Embed Size (px)
Citation preview

Acta histochem. 75, 69~8:3 (1984)
Institut d'Histologie et d'Embryologie generale (Directeur: Prof. Dr. C. CONT'), Universite de Fribourg, Suisse
Analyse de la structure et de la distribution de la chromatine des noyaux en interphase des differents types cellulaires se trouvant
dans un epanchement pleural humain d'origine tumorale Etude par cytophotometrie et microscopie electronique
Analyses of the chromatin's structure and distribution in interphase-nuclei of different cell-types
from a pleural effusion of tumoral origin in man: Cytophotometric and electronmicroscopical studies
Par VASSILIS GOTZOS, BONA CAPPELLI-GoTZOS et GIUSEPPE CONTIl)
Avec 2 Figures at Planche I
(Re<;u Ie 26 Octobre 1983)
Resume
Les auteurs ont etudie la distribution et la structure de la chromatine dans les differents types cellulaires se trouvant dans un epanchcment pleural humain d'origine tumorale (adenocarcinome mammaire). Les resultats montrent que dans les noyaux entiers la distribution et la structure de I" chromatine ne peuvent pas entrc considerees comme elements rnorphologiques caracteristiques de toutes les cellules d'un type cellulaire donne. Ainsi une distinction nette basee uniquement sur ces proprietes de la chromatine, entre cellules en synthese d' ADN ou en G2 et en differenciation est tres discutable. Par contre la distribution et la structure de la chromatine permettent une caracterisation des differents types de cellules se trouvant en G1• Les autenrs discutent la possible contribution de ces donnees au diagnostic cytologique differentiel des frottis provenant d'opanchements humains d'origine tumorale.
Summary
The authors have studied the distribution and the structure of the chromatin in the different cell types from a human pleural effusion of tumoral origin (mammary adenocarcinoma). The results show that in the entire nuclei the distribution and the structure of the chromatin cannot be considered as characteristic morphological features of the cells of the same cellular type. Thus making a clear distinction among differentiating cells and cells in DNA synthesis or in G2 period, only based on these properties of the chromatin, is very debatable. On the other hand, the distribution and the st,ructure of the chromatin permit to characterize the different cell types in G1 period The authors discuss the possible differential cytological diagnosis of the smears from human effusions of tumoral origin.
1) Les auteurs remercient Mile M. VONLANTHEN ct Mile M.-C. POCHON pour leur collaboration technique.

70 V. GOTZOS, B. CAPPELLI-GOTZOS et G. CONTI
Introduction
De nombreux travaux faisant appel it des techniques biochimiques et cytochimiqueS
extremement variees (FRENSTER et al. 1963; SANDRITTER 1970; KIEFER .et al. 1973; PAYS 1975; BRUTLAG 1980) permettent de degager une notion assez precise sur la signification de l'heterochromatine et encore plus de l'euchromatine. L'heterochromatine comprend de 10 it 97 %, selon les auteurs, de l'ADN total de la chromatine et elle apparait comme etant plus dense que l'euchromatine; cette derniere se trouve sous forme diffuse et peut etre definie com me une fraction minoritaire des sequences d'ADN (5 it 20 %), representant la portion active de la chromatine nucIeaire.
Le haut degre stoechiometrique de la reaction de FEULGEN et la densite optique du complexe «colorant + ADN» (KASTEN 1960; DEITCH 1966; PEARSE 1968; GANTER et JOLLES 1969) permettent actuellement par la methode de la cytophotometrie it balayage, de juger l'etat de condensation de la chromatine it travers son constituant Ie plus important, l'ADN, et sa distribution au niveau de chaque noyau considere separement. Cette methode presente d'autres avantages puisque en donnant les resultats cellule par cellule, elle permet de choisir Ie materiel etudie (elimination des cellules en degenerescence), de selectionner les cellules appartenant au meme type cellulaire et aussi d'effectuer une analyse detaillee des images microscopiques; on peut ainsi obtenir pour chaque cellule, des parametres supplementaires tels que la surface de projection, la circonference, Ie diametre.
La structure et la distribution de la chromatine des noyaux en interphase ont ete analysees par cytophotometrie it balayage et au microscope electronique dans des buts differents par plusieurs auteurs. Les resultats de ces travaux ont apporte des renseignements importants en ce qui concerne l'etat de condensation de la chromatine notamment dans les differentes etapes du cycle cellulaire (SAWICKI et al. 1974), dans les cellules en differenciation (ROWINSKI et al. 1972), dans Ie vieillissement cellulaire (JEANNY et al. 1978), dans les cellules tumorales par rapport aux normales du meme type (SANDRITTER et al. 1974), dans les cellules tumorales avant et apres traitement (BARNI et al. 1981).
II est generalement admis que la structure et la distribution de la chromatine representent un element morphologique caracteristique pour chaque type cellulaire, mais les travaux que nous venons de mentionner laissent prevoir qu'elles refletent aussi l'etat fonctionnel, it sa voir, l'etat d'activation nucleaire.
Ce travail est une approche it l'analyse de la structure et de la distribution de la chromatine nuclealre basee sur les proprietes biophysiques de son constituant principal I' ADN, mis en evidence par la reaction de FEULGEN et etudie par des methodes cytochimiques-morphologiques. Tout particulierement notre objectif etait celui de comparer la structure et la distribution de la chromatine dans les noyaux entiers des differents types cellulaires des epanchements pleuraux humains d'origine tumorale pour obtenir des renseignements soit en ce qui concerne l'activation nucleaire pendant la differenciation cellulaire [monocytes-macrophages; CHAPMAN et al. (1967)], soit en ce qui concerne l'etat de la chromatine nucleaire pendant Ie cycle cellulaire (cellules tumorales et autres en multiplication). En outre, l'application de cette technique it notre materiel peut fournir des donnees morphologiques supplement aires pour Ie diagnostic cytologique differentiel des cellules dont la morphologie au microscope optique est semblable (KRIVINKOVA et al. 1976).
Materiel et Methodes :Noire maten'll provient d'nn epanchement pleural chez nne patiente porteuse d'nn adeno
carcinome mammaire. 500 ml dn liqnide d'epanchement ont eM preleves sterilement; apres

Analyse de la structure et de la distribution de la chroma tine 71
centrifugation (10 min a 240g) les ceHules du culot ont He etalees sur frottis et immediatement fixees (ethanol: acetone = 1 : 1, 24 h it 4 DC). Pour la reaction de FEULGEX les frottis ont ete hydrates et hydrolyses dans l'HCL mol/I pendant 20 min a 56 DC, apres avoir ete prechauffes dans de l'eau distillee a 56 DC pendant 2 a. 3 min. Cette faible hydrolyse a He suivie par a lavages a l'eau distiHee a temperature ambiante. L'ADN hydrolyse a ete par la suite mis en evidence par la paranosaniline-SOz (120 min a l'abri de la lumiere et a temperature ambiante). Apres un rigoureux lavage a l'eau-SO" les frottis ont ete deshyrates et montes a l'eukitt.
Les mesures d'extinction "point par point ,) apres rea('tion de FEULGEN ont He effectuees sur les noyaux entiers et it 550 nm (longueur d'onde choisie d'apres la ('ourbe d'extinction de nos preparations) it l'aide cl'un cytophotometre it balayage :\IPV 2, (Leitz) equipe d'une platine it pas fins (O,a/lm) et d'un ordinateur PDP/8 (Digital). L'equipement optique du microscope consistait cI'une lampe it xenon (XBO 150 \V) d'un monochromateur it vi see rectiligne avec echelle graduee pour les valeurs comprises entre 370 nm et 1100 nm, d'un condensateur (20/0.33; s = 150/1,0 Leitz) et d'un objectif (NPI oil, 100/1,30 P, Leitz). Les valeurs d'extinction donnees par l'appareil ont ete converties par l'ordinateur en valeurs d'absorption : des images nucleaires ont ete obtenues d'apres ces mesures d'absorption, grace au programme FF.LD de Leitz. I.e carre do balayage a He toujours Ie meme pour tous los noyaux mesures: pour chaque deplacoment de la platine (0,5 pm) ont ete effectuees 5 mesuros consecutives.
