Embed Size (px)
Citation preview

762 Archives Internationales de Physiologie et de Biochirnie, 1965, 73 ( 5 ) .
ReCu le 20 septembre 1965
TRANSPORT ACTIF ET PERMgABILITB PASSIVE AU NIVEAU DE LA CORNBE
DU CRAPAUD BUFO M A R I N U S L.
PAR
R. R. TERCAFS (l) (Insfitulo de Bioffsica, Universidade do B r a d , R io de Janeiro)
( 1 figure)
Introduction
GRIFFITHS et al. (1964) ont recemment montre que la cornbe isolke de grenouille ktait le sibge d’une diffkrence de potentiel ( d p ) de l’ordre de 5 millivolts, la face interne etant positive. 11s n’ont cependant pas ddtermind quelle Btait I’origine de cette d p . Nous avons effectui: des expkriences h ce sujet sur une autre espbce de Batracien, de plus grande taille, Bufo marinus ; l’intkrht de cette ktude est double : d’une part, la comparaison des caractbres de perm6abilitk de la cornke e t de la peau peut apporter d’utiles renseignements quant B 1’6volution des diffk- rents tissus d’origine embryologiyue analogue mais de fonction diffkrente, e t d’autre part, la determination de ces caractbres de permkabilith est ndcessaire B la comprdhension de la biochimie de l’ceil.
MatCriel et mbthodes
Les Bufo marinus L. utilises dans les expdriences proviennent de la rkgion de Rio de Janeiro.
Les operations suivantes sont effectukes : spinalisation, enlbvement de la paupibre superieure, extraction totale de l’aeil hors de l’orbite, incision autour de la cornke, enlbvement du
(’) Charge de recherche5 du Fonds National de la Recherche Scienlifique de
Adresse permanente : Institut Leon Fredericq (Biochimie) Universitb de I,i+gc, 17, Beigique.
place Delcour, LiBge, Belgique.
Arc
hive
s of
Phy
siol
ogy
and
Bio
chem
istr
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
New
cast
le U
pon
Tyn
e on
12/
20/1
4Fo
r pe
rson
al u
se o
nly.

PERMBAUXLITB DE LA CORNEE DU CRAPAUD 763
cristallin et de la choroi’de. La cornke ainsi isolke est placke entre dcux cadres skparant deux chambres de plexiglass (dis- positif dkcrit par BAILLIEN et SCHOFFENIELS (1960). La fen6tre skparant les deux chambres a une surface utile de 25 mm2. La durkc totale des opkrations est d’environ trois minutes.
Le Ringer (RgCI) baignant la cornke a la composition suivante : 112 mM NaC1, 3.3 mM KC1, 0.45 mM CaC12 et 5mEq PO, pro- venant d’une solution tampon B pH 7.8 de K,HPO, etKH, PO,. Nous utilisons kgalement du Ringer sulfate (RgSO,) qui contient une quantite kquivalente d’ions Na, I< e t Ca sous formc de sul- fates. La solution RgSO, Na 1/10 contient 1 1 mM de Na et la quantitk de saccharosc nkcessaire pour obtenir la miime pression osmotique que le RgSO,. La notation K x 10 indique l’addition de la quantitk de K,SO, nkcessaire pour obtenir une concentration finale en K de 33 mM/l.
Les mesures des flux de Na sont faites en utilisant la technique dcs doubles traceurs (USSING ef aZ., 1951). La dp existant au niveau de la cornbe est abolie pendant ces mesures B l’aide d’une force klcctromotrice extcrne. Lc 22Na provient du Radio- chemical centre, Amersham, England, le 24Na provient de Z’Instilulo da E n o y i a iltdmicn, S i o Paulo, Brasil.
Origine de la diffe‘rence de potentiel au niveau de la corne‘e.
Quand la cornke est baignke de part e t d’autre par une solu- tion de Ringer, il cxiste une diffkrence de potentiel ( d p ) entre les dcux solutions (tableau I) . la solution baignant la face interne est toujours positive. Pour dkterminer l’origine de cette d p , nous allons suivre la mbthode que nous avons utiliske prk- ckdcmnient (TERCAFS, 1065), ct qui consiste B modifier les concentrations ioniques des solutions en contact avec la cornee. Les variations de la d p produitcs par ccs modifications sont mesu- rhes une minute aprbs le changement de solution et interpret6es en tcrmcs de perm8abilitk passive.
