Embed Size (px)
Citation preview

Utilisation d’anticorps bispécifiques en immunothérapie
BILLON Pierre
SAMAIN Florine
ZUBEROGOITIA Sophie
Enseignants : Laurence NIETO Master EGPR Pascal DEMANGE Année 2009-2010

2
Sommaire RESUME
INTRODUCTION..............................................................................................5
RESULTATS………………………………………..…………………………………….8
1. Production d’anticorps bispécifiques ……. ….………..………………8
- Production et purification des anticorps bispécifiques anti-CD3xanti-CD19
- Détermination de l’effet des anticorps sur les cellules cancéreuses
2. Expérimentation animale de l’anticorps bispécifique…………...14
- Expériences d’allogreffes chez la souris
3. Etude de l’effet de cet anticorps chez des patients atteints du
Lymphome B Non Hodgkiniens…………………………………. 16
PROJET DE RECHERCHE.............................................................................18
- Production et purification des anticorps bispécifique anti-gp120Xanti-CD16
- Tests d’affinité et d’activité
CONCLUSION ET PERSPECTIVES…………………………………………. . .23
BIBLIOGRAPHIE…………………………………………………………………24

3
Remerciements
Nous remercions Laurence Nieto et Pascal Demange de nous avoir aidé et guidé dans notre projet tout au long de ces trois mois.
Nous remercions également Fabrice Dumas et Patrice Mascalchi de nous avoir reçus et de nous avoir donné de précieux conseils. Cela nous a permis d’avancer dans notre idée de cibler le HIV.
Merci également à Etienne Joly pour son entretien grâce auquel nous avons décidé de bloquer la protéine gp120 du HIV et aussi de sélectionner les cellules NK.
Merci d’avoir cru en notre projet. Ce fut une courte aventure très encourageante et qui nous a permis de développer une idée que nous pensons réellement applicable. Nous avons partagé une belle expérience humaine. Peut-être verrons-nous un jour prochain une publication avec la même idée…

4
Résumé Au cours des dernières années, de nouveaux agents pharmacologiques, basés sur l'utilisation d'anticorps monoclonaux sont apparus en cancérologie. Ces agents s'appuient sur le fait que les cellules tumorales expriment des cibles antigéniques à des niveaux plus élevés que les cellules normales. Il existe deux types d'anticorps monoclonaux qui peuvent être soit non conjugués, soit conjugués dits «armés» à un agent cytotoxique qui sera apporté directement aux cellules tumorales. Le recours à des anticorps monoclonaux dits «armés» consiste à utiliser les anticorps monoclonaux comme des vecteurs permettant une délivrance ciblée d'autres molécules (toxine, enzyme...) exerçant un effet cytotoxique directement sur leurs sites d'action. D'autres approches sont également envisagées comme le recours à des anticorps monoclonaux particuliers, par exemple des anticorps bispécifiques, des anticorps à action intracellulaire ou encore des fragments d'anticorps recombinants.
Parmi les anticorps monoclonaux non conjugués actuellement très étudiés et utilisés en thérapeutique, il existe deux récepteurs de la famille tyrosine kinase, HER2 (Human Epidermal Growth Factor Receptor-2) : le trastuzumab, EGFR (Epidermal Growth Factor Receptor) : le cetuximab. Un troisième anticorps monoclonal couramment utilisé cible un marqueur transmembranaire du Lymphome Non Hodgkinien B, le CD20 (rituximab). Ces mêmes cellules immunitaires sont la cible d'autres anticorps qui sont en cours d'évaluation clinique, avec des résultats prometteurs. Notamment le Blinatumomab qui est un anticorps bispécifique qui a montré d'excellents résultats pour le traitement du Lymphomes B Non Hodgkinien chez l'Homme. Par son approche bispécifique il a une affinité pour deux antigènes différents : Le CD19 et le CD3. Il permet ainsi d'amener à proximité des cellules cancéreuses (les Lymphocytes B) les cellules effectrices (lymphocytes T cytotoxiques) permettant ainsi l'élimination des cellules cancéreuses. Les auteurs ont dans un premier temps dû passer par une étape d'humanisation de l'anticorps pour pouvoir l'utiliser sur des patients. Dans un second temps, ils ont pu produire et purifier cet anticorps bispécifique. Ensuite, l'efficacité de cet anticorps pour recruter les cellules effectrices et cancéreuses a été testée. Enfin, la lyse des cellules cancéreuses en présence de cet anticorps a été vérifiée. Avant de pouvoir réaliser les tests chez l'homme les auteurs ont testé leur anticorps ex-vivo sur des cellules de patients, puis chez la souris et enfin chez le chimpanzé. Dans ce mémoire, nous avons proposé un projet de recherche : L'utilisation d'anticorps bispécifiques pour le traitement du VIH. Il sera basé sur le même principe que le Blinatumomab. Les objectifs de ce projet seront de créer trois anticorps bispécifiques ayant une affinité à la fois pour les antigènes CD16 des cellules Natural Killer (NK) et pour la gp120 du VIH au niveau de son site d'interaction avec le CD4. Ils permettront de recruter directement les cellules immunitaires NK activées à la surface des cellules gp120+ et induiront l'élimination des cellules infectées.

5
Introduction
Les anticorps sont des glycoprotéines de la superfamille des immunoglobulines. Ils participent activement aux défenses de l’organisme et sont sécrétés par les Lymphocytes B activés, les plasmocytes.
Au cours de la réponse immunitaire les anticorps ont trois fonctions. Ils se lient à l’antigène, activent la cascade du complément et recrutent les cellules immunocompétentes. Les anticorps utilisés en immunothérapie contre le cancer peuvent neutraliser des cellules cancéreuses. Ils se fixent par leur paratope (partie Fab de l’anticorps) sur les épitopes des antigènes situés à la surface des cellules cancéreuses et peuvent déclencher la cascade du complément. Les cellules cancéreuses sont perforées et la phagocytose est facilitée. L’élément pathogène est alors lysé. Les anticorps après avoir reconnus un antigène peuvent également se fixer aux récepteurs Fc (FcR, les récepteurs du fragment constant) des cellules immunocompétentes : macrophages, lymphocytes Natural Killer (NK), afin de déclencher la phagocytose ou le phénomène de cytotoxicité des lymphocytes NK. Les anticorps sont des protéines ayant une structure bien particulière qui est responsable de leur fonction. Ils sont constitués de deux chaînes lourdes (H) et de deux chaînes légères (L) reliées entre elles par des ponts disulfures (Figure 1A). Les chaînes lourdes sont formées de trois ou quatre domaines constants (CH) et d’un domaine variable (VH). Les chaînes légères sont formées d’un domaine constant (CL) et d’un domaine variable (VL). Le domaine variable de la chaîne légère et le domaine variable de la chaîne lourde forment le site de reconnaissance de l’antigène : le paratope (Fab : fragment antigen binding). L’anticorps peut donc lier deux molécules d’antigène. Le fragment constant (Fc) de l’anticorps permet la fixation du complément et la liaison aux récepteurs Fc (FcR) des cellules immunocompétentes. Le paratope est constitué de trois boucles CDR : CDR1, CDR2 et CDR3 (Complementary Determinant Region) par domaines variables. Ces boucles permettent la reconnaissance de l’antigène. Leur structure tridimensionnelle est assurée par quatres régions constantes qui constituent la charpente (Fr : Framework). Les régions Framework sont situées de part et d’autres des séquences CDR. Les CDR sont composés d’acides aminés spécifiques de la reconnaissance de l’antigène appelés SDR (Specificity Determinant Residues) (Figure 1B). La découverte en 1975 des anticorps monoclonaux (1) a permis d’imaginer que l’on pourrait utiliser ces protéines ayant une forte spécificité pour le traitement de maladie et notamment des cancers. En 1982, les premiers succès d’utilisation d’anticorps monoclonaux en clinique ont été publiés (2). Ils ont laissé entrevoir de larges possibilités d’utilisation thérapeutique des anticorps monoclonaux. Cependant, 12 ans plus tard un seul anticorps monoclonal avait reçu une autorisation d’utilisation clinique. Ceci a révélé des échecs répétés dans de nombreux essais cliniques et notamment pour le traitement des cancers. La principale cause de ces nombreux échecs sur les anticorps monoclonaux utilisés en thérapeutique était due à leur origine murine, le patient produisant des anticorps humains anti anticorps murin (AHAM). En effet, dans 84% des cas ces réponses immunes étaient importantes du fait de leur utilisation répétée et à forte dose (3). Ces AHAM induisent une élimination rapide des anticorps murins ainsi que des effets secondaires parfois fatals. De plus, la durée de vie des anticorps murins dans le sérum est courte et ont une capacité limitée pour recruter des effecteurs cellulaires ou les protéines impliquées dans la réponse immune (4). De fait, l’ingénierie des anticorps en vue de transformer progressivement les anticorps murins en anticorps humains s’est développée. La première étape fût de les transformer en anticorps chimériques, dans lesquels les régions constantes des chaînes lourdes et légères des immunoglobulines murines sont remplacées par des régions constantes humaines (5). Cette