Les resultats sont presontes soit sous formo de cartes ("cartes nucleaires ») montrant point par point la densite optique de la surface de projection du noyau, soit sous forme d'histogrammes montrant la distribution de la densite de I'ADN par noyau; sur les cartes, des classes arbitraires sont indiquees par differentes couleurs (fig. 1): blanc, de 14 it 25 unites arbitraires (u. a.); jaune de 26 it 30 u. a.; orange de 31 it 35 u. a.; rose de 36 a 40 u. a.; vert de 41 it 45 u. a.; bleu de 46 it 50 u. a. et enfin nair de 51 u. a. et plus. En se basant sur cette subdivision en classes, nous parlerons de valeurs d'absorption tres importantes pour celles representees par la couleur noir, de valeurs importantes pour celles representees par les couleurs bleu, vert, rose et orange et enfin de valeurs faibles pour celles representees par les couleurs jaune et blanc. Pour cette etude, nous avons retenu et considere comme specifiques a la coloration de l'ADN nucleaire, seulement les valeur d'absorption superieures a 14 u. a. (Ies inferieures etant eliminees par l'ordinateur). Cette valeur (14 u. a.) a ete arbitrairement fixee, avec une marge importante de surete, au double de 7 u. a. (= valeur maximum d'absorption donnee par les artefacts du fond en dehors de la surfaco rle projection nucleaire).
Chaque noyau a ete classe en periode G1 + % S ou % S + G2 du cycle cellulaire d'apres son contenu en ADN: les mesures des noyaux en mitose (prophase et metaphase) ont donne la valeur tetraplojde (4c) de l'espcce. La limite arbitraire entrc cellules en G1 + % Set cellules en % S + G2
a ete fixee it % de cotte valeur tetraploide.
N OU3 avons etudie la distribution de la densite optique de I' ADN nucleaire des types cellulaires suivants: cellules en mitose, petits lymphocytes, lymphocytes transformes, monocytes (histiocytes), macrophages, ceHules geantes mononucleees (macrophages actives ?), cellules mesotheliales et cellules tumorales. Les criteres morphologiques suivants ont He pris on consideration pour differencier au microscope optique les types cellulaires pouvant eire confondus:
1. Pour les monocytes: cellules avec diamCtre entre 15 et 20 /Im, noyau caracteristique reniforme sans signe de phagocytose.
2. Pour les macrophages: cellules avec diametre entre 20 et 30 pm, noyau ovale, rond ou reni· forme pousse en general vers la peripherie cellulaire; presence au niveau du cytoplasme d'un ou de plusieurs globules rouges phagocytes et de vacuoles non rMringeantes (plusieurs petites vacuoles ou une grande: lysosomes).
3. Pour les cellules geantes mononucleees (SATO et al. 1978): cellules avec diametre triple ou plus de celui du macrophage, noyau rond ou legerement ovale de 25 a :30 11m de diametre. On rencontre tr8s souvent des petits lymphocytes attaches it la surface de ces cellules (cellules geantes de type histiocytaire, macrophages actives 1).
4. Pour les cellules mesotheliales: cellules rondes isolees ou groupees (2 it 3) pr6sentant une bordure festonn6e, diametre entre 18 et 22/lm, noyau rond situ6 au centro cle la cellnle ayec 1 ou 2 nucleoles. I.e cytoplasme est abondant, c!epourvu de vacuoles de phagocytcse ou de globules rouges phagocytes.

72 V. GOTZOS, B. CAPPELLI·GOTZOS et G. CONTI
5. Pour les cellules tumorabs : cellules se trouvant en amas, Ie plus souvent de plusieurs clizaines de cellules, ne pr6sentant pas de limite cellulaire nette, avec cytoplasme se colorant dans des differentes intensites de rouge avec la PAPANICOLAOU.
Nous avons etudie plus de 2J noyaux pour chaque type cellulaire et les resultats out ete analyses avec Ie test du x-carre. Les "aleurs de Pont et6 etablies d'apres FISURER et al. (1949).
Les observations morphologiques au micros~ope optique et Ie diagnostic cytologique c!'adenocarcinome ont ete effectues sur des frottis colores par la GIEMSA et la PAPANICOL_\OF.
Une partie des cellules du culot de centrifugation ont ete employees pour I'observation au microscope etectronique: fixation en glutaraldehyde 2,5 % (dans cacodylate, pH = 7,2; 2 h it 4 "C), postfix at ion en OsO. (I % dans l"hLLONlG pH = 7,2, 2 h) et enrobees en Epon 812 d'apres LrFT Les coupes ultrafines (50 um) ont et6 effectu6es avec I'ultrotome III (LKB), contrast6es par I'acetate d'uranile et Ie nitrate de plomb d'aprbs REY-'lOLDS et photographiees au microscope Zeiss E1VI 10 (JR. Des con troles de la ('oloration llucl6aire sur les coupes ultrafines ont eM effectups par digestion avec ADNas8 neutre (vVorthington: 2500 u/mg, 0,2 mg/ml dans tampon phosphate pH = 7,0 con tenant 1ilgCl2, 0,003 mol, 24 h it ;38 'C).
Resultats
A) Analyse des images obtenues sur les cartes nucleaires d'apres la densite optique du complexe «colorant + ADN» de la reaction de FEULGEN
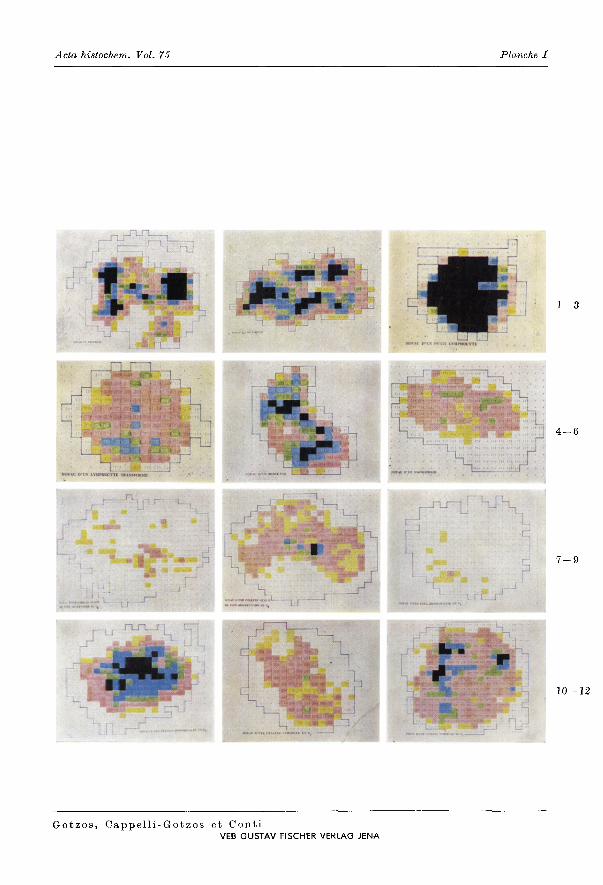
Selon les valeurs d'absorption donnees par la cytophotometrie it balayage, point par point, nous avons obtenu des cartes de distribution de la densite optique du complexe «colorant + ADN» nucleaire et nous avons reporte dans la Planehe lIes images representatives pour chaque type cellulaire pris en consideration. Dans la Fig. 1 (Planche I) Ie noyau d'une cellule en prophase: il existe plusieurs plages d'une absorption tres importante (nair), mais surtout une diversite de valeurs qui s'alternent du blanc au bleu sur tout Ie reste de la surface nucleaire. Dans les noyaux en metaphase (Fig. 2, Planche I) les plages d'absorption tres importantes (nm'r) sont beaucoup plus nombreuses et Ie reste de la surface de projection nucleaire est occupe par des valeurs importantes (surtout rose, vert et bleu) formant une mosaique de differentes intensites d'absorption. Les noyaux des petits lymphocytes (Fig. 3, Planche I) presen tent sur toute la surface de projection nucleaire une distribution uniforme des valeurs d'absorption tres importantes (nair) et