Embed Size (px)
Citation preview

ÉCOLE NATIONALE VÉTÉRINAIRE D’ALFORT
Année 2010
ÉTIOLOGIE DES DIARRHÉES NÉONATALES ET
TRANSFERT COLOSTRAL CHEZ LE VEAU :
ENQUÊTE DANS LA CREUSE
THÈSE
Pour le
DOCTORAT VÉTÉRINAIRE
Présentée et soutenue publiquement devant
LA FACULTÉ DE MÉDECINE DE CRÉTEIL
le……………
par
Paul MAES
Né le 2 Mai 1984 à Reims (Marne)
JURY
Président : M. Professeur à la Faculté de Médecine de CRÉTEIL
Membres
Directeur : Yves Millemann Maître de conférences à l’ENVA
Assesseur : Loïc Desquilbet Maître de conférences à l’ENVA


REMERCIEMENTS
Je remercie le professeur de la faculté de médecine de Créteil qui nous fait l’honneur de
présider notre jury de thèse.
Je tiens à remercier Yves Millemann pour son accompagnement tout au long de cette thèse,
ainsi que pour son sérieux et sa participation.
Je tiens également à remercier Loïc Desquilbet pour avoir accepté l’assessorat alors que la
thèse était déjà commencée, pour sa disponibilité et son implication dans l’analyse et
l’interprétation des données.
Je voudrais aussi remercier Yves Lagalisse (Intervet Schering Plough), Didier Guérin
(GDSCC) et Éric Guillemot (LDA 23), qui ont soutenu cette étude et sans qui cela n’aurait
pas été possible.
Je remercie également le laboratoire Intervet Schering Plough, le GDSCC, le LDA 23 et
l’Unité de Pathologie du Bétail de l’ENVA qui ont financé cette thèse.
Un grand merci à Jérôme Bollache, Boris Boubet et Ludovic Coibion, vétérinaires à la
clinique d’Aubusson qui m’ont accueilli à deux reprises en stage et qui m’ont aidé quant à la
réalisation pratique de cette thèse.
Je remercie également Lionel Steimetz (Intervet Schering Plough) pour s’être penché sur les
résultats et pour avoir soumis des commentaires pertinents.
Je remercie aussi Ludovic Freyburger (unité de pathologie générale, microbiologie,
immunologie) pour son aide dans la traduction de termes d’immunologie.
Je tiens à remercier Jean-Jacques Benet, pour sa sympathie et son aide sur différents plans.
Je remercie Barbara Dufour pour son aide et ses conseils au commencement de cette thèse.

Je remercie les éleveurs des élevages de l’étude, pour leur accueil, leur aide et leur sympathie.
À mes parents, mes frères et sœur, beau-frère et belles-sœurs.
À WB, sans qui ces années d’études n’auraient pas du tout été les mêmes.
À une vraie poulotte, Karine.
À Thibaut, Thibaut, Marco et Pascal pour leur amitié de longue date.
Et surtout, à ma fiancée, Sandrine, que je remercie par-dessus tout, qui a toujours été à mes
côtés, qui m’a aidé, qui m’a permis de me motiver pour débuter cette thèse et qui m’a toujours
soutenu et poussé à aller de l’avant même dans les moments difficiles. Je te remercie d’être là,
pour moi.

TABLE DES MATIÈRES
GLOSSAIRE............................................................................................................................................... 6
LISTE DES FIGURES ET DES TABLEAUX..................................................................................................... 8
INTRODUCTION ..................................................................................................................................... 12
PREMIÈRE PARTIE : étiologie des diarrhées néonatales des veaux et importance du colostrum ........ 14
1/ Étiologie des diarrhées néonatales des veaux ......................................................................... 14
1.1/ Les bactéries incriminées.................................................................................................. 14
1.1.1/ Les colibacilles ........................................................................................................... 14
1.1.1.1/ Étiologie ............................................................................................................. 14
1.1.1.2/ Pathogénie ......................................................................................................... 16
1.1.1.3/ Clinique .............................................................................................................. 16
1.1.2/ Les salmonelles.......................................................................................................... 17
1.1.2.1/ Étiologie ............................................................................................................. 17
1.1.2.2/ Pathogénie ......................................................................................................... 17
1.1.2.3/ Clinique .............................................................................................................. 18
1.2/ Les virus ............................................................................................................................ 21
1.2.1/ Les rotavirus .............................................................................................................. 21
1.2.1.1/ Étiologie ............................................................................................................. 21
1.2.1.2/ Pathogénie ......................................................................................................... 22
1.2.1.3/ Clinique .............................................................................................................. 23
1.2.2/ Les coronavirus.......................................................................................................... 23
1.2.2.1/ Étiologie ............................................................................................................. 23
1.2.2.2/ Pathogénie ......................................................................................................... 25
1.2.2.3/ Clinique .............................................................................................................. 25
1.2.3/ Les torovirus .............................................................................................................. 26
1.2.4/ Autres virus................................................................................................................ 28
1.3/ Les parasites...................................................................................................................... 28
1.3.1/ Cryptosporidium parvum ........................................................................................... 28
1.3.1.1/ Étiologie ............................................................................................................. 28
1.3.1.2/ Pathogénie ......................................................................................................... 30
1.3.1.3/ Clinique .............................................................................................................. 30
1.3.2/ Giardia duodenalis..................................................................................................... 31
1.3.3/ Eimeria bovis et Eimeria zuernii ................................................................................ 31
1

1.4/ Diarrhées nutritionnelles .................................................................................................. 31
2/ Diagnostic, traitements et prévention ..................................................................................... 33
2.1/ Diagnostic étiologique ...................................................................................................... 33
2.1.1/ Bactéries .................................................................................................................... 34
2.1.2/ Virus........................................................................................................................... 35
2.1.2.1/ Coronavirus ........................................................................................................ 35
2.1.2.2/ Rotavirus ............................................................................................................ 36
2.1.2.3/ Torovirus ............................................................................................................ 36
2.1.2.4/ BVD virus ............................................................................................................ 36
2.1.3/ Protozoaires............................................................................................................... 36
2.2/ Traitement ........................................................................................................................ 37
2.3/ Prévention......................................................................................................................... 39
2.3.1/ Facteurs de risque ..................................................................................................... 40
2.3.1.1/ Gestion du troupeau et environnement............................................................ 40
2.3.1.2/ Âge du troupeau ................................................................................................ 41
2.3.1.3/ Complications au vêlage .................................................................................... 42
2.3.1.4/ Gestion colostrale .............................................................................................. 42
2.3.2/ Principes de prévention............................................................................................. 42
2.3.2.1/ Réduction de l’exposition aux agents pathogènes ............................................ 43
2.3.2.2/ Administration du colostrum ............................................................................. 44
2.3.2.3/Amélioration de l’immunité spécifique et non spécifique.................................. 44
3/ Immunité et colostrum ............................................................................................................ 50
3.1/ Rappels sur le statut immunitaire du veau nouveau‐né .................................................. 50
3.2/ Colostrogenèse, composition et rôles du colostrum........................................................ 51
3.3/ Transfert passif de l’immunité : mécanismes et intervalle de transfert .......................... 56
3.4/ Administration du colostrum............................................................................................ 57
3.5/ Facteurs influençant l’administration du colostrum ........................................................ 59
3.6/ Stockage du colostrum ..................................................................................................... 61
3.7/ Évaluation de la qualité colostrale et outils de mesure.................................................... 63
3.8/ Facteurs de modification du colostrum............................................................................ 66
3.9/ Colostrum artificiel ........................................................................................................... 70
2

DEUXIEME PARTIE : étude expérimentale ............................................................................................ 72
1/ Matériels et méthodes............................................................................................................. 72
1.1/ Présentation de l’étude .................................................................................................... 72
1.2/ Choix des élevages............................................................................................................ 73
1.3/ Choix des veaux ................................................................................................................ 74
1.4/ Réalisation des prélèvements........................................................................................... 74
1.4.1/ Prélèvements de fèces .............................................................................................. 74
1.4.2/ Prélèvements sanguins.............................................................................................. 74
1.5/ Matériel de recherche des agents pathogènes ................................................................ 74
1.5.1/ Tetrakits..................................................................................................................... 74
1.5.2/Analyses de laboratoire.............................................................................................. 76
1.5.2.1/ Dénombrement des E. coli dans les fèces.......................................................... 76
1.5.2.2/ Typage des E. coli ............................................................................................... 78
1.5.2.3/ Recherche des salmonelles ................................................................................ 79
1.6/ Dosages des immunoglobulines G et des protéines totales............................................. 79
1.7/ Outils statistiques ............................................................................................................. 80
2/ Résultats................................................................................................................................... 80
2.1/ Partie descriptive .............................................................................................................. 80
2.1.1/ Dosage des IgG et PT ................................................................................................. 80
2.1.2/ Concernant les diarrhées........................................................................................... 82
2.1.2.1/ Description des diarrhées .................................................................................. 82
2.1.2.2/ Agents pathogènes rencontrés .......................................................................... 85
2.1.3/ Concernant les élevages............................................................................................ 87
2.1.3.1/ Bâtiments et environnement............................................................................. 87
2.1.3.2/ Gestion de l’élevage des veaux.......................................................................... 89
2.2/ Partie analytique............................................................................................................. 100
2.2.1/ Expositions individuelles ......................................................................................... 101
2.2.2/ Expositions d’élevage .............................................................................................. 106
3/ Discussion............................................................................................................................... 114
CONCLUSION ....................................................................................................................................... 122
BIBLIOGRAPHIE.................................................................................................................................... 124
3

4

5

GLOSSAIRE
AFNOR : Agence Française de Normalisation
AFSSAPS : Agence Française de Sécurité Sanitaire des Produits de Santé
AINS : Anti‐Inflammatoires Non‐Stéroïdiens
AMM : Autorisation de Mise sur le Marché
ARN : Acide Ribo‐Nucléique
BVD/MD : Bovine Viral Diarrhoea/Mucosal Disease
Cellules M : Cellules spécialisées dans les tissus lymphoïdes intestinaux
COFRAC : Comité Français d'Accréditation
ColV : Colicine V (un type de plasmide de E. coli)
CS31A : Facteur d’attachement de E. coli
DT104 : Lysotype de S. Typhimurium possédant des gènes de multirésistance aux antibiotiques
E. coli : Escherichia coli
ECET : Escherichia coli Entéro‐Toxinogène
ELISA : Enzyme‐Linked ImmunoSorbent Assay
ENVA : École Nationale Vétérinaire d’Alfort
F5 : Facteur d’attachement de E. coli (anciennement K99)
F41 : Facteur d’attachement de E. coli
FcRn : Récepteur néonatal au fragment Fc des immunoglobulines
GDSCC : Groupement de Défense Sanitaire du Cheptel Creusois
GMQ : Gain Moyen Quotidien
IgA : Immunoglobuline A
IgE : Immunoglobuline E
IGF : Insulin Growth Factor
IgG : Immunoglobuline G
6

IgM : Immunoglobuline M
IL‐1 : Interleukine 1
K99 : Facteur d’attachement de E. coli (actuellement F5)
LDA 23 : Laboratoire Départemental Agréé de la Creuse
LMR : Limite Maximale de Résidus
LT : Entérotoxine thermolabile de E. coli
NBVC : Nutrition Biochimie Vétérinaires Consultants
NSP4 : Protéine virale Non Structurale 4
O111 : Souche vérotoxinogène de E. coli
O130 : Souche vérotoxinogène de E. coli
O157:H7 : Souche vérotoxinogène de E. coli
O26 : Souche vérotoxinogène de E. coli
OR : Odds Ratios
PCR : Réaction de Polymérisation en Chaîne
PT : Protéines Totales
RESSAB : Réseau d’Épidémiosurveillance des Salmonelloses Bovines
RID : Immunodiffusion Radiale
RT‐PCR : Retro Transcriptase PCR
STa : Entérotoxine thermostable a de E. coli
STb : Entérotoxine thermostable b de E. coli
TGF : Transforming Growth Factor
TNF‐α : Tumor Necrosis Factor α
UFC : Unité Formant Colonie
7

LISTE DES FIGURES ET DES TABLEAUX
Figure 1 : Représentation schématique d’un E. coli (source : http://en.wikipedia.org/wiki/Bacteria
[en ligne] (consulté le 10 janvier 2010)) ....................................................................................... 15
Figure 2 : Modèle schématique d’un rotavirus (165)............................................................................ 21
Figure 3 : Modèle schématique d’un coronavirus (165) ....................................................................... 24
Figure 4 : Modèle schématique d’un torovirus (165)............................................................................ 26
Figure 5 : Schéma récapitulatif des principaux virus intervenant dans les diarrhées néonatales des
veaux, ainsi que leur mécanisme physiopathologique (165) ....................................................... 27
Figure 6 : Cycle évolutif de Cryptosporidium parvum (26).................................................................... 29
Figure 7 : Début de développement d’un trophozoïte de Cryptosporidium parvum (26) .................... 30
Figure 8 : Trophozoïte de Cryptosporidium parvum développé dans la membrane cytoplasmique
d’un entérocyte (26) ..................................................................................................................... 30
Tableau 1 : Estimation de la déshydratation du veau (20).................................................................... 32
Tableau 2 : Évaluation du degré d’acidose du veau par un examen clinique à distance (20) .............. 33
Figure 9 : Développement de la réponse immunitaire chez le veau : de la conception à la
puberté (29).................................................................................................................................. 51
Tableau 3 : Composition du colostrum, du lait de transition et du lait chez les vaches Holstein (61) . 52
Figure 10 : Survie des veaux en fonction de leur concentration sérique en IgG (116) ......................... 64
Tableau 4 : Présentation de la partie agents pathogènes de l’étude ................................................... 72
Tableau 5 : Présentation de la partie immunité et transfert colostral de l’étude ................................ 73
Photo 1 : Tigettes d’immunochromatographie latérale (en rouge la tigette pour les rotavirus, en
jaune pour les coronavirus, en bleu pour E. coli F5 et en vert pour Cryptosporidium parvum)
(57)................................................................................................................................................ 75
Figure 11 : Lecture du résultat sur la tigette (57).................................................................................. 75
Figure 12 : Mode de fonctionnement des tigettes (57) ........................................................................ 76
Figure 13 : Population totale de l’étude et répartition des veaux en fonction de leur statut malade
ou non malade.............................................................................................................................. 81
Figure 14 : Répartition des taux d’IgG sériques .................................................................................... 81
Figure 15 : Répartition des taux de PT sériques.................................................................................... 82
Figure 16 : Nombre de veaux en fonction de la consistance des diarrhées.......................................... 82
Figure 17 : Nombre de veaux en fonction de la couleur des diarrhées ................................................ 83
Figure 18 : Nombre de veaux en fonction de la température des veaux malades ............................... 83
8

Figure 19 : Nombre de veaux en fonction de l’état d’hydratation des veaux....................................... 84
Figure 20 : Nombre de veaux en fonction de la note de déshydratation des veaux déshydratés........ 84
Figure 21 : Nombre de veaux de la partie agents pathogènes de l’étude selon leur statut................. 85
Figure 22 : Agents pathogènes mis en évidence par le kit rapide......................................................... 85
Figure 23 : Dénombrement des E. coli au laboratoire .......................................................................... 86
Figure 24 : Typage des E. coli au laboratoire......................................................................................... 86
Figure 25 : Répartition des races dans les élevages de l’étude............................................................. 87
Figure 26 : Nombre de bâtiments par élevage...................................................................................... 88
Figure 27 : État de propreté des bâtiments .......................................................................................... 88
Figure 28 : État de la ventilation dans les bâtiments ............................................................................ 89
Figure 29 : Écart d’âge entre les veaux d’un même lot......................................................................... 90
Figure 30 : Modalités de vêlage des veaux malades ............................................................................. 90
Figure 31 : Conditions d’hygiène au vêlage........................................................................................... 91
Figure 32 : Nombre d’élevage selon le type de lieu de vêlages ............................................................ 91
Figure 33 : Hygiène du lieu de vêlage.................................................................................................... 92
Figure 34 : Examens de la mamelle et du lait réalisés avant la tétée colostrale .................................. 92
Figure 35 : Fréquence de lavage des trayons des vaches par l’éleveur ................................................ 93
Figure 36 : Nombre d’élevages selon la fréquence de contrôle de la qualité du colostrum ................ 93
Figure 37 : Nombre d’élevages selon l’isolement ou non de la mère et son veau à la naissance ........ 94
Figure 38 : Nombre d’élevages apportant différents soins aux veaux en néonatalité ......................... 94
Figure 39 : Différents contrôles effectués en néonatalité par les éleveurs .......................................... 95
Figure 40 : Nombre d’élevages selon le type de réponse de l’éleveur face au refus de téter d’un
veau .............................................................................................................................................. 95
Figure 41 : Nombre d’élevages selon le type de réponse de l’éleveur face à une vache sans
colostrum...................................................................................................................................... 96
Figure 42 : Répartition des élevages ayant des veaux voleurs.............................................................. 96
Figure 43 : Nombre d’élevages selon la fréquence de l’isolement des veaux malades........................ 97
Figure 44 : Nombre d’élevages selon le moment de réintroduction des veaux malades (pour les
élevages isolant les veaux malades) ............................................................................................. 97
Figure 45 : Répartition des élevages qui vaccinent ou non leurs vaches contre les agents
pathogènes des diarrhées néonatales.......................................................................................... 98
Figure 46 : Valences des vaccins utilisés par les éleveurs ..................................................................... 98
Figure 47 : Nombre d’élevages selon les modalités de choix des valences des vaccins ....................... 99
Figure 48 : Populations vaccinées par les éleveurs ............................................................................... 99
Figure 49 : Répartition des vaccins utilisés par les éleveurs ............................................................... 100
9

Figure 50 : Répartition des sexes des veaux en fonction de la maladie.............................................. 101
Figure 51 : Répartition des mères des veaux en fonction de la maladie ............................................ 102
Figure 52 : Répartition du taux d’IgG des veaux en fonction de la maladie ....................................... 102
Figure 53 : Répartition du taux de PT des veaux en fonction de la maladie....................................... 103
Figure 54 : Répartition des résultats de détection des rotavirus en fonction de la maladie .............. 103
Figure 55 : Répartition des résultats de détection des coronavirus en fonction de la maladie ......... 104
Figure 56 : Répartition des résultats de détection de Cryptosporidium parvum en fonction de la
maladie ....................................................................................................................................... 104
Figure 57 : Répartition des résultats de détection de E. coli F5 avec le tetrakit en fonction de la
maladie ....................................................................................................................................... 105
Figure 58 : Répartition des résultats de détection de E. coli F5 au laboratoire en fonction de la
maladie ....................................................................................................................................... 105
Figure 59 : Répartition des résultats de détection de E. coli CS31A en fonction de la maladie ......... 106
Figure 60 : Répartition du nombre de vêlages moyen en fonction de la maladie .............................. 107
Figure 61 : Répartition de la qualité de la ventilation en fonction de la maladie ............................... 107
Figure 62 : Répartition du type de sol en fonction de la maladie ....................................................... 108
Figure 63 : Répartition de l'entretien du matériel de vêlage en fonction de la maladie .................... 108
Figure 64 : Répartition de l'hygiène des mains de l'éleveur avant le vêlage en fonction de la
maladie ....................................................................................................................................... 109
Figure 65 : Répartition de l'utilisation de gants de vêlage en fonction de la maladie ........................ 109
Figure 66 : Répartition des lieux de vêlage en fonction de la maladie ............................................... 110
Figure 67 : Répartition de la propreté des lieux de vêlage en fonction de la maladie ....................... 110
Figure 68 : Répartition de l'entretien des lieux de vêlage en fonction de la maladie......................... 111
Figure 69 : Répartition des soins apportés à l'ombilic en fonction de la maladie .............................. 111
Figure 70 : Répartition des examens de la mamelle avant la tétée colostrale en fonction de la
maladie ....................................................................................................................................... 112
Figure 71 : Répartition de l'examen des premiers jets de lait en fonction de la maladie................... 112
Figure 72 : Répartition de la propreté des trayons avant la tétée colostrale en fonction de la
maladie ....................................................................................................................................... 113
Figure 73 : Répartition des veaux voleurs en fonction de la maladie ................................................. 113
Figure 74 : Répartition de la vaccination des mères en fonction de la maladie ................................. 114
10

11

INTRODUCTION
La diarrhée est un syndrome caractérisé par l’émission trop fréquente de fèces trop liquides. La
diarrhée néonatale est encore à ce jour une maladie importante du veau nouveau‐né (135), elle est
la principale cause de maladie chez les veaux. Elle peut toucher de 10% à 80% des veaux suivant les
élevages. Les diarrhées néonatales ont des répercussions économiques importantes de par le coût
des soins à apporter aux veaux et par les mortalités (20).
De nombreux agents pathogènes peuvent causer ces diarrhées, tels que des bactéries, des virus ou
des parasites, chacun ayant une incidence différente selon l’âge du veau atteint (20). Une origine
nutritionnelle est également reconnue comme cause de diarrhées néonatales chez les veaux. Dans
tous les cas, les diarrhées néonatales des veaux sont d’origine multifactorielle, de nombreux facteurs
de risque intervenant. Les principaux agents pathogènes reconnus dans les diarrhées néonatales sont
les rotavirus, les coronavirus, les Escherichia coli entérotoxinogènes (ECET), Cryptosporidium parvum
et les salmonelles. Le torovirus a récemment été associé aux diarrhées néonatales en Amérique du
Nord et en Europe ; le virus du syndrome BVD/MD (Bovine Viral Diarrhoea/Mucosal Disease) est en
revanche peu fréquemment associé aux diarrhées des jeunes veaux (135).
La présence des différents agents pathogènes varie avec l’âge du veau, ce qui est utile pour
déterminer quel est l’agent pathogène probablement en cause. Les agents pathogènes peuvent être
seuls ou en association (135).
Le risque pour un veau de déclarer une diarrhée néonatale ou de mourir suite à une diarrhée
néonatale est fortement lié à l’état sanitaire de l’élevage, par la gestion du troupeau, et notamment
l’efficacité du transfert passif de l’immunité, la nutrition des veaux, l’hygiène des locaux et l’hygiène
au vêlage, le statut de vaccination et la santé des vaches. Le principal facteur de risque est le statut
immunitaire du veau, qui rend compte de l’efficacité du transfert colostral ainsi que de la qualité du
colostrum. Une mortalité inférieure ou égale à cinq pourcents des veaux à cause d’un épisode de
diarrhée est considéré comme acceptable dans un élevage (135). Les veaux malades sont une source
de contamination importante pour l’environnement et représentent donc un facteur de risque pour
les autres veaux.
Les diarrhées sont la conséquence de deux mécanismes différents. Elles peuvent être associées à une
augmentation des sécrétions intestinales suite à l’action d’entérotoxines, dans ce cas on a une
12

diarrhée par hypersécrétion. Elles peuvent également être associées à une diminution de
l’absorption intestinale à la suite de la destruction des villosités par l’agent pathogène ou lors de
fermentations excessives et synthèse de produits de fermentation (comme l’acide lactique) qui par
leur fort pouvoir osmotique vont attirer l’eau vers la lumière intestinale, on parle alors de diarrhée
par malabsorption. Enfin, on peut avoir des diarrhées mixtes (20).
Les pertes en eau qui résultent de la diarrhée peuvent atteindre un à quatre litres par jour. Les
principaux signes cliniques sont la diarrhée, la déshydratation, une hypothermie ou hyperthermie
(plus rarement observée car précoce et fugace (moins de 24h)), un abattement, une diminution du
tonus musculaire, des modifications cardiaques (bradycardie, arythmie), une anurie, une diminution
voire une perte du réflexe de succion. Les infections concomitantes ne sont pas rares (pulmonaires,
ombilicales, articulaires). L’état général du veau peut se dégrader en quelques heures, et cela peut
entraîner sa mort (20).
L’étude s’est déroulée dans la Creuse, dans les environs d’Aubusson, au cours des mois de Janvier et
Février 2009. Le but était de réaliser des prélèvements des fèces sur des veaux de 0 à 15 jours en
diarrhée et sur des veaux sains du même âge dans des élevages qui avaient ou n’avaient pas de
veaux en diarrhée, afin de rechercher des agents pathogènes à l’aide d’un kit de diagnostic rapide,
fourni par Intervet Schering Plough, et également par des analyses classiques de laboratoire faites au
Laboratoire Départemental Agréé de la Creuse (LDA 23), prises en charge pour une partie par le LDA
23 et pour le reste par Intervet Schering Plough.
Le statut immunitaire des veaux a également été mesuré par le laboratoire NBVC (Nutrition
Biochimie Vétérinaires Consultants), suite à une prise de sang sur des veaux en diarrhée ou non, et
âgés de 2 à 8 jours. Ces analyses ont été prises en charge par le GDSCC (Groupement de Défense
Sanitaire du Cheptel Creusois) et par l’Unité de pathologie du bétail de l’ENVA.
Un questionnaire rempli sur place au cours de la visite de l’élevage a également permis de récolter
des informations sur la gestion et l’hygiène des élevages.
L’objectif de cette étude est de mettre en relation le statut immunitaire passif du veau (déterminé
par le dosage des immunoglobulines G (IgG) et des protéines totales sériques) avec la morbidité et la
mortalité des veaux. Des objectifs secondaires sont de mettre en évidence les facteurs de risque des
diarrhées néonatales dans les élevages, et de déterminer les raisons des échecs de transfert passif de
l’immunité.
13

PREMIÈRE PARTIE : Étiologie des diarrhées néonatales des veaux et importance du colostrum
1/ Étiologie des diarrhées néonatales des veaux
De nombreux agents pathogènes sont responsables des diarrhées néonatales. Il peut s’agir de
bactéries, de virus ou de parasites (20).
1.1/ Les bactéries incriminées
Les diarrhées bactériennes sont dues majoritairement aux colibacilles, aux salmonelles et aux
clostridies (20).
1.1.1/ Les colibacilles
1.1.1.1/ Étiologie
Escherichia coli (E. coli) est une entérobactérie anaérobie facultative gram négatif (figure 1), présente
de façon normale dans la flore intestinale d’un veau sain à une concentration de 106 bactéries par
gramme de fèces. Elle colonise l’intestin précocement, dans les huit premières heures de vie de
l’animal (60). Les souches pathogènes d’E. coli possèdent des facteurs de virulence impliqués dans la
pathogénie de la maladie, tels que des adhésines, des entérotoxines et des cytotoxines. Ces souches
pathogènes sont transmises des adultes aux nouveau‐nés par voie fécale‐orale (20, 60). Les
réservoirs de la bactérie sont les individus porteurs sains, ainsi que l’environnement. En effet, les
colibacilles sont capables de survivre plusieurs mois dans le sol, l’eau et les aliments souillés par des
déjections.
14

Figure 1 : Représentation schématique d’un E. coli (source : http://en.wikipedia.org/wiki/Bacteria [en
ligne] (consulté le 10 janvier 2010))
Les ECET possèdent deux catégories de facteurs de virulence, les fimbriae (pili) et les entérotoxines.
Les facteurs d’attachement F5 (anciennement K99) et F41 sont responsables de l’adhérence, et les
entérotoxines thermostables (STa et STb) et thermolabiles (LT) stimulent la sécrétion des cellules des
cryptes. Bien que des ECET produisent des entérotoxines thermolabiles, la plupart des souches
responsables des diarrhées néonatales produisent l’entérotoxine thermostable STa. Cette
entérotoxine ainsi que le facteur d’attachement F5 sont des facteurs de virulence transmis par
l’intermédiaire de plasmides (135).
Les souches vérotoxinogènes (O157:H7, O111, O26 et O130), elles, sont responsables de diarrhées
hémorragiques chez le jeune veau (20, 84).
E. coli CS31A est incriminé dans les gastro‐entérites paralysantes, aussi appelées syndrome
diarrhéique avec ataxie, bien que son rôle soit aujourd’hui mis en doute dans ces diarrhées (20). Les
gastro‐entérites paralysantes sont caractérisées par la discrétion des signes diarrhéiques, l'absence
de déshydratation et la présence signes nerveux dominés par la parésie et de l'ataxie (118).
La maladie est classiquement observée sur des veaux de moins de 4 jours (20, 135), bien que
l’infection concomitante d’un rotavirus puisse étendre cette fenêtre de 7 à 14 jours. Les cellules
intestinales des veaux de plus de 2 jours acquièrent une résistance naturelle à l’adhésion du facteur
d’attachement F5 (20).
15

1.1.1.2/ Pathogénie
La diarrhée est la résultante d’une sécrétion exacerbée, ou d’une baisse de l’absorption dans le tube
digestif. Les colibacilles causent des diarrhées par leur sécrétion d’entérotoxines qui provoquent une
perturbation de la perméabilité cellulaire et entraînent un passage d’eau et d’électrolytes vers la
lumière intestinale (accroissement des sécrétions intestinales) (20, 135).
Ensuite, l’excès de nutriments est fermenté dans le gros intestin, ce qui provoque une prolifération
bactérienne, la formation d’acides organiques et d’autres composés toxiques. L’effet osmotique de
ces nutriments entraîne un appel d’eau vers la lumière du tube digestif, ce qui contribue à la diarrhée
(20).
1.1.1.3/ Clinique
La diarrhée à E. coli est caractérisée par une couleur jaune paille, elle est profuse et très liquide. La
déshydratation est le signe clinique le plus marqué, avec comme conséquences une hypothermie, un
abattement et une hypotension (135).
Les souches vérotoxinogènes se retrouvent chez les veaux de 1 à 4 semaines et se caractérisent par
une diarrhée mucoïde hémorragique.
Les infections à E. coli CS31A présentent un veau mou avec parésie postérieure et fèces pâteux
d’odeur bien particulière de « beurre rance ». L’abdomen est distendu (caillette pleine), la
déshydratation est modérée et la létalité faible. Les signes cliniques pourraient être la conséquence
d’une bactériémie avec endotoxémie colibacillaire subaiguë transitoire, accompagnée d’une acidose
métabolique par les D‐lactates (54, 135).
Quand les pertes hydriques sont supérieures aux apports, des signes de déshydratation et d’acidose
apparaissent (20).
Une cause de mort probable est la défaillance cardiaque, résultant d’un déséquilibre potassique du
myocarde, l’hypothermie contribuant également à cette défaillance cardiaque (20).
16

1.1.2/ Les salmonelles
1.1.2.1/ Étiologie
Les salmonelles sont des entérobactéries gram négatif. Les veaux peuvent être infectés par une large
gamme de sérotypes de Salmonelles dans les heures suivant la naissance (2). Les principaux
sérotypes rencontrés sont S. Typhimurium et S. Dublin. Les réservoirs de la bactérie sont les vaches
adultes, dont l’excrétion de la bactérie est intermittente dans le lait et les fèces chez les porteurs
asymptomatiques, les oiseaux et les rongeurs dont la dissémination de la bactérie se fait par leurs
fèces. La survie de la bactérie est longue : 4 mois dans l’eau, 5 mois dans le sol et 12 mois dans le
fumier. La contamination se fait par voie orale (60), bien que les muqueuses de l’arbre respiratoire
supérieur et les conjonctives soient également des voies de contamination rapportées.
La salmonellose se déclare en général chez des veaux âgés de 1 à 8 jours (60), mais peut se produire
également jusqu’à 28 jours, et même encore chez des veaux plus âgés (2).
L’immunité contre les salmonelles change rapidement au cours des trois premiers mois de la vie du
veau. À deux semaines d’âge la dose létale pour les souches virulentes est de 105 bactéries par
gramme de fèces, vers ses six ou sept semaines de vie, elle est de 107 bactéries par gramme de fèces
et à douze‐quatorze semaines d’âge, elle est de 1010 bactéries par gramme de fèces Les individus
adultes sont aussi bien les réservoirs que les sources de l’infection des veaux par Salmonella Dublin
(20).
L’exposition aux salmonelles peut se produire par du lait ou du colostrum contaminé, par la surface
contaminée des trayons ou du pis de la vache, par le personnel, l’équipement ou l’environnement
(20). Des souches multirésistantes de Salmonella sont fréquemment impliquées dans les maladies
des veaux, et occasionnellement des humains. Au cours d’une suspicion de salmonellose, il est
prudent d’informer les éleveurs du risque zoonotique de la salmonellose pour eux et leur famille.
1.1.2.2/ Pathogénie
Suite à l’ingestion, les salmonelles colonisent le tractus intestinal et envahissent les entérocytes de
l’iléon et les cellules M (qui sont des cellules spécialisées dans les tissus lymphoïdes intestinaux) (74,
139, 183), ainsi que les amygdales (64). Elles se fixent par leurs fimbriae sur des récepteurs
spécifiques et pénètrent dans la cellule par endocytose ce qui laisse des lésions d’effacement et une
17

réorganisation du cytosquelette (60). Elles détruisent les microvillosités intestinales, il en résulte un
défaut d’absorption. Il y a également destruction de la muqueuse et invasion de la sous‐muqueuse.
L’inflammation locale entraîne une augmentation des sécrétions (135).
Dans les tissus lymphoïdes, les salmonelles entrent dans les phagocytes mononucléés et sont
rapidement disséminées dans tout l’organisme via les nœuds lymphatiques (60, 64, 74). La capacité
des salmonelles à infecter les veaux via les amygdales a été démontrée par des études
expérimentales sur des veaux œsophagectomisés (42). Chez ces veaux, Salmonella spp a été isolée
dans les tissus dans les trois heures suivant l’infection orale.
Les mécanismes de virulence de Salmonella spp incluent la capacité d’invasion de la muqueuse
intestinale, de multiplication dans les tissus lymphoïdes, et d’échappement aux systèmes de défense
de l’hôte, entraînant une maladie systémique. On pense que la diarrhée associée à la salmonellose
est due à la réponse inflammatoire à l’infection. Une corrélation positive est retrouvée entre la
sévérité des lésions histopathologiques détectées dans la muqueuse iléale et le volume des
sécrétions de fluides (189). Le relargage d’endotoxines, de prostaglandines, et de cytokines pro‐
inflammatoires (interleukine 1 (IL‐1) et Tumor Necrosis Factor α (TNF‐α)) (166) augmente également
la perméabilité vasculaire et donc l’hypersécrétion. La desquamation des cellules épithéliales
entraîne une hémorragie, une production de fibrine, une maldigestion et une malabsorption (183).
L’état hyperosmotique qui en résulte dans la lumière intestinale appelle les fluides vers la lumière,
contribuant à la perte d’eau, de sodium, de potassium et de bicarbonates. Les dégâts sur la
muqueuse contribuent également à la perte de protéines et donc à l’hypoprotéinémie.
1.1.2.3/ Clinique
Les manifestations de la maladie sont variables, reflet des interactions entre l’immunité de l’hôte, la
dose d’agent pathogène rencontrée et sa virulence. Les diarrhées à salmonelles sont caractérisées
par une diarrhée liquide nauséabonde, une perte d’appétit, un abattement et une hyperthermie
(135).
Peu de signes cliniques de la maladie peuvent être observés chez les veaux souffrant de salmonellose
suraiguë et on les retrouve généralement morts, sans prodromes observés par l’éleveur. Les veaux
atteints peuvent avoir été léthargiques ou sans appétit au cours des quelques repas précédents. Les
18

analyses sanguines montrent une leucopénie, une neutropénie, une hémoconcentration associée à
une déshydratation, une acidose métabolique et une urémie augmentée (104).
L’anorexie et l’abattement sont typiquement les premiers signes cliniques observés dans les
infections aiguës. L’hyperthermie et la diarrhée suivent dans les 48 à 72 heures post‐infection. La
fièvre peut persister jusqu’à 7 jours post‐infection (105, 106). L’absence d’hyperthermie n’exclut pas
la présence de salmonellose, la réponse fébrile étant transitoire et les veaux succombant à l’infection
étant souvent hypothermiques dans les 12‐24 heures précédant la mort (105).
La diarrhée est aqueuse, profuse, et peut être mucofibrineuse et hémorragique. Des différences sont
observées entre les infections par différents sérovars et même des différences entre différentes
souches d’un même sérovar. C’est le résultat de différences entre les facteurs de virulence des
différentes souches (105, 106). S. Typhimurium donne une diarrhée jaune à brunâtre, pouvant
contenir du sang et des débris de muqueuse intestinale. Hyperthermie, déshydratation sévère,
mortalité importante, septicémie fréquente caractérisent une salmonellose à S. Typhimurium. S.
Dublin donne une diarrhée d’odeur fétide, avec parfois du sang et des lambeaux de muqueuse. Les
veaux présentent une anorexie, une hyperthermie et une mortalité en 1 à 2 jours dans 5 à 10% des
cas (60, 64, 133, 155, 183).
S. Typhimurium est communément impliquée dans les pathologies entéritiques des veaux de moins
de deux mois (64, 133, 155, 168, 183, 184). Au contraire, S. Dublin est impliquée dans des maladies
de fréquence similaire chez les jeunes adultes et les adultes (157).
Plusieurs de ces signes cliniques sont associés à une endotoxémie. Les signes cliniques systémiques
de l’endotoxémie sont l’hyperthermie, la tachypnée, la tachycardie, une leucopénie ou leucocytose,
et un abattement profond appelé tuphos. Certains sérotypes, en particulier S. Typhimurium, ont
tendance à induire une inflammation sévère de la muqueuse intestinale. Les pertes hydriques,
électrolytiques et protéiques sont très rapides et peuvent menacer la vie de l’animal si elles ne sont
pas corrigées (20).
Les veaux qui ont survécu à la phase aiguë de la maladie passent par une période de cachexie au
cours de la période de rétablissement. La sévérité et la durée de la phase clinique de la maladie sont
liées à la virulence de la souche, la dose infectante, l’âge du veau, l’efficacité de l’immunité passive,
la nutrition, et le degré de stress environnemental (104).
19

Bien que les pathologistes associent la salmonellose avec des lésions du tube digestif, les infections
suraiguës donnent souvent peu lieu au développement de lésions pathologiques (64, 183).
Lorsqu’elles sont présentes, ces lésions peuvent être une congestion pulmonaire, et des pétéchies
hémorragiques muqueuses et séreuses de multiples organes, dont les intestins et le cœur.
La salmonellose aiguë est typiquement caractérisée par une entérite hémorragique catarrhale diffuse
avec une iléotyphlocolite fibrinonécrotique (99). Le contenu intestinal est aqueux, malodorant, et
peut contenir du mucus ou du sang. L’inflammation de la vésicule biliaire est fréquente, et une
preuve histopathologique de cholécystite fibrineuse est considérée comme pathognomonique d’une
salmonellose aiguë chez le veau (99). Les nœuds lymphatiques mésentériques sont souvent
hypertrophiés, œdémateux et hémorragiques. Des érosions de la muqueuse abomasale peuvent être
observées, en particulier avec S. Dublin. Les salmonelloses chroniques peuvent conduire à un
épaississement de la paroi intestinale avec du matériel nécrotique jaune‐gris en surface d’une
muqueuse rouge (99).
Les E. coli et les salmonelles sont les deux principales bactéries responsables de diarrhées
néonatales chez les veaux. De par les nombreux facteurs de virulence, les E. coli entraînent différents
types de diarrhées, les diarrhées à E. coli F5 étant une des plus fréquentes chez les très jeunes veaux.
Les signes cliniques peuvent être sévères, surtout en cas de diarrhée due à Salmonella qui entraîne
souvent le décès de l’animal.
D’autres bactéries sont soupçonnées d’intervenir dans les diarrhées néonatales, sans que leur
implication ait encore été réellement démontrée. Il s’agit de Clostridium et de Campylobacter.
Les clostridies ne sont pas communément considérées comme des agents pathogènes majeurs de
diarrhée néonatale (20). Elles infectent le veau de 5 à 15 jours, mais peuvent l’infecter avant. Selon le
type de Clostridium, on a des signes cliniques différents (135).
Parmi les clostridies, Clostridium perfringens est l’agent causal le plus important dans les affections
néonatales. Les clostridies produisent des toxines qui sont propres à leur type (135). C. perfringens
type A est associé à une abomasite hémorragique aiguë des nouveau‐nés et une diarrhée mucoïde.
Les signes cliniques comprennent une distension abdominale aiguë, une colique, une dépression, et
une mort subite (20).
L’importance clinique de Campylobacter dans les diarrhées néonatales n’est pas démontrée (20).
20

1.2/ Les virus
Les virus intestinaux se multiplient à l’intérieur des cellules. L’atrophie des villosités se développe au
fur et à mesure de la destruction des cellules épithéliales (20).
1.2.1/ Les rotavirus
1.2.1.1/ Étiologie
Les rotavirus sont la cause la plus commune de diarrhée néonatale du veau. Ils appartiennent à la
famille des Réovirus, ce sont des virus sphériques et nus, avec une double capside autour d’un ARN
bicaténaire ce qui lui confère une grande résistance (figure 2). Ils ont été découverts par Mebus en
1967 par microscopie électronique. On a deux groupes de rotavirus, les groupes A et B. Une étude de
Chinsangaram et al. montre que les rotavirus des groupes A et B sont présents en proportion
équivalente chez les veaux : chez 94% et 81% respectivement des individus de cette étude (36). La
proportion de veaux malades infectés par des rotavirus est de 40 à 50%, et elle est de 10 à 20% chez
les veaux sains (20).
Figure 2 : Modèle schématique d’un rotavirus (165)
La rotavirose touche des veaux âgés de 5 jours à 2 semaines, bien que la maladie puisse se produire à
l’âge de 24 heures, en particulier chez les veaux n’ayant pas pris leur colostrum. On pense que cela
21

touche des veaux de cet âge et n’ayant pas pris leur buvée colostrale car les mères sécrètent dans
leur colostrum des anticorps anti‐rotavirus qui confèrent au veau une protection locale contre les
rotavirus jusqu’à ce que le taux d’anticorps diminue dans le lait entre 48 et 72 heures post‐partum.
Les signes cliniques peuvent également être observés à un âge plus avancé dans le cas où il y a
fection concomitante avec ECET (20).
à l’infection n’est pas fonction de l’âge, contrairement à la résistance à la maladie
linique (20).
mune, avec la vache réservoir infectant son propre
eau, et ces veaux infectant d’autres veaux (20).
neuf
ois et par conséquent peuvent rester dans une zone de vêlage d’une année sur l’autre (20).
1.2.1.2/ Pathogénie
in
La résistance
c
Les rotavirus sont excrétés dans les fèces des animaux infectés, et la transmission est principalement
fécale‐orale (20, 60). Les signes cliniques apparaissent 1 à 3 jours post‐infection et durent de 5 à 9
jours. L’excrétion du virus commence à l’apparition des signes cliniques et continue pendant 3 à 7
jours dans les fèces des veaux malades. Les vaches peuvent être infectées subcliniquement, et
excréter de manière intermittente le virus au cours de la gestation, et principalement au cours de la
parturition. C’est la source d’infection la plus com
v
L’environnement peut également être une importante source d’infection. Les rotavirus peuvent
survivre dans l’eau pendant plus de deux semaines à 23°C, et des mois dans de l’eau ou le sol à une
température inférieure à 5°C. Ils peuvent également survivre dans les fèces ou le fumier jusqu’à
m
Les cellules épithéliales des villosités du duodénum et jéjunum sont les premières cibles des
rotavirus, ensuite ce sont les cellules de la partie distale de l’intestin grêle puis du côlon. Les cellules
épithéliales infectées dégénèrent et finalement se lysent libérant ainsi une grande quantité de
matériel viral dans la lumière intestinale. Ces cellules épithéliales sont remplacées par les cellules des
cryptes, insensibles au virus (20, 60, 135). L’attaque est auto‐limitante étant donné que les cellules
cibles, les entérocytes, sont détruites plus vite qu’elles ne sont remplacées (20, 60). L’immaturité des
cellules des cryptes, incapables de sécréter des enzymes digestives, les empêchent d’absorber les
nutriments, les électrolytes et l’eau, et ne peuvent pas digérer le lactose car ne possèdent pas la
lactase, l’absorption est donc nettement diminuée. Les sécrétions intestinales sont augmentées,
conséquence de l’hyperplasie compensatrice des cellules des cryptes, du déséquilibre osmotique et
22

de l’activité entérotoxinogène de la protéine virale non structurale NSP4 (20). La diarrhée résulte
donc de cet accroissement des sécrétions intestinales, mais aussi de la malabsorption et maldigestion
0, 60). Enfin, les cellules se différencient (60).
1.2.1.3/ Clinique
retrouvent un état
énéral quasiment normal. Le rotavirus seul entraîne donc rarement la mort (60).
(135). Les signes cliniques
ont faiblesse, anorexie, hyperthermie et déshydratation (60).
1.2.2/
1.2.2.1/ Étiologie
des Coronaviridae (38). Les
oronavirus sont des virus sphériques à ARN monocaténaire (figure 3).
(2
La diarrhée peut survenir en 14 à 22 heures, mais les veaux atteints sont généralement âgés de 6 à
10 jours. La diarrhée est généralement transitoire, 3 à 4 jours après, les animaux
g
Les diarrhées dues aux rotavirus sont des diarrhées aqueuses de couleur jaune à blanchâtre, elles
sont moins graves cliniquement que les diarrhées dues aux coronavirus
s
Les coronavirus
Le coronavirus bovin a été découvert par Mebus et al. en 1972, et il est maintenant reconnu comme
une cause importante de diarrhée néonatale des veaux. Le virus peut également infecter le tractus
respiratoire, et est associé à la dysenterie hivernale (winter dysentery) des bovins adultes. Le
coronavirus bovin appartient au groupe antigénique 2 de la famille
c
23