6
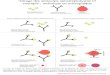
Figure 1 : Structure d’un anticorps, les régions hypervariables CDR et différentes formes d’anticorps humanisés utilisés en immunothérapie.
(A) Structure d’un anticorps, le fragment « antigen binding » Fab permet la reconnaissance de l’antigène via les régions variables V (blanc) formant le paratope et le fragment constant Fc est impliqué dans les fonctions de l’anticorps C (en noir). Sur les régions variables de la chaîne lourde et de la chaîne légère sont situées les trois régions complémentaires déterminantes CDR1, CDR2, CDR3. (B) Les boucles CDR sont les régions hypervariables du paratope impliquées dans la reconnaissance de l’épitope de l’antigène. Les SDR sont les acides aminés critiques du CDR (Specificity Determinant Residues). Les régions FR pour Framework sont les régions charpentes des CDR. (C) Anticorps entier humanisé conjugué à la ricine A sur le Fc. (D) Fragment d’anticorps bispécifique (single-chain bi-specific antibody) formé uniquement des parties variables (VH et VL) de deux anticorps et reconnaissant chacune un antigène spécifique.

7
première étape d’humanisation des anticorps murin a permis une diminution importante des réponses immunes. Entre 1994 et 1998 plusieurs anticorps chimériques ont ainsi pu être utilisés en clinique. Cependant la formation d’anticorps humain dirigés contre les anticorps chimériques (AHAC) est tout de même retrouvée dans 40% des patients soit deux fois moins que pour les anticorps monoclonaux murins (3). Il a donc était nécessaire d’améliorer ces anticorps chimériques en les humanisants encore plus. La première humanisation des anticorps a été décrite en 1986 (6) et consiste à produire des régions variables proches des régions humaines sans que l’anticorps humanisé perde l’affinité et sa spécificité de l’anticorps murin. Il faut transférer dans un cadre humain les acides aminés des régions CDR provenant de l’anticorps monoclonal d’origine murine. Les 6 régions variables des chaînes lourdes et légères contiennent la majorité des acides aminés constituant le site de liaison de l’antigène. Pour ce faire, il faut que la structure du site de liaison de l’antigène ne soit pas affectée, cependant en raison du rôle important de certains acides aminés des régions adjacentes aux CDR (régions charpentes ou FR, framework) dans la structure du site de liaison de l’antigène il est également nécessaire de greffer dans l’anticorps humain les acides aminés de la région charpente de l’anticorps murin (7). Or, certains de ces acides aminés sont potentiellement immunogéniques. Il faut alors rechercher les résidus « atypiques » qui sont rarement retrouvés dans les régions charpentes humaines et les remplacés dans le cadre humain par les résidus les plus fréquemment retrouvés dans les régions charpentes humaines aux positions correspondantes. De même pour éliminer les sites potentiels de N-glycosylation on effectue des changements d’acides aminés susceptibles de ne modifier que légèrement la conformation des régions CDR.
L’ingénierie des anticorps a ainsi permis de modifier et d’optimiser ces anticorps humanisés. Rapidement, des fragments d’anticorps sont développés. Ils sont dépourvus de la région Fc mais capables de lier l’antigène. L’objectif étant d’obtenir des petites molécules qui vont plus facilement accéder et pénétrer à l’intérieur des tumeurs solides. Les premiers fragments qui ont fait l’objet de nombreuses études sont les Fab et F(ab’)2. Ils ont d’abord été obtenus respectivement par digestion à la pepsine et à la papaïne avant d’être synthétisés par génie génétique (8). L’absence de la région Fc pour ces anticorps permet d’éviter une fixation aspécifique sur les tissus sains et diminue les réactions immunitaires de l’organisme. De ces deux fragments ont découlé plusieurs formats d’anticorps possibles dont les Fv (VH associé à un domaine VL) qui sont peu utilisés car n’étant pas reliés par une liaison covalente, ils se dissocient facilement à faible concentration. Ces fragments peuvent être stabilisés par un peptide de liaison qui relie le domaine VL au domaine VH. Ce format appelé scFv (pour single chain Fv) (9) de faible poids moléculaire (55 kDa) est le plus utilisé en thérapie. Ces fragments d’anticorps, de part leur petite taille ont l’avantage de pouvoir détecter de nouveaux épitopes (10).
Les fragments scFv ont été largement exploités pour la conception d’anticorps bispécifiques (Figure 1D). Ces derniers ont la capacité de se lier de façon simultanée à deux antigènes différents. Un site de liaison permet la fixation à un antigène tumoral et l’autre la fixation à des cellules effectrices du système immunitaire telles que les cellules Natural Killer ou les Lymphocytes T (11). La création d’anticorps bispécifiques permet d’apporter des solutions thérapeutiques innovantes en amenant directement des cellules effectrices à leurs cibles.
Afin d’augmenter leur efficacité, les anticorps ou fragments d’anticorps humanisés, principalement les scFv, peuvent également être couplé à des toxines telles que la ricine (Figure 1C), l’exotoxine A, la calicheamicin … ou à des cytokines. On parle alors d’anticorps conjugués ou immunotoxine (12). Lors de la reconnaissance de l’antigène sur la cellule cible, l’anticorps est internalisé, la toxine est libérée et provoque la mort cellulaire. Ce ciblage permet de concentrer

8
spécifiquement l’agent cytotoxique au niveau tumoral en évitant les effets secondaires provoqués lors d’injections systémiques.
L’utilisation des anticorps humanisés en clinique s’est petit à petit accélérée (13, 14, 15) et des résultats très prometteurs sur quelques maladies montrent que cela fonctionne correctement (14, 15). Même si tout n’est pas toujours bien contrôlé (14), les anticorps humanisés seront un traitement de choix dans les années à venir. D’une part grâce à leur faible dangerosité, contrairement à d’autres composés actuellement utilisés en chimiothérapies pour le traitement des cancers et d’autre part pour leur forte spécificité permettant de cibler spécifiquement un type cellulaire et donc un tissu particulier (15).
Résultats
1- Production d’anticorps bispécifiques :
Les Lymphomes Non Hodgkiniens (NHL) sont des tumeurs malignes du système lymphatique (moelle osseuse, la rate, le thymus, les ganglions lymphatiques, vaisseaux sanguins). Il existe deux types de lymphome, une forme ganglionnaire et une forme extra-ganglionnaire. Ces lymphomes peuvent se développer à partir des Lymphocytes T ou Lymphocytes B (16). L’étude utilisant les anticorps bispécifiques sur laquelle nous nous appuierons porte sur des patients atteints des lymphomes les plus répandus (85% des cas) touchant les lymphocytes B (LB).
Les auteurs ont choisi de créer un anticorps fragment bispécifique humanisé appelé
Blinatumomab qui cible le récepteur membranaire CD19. Ce dernier est spécifiquement exprimé par les LB à la plupart des stades de développement (17). C’est un récepteur essentiel pour leur prolifération et son expression est fortement conservée dans les tumeurs impliquant des LB (18).
Le second antigène reconnu par l’anticorps bispécifique cible les effecteurs cellulaires capables de lyser les cellules cancéreuses (LB). Les auteurs utilisent les cellules du corps humain les plus cytotoxiques : les lymphocytes T cytotoxiques (LTc). La majorité des thérapies utilisant un anticorps monoclonal font intervenir le FcγR. L’engagement des LT ne peut pas se faire ainsi car ils ne possèdent pas de FcγR, l’approche bispécifique s’est alors imposée. Le recrutement des LT se fait en ciblant le complexe CD3 qui est un récepteur cellulaire présent à la membrane de tous les LT.
- Production et purification des anticorps bispécifiques anti-CD19xanti-CD3 : La création d’anticorps humanisés bispécifiques est une ingénierie qui se fait en plusieurs étapes. La première consiste à récupérer les ARNm correspondant aux anticorps monoclonaux produits par les hybridomes ayant la meilleure affinité pour l’antigène correspondant (CD3 et CD19 respectivement) (43,19). Par une réaction de reverse transcription utilisant des hexaprimers ou des oligodT, l’ADNc correspondant à l’anticorps murin issu de l’hybridome est obtenu. Puis, uniquement les séquences de l’ADNc correspondant aux domaines variables (scFv) de l’anticorps