seulement it la peripherie on remarque des points d'absorption faible (blanc). II faut retenir que, it part ces dernieres valeurs, l'intensite est comparable aux valeurs les plus hautes (nair) des noyaux des eellules en mitose. La distribution des valeurs d'absorption change profondement dans les noyaux des lymphocytes transformes (Fig_ 4, Planche I). lei, it part la peripherie nucleaire occupee par des valeurs faibles, ladistribution n'est pas uniforme eomn:edans les noyaux des petits lymphocytes, mais des plages de valcurs d'absorption importantes (vert, bleu) s'alternent avec d'autres de valeur moins importante (rose) qui occupent la plus grande partie de la surface de projection nucleaire. II faut souligner que les valeurs d'absorption, mome les plus importantes, des noyaux des lymphocytes transformes sont nettement inferieurs par rapport it celles mesurees dans les noyaux des petits lymphocytes. Si l'on compare maintenant les images des noyaux des monoeytes (Fig. 5, Planche I) et des macrophages en G1 + Y2 S (Fig. 6, Planche I) on constate des differences importantes et de l'intensite d'absorption et de sa distribution. Dans les monocytes on rencontre des points d'absorption tres importants (nair) localises aux 2 poles nucleaires. L'intensite d'absorption de ces points est comparable it eelle des valeurs les plus elevees rencontrees dans les noyaux des cellules en mitose et dans les noyaux des petits lymphocytes. Le reste de la surface de projection nucleaire des monocytes est occupe presque uniquement par des valeurs d'absorption import-

An'tlyse de la structure et de la distribution de la chroma tine 73
antes allant du rose au bleu et rares sont les valeurs d'absorption faibles. Dans les noyaux des macrophages en G1 + Y2 S (Fig. 6, Planche I) par contre on remarque des valeurs d'absorption importantes (rose, vert, bleu) nettement plus nombreuses par rapport it celles des noyaux des monocytes; leur distribution montre des grandes plages (rose, vert, bleu) s'alternant avec des plages d'une intensite tres importante (nair). Les cellules geantes mononucleees de type histiocytaire en G1 + Y2 S sont parmi celles dont les noyaux (Fig. 7, Planche I) presentent les valeurs d'absorption les plus faibles. En effet, on ne note que quelques rares points dont l'intensite d'absorption est importante (rose) tandis que la plus grande partie de la surface nucIeaire est occupee par des valeurs d'absorption faibles (jaune et surtout blanc). Pour les cellules du meme type se trouvant en Y2 S + G2 (Fig. 8, Planche I) on constate une augmentation de l'intensite d'abRorption sur la plus grande partie de la surface de projection nucIeaire. Dans les noyaux des cellules mesotheliales en G1 + Y2 S (Fig. 9, Planche I) les valeurs d'absorption sont parmi les plus faibles (blanc) que nous avons rencontrees. Cependant dans quelques noyaux on rencontre de rares points ayant une intensite d'absorption importante (rose). II faut souligner que frequemment dans les noyaux des cellules mesotheliales en G1 + Y2 S on rencontre des valeurs d'absorption de la meme intensite que celles du fond des preparations et par consequent eliminees par l'ordinateur comme etant non specifiques de l'ADN nucleaire. Les noyaux des cellules mesotheliales en Y2 S + G2 (Fig. 10, Planche I) montrent un aspect completement different de celui des cellules en G1 + Y2 S. Toute la surface nucleaire de ces cellules est occupee par des valeurs d'absorption importantes et on rencontre constamment dans tous les noyaux des plages dont l'intensite d'absorption (tres importante: noir) est comparable it celle des valeurs d'absorption les plus hautei: des noyaux en mitose et des noyaux des petits lymphocytes. Les noyaux des cellules tumorales en G1 + Y2 S (Fig. 11, Planche I) presentent un aspect tout it fait different soit par rapport aux noyaux des cellules geantes mononucleees en G1 + Y2 S, soit par rapport aux noyaux des cellules mesotheliales en G1 + Y2 S. La surface de projection de leurs noyaux est occupee par des plages dont l'intensite d'absorption est tres heterogene, it sa voir des plages dont les valeurs d'absorption sont importantes (rose) s'alternent avec des plages dont les valeurs d'intensite sont faibles (jaune); a ce point de vue les noyaux des cellules tumorales en G1 + Y2S se rapprochent de ceux des macrophages. Dans les noyaux en Y2 S + G2 des cellules tumorales (Fig. 12, Planche I) on constate une augmentation sur toute la surface de projection nucleaire, de l'intensite d'absorption par rapport aux noyaux en G1 + Y2 S. Presque to ute la surface est occupee par des pI ages d'une intensite d'absorption importante (rose, bleu, vert); il existe constamment dans les noyaux des valeurs d'absorption tres importantes (nair) comparables it, celles qu'on rencontre dans les noyaux des cellules en mitose et des petits lymphocytes.
B) Analyse de la frequence des valeurs d'absorption par noyau dans les differents types cellulaires etudies
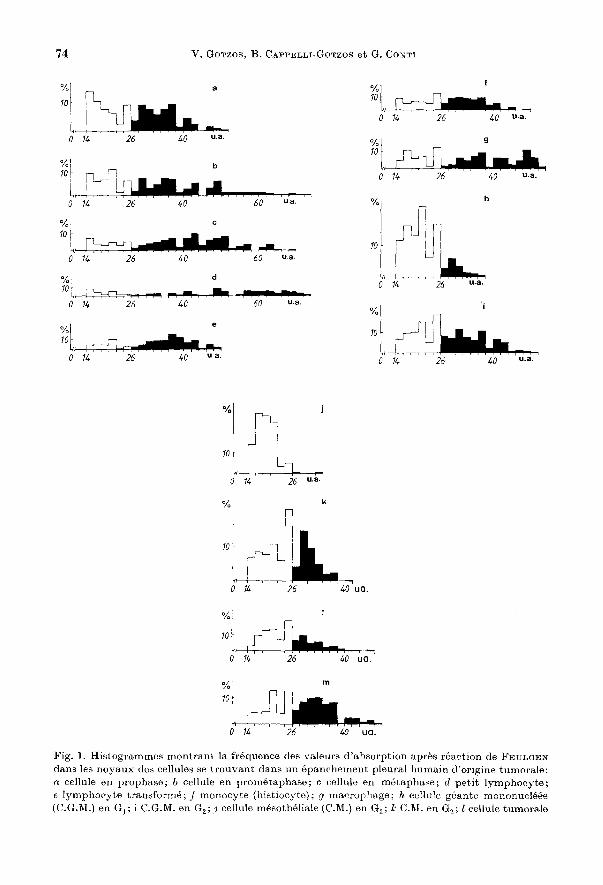
Les histogrammes de la Fig. 1 mont rent les frequences des valeurs d'absorption d'ADN des noyaux des differents types cellulaires. D'apres ces histogrammes les noyaux des cellules en mitose montrent des profils differents selon Ie stade dans lequel ils se trouvent; pendant la prophase (presence des nucIeoles et de l'enveloppe nucleaire) les va leurs sont regroupees entre 14 et 52 unites arbitraires (u. a.) presentant plnsieurs pics, dont les pIns hants sont situes dans les valeurs les plus faibles [(inferieures a 38 u.a.); (Fig.la)]. Par contre, si les noyaux se trouvent dans nn stade plus avance entre prophase et metaphase: prometaphase (Fig. 1 b), Ie profil de distribution est
74 V. GOTZOS, B. CAPPELLI-GOTZOS et G. CONTI
%! a %t IOf [2illw. 10 c=!l n n .. 1""'11 """""I
l 0 14 26 40 U.a.
~
14 26 40 u.a.
