
Schiffers B. (2011). Lutte biologique et production intégrée, manuel 10. PIP éditions,
Bruxelles, Belgique, 128p. http://www.pip.coleacp.org./files/documents/COLEACP.
Manuel consulté le 21-02-2014.
Sesbu Reddy K. V., Prasad J.S. & Sikora R.A. (1998). Biointensive management
of crop borers of banana. ln: S.K. Saini, Proceed. Symp. Biol. Control Trop. Crop
Habitats: Third lot. Conf. Trop. Entornol., 30 October-4 November 1994, Nairobi,
Kenya: ICIPE Science Press, 261-287.
Sholberg P.L., Harlton C., Haag P., Levesque C.A., O'Gorman D. & Seifert K. (2005).
Benzimidazole and diphenylamine sensitivity and ideotity of Penicillium spp. that cause
postharvest blue mold of apples using (3-tubuJin gene sequences. Postharv. Biol. Technol.,
36(1), 41-49.
Simon S. (1994). La lutte intégrée contre le charançon noir des bananiers
Cosmopolites sordidus. Fruits 49: 151-162.
71

Smith J. (2006). Banana bacterial wilt - refining the 'road map' for control. New
Agriculturist. Online . http://www.new-ag.info/en/pov/views.php?a=l080. Consulté le 11
avril 2014.
Spooagel K.W., Diaz F.J. & Cribas A. (1995). El picudo negro del platano,
Cosmopolites sordidus Germar, + plates. La lima, Honduras: FHIA, 35p.
Stierl R. A., Hanke W., Lorenz G., Koelhle B. & Ammermaoo E. (2002).
Nicobifen (BAS 510 F) biological cbaracteristics and redistribution.
Phytopathology 92, S79.
Stover R. H. & Simmonds, N. W. (1987). ). Banana, plantain and abaca disease.
Commonw. Mycology. Instit., Kew, Surrey, UK, 316 p.
Sweeneo R. (1990). Limits of morphotaxonomy: names and synonyms of platains
in Africa and elswere. ln: Identification of genetic diversity in the Genus Musa.
Jarret R.L. (Ed). Proceedings of an International Workshop held at Los Banos,
Philippines, 5-10 September 1988. INIBAP. 172-210.
Temple L. & Tentchou J. (2000). Orientation sectorielle de la politique agricole
sur le bananier plantain au Cameroun. Document CRBP. 207/CRBP/2000.
Tezenas D. M. (1985). Le bananier plantain. Ed. Maisonneuve et Larose. Paris.
140 p.
Tinzaara W., Dicke M., Van Huis A. & Gold C.S. (2002). Use of infochemicals in
pest management with special reference to the banana weevil Cosmopolites
sordidus (Germar) (Coleoptera: Curculionidae): a review. Insect Sei. Appli .. 22:
241-261.
Treverrow N., Peasley D. & Ireland G. (1992). Banana Weevil Borer: A Pest
Management Handbook for Banana Growers. Banana Industry Committee,
NewSouthWales Agriculture, 28p.
Treverrow N.L. & Beddiog R.A. (1993). Development of a system for the
control of the banana weevil borer, Cosmopolites sordidus with entompathogenic
nematodes. ln: R. Bedding, R. Akhurst and H. Kaya. (eds) Nematodes and the
72

Biological Control of Insect Pests. 41-47. Melbourne, Australia: CSIRO.
Turrialba 28, 287-93. Doctor in Sciences tbesis. U. do Sao Paulo, Brazil.
Entomology). IOBC/WPRS Bulletin, Insect pathogens and insect parasitic, 102:
213-215
Tusbemereirwe W., Kangire A., Smith J., Ssekiwoko F., Nakyanzi M.,
Kataama D., Musiitwa C. & Karyaija R. (2003). Epidémie de flétrissement
bactérien sur le bananier en Ouganda. InfoMusa 12. 2: 6-8.
Van Driesche, R.G. & Bellows T.S. Jr. (1996). Biological Control, New York:
Chapman and Hall. 539 p.
Vilardebo A. (1984). Problèmes scientifiques posés par Radopholus similis et
Cosmopolites sordidus en cultures bananières des zones francophones de
production. Fruits 39 : 227-33.
Weddell, J.A. (1945). The ban ana weevil borer. Queensl. Agric. J. 51: 85-91.
Wraight, R. J. & Roberts D. W. (1987). Insect control effort with fungi.
Development Industry. Microbiology. 28: 77-87.
Wyss G.S., Cbarudattan R., Rosskopf E.N. & Littell R. (2004). Effect of
selected pesticides and adjuvants on germination and vegetative growth of
Phomopsis amaranthicola, a biocontrol agent for Amaranthus spp. European Weed
Research Society, Weed Research 44: 469-482
73



![Republique Francaise[1]](https://img.pdfslide.fr/doc/110x75/54878984b47959d80c8b548a/republique-francaise1.jpg)