Embed Size (px)
Citation preview

SCIENCE ACTUELLE 1
SCIENCEACTUELLE
ÉPIGÉNÉTIQUEMARS 2013

2 SCIENCE ACTUELLE
CONTENU
ÉPIGÉNÉTIQUE
4 DE LAMARCK À DARWIN ET RETOUR5 UNE DÉCOUVERTE CHANCEUSE: L’ÉPIGÉNÉTIQUE ET L’HÉRÉDITÉ9 UN REGARD VERS LE FUTUR
BASES DE L’ÉPIGÉNÉTIQUE
Jean-BaptisteLamarck décritla théorie del’évolution
Andrew Fire etCraig C. Mellodémontrentque les ARNsdouble brinsont capablesd’inactiverdes gènes etnomment ceux-ci ARNs inter-férents (ARNi)
Robin Hollidayet Arthur Riggsdécouvrent queles groupesméthyls peu-vent contrôlerl’activité desgènes
Les résultatsdu projetENCODE mont-rent que desportions d’ADNconsidéréesjusqu’à présentsans fonction,sont respon-sables de larégulation desgènes
James D.Watson etFrancis Crickrévèlent lastructure del’ADN
Une équipe derecherche ang-laise prouveque la formedes fleurs de laplante Peloria,découvertepar von Linné,est soumiseà des modifi-cations épigé-nétiques
Richard Jorgen-sen et AntoineStuitje décri-vent le phéno-mène de laco-suppression
1999
2012
1809
19531975
19901998

SCIENCE ACTUELLE 3
Qui connaît les gènes, connaît les humains. C’est du moins ce que croyai-ent les scientifiques et le reste du monde quand, le 26 juin 2000, le géno-
me humain a été déchiffré. Un certain désenchantement s’est cependant fait rapi-dement sentir: tout ce qui avait été découvert, c’est un texte composé d’environ troismilliards de paires de lettres A, C, G et T. Le secret du plan de construction del’être humain n’avait donc pas vraiment été décodé. Depuis le séquençage dugénome humain jusqu’à nos jours, il est alors devenu clair que les gènes, non seu-lement contrôlent, mais sont également contrôlés. Ce n’est que quand nos celluleschoisissent, de manière ciblée, quels gènes lire et quels gènes laisser inactifs que leurdestin prend forme. Le nouveau domaine de recherche appelé épigénétique étudieles raisons pour lesquelles le génome d’un individu ne fixe pas son destin de ma-nière invariable. Ce destin est, au contraire, fortement influençable et désormaisexplorable. L’épigénétique étudie donc comment les facteurs environnementaux etle style de vie peuvent influer l’activité des gènes et comment ces changements peu-vent être transmis d’une génération à l’autre.
D’un point de vue historique, il existe deux définitions différentes del’épigénétique. En 1942, Conrad Waddington décrivit l’épigénétique comme lesinteractions entre les gènes et leurs produits menant au phénotype d’un organisme.De nos jours, c’est cependant la définition d’Arthur Riggs qui est habituellementprise en compte. Elle date de 1996 et décrit l’épigénétique comme les changementstransmissibles de l’activité des gènes, sans que ceux-ci impliquent un changementau niveau de l’ADN. Un grand nombre de caractéristiques biologiques peuventêtre influencées par des mécanismes épigénétiques, comme par exemple, la formedes fleurs chez les plantes et la couleur des yeux chez les drosophiles.
Les processus épigénétiques sont essentiels aussi bien au développement d’unorganisme qu’à la spécialisation des divers types de cellules, comme par exemple,les cellules nerveuses ou les cellules hépatiques. L’état épigénétique d’une celluleou d’un organisme n’est cependant pas figé, il est soumis à des changements occa-sionnés, par exemple, par des influences environnementales ou des processus liésau vieillissement. De récentes découvertes scientifiques démontrent que l’épigénétiquejoue également un rôle dans le développement de maladies.
Durant ces dernières années, notre connaissance des mécanismes moléculairesrégissant les processus épigénétiques n’a cessé de croître. Même si nous sommes loinde connaître tous les aspects de leur fonction et de leur transmission, nous pouvonsdiviser l’épigénétique en trois domaines: les modifications chimiques au niveau del’ADN, les modifications chimiques au niveau des protéines liées à l’ADN, etl’influence exercée par des molécules d’ARN.
Cette édition de « Science Actuelle » met en avant le domaine complexe etpassionnant de l’épigénétique. Elle vous entraîne des débuts de l’épigénétique auxconnaissances actuelles de la recherche, en finissant par lancer un regard vers lefutur. Nous vous souhaitons une très bonne lecture.