9
Figure 2 : Stratégie de construction et de production de l’anticorps bispécifique CD19xCD3.
(A) Etapes de conception de l’ADNc. Après récupération de l’ARNm correspondant aux anticorps monoclonaux anti-CD3 et anti-CD19 dans les hybridomes, une reverse transcription permet d’obtenir l’ADNc correspondant à l’anticorps murin anti-CD3 et l’anti-CD19. Les deux domaines du Fab VH (Chaîne variable lourde) et VL (Chaîne variable légère) sont isolés et amplifiés par PCR, en violet sont représenté de manière schématique les CDRs. (B) Humanisation de l’ADNc murin, contenant uniquement les domaines variables, par alignement de séquence avec la banque de donnée « Kabat data base ». Certains acides aminés charpentes et SDR (représenté de manière schématique en rouge) sont mutés pour humaniser l’anticorps. (C) Clonage des deux séquences en fusion séparées par un linker Gly4-Ser1 ou (Gly4-Ser1)3. Ajout de séquences K (séquence 5’ de Kozak), s.s (séquence de sécrétion eucaryote), F (peptide Flag), séquences H (tag histidine) et tt (terminateur de transcription) permettant sa production. (D) Clonage dans un vecteur d’expression eucaryote sous l’expression du promoteur EF (human elongation factor lα). Expression bicistronique, par la présence de r.r (ribosome réinitiation site), du gène DHFR permettant la sélection.

10
sont amplifiées par PCR (Figure 2A). Lors de cette réaction de PCR, les CDR sont modifiés par mutagenèse dirigée de manière à optimiser les acides aminés importants pour la reconnaissance de l’antigène et les acides aminés charpentes permettant un bon repliement des CDR. L’anticorps est ensuite humanisé pour éviter des réactions immunitaires non souhaitées. L’étape d’humanisation consiste à comparer les séquences issues de l’hybridome murin avec une banque de données de séquences d’anticorps humains « Kabat data base ». Les acides aminés potentiellement immunogènes du fait de leur nature murine sont ensuite modifiés (Figure 2B). Après l’humanisation, les séquences codantes pour les régions variables lourdes (VH) et légères (VL) humanisées dirigées contre les deux antigènes sont clonées en fusion dans le vecteur d’expression eucaryote pEF-DHFR. Comme l’anticorps sera utilisé dans un but thérapeutique, la production est réalisée en cellules de mammifère Chinese Hamster Ovary (CHO). En effet, d’autres modèles de production tels que Escherichia coli (E. coli) produisent des endotoxines dangereuses. De plus, la production par sécrétion fonctionne très bien dans les CHO contrairement à la sécrétion périplasmique chez E.coli. Le système de production en CHO possède surtout le grand avantage de produire l’anticorps bispécifique avec des modifications post-traductionnelles, les plus proches possibles de celles de l’Homme. Un vecteur d’expression eucaryote contenant toutes les séquences nécessaires à une production dans ce système est donc utilisé, ainsi qu’un tag Flag pour la détection. De plus, un linker constitué de GGGGS ou (GGGGS)3 est ajouté entre les séquences codantes des deux scFv. Ce linker permet de rendre flexible les domaines variables de l’anticorps et qu’ils puissent agir indépendamment (Figure 2C). La production se fait dans une souche CHO DHFR- pour sélectionner uniquement les cellules ayant intégrées la construction dans leur génome. En effet, le vecteur est exprimé de façon bicistronique en permettant la synthèse de la DiHydroFolate Reductase (DHFR) pour la complémentation des souches DHFR- et donc leur survie ainsi que la synthèse de l’anticorps (Figure 2D). Les anticorps étant sécrétés dans le surnageant, la purification en est facilitée. Une précipitation des protéines du surnageant est réalisée et une purification sur colonne ni-NTA est effectuée via le tag histidine de notre anticorps. De plus, cette stratégie de construction a les avantages de produire des protéines sécrétées actives et sous forme d’une seule chaîne polypeptidique liant les quatre régions variables. Aucune étape de renaturation et de réassemblement des chaînes lourdes et légères n’est nécessaire. Ainsi, l’anticorps bispécifique ayant à priori de l’affinité pour les deux antigènes CD3 et CD19 a pu être produit à des concentrations de l’ordre du mg/L. Une utilisation thérapeutique de cet anticorps nécessite la vérification de son affinité pour les deux antigènes et son implication dans l’activation des LT induisant la lyse des LB (19).
- Détermination de l’effet des anticorps sur les cellules cibles cancéreuses : L’anticorps bispécifique ainsi produit doit avoir de l’affinité pour les deux antigènes. Plusieurs techniques permettent de vérifier cette double affinité in vitro tout en déterminant la constante d’affinité. Le Biacore est très sensible mais encore peu utilisé. Les techniques les plus utilisées sont l’ELISA (Enzyme-Linked ImmunoSorbent Assay) et la cytométrie en flux (ou FACS) car plus facile à mettre en œuvre. En ELISA les protéines correspondantes aux antigènes (CD3 et CD19) sont fixées au fond des puits. La révélation est faite grâce à un anticorps secondaire anti-Flag couplé à la peroxydase qui reconnait notre anticorps bispécifique,. En cytométrie en flux, il s’agit d’utiliser différentes lignées cellulaires connues pour n’exprimer qu’un seul des deux récepteurs, par exemple les lignées Daudi, Blin-1, Raji, BJAB et SKW6.4 sont connues pour exprimer le récepteur CD19, et la lignée Jurkat le récepteur CD3. Les lignées L363 et NCI n’expriment aucun des deux récepteurs membranaires. Elles sont utilisées comme témoins négatifs.

11
Figure 3 : Détermination par cytométrie de flux de la spécificité de fixation de l’anticorps bispécifique sur ses épitopes à la surface des cellules.
(A) Analyse en FACS de la fixation de l’anticorps bispécifique sur des lignées de cellules CD19+ (BJAB et Raji), CD3+ (Jurkat) et sur des cellules qui n’expriment aucun des deux récepteurs CD3-CD19- (L363) servant de témoin négatif. En pointillé le profil avec un anticorps non spécifique anti-tag histidine-FITC (témoin négatif). Pour les lignées CD19+ et/ou CD3+ un décalage du pic vers la droite est observable par rapport au témoin négatif ce qui indique une forte positivité. L’anticoprs reconnaît donc le CD19 et le CD3. (B) Mesure de la fixation de l’anticorps bispécifique sur des cellules contrôles CHO n’exprimant aucun des deux récepteurs, cellules cancéreuses NALM-6 exprimant le CD19 et sur des LT de patients exprimant le CD3. Les trois profils de FACS correspondent à l’absence (à gauche) d’anticorps bispécifique, à la présence d’anticorps à 0,4µg/mL (au milieu) et avec 10µg/mL d’anticorps (à droite). A 0,4µg d’ancticorps/mL le pic pour les LT est presque confondu avec le témoin négatif. A cette concentration l’anticorps n’est spécifique que pour le CD19 (fort décalage du pic). A 10µg d’anticorps/mL le pic est fortement décalé pour les deux lignées. A cette concentration l’anticorps reconnaît spécifiquement le CD19 et le CD3.