Lc tableau 1 donne les rksultats obtenus; nous pouvons en tircr lcs conclusions suivantes :
1 ) Comparons les expkriences A e t B : si on remplace les solu- tions baignant la membrane par des solutions dc composition cationique identiquc mais ne posskdant pas d’ions C1 (RgSO,), la d p augmente. Ceci s’intcrprbte si I’on considerc que, comme
Arc
hive
s of
Phy
siol
ogy
and
Bio
chem
istr
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
New
cast
le U
pon
Tyn
e on
12/
20/1
4Fo
r pe
rson
al u
se o
nly.

764 R. R. TERCAFS
2.65 f 0.96
5.91 f 1.22
TAULEAL 1. -Difference de potentiel en mV f erreur sur la moyenne mesurde au nweau de la cornde isolee de Bufo marinus. RgCl = Ringer
RgSO, = solution physiologique oh les ions C1 sont remplaces par des ions SO, ; Na 1/10, K x 10, = solutions physiologiques oil la concentration en Ka ou en K est, soit 10 fois plus feible (l/l(J), soit 10 fois plus forte ( x 10) que dans la solution normale; SE = erreur sur la moyenne; P = probabilite a l’bquivalence des moyennes. Mojennes de 10 exphricnces. La difference de potentiel est donnee en millivolts. Le signe se refere ti la solution bsignant la face interne de la cornbe.
DF < 0.001
BG > 0.1
_-____-___ -.
Conditions exp6riment:rles
Sol. externe Sol. interne
R
C
D a-
E
F
_ _
--
WSQ4
RgSO,Nnl/lO
RgSO,Nnl/lO
RgS0,Kn 1/10
RgS0,Na l/lOK x 10
_ _ _ _ ~ . _ _ _
RgSQ4
RgSO4 - RgS0,Na 1/10
RgS0,Nn I/lOK x 10
RgS0,Na 1/10
Iliffkrence de potentiel
l p Moyenne f- S.E.
5.46 i 1.00
7.36 f 1.11 __ -10.98 & 1.45
-4.77 f 1.0%
-10.00 5 1.23
A B 0.1-0.05
BC < 0.001
CD < 0.001
DE < 0.001
pour la peau de Batracien ( KOEFOED-JOHNSEN ef al., 1952), lcs ions SO, sont moins diffusibles que les ions C1. Le court-cir- cuitage de la d p spontanke dQ aux ions C1 est donc diminue en milieu SO4, ce qui a pour resultat une augmentation de la d p .
2) Comparons les cxpkriences B et C : la diminution de la con- centration en Na de la solution externe entraine une inversion de la dp . Ceci indique que la diffusion du Na du comparti- ment extkrieur vers le milieu cellulaire a une influence dkter- minante quand au maintien dc la d p spontanke en RgCl ou en RgSO,. Cette diminution montre dgalement que les poten- tiels de difhsion existant encore B ce moment sont en oppo- sition avec la d p totale primitive.
3) Par contre, la comparaison des experiences B et G montre que la diminution de la concentration en Na dans la solu- tion interne n’a aucune influence sur la d p . La permkabi- lit6 au Na de la face interne est donc trhs faible.
4) Cependant, en apparente contradiction avec cette impermea- bilite au Na de la face interne, il se produit une variation
Arc
hive
s of
Phy
siol
ogy
and
Bio
chem
istr
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
New
cast
le U
pon
Tyn
e on
12/
20/1
4Fo
r pe
rson
al u
se o
nly.

P E R M J ~ A B I L I T ~ DE LA CORNBE I)U CRAPAUD 765
dc la d p quand la concmtration cn Na tie la solution intc*me diminue e t quc la concentration en Na d r la solution externe est Bgalement basse (cxpkricnces C et D). Ccci pcut s’expliqucr par une modification de la perineabilitk de cctte face en fonc- tion de la concentration ionique dii milieu cclfulaire. En effet, la diminution de la concentration (in Na du tiiilicu cxt6rieur provoque vraisemblablrinent drs modifications dans la composition clu milieu c.~llulairc qui pcut rendre corripte de c d t e variation dc pernibahilite dc la facr interne ; ce phkno- mbne de variation des c*aractiws dc perinkabiliti: en fonction dcs concentrations a dej i 6t4 rnis en Cvidenw sur d’autres membranes (CERFIJIIIO ~1 uZ., 1964 ; TERCAE’S, 1965).