Directrice de la Fondation Gen Suisse
PRÉFACE
Dr. Daniela Suter
Directrice de la Fondation Gen Suisse

4 SCIENCE ACTUELLE
Qui ne connaît pas l’histoire de la girafe au coucourt qui, à force d’étirer constamment ce dernier,obtint un long cou lui permettant d’atteindre lesfeuilles des grands arbres? Les girafes au long couont un meilleur accès à la nourriture et peuvent,par le biais de la reproduction, transmettre leuravantage sélectif. Cet exemple illustre la théorie del’évolution de Jean-Baptiste Lamarck datant de1809.
Selon lui, des conditions environnementaleschangeantes peuvent avoir une telle influence surles propriétés d’un organisme que les nouvellescaractéristiques acquises, se transmettant de géné-ration en génération, mènent finalement au chan-gement de l’espèce elle-même. Selon Lamarck,chaque organisme porte en lui une pulsion le pous-sant à se développer.
Durant de nombreuses années, cette théorie n’apas été prise au sérieux dans le monde moderne dela biologie de l’évolution. C’est la théorie pluspopulaire et mieux acceptée scientifiquement deCharles Darwin qui a été suivie. La théorie de lasélection naturelle, élaborée par Charles Darwin en1859, présente une autre vision de la genèse desespèces. Selon Darwin, de nouvelles caractéris-tiques apparaissent au niveau d’un organisme parhasard et l’environnement ne joue alors qu’un rôlesélectif. Si on applique cette théorie à l’exemple citéci-dessus, cela signifie qu’au milieu d’une popula-tion de girafes au cou court se trouvait par hasardune girafe avec un cou plus long. Cette girafe était
alors porteuse d’un avantage sélectif qu’elle trans-mit aux générations suivantes, prenant ainsi le des-sus sur ses congénères au cou court.
La découverte de l’apparition de mutationsspontanées dans le génome est la preuve attestantde la justesse de la thèse de Darwin. Il semble in-vraisemblable que des caractéristiques acquises,telles l’obtention d’un long cou par le biaisd’étirements successifs, aient pu s’introduire dansle génome par des mutations ciblées. Cependant, lathéorie de Darwin n’est pas suffisante à expliquerla vitesse à laquelle l’évolution prend place.
De nos jours, la recherche a réuni d’importantesinformations et connaissances montrant que l’in-formation génétique est modifiable. L’activité desgènes peut être influencée par les conditions envi-ronnementales, sans modification au niveau de laséquence d’ADN. Ce changement de l’activité desgènes peut être transmis aux cellules filles ou à laprochaine génération. On pourrait ainsi dire que« les gènes ont une mémoire ».
Les récentes connaissances acquises dans ledomaine de l’épigénétique font donc revivre l’idée,édifiée par Lamarck, d’une influence de l’en-vironnement transmissible. Cependant, se basantsur l’état actuel de nos connaissances scientifiques,les changements épigénétiques seraient réversibleset il reste encore à découvrir avec quelle stabilitéceux-ci sont hérités.
ÉPIGÉNÉTIQUE
DE LAMARCK À DARWINET RETOUR

SCIENCE ACTUELLE 5
1
2
Au commencement d’une nouvellethéorie se trouve souvent une observa-tion mystérieuse. Le botaniste suédoisCarl von Linné a fait une telle observa-tion au 18e siècle. Il a été le premier àcatégoriser de manière systématique lesplantes et à leur donner leurs nomsactuels. Comme, entre autres, pour lalinaire commune. Dans ce cas-ci, il esttombé sur une variante présentant uneforme de fleur modifiée. Cette forme defleur a été transmise aux générations fu-tures, le reste de la plante restant indif-férenciable de l’espèce d’origine. Linnéa appelé cette variante Peloria, qui sig-nifie monstre en grec.
Afin de résoudre l’énigme, des sci-entifiques ont analysé, dans les années90, le patrimoine génétique des deux va-riétés de linaire commune. Ils se sontalors rendu compte qu’il n’y avait pas dedifférence entre la séquence d’ADN dePeloria et celle de ladite linaire commu-ne. L’énigme n’a tout d’abord pas pu êtrerésolue. C’est en 1999, presque 250 ansaprès la découverte de Carl von Linné,que la raison d’une telle différence phé-notypique a été élucidée par un groupede chercheurs anglais. Ces derniers ontdécouvert que, par le biais de mécanis-mes épigénétiques, un gène nomméLcyc et régulant la forme naturelle de lafleur, avait été inactivé. La raison d’unetelle inactivation est une modificationchimique, transmissible, au niveau del’ADN. L’élément déclencheur de cettemodification, lui, reste une énigme.
