Embed Size (px)
Citation preview

Direction Ressources Vivantes Département Ressources Halieutiques Nathalie Caill-Milly(1)
Marie-Noëlle de Casamajor(2)
Muriel Lissardy(2)
Florence Sanchez(2)
Gilles Morandeau(1)
Septembre 2003 (1) Ifremer Laboratoire Halieutique d'Aquitaine Bidart (2) Cellule d'Études et de Recherches en Environnement estuarien et Côtier en Aquitaine
(CERECA/ADERA)
Évaluation du stock de palourdes du bassin d'Arcachon – Campagne 2003
Septembre 2003

sommaire
Évaluation du stock de palourdes du bassin d’Arcachon –
Campagne 2003
Septembre 2003

sommaire
REMERCIEMENTS 4
1. INTRODUCTION 5
2. BIOLOGIE ET ECOLOGIE DES PALOURDES 7
2.1. POSITION SYSTEMATIQUE 7 2.1.1. Phylogénie 7 2.1.2. Identification spécifique 7
2.2. REPARTITION GEOGRAPHIQUE ET BATHYMETRIQUE 9 2.2.1. Aire de répartition 9 2.2.2. Habitat 9
2.3. BIOLOGIE (D'APRES QUERO ET VAYNE, 1998) 10 2.3.1. Reproduction 10 2.3.2. Alimentation 11 2.3.3. Croissance 11
2.4. PREDATION 14
3. ESTIMATION DU STOCK 2003 14
3.1. MATERIELS ET METHODES 14 3.1.1. Délimitation de l’aire de répartition de la palourde et stratification 14 3.1.2. Échantillonnage aléatoire 16 3.1.3. Nombre de prélèvements par station 16 3.1.4. Méthode de prélèvement 19 3.1.5. Traitement des échantillons à terre 21 3.1.6. Traitement des données 21
3.2. RESULTATS 22 3.2.1. Relations taille – poids frais 22 3.2.2. Estimation des abondances et des biomasses 23 3.2.3. Structure en taille de la population de palourdes japonaises 29
4. DISCUSSION 34
4.1. LA METHODE DE PRELEVEMENT 34 4.2. LA RELATION TAILLE POIDS FRAIS 35 4.3. LES ESPECES 35 4.4. LES DENSITES ET LES EFFECTIFS TOTAUX EN NOMBRE D’INDIVIDUS ET EN POIDS 36 4.5. LA FRACTION EXPLOITABLE 38 4.6. LES ANCIENNES ZONES DE RESERVE 39
5. CONCLUSION ET RECOMMANDATIONS 40
6. BIBLIOGRAPHIE 42
7. ANNEXES 45
Septembre 2003

4
Remerciements
Ils s’adressent à Patrick Frédieu et à l’équipage de la barge ostréicole «Pierren » : Olivier Pruney, Gaby Germain, Yohan Cottenceau, Christophe Rossignol et Franck pour leur accueil et pour leur implication dans la campagne de prélèvement. Leur compétence nous a permis de tenir les objectifs de campagne dans le temps imparti.
Nos remerciements vont aussi à l’ensemble des professionnels qui ont participé activement aux prélèvements de terrain, à Bénédicte Durozoi (CERECA/ADERA Bidart) et à Nicolas Lavesque (Laboratoire d’Océanographie Biologique d’Arcachon) pour les embarquements effectués.
Notre reconnaissance va également à Spyros Fifas (RH/Ifremer Brest) pour son aide dans la mise en place du protocole d’échantillonnage, à Michel Bertignac (RH/Ifremer Lorient) pour son expérience partagée sur l’évaluation de stock de palourdes, à Jérôme Huet (RH/Ifremer Brest) pour ses conseils techniques et le prêt de matériel de prélèvement et de tri, aux membres du comité de pilotage de l’étude pour les avis formulés notamment sur la zone de travail, à Patrick Prouzet (RH/Ifremer Bidart) et à Isabelle Auby (DEL/Ifremer Arcachon) pour leurs conseils sur la rédaction du rapport, à Xavier de Montaudouin (Laboratoire d’Océanographie Biologique d’Arcachon) pour l’aide apportée à cette étude, aux membres du Comité Local des Pêches Maritimes et des Elevages Marins d’Arcachon et en particulier à Céline Laffitte pour l’aide apportée à l’organisation de la campagne et pour le suivi du dossier.
Nous remercions enfin Christian Cantin, Myriam Rumebe et Isabelle Auby (DEL/Ifremer Arcachon), Yoann Cassam Chenaï (CDD/Ifremer Arcachon) et Marie-Céline Sajus (LOB Arcachon) pour leur participation à l’analyse d’une partie des échantillons collectés, François Manaud et Gilles Trut (DEL/Ifremer Arcachon) et Paul Sourcay (Furuno Anglet) pour le prêt de matériel embarqué (GPS) et Claudes Mouches (LEM/UPPA – UFR Sciences et Techniques Côte Basque) pour la mise à disposition du laboratoire pour l’analyse d’échantillons.
Cette étude a été réalisée avec le concours financier : - du Comité Local des Pêches Maritimes et des Elevages Marins du
quartier maritime d’Arcachon ; - du Conseil Général de la Gironde ; - du Conseil Régional d’Aquitaine ; - de l’Ifremer ; - de l’Union Européenne (fonds IFOP).
Septembre 2003

5
1. Introduction
Entre 1980 et 1986, des ostréiculteurs arcachonnais se sont lancés dans l’élevage de la palourde japonaise sur estran (Robert et Deltreil, 1990). Cette production a été de courte durée. La forte chute des cours1 due à la concurrence économique de l’Espagne, du Portugal, de l’Italie et de la Tunisie, l’action des prédateurs tels que le baliste (Popovsky et al., 1999), ajoutées à des mortalités importantes survenues en été et en automne 1992 affectant les 2/3 des parcs, ont conduit à l’abandon de la vénériculture à la fin des années 80.
Dans le bassin d’Arcachon, la population néonaturelle de palourde
japonaise trouve des conditions favorables à un bon développement et s’y répand rapidement. Cette espèce supplante l’espèce locale (palourde européenne) dès le début des années 1990 (Auby, 1993). Sur un secteur de 24 km², Bertignac et al. (2001) montre que 94 % des effectifs et plus de 97 % de la biomasse sont occupés par la palourde japonaise en 2000.
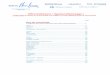
Ce développement génère une activité de pêche importante (essentiellement par des pêcheurs à pied). Les premières statistiques officielles de captures datent de 1992. Elles mettent en évidence une augmentation des apports jusqu’en 1997 (455 tonnes déclarées), puis une diminution pour s’établir autour de 300 tonnes déclarées en 2002 (figure 1). Elles ne concernent que les pêcheurs professionnels et les Conchyliculteurs Petite Pêche (CPP). Par ailleurs, les captures des ostréiculteurs sont estimées par Bobo (1996) à 25 tonnes en 1995. Pour la pêche plaisancière, nous ne disposons pas d’informations.
Par crainte de surexploitation, des mesures de gestion de la ressource sont
prises à partir de 1996. Ces mesures consistent en l’instauration de licences (55 délivrées en 2003) et la fermeture à la pêche de certaines zones (arrêtés préfectoraux de la région Aquitaine n°170 du 30 juin 2000, du 28 juin 2002 et du 13 décembre 2002).
Septembre 2003
1 De 60 F/kg à 30 F/kg en dix ans.
6
050
100150200250300350400450500
1992 1993 1994 1995 1996 1997 1998 1999 2000 2001 2002
Cap
ture
s déc
laré
es e
n to
nnes
0
10
20
30
40
50
60
70
80
Nom
bre
de li
cenc
es
Captures Licences
Figure 1.- Captures de palourdes déclarées sur le bassin d’Arcachon
(sources : CRTS La Rochelle et CLPM Arcachon)
Dans ce contexte, une première campagne d’évaluation du stock de palourdes couvrant plus d’un tiers de l’aire de répartition a été menée par l’Ifremer en 2000. Cette campagne a principalement mis en évidence :
- la prépondérance de la palourde japonaise par rapport aux autres
espèces ; - une abondance et une biomasse supérieures à celles observées sur
d’autres sites de production tels que le golfe du Morbihan et l’étang de Thau ; - une distribution en taille caractérisée par des longueurs inférieures à
celles observées sur les autres sites ; - une biomasse totale pour les 24 km² estimée à 2 900 tonnes.
Les demandes en matière de gestion (nombre de licences, taille de
commercialisation,…) demeuraient nombreuses et il est apparu nécessaire de procéder à l’évaluation complète du stock du Bassin afin d’évaluer l’impact des mesures prises antérieurement et de quantifier la biomasse totale exploitable. En complément, des travaux sur l’âge et sur l’effet densité des gisements pourront être proposés.
L’étude sur l’évaluation de la biomasse a été portée par le CLPM d’Arcachon. Les participants ont été l’Ifremer, le CLPM et l’ADERA/CERECA. Les professionnels se sont fortement associés, notamment au travers du comité de pilotage de l’étude. Ils ont également participé aux prélèvements.
Septembre 2003

7
Le présent rapport est composé de deux parties : une première rappelle les principales caractéristiques de la biologie et de l’écologie de l’espèce exploitée ; la seconde concerne l’évaluation du stock du bassin d’Arcachon. 2. Biologie et écologie des palourdes
2.1. Position systématique
2.1.1. Phylogénie
L'arbre phylogénique des palourdes se décompose de la manière suivante :
Embranchement des Mollusques
Classe des Lamellibranches Sous-classe des Eulamellibranches
Ordre des Vénéroida Famille des Vénéridés
La famille des Vénéridés regroupe à travers le monde plus de 400 espèces
vivantes actuellement. Parmi ces espèces 2 genres et 5 espèces sont exploitées en France :
- la palourde croisée japonaise, Ruditapes philippinarum (Adams &
Reeve, 1850) ; - la palourde croisée européenne, Ruditapes decussatus (Linné, 1758) ; - la palourde jaune, Venerupis aurea (Gmelin, 1791) ; - la palourde rose, Venerupis rhomboïdes (Pennant, 1777) ; - la palourde bleue ou fausse palourde, Venerupis pullastra (Montagu,
1803).
Seules les trois premières espèces ont été recensées lors de la campagne d’évaluation du stock menée en 2000 dans le bassin d’Arcachon (Bertignac et al., 2001).
2.1.2. Identification spécifique
Les palourdes présentent une variabilité morphologique et chromatique
importante. La palourde européenne porte localement le nom de "coutoye".
Septembre 2003
8
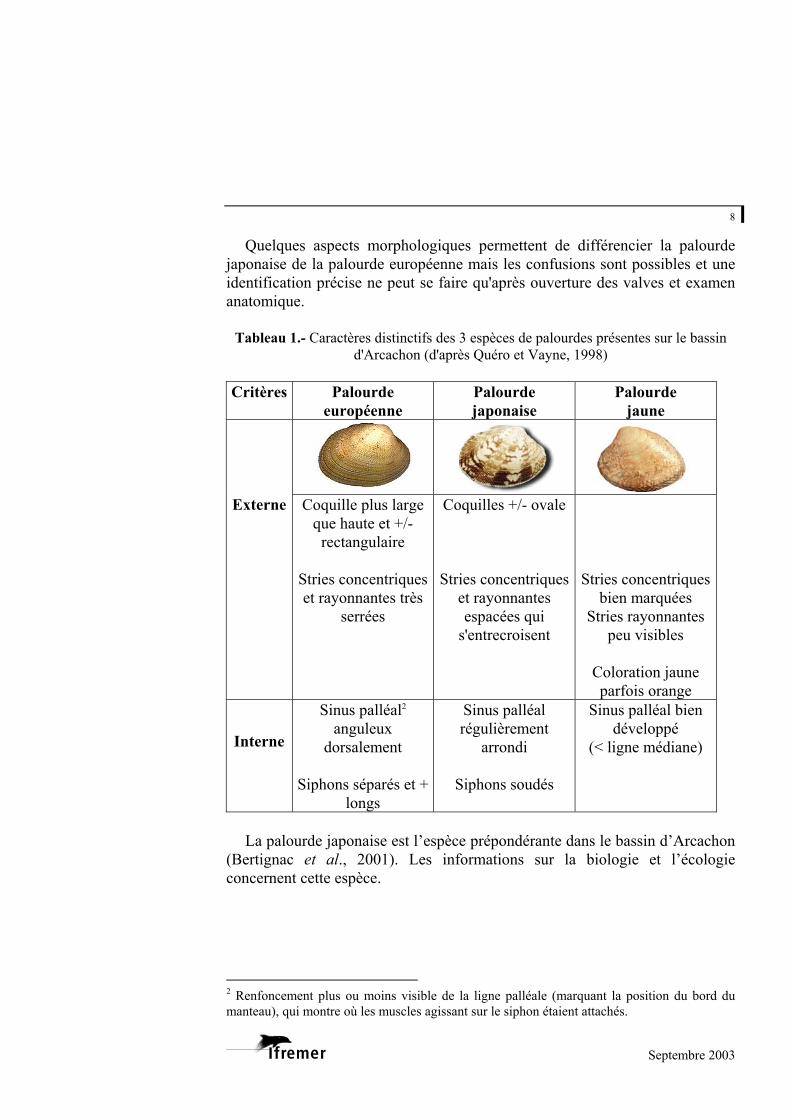
Quelques aspects morphologiques permettent de différencier la palourde japonaise de la palourde européenne mais les confusions sont possibles et une identification précise ne peut se faire qu'après ouverture des valves et examen anatomique.
Tableau 1.- Caractères distinctifs des 3 espèces de palourdes présentes sur le bassin
d'Arcachon (d'après Quéro et Vayne, 1998) Critères Palourde
européenne Palourde japonaise
Palourde jaune
Externe Coquille plus large que haute et +/-
rectangulaire Stries concentriques et rayonnantes très
serrées
Coquilles +/- ovale Stries concentriques
et rayonnantes espacées qui
s'entrecroisent
Stries concentriques bien marquées
Stries rayonnantes peu visibles
Coloration jaune parfois orange
Interne
Sinus palléal2 anguleux
dorsalement
Siphons séparés et + longs
Sinus palléal régulièrement
arrondi
Siphons soudés
Sinus palléal bien développé
(< ligne médiane)
La palourde japonaise est l’espèce prépondérante dans le bassin d’Arcachon
(Bertignac et al., 2001). Les informations sur la biologie et l’écologie concernent cette espèce.
Septembre 2003
2 Renfoncement plus ou moins visible de la ligne palléale (marquant la position du bord du manteau), qui montre où les muscles agissant sur le siphon étaient attachés.

