Embed Size (px)
Citation preview

VETAGRO SUP CAMPUS VETERINAIRE DE LYON
Année 2013 - Thèse n°
PATHOLOGIE CARDIOVASCULAIRE
DES OISEAUX
THESE
Présentée à l’UNIVERSITE CLAUDE-BERNARD - LYON I
(Médecine - Pharmacie)
et soutenue publiquement le 12 Juin 2013
pour obtenir le grade de Docteur Vétérinaire
par
Graham Zoller
Né le 26 Mai 1988
à Annecy (74)

Page | 2
Page | 3
Liste des enseignants du Campus Vétérinaire de Vetagro Sup
Civilité Nom Prénom Unités pédagogiques Grade
M. ALOGNINOUWA Théodore Unité pédagogique Pathologie du bétail Professeur
M. ALVESDEOLIVEIRA Laurent Unité pédagogique Gestion des élevages Maître de conférences
Mme ARCANGIOLI Marie-Anne Unité pédagogique Pathologie du bétail Maître de conférences
M. ARTOIS Marc Unité pédagogique Santé Publique et Vétérinaire Professeur
M. BARTHELEMY Anthony Unité pédagogique Anatomie Chirurgie (ACSAI) Maître de conférences Contractuel
Mme BECKER Claire Unité pédagogique Pathologie du bétail Maître de conférences
M. BELLI Patrick Unité pédagogique Pathologie morphologique et clinique des animaux de compagnie
Maître de conférences Contractuel
Mme BELLUCO Sara Unité pédagogique Pathologie morphologique et clinique des animaux de compagnie
Maître de conférences
Mme BENAMOUSMITH Agnès Unité pédagogique Equine Maître de conférences
M. BENOIT Etienne Unité pédagogique Biologie fonctionnelle Professeur
M. BERNY Philippe Unité pédagogique Biologie fonctionnelle Professeur
Mme BONNETGARIN Jeanne-Marie Unité pédagogique Biologie fonctionnelle Professeur
Mme BOULOCHER Caroline Unité pédagogique Anatomie Chirurgie (ACSAI) Maître de conférences
M. BOURDOISEAU Gilles Unité pédagogique Santé Publique et Vétérinaire Professeur
M. BOURGOIN Gilles Unité pédagogique Santé Publique et Vétérinaire Maître de conférences
M. BRUYERE Pierre Unité pédagogique Biotechnologies et pathologie de la reproduction
Maître de conférences Contractuel
M. BUFF Samuel Unité pédagogique Biotechnologies et pathologie de la reproduction
Maître de conférences
M. BURONFOSSE Thierry Unité pédagogique Biologie fonctionnelle Maître de conférences
M. CACHON Thibaut Unité pédagogique Anatomie Chirurgie (ACSAI) Maître de conférences Contractuel
M. CADORE Jean-Luc Unité pédagogique Pathologie médicale des animaux de compagnie
Professeur
Mme CALLAITCARDINAL Marie-Pierre Unité pédagogique Santé Publique et Vétérinaire Maître de conférences
M. CAROZZO Claude Unité pédagogique Anatomie Chirurgie (ACSAI) Maître de conférences
M. CHABANNE Luc Unité pédagogique Pathologie médicale des animaux de compagnie
Professeur
Mme CHALVETMONFRAY Karine Unité pédagogique Biologie fonctionnelle Maître de conférences
M. COMMUN Loïc Unité pédagogique Gestion des élevages Maître de conférences
Mme DE BOYER DES ROCHES Alice Unité pédagogique Gestion des élevages Maître de conférences Stagiaire
Mme DELIGNETTEMULLER Marie-Laure Unité pédagogique Biologie fonctionnelle Professeur
M. DEMONT Pierre Unité pédagogique Santé Publique et Vétérinaire Professeur
Mme DESJARDINS PESSON Isabelle Unité pédagogique Equine Maître de conférences Contractuel
Mme DJELOUADJI Zorée Unité pédagogique Santé Publique et Vétérinaire Maître de conférences
Mme ESCRIOU Catherine Unité pédagogique Pathologie médicale des animaux de compagnie
Maître de conférences
M. FAU Didier Unité pédagogique Anatomie Chirurgie (ACSAI) Professeur
Mme FOURNEL Corinne Unité pédagogique Pathologie morphologique et clinique des animaux de compagnie
Professeur
M. FRANCK Michel Unité pédagogique Gestion des élevages Professeur
M. FREYBURGER Ludovic Unité pédagogique Santé Publique et Vétérinaire Maître de conférences
M. FRIKHA Mohamed-Ridha Unité pédagogique Pathologie du bétail Maître de conférences
M. GENEVOIS Jean-Pierre Unité pédagogique Anatomie Chirurgie (ACSAI) Professeur
Mme GILOTFROMONT Emmanuelle Unité pédagogique Biologie fonctionnelle Professeur
M. GONTHIER Alain Unité pédagogique Santé Publique et Vétérinaire Maître de conférences
Mme GRAIN Françoise Unité pédagogique Gestion des élevages Professeur
M. GRANCHER Denis Unité pédagogique Gestion des élevages Maître de conférences
Mme GREZEL Delphine Unité pédagogique Santé Publique et Vétérinaire Maître de conférences
M. GUERIN Pierre Unité pédagogique Biotechnologies et pathologie de la reproduction
Professeur
Mme GUERINFAUBLEE Véronique Unité pédagogique Santé Publique et Vétérinaire Maître de conférences
Mme HUGONNARD Marine Unité pédagogique Pathologie médicale des animaux de compagnie
Maître de conférences
M. JUNOT Stéphane Unité pédagogique Anatomie Chirurgie (ACSAI) Maître de conférences
M. KECK Gérard Unité pédagogique Biologie fonctionnelle Professeur
M. KODJO Angeli Unité pédagogique Santé Publique et Vétérinaire Professeur
Mme LAABERKI Maria-Halima Unité pédagogique Santé Publique et Vétérinaire Maître de conférences Stagiaire
M. LACHERETZ Antoine Unité pédagogique Santé Publique et Vétérinaire Professeur
Mme LAMBERT Véronique Unité pédagogique Gestion des élevages Maître de conférences
Mme LE GRAND Dominique Unité pédagogique Pathologie du bétail Maître de conférences
Mme LEBLOND Agnès Unité pédagogique Santé Publique et Vétérinaire Professeur
Page | 4
Mme LEFRANCPOHL Anne-Cécile Unité pédagogique Equine Maître de conférences
M. LEPAGE Olivier Unité pédagogique Equine Professeur
Mme LOUZIER Vanessa Unité pédagogique Biologie fonctionnelle Maître de conférences
M. MARCHAL Thierry Unité pédagogique Pathologie morphologique et clinique des animaux de compagnie
Professeur
Mme MIALET Sylvie Unité pédagogique Santé Publique et Vétérinaire Inspecteur en santé publique vétérinaire (ISPV)
Mme MICHAUD Audrey Unité pédagogique Gestion des élevages Maître de conférences Stagiaire
M. MOUNIER Luc Unité pédagogique Gestion des élevages Maître de conférences
M. PEPIN Michel Unité pédagogique Santé Publique et Vétérinaire Professeur
M. PIN Didier Unité pédagogique Pathologie morphologique et clinique des animaux de compagnie
Maître de conférences
Mme PONCE Frédérique Unité pédagogique Pathologie médicale des animaux de compagnie
Maître de conférences
Mme PORTIER Karine Unité pédagogique Anatomie Chirurgie (ACSAI) Maître de conférences
Mme POUZOTNEVORET Céline Unité pédagogique Anatomie Chirurgie (ACSAI) Maître de conférences Stagiaire
Mme PROUILLAC Caroline Unité pédagogique Biologie fonctionnelle Maître de conférences
Mme REMY Denise Unité pédagogique Anatomie Chirurgie (ACSAI) Professeur
M. ROGER Thierry Unité pédagogique Anatomie Chirurgie (ACSAI) Professeur
M. SABATIER Philippe Unité pédagogique Biologie fonctionnelle Professeur
M. SAWAYA Serge Unité pédagogique Anatomie Chirurgie (ACSAI) Maître de conférences
Mme SEGARD Emilie Unité pédagogique Anatomie Chirurgie (ACSAI) Maître de conférences Contractuel
Mme SERGENTET Delphine Unité pédagogique Santé Publique et Vétérinaire Maître de conférences
Mme SONET Juliette Unité pédagogique Anatomie Chirurgie (ACSAI) Maître de conférences Contractuel
M. THIEBAULT Jean-Jacques Unité pédagogique Biologie fonctionnelle Maître de conférences
M. VIGUIER Eric Unité pédagogique Anatomie Chirurgie (ACSAI) Professeur
Mme VIRIEUXWATRELOT Dorothée Unité pédagogique Pathologie morphologique et clinique des animaux de compagnie
Maître de conférences Contractuel
M. ZENNER Lionel Unité pédagogique Santé Publique et Vétérinaire Professeur

Page | 5
Remerciements
A Monsieur le Professeur Philippe CHEVALIER, De la Faculté de Médecine de Lyon,
Pour nous avoir fait l’honneur d’accepter la présidence de notre jury de thèse, Qu’il reçoive ici l’expression de ma gratitude et de mes hommages respectueux.
-
A Monsieur le Professeur Théodore ALOGNINOUWA, Du Campus Vétérinaire de VetAgroSup,
Qui nous a fait l’honneur d’encadrer ce travail, Qu’il trouve ici l’assurance de notre plus profonde reconnaissance.
-
A Madame le Professeur Jeanne-Marie BONNET, Du Campus Vétérinaire de VetAgroSup,
Qui nous a fait l’honneur de faire partie de notre jury de thèse, Pour avoir accepté de juger ce travail,
Qu’elle trouve ici l’expression de mes remerciements les plus sincères.

Page | 6

Page | 7
A mes parents,
A mes frères,
A mes grand-parents,

Page | 8

Page | 9
Sommaire
TABLE DES ILLUSTRATIONS ............................................................................................................................12
INTRODUCTION ..............................................................................................................................................13
PREMIERE PARTIE : FONDEMENTS STRUCTURELS ET FONCTIONNELS DES AFFECTIONS CARDIO-
VASCULAIRES AVIAIRES ...................................................................................................................................15
I. ELEMENTS ANATOMIQUES D’INTERET DANS LA GESTION DES AFFECTIONS CARDIOVASCULAIRES ..............16
A. CŒUR ............................................................................................................................................................... 16
1. Localisation ................................................................................................................................................ 16
2. Aspect extérieur ......................................................................................................................................... 17
3. Structure interne ........................................................................................................................................ 20
4. Irrigation .................................................................................................................................................... 26
B. VASCULARISATION............................................................................................................................................... 28
1. Réseau artériel ........................................................................................................................................... 28
2. Réseau veineux .......................................................................................................................................... 30
3. Singularités du réseau vasculaire aviaire................................................................................................... 32
II. HISTOLOGIE .................................................................................................................................................34
A. LA PAROI CARDIAQUE ........................................................................................................................................... 34
1. Endocarde .................................................................................................................................................. 34
2. Myocarde ................................................................................................................................................... 34
3. Épicarde et péricarde ................................................................................................................................. 35
4. Tissu de conduction ................................................................................................................................... 35
B. LES VALVES CARDIAQUES ...................................................................................................................................... 36
C. VAISSEAUX SANGUINS .......................................................................................................................................... 37
1. Les artères ................................................................................................................................................. 37
2. Les capillaires ............................................................................................................................................. 39
3. Les veines ................................................................................................................................................... 40
III. PHYSIOLOGIE ...............................................................................................................................................41
A. ROLES DE L’APPAREIL CARDIOVASCULAIRE CHEZ LES OISEAUX ....................................................................................... 41
B. PHYSIOLOGIE CARDIAQUE ..................................................................................................................................... 41
1. Mécanique du cycle cardiaque .................................................................................................................. 41
2. Electrophysiologie ...................................................................................................................................... 43
C. HEMODYNAMIQUE VASCULAIRE ............................................................................................................................. 47
1. Généralités ................................................................................................................................................. 47
2. Particularités de la circulation systémique ................................................................................................ 47
3. Particularités de la circulation pulmonaire ................................................................................................ 48
4. Particularité de la circulation coronaire .................................................................................................... 48
5. Particularité du système porte-rénal ......................................................................................................... 49
D. CONTROLE DES PARAMETRES CARDIOVASCULAIRES .................................................................................................... 50
1. Contrôle mécanique ................................................................................................................................... 50
2. Contrôle nerveux (incluant la réponse surrénalienne) ............................................................................... 50
3. Contrôle humoral ....................................................................................................................................... 51
DEUXIEME PARTIE : ETIOLOGIE ET PATHOGENIE DES AFFECTIONS CARDIOVASCULAIRES AVIAIRES .....................55
I. AFFECTIONS CARDIAQUES ................................................................................................................................56
A. INSUFFISANCE CARDIAQUE CONGESTIVE ........................................................................................................................ 56
B. L’ARRET CARDIAQUE ................................................................................................................................................. 58
C. AFFECTIONS DE LA PAROI CARDIAQUE........................................................................................................................... 59
1. Les affections endocardiques ......................................................................................................................... 59

Page | 10
2. Les affections myocardiques........................................................................................................................... 63
D. AFFECTIONS DU PERICARDIQUE ................................................................................................................................... 78
1. Etiologie .......................................................................................................................................................... 78
2. Démarche diagnostique lors d’affection péricardique ................................................................................... 79
E. MALADIES CARDIAQUES CONGENITALES ........................................................................................................................ 80
1. Facteurs tératogènes ...................................................................................................................................... 80
2. Anomalies congénitales cardio-vasculaires .................................................................................................... 80
3. Symptomatologie ........................................................................................................................................... 80
4. Diagnostic des maladies congénitales ............................................................................................................ 81
II. AFFECTIONS VASCULAIRES ..............................................................................................................................81
A. VASCULOPATHIES METABOLIQUES ............................................................................................................................... 81
B. VASCULOPATHIES NUTRITIONNELLES ............................................................................................................................ 86
C. EMBOLIES VASCULAIRES ............................................................................................................................................ 87
D. VASCULITES ............................................................................................................................................................ 88
E. NEOPLASIES VASCULAIRES : HEMANGIOMES ET HEMANGIOSARCOMES ........................................................................... 89
F. VASCULOPATHIE IDIOPATHIQUE : ........................................................................................................................... 91
III. TROUBLES DE LA PRESSION ARTERIELLE ........................................................................................................94
A. HYPERTENSION ARTERIELLE SYSTEMIQUE ...................................................................................................................... 94
B. HYPOTENSION ARTERIELLE ......................................................................................................................................... 94
TROISIEME PARTIE : DIAGNOSTIC DES AFFECTIONS CARDIOVASCULAIRES AVIAIRES ............................95
I. HISTORIQUE .................................................................................................................................................96
A. COMMEMORATIFS .............................................................................................................................................. 96
B. ANAMNESE ........................................................................................................................................................ 96
II. EXAMEN CLINIQUE ET INTERPRETATION DES TROUVAILLES ........................................................................96
A. OBSERVATION A DISTANCE .................................................................................................................................... 96
B. EXAMEN MANUEL ............................................................................................................................................... 97
1. Mesure de la température cloacale ........................................................................................................... 97
2. Evaluation du système circulatoire périphérique....................................................................................... 97
3. La palpation cœlomique ............................................................................................................................ 98
C. AUSCULTATION................................................................................................................................................... 98
D. RECUPERATION SUITE A LA CONTENTION .................................................................................................................. 98
III. EXAMENS PARACLINIQUES ..........................................................................................................................99
A. PONCTION DES LIQUIDES ...................................................................................................................................... 99
B. MESURE DE LA PRESSION ARTERIELLE .................................................................................................................... 100
C. IMAGERIE MEDICALE .......................................................................................................................................... 102
D. EXAMEN ELECTROCARDIOGRAPHIQUE ................................................................................................................... 115
1. Indications et limites ................................................................................................................................ 115
2. Matériel ................................................................................................................................................... 115
3. Technique ................................................................................................................................................ 116
4. Interprétation .......................................................................................................................................... 117
E. BIOCHIMIE SERIQUE ........................................................................................................................................... 123
F. HEMATOLOGIE ................................................................................................................................................. 125
G. RECHERCHE D’AGENT INFECTIEUX ......................................................................................................................... 125
H. EXAMEN NECROPSIQUE POST-MORTEM ................................................................................................................. 126

Page | 11
QUATRIEME PARTIE : THERAPEUTIQUE CARDIOVASCULAIRE ......................................................... 129
I. LES GRANDES FAMILLES DE MEDICAMENTS ............................................................................................... 130
A. LES MODIFICATEURS DE L’INOTROPISME ................................................................................................................ 130
1. Les inotropes positifs ............................................................................................................................... 130
2. Les inotropes négatifs .............................................................................................................................. 133
B. LES MODIFICATEURS DES VAISSEAUX ............................................................................................................. 133
1. Vasoconstricteurs .................................................................................................................................... 133
2. Vasodilatateurs ........................................................................................................................................ 133
3. Les diurétiques ......................................................................................................................................... 135
II. LE TRAITEMENT DES MALADIES CARDIOVASCULAIRES .............................................................................. 136
A. TRAITEMENT DES SYNDROMES CARDIO-VASCULAIRES ............................................................................................... 136
1. Gestion de l’insuffisance cardiaque ......................................................................................................... 136
2. Gestion des dysrythmies .......................................................................................................................... 137
3. Gestion de l’arrêt cardiaque .................................................................................................................... 137
4. Gestion de l’insuffisance artérielle ........................................................................................................... 138
B. TRAITEMENT DE SUPPORT ................................................................................................................................... 138
1. Fluidothérapie .......................................................................................................................................... 138
2. Alimentation ............................................................................................................................................ 139
3. Chaleur et humidité ................................................................................................................................. 139
4. Oxygénation ............................................................................................................................................ 139
5. Stress ....................................................................................................................................................... 139
6. Résolution des épanchements ................................................................................................................. 139
C. TRAITEMENT ETIOLOGIQUE ................................................................................................................................. 140
1. L’athérosclérose ....................................................................................................................................... 140
2. Les infections ........................................................................................................................................... 140
3. Néoplasie ................................................................................................................................................. 140
4. Les intoxications ...................................................................................................................................... 141
5. Hydropéricarde ........................................................................................................................................ 141
6. La cardiomyopathie dilatée du dindon .................................................................................................... 141
7. Les carences ............................................................................................................................................. 141
CONCLUSION .................................................................................................................................................... 143
ANNEXES ......................................................................................................................................................... 145
ANNEXE I : VALEURS DE REFERENCES POUR L’ECHOCARDIOGRAPHIE CHEZ QUELQUES ESPECES. ............................... 147
ANNEXE II : ELECTROCARDIOGRAMME - VALEURS ET RYTHMES NORMAUX EN DERIVATION II. ................................. 149
ANNEXE III – TROUBLES DU RYTHME DECRITS EN MEDECINE AVIAIRE............................................................................. 151
BIBLIOGRAPHIE .................................................................................................................................................. 153

Page | 12
Table des illustrations
Figure 1 - Localisation du coeur : Vue ventrodorsale (à gauche) et vue latéro-latérale (à droite) .... 17
Figure 2 - Anatomie externe du coeur : Face dorsale (à gauche) et face ventrale (à droite) .............. 19
Figure 3 - Structure péricardique ............................................................................................................ 19
Figure 4 - Structure interne du cœur : détail des atria et des communications atrio-ventriculaires 22
Figure 5 - Structure interne du cœur : détail des ventricules ............................................................... 25
Figure 6 - Irrigation cardiaque : Face ventrale (à droite) et face dorsale (à gauche).......................... 27
Figure 7 - Anatomie du réseau vasculaire .............................................................................................. 31
Figure 8 - Structure histologique des principaux types vasculaires ..................................................... 40
Figure 9 - Système cardionecteur ............................................................................................................ 47
Figure 10 - Electrocardiogramme aviaire normal et mesures d’intérêt............................................. 117
Figure 11 - Construction du système hexaxial de Bailey par combinaison des dérivation triaxiales
bipolaires et unipolaires augmentées ................................................................................................... 121
Tableau 1- Pourcentage cardiaque de la masse corporelle aviaire d’après 1 (NICKEL, et al., 1977), 2
(GRUBB, 1983), 3 (TADJALLI, et al., 2009), 4 (KRAUTWALD-JUNGHANNS, et al., 2004), 5
(HARTMANN, 1961), 6 (BISHOP, et al., 1995) ....................................................................................... 18

Page | 13
INTRODUCTION
Si le vol est la plus remarquable des capacités des oiseaux et la clef de leur grande diversité en terme d’habitat et d’alimentation, les exigences métaboliques qu’il impose sont très élevées et constituent une contrainte majeure à la diversification structurelle au sein de cette classe animale. Ainsi, certains auteurs estiment qu’il y a moins de variations morphologiques parmi les 9700 espèces d’oiseaux que parmi, le seul ordre mammalien des carnivores, lequel ne représente pourtant que 300 espèces environ (MAINA, 1996). En conséquence, le système cardio-vasculaire qui est le garant de l’approvisionnement énergétique et gazeux aux organes du vol d’une part, et de l’élimination des déchets métaboliques que ce mode de déplacement produit en grande quantité d’autre part représente sans doute le système sur lequel repose les plus grandes exigences. Pourtant cet appareil qui est quasiment invariable sur le plan anatomo-physiologique assure d’autres fonctions comme la thermorégulation, et ce dans des situations d’une diversité mirobolante. C’est là la prouesse du système cardio-vasculaire aviaire qui, contraint à une uniformité structurelle et fonctionnelle par les exigences du vol, est doté d’un pouvoir d’adaptation exceptionnelle. Ceci lui permet d’assurer avec constance l’apport énergétique et gazeux nécessaire au bon fonctionnement de l’organisme et ce, dans toutes les situations que les oiseaux peuvent rencontrer, y compris les plus extrêmes comme la nage profonde en eau froide, le vol à hautes altitudes ou encore la course en désert aride. De par son importance capitale lors du processus de vol, l’appareil cardio-vasculaire est un système relativement peu sujet aux modifications d’ordre pathologique en situation naturelle. Néanmoins, tout comme le développement de la civilisation prédispose l’homme à l’apparition de maladies cardio-vasculaires, la domestication des oiseaux tend à augmenter l’incidence de ces affections par la sédentarité qu’elle impose et par les aberrations quantitatives et qualitatives de l’alimentation qui la caractérisent. Ainsi, de récentes études révèlent que 98% des perroquets de compagnie présente une anomalie atteignant le système cardio-vasculaire (KRAUTWALD-JUNGHANNS, et al., 2004). Enfin, si au sens large, le système cardio-vasculaire peut être défini comme un ensemble ‘’ clos de tubes ramifiés (vaisseaux) situés dans le tissu conjonctif et véhiculant les fluides extra cellulaires que sont le sang et la lymphe’’, nous limiterons notre étude au système cardio-vasculaire ‘’sanguin’’. Ainsi, les troubles hématologiques et lymphatiques ne seront pas détaillés. Par ailleurs, bien qu’il soit plus raisonnable de comparer les oiseaux avec les reptiles, puisque ces deux classes animales dérivent d’ancêtres dinosauriens thérodontes, nous préciserons la structure et le fonctionnement du système cardiovasculaire aviaire par comparaison avec celui des Mammifères, probablement mieux connu du lecteur.

Page | 14

Page | 15
PREMIERE PARTIE : FONDEMENTS STRUCTURELS ET FONCTIONNELS DES
AFFECTIONS CARDIO-VASCULAIRES AVIAIRES

Page | 16
I. Eléments anatomiques d’intérêt dans la gestion des affections
cardiovasculaires
A. Cœur
1. Localisation
a. Position
Chez les oiseaux, il n’existe pas de séparation entre la cavité thoracique et la cavité abdominale comparable au diaphragme chez les Mammifères. Cependant, des membranes aponévrotiques, connues sous le nom de septa horizontal et oblique, divisent la cavité thoraco-abdominale en plusieurs compartiments dénommés en fonction des organes qu’ils contiennent. Parmi ces cavités, la cavité cardio-abdominale est limitée dorsalement par la colonne vertébrale et le pelvis, latéralement par les deux septa obliques et ventralement par le sternum ainsi que par la paroi musculaire abdominale. Ce compartiment contient : le cœur dans son péricarde, le foie, le proventricule et la rate (NICKEL, et al., 1977).
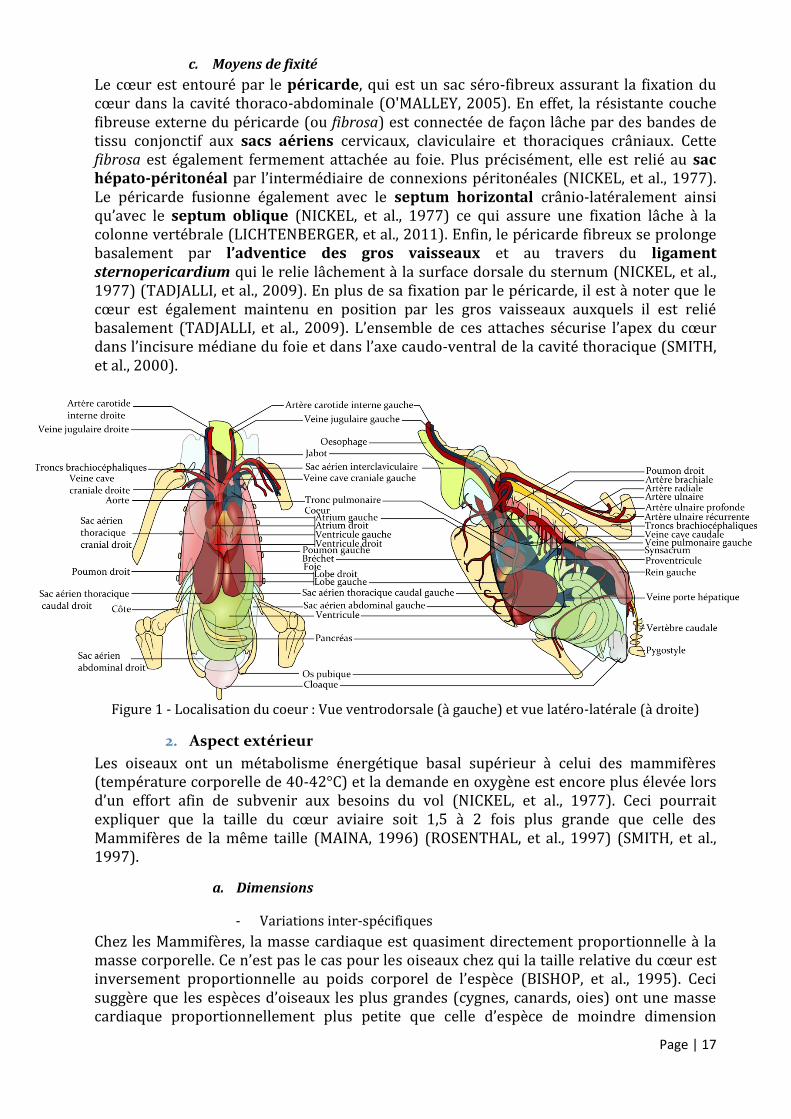
Le cœur est couché sur le plancher thoracique (CHATELAIN, 1992). Il est localisé dans la partie crâniale de la cavité cardio-abdominale (NICKEL, et al., 1977), en position axiale (AKESTER, 1984), parallèlement à la moelle épinière et à proximité du sternum mais son grand axe est légèrement dévié à droite de l’axe médian (PEES, et al., 2006). Il présente une orientation caudo-ventrale. En effet, la base du cœur est dirigée craniodorsalement et se projette à hauteur de la seconde côte alors que l’apex du cœur pointe vers le sternum et est situé sur un plan transverse passant entre la cinquième et la sixième côte (Fig.1) (NICKEL, et al., 1977). Chez l’autruche, il s’étend entre la première et la troisième côte (TADJALLI, et al., 2009). Il est contenu dans un solide sac péricardique fibreux délimitant un espace péricardique qui contient une petite quantité de fluide séreux (PEES, et al., 2006) jouant un rôle de lubrification (LICHTENBERGER, et al., 2011).
b. Rapports anatomiques
Les poumons, les segments crâniaux du septum horizontal, les deux bronches primaires, la syrinx et l’œsophage s’étendent contre la base du cœur, en position dorsale (NICKEL, et al., 1977). Contrairement aux Mammifères, les poumons s’étendent dorsalement au cœur mais ne l’entourent pas (Fig.1) (SMITH, et al., 1997). Les oiseaux n’ont pas de diaphragme (PEES, et al., 2006). En conséquence, les surfaces dorsale et latérales du cœur sont incluses dans le foie (NICKEL, et al., 1977) et l’apex cardiaque est complètement entouré par les lobes hépatiques. Cette inclusion se manifeste radiographiquement comme une superposition et ces deux organes apparaissent ainsi sous la silhouette d’un sablier en vue ventro-dorsale chez de nombreuses espèces de perroquet (O'MALLEY, 2005). Le côté gauche du cœur est en relation avec le proventricule (NICKEL, et al., 1977). La face ventrale du coeur est située à proximité du sternum. Enfin le cœur, ou plus précisément le péricarde, est en relation proche avec les diverticules des sacs aériens cervicaux, calviculaire et thoraciques crâniaux (NICKEL, et al., 1977). La présence de ces structures complique la réalisation d’échocardiographie chez les oiseaux (PEES, et al., 2004).
Page | 17
c. Moyens de fixité
Le cœur est entouré par le péricarde, qui est un sac séro-fibreux assurant la fixation du cœur dans la cavité thoraco-abdominale (O'MALLEY, 2005). En effet, la résistante couche fibreuse externe du péricarde (ou fibrosa) est connectée de façon lâche par des bandes de tissu conjonctif aux sacs aériens cervicaux, claviculaire et thoraciques crâniaux. Cette fibrosa est également fermement attachée au foie. Plus précisément, elle est relié au sac hépato-péritonéal par l’intermédiaire de connexions péritonéales (NICKEL, et al., 1977). Le péricarde fusionne également avec le septum horizontal crânio-latéralement ainsi qu’avec le septum oblique (NICKEL, et al., 1977) ce qui assure une fixation lâche à la colonne vertébrale (LICHTENBERGER, et al., 2011). Enfin, le péricarde fibreux se prolonge basalement par l’adventice des gros vaisseaux et au travers du ligament sternopericardium qui le relie lâchement à la surface dorsale du sternum (NICKEL, et al., 1977) (TADJALLI, et al., 2009). En plus de sa fixation par le péricarde, il est à noter que le cœur est également maintenu en position par les gros vaisseaux auxquels il est relié basalement (TADJALLI, et al., 2009). L’ensemble de ces attaches sécurise l’apex du cœur dans l’incisure médiane du foie et dans l’axe caudo-ventral de la cavité thoracique (SMITH, et al., 2000).
Figure 1 - Localisation du coeur : Vue ventrodorsale (à gauche) et vue latéro-latérale (à droite)
2. Aspect extérieur
Les oiseaux ont un métabolisme énergétique basal supérieur à celui des mammifères (température corporelle de 40-42°C) et la demande en oxygène est encore plus élevée lors d’un effort afin de subvenir aux besoins du vol (NICKEL, et al., 1977). Ceci pourrait expliquer que la taille du cœur aviaire soit 1,5 à 2 fois plus grande que celle des Mammifères de la même taille (MAINA, 1996) (ROSENTHAL, et al., 1997) (SMITH, et al., 1997).
a. Dimensions
- Variations inter-spécifiques
Chez les Mammifères, la masse cardiaque est quasiment directement proportionnelle à la masse corporelle. Ce n’est pas le cas pour les oiseaux chez qui la taille relative du cœur est inversement proportionnelle au poids corporel de l’espèce (BISHOP, et al., 1995). Ceci suggère que les espèces d’oiseaux les plus grandes (cygnes, canards, oies) ont une masse cardiaque proportionnellement plus petite que celle d’espèce de moindre dimension

Page | 18
(SMITH, et al., 2000). La taille du cœur dépend également de la quantité d’énergie aérobie dépensé par l’espèce considérée. Par exemple, un grand oiseau comme le cygne aura un cœur relativement plus petit que celui d’un pigeon de course (MAINA, 1996) (SMITH, et al., 1997).
Espèce Poule 1 Dinde1 Autruche 3 Canard 2 Oie 2 Pigeon 1,6 Perruche ondulée 4
Perruche royale 4
Colibris 5
Pourcentage cardiaque de la masse corporelle
0,5 – 1,42 toutes races confondues
0,5 0,73 0,8 0,8 1,1 – 1,4 1,3 – 1,5 1,4 2,4
Tableau 1- Pourcentage cardiaque de la masse corporelle aviaire d’après 1 (NICKEL, et al., 1977), 2 (GRUBB, 1983), 3 (TADJALLI, et al., 2009), 4 (KRAUTWALD-JUNGHANNS, et al., 2004), 5
(HARTMANN, 1961), 6 (BISHOP, et al., 1995)
Il est à noter que les colibris ont, proportionnellement à leur taille, le cœur le plus gros de tous les oiseaux. Ceci leur permet de répondre à la demande aérobique élevée du vol stationnaire (SMITH, et al., 2000). Par comparaison avec la souris (chez qui le cœur compte pour 0,5% de la masse corporelle), le cœur des colibris représente 2,4%-2,5% de leur poids (HARTMANN, 1961). Toutefois, le poids des oiseaux étant variable au sein d’une même espèce, il semblerait que la longueur du sternum soit un meilleur paramètre à utiliser lors de l’évaluation nécropsique de la dimension cardiaque (KRAUTWALD-JUNGHANNS, et al., 2004).
- Variations inter-individuelles
Deux facteurs ont été reconnus comme influençant les dimensions du cœur chez les oiseaux au sein d’une même espèce : - Exercice physique : Des différences de dimension du muscle cardiaque et du travail cardiaque ont été mises en évidence entre des pigeons de volière non entraînés et des pigeons de course. Le développement d’un cœur ‘’athlétique’’ révèle ainsi la possibilité d’adaptation du cœur aviaire aux exigences requises par l’effort (KRAUTWALD-JUNGHANNS, et al., 2002). - Saison : Nous avons évoqué que, contrairement aux Mammifères, la masse cardiaque était inversement proportionnelle à la masse corporelle chez les oiseaux (LICHTENBERGER, et al., 2011). Cette différence pourrait résulter du fait que le cœur des espèces les plus grandes, comme l’oie de bernache, peut s’hypertrophier préalablement à leur migration de telle sorte que le cœur est alors proportionnel à leur poids (BISHOP, et al., 1995). Ceci laisse à penser que les grands oiseaux volant ont la capacité génétique d’augmenter la taille de leur cœur et par conséquent leur débit cardiaque, par l’intermédiaire de mécanismes humoraux saisonniers, ou à plus long terme, grâce à la sélection naturelle. Cette adaptation du cœur permet de compenser l’hyperviscosité sanguine secondaire à l’hypoxie rencontrée à haute altitude et qui se traduit par des résistances vasculaires à l’écoulement plus forte (SMITH, et al., 2000). Cette cardiomégalie permettrait également de diminuer la fréquence cardiaque lors du vol. En effet, lors d’un vol hors saison de migration, la fréquence cardiaque est plus élevée de 69% par rapport à un vol à la fin de la migration (BUTLER, et al., 2000).
b. Couleur
Le cœur des oiseaux présente une couleur normalement rouge foncée à bleu-rouge (NICKEL, et al., 1977) (TADJALLI, et al., 2009).
c. Structure externe
Page | 19
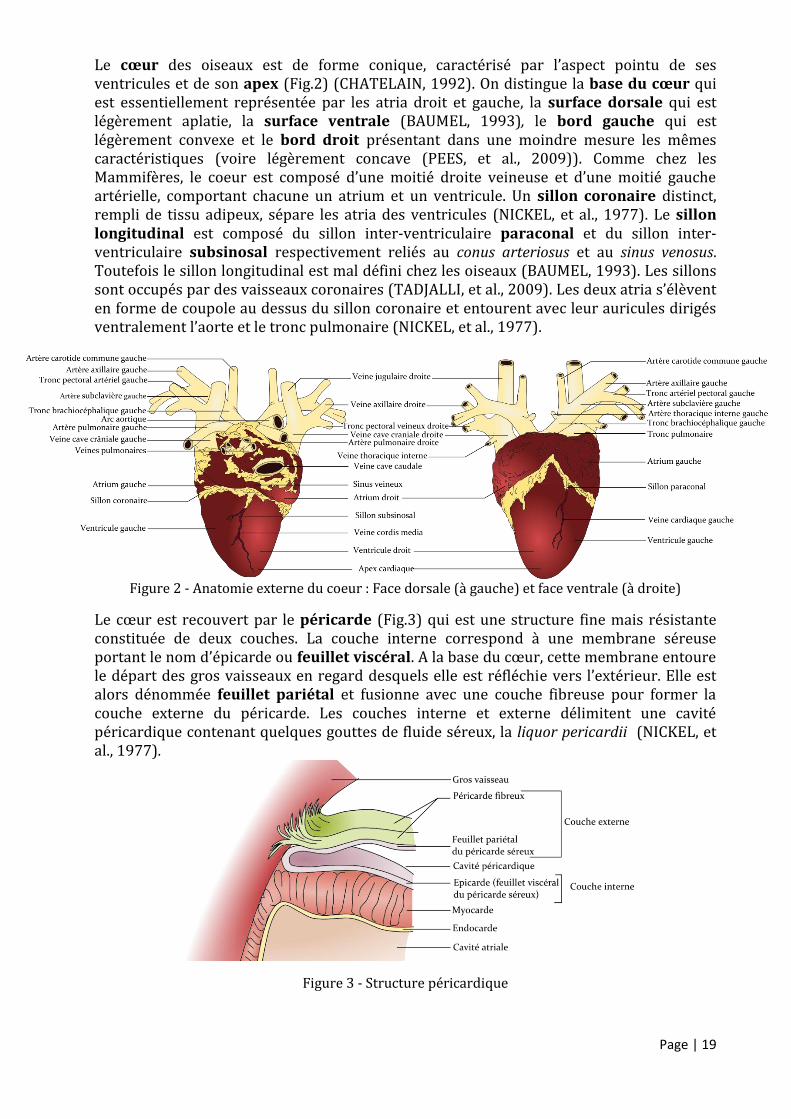
Le cœur des oiseaux est de forme conique, caractérisé par l’aspect pointu de ses ventricules et de son apex (Fig.2) (CHATELAIN, 1992). On distingue la base du cœur qui est essentiellement représentée par les atria droit et gauche, la surface dorsale qui est légèrement aplatie, la surface ventrale (BAUMEL, 1993), le bord gauche qui est légèrement convexe et le bord droit présentant dans une moindre mesure les mêmes caractéristiques (voire légèrement concave (PEES, et al., 2009)). Comme chez les Mammifères, le coeur est composé d’une moitié droite veineuse et d’une moitié gauche artérielle, comportant chacune un atrium et un ventricule. Un sillon coronaire distinct, rempli de tissu adipeux, sépare les atria des ventricules (NICKEL, et al., 1977). Le sillon longitudinal est composé du sillon inter-ventriculaire paraconal et du sillon inter-ventriculaire subsinosal respectivement reliés au conus arteriosus et au sinus venosus. Toutefois le sillon longitudinal est mal défini chez les oiseaux (BAUMEL, 1993). Les sillons sont occupés par des vaisseaux coronaires (TADJALLI, et al., 2009). Les deux atria s’élèvent en forme de coupole au dessus du sillon coronaire et entourent avec leur auricules dirigés ventralement l’aorte et le tronc pulmonaire (NICKEL, et al., 1977).
Figure 2 - Anatomie externe du coeur : Face dorsale (à gauche) et face ventrale (à droite)
Le cœur est recouvert par le péricarde (Fig.3) qui est une structure fine mais résistante constituée de deux couches. La couche interne correspond à une membrane séreuse portant le nom d’épicarde ou feuillet viscéral. A la base du cœur, cette membrane entoure le départ des gros vaisseaux en regard desquels elle est réfléchie vers l’extérieur. Elle est alors dénommée feuillet pariétal et fusionne avec une couche fibreuse pour former la couche externe du péricarde. Les couches interne et externe délimitent une cavité péricardique contenant quelques gouttes de fluide séreux, la liquor pericardii (NICKEL, et al., 1977).
Figure 3 - Structure péricardique

Page | 20
3. Structure interne
La structure interne générale du cœur est relativement similaire à celle des Mammifères. En effet, les quatre chambres du cœur sont complètement individualisées en deux atria et deux ventricules (SMITH, et al., 2000). Ainsi, le cœur est divisé en deux parties de sorte que les sangs oxygénés et désoxygénés ne se mélangent jamais (O'MALLEY, 2005).
a. Atria
- Septum inter-atrial et musculature atriale
La structure des atria aviaires est supportée par une musculature comportant des parties bien distinctes. On note ainsi l’existence de plusieurs arcs musculaires soutenant le toit. L’arc longitudinal dorsal (arc dorsal médian) est incurvé dorsalement, de façon quasiment parallèle à l’attache crâniale du septum inter-atrial et se subdivise à l’approche des ostia des veines pulmonaires. Cet arc est dédoublé dans le cœur des autruches (BAUMEL, 1993). Il émet deux branches latérales : L’arc transverse droit et l’arc transverse gauche (TADJALLI, et al., 2009) qui parcourent le toit des atria droit et gauche. Ils sont orientées à angle droit avec l’arc longitudinal et le septum inter-atrial (SMITH, et al., 2000). Latéralement, chaque arc transverse se ramifie et entre en continuité avec les muscles pectinés généraux (BAUMEL, 1993) qui forment un maillage faisant protrusion dans la lumière atriale (NICKEL, et al., 1977). Une partie des muscles pectinés est en continuité avec la base des valvules sinuatriales (BAUMEL, 1993) mais la majorité des ramifications fusionnent avec le muscle basiannulaire qui est une couche circulaire de muscle entourant la base des atria. Il définit le sillon coronaire et met les atria en contact avec les ventricules (Fig.4) (SMITH, et al., 2000).
Un fin septum inter-atrial sépare l’atrium droit de l’atrium gauche. Au cours du développement embryonnaire et jusqu’à la vie néonatale, ce septum a des perforations en forme de cribles qui autorisent le sang à passer de l’atrium droit à l’atrium gauche (NICKEL, et al., 1977). Fonctionnellement, ces perforations correspondent au foramen ovale fœtaux des mammifères (BAUMEL, 1993). Avec le développement de la respiration pulmonaire, ces ouvertures en forme de fente disparaissent sans laisser de traces (NICKEL, et al., 1977). Chez les oiseaux adultes, il n’y a donc pas d’équivalent de la fossa ovalis décrite chez les Mammifères (BAUMEL, 1993).
- Atrium droit
Organisation générale : L’atrium droit reçoit le sang en provenance des veines caves avant de l’injecter dans le ventricule droit (NICKEL, et al., 1977). Cette chambre a un volume supérieur à celle de gauche chez la plupart des oiseaux (CARUS, 1838) et sa paroi est par certains endroits excessivement fine (NICKEL, et al., 1977). Nous noterons l’existence du récessus gauche de l’atrium droit, un diverticule atrial tubulaire caractéristique du cœur aviaire qui s’étend vers la gauche et dorsalement au bulbe aortique jusqu’à dépasser le plan médian du cœur chez la poule et le canard. Ce récessus est séparé de l’atrium gauche par une partie du septum inter-atrial correspondant à un vestige du septum fœtal (Fig.4) (BAUMEL, 1993).
Abouchement des veines caves et cordis : Les trois veines caves abouchent à la surface atriale. La très large veine cave caudale se projette dans la partie basale de l’atrium droit au travers d’un orifice grossièrement circulaire. Chez les oiseaux, il existe deux veines caves crâniales : la droite entre dans l’atrium au dessus de la terminaison de la veine cave caudale, alors que la gauche entre dans l’atrium droit immédiatement au dessus du sillon coronaire et à la gauche de la veine cave caudale. C’est également à cet endroit que se termine la v. cordis media. La v. cordis magna s’abouche à l’opposé, sur la face ventrale, entre le tronc pulmonaire et l’atrium gauche (NICKEL, et al., 1977).

Page | 21
Chez les taxons Gallus (poule domestique), Struthio (autruche), Corvus (corbeau) et Apteryx (kiwi) le sinus veineux embryonnaire n’est pas complètement incorporé à l’intérieur de la paroi de l’atrium droit (KING, et al., 1984) (BAUMEL, 1993). En conséquence, les veines caves crâniales droite et caudale entrent d’abord dans un sinus veineux avant de se jeter dans l’atrium droit (AKESTER, 1984) (SMITH, et al., 1997) (WEST, et al., 1981). Ce sinus est plus ou moins délimité par un sillon extérieur (BAUMEL, 1993). Toutefois, intérieurement, il est séparé de l’atrium droit par de fines valvules musculaires dites « sinuatriales droite et gauche ». Elles sont également appelées valvule d’Eustache (CRESPEAU, 1992) et interdisent le reflux dans la veine cave (CARUS, 1838). Ces valvules correspondent au muscle pectiné valvulaire (NICKEL, et al., 1977). Le septum du sinus sépare l’ouverture de la veine cave crâniale gauche de l’orifice des deux autre veines (KING, et al., 1984) (TADJALLI, et al., 2009).
Nous noterons que chez certaines espèces, la veine proventriculaire crâniale qui draine le sang veineux de l’estomac glandulaire (ou proventricule) peut rejoindre la veine cave crâniale gauche à proximité de sa terminaison ou alors se vider directement dans l’atrium droit via l’ostium de la veine proventriculaire crâniale (BAUMEL, 1993).
- Atrium gauche
Organisation générale : L’atrium gauche forme la partie caudale de la base cardiaque et s’étend dorsalement au ventricule gauche (TADJALLI, et al., 2009). Il est moins volumineux que le droit, mais sa paroi est plus épaisse. Lui aussi est soutenu par le tronc musculaire qui se poursuit à partir de l’atrium droit, au travers du septum inter-atrial jusqu’au toit de l’atrium gauche, où il se divise également en de nombreux muscles pectinés, se disséminant le long de la paroi (Fig.4) (NICKEL, et al., 1977).
Abouchement des veines pulmonaires : Chez les oiseaux, les veines pulmonaires droite et gauche s’ouvrent dans l’atrium gauche soit ensembles, soit séparément (KING, et al., 1984). Chez les autruches, les veines pulmonaires entrent dans l’atrium gauche indépendamment et sont complètement séparées l’une de l’autre par un septum (TADJALLI, et al., 2009). C’est également le cas des genres Columba et Anas. Chez la poule domestique, les deux veines pulmonaires abouchent généralement séparément à la face atriale mais elles se terminent occasionnellement dans un diverticule de l’atrium gauche. Chez la dinde, les deux veines pulmonaires deviennent confluentes en dehors du cœur et forment une veine pulmonaire dite commune qui abouche dans l’atrium (NICKEL, et al., 1977) ainsi que chez le genre Melopsittacus (BAUMEL, 1993).
Que les veines fusionnent avant ou après avoir pénétré dans l’atrium, elle aboutissent quelque soit l’espèce à la formation d’un seul vaisseau court. Ce vaisseau s’invagine dans l’atrium gauche et fait protrusion en direction de l’ostium atrioventriculaire gauche. Cette veine invaginée est appelée chambre pulmonaire et forme une sous-cavité de l’atrium gauche (Fig.4). Le côté gauche de cette sous-cavité est constitué par une paroi libre qui dirige le sang dans le ventricule gauche, sépare la chambre de la cavité générale de l’atrium gauche (BAUMEL, 1993) et constituerait une valve anti-reflux. Cette valve de la veine pulmonaire est musculaire, en forme de faucille et semble empêcher la régurgitation sanguine du ventricule gauche vers les veines caves (NICKEL, et al., 1977). Elle est beaucoup plus réduite chez les passereaux (KING, et al., 1984).
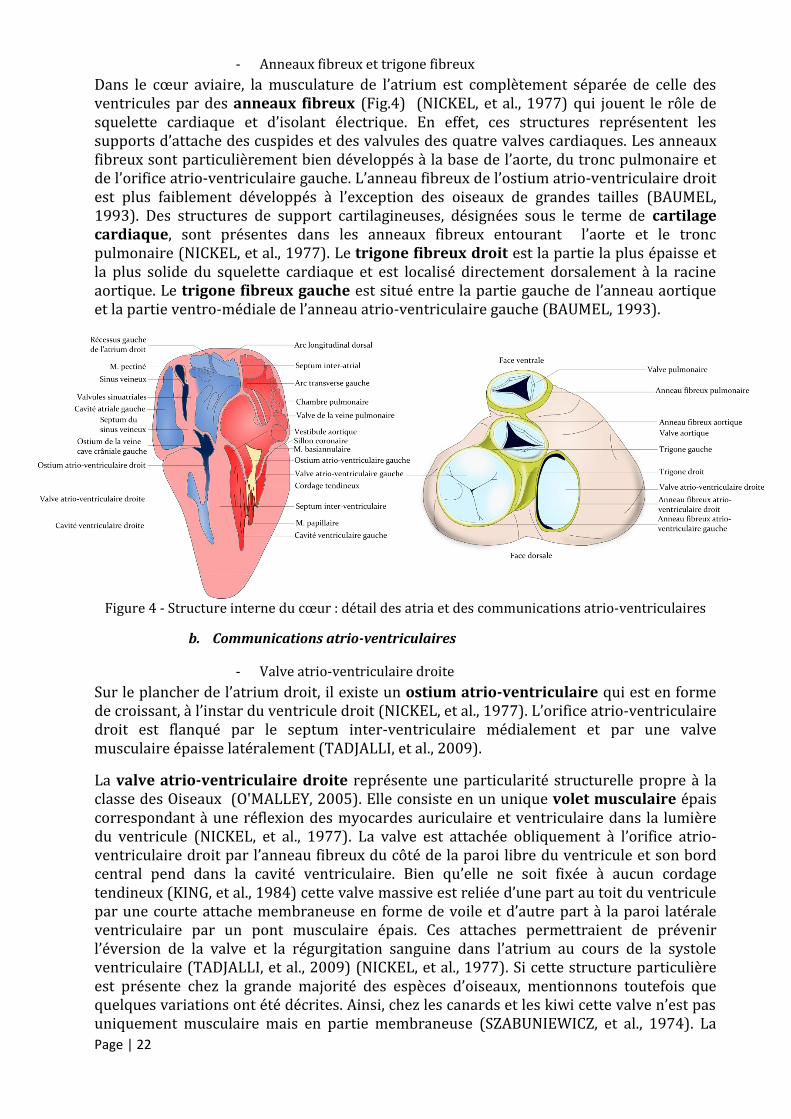
Page | 22
- Anneaux fibreux et trigone fibreux
Dans le cœur aviaire, la musculature de l’atrium est complètement séparée de celle des ventricules par des anneaux fibreux (Fig.4) (NICKEL, et al., 1977) qui jouent le rôle de squelette cardiaque et d’isolant électrique. En effet, ces structures représentent les supports d’attache des cuspides et des valvules des quatre valves cardiaques. Les anneaux fibreux sont particulièrement bien développés à la base de l’aorte, du tronc pulmonaire et de l’orifice atrio-ventriculaire gauche. L’anneau fibreux de l’ostium atrio-ventriculaire droit est plus faiblement développés à l’exception des oiseaux de grandes tailles (BAUMEL, 1993). Des structures de support cartilagineuses, désignées sous le terme de cartilage cardiaque, sont présentes dans les anneaux fibreux entourant l’aorte et le tronc pulmonaire (NICKEL, et al., 1977). Le trigone fibreux droit est la partie la plus épaisse et la plus solide du squelette cardiaque et est localisé directement dorsalement à la racine aortique. Le trigone fibreux gauche est situé entre la partie gauche de l’anneau aortique et la partie ventro-médiale de l’anneau atrio-ventriculaire gauche (BAUMEL, 1993).
Figure 4 - Structure interne du cœur : détail des atria et des communications atrio-ventriculaires
b. Communications atrio-ventriculaires
- Valve atrio-ventriculaire droite
Sur le plancher de l’atrium droit, il existe un ostium atrio-ventriculaire qui est en forme de croissant, à l’instar du ventricule droit (NICKEL, et al., 1977). L’orifice atrio-ventriculaire droit est flanqué par le septum inter-ventriculaire médialement et par une valve musculaire épaisse latéralement (TADJALLI, et al., 2009).
La valve atrio-ventriculaire droite représente une particularité structurelle propre à la classe des Oiseaux (O'MALLEY, 2005). Elle consiste en un unique volet musculaire épais correspondant à une réflexion des myocardes auriculaire et ventriculaire dans la lumière du ventricule (NICKEL, et al., 1977). La valve est attachée obliquement à l’orifice atrio-ventriculaire droit par l’anneau fibreux du côté de la paroi libre du ventricule et son bord central pend dans la cavité ventriculaire. Bien qu’elle ne soit fixée à aucun cordage tendineux (KING, et al., 1984) cette valve massive est reliée d’une part au toit du ventricule par une courte attache membraneuse en forme de voile et d’autre part à la paroi latérale ventriculaire par un pont musculaire épais. Ces attaches permettraient de prévenir l’éversion de la valve et la régurgitation sanguine dans l’atrium au cours de la systole ventriculaire (TADJALLI, et al., 2009) (NICKEL, et al., 1977). Si cette structure particulière est présente chez la grande majorité des espèces d’oiseaux, mentionnons toutefois que quelques variations ont été décrites. Ainsi, chez les canards et les kiwi cette valve n’est pas uniquement musculaire mais en partie membraneuse (SZABUNIEWICZ, et al., 1974). La

Page | 23
structure singulière de cette valve pourrait jouer le rôle de la crista supraventricularis mammalienne dont les oiseaux sont dépourvus (LU, et al., 1993). Elle pourrait également représenter une adaptation à la fréquence cardiaque élevée. En effet, la fermeture rapide de la chambre cardiaque serait facilitée par la contraction active du muscle valvulaire (NICKEL, et al., 1977). Toutefois, aucune de ces hypothèses n’a été prouvée (PEES, et al., 2001).
- Valve atrio-ventriculaire gauche
L’ostium atrio-ventriculaire gauche est entouré par une crête musculaire circulaire et présente une forme d’entonnoir en dépression sur le plancher de l’atrium gauche. Cet orifice est gardé par la valve atrio-ventriculaire gauche. (NICKEL, et al., 1977). La valve atrio-ventriculaire gauche est fine, fibreuse et pratiquement transparente. En cela elle est davantage similaire à celles que l’on retrouve chez les Mammifères (O'MALLEY, 2005) (NICKEL, et al., 1977). Toutefois, celle-ci est généralement tricuspide et non bicuspide, comme c’est le cas chez les Mammifères (AKESTER, 1984) (SMITH, et al., 1997). Les trois cuspides membraneuses sont relativement mal définies mais sont qualifiées de cuspide dorsale (ou antérieure), gauche (ou postérieure) et septale (ou droite, médiale ou aortique) (NICKEL, et al., 1977). Cette dernière est la plus grande (TADJALLI, et al., 2009) et s’attache à la base des valvules sigmoïdes gauche et non-coronarienne de la valve aortique adjacente, par l’intermédiaire d’un tissu fibreux. Le bord libre de la valve est relié à l’endocarde du ventricule gauche par l’intermédiaire de nombreux cordages tendineux inextensibles, prévenant l’éversion de la valve au cours de la systole ventriculaire (SMITH, et al., 2000). La valve atrio-ventriculaire gauche a une base cartilagineuse qui fusionne avec l’élastine des plans plus profond (RANDALL, et al., 1996).
c. Ventricules
- Septum inter-ventriculaire et architecture ventriculaire
L’architecture musculaire des ventricules est plus complexe que celle des atria. On retiendra que le myocarde est composé de plusieurs muscles (SMITH, et al., 2000): - La lame superficielle ou muscle bulbo-spiral superficiel est un fin feuillet musculaire situé sous l’épicarde qui prend origine à partir du bord ventral de la base cardiaque et se dirige vers l’apex en suivant un mouvement spiralé où il forme la corne dorsale. A partir de ce point, les fibres prennent une direction ascendante et se dirigent avec une trajectoire spiralée entre le muscle bulbospiral profond et le muscle sinospiral (cf.infra) jusqu’à la racine aortique chez le poulet (BAUMEL, 1993) et l’autruche (SHANER, 1923). - Le groupe sino-spiral est composé du muscle sino atrial et du muscle de la valve atrio-ventriculaire droite. Le muscle sino-atrial prend son origine en région dorsale de la base du cœur, principalement à partir de l’anneau atrio-ventriculaire droit et est associé à la portion sinusale du cœur, dont il tire son nom. La plus grande partie de ses fibres passe superficiellement et en s’enroulant vers l’apex, où elles forment la corne ventral du vortex. Elles passent ensuite dans le cœur où elles constituent la plus grande partie de la paroi interne du ventricule gauche, et remontent en spirale vers la base du cœur. Les fibres qui ne participent pas à la formation des muscles papillaires se terminent avec l’anneau atrio-ventriculaire gauche. Un petit groupe de fibre, provenant de la même origine que les précédente passent du côté droit en suivant un trajet grossièrement circulaire et constituent la paroi latérale du ventricule droit (SHANER, 1923). En d’autres termes, la paroi libre du ventricule droit correspond à une prolongation de la portion externe de la paroi ventriculaire gauche qui s’enroule autour de la partie droite du cœur pour délimiter une cavité en forme de croissant qui n’atteint pas l’apex du cœur (SMITH, et al., 2000). Le muscle de la valve atrio-ventriculaire droite peut être considéré comme une partie spéciale de ces dernières fibres (SHANER, 1923).

Page | 24
- Le muscle longitudinal du ventricule droit forme au départ la paroi médiale du ventricule droit. Il émerge de la racine aortique s’enroule en direction apicale sous la surface ventrale du ventricule gauche puis rejoint le muscle bulbo-spiral superficiel (SHANER, 1923). - Le muscle bulbo-spiral profond est en grande partie un anneau fermé qui entoure la terminaison des muscles superficiels. Quelque soit son origine, celle-ci se trouve sous le muscle bulbo-spiral superficiel. Comme chez les Mammifères, son développement varie grandement entre les espèces. Ainsi, chez le canard Mallard il forme un anneau très bien délimité, alors que chez de nombreuses espèces de volaille, il est difficile à séparer des autres muscles (SHANER, 1923). La paroi droite du ventricule gauche forme le septum interventriculaire qui sépare les deux chambres (SMITH, et al., 2000). Le septum inter-ventriculaire chez l’autruche est représenté par une cloison musculaire très épaisse entre les ventricules droit et gauche. Ce septum est fortement bombé dans la lumière du ventricule droit (TADJALLI, et al., 2009) (NICKEL, et al., 1977).
- Ventricule droit
Le ventricule droit débute à la face auriculaire du cœur et continue jusqu’à la face atriale en s’étendant contre le ventricule gauche comme une petite poche. Il présente une légère courbure en direction de la pointe du cœur mais s’étend seulement sur les deux tiers basaux de la distance menant à l’apex du ventricule gauche (Fig.5) (NICKEL, et al., 1977)(SMITH, et al., 2000).
Organisation de la chambre : A la gauche de la valve atrio-ventriculaire droite se trouve la chambre d’éjection (ou chambre de chasse) du ventricule droit. Celle-ci se prolonge entre les deux auricules par le cône artériel et l’ostium du tronc pulmonaire (NICKEL, et al., 1977). La chambre de chasse du ventricule droit n’est pas facilement visualisable, mais il est possible de la sonder. Elle s’incurve en suivant une forme spiralée à partir du ventricule droit en longeant le septum inter-ventriculaire crânialement puis pour finir, latéralement aux poumons (KRAUTWALD-JUNGHANNS, et al., 2004). Particularités pariétales : Le ventricule droit présente une bande modératrice tendineuse à proximité de la base du ventricule qui s’étend du septum à la valve musculaire. D’autres bandes modératrices en forme de filament tendineux ou de feuillet mi-tendineux et mi-musculaire sont habituellement retrouvés en région apicale du ventricule droit et s’étendent du septum jusqu’à la paroi externe (TADJALLI, et al., 2009). La paroi externe du ventricule droit porte des petits reliefs musculaires, appelées trabécules charnues (NICKEL, et al., 1977), qui sont plus proéminentes dans la région de l’apex. Il n’y a pas de muscles papillaires ni de cordages tendineux (TADJALLI, et al., 2009).
- Ventricule gauche
Le ventricule gauche a la forme d’un cône creux à paroi épaisse et apparaît en section transversale presque rond. Il s’étend de l’orifice atrio-ventriculaire gauche jusqu’à l’apex du cœur (SMITH, et al., 2000). Organisation de la chambre : La chambre de chasse du ventricule gauche est proéminente et se situe à côté du septum inter-ventriculaire à proximité de la base du ventricule gauche (KRAUTWALD-JUNGHANNS, et al., 2004). L’aorte prend son origine du toit du ventricule gauche, entre l’auricule droit et le cône artériel du tronc pulmonaire. (NICKEL, et al., 1977). Le vestibule de l’aorte correspond à l’espace entre la cuspide septal de la valve atrio-ventriculaire gauche et le septum inter-ventriculaire duquel émerge l’aorte (TADJALLI, et al., 2009).
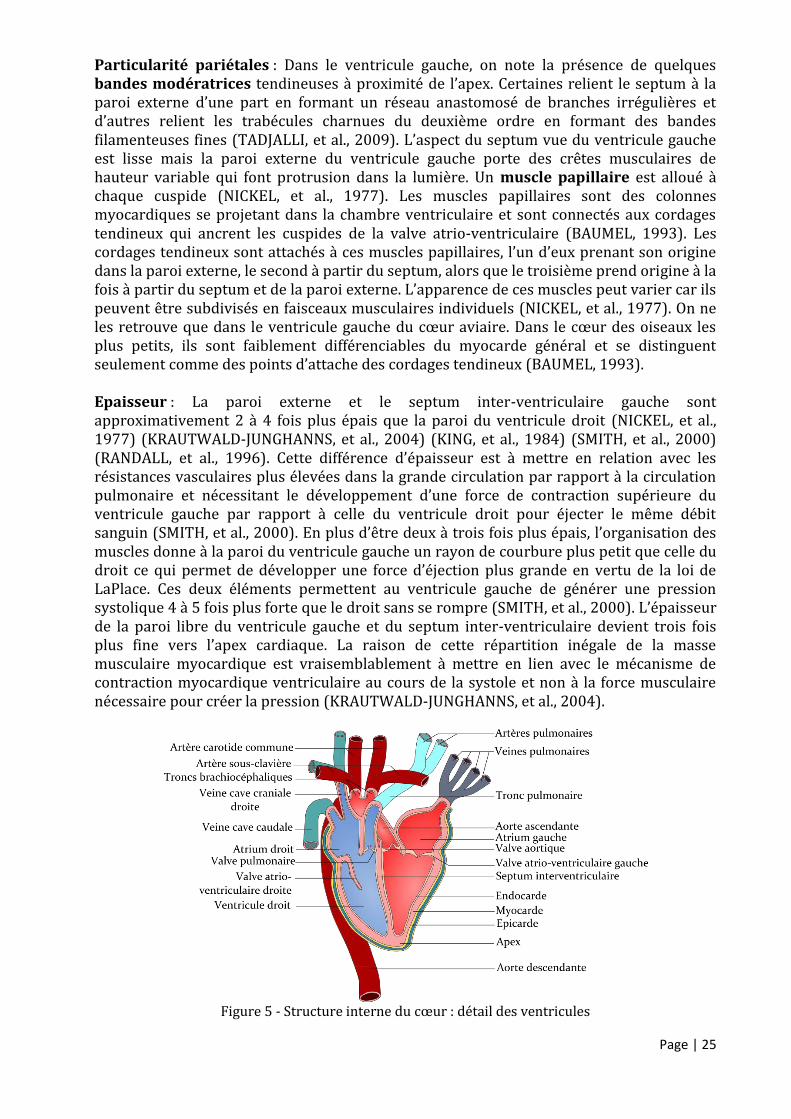
Page | 25
Particularité pariétales : Dans le ventricule gauche, on note la présence de quelques bandes modératrices tendineuses à proximité de l’apex. Certaines relient le septum à la paroi externe d’une part en formant un réseau anastomosé de branches irrégulières et d’autres relient les trabécules charnues du deuxième ordre en formant des bandes filamenteuses fines (TADJALLI, et al., 2009). L’aspect du septum vue du ventricule gauche est lisse mais la paroi externe du ventricule gauche porte des crêtes musculaires de hauteur variable qui font protrusion dans la lumière. Un muscle papillaire est alloué à chaque cuspide (NICKEL, et al., 1977). Les muscles papillaires sont des colonnes myocardiques se projetant dans la chambre ventriculaire et sont connectés aux cordages tendineux qui ancrent les cuspides de la valve atrio-ventriculaire (BAUMEL, 1993). Les cordages tendineux sont attachés à ces muscles papillaires, l’un d’eux prenant son origine dans la paroi externe, le second à partir du septum, alors que le troisième prend origine à la fois à partir du septum et de la paroi externe. L’apparence de ces muscles peut varier car ils peuvent être subdivisés en faisceaux musculaires individuels (NICKEL, et al., 1977). On ne les retrouve que dans le ventricule gauche du cœur aviaire. Dans le cœur des oiseaux les plus petits, ils sont faiblement différenciables du myocarde général et se distinguent seulement comme des points d’attache des cordages tendineux (BAUMEL, 1993). Epaisseur : La paroi externe et le septum inter-ventriculaire gauche sont approximativement 2 à 4 fois plus épais que la paroi du ventricule droit (NICKEL, et al., 1977) (KRAUTWALD-JUNGHANNS, et al., 2004) (KING, et al., 1984) (SMITH, et al., 2000) (RANDALL, et al., 1996). Cette différence d’épaisseur est à mettre en relation avec les résistances vasculaires plus élevées dans la grande circulation par rapport à la circulation pulmonaire et nécessitant le développement d’une force de contraction supérieure du ventricule gauche par rapport à celle du ventricule droit pour éjecter le même débit sanguin (SMITH, et al., 2000). En plus d’être deux à trois fois plus épais, l’organisation des muscles donne à la paroi du ventricule gauche un rayon de courbure plus petit que celle du droit ce qui permet de développer une force d’éjection plus grande en vertu de la loi de LaPlace. Ces deux éléments permettent au ventricule gauche de générer une pression systolique 4 à 5 fois plus forte que le droit sans se rompre (SMITH, et al., 2000). L’épaisseur de la paroi libre du ventricule gauche et du septum inter-ventriculaire devient trois fois plus fine vers l’apex cardiaque. La raison de cette répartition inégale de la masse musculaire myocardique est vraisemblablement à mettre en lien avec le mécanisme de contraction myocardique ventriculaire au cours de la systole et non à la force musculaire nécessaire pour créer la pression (KRAUTWALD-JUNGHANNS, et al., 2004).
Figure 5 - Structure interne du cœur : détail des ventricules

Page | 26
d. Orifices et valves de sortie des ventricules
Les valves situées à la sortie des ventricules sont au premier regard, davantage similaires à celles des Mammifères (O'MALLEY, 2005). L’orifice pulmonaire est circulaire et est gardé par la valve pulmonaire (Fig.5). La valve de sortie pulmonaire est constituée de trois valvules semi-lunaires ainsi organisées que l’une est à la gauche de l’ostium, une seconde à sa droite et la troisième sur le septum inter-ventriculaire (TADJALLI, et al., 2009). Ces valvules sigmoïdes empêchent la régurgitation de l’artère pulmonaire dans le ventricule au cours de la diastole (NICKEL, et al., 1977) (SMITH, et al., 2000). La paroi du tronc pulmonaire forme des renflements en regard de chaque cuspide, appelés sinus pulmonaires (TADJALLI, et al., 2009). L’orifice aortique est gardé par la valve de sortie aortique localisée dans la racine de l’aorte ascendante (NICKEL, et al., 1977). Sa position est marquée extérieurement par le bulbe aortique, la partie proximale distendue de l’aorte ascendante qui héberge les valves aortiques (BAUMEL, 1993) et qui donne naissance aux artères coronaires (TADJALLI, et al., 2009). La valve est composée de trois valvules semi-lunaires qualifiées de droite, gauche et septale (SMITH, et al., 2000). Elles sont épaisses, solides, rigides et attachées plus fermement au myocarde sous-jacent que ne le sont les valvules pulmonaires (BAUMEL, 1993).
4. Irrigation
a. Artères coronaires
Chez les oiseaux, le cœur est perfusé par deux artères coronaires (NICKEL, et al., 1977). Ces artères coronaires assurent un apport nutritif et gazeux au muscle cardiaque et leur lit vasculaire sont les premiers à être irrigué par l’aorte. Le plus fréquemment, on observe l’existence de deux artère coronaires chez les oiseaux mais il peut y en avoir jusqu’à quatre (SMITH, et al., 2000). L’artère coronaire droite prend origine au niveau du sinus de la valve semi-lunaire droite de l’aorte puis après un court trajet, elle se divise en un rameau superficiel et un rameau profond (Fig.6). La branche superficielle court dans le sillon coronaire vers la droite et, en atteignant la surface atriale envoie des branches pour la paroi de l’atrium et du ventricule droits. Les dernières branches irriguant le ventricule entrent rapidement dans la profondeur de la musculature où elles se ramifient davantage (NICKEL, et al., 1977). La branche profonde, très épaisse circule au sein du septum inter-ventriculaire en direction de l’apex du cœur et se divise en deux larges branches. Les subdivisions de ces branches quittent le septum et entrent dans la musculature du ventricule gauche ventralement et dorsalement. Elles s’anastomosent au bord gauche du cœur. L’artère coronaire droite irrigue via ses branches profondes et superficielles une très grande partie du cœur incluant la paroi de l’atrium droit, une grande partie du septum et la paroi du ventricule gauche (NICKEL, et al., 1977). La branche profonde assure la plus grande partie de l’irrigation (BAUMEL, 1993).
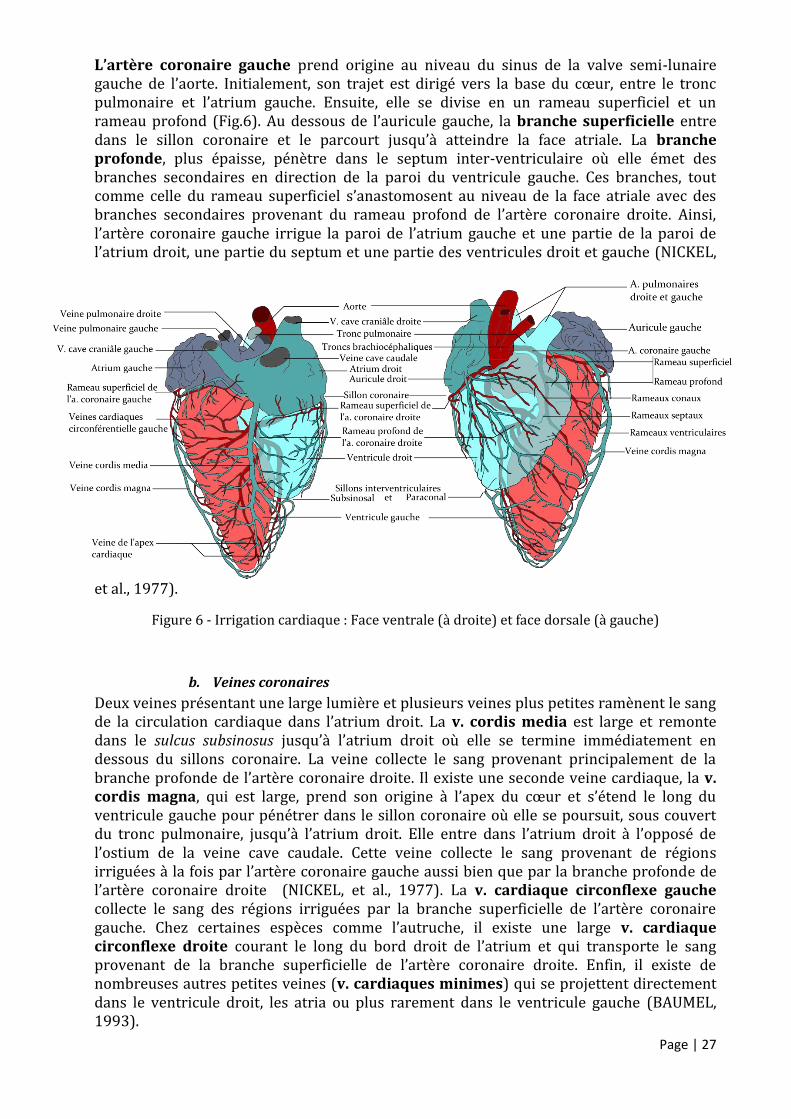
Page | 27
L’artère coronaire gauche prend origine au niveau du sinus de la valve semi-lunaire gauche de l’aorte. Initialement, son trajet est dirigé vers la base du cœur, entre le tronc pulmonaire et l’atrium gauche. Ensuite, elle se divise en un rameau superficiel et un rameau profond (Fig.6). Au dessous de l’auricule gauche, la branche superficielle entre dans le sillon coronaire et le parcourt jusqu’à atteindre la face atriale. La branche profonde, plus épaisse, pénètre dans le septum inter-ventriculaire où elle émet des branches secondaires en direction de la paroi du ventricule gauche. Ces branches, tout comme celle du rameau superficiel s’anastomosent au niveau de la face atriale avec des branches secondaires provenant du rameau profond de l’artère coronaire droite. Ainsi, l’artère coronaire gauche irrigue la paroi de l’atrium gauche et une partie de la paroi de l’atrium droit, une partie du septum et une partie des ventricules droit et gauche (NICKEL,
et al., 1977).
Figure 6 - Irrigation cardiaque : Face ventrale (à droite) et face dorsale (à gauche)
b. Veines coronaires
Deux veines présentant une large lumière et plusieurs veines plus petites ramènent le sang de la circulation cardiaque dans l’atrium droit. La v. cordis media est large et remonte dans le sulcus subsinosus jusqu’à l’atrium droit où elle se termine immédiatement en dessous du sillons coronaire. La veine collecte le sang provenant principalement de la branche profonde de l’artère coronaire droite. Il existe une seconde veine cardiaque, la v. cordis magna, qui est large, prend son origine à l’apex du cœur et s’étend le long du ventricule gauche pour pénétrer dans le sillon coronaire où elle se poursuit, sous couvert du tronc pulmonaire, jusqu’à l’atrium droit. Elle entre dans l’atrium droit à l’opposé de l’ostium de la veine cave caudale. Cette veine collecte le sang provenant de régions irriguées à la fois par l’artère coronaire gauche aussi bien que par la branche profonde de l’artère coronaire droite (NICKEL, et al., 1977). La v. cardiaque circonflexe gauche collecte le sang des régions irriguées par la branche superficielle de l’artère coronaire gauche. Chez certaines espèces comme l’autruche, il existe une large v. cardiaque circonflexe droite courant le long du bord droit de l’atrium et qui transporte le sang provenant de la branche superficielle de l’artère coronaire droite. Enfin, il existe de nombreuses autres petites veines (v. cardiaques minimes) qui se projettent directement dans le ventricule droit, les atria ou plus rarement dans le ventricule gauche (BAUMEL, 1993).