%l 9
10
~ .... 11. 0 14 26 40 U·a.
""' o/QI h 26 60 U.a.
%~ c
(~1. 10 .... • L, ~ ..,. ~ 10 o 14 26 40 60 U·a.
%L d 14 26 u.a.
10~ h=l---n .-- i'SI~ ~ PI!&l 0 14 26 40 60 U.a.
lc~ %t e
10 ~i :IIR ~
0 14 26 40 u a. o 14 26 40 u.a.
% k
10
o 14 26 40 ua.
". 014 26 40 ua.
%1 m
10, CYa~~ o 14 26 40 ua.
Fig. 1. Histogrammes mont rant la frequenee des valeurs d'a1)sorption apres reaction de FEULGEN dans les noyaux des cellules se trouvant dans un epanchement pleural humain d'origine tumorale: ct cellule en prophase; b cellule en prometaphase; c cellule en metaphase; d petit lymphocyte; c lymphoC'yt-e transforme; f monocyte (histiocyte); '1 macrophage; h cellule geante mononucleee (C.G.M.) en G1 ; i C.G.M. en G2 ;.1 cellule mesotheliale (C.M.) en G1 ; 7..' C.1\1. en G2 ; l cellule tumorale

Analyse de la structure et de la chromatine de la distribution 75
different. La gamme des valeurs est en effet tres etalee (entre 14 et 74 u. a.), les pics sont beaucoup plus nombreux et les plus hauts restent, ici aussi, localises dans les valeurs les plus faibles. Les noyaux des ceUules en metaphase presentent egalement un profil tres etale [(entre 14 et 74 u. a.); (Fig. lc)] comparable it celui des noyaux en prophase a vancee, mais ici les pics les plus hauts sont localises dans les valeurs d'absorption les plus elevees. On notera en plus que les pics places dans des valeurs inferieures it 26 u. a. sont considerablement plus bas par rapport it ceux des noyaux en prometaphase. Un comportement analogue it celui des noyaux en metaphase a eM observe dans les noyaux en anaphase (les mesures pour tel., noyaux ont ete effectuees pour ~haque groupe ehromosomique separement). Le profil des valeurs d'absorption d'ADN dans les noyaux des petits lymphocytes leur est typique par rapport aux autres noyaux en interphase (Fig. Id); la distribution des valeurs est tres etalee (entre 14 et 74 II. a.) comparable it ceUe des noyaux des ceUules en prophase avancee (prometaphase) et surtout des noyaux en metaphase et anaphase. Les pics les plus hauts sont localises surtout vers la droite de l'histogramme OU ils forment un plateau entre 56 et 70 u. a. Les noyaux des lymphocytes transformes presentent un profil tout it fait different de celui des petits lymphocytes (Fig. Ie). En effet, les valeurs d'absorption sont plus regroupees (entre 14 et 50 u. a.) et Ie nombre des pies est nettement inferieur. lci aussi, comme dans les petits lymphocytes, les pics les plus hauts sont distribues Oil bien dans les valeurs les plus faibles ou bien dans les plus elevees; ces derniers ne sont pas en plateau comme dans les petits lymphocytes mais ils forment une cloche dont Ie point Ie plus haut se situe entre 36 et 40 u. a. Pour les noyaux des monocytes Ie profil de distribution des va leurs d'absorption d' ADN differe de celui des antres cellules (Fig. If). Dans ce cas, les valeurs sont regroupees (entre 14 et 50 u. a.) comme pour les noyaux des lymphocytes transformes, mais les differents pics sont plus ou moins de la meme hauteur tout Ie long de l'histogramme. Les noyaux des macrophages presentent un profil different de celui des noyaux des monocytes et des autres cellules etudiees dans ce travail (Fig. 1 g): en effet, les pics les plus hauts sont nombreux et distribues tout Ie long de l'histogramme. On remarquera que les valeurs d'absorption sont regroupes entre 14 et 54 u. a.
Les noyalIx des cellules geantes 1110nonucleees en Gl + Y:! S 1110ntrent un profil tout it fait caracteristique (Fig. 1 h). On constate que les valeurs sont groupees it gauche de l'histogral11l11e (entre 14 et 26 u. a.) 011 l'on rencontre les pics les plus hauts. Ce profil est profondement modifie pour les noyaux du l11eme type ceUulaire mais se trouvant en ~ S + G2 (Fig. Ii). Dans ce cas, on note un etalement tres important de l'histogramme vers la droite et l'existence de nombreux pics dont les plus hauts restent localises comme pour les noyaux en Gl + ~ S dans la partie gauche de l'histogramme. Les frequences d'absorption des noyaux des ceUules mesotheliales en Gl + ~ S montrent une accumulation des valeurs dans la partie gauche de l'histogramme (fig. Ij) formant une cloche comprise entre 14 et 26 u. a.; les va leurs les plus elevees ne depassent pas 34 u. a. Si l'on compare cet histogramme avec celui des noyaux des memes ceUules en ~ S + G2 (Fig. lk) on note pour ces dernieres une grande augmentation des frequences vers la droite de l'histogral1lme, l'existence de
(C. T.) en G1 ; m C. T. en G2 • En abscisse,' absorption exprimee en unites arbitraires (u. a.), interyalles dc classe 2 u. a.; en ordonnce,' nombre moyen des points en % dans les differentes classes d'absorption obtenu sur 20 noyaux ct pour chaque type cellulaire represent" par les histogrammeso

76 V. GOTZOS, B. CAPPELLI-GOTZOS et G. CONTI
plusieurs pics nettement separes en 2 groupes et l'etalement des valeurs d'absorption qui arrivent jusqu'a 42 u. a.
Les noyaux des cellules tumorales en G1 + Yz S (Fig. 11) montrent un profil assez caracteristique, forme par 2 groupes de valeurs (a gauche et a droite de I'histogamme) separes par un pic haut, localise entre 24 et 26 u. a. L'histogramme n'est pas tres etale et les valeurs d'absorption les plus elevees ne depassent pas 48 u. a. Dans les cellules tumorales en Yz S + G2 (Fig. 1m) ce profil est profondement modifie montrant une accumulation des valeurs d'absorption dans la moitie droite de l'histogramme avec plusieurs pics assez hauts, notamment entre 28 et 38 u. a.
C) Estimation arbitraire de la chromatine condensee et de la chroma tine diffuse des noyaux en interphase Dans les noyaux interphasiques, la stricte identification, par des methodes mor
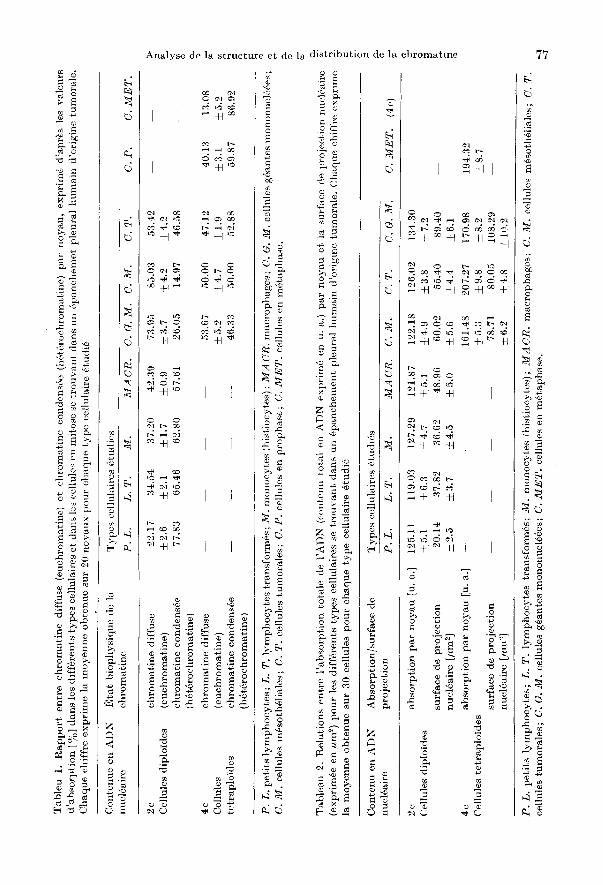
phologiques et cytochimiques, de la chromatine condensee par rapport a la chromatine diffuse dans Ie sens d'heterochromatine et d'euchromatine (respectivement: chromatine inactive et chromatine active, capable de transcription) est it, l'heure actuelle tres discutee. Ceci est dli au fait que Ie concept d'heterochromatine et d' euchromatine est piut6t fonctionnel, tandis que les termes chromatine condensee et diffuse se rMerent a des observations morphologiques qui ne correspondent pas entierement aux concepts fonctionnels. Toutefois la distinction arbitraire entre les 2 etats chromatiniens peut etre consideree valable si on se rMere aces criteres morphofonctionnels detectables par la cytophotometrie (KIEFER et al. 1973). L'etat de la chromatine des chromosomes en metaphase a ete retenu comme critere de distinction. Dans ces derniers en effet soit la morphologie (chromatine hautement spiralisee), soit la fonction (chromatine presque incapable de transcription) representent un etat chromatinien bien determine et facilement detectable. L'absorption moyenne (26 ± 1,57 u. a.) des chromosomes isoles en metaphase nous a servi comme (~valeur de base}) pour la caracterisation de la chromatine des noyaux interphasiques en condensee et diffuse: a savoir nous considerons ici toute valeur d'absorption inferieure it, cette moyenne comme etant de la chromatine diffuse et toute valeur superieure a cette moyenne comme etant de la chromatine condensee.