9
2.2. Répartition géographique et bathymétrique
2.2.1. Aire de répartition
La palourde japonaise est originaire de la province Indo-Pacifique. Elle fait
l’objet d’une pêche importante au Japon notamment, et plus globalement entre les latitudes 25°N et 45°N. Son aire de répartition en Europe est plus limitée aux côtes atlantiques françaises et au nord de la mer Adriatique. Elle a été introduite en Méditerranée dans les années 1980 pour des expérimentations en aquaculture notamment dans la lagune de Venise. Elle a été acclimatée sur les côtes françaises à partir de 1972 pour l’aquaculture. Sur le Bassin d’Arcachon, le gisement naturel de palourde japonaise apparaît dans les années 1980 suite à des essais de vénériculture. Cette espèce a proliféré en dehors des zones d'élevage en raison des conditions écologiques favorables. Elle a, occasionnellement, limité ou remplacé les populations natives de palourde européenne.
2.2.2. Habitat
La palourde japonaise vit aussi bien sur l’estran (à de faibles profondeurs,
généralement de 2,5 à 1 m au-dessus du zéro des cartes) que dans l’étage infratidal. Elle est cependant signalée jusqu'à 10 m. Elle affectionne des milieux abrités et plus particulièrement ceux à salinité variable comme les baies, les lagunes littorales et les « claires ».
On la trouve sur de nombreux types de substrats meubles : vases, sables et graviers, ainsi que ceux à granulométrie mixte. La palourde possède de fortes facultés d'enfouissement dans ces différents types de sédiments, grâce à son pied très développé. La profondeur d'enfouissement est proportionnelle à sa taille et s’élève en moyenne à 7 cm. Elle peut atteindre 10-12 cm lors de très faibles températures hivernales (Le Treut, 1986). L’enfouissement des palourdes est un phénomène rapide. Ainsi en milieu naturel, Maître-Allain (1992) constate qu’une heure après le semis, 90 % des palourdes japonaises sont enfouies. La répartition des palourdes dans le sédiment n'est pas homogène, elle est distribuée en "taches".
Concernant les déplacements latéraux, ils sont qualifiés de très faibles par Le Treut (1986), bien que Tamura (1970) cite des déplacements actifs atteignant expérimentalement 6 mètres par mois.
Septembre 2003

10
2.3. Biologie (d'après Quéro et Vayne, 1998)
1ère maturité sexuelle2 ans
NaissainJuvénile
Émission des gamètesFécondation
Larve véligère
Larve pédivéligère
Métamorphose Fixation au substrat
Figure 2.- Représentation simplifiée du cycle biologique de la palourde
2.3.1. Reproduction
La palourde est gonochorique3, les sexes sont séparés. La maturité apparaît à
partir d'une taille d'une vingtaine de millimètres soit lors de la 2ème année. Le cycle reproducteur présente une forte saisonnalité :
- repos hivernal ; - gamétogenèse printanière et estivale ; - pontes estivales et automnales.
Les facteurs environnementaux jouent un rôle important dans le
déroulement de la reproduction. La vitellogenèse printanière coïncide avec la phase d'allongement de la durée d'éclairement postérieure à l'équinoxe, avec la phase d'augmentation de la température de l'eau ainsi qu'avec le développement des floraisons planctoniques (Laruelle, 1999).
L'apport trophique est primordial dans le bon déroulement de la gamétogenèse. Suivant ces conditions printanière et estivale, elle effectue deux pontes ou une seule ponte estivale. Seuls les plus gros individus (> à 35mm) réalisent ces deux pontes.
Septembre 2003
3 Les gamètes mâles et femelles sont produits par des individus distincts.

11
La reproduction de la palourde japonaise apparaît plus précoce dans la saison que celle de la palourde européenne. Le taux d'occupation gamètique4 est également supérieur : 55% contre 40% chez R. decussatus (Laruelle, 1999).
Les gamètes sont libérés dans le milieu où se produit la fécondation. L'incubation des œufs est rapide et varie en fonction de la température de l'eau (de l'ordre de 24h à 23°c). Le développement larvaire est estimé entre 12 et 15 jours. Les palourdes ont ensuite un mode de vie benthique jusqu'à la fin de leur cycle.
Sur le bassin d'Arcachon, les travaux de Robert et al. (1993) font apparaître une seule période de ponte au lieu de deux à l'automne localement. Compte tenu de la variabilité inter-annuelle des conditions environnementales, de plus amples investigations devraient être réalisées pour valider ces résultats sur une plus longue période.
2.3.2. Alimentation
La palourde se nourrit principalement de phytoplancton et de matières
organiques particulaires. Ces éléments vitaux sont amenés par le courant d’eau créé par le siphon inhalant de la palourde à la surface du sédiment. L'eau ainsi filtrée passe à travers les branchies qui retiennent à la fois l'oxygène dissous dans l'eau et les substances nutritives. Le siphon exhalant assure la fonction d'excrétion.
2.3.3. Croissance
La croissance est fortement ralentie pendant la saison froide. Elle est
maximale au printemps et à l’automne. De fortes variations entre les années sont observées, elles sont liées à la variabilité des conditions climatiques. La croissance est également très variable en fonction de la saison, du site, de la densité et de la nourriture disponible.
Les deux principaux facteurs qui semblent conditionner la croissance de la palourde japonaise sont :
- la température de l'eau, elle agit positivement sur la croissance jusqu'à
une vingtaine de degrés ; - les disponibilités trophiques, la plus forte croissance correspond aux
blooms phytoplanctoniques du printemps et de l'automne (Maître-Allain, 1983).
Septembre 2003
4 Indice sur l’état de maturité des organes reproducteurs.
12
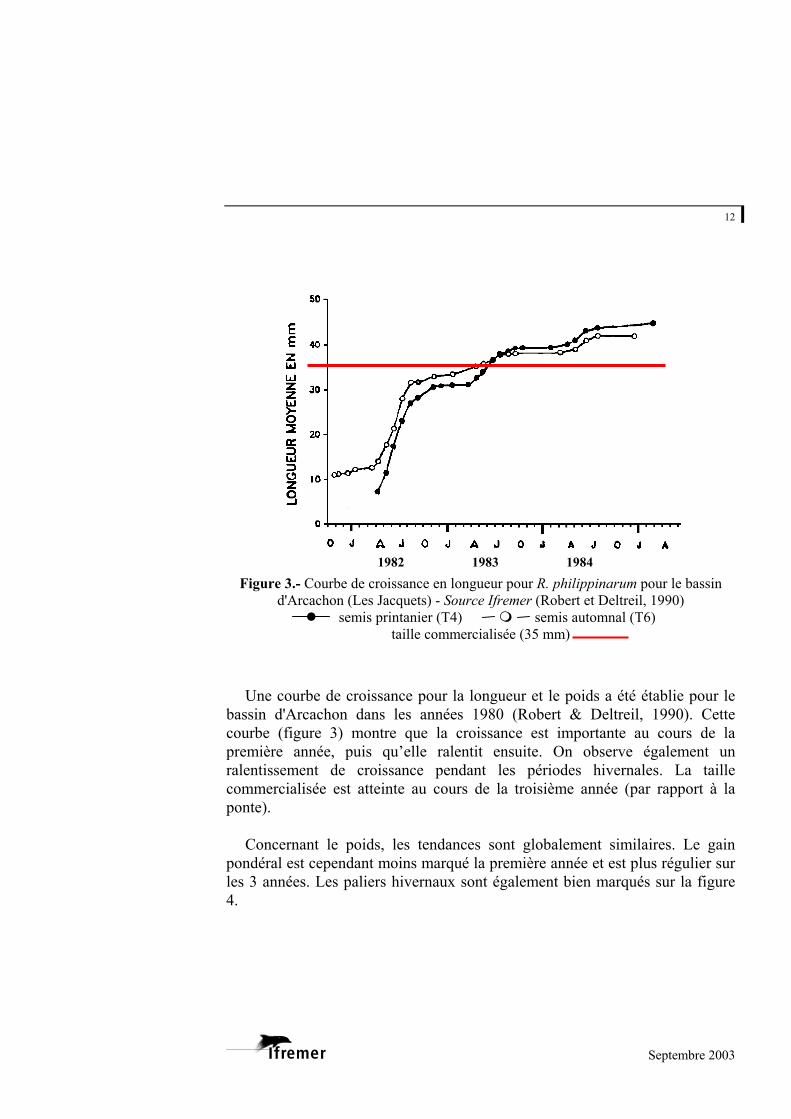
Figure 3.- Courbe de d'Arcachon (Les
semis
Une courbe de croibassin d'Arcachon dacourbe (figure 3) mopremière année, puisralentissement de crcommercialisée est atponte).
Concernant le poid
pondéral est cependanles 3 années. Les palie4.
1982 1983 1984
croissance en longueur pour R. philippinarum pour le bassin Jacquets) - Source Ifremer (Robert et Deltreil, 1990) printanier (T4) semis automnal (T6)
taille commercialisée (35 mm)
ssance pour la longueur et le poids a été établie pour le ns les années 1980 (Robert & Deltreil, 1990). Cette ntre que la croissance est importante au cours de la qu’elle ralentit ensuite. On observe également un oissance pendant les périodes hivernales. La taille teinte au cours de la troisième année (par rapport à la
s, les tendances sont globalement similaires. Le gain t moins marqué la première année et est plus régulier sur rs hivernaux sont également bien marqués sur la figure
Septembre 2003
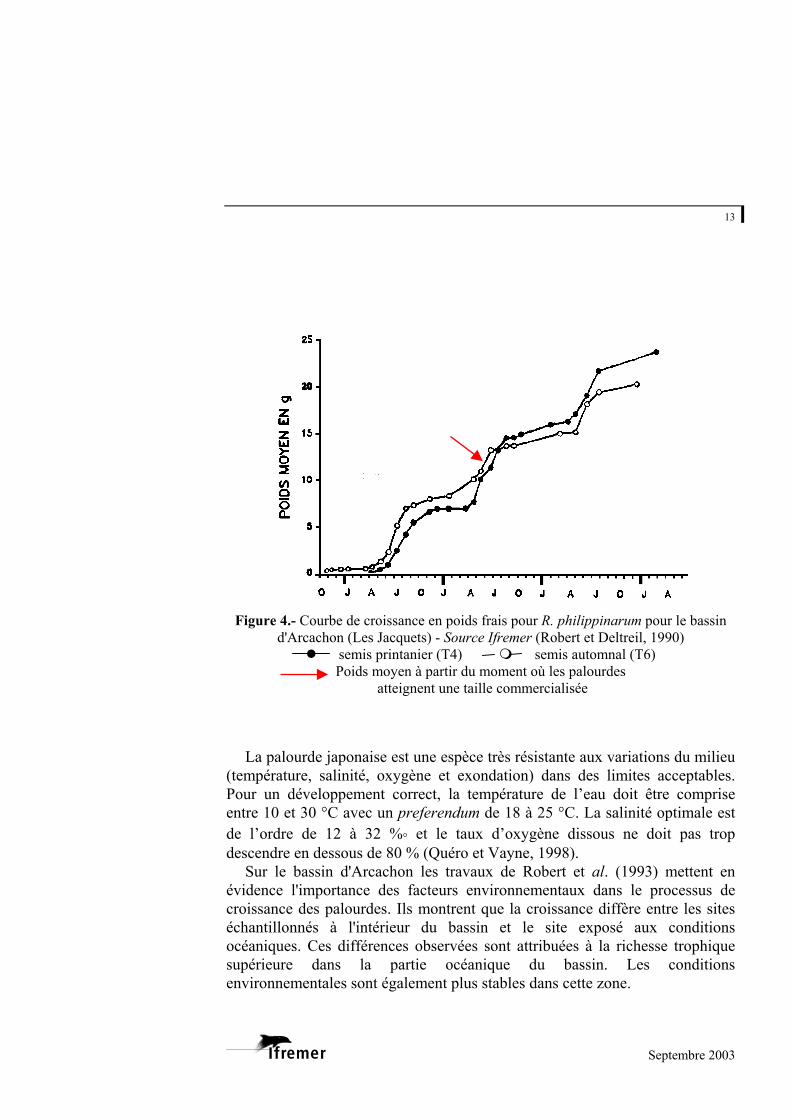
13
Figure 4.- Courbe de croissance en poids frais pour R. philippinarum pour le bassin
d'Arcachon (Les Jacquets) - Source Ifremer (Robert et Deltreil, 1990) semis printanier (T4) semis automnal (T6)
Poids moyen à partir du moment où les palourdes atteignent une taille commercialisée
La palourde japonaise est une espèce très résistante aux variations du milieu
(température, salinité, oxygène et exondation) dans des limites acceptables. Pour un développement correct, la température de l’eau doit être comprise entre 10 et 30 °C avec un preferendum de 18 à 25 °C. La salinité optimale est de l’ordre de 12 à 32 %° et le taux d’oxygène dissous ne doit pas trop descendre en dessous de 80 % (Quéro et Vayne, 1998).
Sur le bassin d'Arcachon les travaux de Robert et al. (1993) mettent en évidence l'importance des facteurs environnementaux dans le processus de croissance des palourdes. Ils montrent que la croissance diffère entre les sites échantillonnés à l'intérieur du bassin et le site exposé aux conditions océaniques. Ces différences observées sont attribuées à la richesse trophique supérieure dans la partie océanique du bassin. Les conditions environnementales sont également plus stables dans cette zone.
Septembre 2003