Figure 3 : Modèle schématique d’un coronavirus (165)
Le coronavirus bovin est présent dans de nombreux pays, et est probablement présent dans le
monde entier. Le virus est bien répandu dans la population bovine, ce qui fait que des anticorps anti‐
coronavirus bovin peuvent être détectés chez la majorité des adultes. Le virus peut être détecté chez
des veaux diarrhéiques mais également des veaux sains, la prévalence allant de 8 à 69% et de 0 à
24% respectivement, avec une prévalence moyenne de 10 à 15% chez les veaux diarrhéiques et de
5% chez les veaux sains (60). Ils sont spécifiques d’espèce et fréquemment isolés avec d’autres
germes, notamment les rotavirus (38, 60). Les veaux atteints ont généralement entre 3 et 15 jours
0). (6
La maladie peut également se déclarer chez des veaux de 24 heures n’ayant pas pris leur colostrum,
mais aussi chez des veaux atteignant un âge de cinq mois. La contamination se fait par voie fécale‐
orale mais peut se faire par voie aérienne (20, 38). L’excrétion fécale commence 3 jours après
l’infection et s’étend sur une semaine, l’excrétion nasale débute 2 jours après l’infection et persiste
pendant deux semaines. Une fois infectés, les veaux excrètent des taux élevés de virus, et sont donc
des sources de contamination. L’infection persiste plusieurs semaines chez des veaux apparemment
guéris, et ceux‐ci continuent à excréter le virus à des taux plus faibles. Les infections persistantes
subcliniques sont communes à la fois chez les veaux nouveau‐nés et chez des veaux plus âgés, et
l’excrétion virale de ces animaux maintient un réservoir d’infection vis‐à‐vis des veaux sensibles. La
prévalence de la maladie est plus importante au cours de l’hiver, cela reflète la capacité du virus à
24

survivre dans des conditions climatiques froides et humides. La diarrhée se déclare souvent plusieurs
années de suite dans un même élevage, cela est dû à la capacité du coronavirus à rester viable dans
l’environnement d’année en année. Le coronavirus bovin est cependant un virus labile et la diarrhée
de l’hiver. Les
eaux nés de mères infectées ont un risque accru de contracter une diarrhée (20, 38).
1.2.2.2/ Pathogénie
ent les cellules épithéliales de la partie distale des villosités intestinales de l’intestin
rêle distal (38).
rrhée,
ntraînant une perte d’eau et d’électrolytes. L’intestin grêle et le côlon sont atteints (38, 135).
1.2.2.3/ Clinique
statut immunologique du veau, et avec la dose infectante et la souche du virus, la diarrhée se
peut se déclarer même si les vaches ont été transférées dans un box de vêlage propre (20, 38).
D’autres sources sont les veaux porteurs sains et les mères porteuses saines (20, 38, 60). Le virus a
été détecté dans les fèces de plus de 70% de vaches saines (20, 38), malgré la présence d’anticorps
spécifiques dans le sérum et les fèces. Les veaux peuvent également s’infecter par l’excrétion des
vaches infectées. Le taux d’excrétion du virus augmente à la parturition et au cours
v
La pathogenèse du coronavirus bovin dans le tractus gastro‐intestinal du veau et les découvertes
pathologiques associées ont été décrites en détail par de nombreux chercheurs dont Mebus.
L’infection virale du tractus digestif débute par la partie proximale de l’intestin grêle et se répand
dans le reste du tube digestif. La réplication virale se déroule à la surface des cellules épithéliales, et
plus particulièrem
g
Les coronavirus abrasent les villosités intestinales, et les entérocytes sont remplacés par les cellules
des cryptes, cellules immatures (60, 135). L’abrasion des villosités due aux coronavirus est bien plus
importante que celle due aux rotavirus, les symptômes observés sont donc plus importants en cas de
coronavirose (60). Dans l’intestin grêle, ces changements conduisent à la fusion de villosités
adjacentes (38). Tout comme avec les rotavirus, c’est la diminution de la digestion et des capacités
d’absorption, avec en plus une hypersécrétion des cellules des cryptes qui conduisent à la dia
e
Les signes cliniques apparaissent après une phase d’incubation de 12 à 36 heures. Les diarrhées à
coronavirus sont des diarrhées aqueuses de couleur jaune à jaune verdâtre avec éventuellement du
mucus ou du sang (38, 60, 135). La gravité de l’entérite à coronavirus bovin varie avec l’âge et le
25

développant plus rapidement et étant plus grave chez les très jeunes veaux et chez les veaux privés
de colostrum (38).
Comme on l’a déjà dit, la maladie due aux coronavirus est souvent plus sévère que la maladie due
aux rotavirus, la principale lésion étant une entérocolite muco‐hémorragique (20). Les signes
cliniques sont une anorexie, une hyperthermie, une acidose, une hypoglycémie et une
déshydratation sévère (38, 60). Les infections sévères peuvent entraîner la mort suite à la
déshydratation, l’acidose, un choc, ou une défaillance cardiaque (20, 38).
1.2.3/ Les torovirus
Le torovirus bovin (aussi appelé Breda virus) a été détecté dans le monde entier, et a récemment été
impliqué en tant que cause importante de diarrhée néonatale du veau (figure 4). Il appartient à la
famille des coronavirus.
Figure 4 : Modèle schématique d’un torovirus (165)
Il n’a été que peu fréquemment rapporté en tant qu’agent causal car il est difficile de le reconnaitre
par microscopie électronique et il ne peut pas pousser sur culture cellulaire, les tests de diagnostic
immunospécifique de routine n’ont pas encore été développés non plus. Les études de laboratoire
utilisant les tests PCR (Polymerase Chain Reaction) l’ont impliqué comme unique agent pathogène
isolé dans 25 à 30% des échantillons fécaux de veaux de moins de six semaines ayant la diarrhée. Il a
aussi été retrouvé dans les fèces d’animaux asymptomatiques, ce qui suggère que l’épidémiologie est
26

relativement similaire aux rotavirus ou aux coronavirus, avec des porteurs asymptomatiques jouant
le rôle de réservoir d’infection au sein d’un troupeau. La transmission est fécale‐orale. C’est
principalement une maladie des veaux de moins de trois semaines, avec une diarrhée débutant 1 à 3
jours après la naissance. Cliniquement, il entraîne une diarrhée bénigne à modérée qui apparaît 24 à
72 heures post‐infection. Le virus infecte l’intestin grêle et le gros intestin, affectant les cellules
épithéliales en différenciation dans les cryptes et les villosités intestinales (20).
Les principaux virus responsables des diarrhées néonatales chez les veaux sont les rotavirus, les
coronavirus, et le torovirus, récemment impliqué en tant qu’agent causal. Leur pathogénie est
quelque peu différente, les rotavirus provoquant des lésions des parties distales des villosités
intestinales, les coronavirus atteignant l’ensemble de la villosité dans l’intestin et le côlon, ce qui
conduit à des diarrhées plus graves qu’avec les rotavirus. Les torovirus, eux, atteignent les cryptes
intestinales du jéjunum, de l’iléon et du côlon (figure 5).
Figure 5 : Schéma récapitulatif des principaux virus intervenant dans les diarrhées néonatales des
veaux, ainsi que leur mécanisme physiopathologique (165)
27

1.2.4/ Autres virus
Le virus de la diarrhée virale bovine (BVD) est une cause occasionnelle de diarrhée et de
thrombocytopénie chez les jeunes veaux non‐infectés in utero. Les anticorps colostraux protègent en
général les veaux contre l’infection par le BVD. On pense que ce virus peut aggraver les infections
d’autres agents pathogènes (20).
C’est un virus épithéliotrope qui se localise au niveau de la muqueuse intestinale principalement de
l’iléon et du côlon. Il provoque des lésions ulcéreuses et nécrotiques et une diarrhée liquide
hémorragique avec perte d’appétit, pétéchies des muqueuses nasales et oculaires, salivation,
ulcérations buccales et podales et larmoiement (135).
Les calicivirus, astrovirus, adénovirus, parvovirus et picobirnavirus ont tous été retrouvés associés à
la diarrhée néonatale des veaux, mais leur pathogénie est incertaine sur le terrain (20).
1.3/ Les parasites
1.3.1/ Cryptosporidium parvum
1.3.1.1/ Étiologie
Les cryptosporidies sont des protozoaires du Sous‐règne Protozoa, Embranchement des
Apicomplexa, Classe des Sporozoea, Sous‐classe des Coccidea, Ordre des Eimeriida, famille des
Cryptosporidiidae, Genre Cryptosporidium (60). Deux espèces de Cryptosporidium sont identifiées
dans les troupeaux : Cryptosporidium parvum dans les intestins et Cryptosporidium andersoni dans
l’abomasum. Il existe de nombreux sous‐génotypes de C. parvum, la majorité sont apparemment
hôte spécifiques et peuvent représenter des espèces distinctes (20).
Ce sont des parasites dont le cycle monoxène comporte trois phases (figure 6) : schizogonie,
gamétogonie et sporogonie. Les ookystes rejetés dans le milieu extérieur sont sporulés et
directement infestants, ils présentent une très grande résistance dans le milieu extérieur (les
ookystes de Cryptosporidium peuvent survivre dans l’eau pendant au moins douze semaines à 4°C)
mais aussi contre les désinfectants habituels, par contre ils sont détruits par le formaldéhyde 10% et
l’ammoniaque 5% après un contact de 18 heures (60).
28

Figure 6 : Cycle évolutif de Cryptosporidium parvum (26)
Les veaux sont généralement infectés entre une et quatre semaines d’âge. La cryptosporidiose se
produit moins fréquemment chez les veaux allaitants en pâture, mais quand ils sont atteints, les
symptômes sont plus sévères que chez les veaux de lait, avec un taux de mortalité allant jusqu’à 30%
(20).
Les anticorps neutralisants présents dans le colostrum ou le lait réduisent l’infection en immobilisant
le parasite, bloquant l’invasion, empêchant l’adhésion aux cellules de l’hôte, ou en ayant une
cytotoxicité directe sur les sporozoïtes. D’importants taux de mortalité ont été rapportés lorsqu’il y a
association de cryptosporidiose avec des taux faibles de sélénium, une mauvaise nutrition, la
présence d’infections intestinales concomitantes. La contamination est fécale‐orale, par ingestion
d’ookystes sporulés. La transmission peut être directe d’un hôte à un autre, par ingestion d’aliment
ou d’eau contaminée, et probablement aussi via les mouches. Les veaux sont sensibles à l’infection
au cours de leur naissance et peu après. Les sources de contamination sont nombreuses : les trayons
souillés des vaches, les vaches infectées, le matériel présent dans les bâtiments (abreuvoirs, seaux…),
mais aussi les contacts avec d’autres veaux (20).
La spécificité d’hôte est très faible, la cryptosporidiose est donc une zoonose (60).
29

1.3.1.2/ Pathogénie
Après ingestion, les ookystes libèrent des sporozoïtes qui se fixent aux microvillosités des cellules
superficielles de la muqueuse intestinale (figure 7) (20, 60). Ceux‐ci se transforment en trophozoïtes
qui sont invaginés par la membrane cytoplasmique et restent donc extracytoplasmiques (figure 8)
(20). Cette invasion entraîne la destruction de l’épithélium et une atrophie bénigne à modérée des
villosités. Cela empêche l’absorption intestinale et entraîne donc une diarrhée par malabsorption des
nutriments et malnutrition (20, 60).
Figure 7 : Début de développement d’un trophozoïte de Cryptosporidium parvum (26)
Figure 8 : Trophozoïte de Cryptosporidium parvum développé dans la membrane cytoplasmique d’un
entérocyte (26)
1.3.1.3/ Clinique
30

Les signes cliniques durent de 4 à 14 jours. La diarrhée qui en résulte est liquide et de couleur jaune
verdâtre à brun verdâtre, parfois muqueuse avec éventuellement du sang, du mucus ou du lait caillé
(60, 135). Les veaux infectés ne montrent souvent aucun autre signe clinique que la diarrhée, mais ils
peuvent présenter une apathie, une déshydratation, une hyperthermie, une faiblesse musculaire et
une anorexie (20, 60). C. parvum atteint principalement la partie distale de l’intestin grêle, mais on
retrouve également des lésions dans le caecum et le côlon, et occasionnellement dans le duodénum
(20). On a en général une faible mortalité mais une forte morbidité (135).
1.3.2/ Giardia duodenalis
Giardia est souvent retrouvé chez les veaux diarrhéiques en association avec d’autres agents
pathogènes, mais son implication en tant qu’agent pathogène isolé n’est pas clairement démontrée.
Les veaux infectés ont au moins deux semaines et même souvent plus d’un mois (20).
1.3.3/ Eimeria bovis et Eimeria zuernii
La coccidiose bovine est due principalement à deux agents pathogènes qui sont Eimeria bovis et
Eimeria zuernii. La transmission est fécale‐orale, l’excrétion débute à un mois et dure trois à quatre
mois (20). La coccidiose étant une maladie du veau plus âgé (de 3 semaines à 6 mois), elle est
simplement citée ici comme agent pathogène conduisant à une diarrhée.
1.4/ Diarrhées nutritionnelles
La diarrhée du veau pourrait également avoir une origine nutritionnelle, bien qu’il n’y ait pas d’étude
contrôlée le démontrant. Malgré tout, le fait de continuer de nourrir de façon normale un veau
infecté par des agents pathogènes intestinaux, entraîne une augmentation de la diarrhée et de
l’abattement (20).
On pense qu’il existe un risque de diarrhée lorsque la vache n’est pas complémentée en minéraux et
donc lorsque le lait a une carence en calcium non lié, ou encore lorsque le lait est trop riche en
31
matières grasses (supérieur à 50 g/l ou supérieur à 35% de la matière sèche), ou quand la
concentration en azote non protéique du lait augmente (135).
Les conséquences de la diarrhée sont une perte d’eau et d’électrolytes, ce qui conduit à une
hypovolémie, une hypoperfusion des tissus périphériques, une acidose par perte de bicarbonates et
production d’acide lactique, et une hypoglycémie par diminution de l’apport énergétique. Il faut faire
en sorte que le veau compense ces pertes, car si les pertes excèdent les apports, la déshydratation et
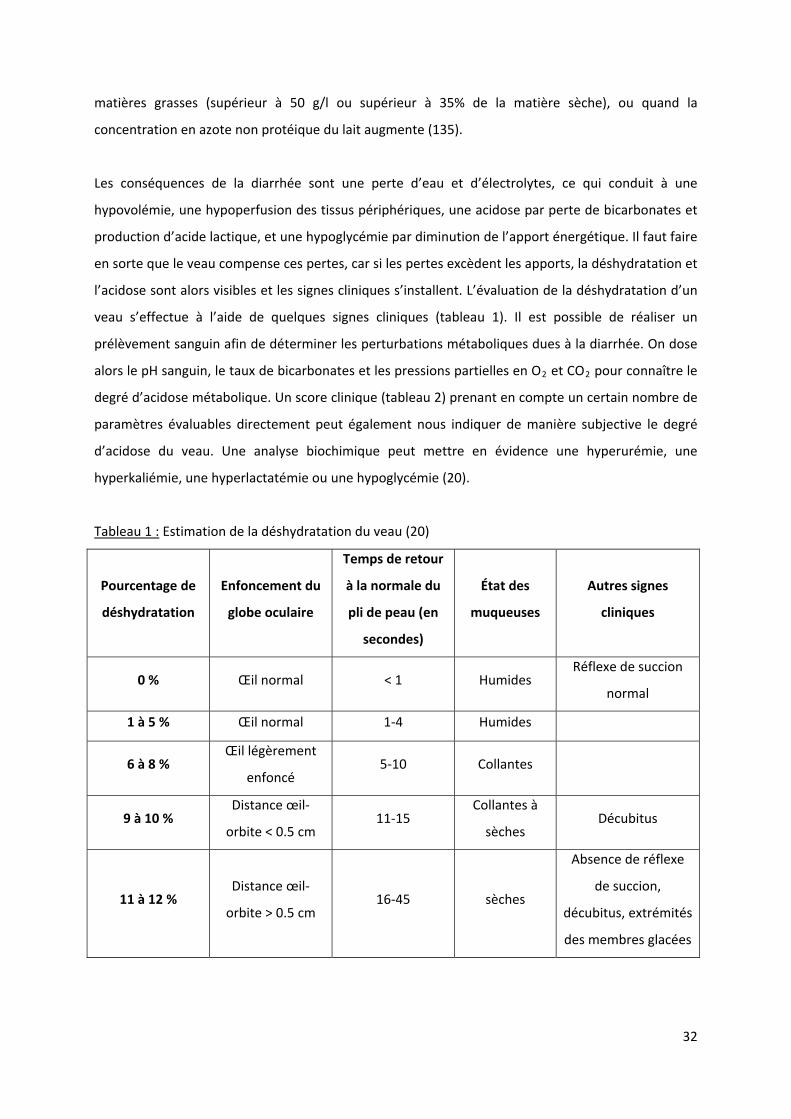
l’acidose sont alors visibles et les signes cliniques s’installent. L’évaluation de la déshydratation d’un
veau s’effectue à l’aide de quelques signes cliniques (tableau 1). Il est possible de réaliser un
prélèvement sanguin afin de déterminer les perturbations métaboliques dues à la diarrhée. On dose
alors le pH sanguin, le taux de bicarbonates et les pressions partielles en O2 et CO2 pour connaître le
degré d’acidose métabolique. Un score clinique (tableau 2) prenant en compte un certain nombre de
paramètres évaluables directement peut également nous indiquer de manière subjective le degré
d’acidose du veau. Une analyse biochimique peut mettre en évidence une hyperurémie, une
hyperkaliémie, une hyperlactatémie ou une hypoglycémie (20).
Tableau 1 : Estimation de la déshydratation du veau (20)
Pourcentage de
déshydratation
Enfoncement du
globe oculaire
Temps de retour
à la normale du
pli de peau (en
secondes)
État des
muqueuses
Autres signes
cliniques
0 % Œil normal < 1 Humides Réflexe de succion
normal
1 à 5 % Œil normal 1‐4 Humides
6 à 8 % Œil légèrement
enfoncé 5‐10 Collantes
9 à 10 % Distance œil‐
orbite < 0.5 cm 11‐15
Collantes à
sèches Décubitus
11 à 12 % Distance œil‐
orbite > 0.5 cm 16‐45 sèches
Absence de réflexe
de succion,
décubitus, extrémités
des membres glacées
32

Tableau 2 : Évaluation du degré d’acidose du veau par un examen clinique à distance (20)
2/ Diagnostic, traitements et prévention
2.1/ Diagnostic étiologique
Le diagnostic étiologique est utile pour traiter et prévenir les infections (régimes spécifiques pour les
infections bactériennes, établissement de méthodes de contrôle et développement de stratégie
vaccinale pour les virus). Le diagnostic de la salmonellose, de la cryptosporidiose ou de la giardiose
est particulièrement important en termes de santé publique (135).
L’épidémiologie et l’examen clinique orientent le clinicien vers tel ou tel agent pathogène.
Cependant, des examens de laboratoires à partir de fèces peuvent être envisagés afin de déterminer
avec certitude l’agent incriminé, et ainsi pouvoir mettre en place des mesures afin de protéger les
futurs veaux. Les techniques immunologiques sont de plus en plus utilisées (immunofluorescences et
ELISA (Enzyme‐Linked Immunosorbent Assay)), surtout si l’on suspecte une infection virale ou
parasitaire. Dans ce cas, on recherche la présence d’antigènes spécifiques dans les fèces. Ces
33

analyses sont réalisées en laboratoire, mais peuvent également être faites au chevet du malade à
l’aide de kits de diagnostic rapide qui permettent de mettre en évidence les coronavirus, les
rotavirus, les Cryptosporidium, les colibacilles F5 et CS31A. Enfin, il est possible d’envoyer au
laboratoire un veau vivant que l’on euthanasiera sur place afin de réaliser des prélèvements
directement après la mort (20).
2.1.1/ Bactéries
E. coli est présent habituellement dans le tractus gastro‐intestinal. Son isolement dans des
échantillons fécaux est donc sans signification sauf si les colibacilles isolés présentent des facteurs de
virulence compatibles avec la clinique (20). ECET peut être identifié par la présence de F5 en utilisant
un dosage immunologique tel que l’agglutination sur latex, le test ELISA, l’immunofluorescence et
l’agglutination sur lame (20).
Il existe de nombreuses méthodes pour détecter les salmonelles, et à l’échelle du troupeau il n’est
pas rare de détecter ou d’isoler la bactérie. On se pose la question de la relation entre la présence de
la bactérie et la déclaration de la maladie. En effet, on a observé que Salmonella peut être retrouvée
dans les fèces de veaux cliniquement sains. L’isolement de salmonelles dans des fèces de veaux
diarrhéiques est donc compatible avec un diagnostic de salmonellose, mais l’isolement seul sans
signe clinique ne permet pas d’établir un diagnostic (20).
Le diagnostic de laboratoire se fait sur des milieux d’enrichissement particuliers, tels que les
bouillons de sélénite ou de tétrathionate, afin de promouvoir la croissance des salmonelles en
inhibant les autres organismes de la flore fécale. Les échantillons sont ensuite mis sur des milieux de
culture spécifiques tels que la xylose lysine désoxycholate ou l’agar vert brillant (175). Les colonies
suspectes sont testées à l’aide d’une série de tests biochimiques et sérogroupées au moyen
d’antisera sérogroupe‐spécifiques. Ce sérotypage est réalisé par des laboratoires de référence.
Les méthodes de diagnostic rapide sont les tests ELISA et les PCR (Réaction de Polymérisation en
Chaîne). Un enrichissement préliminaire est souvent effectué dans les deux méthodes pour
augmenter la sensibilité de la détection. Les tests ELISA ont une sensibilité rapportée de 59% et une
spécificité de 97.6% sur des cultures enrichies (125, 183). La PCR conventionnelle et la PCR en temps
réel ont également été développées pour la détection des salmonelles dans les fèces. La PCR en
temps réel réduit les limites de détection (avec une sensibilité allant de 97.1 à 100% et une spécificité
allant de 91.3 à 100%) et de temps (obtention des résultats en 52 à 54 h) en comparaison avec les
34

techniques de PCR conventionnelle (sensibilité de 87% et spécificité de 95%) et de cultures des fèces
conventionnelles (sensibilité de 98% et spécificité de 99%, et obtention des résultats en 72 à 120 h),
mais la technique est plus compliquée à mettre en œuvre et nécessite donc du personnel qualifié
(18, 53, 186, 187).
Au cours d’un épisode de salmonellose, il n’est pas surprenant de trouver 70 à 80% de veaux
excrétant la bactérie. Une prévalence élevée d’excrétion de salmonelles va dans le sens d’un
diagnostic de salmonellose. Les cultures bactériennes sont préférés aux tests ELISA et PCR car ils
permettent un sérotypage et la réalisation d’un antibiogramme. Les veaux mourant de salmonellose
sont souvent bactériémiques. En conséquence l’isolement de salmonelles dans les organes
systémiques, au cours d’une autopsie est souvent considéré comme une preuve de salmonellose.
2.1.2/ Virus
Les virus sont habituellement identifiés par observation directe des fèces, par des tests
immunologiques, ou des tests aux anticorps fluorescents de la muqueuse intestinale. Le
développement récent de kits de diagnostic rapide relativement peu chers les rend attractifs, mais
selon Bradford et Smith (20), la sensibilité et la spécificité de certains de ces tests en limitent leur
utilisation.
2.1.2.1/ Coronavirus
Les méthodes de mise en évidence des coronavirus comprennent l’isolement sur culture cellulaire, la
microscopie électronique avec ou sans marquage immunologique, des tests immunologiques, des
techniques moléculaires incluant les tests d’hybridation, et la RT‐PCR (Reverse Transcriptase PCR).
L’isolement d’un coronavirus bovin par des techniques de cultures cellulaires est peu pratiqué en
laboratoire, car la technique est compliquée et nécessite d’avoir des virus vivants. La microscopie
électronique quant à elle, a été la technique standard de mise en évidence des coronavirus, mais elle
est difficile.
De nombreux tests ELISA ont été élaborés pour la détection des antigènes de coronavirus dans les
fèces, ce qui implique la commercialisation de nombreux kits de diagnostic rapide. L’utilisation
35

d’anticorps monoclonaux à la place d’anticorps polyclonaux est censée augmenter la sensibilité et la
spécificité de ces tests ELISA. La limite de détection des tests ELISA s’échelonne de 104 à 107 virions
par millilitres de fèces (20).
2.1.2.2/ Rotavirus
Les techniques d’isolement des rotavirus comprennent également la culture cellulaire, la coloration
d’anticorps fluorescent, la microscopie électronique, les tests immunologiques, l’électrophorèse, et
la RT‐PCR. Le rotavirus bovin est difficile à isoler par culture cellulaire à cause de la nature
cytotoxique des fèces et des résidus fécaux, et parce que les effets cytopathiques du virus ne sont
pas constants. La technique des anticorps fluorescents est simple, rapide et spécifique ; bien que les
antigènes des rotavirus soient généralement difficiles à mettre en évidence dans les 24 à 72 heures
après le début de la diarrhée parce que les cellules épithéliales infectées par le rotavirus sont
rapidement éliminées aux extrémités des villosités (20).
2.1.2.3/ Torovirus
Les méthodes diagnostiques pour détecter le virus sont la microscopie électronique,
l’immunofluorescence, les tests ELISA par capture d’antigènes, et la RT‐PCR (20).
2.1.2.4/ BVD virus
Rappelons que le virus BVD est rarement la cause de diarrhées néonatales, sauf cas particuliers de
veaux infectés in utero. On peut le mettre en évidence en isolant le virus, par RT‐PCR,
immunohistochimie, ou des tests ELISA par capture d’antigènes (20).
2.1.3/ Protozoaires
Les Cryptosporidium peuvent être mis en évidence par examen microscopique de frottis fécaux ou
préparation fécales, des tests immunologiques et PCR. L’immunofluorescence indirecte est plus
sensible que le test ELISA à antigène monoclonal (de 103 à 3x105 ookystes par gramme de fèces) (20).
36

2.2/ Traitement
En premier lieu, il est important d’isoler le veau malade, afin de limiter la dissémination des agents
pathogènes dans l’environnement et la contamination d’autres veaux. Le traitement repose sur une
fluidothérapie qui permet de compenser les pertes hydro‐électrolytiques dues à la diarrhée, corriger
l’acidose métabolique, corriger l’hypoglycémie et apporter au veau les besoins énergétiques
nécessaires. La réhydratation peut se faire par voie orale si le réflexe de succion est conservé, ou par
voie intraveineuse. Il peut être recommandé d’arrêter l’alimentation lactée. Les veaux présentant
une diarrhée ont souvent une prolifération de E. coli dans la lumière intestinale (quel que soit l’agent
pathogène responsable de la diarrhée), 30% des veaux présentant une atteinte de l’état général ont
une bactériémie, une antibiothérapie dirigée contre E. coli doit donc être mise en place (39). En cas
de cryptosporidiose, un traitement anticoccidien peut être administré. Un pansement intestinal peut
être donné (kaolin, etc.), afin de diminuer l’absorption des toxines, limiter les pertes hydriques,
ralentir le transit et protéger la muqueuse pour favoriser la cicatrisation. Les probiotiques tels que
Lactobacillus ou d’autres ferments lactiques peuvent être aussi administrés afin d’améliorer la
digestion et l’hygiène intestinale. Enfin on peut donner des anti‐inflammatoires non stéroïdiens
(AINS) pour limiter la production des médiateurs de l’inflammation et réduire les sécrétions
intestinales. Une supplémentation minérale et vitaminique peut être conseillée pour augmenter les
défenses immunitaires du veau (135).
Les salmonelles sont généralement résistantes à la pénicilline, l’érythromycine, et la tylosine. Il y a
une résistance de 60‐70% de S. Typhimurium (sérovar majoritairement identifié dans les élevages
français) à l’ampicilline, 3 à 6% aux aminosides, et une résistance émergente vis‐à‐vis des
céphalosporines de troisième génération et des fluoroquinolones. Parmi le sérovar Typhimurium, le
lysotype DT104 possède la particularité d’être pentarésistant (résistances à l’ampicilline/amoxicilline,
chloramphénicol/florfénicol, streptomycine/spectinomycine, tétracyclines et sulfamides), résistances
conférées par des gènes situés sur un fragment du chromosome appelé locus de multirésistance. Le
RESSAB a récemment détecté certaines souches de S. Typhimurium pentarésistantes ayant une
résistance supplémentaire au Triméthoprime (30). Ces résistances sont plus fréquentes chez les
jeunes veaux que chez les adultes, il faut donc faire une gestion raisonnée de l’utilisation
d’antibiotiques afin de limiter la sélection de bactéries résistantes aux antibiotiques (60). La
salmonellose due à S. Typhimurium DT104 étant une zoonose grave pour l’Homme, ces résistances
peuvent poser des problèmes thérapeutiques en médecine humaine. Pour certains scientifiques,
37

l’utilisation d’antibiotiques dans le traitement de la salmonellose dans les troupeaux est donc
controversée.
En conséquence, un traitement agressif à base d’antibiotiques est recommandé dans les stades
précoces de l’infection (39, 56, 183).
Alors qu’un grand nombre d’antibiotiques à spectre Gram négatif peuvent apparaître appropriés
pour le traitement de la salmonellose néonatale, l’utilisation de la plupart d’entre eux n’est pas
autorisée chez les veaux. Les salmonelles étant des bactéries intracellulaires facultatives, le choix
d’un antibiotique avec une bonne pénétration tissulaire et une action intracellulaire est
recommandé. Des études expérimentales ont montré que l’amoxicilline et le sulfamide
triméthoprime sont efficaces dans le traitement des infections à salmonelles par voie orale,
intraveineuse, et intramusculaire (69). De même, dans une autre étude, l’utilisation hors AMM
(Autorisation de Mise sur le Marché) de ceftiofur à 5 mg/kg a montré une atténuation des signes
cliniques et une réduction de l’excrétion fécale de salmonelles (56).
L’AMM est un accord donné à un médicament par une autorité compétente (Agence Française de
Sécurité Sanitaire des Produits de Santé) suite à l’examen d’un dossier scientifique assurant la
qualité, l’innocuité et l’efficacité des médicaments, pour pouvoir être commercialisé, lorsqu’un
laboratoire pharmaceutique désire mettre en vente un produit de santé. Cependant, il n’existe pas
toujours de médicament vétérinaire autorisé pour toutes les espèces ou toutes les maladies
auxquelles le vétérinaire est confronté. Dans ce cas, l’utilisation hors AMM est autorisée mais sous
certaines conditions. On recourt alors à un arbre décisionnel : on parle du principe de la "cascade".
Lorsqu’aucun médicament autorisé et approprié n’est disponible, il est possible de prescrire :
en première intention : un médicament vétérinaire autorisé pour des animaux d’une autre
espèce dans la même indication thérapeutique ou pour des animaux de la même espèce
mais dans une indication thérapeutique différente.
à défaut : un médicament vétérinaire autorisé pour des animaux d’une autre espèce pour
une indication thérapeutique différente.
à défaut : un médicament autorisé pour l’usage humain.
à défaut : une préparation magistrale vétérinaire (préparation extemporanée réalisée par un
pharmacien ou un vétérinaire).
38

Deux contraintes peuvent éventuellement d’appliquer :
en cas d’administration à des animaux dont la chair ou les produits sont destinés à la
consommation humaine, les substances actives doivent être inscrites dans les annexes I, II ou
III du règlement 2377/90 qui établit les procédures de fixation des limites maximales de
résidus (LMR) : l’usage de la cascade n’est possible que si une LMR a été définie dans le ou
les produits susceptibles d’être commercialisés (muscle, graisse, abats, lait,...).
les délais d’attente ne peuvent pas être inférieurs aux temps d’attente forfaitaires fixés par
l’arrêté ministériel du 16 octobre 2002, soit 28 jours pour la viande, 7 jours pour le lait, et 7
jours pour les œufs.
En ce qui concerne les temps d’attente, il est possible si le médicament prescrit est autorisé pour
l’espèce traitée mais pour une indication différente et si la posologie est inférieure ou égale à celle
prévue dans l’AMM, d’appliquer le délai défini par l’AMM. Dans tous les autres cas, les délais sont
fixés par le vétérinaire prescripteur et sous sa responsabilité (120).
Les AINS sont utilisés pour leur effet analgésique, anti‐inflammatoire et antipyrétique. Mais leur
efficacité thérapeutique dans le traitement de la salmonellose des veaux n’est pas documentée.
L’utilisation hors AMM de flunixine méglumine (2.2 mg/kg IV) et de méloxicam (0.5 mg/kg IV ou SC) a
été rapportée comme améliorant la guérison et réduisant la morbidité des veaux ayant une diarrhée
non spécifique (6, 167). Des précautions doivent être prises quant à l’administration d’AINS aux
veaux déshydratés. En effet, l’hypotension et la diminution de la perfusion rénale augmente le risque
de toxicité, la correction et le maintien de l’hydratation sont donc importants afin de prévenir les
effets secondaires éventuels (65). De plus, les nouveau‐nés ont des systèmes rénal et hépatique qui
sont moins efficaces pour métaboliser et excréter les médicaments.
La guérison est améliorée par un environnement propre, sec, à une température ambiante
convenable, et par une supplémentation nutritionnelle.
2.3/ Prévention
Il est important d’identifier les facteurs de risque, aussi bien pour mettre en place des programmes
de prévention que pour faire face à la maladie. Les diarrhées néonatales sont d’étiologie
multifactorielle, plusieurs facteurs contribuent donc à l’émergence de la maladie et à sa persistance
dans un troupeau (20).
39

2.3.1/ Facteurs de risque
Il existe un grand nombre de facteurs de risque des diarrhées néonatales et de toute maladie
néonatale. Il peut s’agir de problèmes liés directement à la mère comme une perte de lait pré‐
partum (avec perte de colostrum), une maladie de la mère, une malnutrition, un stress important, un
âge avancé de la mère ou au contraire le fait qu’elle soit une primipare. Les problèmes péri‐partum
sont également des facteurs de risque : prématurité, dystocie, gestation prolongée, césarienne,
gémellité. Le post‐partum immédiat est aussi important, notamment si des manœuvres de
réanimation sont nécessaires, si le veau respire mal, s’il refuse de téter, s’il est incapable de se lever
après trois heures de vie, s’il n’élimine pas son méconium ou si la vache n’a pas de lait (135).
2.3.1.1/ Gestion du troupeau et environnement
L’alimentation, l’eau, les engrais naturels (lisier), le troupeau, les introductions, la faune sauvage, les
insectes, les humains ou l’équipement sont autant de sources de contamination. Les pratiques
d’élevage visent à réduire le risque d’exposition en diminuant la quantité de bactéries dans
l’environnement et en améliorant l’immunité de l’hôte. Comme il y a de nombreux points qui
permettent de prévenir l’introduction de salmonelles dans la ferme, il s’avère difficile de réellement
tous les contrôler. Par exemple, l’utilisation de fumier de volailles comme engrais entraîne une
augmentation de l’émergence de sérotypes moins communs de salmonelles dans les élevages de
plein air (104).
Les risques de diarrhées néonatales sont augmentés par une densité élevée d’individus dans un
bâtiment, notamment un nombre élevé de veaux dans un même box. Le climat au moment du vêlage
affecte la survie des agents pathogènes mais également le confort du veau, des abris dans la zone de
vêlage diminuent la mortalité due aux diarrhées des veaux. La plupart des agents pathogènes
peuvent survivre dans l’environnement pendant des mois ou des années dans des conditions
climatiques froides et humides, il faut donc changer d’endroit la zone de vêlage au cours d’une saison
sinon le taux de diarrhées augmente avec l’avancement dans la saison de vêlage (20). Le taux
d’ammoniac est associé au risque de diarrhée, en conséquence d’une mauvaise ventilation ou d’une
quantité insuffisante de paille (135, 151). La présence de moisissures dans l’environnement favorise
la prolifération des agents pathogènes.
40

Les élevages qui achètent des veaux pour en remplacer d’autres ont un taux de mortalité dû aux
diarrhées néonatales plus élevé. Les veaux achetés peuvent introduire de nouveaux agents
pathogènes, le stress dû au transport et à l’arrivée dans un nouvel environnement peut augmenter
l’excrétion, et prédisposer l’animal à contracter la maladie (20).
La gestion des troupeaux et l’environnement sont des facteurs de risque, bien qu’une étude de
Waltner‐Toews de 1986 ne montre pas d’association significative entre les pratiques d’élevage et le
risque de diarrhée. Par contre, des études précédentes ont montré que des veaux élevés en niches
individuelles avaient moins de problèmes de santé que les veaux élevés dans des enclos individuels
plus conventionnels (177).
Le traitement du cordon ombilical ne semble pas avoir d’effet sur la diarrhée (177).
La propreté des vaches est également un facteur de risque. La supplémentation des vaches en
tarissement en vitamines et minéraux est associée à un risque plus élevé de diarrhée (93).
Les élevages qui ne donnent pas de concentrés à leurs vaches ont un risque de diarrhées néonatales
plus élevé que les autres. De plus, l’ensilage de maïs est associé au risque de diarrhée. Cela peut être
expliqué par le fait que l’ensilage de maïs pourrait induire une augmentation transitoire de
triglycérides et d’urée dans le sang, ce qui peut contribuer au risque de diarrhée.
Les élevages ayant eu une forte incidence de diarrhée la saison précédente ont un risque plus élevé
de diarrhée néonatale que les autres. Cela peut s’expliquer par le fait que les conditions d’élevage
n’ont pas évolué entre les deux saisons, et que la concentration d’agents pathogènes demeure
constante dans l’environnement, ou alors par la transmission des agents pathogènes des animaux
plus âgés aux plus jeunes (93).
2.3.1.2/ Âge du troupeau
Les veaux nés d’un premier ou d’un deuxième vêlage ont un risque de mortalité plus élevé comparé à
ceux nés de vaches plus vieilles, et le risque de diarrhée chez les veaux nés de génisses est 3,9 fois
plus élevé que chez les veaux de vaches. Les génisses ont un risque de vêlage dystocique plus élevé,
un colostrum de moins bonne qualité et des aptitudes maternelles moins développées (20).
41

2.3.1.3/ Complications au vêlage
Les dystocies sont associées aux diarrhées néonatales, principalement dans les élevages intensifs et
sont un risque de mortalité en pré‐sevrage (40% des morts en pré‐sevrage sont des veaux nés de
dystocies). La dystocie est une source de stress et affecte la capacité du veau à boire son colostrum,
ce qui entraîne un taux sérique d’IgG plus faible. En conséquence, les veaux dystociques sont deux à
quatre fois plus sensibles aux maladies dans les quarante‐cinq premiers jours de vie. L’œdème de la
tête et de la langue, conséquence fréquente de la dystocie, les gênent pour téter. Ils sont faibles,
fatigués, restent couchés plus longtemps, ce qui augmente leur exposition aux agents pathogènes
fécaux. Les veaux maigres ou gras à la naissance ont un risque de mortalité plus élevé (93, 135).
2.3.1.4/ Gestion colostrale
De nombreuses études ont montré que l’échec du transfert passif de l’immunité conduit à un risque
élevé de diarrhée néonatale. Les immunoglobulines sont capables de passer la barrière intestinale du
veau uniquement pendant une durée limitée après la naissance, et le taux d’IgG sérique est
déterminé par l’état du veau au moment du vêlage, la durée d’ingestion du colostrum, et la quantité
d’immunoglobulines ingérées (20).
Nous reviendrons un peu plus loin sur les propriétés du colostrum, et la gestion de son
administration.
En conséquence de tous les facteurs de risque qui existent, il est difficile d’identifier séparément les
effets individuels de nombreuses pratiques d’élevage, et le plus souvent, plusieurs facteurs sont
associés, ce qui fait que les conseils aux éleveurs sur la gestion de l’élevage doivent être donnés
simultanément.
2.3.2/ Principes de prévention
Les principes de prévention des diarrhées néonatales sont les suivants :
‐ Réduction de l’exposition aux pathogènes
‐ Assurance d’une bonne prise colostrale
‐ Augmentation de l’immunité spécifique et non spécifique
Ces trois principes allant ensemble, il est important de ne pas en négliger un seul.
42

2.3.2.1/ Réduction de l’exposition aux agents pathogènes
Il est indispensable de prévenir l’infection. Cela passe par la gestion de l’hygiène de l’environnement.
Tous les agents pathogènes peuvent survivre dans l’environnement pendant des mois ou des années
dans des conditions d’humidité adéquate. Ils peuvent également survivre sur tout le matériel utilisé
(135).
Il faut donc favoriser un environnement propre et sec, une bonne administration des aliments, avec
des pratiques de stockage et de manipulation, de façon hygiénique (135).
Il est important de disposer d’un local de vêlage qui ne sert que dans ce but. La propreté de l’aire de
vêlage est très importante, la litière doit être changée entre chaque vêlage et le local désinfecté (20,
135). Pour les éleveurs qui mettent leurs vaches sur le point de vêler dans des boxes de vêlage pour
une meilleure surveillance, afin de réduire la contamination des boxes de vêlage, il faut limiter la
durée de séjour des vaches dans les boxes et garder une litière propre. Avant chaque vêlage, le pis et
la région périnéale de la vache doivent être lavés. Lavage et désinfection doivent être effectués entre
chaque lot de veaux, lorsqu’ils sont élevés par lots. Le point important du lavage est le lavage
physique : il faut frotter les surfaces pour en retirer les matières organiques, cela est préféré au
nettoyage à l’eau sous haute pression qui peut créer des aérosols, et donc favoriser la dissémination
et la contamination. Le nettoyage par frottement des surfaces avec de l’eau et du savon élimine 99%
de la charge microbienne sur des surfaces lisses, et 90% sur les surfaces rencontrées habituellement.
L’application de désinfectant après le lavage est indispensable pour éliminer les agents pathogènes
restant et pour prévenir leur prolifération. Le lavage physique ne peut être remplacé par l’application
de désinfectants en grande quantité (135).
Il est préférable de séparer les vaches des génisses jusqu’à ce que leurs veaux aient au moins un
mois. Il faut éloigner et isoler les animaux malades chroniques et les veaux faibles (135). On peut
également isoler les veaux. Cet isolement a pour but de les éloigner de l’exposition aux agents
pathogènes. Ceci, ainsi qu’une bonne gestion de l’environnement améliorent la protection donnée
par l’immunité maternelle en offrant une fenêtre plus large avant que la vaccination soit nécessaire.
Il faut exclure de l’alimentation des veaux le lait inutilisé qui peut rompre cette isolation en
introduisant des agents pathogènes et des antibiotiques qui vont altérer la flore naturelle qui se
développe chez le veau, ce qui les rendrait plus sensibles (29). Il faut isoler les veaux malades des
veaux sains.
43