Comme on peut Ie constater dans la metaphase, par rapport a la prophase, Ie pourcentage de chromatine diffuse est mini me (Tableau 1); la chromatine des noyaux en metaphase est composee donc essentiellement par la forme condensee. Dans les noyaux des petits lymphocytes la chromatine diffuse reste a un pourcentage tres bas, proche de celui des cellules en metaphase et la chromatine condensee constitue plus que Ie 77 % de la chromatine nucleaire. Le pourcentage de la chromatine diffuse augmente de maniere significative au detriment de la chromatine condensee dans les noyaux deE' lymphocytes transformes par rapport aux cellules en metaphase (P < 0.01) ainsi que par rapport aux petits lymphocytes (P < 0.01). Les noyaux des lymphocytes transformes ne montrent pas de difference significative par rapport aux monocytes (P > 0.10) en ce qui concerne les pourcentages respectifs des deux etats chromatiniens; par contre on note une augmentation significative (P < 0.01) du pourcentage de la chromatine diffuse dans les noyaux des macrophages par rapport aux monoeytes et aux lymphocytes transformes. On reneontre un poureentage tres eleve de chromatine diffuse dans les noyaux des cellules geantes mononueJeees et dans les noyaux des eelIules mesotheliales (73,95% et 85,03 % respectivement). La chromatine des noyaux des cellules tumorales est partagee egalement entre les deux formes chromatiniennes (condensee 46,58 %, diffuse 53,42 %). Par contre, Ie pourcentage de chromatine conden see augmente dans les cellules Yz S + G2 par rapport it celles en G1 + Yz S pour tous les types cellulaires consideres.
'J'a
ble
u
1. R
ap
po
rt e
ntr
e e
hro
mati
ne d
iffu
se (
euch
rom
atin
o)
ot
ch
rom
ati
ne c
on
den
s60
(IH
'It{
'roe
hrom
atin
e)
par
no
yau
, ex
pri
m{
d'a
pre
s le
s v
aleu
rs
cl'a
bso
rpi i
on
[%
] Ja
ns
les
dif
fere
nts
ty
po
s ce
llu
lair
os
ot
dan
s le
d co
llu
los
en
mit
ose
se
tro
uv
an
t d
an
s u
n 6
pan
chem
ot
ple
ura
l h
um
ain
d'o
rig
ine
tum
ora
le.
Ch
aqu
e ch
iffr
e ex
pri
me l
a m
oy
en
ne o
bte
nn
e s
ur
20 n
oy
au
x p
ou
r eh
aq
ue t
yp
e e
ellu
lair
e 6
tud
i6
Con
tuIl
uO U
ll A
DN
E
tat
biop
hysi
(IU
O d
o la
T
yp
us
ccll
ulu
iru
s et
ud
il's
n
ucl
6ai
ro
uh
l'o
mat
ine
P.L
. L
. :P
. M
.
2c
chl'
om
atin
c d
iffu
se
Z:U
7
:34.
54
:n.2
0
Cel
lule
s d
iplo
ldes
(e
uch
rom
atin
e)
±2
.6
±2
.1
± 1
.7
eh
rom
ati
ne e
onde
nsec
>
77
.83
6
5.4
6
62
.80
(h
ete
roch
rom
ati
ne)
40
ch
rom
ati
ne d
iffu
se
Cel
lule
s (e
uch
rom
ati
ne)
tetr
ap
loh
les
eh
rom
ati
ne e
on
den
s6e
(hete
roch
rom
ati
ne)
MA
CR
. C
. G
.M.
C.M
.
4:2.
:19
n.9
1;
85
.03
±
0.9
±
3.7
±
4.2
5
7.6
1
26
.05
1
4.9
7
5:3.
67
50
.00
±
5.2
±
4.7
46
.:3:)
5
0.0
0
C.'
1'.
53
.42
±
4.2
4
6.5
8
47
.12
±
l.!
)
52
.88
C.P
.
40.1~
±3
.1
5fU
l7
C.M
E'1
'.
13
.08
±
5.2
86
.H2
P.
L.
peti
ts l
ym
ph
ocy
tes;
L.
T.l
ym
ph
ocy
tes
tran
sfo
rmes
; M
. m
on
ocy
tes
(his
t.io
eyte
s);
MA
CR
. m
aero
ph
ag
es;
C.
G.
M.
cell
ule
s g
{,an
tes
mon
onuP
.!{,
6es;
C
. M
. ee
llu
les
mf.
soth
elia
les;
C.
T.
cell
ule
s t.
um
ora
les;
C.
P.
cell
ule
s en
pro
ph
ase
; C
. M
ET
. ce
llul
eR e
n m
eta
ph
ase
.
Tab
leau
2.
Rela
tio
ns
en
tre I
'ab
sorp
tio
n t
ot.
ale
de
I'A
DN
(co
nt.e
nll
tota
l en
AD
N e
xp
rim
e e
n u
. a.
) p
ur
no
yau
et.
la K
llrf
ace
de
pro
j"cti
on
nu
d,;
ail'
o
(ex
pri
mee
en
,um
2)
po
ur
les
dif
fere
nts
ty
pes
eell
ula
ires
se
tro
m"a
nt
dan
s u
n 6
pan
ch
em
en
t p
leu
rall
mm
ain
d'o
rig
ine
tum
ora
le.
Ch
aqu
e ch
iffr
e o
xp
rim
e la
mo
yen
ne o
bte
nu
e s
ur
30
cel
lule
s p
ou
r eh
aq
ue t
yp
e c
ellu
lair
e etu
die
Co
nte
nu
en
AD
N
Ab
sorp
tio
n/s
urf
ace d
o
Ty
po
s ce
llu
lair
es e
tud
ies
>
::l
>< ''<
en
C!>
0.
C!>
;;"
?4- .., =
~ =
.., C!> ~
0.
c ;;"
8;
?4- .., & =
c+ o· ::l
0
. to
n
ucl
eair
e p
roje
cti
on
P
.L.
L.'
1'.
M
. M
AC
R.
C.M
. C
. '1
'. C
. C/
. M
. C
. M
ET
. (4
0)
;;"
:!c
ab
sorp
tio
n p
ar
no
yau
[u
. a.
] 12
5.11
1
19
.03
1
27
.29
C
ellu
les
dip
loid
es
±5
.1
±6
.3
±4
.7
surf
aee
de
pro
jecti
on
2
0.1
4
37
.82
3
6.6
2
nu
ele
air
e [
11m
2]
±2
.5
±3
.7
±4
.5
4c
ah
sorp
tio
n p
ar
no
yau
[u
. a.