De nos jours, trois mécanismes épi-génétiques sont connus: les modifica-tions chimiques au niveau de l’ADN, lesmodifications chimiques au niveau desprotéines liées à l’ADN, et l’influenceexercée par des molécules d’ARN.
Modifications au niveau del’ADN: la première dimensionépigénétique
En 1975, Robin Holliday et ArthurRiggs ont découvert, indépendammentl’un de l’autre, que l’activité des gènespouvait être contrôlée par des modifica-tions chimiques de l’ADN dues àl’introduction de petites molécules,appelées groupes méthyles, au niveau del’ADN. Ces groupes méthyles peuvent selier à la base d’ADN nommée cytosine etainsi mener à l’inactivation de la porti-on d’ADN touchée.
ADNméthylé
Dans notre génome se trouvent desportions d’ADN, principalement situéesà proximité des gènes, présentant unnombre de cytosines plus élevé que lamoyenne. Ces portions d’ADN particu-lières sont appelées îlots-CpG car unebase cytosine y est suivie d’une base gu-anine. Les îlots-CpG jouent un rôle im-portant dans la régulation de la fonctiondes gènes. Plus une certaine séquenced’ADN est porteuse de groupes méthy-les, moins le gène correspondant sera luet donc traduit en protéine.
La méthylation de l’ADN est essenti-
elle au développement normal d’un
organisme. Elle joue un rôle détermi-nant au niveau de l’hérédité 1 et de laconservation d’un génome intact 2 . Si
UNE DÉCOUVERTE CHANCEUSE:L’ÉPIGÉNÉTIQUE ET L’HÉRÉDITÉ
MARQUE GÉNÉTIQUE
La marque génétique (en ang-lais: genomic imprinting)décrit le phénomène par lequell’activité d’un gène dépend dela provenance de l’allèlecorrespondant, c’est-à-dire dé-pend de si ce dernier a étéhérité du père ou de la mère.Pour les gènes étant soumisà une telle régulation épigéné-tique, soit l’allèle maternel,soit l’allèle paternel est inac-tivé. Aujourd’hui on sait quece sont des méthylationsau niveau de l’ADN qui sonten grande partie responsablesde ce processus et qui mènentà l’inactivation du gène corres-pondant.
PROTECTION CONTRE LES ÉLÉ -
MENTS D’ADN MOBILES
Le séquençage du génome amontré que des segmentsd’ADN mobiles, comme parexemple les transposons, cons-tituent une grande partiede notre patrimoine génétique.Ces éléments mobiles sontcapables, à l’aide de diversprocessus, de « sauter » àun autre endroit dans le géno-me. Etant donné que cela peutaffecter plus ou moins forte-ment l’activité des gèneset ainsi constituer un dangerpour la cellule, les cellules onttrouvé le moyen de rendre ceséléments silencieux. Ceci,par le biais de modificationsau niveau de la chromatine,de méthylation au niveaude l’ADN et de l’utilisation dela machinerie ARN (processusqui conduit chez les eucaryo-tes* à l'inactivation de gènesaprès la transcription**).
* Organismes avec un noyau** Passage de l'ADN en ARN
messager
Pelo r i a
lina ireco
mmune
imag
epe
lori
a:E
mil
V.N
ilss
on

6 SCIENCE ACTUELLE
ACTIFLa phosphorylation et l’acéty-lation des histones favorisent,en règle générale, l’accès àl’ADN: activation del’expression des gènes
les enzymes responsables de la méthyla-tion font défaut à un organisme, de gra-ves maladies se développent. Ces derni-ères années, des modifications de laméthylation de l’ADN ont été déceléesen relation avec divers cancers.
Quels sont donc les éléments déclen-cheurs de modifications épigénétiques ?En plus des signaux délivrés par le corpslui-même durant le développement d’unorganisme, de récentes études ont ap-porté la preuve que des influences exté-rieures peuvent aussi mener à des modi-fications épigénétiques. L’exemplesuivant montre comment la façon de senourrir peut avoir un effet sur l’activitédes gènes et comment cet effet peut êtretransmis sur plusieurs générations.