14
2.4. Prédation
Les palourdes ont de nombreux prédateurs parmi les espèces littorales :
- les oiseaux, avec plus particulièrement des laridés (goéland et
mouette) et un limicole (huîtrier-pie) ; - les poissons parmi lesquels la plie, la daurade royale et le baliste ; - les invertébrés marins : les crustacés comme le crabe vert dont la
prédation est maximale de mai à septembre, la nuit et à marée haute. Les palourdes les plus petites sont particulièrement vulnérables. Les étoiles de mer sont également des consommateurs de palourdes. 3. Estimation du stock 2003
3.1. Matériels et méthodes
Le protocole mis en œuvre reprend celui appliqué en 2000 par Bertignac et
al. (2001) sur le bassin d’Arcachon et par Berthou et al. (1997) pour le golfe du Morbihan. Considérant les résultats obtenus en 2000 et les contacts avec l’Ifremer de Lorient et de Brest, des modifications légères ont été apportées. Cette méthode reste une méthode usuelle appliquée pour l’estimation de stocks de bivalves.
3.1.1. Délimitation de l’aire de répartition de la palourde et stratification
Sur le Bassin, l’aire de répartition de la palourde se situe essentiellement à
l’est de l’axe La Hume/Claouey et couvre environ 93 km². De cet aire, ont été soustraits les zones ne pouvant être échantillonnées (constituées par les herbus5 et par les parcs ostréicoles) et les chenaux principaux. L’étendue ainsi obtenue a été découpée en strates. Définies comme des ensembles spatiaux homogènes, elles intègrent la réglementation en vigueur et les conditions sédimentologiques et bathymétriques de la zone.
Ce découpage en strates a ensuite été présenté au comité de pilotage constitué de professionnels. Leurs connaissances du milieu ont permis de modifier (ils ont notamment fait ajouter une zone autour de l’Ile aux Oiseaux –
Septembre 2003
5 Prés salés.

15
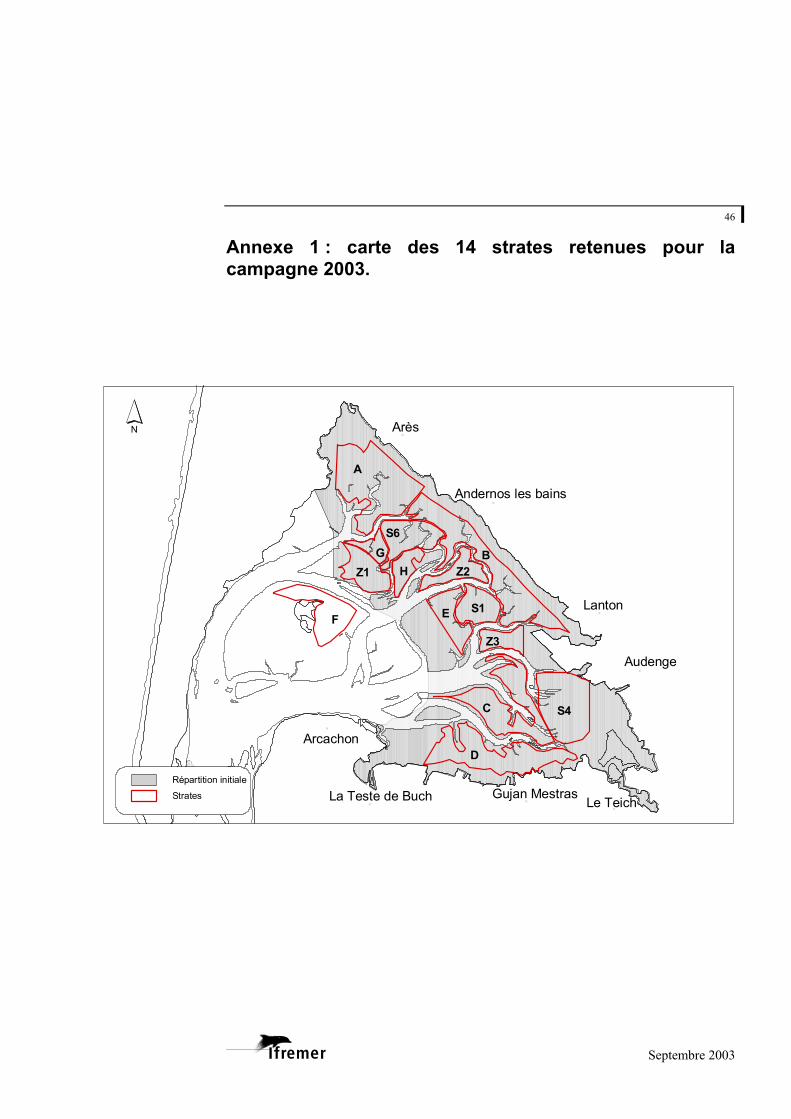
strate F) et de valider une stratification définitive de la zone d’étude. Au total, ce sont 14 strates pour 46,4 km² qui ont été retenues (figure 5).
N
N
N
N
N
NN
N
Arès
Lanton
Audenge
Le Teich
Arcachon
Gujan MestrasLa Teste de Buch
Andernos les bains
B
A
D
C
F
S4
E
S6
Z1
S1
H Z2
Z3
G
N
Répartition initiale
Strates
Figure 5.- Carte des 14 strates retenues pour la campagne 2003 NB : cette carte figure également à l’annexe 1 et peut être dépliée afin de faciliter la lecture des résultats par strate.
Le tableau 2 précise les principales caractéristiques des 14 strates échantillonnées.
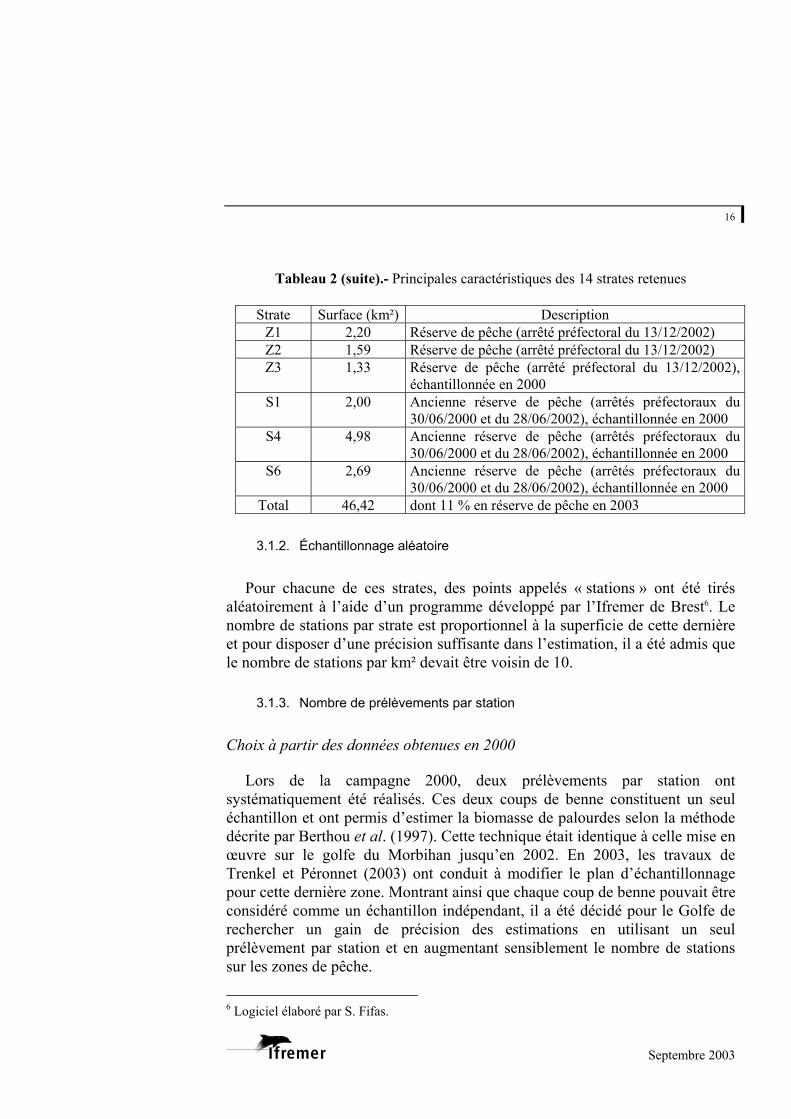
Tableau 2.- Principales caractéristiques des 14 strates retenues
Strate Surface (km²) Description A 7,18 Secteur de pêche B 6,89 Secteur de pêche C 5,10 Secteur de pêche échantillonné pour partie en 2000 D 5,38 Secteur de pêche E 1,96 Secteur de pêche échantillonné en 2000 F 3,08 Secteur de pêche G 0,87 Secteur de pêche H 1,17 Secteur de pêche
Septembre 2003
16
Tableau 2 (suite).- Principales caractéristiques des 14 strates retenues
Strate Surface (km²) Description Z1 2,20 Réserve de pêche (arrêté préfectoral du 13/12/2002) Z2 1,59 Réserve de pêche (arrêté préfectoral du 13/12/2002) Z3 1,33 Réserve de pêche (arrêté préfectoral du 13/12/2002),
échantillonnée en 2000 S1 2,00 Ancienne réserve de pêche (arrêtés préfectoraux du
30/06/2000 et du 28/06/2002), échantillonnée en 2000 S4 4,98 Ancienne réserve de pêche (arrêtés préfectoraux du
30/06/2000 et du 28/06/2002), échantillonnée en 2000 S6 2,69 Ancienne réserve de pêche (arrêtés préfectoraux du
30/06/2000 et du 28/06/2002), échantillonnée en 2000 Total 46,42 dont 11 % en réserve de pêche en 2003
3.1.2. Échantillonnage aléatoire
Pour chacune de ces strates, des points appelés « stations » ont été tirés
aléatoirement à l’aide d’un programme développé par l’Ifremer de Brest6. Le nombre de stations par strate est proportionnel à la superficie de cette dernière et pour disposer d’une précision suffisante dans l’estimation, il a été admis que le nombre de stations par km² devait être voisin de 10.
3.1.3. Nombre de prélèvements par station
Choix à partir des données obtenues en 2000
Lors de la campagne 2000, deux prélèvements par station ont systématiquement été réalisés. Ces deux coups de benne constituent un seul échantillon et ont permis d’estimer la biomasse de palourdes selon la méthode décrite par Berthou et al. (1997). Cette technique était identique à celle mise en œuvre sur le golfe du Morbihan jusqu’en 2002. En 2003, les travaux de Trenkel et Péronnet (2003) ont conduit à modifier le plan d’échantillonnage pour cette dernière zone. Montrant ainsi que chaque coup de benne pouvait être considéré comme un échantillon indépendant, il a été décidé pour le Golfe de rechercher un gain de précision des estimations en utilisant un seul prélèvement par station et en augmentant sensiblement le nombre de stations sur les zones de pêche.
Septembre 2003
6 Logiciel élaboré par S. Fifas.
17
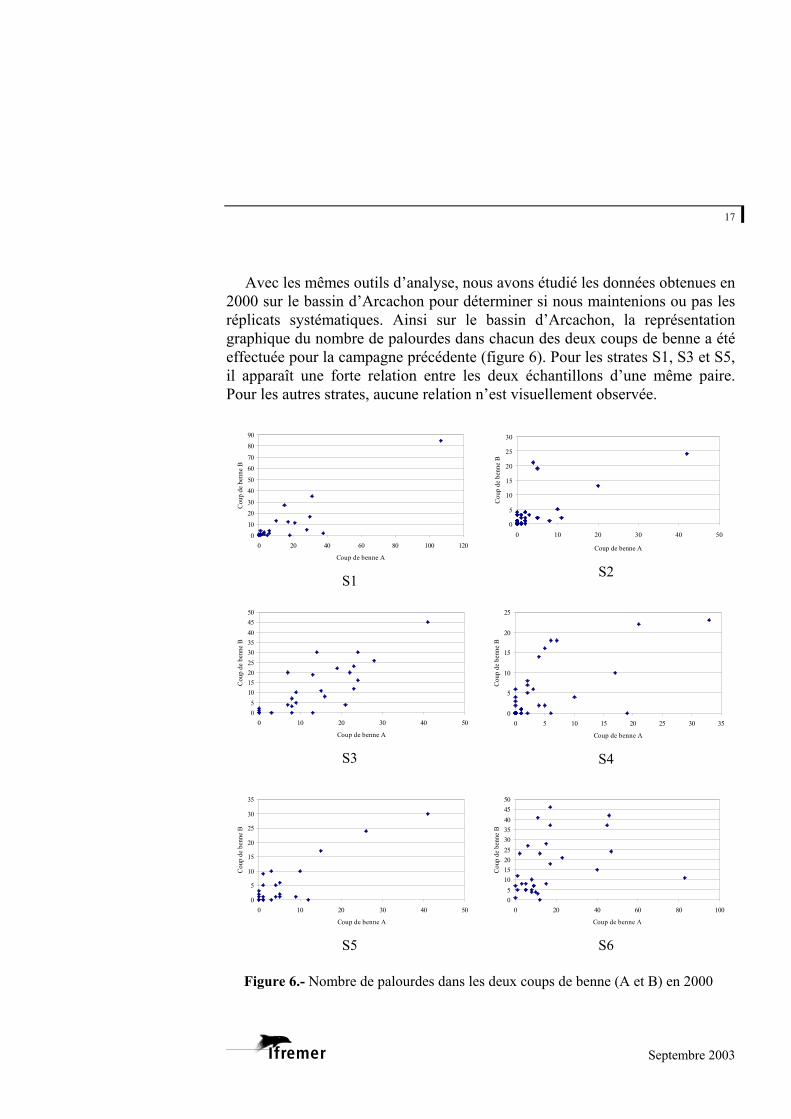
Avec les mêmes outils d’analyse, nous avons étudié les données obtenues en
2000 sur le bassin d’Arcachon pour déterminer si nous maintenions ou pas les réplicats systématiques. Ainsi sur le bassin d’Arcachon, la représentation graphique du nombre de palourdes dans chacun des deux coups de benne a été effectuée pour la campagne précédente (figure 6). Pour les strates S1, S3 et S5, il apparaît une forte relation entre les deux échantillons d’une même paire. Pour les autres strates, aucune relation n’est visuellement observée.
010
2030
4050
6070
8090
0 20 40 60 80 100 120
Coup de benne A
Cou
p de
ben
ne B
S1
0
5
10
15
20
25
30
0 10 20 30 40 5
Coup de benne A
Cou
p de
ben
ne B
0
S2
05
101520253035404550
0 10 20 30 40
Coup de benne A
Cou
p de
ben
ne B
50
S3
0
5
10
15
20
25
0 5 10 15 20 25 30 35
Coup de benne A
Cou
p de
ben
ne B
S4
0
5
10
15
20
25
30
35
0 10 20 30 40 5
Coup de benne A
Cou
p de
ben
ne B
0
S5
05
101520253035404550
0 20 40 60 80 100
Coup de benne A
Cou
p de
ben
ne B
S6
Figure 6.- Nombre de palourdes dans les deux coups de benne (A et B) en 2000
Septembre 2003