12
L’utilisation de l’anticorps non spécifique anti-tag histidine montre la spécificité d’interaction pour l’anticorps bispécifique (Figure 3A). La capacité de l’anticorps bispécifique pour interagir avec ses épitopes a été confirmée in vitro sur des cellules de patients (Figure 3B). Cet anticorps bispécifique a la capacité d’induire la prolifération et l’activation des lymphocytes T qu’il reconnaît via le CD3 sans ajout d’interleukines 2 activatrices de la prolifération des LT. L’induction de la prolifération est vérifiée par essai de relarguage de tritium. Il s’agit de mettre les cellules en présence de thymidine tritiée pendant une durée de 24 ou 48h. La thymidine étant une base de l’ADN, les cellules en prolifération incorporent de la radioactivité dans leur ADN en cours de réplication. Ceci a permis de confirmer que la présence de l’anticorps et des LB permet l’induction de la prolifération des LT (Figure 4C). De plus, un essai de relarguage de chrome radioactif a permis de montrer que la présence de l’anticorps permet l’activation des lymphocytes T et l’induction de la lyse des LB. Pour ce faire, les cellules qui doivent être lysées sont mises en présence de chrome radioactif pendant 2h. Elles incorporent le chrome radioactif naturellement pendant leur croissance. Puis, l’ajout d’anticorps et de cellules effectrices permet le relarguage du chrome dans le milieu suite à la lyse des cellules. S’il n’y a pas lyse, il y a une quantité de chrome relargué équivalente au relarguage spontané, quantité très inférieure à la lyse. Cette expérience a pu mettre en évidence le fait qu’il ne suffisait que de très peu d’anticorps pour permettre la lyse des LB par les LT (Figure 4D). Enfin, pour vérifier que cette lyse est bien induite par l’anticorps bispécifique, Löffler et al. ont fait une compétition de l’anticorps bispécifique avec des anticorps monoclonaux, soit l’anticorps anti-CD3, soit l’anticorps anti-CD19. Ils observent que plus la concentration en anticorps compétiteurs augmente, plus la lyse diminue. Les anticorps monoclonaux se fixent à la place des anticorps bispécifiques. Cela confirme la spécificité des anticorps vis-à-vis de la lyse (Figure 4E) (19).
Ces résultats in vitro ont également été vérifiés sur des cellules de patients et les mêmes résultats ont été obtenus (20,21). Les études in vitro sur des lignées cellulaires exprimant les récepteurs CD3 et CD19 ainsi que sur des cellules sanguines prélevées sur des patients (20,21) ont ainsi permis de montrer que l’anticorps bispécifique CD19xCD3 a une très haute activité de lyse des LB dans une gamme de quelques picogrammes d’anticorps par litres (Figure 4C, D, E) (22).
Pour vérifier le potentiel thérapeutique in vivo de cet anticorps, Dreier et al. ont effectué des
expériences d’allogreffes chez la souris et des tests chez le chimpanzé (23).

13
Figure 4 : Effet de l’anticorps bispécifique sur la prolifération des LT et leur activation pour lyser les LB.
(C) Induction de la prolifération des LT en présence de LB : Essai de prolifération à la thymidine tritiée. L’induction de la prolifération des LT par l’IL-2 est définit comme la valeur 100% d’induction. Les tests sans anticorps (medium) et avec un anticorps non spécifique (bsc17-AxCD3) montrent une faible activation de la prolifération avec (noir) ou sans (gris) la présence de LB. Une forte activation de la prolifération des LT est observée avec 10ng/mL d’anticorps bispécifique et uniquement en présence des LB. (D) Mesure de cytotoxicité des LT par essai de relarguage au chrome radioactif sur différentes lignées cellulaires. Les NCI et L363 n’expriment pas le récepteur CD19 alors que les Daudi (triangle) l’expriment. (E) Panel du haut : Mesure de la spécificité de l’anticorps bispécifique pour le CD19 : Ajout de différentes concentrations d’anticorps monoclonal anti-CD19 avec (triangle) ou sans (losange) anticorps bispécifique. Panel du bas : Mesure de la spécificité de l’anticorps bispécifique pour le CD3 : Ajout de différentes concentrations d’anticorps monoclonal anti-CD3 avec (triangle) ou sans (losange) anticorps bispécifique.

14
2- Expérimentation animale de l’anticorps bispécifique :
- Expériences d’allogreffes chez la souris :
L’efficacité in vivo de l’anticorps bispécifique CD19xCD3 a été testé dans des souris (NOD)/SCID. Ces souris ne possèdent plus de LB, de LT, ni de NK. Elles n’ont plus de réponse immunitaire adaptative (24).
Pour tester le rôle anti-tumoral de cet anticorps, une xénogreffe est faite par injection sous-cutanée de 104 LB cancéreux. A partir de ces cellules injectées une tumeur se développe et cette croissance tumorale est mesurée. Pour cela, une cinétique de la mesure du volume moyen de la tumeur est effectuée. Ainsi, pour étudier l’effet de l’anticorps sur la croissance tumorale, Prochazka et al. l’injectent avec les Peripheral Blood Mononuclear Cell (PBMC) humain (contenant les LT) dans des souris qui développent des tumeurs constituées de LB.
Ils montrent que toutes les souris n’ayant pas reçues de traitement ou un traitement à des
doses faibles développent une tumeur et meurent entre 30 et 40 jours après l’inoculation. Les souris sont euthanasiées lorsqu’elles développent de sévères symptômes neurologiques et montrent des signes de paralysie. Alors qu’à partir d’une dose de 0,1μg d’anticorps injectés, aucune souris ne développe de tumeurs et elles survivent à l’injection de cellules cancéreuses. Ces résultats prouvent l’activité anti-tumorale de l’anticorps bispécifique CD19xCD3 in vivo (Figure 5A).
Afin de vérifier la spécificité de l’anticorps pour l’activité anti-tumorale observée, les souris
sont traitées avec un anticorps bispécifique Ep-CAMxCD3 ayant montré des effets anti cancéreux (25,26). Cet anticorps bispécifique partage le même paratope anti-CD3 que l’anticorps bispécifique CD19xCD3 mais au lieu d’avoir de l’affinité pour le CD19 cet anticorps reconnaît l’Ep-CAM humaine. Cet anticorps étant actif sur les cellules Ep-CAM positives et non actif sur les cellules CD19 positives, il permet donc de contrôler la spécificité. Toutes les souris traitées avec cet anticorps Ep-CAMxCD3 développent une tumeur et meurent de manière comparable aux souris non traitées. Comme il a été montré précédemment, les souris traitées avec l’anticorps bispécifique CD19xCD3 présentent une croissance tumorale inhibée(Figure 5B). Ceci prouve la spécificité de l’anticorps CD19xCD3 dans le traitement des tumeurs dues aux LB chez les souris.
Enfin, la toxicité de l’anticorps a été évaluée, pour cela ils ont réalisé des injections
intraveineuses d’anticorps à des concentration croissantes jusqu’à 3 injections de 30µg d’anticorps. Les souris traitées ne meurent pas, ils en concluent que l’anticorps n’a pas d’effet toxique (Figure 5C).
Ces résultats observés chez la souris ont été confirmés chez un animal très proche de
l’homme : le chimpanzé (27). En injectant l’anticorps dans des chimpanzés, les auteurs ont ainsi pu observer que l’anticorps bispécifique active les LT malgré sa très courte durée de vie dans le sérum. En parallèle une diminution en lymphocytes B est également observable (Figure 5D).
Comme il a été noté in vitro les LT n’ont pas besoin d’être pré-activés par l’IL-2 pour être actifs. En effet, ils s’activent in vivo au contact du LB cancéreux via l’anticorps bispécifique aussi bien in vitro que chez la souris et le chimpanzé. Ces résultats encourageant ont permis à Bargou et al. de débuter les tests cliniques chez des patients atteints de Lymphomes B non Hodgkiniens (15).

15
Figure 5 : Test d’activité de l’anticorps bispécifique CD19xCD3 chez la souris et le chimpanzé.
(A) Effet de l’anticorps bispécifique CD19xCD3 sur la croissance des tumeurs suite à l’injection sous cutanée de cellules LB cancéreuses. Cinétique du volume moyen des tumeurs suite à l’injection des cellules cancéreuses à D=0. Les cinq flèches correspondent aux cinq injections d’anticorps bispécifiques à 1heure, 1jour, 2jours, 3jours et 4jours après l’inoculation aux doses indiquées. La mesure du volume de la tumeur débute à 22 jours. En blanc les souris « témoins négatif » qui n’ont pas reçu d’anticorps (losange blanc) ou de LT (carré blanc). Les carrés et losanges noirs correspondent aux faibles concentrations en anticorps. Les ronds et triangles noirs à des fortes concentrations en anticorps. Le chiffre 0/5 ou 5/5 correspond au nombre de souris en vie à la fin du traitement. (B) Spécificité de l’anticorps bispécifique CD19xCD3. Les carrés et losanges blancs correspondent aux souris « témoins négatif » qui n’ont pas reçu d’anticorps ou de cellules effectrices. Les ronds noirs correspondent à la mesure du volume tumoral suite aux cinq injections de 1µg d’anticorps non spécifique Ep-CAMxCD3. Les triangles noirs correspondent à l’injection de 1µg d’anticorps spécifique CD19XCD3. (C) Effet de l’anticorps sur la survie des souris. Trois injections d’anticorps aux doses indiquées ont été réalisées 1heure, 1 jour et 2 jours après l’inoculation. Les animaux sont euthanasiés quand des signes de paralysie apparaissent. (D) Mesure de l’activité cytotoxique de l’anticorps sur les LB du chimpanzé.