Une équipe de chercheurs a soumispendant plusieurs semaines des rats mâ-les à une diète riche en calories et engraisses. Les rats sont devenus obèses etles premiers signes d’un diabète sontapparus. Leur progéniture femelle a en-suite été étudiée. L’analyse révéla quecelle-ci, bien que nourrie normalement,présentait des symptômes propres audiabète. Une étude plus poussée a alorsmontré que l’activité de plus de 642
gènes différait de celle des animaux con-trôlés dont les pères avaient reçu une di-ète normale. La différence au niveau del’activité de certains gènes provenait demodifications de la méthylation del’ADN.
Protéines se liant à l’ADN: ladeuxième dimension épigénétique
Une autre manière de régulerl’activité des gènes est d’agir au niveaude l’empaquetage de l’ADN. L’ADN setrouve dans le noyau, enroulé autour deprotéines appelées histones et organiséen une structure complexe appelée chro-matine.
D’un point de vue historique, la dé-couverte de modifications au niveau dela chromatine date déjà d’un demi-sièc-le. Cependant, c’est seulement au 21esiècle que les chercheurs ont découvertque des modifications chimiques, iso-lées ou multiples, au niveau des sous-unités de la chromatine, les histones,peuvent réguler l’accès à l’ADN. Cesmodifications (méthylations, acétyla-tions, phosphorylations) mènent auchangement de la structure de la chro-matine et déterminent ainsi si un gèneest actif ou non. La chromatine présen-te dans un état fortement condensé,alors appelée hétérochromatine, est as-sociée à une faible activité génique. Aucontraire, l’euchromatine, terme désig-nant une chromatine moins dense, est,elle, associée à une forte activité gé-nique.
En tenant compte de la diversité desmodifications chimiques possibles ainsique du fait que de courts segmentsd’ADN s’embobinent autour d’octamèresd’histones, eux-mêmes composés decinq différentes histones, la combinai-son des modifications au niveau de la
chromatine est énorme. Cette complexi-té a poussé les chercheurs à soupçonnerl’existence d’un code des histones, pré-servé durant les processus de division etspécialisation cellulaires.
Le concept du code des histonesressemble à une serrure à combinaison:seule la combinaison juste ouvre lecoffre – et mène à l’activité génique. Unedifférence au niveau d’un seul chiffre dela combinaison est assez pour que lecoffre reste fermé – pour que le gène nesoit pas activé.
Les modifications au niveau des his-tones régulent un grand nombred’aspects propres au développementd’un organisme, de la régulation d’unseul gène à celle de processus complexestels la détermination du destin d’unecellule. Des modifications anormales auniveau des histones, dues par exemple àdes facteurs environnementaux, peu-vent conduire au développement de ma-ladies. De telles corrélations ont déjà puêtre démontrées pour la maladie deParkinson, la sclérose en plaques etl’épilepsie.
hist one2A
INACTIFLa tri-méthylation de l’histoneH3 mène à la condensation dela structure de la chromatine:inactivation de l’expressiondes gènes
SCIENCE ACTUELLE 7
3Une récente étude réalisée par deschercheurs américains a mené à l’ob-tention d’importantes connaissances surl’ampleur des influences environnemen-tales sur les modifications au niveau deshistones. Les chercheurs ont exposé desrats en gestation à la neurotoxine conte-nue dans les cigarettes, la nicotine. Lesjeunes rats provenant de ces portées ontultérieurement présenté des modifica-tions au niveau des poumons, sympto-matiques de l’asthme. Encore plussurprenant a été de constater que la pro-géniture de ces jeunes rats, qui n’avait,elle, jamais été exposée à la nicotine,présentait également des dommages auniveau des organes respiratoires. Mêmesi le mécanisme épigénétique derrièretout cela n’a pas encore été complète-ment clarifié, de premières études indi-quent qu’un changement au niveau dela modification des histones dans lescellules sexuelles en serait la cause.
Molécules d’ARN: la troisièmedimension épigénétique
D’un point de vue historique, cemécanisme de régulation épigénétiquese situe au deuxième rang après les mo-difications au niveau de l’ADN et deshistones. Ces dernières années cepen-dant, de nouvelles découvertes scienti-fiques ont changé la donne en révélantune fonction régulatrice, jusqu’ici in-connue, des molécules d’ARN. 3
Cette découverte est en fait due à unhasard. En 1990, les laboratoires deRichard Jorgensen et d’Antoine Stuitjeont fait une découverte d’importance.Ils avaient lancé une expérience consis-tant à introduire dans des pétunias lesgènes responsables de la couleur desfleurs. L’expérience se fit mais les résul-tats obtenus différèrent complètementde ceux attendus. Les pétunias traitésprésentaient, à la place d’une intensecouleur violette, de grands domainessans couleur. Les chercheurs en conclu-rent que le gène déterminant la couleurdes fleurs propre à la plante, ainsi quecelui introduit, avaient été tous deuxinactivés. Ils décrivirent ce phénomènecomme la co-suppression. Le mécanis-me responsable de ce phénomène n’acependant pas pu être expliqué à cemoment-là.