Il faut également augmenter la résistance à l’infection. Cela suppose que la mère ait une nutrition
adéquate au cours de la gestation et notamment lors des deux derniers mois. Les éventuels
déséquilibres en vitamines ou oligo‐éléments doivent être palliés, et les mères doivent être
déparasitées (fasciolose, dicrocoeliose et autres parasitoses). Il faut respecter des bonnes conditions
d’hygiène de l’environnement (gestion de l’humidité, de la ventilation, de la température des locaux
et de la litière) et ne pas mettre en contact des veaux d’âge trop différent (gestion des veaux par lots
homogènes en classe d’âge). Les agents pathogènes étant principalement transmis de manière fécale
orale ou encore par voie aérienne, il faut faire en sorte d’avoir une litière propre, correctement et
régulièrement paillée. De plus les agents pathogènes résistent bien dans l’environnement il est donc
important d’effectuer une désinfection et un vide sanitaire des locaux (cela peut s’avérer intéressant
de connaître les agents pathogènes résidant dans l’élevage pour appliquer les mesures nécessaires à
leur éradication pour ceux qui sont résistants aux désinfectants classiques). Le matériel utilisé doit
aussi être régulièrement désinfecté et nettoyé (135).
2.3.2.2/ Administration du colostrum
Il est important d’administrer au veau un colostrum de bonne qualité et en quantité suffisante dans
les quelques heures suivant la naissance afin d’éviter l’échec du transfert passif de l’immunité. Tout
ce qui concerne le colostrum et son administration vont être vus dans une partie spécifique.
2.3.2.3/Amélioration de l’immunité spécifique et non spécifique
On a longtemps pensé que la vaccination des vaches avant vêlage améliorait les anticorps colostraux
vis‐à‐vis de ces antigènes spécifiques. Cela a d’ailleurs été bien démontré avec les vaccins contre les
diarrhées néonatales, qui permettent une augmentation des anticorps colostraux contre les agents
pathogènes spécifiques de la diarrhée néonatale tels que E. coli, les rotavirus, les coronavirus (112,
146, 147). Quelques recherches ont été faites sur d’autres vaccins, contre le virus du BVD, et leur
impact sur les anticorps colostraux. Bien qu’une étude démontre que les vaches vaccinées avec un
vaccin viral vivant modifié ont une amélioration de leurs anticorps colostraux (52), une étude récente
avec vaccination des vaches avec un virus inactivé ne donne pas les mêmes conclusions (122). Une
étude israélienne démontre même une diminution des anticorps colostraux lorsque les vaches sont
vaccinées avant le vêlage (22). Si les vaccins sont désignés comme une amélioration du transfert
44

colostral en anticorps, des études sont attendues afin de démontrer la capacité des vaccins à
apporter l’effet désiré.
Il y a également un facteur primordial à prendre en compte, il s’agit de la santé de la vache. Il paraît
évident que pour qu’un veau soit bien protégé contre les agents pathogènes, il faut que sa mère soit
en bonne santé, déparasitée, avec un état d’embonpoint suffisant. La gestion de la santé des veaux
passe aussi par la gestion de la santé des mères.
Interférence entre les vaccins et les anticorps maternels :
Une croyance bien établie en immunologie néonatale est que la présence d’anticorps maternels
bloque les réponses immunitaires associées à la vaccination. Cela est basé sur le suivi de l’évaluation
des titres d’anticorps d’animaux vaccinés. Il apparaît clair dans de nombreuses études que si l’animal
est vacciné en présence de haut taux d’anticorps maternels dirigés contre cet antigène, il ne va pas
augmenter son titre en anticorps après vaccination (21, 103). Néanmoins, des études récentes ont
montré la formation d’une réponse des lymphocytes B mémoires et d’une réponse immunitaire à
médiation cellulaire même en présence des anticorps maternels (112) quand ce sont des vaccins
atténués qui sont utilisés. Il apparaît donc que l’interférence des anticorps maternels avec les vaccins
n’est pas absolue. Il a été démontré que les vaccins contre les maladies qui entraînent une réponse
immunitaire à médiation cellulaire sont plus à même de stimuler une réponse immunitaire en
présence des anticorps maternels que ceux qui entraînent une réponse immunitaire à médiation
humorale.
Les anticorps maternels diminuent sévèrement l’efficacité des vaccins contre les rotavirus et les
coronavirus (43, 173).
Impact du stress :
Le stress à un impact sur le système immunitaire du veau, comme chez l’adulte par ailleurs. Il existe
plusieurs facteurs qui affectent le système immunitaire et qui sont spécifiques au veau. Les
conditions de vêlage ont un impact fort sur le système immunitaire du nouveau‐né à cause du
relargage de corticostéroïdes. De plus le nouveau‐né possède un nombre élevé de lymphocytes T
suppresseurs. Ces facteurs, avec d’autres, diminuent dramatiquement les réponses immunitaires
45

systémiques au cours de la première semaine de vie. Des recherches récentes ont démontré que
juste après la naissance, il y a une diminution de la réponse immunitaire jusqu’au troisième jour de
vie, et qu’au cinquième jour de vie la réponse immunitaire revient au niveau qu’elle avait au moment
de la naissance. La vaccination par voie parentérale au cours de cette période est donc déconseillée,
elle peut même avoir des effets indésirables (24). De plus, toute autre source de stress devrait être
proscrite chez le nouveau‐né, tels que la castration, l’écornage, le sevrage, et les déplacements.
La vaccination du jeune veau doit donc être soumise à réflexion. Différents types de vaccination sont
utilisés, sur les veaux ou leurs mères, et leur efficacité est attribuable à une interaction entre
plusieurs facteurs, incluant l’antigène, le type de vaccin (modifié, ou atténué), l’âge du veau, la
présence d’anticorps maternels, les facteurs de stress au moment de la vaccination, et l’exposition
aux agents pathogènes.
Différents types de vaccins :
D’après Pravieux et al. (127), en Amérique du nord et en Europe, de nombreux vaccins pour les
vaches contre les diarrhées néonatales ont été développés. Les vaccins vivants modifiés et les vaccins
inactivés, vaccins utilisés en France, ont montré qu’ils augmentent le titre en anticorps du colostrum
et du lait des vaches vaccinées. Dans la plupart des cas, deux primo‐injections sont faites quelques
semaines avant le vêlage, suivies par un rappel annuel juste avant le vêlage. La sécurité et l’efficacité
pour les vaches gestantes et les nouveau‐nés sont bien établies sur le terrain.
Il existe sept sérogroupes de rotavirus. Deux approches peuvent être envisagées vis‐à‐vis de
l’immunoprophylaxie contre l’infection de ce virus. La première approche consiste en la vaccination
orale des veaux nouveau‐nés, avec un vaccin vivant modifié. Les veaux commencent à avoir un taux
d’IgM détectable à partir de 4 à 6 jours post‐vaccination. Afin d’obtenir une meilleure réponse
immunitaire, le vaccin doit être administré oralement immédiatement après la naissance et avant
que le veau tête car le colostrum de la plupart des vaches contient des anticorps neutralisant les
virus ce qui interférerait avec la vaccination. La seconde approche consiste en la vaccination des
mères avec un vaccin vivant modifié ou un vaccin inactivé afin de stimuler la réponse immunitaire de
la vache et d’obtenir de hauts taux d’anticorps neutralisants spécifiques dans le colostrum et le lait
au cours des premiers jours de vie du veau. Les particules virales sont neutralisées dans la lumière
intestinale, ce qui prévient l’infection des entérocytes des villosités intestinales. Un avantage de cette
immunisation passive est que la protection croisée entre les sérotypes est moins un problème (20).
46

Kapil et al. démontrent dans une étude de 1993 que chez des veaux privés de colostrum et infectés
par un coronavirus virulent ou atténué, la réponse immunitaire (IgM > IgG1 et > IgA) est plus
importante dans le côlon (site primaire de l’infection) que dans l’iléon et le jéjunum, et d’autant plus
importante que la souche est virulente. Selon Kapil et al., ceci montre que la vaccination orale contre
les coronavirus à l’aide d’un vaccin atténué présente des limites et même des échecs (85). En France,
on utilise des vaccins avec des rotavirus vivants atténués pour la vaccination orale des veaux, ou des
vaccins avec des rotavirus vivants atténués ou inactivés pour la vaccination des mères.
Bien que Salmonella spp soit un agent pathogène important chez le veau, il existe peu d’études dans
la littérature autres que l’observation sur le terrain de l’efficacité des vaccins contre les salmonelles.
Il existe trois classes générales pour les vaccins contre les salmonelloses : vaccin tué, vaccin
subunitaire, et vaccin vivant atténué. La plupart des vaccins commercialisés dans le monde sont des
vaccins tués ; en France on utilise un vaccin inactivé.
L’efficacité des vaccins contre la salmonellose est discutée, des études expérimentales ayant montré
des résultats mitigés (40, 75). Les vaccins tués ont comme limite qu’ils ne présentent pas l’antigène
exprimé in vivo, et échouent également dans l’induction d’une immunité cellulaire et mucosale (40,
75, 95, 143). Sur le terrain, l’exposition à l’agent pathogène se fait le plus souvent dans les tout
premiers jours de vie, ce qui limite l’occasion de stimuler les mécanismes immunitaires acquis par la
vaccination des veaux (75). L’immunité passive acquise de vaches vaccinées avec un vaccin tué, par le
transfert de colostrum est limité. Néanmoins, une protection partielle a été rapportée lors de tests
expérimentaux (83, 110). Quelques chocs anaphylactiques ont été rapportés dans la vaccination avec
les vaccins tués. En général, la vaccination contre la salmonellose est effectuée dans des troupeaux
où l’on a une suspicion de salmonellose sur certains animaux.
Le niveau de protection passive des veaux via le colostrum de vaches vaccinées est contesté, de
nombreuses études prouvent qu’il est efficace, d’autres non. La durée de la protection passive liée au
colostrum est relativement courte, mais en considérant que de nombreux veaux sont exposés aux
salmonelles dans leur première semaine de vie, la protection colostrale peut être utile (20).
Il existe donc deux stratégies de vaccination. Soit on cherche à protéger les veaux vis‐à‐vis des agents
pathogènes via le colostrum avec des anticorps spécifiques, on vaccine alors les mères. Ou alors on
vaccine directement les veaux à l’aide d’un vaccin oral. Dans les deux cas il reste des études à faire
afin de prouver la réelle efficacité de ces deux modes de vaccination contre les agents pathogènes les
plus communs.
47

Intérêt de la vaccination :
Les diarrhées à ECET apparaissent au cours des trois premiers jours de vie, les veaux n’ont donc pas
le temps de développer une protection immunitaire en réponse à la vaccination. La protection est
donc apportée par la vaccination des vaches en fin de gestation, ce qui confère au colostrum un haut
taux d’anticorps anti‐E. coli F5. Selon Valente et al. (172), la vaccination des vaches contre E. coli F5
protège les veaux, tandis que les veaux de vaches non vaccinées tombent malades et meurent
beaucoup plus facilement. Il faut donc également s’assurer de la bonne prise de colostrum.
Une étude a montré que des IgG colostrales spécifiques provenant de vaches immunisées avec un
vaccin multivalent de 17 souches de bactéries pathogènes donnant une diarrhée, ont une forte
activité inhibitrice de croissance et de colonisation des pathogènes in vitro en s’agglutinant avec les
bactéries et en détruisant les parois, les IgG provenant de vaches non vaccinées étant incapables de
provoquer les mêmes résultats. En conséquence, les IgG spécifiques colostrales provenant de vaches
immunisées peuvent procurer une protection efficace ou même un traitement contre les diarrhées
bactériennes (185).
Une étude de Castrucci et al. (28) démontre que la contamination expérimentale par des rotavirus de
veaux nourris avec du colostrum de vaches vaccinées contre les rotavirus d’un côté et des veaux
nourris avec du colostrum de vaches non vaccinées de l’autre montre que les veaux dont les mères
sont vaccinées sont protégés contre les rotavirus.
La protection contre la maladie et l’excrétion par le veau débute avant que les IgA sécrétoires ne
soient produites, 10 jours après la vaccination (173), ce qui implique que le vaccin administré par
voie orale peut activer le système immunitaire inné dans le tractus intestinal et donc réduire la
maladie. Ces vaccins seraient d’une bien meilleure utilité dans les élevages dont le colostrum a des
faibles concentrations en anticorps contre les rotavirus et coronavirus.
Une étude de Parreño et al. (124) sur l’évaluation de la protection passive et du développement de la
réponse immunitaire systémique et mucosale des anticorps maternels acquis via le colostrum en
réponse à l’infection à rotavirus montre que les veaux ayant reçu du colostrum provenant de vaches
immunisées contre le rotavirus ont un titre sérique d’IgG1 plus élevé que les veaux ayant reçu du
colostrum de base et que les veaux n’ayant pas reçu de colostrum. De plus ces veaux ont contracté
une diarrhée moins longue que les autres, et une excrétion virale moins importante. Plus les veaux
ont d’IgG1, moins ils ont de cellules sécrétrices d’anticorps, et donc moins la réponse immunitaire
active est importante. Dans les plaques de Peyer, les anticorps sécrétés sont majoritairement les IgM
48

pour les veaux ayant reçu du colostrum et des IgG1 pour les veaux privés de colostrum. Les IgG1
systémiques acquises passivement par le colostrum passent dans la lumière intestinale afin de
procurer une protection passive (124).
Une étude américaine (177) montre que la vaccination des vaches est associée à une augmentation
de la morbidité en général au cours de l’hiver, dont la diarrhée. Il y a également une tendance pour
les élevages vaccinant en routine les veaux contre la diarrhée à avoir plus de veaux à traiter contre la
diarrhée que les autres élevages, mais les veaux sont traités plus jeunes et moins longtemps que les
veaux non vaccinés. Cela peut être dû soit au stress lié à la vaccination, ou simplement à un niveau
de surveillance des animaux plus élevé de la part des éleveurs.
Selon Lorino et al. (93), Bendali et al. montrent dans une étude de 1999 une association négative
entre la vaccination des troupeaux contre E. coli et la diarrhée des veaux, contrairement à d’autres
études plus anciennes (145, 151, 154).
Une étude de Lorino et al. (93) montre également que la vaccination des troupeaux contre le BVD ou
Clostridium perfringens semble diminuer le risque de diarrhée.
Une étude de Martín‐Gómez et al. (94) sur les ovins, dans laquelle des agneaux nourris avec un
colostrum issu de troupeaux immunisés contre la cryptosporidiose et des agneaux témoins sont
infectés expérimentalement avec des oocystes de C. parvum, montre que chez ces agneaux
l’excrétion d’oocystes est moins importante et plus courte que chez les agneaux témoins et que la
diarrhée est également moins sévère. De plus ces agneaux ont un gain de poids de 2 kg par rapport
aux témoins.
Besser dans une étude de 1992 démontre que les anticorps présents dans la lumière intestinale
proviennent du colostrum et lait ingérés mais aussi de la circulation sanguine du veau (IgG1). Les
anticorps sériques ayant une affinité de liaison plus importante que les anticorps colostraux ou
laitiers. Les deux types d’anticorps contribuent de manière significative à la concentration d’anticorps
IgG1 dans la lumière intestinale, mais les concentrations en IgG1 dérivées du lait ne sont pas
maintenues à un niveau constant entre deux repas (10).
De nombreuses études ont donc été réalisées afin de déterminer l’intérêt de la vaccination des
vaches ou des veaux contre les agents des diarrhées néonatales. Certaines sont contradictoires, mais
la plupart s’accordent à démontrer que la vaccination des vaches ou des veaux procure un avantage
pour les veaux pour la prévention et pour la lutte contre les diarrhées néonatales.
49

Gestion de la campagne de vaccination :
En général, il est recommandé de vacciner les mères entre six et trois semaines avant le vêlage, mais
chaque vaccin a son protocole propre (20, 135).
Apport d’anticorps spécifiques :
On peut également apporter un concentré d’anticorps spécifiques, produit commercial, qui confère
une bonne protection contre les entérites néonatales. Bien que des résultats mitigés concernant
l’efficacité des suppléments colostraux aient été observés (66), ils peuvent jouer un rôle important
dans la diminution de la mortalité ou la sévérité de la maladie chez les veaux démunis de colostrum,
mais ils ont un coût élevé, la vaccination des mères est donc en général préférée (20).
La vaccination des mères ne prévient pas l’infection des veaux, mais permet de diminuer les signes
cliniques, le portage et l’excrétion. La valeur acceptable de morbidité est de 10‐15% et la valeur
acceptable de mortalité est de 5% sur une saison de vêlage. On peut également apporter du
colostrum pendant les 3 premières semaines de vie afin de garder un taux d’anticorps intra‐luminal
élevé. Des sérums ou des vaccins peuvent être donnés par voie orale pour protéger la muqueuse
intestinale contre les attaques des agents pathogènes. L’antibioprévention est une autre mesure de
prévention, mais il est important d’en réduire autant que possible l’application afin de prévenir
l’apparition de souches résistantes (135).
3/ Immunité et colostrum
3.1/ Rappels sur le statut immunitaire du veau nouveau‐né
La placentation de la vache est de type syndesmochoriale, les éléments sanguins de la mère et du
fœtus sont séparés, ce qui empêche le passage des immunoglobulines maternelles vers le fœtus au
cours de la gestation. Le veau nait donc dépourvu d’immunité. Celle‐ci devra lui être apportée dès sa
naissance par le biais du colostrum de sa mère, on parle de transfert passif, qui confère une
protection immunologique pendant au moins 2 à 4 semaines de vie, jusqu’à ce que son propre
système immunitaire devienne fonctionnel. L’absorption de colostrum est donc indispensable pour le
50

veau et constitue une étape clé de la néonatalité. De plus, la quantité de colostrum est importante
de même que sa qualité (20).
Une étude de Chigerwe et al. (34) montre que le taux d’IgG sérique du veau avant la première tétée
est détectable (donc > 0.16 g/l) chez plus de la moitié des veaux de l’étude. Il n’y a pas de lien
apparent entre les anticorps sériques dirigés contre les agents infectieux communs qui peuvent être
transmis à travers le placenta et la détection de concentrations mesurables d’IgG sériques. Le sexe
du veau, le poids du veau à la naissance et la saison de vêlage ne sont pas des éléments influant sur
la détection des IgG sériques avant la prise colostrale.
Le développement du système immunitaire se fait petit à petit, à partir de sa conception jusqu’à l’âge
de six mois où il est mature (figure 9) (29).
Figure 9 : Développement de la réponse immunitaire chez le veau : de la conception à la puberté (29)
3.2/ Colostrogenèse, composition et rôles du colostrum
Le colostrum bovin est un mélange de sécrétions lactées et de constituants sériques, et plus
particulièrement d’IgG et autres protéines sériques, qui s’accumulent dans la glande mammaire peu
51
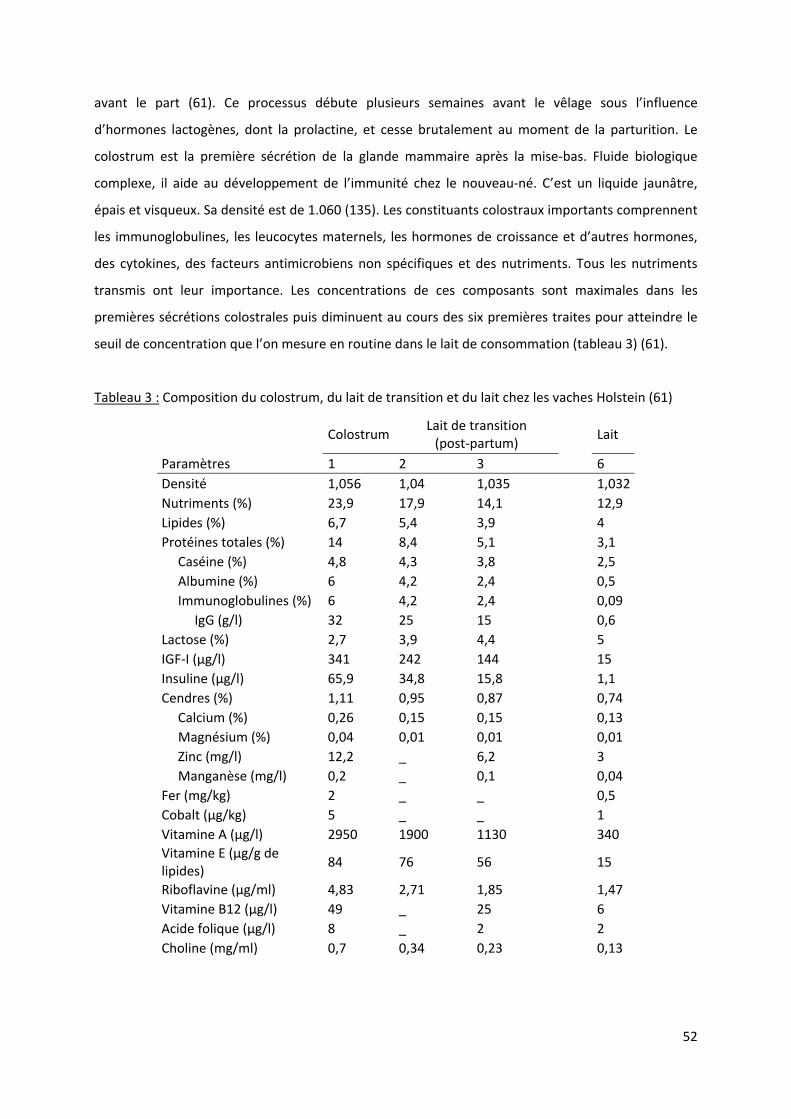
avant le part (61). Ce processus débute plusieurs semaines avant le vêlage sous l’influence
d’hormones lactogènes, dont la prolactine, et cesse brutalement au moment de la parturition. Le
colostrum est la première sécrétion de la glande mammaire après la mise‐bas. Fluide biologique
complexe, il aide au développement de l’immunité chez le nouveau‐né. C’est un liquide jaunâtre,
épais et visqueux. Sa densité est de 1.060 (135). Les constituants colostraux importants comprennent
les immunoglobulines, les leucocytes maternels, les hormones de croissance et d’autres hormones,
des cytokines, des facteurs antimicrobiens non spécifiques et des nutriments. Tous les nutriments
transmis ont leur importance. Les concentrations de ces composants sont maximales dans les
premières sécrétions colostrales puis diminuent au cours des six premières traites pour atteindre le
seuil de concentration que l’on mesure en routine dans le lait de consommation (tableau 3) (61).
Tableau 3 : Composition du colostrum, du lait de transition et du lait chez les vaches Holstein (61)
ColostrumLait de transition (post‐partum)
Lait
Paramètres 1 2 3 6 Densité 1,056 1,04 1,035 1,032 Nutriments (%) 23,9 17,9 14,1 12,9 Lipides (%) 6,7 5,4 3,9 4 Protéines totales (%) 14 8,4 5,1 3,1 Caséine (%) 4,8 4,3 3,8 2,5 Albumine (%) 6 4,2 2,4 0,5 Immunoglobulines (%) 6 4,2 2,4 0,09 IgG (g/l) 32 25 15 0,6 Lactose (%) 2,7 3,9 4,4 5 IGF‐I (µg/l) 341 242 144 15 Insuline (µg/l) 65,9 34,8 15,8 1,1 Cendres (%) 1,11 0,95 0,87 0,74 Calcium (%) 0,26 0,15 0,15 0,13 Magnésium (%) 0,04 0,01 0,01 0,01 Zinc (mg/l) 12,2 _ 6,2 3 Manganèse (mg/l) 0,2 _ 0,1 0,04 Fer (mg/kg) 2 _ _ 0,5 Cobalt (µg/kg) 5 _ _ 1 Vitamine A (µg/l) 2950 1900 1130 340 Vitamine E (µg/g de lipides)
84 76 56 15
Riboflavine (µg/ml) 4,83 2,71 1,85 1,47 Vitamine B12 (µg/l) 49 _ 25 6 Acide folique (µg/l) 8 _ 2 2 Choline (mg/ml) 0,7 0,34 0,23 0,13
52

Les immunoglobulines :
Les IgG, IgA et IgM représentent approximativement 85‐90%, 5% et 7%, respectivement, des
immunoglobulines colostrales, avec les IgG1 représentant à elles seules 80 à 90% du total des IgG
(89). Bien que les taux soient fortement variables entre les vaches et les études, une d’entre elles
rapporte que les concentrations colostrales pour les IgG, IgA et IgM sont respectivement de 7.5 g/l,
0.44 g/l et 0.49 g/l (119). Les IgG et plus particulièrement les IgG1 sont transférées du flot sanguin
vers le colostrum à travers la barrière mammaire par un transport spécifique : les récepteurs des
cellules épithéliales des alvéoles mammaires capturent les IgG1 des fluides extracellulaires par
endocytose, puis elles sont transportées et enfin relarguées dans la lumière des alvéoles (89). Les
cellules épithéliales alvéolaires cessent d’exprimer ce récepteur en réponse à des concentrations
croissantes de prolactine au début de la lactation (7). De petites quantités d’IgA et IgM sont issues
d’une synthèse locale par les plasmocytes de la glande mammaire (89). Bien que l’on ne connaisse
pas encore bien le mécanisme, on a aussi un transfert colostral d’IgE qui se produit et qui pourrait
jouer un rôle important en procurant une protection précoce contre les parasites intestinaux (164).
Les anticorps maternels transférés via le colostrum, activent et régulent la réponse innée des veaux
pour combattre une infection. Cette immunité passive est à double tranchant, car d’un côté elle
protège le nouveau‐né des infections, mais d’un autre côté elle interfère avec la capacité du veau à
développer une immunité face à un antigène vaccinal. La nature des immunoglobulines présentes
dans le colostrum dépend des micro‐organismes présents dans l’environnement et auxquelles les
mères ont été exposées, mais également des vaccinations que la mère aurait subies en fin de
gestation.
Les leucocytes maternels :
Le colostrum bovin d’une vache contient au moins 1.106 cellules/ml de leucocytes maternels
immunologiquement actifs et fonctionnels dès leur absorption par le veau, incluant les macrophages,
les lymphocytes B et T, et les neutrophiles (89, 91, 141). Au moins une partie de ces leucocytes
colostraux sont absorbés intacts à travers la barrière intestinale (150). Une étude de Liebler‐Tenorio
et al. de 2002 (92) montre que la voie préférentielle empruntée par ces leucocytes consiste à passer
à travers les plaques de Peyer dans le jéjunum et l’iléon. Une autre étude, de Reber et al. en 2006
(137), montre que les leucocytes maternels entrent dans la circulation du veau nouveau‐né, puis
circulent vers les tissus non lymphoïdes et tissus lymphoïdes secondaires, pour ensuite disparaître de
53

la circulation 24 à 36 heures après la prise de colostrum. Bien que leur importance fonctionnelle chez
les veaux ne soit pas mesurée en routine, des preuves précoces suggèrent que les leucocytes
colostraux améliorent la réponse des lymphocytes aux mitogènes spécifiques, augmentant la
phagocytose et la capacité d’éliminer les bactéries, et stimulant la réponse immunitaire humorale
(formation d’IgG) chez le veau (48, 50, 91, 136, 140). On présume que ces cellules ne sont pas viables
dans le colostrum pasteurisé ou dans les produits remplaçant le colostrum. Le rôle et la fonction des
leucocytes colostraux sont encore sources de recherches actives.
Les cytokines, facteurs de croissance et autres composants :
D’autres composants colostraux importants sont les facteurs de croissance, les hormones, les
cytokines, et des facteurs antimicrobiens non‐spécifiques. Les composés colostraux bioactifs
possédant une activité antimicrobienne sont la lactoferrine, le lysozyme et la lactoperoxidase (51,
123, 153). La lactoferrine est une protéine chélatrice du fer avec des propriétés antibactériennes et
antivirales. Elle exerce son effet bactériostatique en rendant le fer indisponible pour la multiplication
et la croissance bactérienne. De plus, des études montrent que la lactoferrine possède des propriétés
immunotropiques vis‐à‐vis des lymphocytes B, producteurs d’IgA, en provoquant la maturation de
ces cellules (87, 181). Ces substances biochimiques sont impliquées dans la défense antibactérienne
de la muqueuse intestinale et sont disponibles dans le colostrum. La lactoferrine et la
lactoperoxidase jouent un rôle non négligeable dans la défense immunitaire avec des concentrations
colostrales élevées (6 à 8 g/l) (87). Les oligosaccharides colostraux pourraient protéger contre les
pathogènes par inhibition compétitive aux sites de liaison présents sur la surface épithéliale de
l’intestin (130). De nombreux facteurs de croissance sont contenus dans le colostrum bovin, mais leur
fonction n’est pas totalement connue (voir tableau 1) (123). Le colostrum possède de remarquables
capacités de réparation et de croissance musculo‐squelettique. Des études ont montré que c’est
l’unique source naturelle de deux facteurs de croissance majeurs, les Transforming Growth Factor α
et β (TGF‐α et TGF‐β), et les Insulin Growth Factor (IGF) 1 et 2. Ces facteurs de croissance ont de
multiples effets régénérateurs qui s’étendent à toutes les cellules structurales du corps, comme les
cellules intestinales (171). L’IGF 1 pourrait être un régulateur clé dans le développement du tractus
gastro‐intestinal des veaux nouveau‐nés, stimulant la croissance de la muqueuse, les enzymes de la
bordure en brosse, la synthèse d’ADN intestinal, l’augmentation de taille des villosités, et
l’augmentation de la capture de glucose (9, 15, 25). L’inhibiteur de la trypsine, un composé retrouvé
à des concentrations cent fois plus élevées dans le colostrum que dans le lait, sert à protéger les IgG
et d’autres protéines contre la dégradation protéolytique dans l’intestin des jeunes veaux. Des
54

constituants additionnels du système immunitaire, tels que l’interféron, sont transférés au moyen du
colostrum (77).
Nutriments :
Bien que l’importance immunologique du colostrum soit fréquemment remise en cause, l’utilité
nutritionnelle du premier repas colostral ne devrait pas être contestée. La proportion des nutriments
dans le colostrum et le lait de vaches Holstein est en moyenne de 23.9% et 12.9%, respectivement
(voir tableau 1) (61, 89). Cette différence entre le colostrum et le lait est attribuée à l’augmentation
du contenu protéique du colostrum par rapport au lait, et plus particulièrement des
immunoglobulines et de la caséine (41). Le taux de graisses du colostrum (6.7%) est également plus
élevé que celui du lait (3.6%) (61).
L’énergie provenant des graisses et du lactose dans le colostrum est indispensable à la thermogenèse
et à la régulation de la température corporelle. Certaines vitamines et minéraux, dont le calcium, le
magnésium, le zinc, le manganèse, le fer, le cobalt, la vitamine A, la vitamine E, le carotène, la
riboflavine, la vitamine B12, l’acide folique, la choline et le sélénium sont aussi retrouvés en
quantités plus importantes dans le colostrum que dans le lait (tableau 1) (61, 130).
Outre le passage des anticorps et autres composés colostraux de la mère à son petit via l’absorption
intestinale, le colostrum joue d’autres rôles tels que la stimulation de la motricité intestinale et l’aide
à la colonisation du tube digestif par des bactéries opportunistes qui constitue une barrière
microbienne contre les agents pathogènes (135).
En plus de réduire le risque de morbidité et de mortalité néonatale, des bénéfices à long terme sont
associés au succès du transfert passif de l’immunité, comme la réduction de la mortalité des jeunes
veaux en post‐sevrage, l’augmentation du gain moyen quotidien (GMQ), la réduction de l’âge au
premier vêlage, et l’augmentation de la production laitière en première et seconde lactation (44, 55,
144, 180).
55

3.3/ Transfert passif de l’immunité : mécanismes et intervalle de transfert
Chez les veaux normaux, l’absorption colostrale est faite à travers les cellules intestinales épithéliales
par liaison au FcRn (récepteur néonatal au fragment Fc des immunoglobulines), et pinocytose en
utilisant un transport vacuolaire (5, 76). À partir de là, les immunoglobulines sont transportées à
travers la cellule et relarguées dans les vaisseaux lymphatiques par exocytose, après quoi elles
rentrent dans la circulation générale via le canal thoracique (158). Le mécanisme moléculaire dans
l’intestin du veau nouveau‐né qui conduit au transfert passif des immunoglobulines colostrales de la
lumière intestinale vers la circulation sanguine est également capable de transférer un bon nombre
d’autres macromolécules. La capacité de ce mécanisme est limitée quantitativement, le transfert de
ces macromolécules non immunoglobulines peut donc interférer avec le transfert des
immunoglobulines (11).
La période de transfert de l’immunité est relativement courte. En effet, dès que le tube digestif est
stimulé par l’ingestion de n’importe quel aliment, les entérocytes du veau nouveau‐né sont
remplacés par des cellules épithéliales matures, à partir de ce moment le passage des
immunoglobulines à travers la barrière intestinale n’est plus possible. La capacité d’absorption
commence à diminuer à partir de 6 heures et cesse totalement à 48 heures (5, 148). Selon Rischen, à
6 heures, il ne reste approximativement que 50% de la capacité d’absorption ; à 8 heures il n’en reste
que 33% et à 24h il n’y en a généralement plus (142).
La majeure partie du colostrum doit donc être absorbée dans les 12 premières heures et surtout
dans les 6 premières heures de vie. Dans une étude de Chigerwe et al., il est montré que le volume
de colostrum ingéré au cours du premier repas du veau et l’intervalle entre la naissance et le premier
repas sont des facteurs importants affectant les concentrations sériques en IgG des veaux nourris à la
main : pour les veaux ayant bu autant de colostrum qu’ils pouvaient dans les 4 heures suivant la
naissance, le risque d’échec de transfert passif de l’immunité est réduit. Par contre, ceux qui n’ont
pas ingéré au moins 3 l de colostrum dans les 4 premières heures de vie devraient recevoir du
colostrum par sonde œsophagienne (35). Ce qui est contradictoire avec une étude de Waltner‐Toews
et al., dans laquelle les méthodes d’administration du colostrum et notamment l’heure du premier
repas colostral ne montrent pas de relation avec les maladies des veaux (177).
Ensuite, les immunoglobulines ingérées procurent au veau une protection intestinale locale contre
les agents pathogènes. La durée de vie des IgG dans le sang est de 15 jours, celle des IgM de 4 jours
56

et celle des IgA de 2 jours. Le veau commence à synthétiser ses propres immunoglobulines, mais à
l’âge de 15 jours le taux d’immunoglobulines du veau est au plus bas (les anticorps maternels sont
diminués et les anticorps du veau n’ont pas encore atteint un taux suffisant). C’est une période
critique pendant laquelle le veau est vulnérable aux attaques par les différents agents pathogènes.
Les recommandations sur le plan de l’ingestion de colostrum sont de 6% du poids vif du veau dans les
4 premières heures de vie et de 10 à 15% du poids vif du veau dans les 24 premières heures de vie.
Les jours suivants, l’apport doit être de 10 à 12% du poids vif. Le premier repas doit impérativement
être pris par le veau, que ce soit en tétant directement sa mère, en tétant au biberon le colostrum
préalablement trait ou au moyen d’une sonde œsophagienne. Retarder l’heure de la première prise
colostrale peut seulement légèrement décaler la fermeture de la barrière intestinale à 36 heures
(162). Les éleveurs doivent planifier de nourrir tous les veaux nouveau‐nés dans les 2 premières
heures suivant le vêlage (au moins dans les 6 premières heures). Il est recommandé aux éleveurs
d’avoir une banque de colostrum, c’est‐à‐dire du colostrum de très bonne qualité congelé ou
réfrigéré (respectivement plusieurs mois de conservation et deux à trois jours) pour en avoir toujours
à disposition si un veau en a un jour besoin (135).
Mais l’apport d’immunoglobulines par voie orale reste intéressant après la fermeture de la barrière
intestinale car ils procurent une immunité locale dans la lumière intestinale. Les parties suivantes
discutent des facteurs influençant l’efficacité de l’absorption d’immunoglobulines, dont beaucoup
dépendent directement de la gestion de l’élevage.
3.4/ Administration du colostrum
La méthode de prise colostrale est importante à prendre en compte car cela peut influencer l’heure
du premier repas, le volume colostral consommé et l’efficacité de l’absorption des
immunoglobulines. Un taux élevé d’échecs de transfert passif de l’immunité a été rapporté dans les
élevages où l’on laisse le veau téter seul (12, 23). Cette observation peut être attribuée à un échec de
la prise volontaire de la part du veau d’un volume suffisant de colostrum et à un retard de cette prise
colostrale. Edwards et Broom (50) ont montré que 46% des veaux nés de multipares laitières ne
prenaient pas leur premier repas dans les 6 premières heures de vie, contre 11% pour les veaux nés
de génisses. Ce retard peut être la conséquence de nombreux éléments, comme la faiblesse ou des
blessures des vaches ou des veaux, comme les mammites ou autres maladies, une mamelle très
basse ou ayant des trayons trop larges, ou des vaches ayant des capacités maternelles faibles. C’est
57

pour ces raisons qu’il est recommandé que le veau soit isolé du troupeau au cours des 2 premières
heures suivant le vêlage, et qu’il soit nourri à la main avec un volume de colostrum connu en utilisant
un biberon ou une sonde œsophagienne (100). Dans un sondage, 68.1% en 1992, 70.5% en 1996 et
76.2% en 2002 des éleveurs américains disent utiliser un biberon ou une sonde œsophagienne pour
donner le premier repas au veau (115, 116, 117), indiquant que de plus en plus d’éleveurs font
attention à la prise de colostrum par le veau.
Bien que le repas soit plus rapide avec une sonde œsophagienne, l’administration du colostrum par
biberon est préférée car avec la sonde, le reflexe de déglutition n’est pas déclenché, conduisant à un
dépôt de lait dans les trois pré‐estomacs. Cependant cet élément n’est pas significatif car le passage
du colostrum des pré‐estomacs vers l’abomasum se fait en trois heures (90). Adams et al. (1)
rapportent que les veaux dont le colostrum a été administré au biberon plutôt qu’à la sonde
œsophagienne n’avaient des concentrations en IgG dans leur sérum que légèrement plus élevées,
avec des valeurs numériquement peu différentes et statistiquement non significatives. On conçoit
que les deux méthodes d’administration de colostrum permettent d’obtenir des taux acceptables de
transfert passif lorsqu’un volume suffisant a été délivré (1, 86).
Pour obtenir un bon transfert passif de l’immunité pour un veau Holstein de 43 kg, des chercheurs
ont calculé que l’éleveur doit administrer au moins 100g d’IgG dans la première tétée (41). Pour
connaître la quantité minimum de colostrum à donner, il faut évidemment connaître la
concentration en IgG. Par exemple pour un colostrum contenant 50 g/l d’IgG, l’éleveur doit donner
au moins 1.89 l de colostrum. Si le colostrum n’en contient que 25 g/l, il faut au moins 3.78 l de
colostrum pour le veau. Une étude de Besser et al. (12) démontre qu’il y a seulement 36% des
colostrums testés qui ont une qualité suffisante pour n’avoir besoin de donner à boire que 1.89 l au
veau. Quatre‐vingt cinq pourcents des colostrums testés avaient une qualité suffisante pour apporter
plus de 100 g d’IgG avec un repas de 3.78 l. Dans une étude de Morin et al., la concentration sérique
des veaux en IgG à 24 heures était significativement plus élevée pour des veaux nourris avec 4 l de
colostrum de bonne qualité au cours de la première heure suivant la naissance et avec 2 l à 12h (31.1
g/l) que pour des veaux nourris seulement avec 2 l de colostrum de bonne qualité au cours de la
première heure suivant la naissance et avec 2 l à 12h (23.5 g/l) (109). Une autre étude rapporte que
les veaux Suisse Brune nourris avec 3.78 l (contre 1.89 l) de colostrum au premier repas avaient un
GMQ plus élevé et une production laitière meilleure au cours de leur première et seconde lactation
(55). Dans un sondage, 26.1% en 1992, 35.9% en 1996 et 38.2% en 2002 des éleveurs américains
disent donner au moins 3.89 l de colostrum dans les premières 24 heures (115, 116, 117), montrant
58

que le volume de colostrum administré augmente mais que cela reste un facteur à améliorer pour de
nombreux éleveurs.
Il est important pour les éleveurs de bien penser à stériliser les sondes œsophagiennes (ou à avoir
plusieurs sondes) entre l’administration d’électrolytes à un veau malade et l’administration de
colostrum à un nouveau‐né.
3.5/ Facteurs influençant l’administration du colostrum
Présence du troupeau :
L’efficacité de l’absorption d’immunoglobulines est améliorée quand les veaux sont élevés au milieu
du troupeau (152). Mais la concentration sérique en IgG est suffisamment acceptable lorsqu’ils sont
élevés séparés du reste des animaux, il est donc en pratique recommandé que le veau soit isolé du
reste du troupeau dans les 2 premières heures de vie, ce qui limite les risques d’exposition aux
agents pathogènes du troupeau et de son environnement, et que le colostrum lui soit donné par
l’éleveur (100).
Désordres métaboliques :
Une diminution de l’absorption d’immunoglobulines dans les 12 premières heures a été rapportée
chez les veaux ayant une acidose respiratoire post‐natale, associée à un vêlage prolongé (13). Bien
que chez les veaux hypoxiques la prise de colostrum soit retardée, des études ont montré qu’il n’y a
pas de différence sur la capacité d’absorption entre les veaux hypoxiques ou normaux et qu’il n’y a
pas de différence de concentration sérique en IgG au moment de la fermeture de la barrière
intestinale (48, 169). Weaver et al. (179) suggèrent que l’augmentation du taux d’échecs de transfert
passif de l’immunité observé chez les veaux présentant une acidose métabolique ou respiratoire peut
être la conséquence du retard de la prise colostrale par le veau, et non une capacité d’absorption
réduite.
59