18
L’application du test de corrélation de rang de Spearman, avec hypothèse alternative d’une corrélation positive (Scherrer, 1984), précise ces résultats. Elle montre que pour toutes les strates sauf la strate S2, la relation entre le nombre de palourdes obtenues lors des deux coups de benne est hautement significative (tableau 3). Tableau 3.- Résultats de l’application du Test de corrélation de Spearman aux couples
d’échantillons obtenus en 2000 (seuil de signification 1 %)
Strate Nombre de paires Zrs rsα/2 zα/2 Résultat S1 23 3,001 0,531 H0 rejeté S2 45 2,477 2,575 H0 accepté S3 26 -11,070 0,500 H0 rejeté S4 53 2,920 2,575 H0 rejeté S5 23 1,527 0,500 H0 rejeté S6 31 2,864 2,575 H0 rejeté
Dans ces conditions, il est apparu préférable de maintenir la mise en œuvre
de réplicats pour les strates échantillonnées en 2000 sauf pour la strate S2 qui peut faire l’objet d’un prélèvement simple. A fortiori, l’application de réplicats est à privilégier pour les zones nouvellement prospectées en 2003 puisque nous ne disposons pas d’information sur ces dernières.
Choix à partir des conditions de campagne rencontrées en 2003
Dans la pratique, nous avons réduit le nombre de prélèvements d’autres
strates et ce pour deux raisons. Il s’agit d’une part des modifications apportées par le comité de pilotage sur certaines strates (superficie ou suppression proposée) et d’autre part de la perte de temps provenant de conditions techniques rencontrées lors de l’affrètement d’un premier navire et des conditions météorologiques sur zone par la suite.
Pour faire ce choix, nous avons pris en considération :
- les variabilités intra et inter-stations issues de la campagne 2000. Les variabilités intra-stations étant les plus fortes pour les strates situées le plus au Nord, il a été décidé de conserver l’application des réplicats préférentiellement pour les zones localisées dans la partie Nord du Bassin ;
- les indications fournies par les professionnels sur la pertinence de certaines strates (peu d’intérêt des strates B et D).
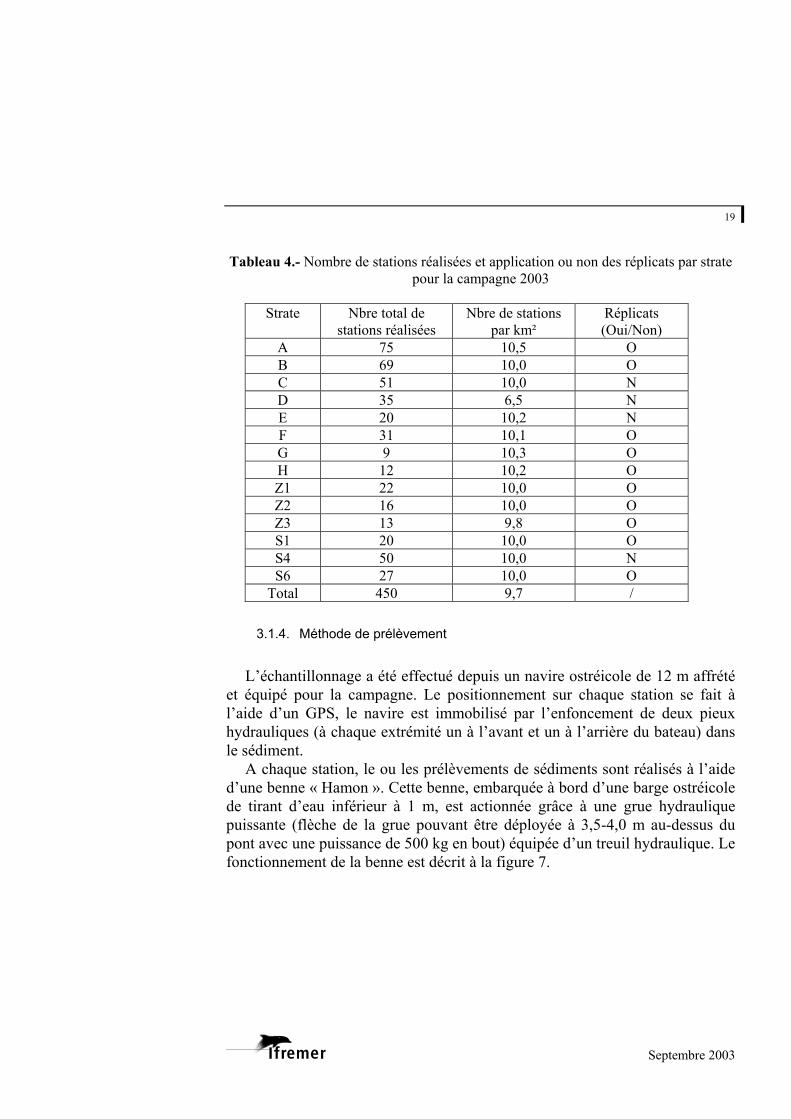
Au final, le tableau 4 présente le nombre de stations réalisées et l’application ou non des réplicats par strate pour la campagne 2003.
Septembre 2003
19
Tableau 4.- Nombre de stations réalisées et application ou non des réplicats par strate
pour la campagne 2003
Strate Nbre total de stations réalisées
Nbre de stations par km²
Réplicats (Oui/Non)
A 75 10,5 O B 69 10,0 O C 51 10,0 N D 35 6,5 N E 20 10,2 N F 31 10,1 O G 9 10,3 O H 12 10,2 O Z1 22 10,0 O Z2 16 10,0 O Z3 13 9,8 O S1 20 10,0 O S4 50 10,0 N S6 27 10,0 O
Total 450 9,7 /
3.1.4. Méthode de prélèvement
L’échantillonnage a été effectué depuis un navire ostréicole de 12 m affrété
et équipé pour la campagne. Le positionnement sur chaque station se fait à l’aide d’un GPS, le navire est immobilisé par l’enfoncement de deux pieux hydrauliques (à chaque extrémité un à l’avant et un à l’arrière du bateau) dans le sédiment.
A chaque station, le ou les prélèvements de sédiments sont réalisés à l’aide d’une benne « Hamon ». Cette benne, embarquée à bord d’une barge ostréicole de tirant d’eau inférieur à 1 m, est actionnée grâce à une grue hydraulique puissante (flèche de la grue pouvant être déployée à 3,5-4,0 m au-dessus du pont avec une puissance de 500 kg en bout) équipée d’un treuil hydraulique. Le fonctionnement de la benne est décrit à la figure 7.
Septembre 2003