5) L’exp6rience E comparkc h I’expCrience D indique que la face interne est perm6ablc aux ions K. En cflet, l’aug- mentation de la d p s’expliquc si on postule 1’cxist(in(*c tl’un gradient de diffusion de K tlu milieu cellulairc vers le corn- partinient internc. Lc potchntirl dc diffusion qui en rPsulte etant cn opposition avcr la difl’C.ronw dc potentiel totalc, sa suppression entralne unci augmmlation de la d p .
6) Conime l’indiquc la cornparaison dcs expkrienccs 1) c t F, la face interne cst egalcment lc siege tl’un poetntipl t l v t l i f - fusion de I<. Sa suppression par augmentation de la conccn- tration d a m la solution &erne augment(* la d p c t rend i nouveau la solution interne positive. En cHct, 6 cc moment ne subsiste que le potentiel de cliflusion en T i du milieu cel- lulaire vers la solution interne.
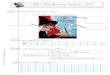
Ces dinerents potentiels de diffusion peu\-c.nt fitre rcprbsentks par un whkma genkral (fig. 1 ) .
Les membranes cellulaircs wrisliluant la face txtcme sont perm6ables aux ions Na, I< ct (2. Les membranes constituant la face interne sont perrnt5ablcs aux ions K et C1. Cctte dispo- sition diffkrente des caractkrcs de pemikabilite sur lcs dcux faces de la cornbe crCe un potcntiel de diffusion du Na A la face cxterne (cxtkrieur nCgatif, milieu cellulaire positif) oppos6 a un potentiel dc diffusion d u K (extbrieur positif, milieu cel- lulaire nCgatif), ainsi qu’un potentiel de diffusion au l< A la face interne (milieu cellulaire negatif, intkrirur positif). Ces trois potentiels ont pour rdsultat la creation d’unc d p (solution externe nkgative, solution interne positive) entre les deux solu-
Arc
hive
s of
Phy
siol
ogy
and
Bio
chem
istr
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
New
cast
le U
pon
Tyn
e on
12/
20/1
4Fo
r pe
rson
al u
se o
nly.

766
0
R. H. TERCAFS
i
+
FIG. 1. - Schema des caracteres de permeabilite :iu niveau de la corji~je de Bufo marinus baignee sur les deux faces par du Ringer. 0 = solution externe ; i = solu- tion interne. Les droites representent des potentiels de diffusion produits par la diffusion des ions d’un milieu concentration Blevbe vers un milieu t~ concentration basse. Les membranes constituant la face externe de la cornbe sont permbables B la fois au Na et au K ; la face interne est permeable aux ions K uniquement. Ces diffbrents potentiels de diffusion creent une diffkrence de potentiel, la solution interne &ant positive. Les gradients nkcessaires sont maintenus par deux transports actifs indkpendants de Na et de K (representes par des cercles). Les ions CI diffnsent passivement a travers la cornee du milieu exterieur vers la solution interne.
tions baignant les deux faces de la cornbe ; cette d p est partiel- lement court-circuitke par la diffusion passive des ions C1 a travers la membrane.
Les potentiels de diffusion mis ainsi en bvidence ne peuvent exister que si la concentration en Na et en K du milieu cellulairc reste diffkrente de la concentration du milieu extbrieur ct inti.- rieur (concentration en Na, basse ; concentration en K, Blevke). On doit postuler la prksence de deux transports actifs de Na et de K ; la position de ces transports actifs ne pcut &re dktcr- minbe par les expkriences prkcbdcntes; en cffct, toutes les deductions sont valables, quc les transports actifs soient loca- lisbs a la face externe ou a la face interne.
Origine des gradients de Concentration.