Stress thermique :
L’absorption d’immunoglobulines peut être diminuée lorsque les veaux nouveau‐nés sont exposés à
des températures extrêmement froides, sûrement en raison des effets directs sur l’absorption
intestinale et le transport des immunoglobulines, et indirectement de la capacité du veau à se lever
et se nourrir (121).
Contamination bactérienne du colostrum :
La présence de bactéries dans le colostrum peut avoir deux effets : soit les bactéries se lient à des
immunoglobulines libres dans la lumière intestinale, ou alors elles bloquent le transport des
immunoglobulines à travers les cellules épithéliales intestinales, ce qui a pour conséquence
d’interférer avec l’absorption passive d’immunoglobulines colostrales (78, 79, 126). Cet effet a été
démontré dans une étude récente contrôlée dans laquelle des veaux nouveau‐nés ont été nourris
soit avec 3.8 l de colostrum pasteurisé (à 60°C pendant 60 minutes) soit avec 3.8 l de colostrum frais,
contenant respectivement 813 Unité Formant Colonie (UFC)/ml et 40 738 UFC/ml (80). Bien que le
volume, l’heure du repas et la qualité du colostrum donné aux deux groupes ne soit pas différent, les
veaux ayant eu le colostrum pasteurisé présentent un taux d’IgG sérique à 24 heures d’âge plus élevé
que ceux ayant eu le colostrum brut (respectivement 22.3 g/l contre 18.1 g/l). Cette amélioration est
attribuée à la diminution des interférences entre les bactéries et les IgG dans le tube digestif,
conduisant à une meilleure efficacité de l’absorption d’IgG dans l’intestin chez les veaux nourris avec
le colostrum pasteurisé à 60°C pendant 60 minutes par rapport au colostrum brut. Des stratégies
visant à prévenir ou réduire la contamination bactérienne du colostrum sont abordées dans le
paragraphe suivant.
Stratégies visant à prévenir ou réduire la contamination bactérienne du colostrum :
Bien que le colostrum soit une importante source de nutriments et de facteurs immunitaires, il peut
aussi représenter un des risques les plus précoces d’exposition aux agents pathogènes tels que
Mycoplasma spp, Mycobacterium avium paratuberculosis, des colibacilles fécaux et des salmonelles
(159, 161, 178). Cette exposition est une préoccupation car des bactéries pathogènes dans le
colostrum peuvent être à l’origine de maladies telles que la diarrhée ou la septicémie. On s’en
préoccupe également car les bactéries contenues dans le colostrum peuvent interférer avec
60

l’absorption d’immunoglobulines (78, 79, 126). Les recommandations sont qu’un colostrum frais doit
contenir moins de 100 000 bactéries par ml et moins de 10 000 colibacilles par ml (100).
Malheureusement, de nombreux comptages bactériens dans des colostrums excèdent souvent ces
recommandations (126, 163). Dans une étude de Poulsen et al. sur un élevage dans le Wisconsin,
82% des échantillons testés dépassaient la limite supérieure de 100 000 UFC/ml (126). Les
paragraphes suivants indiquent les techniques de gestion pour réduire la contamination bactérienne
du colostrum.
Prévention de la contamination au cours du prélèvement du colostrum, de son stockage et
des procédés de son administration :
Les méthodes réduisant le risque de l’exposition des veaux aux agents pathogènes commencent par
l’exclusion des colostrums provenant de vaches infectées, et les mélanges de colostrums bruts. En
plus de cela, tous les éleveurs devraient prendre des précautions en ce qui concerne le prélèvement,
le stockage et l’administration de colostrum. Dans une étude de Quigley et al. (160), le comptage
total bactérien du colostrum prélevé directement dans la glande mammaire était très bas voire nul
(moyenne = 27.5 UFC/ml), la contamination bactérienne avait lieu au cours de la phase de transfert
du colostrum dans le seau (moyenne = 97.724 UFC/ml). Ces résultats démontrent l’importance de la
réduction de la contamination bactérienne du colostrum en préparant proprement les trayons avant
de récolter le colostrum, et d’avoir des seaux et des équipements propres et aseptisés pour contenir
le colostrum.
3.6/ Stockage du colostrum
Afin de réduire le risque de transmission de maladies associé à l’administration de colostrum, il faut
effectuer un nettoyage efficace de l’équipement utilisé dans la collecte et le stockage du colostrum,
être certain que le colostrum ne forme pas de grumeaux, vérifier que les températures des
réfrigérateurs pour la conservation du colostrum soient à des températures adéquates, limiter le
volume de colostrum à 2L par bouteille pour obtenir un refroidissement plus rapide, noter les dates
de récolte sur les colostrums réfrigérés et les jeter après deux ou trois jours de conservation, et avoir
des équipements uniquement dédiés à l’administration de colostrum.
Une étude de Holloway et al. a cherché à savoir si les concentrations sériques en IgG des veaux sont
affectées par la congélation à court terme du colostrum. Les résultats montrent que la
61

consommation de colostrum congelé et de colostrum frais aboutit aux mêmes concentrations
sériques en IgG chez les veaux (73).
Il existe plusieurs mesures visant à réduire la charge bactérienne dans le colostrum. Ce sont
l’utilisation de sorbate de potassium (160), la pasteurisation à la ferme (67), et l’utilisation de
colostrums artificiels.
Il est important de minimiser la croissance bactérienne du colostrum stocké, en effet, les bactéries
peuvent se multiplier rapidement si le colostrum est stocké à des températures ambiantes élevées
(160). Le colostrum doit être congelé ou réfrigéré dans l’heure suivant sa récolte, sauf s’il est donné
directement au veau. On considère que le colostrum peut être conservé congelé un an. Pour
décongeler le colostrum, il faut éviter les températures trop chaudes (ne pas réchauffer à plus de
60°C) afin d’empêcher la dénaturation des immunoglobulines colostrales (101). Un autre choix pour
les éleveurs qui veulent stocker du colostrum frais est la réfrigération, avec ou sans l’utilisation de
conservateurs tels que le sorbate de potassium (160). Les IgG d’un colostrum brut réfrigéré sont
stables au moins une semaine. Par contre, le comptage bactérien dans un colostrum brut réfrigéré
après deux jours de réfrigération peut atteindre des concentrations élevées (> 100 000 UFC/ml), ce
qu’on ne peut pas accepter. En comparaison, le comptage moyen bactérien reste inférieur à 100 000
UFC/ml après six jours de réfrigération quand le colostrum est conservé avec du sorbate de
potassium à 0.5% (160).
Un outil supplémentaire qui peut s’avérer utile afin de réduire la contamination bactérienne du
colostrum est la pasteurisation. Des études précoces ont essayé de pasteuriser le colostrum avec les
méthodes conventionnelles de hautes températures utilisées pour pasteuriser le lait (30 minutes à
63°C ou 15 secondes à 72°C). Ces procédés ne donnaient pas de résultats convenables, entraînaient
un épaississement du colostrum et des grumeaux ainsi que la dénaturation d’approximativement un
tiers des IgG (68). Des études plus récentes ont déterminé que l’utilisation d’une température plus
basse avec une durée plus longue (60 minutes à 60°C) pour pasteuriser le colostrum est suffisant
pour réduire de manière significative les agents pathogènes (dont E. coli, Salmonella Enteritidis,
Mycoplasma bovis et Mycobacterium avium paratuberculosis tout en conservant l’activité des IgG et
les caractéristiques physiques du colostrum (67, 81, 101). Dans une étude terrain récente, les veaux
nourris avec du colostrum pasteurisé (60 minutes à 60°C) ont montré une réduction de l’exposition
aux bactéries colostrales et un taux sérique en IgG à 24 heures significativement plus élevé par
rapport à des veaux nourris avec 3.8 l de colostrum brut (80). S’il est conservé dans un container
propre et fermé, la durée de conservation d’un colostrum pasteurisé et réfrigéré est d’au moins 8 à
62

10 jours (14). Les bénéfices au niveau de la santé et de l’économie à court et long terme de
l’administration de colostrum pasteurisé n’ont pas encore été décrits.
3.7/ Évaluation de la qualité colostrale et outils de mesure
Pour obtenir un très bon transfert passif d’IgG, le veau doit en priorité consommer une quantité
suffisante d’IgG et ensuite doit réussir à en absorber une quantité suffisante vers sa circulation. Les
principaux facteurs affectant la quantité d’IgG sont la quantité de colostrum et la qualité du
colostrum. La qualité du colostrum est la teneur en immunoglobulines du colostrum. Elle dépend de
nombreux facteurs dont la génétique de la vache, l’alimentation et le statut sanitaire. Le principal
facteur affectant la prise colostrale est la faiblesse post‐vêlage du veau, moment où le premier repas
colostral est préconisé.
Bien que l’on sache que le colostrum contient un grand nombre de facteurs immunitaires et nutritifs,
comme la relation entre la concentration en IgG et la santé du veau est mieux comprise, et comme
les IgG représentent plus de 85% des immunoglobulines colostrales totales, la concentration des IgG
colostrales est traditionnellement prise comme critère pour l’évaluation de la qualité du colostrum.
Un colostrum de très bonne qualité a une concentration en IgG supérieure à 50 g/l (100). La
concentration en IgG dans le colostrum varie de manière très importante d’une vache à l’autre. Dans
une étude récente, la moyenne de la concentration en IgG dans le colostrum de vaches Holstein
atteignait 76 g/l, mais les valeurs individuelles s’étalaient de 9 à 186 g/l (163).
De nombreux éleveurs continuent à avoir des pertes significatives de veaux associées à l’échec du
transfert passif de l’immunité. Aux États‐Unis, le taux de mortalité des veaux en pré‐sevrage est
estimé à 8‐11% (115, 116, 117). Une mauvaise gestion de la prise colostrale est le facteur clé
conduisant à ces pertes excessives. Dans une étude, 41% de 2177 veaux prélevés et âgés de 1 à 2
jours avaient un taux d’IgG sériques inférieur à 10 g/l (115). Il a été estimé qu’approximativement
31% des mortalités de veaux en pré‐sevrage durant les 3 premières semaines de vie des veaux
étaient dues à un échec de transfert de l’immunité passive (180). Ces études montrent la nécessité
pour les éleveurs d’améliorer la gestion de la prise colostrale dans les élevages.
On définit que des veaux ont eu un échec de transfert de l’immunité passive si la concentration
sérique en IgG est inférieure à 10 g/l, quand le sang est prélevé entre 24 et 48 heures d’âge (115,
63
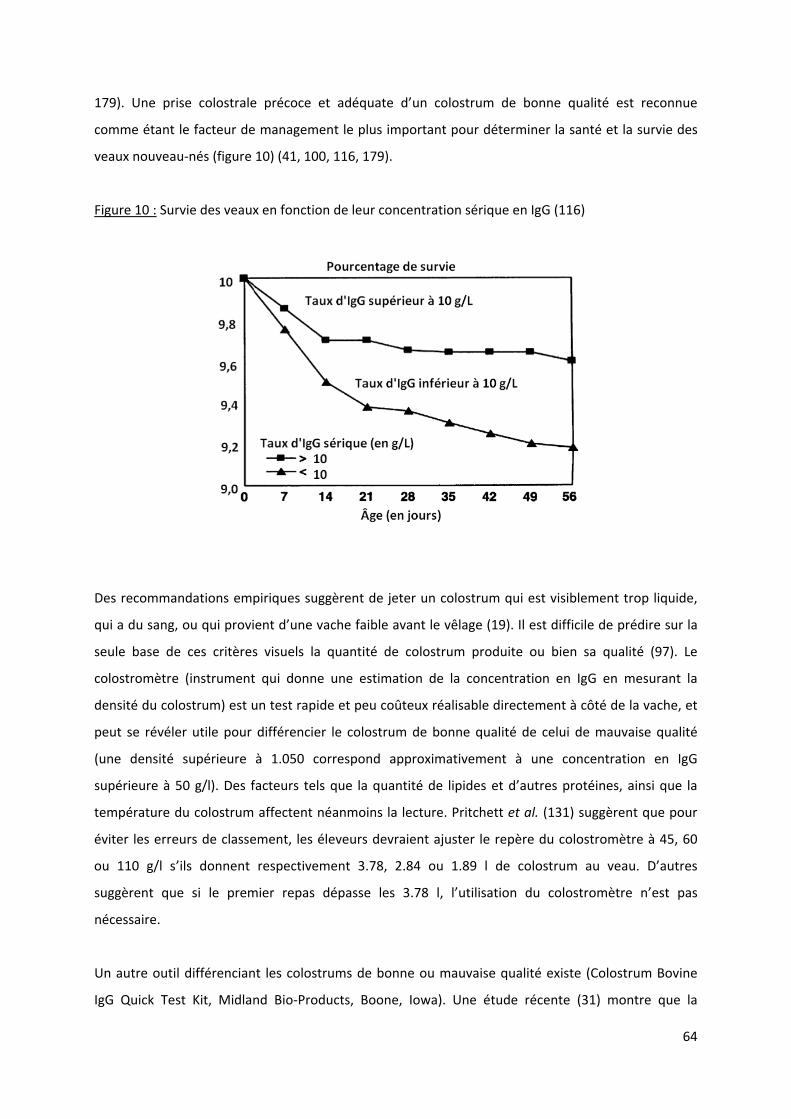
179). Une prise colostrale précoce et adéquate d’un colostrum de bonne qualité est reconnue
comme étant le facteur de management le plus important pour déterminer la santé et la survie des
veaux nouveau‐nés (figure 10) (41, 100, 116, 179).
Figure 10 : Survie des veaux en fonction de leur concentration sérique en IgG (116)
Des recommandations empiriques suggèrent de jeter un colostrum qui est visiblement trop liquide,
qui a du sang, ou qui provient d’une vache faible avant le vêlage (19). Il est difficile de prédire sur la
seule base de ces critères visuels la quantité de colostrum produite ou bien sa qualité (97). Le
colostromètre (instrument qui donne une estimation de la concentration en IgG en mesurant la
densité du colostrum) est un test rapide et peu coûteux réalisable directement à côté de la vache, et
peut se révéler utile pour différencier le colostrum de bonne qualité de celui de mauvaise qualité
(une densité supérieure à 1.050 correspond approximativement à une concentration en IgG
supérieure à 50 g/l). Des facteurs tels que la quantité de lipides et d’autres protéines, ainsi que la
température du colostrum affectent néanmoins la lecture. Pritchett et al. (131) suggèrent que pour
éviter les erreurs de classement, les éleveurs devraient ajuster le repère du colostromètre à 45, 60
ou 110 g/l s’ils donnent respectivement 3.78, 2.84 ou 1.89 l de colostrum au veau. D’autres
suggèrent que si le premier repas dépasse les 3.78 l, l’utilisation du colostromètre n’est pas
nécessaire.
Un autre outil différenciant les colostrums de bonne ou mauvaise qualité existe (Colostrum Bovine
IgG Quick Test Kit, Midland Bio‐Products, Boone, Iowa). Une étude récente (31) montre que la
64

sensibilité et la spécificité de ce test pour identifier les colostrums de mauvaise qualité sont
respectivement de 0.93 et 0.76. Cette mauvaise spécificité de ce test immunologique classerait un
colostrum de bonne qualité sur quatre comme étant de mauvaise qualité, ce qui n’est pas
acceptable. Une autre limite de ce test est qu’il ne donne qu’une réponse positive ou négative, mais
ne donne pas d’information sur la concentration en IgG.
Une étude de Chigerwe et al. montre que l’administration à la sonde œsophagienne de 100g d’IgG
colostrales n’est pas suffisante pour obtenir un transfert passif adéquat des IgG colostrales, le veau
doit avoir ingéré de 150 à 200 grammes d’anticorps au cours de sa première journée de vie pour
avoir un taux sanguin suffisant pour le protéger. De plus, l’utilisation d’une sonde œsophagienne
pour l’administration de 3 l de colostrum aux veaux dans les 2 heures suivant la naissance est
conseillée (32).
On peut aussi déterminer la qualité d’un colostrum à l’aide d’un pèse colostrum dont le marqueur,
en fonction de la densité du colostrum (et donc de sa qualité) flotte plus ou moins et nous indique à
l’aide de graduations si le colostrum est excellent, moyen ou mauvais. Ceci permet à l’éleveur
d’aviser et de prendre les mesures adéquates pour le transfert de l’immunité au veau (135). Les
résultats d’une étude de Chigerwe et al. (33) suggèrent que l’utilisation d’un refractomètre
conventionnel ou électronique est une méthode acceptable pour détecter les colostrums ayant des
faibles concentrations en IgG, bien qu’avec les deux instruments les concentrations soient
surestimées. L’utilisation du pèse‐colostrum n’est pas conseillée à cause de sa faible sensibilité.
On peut également doser directement les immunoglobulines chez le veau. Le dosage des
immunoglobulines sanguines du veau âgé de 2 à 7 jours reflète la quantité d’immunoglobulines bue.
Plusieurs techniques de dosage sont possibles. On peut effectuer un dosage quantitatif des
immunoglobulines sériques en laboratoire par des méthodes sérologiques d’agglutination, de
neutralisation, de fixation du complément ou par des techniques ELISA. On peut également
déterminer la quantité d’immunoglobulines par immunodiffusion radiale (RID : évaluation de la
quantité d’immunoglobulines après diffusion sur gel de gélose à l’aide d’antisera spécifiques). Des
tests semi‐quantitatifs de précipitation des immunoglobulines au sulfate de sodium ou au sulfate de
zinc permettent aussi de connaître la quantité d’immunoglobulines sériques. Il existe également un
test de coagulation au glutaraldéhyde.
On peut aussi doser les protéines totales (PT) et l’albumine sériques, le taux de protéines peut à lui
seul donner une idée du transfert de l’immunité passive (même si la différence avec l’albumine
65

donne un résultat plus précis). Enfin, il est possible de doser les gamma‐glutamyltransférases
sériques qui sont présentes en grande quantité dans le colostrum de bovins (135). La mesure des
protéines totales au refractomètre est un outil simple, rapide et peu coûteux sur du long terme qui
permet aux éleveurs de mieux gérer le colostrum et le transfert passif de l’immunité. Dans une étude
récente sur 185 veaux, la mesure des protéines totales a une bonne corrélation avec la concentration
sérique en IgG (mesurée par RID (R2 = 0.72)) (98). Calloway et al. (27) rapportent que la
concentration en PT mesurée au refractomètre à 50‐52 g/l minimum conduisait à un taux sérique
d’IgG de 10 g/l au moins (Se > 0.8 ; Sp > 0.8 ; proportion de classements corrects > 0.85).
S’il est déterminé qu’un nombre inapproprié de veaux ont un échec de transfert passif de l’immunité,
le vétérinaire et l’éleveur se doivent d’investiguer afin d’identifier et corriger les causes à la base de
cet échec. En plus de tester ses veaux pour vérifier le succès du transfert d’immunité, l’éleveur peut
également périodiquement envoyer les échantillons de colostrum congelé dans un laboratoire pour
analyses. Le but étant d’avoir un comptage bactérien total inférieur à 100 000 UFC/ml et un
comptage total de coliformes inférieur à 10 000 UFC/ml (100).
Un bon transfert passif de l’immunité n’a pas seulement un impact sur la morbidité et la mortalité
chez le jeune veau (17, 142, 144), mais il a aussi un impact positif à long terme sur la santé et la
production (45, 55, 182).
3.8/ Facteurs de modification du colostrum
De nombreux facteurs influencent la quantité et la qualité de colostrum produit. Il peut y avoir une
perte de lait avant le vêlage (veaux voleurs, sphincters lâches) qui entraîne une diminution de la
teneur en immunoglobulines du colostrum à la naissance. De plus, la teneur en immunoglobulines
est plus élevée à partir de la troisième lactation (135). Le niveau de corticostéroïdes néonataux doit
être élevé pour augmenter l’absorption colostrale (148). Le stress lié au froid, la naissance
prématurée, la césarienne et les dystocies inhibent la sécrétion de cortisol néonatal et diminue donc
l’absorption de colostrum. L’administration de corticostéroïdes aux veaux prématurés devrait
augmenter leurs chances de survie (149).
Un colostrum riche en cellules sanguines peut exacerber une diarrhée causée par des bactéries gram
négatif (140). La contamination du colostrum est associée à la récolte et au stockage du colostrum.
66

Le mélange de colostrums augmente le risque de salmonellose en disséminant les bactéries dans un
volume de colostrum plus grand qui va donc contaminer un plus grand nombre de veaux (174). Le
mélange de colostrums réduit également l’efficacité du transfert de l’immunité et compromet donc
ultérieurement l’immunité du veau.
Certains facteurs affectent directement la qualité du colostrum, tels que la race ou l’âge du troupeau,
et sont difficiles à changer pour l’éleveur. Mais de nombreux autres facteurs, tels que la vaccination
des vaches, la durée de la période de tarissement, et le moment du prélèvement du colostrum sont
plus aisément modulables. Cette partie passe en revue les différents facteurs affectant la qualité du
colostrum et discute de l’intérêt des tests rapides de mesure de qualité colostrale.
Race :
Des études comparatives ont démontré qu’il peut y avoir une composante raciale à la qualité du
colostrum (71, 111). Dans une étude de Guy et al., la concentration en IgG1 était plus élevée dans les
troupeaux allaitants (113.4 g/l) que dans les troupeaux laitiers (42.7 g/l) (71). Dans une autre étude,
les Prim’Holstein ont produit un colostrum avec une teneur totale en IgG (5.6%) qui est
numériquement plus faible que chez les Guernesey (6.3%) et les Brunes Suisses, et statistiquement
plus faible que chez les Ayrshire (8.1%) et les Jersey (9.0%) (111).
De manière générale, les vaches allaitantes ont une concentration d’IgG plus importante que les
vaches laitières. Les différences de race peuvent être attribuées à des différences génétiques ou à
des effets de dilution, ou les deux.
Âge du troupeau :
La plupart des études rapportent une tendance pour les vaches plus âgées à produire un colostrum
de meilleure qualité, probablement parce que ces animaux ont eu une plus longue période
d’exposition aux agents pathogènes spécifiques de l’élevage (108, 111, 128, 170). Par exemple, Tyler
et al. (170) rapportent que le taux d’IgG dans le colostrum de vaches en première, deuxième ou
troisième lactation était respectivement de 66, 75 et 97 g/l. Dans la même étude par contre, chez des
vaches de race Guernesey en première, seconde et troisième lactation, il n’y avait pas de différence
(valeurs respectivement de 119, 113 et 115 g/l).
67

Les primipares et les vaches au second vêlage ont une concentration colostrale d’IgG plus faible que
chez les vaches ayant eu plus de veaux, ce qui fait que le taux d’IgG sérique retrouvé chez ces veaux
est inférieur à celui de veaux nés de vaches de troisième vêlage ou plus. L’âge du troupeau peut donc
être un facteur de modification du colostrum, mais les autres facteurs interviennent également.
Alimentation en pré‐partum :
Les études montrent généralement que le taux d’IgG colostraux n’est pas affecté par l’alimentation
en pré‐partum (16). Lacetera et al. (88) rapportent que des vaches supplémentées par injections de
sélénium et vitamine E en fin de gestation produisent un volume de colostrum plus important que
des vaches non supplémentées, quand toutes les vaches sont nourries avec un aliment pré‐partum
déficient en vitamine E et sélénium. Ce traitement n’a cependant pas de répercussion sur le taux
d’IgG.
Saison de vêlage :
Certaines études ont montré une association entre l’exposition à des températures élevées en fin de
gestation avec un colostrum de mauvaise qualité, dont les concentrations en IgG et IgA sont plus
faibles, et dont les taux de protéines totales, caséine, lactalbumine, lipides et lactose sont diminués
(108, 114). Ces observations peuvent être attribuées aux effets négatifs du stress dû à la chaleur sur
la prise alimentaire, conduisant à des restrictions alimentaires, une circulation sanguine réduite dans
la mamelle conduisant à un transfert d’IgG et de nutriments réduit du flot sanguin vers la mamelle,
ou un défaut de réaction immunitaire des plasmocytes de la glande mammaire produisant les IgA
(114).
Quantité de colostrum :
Une étude de Pritchett et al. (128) démontre que les vaches produisant moins de 8,5 kg de colostrum
à la première traite avaient plus tendance à produire un colostrum de bonne qualité (> 50 g/l) que les
vaches qui en produisent plus (> 8.5 kg). Cette observation peut être attribuée à un effet de dilution,
bien que des études plus récentes rapportent qu’il n’y a pas de relation évidente entre la
concentration colostrale en IgG et la quantité de colostrum à la première traite (70, 97).
68

Mammite :
Les infections intra‐mammaires persistantes au cours de la période sèche sont associées à une
diminution du volume de colostrum produit, mais sans altération de la concentration en IgG1 (96).
Vaccination des vaches gestantes :
Bien que toutes les études ne montrent pas de résultats positifs, plusieurs études ont établi que la
vaccination des vaches gestantes trois à six semaines avant le vêlage augmente la concentration des
anticorps colostraux protecteurs, et augmente également le taux d’anticorps maternels chez les
veaux d’élevages vaccinés vis‐à‐vis de certains agents pathogènes dont Mannheimia haemolytica,
Salmonella Typhimurium, Escherichia coli, les rotavirus et coronavirus (3, 72, 83, 113, 176).
Durée de la période de tarissement :
La sécrétion d’IgG au niveau de la barrière de la glande mammaire commence approximativement 5
semaines avant le vêlage. Dans une étude de Pritchett et al., la durée de la période de tarissement
(moyenne de 57.5 ± 11 jours) n’a pas été associée avec des variations de la concentration colostrale
en IgG (128). Dans une autre étude (134), on a également rapporté que la qualité du colostrum
n’était pas différente entre des vaches ayant eu une période de tarissement de 28 ou 56 jours. Par
contre, les vaches ayant une période de tarissement excessivement courte (< 21 jours) ou sans
période de tarissement produisaient un colostrum avec une concentration colostrale en IgG
significativement plus basse (46, 134). De plus, la durée de cette période de tarissement peut
également affecter le volume de colostrum produit : dans une récente étude terrain de Grusenmeyer
et al., les vaches avec une période sèche courte (40 jours) produisaient 2.2 kg de colostrum de moins
que ne le faisaient les vaches avec une période sèche conventionnelle (60 jours) (70).
Prélèvement différé du colostrum :
La concentration colostrale en IgG est plus élevée immédiatement après le vêlage, et commence à
diminuer avec le temps si la prise colostrale est retardée. Dans une étude de Moore et al., retarder le
prélèvement de colostrum de 6, 10 ou 14 heures après le vêlage conduit à une diminution de 17, 27
ou 33% de la concentration colostrale en IgG respectivement (107).
69

Supplémentation des vaches en vitamine E et sélénium :
La question d’une relation entre la supplémentation en vitamine E et sélénium des vaches gestantes
et les IgG colostrales s’est posée. Des études de Bass et al., Arthington, et Zobell ont montré que la
supplémentation orale en vitamine E des vaches en fin de gestation n’affecte pas la concentration
colostrale en IgG, ni les concentrations sériques en IgG des veaux (4, 8, 190).
Une étude de Cipriano et al. montre qu’un déficit en vitamine E n’a pas d’effet sur la réponse du
système immunitaire (37). Par contre une étude de Reffett et al. montre qu’un déficit en sélénium
diminue la réponse du système immunitaire du veau (138).
3.9/ Colostrum artificiel
Les élevages peuvent occasionnellement rencontrer des périodes au cours desquelles une réserve
adéquate de colostrum frais ou stocké et de bonne qualité peut être insuffisante pour nourrir tous
les veaux nouveau‐nés. Pour accentuer ce problème, des éleveurs peuvent mettre de côté du
colostrum provenant de vaches qui ont été testées positives à M. avium paratuberculosis, à la
leucose bovine enzootique ou à mammite à M. bovis. Dans de telles circonstances, l’utilisation de
suppléments colostraux ou de colostrum artificiel permet d’offrir à l’éleveur un moyen convenable
d’augmenter le niveau d’immunité passive chez les veaux tout en réduisant le risque d’exposition aux
agents pathogènes via le colostrum. Les suppléments colostraux ou colostrums artificiels
commerciaux en poudre contiennent des immunoglobulines bovines qui sont dérivées du lait ou du
plasma. Il est recommandé que ces produits soient mélangés dans de l’eau (en accord avec la notice
d’utilisation), et donnés comme des repas séparés après chaque repas de colostrum naturel (100). Il
y a d’importantes différences entre les produits de supplémentation moins coûteux et les colostrums
artificiels qui sont plus onéreux. Les suppléments contiennent typiquement moins de 50 g d’IgG par
dose, ne contiennent pas de nutriments, et sont seulement à but de supplémentation et non de
remplacement du colostrum. S’il est donné seul, un supplément colostral amène à un taux sérique
d’immunoglobulines plus bas et un plus grand risque d’échec de transfert passif de l’immunité chez
les veaux que s’ils étaient nourris avec du colostrum brut (131). Si l’on donne déjà 3 à 4 l de
colostrum bovin de bonne qualité, il n’y a aucun intérêt à utiliser un supplément colostral (63, 188).
Par contre, les colostrums artificiels contiennent au minimum 100 g d’IgG par dose et sont également
une source nutritionnelle de protéines, d’énergie, de vitamines et minéraux et sont indiqués pour
remplacer complètement le colostrum maternel (132).
70

Des résultats d’études effectuées avec des colostrums artificiels démontrent une insuffisance de
transfert d’immunité puisqu’ils ne permettent pas d’atteindre les 10 g/l d’IgG sériques nécessaires au
veau (102, 132, 156, 163). Une étude américaine de Swan et al. (163) rapporte que 239 veaux nourris
avec un colostrum artificiel commercialisé avaient des concentrations sériques en IgG plus basses
(5.8 g/l) que 218 veaux nourris avec le colostrum de leur mère (14.8 g/l). Les taux de morbidité et de
mortalité ne présentent pas de différence significative entre les veaux nourris avec des colostrums
artificiels (morbidité = 59.6% ; mortalité = 12.4%) et ceux nourris avec le colostrum de leur mère
(morbidité = 51.9% ; mortalité = 10.0%). D’autres études ont montré des meilleurs taux de réussite
de transfert passif de l’immunité (concentration sérique moyenne en IgG > 10 g/l), particulièrement
lorsque les veaux ont été nourris avec deux doses de colostrum artificiel (82, 132). Dans une telle
étude, le taux moyen sérique en IgG à 24 heures pour des veaux nourris avec une dose (100 g d’IgG)
ou deux doses (200 g d’IgG) d’un colostrum artificiel, ou avec 3.78 l de colostrum maternel est
respectivement de 11.6, 16.9 et 27.2 g/l (62). L’administration de doses de colostrum artificiel plus
élevées permet d’augmenter le taux de succès de transfert passif de l’immunité, mais le rapport
bénéfice‐coût de cette pratique n’a pas encore été étudié. De façon similaire, l’efficacité et le rapport
bénéfice‐coût de l’utilisation courante de colostrum artificiel dans le contrôle de la paratuberculose
et d’autres maladies infectieuses n’ont pas non plus encore été étudiés. Les performances des
différents produits étant très variables, il revient aux vétérinaires de se tenir informés des études
portant sur les différents produits et leurs performances afin de choisir le plus approprié.
71

DEUXIEME PARTIE : Étude expérimentale
1/ Matériels et méthodes
1.1/ Présentation de l’étude
L’étude est une étude cas‐témoins qui comprend 10 élevages cas et 9 élevages témoins. L’âge des
veaux de l’étude est compris entre 0 et 18 jours avec la majeure partie des veaux entre 0 et 15 jours
(objectif initial). Dans chaque élevage cas, sont réalisés sur un ou deux veaux malades un kit de
diagnostic rapide de quatre agents pathogènes (E. coli F5, rotavirus, coronavirus et Cryptosporidium
parvum), ainsi qu’un prélèvement de fèces qui est envoyé au laboratoire départemental pour
dénombrement et typage de E. coli et recherche de salmonelles. Les mêmes opérations sont
effectuées pour un à deux veaux sains du même élevage. On considère ici un veau sain comme étant
un veau qui n’a jamais eu de diarrhée. Dans les élevages témoins, deux veaux sains sont choisis pour
les mêmes analyses. Ceci est retranscrit dans le tableau 4.
De plus, dans chacun des élevages, une prise de sang est effectuée sur les veaux dont les fèces sont
analysés, mais aussi sur un à trois veaux supplémentaires, sains, dont l’âge correspond aux critères
de départ pour doser les IgG sériques et les PT sériques. Ces éléments sont mentionnés dans le
tableau 5.
Tableau 4 : Présentation de la partie agents pathogènes de l’étude
10 élevages cas 9 élevages témoins
1 à 2 veaux malades 2 veaux sains 2 veaux sains
Kit de diagnostic rapide Kit de diagnostic rapide Kit de diagnostic rapide
Envoi de fèces pour analyse en laboratoire
Envoi de fèces pour analyse en laboratoire
Envoi de fèces pour analyse en laboratoire
72

Tableau 5 : Présentation de la partie immunité et transfert colostral de l’étude
10 élevages cas 9 élevages témoins
1 à 2 veaux malades 2 à 5 veaux sains 2 à 5 veaux sains
Prise de sang pour dosage IgG et PT
Prise de sang pour dosage IgG et PT
Prise de sang pour dosage IgG et PT
Dans chaque élevage également, un questionnaire a été rempli selon les réponses apportées par
l’éleveur ou selon les observations faites au cours de la visite. C’est en fait une visite d’élevage
simplifiée qui a été réalisée.
L’étude s’intéresse donc dans un premier temps à la prévalence des agents pathogènes des diarrhées
néonatales, puis au transfert passif de l’immunité de la mère au veau. L’identification des facteurs de
risque est un objectif secondaire auquel nous allons essayer de répondre.
1.2/ Choix des élevages
Les élevages cas sont des élevages au sein desquels un ou plusieurs veaux ont ou ont eu de la
diarrhée, les élevages témoins sont des élevages dans lesquels aucun veau n’a eu de diarrhée dans la
saison en cours.
La recherche des élevages sains s’est faite à l’aide des conseils des vétérinaires de la clinique
d’Aubusson, qui, connaissant leur clientèle, ont pu m’indiquer des élevages qui n’ont quasiment
jamais de diarrhée. Et c’est par un appel téléphonique que le statut de leur élevage s’est confirmé ou
infirmé. Puis j’ai pris rendez‐vous dans chacun des élevages pour aller y effectuer les prélèvements et
y remplir le questionnaire.
Pour la recherche des élevages cas, deux cas de figure se sont présentés. Soit l’éleveur appelait
directement la clinique, et donc cela me permettait d’aller faire la visite avec le vétérinaire ou bien
d’y aller un peu plus tard, soit j’appelais sur orientation des vétérinaires encore une fois des élevages
qui ont souvent des problèmes de diarrhée, pour leur demander s’ils en avaient à ce moment là.
73

1.3/ Choix des veaux
L’étude des agents pathogènes se limite à des veaux âgés de 0 à 30 jours, avec, quand cela a été
possible, des veaux de 0 à 15 jours. Des veaux diarrhéiques ainsi que des veaux sains sont prélevés.
Les animaux malades choisis doivent être en début d’évolution de la maladie, et ne doivent pas avoir
reçu de traitement antibiotique ni antiparasitaire.
L’étude de l’immunité et du transfert colostral est centrée sur des veaux âgés de 2 à 8 jours en
grande majorité, mais avec des âges de veaux allant de 1 à 18 jours.
1.4/ Réalisation des prélèvements
1.4.1/ Prélèvements de fèces
À l’exception de trois ou quatre individus en diarrhée, j’ai personnellement réalisé tous les
prélèvements de fèces de veaux diarrhéiques ou sains. La démarche a été la suivante : à l’aide de
gants propres en latex, j’ai stimulé le sphincter anal des veaux jusqu’à ce que les fèces arrivent, et je
les récupérais directement dans un pot à prélèvement stérile dès leur sortie.
1.4.2/ Prélèvements sanguins
Pendant que l’éleveur s’occupait de la contention du veau, j’effectuais le prélèvement sanguin sur
chaque veau à la veine jugulaire, à l’aide d’un Vacutainer® sur lequel était montée une aiguille à
prélèvement rose (18G) et un tube sec pour recueillir le sang.
1.5/ Matériel de recherche des agents pathogènes
1.5.1/ Tetrakits
Les tetrakits sont des tigettes tétravalentes (photo 1) Bio K 156 du laboratoire BioX Diagnostics. Elles
permettent de mettre en évidence la présence des rotavirus, coronavirus, de E. coli F5 et de
Cryptosporidium parvum dans les fèces de veaux par immunochromatographie latérale. Le test
74

s’effectue au chevet du veau, et les résultats sont disponibles en maximum 10 minutes, par simple
lecture des bandes de chaque tigette (figure 11).
Photo 1 : Tigettes d’immunochromatographie latérale (en rouge la tigette pour les rotavirus, en
jaune pour les coronavirus, en bleu pour E. coli F5 et en vert pour Cryptosporidium parvum) (57)
Les tigettes sont plongées flèches vers le bas dans un tube dans lequel on a mélangé un échantillon
de fèces à un liquide de migration.
Figure 11 : Lecture du résultat sur la tigette (57)
Si deux bandes apparaissent, le résultat est positif, si une seule bande apparaît le résultat est négatif,
et enfin si aucune bande n’apparaît le test n’est pas interprétable.
75

L’utilisation d’anticorps monoclonaux comme conjugués et pour capturer les agents pathogènes sur
la membrane (figure 12) permet d’obtenir une excellente sensibilité et spécificité (57), ce qui garantit
la fiabilité des résultats. Pour la détection des rotavirus, la spécificité est de 100% et la sensibilité de
96% ; pour les coronavirus la spécificité est de 98.7% et la sensibilité de 88.9%. Les tigettes ont une
spécificité de 98.5% et une sensibilité de 90% pour la mise en évidence de E. coli F5, et une spécificité
de 95.5% et une sensibilité de 94.1% pour Cryptosporidium parvum.
Figure 12 : Mode de fonctionnement des tigettes (57)
1.5.2/Analyses de laboratoire
1.5.2.1/ Dénombrement des E. coli dans les fèces
Le dénombrement des E. coli a été fait par le laboratoire départemental de la Creuse (LDA 23) à
partir des prélèvements de fèces effectués. Ces fèces ont été placées au réfrigérateur le plus
rapidement possible dès leur prélèvement, et conservées au maximum 3 jours avant analyse.
Milieu de culture utilisé :
Le milieu utilisé est la gélose TBX AGAR (Tryptone Bile X‐glucuronide AGAR : peptone, extrait de
viande, extrait de levure, chlorure de sodium), avec de l’eau physiologique stérile ou du Ringer
lactate en tubes de 9 millilitres et de 9,9 millilitres pour les dilutions.
76

Le mode d’action est le suivant : l’enzyme β‐D‐glucuronidase différentie E. coli des autres coliformes.
E. coli absorbe le substrat chromogénique 5‐bromo‐4‐chloro‐3‐indolyl‐β‐D‐glucuronide (X‐β‐D‐
glucuronide). L’enzyme rompt les ponts entre les chromophores 5‐bromo‐4‐chloro‐3‐indolyle et β‐D‐
glucuronide, les colonies d’E. coli sont alors colorées en bleu‐vert. La croissance de la flore
d’accompagnement gram positive est largement inhibée par la présence de sels biliaires.
Technique :
Il faut peser stérilement 1g de fèces préalablement homogénéisé dans 9 ml d’eau physiologique
stérile ou Ringer (dilution au 1/10e), on mélange le tout à l’aide d’un vortex, puis on réalise des
dilutions successives jusqu’à 10‐9 à partir de cette solution mère.
Ensemencement :
Avec une pipette stérile, on dépose dans trois boites de pétri stériles les solutions comme suit :
- Boite 1 : 1 ml de la dilution 10‐5
- Boite 2 : 1 ml de la dilution 10‐7
- Boite 3 : 1 ml de la dilution 10‐9
On ajoute 15 ml de milieu TBX en surfusion refroidi à 45‐50°C dans chacune des boites et on mélange
soigneusement et lentement l’inoculum. Puis on laisse solidifier.
Après refroidissement, on incube les boites retournées 18 à 24 heures à 35‐37°C.
Lecture :
Après 18‐24 heures, les colonies d’E. coli sont colorées en bleu‐vert. Il faut compter, pour chaque
boite de dilution, les colonies caractéristiques bleues, et on retient uniquement les boites contenant
au maximum 150 colonies. Le résultat final est obtenu en multipliant le nombre de colonies par la
dilution correspondante (par exemple si on a 5 colonies à la dilution 10‐5 cela donne 5x105 E. coli par
gramme de fèces).
77

Les exigences du laboratoire en terme de qualité sont telles que le résultat n’est considéré significatif
que si le prélèvement a été conservé à + 4°C et acheminé au laboratoire le jour même, ce qui n’a pas
été possible en pratique.
1.5.2.2/ Typage des E. coli
Milieux et réactifs :
Les milieux sont les géloses Minca et Polyvitex, les réactifs coagglutinés utilisés sont F41, F17 (FY), F5
(K99) et CS31A, et on utilise de l’eau distillée stérile.
Méthode :
Le test est réalisé à partir de colonies fraîches (24 heures) caractéristiques isolées sur boites de pétri
(gélose Hektoën (protéose peptone, extrait de levure, chlorure de sodium, thiosulfate de sodium,
sels biliaires, citrate de fer ammoniacal, salicine, lactose, saccharose, fuchsine acide, bleu de
bromothymol, agar agar) ou gélose Columbia qui est un milieu d’isolement servant de base pour la
préparation des géloses au sang, gélose riche qui peut être additionnée de nombreux suppléments,
et la rendre sélective à certains groupes bactériens (mélange de peptones, amidon, chlorure de
sodium, agar agar).
On repique 2 à 4 colonies sur gélose Minca enrichie et on incube pendant une nuit à 37°C. Après
agitation énergique au vortex et mise à température ambiante des réactifs, on met en contact sur
une lame, une goutte de chacun des 4 réactifs avec 3 ou 4 colonies de la souche à tester puis on
agite. On réalise des témoins à chaque jour d’utilisation : un contrôle réactif (une goutte d’eau
stérile avec une goutte de réactif) et un contrôle souche (une goutte d’eau stérile avec deux ou trois
colonies à tester pour vérifier qu’aucune agglutination n’apparait).
Lecture et interprétation :
- Réaction positive : apparition d’agglutinats en 30 secondes d’agitation avec un des quatre
réactifs et absence d’agglutination pour les deux contrôles.
78

- Réaction négative : absence d’agglutinat après agitation avec les quatre réactifs et absence
d’agglutination pour les deux contrôles.
La réaction est ininterprétable s’il y a apparition d’une réaction positive pour le contrôle souche,
dans ce cas on est en présence d’une souche auto‐agglutinable.
Le sérotype de la souche testée est celui pour lequel des agglutinats apparaissent.
1.5.2.3/ Recherche des salmonelles
La recherche de Salmonella dans les fèces utilise la norme AFNOR (Agence Française de
Normalisation) NFU47‐102 (isolement et identification des principales salmonelles chez les
Mammifères). Le LDA 23 est accrédité COFRAC (Comité Français d'Accréditation) Programme 116
pour cette recherche.
La technique utilise deux moyens de recherche. Une recherche par isolement direct, où l’on dépose
une goutte de fèces sur une gélose Hektoën puis on ensemence par stries ; et une recherche après
enrichissement, on met un gramme de fèces qu’on ensemence dans 10 ml de bouillon
d’enrichissement Müller Kauffmann (bouillon au tétrathionate, qui inhibe la croissance des
coliformes et de la plupart des bactéries du tube digestif, pour l’enrichissement des salmonelles à
partir de la viande, de la charcuterie et d’autres denrées alimentaires (extrait de viande, peptone de
viande, extrait de levure, chlorure de sodium, carbonate de calcium, thiosulfate de sodium, bile de
bœuf séchée)) et on incube 24 heures (à plus ou moins 3 heures) à 41,5°C, puis on repique sur milieu
sélectif (gélose Hektoën).
1.6/ Dosages des immunoglobulines G et des protéines totales
Le dosage des protéines totales est fait sur automate par méthode colorimétrique, selon le test du
Biuret (formation de complexes d’ions cuivriques avec la liaison peptidique, en milieu alcalin). Du
tartrate double de sodium et de potassium est utilisé pour limiter l’autoréduction du cuivre. Une
coloration violette apparait, due au complexe formé. Elle est proportionnelle à la concentration en
protéines dans l’échantillon. La lecture se fait par densité optique (58).
79

L’évaluation des IgG est faite par une électrophorèse des protéines sur gel d’agarose à 80 Volts
pendant 30 minutes. L’évaluation quantitative se fait par lecture de la plaque gel d’agarose à 595 nm.
La méthode est sensible jusqu’à 0.3 g/l (59).
1.7/ Outils statistiques
La partie analytique de l'étude s’attache à comparer la répartition des catégories exposé/non exposé
pour chaque exposition choisie en fonction du statut malade/non malade du veau.
Les outils statistiques utilisés afin de quantifier les associations et les tester statistiquement sont les
Odds Ratios (OR) et le test du Chi‐2. Les tests étaient dits « significatifs » si le p était inférieur au seuil
d’erreur de première espèce fixé à 5%.
2/ Résultats
Dans la suite de la thèse, nous appellerons malades les veaux qui présentent une diarrhée, et sains
les veaux qui ne présentent pas de diarrhée.
2.1/ Partie descriptive
2.1.1/ Dosage des IgG et PT
La partie de l’étude portant sur les immunoglobulines G et les protéines totales sériques des veaux
porte sur 74 veaux. Parmi ces veaux, 12 sont malades et 62 ne présentent pas de diarrhée (figure 13).
80
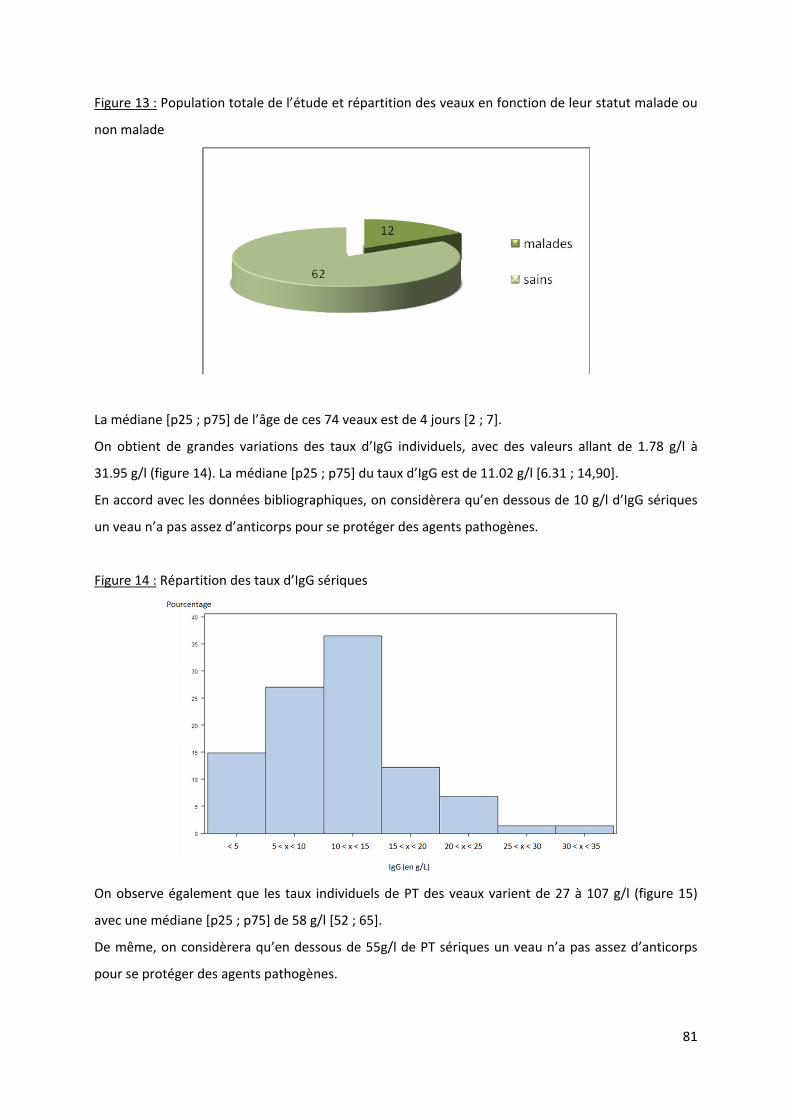
Figure 13 : Population totale de l’étude et répartition des veaux en fonction de leur statut malade ou
non malade
La médiane [p25 ; p75] de l’âge de ces 74 veaux est de 4 jours [2 ; 7].
On obtient de grandes variations des taux d’IgG individuels, avec des valeurs allant de 1.78 g/l à
31.95 g/l (figure 14). La médiane [p25 ; p75] du taux d’IgG est de 11.02 g/l [6.31 ; 14,90].
En accord avec les données bibliographiques, on considèrera qu’en dessous de 10 g/l d’IgG sériques
un veau n’a pas assez d’anticorps pour se protéger des agents pathogènes.
Figure 14 : Répartition des taux d’IgG sériques
On observe également que les taux individuels de PT des veaux varient de 27 à 107 g/l (figure 15)
avec une médiane [p25 ; p75] de 58 g/l [52 ; 65].
De même, on considèrera qu’en dessous de 55g/l de PT sériques un veau n’a pas assez d’anticorps
pour se protéger des agents pathogènes.
81