] C
ellu
les
tetr
ap
loid
es
surf
ace d
e p
roje
eti
on
n
ucle
air
e [
11m
"]
12
1.8
7
12
2.1
8
±5
.1
±4
.!l
48.9
()
()0.
02
±5
.0
±5
.6
16
1.4
8
±5
.:;
78
.71
±
6.2
12
6.0
2
±3
.8
55
.40
±
4.4
20
7.2
7
±9
.8
80
.05
±
1.8
1:~4
.30
±7
.2
89
.40
±
6.1
17
0.9
8
±8
.2
10
8.2
9
± 1
ll.2
194.~2
±8
.7
P.
L.
peti
ts l
ym
ph
oey
tes;
L.
T.
lym
ph
ocy
tes
tran
sfo
rmes
; M
. m
on
oey
tes
(his
tio
cyte
s);
MA
CR
. m
acro
ph
ages
; C
. JJ
;I.
cell
uleR
mes
oth
61
ialc
s;
C.
'1'.
cell
ule
s tu
mo
rale
s; C
. G
. M
. ce
llu
les
gea
nte
s m
on
oll
uc!
eees
; C
. M
El'
. ee
llu
les
en
meta
ph
ase
.
" :=" .., o a ~ § -1
-1

78 v. GOTZOS, B. CAPPELLI·GOTZOS et G. COXTI
D) Relation entre l'absorption totale de l' ADN par noyau et la surface de projection nucleaire d'apres Ie type cellulaire etudie L'absorption totale moyenne par noyau des cellules se trouvant en perio::le G1 +
Y2 S (Tableau 2) montre qu'il n'existe pas de difference signicative entre les differents types cellulaires etudies; par contre la surface de projection nucleaire varie considerablement et de fayon significative d'un type cellula ire a l'autre. En effet, il existe 4 grOllpes de cellules: Ie ler des petits lymphocytes, Ie 2 erne comprenant les lymphocytes transformes et les monocytes, Ie 3eme les macrophages, les cellules mesotheliales et les cellules tumorales et Ie 4eme les cellules geantes mononucleees.
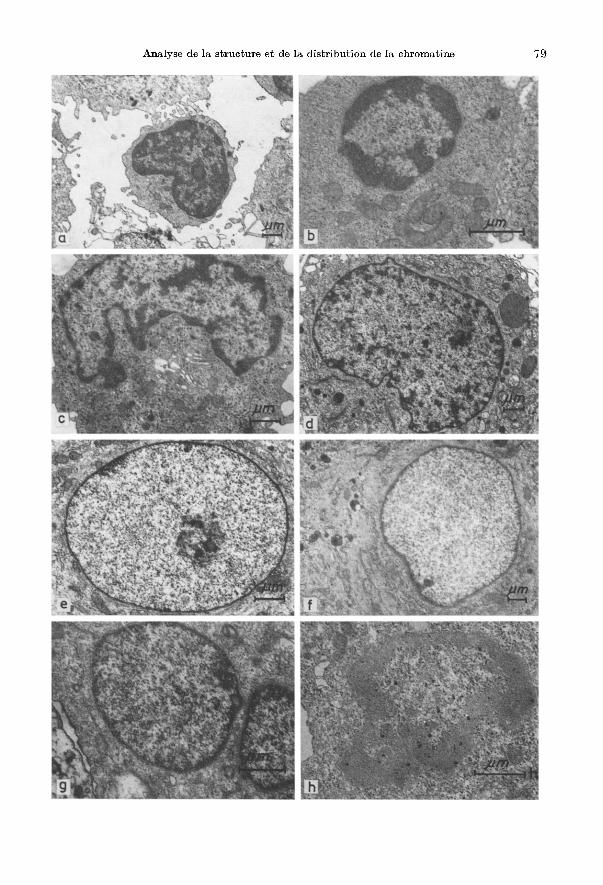
E) Observations au microscope electronique Notre attention s'est portee essentiellement sur l'aspect de la chrolllatine des
noyaux; mais nous n'avons pas neglige pour autant la morphologie particuliere generale de chaque type cellulaire pris en consideration. Au microscope electronique on distingue dans tous les noyaux en interphase des plages ou des points de chromatine dense aux electrons (on parle de chromatine dense ou heterochrOlllatine) et d' autres de chromatine moins dense (on parle alors de chromatine diffuse ou euchromatine). Les formations chromatiniennes denses disparaissent dans les noyaux des differents types cellulaires apres traitement a l'ADNase neutre (Fig. 2h) et a leur place il reste une bible coloration ne differant pas du materiel nucleaire restant. La chromatine moins dense montre aussi, apres traitement a l'ADNase, une diminution d'intensite de coloration par rapport au contr61e. La chromatine dense aux electrons (heterochromatine) semble avoir une distribution caracteristique pour les noyaux de chaque type cellulaire. Dans les petits lymphocytes (Fig. 2a) elle est distribuee surt,out a la peripherie sous forme de grandes plages se prolongeant vers l'interieur OU l'on rencontre peu de chromatine diffuse parsemee de points chromatiniens d'une densiM elevee. Cette cellule montre par ailleurs l'ultrastructure connue: un rapport nucleo-cytoplasmique eleve, des monoribosomes et polyribosomes disperses. II faut souligner que dans Ie materiel etudie les petits lymphocytes se presentent sous deux aspects differents: lymphocytes a cytoplasme dense et lymphocytes a cytoplasme clair. La distribution de la chromatine des noyaux des lymphocytes transformes (Fig. 2b) est semblable a celIe des noyaux des petits lymphocytes mais eUe apparait plus lache et on note une diminution de la surface de projection totale des plages chromatiniennes peripMriques condensees et un nombre plus eleve des points de chromatine parsemant la chromatine diffuse. II faut noter dans ces cellules l'abondance des polyribosomes et des mitochondries. Les noyaux des monocytes (Fig. 2c) presentent une couche de chromatine condensee a la peripherie nucIeaire et un grand nombre de points chromatiniens parsemant la chromatine diffuse qui occupe Ie reste du noyau. On remarquera en outre la presence de citernes du REG, de mitochondries et de granulations delimitees par une membrane (lysosomes primaires). La distribution de la chromatine condensee et diffuse au niveau des noyaux des macrophages (Fig. 2d) se
Fig. 2. Photogrammes, en microscopie electronique a transmission montrant la distribution carac· teristique de la chromatine nucleaire des differents types cellulaires se trouvant dans un epanchement pleural humain d'origine tumorale, avant et apres traitement par l' ADNase. a petit lymphocyte (x 4550); b lymphocyte transforme ( X 14500); c monocyte [(histiocyte); ( X 8360)]; d macrophage (X 4580); e cellule geante mononucleee (x 7330); j cellule mesotheliale (X 4420); " cellule tumorale en amas (X 12700); h noyau de petit lymphocyte apres traitement a I' ADNase (X 13500).