Page | 28
B. Vascularisation
1. Réseau artériel
Comme chez tous les vertébrés, au moins six paires d’arcs aortiques apparaissent lors du développement embryonnaire aviaire, récapitulant ainsi l’évolution de leur ancêtre aquatique. Sur ses six arches, seules trois persistent chez l’adulte et correspondent à l’artère carotide (3ème arc), l’aorte (4ème arc) et l’artère pulmonaire (6ème arc) (SMITH, et al., 2000). Chez les oiseaux, comme chez les Mammifères, on différencie la circulation pulmonaire, ou petite circulation qui conduit le sang veineux du ventricule droit à l’atrium gauche en empruntant le tronc pulmonaire et la circulation systémique ou grande circulation qui conduit le sang artériel du ventricule droit à l’atrium gauche en passant par l’aorte (NICKEL, et al., 1977).
a. Circulation artérielle pulmonaire
Le tronc pulmonaire émerge à partir de la face auriculaire du ventricule droit et est flanqué par les auricules. Le tronc pulmonaire est court et se divise en deux artères pulmonaires. L’artère pulmonaire gauche croise la veine cave crâniale gauche dans une courbe convexe crânialement et entre ventralement dans le poumon gauche, accompagné de la bronche primaire et de la veine pulmonaire. Au sein du poumon, ses rameaux suivent les subdivisions des bronches pour se terminer au sein du réseau capillaire respiratoire situé dans la paroi des parabronches. L’artère pulmonaire droite croise l’aorte, qui se situe à sa droite, pour atteindre le hile du poumon droit. Son trajet ultérieur est similaire à celui de l’artère pulmonaire gauche (NICKEL, et al., 1977).
b. Circulation artérielle systémique
- L’aorte
Comme nous l’avons déjà mentionné, l’aorte dérive du 4ème arc artériel. Toutefois, alors que chez les vertébrés terrestre (hormis les oiseaux et les mammifères), les branches droite et gauche de ce 4ème arc sont conservées et que seul la branche gauche persiste chez les Mammifères, on observe chez les oiseaux la rémanence de la branche droite uniquement. Néanmoins, chez certaines espèces aviaires l’arc aortique gauche peut subsister soit sous la forme d’un solide cordon cellulaire, soit sous la forme d’un vaisseau persistant et fonctionnel malgré son détachement de la crosse de l’aorte. Cette dernière situation est celle du martin pêcheur d’Amérique (Ceryle alcyon) (GLENNY, 1940). La persistance de la branche droite chez les oiseaux pourrait simplement être la conséquence des caractéristiques hémodynamiques de la chambre de chasse du ventricule gauche propres aux oiseaux (SMITH, et al., 2000) (STEPHAN, 1949). L’aorte émerge sous le nom d’aorte ascendante à partir du ventricule gauche (Fig.7). A la base du cœur, elle est entourée par les atria droit et gauche, l’auricule droit et le tronc pulmonaire. A proximité de son origine, elle présente un renflement ; le bulbe aortique (NICKEL, et al., 1977) qui correspond aux trois sinus aortiques situés au dessus des valvules semi-lunaire (BEAUTHIER, et al., 1993). C’est au niveau de ce renflement, à hauteur des valves sigmoïdes que sont émises les artères coronaires droite et gauche. L’aorte s’incurve ensuite vers la droite en formant l’arc aortique et émet deux troncs brachiocéphaliques très larges. En conséquence, son diamètre diminue considérablement, et elle prend le nom d’aorte descendante. Elle parcourt un trajet passant entre le poumon droit et l’œsophage pour atteindre la colonne vertébrale à hauteur de la 4ème ou 5ème vertèbre thoracique environ. Entre les deux poumons, et accompagnée dorsalement par l’œsophage, elle devient l’aorte thoracique puis l’aorte abdominale. Lors de son trajet elle chemine ventralement au rachis et émet des artères paires et impaires de diamètre

Page | 29
variable. Elle se termine à la surface ventrale des vertèbres coccygiennes par la mince a.sacralis media (NICKEL, et al., 1977).
- Irrigation des organes crâniaux au cœur : Les troncs brachiocéphaliques
L’arc aortique s’incurve vers la droite et donne naissance quasiment immédiatement au tronc brachiocéphalique gauche puis immédiatement derrière au tronc brachiocéphalique droit. Ces deux troncs brachiocéphaliques sont très larges (O'MALLEY, 2005) ce qui explique que ces vaisseaux soient souvent vus jusqu’à leur extrémité sur les radiographies ventrodorsale des grands oiseaux (SMITH, et al., 1997). Plus précisément, le diamètre à la base des troncs brachiocéphaliques est généralement égal voire supérieur à celui de l’aorte (KRAUTWALD-JUNGHANNS, et al., 2004). Ceci reflète le débit sanguin plus important dans les vaisseaux brachiocéphaliques que dans l’aorte descendante (SMITH, et al., 2000). En effet, ces troncs permettent la distribution de 75% du débit sanguin aux ailes, muscles du vol (pectoraux) pectoraux et à la tête (AKESTER, 1971) (WEST, et al., 1981). Ces deux artères s’incurvent légèrement en direction de leur côté respectif du corps et croise sur leur trajet les bronches primaires avant de rejoindre l’articulation scapulaire (NICKEL, et al., 1977). Ces vaisseaux donne rapidement naissance aux :
- artères subclavières gauche et droite (Fig.7), qui irriguent les ailes via l’artère axillaire et les muscles du vol via le tronc pectoral (ou artère thoracique externe (BAUMEL, 1993)) (O'MALLEY, 2005).
- artères carotides communes (Fig.7) qui irriguent la tête et le cou. Il existe de grandes variations concernant le ‘’schéma’’ carotidien à proximité du cœur. L’organisation la plus fréquente, par exemple chez la poule, correspond à l’existence de deux vaisseaux de taille égale dont les trajets sont parallèles. En dehors de cet arrangement classique, on peut également retrouver : une seule artère résultant de la fusion des deux carotides (hérons, butors, martin-pêcheurs), une seule artère résultant de la disparition de l’artère carotide droite (passereaux) ou gauche (pluviers) ou deux artères de tailles inégales (flamands roses, cacatoès à huppe jaune, canard (SZABUNIEWICZ, et al., 1974)) (SMITH, et al., 2000). Les artères carotides s’étendent le long d’un sillon ostéo-musculaire (Canalis caroticus cercicalis) situé ventralement à la base des vertèbres cervicales, à proximité de leur axe de rotation. Ceci empêche que les mouvements du cou, qui est très flexible, n’obstrue la lumière vasculaire par compression et arrêtent le flux sanguin en direction du cerveau (KING, et al., 1984) (ROSENTHAL, et al., 1997) (SMITH, et al., 1997). Un autre système de sécurité permet de prévenir l’arrêt de l’irrigation cérébrale et consiste en l’existence d’anastomoses inter-carotidiennes. Ces anastomoses sont retrouvées d’une part entre les carotides et les artères vertébrales et d’autre part entre les artères carotides à la base du cerveau.par des jonctions en X, en I ou en H. Les anastomose inter-carotidiennes sont retrouvées chez tous les oiseaux à l’exception des espèces appartenant au sous-ordre des Tyranni (BAUMEL, et al., 1968). Les oiseaux ne possèdent pas de cercle artériel de Willis comparable à celui des Mammifères mais, étant donné la taille importante des anastomose intercarotidienne dans la majorité des cas, celles-ci assurent une circulation collatérale plus efficace que le cercle artériel des mammifères (SMITH, et al., 2000). Les anastomoses inter-carotides émettent les principales artères de l’hypophyse (BAUMEL, 1993).

Page | 30
- L’aorte descendante et l’irrigation des organes caudaux au cœur
Tout au long de son trajet, l’aorte descendante émet des ramifications paires et impaires permettant l’irrigation des organes abdominaux et des membres pelviens. Ainsi, elle délègue : une artère cœliaque irriguant le foie, la rate, le proventricule succenturié (estomac glandulaire), le ventricule (ou gésier, estomac musculaire), les intestins et le pancréas, une artère mésentérique crâniale irriguant la plus grande portion de l’intestin et le pancréas, des artères rénales irriguant la portion antérieure des reins et les testicules, les artères iliaques externes qui irriguent les muscles abdominaux ventraux (a. pubiennes) et les membres pelviens (a. fémorales & a. fémorales crâniales), des artères ischiatiques irriguant les parties moyennes (a. rénale médiale) et postérieures (a. rénale caudale) des reins, les membres postérieures (a. fémorale caudale, a. poplité, a. tibiales antérieure et postérieure (BOTTE, et al., 1982)) et la partie utérine de l’oviducte, une artère mésentérique caudale irriguant le rectum et le cloaque, des artères iliaques internes irriguant la paroi pelvienne et l’oviducte, l’artère mésentérique caudale irriguant le côlon et s’anastomosant avec l’a. mésentérique crâniale et une artère caudale irriguant la queue (NICKEL, et al., 1977). Nous noterons que trois paires d’artères assurent l’irrigation du rein chez les oiseaux : une paire provenant de l’aorte et deux paires provenant des artères ischiatiques. Toutefois, chez le héron cendré (Ardea cinerea), une paire d’artère rénale provient des artères fémorale plutôt que des artères ischiatiques (SMITH, et al., 2000).
2. Réseau veineux
- Les veines caves crâniales et le drainage des organes crâniaux au cœur
Chez les oiseaux, au contraire de ce que l’on observe chez les mammifères, il existe deux veines caves crâniales (NICKEL, et al., 1977). Ces veines reçoivent le sang en provenance de l’aile (v. axillaire) et de la poitrine (tronc veineux pectoral) via les veines subclavières et en provenance du cou et de la tête via les veines jugulaires (O'MALLEY, 2005). Les veines jugulaires circulent superficiellement sous la peau du cou, de part et d’autre de la trachée et parallèlement aux nerfs vagues et à l’a. comes vagi (NICKEL, et al., 1977). A l’angle de la mâchoire inférieure, une anastomose transverse entre ces veines jugulaires autorisent le passage du sang d’un côté à l’autre lorsque l’une de ces deux veines est comprimée à l’occasion d’un mouvement du cou. Cette anastomose est inclinée caudalement en direction de la veine jugulaire droite et ceci pourrait expliquer pourquoi, dans la plupart des espèces, la veine jugulaire droite est plus grosse que la gauche (BAUMEL, 1993). Chez certains passereaux, la veine jugulaire gauche est complètement absente (O'MALLEY, 2005).
- La veine cave caudale et le drainage des organes caudaux au cœur
La veine cave caudale est un tronc vasculaire court et large drainant le sang en provenance des organes caudaux. Cette veine n’est pas parallèle à la colonne vertébrale mais circule en direction cranio-ventrale pour atteindre l’atrium droit du cœur (BAUMEL, 1993). Elle provient de la fusion des deux veines iliaques communes (NICKEL, et al., 1977) drainant le sang des organes de la partie inférieure du corps, de la partie inférieure de l’intestin et des membres pelviens. Elle collecte ensuite lors de son trajet vers le cœur le sang en provenance du tractus gastro-intestinal, de la rate et du pancréas après son passage par la veine porte et le foie via les veines hépatiques droite, gauche et intermédiaire (O'MALLEY, 2005) ainsi que des surrénales et des ovaires via les veines surrénales et ovariques (NICKEL, et al., 1977). La large veine mésentérique caudale, également appelée veine mésentérique coccygienne est une particularité de la classe des oiseaux. Celle-ci draine le mésentère de la partie inférieure des intestins et connecte la veine porte hépatique à la veine porte rénale (O'MALLEY, 2005).
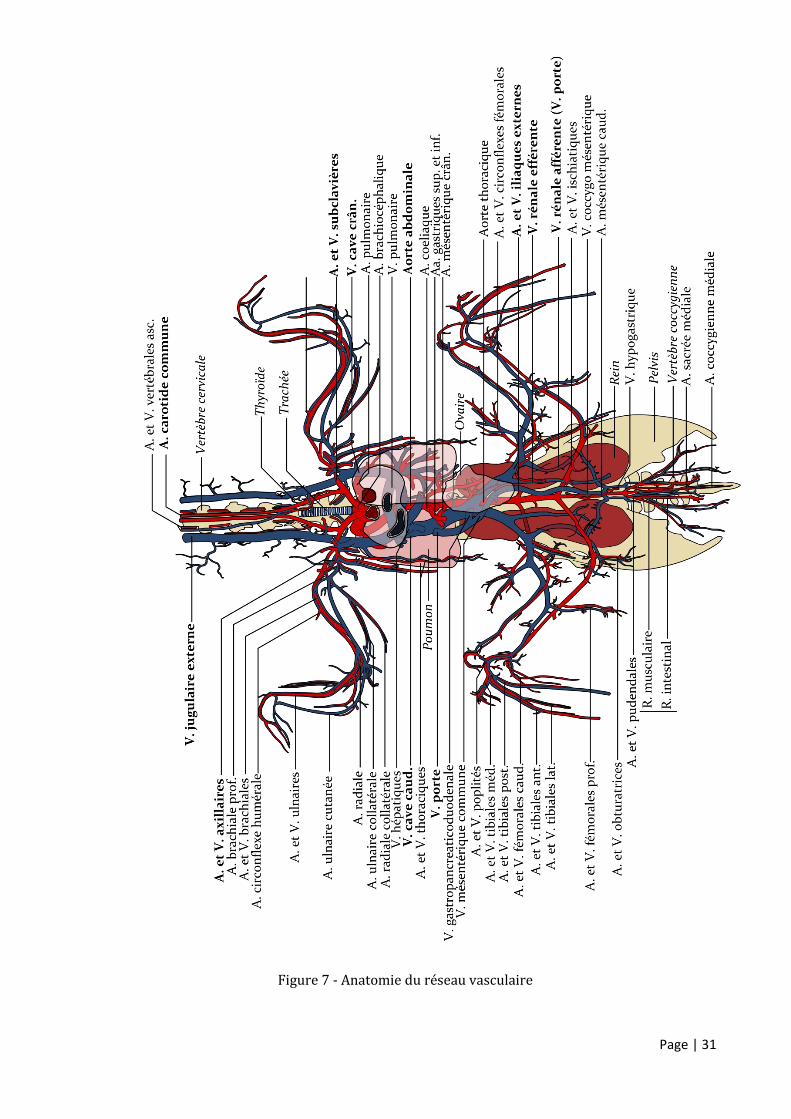
Page | 31
Figure 7 - Anatomie du réseau vasculaire

Page | 32
3. Singularités du réseau vasculaire aviaire
Classiquement, les réseaux capillaires sont compris entre une artère et une veine. Toutefois, il existe des exceptions appelées « réseaux admirables » qui correspondent à un lit capillaire situé entre deux vaisseaux de même nature (deux veines ou deux artères).
a. Les systèmes portes
Les réseaux admirables constituent le fondement des systèmes portes, c'est-à-dire un vaisseau compris entre deux réseaux capillaires (dont l’un est admirable). Les systèmes portes à basse pression sont dits veineux (système porte hypothalamo-hypophysaire, système porte entéro-hépatique, système porte rénal) et ceux à haute pression sont dits artériels (système porte glomérulaire). Les systèmes portes hypophysaire et glomérulaire aviaires étant comparables à ceux des mammifères, nous ne les détaillerons pas.
- Le système porte entéro-hépatique
La veine porte hépatique est le vaisseau fonctionnel du foie, et en tant que telle, collecte le sang provenant du canal gastro-intestinal pour le conduire jusqu’au foie. La confluence des veines suivantes donne naissance au tronc commun de la veine porte. La veine coccygomésenterique qui draine les veines du cloaque ainsi que du côlon, la veine mésentérique inférieure drainant le jéjunum, l’iléum et les caeca et la veine gastro-pancréatico-duodénale, drainant les caeca, l’iléum, le pancréas, le duodenum, la partie droite du ventricule et la veine proventriculo-liénale drainant la rate, le proventricule et la partie gauche du ventricule. La veine porte hépatique draine le sang de ces veines puis bifurque : l’une de ses branches entre dans le lobe hépatique droit alors que les autres pénètrent dans le lobe hépatique gauche (NICKEL, et al., 1977).
- Le système porte rénal
Comme les reptiles, les oiseaux ont un système porte-rénal (ou vascularisation veineuse afférente) (BOUSSARIE, et al., 2010) qui est supporté par l’existence des veines rénales afférentes (ou veines portes rénales) (BAUMEL, 1993). Ces veines sont anastomosées caudalement et forment ainsi un demi-cercle appelé « arc hypogastrique » dans lequel se déverse le contenu des veines iliaques externes, des veines iliaques internes (ou veines hypogastriques), des veines ischiatiques, ovariques, infundibulaire et testicultaires (HOLZ, et al., 2006) (NICKEL, et al., 1977). Cet arc est également en communication avec le sinus veineux vertébral interne et la veine mésentérique caudale mais la direction du flux sanguin dans ces segments est variable. Enfin les branches rénales afférentes crâniales et caudales quittent cette anneau et pénètrent dans le parenchyme rénal ou elles deviennent des veines interlobulaires, qui sont drainées dans les réseaux capillaires péritubulaires. De ce point, le sang entre dans les veines intralobulaires puis les veines efférentes rénales. Sur ces veines, abouchent les veines rénales afférentes pour former les veines iliaques communes qui se rejoignent enfin pour former la veine cave caudale (HOLZ, et al., 2006). L’abouchement de la veine rénale afférente dans la veine rénale efférente crâniale est protégé par une valve musculaire en forme d’entonnoir régulant l’hémodynamique dans le système porte (NICKEL, et al., 1977).

Page | 33
b. Rete mirabile
L’intrication d’un réseau admirable veineux avec un réseau admirable artériel dont les circulations sont à contre-courant porte le nom de « rete mirabile » ou système admirable. Les rete mirabile sont bien décrits chez les oiseaux et jouent un rôle particulier dans la thermorégulation. Nous en mentionnerons quelques uns. Le système admirable temporal (ou ophtalmique) du canard permet de refroidir le sang arrivant au cerveau d’une part et de limiter les pertes de chaleur par les yeux dont la surface est grande chez les oiseaux (BAUMEL, 1993). Le système admirable tibio-tarsal correspond à une réticulation de l’artère poplité localisée sur le bord cranial du tibiotarse et formant un système à contre-courant (CARUS, 1838) permettant de réchauffer le sang à basse température provenant des extrémités chez de nombreuses espèces d’oiseau dont le manchot (SMITH, et al., 1997). Des systèmes équivalents sont retrouvés à la base des pieds, des nageoires et de la queue (DAVENPORT, 1992). Le patch incubateur est retrouvé sous la peau du ventre et fournit du sang en abondance à cette zone glabre destinée à l’incubation des œufs (CARUS, 1838). Enfin, les réseaux faciaux sont des régions cutanées très vascularisées situées en regard de zones de peau glabre. Ces systèmes dériveraient de zones impliquées dans la thermorégulation et dont le rôle actuelle serait davantage lié à la communication inter-individuelle (NEGRO, et al., 2006).
c. Tissus érectiles
Les systèmes admirables peuvent être retrouvés dans des structures associant du tissu conjonctif élastique ainsi que des cellules musculaires et forment ainsi un tissu érectile (NEGRO, et al., 2006). Le plexus artério-veineux sous-cutané en collier est une caractéristique des columbiformes. Il s’agit d’un tissu constitué de veines érectiles sous-cutanées qui s’étend du crâne à la base du cou. Ce plexus est plus développé chez le mâle que chez la femelle (SARABIA, et al., 2008) et serait utilisé à des fins de marquage territorial et sexuel. Il permettrait également de réguler la température corporel en association avec le flutter gulaire (oscillations rapides du plancher buccal et du pharynx) et aux pulsations de l’œsophage supérieur (BAUMEL, et al., 1983). Des lésions du plexus ou des injections réalisées à l’intérieur, particulièrement lorsque celui-ci est développé ou lors de température élevée, peuvent provoquer des hémorragies fatales (VOGEL, et al., 1994) (KALETA, et al., 1989). Des sinus paralinguaux caverneux, larges et bilatéraux s’excavant d’une fosse osseuse médiale de la mandibule ont été identifiés chez les flamands roses. Leur fonction n’a pas été clairement établie. Toutefois, ils pourraient jouer un rôle mécanique lors de la nutrition des flamants, chez qui un système de pompe lingual spécifique est impliqué (HOLLIDAY, et al., 2006). Les crêtes jouent un rôle de communication inter-individuelle chez la dinde (NEGRO, et al., 2006).
d. Le pecten oculi
Le pecten oculi est une structure intraoculaire pigmentée et vasculaire qui se projette à partir du disque optique dans l’humeur vitrée. Elle correspond à un grand conglomérat de vaisseaux sanguins pectinaux de calibre variable, bien ordonné et renforcé par un grand contingent de mélanosomes. Son rôle physiologique est sujet à débat. Les fonctions plausible associées à cette structure serait une augmentation de la sensibilité aux petits objets en mouvement, une régulation de la pression intraoculaire, une réduction de l’éblouissement, une régulation du pH intraoculaire, une stabilisation du corps vitré, un capteur magnétique basé sur l’effet d’orientation grâce au champs magnétique, une hémato-rétinienne ou encore un apport nutritionnel supplémentaire à la rétine. Morphologiquement, le pecten oculi a été classé en trois types : le type « conique » rapporté chez le kiwi, le type « à ailettes » chez l’autruche et le type « plissé » chez la plupart des autres oiseaux (KIAMA, et al., 1998).

Page | 34
II. Histologie
A. La paroi cardiaque
1. Endocarde
L’endocarde est la couche la plus interne du cœur et tapisse complètement la surface des cavités cardiaque atriales et ventriculaires (Fig. 5). Il est donc en contact direct avec le contenu luminal (NICKEL, et al., 1977). Il est constitué d’un tissu conjonctif lâche contenant une grande quantité de fibres élastiques, bordés par une unique couche de cellules squameuses endothéliales (LICHTENBERGER, et al., 2011). Chez les passereaux et les cigognes, le septum inter-atrial est très fin : En certains endroits, il n’est constitué que de deux couches d’endocarde apposées (SMITH, et al., 2000).
2. Myocarde
Le myocarde est la couche musculaire du cœur. Il est située en position intermédiaire. Le myocarde atrial est plus fin que celui des ventricules (SMITH, et al., 2000) et l’épaisseur de la couche myocardique varie en fonction des performances de travail requises (KING, et al., 1984).
a. Myocytes
Les myocytes myocardique aviaires sont comme chez les Mammifères, des fibres musculaires striées et possèdent quelques petites jonction GAP au niveau des disques intercalaires (SCOTT, 1971). Cependant, elles en diffèrent par trois principales caractéristiques concernant la taille ou la structure interne de ces cellules (SOMMER, et al., 1969) (SOMMER, et al., 1970) (HIRAKOW, 1970) :
1. Les fibres myocardiques aviaires sont composées d’un nombre pouvant atteindre 6 cellules (SCOTT, 1971) qui ont un diamètre de 1 à 10 µm (KING, et al., 1984) avec une moyenne de 8um (SCOTT, 1971) ce qui est cinq à dix fois plus petit que celui des cellules musculaires cardiaques mammaliennes (RANDALL, et al., 1996). Ceci signifie que qu’il y a davantage de cellules musculaires dans un volume donné chez les oiseaux que chez les mammifères (SMITH, et al., 2000). Le petit diamètre des fibres musculaires cardiaques serait un facteur permettant d’expliquer que le débit cardiaque aviaire est supérieur à celui des mammifères (PEES, et al., 2009). 2. Contrairement aux Mammifères, le système des tubule T n’existe pas chez les oiseaux. Dans les myocytes mammaliens, le système des tubules T, également appelés tubules transverses correspond à des invaginations tubulaires régulières du sarcolemme perpendiculaires au grand axe de la fibre musculaire. Cette organisation permet d’augmenter l’étendue de la surface par rapport au volume des fibres musculaires et ainsi d’augmenter les flux ioniques transmembranaires. Chez les oiseaux, le petit diamètre des cellules cardiaques s’accompagne d’une grande surface par rapport au volume ce qui compense le besoin de tubules internes pour le couplage excitation-contraction. Comme chez les Mammifères toutefois, le reticulum sarcoplasmique est bien développé (KING, et al., 1984). 3. Les bandes striées remarquables au sein des cellules musculaires striées des Mammifères le sont également chez les oiseaux, à l’exception de la bande M. Chez les mammifères, la bande M correspond à une ligne de protéine qui connecte les filaments de myosine adjacents et forme ainsi une bande observable lorsqu’on l’observe au microscope électronique. Cette bande n’est pas présente chez les oiseaux et la conséquence de cette absence sur les propriétés contractiles des cardiomyocytes aviaires n’est pas connue (SMITH, et al., 2000).

Page | 35
b. Fibroblastes
Les fibroblastes cardiaques forment l’une des plus grande population cellulaire, en terme de nombre de cellules dans le cœur. Les fibroblastes sont retrouvés dans tout le myocarde, entourant les myocytes et comblant les vides entre les couches de tissu myocardique. Ils contribuent au développement cardiaque ainsi qu’aux propriétés structurelles (synthèse de matrice extra-cellulaire), biochimiques (communications inter-cellulaires), mécaniques (élasticité et résistance cardiaque) et électriques (isolant) du myocarde. Des conditions pathologiques sont fréquemment associés avec un remodelage myocardique impliquant une fibrose (CAMELLITI, et al., 2005).
c. Polynucléaires neutrophiles
Il est possible de retrouver des groupes compacts de cellules ayant un cytoplasme éosinophilique s’étendant entre les fibres myocardiques. Cette situation correspond à une granulopoïèse extra-médullaire. Il s’agit d’un phénomène fréquemment rencontré chez les jeunes oiseaux. Leur présence ne doit pas être confondue avec une réponse inflammatoire. La multiplication cellulaire dans ces groupes cellulaires est indiqué par la présence de figures de mitose (RANDALL, et al., 1996).
3. Épicarde et péricarde
L’épicarde, situé en position la plus externe est histologiquement similaire à l’endocarde localisé sur la face interne. En effet, il est constitué d’une couche d’épithélium squameux simple recouvrant un tissu conjonctif lâche sous-jacent contenant une grande proportion de fibres élastiques (SMITH, et al., 2000). L’épicarde correspond en réalité au feuillet viscéral du péricarde séreux. Il se réfléchit à la base des gros vaisseaux en délimitant une cavité (la cavité péricardique) forme alors le feuillet pariétal du péricarde séreux. Un troisième élément entre dans la formation du péricarde : il s’agit d’une couche de tissu conjonctive dense fermement attaché à la couche séreuse pariétale et que l’on nomme péricarde fibreux (TADJALLI, et al., 2009).
4. Tissu de conduction
Le système de conduction électrique cardiaque aviaire fait intervenir quatre types de myocytes (LU, et al., 1993) :
- Cellules pacemaker
Les cellules pacemaker (cellules P) sont petites et de forme sphériques. On les observe dans les nœuds sino-atrial et atrio-ventriculaire. Elles ont la propriété de se dépolariser spontanément et de façon répétitive (SMITH, et al., 2000). Elles contiennent un grand noyau et quelques myofibrilles éparses (LU, et al., 1993). Cette faible quantité de myofibrilles les rends plus pâles histologiquement (NABIPOUR, et al., 2008). Le nœud sino-atrial est constitué d’une association de cellules T et P qui sont parfois contenues dans une lame de tissu conjonctif organisé de façon lâche (HILL, et al., 1980). Le nœud atrio-ventriculaire varie d’un mélange de cellule de Purkinje associées à des cellules myocardiques normales à un tissu constitué de fibres spécialisées contenues dans un tissu conjonctif lâche (PARTO, 2012) et est recouvert par un feuillet de tissu conjonctif, ce qui pourrait être relié à la fréquence cardiaque élevée (QAYYUM, et al., 1976).
- Cellules transitionnelles
Les cellules transitionnelles (cellules T) sont fuselées (LU, et al., 1993) beaucoup plus grosses et ont beaucoup moins de myofibrilles que les cellules musculaires cardiaques. Leur structure est intermédiaire entre les cellules musculaires cardiaques et les fibres de Purkinje. Dans le nœud sino-atrial, les cellules T transmettent l’influx électrique des cellules P aux cellules musculaires atriales ce qui permet leur contraction (SMITH, et al., 2000).

Page | 36
- Cellules de Purkinje
Le faisceau atrio-ventriculaire de His, ses branches ainsi que les réseaux de Purkinje péri-artériel et subendocardique sont constitués de cellules de Purkinje (LU, et al., 1993) (KING, et al., 1984). Les cellules de Purkinje périartérielle comportent jusqu’à 6 cellules alors que les cellules sous-endothéliales en contiennent généralement moins (SCOTT, 1971). Ce sont des cellules larges, allongés, en forme de brique contenant peu de myofibrilles (RANDALL, et al., 1996). Elles peuvent atteindre jusqu’à 5 fois le diamètre des fibres myocardiques. Toutefois, de nombreuses cellules de Purkinje contiennent des fibres longitudinales appelées filaments intermédiaires. Ces derniers font partie du cytosquelette et servent à maintenir la forme des cellules lors des contractions myocardiques (SMITH, et al., 2000). Comme le laisse supposer leur diamètre relativement grand, les cellules de Purkinje ont une vitesse de conduction relativement élevée comparée à celle des cellules myocardiques ordinaires. En conséquence, elles jouent un rôle important dans l’intégration des contraction myocardique et permettent l’efficacité optimum de la pompe. Comme chez les Mammifères, on pense que le couplage électrique entre les cellules myocardiques et les cellules de Purkinje se fait au travers de jonctions GAP dans les disques intercalaires, bien que ces derniers semblent être plus petit et plus rares que chez les Mammifères. (KING, et al., 1984). Toutefois, les jonctions GAP sont plus nombreuses et plus grandes que celles des cardiomyocytes classiques aviaires. Le disque intercalaire n’est pas en marche d’escalier chez les oiseaux (SCOTT, 1971).
B. Les valves cardiaques
Dans la valve atrio-ventriculaire droite, une bande distincte du m. sino-spiral fournit la lamina externe, la plus épaisse de la valve (SHANER, 1923). Une couche fibreuse sépare la lamina externe de la lamina interne plus fine, laquelle dérive d’une invagination de la musculature atriale droite. A son bord crânial le plus épais, cette valve contient l’anneau fibreux droit du squelette cardiaque (BAUMEL, 1993). La composante atriale de la valve s’étend vers l’apex du ventricule sur une distance plus courte que le repli ventriculaire. Ainsi, la valve n’est bilaminaire que dans sa partie supérieure et elle est majoritairement constituée de myocarde ventriculaire (SMITH, et al., 2000). Cette organisation contraste de façon remarquable avec la structure fibreuse caractéristique de la valve tricuspide des mammifères (LU, et al., 1993). Etant un prolongement du myocarde ventriculaire, cette valve s’hypertrophie dans les situations provoquant une hypertrophie ventriculaire droite, ce qui conduit au développement d’une insuffisance cardiaque congestive droite d’autant plus rapide chez les oiseaux, par rapport aux Mammifères (OGLESBEE, et al., 1998). La valve atrio-ventriculaire gauche est constituée d’une double couche d’endocarde et d’un tissu fibreux intermédiaire (NICKEL, et al., 1977). Les cuspides antérieures et postérieures sont petites mais la cuspide médiale (ou aortique) est grande et est attachée à la base des valvules gauche et non-coronarienne de la valve aortique par du tissu fibreux. A la différence de ce que l’on observe chez les mammifères, on observe la présence d’un petit cylindre continu de myocarde parcourant cette jonction fibreuse (LU, et al., 1993). La valve pulmonaire est constituée de trois valvules semi-lunaires. Les valvules de la valve aortique sont plus rigides que les pulmonaires et sont entourées par un anneau de tissu fibreux qui est situé à l’intérieur d’un anneau de cellules myocardiques organisées de façon circonférentielles. L’anneau est complété par une arche de cellules cardiaques qui s’étend entre la cuspide coronaire gauche de la valve de sortie aortique et la valvule aortique de la valve atrio-ventriculaire gauche (le cylindre décrit ci-dessus) (LU, et al., 1993). Cette organisation anatomique contraste clairement avec celle du cœur mammalien, au sein duquel il n’y a que du tissu conjonctif et pas de myocarde, entre la portion de l’anneau musculaire contenu dans la paroi aortique et la valve mitrale adjacente (SMITH, et al., 2000).

Page | 37
C. Vaisseaux sanguins
La paroi des vaisseaux sanguins est constituée des mêmes couches histologiques que le cœur : deux couches endothéliales bordant une couche musculo-conjonctive appelée média (Fig.8). La proportion de ces couches varie légèrement d’un segment vasculaire à l’autre.
1. Les artères
a. Organisation générale de la paroi artérielle
La tunique interne (intima) est la même dans toutes les artères et correspond à la couche la plus mince et la plus interne. C’est donc la couche au contact du sang. Elle est constituée d’un endothélium formé d’une seule couche de cellules endothéliales aplaties (BARONE, 1996). L’intima est le siège des lésions primitives d’athérosclérose (SMITH, et al., 2000). La média est plus épaisse que l’intima. Elle est théoriquement constituée de deux limitantes élastiques bordant une couche musculaire. Les deux limitantes correspondent chacune à une lame de tissu conjonctif fibro-élastique : la membrane élastique interne marque la limite entre l’intima et le bord luminal de la media. Elle est considérée comme appartenant à la média chez les oiseaux, alors qu’elle est généralement considérée comme appartenant à l’intima chez les Mammifères (STOUT, et al., 1980). Toutefois, la limite entre média et intima n’est pas toujours évidente car la membrane élastique interne est absente chez certaines espèces (BEAUFRERE, et al., 2011). Cette limitante est fenêtrée ce qui autorise les échanges entre le plasma sanguin et les parties profondes de la média. La membrane élastique externe, localisée à la jonction avec l’adventice est également fenêtrée et autorise le passage d’éléments vasculaires et nerveux en provenance de l’adventice (BARONE, 1996). Entre les deux limitantes on retrouve une couche constituée de cellules musculaires et de fibres élastiques dont l’arrangement et la composition diffère selon le type d’artère considéré :
- Les artères élastiques ont une paroi principalement constituée d’une alternance de couches élastiques et de couches de muscles lisses enchâssés dans un réseau fin de fibres élastiques, de fibres de collagène, et de la matrice extracellulaire (microfibrilles et protéoglycanes) (BEAUFRERE, et al., 2011) (SMITH, et al., 2000) (HILL, et al., 1997). Il existe également des couches de fibroblastes, propres aux oiseaux (TODA, et al., 1984) (GABELLA, 1997). Une couche, ou unité lamellaire est donc constituée d’une combinaison de fibres élastiques, musculaires et de collagène. La répétition d’un grand nombre de ces unités lamellaires forme la plus grande partie de la paroi des artères élastiques chez les pigeons, les poulets et le cygne tuberculé (Cygnus olor). Nous noterons que les unités lamellaires ne forme pas des cylindres complet autour du vaisseau. Cette particularité est d’autant plus marquée que le segment vasculaire est proche du cœur, où une seule lamelle s’étend tout au plus sur un quart de la circonférence du vaisseau (SMITH, et al., 2000). Il n’existe normalement aucun dépôt lipidique ou calcique dans l’intima ou la media de ces vaisseaux (BEAUFRERE, et al., 2013). La présence de cartilage hyalin à la base de l’aorte constitue une particularité anatomo-histologique remarquable. La fonction de ce cartilage pourrait être de stabiliser les valves aortiques et ainsi, d’assurer une fermeture efficace du repli musculaire formant la valve aortique gauche (KRAUTWALD-JUNGHANNS, et al., 2004). Les artères élastiques sont représentées par les artères situées en position ‘’centrale’’ (SMITH, et al., 2000) : l’arc aortique et ses branches les plus grosses, l’aorte thoracique jusqu’au niveau de l’artère cœliaque environ et les portions extra-pulmonaires des artères pulmonaires.

Page | 38
- Les artères musculaires ont une paroi environ trois fois plus fine que les artères élastiques. Elles sont de structure très différentes puisque leur média est essentiellement constituée de muscles lisses organisé de façon circonférentielle associés à de rares fibres élastiques isolées ou bien associées en un réseau à maille lâche (HODGES, 1974). Nous noterons que chez certaines espèces dont la poule et la dinde, la structure de certaines artères musculaires comme l’artère mésentérique crâniale est le siège de variation notable. En effet, on observe l’existence d’une couche de fibres musculaires orientées de façon longitudinale et dont l’épaisseur est sensiblement égale à celle de la couche des fibres concentriques. La signification fonctionnelle de cette couche musculaire externe n’est pas claire mais elle pourrait servir à raccourcir les vaisseaux afin de supporter les modifications de position provoquée par les mouvements intestinaux (BALL, et al., 1963) Les artères musculaires sont situées en position périphérique et n’apparaissent généralement qu’après le second ordre de ramification (SMITH, et al., 2000). Elles sont représentées par toutes les branches de l’aorte abdominale ainsi que la portion caudale de l’aorte elle-même. - Les artères transitionnelles : Chez les oiseaux et dans la plupart des régions de l’arbre artériel, le changement d’une paroi de type élastique à une media de type musculaire est assez brutal et se rencontre habituellement à un site d’embranchement. Il existe une exception à cette règle, l’aorte elle-même. Au sein de l’aorte, les portions strictement élastiques d’une part et strictement musculaire d’autre part sont séparées par un segment vasculaire s’étendant de l’artère cœliaque jusqu’aux artères ischiatiques et dont la structure pariétal est intermédiaire entre ces deux extrêmes. Nous noterons que cette région de transition est asymétrique chez le pigeon et la dinde, dont la paroi musculaire ventrale est épaisse et la paroi musculaire dorsale, fine (SMITH, et al., 2000).
La média peut être le siège de déformation pathologique. On peut ainsi observer des sténoses ou resserrements, une hypertrophie, ou des dilatations aboutissant à la formation d’anévrysmes qui sont des zones de moindres résistances propices à la rupture. La tunique adventice est la tunique la plus externe. Elle est constituée d’un tissu conjonctif peu organisé et dense qui devient plus lâche à la périphérie et se raccorde par la gaine périartérielle à celui qui enveloppe les organes voisins. Elle est irriguée par des vasa vasorum qui ont un rôle nourricier pour l’adventice elle-même et pour la partie externe de la média. Un réseau de nerfs vasomoteurs non myélinisés rejoint les fibres musculaires lisses de la média (BARONE, 1996). Macroscopiquement, les artères aviaires majeures apparaissent blanches en raison de la présence de fibres de collagène dans la tunique adventice (O'MALLEY, 2005). Ces fibres sont responsable de la rigidité des artères qui permet de lutter contre pression sanguine élevée et d’améliorer le débit sanguin. La contrepartie de cette caractéristique est que les oiseaux sont davantage prédisposés au développement d’athérosclérose de l’aorte et des troncs brachiocéphaliques. (ROSENTHAL, et al., 1997). L’aorte thoracique montre l’existence de deux plexus vasa vasorum : un plexus adventice et un plexus intra mural. Les vaisseaux du plexus adventice sont originaires de la bifurcation des deux artères brachiocéphaliques et des quatre artères intercostales. Ces vaisseaux pénètrent dans la média et nourrissent le tiers extérieur de cette couche, alors que les deux tiers les plus internes restent avascularisés. Le segment aortique abdominal est avasculaire (NEUMANN, et al., 1973).

Page | 39
b. Spécialisation pariétale : les corpuscules
Les corpuscules aortico-pulmonaires se situent dans le tronc pulmonaire et les 2 à 3 mm proximaux des artères pulmonaires ainsi que dans l’aorte ascendante et la crosse aortique, mais pas au-delà du point où l’aorte diminue de diamètre (TAHA, et al., 1986). Les corpuscules carotidiens sont situées entre la division terminale du tronc brachiocéphalique et la glande thyroïde et sont en relation proche avec les glandes parathyroïdes et ultimobranchiale ainsi que le nerf vague (BAUMEL, 1993).
Ils correspondent à des structures vascularisées et innervées que la composition et la localisation permettent de classer en deux groupes histologiques (TAHA, et al., 1986) :
- Les groupes intramuraux, non encapsulés, sont fréquemment retrouvées au sein de la média ou de l’adventice et sont de forme très irrégulières. Ils engendrent une interruption voire une absence de l’alternance de couche élastique et de cellules musculaires lisses et leur organisation rappelle celle des régions baroréceptrices mammaliennes. - Les groupes extramuraux, encapsulés par une fine capsule sont moins fréquents que les groupes intramuraux. Ils sont retrouvés à la surface des vaisseaux ou dans le tissu conjonctif séparant les troncs aortique et pulmonaire à l’extérieur de la paroi (TAHA, et al., 1986).
Si la forme, la taille, la localisation et l’encapsulement diffèrent selon leur localisation au sein de la paroi, les corps aortico-pulmonaire et carotidiens sont tous constituées de cellules de type I granulaires majoritairement situées au centre du glomulus et dont le corps comporte un processus évasé allongé (TAHA, et al., 1986) (HODGES, et al., 1975) contenant des vésicules à centre dense liées à la membrane cellulaire compatible avec une fonction chémoréceptrice ou sécrétoire (HODGES, et al., 1975), de cellules de type II (ou de soutien) engainant les cellules de type I et les axones à la manière des cellules de Schwann (TAHA, et al., 1986) et fournissant un support pour les axones afférents ou efférents des cellules de type (HODGES, et al., 1975) , de terminaisons axoniques qui sont retrouvées à proximité des cellules granuleuses, avec lesquelles forment des synapses de type baroréflexe ou afférent (TAHA, et al., 1986) (TAHA, et al., 1983) et de nombreux capillaires sanguins fenêtrés (TAHA, et al., 1986) (HODGES, et al., 1975). Ils joueraient principalement le rôle de chémorécepteur et serait sensible à la concentration en gaz sanguin (BAUMEL, 1993). Ils pourraient également assurer une fonction de barorécepteur ainsi qu’une fonction sécrétoire (TAHA, et al., 1986) (BUTLER, et al., 1982).
2. Les capillaires
Les capillaires sont, comme chez les mammifères, constitués d’une seule couche de cellules endothéliales entourée par une fine membrane basale. Nous nous intéresserons seulement aux capillaires pulmonaires. Chez les oiseaux, ces capillaires sont situés dans un environnement particulier s’accompagnant de répercussions sur le fonctionnement du système cardiovasculaire (WEST, et al., 2006). En effet, afin de répondre aux exigences du vol, le sang transporté dans les vaisseaux doit être oxygéné de la façon la plus efficace possible. Pour se faire, l’irrigation des poumons possède deux particularités singulières : l’existence d’une circulation à contre-courant permettant une extraction maximale d’oxygène et l’existence d’une barrière sang-air très mince (inférieure à 0.1 µm contre 0.33 µm pour le Mammifère ayant la plus fine barrière) accélérant la vitesse de l’échange (WEST, et al., 2006). Ses exigences fonctionnelles sont supportées par la structure singulière des poumons aviaires. En effet,
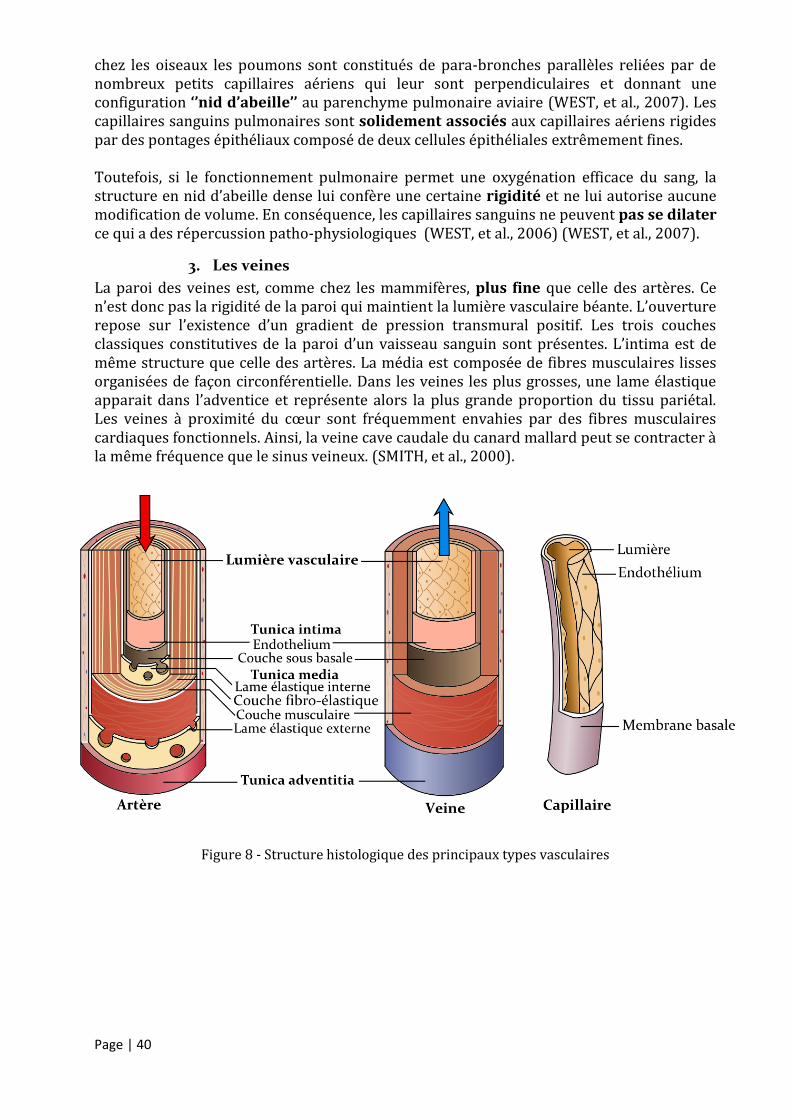
Page | 40
chez les oiseaux les poumons sont constitués de para-bronches parallèles reliées par de nombreux petits capillaires aériens qui leur sont perpendiculaires et donnant une configuration ‘’nid d’abeille’’ au parenchyme pulmonaire aviaire (WEST, et al., 2007). Les capillaires sanguins pulmonaires sont solidement associés aux capillaires aériens rigides par des pontages épithéliaux composé de deux cellules épithéliales extrêmement fines. Toutefois, si le fonctionnement pulmonaire permet une oxygénation efficace du sang, la structure en nid d’abeille dense lui confère une certaine rigidité et ne lui autorise aucune modification de volume. En conséquence, les capillaires sanguins ne peuvent pas se dilater ce qui a des répercussion patho-physiologiques (WEST, et al., 2006) (WEST, et al., 2007).
3. Les veines
La paroi des veines est, comme chez les mammifères, plus fine que celle des artères. Ce n’est donc pas la rigidité de la paroi qui maintient la lumière vasculaire béante. L’ouverture repose sur l’existence d’un gradient de pression transmural positif. Les trois couches classiques constitutives de la paroi d’un vaisseau sanguin sont présentes. L’intima est de même structure que celle des artères. La média est composée de fibres musculaires lisses organisées de façon circonférentielle. Dans les veines les plus grosses, une lame élastique apparait dans l’adventice et représente alors la plus grande proportion du tissu pariétal. Les veines à proximité du cœur sont fréquemment envahies par des fibres musculaires cardiaques fonctionnels. Ainsi, la veine cave caudale du canard mallard peut se contracter à la même fréquence que le sinus veineux. (SMITH, et al., 2000).
Figure 8 - Structure histologique des principaux types vasculaires

Page | 41
III. Physiologie
A. Rôles de l’appareil cardiovasculaire chez les oiseaux
Comme chez les Mammifères, l’appareil cardio-vasculaire joue plusieurs rôles. Leur connaissance permet de comprendre les mécanismes régulant son fonctionnement et les symptômes associés à son dysfonctionnement. Le système cardio-vasculaire permet de réaliser un apport nutritif efficace aux tissus. Les efforts musculaires intenses qu’ont à fournir les oiseaux reposent sur la faculté du système cardio-vasculaire à pouvoir délivrer la quantité suffisante d’oxygène et de nutriments aux organes impliqués. La réalisation de ces efforts nécessite également que le système cardiovasculaire soit capable d’éliminer rapidement les déchets du métabolisme. Chez les oiseaux et les mammifères il est de plus impliqué dans la thermorégulation (SMITH, et al., 2000). En effet, les oiseaux sont des homéothermes et le système cardiovasculaire joue un rôle majeur dans la conservation ou l’élimination de la chaleur. Enfin, chez certaines espèces seulement, le système cardiovasculaire joue un rôle dans la communication inter-individuelle. C’est le cas de certains oiseaux parmi les ordre des struthioniformes, podicipediformes, pelecaniformes, ciconiiformes, falconiformes, galliformes, gruiformes, charadriiformes, columbiformes, psittaciformes, cuculiformes, coraciiformes. Chez ces oiseaux, le plumage laisse apparaître des zones de peau glabre pouvant rougir selon les émotions de l’oiseau (NEGRO, et al., 2006).
B. Physiologie cardiaque
1. Mécanique du cycle cardiaque
a. Hémodynamique intracardiaque
- Diastole ventriculaire
La diastole ventriculaire correspond à la relaxation du myocarde ventriculaire. On distingue deux phases lors de cette diastole. La première phase est la phase de relaxation isovolumétrique et correspond à la chute de pression dans le ventricule sans remplissage. En effet, lorsque le myocarde commence à se relaxer, la pression ventriculaire diminue suffisamment pour provoquer la fermeture des valves aortiques et pulmonaires mais pas suffisamment pour provoquer l’ouverture des valves atrioventriculaires. Lorsque la pression continue de chuter, la pression ventriculaire devient inférieure à la pression atriale et les valves atrio-ventriculaires s’ouvrent. On observe alors la deuxième phase de la diastole ventriculaire qui correspond au remplissage ventriculaire par vidange des atria dans les ventricules. Cette vidange est passive dans un premier temps. En effet les atria accumulent le sang en provenance des veines lorsque les valves atrioventriculaires sont fermées et le gradient de pression provoque leur vidange. Au cours de cette vidange, les pressions atriales et ventriculaires s’équilibrent et provoquent une diminution du flux. On observe alors une phase de remplissage actif correspondant à la contraction des atria. Comme chez les mammifères, ces cavités fonctionnent probablement davantage comme des réservoirs sanguins pour leur ventricule respectif que comme des « remplisseurs » importants. Cette hypothèse est d’autant plus probable que la valve d’Eustache est inexistante chez une grande partie des espèces d’oiseaux de sorte que l’importance de la contraction atriale jouerait un rôle léger dans le remplissage ventriculaire (SMITH, et al., 2000). Les bandes modératrices ventriculaires tendent à limiter une dilatation excessive au cours de la diastole (TADJALLI, et al., 2009).

Page | 42
- Systole ventriculaire
La systole ventriculaire correspond à la contraction du myocarde ventriculaire. On distingue deux phases lors de cette systole. La première phase est la phase de contraction isovolumétrique et correspond à l’augmentation de pression dans les ventricules sans éjection sanguine. En effet, lorsque le myocarde commence à se contracter, la pression ventriculaire augmente suffisamment pour provoquer la fermeture des valves atrioventriculaires (ce qui empêche le reflux de sang dans les atria (NICKEL, et al., 1977)) mais pas suffisamment pour provoquer l’ouverture des valves aortique et pulmonaire. Lorsque la pression continue d’augmenter, la pression intra-ventriculaire devient supérieure à la pression dans l’aorte et le tronc pulmonaire et les valvules sigmoïdes s’ouvrent. On observe alors la deuxième phase de la systole ventriculaire qui correspond à la contraction isotonique (phase d’éjection) par vidange des ventricules dans les gros vaisseaux. Le ventricule droit met sous pression la circulation pulmonaire et le ventricule gauche met sous pression la circulation systémique. Au cours de cette vidange, la pression ventriculaire diminue alors que le volume sanguin à éjecter décroît et ce jusqu’à ce que la pression dans les gros vaisseaux soit supérieure celle des ventricules. On observe alors la fermeture des valves sigmoïdes et un nouveau cycle peut redémarrer. A chaque battement, les ventricules se vident quasiment complètement du sang qu’ils contiennent ce qui est une caractéristique aviaire (STRUNK, 2003).
b. Rôle du péricarde
Souvent considéré comme une simple structure de lubrification et de fixation au sein de la cavité cœlomique, le péricarde joue également un rôle dans le cycle cardiaque. Sa structure le rend relativement non compliant. De ce fait, il est résistant à des augmentations rapide et importante de la taille du cœur, résultant par exemple de la surcharge volumique d’une chambre cardiaque. Cette caractéristique pourrait résulter en un couplage mécanique entre les ventricules via le fluide péricardique de lubrification incompressible qu’il contient. Par exemple, une augmentation de la pression diastolique dans un ventricule pourrait être transmise à l’autre (SMITH, et al., 2000).
c. Débit cardiaque et pression artérielle
Le débit cardiaque correspond à la quantité de sang éjecté par unité de temps. Il correspond donc au volume éjecté lors de chaque contraction cardiaque (appelé volume d’éjection systolique) multiplié par le nombre de contraction dans l’intervalle de temps considéré (appelé fréquence cardiaque) (KING, et al., 1984). Chez les oiseaux, la fréquence cardiaque est plus faible que chez les mammifères de même taille, ce qui améliore la perfusion cardiaque. Elle reste néanmoins très élevée et peut atteindre jusqu’à 1000 battements par minute chez le colibri. De plus, le cœur des oiseaux est proportionnellement plus grand que celui des mammifères de même taille. Par exemple il représente 2,4% du poids corporel chez le colibri. En conséquence, le débit cardiaque (en L/kg/min) a été évalué à environ 7 fois plus important chez une perruche ondulée que chez un homme ou un chien en effort maximum (BRUGER, 1992). Lorsque le sang est éjecté, celui-ci est soumis à des forces de frottement contre les vaisseaux. La résistance à l’écoulement du flux sanguin (appelée résistance périphérique) dépend du diamètre des vaisseaux sanguins et de la viscosité sanguine. Celle-ci est moindre dans la circulation pulmonaire par rapport à celle qui s’exerce dans la circulation systémique. Pour cette raison, le ventricule gauche doit être en mesure de générer une pression systolique 4 à 5 fois supérieure à celle produite par le ventricule droit pour éjecter la même quantité de sang (SMITH, et al., 2000).

Page | 43
2. Electrophysiologie
a. Electrophysiologie cellulaire
- Activité électrique membranaire
Au repos, les cellules sont « polarisées » avec une prédominance de charges positives à l’extérieur et de charges négatives à l’intérieur. La différence de potentiel entre le cytoplasme et l’environnement cellulaire est stable et négatif. On parle de potentiel de repos transmembranaire. Les fibres excitables (neurones, cellules cardiaques) peuvent être stimulées électriquement. Lorsqu’une fibre est stimulée, le potentiel transmembranaire se rapproche de 0. S’il dépasse une valeur seuil, les charges électriques de la cellule sont mises en mouvement et le potentiel transmembranaire varie de façon standardisée. On parle de potentiel d’action. Plus précisément, ces variations se déroulent en trois temps : une première phase dite de dépolarisation (influx de Na+), une seconde phase de repolarisation (efflux de K+) puis une dernière phase dite réfractaire (absolue au départ puis relative) au cours de laquelle la cellule ne peut être excitée.
- Particularités des cellules musculaires cardiaques
Les différents types cellulaires cardiaques décrits au préalable ont des particularités électriques leur conférant un rôle particulier dans le fonctionnement cardiaque. Lorsque les cardiomyocytes classiques sont suffisamment stimulés, un potentiel d’action apparait. Celui-ci présente la particularité que la phase de repolarisation se fait en deux temps entrecoupés par un plateau. L’existence de ce plateau rallonge la période au cours de laquelle la fibre est non stimulable ce qui permet le déroulement de la contraction musculaire sans apparition d’une nouvelle stimulation concomitante. Cela évite que le muscle cardiaque ne se tétanise. Au cours de son trajet, ce potentiel d’action déclenche la libération d’ions calcium par le réticulum sarcoplasmique. Le calcium libéré provoque une modification de la conformation des fibrilles d’actine qui autorise la fixation des têtes de myosine et autorise ainsi la contraction musculaire. Le contrôle du travail cardiaque se fait par la régulation des flux calciques entrant ou sortant du réticulum (LICHTENBERGER, et al., 2011). Les myocytes cardiaques sont des cellules individualisées mais elles se comportent électriquement comme si elle ne faisait qu’une. On parle de syncitium. Cette propriété résulte d’une résistance électrique membranaire faible (existence de canaux) entre les cellules adjacentes. (SMITH, et al., 2000). Nous mentionnerons enfin que la durée des potentiels d’action des cellules ventriculaires est plus longue que celle des cellules atriales (LICHTENBERGER, et al., 2011). Les cellules pacemaker sont caractérisées par un potentiel de repos instable. Celui-ci augmente spontanément suite à la fermeture progressive des canaux de fuite K+ au cours du temps et jusqu’à dépasser la valeur seuil. Cette dépolarisation aboutit à l’émission de potentiels d’actions de façon spontanée et répétée. Selon leur localisation, la fréquence d’émission des potentiel d’action est différente. Ainsi les cellules pacemaker du nœud sino-atrial ont une fréquence de dépolarisation supérieure à celle du nœud atrio-ventriculaire (ZANDVLIET, 2005). Ce sont donc elles qui fixent le rythme cardiaque, subséquemment qualifié de rythme sinusal (KING, et al., 1984). La fréquence de ce rythme est modulé physiologiquement par les catécholamines qui accélèrent la fréquence et l’acétylcholine qui la ralentisse. Le fonctionnement de ce nœud peut par ailleurs être altéré lors de déséquilibre électrolytiques. Pathologiquement, le rythme n’est plus sinusal lorsque le nœud sinusal est déficient ou lorsque d’autres cellules se dépolarisent spontanément plus rapidement que celle du nœud sinusal (SMITH, et al., 2000).

Page | 44
Les cellules de Purkinje conduisent l’influx électrique bien plus rapidement que les myocytes cardiaques. Chez les Mammifères, cette propriété est essentiellement due à l’existence de tubules T dans les cardiomyocytes classiques. Toutefois, chez les oiseaux ce système n’existe pas et l’influx est conduit à grande vitesse du fait que : leur diamètre est beaucoup plus élevé, les cellules sont maintenues circulaires par la présence de filaments intermédiaires, la résistance électrique entre deux cellules est plus faible (jonction GAP plus nombreuses) et possiblement par le fait que les fibres sont finement groupées (SMITH, et al., 2000). Cette grande rapidité leur confère le rôle de cellules conductrices. Pathologiquement la conduction peut être déficiente ce qui entraîne des risques d’arrêt transitoire (syncope) ou permanent (mort subite) de l’activité cardiaque. La durée de dépolarisation des cellules de Purkinje est encore plus longue que celle des cellules ventriculaires, ce qui pourrait prévenir la fibrillation et les extrasystoles ventriculaires chez les espèces aviaires (SMITH, et al., 2000).
b. Trajet de l’influx électrique cardiaque : le système cardionecteur
Toutes les fibres musculaires d’une chambre cardiaque donnée doivent se contracter plus ou moins simultanément alors que, pour assurer un fonctionnement cardiaque normal, il est essentiel que les atria se contractent avant les ventricules (SMITH, et al., 2000) et que l’intervalle entre la contraction de ces deux chambres soit suffisant pour autoriser un remplissage adéquat des ventricules (LICHTENBERGER, et al., 2011). En conséquence, chez les oiseaux comme chez les Mammifères, un tissu de conduction spécialisé est nécessaire pour transporter l’influx électrique des atria aux ventricules, transperçant l’anneau fibreux en un unique point (KING, et al., 1984). A la différence des mammifères, le système de conduction des oiseaux conserve un patron embryonnaire jusqu’à l’âge adulte (ŠOLC, 2007). Ainsi, le système de conduction cardiaque du cœur aviaire, ou système cardionecteur (Fig.10) est constitué par : Le nœud sino-atrial : L’influx électrique est émis physiologiquement par le nœud sino-atrial. L’existence de ce nœud fût l’objet de controverse mais sa présence a été documenté chez le pigeon (DAVIES, 1930), le kiwi, le Gorfou, le poulet (NICKEL, et al., 1977), la poule indienne (PRAKASH, 1956), le moineau domestique, la perdicule rousse gorge, la perruche à collier, les Leghorn, le puffin à bec grêle, le bihoreau gris, le canard, la caille japonaise, les aras, les perruches ondulées, le corbeau à gros bec et la dinde (PARTO, 2010). Sa localisation est variable selon les espèces (SMITH, et al., 2000). La localisation la plus fréquente semble être entre le myocarde de l’atrium droit et l’épicarde, à la base de la valve sinuatrial droite, entre les abouchements de la veine cave craniale droite et de la veine cave caudale (BAUMEL, 1993) (NICKEL, et al., 1977) (poule (LU, et al., 1993), dinde (NABIPOUR, et al., 2008)). Chez la perruche à collier (Psittacula krameri) il est situé vers le côté droit du bord crânial du septum inter-atrial (QAYYUM, et al., 1976) Il peut également se situer dans les valvules et le septum du sinus (autruche) (BAUMEL, 1993) (PARTO, 2010) Il s’étend ensuite en formant des ramification subendocardiques qui assurent un contact avec les cellules myocardiaques ordinaires des parois atriales. La dépolarisation du nœud sino-atrial n’est pas visible sur l’ECG mais engendre une dépolarisation atriale qui elle est visible sous la forme de l’onde P (ZANDVLIET, 2005). Quelque soit sa localisation, le nœud sino-atrial est morphologiquement, et peut-être physiologiquement diffus chez les oiseaux et la région pacemaker électrique primaire semble changer de position spontanément au sein du nœud. Cette dernière caractéristique se traduit par une onde P d’aspect variable et fluctuante au cours du temps chez un même individu (HILL, et al., 1980).

Page | 45
L’existence d’un système de conduction spécialisé au sein des atria des oiseaux est controversé. Des études histologiques prouvent l’existence de fibres de Purkinje réparties diffusément dans le myocarde atrial et des études physiologiques prouvent que l’influx suit leur direction (DAVIES, 1930) (HILL, et al., 1980). D’autre auteurs interprètent ces fibres comme étant les restes d’une formation embryologique concomitante de la formation du système de conduction(SMITH, et al., 2000). Enfin, certains auteurs ont pu mettre en évidence une connexion nerveuse directe entre les nœuds sino-atrial et atrio-ventriculaire et défendent l’hypothèse que l’influx électrique suivrait des fibres musculaires non spécialisés et des fibres nerveuses situées au sein du septum interatrial (QAYYUM, et al., 1976). Chez les vertébrés à sang froid, la musculature des atria est en continuité libre avec celle des ventricules, mais chez les oiseaux (comme chez les Mammifères), des anneaux de tissu fibreux entourent les ouvertures atrio-ventriculaires, aortique et pulmonaire et sépare ainsi les atria des ventricules ce qui permet de prévenir une diffusion généralisée de l’excitation des atria aux ventricules (KING, et al., 1984). Le nœud atrio-ventriculaire : La vague d’excitation qui est initiée au sein du nœud sino-atrial est retardée par le nœud atrio-ventriculaire lors de son trajet vers les ventricules. En effet, alors que l’influx électrique se propage au travers des atria et des ventricules à une vitesse supérieure à 1 m/sec, la vitesse de conduction à travers le nœud atrio-ventriculaire est deux à trois fois plus faible. Ceci autorise les atria à se vidanger avant que la contraction ventriculaire ne débute. (SMITH, et al., 2000). Le nœud atrio-ventriculaire est une structure controversée chez les oiseaux. Des investigations morphologiques ont permis de décrire sa structure chez certaines espèces (par exemple la poule (ŠOLC, 2007), les colombes et les autruches (DAVIES, 1930) (DAVIES, 1930)) mais de nombreuses publications, incluant des analyses morphologiques, histologiques et ultramicroscopiques (SZABO, et al., 1986) ont échoué à le localiser (SANDERS, et al., 1986). En revanche, sa présence a été bien établie suite à la réalisation d’études fonctionnelles (SMITH, et al., 2000). Lorsqu’il peut être identifié, ce nœud est de forme ovale et sa localisation est variable. Il peut être situé dans la partie caudo-dorsale droite du septum inter-ventriculaire (poulet (NICKEL, et al., 1977) (DAVIES, 1930) (LU, et al., 1993), pigeon, cygne noir, moineau domestique (PARTO, 2012), puffin à bec grême, canard mallard, bihoreau gris, caille commune, pigeon biset, marouette brune, gallinule poule d’eau, milan noir, perruche ondulée, capucin domino, corbeau à gros bec, diamant mandarin, ara macao, moineau friquet), dans la jonction atrio-ventriculaire gauche (moineau domestique, poule indienne, bulbul à ventre rouge, moineau domestique, hibou du bengale (PRAKASH, 1956) (SINGH, 1958) (SMITH, et al., 2000)) ou dans la couche endocardique atriale de la valve atrio-ventriculaire droite (autruche (PARTO, 2012)). Le nœud trunco-bulbaire est situé à proximité du bord gauche de la paroi de la racine aortique, sous l’origine de l’artère coronaire gauche (MOORMAN, et al., 1998). L’existence de ce nœud pourrait expliquer la difficulté à identifier un nœud atrio-ventriculaire de structure constante. En effet, des auteurs ont décrits une relation de réciprocité entre la taille des nœuds atrio-ventriculaire et trunco-bulbaire chez différents individus. Plus précisément, lorsque le nœud trunco-bulbaire est large, le nœud atrio-ventriculaire est faiblement développé et vice versa (BAUMEL, 1993). Le nœud trunco-bulbaire s’atténue et forme le faisceau trunco-bulbaire (= branche récurrente du faisceau atrioventriculaire, branche septale (MOORMAN, et al., 1998) ou branche moyenne du faisceau). Ce faisceau se dirige ventralement à travers l’anneau fibreux aortique et tourne caudalement vers la droite dans le myocarde septal. Finalement, il se termine en rejoignant le faisceau atrioventriculaire à son point de bifurcation avec les branches droite et gauche du faisceau (KING, et al., 1984) ou au niveau de la branche gauche du faisceau (NICKEL, et al., 1977). La branche moyenne du faisceau innerve le sphincter musculaire aortique et le rend potentiellement capable de contracter la chambre de chasse du ventricule gauche. Ainsi

Page | 46
innervé, cet anneau musculaire pourrait jouer le rôle de sphincter contrôlant le débit d’éjection du ventricule gauche en modulant les résistance à l’écoulement du flux (LU, et al., 1993) ou pourrait aider à la fermeture de cette valve dont les cuspides sont rigides (SMITH, et al., 2000). L’anneau atrio-ventriculaire droit (ou de Purkinje) est une particularité des oiseaux (KING, et al., 1984). Cette structure relie le nœud atrioventriculaire et le nœud truncobulbaire après avoir encerclé l’orifice atrio-ventriculaire droit. Il forme ainsi une figure en 8 (LU, et al., 1993). Cet anneau passe entre les deux lames myocardiques constituant la valve atrioventriculaire droite (BAUMEL, 1993). Bien que le système de conduction soit isolé par un feuillet de tissu conjonctif lors de son passage dans la valve (VASSALL-ADAMS, 1982) (SZABO, et al., 1986), les preuves actuelles penchent en faveur d’une fermeture active de la valve. Ce mécanisme est alors bien différent de celui observé chez les Mammifères, chez qui les cuspides flottent dans l’orifice et sont déplacées grâce au différentiel de pression généré au cours de la systole (SMITH, et al., 2000). Mentionnons que les pingouins semblent dépourvus d’anneau de Purkinje (ADAMS, 1937) Le faisceau de His et ses branches : Le faisceau de His (= faisceau atrio-ventriculaire) émerge du nœud atrio-ventriculaire et chemine vers l’apex cardiaque en position profonde dans le myocarde du septum inter-ventriculaire. Il perce le tissu fibreux séparant les atria des ventricules puis se divise en branche droite, gauche et moyenne à quelque distance de la crête septale (LU, et al., 1993). Il met ainsi en communication électrique l’étage atrial et l’étage ventriculaire. Généralement, le faisceau atrio-ventriculaire est plus long chez les oiseaux que chez les Mammifères. Toutefois, chez certains oiseaux, cette règle ne se vérifie pas car les branches du faisceau sont émises directement par le nœud atrio-ventriculaire (BAUMEL, 1993). La branche moyenne rebrousse chemin en direction de la racine aortique comme décrit précédemment (LU, et al., 1993). Les branches droite et gauche continuent quant à elles leur descente le long du septum inter-ventriculaire. (KING, et al., 1984). Lors de leur descente, ses branches remontent à la surface pour se positionner en position sous endocardique, de part et d’autre du septum inter-ventriculaire. Ils émettent alors des branches qui diffusent en formant les ramifications sous-endocardiques (BAUMEL, 1993) et se connectent avec le réseau de Purkinje. Peu après son départ, la branche droite émet une ramification parfois appelée branche valvulaire (SMITH, et al., 2000), laquelle remonte jusqu’à la valvule atrio-ventriculaire droite musculaire (NICKEL, et al., 1977). Le réseau de Purkinje : A la différence des carnivores domestiques et de l’homme chez qui le réseau de Purkinje ne pénètre guère plus d’un quart de la paroi musculaire ventriculaire (distribution de type I), les oiseaux ont un système d’activation ventriculaire semblable à celui des ruminants (type II). Ainsi, les fibres de Purkinje pénètrent complètement le myocarde ventriculaire, de l’endocarde à l’épicarde en suivant le trajet des artères coronaires (DAVIES, 1930). En conséquence, l’activation ventriculaire provient à la fois de l’endocarde et de l’épicarde, ce qui est considéré être une adaptation permettant de faciliter le synchronisme des battements à des fréquences élevées (KEENE, et al., 1991). Ce système d’activation à point de départ partiellement sub-épicardique permet d’expliquer la conformation singulière des complexes QRS dits « négatifs » chez les oiseaux (complexes rS et QS majoritaires) (PEES, et al., 2009). Nous signalerons que les bandes modératrices contiennent des fibres de Purkinje et leur permettent ainsi de traverser la cavité cardiaque (TADJALLI, et al., 2009).
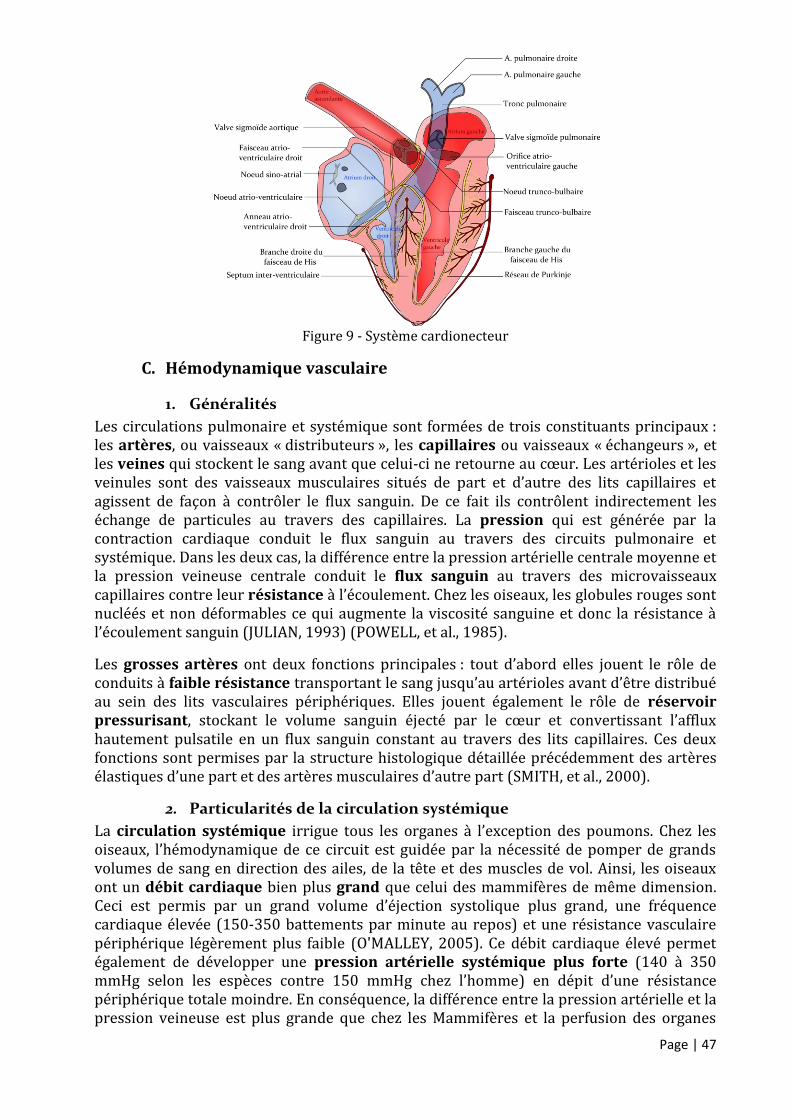
Page | 47
Figure 9 - Système cardionecteur
C. Hémodynamique vasculaire
1. Généralités
Les circulations pulmonaire et systémique sont formées de trois constituants principaux : les artères, ou vaisseaux « distributeurs », les capillaires ou vaisseaux « échangeurs », et les veines qui stockent le sang avant que celui-ci ne retourne au cœur. Les artérioles et les veinules sont des vaisseaux musculaires situés de part et d’autre des lits capillaires et agissent de façon à contrôler le flux sanguin. De ce fait ils contrôlent indirectement les échange de particules au travers des capillaires. La pression qui est générée par la contraction cardiaque conduit le flux sanguin au travers des circuits pulmonaire et systémique. Dans les deux cas, la différence entre la pression artérielle centrale moyenne et la pression veineuse centrale conduit le flux sanguin au travers des microvaisseaux capillaires contre leur résistance à l’écoulement. Chez les oiseaux, les globules rouges sont nucléés et non déformables ce qui augmente la viscosité sanguine et donc la résistance à l’écoulement sanguin (JULIAN, 1993) (POWELL, et al., 1985).
Les grosses artères ont deux fonctions principales : tout d’abord elles jouent le rôle de conduits à faible résistance transportant le sang jusqu’au artérioles avant d’être distribué au sein des lits vasculaires périphériques. Elles jouent également le rôle de réservoir pressurisant, stockant le volume sanguin éjecté par le cœur et convertissant l’afflux hautement pulsatile en un flux sanguin constant au travers des lits capillaires. Ces deux fonctions sont permises par la structure histologique détaillée précédemment des artères élastiques d’une part et des artères musculaires d’autre part (SMITH, et al., 2000).
2. Particularités de la circulation systémique
La circulation systémique irrigue tous les organes à l’exception des poumons. Chez les oiseaux, l’hémodynamique de ce circuit est guidée par la nécessité de pomper de grands volumes de sang en direction des ailes, de la tête et des muscles de vol. Ainsi, les oiseaux ont un débit cardiaque bien plus grand que celui des mammifères de même dimension. Ceci est permis par un grand volume d’éjection systolique plus grand, une fréquence cardiaque élevée (150-350 battements par minute au repos) et une résistance vasculaire périphérique légèrement plus faible (O'MALLEY, 2005). Ce débit cardiaque élevé permet également de développer une pression artérielle systémique plus forte (140 à 350 mmHg selon les espèces contre 150 mmHg chez l’homme) en dépit d’une résistance périphérique totale moindre. En conséquence, la différence entre la pression artérielle et la pression veineuse est plus grande que chez les Mammifères et la perfusion des organes

Page | 48
également. Une perfusion aussi intense permet de répondre à la demande métabolique élevée (KING, et al., 1984). Ces particularités hémodynamiques sont supportées par la rigidité des artères. Celle-ci permettrait d’améliorer le flux sanguin en maintenant une pression artérielle élevée. L’intérêt physiologique d’un niveau élevé de pression artérielle a été mis en relation avec la longueur du cou des oiseaux et à la finesse des carotides (la forte pression serait nécessaire à l’irrigation cérébrale) et avec la structure du rein (BRUGER, 1992). Il existe des différences importantes de pression artérielle systémique selon les espèces, l’âge, le sexe et de ce fait, les valeurs citées n’ont qu’une valeur indicative (BRUGER, 1992). Ce niveau élevé de pression favorise l’apparition d’accidents circulatoires chez les oiseaux tels que des ruptures aortique (BRUGER, 1992) (KRISTA, 1970) ou atriales, des hémorragies ou une insuffisance cardiaque (O'MALLEY, 2005).
3. Particularités de la circulation pulmonaire
Le métabolisme aérobique élevé des oiseaux nécessite une hématose plus efficace que celle des Mammifère. Pour se faire, la structure et le fonctionnement de la circulation pulmonaire présentent quelques singularités. Ainsi, la circulation sanguine se fait à contre-courant du flux aérien chez ces animaux (WEST, et al., 2007). Une telle circulation requiert une ventilation en flux unidirectionel, laquelle est supportée par la juxtaposition de capillaire sanguin et aérien au sein du parenchyme. L’hématose est encore facilitée par la finesse et l’uniformité de la barrière gaz-sang. De telles caractéristiques tendent à rendre les capillaires sanguins pulmonaires très sensibles à une insuffisance de stress (rupture des capillaires lors d’augmentation de la pression sanguine), comme cela est observé chez le cheval, qui a pourtant un interstitium plus développé. Ces complications sont contrecarrées par l’existence de pontages épithéliaux orientés aléatoirement entre les capillaires ainsi que par les faibles variations de volume de ce poumon, qui limitent la distension des vaisseaux (WEST, et al., 2010).
En contrepartie, cette invariabilité du diamètre des capillaires se manifeste par une constance de la résistance périphérique pulmonaire (Rp). La résistance vasculaire totale dans la circulation pulmonaire est moindre que celle de la circulation systémique. La pression artérielle pulmonaire (Pa) étant telle que Pa = Pv + Q x Rp, et le débit (Q) devant être le même dans les circulations pulmonaire et systémique, il apparaît clairement que la circulation pulmonaire se fait à moindre pression. Cette formule montre également que toute augmentation du débit cardiaque Q, provoquera une hypertension pulmonaire. En réalité, la situation est pire puisque R augmente lorsque le débit est très élevé, sans doute suite à la création d’un œdème pulmonaire ou à la création de turbulence à l’écoulement du sang (WEST, et al., 2007). Enfin une augmentation de la pression veineuse (Pv) (insuffisance cardiaque gauche) se répercutera invariablement comme une surcharge de travail du ventricule droit. Cela explique que le ventricule droit soit fréquemment l’objet d’hypertrophie compensatrice (OGLESBEE, et al., 1998).
4. Particularité de la circulation coronaire
Une partie du sang oxygéné provenant du ventricule gauche entre dans les sinus aortiques droit et gauche d’où émergent les artères coronaires droite et gauche respectivement. Comme chez les Mammifères, la perfusion cardiaque se déroule essentiellement au cours de la diastole, quand la lumière vasculaire est béante. La perfusion du muscle cardiaque est plus élevée que ne l’est celle des muscles squelettiques ou tout autre tissu et varie en fonction de la concentration sanguine en O2 et en CO2. La diminution de l’oxygénation sanguine et l’augmentation de la demande en oxygène du myocarde provoquent une hypoxie engendrant une augmentation du flux sanguin coronaire par diminution de la résistance vasculaire. De la même façon, une hypocapnie provoque une augmentation du débit coronaire, ce qui est différent de la dynamique du

Page | 49
flux sanguin coronaire chez les Mammifères, pour qui l’hypocapnie provoque une diminution du flux sanguin. Quand les oiseaux volent à une altitude élevée, la réduction de l’oxygène résulte en une hypoxémie et une augmentation compensatoire de la ventilation. En conséquence, on observe une alcalose sanguine et une hypocapnie. Ainsi lors de vol à haute altitude, la perfusion coronaire augmente (SMITH, et al., 2000).
5. Particularité du système porte-rénal
Les oiseaux possèdent une vascularisation telle que le sang provenant des membres pelviens, de la queue et du tractus digestif inférieur peut rejoindre le cœur selon différentes modalités. Plus précisément, le circuit emprunté par le sang est déterminé par la résistance à l’écoulement sanguin des différents vaisseaux impliqués et notamment par l’existence des valves portales. Ces valves sont situées à la jonction entre la veine iliaque commune et la veine porte rénale crâniale (SMITH, et al., 2000). Elles sont constituées de muscles lisses (O'MALLEY, 2005) et sont innervées à la fois par les systèmes ortho- et parasympathique. Dès lors, une stimulation cholinergique provoque une fermeture des valves et une stimulation adrénergique provoque à l’inverse un relâchement des muscles lisses valvulaires et l’ouverture de la valve (SMITH, et al., 2000) (KING, et al., 1984).
Lorsque les valves sont fermées, le sang emprunte soit la veine porte rénale crâniale, soit la veine porte rénale caudale. Une partie du sang portal rejoint les sinus sanguins péritubulaires où il se mélange avec le sang post-glomérulaire. Il participe alors à la sécrétion et à la réabsorption tubulaire puis est drainé jusqu’au cœur par les veines centrolobulaire qui déversent leur contenu dans les veines rénales efférentes et la veine cave. Toutefois, l’intégralité du sang portal ne participe pas à ces échanges tubulaires. Ainsi, une partie du sang empruntant la veine porte rénale crâniale rejoint le cœur via les sinus veineux vertébraux internes et une partie du sang empruntant la veine porte rénale caudale rejoint le cœur via ces mêmes sinus ou via la veine mésentérique caudale qui rejoint le système porte hépatique (SMITH, et al., 2000). Les valves sont notamment fermées lors du vol, ce qui intensifie la circulation rénale et l’élimination des déchets métaboliques (O'MALLEY, 2005). Lorsque les valves sont ouvertes, le sang veineux s’écoule par les veines iliaques communes pour rejoindre la veine cave caudale et le cœur. Cette situation s’observe lors de stress et permet de détourner le sang rénal vers le cœur et le cerveau (O'MALLEY, 2005).
Au repos, les valves sont partiellement fermées et le système porte fournirait 50 à 70% du débit sanguin rénal (O'MALLEY, 2005). Ce pourcentage dépend notamment de l’espèce, de l’état de stress et de la température (KENDAL, 2002). Physiologiquement, ce système constituerait un mécanisme compensatoire permettant de maintenir le flux sanguin passant par les reins lors d’hypotension ou de déshydratation. En effet, lors de diminution de la perfusion rénale, les artères afférentes pré-glomérulaires des néphrons de type reptilien se contractent et entrainent une baisse du débit de filtration glomérulaire. Dans ce cas, le flux sanguin rénal est maintenu par l’augmentation du flux sanguin dans le système porte rénal crânial et caudal, assurant ainsi un apport nutritionnel aux cellules de ces néphrons (SMITH, et al., 2000).
En pratique, un médicament injecté dans le membre pelvien pourrait donc théoriquement être filtré par le rein avant d’atteindre sa cible et son effet néphrotoxique serait potentialisé. Il convient donc d’injecter les substances dans la partie avant du corps (bréchet)(BOUSSARIE, et al., 2010). De plus, des cellules tumorales ou des agents infectieux situés dans la moitié inférieure du corps peuvent atteindre le foie via la veine mésentérique caudale et/ou le système nerveux via le plexus veineux vertébral interne (CANNY, 1992).