Figure 15 : Répartition des taux de PT sériques
2.1.2/ Concernant les diarrhées
2.1.2.1/ Description des diarrhées
Parmi les 10 élevages cas, 2 élevages avaient 2 veaux simultanément en diarrhée.
Les différentes caractéristiques des diarrhées sont présentées dans les deux graphiques suivants
(figures 16 et 17).
Figure 16 : Nombre de veaux en fonction de la consistance des diarrhées
82
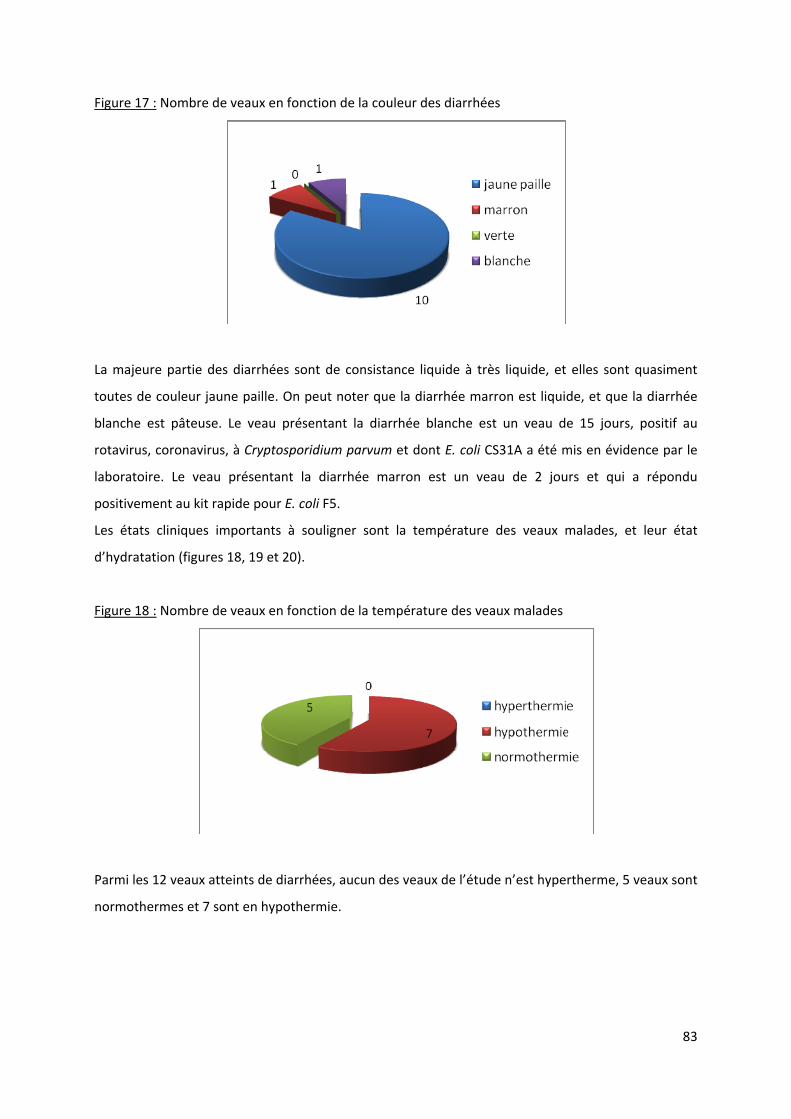
Figure 17 : Nombre de veaux en fonction de la couleur des diarrhées
La majeure partie des diarrhées sont de consistance liquide à très liquide, et elles sont quasiment
toutes de couleur jaune paille. On peut noter que la diarrhée marron est liquide, et que la diarrhée
blanche est pâteuse. Le veau présentant la diarrhée blanche est un veau de 15 jours, positif au
rotavirus, coronavirus, à Cryptosporidium parvum et dont E. coli CS31A a été mis en évidence par le
laboratoire. Le veau présentant la diarrhée marron est un veau de 2 jours et qui a répondu
positivement au kit rapide pour E. coli F5.
Les états cliniques importants à souligner sont la température des veaux malades, et leur état
d’hydratation (figures 18, 19 et 20).
Figure 18 : Nombre de veaux en fonction de la température des veaux malades
Parmi les 12 veaux atteints de diarrhées, aucun des veaux de l’étude n’est hypertherme, 5 veaux sont
normothermes et 7 sont en hypothermie.
83
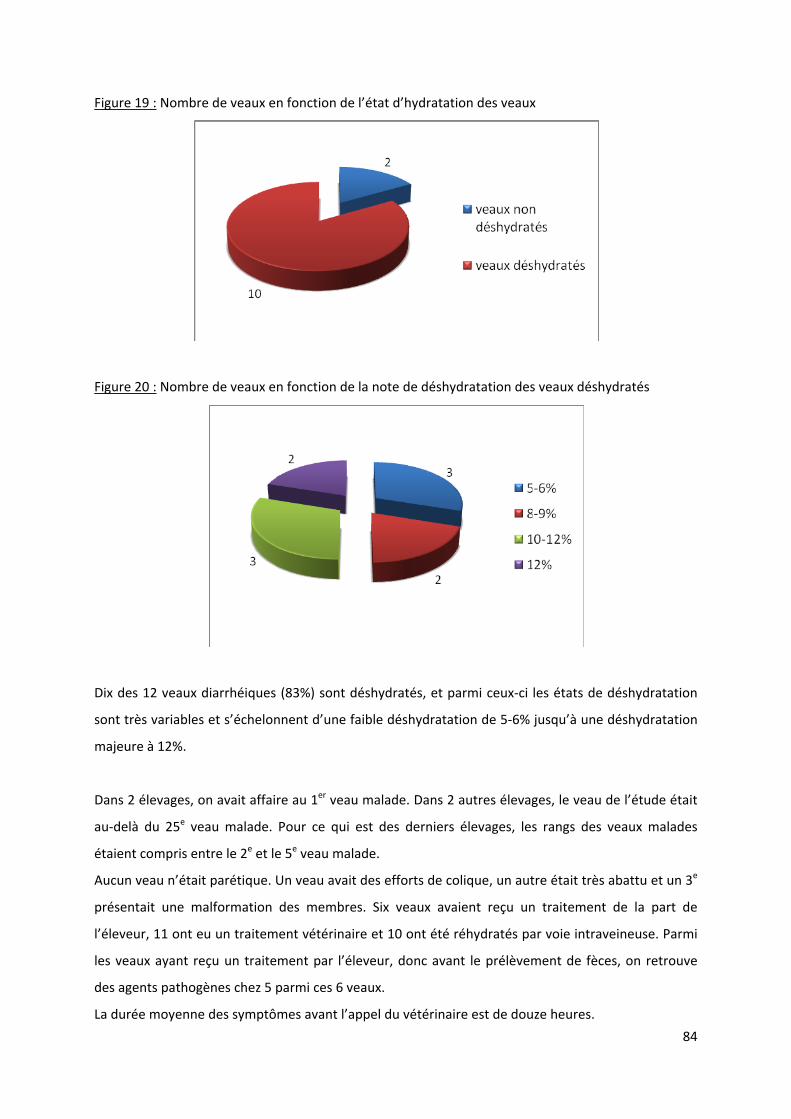
Figure 19 : Nombre de veaux en fonction de l’état d’hydratation des veaux
Figure 20 : Nombre de veaux en fonction de la note de déshydratation des veaux déshydratés
Dix des 12 veaux diarrhéiques (83%) sont déshydratés, et parmi ceux‐ci les états de déshydratation
sont très variables et s’échelonnent d’une faible déshydratation de 5‐6% jusqu’à une déshydratation
majeure à 12%.
Dans 2 élevages, on avait affaire au 1er veau malade. Dans 2 autres élevages, le veau de l’étude était
au‐delà du 25e veau malade. Pour ce qui est des derniers élevages, les rangs des veaux malades
étaient compris entre le 2e et le 5e veau malade.
Aucun veau n’était parétique. Un veau avait des efforts de colique, un autre était très abattu et un 3e
présentait une malformation des membres. Six veaux avaient reçu un traitement de la part de
l’éleveur, 11 ont eu un traitement vétérinaire et 10 ont été réhydratés par voie intraveineuse. Parmi
les veaux ayant reçu un traitement par l’éleveur, donc avant le prélèvement de fèces, on retrouve
des agents pathogènes chez 5 parmi ces 6 veaux.
La durée moyenne des symptômes avant l’appel du vétérinaire est de douze heures.
84
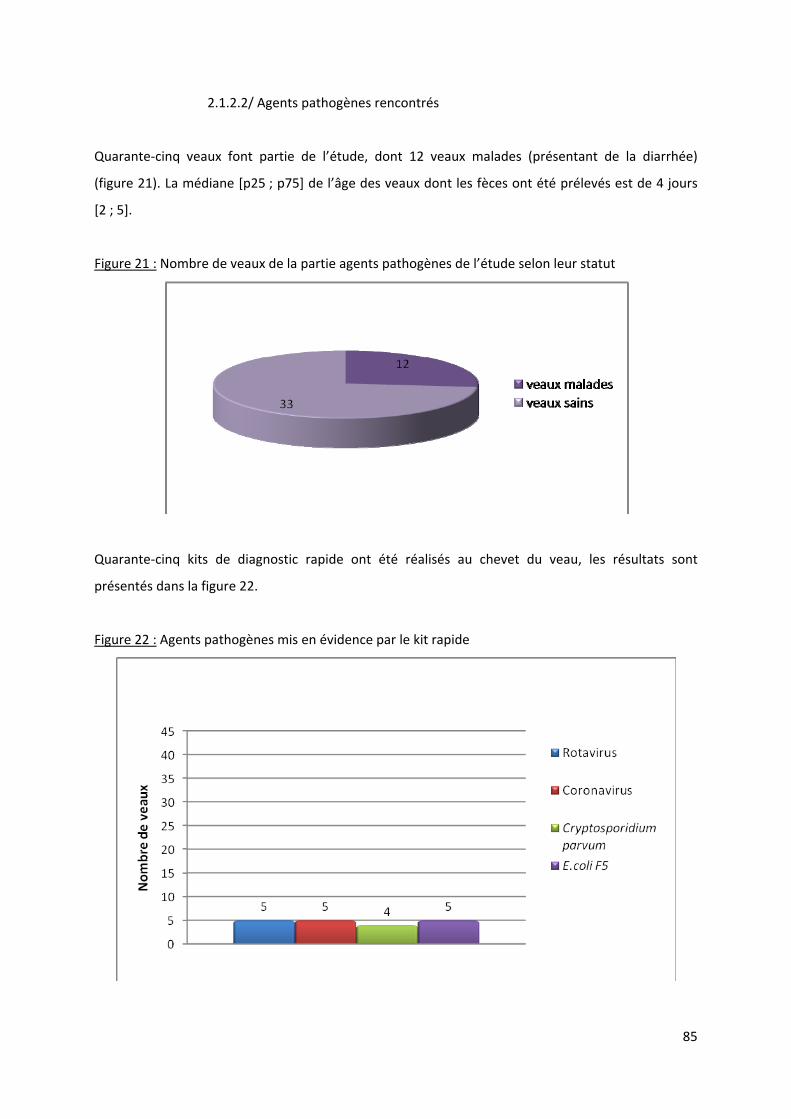
2.1.2.2/ Agents pathogènes rencontrés
Quarante‐cinq veaux font partie de l’étude, dont 12 veaux malades (présentant de la diarrhée)
(figure 21). La médiane [p25 ; p75] de l’âge des veaux dont les fèces ont été prélevés est de 4 jours
[2 ; 5].
Figure 21 : Nombre de veaux de la partie agents pathogènes de l’étude selon leur statut
Quarante‐cinq kits de diagnostic rapide ont été réalisés au chevet du veau, les résultats sont
présentés dans la figure 22.
Figure 22 : Agents pathogènes mis en évidence par le kit rapide
85
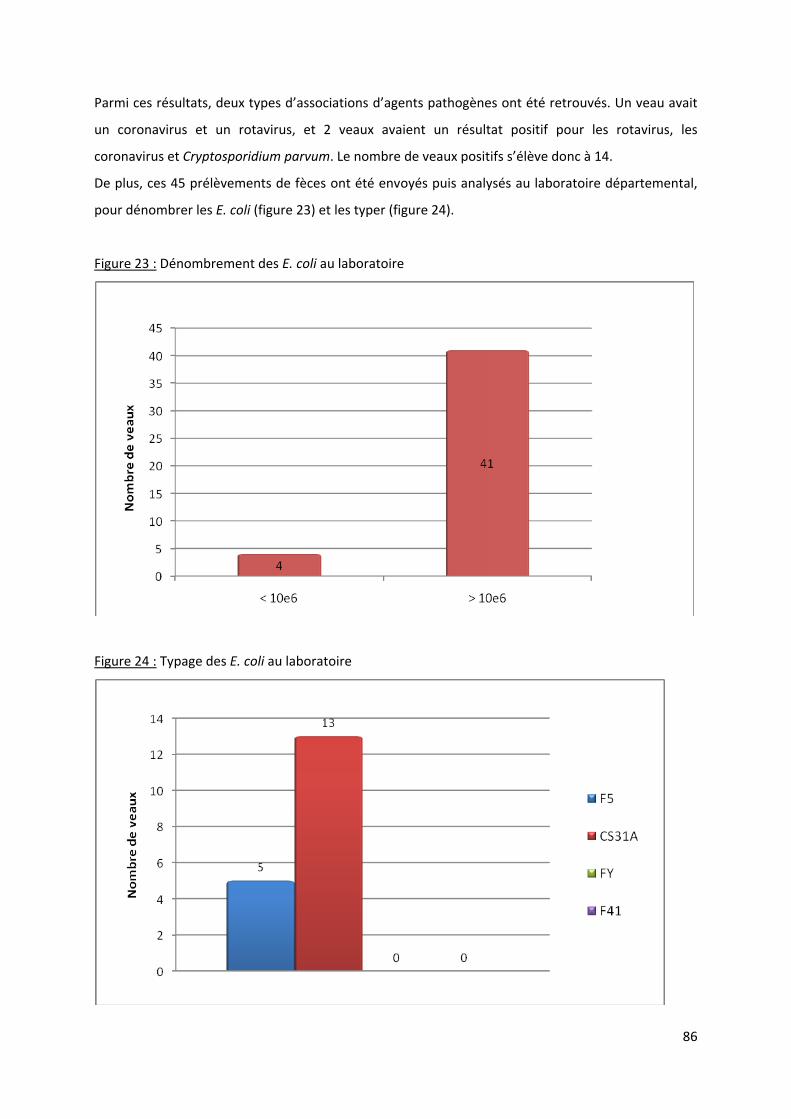
Parmi ces résultats, deux types d’associations d’agents pathogènes ont été retrouvés. Un veau avait
un coronavirus et un rotavirus, et 2 veaux avaient un résultat positif pour les rotavirus, les
coronavirus et Cryptosporidium parvum. Le nombre de veaux positifs s’élève donc à 14.
De plus, ces 45 prélèvements de fèces ont été envoyés puis analysés au laboratoire départemental,
pour dénombrer les E. coli (figure 23) et les typer (figure 24).
Figure 23 : Dénombrement des E. coli au laboratoire
Figure 24 : Typage des E. coli au laboratoire
86

On retrouve ici également 5 veaux positifs à E. coli F5, mais en comparant avec les données des kits
rapides, seulement 3 de ces 5 positifs au laboratoire confirment les résultats des kits rapides. En
effet, 2 prélèvements positifs avec le tetrakit n’ont pas donné de résultat positif au laboratoire, et 2
positifs au laboratoire n’ont pas donné de résultat positif avec les tetrakits. E. coli CS31A est, lui,
détecté chez 13 veaux. Les deux méthodes ne concordent donc pas.
La recherche de salmonelles s’est révélée négative pour tous les prélèvements.
2.1.3/ Concernant les élevages
2.1.3.1/ Bâtiments et environnement
Cette partie de l’étude comprend 10 élevages cas et 9 élevages témoins. La répartition des races de
vaches est présentée dans la figure 25. Le nombre moyen de bovins par élevage est de 160, et le
nombre moyen de vêlages dans une saison est de 80.
Figure 25 : Répartition des races dans les élevages de l’étude
Dix élevages sont purement charolais, 6 sont purement limousins et 3 sont mixtes.
Quatorze élevages (74%) n’ont qu’une seule espèce d’animaux de rente, 4 élevages (21%) ont aussi
des moutons, et 1 élevage (5%) a aussi des poules. Dans ces 5 élevages (26%), les sites d’élevage sont
bien séparés.
Huit élevages de l’étude (42%) sont des élevages principalement de plein air, mais qui rentrent une
partie ou tous leurs animaux au cours de l’hiver. Parmi ces 8 élevages, 4 sont des élevages cas et 4
sont des élevages témoins.
87
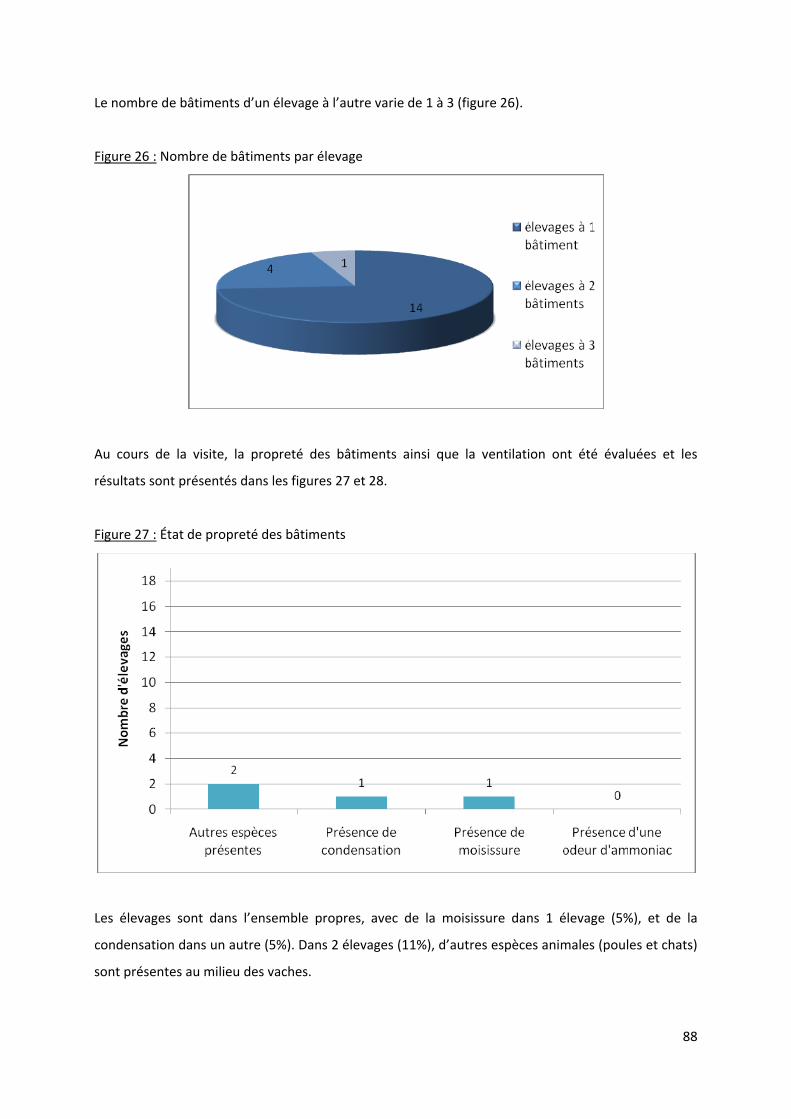
Le nombre de bâtiments d’un élevage à l’autre varie de 1 à 3 (figure 26).
Figure 26 : Nombre de bâtiments par élevage
Au cours de la visite, la propreté des bâtiments ainsi que la ventilation ont été évaluées et les
résultats sont présentés dans les figures 27 et 28.
Figure 27 : État de propreté des bâtiments
Les élevages sont dans l’ensemble propres, avec de la moisissure dans 1 élevage (5%), et de la
condensation dans un autre (5%). Dans 2 élevages (11%), d’autres espèces animales (poules et chats)
sont présentes au milieu des vaches.
88
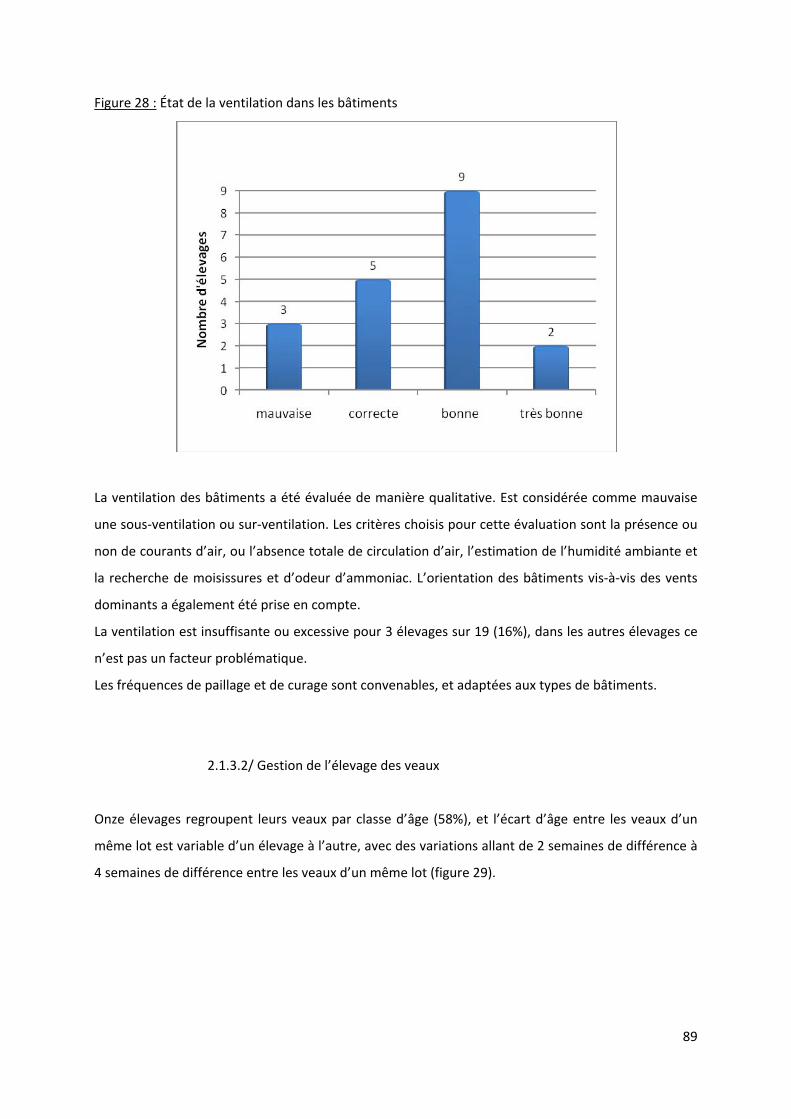
Figure 28 : État de la ventilation dans les bâtiments
La ventilation des bâtiments a été évaluée de manière qualitative. Est considérée comme mauvaise
une sous‐ventilation ou sur‐ventilation. Les critères choisis pour cette évaluation sont la présence ou
non de courants d’air, ou l’absence totale de circulation d’air, l’estimation de l’humidité ambiante et
la recherche de moisissures et d’odeur d’ammoniac. L’orientation des bâtiments vis‐à‐vis des vents
dominants a également été prise en compte.
La ventilation est insuffisante ou excessive pour 3 élevages sur 19 (16%), dans les autres élevages ce
n’est pas un facteur problématique.
Les fréquences de paillage et de curage sont convenables, et adaptées aux types de bâtiments.
2.1.3.2/ Gestion de l’élevage des veaux
Onze élevages regroupent leurs veaux par classe d’âge (58%), et l’écart d’âge entre les veaux d’un
même lot est variable d’un élevage à l’autre, avec des variations allant de 2 semaines de différence à
4 semaines de différence entre les veaux d’un même lot (figure 29).
89
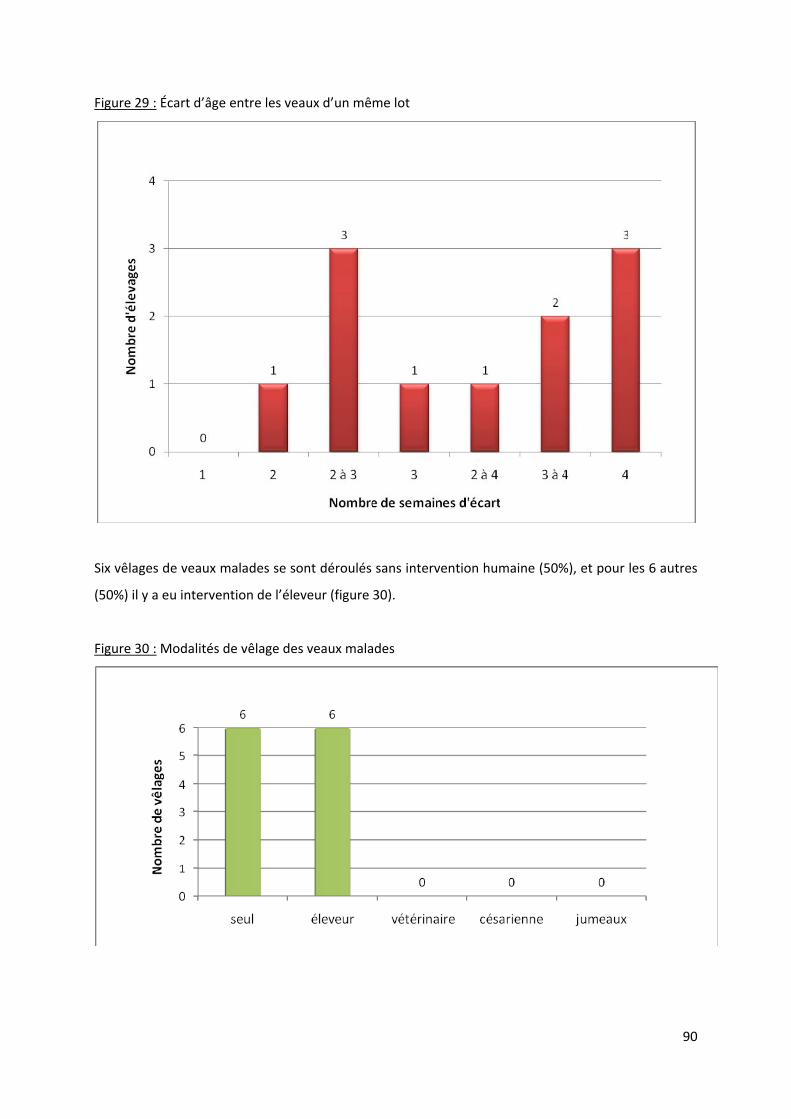
Figure 29 : Écart d’âge entre les veaux d’un même lot
Six vêlages de veaux malades se sont déroulés sans intervention humaine (50%), et pour les 6 autres
(50%) il y a eu intervention de l’éleveur (figure 30).
Figure 30 : Modalités de vêlage des veaux malades
90
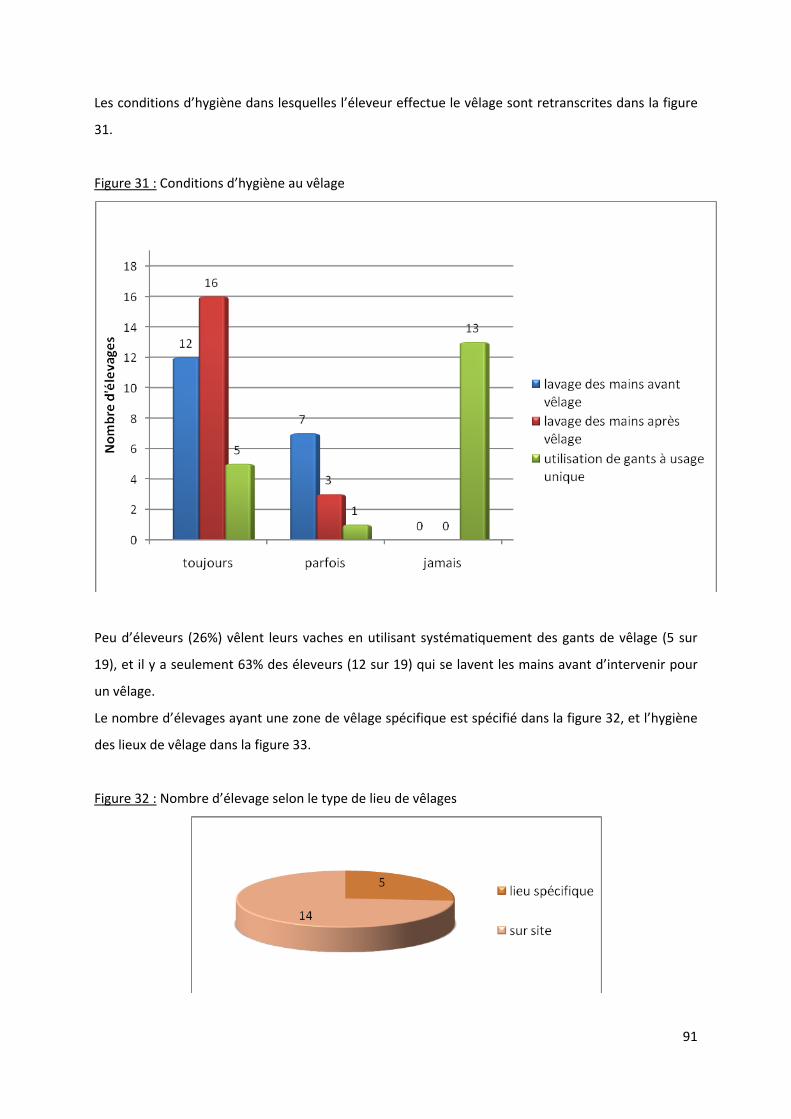
Les conditions d’hygiène dans lesquelles l’éleveur effectue le vêlage sont retranscrites dans la figure
31.
Figure 31 : Conditions d’hygiène au vêlage
Peu d’éleveurs (26%) vêlent leurs vaches en utilisant systématiquement des gants de vêlage (5 sur
19), et il y a seulement 63% des éleveurs (12 sur 19) qui se lavent les mains avant d’intervenir pour
un vêlage.
Le nombre d’élevages ayant une zone de vêlage spécifique est spécifié dans la figure 32, et l’hygiène
des lieux de vêlage dans la figure 33.
Figure 32 : Nombre d’élevage selon le type de lieu de vêlages
91
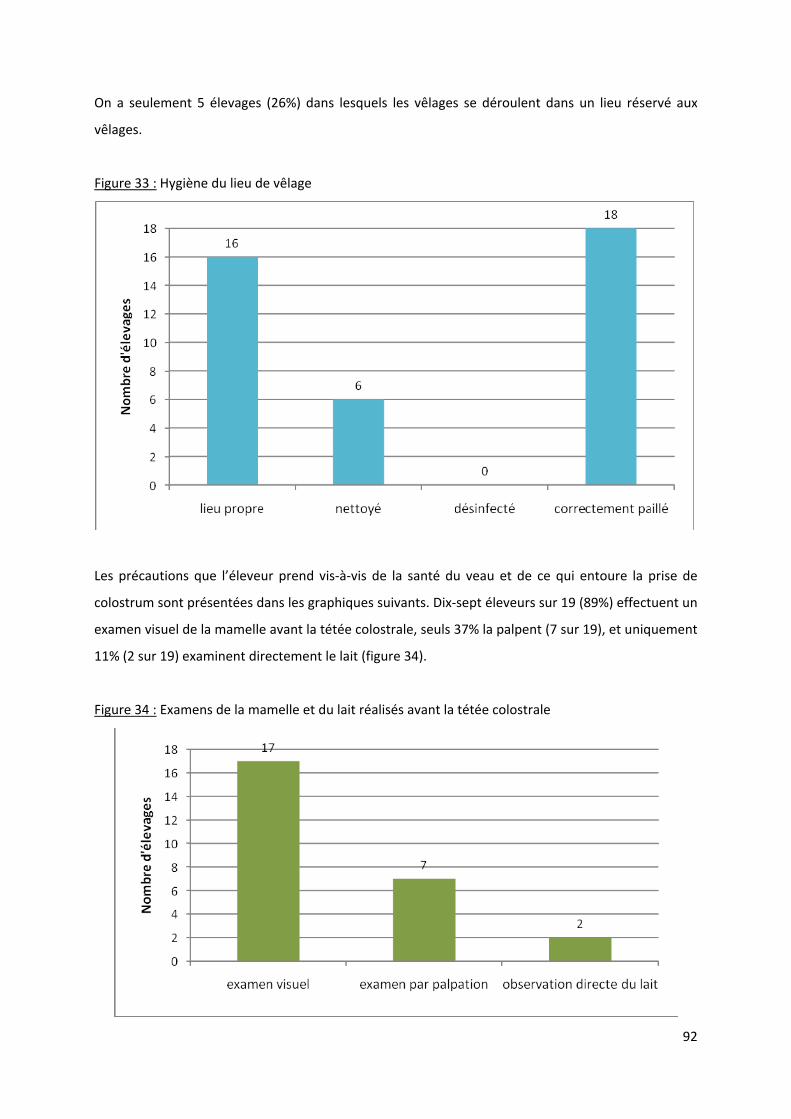
On a seulement 5 élevages (26%) dans lesquels les vêlages se déroulent dans un lieu réservé aux
vêlages.
Figure 33 : Hygiène du lieu de vêlage
Les précautions que l’éleveur prend vis‐à‐vis de la santé du veau et de ce qui entoure la prise de
colostrum sont présentées dans les graphiques suivants. Dix‐sept éleveurs sur 19 (89%) effectuent un
examen visuel de la mamelle avant la tétée colostrale, seuls 37% la palpent (7 sur 19), et uniquement
11% (2 sur 19) examinent directement le lait (figure 34).
Figure 34 : Examens de la mamelle et du lait réalisés avant la tétée colostrale
92
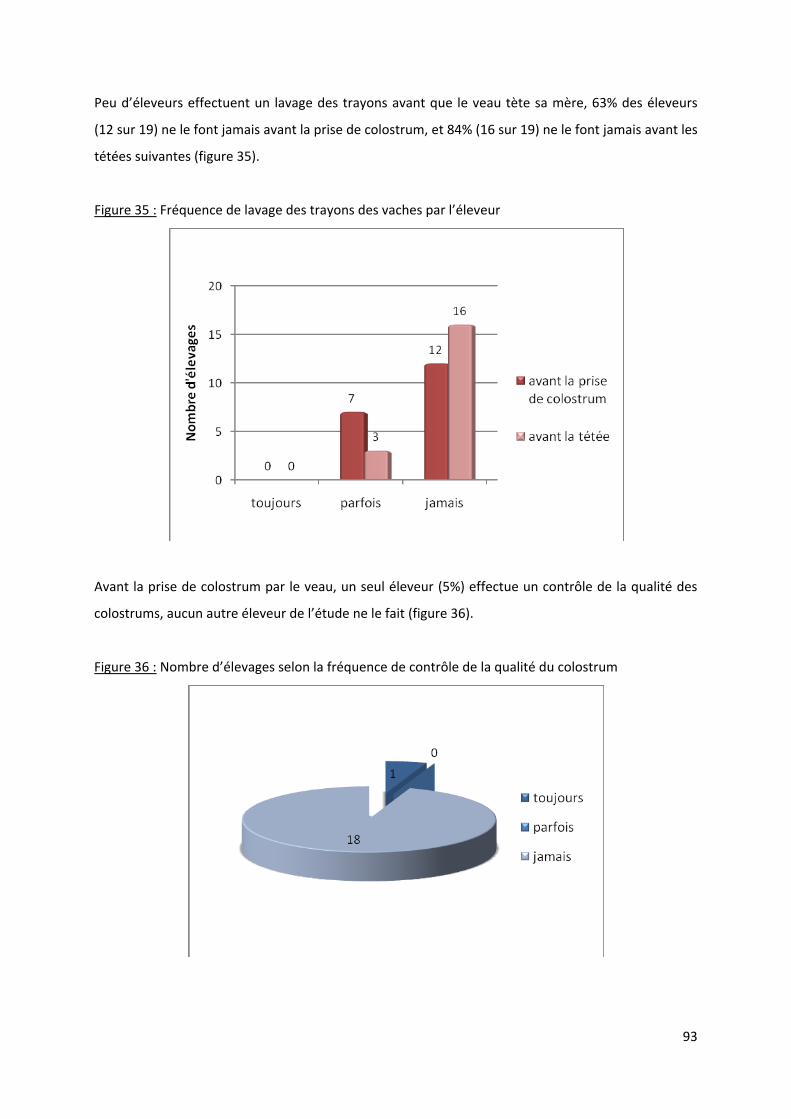
Peu d’éleveurs effectuent un lavage des trayons avant que le veau tète sa mère, 63% des éleveurs
(12 sur 19) ne le font jamais avant la prise de colostrum, et 84% (16 sur 19) ne le font jamais avant les
tétées suivantes (figure 35).
Figure 35 : Fréquence de lavage des trayons des vaches par l’éleveur
Avant la prise de colostrum par le veau, un seul éleveur (5%) effectue un contrôle de la qualité des
colostrums, aucun autre éleveur de l’étude ne le fait (figure 36).
Figure 36 : Nombre d’élevages selon la fréquence de contrôle de la qualité du colostrum
93
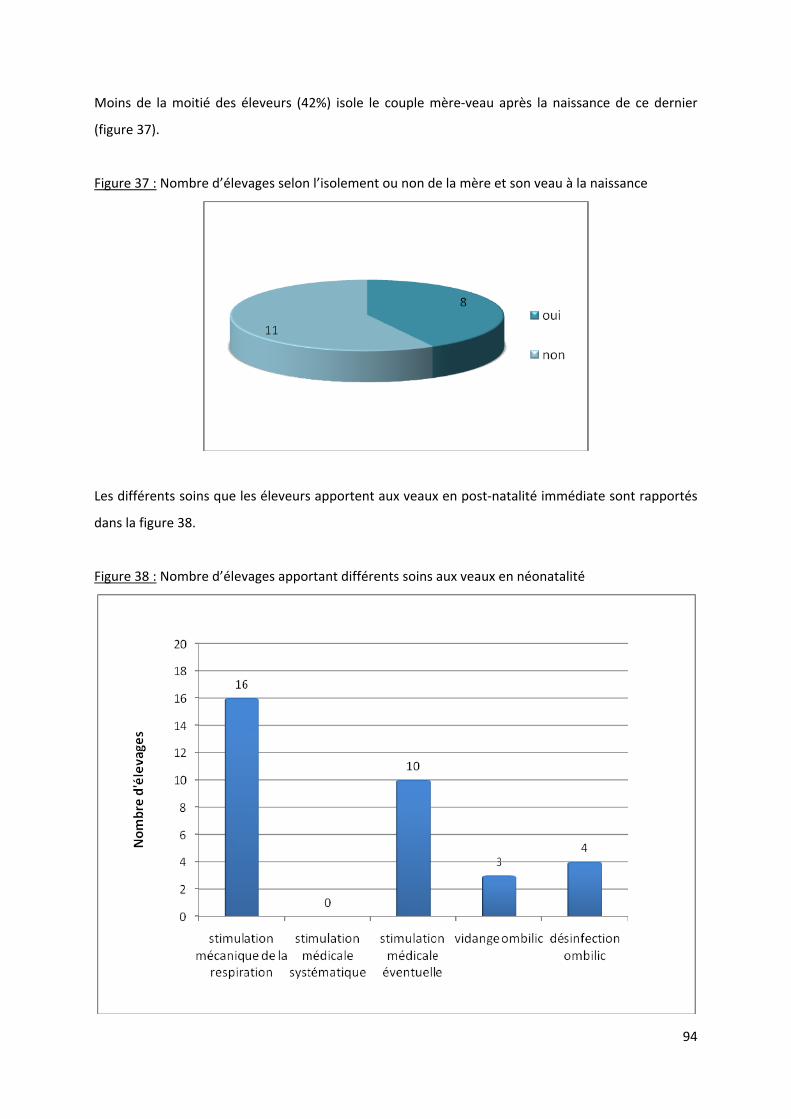
Moins de la moitié des éleveurs (42%) isole le couple mère‐veau après la naissance de ce dernier
(figure 37).
Figure 37 : Nombre d’élevages selon l’isolement ou non de la mère et son veau à la naissance
Les différents soins que les éleveurs apportent aux veaux en post‐natalité immédiate sont rapportés
dans la figure 38.
Figure 38 : Nombre d’élevages apportant différents soins aux veaux en néonatalité
94