20
Figure 7.- Plan de la benne Hamon utilisée pour les prélèvements
Pour accéder à toutes les stations, les prélèvements ont été réalisés à marée
haute pendant les périodes de vives eaux, du 10 au 18 juin 2003 et du 1er au 4 juillet 2003. Les échantillons ont été tamisés sur une table de tri composée de trois tamis de maille carrée de 2 cm, 1 cm et 5 mm, puis conservés dans du formol à 4 %. L’annexe 2 présente quelques étapes de la campagne de prélèvement.
NB : Dès les premiers jours de campagne et pour certaines stations, nous avons observé l’existence d’un bourrelet de vase (d’aspect « crémeux ») au dessus du godet lors du virage. Ce dernier, soit retombait dans l’eau lors de la sortie de la benne, soit était retiré manuellement avant le basculement du godet dans les bacs de prélèvement. Ce phénomène n’ayant pas été décrit pour les autres zones où cette même méthode de prélèvement est pratiquée, nous avons conservé ces « vases supérieures » toutes les cinq stations environ pour discuter de leur incidence éventuelle sur l’estimation du stock.
Septembre 2003

21
3.1.5. Traitement des échantillons à terre
Les échantillons ont été traités en laboratoire. Les espèces ont été déterminées (cf. § 2- Biologie et écologie), la taille [longueur suivant le plus grand axe mesurée au pied à coulisse électronique (+/- 0,01 mm)] et le poids total frais [mesuré à l’aide d’une balance électronique (+/- 0,01 g)] ont été relevés individuellement.
3.1.6. Traitement des données
Relation taille/poids
Pour établir la relation taille et poids frais de la palourde japonaise 4 391 individus récoltés pendant la campagne 2003 ont été pris en compte sur l'ensemble du bassin d'Arcachon. Le nombre de palourdes étudiés varie fortement d'une strate à l'autre (tableau 5).
Tableau 5.- Répartition des palourdes prises en compte dans la relation taille -
poids frais (N : nombre d'individus)
A
B
C
D
E
F
G
H
S1
S4
S6
Z1
Z2
Z3
N
534
982
286
53
227
140
139
218
353
208
571
210
252
218
Afin d'éliminer au mieux la variabilité liée à des problèmes d'échantillonnage un certain nombre d'individus récoltés ont été supprimés :
- dans l'hypothèse où il y aurait une stratification verticale dans le sédiment de la structure en taille, les palourdes issues des points de prélèvement des vases supérieures, au-dessus de la benne et noté "S" n'ont pas été prises en compte ;
- les palourdes cassées ou ouvertes lors de la prise de mesures, pour éliminer les variations de poids ;
- 7 individus sur l'ensemble des strates ont été éliminés car ils présentaient des valeurs aberrantes, probablement liées à des erreurs de mesures.
Évaluation de la biomasse
Pour les strates avec deux prélèvements par station, les calculs de moyennes
et totaux par strate, ainsi que leurs variances associées ont été réalisés selon la
Septembre 2003
22
méthode décrite par Cochran (1997). Le détail des calculs, présenté par Berthou et al. (1997), figure à l’annexe 3.
Pour les strates avec un prélèvement par station, les calculs sont plus simples. Leur détail est également fourni à l’annexe 3.
3.2. Résultats
3.2.1. Relations taille – poids frais
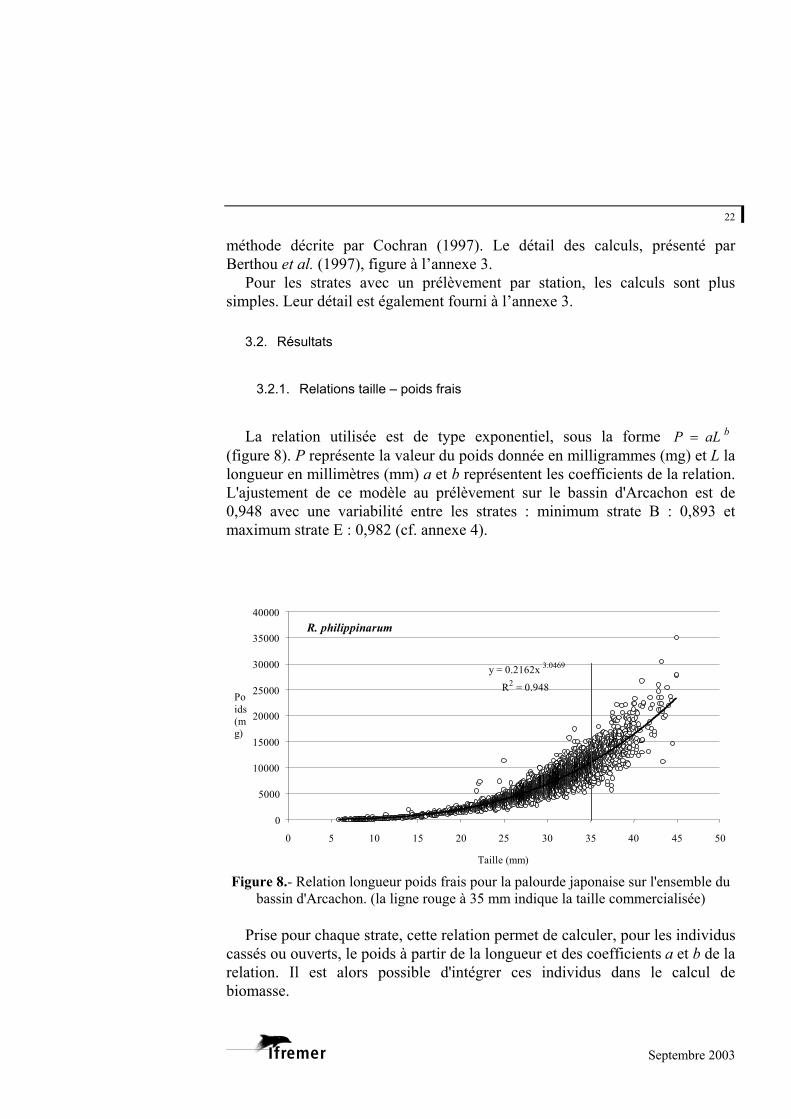
La relation utilisée est de type exponentiel, sous la forme
(figure 8). P représente la valeur du poids donnée en milligrammes (mg) et L la longueur en millimètres (mm) a et b représentent les coefficients de la relation. L'ajustement de ce modèle au prélèvement sur le bassin d'Arcachon est de 0,948 avec une variabilité entre les strates : minimum strate B : 0,893 et maximum strate E : 0,982 (cf. annexe 4).
baLP =
R. philippinarum
y = 0.2162x 3.0469
R2 = 0.948
0
5000
10000
15000
20000
25000
30000
35000
40000
0 5 10 15 20 25 30 35 40 45 50
Taille (mm)
Poids(mg)
Figure 8.- Relation longueur poids frais pour la palourde japonaise sur l'ensemble du bassin d'Arcachon. (la ligne rouge à 35 mm indique la taille commercialisée)
Prise pour chaque strate, cette relation permet de calculer, pour les individus
cassés ou ouverts, le poids à partir de la longueur et des coefficients a et b de la relation. Il est alors possible d'intégrer ces individus dans le calcul de biomasse.
Septembre 2003