Page | 50
D. Contrôle des paramètres cardiovasculaires
La perfusion adéquate des tissus dépend du maintien de la pression artérielle dans des limites relativement strictes. Ainsi, au cours du vol, alors que la fréquence cardiaque des pigeons peut augmenter jusqu’à quatre fois et que le débit cardiaque peut atteindre sept fois la valeur au repos, la pression artérielle sanguine moyenne est maintenue proche de sa valeur au repos (BUTLER, et al., 1977). Inversement, chez les oiseaux plongeurs, la fréquence cardiaque peut diminuer jusqu’à 90% et être accompagnée d’une diminution du débit cardiaque. Pour autant, la pression artérielle moyenne reste dans les limites physiologiques normales (BUTLER, et al., 1997). Ainsi, il apparait que les oiseaux ont un système de contrôle de l’appareil cardiovasculaire supportant leur activité dans des situations aussi diverses que le vol ou la plongée (LUCITTI, et al., 2006). La pression artérielle est régulée par divers agents influençant le travail du cœur et/ou le degré de contraction des vaisseaux.
1. Contrôle mécanique
Le tonus vasculaire dans une partie du système circulatoire peut être défini comme le niveau moyen de contraction des fibres musculaires lisses de la paroi des vaisseaux sanguins de la région considérée. A une pression intraluminale constante et en absence d’influence extrinsèque, les cellules musculaires se contractent spontanément à une fréquence intrinsèque. Une augmentation de la pression artérielle provoque une distension de la paroi vasculaire, augmente le diamètre du vaisseau et provoque une augmentation du débit. Toutefois, la distension du vaisseau augmente la fréquence de contraction des cellules musculaires lisses et cette augmentation de la vasomotricité provoque alors une diminution du diamètre vasculaire, restaurant le flux sanguin initial. Le mécanisme inverse se déroule lors de diminution de la pression artérielle. Ces modifications myogénique du diamètre vasculaire constituent un mécanisme pour autoréguler le flux sanguin localement. Il intervient par exemple pour réguler le débit sanguin glomérulaire rénal lors de modification de la pression artérielle (SMITH, et al., 2000).
2. Contrôle nerveux (incluant la réponse surrénalienne)
Présentation : Le cœur est innervé par des fibres sympathiques accélératrices et par des fibres vagales qui ont une influence inhibitrice sur la fréquence cardiaque. (KING, et al., 1984). L’innervation est constituée de nombreux faisceaux nerveux distinct ainsi que de ganglions situés à la périphérie du noeud sinuatrial (NABIPOUR, et al., 2008). Le degré de contrôle vagal sur le cœur varie selon les espèces d’oiseau. Il est possible de classer les oiseaux en deux grandes catégories : Les oiseaux dont le cœur est relativement gros par rapport à leur poids corporel (pigeon, canard, mouette, accipitridés) et chez qui le système vagal a un effet cardio-inhibiteur puissant (SZABUNIEWICZ, et al., 1974) et les oiseaux dont le cœur est relativement petit par rapport à leur poids corporel (poulet, corbeau freux (Corvus frugilegus), choucas des tours (Corvus monedulla)) chez qui le système vagal a peu d’effet cardio-inhibiteur (SZABUNIEWICZ, et al., 1974).
Tous les vaisseaux pulmonaires et systémiques sont innervés par le système nerveux autonome hormis les lits capillaires. Le degré de vasomotricité dépend de la densité locale de terminaisons effectrices et des récepteurs post-jonctionnels de cette région. La vasomotricité adrénergique est prédominante dans les veines (SMITH, et al., 2000). Les terminaisons nerveuses au sein des artères et des artérioles sont souvent rencontrées dans la tunique adventice externe et s’étendent seulement au niveau de la lame élastique externe. Les terminaisons nerveuses au sein des veines et des veinules sont associées avec les fibres de muscle lisse de la tunique media, en plus de l’adventice externe. La densité de l’innervation varie selon les ordres et possiblement, selon les espèces d’oiseaux et peuvent être la conséquence d’un contrôle neurogénique local de la résistance, tout comme l’est le

Page | 51
réflexe de plongée chez les canards. En effet, certaines espèces de canard ont une densité de terminaison nerveuse augmenté au niveau des vaisseaux sanguins du membre pelvien afin de permettre une vasoconstriction périphérique. Ceci permet de conserver le flux sanguin pour le cœur et le cerveau au cours d’un potentiel événement ischémique, ce qui est le cas lors de plongée (SMITH, et al., 2000).
Stimulation : Comme les mammifères, les oiseaux ont un baroréflexe. Les barorécepteurs sont des terminaisons nerveuses spécialisées immiscées dans les parois artérielles et qui déchargent des influx centraux lorsqu’elles sont stimulées mécaniquement. Certains auteurs pensent que le baroréflexe aviaire proviendrait quasiment exclusivement de l’aorte ascendante. Le processus central du baroréflexe serait sous la dépendance de récepteurs NMDA, comme chez les mammifères (LUCITTI, et al., 2006). Lors d’une stimulation de ce réflexe, les catécholamines sont libérées par les cellules chromaffines de la médullo-surrénale dans le torrent sanguin. Par exemple, en situation de stress, les concentrations en catécholamines plasmatiques peuvent être multipliées par mille ce qui est source de désordre cardiovasculaire comme des arythmies (SMITH, et al., 2000).
Action : L’adrénaline et la noradrénaline induisent toutes une augmentation de la fréquence de dépolarisation du pacemaker. Ceci qui provoque une augmentation de la fréquence cardiaque. De plus, elles provoquent une augmentation de la force de contraction. Il a été prouvé que la adrénaline était aussi efficace à cet égard que la noradrénaline, lors d’expériences menées sur des bandes musculaire isolées de muscle cardiaque aviaire. Ceci diffère des cellules musculaires cardiaques mammalienne chez qui l’adrénaline est plus efficace. Chez les oiseaux comme chez les mammifères, il apparaît que les catécholamines agissent principalement sur des récepteurs adrénergiques beta des cellules myocardiaques et des cellules pacemaker. Les catécholamines provoquent une vasoconstriction généralisée associée à une augmentation de la pression sanguine. Les contractions musculaires des veines médiées par le système sympathique entraînent une augmentation du retour sanguin au cœur (SMITH, et al., 2000).
3. Contrôle humoral
a. Facteurs chimiques
Certains facteurs chimiques affectent directement les myocytes. Ceux-ci incluent la p02, la pCO2, l’acide lactique, d’autres déchets métaboliques, les concentrations en électrolytes et le pH. Par exemple, une accumulation d’acide lactique suite à un effort musculaire engendre une chute du pH et une augmentation de la pCO2. Cela provoque une vasodilatation des lits microvasculaires puisque ces éléments chimiques exercent leur effet directement sur l’appareil contractile des myocytes (SMITH, et al., 2000).
b. Agents vasoactifs produits localements
Les recherches suggèrent que l’oxyde nitrique (NO) et l’endothéline libérés localement par les cellules endothéliales altèreraient le tonus des muscles lisses grâce à un mécanisme reposant sur la fixation à des récepteurs. Comme chez les Mammifères, l’oxyde nitrique est relâché suite au clivage enzymatique de la L-arginine par les cellules de l’endothélium vasculaire. Il exerce un pouvoir vasodilatateur puissant sur le muscle lisse vasculaire. L’acétylcholine (Ach) libérée dans la circulation systémique est à l’origine de la libération de NO, ce qui provoque l’effet vasodilatateur. L’endothéline est un puissant vasoconstricteur chez les mammifères mais son effet n’a pas été évalué chez les oiseaux. Néanmoins, les récepteurs ont été observés sur les myocytes cardiaques aviaires et leur activation provoque une augmentation de la force de contraction (SMITH, et al., 2000).

Page | 52
c. Agents vasoactifs à action systémique
o Le système des kinines
Le système des kinines a reçu peu d’attention à ce jour chez les oiseaux. L’ornithokinine (équivalent de la bradykinine mammalienne) aurait des effets vasodilatateurs. Les kininases seraient présentes et inhibées par certains IECA (PREZOTO, et al., 2009)
o Le système arginine-vasotocine (AVT)
Présentation : L’arginine-vasotocine (AVT) est une hormone peptidique de petite taille libérée par la neuro-hypophyse. La concentration circulante d’AVT est d’environ 10pg/ml chez des oiseaux normalement hydratés (MÖHRING, et al., 1980). C’est l’homologue de l’arginine-vasopressine (AVP ou ADH) des mammifères (NEPHEW, et al., 2005). Sécrétion : Le stimulus primaire engendrant une libération supplémentaire d’AVT est une augmentation de l’osmolalité des fluides extracellulaires, comme une déshydratation. Chez plusieurs espèces, la concentration circulante en AVT augmente de 0.25-2 pg/ml pour chaque mmol/kg d’augmentation de l’osmolalité plasmatique (MÖHRING, et al., 1980). Une diminution du volume de fluide extracellulaire (hémorragie) peut également stimuler la libération d’AVT mais de façon moins sensible que la réponse à des modifications d’osmolalité (GOLDSTEIN, et al., 2000). Action : L’AVT permet de retenir l’eau. Pour ce faire, elle provoque une réduction du débit de filtration glomérulaire, probablement secondaire à une vasoconstriction des artérioles afférentes aboutissant à une réduction voire à un arrêt complet de la filtration de certains néphrons. Les néphrons de type reptiliens (auxquels il manque une anse de Henlé et qui sont donc incapables de produire une urine concentrée) sont plus sensibles à l’AVT (GOLDSTEIN, et al., 2000). L’effet cardiaque de l’AVT chez les oiseaux ne fait à ce jour pas consensus certaines études révèlent une absence d’effet (HASSINEN, et al., 1999) alors que d’autres montrent une diminution ou une augmentation de la fréquence cardiaque (NEPHEW, et al., 2005). Certains auteurs concluent à une absence d’effet sur la pression artérielle (HASSINEN, et al., 1999).
o Le système rénine-angiotensine- aldostérone.
Présentation : L’existence d’un système rénine-angiotensine-aldostérone comparable à celui des Mammifère est bien établi chez les oiseaux (GOLDSTEIN, et al., 2000).
Sécrétion : Lors d’hypovolémie ou de déplétion sodée, les cellules de la macula densa sont stimulées et provoquent la libération de rénine par les cellules juxtaglomérulaires. La rénine clive l’antiotensinogène circulant en angiotensine I, laquelle est convertie en angiotensine II. L’angiotensine II stimule la synthèse d’aldostérone. Contrairement aux Mammifères, l’aldostérone n’est pas libérée en réponse à une hyperkaliémie (DONELEY, 2011).
Action : L’angiotensine II provoque une augmentation de la pression sanguine par augmentation de la fréquence cardiaque et vasoconstriction (HASSINEN, et al., 1999). L’angiotensine II inhiberait l’excrétion sodique par les glandes à sel en provoquant une constriction des vaisseaux de ces glandes par stimulation du système sympathique (SMITH, et al., 2000). L’aldostérone provoque une rétention hydro-sodée par les tubules rénaux (DONELEY, 2011).

Page | 53
o Les facteurs natriurétiques :
Présentation : Alors que l’ANP (atrial natriuretic peptid) est estimé être le facteur le plus important chez les Mammifères, ce facteur ne semble pas exister chez les oiseaux (TRAJANOVSKA, et al., 2008) contrairement à ce qui a été cru (GOLDSTEIN, et al., 2000) (BRAUN, 1998). En revanche, le BNP (brain natriuretic peptid) existe et semble être le facteur natriurétique ayant le plus d’effet (TRAJANOVSKA, et al., 2008). D’autres études mentionnent l’existence d’autres facteurs tels que le CNP 1 et 3 et le RNP (GOLDSTEIN, et al., 2000). Sécrétion : Chez les oiseaux, le niveau plasmatique de BNP est augmenté lors d’hypervolémie chronique et diminué lors d’hémorragie ou chez des oiseaux privés d’eau (GRAY, 1994) (GRAY, et al., 1991). Ces particularités en font un bon marqueur de surcharge cardiaque. Toutefois, chez les Mammifères l’hypoxie, l’ischémie, une endotoxémie ou des états oedemateux non cardiogéniques (néphrose, cirrhose) peuvent également stimuler la libération de BNP. Action : Le BNP serait impliqué dans la régulation de la volémie relative (TAKEI, et al., 2011) en agissant à plusieurs niveau. Tout d’abord, le BNP agit vraisemblablement sur le rein puisque des récepteurs pour le BNP ont été localisés sur les glomérules et les tubules rénaux des canards (SCHUTZ, et al., 1992). A ce niveau, il favoriserait la diurèse et la natriurèse (GRAY, 1994) (GRAY, 1994) mais le mécanisme d’action de l’hormone, incluant la contribution relative des effets glomérulaires et tubulaires reste incompris à ce jour (GOLDSTEIN, et al., 2000). Il a également été prouvé que le BNP stimulait de façon significative la sécrétion par les glandes à sel chez le canard (SCHUTZ, et al., 1990) (GRAY, et al., 1991). De plus le BNP provoquerait une vasodilatation (notamment de l’aorte) (TRAJANOVSKA, 2008). L’effet du BNP sur la pression artérielle ne fait pas consensus : certaines études prouvent qu’il provoquerait une chute de pression (TRAJANOVSKA, et al., 2007) alors que d’autre montre qu’il n’aurait pas d’effet (SCHUTZ, et al., 1992). Enfin, le BNP inhibe l’activité des autres systèmes endocriniens tels que le système rénine-angiotensine-aldostérone et Arginine-Vasotocine, qui engendre une rétention hydro-sodée (GRAY, et al., 1997) (GRAY, et al., 1991) mais cela ne semble pas le cas chez toutes les espèces (KOCSIS, et al., 1995). Le BNP a un effet inotrope négatif sur les myocytes cardiaques aviaires (D'SOUZA, et al., 2004). CONCLUSION : Le système cardiovasculaire aviaire présente donc un certain nombre de
différences anatomiques, histologiques et physiologiques substantielles à comparer de
celui des Mammifères. Ces adaptations contribuent à expliquer les capacités physiques des
oiseaux. En contre-partie, certaines de ces adaptations tels que des capillaires enclavés
dans des poumons rigides ou une tunique adventice épaisse et fibreuse favorisent
l’apparition de troubles cardiovasculaires.

Page | 54

Page | 55
DEUXIEME PARTIE : ETIOLOGIE ET PATHOGENIE DES AFFECTIONS CARDIOVASCULAIRES AVIAIRES

Page | 56
Les affections cardiovasculaires sont des maladies rarement diagnostiquées du vivant de l’oiseau. Néanmoins, elles représentent une dominante pathologique chez les oiseaux gardés en captivité et notamment chez les psittaciformes âgés (BOUSSARIE, et al., 2010) dont le manque d’exercice, l’obésité et les déficit nutritionnels représentent des facteurs de risque (KRAUTWALD-JUNGHANNS, et al., 2004). Il est estimé qu’environ 6% des décès chez les oiseaux sont directement imputables à une maladie cardiaque primitive (OGLESBEE, et al., 1998) et que 10 à 40% des oiseaux de compagnie sont atteints d’une maladie cardiovasculaire (DONELEY, 2011). La forte prévalence des lésions cardiaques aggraverait l’état de santé d’oiseaux atteints par ailleurs de maladies affectant d’autres organes (KRAUTWALD-JUNGHANNS, et al., 2004). En somme, les maladies cardiaques chez les oiseaux de compagnie joue un rôle bien plus grand que celui qui leur était incombé par le passé et l’évaluation du système cardiovasculaire devrait être inclus comme faisant partie intégrante de l’examen physique de routine et des tests diagnostics chez les oiseaux (PEES, et al., 2009).
I. Affections cardiaques
A. Insuffisance cardiaque congestive
Définition : L’insuffisance cardiaque se définit comme une incapacité du cœur à fournir, dans des conditions normales, c'est-à-dire sans hypovolémie, le débit sanguin nécessaire aux besoins métaboliques des différents organes (BOMASSI, et al., 2010) ou lorsque cet apport adéquat n’est possible que lors de pressions de remplissage élevées (DE WIT, et al., 2005). Comme chez les mammifères, l’insuffisance cardiaque congestive chez les oiseaux correspond généralement à une maladie cardiaque en stade terminal (STRUNK, 2003). Epidémiologie : L’insuffisance cardiaque congestive serait retrouvée chez 9.7% des psittaciformes (OGLESBEE, et al., 1998). De nombreux rapports documentent l’existence d’insuffisance cardiaque chez de nombreuses espèces d’oiseaux et chez des individus de tous âges. Chez les oiseaux, l’insuffisance cardiaque congestive a été rapportée chez plusieurs espèces comme les poules (JULIAN, 1993) (WILSON, et al., 1988) (FULTON, 2008), un canard (MITCHELL, et al., 2008), des rapaces (BRANDAO, 2013) (RISI, et al., 2011) (SHRUBSOLE-COCKWILL, et al., 2008) (KNAFO, et al., 2011), les mainates (ROSENTHAL, et al., 1993), une talève sultane (BEEHLER, et al., 1980), et des psittaciformes (ISAZA, et al., 1992) (OGLESBEE, et al., 1998) (JUAN-SALLES, et al., 2011) (PEES, et al., 2006) (SEDACCA, et al., 2009) (JESSUP, 1980) (PHALEN, et al., 1996) Etiologie : L’insuffisance cardiaque n’est pas le diagnostic d’une maladie spécifique mais un syndrome provoqué par un ou plusieurs processus sous-jacents (DE WIT, et al., 2005). Les anomalies majeures sous-jacentes à une insuffisance cardiaque congestive sont :
- Une atteinte myocardique : Elle est caractérisée par une faible contractilité ventriculaire et peut être consécutive à une myocardite, un infarctus myocardique, une cardiomyopathie dilatée, une intoxication à la furazolidone (troubles du flux calcique) (KIM, et al., 2000). - Une surcharge volumique : Les maladies induisant une surcharge en volume incluent les affections valvulaires, les anomalies septales ou des gros vaisseaux. (ZANDVLIET, et al., 2001). - Une surcharge en pression : Elle survient lorsque le ventricule doit générer une pression systolique supérieure à la normale pour éjecter le sang. Cette situation est observée lors de fibrose interstitielle pulmonaire ou de phénomènes athéro-sclérotiques.

Page | 57
- Une altération du remplissage : Les maladies limitant le remplissage ventriculaire peuvent diminuer le débit cardiaque et sont principalement reliées à une maladie péricardique constrictive (DE WIT, et al., 2005).
Physiopathologie : Lorsque la pompe cardiaque dysfonctionne on observe une incapacité du cœur à aspirer le sang en amont associé à une incapacité à fouler le sang en aval. L’insuffisance aval correspond à la diminution du débit sanguin d’éjection systolique. Cette baisse du débit est dans un premier temps corrigée par la mise en place de phénomènes compensateurs. Les barorécepteurs détectent la chute de pression et provoquent une augmentation du tonus sympathique qui stimule l’activité cardiaque et augmente le tonus vasculaire (LUMEIJ, et al., 1994). De plus, la rénine est libérée par l’appareil juxta-glomérulaire et déclenche la cascade angiotensine-aldostérone aboutissant à une rétention hydro-sodée et une vasoconstriction. Cette cascade permet in fine de maintenir un débit normal (DONELEY, 2011). Enfin, le myocarde subit des remaniements structurels telle qu’une hypertrophie excentrique avec dilatation ou une hypertrophie concentrique afin de compenser respectivement une surcharge en volume ou en pression. La phase pendant laquelle la cardiopathie n’entraîne pas de chute du débit cardiaque est la phase de compensation (BOMASSI, et al., 2010). L’activation chronique des mécanismes compensateurs est toutefois délétère pour le cœur. En effet, le cœur malade éjecte déjà avec difficulté le sang. Or, les mécanismes compensateurs mis en place compliquent encore son travail en augmentant la post-charge et en provoquant une fibrose myocardique. Ainsi on aboutit à l’apparition d’un cercle vicieux car l’insuffisance s’aggrave sous l’effet des mécanismes compensateurs. Finalement, l’augmentation du tonus et du volume vasculaire résultent en l’apparition d’un œdème et d’épanchement (DE WIT, et al., 2005). L’insuffisance amont résulte de l’incapacité des ventricules à pomper le sang. En conséquence, on observe une surcharge volumique en amont du ventricule défaillant associée à une augmentation des pressions atriale et veineuse (BOMASSI, et al., 2010). Comme la pression osmotique colloïde du plasma des oiseaux peut être jusqu’à plus de deux fois moins élevée que celle des mammifères, l’existence d’une surcharge volumique provoque plus facilement une extravasation liquidienne que chez les Mammifères (STRUNK, 2003). Nous signalerons que chez les oiseaux, la valve atrio-ventriculaire droite s’hypertrophie en réponse à une surcharge de travail (hypertension pulmonaire), à l’instar du myocarde du ventricule droit dont elle est un prolongement. Il a été supposé que ce changement morphologique prédisposerait les oiseaux à une insuffisance de la valve atrio-ventriculaire droite (régurgitation), une surcharge volumique du ventricule droit et donc une insuffisance cardiaque congestive secondaire (JULIAN, 1993). Lésions : Chez les oiseaux, l’insuffisance cardiaque congestive est communément biventriculaire (OGLESBEE, et al., 1998) (JULIAN, 1993) et on observe une congestion organique associée aux lésions ayant provoqué l’insuffisance (RANDALL, et al., 1996). L’œdème pulmonaire secondaire à une augmentation de la pression veineuse pulmonaire est la trouvaille majeure lors de maladie du ventricule gauche et peut être associée à un épanchement péricardique. La congestion passive chronique du foie, l’ascite et l’œdème systémique due à une pression hydrostatique portale augmentée sont des conséquences potentiels d’une maladie ventriculaire droite (STRUNK, 2003). On observe une hypertrophie des myocytes, avec vacuolisation et dégénérescence (BEEHLER, et al., 1980). Symptômes : Les signes précoces traduisent l’incapacité du système cardiovasculaire à augmenter le débit cardiaque en réponse à un effort. Les symptômes incluent une faiblesse périodique, des syncopes, de la léthargie, une perte de poids, une intolérance à l’exercice

Page | 58
(STRUNK, 2003) et une anorexie partielle (ISAZA, et al., 1992). Lorsque la maladie progresse on observe les symptômes traduisant la congestion des organes. Lors d’une insuffisance du cœur gauche, on observera une modification de la voix, de la toux et de la dyspnée suite à la congestion et l’œdème pulmonaire (ROSENTHAL, et al., 1993). Lors d’insuffisance du cœur droit on observera un hydropéricarde et une distension cœlomique. Cette distension est secondaire à une hépatomégalie ou à la présence de liquide d’ascite lesquels peuvent conduire à une dyspnée par compression des sacs aériens. L’ensemble de ces symptômes seront observés lors d’insuffisance globale (DONELEY, 2011). Diagnostic : Le diagnostic est rarement réalisé ante-mortem. Il repose sur l’existence de symptômes d’insuffisance cardiaque et d’un souffle cardiaque. Une analyse biochimique montrera fréquemment une augmentation modérée des acides biliaires secondaire à la congestion hépatique. Des éléments radiographiques tels qu’une augmentation de la taille de la silhouette cardiaque, un œdème pulmonaire, une congestion hépatique, une compression des sacs aériens abdominaux, une perte de contraste abdominal ou une hyperinsufflation des diverticules axillaires du sac aérien inter-claviculaire seront évocateurs d’une insuffisance cardiaque. L’électrocardiogramme pourra quant à lui révéler une déviation de l’axe électrique moyen, une tachycardie, une modification de l’onde P,….). L’examen échographique est le plus informatif de tous. Il pourra révéler une dilatation des cavités, des modifications pariétales ou valvulaires, un épanchement péricardique, des troubles de la contractilité, une distension des vaisseaux hépatiques, une congestion hépatique,… (LUMEIJ, et al., 1994) Pronostic : Le pronostic à long terme pour des oiseaux atteints d’insuffisance cardiaque décompensée est réservé à sombre (WELLE, et al., 2011).
B. L’arrêt cardiaque
Définition : L’arrêt cardiaque est défini comme un arrêt inattendu de la circulation efficace. Comme les chances de survies sont directement dépendante de la condition sous-jacente, le processus morbide primitif doit toujours être considéré avant de tenter une réanimation cardio-respiratoire. Chez les patients atteints de maladie critique les propriétaires doivent être informés du pronostic de récupération afin de discuter de l’intérêt d’une réanimation (COSTELLO, 2004). Etiologie : L’arrêt cardiaque peut être secondaire à une atteinte non cardiaque (intoxication, hypothermie, traumatisme, anesthésie, déséquilibre électrolytique) ou à une atteinte cardiaque primaire (troubles du rythme). Chez les oiseaux, l'arrêt cardiaque se produit habituellement après l'arrêt respiratoire. Des poulets anesthésiés à l’halotane montrait un délai d'environ 10 minutes entre l'arrêt respiratoire et l'arrêt cardiaque. Habituellement, l’arrêt cardiaque survient plus rapidement (COLES, et al., 2007). Diagnostic : Cliniquement on peut noter une absence de pouls, une pâleur (LIERZ, et al., 2012) et une absence de bruits cardiaques (COSTELLO, 2004). L’électrocardiographie montre une asystole (ligne plate) (LIERZ, et al., 2012) ou des arythmies cardiaques telles qu’une dissociation électro-mécanique (désormais appelé activité électrique sans pouls) ou une fibrillation ventriculaire (COSTELLO, 2004). La reconnaissance du rythme d’arrêt primaire est essentielle pour traiter de façon appropriée ces patients, ainsi que pour évaluer la réponse au traitement (LIERZ, et al., 2012). Pronostic : Le pronostic est réservé à sombre (LIERZ, et al., 2012).

Page | 59
C. Affections de la paroi cardiaque
1. Les affections endocardiques
a. Endocardites:
Définition : Les endocardites correspondent à des inflammations de l’endocarde. Bien que ce terme regroupe tous les types d’inflammation impliquant toute portion de l’endocarde, les rapports en médecine aviaires décrivent uniquement des endocardites végétatives valvulaires infectieuses. Elles correspondent à la cause la plus fréquente des affections endocardiques chez les oiseaux (DE WIT, et al., 2005). Etiologie : Les bactéries les plus communément impliquées comprennent les Streptococcus spp. (faecalis) (SEKIZAKI, et al., 2008) (GROSS, et al., 1962), Staphylococcus spp. (aureus) (GROSS, et al., 1962) (RANDOLPH, et al., 1984), Escherichia Coli, Pasteurella spp. (multocida) (GROSS, et al., 1962), Pseudomonas aeruginosa, Erysipelothrix rhusiopathiae (HARARI, et al., 1983), Enterobacter cloacae (ISAZA, et al., 1992), Enterococcus (RANDALL, et al., 1996), Lactobacillus jensenii (FOLDENAUER, et al., 2009) D’autre causes infectieuses d’endocardite incluent les infections parasitaires comme la trichomoniase (DE WIT, et al., 2005) ou la maladie des vers du cœur (ECHOLS, et al., 2000). Epidémiologie : Des endocardites bactériennes ont été rapportées chez diverses espèces d’oiseaux comme les galliformes (SEKIZAKI, et al., 2008) (RANDALL, et al., 1996) chez qui l’incidence est généralement inférieur à 0,5% (GILKA, et al., 1990), les psittaciformes (ara bleu et or – Ara ararauna (ISAZA, et al., 1992), amazone de Salvin – Amazona autumnalis salvini (FOLDENAUER, et al., 2009)), les threskiornithidés (ibis chauve – Geronticus eremita, ibis sacré – Threskiomis aethiopicus (GREENWOOD, et al., 1996)) (cygne – Cygnus cygnus (HARARI, et al., 1983)), les accipitridés (pygargue à tête blanche – Haliaeetus leucocephalus (JESSUP, 1980) un faucon chiquera (Falco chiquera) (GREENWOOD, et al., 1996)), les casuariformes (RANDOLPH, et al., 1984), une grue antigone (Grus antigone), , un nandou d’amérique (Rhea americana) (GREENWOOD, et al., 1996), un émeu (Dromaius novaehollandiae) (RANDOLPH, et al., 1984) Facteurs favorisants : La plupart des endocardites bactériennes résultent d’une exposition cardiaque à des bactéries véhiculées par le sang. Ces affections sont ainsi plus fréquemment rencontrées chez les oiseaux présentant une bactériémie chronique. Cette situation s’observe lors de septicémie bactérienne chronique (HARARI, et al., 1983), de foyer infectieux chroniques internes (salpingite, hépatite (LUMEIJ, et al., 1994) (DONELEY, 2011), néphrite (TRISTAN, 2010)), de lésions traumatiques (arthrite septique (GREENWOOD, et al., 1996), pododermatite (WALLACH, et al., 1969), angelures (GREENWOOD, et al., 1996)) ou d’immunosuppression conduisant à une translocation bactérienne du tractus gastro-intestinal vers la circulation sanguine (JESSUP, 1980) (FOLDENAUER, et al., 2009). Parallèlement, le cœur peut être plus facilement colonisé lorsqu’il est atteint de myocardite dégénérative ou de lésions congénitales modifiant le flux sanguin (JONES, 2008). Pathogénie : Après que les valves aient été lésées par l’altération du flux sanguin dans le cœur, les surfaces endocardiques des valves peuvent être infectées par les bactéries se trouvant dans le flux sanguin circulant (HARARI, et al., 1983). Les lésions peuvent s’étendre à partir de l’endocarde vers le myocarde (DE WIT, et al., 2005). Les valves mitrales ou aortiques peuvent être affectées (STRUNK, 2003) et une inflammation chronique des valves semi lunaires peut s’accompagner d’une sténose (KRAUTWALD-JUNGHANNS, et al., 2004).

Page | 60
L’insuffisance valvulaire qui résulte de l’atteinte valvulaire provoque habituellement une surcharge volumique se soldant par une insuffisance cardiaque congestive (HARARI, et al., 1983) (GREENWOOD, et al., 1996). La maladie peut être compliquée par l’apparition de thrombo-embolismes secondaires aux lésions cardiaques (RANDOLPH, et al., 1984) ou par l’émission d’emboles septiques dans le système nerveux central, l’estomac (LEMON, et al., 2012), les poumons (JESSUP, 1980), le foie, la rate, le cœur et les reins (JESSUP, 1980). Lésions : Les valves mitrales et tricuspides sont les plus souvent atteintes selon certains auteurs (RANDOLPH, et al., 1984) alors que d’autres estiment que ce sont les valves mitrales et aortiques (RANDALL, et al., 1996). Les lésions consistent en des masses irrégulières multilobulées, molles blanche à jaunâtres sur la surface endocardique (DONELEY, 2011) (HARARI, et al., 1983). Les lésions peuvent également se présenter comme un épaississement végétatif rouge sombre et friable des valves ainsi que par de petits nodules associés aux cordages tendineux correspondant à des thrombi (RANDOLPH, et al., 1984). Il est possible d’observer une dilatation des capillaires autour des artères coronaires dans l’épicarde lors de thrombus coronarien (GREENWOOD, et al., 1996). Microscopiquement on observe une nécrose, de la fibrine et un infiltrat principalement hétérophilique (et moindrement lymphocytaire ou macrophagique (ISAZA, et al., 1992)), contenant des bactéries dans les zones nécrotiques (SEKIZAKI, et al., 2008). Symptômes : A l’examen physique, un souffle systolique peut être audible à gauche lors d’atteinte de la valve mitrale (ISAZA, et al., 1992) (RANDOLPH, et al., 1984) mais ce n’est pas toujours le cas (FOLDENAUER, et al., 2009). Des symptômes peuvent être associés à l’existence d’un thrombo-embolisme et représentent alors les conséquences d’une ischémie. Ainsi, une hypotonie d’une patte, associé à une froideur a été décrit suite à un thrombolembolisme secondaire à une endocardite bactérienne (RANDOLPH, et al., 1984) On peut observer les symptômes d’une insuffisance cardiaque congestive gauche, droite ou globale (DONELEY, 2011) (ISAZA, et al., 1992). Dans la plupart des cas on assiste à une mort subite sans symptômes annonciateurs. (LEMON, et al., 2012) Diagnostic : Dans une étude portant sur des endocardites induite expérimentalement chez des poulets et des dindes, les indicateurs les plus fiables et les plus précoces de l’endocardite étaient la leucocytose hétérophilique, l’hypotension et une culture sanguine positive. Des souffles cardiaques ou d’autres signes de la maladie n’apparaitraient que plus tardivement (GROSS, et al., 1962). Ainsi, on s’intéressera aux éléments suivants : Une anamnèse faisant état d’une apparition soudaine de signes cliniques tels que léthargie, dyspnée, faiblesse, collapsus, syncope. Un examen clinique L’auscultation d’un souffle cardiaque. L’utilisation de culture sanguine pour la détection d’une bactériémie ante-mortem (ISAZA, et al., 1992), bien que des faux positifs puissent exister. Un examen hématologique à la recherche d’une leucocytose hétérophilique (ISAZA, et al., 1992) bien que cette anomalie soit inconstante (RANDOLPH, et al., 1984). L’échocardiographie 2D peut révéler une masse adhérée à la valvule (RANDOLPH, et al., 1984) ou des modifications caractéristiques d’une insuffisance cardiaque. L’échographie Doppler permet de mettre en évidence une régurgitation (PEES, et al., 2006). Pronostic : Le pronostic est réservé. En effet, le traitement de cette maladie est généralement sans succès car les antibiotiques atteignent rarement la concentration efficace au sein du tissu valvulaire infecté. De plus, les lésions de fibrose cicatricielle secondaires à la multiplication bactérienne peuvent avoir de sérieuse répercussion sur l’hémodynamique cardiaque (LEMON, et al., 2012). Pour certains auteur toutefois, le pronostic est bon en cas d’endocardite infectieuse aiguë associée à un traitement agressif.

Page | 61
b. L’endocardiose
Définition : L’endocardiose est un épaississement nodulaire ou une fibrose du bord libre des valves atrio-ventriculaires. Elle est plus fréquemment observée sur la valve atrio-ventriculaire gauche que droite. Occasionnellement elle est retrouvé dans l’endocarde ventriculaire, sous la valve atrio-ventriculaire (CRESPO, et al., 2008). Epidémiologie : Contrairement aux chiens, chez qui l’endocardiose mitrale est considérée être la première cause d’insuffisance cardiaque, la dégénération myxomateuse de la valve mitrale est très peu décrite en médecine aviaire à l’exception des volailles. Chez les volailles, cette lésion est une trouvaille fréquente lors d’autopsie (18% des oiseaux sains) (OLKOWSKI, et al., 1998) qui est plus communément retrouvée chez les oiseaux « âgés » (CRESPO, et al., 2008) atteints du syndrome de l’ascite (OLKOWSKI, et al., 1998). Toutefois, l’endocardiose a été observée chez des poulets commerciaux âgés de 7 jours. (OLKOWSKI, et al., 1998). En dehors des volailles, cette affection a été rapportée chez une perruche à collier (Psittacula krameri) de 27 ans (OGLESBEE, et al., 2001), chez une talève sultane (Porphyrio porphyrio) de 4,5 ans (BEEHLER, et al., 1980) et chez un secrétaire (Sagittarius serpentarius) de 10 ans (WACK, et al., 1994). Pathogénie : Quelqu’en soit l’étiologie, les dommages fonctionnels des valves atrio-ventriculaires conduisent à une insuffisance mitrale régurgitation atriale. Le ventricule associé à la valve affectée fait ainsi l’objet d’une surcharge volumique à laquelle . Ces modifications aboutissent à l’instauration d’une insuffisance cardiaque droite avec congestion hépatique (grande circulation) ou une insuffisance cardiaque gauche avec œdème pulmonaire (petite circulation) pouvant évoluer en insuffisance cardiaque globale (PEES, et al., 2006). Lésions : Macroscopiquement, la valve atrio-ventriculaire est épaissie, opaque, nodulaire et ferme avec une surface lisse. On observe une hypertrophie cardiaque excentrique et des lésions associées à une insuffisance cardiaque congestive (OGLESBEE, et al., 2001). Microscopiquement, l’examen histologique révèle une dégénérescence myxomateuse de la valve (OGLESBEE, et al., 2001), une métaplasie cartilagineuse affectant la valve mitrale, des nodules composés de tissus conjonctif lâche et une substance fondamentale pale à la coloration sur la valve. (RANDALL, et al., 1996), des hémorragies, compatible et des lésions d’insuffisance cardiaque congestive (BEEHLER, et al., 1980). Symptômes : Les symptômes sont non spécifiques et correspondent à une insuffisance cardiaque (OGLESBEE, et al., 2001) (BEEHLER, et al., 1980). Un souffle peut être audible à gauche du sternum, de grade IV/VI holo-systolique et irradiant du côté droit du sternum (OGLESBEE, et al., 2001). Toutefois, cette trouvaille est inconstante (BEEHLER, et al., 1980). Diagnostic : L’examen clinique et la découverte d’un souffle ne peuvent apporter qu’une suspicion diagnostique. Les radiographies montrent un élargissement de la silhouette cardiaque voire un élargissement de l’atrium concerné. L’ECG peut être normal ou montrer une fibrillation atriale. L’échocardiographie peut permettre de confirmer le diagnostic.

Page | 62
c. La fibrose endocardique
Une fibrose endocardique a été décrite chez un secrétaire (Sagittarius serpentarius) mâle de 10 ans, présenté avec des symptômes d’insuffisance cardiaque congestive La fibrose endocardique est associée à une altération de la contractilité du ventricule affecté. Les lésions consistent en une fibrose endocardique du ventricule gauche et une hyperplasie endocardique légère et diffuse. (WACK, et al., 1994). La fibrose endocardique du ventricule gauche de poulet a également été décrite et observée chez des poulets SPF. Les lésions consistent en un stroma lâche de fibroblaste organisé et de fibres de collagènes éparses dans une matrice de coloration pâle, ressemblant au composant stromal des valves atrio-ventriculaires et sont recouvertes, sur leur face luminale par un tissu endothélial intact et hypertrophié et une couche sous-endothéliale de stroma compact. Ces lésions sont localisées de façon circonférentielle autour de la cavité ventriculaire et de façon alignées. Cette disposition remarquable fait penser qu’une origine physique (contact répété des surfaces ventriculaires) ou hémodynamique pourrait être à l’origine de cette modification (DANGLER, et al., 1994).
d. L’insuffisance valvulaire idiopathique
Une insuffisance valvulaire gauche d’origine idiopathique a été décrite chez un mainate de 10 ans présentant des symptômes d’insuffisance cardiaque congestive globale. Les examens cytologique, biochimique, hématologique, radiographiques, électrocardiographiques et échocardiographique n’ont pas permis d’en préciser l’origine. (ROSENTHAL, et al., 1993).
e. Démarche diagnostique lors de suspicion d’une maladie endocardique
Des symptômes non spécifiques liés à l’insuffisance cardiaque (faiblesse, dyspnée et distension abdominale) peuvent être remarquées (PEES, et al., 2006). L’auscultation d’un souffle au cours de l’examen physique peut indiquer une maladie endocardique (DE WIT, et al., 2005). L’examen radiographique ne révèle souvent rien d’anormal jusqu’au développement d’une insuffisance cardiaque congestive où les clichés peuvent alors permettre d’apporter des informations quand à une augmentation de la taille cardiaque ou d’une congestion des organes (DE WIT, et al., 2005). Toutefois le diagnostic radiographique d’altération endocardique n’est pas possible (PEES, et al., 2006). Les observations électrocardiographiques peuvent être normales ou révéler un ECG de bas voltage ou encore une fibrillation atriale (ROSENTHAL, et al., 1993). Le diagnostic des maladies endocardiques infectieuse repose sur une culture sanguine positive associée à l’existence de lésions valvulaires. Il est à noter que l’endocardite reste dans la liste des diagnostic différentiels même lors de culture sanguine négative. L’échocardiographie pourra montrer l’existence d’une régurgitation valvulaire associée à un processus pathologique. L’absence d’anomalie sous-jacente à la régurgitation permet de conclure au diagnostic d’insuffisance valvulaire idiopathique.

Page | 63
2. Les affections myocardiques
Une étude révèle que 15% des psittaciformes autopsiés montrent une hypertrophie ou une dilatation du myocarde ventriculaire (KRAUTWALD-JUNGHANNS, et al., 2004).
a. Cardiomyopathie métabolique
- Syndrome de la mort subite chez la poule
Epidémiologie : Le syndrome de la mort subite (S.M.S.) est une affection cardiovasculaire sporadique décrite primitivement en Australie (KAHN, et al., 2008) et qui est observée chez les poules pondeuses adultes ou juvéniles. Le nombre de fermes d’élevage atteintes est faible mais l’incidence peut atteindre 5% par semaine dans les poulaillers affectés. Les pics de mortalité surviennent pendant les journées très chaudes (TREMBLAY, et al., 1992). Pathogénie : Le S.M.S. est une maladie métabolique de la poule, observée immédiatement avant le démarrage de la ponte. Chez les oiseaux affectés, la kaliémie diminue à la suite d’un apport insuffisant dans la ration, d’un état d’alcalose respiratoire consécutive à l’hyperventilation lors d’un stress thermique et de l’interaction entre le potassium et d’autres minéraux. Le besoin en potassium augmente chez la poule si la ration présente une teneur faible en phosphore. L’hypokaliémie entraîne des troubles de l’excitabilité neuro-musculaire et principalement celle du cœur d’où la mort soudaine de l’oiseau (TREMBLAY, et al., 1992). Signes cliniques : Aucun signe préalable n’est manifesté par les poules. Celles-ci sont calmes et s’alimentent lentement. Dans les poulaillers atteints, la litière est souvent plus humide. Certains oiseaux présentent des convulsions et meurent à la suite d’une période d’excitation, au moment de l’alimentation, lors de combats dans le poulailler ou à la suite d’un stress (TREMBLAY, et al., 1992). Lésions : A l’autopsie on note une congestion de la crête et du cloaque. Les muscles de la poitrine sont très pâles mais le foie, la rate et les poumons sont congestionnés. Le cœur est plus volumineux et hypertrophié avec un liquide péricardique abondant. Les ovaires montrent des signes de développement mais dans la plupart des cas, la maturité sexuelle n’est pas complète (TREMBLAY, et al., 1992). Diagnostic : Un diagnostic différentiel doit être fait avec la maladie de Newcastle, l’influenza aviaire, le choléra aviaire, le stress thermique, les intoxications, le syndrome du foie gras hémorragique et un traumatisme (combats) (TREMBLAY, et al., 1992). Il faut également différencier le SMS de la poule pondeuse d’autre mort brutale chez les poules pondeuses adultes ou en croissance imputable à une hypocalcémie lors de la formation de la coquille de l’œuf. Dans ce dernier cas les poules ont déjà utilisé une grande partie du calcium stocké dans les os et on observe alors des lésions d’ostéoporose (JULIAN, 2005).L’examen biochimique du sang révèle un état d’hypokaliémie (en moyenne 2,64 mmol/L contre 3,32 mmol/L chez des sujets normaux) (TREMBLAY, et al., 1992).
- Hémochromatose
Hémochromatose : Cette maladie affecte essentiellement les oiseaux insectivores et les frugivores maintenus en captivité (mainates, toucans, oiseaux de paradis, loris (DIERENFELD, et al., 1994). On observe une accumulation et un dépôt de fer dans le myocarde ce qui engendre des lésions myocardiques oxydatives et une cardiomyopathie dilatée (ROSENTHAL, et al., 1993) (GOSSELIN, et al., 1983). Les symptômes sont ceux d’une insuffisance cardiaque droite puis gauche (ROSSI, et al., 2005) (CORK, 2000).

Page | 64
- Lipomatosis cordis
L’existence d’une infiltration adipeuse épicardique peut être retrouvée de façon normale chez les oiseaux. Toutefois, elle peut devenir excessive, ce qui est une trouvaille habituellement observée chez les oiseaux obèses. Macroscopiquement, la graisse apparait normale mais histologiquement, on observe une infiltration du myocarde en profondeur (SCHMIDT, et al., 2003). Un tel excès peut provoquer une diminution de la contractilité myocardique (PEES, et al., 2006) et donc favorise le développement l’insuffisance cardiaque. Cette condition est observé chez des oiseaux mourant subitement sans autre modification morphologique pouvant expliquer la mort (SCHMIDT, et al., 2003)
- Transport prolongé
Des oiseaux sauvages ayant été longuement transportés peuvent manifester une nécrose myocardique consécutive à des perturbations métaboliques et se traduisant par l’existence d’arythmies neurogéniques (PEES, et al., 2006).
- Le syndrome de l’ascite ou Syndrôme de l’Hypertension
Pulmonaire (SPH) ou maladie des hautes altitudes
Définition : En médecine aviaire, le syndrome de l’ascite chez le poulet de chair est défini par l’accumulation d’un transsudat non inflammatoire dans la cavité abdominale due à une défaillance du ventricule droit du cœur (TREMBLAY, et al., 1992). On estime que cette condition coûterait environ 1 milliard de dollars à l’industrie du poulet de chair chaque année (WEST, et al., 2007). Epidémiologie : Ce syndrome est de distribution mondiale. De façon générale un pour cent des oiseaux est atteint et l’incidence augmente dans les élevages situés à haute altitude (TREMBLAY, et al., 1992). Le SPH est connu depuis de nombreuses années dans les élevages situés en altitude. Plus récemment, cette condition a été également décrite à basses et moyennes altitudes où elle devient prépondérante. Des études réalisées chez le canard et la dinde ont montré que ces espèces étaient résistantes (JULIAN, et al., 1993). Etio-pathogénie : L’hypothèse majeure à ce jour soutien que l’ascite observée dans ce syndrome résulte d’une insuffisance cardiaque droite secondaire à une hypertension pulmonaire. Cette hypertension résulte à la fois d’une augmentation du débit sanguin pulmonaire en réponse à l’état d’hypoxie provoqué par une demande métabolique élevée (taux de croissance élevé de certaines souches, température faible, alimentation) et d’une augmentation des résistances vasculaires (polycythémie réactionnelle à l’hypoxémie, faible capacité vasculaire des poumons, alimentation riche en sel provoquant une perte de déformabilité des globules rouges). L’hypertrophie myocardique droite qui résulte de cette augmentation de la post-charge provoque alors une hypertrophie de la valve atrio-ventriculaire droite et une insuffisance valvulaire (TREMBLAY, et al., 1992). Une deuxième théorie suppose que l’insuffisance cardiaque droite serait secondaire à une insuffisance contractile due à un manque d’énergie cardiaque (NAIN, et al., 2008) et aggravée par une insuffisance cardiaque gauche secondaire à une insuffisance valvulaire gauche (OLKOWSKI, et al., 1998). Il en résulterait une hypertension pulmonaire sans augmentation du débit sanguin pulmonaire et l’état d’hypoxygénation observé serait alors une conséquence et non la cause de ce syndrome (OLKOWSKI, et al., 1998) (DRISS, et al., 2000). L’éventualité d’une anomalie de la relaxation myocardique secondaire à un hypothyroïdisme a été évoqué mais non investigué (LI, et al., 2006).

Page | 65
Symptômes : Lors de forme aiguë du syndrome d’hypertension pulmonaire, les poulets en croissance meurent subitement d’un œdème pulmonaire (WEST, et al., 2007) ou après un épisode de cyanose et de détresse respiratoire (respiration bec ouvert). Dans les deux cas, les oiseaux sont de taille normale. Lors de forme subaiguë, les oiseaux développent une insuffisance ventriculaire droite et arrêtent de grandir. Ils sont donc plus petits que les autres oiseaux du lot. S’ils survivent quelques jours, on observe l’apparition d’une distension abdominale liée à l’accumulation d’une ascite. Les oiseaux affectés sont apathiques, déprimés, inactifs, cyanosés et peuvent être en détresse respiratoire (TREMBLAY, et al., 1992) (JULIAN, 2005). Lésions : Macroscopiquement, on observe de l’ascite de couleur rosé avec ou sans caillot de fibrine. Le cœur droit est élargi suite à la dilatation du ventricule droit, de l’atrium droit, du sinus veineux et des veines caves mais aussi du fait de l’hypertrophie de la paroi ventriculaire. Des modifications hépatiques variables seront fréquemment observées ainsi qu’une dilatation du ventricule gauche (JULIAN, 2005). Un épaississement nodulaire de la valve atrio-ventriculaire gauche est fréquente. Un hydropéricarde peut être présent (TREMBLAY, et al., 1992). Microscopiquement, les lésions concernant le cœur incluent une désorganisation légère des fibres myocardiques, une dégénérescence myocardique parfois associé à une calcification et des aggrégats lipiques. On observe de l’œdème, une prolifération de tissu conjonctif lâche entre les fibres, des hémorragies focales et une infiltration par des hétérophiles, des éosinophiles et occasionnellement des lymphocytes. Le péricarde est fibrosé (MAXWELL, et al., 1986). Diagnostic : L’électrocardiogramme des poulets atteints du SPH montre une augmentation de voltage du complexe QRS compatible avec une dilatation et une hypertrophie du ventricule droit. Ces modifications de l’ECG sont précédées par des modifications du ventricule gauche compatibles avec un élargissement et une insuffisance progressive du cœur gauche ainsi qu’une hypertrophie du cœur droit compensatoire chez des oiseaux élevées à faible altitude (TREMBLAY, et al., 1992). Une bradycardie est observée (OLKOWSKI, 2007). L’hématologie révèle une polycythémie, une augmentation de l’hémoglobine et de l’hématocrite vraisemblablement secondaire à l’hypoxie. Le comptage leucocytaire montre une leucocytose caractérisée par une hétérophilie, une monocytose et une lymphopénie, compatible avec une formule de stress (MAXWELL, et al., 1986). L’échocardiographie révèle une diminution de la fraction de raccourcissement du ventricule gauche. Une insuffisance de la valve atrio-ventriculaire gauche est fréquemment observée. Un liquide d’ascite peut être observé dans les cas les plus avancés (OLKOWSKI, 2007).

Page | 66
b. Cardiomyopathie toxique
Chez les oiseaux, les cardiomyopathies d’origine toxiques sont essentiellement provoquées par les agents suivants. Le monensin est un antibiotique anti-coccidien de la classe des ionophores. Sa toxicité a été rapportée chez des poulet de chair, des poulets reproducteurs, des dindes, des cailles et des autruches (FULTON, 2008) (ZAVALA, et al., 2011). Il est responsable d’anomalie de la conductance membranaire résultant en une accumulation intracellulaire anormale de sodium et de calcium ainsi qu’en une diminution des réserves en ATP dans les myocytes cardiaques (BHAVSAR, et al., 2010). L’accumulation intracellulaire d’ions provoque un œdème, une dégénérescence et une nécrose des cardiomyocytes (FULTON, 2008). Le cœur se dilate et perd son tonus. On observe alors une insuffisance cardiaque (DOWLING, 1992). La furazolidone est un antibiotique appartenant à la famille des nitrofuranes. Cette intoxication concerne essentiellement les dindes, les canards et les poulets. Les effets toxiques varient fortement avec la dose, l’âge des animaux et il existe également une sensibilité individuelle marquée (FULTON, 2008). La furazolidone provoquerait une ischémie cardiaque chez les oiseaux à forte croissance. Celle-ci perturberait la conductance membranaire et altérerait les propriétés contractiles du myocarde (MIRSALIMI, et al., 1990). Les symptômes et les lésions correspondent à ceux d’une cardiopathie dilatée et d’une insuffisance cardiaque. (MIRSALIMI, et al., 1990) (FULTON, 2008). L’halopéridol est un agent antipsychotique exerçant ses effets via son action antagoniste dopaminergique. Il est utilisé pour traiter les troubles compulsifs (picage) et agressif chez les perroquets. Les effets cardiovasculaires d’une intoxication à l’halopéridol comprennent une hypotension et une bradycardie (MARTIN, 2006). Le moniliformin est une mycotoxine produite par Fusobacterium verticillioides, qui se développe notamment sur les grains de maïs (HOERR, 2008). La toxicité cardiaque du moniliformin est similaire chez les poulets, les dindes, les cailless et le canards et constitue la cause primaire de la mort (SHARMA, et al., 2012). Le moniliformin agirait en inhibant sélectivement l’oxidation mitochondrial du pyruvate et de l’α-ketoglutarate (THIEL, 1978). L’électrocardiographie d’oiseau intoxiqué au moniliformin est caractérisée par une bradycardie avec un allongement des intervalles P-R, Q-T et S-T (NAGARA, et al., 1996). Les lésions sont celles d’une insuffisance cardiaque associée à une dégénérescence et une nécrose myocardique (HOERR, 2008). Les polychlorobiphényls sont des composés produits par les incinérations industrielles. Chez le poulet, la dioxine endommage l’endothélium vasculaire, provoquant des pertes vasculaire et des mouvements de fluides dans les cavités corporelles et le tissu sous-cutané. La dioxine engendre également une dégénérescence du cœur évoluant possiblement vers une insuffisance cardiaque droite avec hypertrophie ventriculaire droite et dilatation des cavités. On observe des symptômes tels que détresse respiratoire, faiblesse, ataxie et œdème (FULTON, 2008). L’acide érucique est un acide gras monoénoïque à longue chaîne qui est retrouvé dans les graines de colza. Sa toxicité a été décrite chez les canetons, les dindes et les poulets. Elle se manifeste par une insuffisance cardiaque avec ascite et hydropéricarde (VAN VLEET, et al., 1986) (FULTON, 2008).

Page | 67
Pois de senteur (Lathyrus spp.) : L’intoxication, également appelée lathyrisme, est liée à l’ingestion de la graine qui contient du beta-aminopropionitrile (BAPN) (FULTON, 2008). On distingue plusieurs formes, d’intoxication : le neurolathyrisme, l’ostéolathyrisme et l’angiolathyrisme qui nous intéresse ici. L’angiolathyrisme se manifeste par une modification de l’élasticité de l’aorte ce qui provoque un anévrisme aortique pouvant se rompre. La manifestation clinique correspond à une diminution de la croissance, des hémorragies sous-cutanées au point de pression (poitrine, jarret) dans 30% des cas et une mort subite secondaire à une hémorragie interne (rupture aortique) dans 60% des cas. Les lésions étant très similaires avec celles liées à une déficience en cuivre, il a été suggéré que le BAPN intervenait sur la même enzyme que le cuivre, diminuait la disponibilité du cuivre (chélation ou augmentation de l’utilisation per se) ou intervenait dans la cascade réactionnelle de l’élastine à un autre endroit (SAVAGE, et al., 1966). Coton (Gossypium spp.) : L’intoxication se fait par ingestion des graines de coton qui contiennent le gossypol. Le gossypol a des effets sur de nombreux systèmes, mais nous nous limiterons à ceux concernant le cœur. Une exposition prolongée peut ainsi provoquer des troubles de la conduction cardiaque semblables à ceux rencontrés lors d’hyperkaliémie. Il en résulte une défaillance cardiaque aiguë se manifestant par une mort d’apparition subite ou faisant suite à des symptômes d’insuffisance cardiaque gauche (FULTON, 2008). Lorsque des lésions sont visibles, elle correspondent à un œdème et une nécrose du myocarde (KAHN, et al., 2008). Café nègre (Cassia occidentalis, cassia obtusifolia, senna occidentalis) : Ce sont les graines qui sont toxiques. On observe une myopathie toxique caractérisée par une dégénérescence et une nécrose myocardiques associées à de l’œdème. Les symptômes sont ; une faiblesse, de l’ataxie, une paralysie, une diminution de la production d’œuf et de la diarrhée. Les muscles apparaissent pâles (FULTON, 2008). Laurier (Nerium oleander), Muguet (Convallaria majalis), Digitale (Digitalis purpurea) et Kalanchoe (Kalanchoe daigremontiana) : Ce sont des plantes d’ornement dont la toxicité est conférée par la présence de digitaliques (= glycosides cardiaques ou hétérosides cardiotoniques) respectivement de type cardénolide (oléandrine, nérine, convalatoxine, digoxine) et bufadienolide (daigremontianine) . Ces digitaliques inhibent la pompe Na+/K+ ATPase. En conséquence, ils ont un effet inotrope positif (le, chronotrope négatif et dromotrope négatif car la durée du potentiel d’action est allongée. De plus, elles ont un effet bathmotrope positif car le potentiel de repos est plus proche de la neutralité et les cellules sont donc plus facilement excitables. L’intoxication se manifeste donc par une augmentation de la contractilité, des arythmies ventriculaire, une bradycardie, un arrêt cardiaque (LIGHTFOOT, et al., 2008), par une dépression, de la faiblesse, de la diarrhée, une gastroentérite, une dégénération hépatique et des symptômes cardiaques et la mort. (STRUNK, 2003) If (Taxus media), Morelle noire (Solanum tuberosum) et autres Solanum spp (pomme de terre) : Les taxines de l’if et la solanine contenue dans les germes et la peau des pommes de terre sont des alcaloïdes cardiotoxiques. Ils perturberaient les flux de calcium ionisé dans les cardiomyocytes et provoqueraient des troubles de la contractilité et des arythmies fatales. Une exposition chronique peut aboutir à une calcification du réseau vasculaire (STRUNK, 2003). Crotalaria spp. (alcaloïdes pyrrolizidine) : Les alcaloïdes sont contenues dans les graines, la tige et les feuilles. Elles provoquent une atteinte du système cardiovasculaire représenté par un hydropéricarde et de l’ascite (FULTON, 2008) .

Page | 68
La persine est un composé retrouvé dans le fruit de l’avocat (Persea americana). La toxicité myocardique a été rapportée lors d’empoisonnement à l’avocat chez des autruches, des perruches ondulés et des canaris (DONELEY, 2011) (KAHN, et al., 2008). Elle provoque une cardiomyopathie caractérisée par une infiltration hétérophilique marquée, une fibroplasie précoce (BURGER, et al., 1994), une dégénération des cardiomyocytes, un hydropéricarde (HARGIS, et al., 1989), de l’œdème interstitiel (FULTON, 2008), une nécrose et des hémorragies myocardiques plus marquées au niveau ventriculaire et du septum inter-ventriculaire (KAHN, et al., 2008). Les lésions provoquent une insuffisance cardiaque congestive globale (BURGER, et al., 1994) (HARGIS, et al., 1989). Chocolat (théobromine, caféine) : L’ingestion de chocolat provoque tachycardie, hypertension et arythmies (STRUNK, 2003) suite à l’action antagoniste des bases méthylxanthines qu’il contient sur les récepteurs cardiaques à l’adénosine (LIGHTFOOT, et al., 2008). Un cas décrit la mort d’un kéa (Nestor notabilis) suite à l’ingestion de 250 mg/kg de théobromine et de 20 mg/kg de caféine (GARTRELL, et al., 2007). Insecticides inhibiteurs des cholinestérases (organophosphorés et carbamates) : Les oiseaux s’intoxiquent en ingérant des proies contaminés tels que des insectes préalablement empoisonnés ou des cadavres eux-mêmes intoxiqués. Ces insecticides agissent en inhibant l’enzyme appelée acétylcholine estérase qui dégrade physiologiquement l’acétylcholine. L’acétylcholine est un neurotransmetteur impliqué dans les synapses nicotiniques et muscariniques. Lorsque ce médiateur n’est pas détruit, il s’accumule dans la synapse et après avoir excité le neurone post-synaptique bloque le passage du message nerveux. Concernant le système cardiovasculaire, cette intoxication provoque une bradycardie. L’intoxication est généralement suraiguë et le diagnostic est réalisé post-mortem (REDIG, et al., 2008). Sodium : L’excès de sodium ionisé provient habituellement du sel (chlorure de sodium) dans l’eau ou l’alimentation. On s’intéresse ici à une intoxication à des doses faibles (mais supérieures à 1200 ppm) qui engendrent chez les poulets et autres volailles des insuffisance cardiaque, de l’œdème et de l’ascite (FULTON, 2008). La sensibilité varie entre les souches et diminue avec l’âge. (MIRSALIMI, et al., 1992). Les oiseaux ont une faible capacité de concentration rénale et éprouvent des difficultés à diminuer l’osmolarité plasmatique par excrétion de l’excès de sodium dans l’eau ce qui les prédispose à la maladie (FULTON, 2008). Mentionnons que certains oiseaux d’eau ont des glandes à sel nasal qui leur permet d’excréter du Na si un excès est ingéré (FULTON, 2008). Le sodium devient toxique lorsqu’il est présent à une concentration telle que le rein reçoit davantage de Na+ que d’eau pour l’éliminer (JULIAN, et al., 1992). Dans cette situation, la rétention hydrosodée provoque une hypervolémie et une diminution de la déformabilité des globules rouges conduisant à une hypertension pulmonaire et une surcharge cardiaque fonctionnelle. En réponse à ces modifications, se développent une hypertrophie et une dilatation ventriculaire sévère puis une insuffisance valvulaire, de l’œdème et de l’ascite (MIRSALIMI, et al., 1993) (JULIAN, et al., 1987) (JULIAN, et al., 1992) (ONDERKA, et al., 1982) (MOHANTY, et al., 1969). Les lésions et les symptômes sont ceux d’une insuffisance cardiaque globale. (DAVISON, et al., 1992). Les facteurs favorisants sont représentés par le stress car les stéroïdes libérés augmentent la rétention hydrosodée (FULTON, 2008) et le froid car celui-ci augmente le débit sanguin et donc l’hypertension pulmonaire (JULIAN, et al., 1992).

Page | 69
Cobalt Des niveaux de cobalt modéré (125 ppm) provoquent une polycythémie et induisent une hypertension pulmonaire. A des doses plus élevées (500 ppm) il provoque une nécrose et une fibrose du muscle cardiaque (DIAZ, et al., 1994). Plomb : Les oiseaux d’eau (canards, cygnes, oies) sont prédisposés à l’intoxication au plomb du fait que ce soient des oiseaux chassés (plomb de chasse) et qu’ils s’alimentent de poissons (plomb de pêche). Certains grands rapaces (pygargue à tête blanche - Haliaeetus leucocephalus) sont également prédisposés car ils se nourrissent des oiseaux précédemment décrits et l’intoxication est d’autant plus rapide que leur pH ventriculaire est faible (environ 2) (JONES, 2008) (LANGELIER, et al., 1991). C’est une maladie qui peut affecter les oiseaux de cage et notamment les psittaciformes, lorsqu’ils sont logés dans des cages recouvertes de peinture au plomb ou que les objets d’enrichissement contiennent du plomb (WESTERHOF, et al., 2011). Le plomb exerce vraisembablement sa toxicité en entrant en compétition avec le calcium (MAUTINO, 1994). L’accumulation de plomb provoque une dégénérescence et une nécrose myocardique (FULTON, 2008) (LANGELIER, et al., 1991) (SILEO, et al., 1973) qui sont fréquemment associées à un infarctus myocardique. Les lésions cardiaques macroscopique sont représentée par une striation ou des foyers pâles du myocarde (LANGELIER, et al., 1991) éventuellement associé à une cardiopathie dilatée (JONES, 2008). Le péricarde peut apparaître marbré sans atteinte du myocarde (LANGELIER, et al., 1991) et il est possible d’observer un hydropéricarde. Les zones de dégénérescence sont associées à la présence d’hétérophiles et de fibroblaste et accompagnée par une fibrose de remplacement (LANGELIER, et al., 1991) Il est également possible d’observer une artérite dégénérative (Service, 1986) associée à une nécrose fibrinoïde de la média des artérioles et des petites artères (KARSTAD, 1971) (FULTON, 2008). En plus des symptômes classiques d’une insuffisance cardiaque congestive, il est possible d’observer des symptômes neurologiques et une anémie. Le diagnostic de suspicion repose sur la symptomatologie, l’existence d’anomalies hématologiques (anémie) (KAHN, et al., 2008) ou électrocardiographiques (augmentation de l’amplitude de R, inversion de l’onde T, modifications de l’élevation du segment S-T (SILEO, et al., 1973), arrêt sinusal, bloc de branche, bradycardie, bas voltage (WESTERHOF, et al., 2011)). La visualisation d’élément d’opacité métalique peut assoir la conviction d’intoxication aux métaux lourds (MAUTINO, 1994). Le diagnostic de certitude est réalisé suite au dosage de la plombémie (KAHN, et al., 2008). Zinc : Les oiseaux sont fréquemment contaminés lors de l’ingestion de pièce contenant du Zinc ou bien en rongeant du grillage galvanisé (MAUTINO, 1994). Les symptômes et la démarche diagnostic sont similaires à ceux de l’intoxation au plomb (DEGERNES, 2008).

Page | 70
c. Cardiomyopathies inflammatoires ou myocardites
- Myocardite non infectieuse : La goutte viscérale
Lors de dépôts d’urate, on observe une forte réponse inflammatoire granulocytaire aiguë comprenant des foyers nécrotiques. Un stade granulomateux peut se développer (RANDALL, et al., 1996). Cette maladie peut apparaître lors de déshydratation, d’ingestion trop importante de calcium, d’urolithiase, de carence en vitamine A, d’intoxication aux aminoglycosides ou aux métaux lourds, ou enfin lors d’insuffisance rénale (infections au virus de la bronchite infectieuse ou au virus de la néphrite aviaire) (KAHN, et al., 2008).
- Myocardites infectieuses
o Myocardites bactériennes
Etiologie : La littérature décrit l’implication des bactéries suivantes lors de myocardite : Staphylococcus aureus (RANDALL, et al., 1996), Listeria monocytogenes (RANDALL, et al., 1996), Pasteurella (RANDALL, et al., 1996), Salmonella typhimurium (RANDALL, et al., 1996), Salmonella pulorum, gallinarum (SHIVAPRASAD, et al., 2009), Erysipelothrix rhusiopathiae (GALINDO-CARDIEL, et al., 2011), Clamydia psittaci (JOHNS, et al., 2009), Listeria monocytogenes (AKANBI, et al., 2008), Mycobacterium spp. (GELIS, et al., 2006), Escherichia Coli (GROSS, 1966), Bacillus cereus (GODOY, et al., 2012), Clostridium piliforme (RAYMOND, et al., 2001), Coxiella burnetti (VAPNIARSKY, et al., 2012). Epidémiologie : Des cas de myocardite bactérienne ont été décrits chez des poules (RANDALL, et al., 1996) (SHIVAPRASAD, et al., 2009) (GROSS, 1966), des pigeons (RANDALL, et al., 1996), des passériformes (canari Serinus serinus (AKANBI, et al., 2008) des psittaciformes ((loriquets Trichoglossus haematodus (RAYMOND, et al., 2001) (GALINDO-CARDIEL, et al., 2011), eclectus (Eclectus roratus) (VAPNIARSKY, et al., 2012) (GALINDO-CARDIEL, et al., 2011), cacatoès Gang gang (Callocephalon fimbriatum) (GELIS, et al., 2006), Ara hyacinthe (Anodorhynchus hyacinthinus), Ara noble (Diopsittaca nobilis) Ara sévère (Ara severa) et Ara bleu et or (Ara ararauna) (GODOY, et al., 2012)), ainsi que chez des rapaces (buse à queue rousse (JOHNS, et al., 2009)). Lésions : Les lésions myocardiques sont dépendantes de l’agent bactérien impliqué. Ainsi, on pourra observer une myocardite à dominante suppurée lors d’infection à S.aureus ou L.monocytogenes et une myocardite à dominante granulomateuse lors d’infection à S.typhymurium (RANDALL, et al., 1996), L. monocytogenes (AKANBI, et al., 2008), Mycobacterium spp. (GELIS, et al., 2006) et lors de coligranulomatosis (AKANBI, et al., 2008). Une myocardite à dominante hémorragique est habituellement décrite lors d’infection à E.rhusiopathiae (GALINDO-CARDIEL, et al., 2011) et une myocardite à dominante lymphocytaire lors d’infection à E.Coli (GROSS, 1966), B.cereus (GODOY, et al., 2012) et C.burnetti (VAPNIARSKY, et al., 2012). On observe souvent une bactériémie concomitante (GODOY, et al., 2012) (GALINDO-CARDIEL, et al., 2011).

Page | 71
o Myocardites virales Etiologie : Les agents viraux responsables de myocardites sont les suivants : Herpesvirus (RANDALL, et al., 1996), Entérovirus (RANDALL, et al., 1996), Virus de l’influenza aviaire (MCKENZIE, et al., 1972), Virus de la maladie de Newcastle (VAN VLEET, et al., 1986), Virus de l’encéphalite de l’est et de l’ouest (VAN VLEET, et al., 1986) (MITCHELL, et al., 1982), Réovirus (chicken reovirus (DAVIS, et al., 2012), turkey reovirus (SHIVAPRASAD, et al., 2009), Virus de la leucose aviaire (GILKA, et al., 1990)), Parvovirus (VAN VLEET, et al., 1986), West Nile Virus, Virus de la maladie de la dilatation du proventricule (BERHANE, et al., 2001) , Circovirus (ANDRAL, et al., 1990) Adénovirus (SHIVAPRASAD, et al., 2004) bunyavirus (SHIVAPRASAD, et al., 2002), d’un togavirus (GUY, et al., 2008) Epidémiologie : Des cas de myocardites virales ont été décrites chez des Ansériformes (Canard (RANDALL, et al., 1996), Oie (VAN VLEET, et al., 1986)), des Galliformes (STEELE, et al., 2000) (Poule (VAN VLEET, et al., 1986) (RANDALL, et al., 1996)) Perdrix choukar (Alectoris chukar) (VAN VLEET, et al., 1986), des Psittaciformes (PALMIERI, et al., 2011) (Cacatoès des moluques (Cacatua moluccensis), Cacatoès à huppe jaune (Cacatua galerita), Cacatoès blanc (Cacatua alba) (BERHANE, et al., 2001), Conure de patagonie (Cyanoliseus patagonus) (CAZAYOUX, 1992), Conure à front bleu (Thectocercus acuticaudata) (PALMIERI, et al., 2011), Perroquet des niam-niams (Poicephalus rubfiventris (BERHANE, et al., 2001)), Papegeais maillés (Deroptyus accipitrinus), Ara bleu et or (Ara ararauna), Conures Jandaya (Aratinga jandaya), Gris d’afrique (Psittacus erithacus) (BERHANE, et al., 2001), Conure à long bec (Enicognathus leptorhynchus) (BERHANE, et al., 2001), Loris et loriquets (Eos, Lorius, Pseudeos, et Trichoglossus spp) (PALMIERI, et al., 2011), Perruches ondulées, (Melopsittacus undulatus) (PALMIERI, et al., 2011), Perruches calopsittes (Nymphicus hollandicus) (PALMIERI, et al., 2011), les platycèrques (Platycercus spp) (PALMIERI, et al., 2011), Amazones (Amazona spp.) (PALMIERI, et al., 2011), Touis (Forpus coelestis, Brotogeris versicolurus), des Passeriformes, des Ciconiiformes, des Pelecaniformes, des Charadriiformes et des Falconiformes, (STEELE, et al., 2000) Lésions : Les lésions macroscopiques sont surtout représentées par l’existences d’une pâleur d’aspect motté, de bandes pâles multifocales ou d’une pâleur généralisée. Ces lésions correspondent à une myocardite lymphoplasmocytaire non suppurée et nécrosante (RANDALL, et al., 1996) (VAN VLEET, et al., 1986) (DAVIS, et al., 2012) (PHALEN, et al., 2004) (FRANCA, et al., 2010) (SHIVAPRASAD, et al., 2009) (GILKA, et al., 1990) (STEELE, et al., 2000) (JONES) (CAZAYOUX, 1992) (BERHANE, et al., 2001) (SAITO, et al., 2007) (VAN VLEET, et al., 1986). Dans certaines situations, on observe une cardiomyopathie dilatée et des lésions d’insuffisance cardiaque (GILKA, et al., 1990) (DAVIS, et al., 2012). (VAN VLEET, et al., 1986), une minéralisation ou des thrombi (O'BRIEN, et al., 2010).
o Myocardites parasitaires Etiologie : La littérature décrit l’implication des parasites suivants lors de myocardite : Sarcocystis falcatula (BOX, et al., 1982) (PAGE, et al., 1989), Toxoplasma gondii (JUNIOR, et al., 2012), Eimeria spp. (NOVILLA, et al., 2004), Leucocytozoon spp. (SIMPSON, 1991) (RAE, 1995) (RANDALL, et al., 1996), Trichomonas spp. (WACK, et al., 1994), Serratospiculum amaculata (BIGLAND, et al., 1964), Filaria spp. (SMITH, 1996), Sarconema eurycerca (WOO, et al., 2010) (EUN-JUNG, et al., 2010) (TEGLAS, et al., 1998), Paronchocerca ciconarum (BARTLETT, 2008).