Deux éléments en post‐natalité sont importants à surveiller. Il s’agit de la prise de colostrum dont
dépend l’apport de l’immunité passive et de nutriments indispensables aux veaux, et de l’élimination
du méconium qui est la première selle du veau. La quasi‐totalité des éleveurs contrôlent
systématiquement la prise colostrale (95%), et près des trois‐quarts des éleveurs vérifient que le
veau a bien éjecté son méconium (74%) comme on peut le voir sur la figure 39.
Figure 39 : Différents contrôles effectués en néonatalité par les éleveurs
Les deux graphiques suivants (figures 40 et 41) nous montrent la façon dont réagit l’éleveur quand le
veau refuse de téter ou quand une vache n’a pas de colostrum afin d’apporter au veau des
immunoglobulines qui lui procureront une immunité passive.
Figure 40 : Nombre d’élevages selon le type de réponse de l’éleveur face au refus de téter d’un veau
95
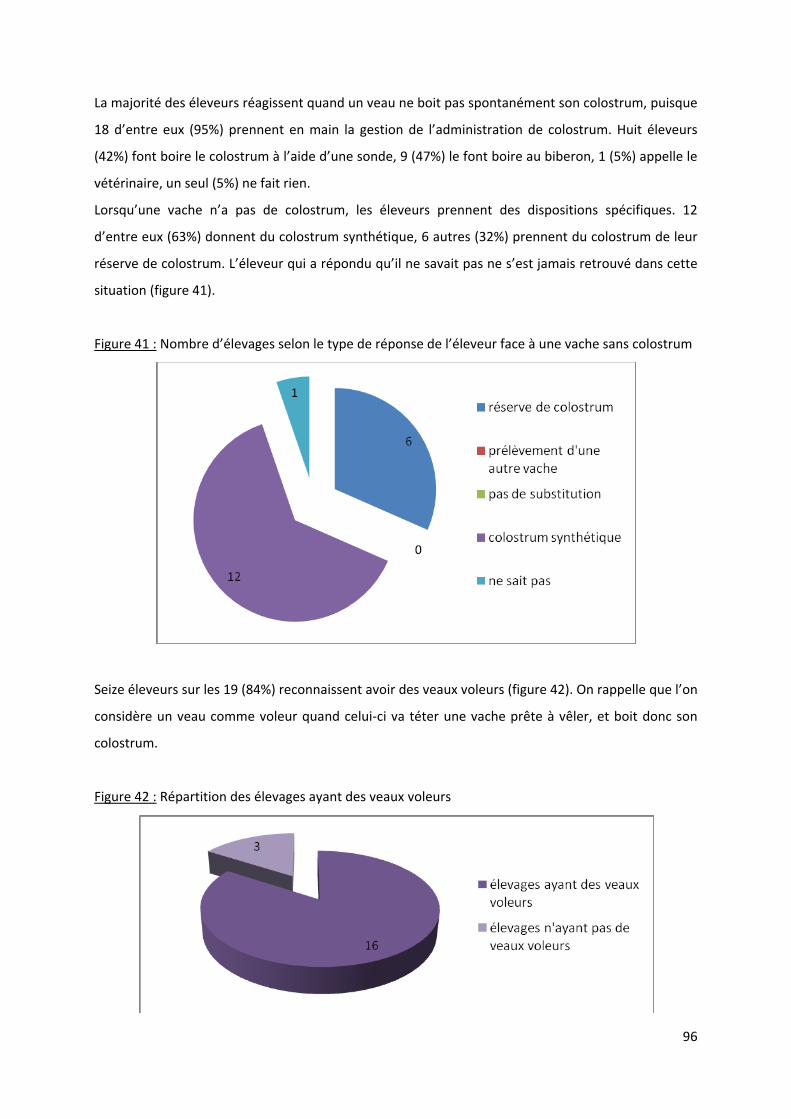
La majorité des éleveurs réagissent quand un veau ne boit pas spontanément son colostrum, puisque
18 d’entre eux (95%) prennent en main la gestion de l’administration de colostrum. Huit éleveurs
(42%) font boire le colostrum à l’aide d’une sonde, 9 (47%) le font boire au biberon, 1 (5%) appelle le
vétérinaire, un seul (5%) ne fait rien.
Lorsqu’une vache n’a pas de colostrum, les éleveurs prennent des dispositions spécifiques. 12
d’entre eux (63%) donnent du colostrum synthétique, 6 autres (32%) prennent du colostrum de leur
réserve de colostrum. L’éleveur qui a répondu qu’il ne savait pas ne s’est jamais retrouvé dans cette
situation (figure 41).
Figure 41 : Nombre d’élevages selon le type de réponse de l’éleveur face à une vache sans colostrum
Seize éleveurs sur les 19 (84%) reconnaissent avoir des veaux voleurs (figure 42). On rappelle que l’on
considère un veau comme voleur quand celui‐ci va téter une vache prête à vêler, et boit donc son
colostrum.
Figure 42 : Répartition des élevages ayant des veaux voleurs
96
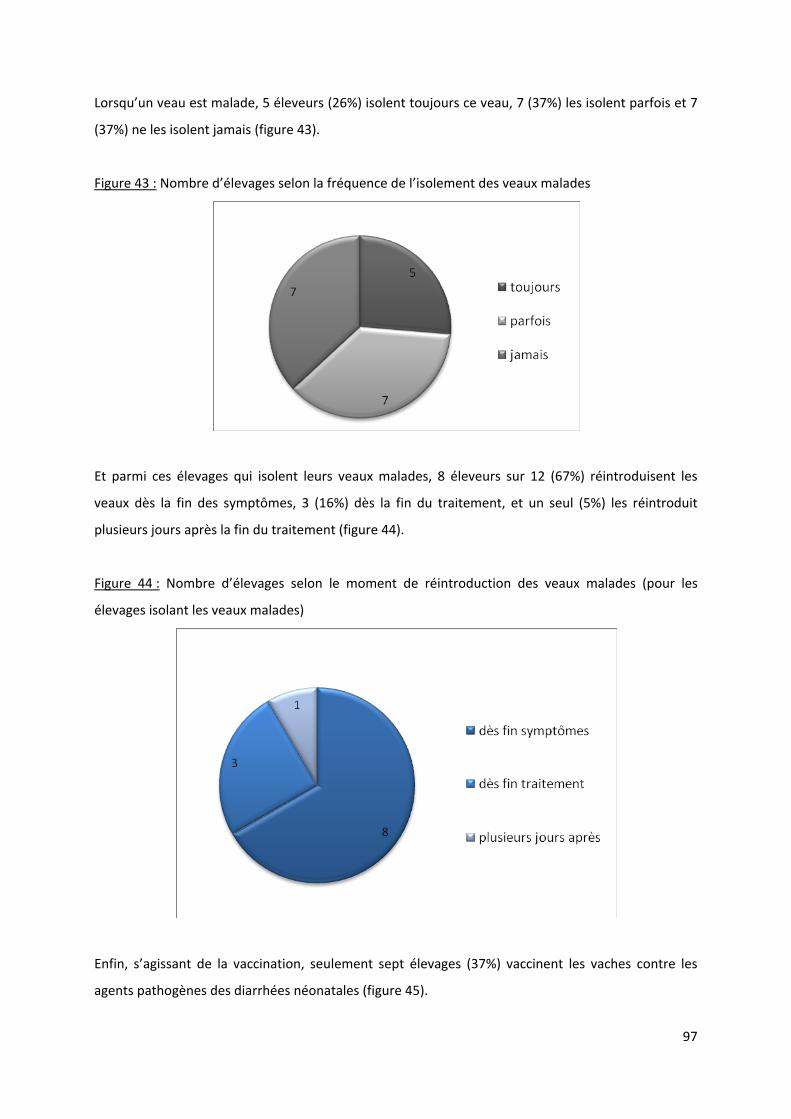
Lorsqu’un veau est malade, 5 éleveurs (26%) isolent toujours ce veau, 7 (37%) les isolent parfois et 7
(37%) ne les isolent jamais (figure 43).
Figure 43 : Nombre d’élevages selon la fréquence de l’isolement des veaux malades
Et parmi ces élevages qui isolent leurs veaux malades, 8 éleveurs sur 12 (67%) réintroduisent les
veaux dès la fin des symptômes, 3 (16%) dès la fin du traitement, et un seul (5%) les réintroduit
plusieurs jours après la fin du traitement (figure 44).
Figure 44 : Nombre d’élevages selon le moment de réintroduction des veaux malades (pour les
élevages isolant les veaux malades)
Enfin, s’agissant de la vaccination, seulement sept élevages (37%) vaccinent les vaches contre les
agents pathogènes des diarrhées néonatales (figure 45).
97
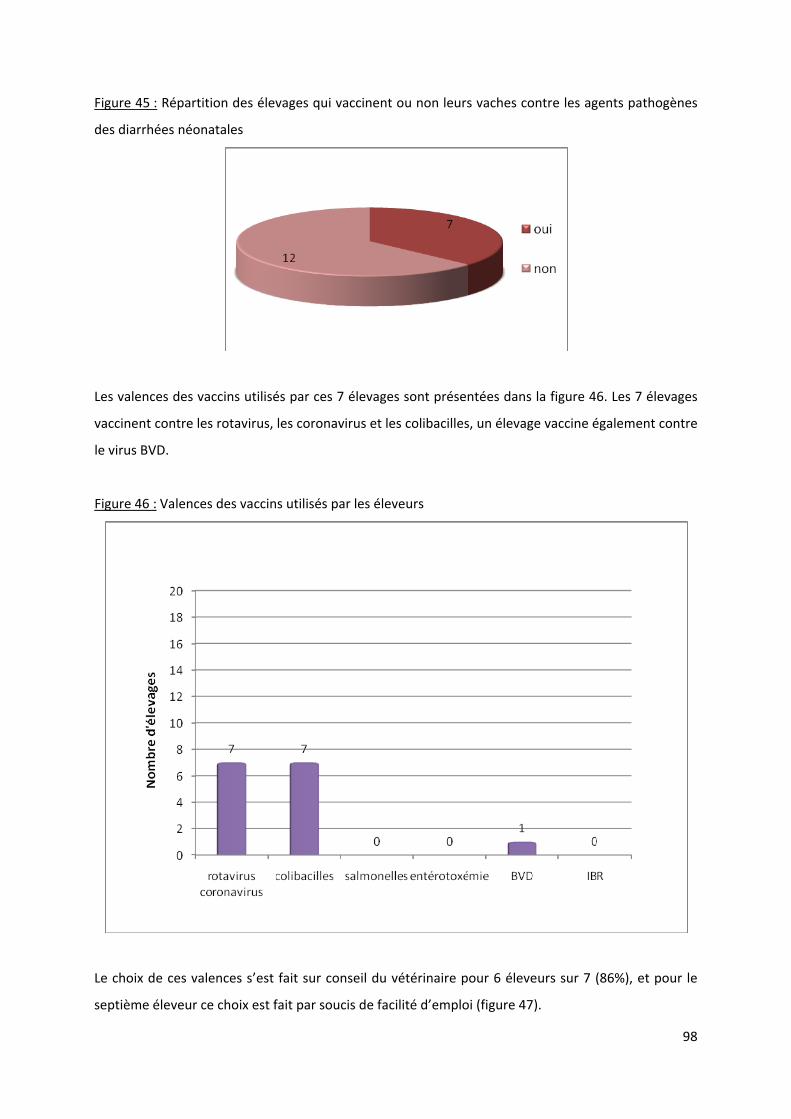
Figure 45 : Répartition des élevages qui vaccinent ou non leurs vaches contre les agents pathogènes
des diarrhées néonatales
Les valences des vaccins utilisés par ces 7 élevages sont présentées dans la figure 46. Les 7 élevages
vaccinent contre les rotavirus, les coronavirus et les colibacilles, un élevage vaccine également contre
le virus BVD.
Figure 46 : Valences des vaccins utilisés par les éleveurs
Le choix de ces valences s’est fait sur conseil du vétérinaire pour 6 éleveurs sur 7 (86%), et pour le
septième éleveur ce choix est fait par soucis de facilité d’emploi (figure 47).
98
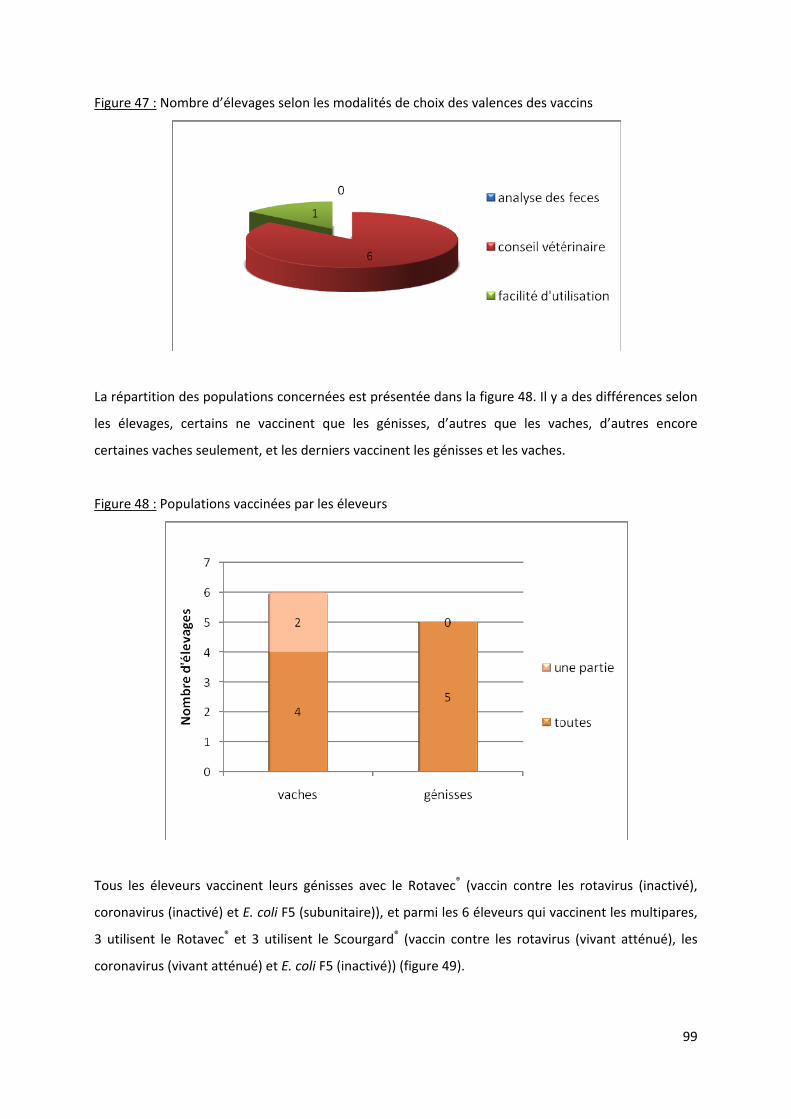
Figure 47 : Nombre d’élevages selon les modalités de choix des valences des vaccins
La répartition des populations concernées est présentée dans la figure 48. Il y a des différences selon
les élevages, certains ne vaccinent que les génisses, d’autres que les vaches, d’autres encore
certaines vaches seulement, et les derniers vaccinent les génisses et les vaches.
Figure 48 : Populations vaccinées par les éleveurs
Tous les éleveurs vaccinent leurs génisses avec le Rotavec® (vaccin contre les rotavirus (inactivé),
coronavirus (inactivé) et E. coli F5 (subunitaire)), et parmi les 6 éleveurs qui vaccinent les multipares,
3 utilisent le Rotavec® et 3 utilisent le Scourgard® (vaccin contre les rotavirus (vivant atténué), les
coronavirus (vivant atténué) et E. coli F5 (inactivé)) (figure 49).
99
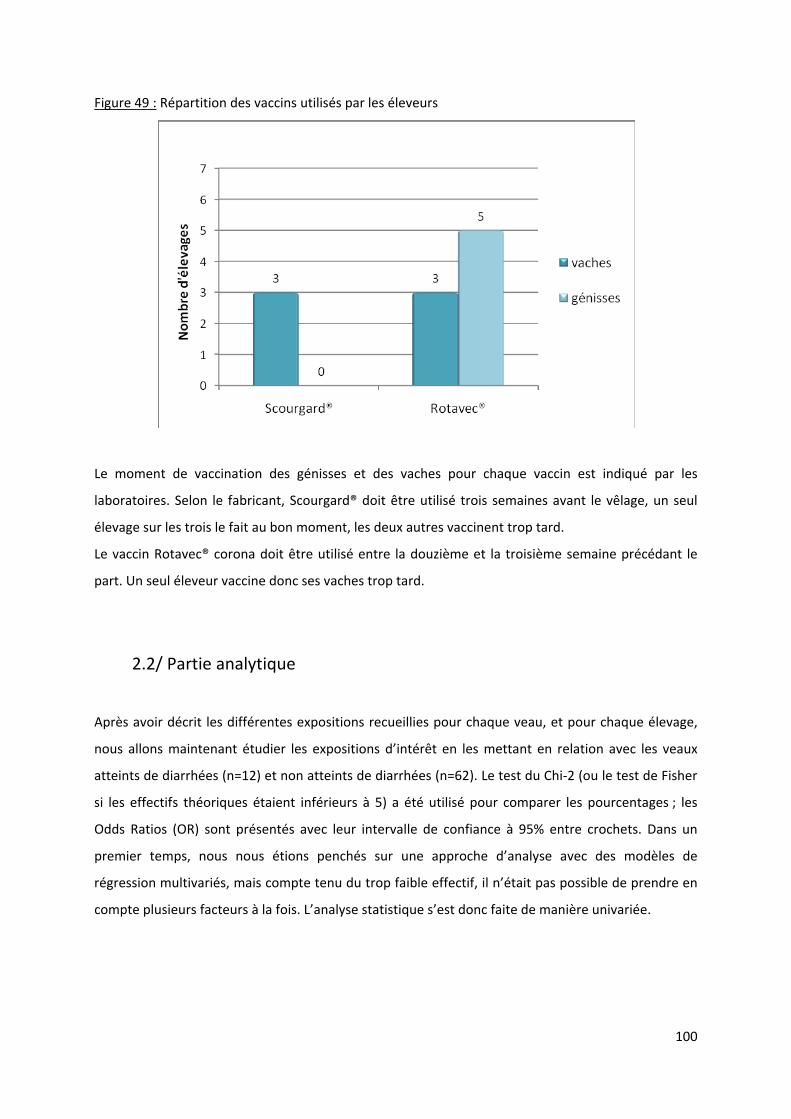
Figure 49 : Répartition des vaccins utilisés par les éleveurs
Le moment de vaccination des génisses et des vaches pour chaque vaccin est indiqué par les
laboratoires. Selon le fabricant, Scourgard® doit être utilisé trois semaines avant le vêlage, un seul
élevage sur les trois le fait au bon moment, les deux autres vaccinent trop tard.
Le vaccin Rotavec® corona doit être utilisé entre la douzième et la troisième semaine précédant le
part. Un seul éleveur vaccine donc ses vaches trop tard.
2.2/ Partie analytique
Après avoir décrit les différentes expositions recueillies pour chaque veau, et pour chaque élevage,
nous allons maintenant étudier les expositions d’intérêt en les mettant en relation avec les veaux
atteints de diarrhées (n=12) et non atteints de diarrhées (n=62). Le test du Chi‐2 (ou le test de Fisher
si les effectifs théoriques étaient inférieurs à 5) a été utilisé pour comparer les pourcentages ; les
Odds Ratios (OR) sont présentés avec leur intervalle de confiance à 95% entre crochets. Dans un
premier temps, nous nous étions penchés sur une approche d’analyse avec des modèles de
régression multivariés, mais compte tenu du trop faible effectif, il n’était pas possible de prendre en
compte plusieurs facteurs à la fois. L’analyse statistique s’est donc faite de manière univariée.
100
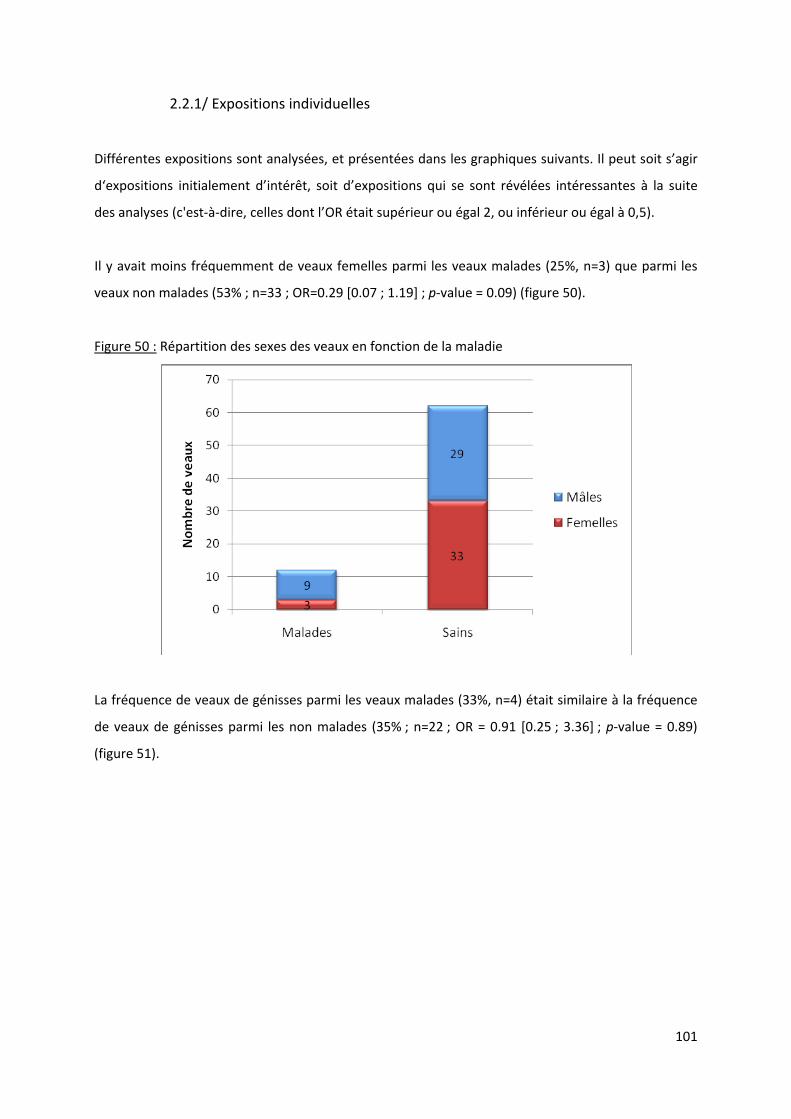
2.2.1/ Expositions individuelles
Différentes expositions sont analysées, et présentées dans les graphiques suivants. Il peut soit s’agir
d‘expositions initialement d’intérêt, soit d’expositions qui se sont révélées intéressantes à la suite
des analyses (c'est‐à‐dire, celles dont l’OR était supérieur ou égal 2, ou inférieur ou égal à 0,5).
Il y avait moins fréquemment de veaux femelles parmi les veaux malades (25%, n=3) que parmi les
veaux non malades (53% ; n=33 ; OR=0.29 [0.07 ; 1.19] ; p‐value = 0.09) (figure 50).
Figure 50 : Répartition des sexes des veaux en fonction de la maladie
La fréquence de veaux de génisses parmi les veaux malades (33%, n=4) était similaire à la fréquence
de veaux de génisses parmi les non malades (35% ; n=22 ; OR = 0.91 [0.25 ; 3.36] ; p‐value = 0.89)
(figure 51).
101
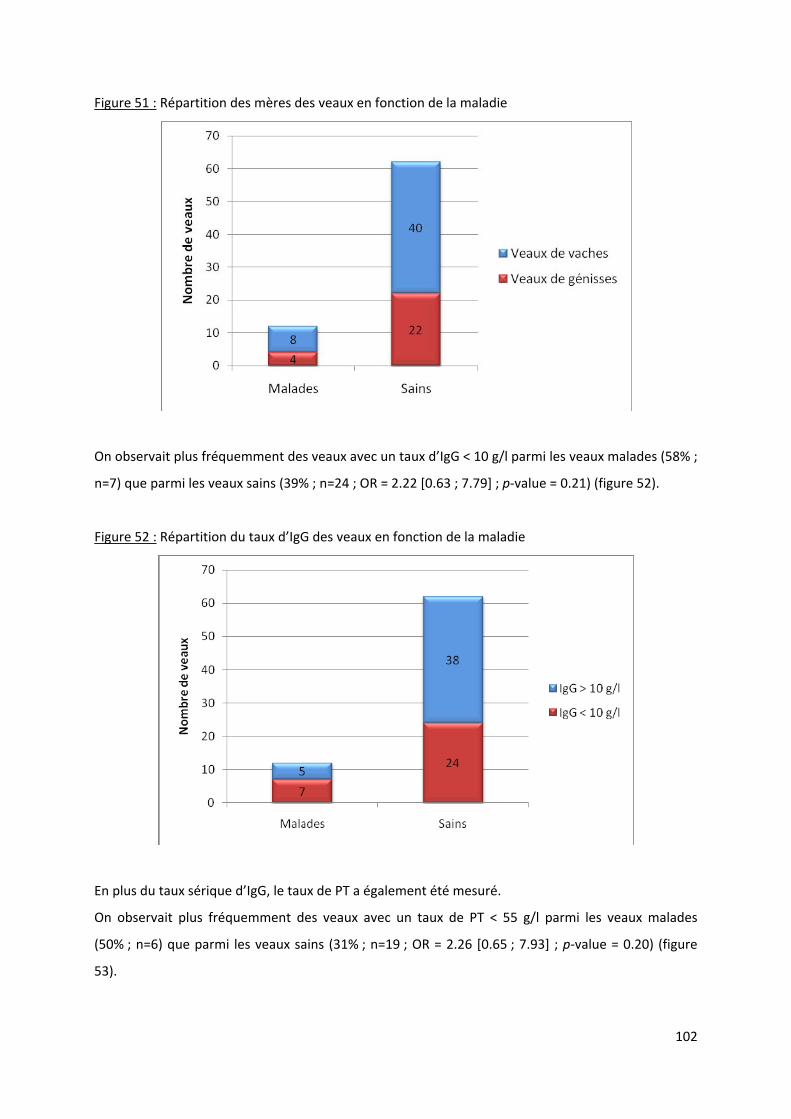
Figure 51 : Répartition des mères des veaux en fonction de la maladie
On observait plus fréquemment des veaux avec un taux d’IgG < 10 g/l parmi les veaux malades (58% ;
n=7) que parmi les veaux sains (39% ; n=24 ; OR = 2.22 [0.63 ; 7.79] ; p‐value = 0.21) (figure 52).
Figure 52 : Répartition du taux d’IgG des veaux en fonction de la maladie
En plus du taux sérique d’IgG, le taux de PT a également été mesuré.
On observait plus fréquemment des veaux avec un taux de PT < 55 g/l parmi les veaux malades
(50% ; n=6) que parmi les veaux sains (31% ; n=19 ; OR = 2.26 [0.65 ; 7.93] ; p‐value = 0.20) (figure
53).
102
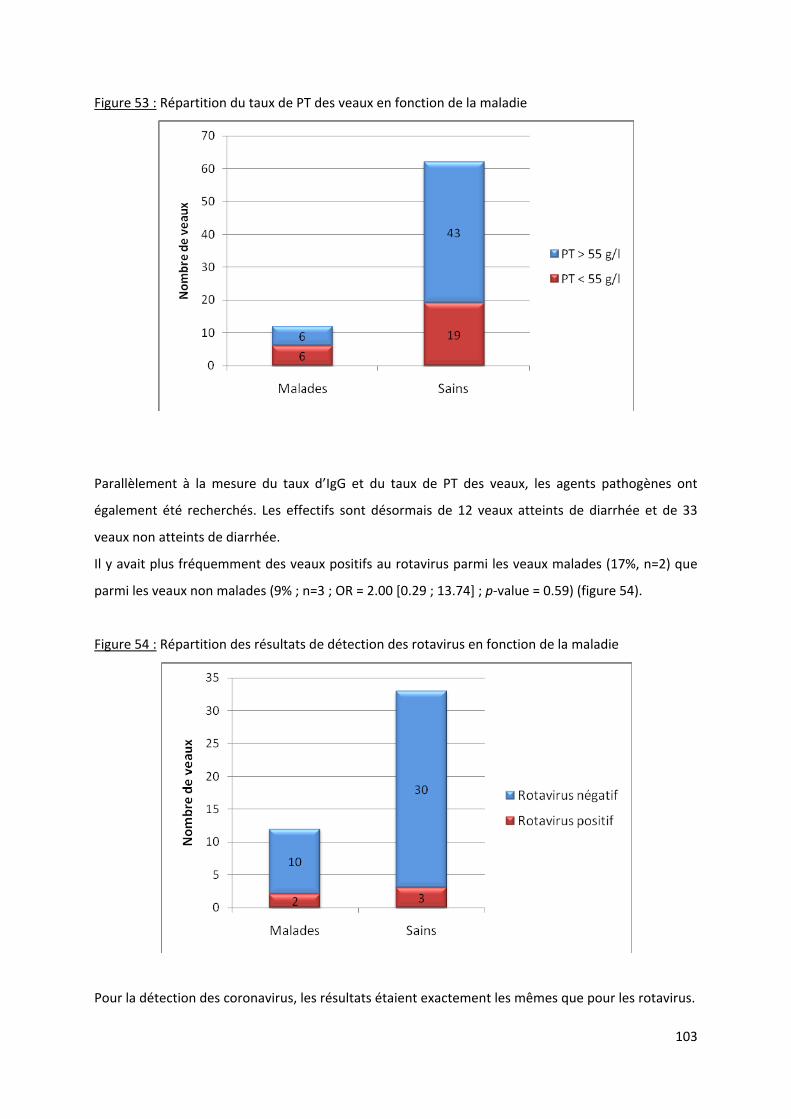
Figure 53 : Répartition du taux de PT des veaux en fonction de la maladie
Parallèlement à la mesure du taux d’IgG et du taux de PT des veaux, les agents pathogènes ont
également été recherchés. Les effectifs sont désormais de 12 veaux atteints de diarrhée et de 33
veaux non atteints de diarrhée.
Il y avait plus fréquemment des veaux positifs au rotavirus parmi les veaux malades (17%, n=2) que
parmi les veaux non malades (9% ; n=3 ; OR = 2.00 [0.29 ; 13.74] ; p‐value = 0.59) (figure 54).
Figure 54 : Répartition des résultats de détection des rotavirus en fonction de la maladie
Pour la détection des coronavirus, les résultats étaient exactement les mêmes que pour les rotavirus.
103
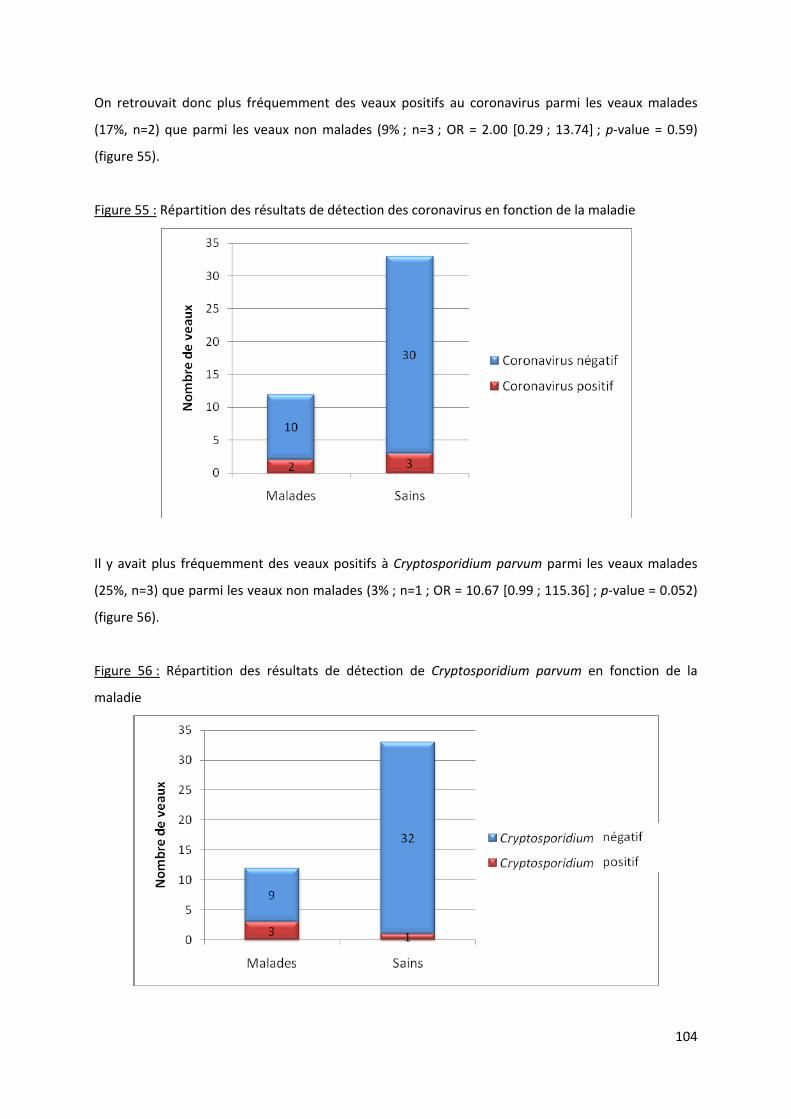
On retrouvait donc plus fréquemment des veaux positifs au coronavirus parmi les veaux malades
(17%, n=2) que parmi les veaux non malades (9% ; n=3 ; OR = 2.00 [0.29 ; 13.74] ; p‐value = 0.59)
(figure 55).
Figure 55 : Répartition des résultats de détection des coronavirus en fonction de la maladie
Il y avait plus fréquemment des veaux positifs à Cryptosporidium parvum parmi les veaux malades
(25%, n=3) que parmi les veaux non malades (3% ; n=1 ; OR = 10.67 [0.99 ; 115.36] ; p‐value = 0.052)
(figure 56).
Figure 56 : Répartition des résultats de détection de Cryptosporidium parvum en fonction de la
maladie
104
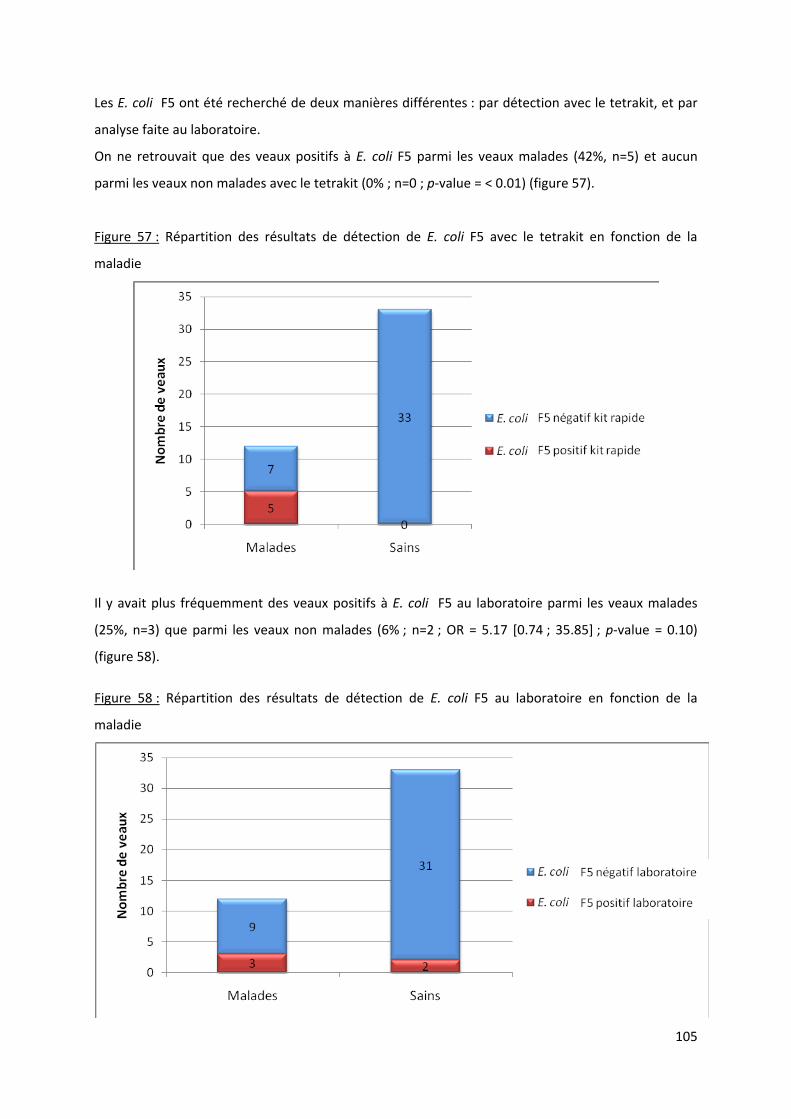
Les E. coli F5 ont été recherché de deux manières différentes : par détection avec le tetrakit, et par
analyse faite au laboratoire.
On ne retrouvait que des veaux positifs à E. coli F5 parmi les veaux malades (42%, n=5) et aucun
parmi les veaux non malades avec le tetrakit (0% ; n=0 ; p‐value = < 0.01) (figure 57).
Figure 57 : Répartition des résultats de détection de E. coli F5 avec le tetrakit en fonction de la
maladie
Il y avait plus fréquemment des veaux positifs à E. coli F5 au laboratoire parmi les veaux malades
(25%, n=3) que parmi les veaux non malades (6% ; n=2 ; OR = 5.17 [0.74 ; 35.85] ; p‐value = 0.10)
(figure 58).
Figure 58 : Répartition des résultats de détection de E. coli F5 au laboratoire en fonction de la
maladie
105
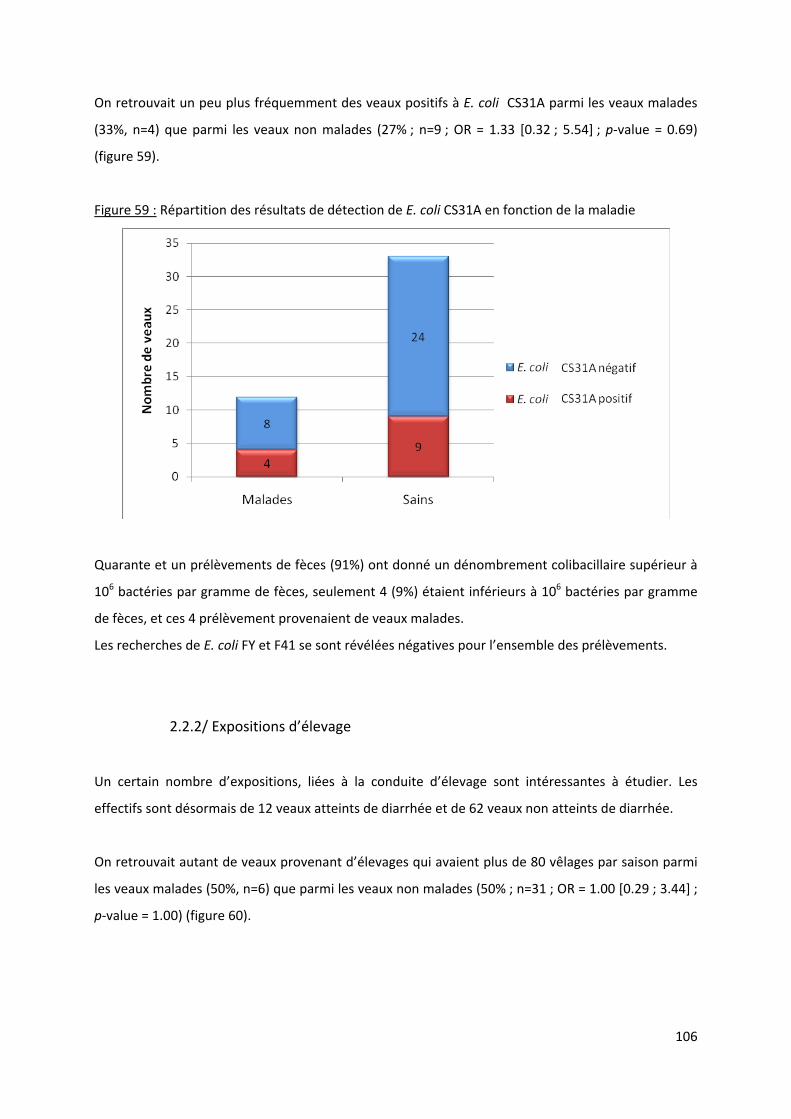
On retrouvait un peu plus fréquemment des veaux positifs à E. coli CS31A parmi les veaux malades
(33%, n=4) que parmi les veaux non malades (27% ; n=9 ; OR = 1.33 [0.32 ; 5.54] ; p‐value = 0.69)
(figure 59).
Figure 59 : Répartition des résultats de détection de E. coli CS31A en fonction de la maladie
Quarante et un prélèvements de fèces (91%) ont donné un dénombrement colibacillaire supérieur à
106 bactéries par gramme de fèces, seulement 4 (9%) étaient inférieurs à 106 bactéries par gramme
de fèces, et ces 4 prélèvement provenaient de veaux malades.
Les recherches de E. coli FY et F41 se sont révélées négatives pour l’ensemble des prélèvements.
2.2.2/ Expositions d’élevage
Un certain nombre d’expositions, liées à la conduite d’élevage sont intéressantes à étudier. Les
effectifs sont désormais de 12 veaux atteints de diarrhée et de 62 veaux non atteints de diarrhée.
On retrouvait autant de veaux provenant d’élevages qui avaient plus de 80 vêlages par saison parmi
les veaux malades (50%, n=6) que parmi les veaux non malades (50% ; n=31 ; OR = 1.00 [0.29 ; 3.44] ;
p‐value = 1.00) (figure 60).
106
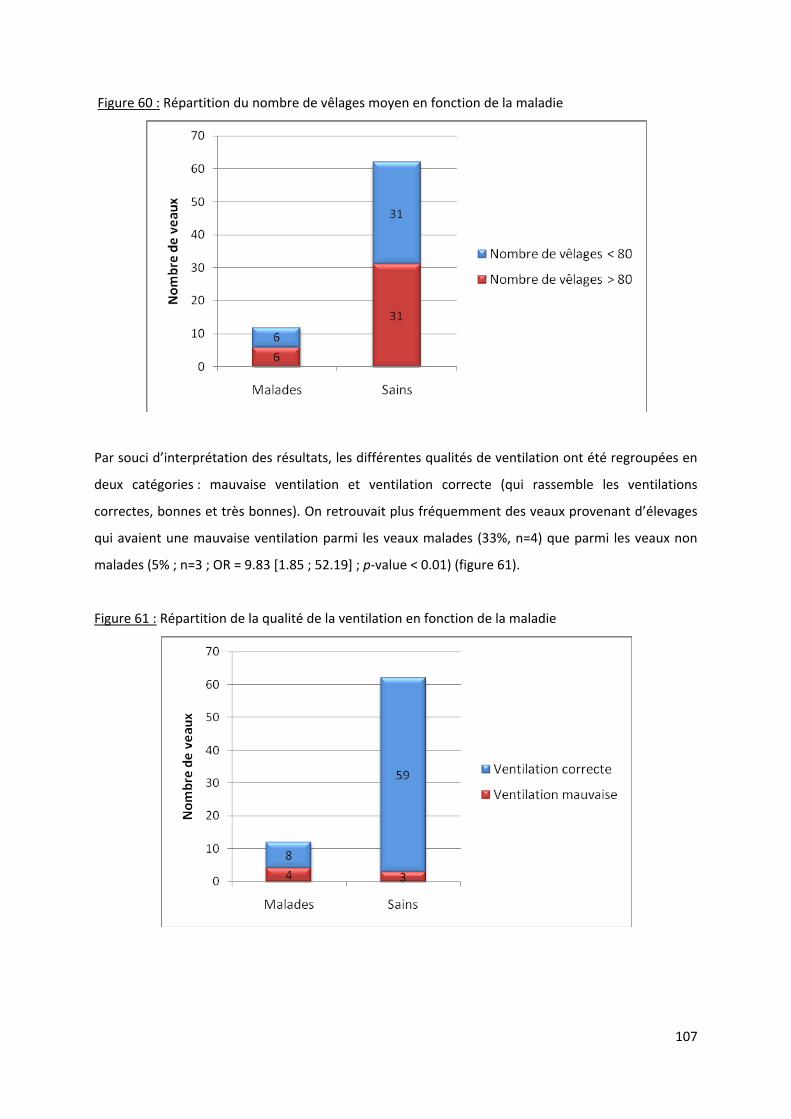
Figure 60 : Répartition du nombre de vêlages moyen en fonction de la maladie
Par souci d’interprétation des résultats, les différentes qualités de ventilation ont été regroupées en
deux catégories : mauvaise ventilation et ventilation correcte (qui rassemble les ventilations
correctes, bonnes et très bonnes). On retrouvait plus fréquemment des veaux provenant d’élevages
qui avaient une mauvaise ventilation parmi les veaux malades (33%, n=4) que parmi les veaux non
malades (5% ; n=3 ; OR = 9.83 [1.85 ; 52.19] ; p‐value < 0.01) (figure 61).
Figure 61 : Répartition de la qualité de la ventilation en fonction de la maladie
107
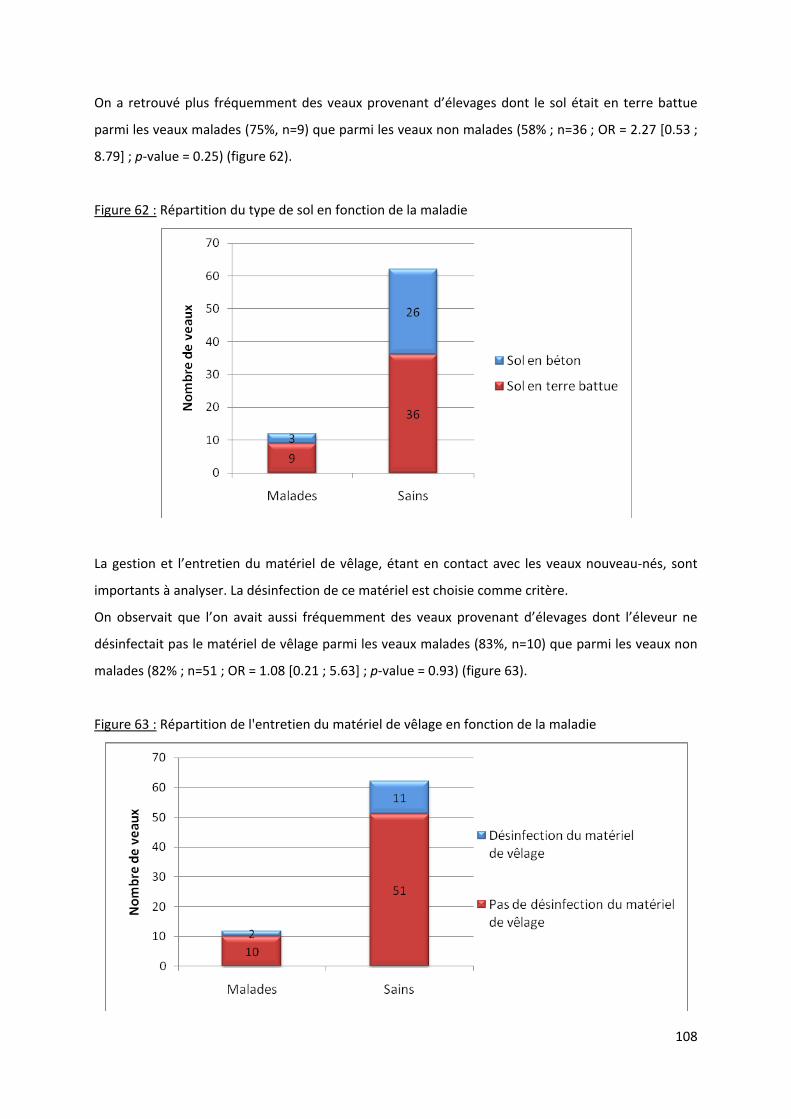
On a retrouvé plus fréquemment des veaux provenant d’élevages dont le sol était en terre battue
parmi les veaux malades (75%, n=9) que parmi les veaux non malades (58% ; n=36 ; OR = 2.27 [0.53 ;
8.79] ; p‐value = 0.25) (figure 62).
Figure 62 : Répartition du type de sol en fonction de la maladie
La gestion et l’entretien du matériel de vêlage, étant en contact avec les veaux nouveau‐nés, sont
importants à analyser. La désinfection de ce matériel est choisie comme critère.
On observait que l’on avait aussi fréquemment des veaux provenant d’élevages dont l’éleveur ne
désinfectait pas le matériel de vêlage parmi les veaux malades (83%, n=10) que parmi les veaux non
malades (82% ; n=51 ; OR = 1.08 [0.21 ; 5.63] ; p‐value = 0.93) (figure 63).
Figure 63 : Répartition de l'entretien du matériel de vêlage en fonction de la maladie
108
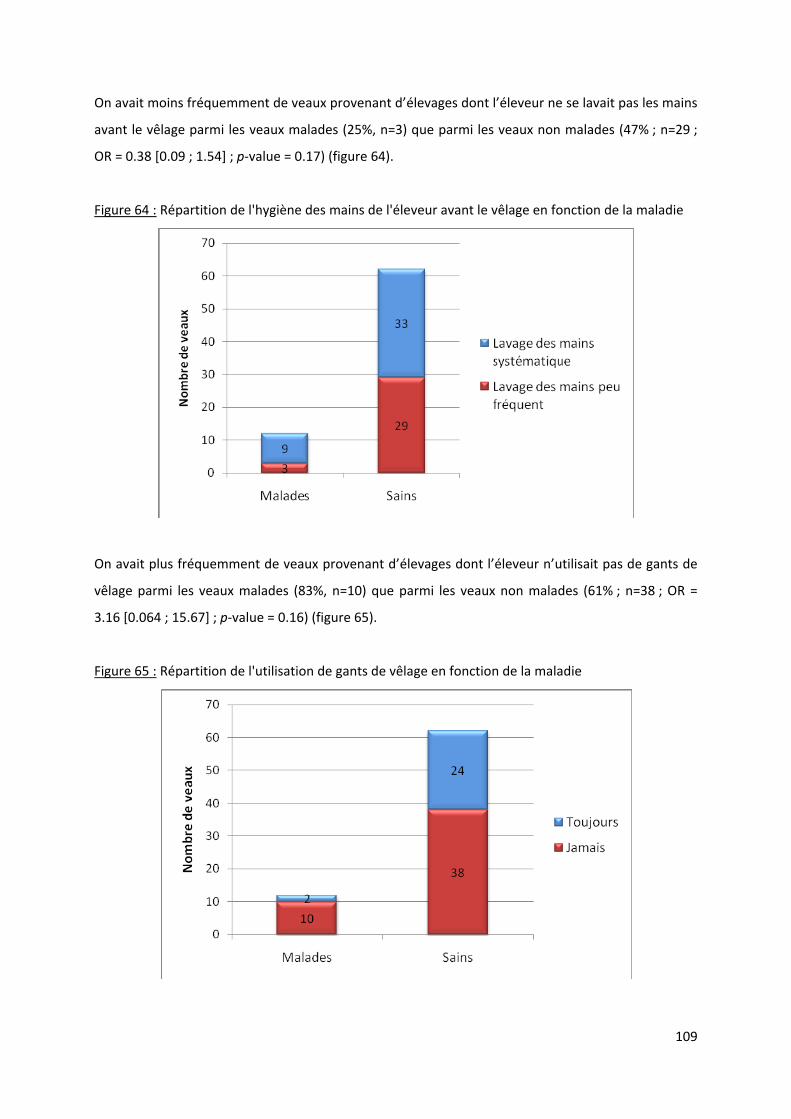
On avait moins fréquemment de veaux provenant d’élevages dont l’éleveur ne se lavait pas les mains
avant le vêlage parmi les veaux malades (25%, n=3) que parmi les veaux non malades (47% ; n=29 ;
OR = 0.38 [0.09 ; 1.54] ; p‐value = 0.17) (figure 64).
Figure 64 : Répartition de l'hygiène des mains de l'éleveur avant le vêlage en fonction de la maladie
On avait plus fréquemment de veaux provenant d’élevages dont l’éleveur n’utilisait pas de gants de
vêlage parmi les veaux malades (83%, n=10) que parmi les veaux non malades (61% ; n=38 ; OR =
3.16 [0.064 ; 15.67] ; p‐value = 0.16) (figure 65).
Figure 65 : Répartition de l'utilisation de gants de vêlage en fonction de la maladie
109
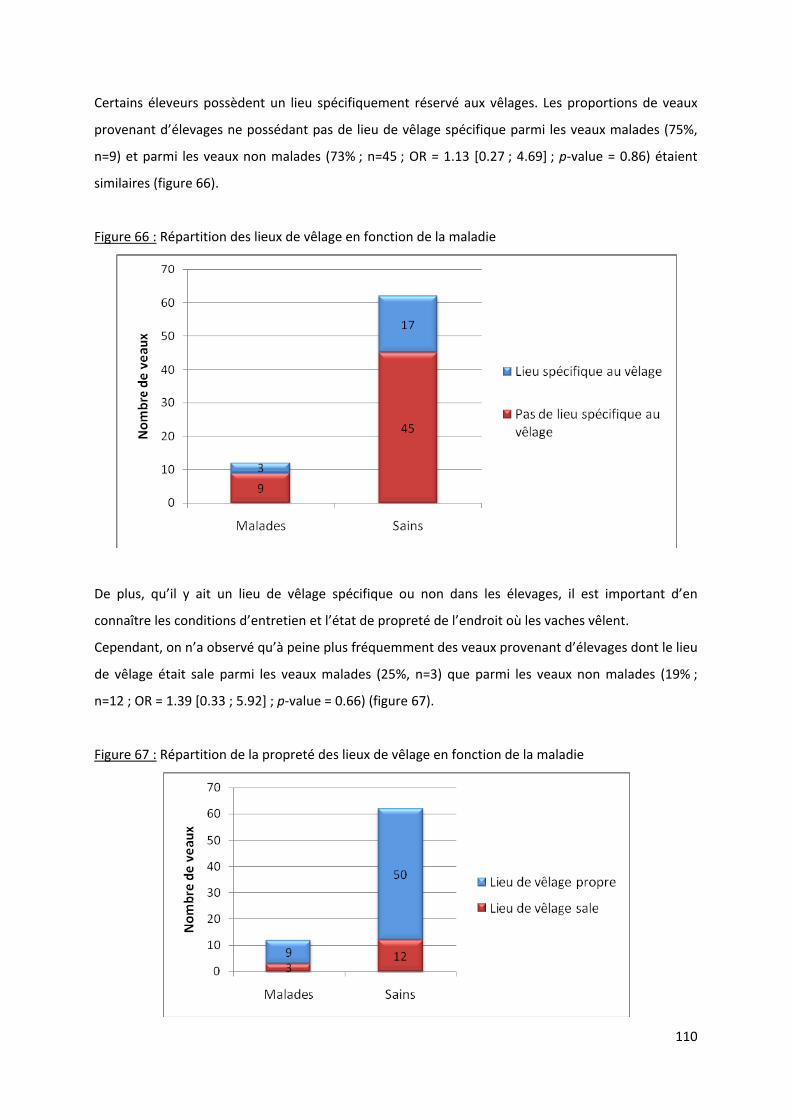
Certains éleveurs possèdent un lieu spécifiquement réservé aux vêlages. Les proportions de veaux
provenant d’élevages ne possédant pas de lieu de vêlage spécifique parmi les veaux malades (75%,
n=9) et parmi les veaux non malades (73% ; n=45 ; OR = 1.13 [0.27 ; 4.69] ; p‐value = 0.86) étaient
similaires (figure 66).
Figure 66 : Répartition des lieux de vêlage en fonction de la maladie
De plus, qu’il y ait un lieu de vêlage spécifique ou non dans les élevages, il est important d’en
connaître les conditions d’entretien et l’état de propreté de l’endroit où les vaches vêlent.
Cependant, on n’a observé qu’à peine plus fréquemment des veaux provenant d’élevages dont le lieu
de vêlage était sale parmi les veaux malades (25%, n=3) que parmi les veaux non malades (19% ;
n=12 ; OR = 1.39 [0.33 ; 5.92] ; p‐value = 0.66) (figure 67).
Figure 67 : Répartition de la propreté des lieux de vêlage en fonction de la maladie
110
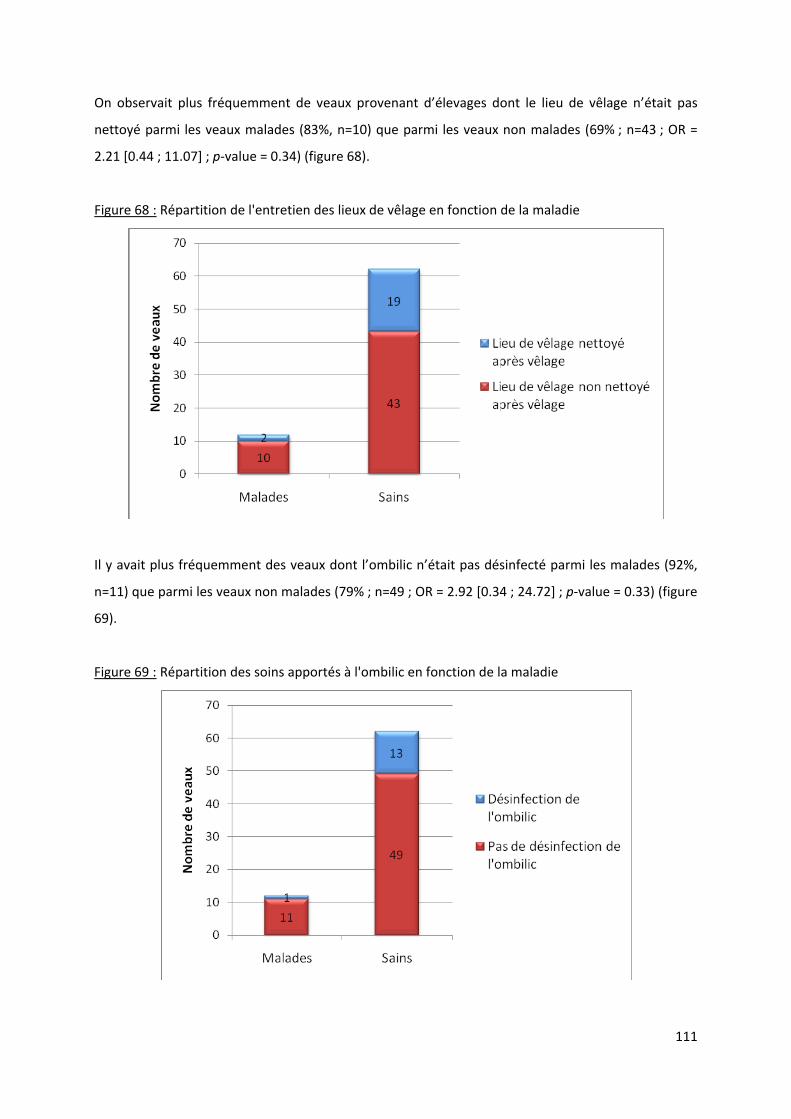
On observait plus fréquemment de veaux provenant d’élevages dont le lieu de vêlage n’était pas
nettoyé parmi les veaux malades (83%, n=10) que parmi les veaux non malades (69% ; n=43 ; OR =
2.21 [0.44 ; 11.07] ; p‐value = 0.34) (figure 68).
Figure 68 : Répartition de l'entretien des lieux de vêlage en fonction de la maladie
Il y avait plus fréquemment des veaux dont l’ombilic n’était pas désinfecté parmi les malades (92%,
n=11) que parmi les veaux non malades (79% ; n=49 ; OR = 2.92 [0.34 ; 24.72] ; p‐value = 0.33) (figure
69).
Figure 69 : Répartition des soins apportés à l'ombilic en fonction de la maladie
111
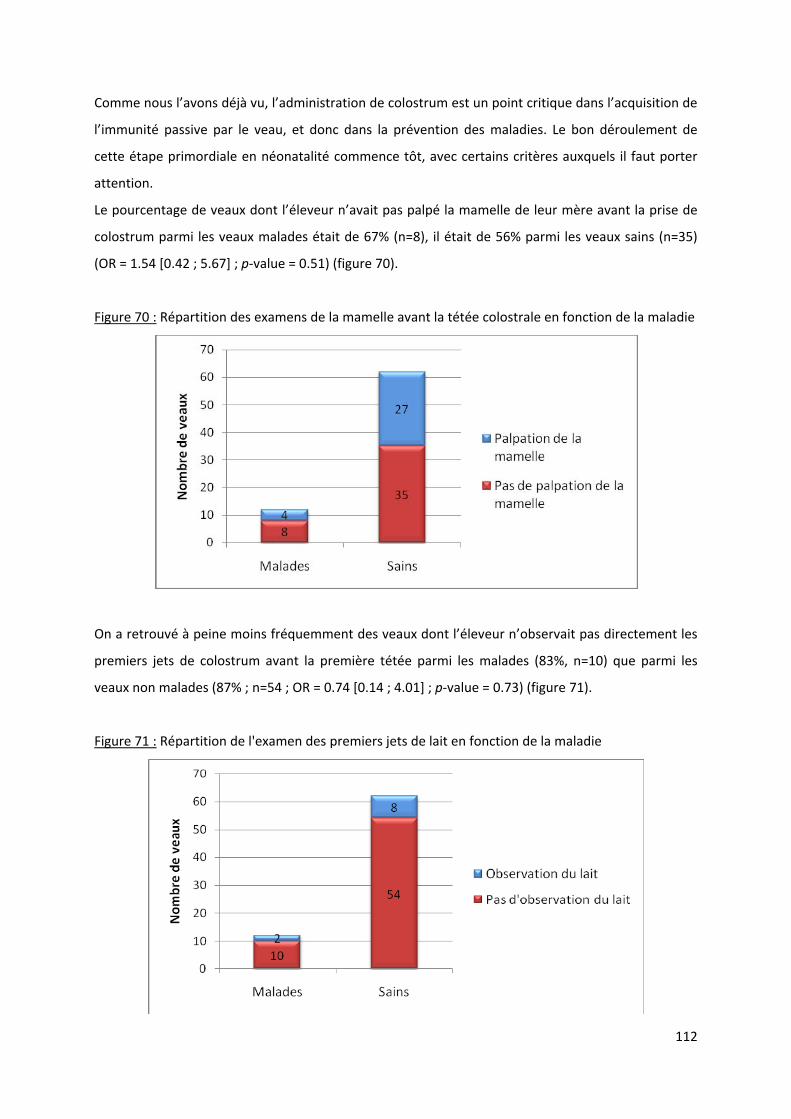
Comme nous l’avons déjà vu, l’administration de colostrum est un point critique dans l’acquisition de
l’immunité passive par le veau, et donc dans la prévention des maladies. Le bon déroulement de
cette étape primordiale en néonatalité commence tôt, avec certains critères auxquels il faut porter
attention.
Le pourcentage de veaux dont l’éleveur n’avait pas palpé la mamelle de leur mère avant la prise de
colostrum parmi les veaux malades était de 67% (n=8), il était de 56% parmi les veaux sains (n=35)
(OR = 1.54 [0.42 ; 5.67] ; p‐value = 0.51) (figure 70).
Figure 70 : Répartition des examens de la mamelle avant la tétée colostrale en fonction de la maladie
On a retrouvé à peine moins fréquemment des veaux dont l’éleveur n’observait pas directement les
premiers jets de colostrum avant la première tétée parmi les malades (83%, n=10) que parmi les
veaux non malades (87% ; n=54 ; OR = 0.74 [0.14 ; 4.01] ; p‐value = 0.73) (figure 71).
Figure 71 : Répartition de l'examen des premiers jets de lait en fonction de la maladie
112
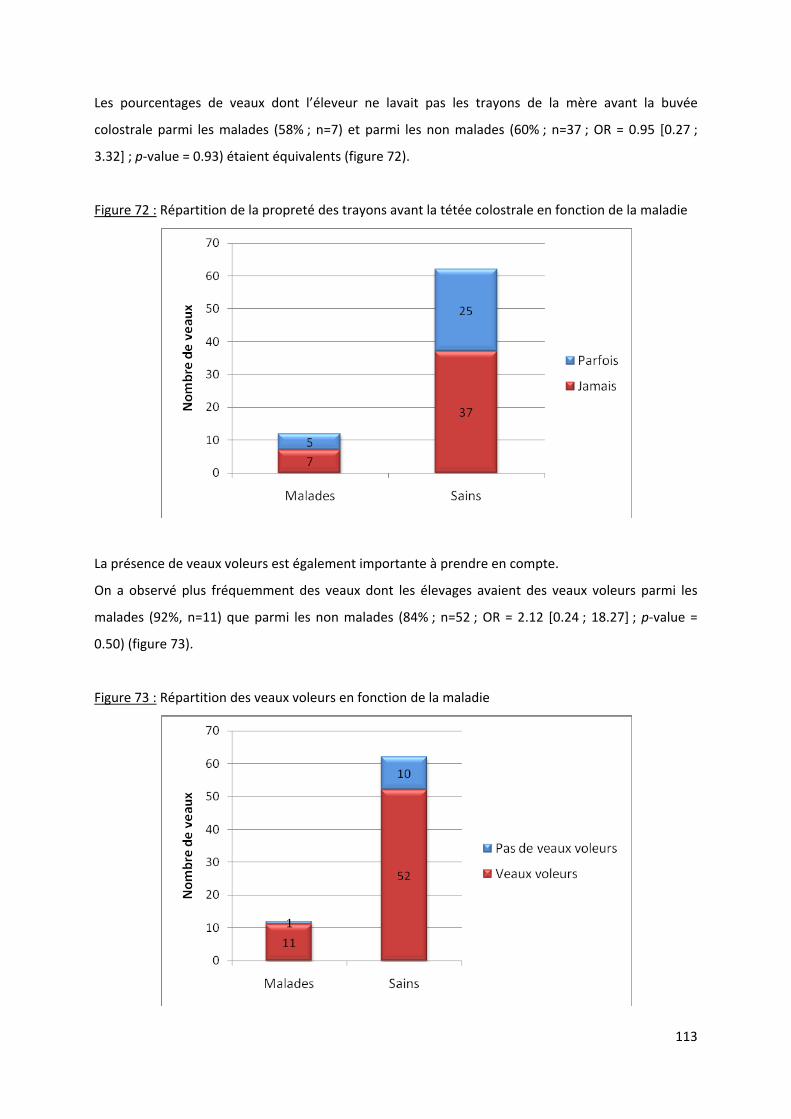
Les pourcentages de veaux dont l’éleveur ne lavait pas les trayons de la mère avant la buvée
colostrale parmi les malades (58% ; n=7) et parmi les non malades (60% ; n=37 ; OR = 0.95 [0.27 ;
3.32] ; p‐value = 0.93) étaient équivalents (figure 72).
Figure 72 : Répartition de la propreté des trayons avant la tétée colostrale en fonction de la maladie
La présence de veaux voleurs est également importante à prendre en compte.
On a observé plus fréquemment des veaux dont les élevages avaient des veaux voleurs parmi les
malades (92%, n=11) que parmi les non malades (84% ; n=52 ; OR = 2.12 [0.24 ; 18.27] ; p‐value =
0.50) (figure 73).
Figure 73 : Répartition des veaux voleurs en fonction de la maladie
113
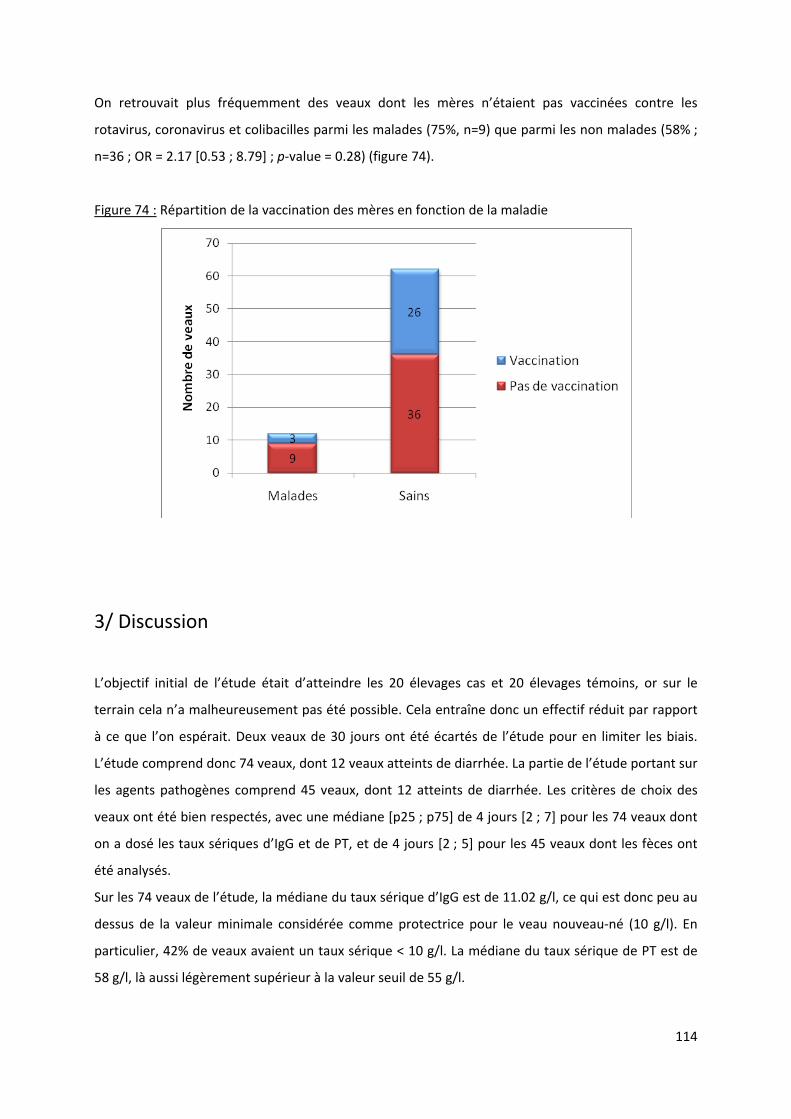
On retrouvait plus fréquemment des veaux dont les mères n’étaient pas vaccinées contre les
rotavirus, coronavirus et colibacilles parmi les malades (75%, n=9) que parmi les non malades (58% ;
n=36 ; OR = 2.17 [0.53 ; 8.79] ; p‐value = 0.28) (figure 74).
Figure 74 : Répartition de la vaccination des mères en fonction de la maladie
3/ Discussion
L’objectif initial de l’étude était d’atteindre les 20 élevages cas et 20 élevages témoins, or sur le
terrain cela n’a malheureusement pas été possible. Cela entraîne donc un effectif réduit par rapport
à ce que l’on espérait. Deux veaux de 30 jours ont été écartés de l’étude pour en limiter les biais.
L’étude comprend donc 74 veaux, dont 12 veaux atteints de diarrhée. La partie de l’étude portant sur
les agents pathogènes comprend 45 veaux, dont 12 atteints de diarrhée. Les critères de choix des
veaux ont été bien respectés, avec une médiane [p25 ; p75] de 4 jours [2 ; 7] pour les 74 veaux dont
on a dosé les taux sériques d’IgG et de PT, et de 4 jours [2 ; 5] pour les 45 veaux dont les fèces ont
été analysés.
Sur les 74 veaux de l’étude, la médiane du taux sérique d’IgG est de 11.02 g/l, ce qui est donc peu au
dessus de la valeur minimale considérée comme protectrice pour le veau nouveau‐né (10 g/l). En
particulier, 42% de veaux avaient un taux sérique < 10 g/l. La médiane du taux sérique de PT est de
58 g/l, là aussi légèrement supérieur à la valeur seuil de 55 g/l.
114