16
3-Etude de l’effet de cet anticorps chez des patients atteints du Lymphome B Non Hodgkiniens :
Après les multiples étapes du développement pré-clinique, les premiers essais thérapeutiques sur l'homme ont pu être réalisés, il s’agit de la phase 1 du développement clinique. Ce dernier a été mené sur des patients atteints du Lymphome B Non Hodgkinien indolent (15). Ces patients avaient été traités au préalable par diverses chimiothérapies et par des injections de Rituximab (87% des patients), un anticorps anti-CD20. Ils présentaient une tumeur d’une taille supérieure ou égale à 1,5 cm évalué par tomographie.
Il a été montré qu’une très faible dose de l’anticorps bispécifique anti-CD19 et anti-CD3 est suffisante pour réduire la tumeur de patients atteints du Lymphome B Non Hodgkiniens. En effet, après différentes doses de Blinatumomab testées, les auteurs ont observé une régression partielle ou totale de la tumeur à partir d’une dose de 0.015 mg/m2/jour chez 21% des patients. A 0.060 mg/m2/jour, tous les patients traités présentaient une régression de la tumeur. Comme prédit par les études pré-cliniques, dès la première injection de Blinatumomab, une disparition des lymphocytes B du sang périphérique a été observé. Dès la fin du traitement, le nombre de lymphocytes B ré augmentent. Le suivi d’un marqueur de l’apoptose (Annexin V) leur a permis d’affirmer que la disparition des lymphocytes B était bien due à leur lyse. De même, le traitement a induit l’expression des marqueurs d’activation (CD69, CD25, HLA-DR+) des lymphocytes T CD8+. Les injections de l’anticorps à faible concentration ont donc permis de recruter les lymphocytes T CD8+ du sang périphérique des patients et de les activer en lymphocytes T cytotoxiques qui ont induit la lyse des lymphocytes B. De plus, les auteurs ont noté que le traitement n’a eu aucune influence sur le nombre de lymphocytes T.
Après avoir démontré l’efficacité du Blinatumomab dans le sang périphérique des patients, des biopsies ont permis de visualiser son effet sur des cellules appartenant à la moelle osseuse ou au foie (Figure 6A). En effet, après une injection de 0.015 mg/m2/jour de Blinatumomab, il a été observé par immunohistochimie et cytométrie en flux une élimination partielle ou totale des cellules tumorales de la moelle épinière dans 9 cas sur 11. Au niveau du foie, un traitement par 0.06 mg /m2/jour de Blinatumomab a permis d’obtenir une élimination totale des cellules tumorales (Figure 6B). Cependant, pour une telle concentration en anticorps des effets secondaires ont été noté, principalement une apparition de lymphopénie, leucopénie, pyrexie, une augmentation de la protéine C réactive, mal de tête, frissons etc. Des symptômes sont également observés au niveau du système nerveux central, ils sont totalement réversibles. En effet, la majorité de ces effets indésirables surviennent durant la première semaine de traitement et diminuent ou disparaissent par la suite.
Les tomographies permettent d’analyser l’efficacité de cet anticorps à l’échelle des organes (Figure 6C). Après 4 semaines de traitement à une dose de 0.015 mg/m2/jour, une régression partielle de la tumeur dans les ganglions lymphatiques a été observée. Par contre, après 8 semaines de traitement la tomographie d’un autre patient a révélé une régression complète de la tumeur dans les ganglions. (Figure 6D). Ainsi, pour une même dose, la durée du traitement influe sur le processus de régression de la tumeur. Cet anticorps termine maintenant sa phase clinique 2 qui étudie son activité sur des patients présentant un Lymphome B Non Hodgkinien aigu.

17
Figure 6 : Activités anti‐tumorales de l’anticorps. (A) Biopsies de moelle osseuse (patient 13), avant le traitement et 15 jours après avoir débuté le traitement avec 0.015mg/m2/jour de blinatumomab, le bleue indique les cellules tumorales (coloration à l’hematoxiline), le marron représente les lymphocytes T (marquage par l’antigène CD3). Une forte diminution du nombre de LB et une augmentation du nombre de LT sont observées. (B) Biopsies de foie (patients 33), avant le traitement et 4 semaines après avoir débuté le traitement avec 0.06mg/m2/jour de blinatumomab, marquage des lymphocytes B par l’antigène CD20. Une disparition des LB est visualisée. (C) Tomographies (patients 16) avant (Baseline) et après 8 semaines de traitement (Treatment) au blinatumomab à une dose de 0.015mg/m2/jour. Les flèches blanches indiquent deux tumeurs des ganglions lymphatiques dans la région pelvienne. Les tumeurs ont disparu de façon complète. (D) Tomographies (patient 15) avant et après 4 semaines de traitement au blinatumomab à une dose de 0.015mg/m2/jour. Les flèches blanches indiquent des tumeurs de ganglions lymphatiques dans le médiastin de la cage thoracique. Les tumeurs ont régressé de façon partielle.

18
Au vu de ces résultats, il serait intéressant d’utiliser la propriété de ces anticorps
bispécifiques pour lyser des cellules infectées par le virus de l’immunodéficience humaine-1 (VIH-1). Notre projet de recherche consistera en la création de trois anticorps humanisés bispécifiques pouvant recruter un effecteur cellulaire capable de lyser les cellules qui sont infectées par ce virus.
Les anticorps devront cibler un antigène de protéines virales exprimés à la membrane des
cellules infectées et un antigène exprimé à la surface d’effecteurs cytotoxique. Il y a deux protéines virales exprimées à la surface des cellules infectées : la gp41 et la gp120 (28). La protéine virale de surface gp120 apparaît comme la cible de choix puisque étant plus accessible aux anticorps que la gp41. Les effecteurs cytotoxiques les plus puissants sont les LT et les Natural Killer (NK). Les LT CD4+ étant les cibles du virus, ils ne pourront pas activer les LT CD8+ en LTc. Les LTc ne pourront pas être recrutés. L’effecteur utilisé sera donc les NK via leur récepteur CD16 (31). Leur potentiel pour lyser les cellules cible a été très étudié pour des anticorps bispécifiques (29, 32). De plus, l’activité cytotoxique anti-tumorale des NK a été démontrée sur le modèle du Lymphome Non-Hodgkinien grâce à l’utilisation d’un anticorps bispécifique CD19xCD16 (41).
Projet de recherche
Le VIH-1 infecte les macrophages présents dans les tissus et les lymphocytes T CD4+ circulants. L’infection se fait selon un mode séquentiel, en effet la protéine membranaire virale gp120 interagit avec le récepteur cellulaire CD4. Cette interaction provoque un changement conformationnel de la gp120 lui permettant d’interagir avec le corécepteur CCR5 ou CXCR4. La présence d’un des deux corécepteur est indispensable à l’entrée du VIH dans la cellule cible. Le trimère formé (CD4-gp120-corécepteur) permet à la gp41 de s’insérer à l’intérieur de la membrane cellulaire provoquant la fusion des membranes aboutissant à l’infection (28).
Naturellement, les cellules NK participent au système immunitaire humain par leur capacité à
détecter les cellules infectées par un virus et à provoquer leur lyse. Ceci est possible grâce à la présence à leur surface du récepteur CD16 (ou FcγRIIIa) qui fixe le fragment Fc des anticorps induisant l’ADCC (Antibody Dependent Cellular Cytotoxicity) (31). Les cellules NK sont dotées de granules cytoplasmiques contenant des granzymes et des perforines dont la libération provoque la perforation de la membrane de la cellule infectée et donc la lyse cellulaire. Les NK sont activés par la reconnaissance des antigènes viraux présentés à la surface des cellules infectées via leurs molécules HLA (ex : HLA B27 présente l’épitope viral gag KRWIIMGLNK). Cependant, la protéine Nef du VIH-1 provoque la réduction de l’expression et l’internalisation de ces protéines HLA. Ainsi, les NK ne reconnaissent plus les cellules infectées de patients.
Le projet de recherche consiste à créer des anticorps bispécifiques ayant une affinité pour les antigènes CD16 des NK et la gp120 virale. Ils permettront de recruter directement les NK activés à la surface des cellules infectés permettant d’induire leur lyse par ADCC.