Analyse de la structure et de la distribution de la chroma tine 79

80 V. GOTZOS, B. CAPPELLI-GOTZOS et G. COXTI
rapproche de celIe que nous venons de decrire pour les monocytes. II faut neanmoins souligner qu'il existe un nombre beaucoup plus important de points denses aux electrons de la peripherie desquels partent des filaments, egalement denses, qui se perdent ensuite dans la chromatine diffuse environnante. Cette cellule est caracterisee par la presence de nombreuses digitations cytoplasmiques, de nombreux organites et du REG surtout it la peripherie et du cote oppose it la zone de GOLGI. On rencontre des vesicules de phagocytose, des lysosomes primaires et des corps residuels. La distribution de la chromatine change dans les noyaux des cellules geantes mononucleees (Fig. 2e). lei, la chromatine condensee est presque exclusivement limitee a une couche mince it la peripherie nucleaire tandis que Ie reste est occupe par des unites granulofilamenteuses d'une densite variable aux electrons. Caracteristique de la distribution de la chromatine de ces noyaux est la presence d'espaces interchromatiniens tres importants. Les caracteristiques ultrastructurales de cette cellule it relever ici sont: Ie rapport nucleo-cytoplasmique petit, Ie presence de nombreuses mitochondnes et de citernes du REG distribuees sans ordre apparent dans tout Ie cytoplasme; il existe une quantite appreciable de faisceaux de microfilaments et un appareil de GOLGI bien developpe. La chromatine des noyaux des cellules mesotheliales (Fig. 2f) se preEente sous forme d'une granulation fine, hogomene ayant u'ne distribution uniforme sur toute la surface de projection nucleaire. Dne tres mince couche de chromatine conden see est adossee au feuillet interne de l'enveloppe nucleaire. L'image cracteristique ultrastructurale du reste de la cellule mesotheliale est la presence de nombreux faisceaux de microfilaments dans la zone perinucleaire. C'est une cellule riche en mitochondries et en REG, ce dernier est plus abondant it la peripherie cellulaire. On remarque la presence de rares microvillosites cytoplasmiques. La distribution chromatinienne qu'on vient de decrire pour les noyaux des cellnles mesotheliales se retrouve aussi dans les noyaux des cellules tumorales (Fig.2g) avec la difference qu'ici on rencontre, au niveau de la chromatine diffuse, de nombreux agregats de petits granules et filaments, beaucoup plus denses aux elec:trons que Ie reste de la chromatine, distribues sans ordre apparent sur toute la surface de projection nucleaire. L'ultrastructure des cellules tumorales en amas (adeno-carcinome mammaire) revele la presence d'un grand nombre de microvillosites cytoplasmiques du cote libre de la cellule (pole apical) de nombreux dispositifs de junction cellulaire (gap junction, desmosomes), de petits faisceaux de microfilaments et la presence de grains de secretion. On trouve egalement un grand nombre de mitochondries et de citernes du REG distribuees sans ordre apparent partout dans Ie cytoplasme.
Discussion et Conclusions
La caracterisation des cellules isolees sur les frottis provenant d'epanchements d'origine tumorale pose des problemes considerables it l'examen par les methodes classiques en microscopie optique. Le probleme a He etudie par plusieurs auteurs ct aborde par differentes methodes, Boit morphologiques en microscopie optique (EFRATI et al. 1976, GOTZOS 1977) soit cytoenzymologiques (WILLIGHAGEN 1973), soit en microscopie electronique (KANESHIMA et al. 1978, Koss et al 1980). Les resultats de la cytophotometrie it balayage reportes dans ce travail, bases sur I'analyse de la structure et de la distribution de la ehromatine et completes par les images en microscopie eleetronique visent entre autre it porter des elements qui puissent contribuer au diagnostic differentiel des cellules des frottis d'epanchement humains d'origine tumorale. ])'apres les images des cartes nucleaires qui refletent l'etat hiophysique de la chromatine, on pout distinguer parmi les cellules en G1 + % S prises en consideration dans ce travail, 2 groupes: Ie ler comprend petits lymphocytes, lymphocytes transformes, monocytes, macrophages et cellules tumorales; Ie 2eme groupe comprend les cellules geantes mononucleees et les mesotheliales. Les noyaux des cellules du ler groupe apparaissent constitues surtout par de la chromatine d'une densite superieure it

Analyse de Itt structure et de Itt distribution de I" chroma tine 81
cplle des chromosomes en metaphase isoles, tandis que ceux des eellules du 2eme groupe sont presque exclusivement constitues par de la chroma tine ayant une densite ega Ie ou inferieure a ces memes chromosomes. A partir des images des cartes nucleaires il est possible de distinguer les differents types cellulaires du leI' groupe. Cela est tres facile pour les petits lymphocytes vis-a-vis de toutes les aut res cellules et aussi des lymphocytes transformes. En effet, leur chromatine presente une densite et une distribution uniforme sur touto la surface de projection nucleaire, contrail'ement aux lymphocytes transformes dans lesquels la distribution de la ehromatine est inegale et ou toutes les valeurs d'absorption sont nettement inferieures par rapport aux petits lymphocytes. Ceci montre que la totalite de la chroma tine des lymphocytes transformes, meme celie dHinie comme «chromatine dense ,) ou dleterochromatine ,) en microscopie 6lectronique, y est moins eondensee. On peut egalement distinguer les lymphocytes transformcs des monocytes gr(LCe it la presence, dans ces derniers, de chromatine dense sous forme de points que l'on ne retrouve pas dans les lymphocytes transformes ou la chroma tine dense est distribuee sous forme de plages. Les macrophages aussi sont reconnaissables gr(iCe it la presence de points de chromatine d'une densite nettement inferieure a celle des points chromatiniens des monocytes. La distinction des cellules tumorales (metastase d'un adeno-carcinome mamm'tire) des aut res cellules ne pose pas de probleme sauf pour les macrophages cn G1 + Yz S. En effet, les images des cartes nucleaires de ces 2 types cellula ires sont semblables, excepte en ce qui concerne Ie nombre de points de chromatine dense qui sont beaucoup plus nombreux et d'une absorption nettement superieura dans les noyaux des macrophages. Par contre en microscopie electronique une distinction entre les 2 types cellulaires est facilitee entre autre par la presence, dans les noyaux des macrophages, d'une large bande de chromatine dense peripherique. Les images des cartes nucleaires des cellules du 2eme groupe (rellules geantes mononucleees et cellules mesotheliales) different des aut res par la presence de chroma tine diffuse distribuee uniformement sur toute la surface de projection nucleaire. Par contre une distinction entre les noyaux de ces 2 types cellulaires uniquemcnt sur la base de la structure pt de la distribution de leur chromatine est rlifficile. Les images des cartes nucleairps des differents types cellulaires sc trouvant en Yz S + G2 ne presentent pas d'element caracteristique du type cellulaire pour permettre nne distinction dans ce sens, pour leur classification ici nous no us sommes bases sur cles caracteres inherants a la morphologie ccllulaire particuliere. En ce qui concerne la distinction de la chromatine diffuse et de la chromatine dense (eu- et hMerochromatine) les rpsultats reportes ici se rapprochent d'une maniere generale a ceux obtenus par des methodes clifferentes, biochimiques notamment (FRENSTER et al. 1963, PAYS 1975). En effet, nous trouvons une plus grande quantite de chromatine dense (heterochromatine) dans les noyaux des eellules en mitose par rapport a C(lUX des cellules en interphase ainsi que dans les noyaux des cellules en Yz S + G2 par rapport a ceux des cellules en G1 + Yz S. D11 point de yue du rapport quantitatif: hMero-cuchromatine, on peut rlistinguer les noyaux des differentes cellules en interphase et en G1 + % S en :1 groupes: 1. Noyaux dont la chromatine se trouve principalement sous forme <l'heterochromatine (petits l~'mphocytes, lymphocytes transformes, monocytes), 2. noyaux dont la chromatine se trouve principalement sous forme d'euchromatine (cellules mesotheIiales, cellules geantes mononucleees) et :1. noyaux clont la chroma tine est plus ou moins egalement partagee entre l'une et I'autre forme (macrophages et ccllules tumorales).
Pour des raisons inherentes it la technique employee ici il est difficile de compareI' I'activite transcriptionelle de la chromatine en se basant sur Ie rapport hetero-euchromattne des different·s types cellulaires en leur attribuant. une activite metabolique plus ou moins importante. Une telle interpretation s'opposerait aussi aux travaux de plusieurs auteurs (HRU 1962, GA"CTIER et al. 1976, BRUTLAG et al. 1977) qui demontrent meme pour I'heterochromatine (sauf pour celie des chromosomes sexuels) une certaine activite transcriptionelle. En tout cas il nous semble important de signaler que I'heterochromatine diminue dans les noyaux des cellules differenciees par rapport aux cellules du meme type non differenciees, comme dans les lymphocytes transformes par rapport aux petits Iympocytes, dans les macrophages par rapport aux monocytes et dans les cellulcs geantes mononucleees (macrophages actives) par rapport aux macrophages.