On remarque que l’on n’a aucun veau en diarrhée qui présente une hyperthermie. Cela peut être
imputé au temps de réaction de l’éleveur pour appeler le vétérinaire, le pic d’hyperthermie pouvant
être fugace, bien que le délai moyen avant appel du vétérinaire soit de 12 heures.
Les agents pathogènes mis en évidence dans l’étude sont les rotavirus, les coronavirus,
Cryptosporidium parvum et E. coli F5 (anciennement K99). Ces 4 agents pathogènes font partie des
agents pathogènes rencontrés le plus fréquemment dans les diarrhées néonatales des veaux.
Le dénombrement des colibacilles au laboratoire nous montre que pour 41 veaux sur 45, les valeurs
obtenues sont supérieures à 106 bactéries/gramme de fèces. Pour les valeurs inférieures à 106
bactéries/gramme de fèces on peut préciser que cela concerne 4 veaux malades. Ces résultats
peuvent être expliqués par le délai d’acheminement des prélèvements vers le LDA 23. En effet, le
laboratoire précise que les résultats ne peuvent être interprétés que si le prélèvement a été conservé
à + 4°C et acheminé au laboratoire le jour même. En pratique, les prélèvements ont été conservés au
réfrigérateur dans les plus brefs délais après collecte, mais ils ont été acheminés vers le laboratoire
avec des délais allant de 12 heures à 4 jours, les prélèvements étant récupérés par un coursier deux
fois par semaine à la clinique.
L’étude a été menée dans la Creuse, en clientèle allaitante dont les deux principales races de vaches
sont les charolaises et les limousines. Les races des élevages n’ont pas été prises en compte dans le
choix des élevages cas et témoin.
Dans l’ensemble, les bâtiments d’élevage étaient propres, avec 5% des élevages qui avaient de la
moisissure dans les bâtiments, 5% des élevages qui avaient de la condensation et 11% dans lesquels
on pouvait trouver d’autres espèces d’animaux présentes (des poules en l’occurrence).
La ventilation dans les bâtiments est plutôt bonne, avec seulement 16% des élevages dont la
ventilation est mauvaise (bâtiments sous‐ventilés ou trop de courants d’air).
Les conditions d’hygiène de vêlage ne sont pas optimales car seulement 63% des éleveurs se lavent
les mains avant de vêler une vache, et uniquement 26% des éleveurs utilisent des gants de vêlage.
On observe également dans cette étude que seulement 26% des élevages possèdent un lieu qu’ils
dédient spécialement et uniquement aux vêlages. Que ce soit sur ces lieux spécifiques ou non, on a
84% des lieux de vêlage qui sont propres, et 32% seulement qui sont nettoyés après un vêlage.
115

L’examen de la mamelle juste avant la première tétée peut donner des indications quant à un
éventuel souci, de mammite par exemple. Quatre‐vingt neuf pourcents des éleveurs disent faire un
examen visuel de la mamelle, 37% disent la palper et 11% seulement observent directement les
premiers jets.
L’état de propreté des mamelles et notamment des trayons avant la prise de colostrum et avant les
tétées ultérieures est important. En effet si les trayons sont souillés, il y a risque de transmission
fécale‐orale d’agents pathogènes. Malgré tout, aucun éleveur ne nettoie les trayons
systématiquement avant la tétée colostrale, 37% les nettoient parfois, et 63% ne le font jamais.
L’isolement des mères avec leur veau permet de mieux surveiller la prise de colostrum, d’être sûr
que seul son veau boira le colostrum. Seulement 42% des éleveurs isolent les veaux avec leur mère
après la naissance.
La quasi‐totalité des éleveurs contrôlent la prise de colostrum (95%), les 5% restant contrôlent
uniquement si le vêlage a été difficile. Lorsqu’un veau refuse de téter, 47% le font boire au biberon,
43% à la sonde, 5% appellent le vétérinaire et 5% ne font rien. Enfin, si une vache n’a pas de
colostrum, 63% des éleveurs donnent du colostrum synthétique, 32% prennent du colostrum dans
leur réserve de colostrum et 5% ne se sont jamais retrouvés dans cette situation.
Quatre‐vingt quatre pourcents des éleveurs reconnaissent avoir des veaux voleurs, cas fréquent dans
des élevages où les vaches prêtes à vêler sont présentes dans les mêmes enclos que les vaches ayant
déjà vêlé. Les veaux voleurs tètent le colostrum des vaches sur le point de vêler, et privent donc le
futur veau de son colostrum, ce qui a pour conséquence de baisser son apport immunitaire passif.
Concernant la gestion des veaux malades, 26% des éleveurs isolent toujours les malades, 37% les
isolent parfois et 37% ne les isolent jamais. Parmi les élevages qui isolent les veaux lorsqu’ils sont
malades, 67% réintroduisent les veaux dès la fin des symptômes, 16% les réintroduisent dès la fin du
traitement et 5% attendent plusieurs jours après la fin du traitement pour remettre les veaux
malades avec les autres animaux.
On n’a que 37% des élevages qui vaccinent les vaches contre les agents des diarrhées néonatales,
tous ces élevages vaccinent contre les rotavirus, coronavirus et colibacilles.
116

Les expositions qui ont été mises en relation avec la présence ou non de diarrhées ont été des
expositions qui pouvaient présenter un intérêt particulier, ou celles qui présentaient un OR > 2 (ou
inférieur à 0,5). Dans la très grande majorité des cas, le p n’était pas significatif (p > 0,05). Par
conséquent, la différence des pourcentages pouvait, dans tous ces cas là, être relativement
facilement attribuée au hasard. Cependant, il nous est paru intéressant de repérer les expositions
dont l’OR était supérieur à 2, car il est possible que dans ces cas là, on ait manqué de puissance
statistique pour mettre en évidence l’association entre l’exposition et la présence de diarrhées.
On observe qu’il y a moins de femelles parmi les veaux malades (25%) que parmi les veaux sains (53%
; OR = 0.29 [0.07 ; 1.19]). Bien que cet OR ne soit pas significatif, on peut néanmoins se poser la
question d’une éventuelle sensibilité liée au sexe, les femelles seraient alors moins sensibles que les
mâles. La même étude avec une population plus importante pourrait répondre à cette question. Il est
également permis de se demander si les éleveurs ne font pas appel au vétérinaire de façon sélective,
en choisissant plus de faire soigner les mâles ayant des valeurs bouchères plus importantes que les
femelles. Dans cette hypothèse, un relevé objectif de tous les individus des élevages de l’étude nous
aurait peut être conduit à un résultat différent.
On remarque que l’on n’obtient pas de différences entre les veaux malades et non malades sur la
proportion de veaux de génisses (33% versus 35%, respectivement), contrairement à ce à quoi on
aurait pu s’attendre, les colostrums de vaches étant en général plus riches en IgG et notamment en
IgG dirigées contre les agents pathogènes de l’élevage. On ne peut pas exclure qu’il y ait eu des biais
de confusion qui ont pu masquer une éventuelle association, on ne peut également pas exclure que
l’on ait obtenu par hasard une absence totale d’association en raison des faibles effectifs de l’étude.
Bien que les tests du χ2 ne soient pas significatifs, on observe que l’on a plus de veaux malades dont
les taux d’IgG sont inférieurs à 10 g/l (58%) que de veaux sains dont les taux d’IgG sont inférieurs à 10
g/l (39% ; OR = 2.22), et on a plus de veaux malades dont les taux de PT sont inférieurs à 55 g/l (50%)
que de veaux sains dont les taux d’IgG sont inférieurs à 55 g/l (31% ; OR = 2.00). Cela paraît logique
étant donné que ces taux reflètent l’immunité des veaux, plus les taux sont bas, moins les veaux sont
protégés. Les veaux ayant des taux bas et n’étant pas malades peuvent être des veaux vivant dans
des élevages dont l’environnement est relativement propre, avec une bonne hygiène d’élevage, tout
en gardant à l’esprit que ces veaux sont plus sensibles à des agents pathogènes que ceux qui ont des
taux élevés en IgG et PT.
117

Les rotavirus, coronavirus et Cryptosporidium parvum sont mis en évidence aussi bien chez les veaux
malades que chez des veaux non malades. En fonction de l’âge de ces veaux porteurs de ces agents
pathogènes mais non malades, deux hypothèses sont possibles : soit les veaux positifs et non
malades sont des porteurs sains, soit ce sont des veaux en phase d’incubation, et ils n’ont donc pas
encore déclaré la maladie.
Les résultats pour la détection de E. coli F5 ont été différents selon le type de détection de la
bactérie. En effet, dans les 2 cas on retrouve 5 prélèvements positifs, mais ils ne concernent pas les
mêmes veaux. Seuls 3 prélèvements donnent un résultat positif à E. coli F5 via le tetrakit et au
laboratoire. Les deux autres prélèvements dont le tetrakit donne un résultat positif sont des fèces de
veaux malades, tandis que les deux autres prélèvements positifs au laboratoire sont des fèces de
veaux sains. La sensibilité des tigettes pour la détection des E. coli F5 du tetrakit est de 90% et la
spécificité de 98.5%. Pour ce qui est de la détection en laboratoire, la spécificité est proche de 100%
(utilisation d’anticorps monoclonaux), la sensibilité par contre est inférieure à 90%, le typage se
faisant après une première culture sur gélose puis repiquage de 2 à 4 colonies, toutes les colonies
d’un prélèvement ne sont pas typées, seules celles qui sont majoritaires ou qui paraissent
intéressantes de par leur aspect le sont. On a donc pu passer à côté du typage de certaines souches
de colibacilles. Les deux veaux positifs au laboratoire et non positifs avec le tetrakit étant des
animaux asymptomatiques au moment du prélèvement, on peut supposer que ce sont des animaux
en incubation ou des porteurs sains. Par contre, la sensibilité des tigettes étant supérieure à la
sensibilité du laboratoire, on peut se poser la question du résultat négatif donné par la tigette. On
peut soit avoir une erreur de lecture, avec éventuellement un résultat qui était très légèrement
positif qui n’a pas été bien lu, ou alors une mauvaise utilisation des tigettes pour ces deux
prélèvements. L’utilisation d’un autre kit de détection rapide aurait peut‐être amené à des résultats
positifs pour ces deux veaux, comme au laboratoire, ou nous aurait peut‐être permis de détecter
d’autres souches de E. coli.
On peut également dire, tout comme pour les agents pathogènes précédemment cités, que les deux
veaux ayant donné un résultat positif mais étant non malades sont probablement soit des porteurs
sains ou des veaux en incubation.
E. coli CS31A est retrouvé chez 29% des veaux, le colibacille est présent chez plus d’un quart des
veaux non malades. Après avoir longtemps pensé que ce colibacille avait un rôle important dans les
gastro‐entérites paralysantes, le nombre d’individus porteurs sains étant toujours relativement
élevé, comme ici, son implication est maintenant mise en doute.
118

La quasi‐totalité des prélèvements avaient un dénombrement de colibacilles supérieur à 106
bactéries par gramme de fèces. On peut imputer ces résultats au délai d’acheminement trop long des
fèces au laboratoire. Les mêmes analyses réalisées dans des conditions de conservation et de
transport adéquats auraient sans aucun doute conduit à des résultats différents.
De manière générale, plus on avance dans la saison de vêlage, plus les veaux nouveau‐nés sont
susceptibles de rencontrer des agents pathogènes. En effet, plus on avance dans la saison de vêlage,
plus les locaux et l’environnement sont censés abriter des agents pathogènes, et de plus les veaux
nés en début de saison de vêlage portent souvent asymptomatiquement des agents pathogènes
qu’ils excrètent dans l’environnement. Le fait que l’on n’observe pas de différences entre les
élevages ayant un faible nombre de vêlages par saison de vêlage et ceux qui en ont beaucoup peut
être attribué au fait que la quasi‐totalité des élevages étaient en début de saison de vêlage au
moment de l’étude. La même étude conduite en fin de saison de vêlage aurait peut‐être apporté des
résultats différents.
On a plus de veaux malades élevés dans des élevages ayant une mauvaise ventilation (sous‐
ventilation ou courants d’airs importants) (33%) que de veaux sains élevés dans des élevages ayant
une mauvaise ventilation (5% ; OR = 9.83 ; p < 0.01). La ventilation semble avoir un rôle en tant que
facteur de risque pour les diarrhées néonatales, avec un OR > 9. On ne peut cependant pas exclure
un biais de confusion, et par conséquent le fait que le véritable OR quantifiant l’association entre la
ventilation et la présence de diarrhées soit éventuellement plus faible.
De même, bien que le χ2 ne soit pas significatif, les veaux qui sont dans des élevages dont le sol est
en terre battue semblent avoir plus de risque d’être malades que les veaux vivant dans des élevages
dont le sol est en béton. On peut penser que la terre battue abrite plus facilement les agents
pathogènes que le béton, et également que les bâtiments dont le sol est en béton sont plus aisément
nettoyés et désinfectés que les bâtiments dont le sol est en terre battue. Néanmoins, on ne peut
exclure que ces résultats soient dus au hasard étant donné la petite taille de l’échantillon.
Les résultats des données concernant l’hygiène de vêlage et l’hygiène des lieux de vêlage ne donnent
pas de résultats présentant des différences significatives, bien que l’on constate par exemple que les
éleveurs qui n’utilisent pas de gants de vêlage sont plus fréquemment confrontés à des veaux
malades que les autres. L’étude manque donc de puissance statistique, on pourrait donc penser que
la même étude réalisée avec une population plus grande conduirait à des résultats significatifs.
119

On constate qu’il y a plus fréquemment des veaux dont l’ombilic n’est pas désinfecté parmi les
malades que parmi les non malades, l’OR s’approchant de 3 (χ2 non significatif). On peut expliquer
ces résultats par le fait que si un cordon ombilical n’est pas désinfecté, il peut représenter une entrée
de germes, et donc affaiblir le veau. Là encore, l’étude manque de puissance statistique pour que
l’on puisse obtenir des résultats significatifs si, au niveau de la population, il existe réellement une
association entre la désinfection de l’ombilic et la présence de diarrhées.
Plusieurs facteurs peuvent donner des indications sur la qualité du colostrum transmis de la mère à
son veau, de même que les conditions d’hygiène dans lesquelles ce transfert est réalisé. On
remarque dans un premier temps que les éleveurs n’accordent pas assez d’importance à cette étape,
pourtant primordiale en néonatalité. En effet, un grand nombre d’éleveurs laissent le veau se
débrouiller et téter seul, alors qu’une surveillance attentive est recommandée afin d’optimiser le
transfert passif de l’immunité de la vache à son veau. Trop peu d’éleveurs observent directement les
premiers jets de colostrum, même si la plupart examinent correctement la mamelle afin de vérifier si
la vache ne présente pas de mammite. De plus, seulement un peu plus de la moitié des éleveurs
nettoient les trayons de la vache avant la première tétée colostrale. Enfin, 85% des éleveurs
reconnaissent avoir des veaux voleurs ce qui peut avoir une incidence sur la transmission de
l’immunité passive des mères aux veaux, car les veaux voleurs pourraient avoir bu les premiers litres
de colostrum avant la naissance d’un veau, et donc priver le nouveau‐né de son colostrum.
L’étude nous montre aussi que l’on a plus de veaux de mères non vaccinées parmi les malades que
parmi les non malades, avec un OR supérieur à 2 (χ2 non significatif). On peut donc dire que l’on a
une tendance à avoir une protection des veaux contre les diarrhées néonatales conférée par la
vaccination des mères, et transmise par l’intermédiaire du colostrum. Mais là encore, on a pu
manquer de puissance statistique pour obtenir des résultats significatifs.
120

121

CONCLUSION
Les diarrhées néonatales sont encore à ce jour une entité pathologique dont l’importance est loin
d’être négligeable pour les élevages. Cette étude avait pour but de mettre en relation le statut
immunitaire passif du veau avec la morbidité et la mortalité des veaux de diarrhée néonatale. Bien
que l’on n’obtienne pas de résultats significatifs avec le dosage des IgG et des PT, on observe qu’elles
ont un rôle dans la protection des nouveau‐nés. On a cherché également à mettre en évidence des
éventuels facteurs de risque des diarrhées néonatales, là encore on n’obtient pas de résultats
significatifs, à part pour la ventilation des bâtiments. On observe que le type de sol, l’absence de
soins aux ombilics des nouveau‐nés, la non vaccination des mères vis‐à‐vis des agents des diarrhées
néonatales, les mauvaises conditions de propreté des locaux, des matériels et des éleveurs au
moment des vêlages auraient tendance à avoir un rôle en tant que facteurs de risque dans les
diarrhées néonatales, on observe également que les conditions de ventilation sont un facteur de
risque. Une étude similaire portant sur des effectifs plus importants et sur une saison de vêlage
entière apporterait des résultats probablement plus intéressants, avec une puissance statistique
suffisante pour obtenir des résultats significatifs et faire ressortir de l’étude les expositions qui sont
réellement des facteurs de risque pour les diarrhées néonatales, grâce en particulier à l’utilisation de
modélisations idoines qui permettent de prendre en compte des biais de confusion.
Cependant, on peut déjà, de par les observations faites au cours de cette étude, mettre en évidence
certaines pratiques d’élevages qui seraient aisément modifiables et qui amélioreraient la gestion et
l’hygiène des élevages et donc mèneraient à des morbidités et mortalités moins importantes pour les
veaux vis‐à‐vis des diarrhées néonatales. Il s’agit principalement de l’hygiène du vêlage. Les éleveurs
n’accordent pas assez d’importance à la propreté des lieux de vêlage et des matériels de vêlages, qui
devraient être nettoyés et désinfectés entre chaque vêlage. De même l’hygiène des éleveurs
assistant les vaches au vêlage reste à améliorer, un lavage des mains avant et après le vêlage, ainsi
que le port de gants de vêlage à utilisation unique se révèlent être des pratiques encore peu
répandues. De plus, les éleveurs n’accordent pas assez de temps aux veaux en post‐natalité
immédiate, la gestion de l’administration du colostrum devrait être prise en main avec nettoyage des
trayons de la mère, traite, contrôle de la qualité du colostrum et administration au veau à l’aide d’un
biberon ou d’une sonde d’une quantité adéquate de colostrum, ou, à défaut, de colostrum pris dans
la réserve de colostrum (dont la mise en place est fortement conseillée afin de pallier tout soucis si
122

une vache n’a pas de colostrum ou si la qualité est insuffisante). Il est important également de pallier
le problème des veaux voleurs, en séparant les vaches ayant vêlé des autres. Bien entendu, une
bonne hygiène de l’environnement est préconisée, avec une désinfection et un vide sanitaire à
effectuer chaque année. Enfin, la vaccination systématique des mères contre les agents des diarrhées
néonatales semble être une aide non négligeable dans la lutte contre cette maladie.
123