Page | 72
Epidémiologie : Des myocardites parasitaires ont été décrites chez des Falconiformes (SMITH, 1996) (épervier d’europe (Accipiter nisus) (JONES, 2008), un pygargue à tête blanche (Haliaeetus leucocephalus) (SZABO, et al., 2004) (CRAWLEY, et al., 1982) Faucon des prairires (BIGLAND, et al., 1964)), des Psittaciformes (lori rouge (Eos bornea), Grand eclectus (Eclectus roratus), Loriquet orné (Trichoglossus ornatus), Amazone de cuba (Amazona leucocephala) (SZABO, et al., 2004), Lori rouge (Eos bornea) (HOWERTH, et al., 1991), Amazone vineuse (Amazona vinacea) (JUNIOR, et al., 2012), Cacatoès des moluques (Cacatua moluccensis) (LATIMER, et al., 1990), (VILLAR, et al., 2008)), des Columbiformes (Pigeon (ECCO, et al., 2008), des Ansériformes (Canard pilet (Anas acuta) (CRAWLEY, et al., 1982), Bernache néné male et femelle (Nesochen sandicensis) (WORK, et al., 2002), Cygne sauvage (Cygnus) (WOO, et al., 2010)), des Suliformes (Fou à pied rouge (Sula sula) (WORK, et al., 2002), des Charadriiformes (Grand Labbe (Stercorarius skua (MUNDAY, et al., 1979)), des Podicipediformes (grèbe argenté (Poliocephalus poliocephalus) (MUNDAY, et al., 1979)), des Passériformes (Pie à dos blanc (Gymnorhina hypoleuca (MUNDAY, et al., 1979)), des Galliformes (Francolin d’Erckel (Francolinus erckelii) (WORK, et al., 2002), des Gruiformes (Grue du Canada (Grus canadiensis) Grue d’Amérique (Grus Americana) (NOVILLA, et al., 2004)), des Ciconiformes (Jabiru d’Afrique (Ephippiorhynchus senegalensis) (ECHOLS, et al., 2000)), Pica pica sericea, Cynopica cyana interposita, corvus torquatus, Ardea sp. Leptoptilos crumeniferus , Ciconia cicona, ciconia episcopus (BARTLETT, et al., 1936) (ENSLEY, 1978) (FAUST, 1966) (PETERS, 1936) Lésions : Lorsqu’elles sont présentes, les lésions cardiaques microscopiques montrent une décoloration focale (WORK, et al., 2002). Microscopiquement, on observe une myocardite nécrosante associée à un infiltrat interstitiel multifocal de type variable : lymphoplasmocytaire (VILLAR, et al., 2008) (SZABO, et al., 2004), lymphohistiocytaire (JUNIOR, et al., 2012) ou mononucléaire (macrophages essentiellement) (WORK, et al., 2002), éventuellement associés à quelques hétérophiles (HOWERTH, et al., 1991) (SZABO, et al., 2004). Cet infiltrat inflammatoire peut envahir l’épicarde (HOWERTH, et al., 1991). Il est à signaler que la présence de kystes de bradyzoïtes ne semble provoquer ni nécrose ni inflammation (JUNIOR, et al., 2012). Les lésions associées à une infection par leucocytozoon consistent en une infiltration granulocytaire (RANDALL, et al., 1996) et une infection par eimeria provoque une myocardite granulomateuse (NOVILLA, et al., 2004)
o Myocardites fungiques Une infection à Candida albicans, a été rapporté chez un canari (Serinus canaria). Cette inflammation était histologiquement caractérisée par une myocardite granulomateuse avec un noyau nécrotique sans calcification dystrophique (MARRUCHELLA, et al., 2011).
- Pathogénie et symptômes des myocardites
Les inflammations du myocarde proviennent soit de l’extension d’une infection des sacs aériens environnants soit d’une dissémination hématogène (MARRUCHELLA, et al., 2011). Les inflammations du myocarde, quelque soit leur nature, s’accompagnent d’une destruction des myocytes. Une telle destruction provoque : un amincissement pariétal qui favorise l’apparition de rupture du myocarde ou de déficits contractiles, une réaction cicatricielle fibreuses provoquant une cardiomyopathie restrictive (RITCHIE, et al., 1995) (ITCHIE, et al., 1995)et des arythmies, secondaires à une atteinte directe du système de conduction ou bien à la libération du potassium intracellulaire (MCKENZIE, et al., 1972). L’ensemble de ces modifications peut alors évoluer en insuffisance cardiaque congestive si les lésions sont peu importantes (RAE, 1995) (SIMPSON, 1991) (WORK, et al., 2002) (O'BRIEN, et al., 2010). Toutefois, les lésions sont généralement trop importantes et provoquent une mort subite sans symptômes précurseurs (MARRUCHELLA, et al., 2011) (GODOY, et al., 2012) (RAYMOND, et al., 2001) (VAN VLEET, et al., 1986).

Page | 73
d. Cardiomyopathie nutritionnelle
- Carence en vitamine E et Sélénium
La Carence en vitamine E et en Sélénium peut se manifester, entre autres, sous une forme dite « cardiomyopathie nutritionnelle ». Elle est caractérisée par une dégénérescence myocardique (TREMBLAY, et al., 1992) (VAN VLEET, et al., 1986) et a essentiellement été rapportée chez les ratites juvéniles (DONELEY, 2011), les perruches calopsittes (WELLE, et al., 2011), les poules (TREMBLAY, et al., 1992), les dindes et les canards (JULIAN, 1993). Etiologie : La carence en vitamine E et en sélénium dans les aliments et la cause primaire de la déficience. Toutefois on notera que certains facteurs favorisent l’apparition de la maladie tels qu’une augmentation de l’activité cellulaire (stress, activité physique, croissance rapide), un apport alimentaire anormalement élevé en acide gras polyinsaturé (acide linolénique) (TREMBLAY, et al., 1992) ou encore l’ingestion d’antagoniste du sélénium (argent, mercure, cuivre, cobalt, cadmium, zinc, étain, tellurium) (VAN VLEET, et al., 1986) Pathogénie : L’oxygène est vital pour l’organisme mais son utilisation s’accompagne de la production de molécules toxiques pour les cellules et que l’on appelle « radicaux libres ». Plusieurs mécanismes protecteurs sont alloués à la gestion de l’oxygène et des radicaux libres. C’est le cas de certaines enzymes (la cytochrome-oxydase captant l’oxygène avant qu’il ne réagisse de façon indésirable avec des substances sensibles, la gluthation peroxydase ou la superoxide dismutase cytoplasmiques et mitochondriales dégradant les radicaux libre directement sur leur site de production) et d’autres molécules comme la Vitamine E (mais aussi le bêta carotène, le gluthation et la vitamine C) qui récupèrent les radicaux libres. Lors de carence en Vitamine E ou en Sélénium (co-facteurs de la gluthation peroxydase), la toxicité des radicaux libres peut s’exprimer et engendrer notamment des lésions du myocarde (TREMBLAY, et al., 1992). Plus précisément, les radicaux libres provoquent la lipoperoxydation des membranes cellulaires. Il se produit une modification de l’architecture membranaire, une inhibition de l’activité enzymatique membranaire et l’accumulation des produits toxiques de certaines réactions métaboliques. Les effets cumulatifs produisent alors une altération pathologique de la perméabilité des membranes (TREMBLAY, et al., 1992) provoquant la mort cellulaire (JULIAN, 2005). Symptômes : Les atteintes des muscles squelettiques, du gésier ou des intestins sont généralement plus remarquable que celles du myocarde (JULIAN, 1993). Les symptomes cardiaques sont des arythmies (STURKIE, et al., 1954). Lésions : Les lésions du muscle cardiaque sont décrites comme des bandes ou des zones blanches associés résultant d’une dégénérescence hyaline et d’une minéralisation focale du myocarde. On observe fréquemment un hydropéricarde (TREMBLAY, et al., 1992). Diagnostic : Les manifestations cliniques d’une carence en sélénium et en vitamine E sont difficiles à différencier des mycotoxicoses, des problèmes de mauvaise régie de troupeau ou des manques d’homogénéité dans la composition de la ration (TREMBLAY, et al., 1992). L’électrocardiographie montre une déviation de l’axe du cœur à droite, une arythmie sinusale, des contractions ventriculaires prématurées et une élévation du segment ST (STURKIE, et al., 1954). Un diagnostic spécifique de la carence requiert l’aide du laboratoire et la confirmation d’un état de carence devrait être mis en évidence de préférence avant de procéder à un traitement quelconque (TREMBLAY, et al., 1992).

Page | 74
- Carence en sodium
Les volailles carencés en sodium montre une diminution des volumes de fluide plasmatique. Le débit cardiaque chute, la pression artérielle moyenne diminue, l’hématocite augmente (KLASING, 2008). Ceci aboutit à une augmentation drastique des niveaux d’acide urique sanguin (KAHN, et al., 2008). Dans ce cas, un état de choc apparait, qui, s’il n’est pas corrigé aboutit à la mort de l’animal (KLASING, 2008).
- Carence en potassium
La carence en potassium provoque une faiblesse cardiaque. Une hypokaliémie peut également être observée lors de stress sévère. En effet, les protéines plasmatiques sont augmentées forçant le rein, sous l’influence d’hormone surrénalienne, à évacuer du potassium dans l’urine. Au cours de l’adaptation au stress, le flux sanguin musculaire s’améliore et les muscle commence à récupérer du potassium (KAHN, et al., 2008)
- Carence en thiamine
La carence en thiamine peut provoquer des lésions cardiaques chez plusieurs espèces d’oiseaux (VAN VLEET, et al., 1986) comme une atrophie (KAHN, et al., 2008). Ces lésions se manifestent par des arythmies sinusales, des extrasystoles ventriculaires et une diminution du segment ST chez les poulets (ESPINO, et al., 2001). Des pigeons atteints de carence chronique en thiamine ont développé une insuffisance cardiaque associée à une nécrose myocardique (VAN VLEET, et al., 1986) (KLASING, 2008).
- Carence en biotine
La carence en biotine se manifeste entre autre par le syndrome d’infiltration graisseuse du foie et du rein qui affecte les poulets de chair âgés d’une semaine ou plus. La carence de la ration en biotine s’accompagne d’une augmentation de la quantité d’acide gras libres (TREMBLAY, et al., 1992) et engendre une infiltration graisseuse du cœur (RANDALL, et al., 1996) sans dégénérescence ni inflammation (TREMBLAY, et al., 1992). Le jeûne aggrave la condition (KLASING, 2008).
e. Cardiomyopathie génétique : La glycogénose
Les altérations myocardiques lors de maladie de stockage du glycogène ont été rapportées chez les cailles. Celle-ci ont une anomalie de stockage du glycogène de type II (maladie de Pompe, caractérisée par un déficit en amylo-1,6 glucosidase ou maltase acide). Dans cette forme, l’accumulation peut se manifester par une insuffisance cardiaque congestive. Le mode de transmission génétique n’a pas été déterminé (VAN VLEET, et al., 1986).
f. Cardiomyopathie idiopathique
- Cardiomyopathie dilatée idiopathique du dindon
Epidémiologie : La cardiopathie dilatée, connue autrefois sous le nom de « maladie du cœur rond », de « cardiomyopathie spontanée du jeune dindon » ou encore du « syndrome cardiohépatique », est une défaillance cardiaque affectant principalement les jeunes dindonneaux mâles âgés de 1 à 4 semaines (TREMBLAY, et al., 1992), le pic de mortalité étant entre 2 et 3 semaines (CRESPO, et al., 2008). Pathogénèse et étiologie : La cause exacte de la cardiomyopathie dilatée du dindon est inconnue. Des épisodes ont été associées à une hypoxie au cours de l’incubation des oeufs (KAHN, et al., 2008). Les dindes atteintes de cardiomyopathie dilatée montrent une

Page | 75
diminution de certaines enzymes impliquées dans l’apport d’énergie (créatinine kinase, lactate déshydrogénase, système de transport calcique) et une diminution de la concentration en acide gras (le substrat principal du métabolisme cardiaque) provoquant une diminution de la concentration d’ATP (CRESPO, et al., 2008). Ces modifications sont compatibles avec un phénomène ischémique (KAHN, et al., 2008). Les dindes domestiques pourraient être prédisposées génétiquement à cette maladie en raison de la structure anormale de leur troponine T (BIESIADECKI, et al., 2002) ce qui a été confirmée par des essais d’hybridation (CRESPO, et al., 2008). Le métabolisme énergétique anormal du cœur et la structure anormale de la troponine pourraient engendrer des altérations de la contractilité cardiaque (CRESPO, et al., 2008). Il en résulte une dilatation passive du ventricule droit et une hypertrophie compensatrice du ventricule gauche. (TREMBLAY, et al., 1992). Les facteurs augmentant la demande métabolique (froid, hypoxie, taux de croissance élevé) seraient des facteurs prédisposants (JULIAN, et al., 1992) Pendant la période du pic de croissance, l’oiseau est incapable de bien oxygéner ses tissus. Un état d’acidose métabolique s’installe avec comme conséquence, une dégénérescence musculaire. Cet état entraîne une séquestration du calcium et secondairement un état d’hypocalcémie entraînant une défaillance cardiaque (CRESPO, et al., 2008). Symptômes : La plupart des volailles meurent subitement. Dans la forme chronique, l’oiseau est éliminé de l’élevage (TREMBLAY, et al., 1992). On observe dans certains cas un retard de croissance aboutissant à des animaux de petites taille, montrant peu de temps avant la mort des ailes pendantes, des plumes ébourriffées (HUNSAKER, et al., 1971) et des difficultés respiratoires (CRESPO, et al., 2008). Lésions : Macroscopiques : Lors de l’examen post-mortem les dindes affectées ont un cœur de taille considérablement augmentée due à la dilatation des deux ventricules. Souvent, le ventricule droit est davantage dilaté et montre un amincissement de la paroi ventriculaire (DONELEY, 2011). On observe également une hypertrophie du ventricule gauche (TREMBLAY, et al., 1992). Un hydropéricarde et de l’ascite peuvent être présents (HUNSAKER, et al., 1971). Les poumons sont généralement congestionnés et oedèmateux. Le foie peut être légèrement oedèmateux avec des bords mousses. Microscopiques : Les modifications sont non spécifiques et incluent une congestion, une dégénération des myofibres, une infiltration lymphocytaire focale. Chez les dindes les plus âgées on observe un épaississement ferme et blanchâtre de l’endocarde du ventricule gauche correspondant à une augmentation de la quantité de tissu fibroélastique (CRESPO, et al., 2008). Celle-ci est vraisemblablement secondaire à une diminution de la perfusion sous-endocardique (VAN VLEET, et al., 1986). On observe un dégénérescence myocardique. Les fibres affectées sont oedemaciées et vacuolisées. (RANDALL, et al., 1996). Diagnostic différentiel : L’intoxication à la furazolidone produit un syndrome similaire à la CMD. Toutefois, la concentration d’acide gras polyinstaurés lors de dilatation induite par la furazolidone est augmentée alors que la concentration de ces acides est notablement diminuée lors de CMD. L’échocardiographie révèle une augmentation du volume télédiastolique et une diminution de la fraction d’éjection ainsi que de la pression sanguine systolique (CRESPO, et al., 2008). Lors d’intoxication à la furazolidone, on observe une dilatation des chambres cardiaques mais sans hypertrophie et sans hyperélastose endocardique (VAN VLEET, et al., 1986). L’intoxication au sodium et au biphényls polychlorés produisent un syndrome similaire (KAHN, et al., 2008).

Page | 76
Diagnostic : Généralement, le diagnostic repose sur l’anamnèse et les trouvailles nécropsiques (KAHN, et al., 2008). A la nécropsie on observe un élargissement cardiaque due à une dilatation du ventricule droit associé à un amincissement de la paroi ventriculaire. Le ratio poids des ventricules/poids du corps est d’ailleurs significativement augmenté (0.80 lors de maladie contre 0.45 chez des sujets sains).. L’électrocardiographie peut être utilisée mais elle est généralement de moindre intérêt (KAHN, et al., 2008). Les modifications sévères de la forme du cœur ainsi que son déplacement provoque les modifications électrocardiographiques suivantes : une onde R et une onde P d’amplitude augmentées associées à une onde T négative dans toutes les dérivations. L’onde S est petite à absente. Toutefois de telles modifications ne sont pas toujours évidentes. L’axe électrique moyen est aux alentours de +70° contre -85 pour des dindes normales (HUNSAKER, et al., 1971). La précision de l’ECG pour la détection d’une maladie du cœur rond est de 80% (CZARNEKI, et al., 1980). L’intérêt de l’échocardiographie a été également démontré (GYENAI, et al., 2012).
- Syndrome de la mort subite du poulet de chair
Epidémiologie : Le syndrome de la mort subite affecte les poulets de chair dès l’âge de deux jours à une incidence de 0,5 à 4% et cela représente 3 à 15% de mortalité totale d’un troupeau. Il atteint un pic vers 2 semaines. Les poulets mâles présentant un fort taux de croissance ont plus de risques à en être affectés (TREMBLAY, et al., 1992). Etio-pathogénie : La mort subite est le résultat d’une arythmie ventriculaire mortelle déclenchée par un stress (bruit, luminosité intense, variation de température) (TREMBLAY, et al., 1992). Toutefois, la cause prédisposant à l’apparition de cette arythmie est inconnue (KAHN, et al., 2008). L’hypothèse principale serait liée à la rapidité de croissance des oiseaux sélectionnée génétiquement. En effet, la croissance rapide provoque une situation d’hypercapnie et d’hypo-oxygénation favorable au développement d’arrythmies cardiaques (OLKOWSKI, 2007). Une maladie métabolique est également suspectée. Le métabolisme des carbohydrates pourrait être altéré, conduisant à une acidose lactique, une perte de l’intégrité de la membrane cellulaire aboutissant à un débalancement électrolytique (KAHN, et al., 2008) (TREMBLAY, et al., 1992). Enfin une augmentation de l’activité de métalloprotéinase provoquerait des adhérences entre le myocarde et le péricarde aggravant les troubles de la conduction du fait de l’existence sous-épicardique du tissu cardionecteur (OLKOWSKI, et al., 2003). Lorsque ces arrythmies deviennent trop sévère, on observe alors une insuffisance cardiaque aiguë et une mort subite (OLKOWSKI, 2007). Symptômes : L’oiseau atteint paraît en bon état général puis, soudainement, il présente des convulsions, se débat et meurt (TREMBLAY, et al., 1992) sur le dos (KAHN, et al., 2008) Diagnostic : La mort subite d’oiseau d’environ 7 jours retrouvés sur le dos associés à une absence de lésions spécifiques (poumons œdematiés et congestionnés, jabot et gésier remplis d’aliments, intestin contenant des matières crémeuses) sont les éléments de base du diagnostic (TREMBLAY, et al., 1992). L’examen histopathologique révèle des modifications dégénératives des cardiomyocytes du ventricule gauche ainsi que du système de His-Purkinje qui sont caractéristiques (KAHN, et al., 2008).

Page | 77
g. Néoplasie myocardique
Les lymphomes cardiaques peuvent être primitifs ou métastatiques. Des lymphomes cardiaques métastatiques ont été observés chez 60% d’une population de poulets atteints de leucose lymphoïde expérimentale. Ces tumeurs semblent s’être développées à partir des nodules lymphoïdes (GILKA, et al., 1990). Des lymphomes cardiaques ont également été observé chez des animaux atteints de maladie de Marek (MAJO, et al., 2012) et de réticuloendothéliose (KAHN, et al., 2008). Dans tous les cas ils se présentent comme des lésions blanche, jaunâtre ou grisâtre diffuses ou multinodulaires constituées de cellules lymphoïdes pléomorphiques mal différenciées (SCHMIDT, et al., 2003). Les rhabdomyomes et rhabdomyosarcomes forment des lésions pâles et fermes pouvant être multiples. Le diagnostic est apporté par l’immunohistochimie qui prouvera que la tumeur provient de cellules striées squelettiques. Les rhabdomyosarcomes infiltrent le myocarde (SCHMIDT, et al., 2003). Le développement de tumeurs musculaire squelettique a été relié à l’infection par des virus oncogénique chez le poulet (DUNCAN, et al., 1997) (LATIMER, 1994). Des fibrosarcomes primaires ont été décrits dans le myocarde. Ces tumeurs prennent l’apparence de masses fermes grises-blanches constituées d’un entrelacement de faisceau de fibroblastes (SCHMIDT, et al., 2003). Des mélanomes malins peuvent être rencontrés dans le cœur aviaire et sont habituellement métastatiques. Ces tumeurs sont marrons-noires, possiblement multiples et sont constituées de mélanocytes mal différenciés formant des amas et des feuillets dans le myocarde. Quelque soit leur nature, les tumeurs cardiaques peuvent diminuer le diamètre de la lumière des chambres cardiaques, interférer avec la contractilité et/ou altérer la conduction et aboutissent à une insuffisance myocardique (LATIMER, 1994).
h. Démarche diagnostique lors d’affection myocardique
Les symptômes observées sont non spécifiques et traduisent, lorsqu’ils existent, l’existence d’une insuffisance cardiaque. Les clichés radiographique peuvent permettre de mettre en évidence des lésions lors de l’existence de granulome. Toutefois comme les granulomes ne se minéralisent pas chez les oiseaux, il peut être difficile de distinguer les lésions provoqué par une mycobactérie de celles produites par de l’aspergillose ou une néoplasie. (DAHLHAUSEN, et al., 2012). Elle peut également révéler des modifications non spécifiques secondaires à une cardiomégalie (DE WIT, et al., 2005). Lors de masses cardiaques, la réalisation d’une cytologie ou d’une histologie est nécessaire pour différencier un granulome d’une néoplasie. L’électrocardiographie pourra révéler l’existence d’un trouble du rythme (GELIS, et al., 2006) (HUNSAKER, et al., 1971) en effet, toutes les conditions aboutissants à une cardiomyopathie peuvent entraîner une augmentation de l’irritabilité cardiaque (JONES, 2008). L’échocardiographie est le meilleur outil diagnostique chez les oiseaux atteints de cardiomyopathies (KRAUTWALD-JUNGHANNS, et al., 1995), puisqu’elle permet d’évaluer la dilatation des chambres cardiaques, l’épaisseur de la musculature ventriculaire et la fonction contractile du myocarde (PEES, et al., 2006). De façon non spécifique, on pourra observer des signes d’insuffisance cardiaque. Une biochimie sanguine évaluant les AsAT, CK, acide urique, LDH et électrolytes peut être réalisée lorsqu’une cardiomyopathie est suspectée (PEES, et al., 2006). Le recours à des méthodes permettant l’identification d’un agent infectieux pourra être utile. Des méthodes immunohistochimiques (SWAYNE, et al., 2001) (RAYMOND, et al., 2001), histopathologiques (JUNIOR, et al., 2012), sérologique, PCR l’utilisation de kit de détection antigénique (ECHOLS, et al., 2000) D’autres techniques comme la PCR (VAPNIARSKY, et al., 2012), la sérologie (AKANBI, et al., 2008), la culture sanguine, ou coloration de gram pourront permettre de préciser l’identité agent étiologique (GALINDO-CARDIEL, et al., 2011), RT-PCR (O'BRIEN, et al., 2010). (PALMIERI, et al., 2011).

Page | 78
D. Affections du péricardique
Les atteintes péricardiques sont essentiellement représentées par les péricardites et les épanchements. En particulier, les néoplasies péricardiques n’ont pas été décrites en médecine aviaire (SCHMIDT, et al., 2003).
1. Etiologie
- Péricardite métabolique
Etiologie : La goutte viscérale est une maladie que l’on peut observer lors d’insuffisance rénale ou lorsque les oiseaux sont privés d’eau (RANDALL, et al., 1996). Pathogénie : L’accumulation d’acide urique dans le plasma aboutit à sa cristallisation et au dépôt de tophi d’urates sur les organes. La goutte viscérale peut engendrer une péricardite non infectieuse (DONELEY, 2011). Si tel est le cas, des cellules inflammatoires nécrotiques sont présentes sous le dépôt. On parle de pericarditis urica. Dans la plupart des cas, la mort de l’animal survient avant l’installation d’une réponse inflammatoire (JONES, 2008). Lésions : Le dépôt d’acide urique dans le péricarde peut être visible lors de goutte viscérale (PEES, et al., 2006). Microscopiquement on observe du matériel basophile duveteux à la surface de l’épicarde qui représente le site de déposition des cristaux d’urate.
- Péricardite infectieuse
Etiologie : Les agents infectieux impliqués peuvent être bactériens, tels que E.Coli (RANDALL, et al., 1996), Streptococcus spp., Riemerella anatipestifer, Listeria spp., Salmonella, Chlamydia (JOHNS, et al., 2009), Mycoplasma, Mycobacterium (GELIS, et al., 2006). Ils peuvent également être mycosiques (aspergillose), parasitaire (trichomonas (CHARLTON, et al., 1991) , microfilaires (GREINER, et al., 1994)) ou viraux (réovirose chez les galliformes et polyomavirose chez les perroquets (DONELEY, 2011)). Epidémiologie : Un enduit séro-fibrineux recouvre le péricarde chez 15% des psittaciformes nécropsiés (KRAUTWALD-JUNGHANNS, et al., 2004). Les péricardites infectieuses ont été décrites chez des psittaciformes (DONELEY, 2011), columbiformes (CHARLTON, et al., 1991), volailles (RANDALL, et al., 1996) et rapaces (JOHNS, et al., 2009). Pathogénie : L’infection du péricarde peut se faire par extension locale d’une myocardite ou d’une infection de l’appareil aérien (aspergillose, E. Coli) ou bien par diffusion hématogène d’une maladie systémique (trichomonas) (PEES, et al., 2006). Selon l’agent considéré l’inflammation du péricarde pourra être sèche, fibrineuse, séro-fibrineuse ou séreuse. Les péricardite fibrineuse, fibrino-purulente ou séro-fibrineuse sont les plus fréquemment rencontrées. L’inflammation du péricarde peut engendrer une péricardite constrictive. La péricardite constrictive correspond à un ensemble de lésion perturbant la relaxation ventriculaire et le remplissage cardiaque. Chez les oiseaux, deux types de lésions constrictives ont été décrites. La péricardite fibreuse chronique constrictive correspond à un épaississement fibreux du péricarde secondaire à une inflammation chronique. La péricardite adhésive constrictive correspond à la formation de brides fibreuses entre les feuillets péricardiques suite à un exsudat purulent, fibrineux ou sanguin (STRUNK, 2003). Symptômes : Une péricardite constrictive se manifeste par les symptômes classiques d’une insuffisance cardiaque (LUMEIJ, et al., 1994). Lésions : L’apparence macroscopique dépend de la composition du fluide. Après Quelques heures après la mort, un épanchement riche en protéine devient gélatineux (SCHMIDT, et al., 2003)

Page | 79
- Epanchement péricardique
Etiologie : Un épanchement péricardique hémorragique (hémopéricarde) peut résulter d’un trauma, d’une coagulopathie ou d’une rupture myocardique, elle-même secondaire à une myocardiopathie (néoplasie (DONELEY, 2011), myocardite (JONES, 2008), dégénérescence (DOMINGO, et al., 1991) (SATO, et al., 2000)) ou idiopathique (BOUGIOUKLIS, et al., 2005). Un épanchement péricardique séreux (hydropéricarde) peut être secondaire à une insuffisance cardiaque congestive, une péricardite infectieuse virale (fowl adenovirus, reovirus, polyomavirus), une intoxication à l’avocat (HARGIS, et al., 1989) ou à une hypoalbuminémie (DONELEY, 2011). Dans certains cas, aucune cause sous-jacente n’est mise en évidence et on parle d’épanchement péricardique idiopathique. Epidémiologie : Une étude révèle que 6% des psittaciformes nécropsiés présentent un épanchement péricardique (KRAUTWALD-JUNGHANNS, et al., 2004). Un hémopéricarde a été observé chez des poussins suite à une rupture atriale (BOUGIOUKLIS, et al., 2005) et chez des tétras (Tetrao urogallus) suite à une rupture de l’apex cardiaque (SATO, et al., 2000). Pathogénie : L’existence d’un épanchement peut provoquer une tamponnade cardiaque. La tamponade correspond à une accumulation aiguë de fluide dans la cavité péricardique. Il peut s’agir d’un épanchement mécanique, inflammatoire, sanguin ou idiopathique. (LUMEIJ, et al., 1994) (DE WIT, et al., 2005) (PEES, et al., 2006). Une telle accumulation provoque une compression externe sur le muscle cardiaque ce qui limite progressivement le remplissage, et diminue en conséquence le débit cardiaque. On parle de tamponnade cardiaque et les symptômes sont ceux d’une insuffisance cardiaque (DE WIT, et al., 2005). Symptômes : Les symptômes d’un épanchement péricardique sont typiquement non spécifiques. Lorsque l’affection provoque une tamponnade, il est alors possible d’observer un état léthargique, une moindre tolérance à l’exercice et une dyspnée (STRUNK, 2003). Lésions : On observe un épanchement péricardique dont les caractéristique dépendent de la cause sous-jacente éventuellement associée à des lésions d’insuffisance cardiaque.
2. Démarche diagnostique lors d’affection péricardique
Les symptômes cliniques de péricardite sont non spécifiques. La radiographie peut révéler un élargissement de la silhouette cardiaque ainsi qu’une perte de détail (STRAUB, et al., 2001). L’échocardiographie est alors utilisée afin de différencier un épanchement péricardique d’une cardiomégalie (l’hydropéricarde peut être visualisé comme une région anéchogène entre le myocarde et le péricarde) (DE WIT, et al., 2005). Toutefois elle est d’intérêt limité en l’absence d’épanchement péricardique. L’endoscopie permet d’évaluer l’aspect du péricarde et de réaliser des prélèvements. C’est l’outil diagnostic le plus fiable ante mortem, bien qu’il y ai un risque accru au cours de l’anesthésie (FITZGERLAD, et al., 2011). En cas de goute viscéral, les niveaux biochimiques d’acide urique plasmatique peuvent être augmentés (PEES, et al., 2006). L’électrocardiographie peut révéler un ECG de bas voltage chez de tels patients (DE WIT, et al., 2005) mais n’apporte qu’un diagnostic de suspicion (le dépôt de tissu adipeux ou un processus volumineux comme une tumeur peut également provoquer une diminution du voltage de l’ECG) (PEES, et al., 2006). Pour davantage d’examen (cytologie, microbiologie) une aspiration échoguidée du sac péricardique peut être réalisée. Dans la plupart des cas, l’épanchement péricardique est non infectieux et d’aspect clair. Néanmoins, la présence de cellules inflammatoires peuvent indiquer une infection et la culture permettra d’identifier la cause bactérienne ou fongique sous jacente. Le diagnostic est généralement confirmé par nécropsie (PEES, et al., 2006).

Page | 80
E. Maladies cardiaques congénitales
1. Facteurs tératogènes
Les anomalies cardiaques chez les oiseaux ont été décrites comme étant secondaires à une diminution de la température ou à une hypoxie au cours de l’incubation. L’hypoxie pourrait engendrer l’apparition d’anomalies cardiaques en augmentant l’expression du facteur de croissance endothélial vasculaire (VEGF) et les températures froides pourraient engendrer des anomalies congénitales en augmentant le métabolisme basal et donc le fonctionnement cardiaque (RISI, et al., 2011). Ces anomalies pourraient également être secondaires à des déficiences nutritionnelles maternelles (acide nicotinique, vitamine B12, vitamine D) (PENCE, 1996). L’exposition à certaines molécules (toxiques comme les organochlorés, médicaments) provoque également des anomalies congénitales cardiaques (EVANS, et al., 2001).
2. Anomalies congénitales cardio-vasculaires
Les communications inter-ventriculaires sont les anomalies les plus fréquemment rapportées. Des cas ont été décrits chez des cacatoès blancs de 13 semaines (EVANS, et al., 2001), une outarde houbara juvénile (Chlamydotis undulata macqueenii) (BAILEY, et al., 201), des poulets (SILLER, 1958), des dindes (EINZIG, et al., 1972) et un cygne de Bewick de 7 ans (Cygnus columbianus) (HARARI, et al., 1983). Un anévrisme cardiaque congénital vrai (défaut musculaire) du ventricule droit a été décrit chez un pigeon de 7 mois (GAL, et al., 2012). Un shunt artério-veineux entre l’aorte et le ventricule droit a été rapporté chez une autruche adulte (BAPTISTE, et al., 1997). Une persistance du foramen ovale a été décrite chez une dinde (EINZIG, et al., 1972) et une communication inter-atrial a été observée chez un vautour griffon de 6 semaines, des pigeons et des poulets (RISI, et al., 2011). Une sténose mitrale subvalvulaire a été décrite chez un canard de 2,6 ans. Des duplicita cordis, multiplicatis cordis et ectopia cordis ont également été rapportés (DONELEY, 2011). Ont été décrits associées à des communications inter-ventriculaires une persistance du tronc artériel ou une hypoplasie aortique chez des cacatoès (EVANS, et al., 2001) Les turbulences associées aux anomalies congénitales peuvent provoquer des lésions endothéliales et favoriser l’apparition d’endocardite bactérienne lors de bactériémie (HARARI, et al., 1983) (SILLER, 1958).
3. Symptomatologie
La plupart des cas se soldent par la mort à un âge jeune (STRUNK, 2003). En effet, du fait de la charge de travail physiologique élevée chez les oiseaux, les anomalies congénitales sont rarement viables et l’on estime qu’elles sont la cause probable d’une mortalité embryonnaire précoce ou juvénile (KRAUTWALD-JUNGHANNS, et al., 2004). Toutefois, dans le cas d’anomalie peu importante, chez des oiseaux de compagnie sédentaire, la maladie peut passer inaperçue pendant un certain nombre d’année. A terme, elle peut s’exprimer lorsque le cœur devient insuffisant (PEES, et al., 2001). La symptomatologie correspond habituellement à une insuffisance cardiaque congestive (LUMEIJ, et al., 1994). L’examen physique révélera fréquemment l’existence d’un souffle dont les caractéristiques pourront orienter le clinicien vers une anomalie du cœur. Ainsi ont été décrits un souffle IV/VI systolique à gauche lors de sténose mitrale (MITCHELL, et al., 2008), un souffle de grade IV/VI systolique audible à gauche et moindrement à droite lors de shunt artério-veineux (BAPTISTE, et al., 1997) et un souffle de grade III-IV/VI systolique apexien gauche lors de communication inter-ventriculaire (EVANS, et al., 2001). Des anomalies pulsatiles comme un pouls bondissant ont été rapportées lors de communication inter-ventriculaire (EVANS, et al., 2001).

Page | 81
4. Diagnostic des maladies congénitales
Le diagnostic est réalisé à l’aide d’une échographie transcoelomique (turbulence visibles en mode doppler couleur, modification de la vélocité détectée avec le mode doppler spectral, signes de congestion avec le mode B) (MITCHELL, et al., 2008) et peut être confirmé par la nécropsie (BAPTISTE, et al., 1997). La radiographie montre fréquemment des signes d’insuffisance cardiaque. L’ECG, les bilans hématologique et biochimique sont habituellement normaux (MITCHELL, et al., 2008).
II. Affections vasculaires
A. Vasculopathies métaboliques
- Athérosclérose Définition : L’athérosclérose est une maladie vasculaire métabolique, inflammatoire et dégénérative (BEAUFRERE, et al., 2011). L’athérosclérose est probablement l’affection cardio-vasculaire la plus fréquente et correspond à l’une des maladies ayant les plus grave répercussions sur la santé des individus adultes à âgés (WELLE, et al., 2011). Sa prévalence et sa gravité chez les oiseaux semblent être supérieures à celle de nimporte quelle mammifère, à l’exception de l’homme (BAVELAAR, et al., 2004). Etiologie : Aucun facteur déterminant n’a été mis en évidence et peu d’éléments sont connus relativement aux facteurs de risque chez les perroquets (BAVELAAR, et al., 2004). Epidémiologie analytique et facteurs de risques : Influence de l’espèce : Les perroquets comme, le gris du Gabon (Psittacus erithacus), les Amazones (Amazona spp.) et les perruches callopsites (Nymphicus hollandicus) semblent prédisposés au développement de lésions athérosclérotiques. Ceci est en faveur d’un rôle de l’hérédité dans l’apparition des lésions (BEAUFRERE, et al., 2013). Ce facteur héréditaire est appuyé par le fait que la dinde et le poulet seraient prédisposés à développer spontanément des lésions d’athérosclérose en l’absence d’apport de cholestérol dans l’alimentation. De même, il existe des souches résistantes et des souches sensibles à l’athérosclérose chez la caille japonaise et le pigeon (SAINT CLAIR, 1998). Bien que certaines espèces soient prédisposées, cette affection a été décrite chez un grand nombre d’oiseaux tels que des psittaciformes (BAVELAAR, et al., 2003) (BOHORQUEZ, et al., 1972) (GRINER) (JOHNSON, et al., 1992) (PHALEN, et al., 1996), des cuculiformes (WAINE, et al., 2002), des falconiformes et des strigiformes (SHRUBSOLE-COCKWILL, et al., 2008) (BOHORQUEZ, et al., 1972) (COOPER, et al., 1982) (JOHNSON, et al., 1992), des struthioniformes, des piciformes (BOHORQUEZ, et al., 1972) (JOHNSON, et al., 1992), des passériformes (JOHNSON, et al., 1992), des columbiformes (LEWIS, et al., 1997), des sphénisciformes, des pelecaniformes, des casuariformes, des rheiformes, des ciconiiformes, des ansériformes, des gruiformes, des coraciiformes et des galiformes (BOHORQUEZ, et al., 1972). Influence de l’âge : Les oiseaux âgés seraient davantage prédisposés à développer de l’athérosclérose (BAVELAAR, et al., 2004). Le risque augmente considérablement aux environs de 20-30 ans (BEAUFRERE, et al., 2013). Influence du sexe : Les femelles sont 4 fois plus prédisposées au développement des lésions que ne le sont les mâles (BEAUFRERE, et al., 2013). Influence des comportements sociaux : L’inactivité chez les poulets a été corrélée à une augmentation de l’athérosclérose alors qu’un exercice forcé semble protéger contre le développement de lésions (WARNOCK, et al., 1957). Par ailleurs, le stress social provoque un débalancement de la sécrétion adrénergique qui favoriserait le développent

Page | 82
d’athérosclérose (RATCLIFFE, 1966). Les grands perroquets en zoo développeraient davantage de lésions par rapport aux petits, en relation avec le stress social résultant des difficultés qu’ils éprouvent à exprimer un comportement naturel (RATCLIFFE, et al., 1958). Influence de l’alimentation : Trois composants alimentaires entrent en jeu dans la formation d’athérosclérose : le cholestérol, les acides gras et les antioxydants. Plus précisément, des auteurs ont mis en évidence l’existence d’une relation linéaire entre le cholestérol plasmatique et la sévérité des lésions d’athérosclérose (BAVELAAR, et al., 2004). Une diète alimentaire riche en acides gras saturés augmente significativement le niveau de cholestérol plasmatique par rapport à une alimentation à une alimentation pauvre en graisse ou riche en acide gras n-6 polyinsaturés (BAVELAAR, 2003). Une carence en vitamine A augmente la sévérité et l’incidence des lésions athérosclérotiques (VINK-NOOTEBOOM, et al., 1998). Influence de maladies concomitantes : Certaines études montrent que l’athérosclérose pourrait être secondaire à une infection par un herpèsvirus chez les volailles (virus de la maladie de Marek chez les volailles) (FABRICANT, et al., 1999). Aucun agent étiologique n’a été mis en évidence chez les psittacidés (BEAUFRERE, et al., 2010). En particulier, l’implication de C.psittaci dans l’apparition d’athérosclérose a été réfuté (SCHENKER, et al., 2007). Certains auteurs rapportent une association avec les pontes chroniques (BAINE, 2012) et l’hypertension (MILLER, 1985). Pathogénie : Les lésions athérosclérotiques résultent d’un déséquilibre du flux de cholestérol trans-pariétal caractérisé par des afflux supérieurs aux efflux. (BAVELAAR, et al., 2004). Ce déséquilibre est initié par un dysfonctionnement endothélial associé aux facteurs de risque décrits précédemment (BEAUFRERE, et al., 2011) et engendre un dépôt de fibrinogène et de lipoprotéines de faible densité dans la paroi artérielle. On observe alors un épaississement de l’intima secondaire à l’accumulation. Les lipoprotéines sont alors oxydées par les déchets du métabolisme des cellules de la paroi artérielle. Ces lipoprotéines oxydées engendrent alors le recrutement de cellules inflammatoires (macrophages) pour lesquelles elles sont toxiques ce qui aboutit à l’accumulation de cellules spumeuses (BERLINER, 1995). A ce moment, on parle du stade de « strie lipidique ». Les macrophages produisent divers cytokines à l’origine d’une inflammation chronique et d’une destruction de la matrice extracellulaire (PAUL, et al., 2009). L’accumulation de cellules spumeuses conduit à la formation d’un noyau lipique (le centre athéromateux) recouvert d’une chape fibreuse constituée de cellules musculaires lisses dédifférenciées sécrétant des protéines de la matrice extracellulaire. Ce stade est dénommée « plaque fibro-lipidique ». La nécrose et l’apoptose des macrophages contribue à la croissance du noyau. L’évolution de la plaque peut engendrer :
Une sténose vasculaire : Initialement, le développement de la plaque n’entraîne pas de diminution du diamètre luminal car il est compensé par une dilatation des artères (TOUSSAINT, et al., 2003). Cette compensation est efficace tant que la masse intimale reste inférieure à 40% de la surface pariétale (PAUL, et al., 2009). Un anévrisme : Dans certains cas, il peut se produire une destruction des composants de la matrice extracellulaire de la plaque. Dans ce cas, les portions de vaisseaux atteintes évoluent vers la formation d’anévrisme. L’anévrisme peut comprimer des structures environnantes notamment nerveuses. Il pourra également se rompre et provoquer une hémorragie (BAVELAAR, et al., 2004). Une thrombose : Lorsque la plaque se rompt ou que l’endothélium qui la recouvre est érodé le sang est en contact avec du matériel thrombogène ce qui aboutit à la formation d’un thrombus luminal. Le thrombus peut être sub-occlusif auquel cas il est asymptomatique ou occlusif aigu, entraînant alors une ischémie aiguë en l’absence d’irrigation collatérale. Il peut également aboutir à la formation d’une embolie.

Page | 83
Ulcère et hémorragies : Les ulcères ont les mêmes conséquences thrombotiques que la rupture. Les hémorragies entrainent une augmentation brutale du volume de la plaque et donc un rétrécissement de la lumière artérielle. Calcification : Au cours de leur évolution, le noyau lipidique ou le tissu conjonctif des plaques peuvent subir une calcification dystrophique par de l’hydroxyapatite. On parle alors de plaque fibrocalcaire. La calcification rend la plaque rigide, peu déformable et davantage susceptible de se rompre.
Symptômes : Lors d’athérosclérose, les symptômes sont rarement rapportés car la manifestation la plus fréquente est la mort subite (BAVELAAR, et al., 2004). Cette mort est secondaire à la rupture d’un anévrisme disséquant chez les poules et les canards hypertendus (DONELEY, 2011). Elle peut également faire suite à une fibrillation cardiaque consécutive à une hypoxie myocardique elle-même secondaire à une sténose ou à un infarctus (embole athéromateuse se détachant des plaques) (MANS, et al., 2007). Toutefois, lorsqu’il y a des symptômes cliniques, ceux-ci sont souvent subtils, intermittents et peuvent être attribués :
- A une diminution du flux sanguin aux tissus : Selon la localisation des lésions, la diminution de la perfusion périphérique se manifeste essentiellement par des déficits neurologiques tels que de l’ataxie (BEAUFRERE, et al., 2011), des pertes d’équilibre, des chutes, une parésie (PHALEN, et al., 1996) pouvant évoluer en paralysie (BAVELAAR, et al., 2004), des collapsus (VINK-NOOTEBOOM, et al., 1998), des crises convulsives (JOHNSON, et al., 1992), de l’opisthotonos, de la cécité (DELK, 2012) ou des tremblements (DE WIT, et al., 2005) mais aussi par le syndrome de claudication intermittente (BEAUFRERE, et al., 2011). Le syndrome de claudication intermittente représente une boiterie répétable, apparaissant à l’effort et disparaissant au repos. En médecine humaine, ce syndrome est un signe d’appel de sténose sévère (80-90% chez les oiseaux) de l’aorte, de l’iliaque ou des artères fémorales et consiste un facteur pronostic négatif. Dans cette situation, l’hypoperfusion musculaire entraîne douleur, crampe et faiblesse (BEAUFRERE, et al., 2011). - A une insuffisance cardiaque congestive gauche (PHALEN, et al., 1996) (MANS, et al., 2007) (VINK-NOOTEBOOM, et al., 1998) ou plus rarement globale (MANS, et al., 2007) résultant de l’augmentation de la charge cardiaque (BAVELAAR, et al., 2004) (PEES, et al., 2006) (SEDACCA, et al., 2009) (SIMONE-FREILICHER, 2007) - A la compression veineuse par un anévrisme. Ainsi, un anévrisme de l’artère coronaire droite a provoqué la compression de la veine cave caudale d’un cacatoès aboutissant à une congestion de cette veine et donc à un épanchement péricardique et de l’ascite. Il s’agit d’une situation plus occasionnelle (VINK-NOOTEBOOM, et al., 1998).
Signalons que la grande majorité des oiseaux de compagnie étant confiné dans des cages, il n’est pas surprenant que ceux-ci soient asymptomatiques jusqu’à ce qu’ils soient stressés et décompensent de façon aiguë (BAVELAAR, et al., 2004). Lésions : Aspect des lésions : Macroscopiquement la paroi artérielle des grosses artères et de leur branches principales (jusqu’aux artères fémorales) atteintes d’athérosclérose apparaissent irrégulièrement épaissies et de couleur jaune pâle avec une diminution de la taille de la lumière (BEAUFRERE, et al., 2011). La paroi contient des plaques intimales grossières et jaunes (SCHMIDT, et al., 2003). Les lésions sont plus diffuse chez les oiseaux que chez l’homme. Les lésions compliquées sont rares et les ruptures de plaques restent exceptionnelles. Il semblerait que les perroquets ne montrent que peu d’anévrismes (St

Page | 84
LEGER, 2007), d’érosion et que la thrombose n’existe pas (BAVELAAR, et al., 2004). Certains auteurs estiment que les thrombi muraux sont rares chez les oiseaux ce qui pourrait expliquer l’absences de complications ischémiques chez ces espèces (BOHORQUEZ, et al., 1972). Localisation des lésions : Macroscopiques : Les oiseaux sont sujet à une forme centrale d’athérosclérose, ce qui signifie que les processus athérosclérotiques apparaissent et se limitent aux portions de la circulation systémique les plus proches du cœur (DE WIT, et al., 2005)) (St LEGER, 2007). Ainsi, le site de prédilection de l’athérosclérose, est l’aorte à la base du cœur (St LEGER, 2007) (BAVELAAR, et al., 2004) (PILNY, 2004) et l’aorte descendante est habituellement dépourvue de lésion athérosclérotique (BAVELAAR, et al., 2004). Si les lésions modérées à avancées sont effectivement essentiellement retrouvées à la base de l’aorte, elles peuvent toutefois s’étendre dans les artères brachiocéphaliques et les artères coronaires voire dans les carotides, les artères pulmonaires (SEDACCA, et al., 2009) et les artères émises en descendant le long des vaisseaux thoraciques et abdominaux (artères cœliaque, artères iliaques, artères fémorales, artères spléniques, artères gastriques (JOHNSON, et al., 1992)) système coronarien (BAVELAAR, et al., 2004) (SHRUBSOLE-COCKWILL, et al., 2008) où elles sont généralement de gravité plus faible (BEAUFRERE, et al., 2011). Toutefois, cette distribution générale connait certaines exceptions. Ainsi, chez le pigeon, l’athérosclérose se développe d’abord dans l’aorte, au niveau de la bifurcation avec l’artère cœliaque puis d’autres sites peuvent être atteints secondairement, notamment le système coronarien. Toutefois, ce sont les petites artères intramyocardiques qui sont affectées plutôt que les branches principales de la base du cœur comme c’est le cas chez l’homme. Chez la dinde, l’aorte abdominale est davantage affectée par les lésions athérosclérotiques. Ces variabilités de site par rapport au schéma général pourraient suggérer l’existence de causes différentes (SAINT CLAIR, 1998). Les dindes sont également prédisposées à faire des anévrismes aortiques secondaire à l’athérosclérose (St LEGER, 2007). L’athérosclérose des artères coronaires, des carotides et des autres artères périphériques a été rarement documenté chez les espèces aviaires (MUELLER, et al., 1980) (VINK-NOOTEBOOM, et al., 1998) (BENETT, 1994). Les variations de perméabilité vasculaire et de pression artérielle sanguine pourraient expliquer la prédisposition de certaines régions au développement de lésions athérosclérotiques (DONELEY, 2011). Microscopique : La nécropsie révèle des modifications dégénératives de la paroi artérielle associée à une diminution de la lumière, avec dépôt de collagène, cholestérol, calcium parfois même en l’absence de lésions macroscopiques(SHRUBSOLE-COCKWILL, et al., 2008). Bien qu’il n’y ai pas de réel consensus, les dernières études classent les lésions microscopiques en 7 catégories, selon leur importance. Le type I est caractérisé par une augmentation de la présence des macrophages et l’existence de cellules spumeuses isolées, ainsi que de la quantité de matrice extra-cellulaire de l’intima. On observe de plus une vacuolisation des cellules endothéliales, la présence de lipides dans l’intima et la présence de cellules musculaires lisses riches en RER. Le type II est défini par l’accumulation de macrophages et de cellules spumeuses qui se répartissent en feuillet ainsi que d’une petite quantité de lipides extracellulaires. Les lésions modérées de type III sont caractérisées par l’existence de ‘’pré-athérome’’. On observe ainsi l’augmentation de lipides extracellulaires. Lésions avancées de type IV et V : le type IV est caractérisé par la présence d’athérome alors que le type V est caractérisé par la présence de fibroathérome. Les lésions compliquées correspondent au type VI. Lorsqu’elles sont très avancée, on parlera de type VII (BEAUFRERE, et al., 2011).

Page | 85
Diagnostic : Les signes cliniques sont souvent absents, subtiles (DE WIT, et al., 2005) ou masqués par l’existence d’une maladie concomitante dans 50% des cas (BAVELAAR, et al., 2004). En conséquence, le diagnostic d’athérosclérose est très difficile du vivant de l’animal (BEAUFRERE, et al., 2010) et celui-ci est le plus souvent réalisé post-mortem (PHILILIPS, 1986). Le diagnostic n’est réalisé de façon ante-mortem que lors de cas avancé avec une calcification des artères ou en association avec une insuffisance cardiaque congestive (MANS, et al., 2007) (SEDACCA, et al., 2009) (SIMONE-FREILICHER, 2007). Auscultation : On peut entendre un souffle ou des irrégularités du rythme cardiaque, bien que ces éléments soient rarement rapportés chez les oiseaux. Un souffle cardiaque peut être confirmé avec l’aide de la phonocardiographie (BAVELAAR, et al., 2004). Biochimie : La réalisation d’un bilan lipidique peut aider à mettre en évidence l’existence de facteurs de risques, mais ne peut en aucun cas diagnostiquer les lésions vasculaires (WELLE, et al., 2011). Lors d’athérosclérose, les niveaux de cholestérol peuvent être augmentés ou normaux (DONELEY, 2011). Une étude de PILNY (2004) rapporté par (MANS, et al., 2007) aurait révélé que les oiseaux atteints d’athérosclérose auraient des concentrations sérique en AsAT, Créatinine phosphokinase, acide urique et acides biliaires élevée. Electrocardiographie : L’aorte est le vaisseau le plus fréquemment affecté par l’athérosclérose. Puisqu’une sténose de cette artère provoque une augmentation de la post-charge cardiaque, il est possible d’observer des modifications électrocardiographiques compatibles avec une hypertrophie ventriculaire gauche (BAVELAAR, et al., 2004). Radiographie : Les radiographies peuvent révéler une augmentation de la densité et de la taille des vaisseaux affectés (DE WIT, et al., 2005). Occasionnellement, la paroi vasculaire peut être le siège de minéralisation. Dans ces cas, des clichés radiographiques peuvent révéler l’existence de vaisseaux de densité minérale. En l’absence de minéralisation, l’évaluation de la vascularisation par radiographie reste d’interprétation subjective et peut mener soit à des diagnostics par excès, soit à des diagnostics par défaut (MANS, et al., 2007). En cas d’athérosclérose, on pourra parfois objectiver une cardiomégalie. Toutefois, l’anatomie complexe de la région thoracique crâniale des oiseaux rend l’identification des lésions difficiles chez la plupart des espèces. Echographie : L’échographie peut permettre de détecter des processus athérosclérotiques proches du cœur (DE WIT, et al., 2005). Angiographie avec produit de contraste : L’angiographie a été utilisé pour détecter un anévrisme de l’artère coronaire droite chez un cacatoès (VINK-NOOTEBOOM, et al., 1998) et une dilatation cardiaque associée à une réduction du flux sanguin dans l’aorte et les artères brachiocéphaliques chez un ara (PHALEN, et al., 1996). Dans les deux cas, les oiseaux étaient atteints d’athérosclérose. Histologie : L’athérosclérose ne peut être diagnostiquée uniquement sur des éléments macroscopiques. En conséquence, seule l’histologie permet un diagnostic de certitude (KRAUTWALD-JUNGHANNS, et al., 2004). Mesure de la distance de boiterie : Comme son nom l’indique, la distance de boiterie est la distance à partir de laquelle on observe une boiterie. Les résultats sont associés à des pronostics différents en médecine humaine. Toutefois, il n’existe pas de valeurs de référence à ce jour en médecine aviaire (BEAUFRERE, et al., 2011). Mesure de pression : La mesure de l’index de pression cheville-bras est utilisé chez les hommes pour caractériser une claudication intermittente. Toutefois, les valeurs de référence n’existent pas en médecine aviaire et les mesures de pression sont jugées à ce jour trop incertaines pour utiliser ce test (BEAUFRERE, et al., 2011).

Page | 86
- Amyloïdose
Chez les oiseaux de compagnie l’amyloïdose est plus courramment rencontrée chez les petites espèces de passereaux. Elle peut être occasionnellement rencontrée chez les autres oiseaux également. La substance amyloïde se dépose dans un grand nombre de tissu mou et, dans certains cas, est retrouvée dans les parois des vaisseaux sanguins. Cette affection est habituellement non détectée macroscopiquement. Histologiquement, les vaisseaux affectés ont une media épaissie, qui apparaît anamorphe et lisse et qui présente une coloration éosinophilique ou amphophilique à la coloration hematoxyline-éosine. Comme chez les Mammifères, la substance amyloïde est biréfringente lorsqu’elle est colorée avec du rouge Congo et observée avec une lumière polarisée (SCHMIDT, et al., 2003). Elle a été détectée chez un nandou américain (Rhea americana) associée avec une inflammation chronique (RANDALL, et al., 1996).
- Minéralisation
La minéralisation des vaisseaux sanguins sans autre modification morphologique est observée dans les cas d’insuffisance rénale sévère, de ration dont le rapport phospho-calcique est déséquilibré chroniquement et lors d’intoxication à la vitamine D. Les modifications sont typiquement dans les artères ou les artérioles et peuvent être observées dans nimporte quelle organe ou tissu. La seule répercussion macroscopique consiste parfois en une sensation graveleuse du tissu si la lésion est largement étendue ou associée avec d’autres minéralisations des tissus mous. Dans les plus grosses artères il est possible d’observer des plaques grises-blanches, brillantes, fermes, irrégulières et surrélevées. Histologiquement, la paroi du vaisseau peut être affectée entièrement ou en partie seulement (SCHMIDT, et al., 2003).
B. Vasculopathies nutritionnelles
- Carence en vitamine E et en Sélénium
La carence en vitamine E et en Selenium peut se manifester, entre autres, sous une forme dite « diathèse exsudative », caractérisée par une atteinte vasculaire (TREMBLAY, et al., 1992). Elle a été rapportée chez les volailles (KAHN, et al., 2008). Elle apparaît dans les même circonstances que la forme dite « cardiomyopathie nutritionnelle » mais est caractérisée par une augmentation marquée de la perméabilité des capillaires. Ainsi, les symptômes sont représentés essentiellement par de l’œdème (KAHN, et al., 2008). Les lésions de l’endothélium vasculaire sont suivies par l’apparition de thrombi fibrineux, d’hémorragie et d’une nécrose ischémique (TREMBLAY, et al., 1992). Les examens sanguins montrent une hypoalbuminémie alors que le fluide exsudé est riche en protéine (KAHN, et al., 2008).

Page | 87
- Carence en Cuivre et en Fer
La déficience en cuivre est considérée rare chez les oiseaux autres que les dindes. Elle est supposée être une cause d’anévrisme disséquant (SCHMIDT, et al., 2003) (FERRERAS, et al., 2001) (BEAUFRERE, et al., 2010). En effet, le cuivre est un composant qui permet la réalisation de réactions redox et assure ainsi le fonctionnement de plusieurs enzymes. Il est en particulier nécessaire pour assurer le bon fonctionnement de la lysyl oxidase. La lysyl oxidase est une enzyme contenant du cuivre qui assure la réticulation du tissu conjonctif (GAL, et al., 2012). Plus précisément, le cuivre permet de catalyser l’oxidation des résidus de lysines au cours de la formation de la structure réticulaire qu’est la desmosine. La desmosine assure des interconnexions par liaison covalente entre des fibres d’élastine et leur confère leur propriétés mécaniques. Une déficience en cuivre aboutit ainsi à une diminution des pontages ce qui affaiblit la structure de l’élastine et conduit à une rupture aortique chez la volaille (KLASING, 2008). Un anévrisme aortique disséquant vraisemblablement secondaire à une carence en cuivre a été décrit chez une autruche (Struthio camelus) (BAPTISTE, et al., 1997).
C. Embolies vasculaires
Définition : Les embolies artérielles sont peu décrites chez les oiseaux bien que celles-ci puissent avoir des répercussions significatives sur la santé de ces animaux (BEAUFRERE, et al., 2010). Etiologie : Les emboles peuvent être représentées par des thrombi aseptiques émis lors de rupture de plaque d’athérome, d’une intoxication au métaux lourds (DONELEY, 2011) ou d’un phénomène traumatique. Des thrombi septiques sont fréquemment associés à une endocardite valvulaire bactérienne (RANDALL, et al., 1996) (RANDOLPH, et al., 1984), une septicémie (RANDALL, et al., 1996) ou une infection aspergillaire des sacs aériens. (SCHMIDT, et al., 2003) (JESSUP, 1980). Des embolies graisseuses ont été rapportées chez un balbuzard pêcheur (DENNIS, et al., 2000). Des embolies de jaune d’œuf et des embolies fibrocartilagineuses en provenance du cartilage articulaire intervertébral ont été décrite dans des vaisseaux de la moelle épinière et/ou des leptoméninges chez des dindes (STEDMAN, et al., 1998). Pathogénie : L’émission d’embole peut aboutir à l’obstruction mécanique des vaisseaux, ce qui provoque un infarcissement lors d’obstruction veineux ou une ischémie lors d’obstruction artérielle. L’embolisation de matériel septique peut engendrer la dissémination d’un phénomène infectieux (DONELEY, 2011). Symptômes : Les symptômes sont en relation avec la défaillance de l’organe dont l’irrigation a été perturbée. Ainsi, on pourra observer des symptômes nerveux lors d’accident ischémique cérébral (BENETT, 1994) (BEAUFRERE, et al., 2010) ou d’emboli fibrocartilagineuse médullaire (STEDMAN, et al., 1998), une mort subite lors d’ischémie myocardique (DONELEY, 2011). Lors d’embole fibrocartilagineuse les animaux peuvent récupérer des fonctions nerveuses avec le temps (STEDMAN, et al., 1998). Diagnostic : Selon la localisation de l’embole, le diagnostic reposera sur l’observation clinique d’une froideur du membre, éventuellement confirmée par un examen de thermographie. Une angiographie fluorescéine pourra être utilisée. En cas d’atteinte d’un vaisseau profond, le diagnostic reposera sur l’observation de lésions ischémiques lors d’un examen d’imagerie par résonnance magnétique (BEAUFRERE, et al., 2010).

Page | 88
D. Vasculites
Définition : Les vasculites (artérite, phlébite, lymphangite) sont des affections inflammatoires des vaisseaux (respectivement des artères, veines, vaisseaux lymphatiques). Chez les oiseaux, seuls des causes infectieuses ont été décrites. Etiologie et épidémiologie : Les vasculites sont des affections relativement peu fréquentes chez les oiseaux (BEAUFRERE, et al., 2010). Elle résultent de l’infection par :
Des bactéries telles que Mycobacterium spp. (SCHMIDT, et al., 2003), Escherichia coli (Poule – Gallus gallus (RANDALL, et al., 1996)) ou Staphylococcus aureus (Poule – Gallus gallus (RANDALL, et al., 1996)) Des virus tels que des adénovirus (Crécerelle – Falco punctatus (RANDALL, et al., 1996)), l’influenza aviaire (Autruche – Struthio camelus (RANDALL, et al., 1996)) Paramyxovirus 1 (PMV-1) et des togavirus (WEE, encéphalite équine de l’ouest) (SCHMIDT, et al., 2003) Des champignons comme Aspergillus spp. (JUNG, et al., 2009) (Amazone à front bleu – Amazona aestiva (RANDALL, et al., 1996)) Des parasites tels que Haemoproteus spp. (Pigeon – Columba livia) (Emeu – Dromaius novaehollandiae (RANDALL, et al., 1996)) ou les vers filaroïdes des genres Splendidofilaria (rencontrés chez les anatidés, corneille d’amérique - Corvus brachyrhynchos, Pie d’amérique - Pica hudsonia, moineau domestique - Passer domesticus), Sarconema (Anatidés), Paronchocerca (Ciconiidés, Odontophoridés, Otididés – Eupoditis ruficrista (NICHOLLS, et al., 1995)), Cardiofilaria (Cacatuidées, oiseaux de proie) ainsi que Chandlerella (Cacatuidés, Corvidés) (BARTLETT, 2008), Schistosoma spp. (Cygne – Cygnus olor (RANDALL, et al., 1996) (AKAGAMI, et al., 2010))
Pathogénie : Des infections organiques peuvent provoquer une inflammation des vaisseaux de l’organe atteints par extension (SCHMIDT, et al., 2003). C’est notamment le cas d’Aspergillus qui colonise la vascularisation adjacente aux sacs aériens (JUNG, et al., 2009). Secondairement, les agents pathogènes peuvent migrer par voie hématogène. Dès lors on peut observer le développement de vasculite dans nimporte quel organe (SCHMIDT, et al., 2003). Lésions : Lors de vasculite bactérienne on observe une nécrose de la paroi vasculaire et une réponse faisant intervenir primitivement des hétérophiles (SCHMIDT, et al., 2003). Lors d’infection aspergillaire, on observe une vasculite granulomateuse (JUNG, et al., 2009). Une étude rétrospective post-mortem a montré que ces artérites granulomateuses mycobactériennes affectaient en premier lieu les artères majeures et coronaires chez 6 espèces de perroquets (perruches ondulées, perruches callopsites, cacatoès des moluques, toui flamboyant, pione à couronne blanche et platycercus spp.) (REAVILL, et al., 2009). Il est également possible d’observer des anévrismes mycosiques (dits également anévrismes infectés) (COURCHESNE, et al., 2009). Lors d’infections virales on observe des hémorragies, particulièrement dans les séreuses du tractus gastro-intestinal. Histologiquement, on observe des hémorragies associé à de l’œdème, une hyalinisation et une dégénération de la paroi des vaisseaux sanguins. Il est également possible de rencontrer une nécrose endothéliale, une thrombose ainsi qu’un infiltrat inflammatoire mononucléaire variable (SCHMIDT, et al., 2003). Les infections par des nématodes s’accompagnent d’une hypertrophie de la média (RANDALL, et al., 1996). Lors d’haemoproteus on observe une vasculite granulomateuse (RANDALL, et al., 1996)

Page | 89
E. Néoplasies vasculaires : Hémangiomes et hémangiosarcomes
Définition : Les hémangiomes sont des tumeurs bénignes de l’endothélium vasculaire. Les hémangiosarcomes sont également connus sous le nom d’hémangioendothéliomes malins ou d’angiosarcomes. Ces derniers correspondent à la version malignes des hémangiomes (REAVILL, 2001) et représentent donc des néoplasies malignes se développant à partir de cellules endothéliales vasculaires (NAKANO, et al., 2012). Ces tumeurs sont caractérisées par une angiogénèse abondante, une capacité à métastaser rapidement et un haut degré de malignité. Etiologie et facteurs de risques : Causes infectieuses : Il a été rapporté que le rétrovirus de la leucose/sarcome aviaire (ALSV sous groupe J) pouvait induire la formation multifocale de tumeurs vasculaires (LAI, et al., 2011) (NAKANO, et al., 2012) (BURSTEIN, et al., 2007). Un autre rétrovirus, dit de l’hémangiome aviaire (AHV) a également été impliqué dans la formation de telles tumeurs chez les poulets (SOFFER, et al., 1990) (SOLA, et al., 2007) (SOFFER, et al., 1990) (VENUGOPAL, et al., 2007). Enfin, l’injection du rétrovirus du faisan à collier (RPV) chez des poussins aboutit à la formation d’hémangiomes pulmonaires dans 88% des cas (CARTER, et al., 1983). Causes environnementales : Une exposition prolongée aux rayons ultra-violets pourrait constituer un facteur de développement de tumeurs vasculaires. En effet, chez les oiseaux d’eau les hémangiomes et hémangiosarcomes se développent le plus fréquemment dans des zones où le plumage est de faible densité (REAVILL, 2001). Par ailleurs, l’implication d’une irritation cutanée traumatique chronique, également décrite en médecine humaine, est un facteur de risque suspecté par certains auteurs (DE LAS MULAS, et al., 1993). Causes iatrogènes : Comme en médecine humaine, les hémangiosarcomes hépatiques seraient reliés à l’utilisation de stéroïdes anabolisants androgènes (FALK, et al., 1979). Un cas décrit le développement d’un hémangiosarcome hépatique chez un amazone recevant un traitement à long-terme de testostérone pour perte de plume (ROSSI, 1998). Causes héréditaire : Dans certains cas, aucune particule virale n’a été mise en évidence ni aucun facteur de risque. Ceci pourrait laisser penser à une prédisposition génétique (GARDNER, et al., 1981). Epidémiologie : Les tumeurs vasculaires sont relativement rares chez les oiseaux (BEAUFRERE, et al., 2010). Toutefois, le groupe des hémangiomes-hémangiosarcomes représentent la deuxième tumeur la plus fréquente (20%) des tumeurs non lymphoïdes. (REECE, 1996). Les hémangiomes sont plus fréquemment rapportés chez les perruches ondulées (Melopsittacus undulatus) que chez les autres oiseaux (REAVILL, 2001). Des cas sporadiques ont été décrits chez des autruches en association avec de l’aspergillose (SHATHELE, et al., 2009), chez un faucon pèlerin de 15 ans (FORBES, et al., 2000), chez des cailles sauvages (REECE, 2007). Une épidémie d’hémangiome chez des poulets en Israël a engendré une mortalité de 20% de la bande (BURSTEIN, et al., 2007). Chez les poulets, il n’existe pas de prédisposition sexuelle (CHENG, et al., 2010) (BURSTEIN, et al., 2007). L’âge moyen de survenu est de 10.8 ans (écart de 3 à 20 ans) (REAVILL, 2001).

Page | 90
Les perruches callopsittes (Nymphicus hollandicus) (SLEDGE, et al., 2006) sont en tête de liste parmi les espèces affectées par les hémangiosarcomes, laquelle inclue également les poulets (Gallus gallus domesticus) (BURSTEIN, et al., 2007) (NAKAMURA, et al., 1988), les faisans (NAKANO, et al., 2012) (SUEDMEYER, et al., 2001), les autruches (HEADLEY, 2005) (SHATHELE, et al., 2009), les cygnes, les amazones (Amazona sp.) (HANLEY, et al., 2005) (ROSSI, 1998) (MICKLEY, et al., 2009), les inséparables (Agapornis sp.) les gris d’Afrique (Psittacus erithacus), les piones (Pionus sp.), les perruches ondulées (FREEMAN, et al., 1999) (PETRAK, et al., 1982), les canaris (Serinus canarius), les fringillidés (REAVILL, 2001), le Padda de Java (Padda oryzivora) (NAKANO, et al., 2012) et la perdrix rouge (DE LAS MULAS, et al., 1993). L’écart d’âge affecté par ces tumeurs est similaire à celui des hémangiomes et les sexes sont également représentés (FREEMAN, et al., 1999). Lésion : Macroscopique : Localisation : Bien que les hémangiomes et les hémangiosarcomes sont plus fréquents dans leur forme cutanée, ils peuvent également se développer sur les artères principales (SCHMIDT, et al., 2003)(BEAUFRERE, et al., 2010) et particulièrement dans les reins (SILLER, 1981). Les hémangiomes se rencontrent habituellement au niveau cutané (pieds, région inguinale, cloaque, sur le côté du cou (CERRUTI SOLA, et al., 1997) et les ailes (REECE, 1996)), dans la rate (REAVILL, 2001) ou le foie (REECE, 2007). Plus rarement, il est possible de les rencontrer sur le cœur et les intestins (CHENG, et al., 2010). Le site d’apparition primaire des hémangiosarcomes peut-être sous-cutanée (PETRAK, et al., 1982) (SLEDGE, et al., 2006), splénique (PETRAK, et al., 1982), hépatique (PETRAK, et al., 1982) (ROSSI, 1998), interclaviculaire à partir de l’artère carotide droite (HANLEY, et al., 2005), articulaire (carpal droite) (FREEMAN, et al., 1999), ovarien (MICKLEY, et al., 2009), intra-thoracique, adhérent au cœur et poumon (HEADLEY, 2005) et pulmonaire (SHATHELE, et al., 2009). Les sites cutanés d’hémangiosarcomes les plus fréquents sont représentés par le bec, les ailes (propatagium (SUEDMEYER, et al., 2001)), les pieds, les pattes (SLEDGE, et al., 2006) et le cloaque (REAVILL, 2001). Les hémangiosarcomes sont localement invasifs et multicentriques (FREEMAN, et al., 1999). Aspect Les hémangiomes sont habituellement rouge foncés à noir, pédonculés et circonscrits (REAVILL, 2004). Ils ont une surface lisse et apparaissent secs à la coupe (HEADLEY, 2005). Occasionnellement, ils sont associés à une prolifération de tissu adipeux conduisant à une tumeur qui a été appelée hémangiolipome (SCHMIDT, et al., 2003). Les hémangiosarcomes peuvent être rouge-marrons à grisâtres et ont une apparence grossière avec des bords mal délimités. Lors de localisation cutanées, les tumeurs tendent à apparaître enflammées et nécrotiques (FREEMAN, et al., 1999). Il correspondent à des masses plus ou moins molles à fibreuses. Elles présentent un aspect spongieux friable à la coupe (HEADLEY, 2005)(SUEDMEYER, et al., 2001). Microscopique : Les hémangiomes sont composés de nombreux canaux vasculaires plus ou moins réguliers délimités par un endothélium bien différencié. Les hémangiosarcomes sont quant à eux constitués de canaux vasculaires irréguliers, d’un endothélium modérément indifférencié ou pauvrement différencié et de foyers solides (SCHMIDT, et al., 2003). Il est possible d’observer des inclusions dans les cardiomyocytes lors d’infection par le virus de la leucose lymphoïde (NAKAMURA, et al., 1988). Symptômes : On observe généralement une mort subite (ROSSI, 1998) ou survenant rapidement après un épisode de détresse respiratoire et une anémie (SUEDMEYER, et al., 2001). Ces symptômes sont secondaires à l’apparition d’un hémopéricarde, d’un hémothorax ou d’un hémoabdomen provoquant un collapsus circulatoire et un choc hypovolémique (HEADLEY, 2005). Il est possible d’observer du picage lors de néoplasie

Page | 91
hépatique (ROSSI, 1998). Lorsque ces néoplasie n’affectent pas des organes internes, elle se présentent comme des nodules cutanés saignant facilement (DE LAS MULAS, et al., 1993). Diagnostic différentiel : Les tumeurs endothéliales doivent être différenciés d’autres malformations vasculaires tels que les fistules artério-veineuses, les anévrismes, les hématomes et les tissus de granulation fortement vascularisés (REAVILL, 2004). Les mélanomes malins, bien que rares chez les oiseaux domestiques, peuvent avoir la même apparence que les hémangiomes (REAVILL, 2004). Pronostic : Le pronostic des hémangiosarcomes est sombre car il s’agit de processus hautement infiltrant et que la rupture de la tumeur peut provoquer des hémorragies fatales (HEADLEY, 2005). Chez les mammifères, on considère habituellement des hémorragies brutales mais la littérature décrit également des hémorragies chroniques continuelles provoquant des anémies. La chute de l’osmolalité plasmatique peut provoquer des épanchements péricardiques et s’accompagner d’un choc hypovolémique mortel (NAKANO, et al., 2012).
F. Vasculopathie idiopathique :
- L’anévrisme disséquant spontané de la dinde
Définition : Un anévrisme est une dilatation localisée ou une évagination d’une portion fine et peu résistante d’un vaisseau, remplie de sang et communiquant avec la lumière artérielle (GAL, et al., 2012). Comme nous l’avons déjà vu, les ruptures artérielles (souvent aortiques) représentent un syndrome observé lors de l’évolution naturelle de plusieurs maladies chez les oiseaux. Ainsi, un processus athérosclérotique (VINK-NOOTEBOOM, et al., 1998), une intoxication au pois de senteur, une infiltration infectieuse de la paroi vasculaire (COURCHESNE, et al., 2009) ou une carence nutritionnelle en cuivre peuvent provoquer l’apparition d’anévrismes pouvant se rompre. Toutefois, chez la dinde, il existe un syndrome multifactoriel d’anévrisme artériel dissécant touchant l’aorte ou les coronaires (TREMBLAY, et al., 1992). Epidémiologie : Cette affection a été décrite chez les dindons mâles âgés de 7 à 24 semaines, avec un pic de mortalité entre 12 et 16 semaines (TREMBLAY, et al., 1992). Les femelles sont également affectées mais l’incidence est moindre (KAHN, et al., 2008)La rupture aortique a également été décrite chez les autruches et les émeus (STEWART, 1994). Les anévrismes aortiques sont également retrouvés occasionnellement chez les volailles, et plus spécifiquement chez les poulets, les dindes, et de façon plus anecdotique, chez les autruches (SCHMIDT, et al., 2003) (FERRERAS, et al., 2001). Etio-Pathogénie : L’étiologie sous-jacente des anévrismes artériels pourrait être soit une faiblesse de la paroi artériel, ou bien une interruption locale de la paroi artérielle qui dans les deux cas peut se rompre (FOX, et al., 2005) (SCHMIDT, et al., 2003). La cause de cette faiblesse est encore peu comprise et vraisemblablement multifactorielle. Les dinde mâles auraient une prédisposition génétique à l’origine de trouble des tissus conjonctifs relatif à l’élastine ou au collagène (CRESPO, et al., 2008). Cette prédisposition consisterait soit à un défaut d’absorption ou à un excès d’élimination du cuivre, conduisant à une hypocuprémie non nutritionnelle (SHIVAPRASAD, et al., 2004) soit à un défaut de lysyl oxidase (KAHN, et al., 2008). En effet, la lysyle, oxidase isolée d’aorte de dinde est active sur le tropelaston et les pontages entre les fibres de collagène semble être moins élevé chez les mâles que chez les femelles. Ceci pourrait être un facteur de développement d’anévrismes aortique spontané chez les dindons (KAHN, et al., 2008). La pression artérielle systolique supérieure à 250 mmHg (KRISTA, et al., 1965) (hypertension chronique, aggravée lors de

Page | 92
stress (TREMBLAY, et al., 1992)) favoriserait l’apparition d’artérosclérose. L’arterosclérose est caractérisée par une rigidité et donc une perte d’élasticité des artères s’accompagnant secondairement d’une augmentation de l’amplitude de la pression pulsatile. L’amplitude des ondes de pression augmente alors le long de l’aorte jusqu’à faire un pic en région ischiatique. C’est à cette région que l’endothélium a alors le plus de chance d’être lésée et de former des lésions athérosclérotique. (RUIZ-FERIA, et al., 2004) Cette artérosclérose pourrait être encore aggravé par l’existence de taux lipidiques sanguins élevé (principalement le cholestérol) (PRITCHARD, et al., 1958) favorisant le développement de plaque d’athérome (GRESHAM, et al., 1961).En plus d’atteinte de l’élasticité tissulaire, l’absence de vasa vasorum intra mural au niveau de l’aorte abdominale pourrait expliquer une moins bonne nutrition de la paroi vasculaire et à une dégénérescence de la média (NEUMANN, et al., 1973). Quelqu’en soit la cause, le tissu élastique de l’aorte dans sa région distale, entre l’artère iliaque externe et l’artère sciatique, est remplacé par du tissu fibreux entraînant une élévation de la pression artérielle, une rupture de l’intima et la formation d’un anévrisme dans la paroi du vaisseau. La rupture de l’anévrisme provoque une hémorragie abdominale massive et fatale (TREMBLAY, et al., 1992). Lésions : Macroscopiques : On observe une anémie de la tête, de la peau et de la musculature. Occasionnellement, le sang peut s’écouler par la bouche. De gros caillot sanguin sont retrouvés dans la cavité abdominale et en position sous-capsulaire rénale. Les caillot peuvent être présents dans le sac péricardique, les poumons et les muscles des jambes (McSHERRY, et al., 1954) (PRITCHARD, et al., 1958). Les anévrismes et rupture d’autres artères comme les coronaires ont été décrits (SHIVAPRASAD, et al., 2004). On observe l’existence d’une fente longitudinale entre les artères iliaque externe et ischiatiques. Dans cette région, l’aorte est dilatée, la paroi est fine et a perdu de son élasticité. Microscopiques : Les tuniques intima et media peuvent être partiellement séparées de l’adventice. Les fibres de la tunica media peuvent montrer des modifications dégénératives sévères et peut être infiltrée par des hétérophiles et des macrophages. La média peut être épaissie suite à une augmentation de la substance fondamentale et une prolifération fibroblastique. La laminae elastique peut avoir disparu au site de rupture. Des modifications dégénératives et des surfaces d’érosion associée à une infiltration cellulaire peuvent être présentes dans l’adventice. La région de rupture est également caractérisée par un épaississement intimal marqué ou par l’existence d’une grande plaque fibreuse (CRESPO, et al., 2008). Symptômes : La mort subite est secondaire à une hémorragie intra-abdominale fatale
(TREMBLAY, et al., 1992).
Diagnostic : Les commémoratifs, la mort subite des dindons mâles surtout et quelques signes cliniques avec une peau très pâle et la présence de sang dans le bec sont les éléments du diagnostic (TREMBLAY, et al., 1992). Prévention : On peut prévenir la maladie par la réduction de la vitesse de croissance, l’élimination des oiseaux reproducteurs présentant une pression systolique élevée, l’utilisation de médicaments hypotenseurs et la supplémentation de la ration en cuivre jusqu’à 200 ppm (TREMBLAY, et al., 1992).