pétun ia
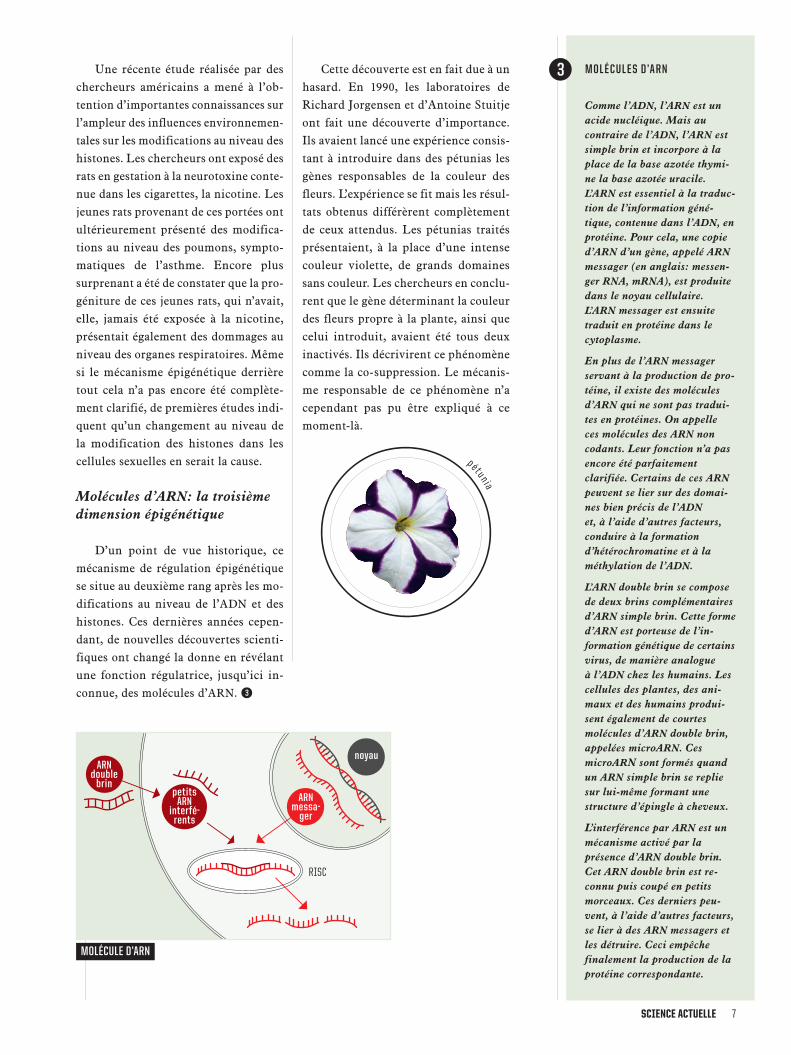
MOLÉCULE D’ARN
ARNdoublebrin
noyau
RISC
MOLÉCULES D’ARN
Comme l’ADN, l’ARN est unacide nucléique. Mais aucontraire de l’ADN, l’ARN estsimple brin et incorpore à laplace de la base azotée thymi-ne la base azotée uracile.L’ARN est essentiel à la traduc-tion de l’information géné-tique, contenue dans l’ADN, enprotéine. Pour cela, une copied’ARN d’un gène, appelé ARNmessager (en anglais: messen-ger RNA, mRNA), est produitedans le noyau cellulaire.L’ARN messager est ensuitetraduit en protéine dans lecytoplasme.
En plus de l’ARN messagerservant à la production de pro-téine, il existe des moléculesd’ARN qui ne sont pas tradui-tes en protéines. On appelleces molécules des ARN noncodants. Leur fonction n’a pasencore été parfaitementclarifiée. Certains de ces ARNpeuvent se lier sur des domai-nes bien précis de l’ADNet, à l’aide d’autres facteurs,conduire à la formationd’hétérochromatine et à laméthylation de l’ADN.