BIBLIOGRAPHIE
1 Adams GD, Bush LJ, Horner JL, et al. Two methods for administering colostrum to newborn calves. J. Dairy Sci., 1985, 68, 773‐775.
2 Anderson RJ, House JK, Smith BP, et al. Epidemiologic and biological characteristics of salmonellosis in three dairy herds. J. Am. Vet. Med. Assoc., 2001, 219, 310‐322.
3 Archambault D, Morin G, Elazhary Y, et al. Immune response of pregnant heifers and cows to bovine rotavirus inoculation and passive protection to rotavirus infection in newborn calves fed colostral antibodies or colostral lymphocytes. Am. J. Vet. Res., 1988, 49, 1084‐1091.
4 Arthington JD. Effects of dietary copper, chromium, and vitamin E on measures of stress and immune competence in growing cattle. PhD thesis, Department of Animal Sciences and Industry, College of Agriculture, Kansas State University, Manhattan, Kan, 1995.
5 Baintner K. Transmission of antibodies from mother to young : evolutionary strategies in a proteolytic environment. Vet. Immunol. Immunopathol., 2007, 117, 153‐161.
6 Barnett SC, Sischo WM, Moore DA, et al. Evaluation of flunixin meglumine as an adjunct treatment for diarrhea in dairy calves. J. Am. Vet. Med. Assoc., 2003, 223, 1329‐1333.
7 Barrington GM, Besser TE, Gay CC, et al. Effect of prolactin on in vitro expression of the bovine mammary immunoglobulin G1 receptor. J. Dairy Sci., 1997, 80, 94‐100.
8 Bass RT, Swecker WSJr, Eversole DE. Effects of oral vitamin E supplementation during late gestation in beef cattle that calved in late winter and late summer. Am. J. Vet. Res., 2001, 62, 921‐927.
9 Baumrucker CR, Hadsell DL, Blum JW. Effects of dietary insulin‐like growth factor I on growth and insulin‐like growth factor receptors in neonatal calf intestine. J. Anim. Sci., 1994, 72, 428–433.
10 Besser TE. Concentrations of passively acquired IgG1 antibodies in the intestinal lumen of the neonatal calf. Vet. Immunol. and lmmunopathol., 1993, 38, 103‐112.
11 Besser TE and Osbom D. Effect of bovine serum albumin on passive transfer of immunoglobulin Gl to newborn calves. Vet. Immunol. Immunopathol., 1993, 37, 321‐327.
12 Besser TE, Gay CC, Pritchett L. Comparison of three methods of feeding colostrum to dairy calves. J. Am. Vet. Med. Assoc., 1991, 198, 419‐422.
13 Besser TE, Szenci O, Gay CC. Decreased colostral immunoglobulin absorption in calves with postnatal respiratory acidosis. J. Am. Vet. Med. Assoc., 1990, 196, 1239‐1243.
14 Bey R, Godden S, Lillegaard H, et al. Improving cleanliness and shelf‐life of refrigerated colostrum using heat‐treatment and chemical preservatives. In : Proc. Annu. Meet. Minnesota Dairy Health Management Conference. St. Paul, Minnesota, USA, May 2007, 15–17.
15 Bird AR, Croom WJ, Fan YK, et al. Peptide regulation of intestinal glucose absorption. J. Anim. Sci., 1996, 74, 2523‐2540.
16 Blecha GK, Bulls RC, Olson DP. Effects of prepartum protein restriction in the beef cow on immunoglobulin content in blood and colostral whey and subsequent immunoglobulin absorption by the neonatal calf. J. Anim. Sci., 1981, 53, 1174‐1180.
17 Boland W, Cortese VS, Steffen D. Interactions between vaccination, failure of passive transfer, and diarrhea in beef calves. Agri. Practice., 1995, 16, 25‐28.
18 Boraychuk VM, Gensler GE, McFall ME, et al. A real‐time PCR assay for the detection of Salmonella in a wide variety of food and food‐animal matrices. J. Food Prot., 2007, 70, 1080‐1087.
124

19 Bovine Alliance on Management and Nutrition (BAMN). A guide to colostrum and colostrums management for dairy calves. [pdf]. Mis à jour en 2001. [http://www.aphis.usda.gov/vs/ceah/ncahs/nahms/dairy/bamn/BAMNColostrum.pdf] (consulté le 05 janvier 2010).
20 Bradford P, Smith. Large Animal Internal Medicine. 4th edition. Mosby, 2008, 1872p. 21 Brar JS, Johnson DW, Muscoplat CC, et al. Maternal immunity to infectious bovine rhinotracheitis
and bovine viral diarrhea : duration and effect on vaccination in young calves. Am. J. Vet. Res., 1978, 39, 241‐244.
22 Brenner J, Samina I, Machanai B, et al. Impact of vaccination of pregnant cows on colostral IgG levels and on term of pregnancy. Field observations. Israel J. of Vet. Med., 1997, 52, 56‐59.
23 Brignole TJ, Stott GH. Effect of suckling followed by bottle feeding colostrum on immunoglobulin absorption and calf survival. J. Dairy Sci., 1980, 63, 451‐456.
24 Bryan LA. Fatal, generalized bovine herpesvirus type‐1 infection associated with a modified‐live infectious bovine rhinotracheitis/parainfluenza‐3 vaccine administered to neonatal calves. Can. Vet. J., 1994, 35, 223‐228.
25 Bühler C, Hammon H, Rossi GL. Small intestinal morphology in eight‐day‐old calves fed colostrum for different durations or only milk replacer and treated with long‐R3‐insulinlike growth factor I and growth hormone. J. Anim. Sci., 1998, 76, 758‐765.
26 Bussiéras J., Chermette R. Abrégé de parasitologie vétérinaire. Fascicule 2. Protozoologie vétérinaire. École Nationale Vétérinaire d’Alfort. Unité Pédagogique de Parasitologie et Maladies Parasitaires. 1992, 186p.
27 Calloway CD, Tyler JW, Tessman RK, et al. Comparison of refractometers and test endpoints in the measurement of serum protein concentration to assess passive transfer status in calves. J. Am. Vet. Med. Assoc., 2002, 221, 1605‐1608.
28 Castrucci G, Frigeri F, Ferrari M, Cilli V, Caleffi F, Aldrovandi V and Nigrelli A. The efficacy of colostrum from cows vaccinated with rotavirus in protecting calves to experimentally induced rotavirus infection. Comp. lmmun. Microbiol. infect. Dis., 1984, 7(1), 11‐18.
29 Chase CCL, Hurley DJ, Reber AJ. Neonatal immune development in the calf and its impact on vaccine response. Vet. Clin. Food Anim., 2008, 24, 87‐104.
30 Chazel M. Le réseaux d'épidémiosurveillance des salmonelloses bovines ‐ RESSAB. Bulletin épidémiologique,2004, 13, 5‐6.
31 Chigerwe M, Dawes ME, Tyler JW, et al. Evaluation of a cow‐side immunoassay kit for assessing IgG concentration in colostrum. J. Am. Vet. Med. Assoc., 2005, 227, 129‐131.
32 Chigerwe M, Tyler JW, Schultz LG, Middleton JR, Steevens BJ, Spain JN. Effect of colostrum administration by use of oroesophageal intubation on serum IgG concentrations in Holstein bull calves. Am. J. Vet. Res., 2008, 69, 1158‐1163.
33 Chigerwe M, Tyler JW, Middleton JR, Spain JN, Dill JS, Steevens BJ. Comparison of four methods to assess colostral IgG concentration in dairy cows. J. Am. Vet. Med. Assoc., 2008, 233, 761‐766.
34 Chigerwe M, Tyler JW, Nagy DW, Middleton JR. Frequency of detectable serum IgG concentrations in precolostral calves. Am. J. Vet. Res., 2008, 69, 791‐795.
35 Chigerwe M, Tyler JW, Summers MK, Middleton JR, Schultz LG, Nagy DW. Evaluation of factors affecting serum IgG concentrations in bottle‐fed calves. J. Am. Vet. Med. Assoc., 2009, 234, 785‐789.
36 Chinsangaram J, Schore CE, Guterbock W, Weaver LD, Osburn BI, Prevalence of groupe A and group B rotaviruses in the feces of neonatal dairy calves from California. Comp. Immun. Microbiol. Infect. Dis., 1995, 18(2), 93‐103.
37 Cipriano E, Morrill JL, Anderson NV. Effect of dietary vitamin E on the immune responses of calves. J. Dairy Sci., 1982, 65, 2357‐2365.
125

38 Clark. Bovine Coronavirus. Br. Vet. J., 1993, 51, 149. 39 Constable P. Antimicrobial use in the treatment of calf diarrhea. J. Vet. Intern. Med., 2004, 18, 8‐17.40 Curtiss R, Kelly SM, Hassan JO. Live oral avirulent Salmonella vaccines. Vet. Microbiol., 1993, 37,
397‐405.
41 Davis CL, Drackley JK. In: The development, nutrition, and management of the young calf. 1st edition. Ames (IA): Iowa State University Press, 1998, 179‐206.
42 De Jong H, Ekdahl MO. Salmonellosis in calves ‐ the effect of dose rate and other factors on the transmission. N. Z. Vet. J., 1965, 13, 59‐64.
43 De Leeuw PW, Ellens DJ, Talmon FP, et al. Rotavirus infections in calves: efficacy of oral vaccination in endemically infected herds. Res. Vet. Sci., 1980, 29, 142‐147.
44 De Nise SK, Robison JD, Stott GH, et al. Effects of passive immunity on subsequent production in dairy heifers. J. Dairy Sci., 1989, 72, 552‐554.
45 Dewell RD, Hungerford LL, Keen JE, et al. Association of neonatal serum immunoglobulin G1 concentration with health and performance in beef calves. J. Am. Vet. Med. Assoc., 2006, 228, 914‐921.
46 Dixon FJ, Weigle WO, Vasquez JJ. Metabolism and mammary secretion of serum protein in the cow. Lab. Invest., 1961, 10, 216‐237.
47 Donovan D, Reber A, Gabbard J, et al. Effect of maternal cells transferred with colostrums on cellular response to pathogen antigens in neonatal calves. Am. J. Vet. Res., 2007, 68, 778‐782.
48 Drewry JJ, Quigley JD, Geiser DR. Effect of high arterial carbon dioxide tension on efficiency of immunoglobulin G absorption in calves. Am. J. Vet. Res., 1999, 60, 609‐614.
49 Duhamel GE. Characterization of bovine mammary lymphocytes and their effects on neonatal calf immunity. Ann Arbor (MI): UMI Dissertation Services, 1993.
50 Edwards SA, Broom DM. The period between birth and first suckling in dairy calves. Res. Vet. Sci., 1979, 26, 255‐256.
51 Elfstrand L, Lindmark‐Månsson H, Paulsson M, et al. Immunoglobulins, growth factors and growth hormone in bovine colostrum and the effects of processing. Int. Dairy J., 2002, 12, 879‐887.
52 Ellis JA, Hassard LE, Cortese VS, et al. Effects of perinatal vaccination on humoral and cellular immune responses in cows and young calves. J. Am. Vet. Med. Assoc., 1996, 208, 393‐399.
53 Eriksson E, Aspan A. Comparison of culture, ELISA and PCR techniques for Salmonella detection in faecal samples for cattle, pig and poultry. BMC Vet. Res., 2007, 3, 21.
54 Espinasse J, Navetat H, Contrepois G, Baroux D, Schelcher F. A new diarrhoeic syndrome with ataxia in young Charolais calves : clinical and microbiological studies. The Veterinary Record, 1991, 128 (18), 422‐425.
55 Faber SN, Faber NE, McCauley TC, et al. Effects of colostrum ingestion on lactational performance. Prof. Anim. Sci. 2005, 21, 420‐425.
56 Fecteau M, House JK, Kotarski SF, et al. Efficacy of ceftiofur for treatment of experimental salmonellosis in neonatal calves. Am. J. Vet. Res., 2003, 64, 918‐925.
57 Fiche technique Bio‐X Diagnostics. Tigettes tétravalentes (Rota, Corona, E. coli F5, Cryptosporidium) BIO K 156. [pdf]. [http://www.biox.com/UDTData/13/UDTFrenchDatasheet/TetraStrips BIO K 156 (F) DS.pdf], (consulté le 05 janvier 2010).
58 Fiche technique Protéines‐Kit (PROT), REF 61 602, (document interne), 2007 ; bioMérieux. 59 Fiche technique SAS‐MX Serum Protein, REF 100 100, (document interne), 2007 ; Helena
Biosciences Europe.
60 Fichou E. Enquête de terrain sur l'étiologie microbienne des diarrhées néonatales de veaux et sur la sensibilité aux anti‐infectieux des colibacilles isolés. Thèse Méd. Vét., Nantes, 2003, n°92, 104p.
61 Foley JA, Otterby DE. Availability, storage, treatment, composition, and feeding value of surplus colostrum: a review. J. Dairy. Sci., 1978, 61, 1033‐1060.
126

62 Foster DM, Smith GW, Sanner TR, et al. Serum IgG and total protein concentrations in dairy calves fed two colostrum replacement products. J. Am. Vet. Med. Assoc., 2006, 229, 1282‐1285.
63 Francisco SFA, Quigley JD. Serum immunoglobulin concentrations after feeding maternal colostrum or maternal colostrum plus colostral supplement to dairy calves. Am. J. Vet. Res., 1993, 54, 1051‐1054.
64 Gelberg HB. Alimentary system 1. In: McGavin MD, Carlton WW, Zachary JF, editors. Thomson’s special veterinary pathology. 3rd edition. St. Louis (MO): Mosby, Inc., 2001, 1‐79.
65 George LW. Pain control in food animals. In: Steffey EP, editor. Recent advances in anesthetic management of large domestic animals. Ithaca (NY): International Veterinary Information Service, 2003.
66 Godden S. Colostrum management for dairy calves. Vet. Clin. North Am. Food Anim. Pract., 2008, 24, 19‐39.
67 Godden SM, McMartin S, Feirtag JM. Heat‐treatment of bovine colostrum II: effects of heating duration on pathogen viability and immunoglobulin G. J. Dairy Sci., 2006, 89, 3476‐3483.
68 Godden SM, Smith S, Feirtag JM, et al. Effect of on‐farm commercial batch pasteurization of colostrum on colostrum and serum immunoglobulin concentrations in commercial dairy calves. J. Dairy Sci., 2003, 86, 1503‐1512.
69 Groothuis DG, van Miert AS. Salmonellosis in veal calves: some therapeutic aspects. Vet. Q., 1987, 9, 91‐96.
70 Grusenmeyer DJ, Ryan CM, Galton DM, et al. Shortening the dry period from 60 to 40 days does not affect colostrum quality but decreases colostrum yield by Holstein cows. J. Dairy Sci., 2006, 89(Suppl 1), 336.
71 Guy MA, McFadden TB, Cockrell DC, et al. Regulation of colostrum formation in beef and dairy cows. J. Dairy Sci., 1994, 77, 3002‐3007.
72 Hodgins DC, Shewen PE. Preparturient vaccination to enhance passive immunity to the capsular polysaccharide of Pasteurella haemolytica A1. Vet. Immunol. Immunopathol., 1996, 50, 67‐77.
73 Holloway NM, Tyler JW, Lakritz J, Carlson SL, Holle J. Serum immunoglobulin G concentrations in calves fed fresh and frozen colostrums. J. Am. Vet. Med. Assoc., 2001, 219(3).
74 Holt P. Host susceptibility, resistance and immunity to Salmonella in animals. In: Wray C, Wray W, editors. Salmonella in domestic animals. New York : CABI Publishing, 2000.
75 House JK, Ontiveros MM, Blackmer NM, et al. Evaluation of an autogenous Salmonella bacterin and a modified live Salmonella serotype Choleraesuis vaccine on a commercial dairy farm. Am. J. Vet. Res., 2001, 62, 1897‐1902.
76 Israel EJ, Patel VK, Taylor SF, et al. Requirement for a beta 2‐microglobulin‐associated Fc receptor for acquisition of maternal IgG by fetal and neonatal mice. J. Immunol., 1995, 154, 6246‐6251.
77 Jacobsen KL, Arbtan KD. Interferon activity in bovine colostrum and milk. In: Proceedings of the XVII World Buiatrics/XXV American Association of Bovine Practitioners Congress. Minneapolis (MN), 1992, 1‐2.
78 James RE, Polan CE. Effect of orally administered duodenal fluid on serum proteins in neonatal calves. J. Dairy Sci., 1978, 61, 1444‐1449.
79 James RE, Polan CE, Cummins KA. Influence of administered indigenous microorganisms on uptake of [iodine‐125] gamma‐globulin in vivo by intestinal segments of neonatal calves. J. Dairy Sci., 1981, 64(1), 52‐61.
80 Johnson J, Godden S, Molitor T, et al. The effect of feeding heat‐treated colostrum on passive transfer of cellular and humoral immune parameters in neonatal dairy calves. J. Dairy Sci., 2007, 90, 5189‐5198.
81 Johnston WS, MacLachlan GK, Hopkins GF. The possible involvement of seagulls (Larus sp) in the transmission of salmonella in dairy cattle. Vet. Rec., 1979, 105, 526‐527.
127

82 Jones CM, James RE, Quigley JD, et al. Influence of pooled colostrum or colostrums replacement on IgG and evaluation of animal plasma in milk replacer. J. Dairy Sci., 2004, 87, 1806‐1814.
83 Jones PW, Collins P, Aitken MM. Passive protection of calves against experimental infection with Salmonella typhimurium. Vet. Rec., 1988, 123, 536‐541.
84 Jonsson ME, Eriksson E, Boqvist S, Urdahl AM, Aspán A. Experimental infection in calves with a specific subtype of verocytotoxin‐producing Escherichia coli O157:H7 of bovine origin. Act. Vet. Scan., 2009, 51:43 doi:10.1186/1751‐0147‐51‐43.
85 Kapil S, Trent AM, Goyal SM. Antibody responses in spiral colon, ileum, and jejunum of bovine coronavirus‐infected neonatal calves. Comp. Immun. Microbiol. Infect. Dis., 1994, 17(2), 139‐149.
86 Kaske M, Werner A, Schuberth HJ, et al. Colostrum management in calves : effects of drenching vs. bottle feeding. J. Anim. Physiol. Anim. Nutr., 2005, 89, 151‐157.
87 Kussendrager K. Lactoferrin and lactoperoxydase. Bioactive milk proteins. Intl. Food Ingredients 1993, 6, 17‐21.
88 Lacetera N, Bernabucci U, Ronchi B, et al. Effects of selenium and vitamin E administration during a late stage of pregnancy on colostrum and milk production in dairy cows, and on passive immunity and growth of their offspring. Am. J. Vet. Res. 1996, 57, 1776‐1780.
89 Larson BL, Heary HL Jr, Devery JE. Immunoglobulin production and transport by the mammary gland. J. Dairy Sci., 1980, 63, 665‐671.
90 Lateur‐Rowet HJM, Breukink HJ. The failure of the oesophageal groove reflex, when fluids are given with an oesophageal feeder to newborn and young calves. Vet. Q., 1983, 5, 68‐74.
91 Le Jan C. Cellular components of mammary secretions and neonatal immunity : a review. Vet. Res., 1996, 27, 403‐417.
92 Liebler‐Tenorio EM, Riedel‐Caspari G, Pohlenz JF. Uptake of colostral leukocytes in the intestinal tract of newborn calves. Vet. Immunol. Immunopathol., 2002, 85, 33‐40.
93 Lorino T, Daudin JJ, Robin S, Sanaa M. Factors associated with time to neonatal diarrhoea in French beef calves. Prev. Vet. Med., 2005, 68, 91‐102.
94 Martín‐Gómez S, Álvarez‐Sánchez MA, Rojo‐Vázquez FA. Immunization protocols against Cryptosporidium parvum in ovines : protection in suckling lambs. Vet. Parasitol., 2005, 129, 11‐20.
95 Mastroeni P, Chabalgoity JA, Dustan SJ, et al. Salmonella : immune responses and vaccines. Vet. J., 2000, 162, 132‐164.
96 Maunsell FP, Morin DE, Constable PD, et al. Effects of mastitis on the volume and composition of colostrum produced by Holstein cows. J. Dairy Sci., 1998, 81, 1291‐1299.
97 Maunsell FP, Morin DE, Constable PD, et al. Use of mammary gland and colostral characteristics for prediction of colostral IgG1 concentration and intramammary infection in Holstein cows. J. Am. Vet. Med. Assoc., 1999, 214, 1817‐1823.
98 McBeath DG, Penhale WJ, Logan EF. An examination of the influence of husbandry on the plasma immunoglobulin level of the newborn calf, using a rapid refractometer test for assessing immunoglobulin content. Vet. Rec., 1971, 88, 266‐270.
99 McGavin M, Carlton W, Zachary J. Thompson's special veterinary pathology. 3rd edition. Mosby Inc, 2001, 755p.
100 McGuirk SM, Collins M. Managing the production, storage and delivery of colostrum. Vet. Clin. North Am. Food Anim. Pract., 2004, 20(3), 593‐603.
101 McMartin S, Godden S, Metzger L, et al. Heat treatment of bovine colostrum. I: effects of temperature on viscosity and immunoglobulin G level. J. Dairy Sci., 2006, 89, 2110‐2118.
102 Mee JF, O’Farrell KJ, Reitsma P, et al. Effect of a why protein concentrate used as a colostrums substitute or supplement on calf immunity, weight gain, and health. J. Dairy Sci., 1996, 79, 886‐889.
128

103 Menanteau‐Horta AM, Ames TR, Johnson DW, et al. Effect of maternal antibody upon vaccination with infectious bovine rhinotracheitis and bovine virus diarrhea vaccines. Can. J. Comp. Med., 1985, 49, 10‐14.
104 Mohler VL, Izzo MM, House JK. Salmonella in Calves. Vet. Clin. Food Anim., 2009, 25, 37‐54. 105 Mohler VL, Heithoff DM, Mahan MJ, et al. Cross‐protective immunity in calves conferred by a DNA
adenine methylase deficient Salmonella enterica sérovar Typhimurium vaccine. Vaccine, 2006, 24, 1339‐1345.
106 Mohler VL, Heithoff DM, Mahan MJ, et al. Cross‐protective immunity conferred by a DNA adenine methylase deficient Salmonella enterica serovar Typhimurium vaccine in calves challenged with Salmonella serovar Newport. Vaccine, 2008, 26, 1751‐1758.
107 Moore M, Tyler JW, Chigerwe M, Dawes ME and Middleton JR. Effect of delayed colostrum collection on colostral IgG concentration in dairy cows. J. Am. Vet. Med. Assoc., 2005, 226, 1375‐1377.
108 Morin DE, Constable PD, Maunsell FP, et al. Factors associated with colostral specific gravity in dairy cows. J. Dairy Sci., 2001, 84, 937‐943.
109 Morin DE, McCoy GC, Hurley WL. Effects of quality, quantity, and timing of colostrums feeding and addition of a dried colostrum supplement on immunoglobulin G1 absorption in Holstein bull calves. J. Dairy Sci., 1997, 80, 747‐753.
110 Mortola ME, Pennimpede PE, Arauz PM, et al. Calf salmonellosis : prophylaxis by maternal immunization. Avances en Ciencias Veterinarias 1992, 7, 203‐208.
111 Muller LD, Ellinger DK. Colostral immunoglobulin concentrations among breeds of dairy cattle. J. Dairy Sci., 1981, 64, 1727‐1730.
112 Murakami T, Hirano N, Inoue A, et al. Transfer of antibodies against viruses of calf diarrhea from cows to their offspring via colostrum. Jap. J. Vet. Sci., 1985, 47, 507‐510.
113 Myers LL, Snodgrass DR. Colostral and milk antibody titers in cows vaccinated with a modified live rotavirus‐coronavirus vaccine. J. Am. Vet. Med. Assoc., 1982, 181, 486‐488.
114 Nardone A, Lacetera N, Bernabucci U, et al. Composition of colostrum from dairy heifers exposed to high air temperatures during late pregnancy and the early postpartum period. J. Dairy Sci., 1997, 80, 838‐844.
115 National Animal Health Monitoring System (NAHMS). Dairy heifer morbidity, mortality, and health management focusing on preweaned heifers. [pdf]. In : National dairy heifer evaluation project. 1996. [http://www.aphis.usda.gov/vs/ceah/ncahs/nahms/dairy/ndhep/hepdes2.pdf], (consulté le 05 janvier 2010).
116 National Animal Health Monitoring System (NAHMS). Dairy herd management practices focusing on preweaned heifers. [pdf] In : National dairy heifer evaluation project. 1993. [http://www.aphis.usda.gov/vs/ceah/ncahs/nahms/dairy/ndhep/dr91des1.pdf], (consulté le 05 janvier 2010).
117 National Animal Health Monitoring System (NAHMS). Part 1: reference of dairy health and management in the United States. [pdf]. In : National dairy heifer evaluation project. 2002. [http://ageconsearch.umn.edu/bitstream/32775/1/pa02da01.pdf], (consulté le 05 janvier 2010).
118 Navetat H, Contrepois M, Schelcher, et al.Les gastro‐entérites paralysantes : une revue. Bulletin des GTV, 1996, 1, 7‐14.
119 Newby TJ, Stokes CR, Bourne FJ. Immunological activities of milk. Vet. Immunol. Immunopathol., 1982, 3, 67‐94.
120 Note de service DGAL/SDSPA/N2004‐8185 du 16 juillet 2004. [en ligne]. [pdf]. [http://agriculture.gouv.fr/IMG/pdf/dgaln20048185.pdf] (consulté le 05 janvier 2010).
121 Olson DP, Bull RC, Woodward LF, et al. Effects of maternal nutritional restriction and cold stress on young calves : absorption of colostral immunoglobulins. Am. J. Vet. Res., 1981, 42, 876‐880.
129

122 Osterstock JB, Callan RJ, Van Metre DC. Evaluation of dry cow vaccination with a killed viral vaccine on post‐colostral antibody titers in calves. In: Proceedings of the American Association of Bovine Practitioners. Columbus (OH). 2003, 163‐164.
123 Pakkanen R, Aalto J. Growth factors and antimicrobial factors of bovine colostrum. Int. Dairy J., 1997, 7, 285‐297.
124 Parreño V, Béjar C, Vagnozzi A, Barrandeguy M, Costantini V, Craig MI, Yuan L, Hodgins D, Saif L, Fernández F. Modulation by colostrum‐acquired maternal antibodies of systemic and mucosal antibody responses to rotavirus in calves experimentally challenged with bovine rotavirus. Vet. Immunol. Immunopathol., 2004, 100, 7‐24.
125 Poppe C, Duncan CL. Comparison of detection of Salmonella by the Tecra® Unique ™. Salmonella test and the modified Rappaport Vassiliadis medium. Food Microbiol., 1996, 13, 75‐81.
126 Poulsen KP, Hartmann FA, McGuirk SM. Bacteria in colostrum : impact on calf health [abstract 52] In : Proc. 20th American College of Internal Veterinary Medicine. St. Luois (MO): Mira Digital Publishing, 2002, 773.
127 Pravieux JJ, Poulet H, Charreyre C, Juillard V. Protection of Newborn Animals through Maternal Immunization. J.Comp. Path., 2007, 137, 32‐34.
128 Pritchett LC, Gay CC, Besser TE, et al. Management and production factors influencing Immunoglobulin G1 concentration in colostrum from Holstein cows. J. Dairy Sci., 1991, 74, 2336‐2341.
129 Pritchett LC, Gay CC, Hancock DD, et al. Evaluation of the hydrometer for testing immunoglobulin G1 concentrations in Holstein colostrum. J. Dairy Sci., 1994, 77, 1761‐1767.
130 Przybylska J, Albera E, Kankofer M. Antioxidants in bovine colostrum. Reprod. Domest. Anim., 2007, 42, 402‐409.
131 Quigley JD, Fike DL, Egerton MN, et al. Effects of a colostrum replacement product derived from serum on immunoglobulin G absorption by calves. J. Dairy Sci., 1998, 81, 1936‐1939.
132 Quigley JD, Strohbehn RE, Kost CJ, et al. Formulation of colostrum supplements, colostrums replacers and acquisition of passive immunity in neonatal calves. J. Dairy Sci., 2001, 84, 2059‐2065.
133 Quinn PJ, Markey BK, Carter ME, et al. Enterobacteriaceae. In: Veterinary microbiology and microbiology disease. Carlton (Victoria, Australia): Blackwell Science Ltd. 2002, 106‐123.
134 Rastani RR, Grummer RR, Bertics SJ, et al. Reducing dry period length to simplify feeding transition cows : Milk production, energy balance and metabolic profiles. J. Dairy Sci., 2005, 88, 1004‐1014.
135 Ravary B, Sattler N. Néonatalogie du veau. 1ère édition. Les éditions du point vétérinaire, 2006, 265p.
136 Reber AJ, Hippen AR, Hurley DJ. Effects of the ingestion of whole colostrum or cell‐free colostrum on the capacity of leukocytes in newborn calves to stimulate or respond in oneway mixed leukocyte cultures. Am. J. Vet. Res., 2005, 66, 1854‐1860.
137 Reber AJ, Lockwood A, Hippen AR, et al. Colostrum induced phenotypic and trafficking changes in maternal mononuclear cells in a peripheral blood leukocyte model for study of leukocyte transfer to the neonatal calf. Vet. Immunol. Immunopathol., 2006, 109, 139‐150.
138 Reffett JK, Spears JW, Brown TT. Effect of dietary selenium on the primary and secondary response in calves challenged with infectious bovine rhinotracheitis virus. J. Nut., 1988, 118, 229‐235.
139 Reis BP, Zhang SP, Tsolis RM, et al. The attenuated sopB mutant of Salmonella enterica serovar Typhimurium has the same tissue distribution and host chemokine response as the wild type in bovine Peyer’s patches. Vet. Microbiol., 2003, 97, 269‐277.
140 Riedel‐Caspari G. The influence of colostral leukocytes on the course of an experimental Escherichia coli infection and serum antibodies in neonatal calves. Vet. Immunol. Immunopathol., 1993, 35, 275‐288.
130

141 Riedel‐Caspari G, Schmidt FW. The influence of colostral leukocytes on the immune system of the neonatal calf. I. Effects on lymphocyte responses (p. 102‐107). II. Effects on passive and active immunization (p. 190‐194). III. Effects on phagocytosis (p. 330‐334). IV. Effects on bactericidity, complement and interferon. Synopsis (p. 395‐398). Dtsch Tierarztl Wochenschr., 1991, 98, 102‐398.
142 Rischen CG. Passive immunity in the newborn calf. Iowa State Univ. Vet., 1981, 12(2), 60‐65. 143 Robertsson JA, Lindberg AA, Hoiseth S, et al. Salmonella typhimurium infection in calves :
protection and survival of virulent challenge bacteria after immunization with live or inactivated vaccines. Infect. Immun., 1983, 41, 742‐750.
144 Robison AD, Stott GH, DeNise SK. Effects of passive immunity on growth and survival in the dairy heifer. J. Dairy Sci., 1988, 71, 1283‐1287.
145 Roy JHB. The Calf, 5th ed., vol. 1, Management of Health. British Library Cataloguing in Publication Data, 1990, 1‐117.
146 Saif LJ, Redmen DR, Smith KL, et al. Passive immunity to bovine rotavirus in newborn calves fed colostrum supplements from immunized or nonimmunized cows. Infect. Immun., 1983, 41, 1118‐1131.
147 Saif LJ, Smith KL, Landmeier BJ, et al. Immune response of pregnant cows to bovine rotavirus immunization. Am. J. Vet. Res., 1984, 45, 49‐58.
148 Sangild PT. Uptake of colostral immunoglobulins by the compromised newborn farm animal. Acta. Vet. Scand. Suppl., 2003, 98, 105‐122.
149 Sangild PT, Schmidt M, Petersen YM. The premature newborn calf : how to use steroids to improve survival ? Acta. Vet. Scand. Suppl., 2003, 98, 280.
150 Schnorr KL, Pearson LF. Intestinal absorption of maternal leucocytes by newborn lambs. J. Reprod. Immunol., 1984, 6, 329‐337.
151 Schumman FJ, Townsend HGG, Naylor JM. Risk factors for mortality from diarrhoea in beef calves in Alberta. Can. J. Vet. Res., 1990, 54, 336‐372.
152 Selman IE, McEwan AD, Fisher EW. Studies on dairy calves allowed to suckle their dams at fixed times postpartum. Res. Vet. Sci., 1971, 12, 1‐6.
153 Shah NP. Effects of milk‐derived bioactives: an overview. Br. J. Nutr., 2000, 84(1), 3‐10. 154 Sivula NJ, Ames TR, Marsh WE, Werdin RE. Descriptive epidemiology of morbidity and mortality in
Minnesota dairy heifer calves. Prev. Vet. Med., 1996, 27, 155‐171.
155 Smith BP. Diseases of the alimentary tract. Salmonellosis in ruminants. In: Large animal internal medicine. 3rd edition. St. Louis (MO): Mosby, Inc., 2002, 775‐779.
156 Smith GW, Foster DM. Short Communication : absorption of protein and immunoglobulin G in calves fed a colostrum replacer. J. Dairy Sci., 2007, 90, 2905‐2908.
157 Sojka WJ, Field HI. Salmonellosis in England and Wales 1958–1967. Vet. Bull., 1970, 40, 515‐531. 158 Staley TE, Corles CD, Bush LJ, et al. The ultrastructure of neonatal calf intestine and absorption of
heterologous proteins. Anat. Rec., 1972, 172, 559‐579.
159 Steele ML, McNab WB, Poppe C, et al. Survey of Ontario bulk tank raw milk for foodborne pathogens. J. Food Prot., 1997, 60(11), 1341‐1346.
160 Stewart S, Godden S, Bey R, et al. Preventing bacterial contamination and proliferation during the harvest, storage and feeding of fresh bovine colostrum. J. Dairy Sci., 2005, 88, 2571‐2578.
161 Streeter RN, Hoffsis GF, Bech‐Nielsen S, et al. Isolation of Mycobacterium paratuberculosis from colostrum and milk of subclinically infected cows. Am. J. Vet. Res., 1995, 56(10), 1322‐1324.
162 Stott GH, Marx DB, Menefee BE, et al. Colostral immunoglobulin transfer in calves I. Period of absorption. J. Dairy Sci., 1979, 62, 1632‐1638.
163 Swan H, Godden S, Bey R, et al. Passive transfer of immunoglobulin g and preweaning health in Holstein calves fed a commercial colostrum replacer. J. Dairy Sci., 2007, 90, 3857‐3866.
131

164 Thatcher EF, Gershwin LJ. Colostral transfer of bovine immunoglobulin E and dynamics of serum IgE in calves. Vet. Immunol. Immunopathol., 1989, 20, 325‐334.
165 Thiry E. Maladies virales digestives des bovins. In : Pathologies des Maladies Virales (chapitre 4). [en‐ligne], Novembre 2009, Université de Liège, [http://www.dmipfmv.ulg.ac.be/virovet/m/bovins/bovin_dig.pdf]
166 Tizard IR. Veterinary immunology : an introduction. 6th edition. Philadelphia: W.B. Saunders Company, 2000.
167 Todd CG, McKnight DR, Millman ST, et al. An evaluation of meloxicam (Metacam®) as an adjunctive therapy for calves with neonatal calf diarrhea complex. J. Anim. Sci., 2007, 85, 369 [abstract].
168 Tsolis RM, Adams LG, Ficht TA, et al. Contribution of Salmonella typhimurium virulence factors to diarrheal disease in calves. Infect. Immun., 1999, 67, 4879‐4885.
169 Tyler H, Ramsey H. Hypoxia in neonatal calves : Effect on intestinal transport of immunoglobulins. J. Dairy Sci., 1991, 74, 1953‐1956.
170 Tyler JW, Steevens BJ, Hostetler DE, et al. Colostral immunoglobulin concentrations in Holstein and Guernsey cows. Am. J. Vet. Res., 1999, 60, 1136‐1139.
171 Uruakpa FO, Ismond MAH, Akobundu ENT. Colostrum and its benefits : a review. Nut. Res., 2002, 22, 755‐767.
172 Valente C, Fruganti G, Tesei B, CIORBA A, Cardaras P. Vaccination of pregnant cows with k99 antigen of enterotoxigenic Escherichia coli and protection by colostrum in newborn calves. Comp. lmmun. Microbiol. Infect. Dis., 1996, 11(3‐4).
173 van Zaane D, Ijerman J, de Leeuw PW. Intestinal antibody response after vaccination and infection with rotavirus of calves fed colostrum with or without rotavirus antibody. Vet. Immunol. Immunopathol., 1986, 11, 45‐63.
174 Veling J, Wilpshaar H, Frankena K, et al. Risk factors for clinical Salmonella enterica subsp. enterica serovar Typhimurium infection on Dutch dairy farms. Prev. Vet. Med., 2002, 54, 157‐168.
175 Waltman WD. Methods for the cultural isolation of Salmonella. In: Wray C, Wray A, editors. Salmonella in domestic animals. Wallingford (UK): CABI Publishing. 2000, 355‐372.
176 Waltner‐Toews D, Martin SW, Meek AH, et al. A field trial to evaluate the efficacy of a combined rotavirus‐coronavirus Escherichia coli vaccine in dairy cattle. Can. J. Comp. Med., 1985, 49, 1‐9.
177 Waltner‐Toews D, Martin SW, Meek AH. Dairy calf management, morbidity and mortality in Ontario Holstein herds. Prev. Vet. Med., 1986, 4, 103‐171.
178 Walz PH, Mullaney TP, Render JA, et al. Otitis media in preweaned Holstein dairy calves in Michigan due to Mycoplasma bovis. J. Vet. Diagn. Invest., 1997, 9, 250‐254.
179 Weaver DM, Tyler JW, VanMetre DC, et al. Passive transfer of colostral immunoglobulins in calves. J. Vet. Intern. Med., 2000, 14, 569‐577.
180 Wells SJ, Dargatz DA, Ott SL. Factors associated with mortality to 21 days of life in dairy heifers in the United States. Prev. Vet. Med., 1996, 29, 9‐19.
181 Wilson J. Immune system breakthrough : colostrum. J. Longevity Res., 1997, 3, 7‐10. 182 Wittum TE, Perino LJ. Passive immune status at postpartum hour 24 and longterm health and
performance of calves. Am. J. Vet. Res., 1995, 56, 1149‐1154.
183 Wray C, Davies R. Salmonella infections in cattle. In: Wray C, Wray W, editors. Salmonella in domestic animals. New York : CABI Publishing. 2000, 169‐190.
184 Wray C, Todd N, Hinton MH. The epidemiology of Salmonella typhimurium infection in calves : excretion of S. typhimurium in faeces of calves in different management systems. Vet. Rec., 1987, 121, 293‐296.
185 Xu LB, Chen L, Gao W, Du KH. Bovine immune colostrums against 17 strains of diarrhea bacteria and in vitro and in vivo effects of its specific IgG. Vaccine,2006, 24, 2131‐2140.
132

133
186 Zahraei ST, Mahzounieh MR, Vatankhah J. Identification and diagnosis of Salmonella serotypes in cow’s faecal samples by polymerase chain reaction (PCR). University of Tehran. J. Fac. Vet. Med., 2006, 61, 243‐247.
187 Zahraei ST, Tadjbakhsh TH, Atashparvar N, et al. Detection and identification of Salmonella Typhimurium in bovine diarrhoeic fecal samples by immunomagnetic separation and multiplex PCR assay. Zoonoses Public Health 2007, 54, 231‐236.
188 Zaremba W, Guterbock WM, Holmberg CA. Efficacy of a dried colostrum powder in the prevention of disease in neonatal Holstein calves. J. Dairy Sci., 1993, 76, 831‐836.
189 Zhang SP, Santos RL, Tsolis RM, et al. The Salmonella enterica serotype typhimurium effector proteins SipA, SopA, SopB, SopD, and SopE2 act in concert to induce diarrhea in calves. Infect. Immun., 2002, 70, 3843‐3855.
190 Zobell DR, Schafer AL, LePage PL. Gestational vitamin E supplementation in beef cows : effects on calf immunological competence, growth and morbidity, in Proceedings. Western Sect ASAS. 1995, 46, 464‐466.

ÉTIOLOGIE DES DIARRHÉES NÉONATALES ET TRANSFERT
COLOSTRAL CHEZ LE VEAU : ENQUÊTE DANS LA CREUSE
NOM et Prénom : MAES Paul
Résumé :
Les diarrhées néonatales sont la principale cause de maladie chez les veaux. Elles peuvent toucher de 10% à 80% des veaux suivant les élevages. Elles ont des répercussions économiques importantes de par le coût des soins à apporter aux veaux et par les mortalités. Les agents pathogènes les plus communs sont les rotavirus, les coronavirus, E. coli F5 et Cryptosporidium parvum. Plusieurs facteurs de risque interviennent, notamment l’hygiène de l’environnement et la conduite d’élevage, et plus particulièrement le transfert colostral qui est un point critique en néonatalité. Cette étude, menée dans la Creuse au cours de l’hiver 2008‐2009, avait pour objectifs d’évaluer l’importance des principaux pathogènes dans les diarrhées et de mettre en relation le statut immunitaire passif du veau nouveau‐né (déterminé par le dosage des immunoglobulines G et des protéines totales sériques) avec les diarrhées néonatales. D’autres facteurs de risque ont également été recherchés. De faibles taux d’immunoglobulines et de protéines totales ont été mis en relation avec la maladie, sans toutefois obtenir de résultats significatifs. L’échantillon n’était probablement pas de taille suffisante, et l’analyse a pu manquer de puissance statistique. Il est montré également que de mauvaises conditions d’hygiène au vêlage, des mauvaises conditions environnementales (notamment une sur ou sous‐ventilation), et une mauvaise gestion de l’administration de colostrum sont des facteurs de risque des diarrhées néonatales.
Mots clés : DIARRHÉE NÉONATALE, COLOSTRUM, IMMUNOGLOBULINE, IMMUNITÉ PASSIVE,
ROTAVIRUS, CORONAVIRUS, E. COLI F5, CRYPTOSPORIDIUM PARVUM, ANIMAUX JEUNES, BOVIN,
VEAU, CREUSE
Jury :
Président : Pr. Directeur : Dr. Yves MILLEMANN Assesseur : Dr. Loïc DESQUILBET
Adresse de l’auteur :
Mr Paul MAES
6, rue Pierre Curie
94700 MAISONS‐ALFORT

NEONATAL CALF DIARRHOEA ETIOLOGY AND COLOSTRAL
PASSIVE TRANSFER: A SURVEY IN THE DEPARTMENT OF
CREUSE
SURNAME: MAES
Given name: Paul
Summary:
Neonatal calf diarrhoea is the main cause of calves illness. It can reach 10 to 80% of calves, depending on farms. It has an important economic impact, due to the cost of medical treatments of calves and to neonatal death losses. Rotavirus, coronavirus, E. coli F5 and Cryptosporidium parvum are recognized as the major pathogens associated with diarrhoea in calves. Several risk factors are involved, including environmental and herd management, and most likely passive transfer which is a critical point in neonatality. The present study was carried out in the department of Creuse during winter 2008‐2009. Its aims were to evaluate the importance of the main pathogens in diarrhoea and to relate the passive immune status of neonate calves (determined by serum immunoglobulin G and total solids levels) with neonatal calves diarrhoea. Other risk factors were also investigated. Low immunoglobulin and total proteins levels were associated to presence of diarrhoea. The association was however not significant, and we may have lacked some statistical power in order to provide significant results, because of the small sample size. In this study, it has also been shown that poor calving hygiene, poor environmental management conditions (particularly under or over‐ventilation), and poor colostrums management are risk factors of neonatal calf diarrhoea.
Keywords : NEONATAL DIARRHOEA, COLOSTRUM, IMMUNOGLOBULIN, PASSIVE IMMUNITY,
ROTAVIRUS, CORONAVIRUS, E. COLI F5, CRYPTOSPORIDIUM PARVUM, YOUNG ANIMALS, BOVINE,
CALF, CREUSE.
Jury:
President: Pr. Director: Dr. Yves MILLEMANN Assessor: Dr. Loïc DESQUILBET
Author’s address:
Mr Paul MAES
6, rue Pierre Curie
F‐94700 MAISONS‐ALFORT





