Pour verifier les hypothbses prbcbdentcs, nous avons effectub des expbriences 5 l’aide de la technique des doubles traceurs,
Arc
hive
s of
Phy
siol
ogy
and
Bio
chem
istr
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
New
cast
le U
pon
Tyn
e on
12/
20/1
4Fo
r pe
rson
al u
se o
nly.

PERMEABILITE DE LA CORNBE DU CRAPAUD 767
Oulffux pEq/h/cm2
1.028
k0.187
en utilisant du 22Na e t du 2QNa. La d p existant au niveau de la cornke isolbe est abolie a l’aide d’une force klectromotrice (USSING et al., 1951). Les rksultats obtenus sont repris dans le tableau 11.
Influx pEq/h/cm2
7.129
12.048
TABLEAU 11. - Flux de N u d frauers la cornee de Rufo marinus, mesure a l’aide de la technique des doubles fraceurs. Outflux = flux de la solution interne vers la solu- tion externe ; influx = flux de la solution externe vers la solution interne. CCC = cou- rant de court-circuitage necessaire d l’abolition de la dp. mcb = millicoulombs. Valeurs
moyennes de 16 experiences (periodes d’une heure), f erreur sur la moyenne.
6.101
1.897
588.74 17.12 61.63
183.03 & 1.95 & 7.01
Les rksultats montrent clairement qu’il y a un flux net de Na de la solution externe vers la solution interne. La diffkrence entre l’outflux et l’influx est hautement significative (P <0.01). I1 existe donc bien un transport actif de Na que nous pouvons localiser a la face interne de la cornke. Si on exprime le flux net mesurk 4 l’aide d’isotopes dans les m&mcs unites que celles utilisees pour la mesure du courant de court-circuitage, on constate une grande difference : le flux de Na est plus eleve que la quantitk de courant necessaire a l’abolition de la dp . I1 n’en serait pas ainsi si le Na etait le seul ion transporte active- ment. Mais si l’on retourne au schema des caractkres de permkabi- lit& (fig. I) , on voit qu’il existe un flux net de I( opposi: B celui du Na. La diffkrence entre le courant de court-circuitage e t le flux de Na est donc produit par ce flux dc K dont on peut cal- culer la valeur a partir des donnbes mesurbes, c t qui est donc de 588.74-61.63 = 527.11 mcb/h/cm2. S’il en est bien ainsi, nous devons constater une augmentation du courant de court- circuitage si nous rkduisons le gradient de diffusion du K A la face externe en augnientant la concentration du K dans le milieu extbrieur. Le tableau I11 donne les valeurs obtenues. On constate en effet une augmentation du courant de court- circuitage quand la concentration en K du milieu exthrieur est multiplihe par 10. Thkoriquement, si le gradient de K est totalement supprime a la face externe, nous devons constater
Arc
hive
s of
Phy
siol
ogy
and
Bio
chem
istr
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
New
cast
le U
pon
Tyn
e on
12/
20/1
4Fo
r pe
rson
al u
se o
nly.

768 R. R. TERCAFS
Solution exlerne 1 Solution interne
une equivalence entre le flux net de Na et le courant de court- circuitage, inais il ne faut pas ouhlier que, d’abord, nous ne connaissaons pas exactement la concentration en I< du milieu cellulaire, e t ensuite la variation de concentration en K du milieu extkrieur modifie ccrtainement la pcrmeabilitk au K et au Na de la membranc.
l p bloyenne f SE
TAnLEarr 111. - Courant de court-circuitage (C.C.C.) exprime en pA/cm* au niveau de lu cornee de Rrifo mnrinus
Moyennes de 3 experiences f erreur sur la moyenne. RgCl = Ringer ; K x 10 = solution oh la concentration en K est 10 fois plus hlevt!e que dans le Ringer normal. P = probabilitt! a l’equivalence des moyennes.
Conditions expt!rimentoIes C.C.C. I I
RgCI 39.3 * 0.5 1 <0.001 I 1 RgCl/K x 10
La demonstration de l’existence d’un transport actif de Na nous permet de supposer I’existence, sans doute aussi au nivcau de la face interne, d’un transport actif de K, maintenant une concentration &levee dans le milieu cellulaire.