Page | 93
- Le syndrome d’hémorragie péri-rénale de la dinde
Définition : Le syndrome d’hémorragie péri-rénale est également appelé syndrome de la mort subite de la dinde ou angiopathie hypertensive. Il s’agit d’un trouble cardiovasculaire non infectieux caractérisé par une mort subite associée à une hémorragie péri-rénale et une cardiomyopathie hypertrophique (KAHN, et al., 2008). Epidémiologie : La mort subite de la dinde associée à une hémorragie péri-rénale a été reconnue comme étant une cause de mortalité significative chez les dindes mâles entre 8 et 14 semaines d’âge en Amérique du Nord (CRESPO, et al., 2008). Les troupeaux à croissance rapide, dans un environnement froid, stressant avec une alternance jour/nuit fréquente sont prédisposés à développer cette affection (FRANK, et al., 1990). La mortalité est de 0,5 à 2% et il n’y a pas de morbidité (KAHN, et al., 2008). Etio-pathogénie : L’étio-pathogénie est inconnue à ce jour. L’implication de phénomènes hypertensif (LAROCHELLE, et al., 1992) ou infectieux (MUTALIB, et al., 1990) a été éliminée. L’hypothèse recevant actuellement le plus de crédit implique une insuffisance cardiaque congestive aiguë déclenchée à l’occasion d’un exercice (KAHN, et al., 2008). Cette insuffisance serait secondaire à une myocardiopathie hypertrophique, elle-même provoquée par un fonctionnement thyroïdien excessif primaire ou secondaire (lumière, stress, température, trouble métabolique) (CRESPO, et al., 2008). Elle s’accompagne d’une sévère hypotension, d’une arythmie ventriculaire et d’une mort subite (LAROCHELLE, et al., 1992). Dans tous les cas, l’hémorragie péri-rénale serait la conséquence d’une congestion passive sévère qui pourrait être aggravé par la fermeture de la valve rénale, de la circulation portale rénale (CRESPO, et al., 2008). Cette hémorragie n’est surement pas la cause de la mort car cette hémorragie est inconstante et souvent modérée (KAHN, et al., 2008). Symptômes : On observe une mort subite sans symptôme précurseur (CRESPO, et al., 2008). Lésions : Macroscopiques : Les animaux sont en bon état général (CRESPO, et al., 2008). On observe une hypertrophie cardiaque affectant le ventricule gauche et le septum inter-ventriculaire. Associée à cela, on observe des caillots sanguins recouvrant tout ou au moins une partie du rein ainsi que des signes de congestion sanguine (poumon congestionné et œdèmateux, splénomégalie, foie congestionné). Les hémorragies péri-rénales sont une trouvaille inconstante. Microscopiques : Les lésions microscopiques correspondent à des signes de surcharge dans le poumon ainsi que des hémorragies pulmonaires et rénale. Les lésions artérielles correspondent à une vacuolisation interne et une hyperplasie médiale (bien que ces éléments aient également été retrouvé chez des dindes normales). Une hyperplasie de l’épithélium et une diminution des colloïdes des glandes thyroïdiennes ont été remarquées (CRESPO, et al., 2008). On ne remarque pas de lésions compatibles avec une hypertension artérielle (LAROCHELLE, et al., 1992)

Page | 94
III. Troubles de la pression artérielle
A. Hypertension artérielle systémique
Définition : Des pressions artérielles systoliques (mesurées avec un Doppler) supérieures à 200 mmHg chez des oiseaux vigiles ou supérieures à 160 mmHg chez des oiseaux anesthésiés sont considérées être indicatives d’une hypertension chez les perroquets. Chez les rapaces la valeur pourrait être plus élevée et l’hypertension considérée à partir de 240 mmHg (BOWLES, et al., 2007). Etio-pathogénie : Les tumeurs rénales, fréquentes chez les perroquets, pourraient induire une hypertension systémique en agissant par l’intermédiaire du système rénine-angiotensine-aldostérone. Une association entre tumeur rénale et hypertension a été documentée chez les perroquets. L’athérosclérose est également responsable d’hypertension chez les oiseaux (BOWLES, et al., 2007) (SEDACCA, et al., 2009) ainsi que la douleur aiguë (LIERZ, et al., 2012) (GENTLE, 2011) Un choc distributif en phase compensée (perte de moins de 20% du volume sanguin) peut s’accompagner d’une hypertension artérielle secondaire à la stimulation du baroréflexe (LICHTENBERGER, 2004). Les lésions apparaissent lorsque les mécanismes compensateurs autorégulateurs de contrôle de la perfusion de chaque organe sont dépassés par une pression artérielle trop élevée (BOWLES, et al., 2007). Symptômes : Les signes cliniques incluent des lésions oculaires (hémorragies rétiniennes, détachement de la rétine ou un papillœdème), des troubles neurologiques (dépression, confusion, crises convulsives), des signes cardiovasculaires (hypertrophie ventriculaire concentrique et insuffisance cardiaque congestive, accident vasculaire) et des anomalies du système rénal (glomérulonéphrite) (DONELEY, 2011). Diagnostic : Le diagnostic repose sur les signes cliniques et la mesure indirecte de la pression sanguine (méthode Doppler) (DONELEY, 2011).
B. Hypotension artérielle
Définition : Des pressions artérielles systoliques (mesurées avec un Doppler) inférieure à 90 mmHg chez des oiseaux vigiles sont considérées être indicatives d’une hypotension chez les oiseaux (LICHTENBERGER, 2005). Etio-pathogénie: Certains agents anesthésiques (halothane, isoflurane) peuvent engendrer une dépression cardiovasculaire et provoquer une hypotension artérielle (GOELZ, et al., 1990). On peut également observer une hypotension lors d’arythmies cardiaques, ou lors d’un choc distributif secondaire à un sepsis, une déshydratation ou une hémorragie en phase décompensée précoce ou en phase terminale (LICHTENBERGER, 2005). Le différentiel de pression entre les artère et les veines est le moteur de la perfusion capillaire. Ainsi, lors d’hypotension artérielle on observe une hypoperfusion des reins, du GIT, de la peau et des muscles. La distribution du volume circulant est inégale et les tissus fonctionnent en anaérobie ce qui provoque une acidose (LICHTENBERGER, 2004). Si cette hypoperfusion se maintient les organes commencent à défaillir (SCHNELLBACHER, et al., 2012).
Symptômes : Lors de choc, on observera une hypothermie, des extrémités froides, une tachycardie, une pression sanguine diminuée, des muqueuses pâles, un TRC augmenté ainsi qu’une dépression mentale (LICHTENBERGER, 2004). Diagnostic : Le diagnostic repose sur les signes cliniques, une anesthésie concomitante et la mesure indirecte de la pression sanguine (méthode Doppler) (DONELEY, 2011).

Page | 95
TROISIEME PARTIE : DIAGNOSTIC DES AFFECTIONS CARDIOVASCULAIRES
AVIAIRES

Page | 96
I. Historique
A. Commémoratifs
Les commémoratifs sont récoltés classiquement. Toutefois, une attention particulière sera porté au signalement de l’espèce. Ainsi les mainates (Gracula religiosa) sont prédisposés à développer de l’hémochromatose (DE WIT, et al., 2005) alors que les amazones (Amasona spp.), les gris du Gabon (Psittacus erithacus) et les perruches callopsittes (Nymphicus hollandicus) sont prédisposés à développer de l’athérosclérose (BEAUFRERE, et al., 2013). L’âge est aussi une information intéressante, puisque les oiseaux les plus âgés sont davantage susceptibles de développer des cardiomyopathies (KENNETH, et al., 2011). Une étude a prouvé que 19,5% des gris du gabon de 30 à 55 ans, 15% des ara de 30 à 60 ans et 11% des inséparable de 6 à 18 ans montraient des lésions cardiaques (REAVILL, et al., 2010). Par exemple l’athéroclérose est une affection se déclarant généralement après 15 ans chez les psittacidés prédisposés (DE WIT, et al., 2005). Des renseignements concernant l’alimentation et l’environnement peuvent permettre de mettre en évidence un régime riche en graine de tournesol prédisposant à l’athérosclérose ou une déficience en cuivre favorisant les ruptures aortiques (STRUNK, 2003). L’exposition à la fumée de cigarette, à d’autres toxines ou la prise de médicaments peut constituer un facteur important de maladie cardiovasculaire. Le contact avec des vecteurs potentiels de maladie (insectes, souris, autres oiseaux) ou avec un environnement possiblement source d’infection doit être noté (KENNETH, et al., 2011) (DE WIT, et al., 2005).
B. Anamnèse
Le propriétaire sera questionné quant au niveau d’activité de l’oiseau, à ses capacités respiratoire, à la présence de toux, (DE WIT, et al., 2005), à la tolérance à l’exercice et à la force du patient en général. Toute crise convulsive ou épisode de syncope doit être rapporté. Toute modification du comportement, de l’appétit, des vocalisations ou de la coordination motrice peut se révéler importante.
II. Examen clinique et interprétation des trouvailles
A. Observation à distance
L’examen physique doit commencer par une évaluation visuelle de l’oiseau dans son enclos, sa volière ou sa cage (TRISTAN, 2010) (DE WIT, et al., 2005). On s’attachera à rechercher des signes généraux tels que de la faiblesse intermittente et de la léthargie (PEES, et al., 2006) (BAINE, 2012) (MITCHELL, et al., 2008), de l’anorexie (BAINE, 2012), une intolérance à l’exercice. Il faudra ensuite évaluer et rechercher des troubles neurologiques (BAVELAAR, et al., 2004) en évaluant l’état mental. La fréquence respiratoire est mesurée (TRISTAN, 2010) et on s’attachera à rechercher une dyspnée. Nous noterons que si la dyspnée peut exister chez les oiseaux (PEES, et al., 2006) (BAINE, 2012) (MITCHELL, et al., 2008), celle-ci peut ne pas être observée car la surface des conduits aériens aviaires est recouverte par une substance trilaminaire qui limite le développement d’œdème pulmonaire (REMBERT, et al., 2008). En cas de dyspnée sévère, la mise dans une cage à oxygène peut être indiquée avant la poursuite de l’examen (DE WIT, et al., 2005). Concernant la toux (MITCHELL, et al., 2008), celle-ci évoquera une maladie du cœur droit contrairement aux Mammifères chez qui elle évoque une atteinte à gauche. En effet, l’aorte passe à droite et ne provoque donc pas compression de la bronche gauche entre l’aorte et l’élargissement cardiaque gauche (REMBERT, et al., 2008).

Page | 97
B. Examen manuel
En cas de problème circulatoire aigu, les oiseaux doivent être traités en urgence. La manipulation de l’animal doit se limiter au minimum et l’oiseau doit être maintenu en position debout pour prévenir toute insuffisance cardiaque (PEES, et al., 2006). Dans tous les cas, le vétérinaire doit être en connaissance de la réponse de l’oiseau à la manipulation et doit être prêt à arrêter son examen si l’état du patient se détériorait. Une dyspnée, une tachypnée, un collapsus, des syncopes ou des crises convulsives doivent nécessiter un arrêt de l’examen (TRISTAN, 2010).
1. Mesure de la température cloacale
La mesure de la température cloacale permet d’évaluer l’existence d’une hypothermie, observée lors d’un état de choc. L’hypothermie peut également être responsable d’arythmie cardiaque. (LICHTENBERGER, et al., 2007). La façon la plus précise de mesurer la température chez les oiseaux et d’utiliser une sonde thermique flexible œsophagienne et de mesurer la température en regard du cœur (COSTELLO, 2004).
2. Evaluation du système circulatoire périphérique
Couleur des muqueuses : La couleur des muqueuses constitue une évaluation de la perfusion périphérique des tissus (LICHTENBERGER, et al., 2007). Elle peut indiquer l’existence d’un problème cardio-vasculaire (KENNETH, et al., 2011) (PEES, et al., 2006) (BAINE, 2012). Les muqueuses utilisées lors de l’examen clinique sont la muqueuse buccale (généralement pigmentée (DE WIT, et al., 2005)), la muqueuse sclérale (LICHTENBERGER, et al., 2007) et la muqueuse cloacale (WELLE, et al., 2011). Les muqueuses sont roses chez les oiseaux ayant une bonne perfusion des tissus périphériques et montreront une pâleur voire une cyanose lors de déficit perfusionel (LICHTENBERGER, et al., 2007). Certains auteurs préconisent d’évaluer l’existence d’une cyanose en jugeant de la couleur de la peau périorbitaire si les lits capillaires sont visibles à travers la peau de la face (PEES, 2005) ou à travers la kératine des ongles ou de la rhinothèque (STRUNK, 2003). Pouls périphérique : Le pouls périphérique est palpable sur la face médiale de l’aile, au niveau de l’artère ulnaire. Toutefois la palpation du pouls n’est pas réalisable chez tous les oiseaux (PEES, et al., 2006). Il sera alors possible de détecter le pouls à l’aide d’un boîtier Doppler (HURLEY-SANDERS, et al., 2012). La palpation du pouls est utile lors de l’évaluation de la perfusion périphérique (DE WIT, et al., 2005) : Un pouls bondissant pourra évoquer une communication inter-ventriculaire (EVANS, et al., 2001) alors qu’une absence de pouls, est indicative d’un trouble circulatoire telle qu’une ischémie thromboembolique (HURLEY-SANDERS, et al., 2012). Les extrémités (en particulier les tarses, les extrémités digitées ou la tête) peuvent être palpées à la recherche d’œdème notamment chez les rapaces (TRISTAN, 2010). En effet, cette situation se produit rarement chez les autres oiseaux atteints de maladies cardiovasculaires (DE WIT, et al., 2005) (PEES, et al., 2009). Le temps de recoloration capillaire (temps de remplissage capillaire) peut être mesuré en appliquant une pression légère sur une rhampothèque ou sur des griffes translucides (DE WIT, et al., 2005) et en mesurant le temps de recoloration. La rampothèque est une enveloppe cornée recouvrant le bec, séparée des couches plus profondes du derme par une couche vasculaire. Le temps de recoloration doit être inférieur à 2 secondes. Une augmentation sera observée chez les patients déshydratés à plus de 7%, avec une mauvaise perfusion périphérique ou avec un mauvais retour veineux (STRUNK, 2003). Le statut d’hydratation interstitielle des oiseaux peut être évalué en jugeant de l’humidité des muqueuses, de l’enfoncement du globe oculaire et en exerçant une traction sur la peau des pattes ou de la paupière supérieure (DE WIT, et al., 2005). Lorsque la déshydratation est supérieure à 8% on peut s’attendre à observer une hypovolémie (LICHTENBERGER, 2004).

Page | 98
Etat mental : On déterminera le degré de conscience et si l’animal est stuporeux (réactif lors de stimuli nociceptifs) ou comateux (non réactif aux stimuli nociceptifs). Une absence mentale pourra suggérer une hypoperfusion cérébrale ou une atteinte primitive du cerveau. On réalisera alors un examen neurologique complet (LICHTENBERGER, et al., 2007). Il est possible d’observer une distension et un aspect sinueux de la veine jugulaire droite externe (KENNETH, et al., 2011) ainsi qu’une dépression de la région périorbiaire (PEES, et al., 2009) lors d’insuffisance cardiaque congestive (ENSLEY, et al., 1979).
3. La palpation cœlomique
Une distension cœlomique peut être observée chez les oiseaux atteints de maladie cardiovasculaire (PEES, et al., 2006) en relation avec la présence d’ascite (KENNETH, et al., 2011) (BAINE, 2012) (MITCHELL, et al., 2008) ou d’une organomégalie due à une congestion. L’ascite peut être observé lors d’insuffisance cardiaque congestive droite ou généralisée (DE WIT, et al., 2005). L’ascite est le plus souvent secondaire à un trouble hépatique chez les oiseaux et peut également être secondaire à l’existence d’une péritonite (STRUNK, 2003).
C. Auscultation
L’auscultation cardiaque a été décrite comme étant difficile du fait de la fréquence cardiaque élevée (DE WIT, et al., 2005) (TRISTAN, 2010) (et d’autant plus lors du stress de la contention (PEES, et al., 2006) (BOUSSARIE, et al., 2010)). En revanche, elle est simplifiée par le fait que les nombreuses cavités corporelles fournissent de nombreuses chambres de résonnance. Ainsi le cœur est audible plus ou moins partout chez les petites espèces. (WELLE, et al., 2011) mais l’auscultation est plus facile à réaliser à la base du sternum, du côté ventral droit ou à gauche du thorax (DE WIT, et al., 2005). Un bref passage sous anesthésie à l’isoflurane a été suggérée pour diminuer la fréquence cardiaque et faciliter l’évaluation cardiaque mais dans la plupart des cas, l’auscultation d’oiseau vigile apporte les informations nécessaires. L’auscultation pourra alors révéler l’existence d’arythmies, d’un souffle cardiaque ou une atténuation des bruits. (DE WIT, et al., 2005). Le « woosh » des souffles mammaliens est également caractéristique des souffles aviaires. Une attention particulière sera portée à confirmer que le bruit est associé aux battements cardiaque et non à la respiration. Le système de gradation utilisé chez les mammifères peut être utilisé chez les oiseaux (WELLE, et al., 2011). Un souffle traduira l’existence de turbulence du flux sanguin secondaire à une anomalie du cœur (insuffisance valvulaire, anomalie congénitale) ou du sang (anémie). Une atténuation des bruits cardiaques pourra être secondaire à un hydropéricarde ou, caractéristique uniquement aviaire, une hépatomégalie. En effet une hépatomégalie peut provoquer un étouffement des bruits cardiaques puisque un foie de taille augmenté aura tendance à envelopper davantage le cœur (DONELEY, 2011). L’auscultation pulmonaire est réalisée par le dos, entre les omoplates. L’accumulation de fluide pleuro-péritonéal ou (moins probablement) pulmonaire peut provoquer un étouffement des bruits pulmonaires ou des râles chez l’oiseau (DONELEY, 2011). Des craquements surajoutés aux bruits pulmonaires peuvent indiquer une affection cardiaque (KENNETH, et al., 2011) (MCMILLAN, 1994).
D. Récupération suite à la contention
Certains auteurs estiment que le temps de récupération respiratoire d’un oiseau normal devrait inférieur à 2 minutes après l’examen clinique (OGLESBEE, et al., 2001) mais peut être considéré normal jusqu’à 5 minutes car cette valeur peut varier selon l’état de stress du patient. Au delà ce temps sera évocateur d’une intolérance à l’effort. (BAINE, 2012) (MCMILLAN, 1994) (KENNETH, et al., 2011) (PEES, et al., 2006) (MITCHELL, et al., 2008).

Page | 99
III. Examens paracliniques Les examens complémentaires doivent être utilisés afin d’investiguer l’existence d’une affection cardiovasculaire primaire, mais aussi afin de rechercher des affections pouvant résulter secondairement en une atteinte cardiaque comme c’est le cas de la goutte et de la septicémie (TRISTAN, 2010). En cas de troubles circulatoires aigus, les procédures diagnostiques doivent être sélectionnées avec précaution. Ainsi, afin de limiter le stress de l’animal et lui assurer une position debout, l’échographie sera davantage indiquée chez ces patients qu’un examen radiographique (TRISTAN, 2010).
A. Ponction des liquides
Indication : En cas d’épanchement abdominal sévère, l’abdominocentèse peut être indiquée avant la réalisation d’autres procédures diagnostiques pour stabiliser le patient (DE WIT, et al., 2005). La nature de tout épanchement abdominal devrait être identifiée par ponction. Technique :
Coeliocentèse : Une aiguille ou un cathéter papillon (21 à 25 G) est inséré caudalement à la base du sternum sur la ligne ventrale en direction de la droite de l’oiseau afin d’éviter la ponction du ventricule (STRUNK, 2003). Le volume de fluide aspiré doit être suffisant pour supprimer les signes cliniques secondaires à la compression des sacs aériens et du cœur par l’accumulation de fluide. Toutefois, le retrait d’un plus grand volume doit être évité puisqu’une telle manœuvre pourra résulter en une hypoprotéinémie et une hypovolémie. Signalons que dans certains cas d’ascite, l’aspiration de volume plus grand pourra représenter un traitement palliatif si les autres thérapies habituelles échouent (diurétiques). Il est rapporté qu’il est possible de ponctionner 60 à 80 ml de fluide deux fois par semaine sans répercussion apparente sur la santé de l’oiseau (DE WIT, et al., 2005). Ponction péricardique : Après préparation aseptique du site de ponction, la péricardiocentèse est réalisée sous contrôle échographique. Une aiguille de 25G est placée dans la cavité péricardique à partir de la paroi abdominale ventro-médiane. C’est une manœuvre facile et qui peut s’accompagner de succès mais la ponction doit être très lente. Elle peut être réalisée sans anesthésie et comporte moins de risque qu’une fenestration péricardique. Il peut ne pas être possible de retirer l’intégralité du fluide contenu dans la cavité péricardique. Toutefois, il est probable que la diminution de pression engendrée par cet acte assure les conditions nécessaires au retrait des fluides restant par traitement médicamenteux (STRAUB, et al., 2003).
Interprétation : Quel qu’en soit sa provenance (ascite, épanchement péricardique, épanchement pleural), on procédera à la mesure de la quantité de fluide aspiré ainsi qu’à l’évaluation de son aspect macroscopique. Ces examens seront suivis par la mesure de la densité et du pH ainsi que par la réalisation d’une cytologie et d’une culture. L’ensemble de ces éléments pourra aider à préciser le type de liquide ainsi que son origine et ainsi orienter le diagnostic de la maladie (DE WIT, et al., 2005).

Page | 100
B. Mesure de la pression artérielle
Indication : La pression sanguine artérielle systémique est le produit du débit cardiaque et de la résistance vasculaire périphérique totale (STEPIEN, 2005). La pression sanguine systolique correspond alors à la pression exercée par le sang contre la paroi vasculaire au cours de la systole (DONELEY, 2011). La mesure de cette pression a un intérêt pour le praticien et constitue une indication dans les cas suivants : Surveillance cardio-vasculaire des patients anesthésiés (ACIERNO, et al., 2008), identification et suivi d’une hypertension (ACIERNO, et al., 2008), détection d’une hypotension lors de choc, évaluation des conséquences hémodynamiques d’une maladie cardiaque (LICHTENBERGER, 2005). Certains auteurs conseillent de réaliser des mesures indirectes de la pression artérielle uniquement chez des patients de plus de 70 grammes (LICHTENBERGER, 2005). En contexte clinique, compte tenu de la différence significative de la valeur de pression mesurée lors de la répétition de la mesure sur un même individu, avec un même brassard replacé au même endroit, certains auteurs suggèrent que la mesure de pression de façon indirecte ne peut être utilisée de manière fiable que dans le cadre d’un suivi de pression per-anesthésique (sans replacement du brassard) (JOHNSTON, et al., 2011). Technique : Il existe plusieurs techniques de mesure de la pression artérielle :
- Les méthodes de mesure directe consistent à placer un cathéter directement dans une artère appropriée (26 gauge et 19 mm dans l’artère ulnaire d’un Amazone par exemple (ACIERNO, et al., 2008)) et à connecter ce cathéter à un transducteur de pression électronique. Bien que ces techniques de mesure de la pression artérielle soient considérées comme étant le gold standard, elles ont l’inconvénient de nécessiter des capacités techniques spécifiques, un investissement coûteux dans l’équipement, une procédure invasive, des soins de maintenance du cathéter (héparinisation fréquente) et s’accompagnent d’un risque de thrombose et d’infection (JOHNSTON, et al., 2011). - Les méthodes de mesure indirecte sont décrites chez les oiseaux. Comme chez les Mammifères, le principe repose sur la détection d’un flux sanguin passant sous un brassard gonflé à l’aide d’un détecteur de flux à ultrasons Doppler qui le convertit en un signal audible (DE LAFORCADE, et al., 2001). L’utilisation des méthodes sphygmomanométrique simple et oscillométrique est peu décrite dans la littérature mais semble infructueuse (ACIERNO, et al., 2008) (LICHTENBERGER, 2005). L’avantage majeur de ces techniques réside en son caractère non invasif et sa moindre technicité dans le cadre d’une activité de routine (LICHTENBERGER, 2005). Ainsi il est possible d’évaluer la pression artérielle au niveau des ailes ou des pattes. Les deux sites sont équivalents quant à la précision des mesures (JOHNSTON, et al., 2011) :
- Pour mesurer la pression artérielle à l’aile chez des espèces de taille moyenne telles que des Amazones, on pourra utiliser un brassard néonatal de taille 1 autour de l’humérus (ACIERNO, et al., 2008). Idéalement, la taille du brassard devrait être telle que sa largeur mesure 30 à 50% de la circonférence de l’aile (LICHTENBERGER, 2005), (ZEHNDER, et al., 2009). On prendra soin de placer les plumes de sorte à ce qu’elles ne soient pas comprimées entre le brassard et l’aile. Si cela n’est pas possible, il faudra envisager d’enlever les plumes réfractaires. Le détecteur Doppler sera quant à lui placé en regard de l’artère ulnaire superficielle distale ou de l’artère ulnaire (LICHTENBERGER, 2005). Les plumes recouvrant la surface d’examen ultrasonographique peuvent être humidifiées avec de l’éthyl alcool avant d’appliquer le gel de contact (JOHNSTON, et al., 2011).

Page | 101
- Pour mesurer la pression artérielle sur une patte chez des espèces de taille moyenne telles que des Amazones, on pourra utiliser un brassard néonatal de taille 1 autour de la portion proximal du tibiotarse. Idéalement, la taille du brassard devrait être telle que sa largeur mesure 30 à 40% de la circonférence de la patte. Le détecteur Doppler sera cette fois-ci placé en regard du segment distal de l’artère ischiatique (ACIERNO, et al., 2008). Il est également possible de placer le brassard autour de la partie distale du fémur. Dans cette seconde option, il faudra alors placer le détecteur Doppler en regard de l’artère tibial crâniale (JOHNSTON, et al., 2011).
Le brassard est alors gonflé jusqu’à ce que l’artère concernée collabe ce qui se manifeste par un arrêt de la circulation en aval. Une fois que le flux est arrêté, le brassard est dégonflé progressivement jusqu’à ce que la pression artérielle lui devienne égale. A ce moment, le sang circule de nouveau dans les artères et il est possible d’entendre le flux de celui-ci à l’aide du détecteur Doppler. La valeur de pression indiquée sur la manomètre correspond alors à la valeur de la pression sanguine systolique (LICHTENBERGER, 2005).
Interprétation : Chez les chats et les chiens, la mesure de pression artérielle de façon indirecte est largement utilisée. Toutefois, l’interprétation de ces techniques est à ce jour encore controversée en médecine aviaire (JOHNSTON, et al., 2011). En effet, certaines études montrent que les valeurs de pression mesurées par méthode ultrasonique sont corrélées à la pression artérielle moyenne (ZEHNDER, et al., 2009) alors que d’autres montrent une corrélation meilleure avec la pression artérielle systolique (LICHTENBERGER, et al., 2004). D’autre auteurs estiment que la mesure de pression par méthode ultrasonique n’est pas suffisamment sensible et n’est pas significativement corrélées aux valeurs de la pression artérielle mesurée par méthode directe. Cette technique a donc été déconseillée par certains auteurs (ACIERNO, et al., 2008). Toutefois, on considère habituellement que la pression artérielle doit se situer entre 90 et 180 mmHg (LICHTENBERGER, 2005). Ainsi :
- Les oiseaux dont la pression artérielle se situe en dessous de 90 mmHg seront considérés comme étant hypotendus (LICHTENBERGER, 2005). Les causes de cette hypotension peuvent être une hypovolémie (secondaire à une perte de sang ou à une déshydratation), un sepsis, des arythmies ou encore une dépression cardio-vasculaire provoquée par des agents anesthésiques (association de kétamine et diazépam, benzodiazépines, alpha-2 agonistes par exemple (LIERZ, et al., 2012)) (LICHTENBERGER, 2004). - L’hypertension est une affection dont la gestion est plus complexe. En effet, la fréquence, les causes et le traitement de l’hypertension sont peu renseignées à ce jour (LICHTENBERGER, 2005). On considère qu’une pression artérielle systolique supérieure à 200 mmHg est trop élevée. Cela peut évoquer l’existence d’une affection rénale (BOWLES, et al., 2007), d’athérosclérose ou de douleur par exemple (SEDACCA, et al., 2009).
On notera que la pression sanguine est influencée par l’âge, le sexe (supérieur chez le mâle), l’environnement et le régime alimentaires (STRUNK, 2003).

Page | 102
C. Imagerie médicale
Les techniques d’imagerie médicale permettent d’évaluer facilement le cœur ainsi que les troncs brachiocéphaliques, l’aorte ascendante, l’aorte abdominale, les veines et artères pulmonaires ainsi que les artères carotides. Occasionnellement, les veines jugulaires, les veines caves crâniales et caudale, l’artère mésentérique et certaines artères viscérales peuvent également être visualisées (BEAUFRERE, et al., 2010).
a. Examen radiographique Indication : La radiographie est indiquée en première intention lors d’anomalie faisant penser à l’existence d’un problème cardiaque. En effet, les signes radiographiques sont non spécifiques et ne peuvent apporter que des éléments de suspicion quant à l’existence d’une affection cardiaque (COLES, et al., 2007). Elle est peu indiquée lors d’atteinte vasculaire car cette technique est relativement peu sensible pour détecter les affection des vaisseaux (BEAUFRERE, et al., 2010). Technique : Il faudra réaliser des clichés radiographiques en vue ventro-dorsale et latéro-latérale afin d’évaluer la taille et la forme du cœur (DE WIT, et al., 2005). Il est important de noter que la technique influe sur l’interprétation des images obtenues. Ainsi, les valeurs de références sont valables et comparables aux mesures réalisés si et seulement si les clichés sont réalisés en suivant le même protocole. En particulier, la distance focale du film et la distance entre le sujet radiographié et le film influent sur les mesures (RETTMER, et al., 2011). Les constantes suivantes ont été utilisés chez des perroquets de taille moyenne (Psittacus erithacus, Poicephalus senegalis, Amazona amazonica) : Distance du film 80cm, exposition 45kV et 10.0 mAs (STRAUB, et al., 2002) ou Distance focale-film 75 cm, exposition 40 kV pour 0.08 sec (RETTMER, et al., 2011). Interprétation : Une radiographie ventro-dorsale est considérée de qualité adéquate lorsque la carène sternale et la moelle épinière sont surperposées et les ailes symétriques. Un mauvais placement ne garantit pas une bonne interprétation radiographique (STRAUB, et al., 2002). Anatomie radiographique : Chez les oiseaux, les sacs aériens engendrent un contraste négatif facilitant l’évaluation des organes de la cavité cœlomique (STRAUB, et al., 2002). En général, la base du cœur est dirigée cranio-dorsalement et est localisée en regard de la deuxième côte. L’apex est dirigé en direction caudo-ventral et s’étend entre les cinquièmes et sixièmes côtes. La taille et la forme de la silhouette cardiaque varient selon les espèces (MCMILLAN, 1994). Certains auteurs considèrent que les phases respiratoires, modifiant la dimension des sacs aériens, influent sur la taille du cœur (MCMILLAN, 1994) alors que d’autres estiment que les dimensions cardiaques ne sont pas significativement modifiées par le cycle respiratoire (STRAUB, et al., 2002). Les bords latéraux du cœur et d’un foie normaux créent une forme de sablier sur les radiographies ventro-dorsales chez de nombreuses espèces de perroquet (SMITH, et al., 1997). La largeur de la silhouette cardiaque mesurée sur une vue ventro-dorsale à la hauteur des atria devrait être d’environ 50% de la profondeur de la cavité cœlomique mesurée à la hauteur de la 5ème vertèbre thoracique. (MCMILLAN, 1994). Les artères centrales peuvent habituellement être bien visualisées et délimitées. Les troncs brachiocéphaliques, l’aorte, les artères pulmonaires et les veines pulmonaires peuvent habituellement être visualisées. En vue ventro-dorsale, l’aorte ascendante peut être visualisée à la droite de l’axe de symétrie central. Les vaisseaux centraux sont davantage visibles chez les grands oiseaux et les oiseaux dont les sacs aériens sont hypergonflés (BEAUFRERE, et al., 2010).

Page | 103
Lésions radiographiques : Le cœur : L’évaluation cardiaque repose sur l’évaluation de la taille, de la forme et de la radiodensité du cœur (COLES, et al., 2007). Dimensions : Evaluation subjective : L’élongation de la silhouette cardiaque, la perte des étranglements crânial et caudal ainsi qu’une perte de l’indentation à la jonction entre le cœur et le foie indiquent subjectivement une augmentation de la taille du cœur (MCMILLAN, 1994). On retiendra que plus les oiseaux sont petits, plus le cœur est grand en proportion (HANLEY, et al., 1997). Les aras ont un foie relativement petit, qui peut conduire à un diagnostic de cardiomégalie par excès (SMITH, et al., 1997). Evaluation objective : Quelques mesures standardisées ont été publiées et peuvent être utilisées afin de garantir une approche objective (RETTMER, et al., 2011). La largeur du cœur peut être mesurée sur une vue ventro-dorsale de façon précise mais pas sur la vue latéro-latérale compte tenu de la superposition du proventricule et du sternum (STRAUB, et al., 2002). De plus, la mesure de la longueur cardiaque est rendue impossible par son enclavement au sein des lobes hépatiques. Les cacatoès font exception, puisque la position des sacs aériens thoraciques crâniaux environnants fait un contraste négatif sur la radiographie et conduit à une bonne visualisation cardiaque (PEES, et al., 2009). Compte tenu des erreurs de mesure et du degré de corrélation avec la dimension du cœur, il est recommandé d’utiliser préférentiellement le ratio Largeur de la silhouette cardiaque : longueur sternale (intervalle de référence [35-41%]) puis le ratio Largeur de la silhouette cardiaque : épaisseur thoracique (intervalle de référence [51-61%] (STRAUB, et al., 2002). La mesure du sternum n’étant pas toujours évidente sur les clichés radiographiques, il est recommandé de la mesurer directement sur l’oiseau (STRAUB, et al., 2002). Ces valeurs sont toutefois à considérer avec du recul car aucune étude n’a été réalisée afin de déterminer dans quelle mesure ces ratios permettaient de discriminer une cardiomégalie d’un cœur normal (STRAUB, et al., 2002). En conséquence, une silhouette cardiaque dont les dimensions sont en dehors des valeurs de référence doit être interprétée avec précaution puisque seuls des écarts majeurs sont d’indication diagnostique (RETTMER, et al., 2011). Ainsi, toute suspicion doit inciter à réaliser d’autres examens cardiaques (mesures d’ECG, échocardiographie) (STRAUB, et al., 2002) (COLES, et al., 2007). Un élargissement de la silhouette cardiaque peut être secondaire à un certain nombre de causes différentes telles que : une dilatation des cavités, une hypertrophie pariétale, un épanchement péricardique ou des tumeurs, mais il n’est pas possible de les différencier sur la seule base des images radiographiques (PEES, et al., 2001). En cas de cardiomégalie, l’élargissement du cœur est habituellement assymétrique. La cardiomégalie peut être la résultante d’une maladie cardiaque primaire (anomalie congénitale) ou bien être secondaire à une poxvirose (rapporté chez des aras), une dégénération valvulaire myxomateuse, une endocardite, une hémochromatose, une anémie chronique ou une compression par une masse extrinsèque (MCMILLAN, 1994). La microcardie est associée à une hypovolémie due à une perte volumique aiguë ou à un choc endotoxique. On observe alors une rétraction du cœur à partir de sa localisation entre les lobes hépatiques, une apparence davantage angulaire et une diminution de la dimension trans-atriale. Quel qu’en soit la cause, la microcardie suggère un état critique et l’administration d’un volume de remplacement approprié devrait être réalisé en urgence (MCMILLAN, 1994). L’épanchement péricardique est reconnue radiographiquement comme un élargissement symétrique et globoïde de la silhouette cardiaque et peut être observé chez des oiseaux atteints de chlamydiose, de polyomavirose, de tuberculose ou de néoplasie (MCMILLAN, 1994).

Page | 104
Forme : Les altérations de la forme du cœur peuvent suggérer une dilatation des chambres cardiaques ou des lésions granulomateuses (d’origine mycosique ou mycobactérienne) (COLES, et al., 2007). Radiodensité : Une augmentation de la radiodensité cardiaque est habituellement un signe d’inflammation ou de congestion (COLES, et al., 2007).
Les vaisseaux : Il n’existe pas de méthode objective d’évaluation des vaisseaux sanguins chez les oiseaux. L’interprétation se limite donc à l’évaluation subjective de variations de la taille ou de la radio-opacité des vaisseaux. Radiodensité : Les altérations des gros vaisseaux sont généralement visualisables comme une augmentation de la radiodensité (MANS, et al., 2007). Une telle augmentation se manifeste par l’existence de silhouettes circulaires superposées à la silhouette cardiaque en projection ventrodorsale et comme une silhouette radiodense en projection latéro-latérale (PEES, et al., 2009). Une augmentation de la radiodensité d’un gros vaisseaux indique, en particulier chez les perroquets très âgés, une calcification et possiblement un processus athérosclérotique (COLES, et al., 2007) (RITCHIE, et al., 1994). Cette modification radiographique n’est pas spécifique car une minéralisation de l’aorte doit également faire penser à une insuffisance rénale sévère ou une hypervitaminose D (PEES, et al., 2009). Elle n’est pas sensible puisque des cas d’infarctus aigus du myocarde, des syncopes et des crises convulsives ont été décrits chez des oiseaux avec de l’athérosclérose en l’absence de lésions radiographiques (MCMILLAN, 1994). Dimension : Une dilatation vasculaire peut être le reflet d’un processus athérosclérotique (TRISTAN, 2010), d’un anévrisme (anévrisme mycotique chez un eider (COURCHESNE, et al., 2009)) ou encore d’une néoplasie vasculaire (hémangiosarcome chez un amazone (HANLEY, et al., 2005)). Les dilatations apparaissent généralement sous la forme de large plage d’opacité de tissu mou. Il faut prendre garde à ne pas confondre les troncs brachiocéphaliques avec des granulomes fongiques (SMITH, et al., 1997). Le foie : Il est possible d’observer des modifications hépatiques telles qu’une hépatomégalie compatible avec une atteinte cardiaque (TRISTAN, 2010). L’ascite : La radiographie permet de mettre en évidence l’existence d’un liquide d’ascite, éventuellement secondaire à une affection cardio-vasculaire (PEES, et al., 2001). Les poumons : Des modifications des poumon, des sacs aériens (TRISTAN, 2010) tels qu’une congestion des vaisseaux pulmonaires, de l’œdème pulmonaire, un épanchement pleural sont des modifications compatibles avec une atteinte cardiaque (DONELEY, 2011).

Page | 105
b. Examen angiographique par fluoroscopie(angiocardiographie)
Indication : L’angiographie par fluoroscopie peut permettre la visualisation du cœur et de l’arbre vasculaire en temps réel. Il s’agit d’une procédure facile, peu coûteuse et qui peut être enregistrée en vue de la réalisation de mesures et d’analyses plus poussées (BEAUFRERE, et al., 2010). Bien qu’il existe peu d’études sur les avantages de l’angiocardiographie chez les oiseaux, cette technique pourrait avoir un grand intérêt lors de l’évaluation du système vasculaire d’une part mais aussi du cœur. En effet, si l’angiocardiographie ne peux pas remplacer l’échocardiographie, celle-ci peut apporter des informations supplémentaires, en particulier chez les oiseaux dont l’examen échocardiographique n’a pas pu être suffisamment détaillé (PEES, et al., 2006). Technique : Sous anesthésie générale, l’oiseau est positionné initialement en décubitus latéral gauche sur une table de fluoroscopie. Un bolus de 500 mg/kg d’agent de contraste iodé non ionique (soit deux fois la dose utilisée chez les Mammifères), est injecté à la vitesse de 1-2 ml/kg/s par un cathéter inséré dans la veine ulnaire, métatarsienne médiale (BEAUFRERE, et al., 2010), basilique ou jugulaire (PEES, et al., 2006) au cours de l’enregistrement vidéo. L’acquisition des images est réalisée avec une fréquence de 30 images par seconde pour une résolution optimale. Le même bolus est répété afin d’obtenir la vue ventrodorsale. Un marqueur calibré devra être gardé dans le champs au cours de l’enregistrement de la fluoroscopie, afin de pouvoir réaliser des mesures sans biais de grossissement (BEAUFRERE, et al., 2010). L’angiographie par soustraction digitale est une technique fluoroscopique utilisée en radiologie interventionnelle afin de visualiser clairement les vaisseaux sanguins situés au sein de tissus mous denses ou dans un environnement osseux. Les images sont produites en soustrayant de façon digitale une image dite ‘’pré-contraste’’ réalisée sans moyen de contraste aux images obtenues ultérieurement, une fois que le produit de contraste a été introduit dans le système vasculaire. Les bolus sont administrés de la même façon et en suivant les même dosage du produit de contraste que pour une angiographie fluoroscopique traditionnelle mais une option spéciale est sélectionnée sur la machine. Il en résulte une visualisation des vaisseaux contenant le produit de contraste seuls, sans arrière plan. Cette technique permet ainsi d’accentuer considérablement les contours des artères. Elle autorise une meilleure détection des artères les plus petites, non visibles avec une angiographie conventionnelle et plus particulièrement celles situées aux extrémités comme les pattes, les ailes et la tête. En revanche, la qualité des images a tendance à être facilement dégradée par de petits mouvement et du bruit (BEAUFRERE, et al., 2010). Interprétation : Cette technique permet d’observer les troncs brachiocéphaliques, l’aorte, les artères pulmonaires, les veines pulmonaires ainsi que la veine cave caudale. Les troncs brachiocéphaliques et l’aorte peuvent être vus en train de pulser avec les battements cardiaques. Des modifications luminales marquées peuvent être observées au cours du cycle cardiaque (BEAUFRERE, et al., 2010). Du fait de la rapidité de la fréquence cardiaque chez les oiseaux, l’évaluation de la contractilité peut être difficile. En revanche, il est possible de détecter une hypertrophie ventriculaire, une dilatation ventriculaire ou atriale, une sténose des vaisseaux ou des valves cardiaques, ainsi que des anévrismes vasculaires. (PEES, et al., 2006). Ainsi, la présence d’un anévrisme de l’artère coronaire droite a été mise en évidence chez un cacatoès blanc (Cacatua alba) grâce à l’angiocardiographie (VINK-NOOTEBOOM, et al., 1998). Cette technique a également été utilisé sur des aras bleu et or (Ara ararauna) (PHALEN, et al., 1996) ainsi que sur un cygne chanteur (Cygnus cygnus) (FISCHER, et al., 2005).

Page | 106
c. Angiographie fluorescéine Indication : L’angiographie fluorescéine est indiquée dans le diagnostic d’ischémie thrombo-embolique et peut aider à définir un site d’amputation afin de sectionner le membre dans une zone vascularisée. Elle sera déconseillée si le réseau vasculaire est peu accessible (HURLEY-SANDERS, et al., 2012). Technique : C’est une procédure invasive pouvant nécessiter le recours à une anesthésie générale mais l’examen peut également être réalisé sur animal vigile. De la fluorescéine sodium est injectée dans la veine métacarpale médiale gauche et le corps de l’oiseau est éclairé immédiatement après avec une lumière ultra-violette (HURLEY-SANDERS, et al., 2012). Interprétation : La peau doit fluorescer. Si tel n’est pas le cas, il est alors conclu à un trouble circulatoire. L’angiographie fluorescéine a été réalisée chez un flamant du Chili (Phoenicopterus chilensis) et a permis de diagnostiquer l’existence d’un trouble circulatoire compatible avec un trauma vasculaire ou une obstruction (HURLEY-SANDERS, et al., 2012).
d. Thermographie Indication : La thermographie est indiquées dans le diagnostic d’ischémie thrombo-embolique. C’est une technique rapide, non invasive mais subjective. Elle est limitée à l’évaluation des structures vasculaires superficielles (HURLEY-SANDERS, et al., 2012). Technique : Les images thermographiques peuvent être obtenues avec une caméra infra-rouge portable. La thermographie a été réalisée avec succès dans une pièce fermée dont la température était de 30.6°C (HURLEY-SANDERS, et al., 2012). Interprétation : La comparaison des images thermographiques permet d’évaluer la radiation de chaleur des différents endroits du corps et de déceler des anomalies de la circulation sanguine. La thermographie évalue les répercussions en surface de l’apport sanguin calorifique aux tissus. En conséquence c’est une technique peu sensible lors de l’évaluation de la viabilité des structures vasculaires profondes nichées au sein de tissus épais. Enfin, il faut prendre garde aux artefacts de lecture dûs à des courants d’air, aux positions et mouvements du patient, à l’existence de traitements médicaux, aux variations d’emplumement de la surface cutanée ou à toute autre raison exogène pouvant provoquer un réchauffement ou un refroidissement de zones corporelles. L’angiographie fluorescéine a été réalisée chez un flamant du Chili (Phoenicopterus chilensis) et a permis de diagnostiquer l’existence d’un trouble circulatoire compatible avec un trauma vasculaire ou une obstruction (HURLEY-SANDERS, et al., 2012).

Page | 107
e. Examen échographique En raison de l’amélioration des progrès techniques au cours des dernières années, l’échocardiographie chez les oiseaux est devenu un outil diagnostique valable et important. Il est sécuritaire et simple à réaliser (PEES, 2005). Indications : L’examen échocardiographique est indiqué lorsqu’une cardiomégalie est objectivée. Il permet de faire la distinction entre dilatation, hypertrophie, épanchement péricardique ou masse cardiaque (STRUNK, 2003). L’échocardiographie est également indiquée lorsqu’un souffle cardiaque est entendu, lors d’épanchement cœlomique (DE WIT, et al., 2005), d’arythmie cardiaque ou d’une cyanose (PEES, 2005). Matériel : La fréquence cardiaque élevée et la petite taille des structures internes du cœur aviaire imposent des exigences élevées sur l’équipement technique (PEES, 2005). La sonde : Des sondes avec de petites surfaces couplantes (sonde microconvexe ou sonde à balayage électronique sectoriel par déphasage) et des fréquences d’au moins 7.5 MHz sont nécessaires pour réaliser une échocardiographie correcte chez les oiseaux pesant jusqu’à 1000 grammes. L’utilisation d’un transducteur multi-élément de 8 à 11 MHz (BEAUFRERE, et al., 2010) est possible. Toutefois si une fréquence ultrasonore plus élevée améliore la résolution de l’image, elle provoquera également une diminution de la fréquence d’image et la profondeur maximal d’examen. La sonde devra être équipée d’une fonction Doppler spectral et couleur (PEES, 2005). Le système échographique : L’échographe devra être capable de produire au moins 100 images par seconde (BEAUFRERE, et al., 2010). La haute fréquence d’image est nécessaire pour obtenir des images du cycle cardiaque précise (fin de systole et de diastole par exemple). En effet, chez les oiseaux ayant une fréquence cardiaque pouvant atteindre 600 bpm (10 battements par seconde), une fréquence d’image de 100 images par seconde fournie seulement 6 images par cycle. Le système devra posséder un système d’enregistrement et une fonction gâchette (enregistrement électrocardiographique). La fonction gâchette est utile pour corréler les images et mesures cardiaques avec les différentes étapes du cycle cardiaque (holodiastolique, holosystolique) (PEES, 2005). L’animal : Un tractus gastro-intestinal rempli peut perturber la pénétration des ondes ultrasons. Les oiseaux devront donc être mis à jeûn avant l’examen. Chez les psittacidés, un jeûne de 2 à 4 heures est recommandé. Les pigeons devront jeûner 12 heures et les rapaces jusqu’à 48 heures (PEES, 2005). Chez la plupart des oiseaux, l’échographie peut être réalisée sous contention manuelle (DE WIT, et al., 2005). Une analyse en mode B ne nécessitera une anesthésie que pour les oiseaux les plus stressés. Un examen Doppler devra être réalisé sous anesthésie générale afin de réduire l’influence de la manipulation et de la contention sur la vitesse du flux sanguin. Le patient sera positionné aussi droit que possible afin de limiter l’influence de la position sur le fonctionnement cardiaque. En particulier, le décubitus dorsal provoque des problèmes circulatoires importants chez les oiseaux atteints de maladie cardiaque. Les gels acoustiques hydrosolubles commerciaux sont bien tolérés chez les oiseaux (PEES, 2005). On évitera par ailleurs l’utilisation d’alcool chez les oiseaux anesthésiés afin de limiter l’hypothermie. Le plus souvent, le décubitus ventral autorise la réalisation des meilleures images. Toutefois, si le cœur n’est pas visible de façon satisfaisante, il est possible de modifier le décubitus de l’oiseau (latéral voire dorsal) (STRAUB, et al., 2003).

Page | 108
Technique : Des protocoles d’échographie transcœlomique et transœsophagienne ont été bien établis (BEAUFRERE, et al., 2010) (PEES, 2005) (PEES, et al., 2006) (PEES, et al., 2004). Echographie trans-cœlomique (ou trans-cutanée) : L’abord transcutané est compliqué par la présence du squelette (PEES, et al., 2004) et des sacs aériens. Cette technique gagne en intérêt lors d’hépatomégalie ou d’épanchement, car il n’y a alors plus d’air entre la sonde et le cœur (OGLESBEE, et al., 2001). En raison des particularités anatomiques du cœur aviaire et de son environnement dans la cavité cœlomique, les vues standardisées recommandées chez les Mammifères ne peuvent être utilisées chez les Oiseaux. En échographie transcutanée les modes utilisables sont le mode B, le Doppler couleur et le Doppler pulsé. A ce jour, le mode B (échocardiographie 2D) est une technique d’examen bien établie. Des études récentes ont prouvé l’intérêt du mode Doppler comme une technique fiable d’évaluation du flux sanguin cardiaque chez les patients aviaires. En particulier, le doppler pulsé a été étudié et des cas cliniques rapportent l’utilisation du mode Doppler couleur (PEES, 2005). Le mode M (ou TM) est un outil utilisable pour évaluer l’épaisseur de la paroi et sa contractilité chez les mammifères mais elle n’est pas possible par abord transcutané chez les oiseaux car le cœur ne peut être visualisé qu’en vue longitudinale et semi-transversale. La visualisation du cœur est obtenue par deux approches possible (OGLESBEE, et al., 2001) : - L’approche ventro-médiane : La sonde est située dans le plan sagittal, juste caudalement au sternum et le plan de coupe et dirigé cranio-dorsalement vers le cœur (PEES, et al., 2004). Selon les cas, il pourra être nécessaire de retirer des plumes afin de permettre un meilleur contact entre la sonde et le patient (TRISTAN, 2010). Toutefois, cette zone est une aptérie chez de nombreuses espèces (PEES, 2005). Sous cette approche, le foie sert de fenêtre acoustique et les poumons sont situés dorsalement. On réalise habituellement deux plans de coupe : une vue horizontale et une vue verticale. La vue horizontale dite « apicale quatre chambres » permet de visualiser les atria, la racine aortique, les deux ventricules, les deux atria, les deux valves atrio-ventriculaires ainsi que le septum inter-ventriculaire mais il peut être difficile de délimiter les atria (PEES, et al., 2006). La vue verticale dite « apicale deux chambres » permet de visualiser les cavités du cœur gauche, la valve atrio-ventriculaire gauche et l’aorte (KRAUTWALD-JUNGHANNS, et al., 1995). Le ventricule droit peut être observé mais les images ne sont pas de qualité suffisante pour permettre la réalisation de mesure. Dans cette seconde vue, des artefacts de réflexion liés à la présence du sternum peuvent produire un « second cœur » localisé en dehors de la cavité thoraco-abdominale (artefact d’image en miroir) (PEES, et al., 2006).
- L’approche para-sternale : Cette approche complémentaire est utilisable chez les oiseaux ayant un espace suffisant entre la dernière côte et l’os pelvien (par exemple chez les pigeons et certaines espèces de rapaces). La sonde est positionnée du côté droit avec la patte étirée soit en arrière soit en avant et le plan du faisceau est dirigé cranio-médialement (PEES, 2005). La sonde doit être légèrement pressée contre la paroi corporelle pour comprimer les sacs aériens sous-jacents (COLES, et al., 2007). Cette approche montre le cœur en vue semi-transverse et peut être utile pour examiner les valves et les vaisseaux (PEES, 2005). On distingue deux vues. La vue « grand axe para-sternale droite » permet, en étudiant différentes section par balayage du cœur de visualiser les deux ventricules et leur paroi, le septum inter-ventriculaire, les valves atrio-ventriculaires, l’atrium droit, l’aorte et ses valves ainsi que l’atrium gauche. La vue « petit axe para-sternale droite » permet de visualiser, par l’étude de différents plans, l’aorte, les artères pulmonaires, les valves atrio-ventriculaires, les deux ventricules et l’atrium gauche (KRAUTWALD-JUNGHANNS, et al., 1995).

Page | 109
Echographie trans-œsophagienne : L’échographie trans-œsophagienne constitue une alternative à l’abord trans-cœlomique et permet d’obtenir de meilleures images. Il convient d’utiliser une sonde d’échographie trans-œsophagienne (sonde échographique multi-plans à balayage électronique contenu dans l’extrémité d’un tube flexible) pédiatrique (3-7 MHz) possédant les modes M, B et dopplers (pulsé, continu et couleur). Dans cette approche, la proximité du cœur avec l’œsophage et le proventricule permet d’éviter les contraintes liées à la présence des sacs aériens chez les oiseaux. La sonde est introduite dans l’œsophage du patient et ajustée au niveau du cœur. Cette manœuvre est plus facile à réaliser chez les oiseaux ne possédant pas de jabot (oies, canards, chouettes rayées) car le passage de l’entrée thoracique est plus aisé. Dans tous les cas, une extension du cou, des connaissances sur les caractéristiques anatomiques de l’œsophage et une redirection de la sonde par palpation et manipulation au travers de l’œsophage devraient aider à positionner convenablement la sonde. Cette approche est la seule permettant de réaliser des échographies en mode M du ventricule gauche et de la racine aortique. Les vues transverses et longitudinales obtenues par échocardiographie trans-œsophagienne avec la sonde placée en position crâniale fournissent des détails suffisants de la racine aortique chez la plupart des espèces (BEAUFRERE, et al., 2010). Limitée par la taille de l’oiseau (taille du bec et de l’entrée de la poitrine) cette technique est déconseillée chez les psittacidés pesant moins de 900 grammes. Elle est peu invasive mais nécessite une anesthésie générale et donc un oiseau cliniquement stable. Elle est par ailleurs impossible chez les pélicanidés dont les sacs aériens sont trop développés (BEAUFRERE, et al., 2010). On décrit trois positions :
La position caudale : La sonde est placée dans la lumière du proventricule puis fléchie ventralement pour imager le cœur au travers du foie. Il est alors possible d’obtenir des coupes transverses des ventricules à différents niveaux et des coupes longitudinales du cœur incluant l’apex cardiaque. La vue transverse obtenue en position caudale fournit une section en tranche du ventricule gauche similaire à celle obtenue par abord trans-thoracique para-sternale droit, c'est-à-dire une vue petit-axe. Cette vue n’est pas visualisable par approche ventromédiane. Les muscles papillaires du ventricule gauche ne sont pas proéminents chez les oiseaux (à l’exception des canards et du ara militaire). En conséquence, il n’est pas possible de réaliser des mesures à hauteur de ces muscles (épaisseur de la paroi, dimension de la chambre) comme chez les mammifères. Il a été proposé en guise d’alternative de balayer le ventricule gauche de l’apex à la pointe des valvules constituants la valve auriculo-ventriculaire et de sélectionner l’image montrant le plus grand diamètre de la cavité cardiaque pour y réaliser les mesures. Le mode M a ainsi été utilisé pour obtenir les mesures systoliques et diastoliques de la paroi libre du ventricule gauche, de l’épaisseur du septum inter-ventriculaire et du diamètre interne du ventricule gauche (BEAUFRERE, et al., 2010). La position moyenne : La sonde est placée en position droite (neutre) puis est légèrement retirée crânialement dans l’œsophage thoracique pour être placée crânialement à la base du cœur. Une vue transverse à ce niveau montre les valves atrio-ventriculaires et fournit des images similaires à une coupe 4-cavités mammalienne. La valve atrio-ventriculaire droite est clairement visualisable. Une coupe longitudinale est également réalisable à ce niveau et montre la racine aortique. Il est alors possible de mesurer la dimension de la lumière aortique. Le doppler couleur permet d’évaluer le fonctionnement des valves atrio-ventriculaires. Le doppler spectral permet de mesurer les flux atrio-ventriculaires sanguins (BEAUFRERE, et al., 2010).

Page | 110
La position crâniale : La position crâniale est obtenue en déplaçant la sonde crânialement et en réalisant une légère dorso-flexion. Là encore, il est possible de réaliser une vue transverse et une vue longitudinale. La vue crâniale est difficile à obtenir chez les grands anatidés compte tenu de la présence des sacs aériens. La vue longitudinale obtenue en position crâniale, permet de visualiser clairement la base de l’aorte ascendante. Des mesures du diamètre et de la paroi aortiques sont obtenues à l’aide du mode M. Cette vue permet de nouvelles évaluations de la fonction systolique, de l’épaisseur de la paroi ventriculaire gauche, de la taille des cavités atriales et ventriculaires, du diamètre de l’aorte ainsi que de la présence de lésions athérosclérotiques. Le mode doppler couleur permet d’évaluer le fonctionnement de la valve aortique en coupe transverse. Le doppler spectral permet de mesurer les flux d’éjections aortique et pulmonaire (BEAUFRERE, et al., 2010).
Cette technique comporte certains risques. Ainsi, il est possible de déclencher une arythmie cardiaque autolimitante lors de la pression de la sonde sur la base du cœur à l’occasion d’une ventroflexion. Il est également possible de perforer l’œsophage à la hauteur de l’entrée de la poitrine. Toutefois, bien que plus risquée qu’une approche transcutanée, cette technique offre la possibilité de réaliser davantage de vue, des images de meilleure qualité, des enregistrements en mode-M ainsi que des observations plus détaillées des valves. Elle pourrait se révéler plus intéressante dans le diagnostic d’athérosclérose. Toutefois sa réalisation est compliquée chez les perroquets dont l’entrée thoracique semble être relativement petite. Cette difficulté s’accompagne de risque de perforation œsophagienne ou d’éversion du jabot dans le proventricule (BEAUFRERE, et al., 2010). Echographie avec produit de contraste : L’échocardiographie avec produit de contraste a été évaluée chez plusieurs espèces d’oiseaux (BEAUFRERE AND PARIAUT, unpublished data). Cette technique repose sur l’injection lente par voie intraveineuse de microsphères lipidiques contenant du perflutren à la dose de 0,1ml/oiseau après avoir agité la solution à l’aide d’un dispositif d’activation mécanique. Au cours de l’examen échographique, une puissance acoustique faible doit être utilisée (index mécanique de 0.2 à 0.3) afin de limiter la perturbation des microsphères intravasculaires. La petite taille des microsphères autorise celle-ci à traverser les lits capillaires pulmonaires. Elles peuvent, en conséquence, être visualisées dans les chambres cardiaques gauches. Cela représente un avantage par rapport à l’injection d’un petit volume de solution saline agité, qui opacifie seulement le ventricule droit (ceci permet toutefois de mettre en évidence des shunts intra-cardiaque et extra-cardiaque droit-gauche). L’addition d’un produit de contraste peut aider à mieux délimiter le bord de l’endocarde du ventricule gauche et fournit des mesures plus précises de la racine aortique et du diamètre ventriculaire interne systolique. Le produit de contraste est injecté lentement à l’aide d’un cathéter périphérique alors que le cœur est observé par échographie transcoelomique ou transoesophagienne. Le contraste dure quelques minutes et les microsphères peuvent être détruites en augmentant le puissance acoustique si cela est jugé nécessaire. Aucun effet secondaire n’a été observé (BEAUFRERE, et al., 2010).

Page | 111
Interprétation : Dans de nombreux cas, la visualisation échographique du cœur chez les oiseaux atteints de maladie cardiaque est bien plus simple que chez les oiseaux sains. En effet, l’augmentation de la taille des organes (cœur, foie) et l’accumulation de liquide dans la cavité thoraco-abdominale améliore la qualité des images. En cas de distension abdominale, des sondes plus grosses que celles recommandées peuvent être utilisées pour l’examen (PEES, 2005). L’utilisation des différents modes échocardiographiques dans les différents abords permet d’évaluer à la fois la structure (dimension des cavités, structure pariétale), le fonctionnement (flux sanguins, fonctionnement valvulaire) et les mouvements du cœur (contractilité)(TRISTAN, 2010). Elle permet également d’évaluer la paroi des vaisseaux centraux et la structure péricardique (BEAUFRERE, et al., 2010). Interprétation du Mode B dit échocardiographie 2-dimensions Le mode B fournit des coupes du cœur et permet l’évaluation anatomique des structures cardiaques (DONELEY, 2011). On évaluera la taille des cavités, la relation entre les ventricules, l’épaisseur du septum inter-ventriculaire et la contractilité des ventricules. De façon complémentaire, la morphologie et le fonctionnement de la valve atrio-ventriculaire gauche, de la valve aortique et de la valve atrio-ventriculaire droite musculaire peuvent être évaluées si la qualité de l’image est suffisante (PEES, 2005). Echogénicité : Les vaisseaux peuvent être évalués et l’échocardiographie d’avoir un intérêt pour diagnostiquer l’existence d’un processus athérosclérotique. Les vaisseaux apparaissent alors épaissis et hyperéchogènes. Lors d’inflammation, l’évaluation échographique montre une augmentation de l’échogénicité. L’épanchement péricardique est clairement visualisable comme une structure anéchogène entre le cœur et le péricarde (PEES, et al., 2009). Mesures : Puisque les coupes du cœur en mode M sont difficilement réalisables en pratique courante, les mesures doivent être réalisées à partir d’images 2D (PEES, et al., 2006). Pour mesurer les paramètres cardiaques spécifiques, la position de la sonde doit être ajustée jusqu’à observer l’expansion maximale des chambres. Les paramètres doivent être mesurés en diastole et en systole au point le plus large. Les bords externes de la paroi du cœur sont souvent difficiles à distinguer des tissus sous-jacents. On utilisera donc la méthode du bord interne. Il conviendra de réaliser trois mesures lors de trois cycles distincts dont on calculera la moyenne. Les valeurs sont alors comparées aux intervalles de référence (cf. Annexe I). Le diamètre de la racine aortique a été documenté (PEES, 2005) (PEES, et al., 2006) (PEES, et al., 2004). Les mesures du ventricule gauche sont possibles dans toutes les vues mais sont plus faciles à obtenir en vue verticale. Chez les perroquets, il y a une forte corrélation entre la longueur sternal et la longueur du ventricule gauche : LVG = 0.5 + 0.33 x longueur sternale. La mesure de la longueur du ventricule droit est plus difficile car il est visualisé comme un triangle aigu. De plus, la valve atrioventriculaire musculaire est, en raison de ses mouvements un bord imprécis lors de la réalisation des mesures (PEES, et al., 2006). L’épaisseur du SIV peut donner des informations concernant une hypertrophie ventriculaire. La mesure des atria est imprécise et donc d’intérêt limité. De plus, l’importance de la contraction atriale lors du remplissage ventriculaire est mineure et l’évaluation atriale peu importante (PEES, et al., 2006). Une augmentation de la pression sanguine dans la grande circulation conduit souvent à une congestion hépatique (dilatation visible des vaisseaux hépatiques) et à de l’ascite. Les mesures des paramètres cardiaques étant effectuées sur des cœurs de petite taille à fréquence rapide à partir de plan d’imagerie peu standardisées, le suivi de l’évolution doit être réalisé par le même opérateur et la même technique d’imagerie. Une différence d’au moins 20% sera nécessaire pour attester d’une évolution (BEAUFRERE, et al.).

Page | 112
Calculs : Les mesures obtenues permettent de calculer la fraction de raccourcissement (FR). Il s’agit d’un paramètre fonctionnel permettant d’évaluer l’efficacité de la contraction ventriculaire. Elle est calculée comme la réduction relative de la largeur transversale du diamètre du ventricule gauche au cours de la systole, en relation avec le diamètre en diastole (%FR = (diastole-systole)x100 / diastole). Les valeurs obtenues diffèrent de celles des Mammifères ce qui pourrait être dû à des particularités anatomiques. La contractilité du ventricule droit est significativement plus élevée que celle du gauche. Pour les perroquets, la valeur est relativement constante (PEES, et al., 2006). L’échocardiographie Temps-Mouvement ou mode M Le mode M produit des images perpendiculaires au mode B, autorisant ainsi la quantification des dimensions cardiaques et des événements cardiaques au cours du temps (DONELEY, 2011). Des études ont prouvé la possibilité de sa réalisation mais aucune valeur de référence n’a été publiée (BEAUFRERE, et al., 2010). L’échocardiographie Doppler ou mode Doppler Le doppler couleur facilite l’identification des turbulences. Il peut être utilisé pour évaluer la présence d’insuffisance valvulaire, de sténose, de shunt (DONELEY, 2011), de fluide péricardique, la contractilité myocardique, l’épaisseur de la paroi ventriculaire, la taille des chambres ou encore l’existence d’anévrisme (DE WIT, et al., 2005). Il est également utile pour positionner la fenêtre en vue d’une analyse doppler spectral afin de déterminer le degré d’une insuffisance valvulaire. Le Doppler couleur a ainsi été utilisé avec succès lors du diagnostic d’anévrisme d’une artère coronaire chez un oiseau. Avec l’approche ventromédiane standard, l’influx diastolique dans les ventricules et affiché en rouge (flux vers le la sonde) et le flux de sortie systolique est bleu (flux s’éloignant de la sonde) (PEES, et al., 2006). Toutefois, la fréquence des images diminue lors de l’utilisation de ce mode. Ce problème limite significativement son intérêt en échocardiographie aviaire (PEES, 2005). Le doppler spectral : C’est le gold standard pour mesurer le flux sanguin de façon non invasive en médecine mammalienne. Les signaux sont affichés dans des graphiques en deux dimensions affichant le temps en abscisse et la vélocité en ordonnée. Deux types de doppler spectraux présentent un intérêt : les doppler pulsé ou continu.
Le doppler pulsé est limité à la mesure de vitesse inférieur à 2 m/s (PEES, et al., 2006) mais autorise les mesures dans un volume défini par l’opérateur. Pour mesurer la vitesse, le flux sanguin doit être aussi parallèle que possible à la direction des ondes ultrasons. Pour cette raison, l’approche ventromédiane est idéale (PEES, 2005). L’influx diastolique dans les ventricules droit et gauche ainsi que l’effux aortique systolique a été mesuré chez les perroquets et les rapaces (annexe I) (PEES, 2005) (PEES, et al., 2006) (STRAUB, et al., 2004). Lors de l’influx mitral, deux pics peuvent être enregistrés, une onde E correspondant au remplissage ventriculaire passif, et l’onde A correspondant à la contraction atriale). E :A était positive chez les aras et les gris du Gabon mais négative chez les cacatoès (PEES, et al., 2006). Les ondes E et A ne sont pas toujours différenciables (STRAUB, et al., 2004). Le flux sanguin dans l’artère pulmonaire n’a pu être mesuré que dans de rares cas (PEES, et al., 2006). Nous signalerons que le flux sanguin intra-cardiaque augmente significativement chez les oiseaux stressés (STRAUB, et al., 2003). Dès lors, il est recommandé de réaliser l’échocardiographie Doppler sous anesthésie générale (PEES, et al., 2006). Le doppler continu est utile pour détecter des flux sanguins de très grande rapidité mais ne permet pas de mesurer dans certaines régions. Aucune étude n’a été publiée concernant l’utilisation du Doppler continu à ce jour.