19
Figure 7 : Construction de l’anticorps bispécifique anti‐CD16 et anti‐gp120.
(A) Première PCR pour assembler le domaine variable de la chaine lourde avec le domaine variable de la chaine légère par ajout d’un linker (Gly4Ser1). (B) Deuxième PCR permettant l’ajout des sites de restriction et d’un linker en amont du cDNA. (C) Clonage de l’anticorps bispécifique chez E. coli. (D) Après amplification chez E. coli, la séquence codante pour les deux scFv est clonée dans un vecteur d’expression eucaryote pEF DHFR (cf. légende figure 2). Les cellules CHO DG44 (DHFR-) sont transfectées de façon stable par le vecteur d’expression. L’anticorps bispécifique est directement sécrété.

20
1- Production et purification des anticorps bispécifiques anti-CD16xanti-gp120 :
Nous allons produire trois anticorps qui cibleront simultanément le CD16 et un des épitopes de
la gp120. La gp120 est une protéine très variable qui permet au VIH-1 d’échapper au système immunitaire. Cependant, quelques anticorps monoclonaux sont actuellement utilisés pour neutraliser le virus dans le sang via la partie constante de la gp120. Notre choix se porte sur les anticorps monoclonaux reconnaissant le site de liaison de la gp120 avec le CD4 du lymphocyte T lors de l’infection. Ce site varie très peu afin que le virus puisse conserver son affinité pour le récepteur CD4. Les anticorps b12, F105 et b13 provenant de sérum de patients infectés par le VIH-1, reconnaissent différents épitopes de cette région et ont été cristallisés en interaction avec la gp120 (30). Nous utiliserons donc leur paratope pour la construction de nos anticorps bispécifiques anti-CD16xb12, anti-CD16xb13 et anti-CD16xF105. Pour cela, nous contacterons les équipes qui ont crée ces anticorps afin de récupérer le plasmide codant pour l’anticorps monoclonal humanisé correspondant. Le vecteur contenant la séquence de l’anticorps b12 pourra nous être fourni par Raju Koduri et Dean Sauer (34, 35). Le vecteur de l’anticorps b13 nous sera fourni par Barbas et al. (36) Celui de F105 nous sera donné par Marchal Posner (37). Le vecteur codant pour le scFv de l’anticorps anti-CD16 sera fourni par Arndt et al. (38).
A partir des vecteurs codant pour la chaine légère et la chaine lourde des anticorps monoclonaux anti-gp120 et grâce aux analyses structurales qui ont été faites nous allons pouvoir récupérer par PCR uniquement les domaines variables VH et VL. En utilisant le logiciel « Kabat data base », un alignement bioinformatique de la séquence nucléotidique ou peptidique des anticorps avec une banque d’ADNc d’IgG humain permet de délimiter les zones variables. Une deuxième PCR permettra d’assembler les cDNA de la chaine lourde (VH) avec ceux de la chaine légère (VL) par ajout d’un linker GGGGS (33) (figure 7 A). La séquence codante pour le scFv anti-gp120 (b12, b13 ou F105) sera ainsi obtenue. Avant de passer à l’étape suivante, le scFv peut être produit chez E. coli afin de vérifier par un test ELISA qu’il a conservé son affinité pour la gp120.
Nous pourrons ensuite insérer cette séquence dans un vecteur contenant déjà le cDNA des domaines variables (scFv) anti-CD16 (figure 7 C). Pour cela une PCR sera réalisée afin d’insérer des sites de restriction ainsi qu’un linker GGGGS en amont du cDNA (figure 7 B). Ce linker permettra de lier les deux scFv (anti-CD16 et anti-gp120) pour obtenir des anticorps bispécifiques flexibles dont la reconnaissance de leur antigène se fait de façon indépendante. Il faudra effectuer un séquençage des plasmides.
Après amplification chez E. coli, le cDNA codant pour les deux scFv sera cloné dans un vecteur
d’expression eucaryote pEF DHFR (figure 7 D). Ce vecteur sera identique à celui utilisé pour produire l’anticorps bispécifique anti-CD19xanti-CD3 (39). Il comprendra toutes les séquences nécessaires à une production en système CHO ainsi qu’une séquence de sécrétion. Un Tag histidine et un Tag Flag seront également présents pour faciliter la purification et permettre de réaliser les tests ELISA par la suite. La production se fera dans la souche CHO DG44 (DHFR-) (figure 7 E). Ces cellules seront transfectées de façon stable et l’intégration du vecteur se fera au hasard. Le vecteur est exprimé de façon bicistronique, notre anticorps sera donc exprimé en même temps que la protéine DHFR. Cette protéine permet l’acquisition de la résistance au methotrexate. Nous traiterons donc les cellules au methotrexate (20 nM) pour sélectionner uniquement celles qui ont intégré notre séquence et qui pourront produire l’anticorps anti-CD16xanti-gp120 (39).

21
Figure 8 : Mesure de l’affinité et de l’activité de l’anticorps bispécifique.
(A) Mesure par cytométrie de flux de l’interaction de l’anticorps bispécifique avec les récepteurs cellulaires. Profil attendu en cytométrie de flux. On mesure le nombre de cellules en fonction de la fluorescence émise par l’anticorps fixé à son récepteur. Le profil en rouge correspond au comptage des cellules sans anticorps bispécifique. Le profil en vert correspond aux nombres de cellules avec incubation de l’anticorps. (B) Profil attendu sans anticorps bispécifiques (rouge) ou avec l’anticorps bispécifique non spécifique CD19xCD3, ou avec préneutralisation des récepteurs cellulaires avec les anticorps monoclonaux anti-CD16 et anti-gp120 (en vert) (C) Profil de cytométrie de flux double dimension contenant la population de cellules effectrices CD16 (entourés en rose) et la population de cellules cible gp120+ (entourés en rouge). La population de cellules entourées en vert correspond à d’autres cellules issues des purifications. (D) Mesure par cytométrie de flux double dimension de la lyse des cellules cibles par les NK via l’anticorps bispécifique. L’ajout de l’anticorps bispécifique induit la lyse des cellules cibles détectable par la disparition de la population entourée en rouge.