On peut Cgalement mettre en kvidence ccs transports actifs d’une fagon indirecte, en etudiant l’action du 2-4-Dinitrophbnol. Cette substance inhihe notamment le transport actif de Na au niveau de la peau de grenouille (SCHOFFENIELS, 1955), de l’intestin e t du cijlon de tortue (BAILLIEN ef al., 1961). Dans le cas de la cornee de crapaud, nous devons nous attendre A une diminution de la d p , par suite de l’abolition des potentiels de diffusion clus A la presence des transports actifs, aprks l’addition de 2-4-DinitrophCtnol. C’est bien, en effet, ce que l’on observe (tableau IV). A la concentration de 0.1 mM/1, la d p est annihilee une demi-heure aprbs l’addition de 2-4-DNP dans les deux solu- tions.
La disposition des caractkres de permbabilitk rendant compte de la d p observee est maintenant complkte (fig. 1). L’Bquation genbrale exprimant la valeur de la d p observe sera de la forme (GOLDMAN, 1943) :
Arc
hive
s of
Phy
siol
ogy
and
Bio
chem
istr
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
New
cast
le U
pon
Tyn
e on
12/
20/1
4Fo
r pe
rson
al u
se o
nly.

PERMBARILITB DE LA CORNBE D U CRAPAIJD
~~~~ ~ -. .. _____~
r1gC:l
Difference de potentiel
769
~~ ~~ ~ .. . . .~ ~- ~ ~ ~- .
b!-~t-rjN I’
(:oiicentr:ilion 1 Diff6rwce de l’oteiitiel j I
Oh Eexta - Eceii. = d p cntrc la solution cxtbrieurc et le milieu
Ecell. - Eint, = dp entre lc niilieu ccllrilairc (4 la solution cellulairc.
intcrn e.
6.21 i 1.46 0.1 mM/1 \6!
- 0.10 c 0.23 <n.ooi (6)
R = constante des gaz. T = tempkrature absolue. I , = c.liarge de l’ion. F = nombre do Faraday. In = logarithmc nbpbrien.
pext . P=t.I(, pext. c1 = coefficient de pcrrn6abilite pour Na’ les ions Na, I< e t C1 des membranes cellulaircs formant la face externe.
T A n m A u IV. -- InJluenee drc 2--~-J)inifrr~phCno/ ( 2 4 - D N t ’ ) S U P la ililrerence de potentiel uu nioeau de la corne‘e de Bufo marinus. Dilference de potentiel en milliruolts,
erreur sur lu moyenne ; nomhre d’experiences enire parenth6srs. Le siqne s e refere u la snlntion interne. Mevures effectiides li.2 hrrrre nprc‘s l’nrldilinn rle 2-4-I)AVP dans
IPS solutions interne el estcrne
Arc
hive
s of
Phy
siol
ogy
and
Bio
chem
istr
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
New
cast
le U
pon
Tyn
e on
12/
20/1
4Fo
r pe
rson
al u
se o
nly.

770 R. R. TERCAFS
Pint. et Pint. = coefficient, de permkahilite pour les ions
K et C1 des membranes cellulaires formant la face interne.
(Na), (K), C1) represente les concentrations des ions de la soiu- tion externe (ext.), interne (int.), e t du compartiment cellulaire (cell.),
K c1
Discussion
Nos resultats montrent que la cornbe de Bufo marinus est le sibge d’un transport actif responsable d’un flux net de Na de la solution externe vers la solution interne ; nous constatons kgalement un flux net de K dans l’autre sens. Les membranes constituant la face externe sont permkables au Na, au K e t au C1, e t cclles de la face interne sont permeables au K et au C1; c’est Bgalement A ce niveau que se situent les transports actifs de Na et vraisemblablement de K.
Comparons cette disposition spatiale des caractbres de per- meabilite a celle presente au niveau de la peau du m&me animal. On sait, notamment d’aprbs les travaux d’USSING e t de ses collahorateurs (KOEFOED-JOHNSEN et al., 1958), que la peau de grenouille (Rana temporaria, Rana esculenta) , prksente les caractbres de permeabiliti: suivants : face externe permeable au Na e t au C1, face interne permeable au K et au C1, transports actifs de Na du milieu cellulaire vers la solution interne et de K dans I’autre sens, au nivcau de la face interne. Cettc dispo- sition a pour rksultante, en RgCl, une diffBrence de potentiel, la solution interne etant positive. Une disposition identique s’observe au niveau de la peau dc Bufo marinus, comme nous l’avons verifik; a titrc d’exeniple, en RgCl, la d p cst de 59.5 & 9.3 mV, en RgSO,, de 88.5
L,a cornhe prksente donc, a la face externe, un caractbre de permeabilitk supplementaire ; une permeabilite passive aux ions K. Le transport actif de Na est kgalement plus eleve qu’au niveau de la peau.