L’ARN double brin se composede deux brins complémentairesd’ARN simple brin. Cette formed’ARN est porteuse de l’in-formation génétique de certainsvirus, de manière analogueà l’ADN chez les humains. Lescellules des plantes, des ani-maux et des humains produi-sent également de courtesmolécules d’ARN double brin,appelées microARN. CesmicroARN sont formés quandun ARN simple brin se repliesur lui-même formant unestructure d’épingle à cheveux.
L’interférence par ARN est unmécanisme activé par laprésence d’ARN double brin.Cet ARN double brin est re-connu puis coupé en petitsmorceaux. Ces derniers peu-vent, à l’aide d’autres facteurs,se lier à des ARN messagers etles détruire. Ceci empêchefinalement la production de laprotéine correspondante.
petitsARN
interfé-rents
ARNmessa-ger

8 SCIENCE ACTUELLE
L’explication relative à ce phé-nomène fut obtenue grâce au travail surles nématodes d’Andrew Fire et Craig C.Mello. Ils injectèrent des moléculesd’ARN synthétiques d’un gène muscu-laire dans les nématodes. Ils utilisèrent,au contraire de l’ARN cellulaire, del’ARN double brin (dsRNA). Ce traite-ment eut pour conséquence le tremble-ment incontrôlé des vers, un comporte-ment semblable à celui de nématodeschez qui ce gène est inactivé. Mais ques’était-il passé?
En réaction à l’injection des molécu-les d’ARN, les cellules des nématodesavaient détruit leur propre ARN (ARN
messager) ainsi que l’ARN synthétiquereçu correspondant au gène musculaire.La protéine codée par cet ARN n’avaitainsi pas pu être produite.
nématode
Après une importante série d’ex-périences, Fire et Mello parvinrent à laconclusion suivante: l’ARN double brinest capable d’inactiver des gènes, cesARN interférents (ARNi) sont spéci-fiques aux gènes présentant la mêmeséquence que l’ARN injecté, et ces ARNipeuvent se propager aux cellules filles etsont même héritables.
En 2006, Andrew Fire and Craig C.Mello reçurent le prix Nobel de méde-
cine pour leur découverte. Leur recher-che permet aujourd’hui aux chercheursd’utiliser un mécanisme naturel pourinactiver des gènes de manière spéci-fique. Si on connaît la séquence del’ARN messager d’origine, on peut pro-duire l’ARN double brin correspondant,l’introduire dans les cellules et ainsiinactiver le gène d’intérêt. 4
Ces dernières années, notre con-naissance des mécanismes de la régula-tion épigénétique par le biais de molé-cules d’ARN a beaucoup augmenté. Lesplantes, comme les animaux, utilisent ceprocessus comme mécanisme de défen-se contre le matériel génétique intrusif,comme par exemple celui des virus.
De récentes études montrent égale-ment que les cellules peuvent générer el-les-mêmes de l’ARN double brin et ainsiréguler l’activité de gènes en fonction designaux cellulaires ou d’influencesexternes. Ce type d’ARN double brin estappelé microARN (miARN). Les mic-roARN sont présents chez les plantes,les animaux et les hommes et jouent unrôle déterminant dans le développementd’un organisme. Ils régulent l’activitéd’un grand nombre de gènes et ainsi,presque tous les processus cellulaires,allant de la division cellulaire à la mortcellulaire contrôlée. Tout comme pourla méthylation de l’ADN et la modifica-tion des histones, un déséquilibre auniveau de ces molécules d’ARN peutconduire au développement de maladies. ADN
(ACIDE DÉSOXYRIBONUC-LÉIQUE)Description de la substance chi-mique constituant les gènes. L’ADNse compose de deux brins consti-tués par la combinaison des basesadénine (A), thymine (T), guanine(G) et cytosine (C) et enrouléssur leur propre axe.
ARN(ACIDE RIBONUCLÉIQUE)Description de la substance chi-mique constituant les copiesdes gènes. Les copies des gènes sontcomposées à partir des quatrebases A, U, G et C. Ce qui signifieque la base thymine (T), présenteau niveau de l’ADN, est dansce cas-ci remplacée par une baseuracile (U).