22
Chaque anticorps bispécifique sera produit par sécrétion grâce à l’ajout de la séquence de
sécrétion eucaryote (40). Les surnageants de culture seront précipités au sulfate d’ammonium. La purification des protéines du surnageant se fera en trois étapes. La première consistera à réaliser une chromatographie d’affinité IMAC pour fixer notre anticorps bispécifique taggué Histidine sur une colonne Ni-NTA (Quiagen). L’élution se fera avec de l’imidazole à 200mM. La seconde étape consistera à passer l’éluat de la première chromatographie sur une colonne d’exclusion Amicon YM10 ayant un cut-off de 30 kDa (la masse de notre protéine sera entre 55 000 et 60 000). A l’issue de la chromatographie nous ferons une dialyse afin de purifier notre protéine dans du tampon phosphate (PBS). Après chaque étape de purification nous effectuerons un Western Blot avec l’anticorps anti-Flag pour suivre notre anticorps bispécifique. Nous mesurerons la concentration en anticorps anti-CD16 et anti-gp120 par la méthode de Bradford (42).
2- Tests d’affinité et d’activité des anticorps : L’anticorps ainsi produit doit être capable de cibler simultanément le CD16 des NK et la gp120
à la surface des cellules infectées. Pour vérifier cette double spécificité, nous utiliserons des cellules HeLa gp120+ gp41+ qui exprime à leur surface les protéines virales gp120 et gp41. Les cellules NK purifiées nous permettront de montrer l’affinité de l’anticorps avec le récepteur CD16 (44).
En cytométrie de flux l’interaction entre les cellules et l’anticorps se fera dans du milieu de
culture avec 10% FCS. Nous incuberons 5.105 cellules avec 10 µg/mL d’anticorps. Un anticorps secondaire marqué au FITC permettra de révéler l’interaction, la fluorescence sera detectée par le FACS (FACScan flow cytometer de Becton Dickinson, Mountain View, CA). Nous comparerons les profils de cytométrie en flux en présence ou non d’anticorps bispécifique. Si l’interaction se fait correctement nous apercevrons un décalage du pic des cellules NK et des cellules HeLa en présence d’anticorps (Figure 8A).
Pour s’assurer de la spécificité de l’interaction, nous ferons un contrôle positif. Il consistera à
utiliser des anticorps monoclonaux anti-CD16 dirigés contre les NK et des anticorps monoclonaux anti-gp120 spécifique des cellules Hela avec lesquels nous devrions obtenir le même décalage au niveau du profil de cytométrie en flux. En tant que contrôle négatif, nous utiliserons l’anticorps bispécifique CD19xCD3 avec lequel l’interaction ne devrait pas se faire. En effet cet anticorps ne reconnait que le CD3 présent à la surface des lymphocytes T et le CD19 présent à la surface des lymphocytes B. (Figure 8B).
L’activité de l’anticorps bispécifique permettant aux NK de lyser les cellules infectées sera déterminée en utilisant les NK comme effecteurs et la lignée CEM-NKR comme cellules cibles (43). Les CEM-NKR sont des lymphocytes T sensibles à la lyse par les NK mais insensibles à la lyse spontanée des NK via les récepteurs activateurs et inhibiteurs KIR/KAR. Nous détecterons la lyse des cellules cibles par cytométrie de flux double dimensions (Figure 8C). Le profil du cytomètre lorsqu’il n’y a pas lyse sera représentatif de la population d’effecteurs et de cellules cibles. Si l’anticorps induit la lyse, la population correspondant aux cellules cibles ne sera plus présente sur le profil de FACS (Figure 8D). La lyse se fera en incubant les cellules NK avec les cellules CEM-NKR suivit de l’ajout ou non d’anticorps bispécifiques. Le ratio « Cellules effectrices/Cellules cibles » devra être déterminé. Pour cela, nous allons faire une gamme de différentes concentrations en effecteurs et mesurer l’impact sur la lyse.

23
Enfin, pour prouver que cette lyse est bien engendrée par la présence de l’anticorps bispécifique des contrôles seront également réalisés. Ils consisteront à ajouter l’anticorps bispécifique et des anticorps monoclonaux anti-CD16 ou anti-gp120 en excès de manière à neutraliser les récepteurs des cellules. Dans ce cas, l’ajout de l’anticorps bispécifique à une concentration permettant la lyse n’aura aucun effet sur les cellules cibles. Des contrôles négatifs seront aussi faits avec l’anticorps bispécifique CD16xCD3 qui pourra se fixer sur les NK mais pas sur la gp120 et qui sera donc incapable de lyser les cellules cibles. Enfin de la même manière que pour déterminer l’interaction, nous utiliseront l’anticorps bispécifique CD3xCD19.
Les cytokines ont un rôle prépondérant pour augmenter l’effet cytotoxique des cellules du
système immunitaire. C’est pourquoi nous devrons tester si l’ajout de cytokines (l’IL2 ou l’IL12) améliorera l’activité lytique des NK. Ainsi nous pourrions l’ajouter en tant qu’adjuvant dans les traitements, en prenant en compte les effets secondaires possibles.
Une fois les conditions de lyse mises au point nous déterminerons l’EC50 correspondant à la
concentration en anticorps permettant une lyse de 50% des cellules cibles. Pour cela il faudra mesurer le pourcentage de lyse en fonction d’une gamme de concentration croissante en anticorps.
PERSPECTIVES Une fois ces tests réalisés in vitro sur des cellules modèles, il faudra confirmer ces résultats sur
des cellules de patients (ex-vivo). Puis, les trois anticorps bispécifiques produits seront ensuite testés chez un modèle de souris NOD-SCID qui est génétiquement modifié pour exprimer les protéines d'enveloppes virale gp120 et gp41. La validation de l’effet thérapeutique chez la souris permettra d’étudier ces anticorps chez d'autres mammifères qui possèdent leur propre virus du SIDA (félin FIV, macaque SIV...). Enfin, des essais cliniques chez l’Homme pourraient être réalisés avec l’anticorps le plus efficace qui permet l'élimination des cellules infectées et dans les meilleures conditions expérimentales. Il serait même possible d’envisager une utilisation des trois anticorps simultanément si cela permet d’obtenir de meilleurs résultats. Les thérapies actuelles bloquent le cycle viral mais n'attaquent pas les cellules infectées. C'est pourquoi l'action combinée de ces thérapies bloquant le cycle viral avec l'immunothérapie en utilisant nos anticorps permettrait de bloquer la multiplication du virus et, en parallèle, d'attaquer le virus directement là où il se cache.

24
Bibliographie (1) Kohler G, Milstein C. Continuous cultures of fused cells secreting antibody of predefined specificity. Nature 1975 ; 256 : 495-7. (2) Miller RA, Maloney DG, Warnke R, Levy R. Treatment of B-cell lymphoma with monoclonal anti-idiotype antibody. N Engl J Med 1982 ; 306 : 517- 22. (3) Hwang WY, Foote J. Immunogenicity of engineered antibodies. Methods 2005 ; 36 : 3-10. (4) Qu Z Zhengxing Qu, Gary L. GriYths, William A. Wegener, Chien-Hsing Chang, Serengulam V. Govindan, Ivan D. Horak, Hans J. Hansen, David M. Goldenberg Development of humanized antibodies as cancer therapeutics. Methods 2005 ; 36 : 84-95. (5) Morrison SL, Johnson MJ, Herzenberg LA, Oi VT. Chimeric human antibody molecules: mouse antigen-binding domains with human constant region domains. Proc Natl Acad Sci USA 1984 ; 81 : 6851-5. (6) Jones PT, Dear PH, Foote J, Neuberger MS, Winter G. Replacing the complementaritydetermining regions in a human antibody with those from a mouse. Nature 1986 ; 321 : 522-5. (7) Tsurushita N, Hinton PR, Kumar S. Design of humanized antibodies: from anti-Tac to Zenapax. Methods 2005 ; 36 : 69-83 (8) Better M, Chang CP, Robinson RR, Horwitz AH. Escherichia coli secretion of an active chimeric antibody fragment. Science 1988 ; 240 : 1041-1043. (9) Bird RE, Walker BW, Single chain antibody variable regions. Trends Biotechnol 1991; 9 : 132-7. (10) Ward ES, Güssow D, Griffiths AD, Jones PT, Winter G, Binding activities of a repertoire of single immunoglobulin variable domains secreted from Escherichia coli Nature 1989; 341 : 544-546. (11) Baeuerle P. and Reinhardt Carsten, Bispécific T-Cell Engaging Antibodies for Cancer Therapy, Cancer Res 2009; 69 (12). (12) Brinkmann U and Pastan I, immunotoxins against cancer. Biochem Biophys Acta, 1994; 1198 : 27-45. (13) Aaron L Nelson and Janice M Reichert Development trends for therapeutic antibody fragments. Nature biotechnology 2009 ; 4 : 331-7. (14) Chen Z, Moayeri M, Zhao H, Crown D, Leppla SH, Purcell RH. Potent neutralization of anthrax edema toxin by a humanized monoclonal antibody that competes with calmodulin for edema factor binding. PNAS 2009 ; 106(32) : 13487-92. (15) Bargou R, Leo E, Zugmaier G, Klinger M, Goebeler M, Knop S, Noppeney R, Viardot A, Hess G, Schuler M, Einsele H, Brandl C, Wolf A, Kirchinger P, Klappers P, Schmidt M, Riethmüller G, Reinhardt C, Baeuerle PA, Kufer P. Doses of a T Cell_Engaging Antibody Tumor Regression in Cancer Patients by Very Low. Science 2008 ; 321(5891) : 974-7. (16) Kuppers R, Schwering I, Brauninger A, Rajewsky K, Hansmann ML. Biology of Hodgkin’s lymphoma. Ann Oncol. 2002 ; 13(suppl 1) : 11-18.