Page | 113
f. Examen angiographique par tomodensitométrie Indication : L’angiographie par tomodensitométrie permet une excellente évaluation de tous les vaisseaux principaux et sera indiquée lors de vasculopathie. L’adjonction d’un agent de contraste améliore grandement la visualisation des artères, des veines ainsi que de leur lumière. Aucun cas clinique ne documente l’utilisation du CT-Angiographie dans un contexte clinique chez les oiseaux. Toutefois, son utilisation pourrait se révéler inestimable dans l’évaluation de sténose luminale artérielle, d’anévrisme, de dilatation artérielle et de calcification des parois. Elle est peu informative pour l’évaluation cardiaque, car sa fréquence de battement rend les images difficilement interprétables (BEAUFRERE, et al., 2010). Le CTA haute résolution a également été utilisée chez les oiseaux pour décrire leur anatomie vasculaire normale (HOLLIDAY, et al., 2006). Technique : Le patient doit être anesthésié afin de placer un cathéter en vue de l’administration du produit de contraste. Le protocole nécessite 4 scans : Préparatifs : On réalisera un scan de localisation corps entier en plan orthogonaux suivi par un scan pré-contraste en utilisant un protocole de scan abdominal prédéfini. On déterminera ensuite le temps d’arrivée du produit de contraste en réalisant un scan axial dynamique à une coupe prédéterminée (au niveau de l’aorte abdominal, dorsalement au proventricule) avec un bolus initial unique de iohexol de 1 ml/kg sur une seconde. Le moment du démarrage du scan dynamique doit coïncider avec l’initiation de l’administration du produit de contraste (BEAUFRERE, et al., 2010). L’angiographie par CT est ensuite réalisée en utilisant les mêmes paramètres de scan que pour les séries d’étude pré-contrastes avec 3 ml/kg d’agent de contraste administré sur 3 secondes. Une pompe à perfusion peut être utilisée mais peut être problématique en regard des petits volumes d’injection requis chez les oiseaux. Aussi, l’injection manuelle est préférée par certains auteurs (BEAUFRERE, et al., 2010). Le début de l’administration du produit de contraste doit précéder le début du scan par le temps du pic d’amplification préalablement déterminé (Estimating intrathoracic arterial diameter using CT angiography in Hispaniolan Amazon Parrots (Amazona ventralis) in C.R. Congrès ''AAV Conference'', 2009). En pratique, chez les oiseaux de taille petite à moyenne, le temps nécessaire à l’obtention du pic d’amplification de l’aorte est très court en raison de la vitesse de circulation du contraste dans l’arbre vasculaire et il est recommandé de réaliser l’angiographie par CT immédiatement après l’administration du produit de contraste. Des séries de reconstruction additionnelles avec des coupes plus fines (par exemple 0.625mm) et des algorithmes pour tissus-mous sont recommandés. Interprétation : L’angiographie par tomodensitométrie permet la visualisation de toutes les artères principales et de leur anatomie chez les psittacidés et les rapaces. Des valeurs de références ont été publiées pour les diamètres artériels des troncs brachiocéphaliques, de l’aorte ascendante, de l’aorte abdominale et des artères pulmonaires. L’utilisation d’une fenêtre médiastinale ou d’une fenêtre d’angiographie manuelle est recommandée pour obtenir des mesures plus précises et répétables (Estimating intrathoracic arterial diameter using CT angiography in Hispaniolan Amazon Parrots (Amazona ventralis) in C.R. Congrès ''AAV Conference'', 2009). Bien que les valeurs de références n’existent pas à ce jour, cette technique permet également la visualisation des veines caves crâniale et caudale, des artères carotides, de l’artère mésentérique ainsi que certaines de ses ramifications, et plusieurs des artères plus petite de l’organisme aviaire. Le reformatage 3D du cœur et des artères centrales sont également possibles et peuvent aider au diagnostic d’anévrisme et d’anomalie vasculaire congénitale (BEAUFRERE, et al., 2010).
g. Imagerie par résonnance magnétique

Page | 114
L’angiographie par résonnance magnétique n’a pas été évaluée en contexte clinique chez les oiseaux à ce jour. Une étude relative à la réalisation d’IRM chez des pigeons normaux a été réalisée. Celle-ci révèle que les vascularisations hépatique et rénale sont bien visibles (ROMAGNANO, et al., 1996). Toutefois, les artefacts de mouvements empêche la réalisation d’images adéquates du cœur. De plus, l’imagerie par résonnance magnétique ne s’est pas révélée être d’une bonne valeur diagnostique pour le système vasculaire, du fait de la circulation rapide du produit de contraste (gadolinium) à travers le réseau de vaisseau d’une part, et la petite taille des vaisseaux à investiguer d’autre part. La réalisation d’images diagnostiques de qualité pour des vaisseaux aussi petits nécessiterait des champs magnétiques plus élevés (3 Teslas ou plus). Cette exigence technique limiterait encore plus son utilisation clinique (BEAUFRERE, et al., 2010).
2. Examen endoscopique Indication : La cœlioscopie permet d’observer directement le cœur et certaines artères. La recherche d’anomalie macroscopique visible à la surface du cœur (suspicion de goutte viscérale (PEES, et al., 2006) ou d’hémangiosarcome (HANLEY, et al., 2005)) sont des indications à la réalisation d’une endoscopie. Technique : La coelioscopie est un acte chirurgical réalisé sous anesthésie générale. Les oiseaux devront être mis à jeun de sorte que le tractus digestif soit vide lors de la procédure. En effet, si tel n’est pas le cas, les organes digestifs remplis limiteront la visualisation de certaines parties du coelome. L’oiseau est placé en décubitus latéral vrai, avec les ailes rabattues dorsalement. On utilisera un endoscope rigide, de 170 à 190 mm de long et dont le diamètre sera adapté à l’espèce (1.9 mm chez des oiseaux de moins de 100g, 2.5 mm chez des oiseaux de 100g à 4kg et 4 mm chez des oiseaux de plus de 4 kg). La peau est préparée de façon chirurgicale avant d’insérer l’endoscope. Quatorze abords endoscopiques ont été décrits mais seuls trois présentent un intérêt lors de l’évaluation du système cardiovasculaire. Il sera possible de réaliser des biopsies ou des ponctions au cours de la procédure (TAYLOR, 1994): - les abords latéraux standards (HERNANDEZ-DIVERS, et al., 2004) (TAYLOR, 1994) permettent de visualiser les vaisseaux assurant l’apport sanguin aux organes abdominaux, en se situant dans les sacs aériens abdominaux. Il est ainsi possible d’observer l’aorte abdominale, les artères iliaques, l’artère mésentérique, les artères rénales, la veine cave caudale ainsi que les veines ischiatiques. De plus, les artères et veines pulmonaires peuvent être observées à proximité de la base du cœur, en positionnant l’endoscope dans les sacs aériens thoraciques crâniaux gauche et droit. - l’abord interclaviculaire (TAYLOR, 1994) (HERNANDEZ-DIVERS, et al., 2004) est celui qui est le moins fréquemment utilisé. Cependant, il permet d’accéder à la base du cœur en passant par le sac aérien inter-claviculaire et devrait être considéré comme un abord de choix afin d’évaluer les grosses artères, les carotides ainsi que la base du cœur. A cet endroit, les artères crâniales et les artères majeures peuvent toutes être facilement identifiées, à l’instar des troncs brachiocéphaliques, de l’aorte ascendante, des artères brachiales, des artères carotides, des artères pulmonaires et des veines jugulaires. L’accumulation de graisse à la base du cœur peut rendre significativement plus difficile la visualisation des artères par l’abord interclaviculaire (BEAUFRERE, et al., 2010). - l’abord par la ligne ventrale permet de passer par les sacs hépato-péritonéaux ventraux et sera à privilégier lors d’ascite, de sorte à éviter toute fuite dans les sacs aériens. Interprétation : Comme lors d’un examen nécropsique, on s’attachera à évaluer les variations de taille, de forme, de couleur et de consistance des organes. Il sera ainsi possible d’observer une dilatation cardiaque, un épanchement péricardique, la présence de néoplasie, de granulome ou une décoloration artérielle.

Page | 115
D. Examen électrocardiographique
1. Indications et limites
L’électrocardiographie présente de nombreuses indications. Elle permet de mesurer la fréquence cardiaque chez des espèces pour lesquelles ce paramètre peut être difficilement calculable à l’auscultation et le suivi anesthésique cardiaque avant, pendant et après une opération chirurgicale. Elle renseigne également sur les modifications structurelles du myocarde telles qu’un élargissement cardiaque (cardiomégalie) secondaire à l’hypertrophie de l’une des chambres cardiaques, un épanchement pleural ou péricardique. C’est une technique de choix pour le diagnostic des arythmies cardiaques, le suivi de ces maladies et l’évaluation des effets de molécules cardioactives. Elle peut être utilisée afin de repérer la diastole et la systole lors d’une échocardiographie (PEES, et al., 2009). Enfin, le myocarde étant très sensible à l’hypoxie, l’électrocardiographie peut être un indicateur fiable de l’oxygénation (DONELEY, 2011) (ZANDVLIET, 2005). Toutefois, et bien qu’abondamment décrite chez les oiseaux l’électrocardiographie est peu utilisée en pratique courante à ce jour en médecine aviaire du fait de ces limites. La première limite est la difficulté d’interprétation : d’une part il existe des arythmies physiologiques (provoquées par le stress de l’examen) et d’autre part, la différence entre un ECG normal et anormal est parfois floue (PEES, et al., 2001). De plus, les informations obtenues lors d’un ECG proviennent directement des phénomènes électriques myocardiques. En conséquence, cet examen ne peut être utilisé pour évaluer les performances mécaniques du cœur et ne permet pas d’évaluer les structures cardiaques telles que les valves et l’endocarde, si ce n’est de façon indirecte. En outre, des trouvailles ECG isolées permettront difficilement de diagnostiquer une maladie cardiaque précise et il faudra les mettre en relation avec les résultats d’autres examens (ZANDVLIET, 2005). Enfin, si la plupart des affections cardiaques se manifestent par des anomalies électrocardiographiques (LUMEIJ, et al., 1994), un ECG normal n’exclut pas l’existence d’une maladie cardiaque (ZANDVLIET, 2005).
2. Matériel
Volt-mètre : En plus des conditions standards relatives à une machine à ECG (sécurité de l’animal et de l’opérateur, réglage de la sensibilité) il est de la plus grande importance que la vitesse de déroulement du papier soit suffisante. Compte tenu de la fréquence cardiaque élevée, le papier doit être déroulé à 50mm/sec au minimum et certains auteurs conseillent de ne pas réaliser d’enregistrement en dessous de 100 mm/sec. Pour les espèces plus petite, il est possible de devoir calibrer la vitesse de défilement à 200mm/sec. L’interprétation des tracés sera plus fiable si la machine est capable d’enregistrer les dérivations bipolaires et les dérivations unipolaires augmentées en même temps (ZANDVLIET, 2005). L’enregistrement est réalisé en routine avec une sensibilité de 10 mm/mV (SCHOEMAKER, et al., 2005). Electrodes : Il est possible d’utiliser des pinces crocodiles atraumatiques (émoussées) avec du gel échographique (UZUN, et al., 2004) ou de l’alcool (HASSANPOUR, et al., 2010) pour augmenter la conduction électrique. Toutefois des électrodes-aiguilles placées en position sous-cutanées sont plus intéressantes que les pinces alligator en médecine aviaire (DONELEY, 2011). Des patchs ECG peuvent également être utilisés (SCHOEMAKER, et al., 2005).

Page | 116
Animal : Les tracés sont dépendants de la position du patient et ce d’autant plus chez les oiseaux que leur thorax n’est pas du tout cylindrique. Ainsi, l’utilisation de l’ECG à des fins diagnostiques nécessite que l’animal soit placé dans une position standard (SCHOEMAKER, et al., 2005). Dans la plupart des cas, le positionnement, le temps nécessaire à l’enregistrement et la prévention des artefacts (tremblements, mouvements) ne peuvent être obtenus que sous anesthésie (OGLESBEE, et al., 2001). Toutefois, il doit être signalé que l’anesthésie peut provoquer des artefacts sur le tracé ECG. On pourra ainsi observer une modification de la fréquence cardiaque et une prolongation de l’intervalle Q-T. Dépendamment du type d’agent anesthésique utilisé, il sera possible d’observer des arythmies (SCHOEMAKER, et al., 2005) telles que des blocs atrio-ventriculaires du second et du troisième degré, un arrêt sinusal, une dépression de l’onde T ou des contractions atriales prématurées avec l’isoflurane (PEES, et al., 2009). Il est néanmoins possible de réaliser des ECG de qualité convenable en utilisant des supports en V (STURKIE, 1949), la télémétrie (SAVORY, et al., 1997) (SAWBY, et al., 1974) ou même la contention manuelle (utilisée chez les rapaces, qui développent une réaction d’immobilité tonique ou des patients léthargiques) (RODRIGUEZ, et al., 2004).
3. Technique
Principe : L’activité électrique enregistrée résulte de la dépolarisation de cardiomyocytes individuels donnant naissance à de petites modifications du potentiel extracellulaire. Lorsque deux électrodes sont attachées à la surface du corps, un circuit est formé, et l’utilisation d’un amplificateur associé à un volt-mètre permet de mesurer les changements de voltage entre ces deux électrodes. L’enregistrement du voltage de ces signaux en continu, aboutit à la production d’un ECG. L’ensemble formé par les deux électrodes est appelé une dérivation (SCHOEMAKER, et al., 2005). Le mouvement des charges électriques dans le cœur est un événement se déroulant dans les trois dimensions de l’espace et la charge résultante nette à tout moment du cycle cardiaque peut être représentée par un vecteur ayant une direction, un sens et une amplitude. Ce vecteur cardiaque est représenté par une flèche décrivant ses caractéristiques : la direction est représentée par la direction de l’axe, la polarité par la tête de la flèche et la magnitude par la longueur de la flèche. L’ECG de surface est alors la projection du vecteur cardiaque dans un plan déterminé par la position des électrodes (SCHOEMAKER, et al., 2005).
Les sytèmes de dérivation : L’électrocardiogramme est une projection du vecteur cardiaque dans un plan déterminé par la position des électrodes. Différents systèmes de dérivation sont utilisés (DONELEY, 2011): L’ECG standard à 6 dérivations est enregistré à l’aide de 4 électrodes (3 électrodes de dérivation et une dérivation reliée à la terre) qui doivent être attachées directement à la peau de l’animal sur des extrémités appropriées (SCHOEMAKER, et al., 2005). Les dérivations sont attachées aux deux ailes (propatagium) et aux deux pattes (pli du grasset). On forme ainsi : - Le système de dérivation bipolaire standard : DI, DII, DIII - Le système de dérivation augmenté, ou unipolaire : avR, avL, avF Il existe une façon de représenter les 6 champs frontaux (I, II, III, aVL, aVR et aVF) : le système hexa-axial dit de Bailey. Dans ce système, les 6 axes frontaux sont superposés de sorte qu’ils passent tous par le même zéro. Il en résulte un champs circulaire fait de 12 branches provenant d’un point médian commun, chaque ligne étant séparée avec un angle de 30° de sa branche voisine (ZANDVLIET, 2005).

Page | 117
4. Interprétation
Un enregistrement ECG complet devrait inclure un minimum de 4 à 5 beaux complexes, enregistrés sur les 6 dérivations périphériques (I, II, III, aVL, aVR, aVF). Ceci est nécessaire pour mesurer les différents complexes et les intervalles. Un enregistrement plus long (10 à 20 complexes) en dérivation II est nécessaire pour évaluer la fréquence, le rythme et la régularité. Si des tremblements musculaires sont présents et engendrent un tremblement de la ligne de base excessif, l’utilisation d’un filtre peut être envisagé, bien que cela réduise l’amplitude des complexes. Si les complexes ne sont pas de taille acceptable, la sensibilité doit être ajusté (de 5 à 20 mm/mV). Lors de suspicion d’arythmie intermittente, il est conseillé d’enregistrer en dérivation II pendant au moins deux minutes (SCHOEMAKER, et al., 2005). Les artefacts sont le résultat de problèmes techniques ou dépendant de l’animal, qui résultent en un tracé ECG anormal. Les problèmes techniques ont pour origine un placement incorrect des dérivations, des interférences électriques, des câbles défectueux, des électrodes sales ou des réglages inadaptés. Les causes reliées à l’animal comprennent des tremblements musculaires, les mouvements respiratoires et les mouvements volontaires de l’animal. Différencier des artefacts d’anomalie peut être difficile et il n’existe pas de critère définis. Toutefois la plupart des artefacts apparaissent généralement de façon irrégulière et n’interrompent pas le rythme cardiaque sous-jacent. L’ECG doit être interprété en suivant un protocole précis pour n’oublier aucune information importante. On pourra suivre le plan d’analyse suivant :
a. Identification, mesures de l’amplitude et de la durée des déflections et des
segments en dérivation II
Analyse des déflections du patient L’électrocardiogramme obtenu correspond à l’enregistrement de la séquence des dépolarisations et repolarisations cardiaques. Il est ainsi possible d’identifier (Fig. 10) (DONELEY, 2011) : - l’onde P qui reflète la dépolarisation atriale et traduit (du moins en situation normale) la contraction du myocarde des atria (DONELEY, 2011). La partie initiale de l’onde P représente la dépolarisation atriale droite et la deuxième partie représente la dépolarisation de l’atrium gauche (ZANDVLIET, 2005). Cette onde est positive et de morphologie constante en dérivation DII. On mesurera l’amplitude et la durée de l’onde P comme indiquée sur le schéma (NAP, et al., 1992). Une onde P de grande amplitude et étroite sera caractéristique d’une hypertrophie atriale droite (maladie respiratoire chronique, anomalies congénitales) alors qu’une onde P longue et élargie sera associée à une hypertrophie atriale gauche (insuffisance mitrale, communication inter-ventriculaire)(ZANDVLIET, 2005). Enfin une onde P grande et large sera évocatrice d’un élargissement biatrial (MCKENZIE, et al., 1972) (DONELEY, 2011).
Figure 10 - Electrocardiogramme aviaire normal et mesures d’intérêt

Page | 118
- L’onde Ta (T atriale) correspond à la repolarisation atriale. Elle est visible physiologiquement seulement chez certains groupes d’oiseaux comme les pigeons (LUMEIJ, et al., 1994), certains gallinacées (BOULIANNE, et al., 1994) et les perroquets (ZANDVLIET, 2005). C’est une déflection négative en DII. Chez les autres oiseaux, la repolarisation atriale coïncide avec la dépolarisation ventriculaire est n’est donc habituellement pas visible. Sa présence est alors caractéristique d’une hypertrophie atriale droite (ZANDVLIET, 2005). - Le segment P-R reflète le trajet de l’influx entre les oreillettes et les ventricules. Il témoigne notamment du ralentissement du signal lors de son passage dans le nœud atrio-ventriculaire et correspond à la fin du remplissage ventriculaire (DONELEY, 2011). Ce trajet apparaît comme une période isoélectrique (ZANDVLIET, 2005). Un intervalle P-R trop court suggère une augmentation du tonus sympathique, une formation ectopique de l’onde P ou un phénomène de ré-entrée. Un intervalle P-R prolongé est observé lors d’intoxication à la digoxine et lors de blocs atrio-ventriculaires (ZANDVLIET, 2005).
- Le complexe rS correspond à la dépolarisation ventriculaire et est normalement suivi de la contraction des ventricules (ZANDVLIET, 2005). L’onde R est la première déflection positive et l’onde S est la première déflection négative suivant l’onde R, en dérivation II. Par nomenclature, l’onde la plus grande dans le complexe QRS est écrite en lettre majuscule. Chez les oiseaux il est du type rS et est négatif en DII (DONELEY, 2011). Comme la masse ventriculaire gauche est plus importante que celle du droit, l’influx électrique produit par cette partie du cœur est également plus important et l’activité de ce ventricule gauche prédomine sur le tracé ECG (ZANDVLIET, 2005). Deux mesures sont réalisées sur le complexe rS ; 1. La durée qui est mesurée du début de l’onde R à la fin de l’onde S, 2. L’amplitude de l’onde S qui est mesurée à partir de la ligne de base (DONELEY, 2011). L’enregistrement d’une alternance entre des complexe QRS de grande et de faible amplitude (connu sous le nom d’alternance électrique) est une trouvaille fortement suggestive d’un épanchement péricardique. L’ECG de bas voltage est fréquemment observé chez les oiseaux et doit suggérer fortement l’existence d’un épanchement péricardique (insuffisance cardiaque, infections, hypoalbuminémie, épanchement péricardique idiopathique) (ZANDVLIET, 2005). Chez les oiseaux, une onde R proéminente a été associée avec une hypertrophie ventriculaire droite (CZARNEKI, et al., 1980) (HUNSAKER, et al., 1971) et un complexe rS profond ou élargit peut suggérer une hypertrophie ventriculaire gauche (ZANDVLIET, 2005) (DONELEY, 2011). - Le segment S-T peut être très court voire absent chez des perroquets sains (ZANDVLIET, 2005) et des pigeons (LUMEIJ, et al., 1985). Dans ce second cas, l’onde S émerge alors directement dans l’onde T. On parle de « ST-slurring » (DONELEY, 2011). Lorsqu’il est présent, ce segment est normalement isoélectrique. On mesure la durée de ce segment et on portera une attention à son inclinaison. Une dépression en DII peut être expliquée par une hyper- ou une hypokaliémie, une intoxication à la digoxine ou un traumatisme cardiaque (ZANDVLIET, 2005). Contrairement à ce que l’on observe chez les Mammifères, une élévation de ce segment en dérivation II est fréquente chez des oiseaux sains. Elle ne doit pas être interprétée comme un trouble électrique (OGLESBEE, et al., 2001) (LUMEIJ, et al., 1994).

Page | 119
- L’intervalle Q-T peut être allongé de façon physiologique chez les gris d’afrique (Psittacus erithacus) et les amazones (Amazona spp.) (DONELEY, 2011). L’intervalle Q-T est rarement mesuré et est fréquence dépendant. Toutefois, un intervalle Q-T prolongé peut être observé lors d’hypocalcémie, d’hypokaliémie, de défaut de conduction inter-ventriculaire, d’hypertrophie ventriculaire gauche, d’exercice intense, d’hypothermie et de désordre du système nerveux central. Chez les oiseaux, l’anesthésie résulte en un allongement de l’intervalle QT. Un intervalle QT raccourci peut être observé lors d’hypocalcémie, d’hyperkaliémie ou d’intoxication à la digoxine (ZANDVLIET, 2005). - L’onde T correspond à la repolarisation ventriculaire. L’onde T peut être positive, négative ou biphasique mais doit être de morphologie constante. Son amplitude devrait être inférieure à 25% de l’onde S. Les ondes T et P se chevauchent fréquemment et de façon physiologique chez les amazones et certains gris d’afrique. On parle de phénomèneP sur T et cela correspond au fait que les atria se dépolarisent avant que les ventricules ne soient repolarisés (ZANDVLIET, 2005). Les poules semblent davantage prédisposées au phénomène P sur T que les coq et cette différence à été attribuée à l’influence œstrogènique (STURKIE, 1986). On mesure l’amplitude de cette onde. Lorsque l’onde T change de polarité ou augmente progressivement d’amplitude, cela suggère une hypoxie myocardique. Une augmentation de l’amplitude de l’onde T est observée lors d’hyperkaliémie (DONELEY, 2011). Des modifications de l’onde T sont également observées en relation avec un défaut de conduction inter-ventriculaire, des déséquilibres électrolytiques (hypocalcémie, hyperkaliémie), des maladies métaboliques (anémie, choc, urémie, acido-cétose, hypoglycémie et fièvre) et des intoxications médicamenteuses (digoxine et procainamide). (ZANDVLIET, 2005). - l’onde U a été observée chez des cailles et ce, de façon physiologique. L’origine de l’onde U n’est pas bien comprise, y compris en médecine humaine. Les hypothèses les plus probables interprètent cette onde comme l’onde T de repolarisation des cellules de Purkinje, des post-dépolarisations tardives ou la repolarisation tardive des cellules du myocarde-moyen (“M cells”) ou des muscles papillaires. Elle est particulièrement visible lors de ralentissement cardiaque (SZABUNIEWICZ, et al., 1974). Caractéristiques des déflections et des segments de l’ECG normal de l’espèce : Pour évaluer les caractéristiques des déflections et des segments de l’ECG de l’espèce considérée, on aura recours au valeurs fournies par la littérature (cf. tableau). Conclusion de l’analyse : On comparera alors les mesures réalisées sur l’ECG du patient avec les valeurs de références de l’espèce considérée afin de déterminer l’origine du rythme. De façon générale, plus la fréquence cardiaque augmente, plus les paramètres mesurés sont faibles. Nous noterons de plus que les amplitudes mesurées sur l’ECG aviaire sont faibles par rapport à celle observée chez les Mammifères car ils ont un sternum rigide associé à des muscles pectoraux volumineux (MCKENZIE, et al., 1971). L’origine d’un rythme peut être établie en regardant la morphologie des complexes QRS. Lorsqu’un complexe QRS apparaît normal pour l’espèce considérée, il a une origine supra-ventriculaire (atriale ou jonctionelle/nodale). En effet, la dépolarisation ventriculaire suit son cours normal ce qui engendre un complexe QRS de morphologie normale. Ceci contraste avec un complexe QRS d’origine ventriculaire pour lequel la dépolarisation initiale provient du myocarde ventriculaire, se propage lentement par une conduction de cellules musculaire à cellule musculaire et suit un trajet de dépolarisation ventriculaire anormal. Cela engendre un complexe QRS d’apparence large et bizarre, d’amplitude plus grande que la normale et une onde T qui est directement opposé au complexe QRS (ZANDVLIET, 2005). L’analyse conclura alors à l’origine du rythme : supra-ventriculaire ou ventriculaire qui est un élément indispensable à l’évaluation de l’arythmie.

Page | 120
b. Détermination de la fréquence (ventriculaire) cardiaque
Fréquence cardiaque du patient : Si le rythme est régulier, on peut multiplier la vitesse de défilement (mm/s) par 60 et diviser le résultat par la distance en millimètre entre deux ondes R, ce qui donne la fréquence en battements par minute (SCHOEMAKER, et al., 2005). Si le rythme est irrégulier, cette première méthode n’est pas valide. Il faut alors compter le nombre de complexes QRS qui apparaissent en 3 secondes et multiplier ce nombre par 20 pour obtenir le nombre de battement par minute (DONELEY, 2011). Fréquence cardiaque normale de l’espèce : Pour évaluer la fréquence normale d’un oiseau, on pourra utiliser les valeurs de références décrites dans la littérature (cf. annexe II) ou bien utiliser la formule suivante si aucune étude n’a été réalisée sur l’espèce considérée : Fréquence cardiaque (min-1) = 241 x Mc -0.25 où Mc est la masse corporelle en kg. Une fréquence cardiaque 20% plus élevée ou plus basse que la fréquence calculée sera évocatrice respectivement d’une tachycardie ou d’une bradycardie (LICHTENBERGER, et al., 2007). Conclusion de l’analyse : On se posera alors la question de savoir si la fréquence cardiaque du patient est normale ou anormale pour son espèce. Le caractère normocarde, tachycarde ou bradycarde de l’arythmie constitue la première étape de l’interprétation (cf. tableau des arythmies) (DONELEY, 2011)
c. Détermination du rythme cardiaque
Analyse du rythme du patient : L’analyse du rythme doit répondre aux questions suivantes : 1. Le rythme est-il régulier ou irrégulier ? Une variation d’un intervalle R-R à l’autre de moins de 10% doit amener à considérer le rythme comme étant régulier (ZANDVLIET, 2005). 2. Y a-t-il des ondes P et sont-elles ressemblantes entre elles ? On identifiera les ondes P et on évaluera leur uniformité ainsi que leur régularité. Lorsque les ondes P ne sont pas facilement identifiables, il peut être utile de doubler la sensibilité de l’électrocardiographe (ZANDVLIET, 2005). 3. Y a-t-il des complexes QRS et sont-ils ressemblants entre eux ? On identifiera les complexes QRS et on évaluera leur uniformité ainsi que leur régularité (ZANDVLIET, 2005). 4. Y a-t-il une relation entre les ondes P et les complexes QRS ? On évaluera pour répondre à cette question si chaque onde P est suivie par l’apparition d’un complexe QRS dans un délai normal et si chaque complexe QRS succède à une onde P (mesure du ratio P : QRS) (DONELEY, 2011). Rythme normal aviaire : Le rythme sinusal est le rythme le plus fréquemment rencontré dans la plupart des espèces pour une fréquence cardiaque normale. Il est caractérisé par des ondes P-QRS-T régulières, avec une onde P positive dans les dérivations II, III, aVF et un intervalle PR relativement constant (ZANDVLIET, 2005). Conclusion de l’analyse : Si le rythme ne correspond pas à un rythme considéré physiologique chez les oiseaux, l’analyse devra alors préciser le rythme : Régulier, régulièrement irrégulier ou irrégulier. L’analyse devra distinguer si le trouble correspond à un battement prématuré ou un rythme d’échappement. Un rythme d’échappement ou un battement d’échappement ont tous les deux une origine ectopique et apparaissent à des fréquences cardiaques faibles pour l’espèce considérée, et ce afin d’empêcher une asystolie. Un battement prématuré (ou extrasystole) est un battement cardiaque supplémentaire se surajoutant à une fréquence cardiaque normale ou haute. Les rythmes prématurés apparaissent plus tôt que prévu, ce qui signifie que l’intervalle R-R est plus court que l’intervalle R-R du cycle précédant et que cela engendre un rythme irrégulier. Les battements prématurés sont soit d’origine supraventriculaire (atriale ou jonctionnelle) ou ventriculaire, la différence étant basée sur la morphologie des complexes QRS (cf. supra). La différence entre un battement prématuré supra-ventriculaire d’origine atriale ou jonctionnelle est difficile à établir et n’a pas d’intérêt clinique majeur (ZANDVLIET, 2005).

Page | 121
d. Détermination de l’axe électrique moyen ventriculaire
Détermination de l’axe électrique moyen ventriculaire du patient : L’axe électrique moyen (AEM) est mesuré dans le plan frontal et donne la direction ainsi que l’activité électrique moyenne au cours du cycle cardiaque. Il est possible de déterminer cet axe soit par construction soit par estimation en utilisant le système de dérivation hexa-axial de Bailey (Fig.11) (ZANDVLIET, 2005).
- Par estimation : Pour estimer rapidement l’axe électrique moyen du complexe QRS, il faut trouver une dérivation dans laquelle le complexe QRS est isoélectrique (c'est-à-dire la dérivation pour laquelle l’amplitude des QRS est nulle). On utilise ensuite le diagramme de Bailey qui représente le système de référence à 6 axes (DI, DII, DII, avR, avL, avF) et l’on trouve quelle dérivation est perpendiculaire à la dérivation isoélectrique. Par exemple, si la dérivation I est isoélectrique, alors l’axe électrique moyen doit s’étendre le long de l’axe aVF. Il faut enfin déterminer si la dérivation perpendiculaire est positive ou négative sur le tracé et examiner la valeur de l’angle sur le système de référence à 6 axes (DONELEY, 2011). En absence de dérivation isoélectrique, il est possible d’estimer l’AEM en sélectionnant la dérivation qui a le complexe QRS le plus grand. Si la déflection est positive, l’AEM est dirigé vers le pôle positif de l’axe de la dérivation et vers le pole négatif en cas de déflection négative. Si deux dérivations ont des déflections égales, l’axe électrique se situe à mi-chemin entre ces deux dérivations (ZANDVLIET, 2005).
Figure 11 - Construction du système hexaxial de Bailey par combinaison des dérivation triaxiales
bipolaires et unipolaires augmentées
- Par construction : C’est la méthode la plus précise pour déterminer l’axe électrique moyen. Cette méthode nécessite de mesurer la hauteur des déflections dans deux dérivations (en général DI et DIII) puis de reporter ces valeurs dans le système triaxial, sur les axes des dérivations correspondantes, le zéro correspondant à l’intersection des trois dérivations. Des lignes perpendiculaires sont tracées à partir de ces deux points jusqu’à ce qu’elles se coupent. Une ligne est alors tracée entre le zéro et ce point d’intersection. L’axe électrique moyen est l’angle entre l’axe de la dérivation I et cette ligne dernièrement tracée (ZANDVLIET, 2005).

Page | 122
Axe électrique moyen ventriculaire normal de l’espèce : Pour évaluer l’angulation de l’axe électrique moyen de l’espèce considérée, on pourra utiliser les valeurs de références décrites dans la littérature (cf. tableau). Si aucune valeur n’existe dans la littérature, on estimera que l’axe électrique moyen est situé entre 83° et -162° chez les oiseaux (DONELEY, 2011). L’axe électrique moyen lors de la dépolarisation ventriculaire est négatif chez les oiseaux pour deux raisons : d’une part car la vague de dépolarisation démarre en position sous-épicardique et diffuse ensuite au travers du myocarde vers l’endocarde (DONELEY, 2011) et d’autre part car l'anneau atrioventriculaire de Purkinje entraîne une orientation de l'axe électrique moyen du complexe QRS à près de -90° (c'est à dire cranialement le long du grand axe du corps plutôt que caudalement comme c'est typiquement le cas chez les Mammifères) (TULLY, et al., 2009). Conclusion de l’analyse : On comparera ensuite l’axe électrique moyen du patient avec la valeur attendue. On pourra alors conclure à une absence de déviation, une déviation vers la droite ou une déviation vers la gauche. L’axe électrique moyen peut subir une déviation vers la droite lors d’hypertrophie ventriculaire droite marquée (sténose pulmonaire, tétralogie de Fallot, maladie des vers du cœur, insuffisance valvulaire tricuspidienne, cœur pulmonaire aigu, maladie pulmonaire diffuse chronique, embolisme pulmonaire) et lors d’anomalie de conduction (bloc de branche droite) (ZANDVLIET, 2005). L’axe électrique moyen peut subir une déviation vers la gauche lors d’hypertrophie ventriculaire gauche marquée (persistance du canal artériel, défaut du septum ventriculaire et sténose aortique, cardiomyopathie dilatée) et lors d’anomalie de conduction (bloc de branche gauche) (ZANDVLIET, 2005). Chez les oiseaux, l’élargissement biventriculaire est difficile à diagnostiquer mais est suggéré par la combinaison d’un élargissement ventriculaire gauche (Q profond ou ondes R de grande amplitude) et une déviation de l’axe vers la droite (ZANDVLIET, 2005).
e. Identification des causes de l’arythmie et démarche diagnostic
Les conclusions établies précédemment permettent de caractériser les anomalies de fréquence, de régularité, d’amplitude et d’intervalles du patient. Il faut garder à l’esprit qu’il existe des différences entre les espèces, les races et parfois même les sexes, ce qui explique les petites différences de valeurs entre les différentes études publiées. Encore une fois, le clinicien doit prendre garde de ne pas surinterpréter des anomalies mineures (SCHOEMAKER, et al., 2005). La deuxième étape consiste à associer ces anomalies avec l’existence d’une maladie cardiaque (Cf. Annexe III). Il faut garder en tête que les arythmies peuvent être d’origine cardiaque, extra-cardiaque (anomalies métaboliques, modifications du tonus autonome). Il est donc important d’éliminer les anomalies métaboliques en mesurant les concentrations en sodium, potassium, calcium, l’hématocrite (hypoxie) et si possible le statut acido-basique voire le glucose. En absence d’hypothermie, de fièvre, d’anomalie encéphalique, de maladie respiratoire ou gastrointestinale, la cause de l’arrythmie est considérée être d’origine cardiaque. Il faut également prendre en compote les effets cardioactifs de certaines molécules parmi lesquelles, la digoxine, les beta bloquants, les diurétiques, les vasodilatateurs et divers agents anesthésiques (ZANDVLIET, 2005). Conclusion : L’ECG est donc une technique facilement réalisable et pouvant apporter des informations inestimables sur les troubles du rythme (anomalie de formation ou de conduction du signal électrique) ou sur la structure du cœur (hypertrophie cardiaque, distension des cavités). C’est également une technique permettant une mesure précise de la fréquence cardiaque chez des espèces pour lesquelles cette dernière peut être élevée.

Page | 123
E. Biochimie sérique
Indication : L’apport diagnostic de la biochimie clinique en médecine aviaire répond à trois objectifs :
- Objectiver l’existence d’une lésion cardiaque (diagnostic lésionnel) par le dosage de marqueurs de la cytolyse cardiaque. - Objectiver les répercussions fonctionnelles de l’atteinte cardiaque (pronostic) par le dosage de marqueurs de l’insuffisance cardiaque et un bilan biochimique classique. - Investiguer une cause sous-jacente à l’apparition de la maladie (diagnostic étiologique) par la réalisation d’un dosage des paramètres biochimiques habituels et d’une électrophorèse des protéines.
Technique : Lors de la mesure du potassium, il est important d’avoir à l’esprit que le stockage du sang après le prélèvement a une grande influence sur la concentration de cet électrolyte en particulier. Un stockage au froid diminuera la concentration par plus de 10% alors qu’un stockage à température ambiante augmentera la concentration jusqu’à 20%. Ces modifications ne sont pas visibles lorsque le plasma et les cellules sont séparés immédiatement après le prélèvement (DE WIT, et al., 2005). Interprétation : Les marqueurs de la cytolyse cardiaque : Nous classerons ces enzymes par ordre de spécificité croissant. - L’aspartate amino-transférase (AST, SGOT) : Les valeurs plasmatiques normales en AST devraient, pour la plupart des oiseaux, se trouver dans l’intervalle 52 à 270 UI/L. Chez l’autruche les valeurs normales vont de 100 à 160 UI/L. Cette enzyme n’est pas spécifique d’organe. Après le foie, ces enzymes sont retrouvées dans le muscle cardiaque, le muscle squelettique, les cellules gastro-intestinales, le rein et le cerveau. Comme c’est le cas pour la biochimie d’un grand nombre de constituants sanguins, les niveaux varient avec l’âge et l’activité saisonnière. Une augmentation de la concentration sérique en AST indiquera en priorité une atteinte hépatique. Néanmoins, toute lésion musculaire et tout dommage infligé aux tissus mous, incluant les injections intra-musculaires peuvent provoquer une augmentation des AST plasmatiques (COLES, et al., 2007). Quand une élevation des AST est observée, il convient donc de réaliser un dosage biochimique de la créatinine kinase (CK). Une élévation des AST sans augmentation simultanée de CK est hautement suggestif d'une perturbation hépato-cellulaire (TULLY, et al., 2009).

Page | 124
- Lactate déshydrogénase (LDH) : Chez la plupart des oiseaux normaux, les taux sériques de LDH sont entre 46-442 UI/L. Chez les autruches, ils peuvent être de 1000-2000 UI/L et chez les canaris, ils sont d’approximativement 1582 UI/L (COLES, et al., 2007). Bien que cette enzyme ne soit pas spécifique d’organe (elle est trouvée dans les muscles squelettiques et cardiaque, le foie, les reins, les os et les érythrocytes (TULLY, et al., 2009)), une augmentation de la concentration sérique est communément observée lors d’hépatopathie et de myopathie. Toutefois, une augmentation peut également être observée en cas d'hémolyse ce qui la rend extrêmement peu spécifique (TULLY, et al., 2009). Comme pour les AST, la valeur de la LDH doit toujours être analysée de paire avec celle de la CPK pour juger de son origine musculaire ou autre (COLES, et al., 2007). - Créatinine kinase (CK, CPK) : Les valeurs normales des CPK chez les oiseaux varient entre 110-480 UI/L. Chez l’autruche, elles peuvent être plus élevées, entre 400-900 UI/L. Bien que cette enzyme ne soit pas spécifique d’organe (on en retrouve dans le muscle squelettique, le muscle cardiaque et le tissu nerveux (TULLY, et al., 2009)), une augmentation de sa concentration sérique est en pratique spécifique d’un trauma musculaire. Elle est en ce sens un guide très utile pour différencier des dommages musculaires de dommages hépatiques lorsque les niveaux d’AST et de LDH sont augmentés. Si les CK sont normales, il est relativement sûr de conclure que le foie est la source de l'élévation des AST. Si les CK sont élevées, le muscle cardiaque (HOCHLEITHNER, 1994) ou les muscles squelettiques doivent être considérés comme une source possible de l'élévation des AST (TULLY, et al., 2009). Il faut toutefois noter que des injections intramusculaires réalisés avant la réalisation du prélèvement sanguin peuvent engendrer une augmentation de la valeur de la CPK. De plus, le taux de CPK est supposé augmenter lors de neuropathies associées à des convulsions, d’intoxication au plomb, de chlamydiose, de septicémie bactérienne et de carence en vitamine E (COLES, et al., 2007). Il a été montré que les niveaux de l’iso-enzyme cardiaque de la créatinine kinase augmentaient secondairement à la toxicité cardiaque du furazolidone chez les canards de Pékin (Anas platyrhynchos) (WEBB, et al., 1991). - Les troponines : Les troponines T et I sont deux protéines constitutives des filaments fin régulant la contraction des cellules musculaires striées et ont des isoenzymes spécifiques du cœur (cTnI et cTnT). Les études révèlent que les dosages immunochimiques de la cTnI et de la cTnT peuvent être utilisés de façon sensible et spécifique pour détecter une lésion cardiaque chez la dinde et la poule. Toutefois l’évaluation des cTnT sérique est limité à un processus lésionnel cardiaque aigü ou en cours. L’interprétation doit également être précautionneuse en cas d’atteinte musculaire squelettique sévère car ces muscles contiennent, bien qu’en quantité très minime de la cTnT. Le résultat des CK devra alors être pris en considération. La cTnI est retrouvée en quantité 230 fois plus grande dans le cœur que dans les muscles squelettiques (contre 1800 chez le chien). Ainsi, la cTnI est un biomarqueur spécifique de lésions cardiaques chez les oiseaux et le dosage des cTnI est 5 à 10 fois plus spécifique que celui de la cTnT (APPLE, 1999) (BOGIN, et al., 1976). Les marqueurs de l’insuffisance cardiaque : Contrairement à ce qui est réalisé chez l’homme et les carnivores domestiques, la faisabilité et l’intérêt du dosage du NT-pro BNP chez les oiseaux n’ont pas été évalué à ce jour. Toutefois, compte tenu de la similarité des facteurs déclenchant la libération de cette hormone chez les oiseaux et les mammifères il est probable qu’une insuffisance cardiaque, accompagnée d’une surcharge se manifeste par une augmentation de la concentration sérique en NT-pro BNP.

Page | 125
Paramètres d’intérêt d’un bilan biochimique classique : Les enzymes hépatiques peuvent être augmentées suite à une congestion hépatique elle-même secondaire à une insuffisance cardiaque droite ou à un dommage hépato-cellulaire hypoxique associé à une insuffisance cardiaque (WEBB, et al., 1991), (O’BRIEN, et al., 1993). Il est fréquent d’observer une augmentation des acides biliaires lors de congestion hépatique. Chez les dindes atteintes de cardiomyopathie induite par la furazolidone, l’hypoprotéinémie caractérisée par une hypoalbuminémie et une hypoglobulinémie est expliquée par une expansion du volume sanguin et secondairement par une inanition, voire possiblement des pertes transsudatives dans les cavités corporelles (O’BRIEN, et al., 1993). L’hyperuricémie peut conduire à l’apparition d’une goutte péricardique. Les désordres éléctrolytiques (hypokaliémie et hypocalcémie notamment) sont des causes communes de bradycardie et d’arythmie. Les perroquets gris d’afrique sont particulièrement sensibles à l’hypocalcémie, laquelle peut être exacerbée par une anesthésie à l’isoflurane (GOODMAN, 1996). Le dosage de la Vitamine E et du Sélénium pourra être intéressant si une carence est suspectée. Des modifications de l’électrophorèse protéiques peuvent indiquer une cause infectieuse. Le cholestérol plasmatique peut orienter vers un diagnostic d’athérosclérose (DE WIT, et al., 2005).
F. Hématologie
Indication : La réalisation d’un examen hématologique est motivé par la recherche d’une origine et des répercussions fonctionnelles de la maladie cardiaque (DE WIT, et al., 2005). Technique : La ponction de 0,3 ml de sang à l’aide d’une aiguille 22-25 G seront suffisants. Le prélèvement sera stocké dans un tube EDTA. Les automates n’étant pas adaptés au comptage des cellules aviaires, il faudra réaliser un comptage manuel (CAPITELLI, et al., 2013). Interprétation : Une numération-formule peut révéler des modifications dans le comptage des cellules rouge ou blanche, lesquelles peuvent appuyer un diagnostic de maladie cardiaque (STRUNK, 2003). La polycythémie est une trouvaille commune lors d’hypoxie chronique. C’est le cas chez les dindes et les poulets en croissance qui développent une insuffisance cardiaque chronique (RIDDELL, 1997). Un hématocrite supérieur à 55% a été décrit chez un amazone âgé présentant une insuffisance cardiaque droite associée à une fibrose pulmonaire interstitielle chronique (DE WIT, et al., 2005). Une leucocytose appuiera l’hypothèse infectieuse de la cardiopathie (endocardite, myocardite)(ISAZA, et al., 1992) (CAMPBELL, 1994). Des infections bactériennes très intenses ou virales pourront à l’inverse engendrer une leucopénie (CAMPBELL, 1994).
G. Recherche d’agent infectieux
Indication : La réalisation de tests spécifiques d’agent infectieux permet de rechercher la cause de la maladie cardiaque. On recherchera les organismes classiquement impliqués dans l’apparition d’infections cardiaques (DE WIT, et al., 2005). Technique : La culture sanguine est particulièrement indiquée si une endocardite bactérienne est suspectée (apparition subite d’un souffle cardiaque) (STRUNK, 2003)(DE WIT, et al., 2005). Une diversité de test (polymerase chain reaction, sérologie, neutralisation virale, etc.) peut être utilisée pour appuyer un diagnostic de maladie cardiaque due à des processus infectieux spécifiques. En plus d’un frottis sanguin direct et de test de filtration sanguine, un test commercial ELISA contenant des antigène de ver du cœur canin a été utilisé pour soutenir le diagnostic de microfilaires (Paronchocerca ciconarum) chez deux Jabirus d’Afrique (Ephippiorhynchus senegalensis) (ECHOLS, et al., 2000). La spécificité et la sensibilité de ces tests pour les filaires aviaires sont inconnus, mais les données de ce cas suggèrent que des microfilaires aviaires et mammaliennes pourraient avoir conservé des antigènes communs (ECHOLS, et al., 2000) (STRUNK, 2003). Interprétation : La mise en évidence d’un agent infectieux associé à des modifications du leucogramme conforteront l’origine microbienne de l’affection cardiovasculaire.

Page | 126
H. Examen nécropsique post-mortem
Indication : Toute suspicion d’une affection cardio-vasculaire pourra justifier la réalisation d’un examen nécropsique (MAJO, et al., 2012). Technique : Après avoir ouvert la cavité thoraco-abdominale, l'aspect extérieur du cœur ainsi que le péricarde seront évalués (COLES, et al., 2007). En cas d’épanchement péricardique, celui-ci sera ponctionné et fera l’objet d’un examen cytologique et bactériologique. Avant de retirer le cœur de la cavité thoracique, le sang qu’il contient peut être prélevé à l’aide d’une seringue et d’une aiguille stériles pour la réalisation d’un examen bactériologique et d’un frottis. Le frottis sera coloré avec une coloration de Wright en vue de la recherche de parasites (hémoprotozoaire, microfilaire) ou coloré par une coloration de Gram en vue de la recherche de bactéries (RAE, 2003). Le cœur est extrait de la cavité cœlomique avec l’ensemble représenté par l’œsophage, le jabot, la trachée et les poumons. Le cœur est ensuite isolé. Le péricarde est incisé puis la quantité et l’aspect du liquide péricardique sont évalués. Le cœur est ensuite incisé en coupe transversale afin d’évaluer la taille de la lumière ventriculaire ainsi que l’épaisseur du myocarde. On procédera à l’ouverture du cœur dans la direction du flux sanguin, tout en utilisant de l’eau pour rincer le sang et les caillots. La surface endocardique, les rapports entre les cavités et le myocarde seront alors évalués. On procédera ensuite à l’ouverture des gros vaisseaux. Il est conseillé de placer la plus grande partie du cœur dans le formalin, de sorte à pouvoir réaliser plusieurs sections en vue d’une évaluation histologique (RAE, 2003). Interprétation : Lésions endocardiques : On recherchera l’existence d’ulcérations endothéliales éventuellement recouvertes de thrombi. De telles ulcérations sont compatibles avec une endocardite valvulaire thrombotique, une sténose valvulaire consécutive à une inflammation chronique, une septicémie ou une maladie métabolique. Il sera également possible d’observer des nodules fibreux localisés sur les valves évoquant une endocardiose valvulaire (KRAUTWALD-JUNGHANNS, et al., 2004). Lésions péricardiques : On évaluera tout d’abord l’opacité du péricarde. Une opacification due à l’existence d’exsudat fibrineux ou fibrino-purulent pourra être observé lors de péricardite suppurée (KRAUTWALD-JUNGHANNS, et al., 2004), fréquemment rencontrée lors d’une septicémie généralisée à E. coli, (MAJO, et al., 2012), Pasteurella spp. ou Chlamydia psittaci. La présence d’un dépôt granuleux, mucoïde et blanchâtre évoquera une précipitation d’urate (goutte viscérale). La confirmation de la présence de cristaux d’acide urique peut être réalisée en observant un frottis sous lumière polarisée. La fixation dans le formalin peut dissoudre les critaux d’acide urique qui peuvent ne pas être visualisés lors de l’examen histopathologique. En revanche, ils ne sont pas dissous dans l’éthanol (RAE, 2003). On s’attachera également à évaluer la quantité de liquide dans le sac péricardique, celle-ci étant normalement imperceptible (COLES, et al., 2007). L’existence d’un épanchement péricardique séreux (hydropéricarde) s’observe le plus souvent lors d’insuffisance cardiaque mais peut aussi être observé lors d’hépatite à corps d’inclusion provoquée par un adénovirus (RAE, 2003) (BISWAS, et al., 2002), une insuffisance hépatique (intoxication à l’avocat chez le canari) (HARGIS, et al., 1989) ou une polyomavirose (mue française) (RAE, 2003) (BERNIER, et al., 1981) (DAVIS, et al., 1981)). Un épanchement hémorragique pourra être remarqué lors de rupture myocardique. Des pétéchies ou des hémorragies épicardiques ecchymotiques sont fréquemment observées en cas de mort subite (RAE, 2003).

Page | 127
Lésions myocardiques : La forme globale du cœur est évaluée en premier lieu. Le cœur normal d'un oiseau est de forme grossièrement triangulaire et toute évolution vers une forme sphérique est anormale (COLES, et al., 2007). On évaluera ensuite la taille des cavités ainsi que l’épaisseur du myocarde. Les cavités ventriculaires sont normalement pratiquement virtuelles chez les oiseaux (MAJO, et al., 2012) et des études ont été publiées quant à la morphométrie du cœur aviaire. Les lésions macroscopiques les plus fréquemment rencontrées sont une augmentation de la taille du cœur associée à une dilatation et une hypertrophie du ventricule droit (MAJO, et al., 2012) (KRAUTWALD-JUNGHANNS, et al., 2004). Lors de l’examen de l’épaisseur pariétal, il faut garder à l’esprit qu’une disparité d’épaisseur entre les deux ventricules peut être due à l’angle de transsection (RANDALL, et al., 1996). On s’attachera ensuite à évaluer l’aspect du myocarde : l’existence de masse ou de nodules blanchâtre pourront évoquer une tumeur myocardique (lymphome secondaire à une maladie de Marek (MAJO, et al., 2012)), une infiltration graisseuse pourra évoquer une lipomatosis cordis, des foyers ou des bandes pâles indiqueront quant à eux une myopathie dégénérative en relation avec une carence en vitamine E/Selenium ou une myocardite en association avec une septicémie, une maladie virale (RAE, 2003). Occasionnellement, il est possible d'observer une rupture de l'atrium droit consécutive à une dilatation massive au cours d'une insuffisance circulatoire produite par une maladie grave (COLES, et al., 2007). Lésions vasculaires : On recherchera une décoloration jaunâtre, une induration éventuellement secondaire à une calcification ou une métaplasie chondroïde (KRAUTWALD-JUNGHANNS, et al., 2004) ainsi qu’une obstruction luminale pouvant évoquer un processus athérosclérotique (COLES, et al., 2007). Les calcifications sont également compatibles avec une maladie rénale ou bien une hypervitaminose D (RAE, 2003). Sporadiquement, il est possible d’observer un dépôt d’urate dans l’aorte (KRAUTWALD-JUNGHANNS, et al., 2004). Des hémorragies pétéchiales pourront être un signe de septicémie ou de agonie violente précédant la mort (COLES, et al., 2007). Les jeunes dindes souffrent communément d'anévrismes disséquant des artères (COLES, et al., 2007), lesquels peuvent se rompre (MAJO, et al., 2012). Lésions associées à une affection cardiovasculaire : Lors de l’examen post-mortem, les fluides et la fibrine sont retrouvés primitivement dans les deux espaces hépato-péritonéaux ventraux (JULIAN, 1993). Lors d’ascite, il est important de bien rechercher la cause sous jacente cardiaque ou hépatique de l’augmentation de la pression portale (JULIAN, 2005). CONCLUSION : Pour diverses raisons d’ordre physiologique ou anatomique, l’examen physique reste limité chez les oiseaux, à comparer de celui réalisable chez les chiens ou les chats (STRAUB, et al., 2002). De plus, la symptomatologie des maladies cardiaques est typiquement non spécifique. Leur diagnostic est rendu encore plus difficile par le fait que ces maladies sont fréquemment associées à d’autres maladies internes chez les perroquets ce qui fausse le tableau clinique (PEES, et al., 2001). Si l’examen clinique apporte des éléments amenant à inclure une maladie cardio-vasculaire dans le diagnostic différentiel il sera indiqué de réaliser des procédures diagnostiques plus poussées (PEES, et al., 2006).

Page | 128

Page | 129
QUATRIEME PARTIE : THERAPEUTIQUE CARDIOVASCULAIRE

Page | 130
I. Les grandes familles de médicaments Il existe de nombreuses molécules modifiant le fonctionnement du cœur et des vaisseaux. Ces agents thérapeutiques agissent sur des récepteurs (adrénergiques α, β), des canaux (calciques voltage dépendant), des systèmes de transport ionique (ATPase Na+/K+) ou des enzymes (phosphodiestérase, enzyme de conversion de l’angiotensine). Leurs effets sur l’appareil cardiovasculaire dépendent à la fois de leur(s) cible(s) moléculaire(s) et de leur(s) organe(s) cible(s) (cœur et/ou vaisseaux). Ainsi, au sein d’une même classe thérapeutique, certains agents auront des effets vasculaires lorsque d’autres auront des effets cardiaques. Nous classerons donc ces molécules selon leur effet clinique.
A. Les modificateurs de l’inotropisme
1. Les inotropes positifs
a. Les digitaliques : la digoxine
Pharmacocinétique : Le temps de demi-vie de la digoxine est d’environ 24 heures chez les dindes, les rapaces et les psittacidés (21 à 26 heures selon l’espèce et la posologie choisie). Ceci justifie une administration q24h pour maintenir un niveau plasmatique considéré stable (HAMLIN, et al., 1987) (WILSON, et al., 1989). Mode d’action : La digoxine agit en bloquant la pompe ATPase Na+/K+. En conséquence, le fonctionnement du transporteur Na+/Ca2+ s’inverse et provoque une accumulation de Ca2+ dans la cellule. Ainsi, au niveau myocardique, la digoxine améliore la contraction myocardique (inotropisme positif) ce qui facilite la résorbtion des oedèmes et de l’ascite (JONES, 2008). Le blocage de l’ATPase Na+/K+ des neurones des systèmes nerveux central et périphérique provoque une augmentation du tonus parasympathique sur les nœuds sinusal et atrio-ventriculaire. En conséquence, on observe un ralentissement de la fréquence sinusale (chronotropisme négatif) et une diminution de la vitesse de conduction (dromotropisme négatif). Ceci lui confère des effets anti-arrythmiques (ROSENTHAL, et al., 1997).
Indication : L’utilisation de la digoxine est indiquée lors de diminution de la contractilité (cardiomyopathie dilatée (PEES, et al., 2006)), de surcharge volumique chronique (insuffisance chronique valvulaire) (DONELEY, 2011), de tachycardie supraventriculaire (fibrillation ou un flutter atrial)(PEES, et al., 2006). Certains auteurs conseillent d’utiliser cette classe thérapeutique pour stabiliser un oiseau en urgence lors d’insuffisance cardiaque terminale (BEEHLER, et al., 1980) plutôt qu’en vue d’une utilisation à long-terme (JONES, 2008). Contre-indication : Les digitaliques sont contre-indiqués lors de cardiomyopathie hypertrophique, de tachycardie ventriculaire, de maladie sinusale, de bloc atrio-ventriculaire (DONELEY, 2011), d’hypercalcémie, d’hypokaliémie (JONES, 2008) et de sténose valvulaire (PEES, et al., 2006). La digoxine ne doit pas non plus être utilisée dans le traitement de l’épanchement péricardique. En effet, l’accumulation de fluide au sein de la cavité péricardique provoque une insuffisance cardiaque diastolique qui correspond à un remplissage insuffisant des ventricules en diastole, secondaire à la compression des atria. Les glycosides cardiaques sont donc absolument contre-indiqués car il n’y a pas d’intérêt à augmenter la contractilité myocardique laquelle conduit à une augmentation de la consommation en oxygène pour un volume de sang ventriculaire diminué (STRAUB, et al., 2003).

Page | 131
Interactions médicamenteuses : L’utilisation de la digoxine en association avec d’autres molécules (tels que l’itraconazole, fréquemment utilisée car les psittaciformes atteints de troubles cardiaques sont fréquemments atteints d’aspergillose) peut aboutir à une toxicité de la digoxine. En effet, bien que leur mode d’élimination soient différents, les deux sont fixés aux protéines plasmatiques (99% pour l’itraconazole et 20% pour la digoxine) (STRAUB, et al., 2003). Si une toxicité est suspectée, le traitement doit être arrêté 1 à 2 jours puis ré-instauré à la moitié du dosage initial (DE WIT, et al., 2005).
Effet secondaire : Les marges thérapeutiques sont très faibles et les effets secondaires apparaissent à partir d’un taux sérique de 2.4 ng/ml (ROSENTHAL, et al., 1993). Les effets secondaires correspondent à une toxicité myocardique (DONELEY, 2011) se manifestant par de la léthargie, une anorexie, des vomissements, de la diarrhée, de l’ascite, des oedèmes (PEES, et al., 2009) (JONES, 2008), un bloc de branche du second degré (ROSENTHAL, et al., 1993) et un intervalle P-R prolongé (DONELEY, 2011). Posologie recommandée : Plusieurs protocoles d’utilisation de la digoxine ont été proposés (ROSENTHAL, et al., 1993) (WILSON, et al., 1989) (HAMLIN, et al., 1987) (OGLESBEE, et al., 2001). Les doses initiales suggérées sont de 0.02 mg/kg (ROSENTHAL, et al., 1993) à 0.05 mg/kg (WILSON, et al., 1989) PO q12h les 2-3 premiers jours, puis de diminuer à 0.01 mg/kg q12-24 ensuite (PEES, et al., 2009) (JONES, 2008). (OGLESBEE, et al., 2001) (ROSENTHAL, et al., 1993) (WILSON, et al., 1989). Toutefois, il a été prouvé qu’à cette posologie pouvait s’exercer des effets toxique chez le mainate (ROSENTHAL, et al., 1993). Ainsi, certains auteurs recommandent une dose initiale de 0.01 mg/kg, suivi par un dosage de la digoxinémie 4 à 6 semaines plus tard justifiant une augmentation de la posologie de 0.005 mg/kg si la concentration plasmatique en digoxine est inférieur à l’intervalle considéré thérapeutique chez les oiseaux (OGLESBEE, et al., 2001), soit 0.8-2.4 ng/ml 8-10 heures après la dernière administration. Il est important d’insister sur le fait que cet intervalle provient de la littérature mammalienne et que sa pertinence n’a pas été évaluée chez les oiseaux (ROSENTHAL, et al., 1993). Les concentrations sériques doivent être mesurées après une semaine de thérapie ou une semaine après une modification de la posologie (DONELEY, 2011).
b. Les sensibilisateurs du calcium et inhibiteur des phosphodiestérases : Le
pimobendane
Pharmacocinétique : La pharmacocinétique du pimobendane n’a pas été étudiée chez les oiseaux (BEAUFRERE, et al., 2009). Des études sont en cours (GUZMAN, et al., 2012). Mode d’action : Le pimobendane est un derivé benzimidazole-pyridazone. Il appartient à la classe médicamenteuse des sensibilisateurs calciques car il augmente l’affinité des sites de liaison calcique de la troponine C au calcium. Ainsi, en présence de pimobendane, les myofibrilles se contractent pour une concentration calcique plus faible. Contrairement aux glycosides cardiaques et à la plupart des inotropes, le pimobendane n’entraine donc pas d’augmentation de la consommation en oxygène du myocarde. Cette molécule exerce également une action inhibitrice sur la phosphodiestérase III cardiaque. En conséquence, l’AMPc n’est plus détruite et s’accumule dans la cellule. Ceci augmente l’inotropisme, mais de façon mineure. Toutefois, elle tend à augmenter légèrement la fréquence cardiaque et la vitesse de conduction cardiaque. De plus, l’AMPc favorise la capture diastolique du calcium et donc la relaxation myocardique. Enfin, le pimobendane inhibe également les phosphodiestérases III et V vasculaires ce qui engendre une vasodilatation. En conclusion, le pimobendane est un inotrope positif, un lusitrope positif, et un vasodilatateur périphérique. On le qualifie d’inodilatateur (BEAUFRERE, et al., 2009) (MADDISON, et al., 2008).

Page | 132
Indication et contre-indications : L’effet du pimobendane est similaire à celui de la digoxine mais comporte moins de d’effet indésirable. Il a les mêmes indications et contre-indications (JONES, 2008). Posologie recommandée : Le pimobendane a été utilisé occasionnellement chez des psittacidés et a occasionnellement permis une amélioration des symptômes cliniques. Des études pharmacocinétique ont conclu à une posologie de 10 mg/kg PO q12h chez les amazones (GUZMAN, et al., 2012). Toutefois la durée de survie après le diagnostic d’insuffisance cardiaque congestif a toujours été courte (SEDACCA, et al., 2009) (PHALEN, et al., 1996) (PEES, et al., 2001) (PEES, et al., 2006). Les doses empiriques conseillées vont de 0.15mg/kg PO q12h (DONELEY, 2011) à 0.25-0.5 mg/kg q24h (PEES, et al., 2009).
c. Les agonistes adrénergique β-mimétiques
- La dopamine La dopamine est une catécholamine endogène dont la demi-vie très courte (2 minutes) nécessite une utilisation en perfusion continue. Elle exerce ses effets sur le système cardiovasculaire par la stimulation des récepteurs α-adrénergique (vasoconstricteur), β-adrénergique (inotrope et chronotrope) et dopaminergique (vasodilatateur). L’effet global de la dopamine est dépendant de nombreux facteurs intrinsèques et extrinsèque dont la posologie. En effet, l’affinité de la dopamine pour chacun des récepteur est dépendante de sa concentration plasmatique. Une utilisation à 10 µg/kg/min est recommandée. (SCHNELLBACHER, et al., 2012).
- La dobutamine La dobutamine est une catécholamine sympathomimétique de synthèse. Sa demi-vie est très courte (2 à 3 minutes) et justifie une utilisation en perfusion continue. Son action sur le système cardiovasculaire est essentiellement reliée à son action agoniste sur les récepteurs β1-adrénergiques. Elle possède également un effet β2 agoniste faible et aurait un effet agoniste sur les récepteurs α1-adrénergiques. A faible dose, elle améliore la contractilité, le volume d’éjection systolique sans augmentation de la fréquence cardiaque. A doses fortes, elle induit une tachycardie, une vasoconstriction, de l’hypertension et des arythmies. La dobutamine est indiquée lors de dysfonctionnement systolique et de choc cardiogénique. Des arythmies sont observées à des doses de 15 µg/kg/min et correspondent à une tachycardie ainsi qu’un BAV du second ordre. Ces arythmies sont vraisemblablement secondaires à l’hypoxie myocardique induite par la tachycardie (SCHNELLBACHER, et al., 2012).
- L’adrénaline L’adrénaline est un agent α et β-adrénergique dont les effets alpha sont les plus marquées. Elle stimule l’activité cardiaque tout en provoquant une vasoconstriction périphérique, une augmentation du flux sanguine coronaire (suite à une augmentation de la pression diastolique aortique) et une augmentation du flux sanguin cérébral (COSTELLO, 2004). Elle permet donc d’améliorer la perfusion cérébrale et myocardique. Elle est utilisée lors de réanimation cardiaque ou de bradycardie à la posologie de 0.1 mg/kg IC ,IM, IV, IO, IT CPR (RITCHIE, et al., 1994)(DONELEY, 2011) (COLES, et al., 2007). En conclusion, les inotropes positifs sont utilisés afin d’améliorer la contractilité cardiaque. Dans la plupart des cas, ils ont le désavantage d’augmenter la consommation myocardique en oxygène et de provoquer des arythmies. Ils sont contre-indiqués lors de cardiomyopathie hypertrophique.

Page | 133
2. Les inotropes négatifs
Les inotropes négatifs sont théoriquement représentés par les antagonistes calciques cardiospécifiques et par les antagonistes β-adrénergiques. L’effet des antagonistes calciques sur l’inotropisme cardiaque n’a pas été évalué chez les oiseaux et nous détaillerons seulement l’utilisation des antagonistes β-adrénergiques. Le propranolol (antagoniste non spécifique β1- β2) a une marge thérapeutique faible et une demi-vie courte (PEES, et al., 2009). Il provoque une diminution de la pression artérielle (KUO, et al., 2001) (KAMIKURA, et al., 1995) et préviendrait le remodelage cardiaque lors d’intoxication à la furazolidone (STRUNK, 2003). L’atenolol et le practolol (β1 sélectifs) diminuent la fréquence cardiaque ainsi que la pression artérielle moyenne chez les poulets (KAMIKURA, et al., 1995). Le carteolol semble intéressant pour inverser le remodelage cardiaque observé lors d’intoxication à la furazolidone chez les dindes. Le labetalol (alpha et bêta bloquant) provoque une diminution significative de la fréquence cardiaque et de la pression artérielle chez les dindes. Des études complémentaires sont nécessaires pour évaluer la sécurité à long terme de l’utilisation des bêta-bloquants chez les oiseaux (STRUNK, 2003).
B. Les modificateurs des vaisseaux
1. Vasoconstricteurs
L’adrénaline et la dopamine décrites précédemment ont des effets vasoconstricteurs. C’est également le cas de la noradrénaline. Les effets de la noradrénaline s’installent en 1 minute et disparaissent en 2-3 minutes chez les amazones. La noradrénaline agit comme un agoniste α-adrénergique et provoque une vasoconstriction. Elle peut être utilisée en CRI à une posologie variant de 0,4-5 µg/kg/min. Des effets secondaires (bradycardie) correspondant vraisemblablement à un choc vagal ont été observés à partir de 3 µg/kg/min (ZEHNDER, et al., 2009).
2. Vasodilatateurs
a. Les inhibiteurs de l’enzyme de conversion (IECA)
Pharmacocinétique : La pharmacocinétique n’a pas été documentée chez les oiseaux. Il semblerait que le captopril ai une durée de vie inférieur à celle de l’enalapril et du benazepril. L’utilisation de ces derniers est donc préférée (PEES, et al., 2009). Mode d’action : Les IECA agissent en bloquant la formation de l’angiotensine II. Ainsi, ils bloquent le système rénine-angiotensine-aldostérone ce qui empêche la vasoconstriction artérielle et veineuse ainsi que la rétention hydro-sodée normalement engendrées par ce système. Une telle inhibition aboutit à une amélioration de la fonction rénale, à une augmentation de la diurèse ainsi qu’à une diminution de la pression sanguine. En agissant de la sorte, ils diminuent la pré- et la post-charge du cœur, autorisant un repos des cardiomyocytes(PEES, et al., 2005). L’utilisation de captopril chez des cailles a provoqué une diminution de la consommation d’eau. Les IECA ne provoquent pas de baisse majeur de la pression artérielle moyenne (PEES, et al., 2006). Indications Les principales indications de l’utilisation des IECA sont, selon certains auteurs, les altération du ventricule droit (hypertrophie, dilatation, insuffisance de la valve atrioventriculaire droite) et la congestion qui leur est secondaire au sein de la grande circulation (congestion hépatique, hydropéricarde et ascite). Ils seraient également indiqués lors de dysfonctionnement ventriculaire gauche et lors d’hydropéricarde idiopathique (PEES, et al., 2005).

Page | 134
Effets secondaires Les IECA semblent relativement sécuritaires (PEES, et al., 2005). En effet, des pigeons surdosés à 10 mg/kg/jr pendant 3 semaines n’ont montré aucune altération clinique ni dégradation des paramètres de laboratoire (PEES, et al., 2009). Les effets secondaires potentiels peuvent inclurent une hypotension, une tachycardie réflexe, une déshydratation, des troubles gastro-intestinaux, un dysfonctionnement rénal et une hyperkaliémie (DONELEY, 2011) (DE WIT, et al., 2005). Toutefois, le seul effet secondaire observé cliniquement au court d’une thérapie à long terme à une dose de 5mg/kg/jour par voie orale a été une augmentation de l’hématocrite, possiblement causée par une augmentation de la diurèse. La déshydratation pouvant être sévère, l’utilisation d’enalapril à long terme doit être suivie attentivement à l’aide d’examen sanguins. Une réduction de la dose après amélioration de l’état de santé général est conseillé (PEES, et al., 2005). En conséquence, son utilisation est non recommandée si le patient est atteint de maladie rénale (COLES, et al., 2007). Posologie recommandée : Le médicament est généralement administré par voie orale, en diluant des comprimés dans de l’eau. Ceci facilite le dosage et l’administration. L’acceptance de la molécule est décrite comme extraordinairement bonne par certains auteurs ce qui semble dû à une bonne appétence de la solution. Ceci permet une bonne observance (PEES, et al., 2005).
Enalapril : Les doses utilisées varie de 0.25 mg/kg PO q24-48h (COLES, et al., 2007) à 1.25 mg/kg PO q12-24h (PEES, et al., 2009) (TRISTAN, 2010). Certains auteurs conseillent une dose d’attaque de 5mg/kg q24h diminuée à 1 mg/kg q24h lorsque la fonction cardiaque s’améliore (JONES, 2008)(STRAUB, et al., 2003). Benazepril : Les doses empiriques suivantes ont été utilisés dans la littérature : 0.5 mg/kg PO q12-24h (DONELEY, 2011) (BOWLES, et al., 2007) (SEDACCA, et al., 2009), 5mg/kg PO q24h (STRAUB, et al., 2003), 1.25mg/kg (PEES, et al., 2009) Captopril : 0.1 mg/kg IV (PREZOTO, et al., 2009). Imidapril : 0,1 ml/kg/j ou 1 goutte pour 250g (Prilium® Chiens nains, 75mg Suspension buvable) (BOUSSARIE, et al., 2010).
b. Les agonistes β-adrénergiques
L’isoxsuprine est un agoniste β2-adrénergique et provoque une vasodilatation périphérique (COLES, et al., 2007). Elle a été utilisée à la posologie de 5-10mg/kg PO q24h chez des rapaces atteints d’œdème de la pointe alaire (LEWIS, et al., 1993) et chez un perroquet atteint d’athérosclérose. Chez le perroquet, le traitement a été poursuit pendant prêt de 2 ans et les tentatives d’arrêt provoquaient la réapparition des symptomes. Aucun effet secondaire n’a été remarqué (SIMONE-FREILICHER, 2007).
c. Les antagonistes calciques
La pranidipine est un antagoniste des canaux du calcium de type dihydropyridine. Elle a une longue durée d’action et agit spécifiquement sur les vaisseaux. Utilisée à 10 mg/kg PO q24h elle diminue la post-charge et limite les remodelages ventriculaires chez des dindes ayant une cardiopathie induite expérimentalement avec de la furazolidone (STRUNK, 2003).
d. Les dérivés xanthiques
La propentofylline peut être utilisée à 25 mg/kg PO q48h. Elle facilite l’oxygénation cérébrale chez les perroquets séniles atteint de cardiopathie. Elle a également été utilisée pour le traitement de l'oedème de la pointe des ailes (COLES, et al., 2007).

Page | 135
3. Les diurétiques
Les diurétiques sont utilisés afin de diminuer la surcharge volumique, la congestion organique, les œdèmes et les épanchements. Toutefois, la diminution du volume sanguin vasculaire peut également engendrer une diminution délétère du débit cardiaque. En conséquence, l’intérêt d’utiliser ces agents devra être mesuré.
a. Les diurétiques de l’anse : le furosémide
Pharmacocinétique : La pharmacocinétique du furosémide n’a pas été étudiée chez les oiseaux. Mode d’action : Les mécanismes pharmacodynamiques exacts du furosémide ne sont pas connus chez les oiseaux. Il semble induire une augmentation de la diurèse en intervenant sur les anses de Henlé des néphrons mammaliens (JONES, 2008), bien que ceux-ci ne représentent que 10 à 30% des néphrons du rein des oiseaux (GOLDSTEIN, et al., 2000). Indications : Le furosémide est indiqué lorsque la surcharge volumique est un problème et notamment lors d’œdème (JONES, 2008) et d’ascite (DONELEY, 2011). Il permet de faciliter le travail cardiaque en diminuant la précharge et la postcharge (PEES, et al., 2009). Il est donc indiqué lors d’insuffisance cardiaque congestive (BOUSSARIE, et al., 2010). Certains auteurs le recommandent également lors d’épanchement péricardique (JONES, 2008), mais cela ne fait pas consensus (DONELEY, 2011). Contre-indications Le furosémide semble être de moindre intérêt dans le traitement médical de l’hydropéricarde car son efficacité serait limitée. De plus, lors de tamponnade les ventricules sont peu remplis, et une diminution de la volémie pourrait être dommageable (STRAUB, et al., 2003). Effets secondaires : Les loriquets sont réputés être très sensibles à cette molécule et elle doit donc être utilisée avec précaution chez ces patients (HAWKINS, 2013). Un surdosage peut entraîner une déshydratation et des anomalies électrolytiques (DONELEY, 2011). Comme chez les mammifères, un traitement au long terme peut provoquer un déficit potassique et une déshydratation, ce qui justifie un monitoring étroit (pesée quotidienne pour évaluer la perte fluidique, mesure dee l’urée et de l’acide urique)(JONES, 2008). Les effets secondaires des traitements diurétiques incluent anorexie, vomissements, hypovolémie et hypokaliémie (STRUNK, 2003). Dose recommandée : Le furosémide est habituellement utilisé à la dose de 0.15-2mg/kg IM, SC ou PO q12-24h (DONELEY, 2011) (DE MATOS, et al., 2005) (MITCHELL, et al., 2008) (HARRISON, et al., 2006) (PEES, et al., 2009). Toutefois, il est possible de monter la posologie jusqu’à 2.2mg/kg PO BID (GRAHAM, et al., 2007)(OGLESBEE, et al., 2001) et certains auteurs ont administré des doses telles que 2,5mg/kg IM q12-24h ou 5 mg/kg PO q12-24h (ESFANDIARY, et al., 2010).
b. Les diurétiques thiazidiques : l’hydrochlorthiazide
Une étude révèle que l’utilisation d’hydrochlorthiazide à 10 mg/kg PO n’induit pas une diurèse significativement différente de celle d’un animal non traité (ESFANDIARY, et al., 2010).
c. Les diurétiques distaux : la spironolactone
La spironolactone est un antagoniste de l’aldostérone. C’est un diurétique épargnant le potassium qui a été utilisé avec succès à une dose de 1-2 mg/kg PO q12-24h chez des perroquets et des poulets.. Toutefois, une étude révèle que l’utilisation de spironolactone à 1 mg/kg n’induit pas une diurèse significativement différente que chez un animal non traité (ESFANDIARY, et al., 2010). En pratique, le traitement est initié avec du furosémide et une fois que le patient est stable, la spironolactone est ajoutée puis la dose de furosémide est alors diminuée. Les niveau électrolytiques (K+ en particulier) doivent être monitoré avec précaution (STRUNK, 2003).

Page | 136
II. Le traitement des maladies cardiovasculaires Il est important de signaler que les cas rencontrés en cardiologie aviaires correspondent généralement à des altérations de grande intensité, diagnostiquées alors que la maladie est déjà à un stade de décompensation plus ou moins avancé. Ainsi tous les oiseaux présentant des problèmes cardiaques devraient être considérés comme des urgences vitales. De manière générale, la thérapeutique cardiovasculaire aura trois objectifs : 1/ Rétablir médicalement une fonction circulatoire correcte 2/Apporter des soins de support adéquat 3/ Traiter la cause sous jacente de la maladie si cela est possible (PEES, et al., 2009).
Chez les oiseaux de compagnie, le traitement visera à améliorer la qualité et la durée de vie. La décision d’initier un plan thérapeutique chez les rapaces (sauvages ou utilisés en fauconnerie) doit prendre en grande considération le pronostic de retour à des capacités de vol parfaitement normal, nécessaire à la chasse. Bien que certaines conditions (endocardite bactérienne) puissent correctement répondre au traitement, l’euthanasie est indiquée pour des raisons éthiques dans de nombreux cas de maladie cardiaque avancée (JONES, 2008). Chez les volailles d’élevage, le coût des traitements et le temps nécessaire à la réalisation des soins rendent généralement impossible l’instauration d’un traitement.
A. Traitement des syndromes cardio-vasculaires
Lors de dysfonctionnement cardiaque, le foie peut être congestionné et le flux sanguin rénal diminué. Ceci peut entraîner un dysfonctionnement voire des lésions de ces organes (WELLE, et al., 2011) associées à une augmentation des métabolites toxiques. Ces derniers pourront alors affecter l’état général de l’oiseau. De plus les poumons peuvent être congestionnés ce qui réduit leur efficacité. En conséquence, il faudra tenter de rétablir un fonctionnement normal du système cardiovasculaire (PEES, et al., 2006).
1. Gestion de l’insuffisance cardiaque
Les stratégies thérapeutiques de l’insuffisance cardiaque congestive ont pour but de bloquer l’activation hormonale excessive, d’améliorer le débit cardiaque, de supporter le fonctionnement myocardique et de contrôler les signes de congestion (LUMEIJ, et al., 1994). Ceci est permis par la mise en place du triptique thérapeutique suivant : Inhibiteurs de l’enzyme de conversion de l’angiotensine : En bloquant la formation de l’angiotensine II, les IECA diminuent les effets de l’activation neurohormonale excessive et favorisent la vasodilatation ainsi que la diurèse. Si l’oiseau recoit un traitement diurétique, il peut être préférable de d’instaurer les IECA à une dose faible tout en diminuant l’autre thérapie diurétique (DE WIT, et al., 2005). Cela permet de réduire les risques d’hypotension et de tachycardie réflexe (les IECA ont un effet diurétique). Dans un second temps, la dose d’IECA sera progressivement augmentée (DONELEY, 2011). Thérapie diurétique : La thérapie diurétique est indiquée pour contrôler l’œdème pulmonaire cardiogénique et les épanchements. Un surdodage de diurétique peut engendrer une perte volumique excessive et activer la cascade-rénine-angiotensine-aldostérone. En conséquence, la dose effective la plus basse doit toujours être utilisée (RITCHIE, et al., 1994). Soutien de la fonction cardiaque : La digoxine permet d’améliorer le fonctionnement myocardique et donc de soutenir le débit cardiaque (LUMEIJ, et al., 1994). Le pimobendane est une alternative plus sécuritaire (BEAUFRERE, et al., 2009).