I1 est bien connu que la cornee est d’origine epidermique, e t il est remarquable qu’il existe une grande analogie entre les caractkres de permeabilite de ce tissu et ceux de la peau. Malgre des fonctions diffbrentes, ces deux tissus d’origine commune ont conservk un certain nomhre de caractkres de permeabilite
10.4 mV.
Arc
hive
s of
Phy
siol
ogy
and
Bio
chem
istr
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
New
cast
le U
pon
Tyn
e on
12/
20/1
4Fo
r pe
rson
al u
se o
nly.

P E R M B A B I L I T ~ DE LA CORNBE DU CRAPAUD 771
communs. I1 est donc possible d’envisager une evolution dcs caractbrcs de permbabilitk d’une membrane vivante, comme on considbre l’bvolution dc sa structure morphologique ; cette Bvolution est en fait un aspect de l’evolution biochimique des cellules, car ces caractkres de permbabilit8 sont sous la dbpendance dc structures biochimiques.
Envisageons maintenant le r61e des caractbres de permeabilitb observks au niveau de la cornke, dans la physiologie de l’ceil.
La composition chimique des larmes humaines est de 146 3 10 mEq/l de Na, 128 f mEq/l C1 ct 2.42 & mEq/l de K (THAYSEN el al., 1954). On remarque que la concentration en Na e t en C1 est A peu pres identique a celle du sang, mais que la concentration en K est 5.6 fois plus blevke. La composition des larmes dc batraciens n’est pas connue. Supposons qu’elle soit semblablc; par le jeu de la paupiere, cette solution est Btcnduc en permanence sur la cornbe ; ccpcndant, I’kvaporation de l’eau sur cette surface tend augnienter constamment la concentration en ions dc cette solution. Cette augmentation doit &re maintenue a un taux compatible avec la vie des cellules par un mbcanisme rbgulateur de rkabsorption ; nos rbsultats en mon- trent un : les ions Na sont rkabsorbks par un mkcanisme de transport actif, de l’extkrieur de l’ceil vers l’humeur aqueuse, B travers la cornkr. Les ions C1 diffusent passivement dans le m6me sens, gr2ce A l’existence d’une diffkrence de potentiel, (la solution interne btant positive), et d’une permbabilitb pas- sive au C1 des deux faces de la cornbe. La rkabsorption du K ne s’effectuc pas a ce niveau ; au contraire, nous mesurons un flux de I< de l’intkrieur de I’ceil vcrs l’cxterieur. On peut imaginer que cette reabsorption se passe au niveau dc la paupibre. Nous avons l’intention dc verifier prochaincment cctte hypothese.
Conclusion
De cettc etude, nous pouvons tirer les conclusions suivantes : 1. La cornee isolee de Bufo rnarinus est lc sibge d’une diffe-
rence de potentiel ( d p ) de 5.46 f 1.00 millivolts, la solution en contact avec la face interne ktant positive.
2. Les membranes constituant la face externe de la cornbe sont permbables aux ions Na, K et C1. Les membranes constituant la face interne sont permbables aux ions K e t C1. I1 existe
Arc
hive
s of
Phy
siol
ogy
and
Bio
chem
istr
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
New
cast
le U
pon
Tyn
e on
12/
20/1
4Fo
r pe
rson
al u
se o
nly.

772 R. R . TERCAFS
un transport actif de Na produisant un flux net de Na du milieu exterieur vers la solution interne; ce transport est situe au niveau de la face interne. Un transport actif de K, independant, est situk vraisemblablcment B cette face &gale- ment. I1 existe cependant un flux net de K de la solution interne vers la solution externe. La disposition dt. ces carac- tkrtls permet tl’clxpliqucr la d p ohscrvke.