23
La palourde jaune et la palourde européenne présentent un faible nombre d'individus sur l'ensemble des strates, mais font également l'objet d'une relation taille - poids frais donnée à titre indicatif en annexe 5.
3.2.2. Estimation des abondances et des biomasses
Les résultats sont détaillés à l’annexe 6
Pour l’ensemble de la zone échantillonnée
L’abondance moyenne, toutes espèces confondues, est de 6,08 palourdes par
unité d’échantillonnage (0,25 m²), soit 24,32 individus/m². La biomasse moyenne en poids frais est de 43,59 g/0,25 m², soit 174g/m².
Pour l’ensemble de la zone échantillonnée, l’abondance totale est estimée à 1 129 (+/- 167) millions d’individus pour une biomasse totale de 8 095 (+/- 1 046) tonnes.
Les trois espèces habituellement répertoriées sur le bassin ont été récoltées
lors de la campagne, à savoir la palourde européenne (Ruditapes decussatus), la palourde japonaise (Ruditapes philippinarum) et la palourde jaune (Venerupis aurea). La palourde japonaise domine très largement puisqu’elle représente 98 % des effectifs et 99 % de la biomasse. Les biomasses de palourde européenne et de palourde jaune sont ainsi estimées à seulement 63 et 39 tonnes pour l’ensemble de la zone d’étude.
Par strate
Comme en 2000, les abondances et les biomasses varient fortement selon
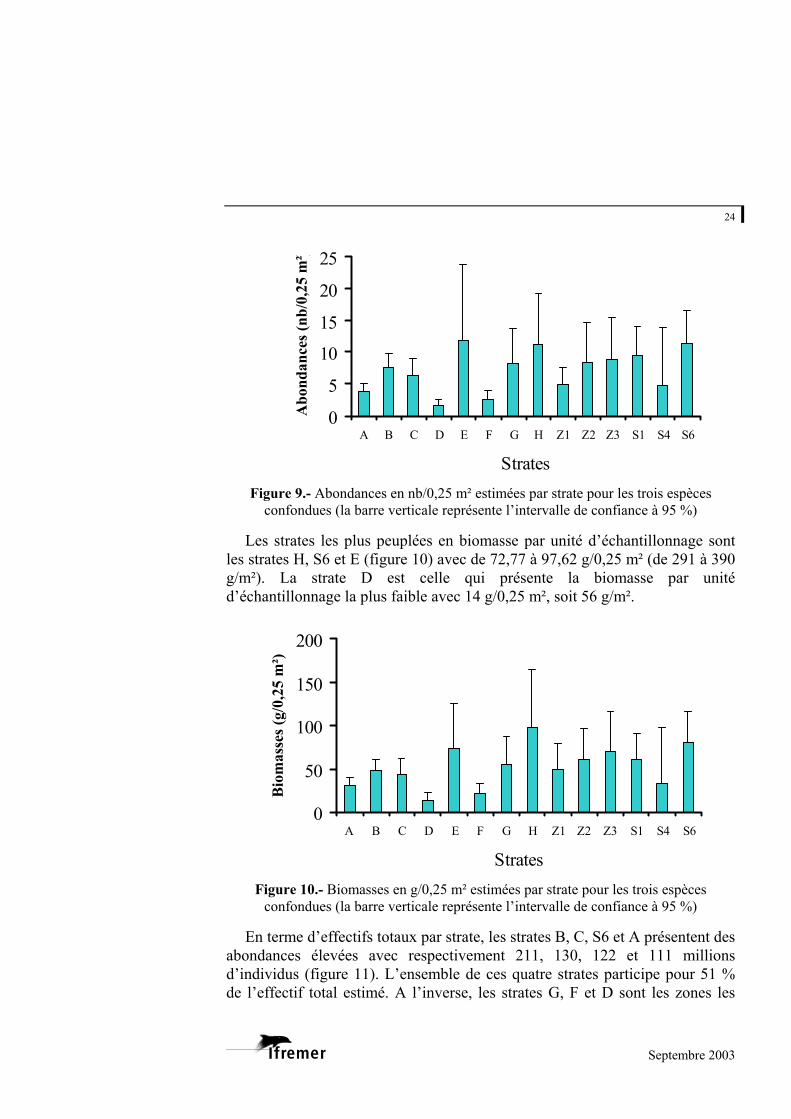
les strates. Les strates les plus densément peuplées par unité d’échantillonnage sont les
strates E, S6 et H (figure 9) avec de 11,23 à 11,95 palourdes/0,25 m² (soit 44,91 à 47,80 palourdes/m²). La strate la moins densément peuplée est la strate D, avec 1,66 palourdes/0,25 m² (6,63/m²). Remarque : il convient de noter la faible précision obtenue pour la strate E.
Septembre 2003
24
0
5
10
15
20
25
A B C D E F G H Z1 Z2 Z3 S1 S4 S6
Strates
Abo
ndan
ces (
nb/0
,25
m²)
Figure 9.- Abondances en nb/0,25 m² estimées par strate pour les trois espèces
confondues (la barre verticale représente l’intervalle de confiance à 95 %)
Les strates les plus peuplées en biomasse par unité d’échantillonnage sont les strates H, S6 et E (figure 10) avec de 72,77 à 97,62 g/0,25 m² (de 291 à 390 g/m²). La strate D est celle qui présente la biomasse par unité d’échantillonnage la plus faible avec 14 g/0,25 m², soit 56 g/m².
0
50
100
150
200
A B C D E F G H Z1 Z2 Z3 S1 S4 S6
Strates
Bio
mas
ses (
g/0,
25 m
²)
Figure 10.- Biomasses en g/0,25 m² estimées par strate pour les trois espèces
confondues (la barre verticale représente l’intervalle de confiance à 95 %)
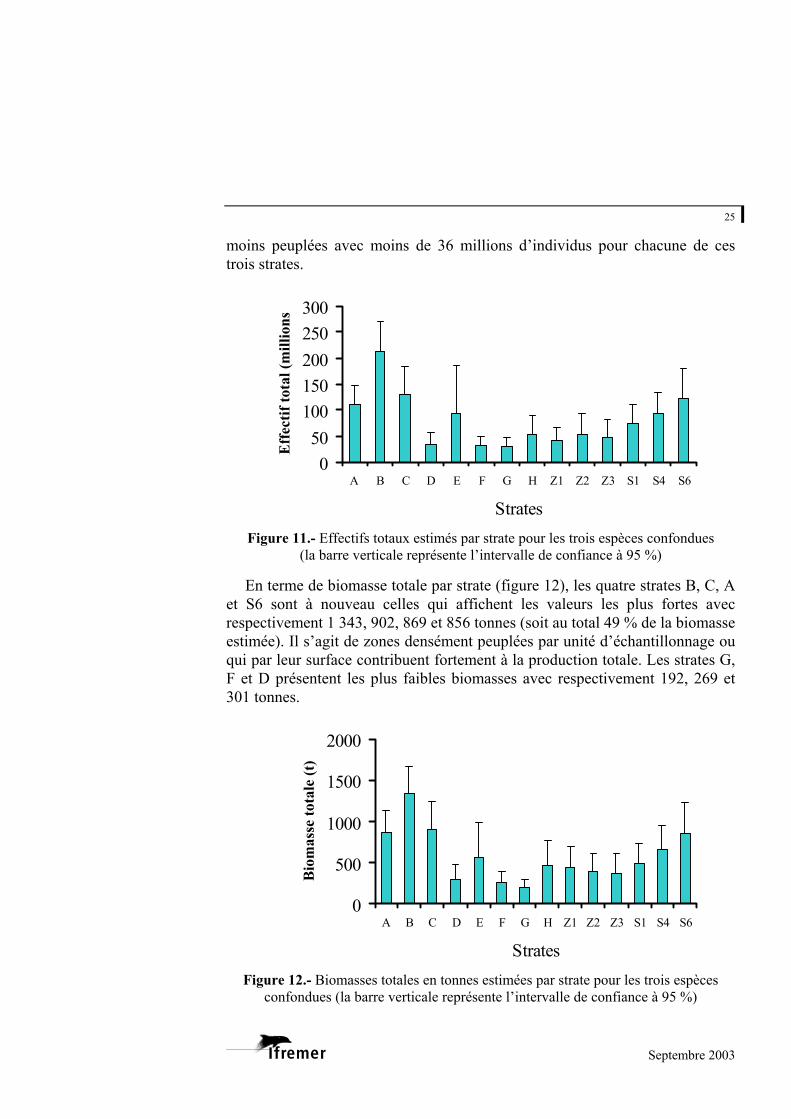
En terme d’effectifs totaux par strate, les strates B, C, S6 et A présentent des abondances élevées avec respectivement 211, 130, 122 et 111 millions d’individus (figure 11). L’ensemble de ces quatre strates participe pour 51 % de l’effectif total estimé. A l’inverse, les strates G, F et D sont les zones les
Septembre 2003
25
moins peuplées avec moins de 36 millions d’individus pour chacune de ces trois strates.
050
100150200250300
A B C D E F G H Z1 Z2 Z3 S1 S4 S6
Strates
Eff
ectif
tota
l (m
illio
ns)
Figure 11.- Effectifs totaux estimés par strate pour les trois espèces confondues
(la barre verticale représente l’intervalle de confiance à 95 %)
En terme de biomasse totale par strate (figure 12), les quatre strates B, C, A et S6 sont à nouveau celles qui affichent les valeurs les plus fortes avec respectivement 1 343, 902, 869 et 856 tonnes (soit au total 49 % de la biomasse estimée). Il s’agit de zones densément peuplées par unité d’échantillonnage ou qui par leur surface contribuent fortement à la production totale. Les strates G, F et D présentent les plus faibles biomasses avec respectivement 192, 269 et 301 tonnes.
0
500
1000
1500
2000
A B C D E F G H Z1 Z2 Z3 S1 S4 S6
Strates
Bio
mas
se to
tale
(t)
Figure 12.- Biomasses totales en tonnes estimées par strate pour les trois espèces
confondues (la barre verticale représente l’intervalle de confiance à 95 %)
Septembre 2003
26
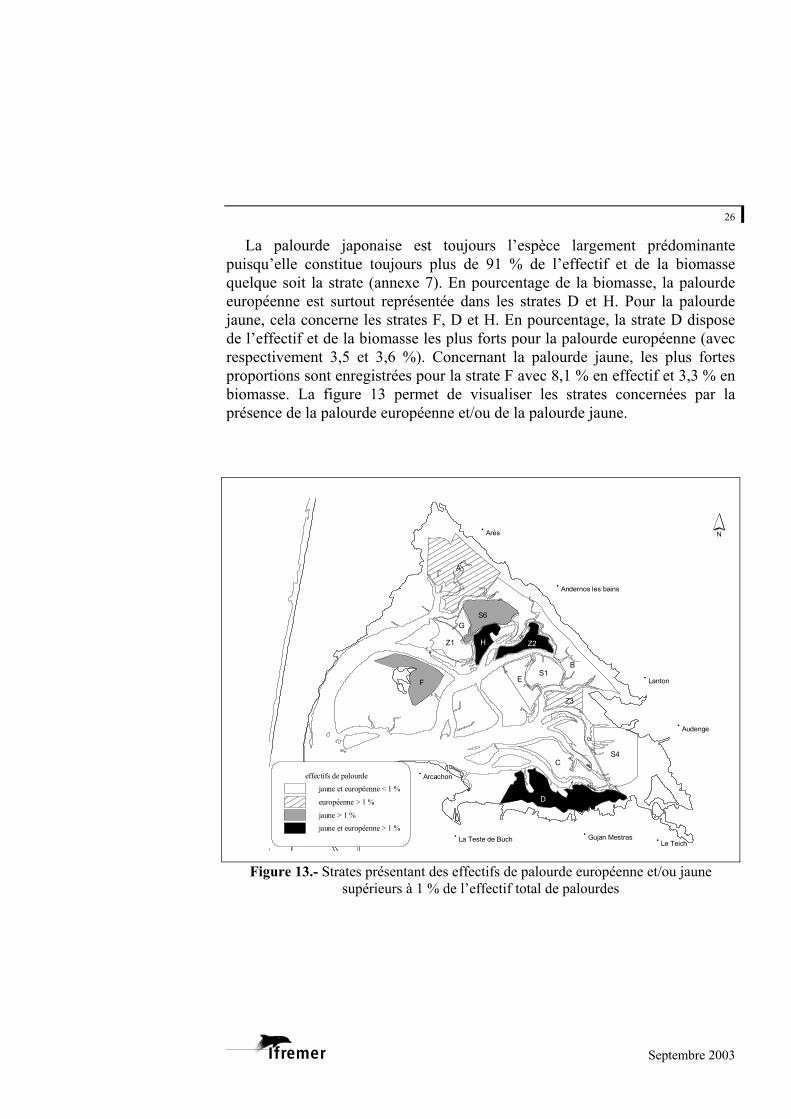
La palourde japonaise est toujours l’espèce largement prédominante puisqu’elle constitue toujours plus de 91 % de l’effectif et de la biomasse quelque soit la strate (annexe 7). En pourcentage de la biomasse, la palourde européenne est surtout représentée dans les strates D et H. Pour la palourde jaune, cela concerne les strates F, D et H. En pourcentage, la strate D dispose de l’effectif et de la biomasse les plus forts pour la palourde européenne (avec respectivement 3,5 et 3,6 %). Concernant la palourde jaune, les plus fortes proportions sont enregistrées pour la strate F avec 8,1 % en effectif et 3,3 % en biomasse. La figure 13 permet de visualiser les strates concernées par la présence de la palourde européenne et/ou de la palourde jaune.
N#
#
#
#
#
##
#
Z1
S6
Z2
S1
Z3
CS4
E
G
H
A
F
B
D
Arès
Andernos les bains
Lanton
Audenge
Le TeichGujan MestrasLa Teste de Buch
Arcachoneffectifs de palourde
jaune et européenne < 1 %
européenne > 1 %
jaune > 1 %
jaune et européenne > 1 %
Figure 13.- Strates présentant des effectifs de palourde européenne et/ou jaune supérieurs à 1 % de l’effectif total de palourdes
Septembre 2003