Page | 137
2. Gestion des dysrythmies
Si les arythmies causent une instabilité hémodynamique chez l’oiseau, le traitement est nécessaire. Le traitement des arythmies est guidé par la cause sous-jacente et il est possible d’ajouter au plan thérapeutique des molécules anti-arythmiques ou des agents médicamenteux soutenant la fonction cardiaque. Puisqu’il n’y a pas ou peu de références quant à l’utilisation clinique des molécules anti-arythmiques en médecine aviaire, la plupart des protocoles proposés sont extrapolés des thérapies anti-arthmiques utilisées chez le chien et le chat (DE WIT, et al., 2005). Avant de traiter une arythmie, l’existence de déséquilibres électrolytiques doit être exclue.
-Tachycardie supra-ventriculaire (fibrillation atriale) : Le traitement vise à diminuer la vitesse de conduction atrio-ventriculaire, laissant ainsi davantage de temps au ventricule pour se remplir, ce qui réduit l’impact relatif des contractions atriales. La digoxine est la molécule à utiliser en première intention mais un bêta-bloquant peut-être utilisé afin de réduire davantage la conduction atrio-ventriculaire (DE WIT, et al., 2005). - Tachycardie ventriculaire : Puisque la digoxine prédispose à l’apparition d’arythmies ventriculaires, celle-ci ne représente par la molécule à utiliser en première intention. En revanche, les bêta-bloquants peuvent être utilisés. - Bradycardie et blocs atrio-ventriculaires : Les options thérapeutiques pharmacologiques sont d’intérêt limité. Toutefois, les agents anticholinergiques et sympathomimétiques sont parfois administrés pour essayer d’augmenter la fréquence de réponse ventriculaire et de diminuer les signes associés aux bradyarythmies légères. L’administration d’atropine corrige habituellement la bradycardie et les blocs atrio-ventriculaires qui sont reliés à un tonus vagal élevé. Un gris du gabon (Psittacus erithacus) souffrant de bradycardie et de BAV II a été traité avec de l’éphédrine (0.5 mg/kg q8h) sans effets indésirables pendant plus d’un an (DE WIT, et al., 2005)(LIERZ, et al., 2012).
3. Gestion de l’arrêt cardiaque
Le massage cardiaque est difficile à réaliser chez les oiseaux car le cœur est contenu dans une boîte osseuse formée par les côtes, la colone vertébrale et le bréchet. Cette boîte rend la compression des chambres cardiaques difficile, si ce n’est impossible (DE MATOS, et al., 2005). De plus, les oiseaux n’ont pas de diaphragme ce qui rend impossible l’utilisation du mécanisme de pompe thoracique lors de la compression à thorax fermé. Ainsi il n’est pas possible de diminuer la pression intrathoracique négative (LICHTENBERGER, 2005). Le massage cardiaque est encore plus inefficace en cas d’hémopéricarde ou de tamponnade cardiaque (COSTELLO, 2004). Le praticien peut toutefois tenter d'exercer une pression digitale intermittente sur le sternum. Les pressions sur le bréchet seront alternées avec une pression coelomique caudale pour simuler la fréquence respiratoire normale. Toutefois, cette pratique n’est généralement pas très fructueuse (LIERZ, et al., 2012). Chez les oiseaux d'environ 350-400g (comme les gris du gabon ou les amazones) un massage cardiaque direct en utilisant les doigts à travers une incision abdominale ou en utilisant un coton-tige humidifié peut être entrepris mais n'est généralement guère plus concluant (COLES, et al., 2007). La compression doit être réalisée avec une fréquence rapide, le but étant de réaliser des compressions efficaces à la fréquence approchant celle du patient. Une sonde doppler peut être placé en regard de l’artère digitale pour évaluer l’efficacité du massage (COSTELLO, 2004).

Page | 138
L’atropine et le glycopyrrolate sont des agents anticholinergiques recommandés lors de bradycardie, d’intoxication aux organophosphorés (GRAHAM, et al., 2007), d’arrêt cardiaque liée à une stimulation du vague, d’asystole ou d’activité électrique sans pouls (COSTELLO, 2004). L’atropine est utilisée à la dose de 0.01-0.02 mg/kg IM,SC (RITCHIE, et al., 1994) jusqu’à 0.2–0.5 mg/kg IM, IV, IO, IT (GRAHAM, et al., 2007) (DE MATOS, et al., 2005). Bien que l’administration d’atropine et d’adrénaline en intra-cardiaque a été recommandée par certains auteurs, cette pratique ne devrait plus être réalisée en raison des nombreux risques de lacération des artères coronaires, d’hémopéricarde et d’arythmies intraitables que cette voie comporte (COSTELLO, 2004). Ces médicaments peuvent être administrés par voie intra-veineuse ou par voie intra-osseuse. La voie d’administration intra-trachéale peut être utilisée pour l’atropine et l’adrénaline. Il faut alors multiplier les doses par 2 à 2.5 et diluer la molécule dans de l’eau saline stérile pour obtenir un volume de 2-4 ml et faciliter l’absorption (COSTELLO, 2004).
4. Gestion de l’insuffisance artérielle
Peu de données sont disponibles à ce jour. Toutefois, l’isoxuprine, un vasodilatateur périphérique, a été utilisé avec succès dans la gestion des signes clinique d’un perroquet suspecté d’être atteint d’athérosclérose (SIMONE-FREILICHER, 2007).
B. Traitement de support
Nous avons déjà vu comment les médicaments agissant sur le système cardio-vasculaire permettait de diminuer la congestion et d’améliorer le débit cardiaque. La mise en place d’une thérapie adjuvante est essentielle pour maintenir le fonctionnement des organes et l’activité circulatoire. Les mesures de support décrites dans ce paragraphe devraient également être instaurées (PEES, et al., 2009).
1. Fluidothérapie
L’administration de fluide est essentielle lors de la thérapie. Elle permet : - de prévenir ou de corriger l’apparition d’une déshydratation extracellulaire lors d’utilisation de diurétiques. Ainsi, l’existence d’une azotémie justifiera l’administration de fluide. Les cristalloïdes sont excellents en tant que fluide de réhydratation. Le déficit de fluide (pourcentage de déshydratation x poids corporel) doit être corrigé sur 12 à 24 heures chez des oiseaux sans atteintes cardiopulmonaire ou sur 24 à 36 heure si un tel trouble existe. A ce volume doit être ajouté des fluides de maintien à une dose de 2 (DE MATOS, et al., 2005) à 4 ml /kg/jour (LICHTENBERGER, 2004). - de corriger les déficits de perfusion lors de mesures de réanimation (collapsus circulatoire, choc). La thérapie devrait être initiée par voie intra-veineuse ou intra-osseuse puis l’administration en maintenance peut être relayée par une réhydratation par voie sous-cutanée ou orale. Un suivi attentif devra être réalisé afin que le patient ne soit pas sujet à une surcharge volumique ce qui augmenterait la congestion (DONELEY, 2011). De façon générale, les oiseaux montrant un choc hypovolémique doivent recevoir des colloïdes tièdes à la dose de 10 ml/kg, la dose étant répétée jusqu’à ce que que le patient montre une amélioration clinique. Des cristalloïdes peuvent également être administrés à la dose de 30 à 60 ml/kg/h en cas de faible perfusion. Lors d’hypotension associée à une hypovolémie, il sera possible d’utiliser des bolus de cristalloïdes à 10 ml/kg et de colloïdes à 5 ml/kg sur une minute jusqu’à rétablissement d’une pression normale. Généralement un ou deux bolus suffisent à traiter une hypotension secondaire à une hypovolémie chez les oiseaux (LICHTENBERGER, 2005) (LICHTENBERGER, 2004).

Page | 139
2. Alimentation
L’ajout ou la substitution d’électrolytes, de vitamines et d’acides aminés peut être indiquée. En cas d’athérosclérose, l’ajout de Vitamine A, B, C, E et de Sélénium est recommandée (PEES, et al., 2009) (DONELEY, 2011)
3. Chaleur et humidité
Les oiseaux doivent être place dans un incubateur chaud (25°C au minimum, idéalement entre 30-32°C) (DONELEY, 2011) (LICHTENBERGER, 2004) et humide (60% ou plus) afin d’améliorer la circulation sanguine et de diminuer les perte d’énergie. De plus la chaleur pourra permettre de limiter l’apparition d’arythmie. (WELLE, et al., 2011). Ainsi, la tempértature cloacale sera suivie avec attention (COSTELLO, 2004).
4. Oxygénation
Afin de faciliter l’hématose en cas de congestion pulmonaire, les oiseaux seront oxygénés dans une cage riche en O2 pendant 6 à 8 heures (DONELEY, 2011).
5. Stress
Afin de limiter l’effet du stress sur le système cardiovasculaire, les manipulations devront être réduite au minimum. Le positionnement du patient en décubitus ventrodorsal pour une radiographie est une pratique risquée et doit être considérée avec attention. L’environnement doit être calme et temporairement assombri. Enfin, une sédation légère peut aider à réduire le niveau de stress des patients anxieux. (DONELEY, 2011).
6. Résolution des épanchements
La péricardiocentèse devrait être réalisée chez des oiseaux ayant un épanchement péricardique de haut grade. Le retrait du fluide péricardique améliore le fonctionnement cardiaque. La procédure, réalisée sous contrôle échographique est facile et sécuritaire. Après avoir visualisé la cavité péricardique, l’aiguille est placée en regard de la ligne blanche caudalement au sternum et insérée craniodorsalement vers le cœur. La quantité de fluide ponctionnée ne devrait pas dépassé 10 ml/kg de masse corporelle par jour afin de prévenir l’apparition d’un choc hypovolémique. La ponction du foie est nécessaire lors de cette procédure mais ne semble pas avoir d’effet néfaste. Cette procédure devrait toujours être réalisée sous anesthésie à l’isoflurane afin de diminuer le risque de mouvement (PEES, et al., 2009). Une abdominocentèse peut être réalisée afin de diminuer la pression exercée sur les sacs aériens (BAINE, 2012).

Page | 140
C. Traitement étiologique
1. L’athérosclérose
Le traitement de l’athérosclérose représente un défi (WELLE, et al., 2011). Une grande diversité de traitement a été préconisé pour diminuer les niveaux de cholestérol mais aucun ne semble d’efficacité constante (DONELEY, 2011) : Les acides gras (Omega 3, acide alpha-linolénique), le chitosan (un produit fibreux permettant la fixation des lipides,), la statine (inhibiteurs de la HMG-CoA réductase à effet hypolipidémiant), la niacine (Viatmine B3) ont été utilisés chez les humains et leur utilisation a été proposée chez les oiseaux (BOWLES, et al., 2007). Des traitements anti-oxydants tels que de la vitamine C ou l’administration de gel d’aloe vera ont été proposés par certains auteurs. La supplémentation en vitamine A et E ne semble pas avoir d’effet (MANS, et al., 2007). La crocine, un composant chimique naturel de type caroténoïde existant chez les crocus, inhibe la formation d’athérosclérose chez les cailles. La crocine exerce ses effets anti-athérosclérotiques en diminuant le niveau de lipo-protéine de faible densité modifiées par oxydation, et en limitant l’augmentation de calcium intra-cellulaire, un médiateur important lors de la multiplication des cellules musculaires lisses (HE, et al., 2005). Chez les volailles, l’administration de bêta-bloquants (oxprenolol, propranolol) a montré un effet protecteur vis-à-vis du développement des plaques athérosclérotiques mais ne permet pas leur régression (PAULETTO, et al.) (BAINE, 2012). L’utilisation d’inhibiteurs calciques (verapamil, nifedipine, diltiazem) peut diminuer l’extension et la distribution des lésions athérosclérotiques (GARCIA PEREZ, et al., 2003). Le pravastin (inhibiteur de la coenzyme A réductase) associé à du quinapril (IECA) a permis de diminuer la surface des plaques en formation chez des oiseaux dont l’athérosclérose est induite (STRUNK, 2003). Davantage d’études sont nécessaires pour préciser les posologies et l’efficacité de ces pistes thérapeutiques.
2. Les infections
Une thérapie agressive avec des antibiotiques bactéricides (enrofloxacine, amoxicilline-acide clavulanique par exemple) est indiquée chez les oiseaux atteints d’endocardite infectieuse ou de myocardite (DE WIT, et al., 2005). Le choix de l’antibiotique doit idéalement être guidé par la culture sanguine et les résultats des tests de sensibilité (FOLDENAUER, et al., 2009). En cas d’endocardite aiguë, un traitement approprié peut permettre une rémission complète (STRUNK, 2003).
3. Néoplasie
L’excision chirurgical complète est curative pour les cas d’hémangiomes cutanés (REAVILL, 2004). En revanche, dans le cas d’hémangiosarcomes la majorité des tumeurs réapparaissent localement sous quelques jours à quelques mois après excision chirurgicale (SLEDGE, et al., 2006) (SUEDMEYER, et al., 2001). Comme chez les chiens (et basé sur un rapport de guérison chez une perruche ondulée), l’excision chirurgicale complète suivi de radiation, de chimiothérapie ou des deux est recommandée lors d’hémangiosarcome cutané (FREEMAN, et al., 1999).

Page | 141
4. Les intoxications
Lors d’intoxication, la démarche thérapeutique sera la même que celle utilisée chez les Mammifères, avec certaines particularités aviaires. Les mesures à mettre en place se découpent en 4 points à associer aux traitements de support déjà décrits : Etape 1 - Stabiliser le patient : Cette étape consiste à mettre en place une thérapie d’urgence pour maintenir les fonctions circulatoires, respiratoire, neurologiques et métaboliques. Etape 2 - Empêcher la prolongation de l’exposition : Selon les cas il conviendra d’enlever tous les oiseaux de l’environnement affecté, de ventiler la zone, de rincer ou de nettoyer la peau et les plumes de l’oiseau pour retirer les toxines, de diminuer le pourcentage d’isoflurane,… Etape 3 - Retarder l’absorption : Les émétiques étant contre-indiqués chez les oiseaux, l’absorption des toxines est retardée grâce à un lavage du jabot ou du proventricule, à l’utilisation d’adsorbant ou de cathartiques, ou par retrait chirurgical. L’intubation d’un oiseau anesthésié est nécessaire pour la réalisation du lavage ingluvial ou proventriculaire. Etape 4 - Instituer une thérapie à l’aide d’un antagoniste physiologique. Lorsqu’il existe on utilisera l’antidote de la toxine incriminée (LABONDE, 1995). Par exemple, la naloxone antagonise les opioïde, le flumazénil antagonise les benzodiaépine et la yohimbine antagonise les adrénergiques alpha-2 (COSTELLO, 2004).
5. Hydropéricarde
Le traitement doit avoir pour but de gérer la cause initiale. Les diurétiques sont contre-indiqués à cause de l’hypotension sévère observée chez de nombreux patient avec une pré-charge cardiaque réduite. Si l’épanchement péricardique évolue en tamponnade cardiaque, la péricardiocentèse est le traitement de choix (DE WIT, et al., 2005). La fenêstration endoscopique du péricarde est recommandée afin de prélever du fluide péricardique et peut permettre d’empêcher le re-remplissage du péricarde. L’approche endoscopique du cœur doit se faire par les cavités hépato-péritonéales afin de prévenir le drainage des fluides dans les sacs aériens. Du fait que le péricarde ne peut pas être entièrement retiré, il est possible que l’ouverture chirurgicale du péricarde se referme. Cette fermeture provoquera souvent une récidive de l’épanchement péricardique (FITZGERLAD, et al., 2011).
6. La cardiomyopathie dilatée du dindon
L’administration quotidienne de propranolol chez des dindonneaux de souche fortement atteinte par cette maladie permet de retarder la mortalité mais pas de prévenir la maladie (TREMBLAY, et al., 1992). Il est probable que le retard de la mortalité soit secondaire à une amélioration temporaire du transport du calcium ainsi qu’à un contrôle des tachy-arythmies engendrées par les dilatations des cavités cardiaques (VAN VLEET, et al., 1986). L’administration de pranidipine permet de prévenir la dilatation ventriculaire progressive gauche ainsi que l’amincissement pariétal. De plus, microscopiquement, cette molécule engendre une régression de l’hypertrophie des myocytes (KIM, et al., 1999).
7. Les carences
Afin de prévenir l’apparition d’affection cardiovasculaires, les rations doivent être équilibrée de sorte à fournir un apport en minéraux et vitamine adéquat. En cas de carence, le traitement consistera à complémenter la ration (TREMBLAY, et al., 1992).
CONCLUSION : Le traitement spécifique des maladies cardiovasculaires repose sur des mesures visant à moduler le fonctionnement du cœur, des vaisseaux et de la volémie. La pauvreté des données pharmacocinétique et pharmacodynamique des agents utilisés chez les oiseaux incitent à réaliser un suivi de l’état des patients traités médicalement.Des mesures non spécifiques devront compléter le plan thérapeutique.

Page | 142

Page | 143
CONCLUSION

Page | 144

Page | 145
ANNEXES

Page | 146
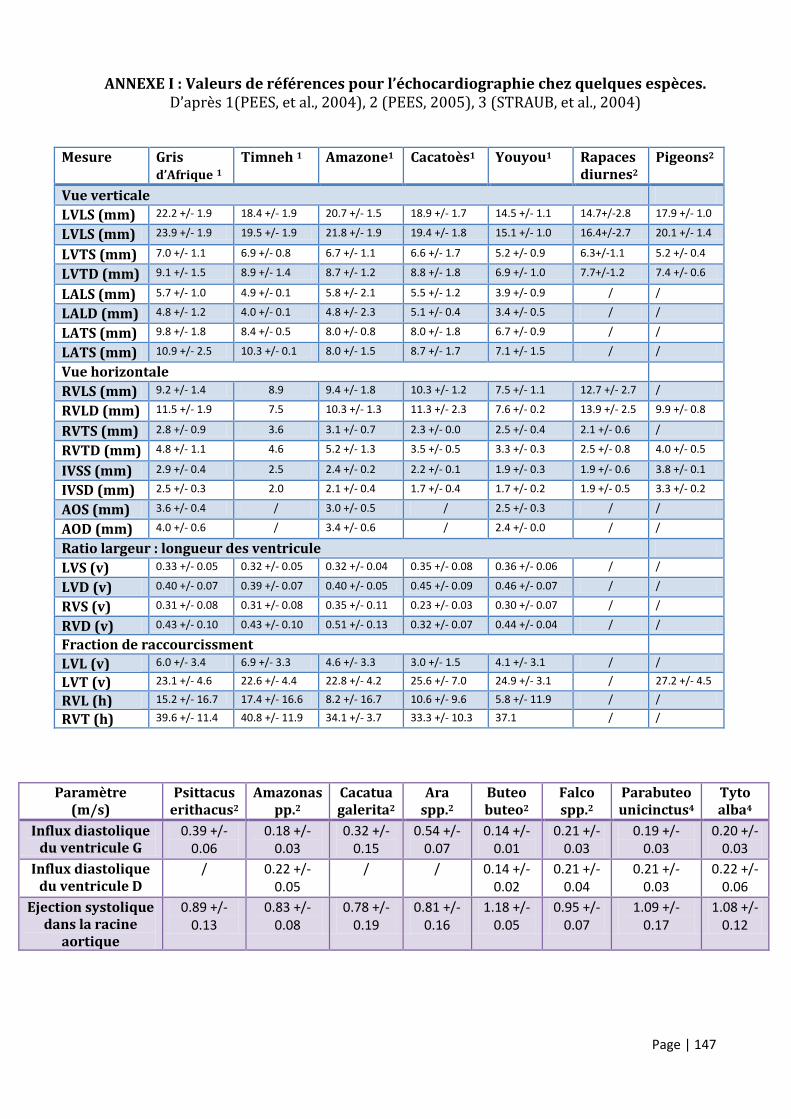
Page | 147
ANNEXE I : Valeurs de références pour l’échocardiographie chez quelques espèces.
D’après 1(PEES, et al., 2004), 2 (PEES, 2005), 3 (STRAUB, et al., 2004)
Mesure Gris d’Afrique 1
Timneh 1 Amazone1 Cacatoès1 Youyou1 Rapaces diurnes2
Pigeons2
Vue verticale
LVLS (mm) 22.2 +/- 1.9 18.4 +/- 1.9 20.7 +/- 1.5 18.9 +/- 1.7 14.5 +/- 1.1 14.7+/-2.8 17.9 +/- 1.0
LVLS (mm) 23.9 +/- 1.9 19.5 +/- 1.9 21.8 +/- 1.9 19.4 +/- 1.8 15.1 +/- 1.0 16.4+/-2.7 20.1 +/- 1.4
LVTS (mm) 7.0 +/- 1.1 6.9 +/- 0.8 6.7 +/- 1.1 6.6 +/- 1.7 5.2 +/- 0.9 6.3+/-1.1 5.2 +/- 0.4
LVTD (mm) 9.1 +/- 1.5 8.9 +/- 1.4 8.7 +/- 1.2 8.8 +/- 1.8 6.9 +/- 1.0 7.7+/-1.2 7.4 +/- 0.6
LALS (mm) 5.7 +/- 1.0 4.9 +/- 0.1 5.8 +/- 2.1 5.5 +/- 1.2 3.9 +/- 0.9 / /
LALD (mm) 4.8 +/- 1.2 4.0 +/- 0.1 4.8 +/- 2.3 5.1 +/- 0.4 3.4 +/- 0.5 / /
LATS (mm) 9.8 +/- 1.8 8.4 +/- 0.5 8.0 +/- 0.8 8.0 +/- 1.8 6.7 +/- 0.9 / /
LATS (mm) 10.9 +/- 2.5 10.3 +/- 0.1 8.0 +/- 1.5 8.7 +/- 1.7 7.1 +/- 1.5 / /
Vue horizontale
RVLS (mm) 9.2 +/- 1.4 8.9 9.4 +/- 1.8 10.3 +/- 1.2 7.5 +/- 1.1 12.7 +/- 2.7 /
RVLD (mm) 11.5 +/- 1.9 7.5 10.3 +/- 1.3 11.3 +/- 2.3 7.6 +/- 0.2 13.9 +/- 2.5 9.9 +/- 0.8
RVTS (mm) 2.8 +/- 0.9 3.6 3.1 +/- 0.7 2.3 +/- 0.0 2.5 +/- 0.4 2.1 +/- 0.6 /
RVTD (mm) 4.8 +/- 1.1 4.6 5.2 +/- 1.3 3.5 +/- 0.5 3.3 +/- 0.3 2.5 +/- 0.8 4.0 +/- 0.5
IVSS (mm) 2.9 +/- 0.4 2.5 2.4 +/- 0.2 2.2 +/- 0.1 1.9 +/- 0.3 1.9 +/- 0.6 3.8 +/- 0.1
IVSD (mm) 2.5 +/- 0.3 2.0 2.1 +/- 0.4 1.7 +/- 0.4 1.7 +/- 0.2 1.9 +/- 0.5 3.3 +/- 0.2
AOS (mm) 3.6 +/- 0.4 / 3.0 +/- 0.5 / 2.5 +/- 0.3 / /
AOD (mm) 4.0 +/- 0.6 / 3.4 +/- 0.6 / 2.4 +/- 0.0 / /
Ratio largeur : longueur des ventricule
LVS (v) 0.33 +/- 0.05 0.32 +/- 0.05 0.32 +/- 0.04 0.35 +/- 0.08 0.36 +/- 0.06 / /
LVD (v) 0.40 +/- 0.07 0.39 +/- 0.07 0.40 +/- 0.05 0.45 +/- 0.09 0.46 +/- 0.07 / /
RVS (v) 0.31 +/- 0.08 0.31 +/- 0.08 0.35 +/- 0.11 0.23 +/- 0.03 0.30 +/- 0.07 / /
RVD (v) 0.43 +/- 0.10 0.43 +/- 0.10 0.51 +/- 0.13 0.32 +/- 0.07 0.44 +/- 0.04 / /
Fraction de raccourcissment
LVL (v) 6.0 +/- 3.4 6.9 +/- 3.3 4.6 +/- 3.3 3.0 +/- 1.5 4.1 +/- 3.1 / /
LVT (v) 23.1 +/- 4.6 22.6 +/- 4.4 22.8 +/- 4.2 25.6 +/- 7.0 24.9 +/- 3.1 / 27.2 +/- 4.5
RVL (h) 15.2 +/- 16.7 17.4 +/- 16.6 8.2 +/- 16.7 10.6 +/- 9.6 5.8 +/- 11.9 / /
RVT (h) 39.6 +/- 11.4 40.8 +/- 11.9 34.1 +/- 3.7 33.3 +/- 10.3 37.1 / /
Paramètre (m/s)
Psittacus erithacus2
Amazonas pp.2
Cacatua galerita2
Ara spp.2
Buteo buteo2
Falco spp.2
Parabuteo unicinctus4
Tyto alba4
Influx diastolique du ventricule G
0.39 +/- 0.06
0.18 +/- 0.03
0.32 +/- 0.15
0.54 +/- 0.07
0.14 +/- 0.01
0.21 +/- 0.03
0.19 +/- 0.03
0.20 +/- 0.03
Influx diastolique du ventricule D
/ 0.22 +/- 0.05
/ / 0.14 +/- 0.02
0.21 +/- 0.04
0.21 +/- 0.03
0.22 +/- 0.06
Ejection systolique dans la racine
aortique
0.89 +/- 0.13
0.83 +/- 0.08
0.78 +/- 0.19
0.81 +/- 0.16
1.18 +/- 0.05
0.95 +/- 0.07
1.09 +/- 0.17
1.08 +/- 0.12

Page | 148
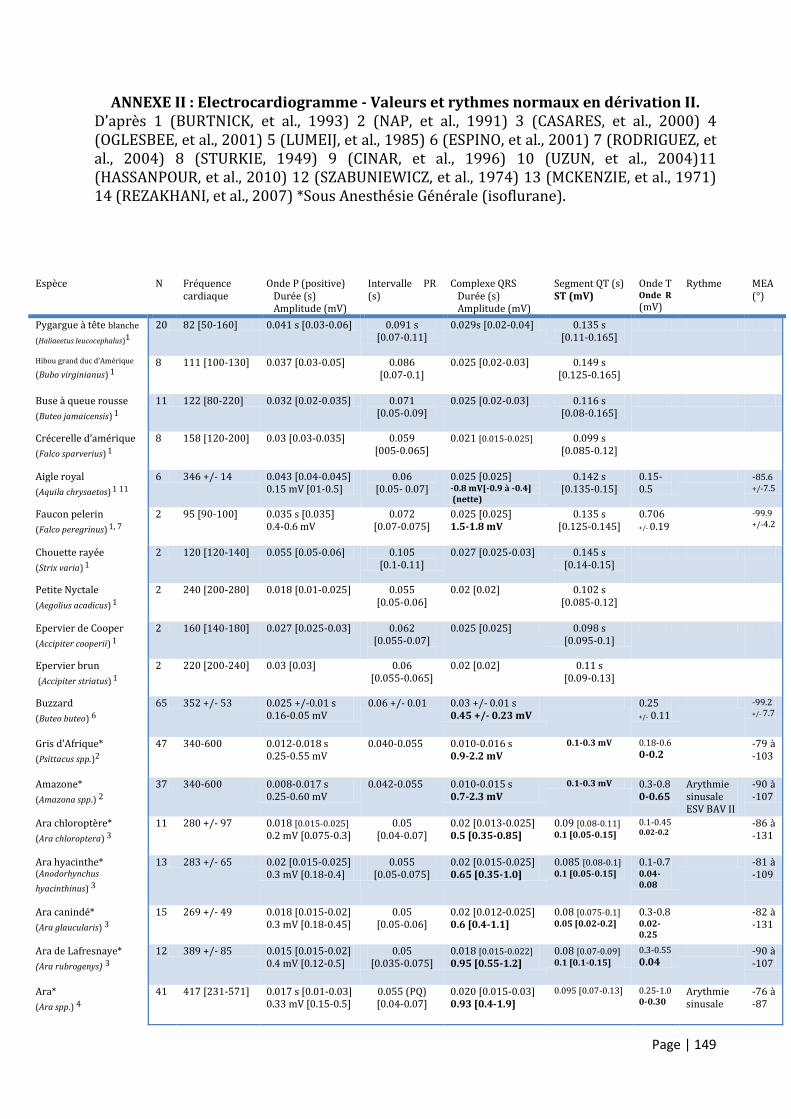
Page | 149
ANNEXE II : Electrocardiogramme - Valeurs et rythmes normaux en dérivation II. D’après 1 (BURTNICK, et al., 1993) 2 (NAP, et al., 1991) 3 (CASARES, et al., 2000) 4 (OGLESBEE, et al., 2001) 5 (LUMEIJ, et al., 1985) 6 (ESPINO, et al., 2001) 7 (RODRIGUEZ, et al., 2004) 8 (STURKIE, 1949) 9 (CINAR, et al., 1996) 10 (UZUN, et al., 2004)11 (HASSANPOUR, et al., 2010) 12 (SZABUNIEWICZ, et al., 1974) 13 (MCKENZIE, et al., 1971) 14 (REZAKHANI, et al., 2007) *Sous Anesthésie Générale (isoflurane).
Espèce N Fréquence cardiaque
Onde P (positive) Durée (s) Amplitude (mV)
Intervalle PR (s)
Complexe QRS Durée (s) Amplitude (mV)
Segment QT (s) ST (mV)
Onde T Onde R (mV)
Rythme MEA (°)
Pygargue à tête blanche
(Haliaeetus leucocephalus)1
20 82 [50-160] 0.041 s [0.03-0.06] 0.091 s [0.07-0.11]
0.029s [0.02-0.04] 0.135 s [0.11-0.165]
Hibou grand duc d’Amérique
(Bubo virginianus) 1 8 111 [100-130] 0.037 [0.03-0.05] 0.086
[0.07-0.1] 0.025 [0.02-0.03] 0.149 s
[0.125-0.165]
Buse à queue rousse
(Buteo jamaicensis) 1
11 122 [80-220] 0.032 [0.02-0.035] 0.071 [0.05-0.09]
0.025 [0.02-0.03] 0.116 s [0.08-0.165]
Crécerelle d’amérique
(Falco sparverius) 1
8 158 [120-200] 0.03 [0.03-0.035] 0.059 [005-0.065]
0.021 [0.015-0.025] 0.099 s [0.085-0.12]
Aigle royal
(Aquila chrysaetos) 1 11
6 346 +/- 14 0.043 [0.04-0.045] 0.15 mV [01-0.5]
0.06 [0.05- 0.07]
0.025 [0.025] -0.8 mV[-0.9 à -0.4] (nette)
0.142 s [0.135-0.15]
0.15-0.5
-85.6 +/-7.5
Faucon pelerin
(Falco peregrinus) 1, 7
2 95 [90-100] 0.035 s [0.035] 0.4-0.6 mV
0.072 [0.07-0.075]
0.025 [0.025] 1.5-1.8 mV
0.135 s [0.125-0.145]
0.706 +/- 0.19
-99.9 +/-4.2
Chouette rayée
(Strix varia) 1
2 120 [120-140] 0.055 [0.05-0.06] 0.105 [0.1-0.11]
0.027 [0.025-0.03] 0.145 s [0.14-0.15]
Petite Nyctale
(Aegolius acadicus) 1
2 240 [200-280] 0.018 [0.01-0.025] 0.055 [0.05-0.06]
0.02 [0.02] 0.102 s [0.085-0.12]
Epervier de Cooper
(Accipiter cooperii) 1
2 160 [140-180] 0.027 [0.025-0.03] 0.062 [0.055-0.07]
0.025 [0.025] 0.098 s [0.095-0.1]
Epervier brun
(Accipiter striatus) 1
2 220 [200-240] 0.03 [0.03] 0.06 [0.055-0.065]
0.02 [0.02] 0.11 s [0.09-0.13]
Buzzard
(Buteo buteo) 6
65 352 +/- 53 0.025 +/-0.01 s 0.16-0.05 mV
0.06 +/- 0.01 0.03 +/- 0.01 s 0.45 +/- 0.23 mV
0.25 +/- 0.11
-99.2 +/- 7.7
Gris d’Afrique*
(Psittacus spp.)2
47 340-600 0.012-0.018 s 0.25-0.55 mV
0.040-0.055 0.010-0.016 s 0.9-2.2 mV
0.1-0.3 mV 0.18-0.6
0-0.2 -79 à
-103
Amazone*
(Amazona spp.) 2
37 340-600 0.008-0.017 s 0.25-0.60 mV
0.042-0.055 0.010-0.015 s 0.7-2.3 mV
0.1-0.3 mV 0.3-0.8 0-0.65
Arythmie sinusale ESV BAV II
-90 à -107
Ara chloroptère*
(Ara chloroptera) 3
11 280 +/- 97 0.018 [0.015-0.025]
0.2 mV [0.075-0.3] 0.05
[0.04-0.07] 0.02 [0.013-0.025] 0.5 [0.35-0.85]
0.09 [0.08-0.11] 0.1 [0.05-0.15]
0.1-0.45 0.02-0.2
-86 à -131
Ara hyacinthe* (Anodorhynchus
hyacinthinus) 3
13 283 +/- 65 0.02 [0.015-0.025] 0.3 mV [0.18-0.4]
0.055 [0.05-0.075]
0.02 [0.015-0.025] 0.65 [0.35-1.0]
0.085 [0.08-0.1] 0.1 [0.05-0.15]
0.1-0.7 0.04-0.08
-81 à -109
Ara canindé*
(Ara glaucularis) 3
15 269 +/- 49 0.018 [0.015-0.02] 0.3 mV [0.18-0.45]
0.05 [0.05-0.06]
0.02 [0.012-0.025] 0.6 [0.4-1.1]
0.08 [0.075-0.1] 0.05 [0.02-0.2]
0.3-0.8 0.02-0.25
-82 à -131
Ara de Lafresnaye*
(Ara rubrogenys) 3
12 389 +/- 85 0.015 [0.015-0.02] 0.4 mV [0.12-0.5]
0.05 [0.035-0.075]
0.018 [0.015-0.022] 0.95 [0.55-1.2]
0.08 [0.07-0.09] 0.1 [0.1-0.15]
0.3-0.55
0.04 -90 à
-107
Ara*
(Ara spp.) 4
41 417 [231-571] 0.017 s [0.01-0.03] 0.33 mV [0.15-0.5]
0.055 (PQ) [0.04-0.07]
0.020 [0.015-0.03] 0.93 [0.4-1.9]
0.095 [0.07-0.13] 0.25-1.0 0-0.30
Arythmie sinusale
-76 à -87
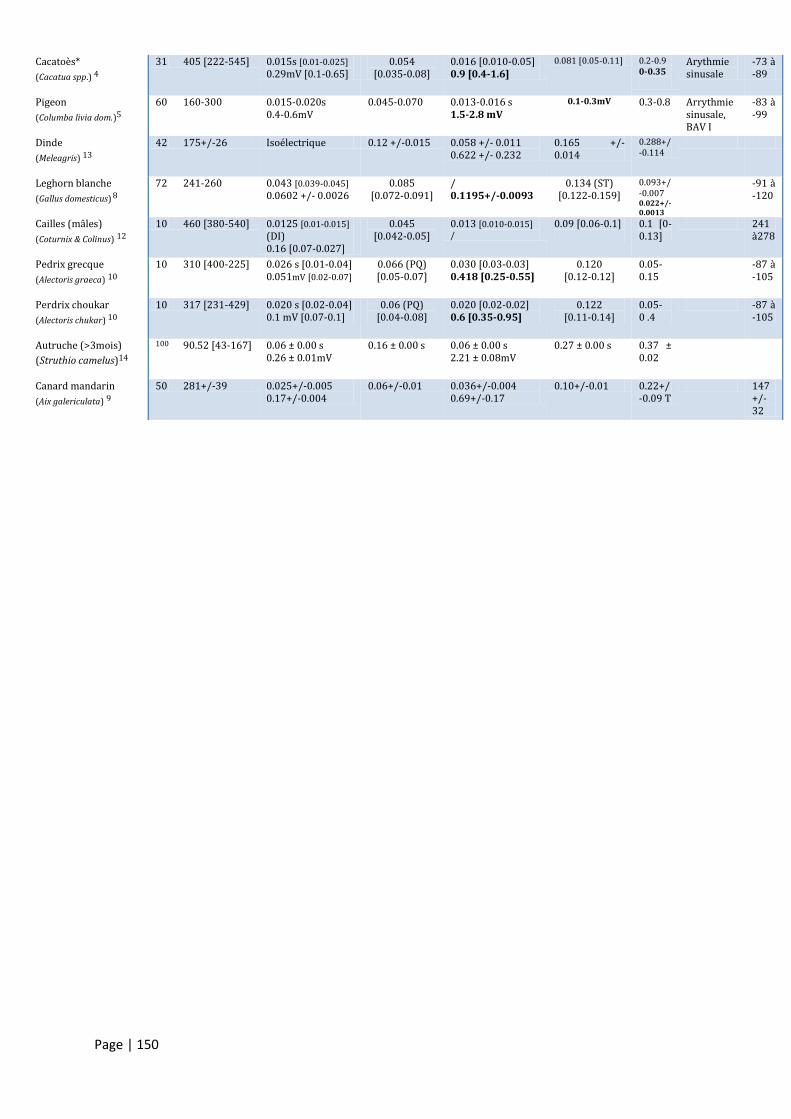
Page | 150
Cacatoès*
(Cacatua spp.) 4
31 405 [222-545] 0.015s [0.01-0.025] 0.29mV [0.1-0.65]
0.054 [0.035-0.08]
0.016 [0.010-0.05] 0.9 [0.4-1.6]
0.081 [0.05-0.11] 0.2-0.9 0-0.35
Arythmie sinusale
-73 à -89
Pigeon
(Columba livia dom.)5
60 160-300 0.015-0.020s 0.4-0.6mV
0.045-0.070 0.013-0.016 s 1.5-2.8 mV
0.1-0.3mV 0.3-0.8 Arrythmie sinusale, BAV I
-83 à -99
Dinde
(Meleagris) 13
42 175+/-26 Isoélectrique 0.12 +/-0.015 0.058 +/- 0.011 0.622 +/- 0.232
0.165 +/- 0.014
0.288+/-0.114
Leghorn blanche
(Gallus domesticus) 8
72 241-260 0.043 [0.039-0.045]
0.0602 +/- 0.0026 0.085
[0.072-0.091] / 0.1195+/-0.0093
0.134 (ST) [0.122-0.159]
0.093+/-0.007 0.022+/-0.0013
-91 à -120
Cailles (mâles)
(Coturnix & Colinus) 12
10 460 [380-540] 0.0125 [0.01-0.015]
(DI) 0.16 [0.07-0.027]
0.045 [0.042-0.05]
0.013 [0.010-0.015] /
0.09 [0.06-0.1]
0.1 [0-0.13]
241 à278
Pedrix grecque
(Alectoris graeca) 10
10 310 [400-225] 0.026 s [0.01-0.04] 0.051mV [0.02-0.07]
0.066 (PQ) [0.05-0.07]
0.030 [0.03-0.03] 0.418 [0.25-0.55]
0.120 [0.12-0.12]
0.05-0.15
-87 à -105
Perdrix choukar
(Alectoris chukar) 10
10 317 [231-429] 0.020 s [0.02-0.04] 0.1 mV [0.07-0.1]
0.06 (PQ) [0.04-0.08]
0.020 [0.02-0.02] 0.6 [0.35-0.95]
0.122 [0.11-0.14]
0.05-0 .4
-87 à -105
Autruche (>3mois)
(Struthio camelus)14
100 90.52 [43-167] 0.06 ± 0.00 s 0.26 ± 0.01mV
0.16 ± 0.00 s 0.06 ± 0.00 s 2.21 ± 0.08mV
0.27 ± 0.00 s 0.37 ± 0.02
Canard mandarin
(Aix galericulata) 9
50 281+/-39 0.025+/-0.005 0.17+/-0.004
0.06+/-0.01 0.036+/-0.004 0.69+/-0.17
0.10+/-0.01 0.22+/-0.09 T
147+/-32
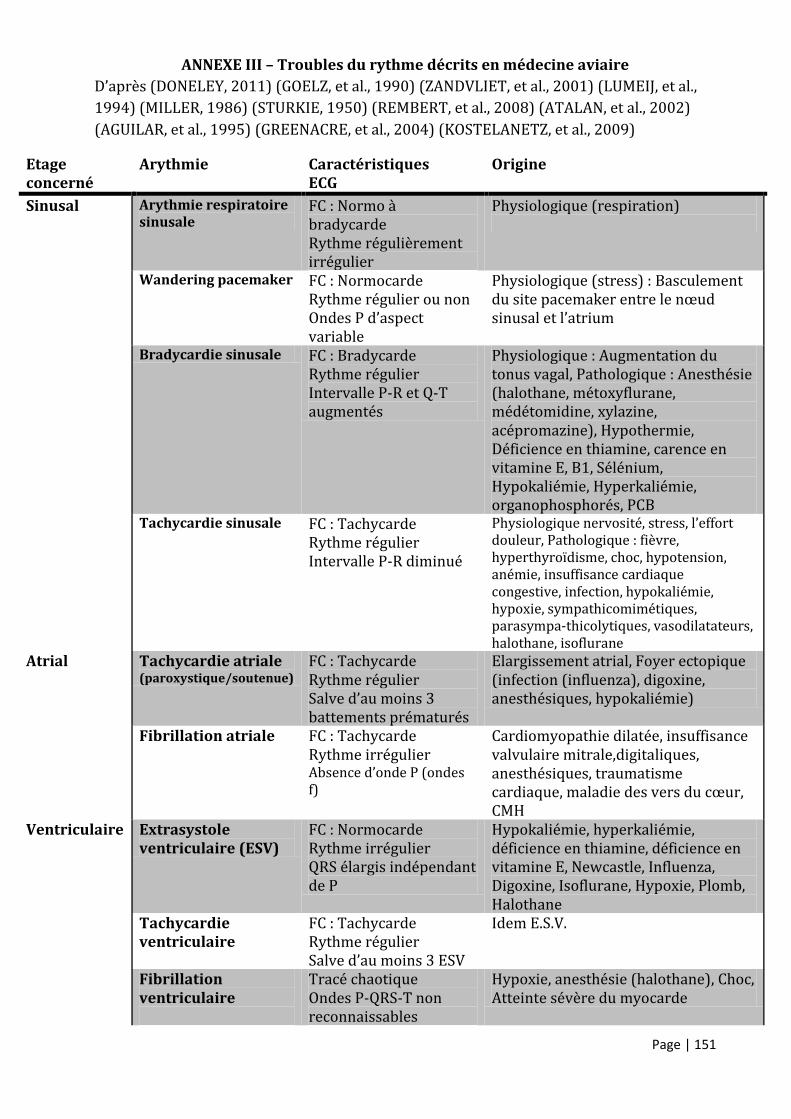
Page | 151
ANNEXE III – Troubles du rythme décrits en médecine aviaire
D’après (DONELEY, 2011) (GOELZ, et al., 1990) (ZANDVLIET, et al., 2001) (LUMEIJ, et al.,
1994) (MILLER, 1986) (STURKIE, 1950) (REMBERT, et al., 2008) (ATALAN, et al., 2002)
(AGUILAR, et al., 1995) (GREENACRE, et al., 2004) (KOSTELANETZ, et al., 2009)
Etage concerné
Arythmie Caractéristiques ECG
Origine
Sinusal Arythmie respiratoire sinusale
FC : Normo à bradycarde Rythme régulièrement irrégulier
Physiologique (respiration)
Wandering pacemaker FC : Normocarde Rythme régulier ou non Ondes P d’aspect variable
Physiologique (stress) : Basculement du site pacemaker entre le nœud sinusal et l’atrium
Bradycardie sinusale FC : Bradycarde Rythme régulier Intervalle P-R et Q-T augmentés
Physiologique : Augmentation du tonus vagal, Pathologique : Anesthésie (halothane, métoxyflurane, médétomidine, xylazine, acépromazine), Hypothermie, Déficience en thiamine, carence en vitamine E, B1, Sélénium, Hypokaliémie, Hyperkaliémie, organophosphorés, PCB
Tachycardie sinusale FC : Tachycarde Rythme régulier Intervalle P-R diminué
Physiologique nervosité, stress, l’effort douleur, Pathologique : fièvre, hyperthyroïdisme, choc, hypotension, anémie, insuffisance cardiaque congestive, infection, hypokaliémie, hypoxie, sympathicomimétiques, parasympa-thicolytiques, vasodilatateurs, halothane, isoflurane
Atrial Tachycardie atriale (paroxystique/soutenue)
FC : Tachycarde Rythme régulier Salve d’au moins 3 battements prématurés
Elargissement atrial, Foyer ectopique (infection (influenza), digoxine, anesthésiques, hypokaliémie)
Fibrillation atriale FC : Tachycarde Rythme irrégulier Absence d’onde P (ondes f)
Cardiomyopathie dilatée, insuffisance valvulaire mitrale,digitaliques, anesthésiques, traumatisme cardiaque, maladie des vers du cœur, CMH
Ventriculaire Extrasystole ventriculaire (ESV)
FC : Normocarde Rythme irrégulier QRS élargis indépendant de P
Hypokaliémie, hyperkaliémie, déficience en thiamine, déficience en vitamine E, Newcastle, Influenza, Digoxine, Isoflurane, Hypoxie, Plomb, Halothane
Tachycardie ventriculaire
FC : Tachycarde Rythme régulier Salve d’au moins 3 ESV
Idem E.S.V.
Fibrillation ventriculaire
Tracé chaotique Ondes P-QRS-T non reconnaissables
Hypoxie, anesthésie (halothane), Choc, Atteinte sévère du myocarde
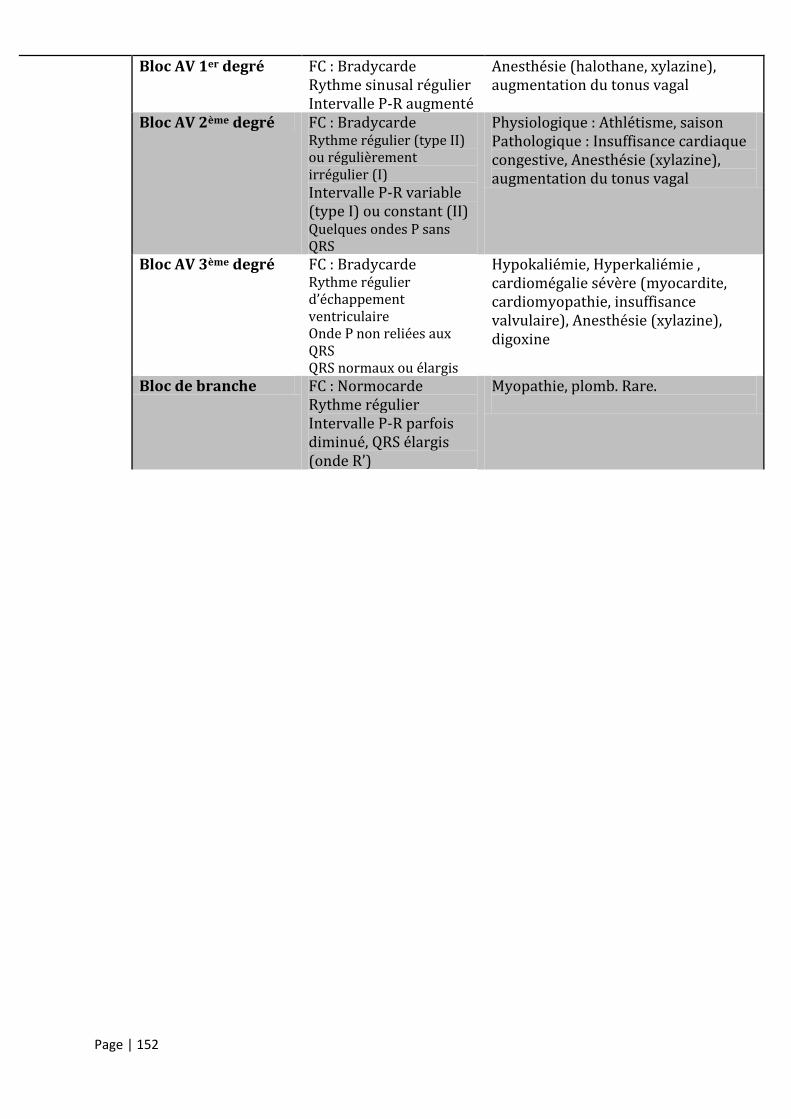
Page | 152
Bloc AV 1er degré FC : Bradycarde Rythme sinusal régulier Intervalle P-R augmenté
Anesthésie (halothane, xylazine), augmentation du tonus vagal
Bloc AV 2ème degré FC : Bradycarde Rythme régulier (type II) ou régulièrement irrégulier (I) Intervalle P-R variable (type I) ou constant (II) Quelques ondes P sans QRS
Physiologique : Athlétisme, saison Pathologique : Insuffisance cardiaque congestive, Anesthésie (xylazine), augmentation du tonus vagal
Bloc AV 3ème degré FC : Bradycarde Rythme régulier d’échappement ventriculaire Onde P non reliées aux QRS QRS normaux ou élargis
Hypokaliémie, Hyperkaliémie , cardiomégalie sévère (myocardite, cardiomyopathie, insuffisance valvulaire), Anesthésie (xylazine), digoxine
Bloc de branche FC : Normocarde Rythme régulier Intervalle P-R parfois diminué, QRS élargis (onde R’)
Myopathie, plomb. Rare.

Page | 153
Bibliographie ACIERNO, M.J., et al. 2008. Agreement between direct and indirect blood pressure measurements
obtained from anesthetized Hispaniolan Amazon parrots. Journal of the American Veterinary Medical
Association. 2008, Vol. 233, 10, pp. 1587-1590.
ADAMS, W.E. 1937. A Contribution to the Anatomy of the Avian Heart as seen in the Kiwi (Apteryx
australis) and the Yellow-crested Penguin (Megadyptes antipodum). Proceedings of the Zoological
Society of London. September, 1937, Vol. B107, 3, pp. 417-441.
AGUILAR, R.F., et al. 1995. Arrythmias associated with isoflurane anesthesia in bald eagles (Haliaeetus
leucocephalus). J Zoo Wildl Med. 1995, Vol. 24, pp. 508-516.
AKAGAMI, M., et al. 2010. Pathogenesis of venous hypertrophy associated with schistosomiasis in
Whooper swans (cygnus cygnus) in Japan. Avian diseases. Mach, 2010, Vol. 54, 1, pp. 146-150.
AKANBI, O.B., et al. 2008. Systemic listeriosis in caged canaries (Serinus canarius). Avian pathology. June
2008, Vol. 37, 3, pp. 329-332.
AKESTER, A.R. 1971. The blood vascular system. [book auth.] D.J. BELL and B.M. FREEMAN. Physiology
and biochemistry of the domestic fowl. London : Academic Press, 1971, Vol. 2, pp. 783-837.
—. 1984. The cardiovascular system. [book auth.] B.M. FREEMAN. Physiology and biochemistry of the
domestic fowl. London : Academic Press, 1984, Vol. 5, pp. 172-257.
ANDRAL, B., et al. 1990. Picorna-like viruses of young turkeys: pathogenesis of a disease of poults
caused by a picorna-like virus. Avian pathology. 1990, Vol. 19, pp. 245-254.
APPLE, F.S. 1999. The Specificity of Biochemical Markers of Cardiac Damage: a problem solved. Clin
Chem Lab Med. 1999, Vol. 37, 11, pp. 1085-1089.
ATALAN, G., UZUN, M. and al, DEMIRKAN I. et. 2002. Effect of medetomidine-butorphanol-ketamine
anaesthesia and atipamezole on heart and respiratory rate and cloacal temperature of domestic
pigeons. J Vet Med A Physiok Pathol Clin Med. 2002, Vol. 49, pp. 281-285.
BAILEY, T.A. and KINNE, J. 201. Ventricular septal defect in a Houbara Bustard (Chlamydotis undulata
macqueenii). Avian diseases. 201, Vol. 45, pp. 229-233.
BAINE, K. 2012. Management of the geriatric psittacine patient. Journal of Exotic Pet Medicine. 2012,
Vol. 21, pp. 140-148.
BALL, et al. 1963. Morphological characteristics of the anterior mestenteric artery of the fowl. Anatomy
record. 1963, 146, pp. 251-256.
BAPTISTE, K.E., et al. 1997. Dissecting aortic aneurysm associated with a right ventricular arterioveinous
shunt in a mature Ostrich (Struthio camelus). Journal of Avian Medicine and Surgery. September, 1997,
Vol. 11, 3, pp. 194-200.
BARONE, R. 1996. Artères-Caractères généraux. Anatomie comparée des animaux domestiques Tome 5
Angiologie. Paris : Vigot, 1996, pp. 103-122.

Page | 154
BARTLETT, C.M. and ANDERSON, R.C. 1936. Paronchocerca struthionus n. sp. (Nematoda Filaroidea)
from ostriches (Struthio camelus), with a redescription of Paronchocerca ciconiarum Peters. Can J Zool.
1936, Vol. 64, pp. 2480-2491.
BARTLETT, C.M. 2008. Filarioid nematodes. [book auth.] C.T. ATKINSON, N.J. THOMAS and D.B. HUNTER.
Parasitic diseases of wild birds. Ames, Iowa : Wiley-Blackwell, 2008, pp. 439-462.
BAUMEL, J.J. and GERCHMANN, L. 1968. The avian intercarotid anastomosis and its homologue in other
vertebrates. American Journal of Anatomy. 1968, Vol. 128, pp. 1-18.
BAUMEL, J.J. 1993. Systema cardiovasculare. Handbook of avian anatomy : nomina anatomica avium
2nd edition. Cambridge, MA : Nuttall Ornithological Club, 1993, pp. 407-476.
BAUMEL, J.J., DALLEY, A.F. and QUINN, T.H. 1983. The collar plexus of subcutaneous thermoregulatory
veins in the pigeon, Columba livia ; its association with esophageal pulsation and gular flutter.
Zoomorphology. 1983, Vol. 102, pp. 215-239.
BAVELAAR, F.J. and BEYNEN, A.C. 2004. Atherosclerosis in parrots. A review. Vet Q. 2004, Vol. 26, 2, pp.
50-60.
—. 2003. Severity of atherosclerosis in parrots in relation to the intake of alpha-linolenic acid. Avian
diseases. 2003, Vol. 47, pp. 566-577.
BAVELAAR, F.J. BEYNEN, A.C. 2003. Influence of amount and type of dietary fat on plasma cholesterol
concentrations in African Grey Parrots. The Journal of Applied Research in Veterinary Medicine. Winter,
2003, Bd. 1, 1, S. 1-8.
BEAUFRERE, H. and PARIAUT, R. 2009. Therapeutic review. Pimobendan. J. exot. Pet Med. PRECISER LE
MOIS, 2009, Vol. 18, 4, pp. 311-313.
BEAUFRERE, H., et al. 2010. Avian vascular imaging - a review. Journal of avian medicine and surgery.
September, 2010, Vol. 24, 3, pp. 174-184.
BEAUFRERE, H., et al. 2011. Characterization and classification of psittacine atherosclerotic lesions by
histopathology, digital image analysis, and electron microscopy. Avian Pathology. October 2011, Vol. 40,
5, pp. 531-544.
BEAUFRERE, H., et al. Comparison of Transcoelomic, Contrast, and Transesophageal Echocardiography
in Anesthetized red-tailed hawk. In Press.
BEAUFRERE, H., et al. 2010. Feasibility of transesophageal echocardiography in birds without cardiac
disease. Journal of American Veterinary Medical Association. March, 2010, Vol. 236, 5, pp. 540-547.
BEAUFRERE, H., et al. 2011. Intermittent claudication-like syndrome secondary to atherosclerosis in a
Yellow-naped amazon parrot (Amazona ochrocephala auropalliata). Journal of avian medicine and
surgery. 2011, Vol. 25, 4, pp. 266-276.
BEAUFRERE, H., et al. 2013. Prevalence and risk factors in psittacine atherosclerosis: a multicenter case-
control study. Journal of the american veterinary association. 2013.
BEAUFRERE, H., NEVAREZ, J.G. and GASCHEN, L. et al. 2010. Presumed acute ischemic stroke in an
African grey parrot (Psittacus erithacus erithacus) : diagnosis and seizure treatment. Proceeding of the
Annual Association of Avian Veterinarians. 2010.

Page | 155
BEAUTHIER, J.P. and LEFEVRE, P. 1993. Traité d'anatomie : de la théorie à la pratique palpatoire.
Bruxelles : De boeck université, 1993. 2 8041 1500 3.
BEEHLER, B.A., MONTALI, R.J. and BUSH, M. 1980. Mitral valve insufficiency with congestive heart
failure in a pukeko. Journal of American Veterinary Medical Association. 1980, Vol. 177, pp. 934-937.
BENETT, R.A. 1994. Neurology. [book auth.] G.J. HARRISON, L.R. HARRISON and B.W. RITCHIE. Avian
Medicine : Principles and Application. Lake Worth (FL) : Wingers Publishing, 1994, pp. 723-747.
BERHANE, Y., et al. 2001. Peripheral neuritis in psittacine birds with proventricular dilatation disease.
Avian pathology. 2001, 30, pp. 563-570.
BERLINER, J.A. et al. 1995. Atherosclerosis : basic mechanisms. Oxidation, inflammation, and genetics.
Circulation. 91, 1995, Vol. 9, pp. 2488-2496.
BERNIER, G., MORIN, M. and MARSOLAIS, G. 1981. A generalized inclusion body disease in the
budgerigar (Melopsittacus undulatus) caused by a papovavirus like agent. Avian Diseases. 1981, 25, pp.
1083-1092.
BHAVSAR, S.K., et al. 2010. Monensin Induced Suicidal Erythrocyte Death. Cell Physiol Biochem. 2010,
Vol. 25, pp. 745-752.
BIESIADECKI, B.J. and JIN, J-P. 2002. Exon Skipping in Cardiac Troponin T of Turkeys with Inherited
Dilated Cardiomyopathy. The journal of biological chemistry. May 24, 2002, Vol. 277, 21, pp. 18459-
18458.
BIGLAND, C.H., LNI, S.K. and PERRY, M.L. 1964. Five cases of Serratospiculure amaculata (Nematoda:
Filaroidea) infection in prairie falcons (Falco mexicanus). Avian Diseases. 1964, Vol. 8, pp. 412-419.
BISHOP, C.M. and BUTLER, P.J. 1995. Physiological modelling of oxygen consumption in birds during
flight. The journal of experimental biology. 1995, 198, pp. 2153-2163.
BISWAS, P.K., et al. 2002. Adenovirus induced hydropericardium-hepatitis syndrome in broiler parent
chickens in chittagong, Bangladesh. Pakistan journal of biological sciences. 2002, Vol. 5, 9, pp. 994-996.
BOGIN, E. and ISRAELI, B. 1976. Enzyme profile of heart and skeletal muscles, liver and lung of roosters
and geese. Zentralbl Veterinarmed A. 1976, Vol. 23, pp. 152-157.
BOHORQUEZ, F. and STOUT, C. 1972. Aortic atheroclerosis in exotic avians. Experimental and molecular
pathology. 1972, Vol. 17, pp. 261-273.
BOMASSI, E., CORLOUER, J-P. and ROUSSELOT, J-F. 2010. La cardiologie au quotidien Guide pratique.
France : Selecta Impression, 2010.
BOTTE, V. and PELAGALLI, G.V. 1982. Anatomia funzionale degli uccelli domestici . Milano : Edi.Ermes,
1982. 88-85019-19-6.
BOUGIOUKLIS, P.A., BRELLOU, G. and GEORGOPOULOU, I. 2005. Rupture of the right auricle in brouiler
chickens. Avian Pathology. 2005, Vol. 34, pp. 388-391.
BOULIANNE, M., HUNTER, D.B. and JULIAN, R.J. et al. 1994. Cardiac muscle mass distribution in the
domestic turkey and relationship to electrocardiogram. Avian diseases. 1994, Vol. 36, pp. 582-589.

Page | 156
BOUSSARIE, D. and RIVAL, F. 2010. Planète NAC. s.l. : Vétoquinol, 2010.
BOWLES, H., LENNOX, A. and LICHTENBERGER, M. 2007. Emergency and critical care in pet birds.
Veterinary Clinics of North America : Exotic Animal Practuce. 2007, 10, pp. 345-494.
BOX, E.D. and SMITH, J.H. 1982. The intermediate host-spectrum in a sarcocystis species of birds. J.
Parasitol. 1982, Vol. 68, pp. 668-679.
BRANDAO, J. 2013. Congestive Heart Failure in a Harris Hawk (Parabuteo Unicinctus). In 12th Proc Annu
Conf Europ Assoc Avian Vet. 2013.
BRAUN, E.J. 1998. Comparative renal function in reptiles, birds and Mammals. Seminars in Avian and
Exotic Pet Medicine. Avril, 1998, Vol. 7, 2, pp. 62-71.
BRUGER, H. 1992. Particularités de la physiologie des oiseaux. [book auth.] J. BRUGERE-PICOUX and A.
SILIM. Manuel de pathologie aviaire. MAISONS-ALFORT : Imprimerie du Cercle des Elèves de l'Ecole
Nationale Vétérinaire d'Alfort, 1992, pp. 15-24.
BURGER, W.P., et al. 1994. Cardiomyopathy in ostriches (Struthio camelus) due to avocado (Persea
americana var. guatemalensis) intoxication. J S Afr Vet Assoc. September, 1994, Vol. 65, 3, pp. 113-118.
BURSTEIN, H., et al. 2007. Viral aetiology of haemangiosarcoma outbreaks among layer hens. Avian
pathology. 2007, Vol. 13:4, pp. 715-726.
BURTNICK, N.L. and DEGERNES, L.A. 1993. Eletrocardiography on fifty-nine anesthetized convalescing
raptors. [book auth.] P.T. REDIG, J.E. COOPER and J.D. et al REMPLE. Raptor biomedicine. Minneapolis :
University of Minnesota Press, 1993, pp. 111-121.
BUTLER, P., WEST, N. and JONES, D. 1977. Respiratory and cardiovascular responses of the pigeon to
sustained level flight in a wind-tunnel. J Exp Biol. 1977, Vols. 7-17.
BUTLER, P.J. and JONES, D.R. 1997. Physiology of Diving of Birds and Mammals. Physiological Reviews.
1997, Vol. 77, 3, pp. 837-899.
BUTLER, P.J. and WOAKS, A.J. 1982. Control of heart rate by carotid body chemoreceptors during diving
in tufted ducks. Journal of applied physiology. 1982, Vol. 53, pp. 1405-1410.
BUTLER, P.J., et al. 2000. Heart rate and rate of oxygen consumption during flight of Barnacle Goose.
Comparative biochemistry and physiology part A. 2000, Vol. 1266, pp. 379-385.
CAMELLITI, P., BORG, T.K. and KOHL, P. 2005. Structural and functional characterisation of cardiac
fibroblasts. Cardiovascular Research. 2005, Vol. 65, pp. 40-51.
CAMPBELL, T.W. 1994. Hematology. [book auth.] B. RITCHIE, G. HARRISON and L. editors HARRISON.
Avian Medicine, Principles and Applications. Lake Worth (FL) : Wingers Publishing, 1994, pp. 176-198.
CANNY, C. 1992. Gross anatomy and imaging of the avian and reptilian urinary system. Seminars in
Avian and Exotic Pet Medicine. April, 1992, Vol. 7, 2, pp. 72-80.
CAPITELLI, R. and CROSTA, L. 2013. Overview of Psittacine blood analysis and comparative retrospective
study of clinical diagnosis, hematology and blood chemistry in selected psittacine species. Veterinary
Clinic Exotic Animal. 2013, Vol. 16, pp. 71-120.

Page | 157
CARTER, J.K., PROCTOR, S.J. and SMITH, R.E. 1983. Induction of angiosarcoma by Ring-Necked pheasant
virus. Infectious immunology. 1983, Vol. 40, pp. 310-319.
CARUS, C.G. 1838. Traité élémentaire d'anatomie comparée. Bruxelles : Meline, Cans et compagnie,
1838.
CASARES, M., ENDERS, F. and MONTOYA, J.A. 2000. Comparative electrocardiography in four species of
macaws (genera Anodorhynchus and Ara). J Vet Med A Physiol Pathol Clin Med. 2000, Vol. 47, pp. 277-
281.
CAZAYOUX, C.A. 1992. Myocarditis as a component of psittacine proventricular dilatation syndrome in a
Patagonian Conure. Avian Diseases. 1992, Vol. 36, pp. 1117-1119.
CERRUTI SOLA, S., BORELLO, B. and CASTAGNARO, M. 1997. Occurrence of cutaneous haemangiomas
in chickens : Morphological aspects. Avian Pathology. 1997, Vol. 26, 3, pp. 501-510.
CHARLTON, B.R., et al. 1991. Systemic trichomoniasis in a squab operation. Avian Diseases. 1991, Vol.
35, pp. 426-432.
CHATELAIN, E. 1992. L'anatomie des oiseaux. [book auth.] J. BRUGER-PICOUX and A. SILIM. Manuel de
pathologie aviaire. MAISONS-ALFORT : Imprimer du Cercle des Elèves de l'Ecole Nationale Vétérinaire
d'Alfort, 1992, pp. 25-36.
CHENG, Z., LIU, J. and ZHANG, L. 2010. Tumors associated with avian leukosis subgroup J in layers Hens
during 2007 to 2009 in Chna. Journal of Veterinary Medical Science. 2010, Vol. 72, 8, pp. 1027-1033.
CINAR, A., et al. 1996. The electrocardiogram of the Pekin duck. Avian Diseases. October-December,
1996, Vol. 40, 4, pp. 919-923.
COLES, B.H., et al. 2007. Essentials of avian medicine and surgery. 3rd edition. Ames : Blackwell
Publishing, 2007. 978-1-4051-5755-1.
COOPER, J.E. and POMERANCE, A. 1982. Cardiac lesions in birds of prey. Journal of comparative
pathology. 1982, Vol. 92, pp. 161-168.
CORK, S.C. 2000. Iron storage diseases in birds. Avian pathology. 2000, Vol. 29, pp. 7-12.
COSTELLO, M.F. 2004. Principles of Cardiopulmonary Cerebral Resuscitation in Special species. Seminars
in Avian and Exotic Pet Medicine. July 2004, Vol. 13, 3, pp. 132-141.
COURCHESNE, S. and GARNER, M. 2009. What is your diagnosis. Journal of avian medicine and surgery.
2009, Vol. 23, 1, pp. 69-73.
CRAWLEY, R.R., ERNST, J.V. and MILTON, J.L. 1982. Sacocystis in a bald eagle (haliaeetus
leucocephalus). Journal of Wildlife Diseases. April, 1982, Vol. 18, 2, pp. 253-255.
CRESPEAU, F. 1992. L'autopsie des oiseaux. [book auth.] J. BRUGER-PICOUX and A. SILIM. Manuel de
patholgie aviaire. MAISONS-ALFORT : Imprimerie du Cercle des Elèves de l'Ecole Nationale Vétérinaire
d'Alfort, 1992.
CRESPO, R. and SHIVAPRASAD, H.L. 2008. Developmental, metabolic, and other noninfectious
disorders. [book auth.] Y.M. SAIF. Diseases of poultry 12th edition. Ames, Iowa : Blackwell Publishing,
2008, pp. 149-1170.

Page | 158
CZARNEKI, C.M. and GOOD, A.L. 1980. Electrocardiographic technic for identifying developing
cardiomyopathies in young turkey poults. Poultry Science. 1980, Vol. 59, pp. 1515-1520.
DAHLHAUSEN, B., SOLER-TOVAR, D. and SAGGESE, M.G. 2012. Diagnosis of mycobacterial infections in
the Exotic Pet Patient with emphasis on birds. Veterinary Clinics of North America : Exotic Animal
Practice. 2012, Vol. 15, 1, pp. 71-83.
DANGLER, C.A. and NJENGA, M.K. 1994. Left ventricular endocardial proliferation in chickens.
Veterinary Pathology. 1994, Vol. 31, pp. 488-491.
DAVENPORT, J. 1992. Animal life at low temperature. London : Chapman & Hall, 1992. p. 101.
DAVIES, F. 1930. Further studies of the conducting system of the bird’s heart. J Anat. 1930, Vol. 64, pp.
319-324.
—. 1930. The conducting system of the bird’s heart. J. Anat. 1930, Vol. 64, pp. 129-146.
DAVIS, J.F., KULKARNI, A. and FLETCHER, O. 2012. Myocarditis in 9- and 11-day-old broiler breeder
chicks associated with a reovirus infection. Avian Diseases. December, 2012, Vol. 56, 4, pp. 786-790.
DAVIS, R.B., BOZEMAN, L.H. and GAUDRY, D. et al. 1981. A viral disease of fledgling budgerigars. Avian
Diseases. 1981, Vol. 25, pp. 179-183.
DAVISON, S. and WIDEMAN, R.F. 1992. Excess sodium bicarbonate in the diet and its effect on leghorn
chickens. Br Poult Sci. 1992, Vol. 33, pp. 859-870.
DE LAFORCADE, A.M. and ROZANSKI, E.A. 2001. Central venous pressure and arterial blood pressure
measurements. Veterinary Clinic of North America Small Animal Practice. 2001, Vol. 31, 6, pp. 1163-
1174.
DE LAS MULAS, J.M., et al. 1993. Multiple cutaneous capillary haemangioma in a red partridge
(Alectoris rufa). Avian Pathology. 1993, Vol. 22, pp. 637-642.
DE MATOS, R. and MORRISEY, J.K. 2005. Emergency and critical care of small psittacine and passerines.
Seminars in avian and exotic pet medicine. Avril, 2005, Vol. 14, 2, pp. 90-105.
DE WIT, M. and SCHOEMAKER, N.J. 2005. Clinical approach to avian cardiac disease. Seminars in Avian
and Exotic Pet Medicine. January, 2005, Vol. 14, 1, pp. 6-13.
DEGERNES, L.A. 2008. Waterfowl Toxicology: A Review. Vet Clin Exot Anim. 2008, Vol. 11, pp. 283-300.
DELK, K. 2012. Clinical management of seizures in avian patients. Journal of Exotic Pet Medicine. 2012,
Vol. 21, pp. 132-139.
DENNIS, P.M., HEARD, D.J. and CASTLEMAN, W.L. 2000. Respiratory distress associated with pulmonary
fat emboli in an Osprey (Pandion haliaetus). Journal of Avian Medicine and Surgery. December, 2000,
Vol. 14, 4, pp. 264-267.
DIAZ, G.J. and JULIAN, R.J., SQUIRES, E.J. 1994. Lesion in broiler chickens following experimental
intoxication with cobalt. Avian Diseases. 1994, Vol. 38, pp. 308-316.
DIERENFELD, E.S., PIOIS, M.T. and SHEPPARD, C.D. 1994. Hemosiderosis and dietary iron in birds. J Clin.
1994, Vol. 124, pp. 26585-26868.

Page | 159
DOMINGO, M., et al. 1991. Heart rupture and haemopericardium in capercaillie (Tetrao urogallus)
reared in captivity. Avian Pathology. 1991, Vol. 20, pp. 363-366.
DONELEY, B. 2011. Disorders of the cardiovascular system. Avian Medicine and Surgery in Practice -
Companion and aviairy birds. s.l. : Manson Publishing, 2011, pp. 191-200.
DOWLING, L. 1992. Ionophore toxicity in chickens: A review of pathology and diagnosis. Avian
Pathology. 1992, Vol. 21, 3, pp. 355-368.
DRISS, A.B., et al. 2000. Haemodynamic stresses induce endothelial dysfunction and remodeling of
pulmonary artery in experimental compensated heart failure. Circulation. 2000, Vol. 101, pp. 2764–
2770.
D'SOUZA, S.P., DAVIS, M. and BAXTER, G.F. 2004. Autocrine and paracrine actions of natriuretic
peptides in the heart. Pharmacology & Therapeutics 101 (2004) 113– 129. 2004, Vol. 101, pp. 113-129.
DUNCAN, A.E. and FITZGERALD, S.D. 1997. Clinical challenge. J Zoo Wildl Med. 1997, Vol. 28, 4, pp. 501-
503.
ECCO, R., et al. 2008. An outbreak of sarcocystosis in psittacines and a pigeon in a zoological collection
in brazil. Avian diseases. December, 2008, Vol. 52, 4, pp. 706-710.
ECHOLS, M.S., CRAIG, T.M. and SPEER, B.L. 2000. Heartworm (Paronchocerca ciconarum) infection in
two Saddle-Billed Storks (Ephippiorynchus senegalensis). Journal of Avian Medicine and Surgery. March,
2000, Vol. 14, 1, pp. 42-47.
EINZIG, S., JANKUS, E.F. and MOLLER, J.H. 1972. Ventricular septal defect in turkeys. American Journal
of Veterinary Research. 1972, 33, pp. 563-566.
ENSLEY, P.K. 1978. Levamisole hydrochloride for treatment of Paronchocerca ciconarum microfilariae in
a marabou stork. J Am Vet Med Assoc. 1978, Vol. 173, pp. 1246-1248.
ENSLEY, P.K., HATKIN, J. and SILVERMAN, S. 1979. Congestive heart failure in a greater hill mynah.
Journal of the American Veterinary Medical Association. 1979, Vol. 175, pp. 1010-1013.
ESFANDIARY, A., et al. 2010. Diuretic effects of several chemical and herbal compounds in adult laying
hens. International Journal of Poultry Science. 2010, Vol. 9, 3, pp. 247-253.
ESPINO, L., et al. 2001. Electrocardiogram reference values for the buzzard in Spain. Journal of Wildlife
Diseases. October, 2001, Vol. 37, 4, pp. 680-685.
Estimating intrathoracic arterial diameter using CT angiography in Hispaniolan Amazon Parrots
(Amazona ventralis) in C.R. Congrès ''AAV Conference''. BEAUFRERE, H., et al. 2009. Milwaukee,
Wisconsin : s.n., 2009.
EUN-JUNG, B., et al. 2010. Myocarditis caused by nematode infection, presumptively sarconema
eurycerca, in a wild whooper swan (cygnus cygnus) in korea. Proceeding de l'ESVP-ECVP. 2010, p. 349.
EVANS, D.E., et al. 2001. Congenital cardiovascular anomalies, including ventricular septal defects, in
two cockatoos. Journal of Avian Medicine and Surgery. June, 2001, Vol. 15, 2, pp. 101-106.
FABRICANT, C.G. and FABRICANT, J. 1999. Atherosclerosis induced by infection with Marek's disease
herpesvirus in chickens. American Heart Journal. November 1999, Vol. 138, 5 Pt 2.