3. La comparaison cntre ces caracti.rcs c t ceux qui existent au nivcau d e la peau montre que, malgrk des fonctions diffe- rentes, ces deux tissus d’origine embryologique idcntique ont conserve un certain nombrc tle caract6rc.s rle permkahi- lit6 communs.
4. Le transport actif dcl Na au nivcau dc la cornee joue cer- taincment un r6le important quant au rnaintien d’une concen- tration compatible avec la vie des cellules dans la solution baignant l’ceil. La reabsorption du I< ne se fait pas a ce nivcau ; elle s’vffectuc put-i.tre au niveau de la paupikrc
Resume
Lcs caractkres de permkahilite suivants ont ete mis cn evi- dence au niveau dr la cornkc isolkr de Bufo marincis : 1 . Au niveau des membranes constituant la face extcrne : per-
meabilite passivc aux ions Na, K e t CI. 2 . Au niveau des membranes constituant la face interne :
perm6abilite passive aux ions K (4. C1 ; transport actif de Na de la solution intcrnr le conipartiment cellulaire. Le transport actif de Na a et6 mesurk B l’aide de la technique des doubles traceurs, en utilisant du 22Na c t du 24Na. Ces differents caractercs de permeahiliti: detcrminent une
difference dr potentiel de 5.46 & 1.00 millivolts. La solution interne est positive quand la corn& est baignee sur s?s deux faces par du Ringer. Les ions C1 difyusent passivemcnt de la solution externc vers la solution intcrne.
Ces caractkres, comparks A ceux de la peau du meme animal, montrent que les deux tissus ont conserve un certain nombre de caractkres de permkabilitk identiques.
Le r61e dc ces caractkres dans la biochimie de l’czil sont discut 6s.
Arc
hive
s of
Phy
siol
ogy
and
Bio
chem
istr
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
New
cast
le U
pon
Tyn
e on
12/
20/1
4Fo
r pe
rson
al u
se o
nly.

PERMEABILITE DE LA CORNEE D U CRAPAUD 773
Remerciements
J e remercie vivement M. le Professeur CHAGAS, qui m’a accord6 les pluu grandes facilitCs pour effectuer mes recherches, et le Minisldre belge de 1’Edricntion Nationale et de la Culture (Direction des Relations culturelles e t de 1’U.N.E.S.C.O.) dont l’aide matkrielle a permis mon skjour au Brksil. J e remercie egalement M. le Docteur E. Sr:HoFFmmi,s , Professeur associC, pour ses conseila lors de la discussiond e ce travail.
BIRLIOGRAPIIIE
UAII.I,IE;Y, M. e t SCIIOFFP~NIELS, E. (1‘360). - Arch. inferriat. I’hysiol. el Biochim., 68,
BAILLIEN, M . et SCIIOFFENIELS, 15. (1961). - Biochim. Biophys. Ada, 53, 737. CEHEJEIDO, M . , HERRERA, F. C., FLANIGAN, W’. . J , e t CURRAN, P. F. (1964). -
J . gen. Physiol., 47, 879. GOLDMAN, D. E. (1943). - J . gen. Physiol., 27, 37. GRIFFITIIS, R. S. et HOARD, J. R. (1964). - Biophgs. Soc., 8th Ann. Meet., Abstracts,
KOEIIFOED-JOEINSEN, V., LEVI, 11. et USSING, 11. 11. (1952). - Acta Phpiol. Scand.,
KOEDFOED-JOIINSEN, V. et USSING, I I . H . (1958). - Acta Physiol . Scand., 4 2 , 298. SCHOFFENIBLS, E. (1955). - Arrh. internat. Physiol. Biochern., 63, 361. TERCAFS, R. R. (1965). -- Acta Physiologica Latino-Americana, 15 206. THAYSEN, J . 11. et THORN, N. A . (1961). - Am. J. Physiol., 178, 160. Ussrxc, €-I. 11. et ZERAHN, K . (1951). - Acta Physiol. Smnd. , 23, 110.
710.
TD 12, Chicago.
25, 150.
Arc
hive
s of
Phy
siol
ogy
and
Bio
chem
istr
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
New
cast
le U
pon
Tyn
e on
12/
20/1
4Fo
r pe
rson
al u
se o
nly.