27
Cas des zones fermées à la pêche
Pour les trois zones récemment fermées à la pêche (Z1, Z2 et Z3), la
biomasse totale est estimée à 1 192 tonnes, soit 15 % de la biomasse estimée pour l’ensemble de la zone d’étude. La strate Z1 apparaît faiblement peuplée en effectif par unité d’échantillonnage (4,8 palourdes/0,25 m²), alors que les strates Z2 et Z3 se situent au dessus de la moyenne (avec respectivement 8,4 et 8,8 palourdes/0,25 m²). En biomasse par unité d’échantillonnage, ces trois strates présentent des valeurs supérieures à la moyenne avec 49 à 69 g/0,25 m².
Cas des anciennes zones fermées à la pêche
Pour les trois zones anciennement fermées à la pêche (S1, S4 et S6), la
biomasse totale est estimée à 2 000 tonnes, soit 25 % de la biomasse estimée pour l’ensemble de la zone d’étude. Les strates S1 et S6 présentent des abondances et des biomasses élevées. A l’opposé, la strate S4 affiche des valeurs très moyennes comparativement à celles obtenues sur l’ensemble de la zone d’étude. Cas des vases supérieures
Sur les 70 prélèvements de « vases supérieures » collectées, 30 échantillons étaient vides et le nombre moyen de palourdes par échantillon est de 2,9 individus. En effectif et pour les stations concernées, le nombre global de palourdes contenu dans ces vases supérieures correspond à près de 29 % de l’effectif global dénombré dans les prélèvements par godet.
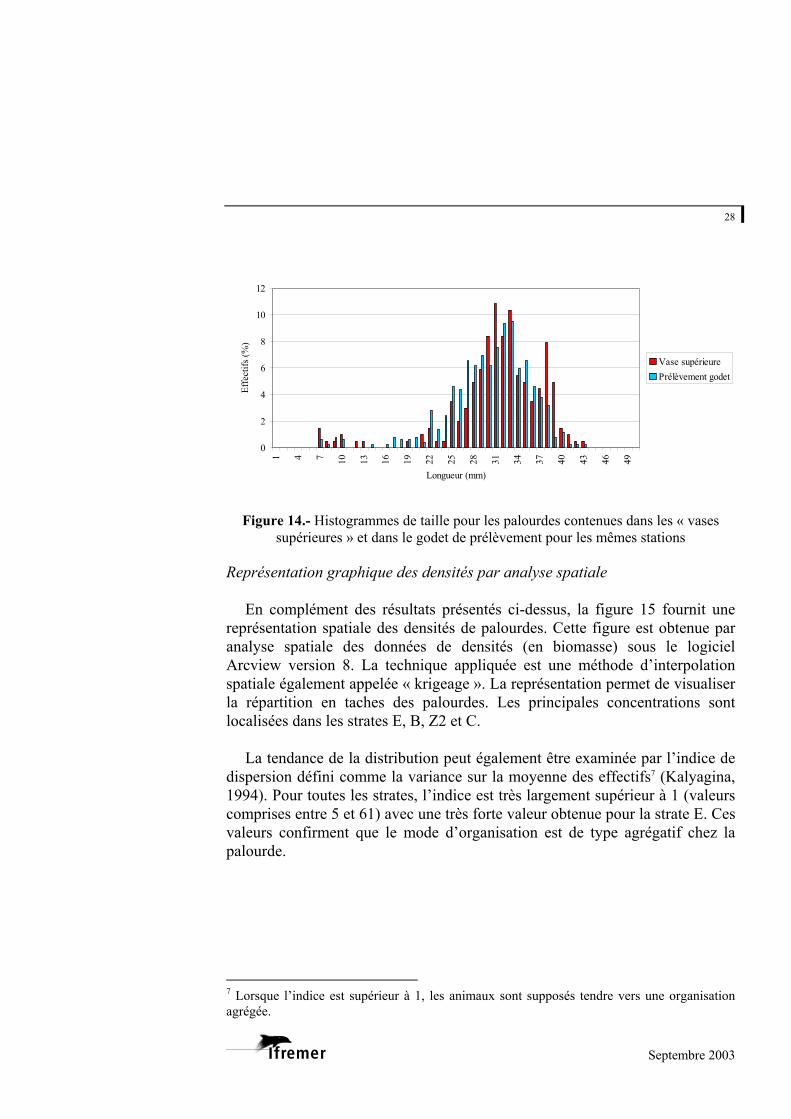
Aucune différence de longueur n’est constatée entre les histogrammes de
taille (figure 14) des palourdes présentes dans les vases supérieures et celles présentes dans le godet de prélèvement.
Septembre 2003
28
0
2
4
6
8
10
121 4 7 10 13 16 19 22 25 28 31 34 37 40 43 46 49
Longueur (mm)
Effe
ctifs
(%)
Vase supérieurePrélèvement godet
Figure 14.- Histogrammes de taille pour les palourdes contenues dans les « vases
supérieures » et dans le godet de prélèvement pour les mêmes stations Représentation graphique des densités par analyse spatiale
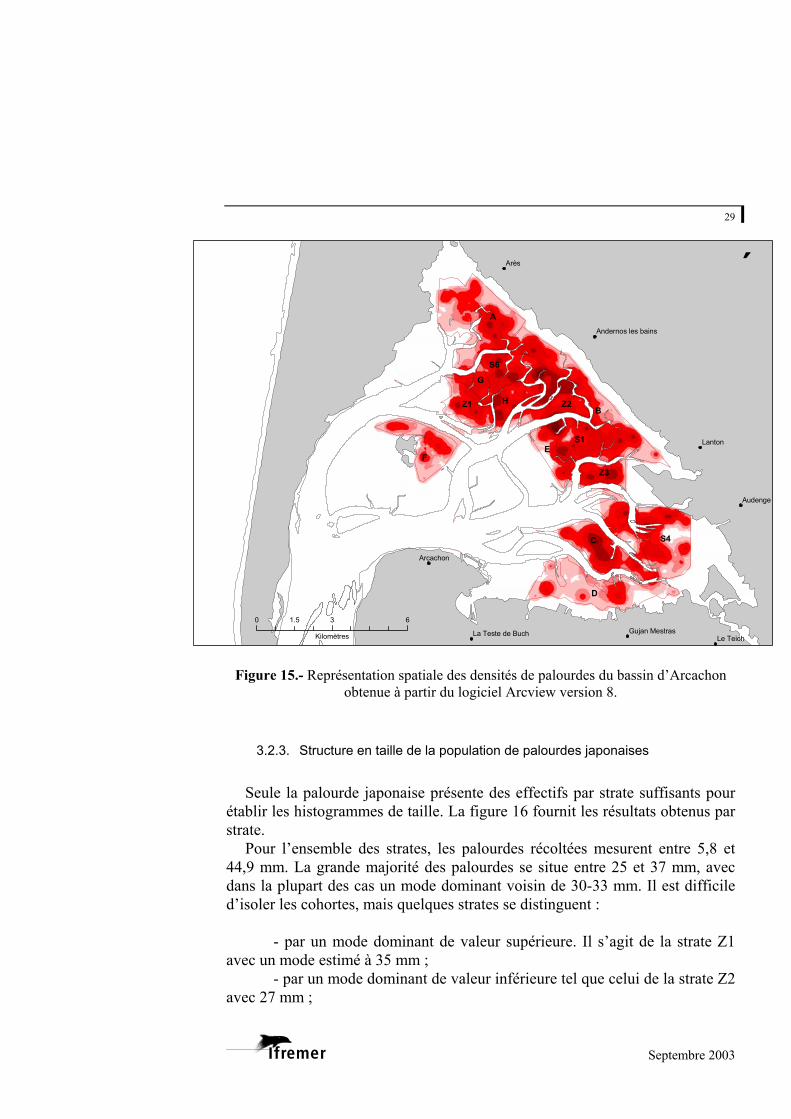
En complément des résultats présentés ci-dessus, la figure 15 fournit une représentation spatiale des densités de palourdes. Cette figure est obtenue par analyse spatiale des données de densités (en biomasse) sous le logiciel Arcview version 8. La technique appliquée est une méthode d’interpolation spatiale également appelée « krigeage ». La représentation permet de visualiser la répartition en taches des palourdes. Les principales concentrations sont localisées dans les strates E, B, Z2 et C.
La tendance de la distribution peut également être examinée par l’indice de
dispersion défini comme la variance sur la moyenne des effectifs7 (Kalyagina, 1994). Pour toutes les strates, l’indice est très largement supérieur à 1 (valeurs comprises entre 5 et 61) avec une très forte valeur obtenue pour la strate E. Ces valeurs confirment que le mode d’organisation est de type agrégatif chez la palourde.
Septembre 2003
7 Lorsque l’indice est supérieur à 1, les animaux sont supposés tendre vers une organisation agrégée.
29
A
B
D
C
F
S4
E
S6
Z1
S1
H Z2
Z3
G
Arès
Lanton
Audenge
Arcachon
Le TeichGujan MestrasLa Teste de Buch
Andernos les bains
´
0 3 61.5
Kilomètres
Figure 15.- Représentation spatiale des densités de palourdes du bassin d’Arcachon
obtenue à partir du logiciel Arcview version 8.
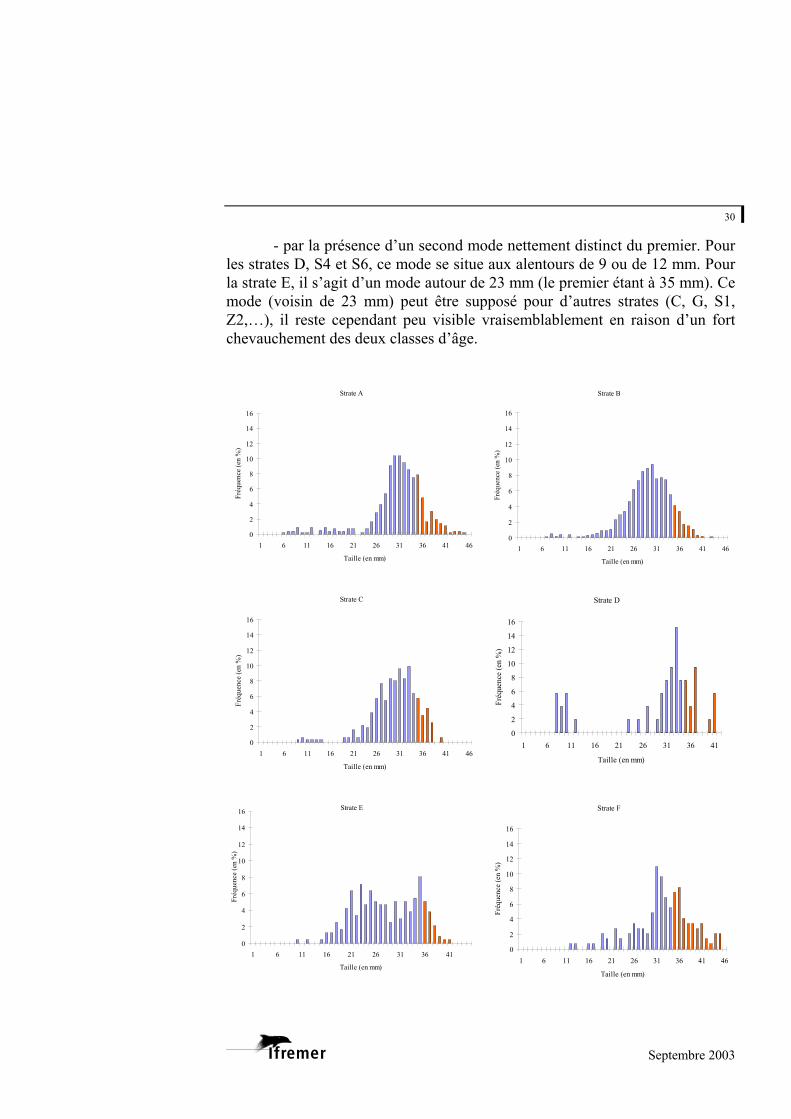
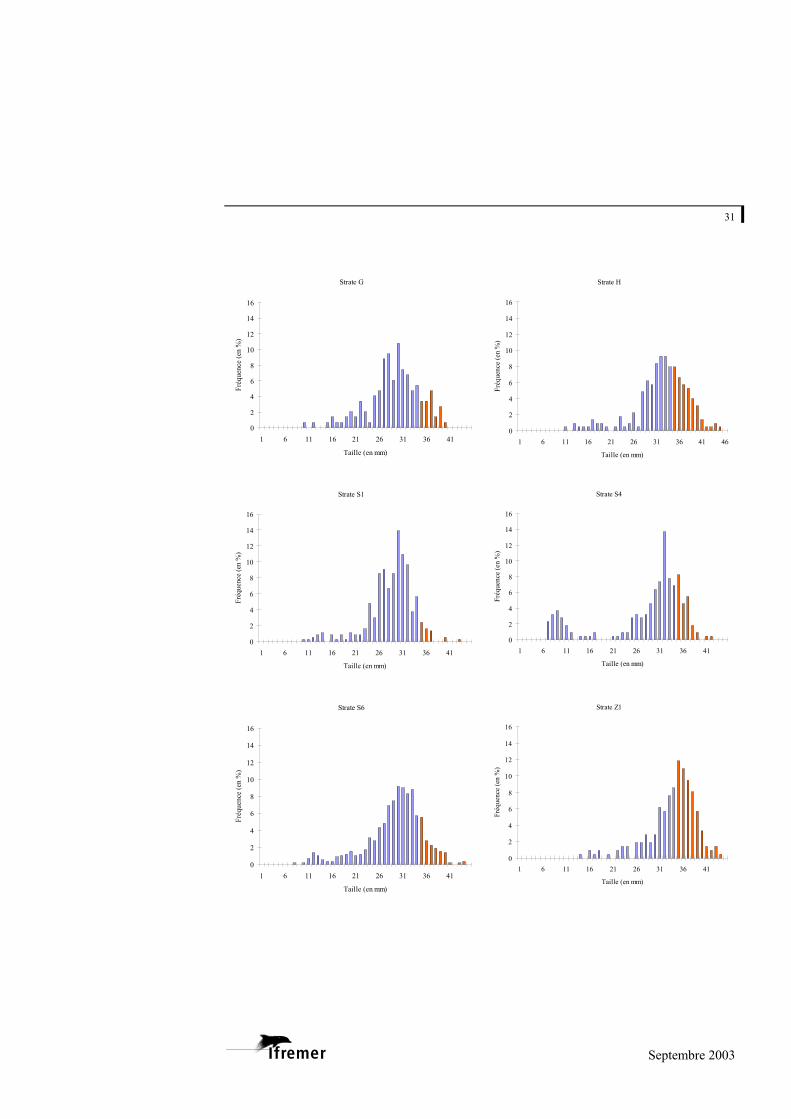
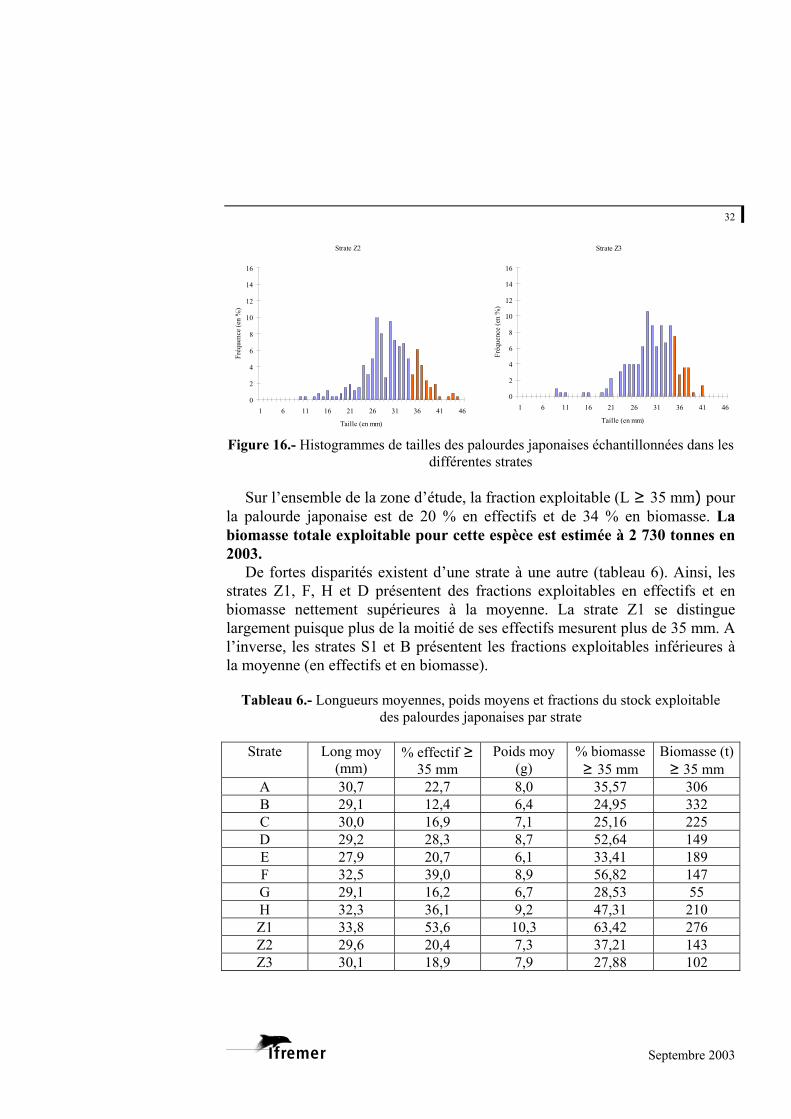
3.2.3. Structure en taille de la population de palourdes japonaises
Seule la palourde japonaise présente des effectifs par strate suffisants pour
établir les histogrammes de taille. La figure 16 fournit les résultats obtenus par strate.
Pour l’ensemble des strates, les palourdes récoltées mesurent entre 5,8 et 44,9 mm. La grande majorité des palourdes se situe entre 25 et 37 mm, avec dans la plupart des cas un mode dominant voisin de 30-33 mm. Il est difficile d’isoler les cohortes, mais quelques strates se distinguent :
- par un mode dominant de valeur supérieure. Il s’agit de la strate Z1
avec un mode estimé à 35 mm ; - par un mode dominant de valeur inférieure tel que celui de la strate Z2
avec 27 mm ;
Septembre 2003
30