25
(17) Fujimoto M, Poe JC, Inaoki M, Tedder TF (1998) CD19 regulates B lymphocyte responses to transmembrane signals. Semin. Immunol. 1998 ; 10 : 267–277 (18) Scheuermann, R. H., and E. Racila. 1995. CD19 antigen in leukemia and lymphoma diagnosis and immunotherapy. Leuk. Lymphoma 1995 ; 18 : 385. (19) Löffler A, Kufer P, Lutterb¨use R, Zettl F, Daniel PT, Schwenkenbecher JM, Riethmuller G, D¨orken B, Bargou RC. A recombinant bispecific single-chain antibody, CD19xCD3, induces rapid and high lymphoma-directed cytotoxicity by unstimulated T lymphocytes. Blood 2000 ; 95 : 2098–103. (20) Dreier T, Lorenczewski G, Brandl C, Hoffmann P, Syring U, Hanakam F, Kufer P, Riethmuller G, Bargou R, Baeuerle PA. Extremely potent, rapid and costimulation-independent cytotoxic T-cell response against lymphoma cells catalyzed by a single-chain bispecific antibody. Int. J. Cancer 2002 ; 100 : 690-7.
(21) Löffler A, Gruen M, Wuchter C, Schriever F, Kufer P, Dreier T, Hanakam F, Baeuerle PA, Bommert K, Karawajew L, Dörken B, Bargou RC. Efficient elimination of chronic lymphocytic leukaemia B cells by autologous T cells with a bispecific anti-CD19/anti-CD3 single-chain antibody construct. Leukemia 2003 ; 17: 900-909. (22) Mølhøj M, Crommer S, Brischwein K, Rau D, Sriskandarajah M, Hoffmann P, Kufer P, Hofmeister R, Baeuerle PA CD19-/CD3-bispecific antibody of the BiTE class is far superior to tandem diabody with respect to redirected tumor cell lysis. Mol Immunol 2007 ; 44 : 1945–1953. (23) Dreier T, Baeuerle PA, Fichtner I, Grün M, Schlereth B, Lorenczewski G, Kufer P, Lutterbüse R, Riethmüller G, Gjorstrup P, Bargou RC. T cell costimulus-independent and very efficacious inhibition of tumor growth in mice bearing subcutaneous or leukemic human B cell lymphoma xenografts by a CD19-/CD3- bispecific single-chain antibody construct. J Immunol 2003 ; 170 : 4397-402. (24) Prochazka, M., Gaskins, H.R., Shultz, L.D., and Leiter, E.H. The nonobese diabetic scid mouse : model for spontaneous thymomagenesis associated with immunodeficiency. Proc. Natl. Acad. Sci. 1992; 89 : 3290–3294. (25) Schlereth B, Fichtner I, Lorenczewski G, Kleindienst P, Brischwein K, da Silva A, Kufer P, Lutterbuese R, Junghahn I, Kasimir-Bauer S, Wimberger P, Kimmig R, Baeuerle PA Eradication of Tumors from a Human Colon Cancer Cell Line and from Ovarian Cancer Metastases in Immunodeficient Mice by a Single-Chain Ep-CAM-/CD3-Bispecific Antibody Construct. Cancer Res. 2005 ; 65 :2882-2889. (26) Wimberger P, Xiang W, Mayr D, Diebold J, Dreier T, Baeuerle PA, Kimmig R. Efficient tumor cell lysis by autologous, tumor-resident T lymphocytes in primary ovarian cancer samples by an EP-CAM-/CD3-bispecific antibody. Int J Cancer 2003 ; 105: 241-8. (27) Schlereth B, Quadt C, Dreier T, Kufer P, Lorenczewski G, Prang N, Brandl C, Lippold S, Cobb K, Brasky K, Leo E, Bargou R, Murthy K, Baeuerle PA. T-cell activation and B-cell depletion in chimpanzees treated with a bispecific anti-CD19/anti-CD3 single-chain antibody construct. Cancer Immunol Immunother 2006 ; 55 : 503–14. (28) P. R. Clapham and A. McKnight, Cell surface receptors, virus entry and tropism of primate lentiviruses J. Gen. Virol. 2002 ; 83 : 1809-1829. (29) Ruggeri, L., Capanni, M., Urbani, E., Perruccio, K., Shlomchik, W.D., Tosti, A., Posati, S., Rogaia, D., Frassoni, F., Aversa, F., Martelli, M.F., Velardi, A., 2002. Effectiveness of donor natural killer cell alloreactivity in mismatched hematopoietic transplants. Science 295 (5562) : 2097–2100.

26
(30) Chen L, Do Kwon Y, Zhou T, Wu X, O'Dell S, Cavacini L, Hessell AJ, Pancera M, Tang M, Xu L, Yang ZY, Zhang MY, Arthos J, Burton DR, Dimitrov DS, Nabel GJ, Posner MR, Sodroski J, Wyatt R, Mascola JR, Kwong PD Structural basis of immune evasion at the site of CD4 attachment on HIV-1 gp120. Science. 2009. 326 (5956) : 1123-7 (31) Vivier, E., N. Rochet, M. Ackerly, J. Petrini, H. Levine, J. Daley, and P. Anderson. Signaling function of reconstituted CD16:z:γ receptor complex isoforms. Int. Immunol. 1992 ; 4 : 1313–1323. (32) R Zeidler, J Mysliwietz, M Csánady, A Walz, I Ziegler, B Schmitt, B Wollenberg and H Lindhofer. The Fc region of a new class of intact bispecific antibody mediates activation of accessory cells and NK cells and induces direct phagocytosis of tumour cells British Journal of Cancer 2000 ; 83(2) : 261–266
(33) Wang S, Zheng C, Liu Y, Zheng H, Wang Z. Construction of multiform scFv antibodies using linker peptide. J. Genet Genomics 2008 ; 35(5) : 313-6. (34) MARJAN HEZAREH, ANN J. HESSELL, RICHARD C. JENSEN, JAN G. J. VAN DE WINKEL, AND PAUL W. H. I. PARREN Effector Function Activities of a Panel of Mutants of a broadly neutralizing Antibody against Human Immunodeficiency Virus Type 1. Journal of Virology 2001 ; 75 : 12161-12168. (35) Burton DR, Pyati J, Koduri R, Sharp SJ, Thornton GB, Parren PW, Sawyer LS, Hendry RM, Dunlop N, Nara PL, et al. Efficient neutralization of primary isolates of HIV-1by a recombinant human monoclonal antibody. Science 1994 ; 266(5187) : 1024-7. (36) Barbas CF 3rd, Collet TA, Amberg W, Roben P, Binley JM, Hoekstra D, Cababa D, Jones TM, Williamson RA, Pilkington GR. Molecular profile of an antibody response to HIV-1 as probed by combinatorial libraries. J. Mol. Biol. 1993 ; 230(3) : 812-23.
(37) Posner MR, Cavacini LA, Emes CL, Power J, Byrn R. Neutralization of HIV-1 by F105, a human monoclonal antibody to the CD4 binding site of gp120. J Acquir Immune Defic Syndr. 1993 ; 6(1) : 7-14. (38) Arndt, M.A., J. Krauss, S.M. Kipiriyanov, M. Pfreundschuh, M. Little. A bispecific diabody that mediates natural killer cell cytotoxicity against xenotransplanted human Hodgkin’s tumors. Blood 1999 ; 94: 2562-2568.
(39) Mack M, Riethmüller G, Kufer P. A small bispecific antibody construct expressed as a functional single-chain molecule with high tumor cell cytotoxicity. Proc. Natl. Acad. Sci. 1995 ; 92 : 7021-7025.
(40) Von Heijne G. A new method for predicting signal sequence cleavage sites. Nucleic. Acids Res. 1986 ; 14 : 4683-4690. (41) Sergey M. Kipriyanov, Björn Cochlovirus, Holger J. Schäfer, Gerhard Moldenhauer, Alexandra Bähre, Fabrice Le Gall, Stefan Knackmuss, Melvyn Little. Synergistic antitumor effects of bispecific CD19xCD3 and CD19xCD16 diabodies in a preclinical model of Non-Hodgkin’s Lymphoma J. Immunol. 2002 ; 169 : 137-144. (42) Kruger NJ. The Bradford method for protein quantitation. Methods Mol Biol. 1994 ; 32 : 9-15. (43) Kantakamalakul et al.A novel EGFP-CEM-NKr flow cytometric method for measuring antibody dependent cell mediated-cytotoxicity (ADCC) activity in HIV-1 infected individuals J. of Immunol. Methods 315 (2006) 1–10

27
(44) Timonen T, Saksela E Isolation of human NK cells by density gradient centrifugation Immunol Methods. 1980; 36(3-4):285-91