Nous conclurons en retenant de ce travail que la structure et la rlistribution de la chromatine etuciiees sur les images nucleaires obtenues par cytophotometrie a balayage apres roaction au FEUL
Gl<JN, sont caracteristiques pour les noyaux clu meme type cellulaire pourvu que les cellules se trouvent dans des conditions morpho-fonctionelles semblables. Des changements importants en
6 Acta histochem., Bd. 75

82 V. GOTZOS, B. CAPPELLI-GOTZOS et G. CONTI
resultent, pour la structure et la distribution de la chromatine nucleaire des cellules du meme type, lorsque les cellules se trouvent en differenciation ou dans des differentes peri odes du cycle cellulaire. Tels changements ne sont pas synchronises dans l'ensemble des cellules de la meme population de fa<,lon qu'ils ne permettent pas l'obtention d'un modde nucleaire caracteristique (image nucleaire de la chromatine) d'un type cellula ire donne. Nos resultats montrent done clairement que la cytophotometrie it balayage, basee sur les caracteristiques biophysiques de la chroma tine ne peut pas toujours Hre consideree comme un element sur de diagnostic cytologique differentiel, mais par contre elle peut contribuer it preciser davantage la nature d'un type cellulaire donne.
Bibliographie
BARNI, S., NOVELLI, G., ZANOIO, L., GERZELI, G., and VECCHIETTI, G., Chromatin analysis in human endometrial adenocarcinoma before an after treatment with G-methyl-17 -hydroxyprogesterone acetone (MPA). Virchows Arch. (Cell Pathol.) 37, 167-177 (1981).
BRUTLAG, D. L., Molecular arrangement and evolution of heterochromatic DNA. Ann. Rev. Genet. 14, 121-144 (1980).
- ApPEL", R., DENNIS, E. S., and PEACOCK, W. J., The highly repeated DNA sequences of Dro8ophila melanoga8ter. J. Mol. BioI. 112, 31-47 (1977).
CHAPMAN, J. A., GOUGH, J., and ELVES, M. W., An electron microscope study of the in vitro transformation of human leucocytes II. Transformation to macrophages. J. Cell Sci. 2, 371-180 (1967).
DEITCH, A. D., Cytophotometry of nucleic acids. In: Introduction to quantitative cytochemistry. Ed.: G. L. WIED. Academic Press, New York-London 1966, pp. 327-354.
EFRATI, P, and NIR, E., Morphological and cytochemical investigation of human mesothelial cells from pleural and peritoneal effusions. Israel J. med. sci. 7, 662-673 (1976).
FISCHER, R. A., and YATES, F., Statistical tables for biological, agricultural and medical research. Oliver & Boyd, London-Edinburgh 1949.
FRENSTER, J. H., ALLFREY, V. G., and MIRSKY, A. E., Repressed and active chromatin isolated from interphase lymphocytes. Proc. Nat. Acad. Sci. USA 50, 1026-1032 (1963).
GANTER, P., et JOLLES, G., Histochimie normale et pathologique. Vol. 1. Gauthier-Villars, Paris 1969, 356-366.
GAUTIER, F., MAYER, H., and GOEBEr" W., Cloning of calf thymus satellite DNA in 7JJ. coli. Mol. Gen. Genet. 149, 23-31 (1976).
GOTZOS, V., Culture in vitro des ceIlules de liquide peritoneal humain. Acta anat. 98, 281-294 (1977).
Hsu, T. C., Differential rate in RNA synthe.3is betwe~m euchromatin and heterochromatin. Exp. Cell Res. 27, 332-334 (1962).
JE.ANNY, J. C., et GONTCHAROFF, M., Etude en microscopie electronique et par cytophotometrie it balayage de la structure et de la distribution de la chromatine dans les noyaux des cellules cartilagineuses de Trituru8 cri8tatu8 au cours de la senescence. Bioi. Cell 32, 233-244 (1978).
KANE.SHIMA, S., KIYASU, Y., KUDO, R., KOGA, S., and TANAKA, K., An application of scanning electron microscopy to cytodiagnosis of pleural and peritoneal fluids. Acta Cytol. 22, 490-499 (1978).
KASTE.N, F. R., The chemistry of SCHIFF'S reagent. Int. Rev. Cytol. 1-56 (1960). KIEFER, R., KIEFER, G., SALM, R., ROSSNER, R., and SANDRITTER, W., A method for the quanti.
tative evaluation of eu- and heterochromatin in interphase nuclei using cytophotometry and pattern analysis. Beitr. Path. 150, 163-173 (1973).
Koss, L. G., and DOMAGALA, W., Configuration of surfaces of human cancer cells in effusions. A review. Seann. Elect. Microsc. 3, 89-100 (1980).
KRIVINKOVA, H., PONTEN, J., and BLONDAL, T., The diagnosis of cancer from body fluids. Acta path. micro bioI. scand. A 84, 455-467 (1976).
PAYS, E., Structure et activite de la chromatine in vitro. Ann. BioI. 14,501-563 (1975). PEARSE, A. G. E., Histochemistry Theoretical and Applied. Vol. 1. J. & A. Churchill, London
1968, pp. 269-264.

Analyse de la structure et de la distribution de la chroma tine 83
ROWIXSKI, J., PIEXKOWSKI, :\I., and ABRAl\ICZUK, J., Area representation of optical density of chromatin in resting and stimulated lymphocytes as measured by means of Quantimet. Histochemic 32, 75-80 (197~).
SAXDRITTER, 'V., Funktionsstrukturen des Zellkerns: Eu- und Heterochromatin. Med. Welt 21, 1-12 (1970)_ KIEFER, G_, KIEFER, R.o SALM, R., :\IoORE, G. 'V., and GRIMM, H., DNA in heterochromatin cytophotomctric pattern recognition image analysis among cell nuclei in duct epithelium and carcinoma of the human breast. Beitr. Path. 151, 87-96 (1974).
SATO, S., ITO, :\1., and NAGASE, C., Morphometric and cytological studies of giant cell carcinoma cells in the pleural fluid. Chest Disease Res. Inst. Kyoto Univ. 10, 38-43 (1978).
SA WICK I, 'V., ROWINSKI, J., and ABRAl\ICZ l.-K, J., Image analysis of chromatin in cells of preimplantation mouse embryos .. J. Cell BioI. 63, 227-233 (1974).
"-ILLIGHAGEN, R. G .. J., Enzymhistochemie in del' Diagnostik maligner Tumoren. Verh. dtsch. pathol. Ges. 57, 85-88 (19n).
Adresse: Dr. VASSILIS GOTZO>', PD, Institut d'Histologie et d'Embryologie generale, Universit& de Fribourg, Perolles, CH - 1700 Fribourg.
Explications de Planche
Planche I
Distribution des valeurs d'absorption dans les noyaux (cartes nucleaires) des cellules se trouvant dans un epanchement pleural humain d'origine tumorale, d'apres la densite optique du complexe «colorant (FEULGEN) + ADN»: 1 prophase; 2 metaphase; 3 petit lymphocyte; 4 lymphocyte transforme; 5 monocyte (histiocyte); 6 macrophage; 7 cellule geante mononucleee (C.G.M.) en G1 ; 8 C.G.M. en G2 ; 9 cellule mesotheliale (C.M.) en G1 ; 10 C.M. en G2 ; 11 cellule tumorale (C.T.) en G1 ; 12 C.T. en G2• Les differentes couleurs indiquent des classes de densite optique croissante, allant du blanc (densite optique faible) au noir [(densite optique forte); (voir chapitre: materiel et methodes)].
Acta histochem. Vol. 7.5 P lanche I
1- 3
4- 6
7-9
10 - 12
Gotzos, Cappe lli·Gotzos et Conti VEB GUSTAV FISCHER VERLAG lENA