250Il a fallu presque 250 ans jusqu’à ce que le secret de la plante Peloria de von Linné soitpercé. La cause de la modification de cette plante est une modification chimiquehéréditaire de l’ADN
4 PRIX NOBEL DE MÉDECINE 2006ANDREW FIRE & CRAIG C. MELLO
L’interférence par ARN commemoyen d’inactiver de manièreciblée des gènes et des pro-téines spécifiques
La façon usuelle de produiredes protéines dans une cellule
Interruption ciblée de laproduction de protéines dansune cellule
Patrimoinehéréditaire: l’ADN
noyau cellulaire
ARN messager
destruction del’ARN messager
molécules d’ARNsynthétiques
PAS DE protéine
transport endehors du noyau
production deprotéine
protéine

SCIENCE ACTUELLE 9
Le domaine de l’épigénétique a tenu, ces der-nières années, les chercheurs en haleine. De nou-velles découvertes sont régulièrement faites, nousaidant à mieux comprendre la régulation de notregénome, un phénomène bien plus complexequ’imaginé seulement quelques années auparavant.
En 2001, le Projet Génome Humain nous aapporté une multitude de nouvelles connaissanceset a ainsi ouvert une nouvelle dimension dans lesdomaines de la recherche fondamentale et de la re-cherche appliquée à la médecine. Cependant aucu-ne fonction n’avait pu être attribuée à plus de 90pourcent de notre séquence d’ADN.
Afin de combler cette lacune, un consortiumscientifique international, composé de plus de 440
chercheurs provenant de 32 laboratoires à traversle monde, a été créé. Ce projet a reçu le nom deENCODE (the ENCyclopedia Of Dna Elements).Les résultats obtenus jusqu’à présent montrentqu’une bonne partie de d’ADN considéré alors sansfonction, est responsable de réguler l’activité desgènes. Ceci explique, entre autres, comment il estpossible qu’une cellule hépatique ou une cellulenerveuse se différencie à partir d’une même celluleprécurseur. La différence n’est pas contenue dansl’information génétique en soi, mais dans la régu-lation de celle-ci.
«Grâce à ENCODE, nous avons obtenu un atlas
des séquences régulatrices présentes dans notre
génome. Nous, chercheurs, avons maintenant à dis-
position un ouvrage de référence sur la base duquel
nous pouvons chercher des réponses à nos questi-
ons spécifiques», explique Susanne Gasser, direc-
trice de Friedrich Miescher Institute for Biomedi-
cal Research à Bâle.
La régulation de l’activité des gènes est deve-nue plus complexe. Une analyse de la séquenced’ADN ne suffit plus. Les modifications épigéné-tiques, causées par des molécules d’ARN régula-trices, la méthylation de l’ADN ou encore desmodifications au niveau des histones, doivent êtreintégrées dans le tableau actuel. Les chercheurs sesont donc fixé comme but de décoder ce «deuxièmegénome». Ils espèrent ainsi, par l’analyse des mo-difications épigénétiques présentes dans des orga-nismes modèles comme la souris ou la mouche,mieux comprendre l’évolution et le développementde maladies.
UN REGARD VERSLE FUTUR
ENCODE«est la plusgrande collectionde données épigé-nétiques obtenues àpartir de lignéescellulaires»
Enc
ode
artw
ork
avec
lape
rmis
sion
deC
arld
eTo
rres
Gra
phic
Des
ign

10 SCIENCE ACTUELLE
Des modifications épigénétiques particulières peuventmener au développement de maladies telles que le cancer. Aucontraire des modifications génétiques, les processus épigé-nétiques sont réversibles. Pour cette raison, ces processus con-stituent d’intéressantes cibles thérapeutiques. Dans le futur,la médecine personnalisée ne prendra donc pas seulement encompte les mutations génétiques mais également les différen-ces au niveau du profil épigénétique de chaque patient, ceciafin de choisir une thérapie appropriée. Susanne Gasser note:
«Dans le cas de maladies où les cellules se différencient de
manière inappropriée, telles les leucémies ou d’autres types
de cancer, une telle approche est envisageable. Par contre, dans
le cas de maladies telles Alzheimer, notre connaissance des
modifications épigénétiques est, pour le moment, trop limitée
pour influencer le traitement à suivre».
groupeméthy l é
Les premiers médicaments agissant sur des processus épi-génétiques, comme par exemple ceux modifiant la méthylati-on de l’ADN, ont déjà été approuvés pour le traitement decertains cancers. Le nouvel espoir des scientifiques reposemaintenant sur des molécules modifiant les histones. Lespremiers médicaments candidats de ce genre sont déjà en dé-veloppement clinique. «Les thérapies utilisant des molécules
modifiant les histones sont beaucoup plus prometteuses car le
nombre de protéines modifiant les histones est plus important.
Il existe donc plus de points de départ ainsi qu’une multitude
d’approches différentes», souligne Susanne Gasser.