Page | 160
FALK, H., et al. 1979. Hepatic angiosarcoma associated with androgenic-anabolic steroids. Lancet. 1979,
Vol. 2, 8152, pp. 1120-1123.
FAUST, E.C. 1966. Paronchocerca mansoni sp. nov. from Pica pica sericea (Gould) and Caynopica cyana
interposita Hartert, Peking, China with observations on the genus Paronchocerca Peters. J Helminthol.
1966, Vol. 40, pp. 53-62.
FERRERAS, M.C., GONZALEZ, J. and PEREZ, V. 2001. Proximal aortic dissection (dissecting aortic
aneurysm) in a mature ostrich. Avian Diseases. January-March, 2001, Vol. 45, 1, pp. 251-256.
FISCHER, I., CHRISTEN, C. and SCHARF, G. et al. 2005. Cardiomegaly in a whooper swan (Cygnus cygnus).
Veterinary Record. 2005, Vol. 156, 6, pp. 178-182.
FITZGERLAD, B.C. and SPEER, B.L. 2011. Endoscopic partial pericardectomy for resolution of pericardial
effusion in a jenday conure in C.R. Congrès ''11th EAAV Conference''. Madrid : Proc Annu Conf Europ
Assoc Avian Vet, 2011. p. 103.
FOLDENAUER, U., et al. 2009. Endocarditis due to lactobacillus jensenii in a Salvin's amazon parrot
(Amazona autumnalis salvini). Avian pathology. 2009, Vol. 38, 1, pp. 55-58.
FORBES, N.A., COOPER, J.E. and HIGGINS, R.J. 2000. Neoplasms in birds of prey. [book auth.] J.T.
LUMEIJ, J.D. REMPLE and P.T. et al. REDIG. Raptor biomedicine III. Lake Worth (FL) : Zoological Education
Network, 2000, pp. 127-146.
FOX, P.R., PETRIE, J.P. and HOHENHAUS, A.E. 2005. Peripheral vascular disease. [book auth.] S.J.
ETTINGER and E.C. FELDMAN. Textbook of Veterinary Internal Medicine. s.l. : Elsevier-Saunders, 2005,
pp. 1145-1167.
FRANCA, M., et al. 2010. Retrospective stidu of myocarditis associated with reovirus in turkeys. Avian
diseases. September, 2010, Vol. 54, 3, pp. 1026-1031.
FRANK, R.K., et al. 1990. Perirenal hemorrhage syndrome in market turkey toms: effect of management
factors. Avian Diseases. Oct-Dec, 1990, Vol. 34, 4, pp. 833-842.
FREEMAN, K.P., et al. 1999. Radiation therapy for hemangiosarcoma in a budgerigar. Journal of Avian
Medicine and Surgery. 1999, Vol. 13, 1, pp. 40-44.
FULTON, R.M. 2008. Other toxins and poisons. [book auth.] Y.M. SAID. Diseases of poultry 12th edition.
Ames, Iowa : Blackwell Publishing, 2008, pp. 1231-1258.
GABELLA, G. 1997. Morphology of smooth muscle. [book auth.] C.Y. KAO and M.E. KARSTEN. Cellular
aspect of smooth muscle function. Cambridge : Cambridge University Press, 1997, pp. 1-47.
GAL, A.F., et al. 2012. The first description of a congenital right ventricular cardiac aneurysm in a Pigeon
(Columbia livia domestica, Cluj Blue Tumbler Pigeon). Avian Diseases. December, 2012, Vol. 56, 4, pp.
778-780.
GALINDO-CARDIEL, I., et al. 2011. Outbreak of Mortality in Psittacine Birds in a Mixed-Species Aviary
Associated With Erysipelothrix rhusiopathiae infection. Veterinary pathology. 2011, Vol. 49, 3, pp. 498-
502.

Page | 161
GARCIA PEREZ, B., et al. 2003. Planimetric and histological study of the aortae in atherosclerotic
chickens treated with nifedipine, verapamil and diltiazem. Histol Histopathol. 2003, Vol. 18, pp. 1027-
1033.
GARDNER, M.B., et al. 1981. Electron microscopic search for retrovirus in spontaneous tumors of the
parakeet. Veterinary pathology. 1981, Vol. 18, pp. 700-703.
GARTRELL, B.D. and REID, C. 2007. Death by chocolate: A fatal problem for an inquisitive wild parrot.
New Zealand Veterinary Journal. 2007, Vol. 55, 3, pp. 149-151.
GELIS, S., et al. 2006. Mycobacteriosis in Gang gang cockatoos (Callocephalon fimbriatum). Veterinary
clinics, Exotic Animal Practice. 2006, Vol. 9, pp. 487-494.
GENTLE, M.J. 2011. Pain issue in poultry. Applied Animal Behaviour Science. 2011, Vol. 135, pp. 252-258.
GILKA, F. and SPENCER, J.L. 1990. Chronic myocarditis and circulatory syndrome in a White Lehgorn
strain induced by an Avian Leukosis Virus : Light and Electron Microscopic study. Avian Disease. 1990,
Vol. 34, pp. 174-184.
GLENNY, F.H. 1940. A systematic study of the main arteries in the region of the heart - Aves. Anatomical
record. 1940, Vol. 76, pp. 371-380.
GODOY, S.N., et al. 2012. Bacillus cereus infection outbreak in captive Psittacines. Veterinary
Microbiology. December, July 2012, Vols. 161 (1-2), pp. 213-217.
GOELZ, M.F., HAHN, A.W. and KELLEY, S.T. 1990. Effects of halothane and isoflurane on mean arterial
blood pressure, heart rate, and respiratory rate in adult Pekin ducks. American Journal of Veterinary
Research. 1990, Vol. 51, pp. 458-460.
GOLDSTEIN, D.L. and SKADHAUGE, E. 2000. Renal and extrarenal regulation of body fluid composition.
[book auth.] J.C. WHITTOW. Sturkie's Avian Physiology. s.l. : Academic Press, 2000, pp. 265-297.
GOODMAN, G.J. 1996. Metabolic disorders. [book auth.] W.J. ROSSKOPF and R.W. editors WOERPEL.
Diseases of cage and aviary birds. 3rd edition. 3rd. Baltimore : Williams & Wilkins, 1996, pp. 470-479.
GOSSELIN, S.J. and KRAMER, L.W. 1983. Pathophysiology of excessive iron storage in mynah birds.
Journal of the American Veterinary Medical Association. December, 1983, Vol. 183, 11, pp. 1238-1240.
GRAHAM, J.E. and HEALTEY, J.J. 2007. Emergency care of raptors. Vet Clin Exot Anim. 2007, Vol. 10, pp.
395-418.
GRAY, D.A. 1994. Atrial natriuretic peptide suppresses osmotically stimulated plasma arginine vasotocin
levels in the Pekin duck. Gen. Comp. Endocrinol. 1994, Vol. 93, pp. 122-127.
—. 1994. Role of endogenous atrial natriuretic peptide in volume expansion diuresis and natriuresis of
the Pekin duck. J. Endocrinol. 1994, Vol. 140, pp. 85-90.
GRAY, D.A., DOWNING, C. and SAYED, N. 1997. Endogenous plasma atrial natriuretic peptide and the
control of salt gland function in the Pekin duck. Am. J. Physiology. 1997, Vol. 273, pp. R1080-R1085.
GRAY, D.A., SCHUTZ, H. and GERSTBERGER, R. 1991. Interaction of atrial natriuretic factor and
osmoregulatory hormones in the Pekin duck. Gen. Comp. Endocrinol. 1991, Vol. 81, pp. 246-255.

Page | 162
—. 1991. Plasma atrial natriuretic factor responses to blood volume changes in the Pekin duck.
Endocrinology. 1991, Vol. 128, pp. 1655-1660.
GREENACRE, C.B. and LUSBY, A.L. 2004. Physiologic responses of Amazon parrots (Amazona species) to
manual restraint. Journal of Avian Medicine and Surgery. March, 2004, Vol. 18, 1, pp. 19-22.
GREENWOOD, A.G., MARSHALL, J. and TINSLEY, E.G.F. 1996. Vegetative andocarditis in a Waldrapp ibis.
Avian Pathology. 1996, Vol. 25, pp. 387-391.
GREINER, E.C. and RITCHIE, B.W. 1994. Parasites. [book auth.] B.W. RITCHIE, G.J. HARRISON and L.R.
HARRISON. Avian Medicine : Principles and application. Lake Worth, Florida, USA : Wingers Publishing,
Inc., 1994, pp. 1007-1029.
GRESHAM, G.A. and HOWARD, A.N. 1961. Aortic rupture in the turkey. Journal of atherosclerosis
research. 1961, 1, pp. 75-80.
GRINER, L.A. Pathology of zoo animals; a review of necropsies conducted over a 14-year period at the
San Diego zoo an San Diego wild animal park. Zoological society of san diego. pp. 222-226.
GROSS, W.B. and DOMERMUTH, C.H. 1962. Bacterial endocarditis of poultry. American Journal of
Veterinary Research. 1962, Vol. 23, pp. 320-329.
GROSS, W.B. 1966. Electrocardiographic changes of Escherichia coli infected brds. American Journal of
Veterinary Research. 1966, Vol. 27, pp. 1427-1436.
GRUBB, B.R. 1983. Allometric relations of cardiovascular function in birds. American Journal of
Physiology. 1983, Vol. 245, 4, pp. 567-572.
GUY, J.S. and MALKINSON, M. 2008. Arbovirus infections. [book auth.] Y.M., FADLY, A.M. SAIF, et al.
Diseases of poultry 12th edition. Ames, Iowa : Blackwell Publishing, 2008, pp. 414-425.
GUZMAN, D., et al. 2012. Pharmacokinetics of a single oral dose of pimobendan in Hispaniolan Amazon
parrot (Amazona ventralis). In Proc Ass Avian Vet. Louisville : s.n., 2012, pp. 5-6.
GYENAI, K., et al. 2012. An assessment of echocardiography as a diagnostic tool for dilated
cardiomyopathy in turkey (Meleagris gallopavo). American Journal of Animal and Veterinary Sciences.
2012, Vol. 7, 3, pp. 120-125.
HAMLIN, R.L. and STALNAKER, P.S. 1987. Basis for use of digoxin in small birds. Journal of Veterinary
Pharmacology and Therapeutics. 1987, Vol. 10, pp. 354-356.
HANLEY, C.S., et al. 1997. Establishing cardiac measurement standards in three avian species. Journal of
Avian Medicine and Surgery. March, 1997, Vol. 11, 1, pp. 15-19.
HANLEY, C.S., WILSON, G.H. and LATIMER, K.S. et al. 2005. Interclavicular hemangiosarcoma in a
double yellow-headed Amazon parrot (Amazona ochrocephala oratrix). Journal of Avian Medicine and
Surgery. 2005, Vol. 19, 2, pp. 130-137.
HARARI, J. and MILLER, D. 1983. Ventricular septal defect and bacterial endocarditis in a whistling swan.
Journal of Veterinary Medical Association. 1983, 183, pp. 1296-1297.
HARGIS, A.M., et al. 1989. Avocado (Persea americana) intoxication in caged birds. Journal of American
Veterinary Medicine Association. 1989, 194, pp. 64-66.

Page | 163
HARRISON, G.J. and T.L., LIGHTFOOT. 2006. Clinical avian medicine. Palm Beach, FL : Spix, 2006.
HARTMANN, F.A. 1961. Locomotor mechanisms of birds. Smithsonian miscellaneous collection. 1961,
Vol. 143, 1, pp. 1-91.
HASSANPOUR, H., MOGHADDAM, A.K.Z. and BASCHI, M.C. 2010. The normal electrocardiogram of
conscious golden eagles (aquila chrysaetos). Journal of Zoo and Wildlife Medicine. September, 2010, Vol.
41, 3, pp. 426-431.
HASSINEN, E., PYORNILA, A. and HISSA, R. 1999. Cardiovascular and thermoregulatory responses to
vasotocin and angiotensin II in the pigeon. Comparative biochemistry and physiology. 1999, Vol. 123, pp.
279-285.
HAWKINS, M. et al. 2013. Birds. [book auth.] J. CARPENTER and C. MARION. Exotic animal formulary. St
Louis, MO : Elsevier, 2013, pp. 184-438.
HE, S-Y., et al. 2005. Effect of crocin on experimental atherosclerosis in quails and its mechanisms. Life
Sciences. 2005, Vol. 77, pp. 907-921.
HEADLEY, S.A. 2005. Intrathoracic haemangiosarcoma in an ostrich (Struthio camelus). Veterinary
record. 2005, Vol. 156, pp. 353-354.
HERNANDEZ-DIVERS, S.J. and HERNANDEZ-DIVERS, S.M. 2004. Avian diagnostic endoscopy.
Compendium. 2004, Vol. 26, 11, pp. 839-582.
HILL, C.H., et al. 1997. Dietary iron deficiency compromises normal development of elastic fibers in the
aorta and lungs of chicks. The journal of nutrition. August 1997, Vol. 137, 8, pp. 1895-1900.
HILL, J.R. and GOLDBERG, J.M. 1980. P-wave morphology and atrial activation in the domestic fowl.
American Journal of Physiology. November, 1980, Vol. 239, 5, pp. R283-R288.
HIRAKOW, R. 1970. Ultrastructural characteristics of the mammalian and sauropsidian heart. American
Journal of Cardiology. 1970, Vol. 25, p. 195.
HOCHLEITHNER, M. 1994. Biochemistries. [book auth.] B. RITCHIE, G. HARRISON and L. editors
HARRISON. Avian medicine, Principles and Applications. Lake Worth (FL) : Wingers Publishing, 1994, pp.
223-245.
HODGES, R.D. 1974. The histology of the domestic fowl. London : Academic Press, 1974.
HODGES, R.D., et al. 1975. The general ultrastructure of the carotid body of the domestic fowl. Cell ant
Tissue Research. October 27, 1975, Vol. 162, 4, pp. 483-497.
HOERR, F.J. 2008. Mycotoxicoses. [book auth.] Y.M. SAIF. Diseases of poultry. Ames, Iowa : Blackwell
Publishing, 2008, pp. 1197-1229.
HOLLIDAY, C.M., RIDGELY, R.C. and BALANOFF, A.M. et al. 2006. Cephalic vascular anatomy in
flamingos (phoenicopterus ruber) based on novel vascular injection and computed tomography imaging
analyses. Anat Re A Disc Mol Cell Evol Biol. 2006, Vol. 288A, 10, pp. 1031-1041.
HOLZ, P.H. and RAIDAL, S.R. 2006. Comparative renal anatomy of exotic species. Vet Clin Exot Anim.
2006, Vol. 9, pp. 1-11.

Page | 164
HOWERTH, E.W., RICH, G. and DUBEY, J.P. et al. 1991. Fatal toxoplasmosis in a red lorry (Eos bornea).
Avian Diseases. 1991, Vol. 35, pp. 642-646.
HUNSAKER, W.G., ROBERTSON, A. and MAGWOOD, S.E. 1971. The effect of round heart disease on the
electrocardiogram and heart weight of turkey poults. Poultry Science. 1971, Vol. 50, pp. 1712-1720.
HURLEY-SANDERS, J.L., et al. 2012. Use of thermography and fluorescein angiography in the
management of a chilean flamingo with avascular necrosis of the wing. Journal of Avian Medicine and
Surgery. December, 2012, Vol. 26, 4, pp. 255-257.
ISAZA, R., BUERGELT, C. and KOLLIAS, GV. 1992. Bacteremia and vegetative endocarditis associated
with a heart murmur in a blue-and-gold macaw. Avian diseases. Oct-Dec, 1992, Vol. 36, 4, pp. 1112-
1116.
ITCHIE, B.W. and CARTER, K. 1995. Avian viruses. Lake Worth, Fl : Wingers Publishing, 1995.
JESSUP, D.A. 1980. Valvular endocarditis and bacteriemia in a bald eagle. Modern Veterinary Practice.
January, 1980, Vol. 61, 1, pp. 49-51.
JOHNS, J.L., et al. 2009. What is your diagnosis? Blood smear from an injured red-tailed hawk.
Veterinary clinic pathology. June 2009, Vol. 38, 2, pp. 247-252.
JOHNSON, J.H., PHALEN, D.N. and KONDIK, V.H. et al. 1992. Atherosclerosis in psittacine birds.
Proceeding of the Association of Avian Veterinarians Conference. 1992.
JOHNSTON, M.S., et al. 2011. Precision of repeated, doppler-derived indirect blood pressure
measurements in conscious psittacine birds. Journal of Avian Medicine and Surgery. 2011, Vol. 25, 2, pp.
83-90.
JONES, M.P. Selected Infectious Diseases of Birds of Prey. Journal of exotic pet medicine. Vol. 15, 1, pp.
5-17.
JONES, R. 2008. Raptors : systemic and non-infectious diseases. [book auth.] J. CHITTY and M. LIERZ.
BSAVA Raptors, pigeons and passerine birds. BSAVA : s.n., 2008.
JUAN-SALLES, C., et al. 2011. Congestive heart failure in 6 African Grey parrots (Psittacys e erithacus).
Veterinary pathology. 2011, Vol. 48, 3, pp. 691-697.
JULIAN, J.R., MIRSALIMI, S.M. and SQUIRES, E.J. 1993. Effect of hypobaric hypoxia in diet on blood
parameters and pulmonary hypertension-induced right ventricular hypertrophy in turkey poults and
ducklings. Avian pathology. 1993, Vol. 22, pp. 683-692.
JULIAN, R.J. 1993. Ascites in poultry. Review article. Avian Pathology. 1993, Vol. 22, pp. 419-454.
—. 2005. Production and growth related disorders and other diseases of poultry - A review. The
Veterinary Journal. 2005, Vol. 169, pp. 350-369.
JULIAN, R.J., CASTON, L.J. and LEESON, S. 1992. The effect of dietary sodium on right ventricular failure-
induced ascites, gain and fat deposition in meat-type chickens. Can J Vet Res. 1992, Vol. 56, pp. 214-219.
JULIAN, R.J., et al. 1992. Effect of hypoxia and diet on spontaneous turkey cardiomyopathy (round-heart
disease). Avian diseases. 1992, Vol. 36, 4, pp. 1043-1047.

Page | 165
JULIAN, R.J., et al. 1987. The relationship of right ventricular hypertrophy, right ventricular failure, and
ascites to weight gain in broiler and roaster chickens. Avian diseases. 1987, Vol. 31, pp. 130-135.
JUNG, K., et al. 2009. Aspergillus fumigatus infection in two wild Eurasian black vultures (Aegypius
monachus Linnaeus) with carbofuran insecticide poisoning A case report. Veterinary journal. February
2009, Vol. 179, 2, pp. 307-312.
JUNIOR, F.C.F., et al. 2012. Fatal toxoplasmosis in a Vinaceous Amazon parrot (Amazona vinacea). Avian
Diseases. December, 2012, Vol. 56, 4, pp. 774-777.
KAHN, C.M. and LINE, S. 2008. The Merck Veterinary Manual. Whitehouse station, N.J. : Merck Sharp &
Dohme Corp.,, 2008.
KALETA, E.F., BRUCKNER, D. and GOLLER, H. 1989. Acute fatalities following subcutaneous injection of
paramyxovirus type 1 vaccines in pigeons. Avian Pathology. 1989, Vol. 18, pp. 203-210.
KAMIKURA, K., NISHIMURA, H. and BAILEY, J.R. 1995. Blockade of beta-adrenoceptor in control of
blood pressure in fowl. American Journal of Physiology. October, 1995, Vol. 269, 4 Pt 2, pp. R 914-922.
KARSTAD, L. 1971. Angiopathy and cardiopathy in wild waterfowl from ingestion of lead shot. Conn
Med. 1971, Vol. 35, pp. 355-360.
KEENE, B.W. and FLAMMER, K. 1991. ECG of the month. Journal of the American Veterinary Medical
Association. 1991, Vol. 198, pp. 408-409.
KENDAL, E.H. 2002. Clinical Chemistry of Companion Avian Species: A Review. Vet Clin Pathol. 2002, Vol.
31, 3, pp. 140-151.
KENNETH, R.W., LIGHTFOOT, T. and REAVILL, D. 2011. Cardiovasculair disease in aging birds. [book
auth.] Association of avian veterinarians. Proceeding of the Annual Association of avian veterinarians
conference. Seattle : s.n., 2011, pp. 11-15.
KIAMA, S.G., et al. 1998. A scanning electron microscope study of the luminal surface specializations in
the blood vessels of the pecten oculi in a diurnal bird, the black kite (Milvus migrans). Annals of
anatomy. 1998, Vol. 180, pp. 455-460.
KIM, C.S., DOVE, A.A and GWATHMEY, J.K. 2000. Intracellular calcium and the relationship to
contractility in an avian model of heart failure. Journal of Comparative Physiology B. 2000, Vol. 170, 4,
pp. 295-306.
KIM, C.S., et al. 1999. Effects of Pranidipine, a Calcium Channel Antagonist, in an Avian Model of Heart
Failure - C.S. Kim. [ed.] Fluwer Academic Publisher. Cardiovascular Drugs and Therapy. 1999, Vol. 13, pp.
455-463.
KING, A.S. and McLELLAND, J. 1984. Cardiovascular system. Birds : their structure and function 2nd
Edition. Philadelphia : Bailliere Tindall, 1984, pp. 214-228.
KLASING, K.C. 2008. Nutritional diseases. [book auth.] Y.M. SAIF. Diseases of poultry 12th edition. Ames,
Iowa : Blackwell publishing, 2008, pp. 1121-1148.
KNAFO, E., et al. 2011. Cardiomyopathy and Right-sided Congestive Heart Failure in a Red-tailed Hawk
(Buteo jamaicensis). Journal of Avian Medicine and Surgery. 2011, Vol. 25, 1, pp. 32-39.

Page | 166
KOCSIS, J.F., MCILROY, P.J. and CARSIA, R.V. 1995. Atrial natriuretic peptide stimulates aldosterone
production by turkey (Meleagris gallopavo) adrenal steriodogenic cell. Gen. Comp. Endocrinol. 1995, Vol.
99, pp. 364-372.
KOSTELANETZ, S., DICKENS, M.J. and ROMERO, L.M. 2009. Combined effects of molt and chronic stress
on heart rate, heart rate variability, and glucocorticoid physiology in European Starlings. Comparative
biochemistry and physiology. 2009, Vol. 154, pp. 493-501.
KRAUTWALD-JUNGHANNS, M.E., et al. 2004. Research on the anatomy and pathology of the psittacine
heart. Journal of Avian Medicine and Surgery. March, 2004, Vol. 18, 1, pp. 2-11.
KRAUTWALD-JUNGHANNS, M.E., PEES, M. and SCHUTTERLE, N. 2002. Echocardiographic examinations
in unsedated racing pigeons (Columbia livia forma domestica) with special consideration for the physical
training. Berl Munch Tierarztl Wochenschr. 2002, Vol. 115, 5-6, pp. 221-224.
KRAUTWALD-JUNGHANNS, M.E., SCHULZ, M. and HAGNER, D. et al. 1995. Transcoelomic two-
dimensional echocardiography in the avian patient. Journal of Avian Medicine and Surgery. 1995, Vol. 9,
pp. 19-30.
KRISTA, L.M. et al. 1970. Comparison of electrocardiograms of hypertensive and hypotensive male
turkeys. Poultry Science. 1970, 49, pp. 700-703.
KRISTA, L.M., WAIBEL, P.E. and BURGER, R.E. 1965. The influence of dietary alterations, hormones, and
blood pressure on the incidence of dissecting aneurysm in turkeys. Poultry Science. 1965, Vol. 44, pp.
15-22.
KUO, A.Y., et al. 2001. Differential cardiovascular effects of pharmacological agents in chickens selected
for high and low body weight. Physiology & Behavior. 2001, Vol. 74, pp. 573-579.
LABONDE, J. 1995. Toxicity in pet avian patients. Seminars in Avian and Exotic Pet Medicine. January,
1995, Vol. 4, 1, pp. 23-31.
LAI, H., et al. 2011. Isolation and characterization of emerging subgroup J avian leukosis virus associated
with hemangioma in chickens. Veterinary Microbiology. 2011, Vol. 151, pp. 275-283.
LANGELIER, K.M., et al. 1991. Lead poisoning in bald eagles in British Columbia. Canadian Veterinary
Journal. 1991, Vol. 32, pp. 108-109.
LAROCHELLE, D., MORIN, M. and BERNIER, G. 1992. Sudden death in turkeys with perirenal
hemorrhage: pathological observations and possible pathogenesis of the disease. Avian Diseases. Jan-
Mar, 1992, Vol. 36, 1, pp. 114-124.
LATIMER, K.S. 1994. Oncology. [book auth.] B. RITCHIE, G. HARRISON and L. HARRISON. Avian medicine :
principles and applications. Lake Worth (FL) : Wingers Publishing, 1994, pp. 640-672.
LATIMER, K.S., PERRY, R.W. and MO, I.P. 1990. Myocardial sarcocystis in a grand Eclectus parrot
(Eclectus roratus) and a Moluccan cockatoo (Cacatua moluccensis). Avian Diseases. 1990, Vol. 34, 1, pp.
501-505.
LEMON, M.J., PACK, L.A. and FORZAN, M.J. 2012. Valvular endocarditis and septic thrombosis
associated with a radial fracture in a red-tailed hawk (Buteo jamaicensis). Canadian Veterinary Journal.
2012, Vol. 53, pp. 79-82.

Page | 167
LEWIS, J.C. and KOTTKE, B.A. 1997. Endothelial damage and thrombocyte adhesion in pigeon
atherosclerosis. Science. 1997, Vol. 196, pp. 1007-1009.
LEWIS, J.C., STORM, J. and GREENWOOD, A.G. 1993. Treatment of wing tip oedema in raptors. Vet Rec.
Septembre, 1993, Vol. 133, 13, p. 328.
LI, K., et al. 2006. Increased Calcium Deposits and Decreased Ca2+-ATPase in Right Ventricular
Myocardium of Ascitic Broiler Chickens. Journal of Veterinary Medicine. 2006, Vol. A53, pp. 458-463.
LICHTENBERGER, M. and KO, J. 2007. Critical Care Monitoring. Veterinary Clinics - Exotic Animal
Practice. 2007, Vol. 10, pp. 317-344.
LICHTENBERGER, M. and OROSZ, S.E. 2011. The avian cardiovascular system in C.R. Congrès ''11th EAAV
Conference''. Proc Annu Europ Assoc Avian Vet. Madrid : s.n., 2011, p. 319.
LICHTENBERGER, M. 2005. Determination of indirect blood pressure in the companion bird. Seminar
Avian Exotic Pet Medicine. 2005, Vol. 14, pp. 149-152.
—. 2004. Principles of Shock and Fluid Therapy in Special Species. Seminars in Avian and Exotic Species.
July 2004, Vol. 13, 3, pp. 142-153.
—. 2005. Shock, fluid therapy and CPCR for the avian patient. [book auth.] Association of the Avian
Veterinarian. Proceeding of the Association of the Avian Veterinarian Conference. Arles : s.n., 2005, pp.
374-387.
LICHTENBERGER, M.L., CHAVEZ, W. and BRUNSEN, D. et al. 2004. Direct versus indirect blood pressure
monitoring during acute blood loss in pecking ducks. [book auth.] Association of Avian Veterinarians.
Scientific proceedings, 25th Annual Conference and Expo. New Orleans, Louisiana : s.n., 2004, p. 3.
LIERZ, M. and KORBEL, R. 2012. Anesthezia and analgesia in birds. Journal of Exotic Pet Medicine. 2012,
Vol. 21, pp. 44-56.
LIGHTFOOT, T.L. and YEAGER, J.M. 2008. Pet bird toxicity and related environmental concerns.
Veterinary Clinics Exotic Animal Practice. 2008, Vol. 11, pp. 229-259.
LU, Y., et al. 1993. Cardiac conduction system in the chicken: gross anatomy plus light and electron
microscopy. Anat rec. July, 1993, Vol. 236, 3, pp. 493-510.
LU, Y., et al. 1993. Histological organization of the right and left atrioventricular valves of the chicken
heart and their relationship to the atrioventricular Purkinje ring and the middle bundle branch. Anatomy
record. January, 1993, Vol. 235, 1, pp. 74-86.
LUCITTI, J.L. and HEDRICK, M.S. 2006. Characterization of baroreflex gain in the domestic pigeon
(Columba livia). Comparative biochemistry and physiology. 2006, Vol. 143, pp. 103-111.
LUMEIJ, J.T. and B.W., RITCHIE. 1994. Cardiology. [book auth.] B.W. RITCHIE, G.J. HARRISON and L.R.
HARRISON. Avian medicine: Principles and application. Lake Worth (FL) : Wingers Publishing, 1994, pp.
695-722.
LUMEIJ, J.Y. and STOKHOF, A.A. 1985. Electrocardiogram of the racing pigeon (Columbia livia
domestica). Res Vet Sci. 1985, Vol. 38, pp. 275-278.
MADDISON, J. and CHURCH, D. 2008. Small animal clinical phamacology. s.l. : Saunders, 2008.

Page | 168
MAINA, J.N. 1996. Perspectives on the structure and function of birds. [book auth.] W. ROSSKOPF and R.
WOERPEL. Diseases of cage and aviary birds. 3rd edition. Baltimore : William & Willkins, 1996, pp. 163-
217.
MAJO, N. and DOLZ, R. 2012. Autopsie des volailles. RUEIL-MALMAISON : Les éditions du Point
vétérinaire, 2012. 978-2-86326-314-3.
MANS, C. and BROWN, C.J. 2007. Radiographic evidence of atherosclerosis of the descending aorta in a
Grey-cheeked parakeet (Brotogeris pyrrhopterus). Journal of Avian Medicine and Surgery. March, 2007,
Vol. 21, 1, pp. 56-62.
MARRUCHELLA, G., et al. 2011. Granulomatous myocarditis caused by candida albicans in a canary
(Serinus canaria). Journal of Avian Medicine and Surgery. 2011, Vol. 25, 3, pp. 205-209.
MARTIN, K. 2006. Psittacine behavioral pharmacotherapy. [book auth.] A. LUESCHER. Manual of parrot
behavior. Ames, IA : Blackwell Publishing, 2006, pp. 267-279.
MAUTINO, M. 1994. Lead and Zinc Intoxication in Zoological Medicine: A Review. Journal of Zoo and
Wildlife Medicine. March, 1994, Vol. 28, 1, pp. 28-35.
MAXWELL, M.H., ROBERTSON, G.W. and MOSELEY, D. 1986. Studies on an ascitic syndrome in young
broilers. 1. Haematology and pathology. Avian pathology. 1986, Vol. 15, pp. 511-524.
MCKENZIE, B., WILL, E.J. and HARDIEN, A. 1971. The electrocardiogram of turkey. Avian diseases. 1971,
Vol. 15, pp. 737-744.
MCKENZIE, B.E. and WILL, J.A. 1972. Electrocardiographic changes following influenza infection in
turkeys. Avian diseases. 1972, Vol. 16, pp. 308-318.
MCMILLAN, M. 1994. Imaging techniques. [book auth.] B.W. RITCHIE, G.J. HARRISON and L.R.
HARRISON. Avian medicine principles and application. Lake Worth, Fla : Wingers Publishing, 1994, pp.
246-326.
McSHERRY, B.J., FERGUSON, A.E. and BALLANTYNE, J. 1954. A dissecting aneurism in internal
hemorrhage in turkeys. Journal of the American Veterinary Medical Association. 1954, Vol. 124, pp. 279-
283.
MICKLEY, K., et al. 2009. Ovarian hemangiosarcoma in an Orange-winged Amazon parrot (Amazona
amazonica). Journal of avian medicine and surgery. 2009, Vol. 23, pp. 29-35.
MILLER, L.E. et al. 1985. Effects of caponization on body weight, atherosclerosis, and cardiovascular
variables in hypertensive and hypotensive turkey lines. Poultry Science. 1985, Vol. 64, 5, pp. 1002-1014.
MILLER, M. 1986. Electrocardiography. [book auth.] G.J. HARRISON and L.R. HARRISON. Clinical avian
medicine and surgery. Philadelphia, PA : Saunders, 1986, pp. 286-292.
MIRSALIMI, S.F., et al. 1990. Myocardial Biochemical Changes in Furazolidone-induced Cardiomyopathy
of Turkeys. Journal of comparative pathology. 1990, Vol. 102, pp. 139-147.
MIRSALIMI, S.M., O'BRIEN, P.J. and JULIAN, R.J. 1992. Blood volume in salt-induced pulmonary
hypertension heart failure and ascites in broiler and White Leghorn chickens. Can J Vet Res. 1992, Vol.
57, pp. 110-113.

Page | 169
—. 1993. Blood volume increase in salt-induced pulmonary hypertension, heart failure and ascites in
broiler and White Leghorn chickens. Avian Diseases. 1993, Vol. 53, pp. 2353-2369.
MITCHELL, B.W. and BRUGH, M. 1982. Comparison of electrocardiograms of chickens infected with
viscerotropic velogenic Newcastle disease virus and virulent avian influenza virus. American journal of
veterinary research. 1982, Vol. 43, pp. 2274-2278.
MITCHELL, E.B., HAWKINS, M.G. : ORVALHO, J.S. and THOMAS, W.P. 2008. Congenital mitral stenosis,
subvalvular aortic stenosis, and congestive heart failure in a duck. Journal of Veterinary cardiology.
2008, Vol. 10, pp. 67-73.
MOHANTY, G.C. and WEST, J.L. 1969. Pathologic features of experimental sodium chloride poisoning in
chicks. Avian Diseases. 1969, Vol. 13, pp. 762-773.
MÖHRING, J. SCHOUN, J. SIMON-OPPERTMAN, C. and SIMON, E. 1980. Radioimmunoassay for
arginine-vasotocin (AVT) in serum of Pekin Ducks: AVT concentrations after adaptation to freshwater
and salt water. Pflügers Arch/. 1980, Vol. 387, pp. 91-97.
MOORMAN, A.F., et al. 1998. Development of the cardiac conduction system. Circ Res. 1998, Vol. 82,
pp. 629-644.
MUELLER, R.W., RAPLEY, W.A. and K.G., MEHREN. 1980. Pathology of thyroid disease and
atheriosclerosis in captive wild birds. [book auth.] R.J. MONTALI. Comparative pathology of zoo animals.
Washington D.C. : Smithsonian Institution Press, 1980, pp. 523-537.
MUNDAY, B.L., et al. 1979. Sarcocystis and related organisms in autralian wildlife II : Survey findings in
birds, reptiles, amphibians and fish. Journal of Wildlife Diseases . January, 1979, Vol. 15, 1, pp. 57-73.
MUTALIB, A.A. and HANSON, J.A. 1990. Sudden death in turkeys with perirenal hemorrhage : Field and
laboratory findings. Canadian Veterinary Journal. September, 1990, Vol. 31, pp. 637-642.
NABIPOUR, A. and TAHOONGHI, M. 2008. Histological Study on the Sinu-Atrial Node of Turkey. World
Journal of Zoology. 2008, Vol. 3, 1, pp. 8-12.
NAGARA, R.Y., et al. 1996. Acute cardiotoxicity of moniliformin in broiler chickens as measured by
electrocardiography. Avian diseases. 1996, Vol. 40, pp. 223-227.
NAIN, S., et al. 2008. The role of oxidative stress in the development of congestive heart failure in a
chicken genotype selected for rapid growth. Avian pathology. 2008, Vol. 37, 4, pp. 367-373.
NAKAMURA, K., ABE, F. and TANIGUCHI, T. 1988. Myocardial cytoplasmic inclusions in chickens with
haemangioma and lymphoid leukosis. Avian pathology. 1988, Vol. 17, 1, pp. 3-10.
NAKANO, Y. and UNE, Y. 2012. Hemangiosarcoma with widespread metastasis that originated on the
metatarsal pad of a java sparrow (Padda oryzivora). Journal of Veterinary Medical Science. 2012, Vol. 74,
5, pp. 621-623.
NAP, A.M. and LUMEIJ, J.T. 1991. Cardiology in Birds. Tijdschr Diergeneeskd. 1991, Vol. 116, 1, pp. 83S-
84S.
NAP, A.M.P., LUMEIJ, J.T. and STOKHOF, A.A. 1992. Electrocardiogram of the African grey (psittacus
erithacus) and Amazon (Amazona spp.) parrot. Avian pathology. 1992, Vol. 21, 1, pp. 5-53.

Page | 170
NEGRO, J.J., et al. 2006. Function and occurence of facial flushing in birds. Comparative biochemistry
and physiology, Part A. 2006, Vol. 143, pp. 78-84.
NEPHEW, B.C., et al. 2005. A potential cardiovascular mechanism for the behavioral effects of central
and peripheral arginine vasotocin. General and comparative pathology. 2005, Vol. 144, pp. 156-166.
NEUMANN, F., et al. 1973. Spontenous aortic rupture in turkeys and the vascularization of the aortic
wall. Can Vet J. 1973, Vol. 14, pp. 136-138.
NICHOLLS, P.K., et al. 1995. Parasitic infection in a flock of rufous-crested bustards (eupoditis ruficrista)
in the united arab emirates. Journal of Zoo and Wildlife Medicine. 1995, Vol. 26, 4, pp. 590-596.
NICKEL, R., SCHUMMER, A. and SEIFERLE, E. 1977. Anatomy of the domestic birds. Berlin : Verlag Paul
Parey, 1977. 3-489-55418-3.
NOVILLA, M.N. and CARPENTER, J.W. 2004. Pathology and pathogenesis of disseminated visceral
coccidiosis in cranes. Avian patholgy. June 2004, Vol. 33, 3, pp. 275-280.
O’BRIEN, P.J., O'GRADY, M. and LUMSDEN, J.H. 1993. Clinical pathologic profiles of dogs and turkeys
with congestive heart failure, either non-induced or induced by rapid ventricular pacing,. American
Journal of Veterinary Research. 1993, Vol. 54, 1, pp. 60-68.
O'BRIEN, V.A., et al. 2010. Prevalence and Pathology of West Nile Virus in Naturally Infected House
Sparrows, Western Nebraska, 2008. The american journal of tropical medicine and hygiene. 2010, Vol.
85, 2, pp. 937-944.
OGLESBEE, B.L. and LEHMKUHL, L. 2001. Congestive heart failure associated with myxomatous
degeneration of the left atrioventricular valve in a parakeet. Journal of American Veterinary Medical
Association. February, 2001, Vol. 218, 3, pp. 376-380.
OGLESBEE, B.L. and OGLESBEE, M.J. 1998. Results of post-mortem examination of psittacine birds with
cardiac disease : 26 cases (1991-1995). Journal of the American Veterinary Medical Association. 1998,
212, pp. 1737-1742.
OGLESBEE, B.L., et al. 2001. Electrocardiographic reference values for Macaws (Ara species) and
Cockatoos (Cacatua species). Journal of Avian Medicine and Surgery. March, 2001, Vol. 15, 1, pp. 17-22.
OLKOWSKI, A.A. 2007. Pathophysiology of Heart Failure in Broiler Chickens Structural, Biochemical, and
Molecular Characteristics. Poultry Science. 2007, Vol. 86, pp. 999-1005.
OLKOWSKI, A.A., CLASSEN, H.L. and KUMOR, L. 1998. Left atrio-ventricular valve degeneration, left
ventricular dilation and right ventricular failure : A possible association with pulmonary hypertension
and aetiology of ascites in broiler chickens. Avian pathology. 1998, Vol. 27, pp. 51-59.
OLKOWSKI, A.A., et al. 2003. Lesions of the pericardium and their significance in the aetiology of heart
failure in broiler chickens. Research in Veterinary Science. 2003, Vol. 74, pp. 203-211.
O'MALLEY, B. 2005. Clinical anatomy and physiology of Exotic species. Germany : Elsevier Saunders,
2005. pp. 113-116. 0 7020 2782 0.
ONDERKA, D.K. and BHATNAGAR, R. 1982. Ultrastructural changes of sodium chloride-induced
cardiomyopathy in turkey poults. Avian Diseases. 1982, Vol. 26, pp. 835-841.

Page | 171
PAGE, C.D., SCHMIDT, R.E. and HUBBARD, G.B. et al. 1989. Sarcocystis myocarditis in a red lory (Eos
bornea). J Zoo Wildl Med. 1989, Vol. 20, pp. 461-464.
PALMIERI, C., et al. 2011. Pathology and Immunohistochemical Findings of West Nile Virus Infection in
Psittaciformes. Veterinary pathology. 2011, Vol. 48, 5, pp. 975-984.
PARTO, P. 2010. Macroscopic and microscopic studies of sino-atrial node in the heart of ostrich (stuthio
camelus). World Journal of Zoology. 2010, Vol. 5, 2, pp. 90-95.
—. 2012. The Structure of the Atrioventricular Node in the Heart of the Female Laying Ostrich (Struthio
camelus). Anat. Histol. Embryol. 2012, Vol. 41, pp. 75-78.
PAUL, J.L. and BAUDIN, B. 2009. Pathophysiology of atherosclerosis and markers for early detention.
Revue francophone des laboratoires. 2009, Vol. 39, 409, pp. 41-50.
PAULETTO, P., et al. Evidence of reduced atherosclerotic lesions in broad breasted white turkeys treated
with oxprenolol. Artery. 12, pp. 220-233.
PEES, M. and KRAUTWALD-JUNGHANNS, M.E. 2009. Cardiovascular physiology and diseases of pet
birds. Veterinary clinic of North America - Exotic Animal Practice. January 2009, Vol. 12, 1, pp. 81-97.
PEES, M. 2005. Avian echocardiography. Seminars in Avian and Exotic Pet Medicine. January, 2005, Vol.
14, 1, pp. 14-21.
PEES, M., KRAUTWALD-JUNGHANNS, M-E. and STRAUB, J. 2006. Evaluating and treating the
cardiovascular system. [book auth.] B.W. RITCHIE, G.J. HARRISON and L.R. HARRISON. Clinical avian
medicine. Palm beach, FL : Spix Publishing, 2006, pp. 379-394.
PEES, M., KUHRING, K. and KRAUTWALD-JUNGHANNS, M.E. 2005. The use of enalapril in birds :
indications, clinical experiences, pharmakokinetics and possible side effects. Arles : European
Association of the Avian Veterinarians, 2005.
PEES, M., SCHMIDT, V. and COLES, B. et al. 2006. Diagnosis and long-term therapy of right-sided heart
failure in a yellow-crowned amazon (Amazona ochrocephala). Veterinary Records. 2006, Vol. 158, 13,
pp. 445-447.
PEES, M., STRAUB, J. and KRAUTWALD-JUNGHANNS, M.E. 2004. Echocardiographic examinations of 60
African grey parrots and 30 other psittacine birds. Veterinary Records. 2004, Vol. 155, 3, pp. 73-76.
—. 2001. Insufficiency of muscular atrioventricular valve in the heart of a blue-fronted amazon
(Amazona aestiva aestiva). Veterinary Record. April, 2001, Vol. 148, 17, pp. 540-543.
PENCE, P.A. 1996. Developmental abnormalities. [book auth.] W.J. ROSSKOPF and R.W. WOERPEL.
Diseases of cage and aviary birds. Baltimore, MD : Williams and Wilkins, 1996, pp. 464-469.
PETERS, B.G. 1936. Paronchocerca ciconiarum n. g. sp. from the saddle-billed stork in West Africa. J
Helminthol. 1936, Vol. 14, pp. 1-10.
PETRAK, L.M. and GILMORE, C.E. 1982. Neoplasms. Diseases of Cage and aviary birds 2nd ed. 1982, pp.
606-637.
PHALEN, D.N. and DAHLHAUSEN, B. 2004. West Nile Virus. Sem Avian Exotic Pet Med. 2004, Vol. 13, pp.
67-78.

Page | 172
PHALEN, D.N., et al. 1996. Heart failure in a macaw with atherosclerosis of the aorta and
brachiocephalic arteries. Journal of American Veterinary Medical Association. 1996, Vol. 209, 8, pp.
1435-1440.
PHILILIPS, I.R. 1986. Parrots encountered in practice : a survey of one hundred and twelve cases. Journal
of Small Animal Practice. 1986, Vol. 42, pp. 523-535.
PILNY, A.A. 2004. Retrospective of Atherosclerosis in Psittacine Birds: Clinical and Histopathologic
Findings in 31 Cases. Proceeding of the AAV conference. 2004, pp. 349-351.
POWELL, F.L., HASTINGS, R.H. and MAZZONE, R.W. 1985. Pulmonary vascular resistance during
unilateral pulmonary arterial occlusion in ducks. American journal of Physiology. 1985, 249, pp. 39-43.
PRAKASH, R. 1956. The heart and its conducting system in the common Indian fowl. Proc. Nat. Inst. Sci.
India. 1956, Vol. 22, pp. 22-27.
PREZOTO, B.C., et al. 2009. Cardioprotective effect of ornitho-kinin in an anesthetized, open-chest
chicken model of acute coronary occlusion. Brazilian journal of medical and biological research.
September 2009, Vol. 42, 9, pp. 824-830.
PRITCHARD, W.R., HENDERSON, W. and BEALL, C.W. 1958. Experimental production of dissecting
aneurysms in turkey. American Journal of Veterinary Research. 1958, Vol. 19, pp. 696-705.
QAYYUM, M.A. and SHAAD, F.U. 1976. Anatomical and neurohistological observations on the heart of
the rose ringed parakeet, Psittacula krameri. Anat Anz. 1976, Vol. 140, 1-2, pp. 42-51.
RAE, M. 1995. Hemoprotozoa of Caged and Aviary Birds. Seminars in Avian and Exotic Pet Medicine. uly,
1995, Vol. 4, 3, pp. 131-137.
RAE, M.A. 2003. Practical avian necropsy. Seminars in Avian and Exotic Pet Medicine. April, 2003, Vol.
12, 2, pp. 62-70.
RANDALL, C.J. and REECE, R.L. 1996. Color atlas of avian histopathology . London : Mosby Wolfe
Publishing, 1996. 0-7234-2087-4.
RANDOLPH, J.F., MOISE, N.S. and GRAHAM, D.L. MURPHY, J. 1984. Bacterial endocarditis and
thromboembolism of a pelvic limb in an emu. JAVMA. December, 1984, Vol. 185, 11, pp. 1409-1410.
RATCLIFFE, H.L. and CRONIN, M.T.I. 1958. Changing frequency of arteriosclerosis in mammals and birds
at the Philadelphia Zoological Garden. Circulation. 1958, Vol. 18, pp. 41-52.
RATCLIFFE, H.L. 1966. Arterial lesions of zoo birds : response to environmental factors. Acta Zoologie et
Pathologie Antverpens. 1966, Vol. 39, pp. 3-26.
RAYMOND, J.T., et al. 2001. Tyzzer's Disease in a Neonatal Rainbow Lorikeet (Trichoglossus
haematodus). Veterinary pathology. 2001, 38, pp. 326-327.
REAVILL, D. and SCHMIDT, R. 2009. Mycobacterial granulomatous arteritis in pet birds. Proceeding of
the Association of Avian Veterinarians Conference. 2009, pp. 63-64.
REAVILL, D.R. and DORRESTEIN, G.M. 2010. Pathology of aging psittacines. Veterinary clinics of North
America : Exotic Animal Practice. 2010, Vol. 13, 1, pp. 135-150.

Page | 173
REAVILL, D.R. 2001. Pet bird oncology. [book auth.] Association of Avian Veterinarians. Proceedings
Association of Avian Veterinarians, Avian Speciality advanced program. Orlando, FL : s.n., 2001, pp. 29-
43.
—. 2004. Tumor of pet birds. Veterinary clinics Exotic Animal Practice. 2004, Vol. 7, pp. 537-560.
REDIG, P.T. and ARENT, L.R. 2008. Raptor toxicology. Vet Clin Exot Anim. 2008, Vol. 1, pp. 261-282.
REECE, L.R. 1996. Some observations on naturally occuring neoplasms of domstic fowl in the State of
Victoria, Australia (1977-87). Avian pathology. 1996, Vol. 25, 3, pp. 407-447.
REECE, R.L. 2007. Observations on naturally occuring neoplasms in birds in the state of Victoria,
Australia. Avian pathology. 2007, Vol. 21, 1, pp. 3-32.
REMBERT, M.S., et al. 2008. Intermittent bradyarrythmia in a hispaniolan amazon parrot (amazona
ventralis). Journal of Avian Medicine and Surgery. March, 2008, Vol. 22, 1, pp. 31-40.
RETTMER, H., et al. 2011. Radiographic measurement of internal organs in Spix's Macaws (Cyanopsitta
spixii). Journal of Avian Medicine and Surgery. 2011, Vol. 25, 4, pp. 254-258.
REZAKHANI, A., et al. 2007. A preliminary study on normal electrocardiographic parameters of ostriches
(Struthio camelus). Journal of the South African Veterinary Association. 2007, Vol. 78, 1, pp. 46-48.
RIDDELL, C. 1997. Developmental, metabolic, and other non-infectious disorders. [book auth.] B.
CALNEK, H. BARNES and C. et al. editors. BEARD. Diseases of poultry. 10th edition. Ames (IA) : Iowa State
University Press, 1997, pp. 19-950.
RISI, E., et al. 2011. Cardiomyopathy on a griffon vulture (gyps fulvus). In 11th Proc Annu Conf Europ
Assoc Avian Vet. Madrid : s.n., 2011, pp. 244-249.
RITCHIE, B.W. and CARTER, K. 1995. Avian viruses. Lake Worth, Fl : Wingers Publishing, 1995.
RITCHIE, B.W. and HARRISON, G.J. 1994. Formulary. [book auth.] B.W. RICHIE, G.J. HARRISON and L.R.
HARRISON. Avian medicine : Principles and application. Lake Worth, FL : Wingers Publishing, 1994, pp.
457-477.
RITCHIE, B.W., HARRISON, G.J. and HARRISON, L.R. 1994. Avian medicine : principles and application.
Lake worth : Wingers Publishing Inc, 1994. 0-9636996-0-1.
RODRIGUEZ, R., PRIETO-MONTANA, F. and MONTES, A.M. 2004. The normal electrocardiogram of the
unanesthetized peregrine falcon (Falco peregrinus brookei). Avian Diseases. 2004, Vol. 48, pp. 405-409.
ROMAGNANO, A., SHIROMA, J.T. and HEARD, D.J., et al. 1996. Magnetic Resonance imaging of the
brain and coelomic cavity of the domestic pigeon (Columbia livia domestica). Vet Radiol Ultrasound.
1996, Vol. 37, 6.
ROSENTHAL, K. and STAMOULIS, M. 1993. Diagnosis of congestive heart failure in an Indian Hill Mynah
(Gracula religiosa). Journal of the Association of Avian Veterinarians. 1993, Vol. 7, pp. 27-30.
ROSENTHAL, K., et al. 1997. The cardiovascular system. [book auth.] R.B. ALTMAN, et al. Avian medicine
and surgery. Philadelphia : WB Saunders, 1997, pp. 489-500.

Page | 174
ROSSI, G. 1998. A poorly-differenciated haemangiosarcoma in an Amazon farinosa parrot. Avian
pathology. 1998, Vol. 27, 4, p. 427.
ROSSI, G., et al. 2005. Bio-molecular study of a progression of haemochromatosis in hill mynahs
(Gracula religiosa). [book auth.] European Association of Avian Veterinarians. Proceeding of the
Conference of the European Association of Avian Veterinarians. Arles : EAAV, 2005, pp. 141-147.
RUIZ-FERIA, C.A., et al. 2004. Age- and sex-dependent changes in pulse pressure in fowl aorta.
Comparative Biochemistry and Physiology Part A. 2004, Vol. 137, pp. 311–320.
SAINT CLAIR, R.W. 1998. The contribution of Avian Models to our understanding of atherosclerosis and
their promise for the future. Laboratory animal science. 1998, Vol. 48, 6, pp. 565-568.
SAITO, E.K., et al. 2007. Raptor mortality due to West Nile Virus in the United States, 2002. Journal of
Wildlife diseases. 2007, Vol. 43, 2, pp. 206-213.
SANDERS, E., DE GROOT, I.J. and GEERTS, W.J. et al. 1986. The local expression of adult chicken heart
myosins during development. II. Ventricular conducting tissue. Anat Embryol (Berl). 1986, Vol. 174, pp.
187-193.
SARABIA, J., et al. 2008. Lesions associated with the plexus venosus subcutaneus collaris of pigeon with
chlorophacinon toxicosis. Avian Diseases. September, 2008, Vol. 52, 3, pp. 540-543.
SATO, Y., AOYAGI, T. and MIYANO, N. 2000. An occurence of cardiac rupture in a capercaillie (Tetrao
urogallus) probably induced by myocardial infarction. Avian diseases. 2000, Vol. 44, pp. 231-233.
SAVAGE, J.E., et al. 1966. Comparison of copper deficiency and lathyrism in turkey poults. The Journal of
Nutrition. January, 1966, Vol. 88, 1, pp. 15-25.
SAVORY, C.J. and KOSTAL, L. 1997. Application of a radiotelemetry system for Chronic Measurement of
Blood Pressure, Heart Rate, EEG, and Activity in the Chicken. Physiology & Behavior. 1997, Vol. 61, 6, pp.
963–969.
SAWBY, S.W. and GESSAMAN, J.A. 1974. Telemetry of electrocardiograms from free-living birds: a
method of electrode placement. Condor. 1974, Vol. 76, pp. 479-481.
SCHENKER, O.A. and HOOP, R.K. 2007. Chlamydiae and atherosclerosis: can psittacine cases support the
link? Avian Diseases. 2007, Vol. 51, 1, pp. 8-13.
SCHMIDT, R.E., REAVILL, D.R. and PHALLEN, D.N. 2003. Pathology of pet and aviary birds. Ames : Iowa
State Press, 2003. pp. 3-16. 0-8138-0502-3.
SCHNELLBACHER, R.W., et al. 2012. Effects of dopamine and dobutamine on isoflurane-induced
hypotension in Hispaniolan Amazone parrot (Amazona ventralis). American Journal of Veterinary
Research. July, 2012, Vol. 73, 7, pp. 952-958.
SCHOEMAKER, N.J. and ZANDVLIET, M.J.M. 2005. Electrocardiograms in selected species. Seminars in
Avian and Exotic Pet Medicine. January, 2005, Vol. 14, 1, pp. 26-33.
SCHUTZ, H. and GERSTBERGER, R. 1990. Atrial natriuretic factor controls salt gland secretion in the
Pekin duck (Anas platyrhynchos) through interaction with high affinity receptors. Endocrinology. 1990,
Vol. 127, pp. 1718-1726.

Page | 175
SCHUTZ, H., GRAY, D.A. and GERSTBERGER, R. 1992. Modulation of kidney function in counscious Pekin
Ducks by atrial natriuretic factor. Endocrinology. 1992, Vol. 130, pp. 678-684.
SCOTT, T.M. 1971. The ultrastructure of ordinary and Purkinje cells of the fowl heart. J. Anat. 1971, Vol.
110, 2, pp. 259-273.
SEDACCA, C.D., CAMPBELL, T.W. and BRIGHT, J.M. et al. 2009. Chronic cor pulmonale secondary to
pulmonary atherosclerosis in an African Grey Parrot. Journal of American Veterinary Medicine
Association. 2009, Vol. 234, 8, pp. 1055-1059.
SEKIZAKI, T., et al. 2008. Endocarditis in chickens caused by subclinical infection of Streptococcus
gallolyticus subsp. gallolyticus. Avian diseases. March, 2008, Vol. 52, 1, pp. 183-186.
Service, U.S. Fish and Wildlife. 1986. Final supplemental environnemental impact statement on the use
of lead shot for hunting migratory birds in the united states. Fish and Wildlife service, Washington
DC:US. Publication no. FES 86-16, 1986.
SHANER, R.F. 1923. On the muscular anatomy of the Vertebrate ventricle. Journal of Anatomy. 1923, 58,
pp. 59-69.
SHARMA, D., et al. 2012. Toxic interaction between fumonisin B1 and Moniliformin for cardiac lesions in
japanese quail. Avian Diseases. September, 2012, Vol. 56, 3, pp. 545-554.
SHATHELE, M.S., FADLELMULA, A. and AL-HIZAB, F.A. et ZAKI, M.M. 2009. Fatal aspergillosis in an
Ostrich (Struthio camelus) predisposed by pulmonary haemangioma in the Kingdom of Saudi Arabia.
International journal of zoological research. 2009, Vol. 5, 2, pp. 80-85.
SHIVAPRASAD, H.L., CRESPO, R. and PUSCHNER, B. 2004. Coronary artery rupture in male commercial
tukey. Avian pathology. 2004, Vol. 33, pp. 226-232.
SHIVAPRASAD, H.L., et al. 2004. Circovirus infection in a Gouldian finch (Chloebia gouldiae). Avian
pathology. 2004, Vol. 33, 5, pp. 525-529.
SHIVAPRASAD, H.L., et al. 2009. Myocarditis associated with reovirus in turkey poults. Avian diseases.
December, 2009, Vol. 53, 4, pp. 523-532.
SHIVAPRASAD, H.L., et al. 2002. Turlock-like bunyavirus associated with encephalomyelitis and
myocarditis in ostrich chick. Journal of Veterinary Diagnostic Investigation. 2002, Vol. 14, 363, pp. 363-
370.
SHRUBSOLE-COCKWILL, A., WOJNAROWICZ, C. and PARKER, D. 2008. Atherosclerosis and ischemic
cardiomyopathy in a captive, adult red-tailed hawk (Buteo jamaicensis). Avian diseases. September,
2008, Vol. 52, 3, pp. 537-539.
SILEO, L., JONES, R.N. and HATCH, R.C. 1973. The effect of ingested lead shot on the electrocardiogram
of Canada geese. Avian diseases. 1973, Vol. 17, pp. 308-313.
SILLER, W.G. 1981. Renal pathology of the fowl - a review. Avian pathology. November 12, 1981, Vol. 10,
3, pp. 187-262.
—. 1958. Ventricular septal defects in the fowl. Journal of pathology and bacteriology. 1958, 76, pp.
413-440.

Page | 176
SIMONE-FREILICHER, E. 2007. Use of isoxsuprine for treatment of clinical signs associated with
presumptive atherosclerosis in a yellow-naped Amazone parrot (Amazona ochrocephala auropalliata).
Journal of Avian Medicine and Surgery. September 2007, Vol. 21, 3, pp. 215-219.
SIMPSON, V.R. 1991. Leucocytozoon-like infection in parakeets, budgerigars and a common buzzard.
Vet Rec. July, 1991, Vol. 129, 2, pp. 30-32.
SINGH, T. 1958. Histological study of the specialised tissue of the avian heart. Proc. Nat. Inst. Sci. India
24, 140–144. 1958, Vol. 24, pp. 140-144.
SLEDGE, D.G., et al. 2006. Subcutaneous haemangiosarcoma in a cockatiel (Nymphicus hollandicus).
Journal of veterinary medicine. A, physiology, pathology, clinical medicine. 2006, Vol. 53, pp. 293-295.
SMITH, B.J. and SMITH, S.A. 1997. Radiology. [book auth.] R.B. ALTMAN, et al. Avian medicine and
surgery. Philadelphia : WB Saunders, 1997, pp. 170-200.
SMITH, F.M., WEST, N.H. and JONES, D.R. 2000. The cardiovascular system. [book auth.] G.C. WHITOW.
Surkie's avian physiology. 5th edition. London : Academic Press, 2000, 9, pp. 141-232.
SMITH, S.A. 1996. Parasites of birds of prey : their diagnosis and treatment. Seminars in Avian and
Exotic Pet Medicine. April, 1996, Vol. 5, 2, pp. 97-105.
SOFFER, D., RESNICK-ROGUEL, N. and ELDOR, A. et al. 1990. Multifocal vascular tumors in fowl induced
by a newly isolated retrovirus. Cancer research. 1990, Vol. 50, 15, pp. 4787-4793.
SOLA, S.C., BORELLO, B. and CASTAGNARO, M. 2007. Occurence of cutaneous haemangiomas in
chickens: Morphological aspects. Avian pathology. 2007, Vol. 26, 3, pp. 501-510.
ŠOLC, D. 2007. The heart and heart conducting system in the kingdom of animals: A comparative
approach to its evolution. Exp Clin Cardiol. 2007, Vol. 12, 3, pp. 113-118.
SOMMER, J.R. and JOHNSSON, E.A. 1969. Cardiac muscle : A comparativ ultrastructural study with
special reference to frog and chicken hearts. Z. Zellforsch Mikroskopik Anatomie. 1969, 98, pp. 437-468.
—. 1970. Comparative ultrastructure of cardiac cell membrane specialization: A review. American
Journal of Cardiology. 1970, Vol. 25, pp. 184-194.
St LEGER, J. 2007. Avian atherosclerosis. [book auth.] M.E. FOWLER and R.E. MILLER. Medicine, Zoo and
Wild Animal, Current therapy VI. 2007.
STEDMAN, N.L., BROWN, T.P. and ROWLAND, G.N. 1998. Intravascular cartilaginous emboli in the
spinal cord of turkeys. Avian diseases. 1998, Vol. 42, pp. 423-428.
STEELE, K.E., et al. 2000. Virus infections in native and exotic birds during the 1999 outbreak in New ork
City, New York. Veterinary Pathology. 2000, Vol. 37, pp. 208-224.
STEPHAN, F. 1949. Les suppliances obtenues expérimentalement dans le système des arcs aortiques de
l'embryon d'oiseau. C.R. de la société d'anatomie. 1949, Vol. 36, p. 647.
STEPIEN, R.L. 2005. Blood pressure assessment. [book auth.] S.J. ETTINGER and E.C. FELDMAN. Textbook
of Veterinary Internal Medicine. St Louis : MO: Saunders/Elsevier, 2005, pp. 470-472.

Page | 177
STEWART, J. 1994. Ratites. [book auth.] B.W. RITCHIE, G.J. HARRISON and L.R. HARRISON. Avian
Medicine: Principles and applications. Lake Worth, Florida : Wingers Publishing, 1994.
STOUT, L.C. and THORPE, L.W. 1980. Histology of normal aortas in birds with emphasis on intimal
thickening. Atherosclerosis. 1980, 36, pp. 545-558.
STRAUB, J, PEES, M. and KRAUTWALD-JUNGHANNS, M.E. 2001. Diagnosis of pericardial effusion in
birds by ultrasound. Veterinary record. 2001, Vol. 149, pp. 86-88.
STRAUB, J., et al. 2003. Pulsed-wave Doppler echocardiography in birds of prey. Veterinary record.
2003, Vol. 153, pp. 742-746.
STRAUB, J., et al. 2004. Pulsed-wave Doppler-derivated velocity of diastolic ventricular inflow and
systolic aortic outflow in raptors. Veterinary Record. 2004, Vol. 154, pp. 145-147.
STRAUB, J., FORBES, N. and PEES, M. et al. 2003. Effet of handling-induced stress on the results of
spectral Doppler-echocardiography in falcons. Res Vet Sci. 2003, Vol. 74, pp. 119-122.
STRAUB, J., PEES, M. and ENDERS, F. 2003. Pericardiocentesis and the use of enalapril in a fischer's
lovebird. Veterinary Record. 2003, Vol. 152, pp. 24-26.
STRAUB, J., PEES, M. and KRAUTWALD-JUNGHANNS, M.E. 2002. Measurement of the cardiac
silhouette in psittacines. Journal of American Veterinary Medical Association. January, 2002, Vol. 221, 1.
STRUNK, A. 2003. Avian cardiology. Veterinary Clinics of North America: Exotique animal practice. 2003,
Vol. 1, 28, pp. 1-28.
STURKIE, P.D. 1950. Abnormal electrocardiograms of chickens produced by potassium deficiency and
effects of certain drugs on the abnormalities. American Journal of Physiology. 1950, Vol. 162, pp. 538-
544.
—. 1986. Heart : Contraction, conduction, and electrocardiography. Avian Physiology. 4th edition. New
York : Springer-Verlag, 1986, pp. 167-190.
—. 1949. The electrocardiogram of the chicken. American Journal of Veterinary Research. 1949, Vol. 10,
pp. 168-175.
STURKIE, P.D., SINGSEN, E.P. and MATTERSON, L.D. et al. 1954. The effects of dietary deficiencies of
vitamin E and the B complexe vitamins on the electrocardiogram of chickens. American Journal of
Veterinary Research. 1954, Vol. 162, pp. 457-462.
SUEDMEYER, W.K., WITTER, R.L. and BERMUDEZ, A. 2001. Hemangiosarcoma in a Golden Pheasant
(Chrysolophus pictus). Journal of Avian Medicine and Surgery. June, 2001, Vol. 15, 2, pp. 126-130.
SWAYNE, D.E., BECK, J.R. and SMITH, C.S. 2001. Fatal Encephalitis and Myocarditis in Young Domestic
Geese (Anser anser domesticus) caused by West Nile Virus. Emerging Infectious Diseases. July-August,
2001, Vol. 7, 4, pp. 751-753.
SZABO, E., VIRAGH, S. and CHALLICE, C.E. 1986. The structure of the atrioventricular conducting system
in the avian heart. Anat rec. 1986, Vol. 215, pp. 1-9.
SZABO, K.A., et al. 2004. Fatal Toxoplasmosis in a Bald Eagle (Haliaeetus leucocephalus). Journal of
parasitology. 2004, Vol. 90, 4, pp. 907-908.

Page | 178
SZABUNIEWICZ, M. and MCCRADY, J.D. 1974. Some aspects of the cardiovascular physiology of the
duck. ZENTRALBLATT FUR VETERINARMEDIZIN REIHE A. 1974, Vol. 21, pp. 824-833.
—. 1974. The electrocardiogram of the Japanese (Coturnix coturnix japonica) and Bobwhite (Colinus
virginianus) Quail. Zentralblatt für Veterinärmedizin Reihe A. Avril, 1974, Vol. 21, 3, pp. 198-207.
TADJALLI, M., GHAZI, S.R. and PARTO, P. 2009. Gross anatomy of the heart in Ostrich (Struthio
Camelus). Iranian Journal of Veterinary research. November 26, 2009, Vol. 10, 1, pp. 21-27.
TAHA, A.A.M. and KING, A.S. 1986. Aortico-pulmonary bodies in the domestic fowl : ultrastructure,
innervation and secretion. Journal of Anatomy. 1986, 149, pp. 41-53.
TAHA, A.A.M., ABDEL-MAGIED, E.M. and KING, A.S. 1983. Ultrastructure of aortic and pulmonray
receptors in the domestic fowl. Journal of anatomy. 1983, 137, pp. 197-207.
TAKEI, Y., et al. 2011. B-type natriuretic peptide (BNP), not ANP, is the principal cardiac natriuretic
peptide in vertebrates as revealed by comparative studies. General and Comparative Endocrinology.
2011, Vol. 171, pp. 258-266.
TAYLOR, M. 1994. Endoscopic examination and biopsy techniques. [book auth.] B. RITCHIE, G.J.
HARRISON and L.R. HARRISON. Avian Medicine: Principles and Applications. Lake worth, FL : Blackwell
Publishing, 1994, pp. 327-354.
TAYLOR, W.M. 1994. Endoscopic examination and biopsy thechniques. [book auth.] B.W. RITCHIE, G.J.
HARRISON and HARRISON L.R. [ed.] eds. Harrison. Avian medicine : Principles and application. Lake
Worth : FL: Wingers Publishing, 1994, pp. 327-354.
TEGLAS, M.B., et al. 1998. Sarcocystis-associated encephalitis and myocarditis in a wild turkey (Mealgris
gallopavo). J. Parasitol. 1998, Vol. 86, pp. 661-663.
THIEL, P.G. 1978. A molecular mechanism for the toxic action of moniliformin, a mycotoxin produced by
fusarium moniliforme. Biochemical Pharmacology. February, 1978, Vol. 4, 15, pp. 483-486.
TODA, T., TODA, Y. and KUMMEROW, F.A. 1984. Electron microscopic comparison of the tunica media
of the thoracic aorta between species. The journal of experimental medicine. 1984, 143, pp. 141-147.
TOUSSAINT, J.F., et al. 2003. L'athérosclérose - Physiologie, diagnostics, thérapeutiques. Paris :
MASSON, 2003. p. 776 p.
TRAJANOVSKA, S. and DONALD, J.A. 2008. Molecular cloning of natriuretic peptides from the heart of
reptiles : loss of ANP in diapsid reptiles and birds. General and Comparative Endocrinology. 2008, Vol.
156, pp. 339–346.
TRAJANOVSKA, S. 2008. Molecular biology of natriuretic peptides in reptiles and birds, Pd.D. Thesis.
Australie : Deakin University, 2008.
TRAJANOVSKA, S., et al. 2007. Genomic analyses and cloning of novel chicken natriuretic peptide genes
reveal new insights into natriuretic peptide evolution. Peptide 28. 2007, pp. 2155-2163.
TREMBLAY, A. and BERNIER, G. 1992. Maladies d'origine nutritionnelle et métabolique. [book auth.] J.
BRUGERE-PICOUX and A. SILIM. Manuel de pathologie aviaire. MAISONS-ALFORT : Imprimerie du Cercle
des Elèves de l'Ecole Nationale Vétérinaire d'Alfort, 1992, pp. 343-354.

Page | 179
TRISTAN, T. 2010. The aging rapor. Veterinary Clinic of Norton America : Exotic Animal Practice. 2010,
Vol. 13, pp. 51-84.
TULLY, T.N., DORRESTEIN, G.M. and JONES, A.K. 2009. Handbook of avian medicine. 2nd edition.
Edinburgh : Saunders-Elsevier, 2009. 978-0-7020-2874-8.
UZUN, M., YILDIZ, S. and ONDER, F. 2004. Electrocardiography of rock partridges (alectoris graeca) and
chukar partridges (alectoris chukar). Journal of Zoo and Wildlife Medicine. December, 2004, Vol. 35, 4,
pp. 510-514.
VAN VLEET, J.F. and FERRANS, V.J. 1986. Myocardial diseases of animals. American journal of
pathology. 1986, Vol. 1, 124, pp. 98-178.
VAPNIARSKY, N., BARR, B.C. and MURPHY, B. 2012. Systemic Coxiella-like infection with myocarditis
and hepatitis in an Eclectus Parrot (Eclectus roratus). Veterinary pathology. 2012, Vol. 49, 4, pp. 717-
722.
VASSALL-ADAMS, P.R. 1982. The development of the atrioventricular bundle and its branches in the
avian heart. J Anat. 1982, Vol. 134, pp. 169-183.
VENUGOPAL, K., et al. 2007. Characteristics of an avian retrovirus isolated from an outbreak of
haemangioma among layers. Avian pathology. 2007, Vol. 27, 51.
VILLAR, D., et al. 2008. Clinical presentation and pathology of sarcocystosis in Psittaciform birds : 11
cases. Avian diseases. March, 2008, Vol. 52, 1, pp. 187-194.
VINK-NOOTEBOOM, M., et al. 1998. Clinical diagnosis of aneurysm of the right coronary artery in a
white cockatoo (Cacatua alba). Journal of Small Animal Pratice. 1998, 39, pp. 533-537.
VOGEL, C., GERLACH, H. and LÖFFLER, M. 1994. Columbiformes. [book auth.] B.W. RITCHIE, G.J.
HARRISON and L.R. HARRISON. Avian Medicine : Principles and Application. Lake Worth, FL : Wingers
Publishing, 1994, pp. 1200-1217.
WACK, R., KRAMER, L. and ANDERSON, L. 1994. Cardiomegaly and endocardial fibrosis in a secretary
bird (Sagittarius serpentarius). J Assoc Avian Vet. 1994, Vol. 8, pp. 76-80.
WAINE, J.C. and PATTERSON-KING, J.C. 2002. Atherosclerosis in a red-crested turaco (Tauracp
erythorlophus). Veterinary Record. December, 2002, Vol. 151, 25, p. 744.
WALLACH, J.D. and FLIEG, G.M. 1969. Frostbite and its sequelae in captive exotic birds. Journal of the
American Veterinary Medical Association. 1969, Vol. 155, 7, pp. 1035-1037.
WARNOCK, N.H., CLARKSON, T.B. and STEVENSON, R. 1957. Effects of exercise on blood coagulation
time and atherosclerosis of cholesterol-fed cockerels. Circulation research. 1957, Vol. 5, p. 478.
WEBB, D.M., DE NICOLA, D.B. and VAN VLEET, J.F. 1991. Serum chemistry alterations, including
creatine kinase isoenzymes, in furazolidone toxicosis of ducklings: preliminary findings. Avian diseases.
1991, Vol. 35, pp. 662-667.
WELLE, K.R., LIGHTFOOT, T. and REAVILL, D. 2011. Cardiovascular disease in aging birds. Proceedings of
the 2011 Association of Avian Veterinarians Annual Confererence. 2011, pp. 9-15.

Page | 180
WEST, J.B., et al. 2010. Structure-function studies of blood and air capillaries in chicken lung using 3D
electron microscopy. Respiratory Physiology & Neurobiology. 2010, Vol. 170, pp. 202-209.
WEST, J.B., WATSON, R.R. and FU, Z. 2006. The honeycomb-like structure of the bird lung allows a
uniquely thin blood-gaz barrier. Respiratory Physiology & Neurobiology. 2006, Vol. 152, pp. 115-118.
WEST, J.B., WATSON, R.R. and ZHENXING, F. 2007. Major differences in the pulmonary circulation
between birds and mammals. Respiratory Physiology & Neurobiology. 2007, Vol. 157, pp. 382-390.
WEST, N.H., LOWELL LANGILLE, B. and JONES, D.R. 1981. Cardiovascular system. [book auth.] A.S. KING
and J. McLELLAND. Form and function in birds. London : Academic Press, 1981, pp. 235-341.
WESTERHOF, I., M.D., VAN DE WAL and J.T., LUMEIJ. 2011. Electrocardiographic changes in a galah
(elophus roseicapilla) with lead poisonning in C.R. Congrès ''1st Scientific ECZM Meeting''. In Proc Annu
Conf Europ Assoc Avian Vet. Madrid : s.n., 2011, p. 59.
WILSON, J.B., JULIAN, R.J. and BARKER, I.K. 1988. Lesions of right heart failure and ascites in broiler
chickens. Avian diseases. 1988, Vol. 32, pp. 246-261.
WILSON, R.C., ZENOBLE, R.D. and HORTON, C.R. et al. 1989. Single dose digoxin pharmakokinetics in
the Quaker Conure (Myopsitta monachus). Journal of Zoo and Wildlife Medicine. 1989, Vol. 20, pp. 432-
434.
WOO, G-H., et al. 2010. Myocarditis by Nematodes Infection, Presumably Sarconema eurycerca, in a
wild whooper swan (cygnus cygnus) in korea. Journal of Veterinary Medicine Sciences. 2010, Vol. 72, 9,
pp. 1233-1235.
WORK, T.M., et al. 2002. Toxoplasmosis in three species of native and introduced Hawaiian birds. The
Journal of Parasitology. October, 2002, Vol. 88, 5, pp. 1040-1042.
ZANDVLIET. 2005. Electrocardiography in psittacine birds and ferrets. Seminars in Avian and Exotic Pet
Medicine. January, 2005, Vol. 14, 1, pp. 34-51.
ZANDVLIET, M.M.J.M., DORRESTEIN, G.M. and VAN DER HAGE, M.H. 2001. Chronic pulmonary
interstitial fibrosis in Amazon parrots. Avian pathology. 2001, Vol. 30, pp. 517-524.
ZAVALA, G., et al. 2011. Acute monensin toxicosis in broiler breeder chickens. Avian diseases.
September, 2011, Vol. 55, 3, pp. 516-521.
ZEHNDER, A.M., et al. 2009. Evaluation of indirect blood pressure monitoring in awake and
anesthetized red-tailed hawks (Buteo jamaicensis): effects of cuff size, cuff placement, and monitoring
equipment. Veterinary Anaesthesia and Analgesia. 2009, Vol. 36, pp. 464-479.
ZEHNDER, A.M., HAWKINS, M.G. and PSCOE, P.J. et al. 2009. evaluation of indirect blood pressure
monitoring in awake and anesthetized red-tailed hawk (Buteo jamaicensis): effects of cuff size, cuff
placement, and monitoring equipment. Veterinary Anaesthesia and Analgesia. 2009, Vol. 36, 5, pp. 464-
479.

Page | 181

Page | 182
NOM PRENOM : ZOLLER GRAHAM
TITRE : PATHOLOGIE CARDIOVASCULAIRE DES OISEAUX
Thèse d’Etat de Doctorat Vétérinaire : Lyon, le 12 Juin 2013
RESUME :
Qu’ils soient sauvages, d’élevage, de compagnie ou maintenus au sein de collections
zoologiques, les oiseaux ont un système cardiovasculaire structurellement uniforme et
physiologiquement adapté à l’approvisionnement énergétique intense des muscles du vol ou de
la course et ce, quelles que soient les conditions environnementales. Ces particularités, associées
au mode de vie de ces oiseaux permettent d’expliquer en partie l’apparition des lésions du cœur
et des vaisseaux. Néanmoins, quelle qu’en soit l’origine, les atteintes du système
cardiovasculaire s’installent généralement de façon insidieuse et se manifestent par des
symptômes relativement non spécifique, lorsqu’elles ne sont pas fulgurantes et mortelles. Ces
tableaux cliniques frustes rendent la réalisation d’examens complémentaires nécessaire à
l’établissement d’un diagnostic de certitude. L’approche thérapeutique des affections
cardiovasculaires vise à soutenir de façon hygiénique, médicale et chirurgicale l’hémodynamique
dans le système circulatoire et à traiter la cause de la maladie.
MOTS CLES :
- Nouveaux animaux de compagnie
- Oiseaux
- Cardiologie
- Maladies
JURY :
Président : Monsieur le Professeur CHEVALIER Philippe
1er Assesseur : Monsieur le Professeur ALOGNINOUWA Théodore
2ème Assesseur : Madame le Professeur BONNET-GARIN Jeanne-Marie
DATE DE SOUTENANCE : 12 Juin 2013
ADRESSE DE L’AUTEUR :
Lieu-dit : « La Vieille Eglise »
73410 Saint-Girod





