De nos jours, trois mécanismes épigénétiques sont connus:les modifications chimiques au niveau de l’ADN ou au niveau desprotéines liées à l’ADN ainsi que l’influence exercée par desmolécules d’ARN
3
i INACTIVATION
DU CHROMOSOME X
L’inactivation du chromosomeX décrit un phénomène bienconnu chez les mammifères.Les femelles sont porteuses dedeux copies du chromosome X,alors que les mâles sontporteurs d’une copie du chro-mosome X et d’une copiedu chromosome Y. Afin que ladose génique soit la même chezles deux sexes, une des deuxcopies du chromosome X estinactivée chez la femelle. Lechoix de quelle copie serainactivée est aléatoire. Cetteinactivation implique touteune série de processus épigéné-tiques. Ces processus sontdéclenchés par une longuemolécule d’ARN, Xist, dontl’activité est elle-même réguléede manière épigénétique. Laliaison de cet ARN et d’autresfacteurs déclenche une cascademenant à l’inactivation d’unegrande partie des gènes pré-sents sur la deuxième copie duchromosome X. Des modi-fications au niveau de la chro-matine, la formation d’hétéro-chromatine ainsi que desméthylations au niveau del’ADN font partie de cettecascade. L’emballage compactde la chromatine et les change-ments structurels en résultantont donné au chromosome Xinactif le nom de corpuscule deBarr.
Un exemple illustrant lamarque génétique est la pré-sence de patchs de fourrurede couleurs différentes chez leschats femelles.

SCIENCE ACTUELLE 11
Beisel Ch. & Paro R., 2011Silencing chromatin: compa-ring modes and mechanisms;Nature Reviews Genetics;12: 123-135.
Bird A., 2007Perceptions of epigenetics;Nature, 447, 396-398.
Chahwan R. et al., 2011The multidimensional natureof epigenetic information andits role in disease; Discov.Med.; 11(58):233-43.
Choudhuri S., 2011From Waddington`s epigene-tic landscape to small non-coding RNA: some importantmilestones in the historyof epigenetics research,Toxicology; Mechanisms andMethods, 21, 252-274.
Crews D., et al., 2012Epigenetic transgenerationalinheritance of altered stressresponses; PNAS, 109(23),9143-9148.
Cubas P. et al., 1999An epigenetic mutation res-ponsible for natural variationin floral asymmetry; Nature,401, 157-161.
Dawson M. A. & Kouzarides T.,2012Cancer epigenetics: Frommechanism to therapy;Cell; 150: 12-27.
Eccleston et al., 2007Epigenetics; Nature; 447, 395.
Feinberg A.P., 2007Phenotypic plasticity and theepigenetics of humandisease; Nature; 24(447),433-440.
Fraga M.F. et al., 2005Epigenetic differences ariseduring the lifetime of mono-zygotic twins; PNAS;102(30):10604-10609.
Holliday R. et al., 1975DNA modification mechanismsand gene activity duringdevelopment; Science,187, 226-232.
Jenuwein T. & Allis D., 2001Translating the histone code;Science; 293:1074-1080.
Jones P. A., 2012Functions of DNA methylation:islands, start sites, genebodies and beyond; Nature;13: 484-492.
Kouzarides T., 2007Chromatin modifications andtheir function; Cell; 128:693-705.
Mack G.S., 2006Epigenetic cancer therapymakes headway; J. NatlCancer Inst, 98, 1443-1444.
Marx V., 2012Reading the second genomiccode; Nature, 491, 143-147.
Ng S.-F. et al., 2010Chronic high-fat diet infathers programs beta-celldysfunction in female ratoffspring; Nature, 467,963-967.
Payer B. und Lee J.T., 2008X chromosome dosagecompensation: how mammalskeep the balance; NatureReviews Genetics; 42:733-772.
Rehan V.K., 2012Perinatal nicotine exposureinduces asthma in secondgeneration offspring, BioMedCentral; Medicine, 10, 129.
Riggs A, 1975X-Inactivation, differentiati-on, and DNA methylation;Cell Genet., 14, 9-25.
Russo V. et al., 1996Epigenetic Mechanisms ofGene Regulation; Cold SpringHarbor Laboratory Press,Woodbury.
Strahl B.D. et al., 2000The language of covalenthistone modifications;Nature, 403, 41-45.
Waddington C.H., 1957The strategy of the genes;Allen & Unwin.
L I T TÉRATURE

«Le dialogue est notre objectif»Fondation Gen SuisseAarbergergasse 29CH-3011 BerneT +41 (0)31 356 73 84F +41 (0)31 356 73 [email protected]