- par la présence d’un second mode nettement distinct du premier. Pour les strates D, S4 et S6, ce mode se situe aux alentours de 9 ou de 12 mm. Pour la strate E, il s’agit d’un mode autour de 23 mm (le premier étant à 35 mm). Ce mode (voisin de 23 mm) peut être supposé pour d’autres strates (C, G, S1, Z2,…), il reste cependant peu visible vraisemblablement en raison d’un fort chevauchement des deux classes d’âge.
Strate A
0
2
4
6
8
10
12
14
16
1 6 11 16 21 26 31 36 41 46
Taille (en mm)
Fréq
uenc
e (e
n %
)
Strate B
0
2
4
6
8
10
12
14
16
1 6 11 16 21 26 31 36 41 46
Taille (en mm)
Fréq
uenc
e (e
n %
)
Strate C
0
2
4
6
8
10
12
14
16
1 6 11 16 21 26 31 36 41 46
Taille (en mm)
Fréq
uenc
e (e
n %
)
Strate D
0
2
4
6
8
10
12
14
16
1 6 11 16 21 26 31 36 41
Taille (en mm)
Fréq
uenc
e (e
n %
)
Strate E
0
2
4
6
8
10
12
14
16
1 6 11 16 21 26 31 36 41
Taille (en mm)
Fréq
uenc
e (e
n %
)
Strate F
0
2
4
6
8
10
12
14
16
1 6 11 16 21 26 31 36 41 46
Taille (en mm)
Fréq
uenc
e (e
n %
)
Septembre 2003
31
Strate G
0
2
4
6
8
10
12
14
16
1 6 11 16 21 26 31 36 41
Taille (en mm)
Fréq
uenc
e (e
n %
)
Strate H
0
2
4
6
8
10
12
14
16
1 6 11 16 21 26 31 36 41 46
Taille (en mm)
Fréq
uenc
e (e
n %
)Strate S1
0
2
4
6
8
10
12
14
16
1 6 11 16 21 26 31 36 41
Taille (en mm)
Fréq
uenc
e (e
n %
)
Strate S4
0
2
4
6
8
10
12
14
16
1 6 11 16 21 26 31 36 41
Taille (en mm)
Fréq
uenc
e (e
n %
)
Strate S6
0
2
4
6
8
10
12
14
16
1 6 11 16 21 26 31 36 41
Taille (en mm)
Fréq
uenc
e (e
n %
)
Strate Z1
0
2
4
6
8
10
12
14
16
1 6 11 16 21 26 31 36 41
Taille (en mm)
Fréq
uenc
e (e
n %
)
Septembre 2003
32
Strate Z2
0
2
4
6
8
10
12
14
16
1 6 11 16 21 26 31 36 41 46
Taille (en mm)
Fréq
uenc
e (e
n %
)
Strate Z3
0
2
4
6
8
10
12
14
16
1 6 11 16 21 26 31 36 41 46
Taille (en mm)
Fréq
uenc
e (e
n %
)
Figure 16.- Histogrammes de tailles des palourdes japonaises échantillonnées dans les différentes strates
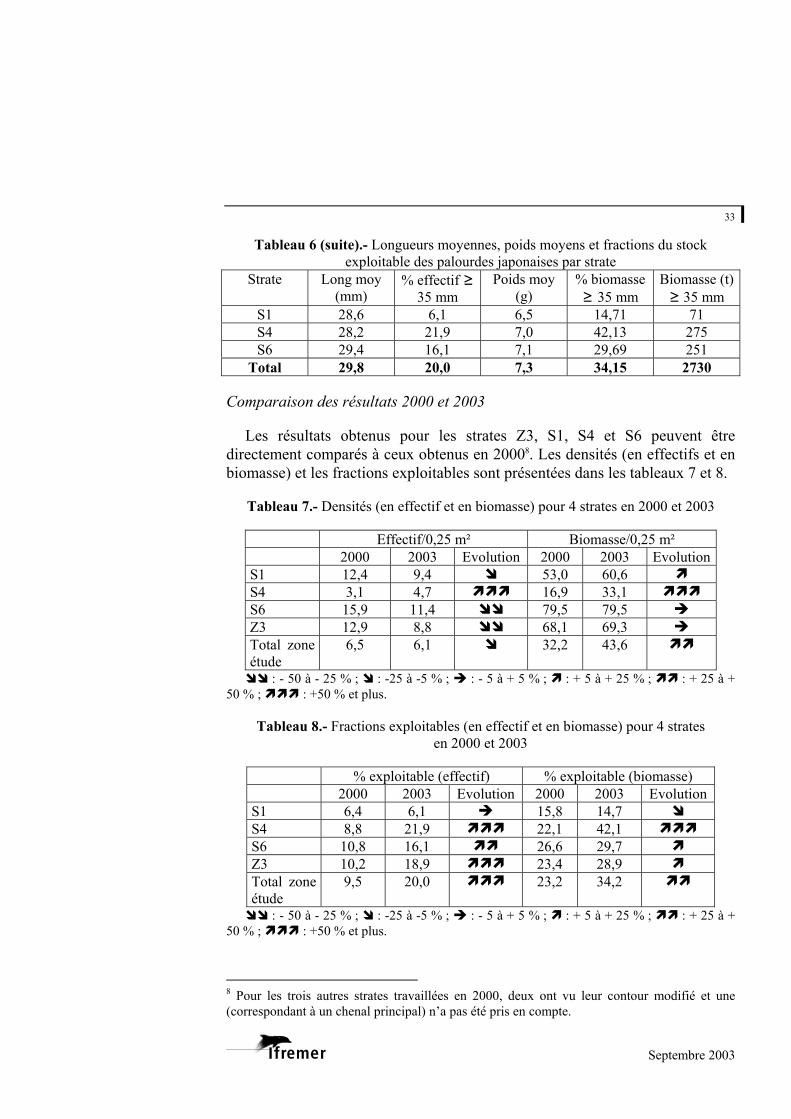
Sur l’ensemble de la zone d’étude, la fraction exploitable (L ≥ 35 mm) pour
la palourde japonaise est de 20 % en effectifs et de 34 % en biomasse. La biomasse totale exploitable pour cette espèce est estimée à 2 730 tonnes en 2003.
De fortes disparités existent d’une strate à une autre (tableau 6). Ainsi, les strates Z1, F, H et D présentent des fractions exploitables en effectifs et en biomasse nettement supérieures à la moyenne. La strate Z1 se distingue largement puisque plus de la moitié de ses effectifs mesurent plus de 35 mm. A l’inverse, les strates S1 et B présentent les fractions exploitables inférieures à la moyenne (en effectifs et en biomasse).
Tableau 6.- Longueurs moyennes, poids moyens et fractions du stock exploitable des palourdes japonaises par strate
Strate Long moy
(mm) % effectif ≥
35 mm Poids moy
(g) % biomasse ≥ 35 mm
Biomasse (t) ≥ 35 mm
A 30,7 22,7 8,0 35,57 306 B 29,1 12,4 6,4 24,95 332 C 30,0 16,9 7,1 25,16 225 D 29,2 28,3 8,7 52,64 149 E 27,9 20,7 6,1 33,41 189 F 32,5 39,0 8,9 56,82 147 G 29,1 16,2 6,7 28,53 55 H 32,3 36,1 9,2 47,31 210 Z1 33,8 53,6 10,3 63,42 276 Z2 29,6 20,4 7,3 37,21 143 Z3 30,1 18,9 7,9 27,88 102
Septembre 2003
33
Tableau 6 (suite).- Longueurs moyennes, poids moyens et fractions du stock exploitable des palourdes japonaises par strate
Strate Long moy (mm)
% effectif ≥ 35 mm
Poids moy (g)
% biomasse ≥ 35 mm
Biomasse (t) ≥ 35 mm
S1 28,6 6,1 6,5 14,71 71 S4 28,2 21,9 7,0 42,13 275 S6 29,4 16,1 7,1 29,69 251
Total 29,8 20,0 7,3 34,15 2730
Comparaison des résultats 2000 et 2003 Les résultats obtenus pour les strates Z3, S1, S4 et S6 peuvent être
directement comparés à ceux obtenus en 20008. Les densités (en effectifs et en biomasse) et les fractions exploitables sont présentées dans les tableaux 7 et 8.
Tableau 7.- Densités (en effectif et en biomasse) pour 4 strates en 2000 et 2003 Effectif/0,25 m² Biomasse/0,25 m² 2000 2003 Evolution 2000 2003 EvolutionS1 12,4 9,4 53,0 60,6 S4 3,1 4,7 16,9 33,1 S6 15,9 11,4 79,5 79,5 Z3 12,9 8,8 68,1 69,3 Total zone étude
6,5 6,1 32,2 43,6
: - 50 à - 25 % ; : -25 à -5 % ; : - 5 à + 5 % ; : + 5 à + 25 % ; : + 25 à + 50 % ; : +50 % et plus.
Tableau 8.- Fractions exploitables (en effectif et en biomasse) pour 4 strates
en 2000 et 2003 % exploitable (effectif) % exploitable (biomasse) 2000 2003 Evolution 2000 2003 Evolution S1 6,4 6,1 15,8 14,7 S4 8,8 21,9 22,1 42,1 S6 10,8 16,1 26,6 29,7 Z3 10,2 18,9 23,4 28,9 Total zone étude
9,5 20,0 23,2 34,2
: - 50 à - 25 % ; : -25 à -5 % ; : - 5 à + 5 % ; : + 5 à + 25 % ; : + 25 à + 50 % ; : +50 % et plus.
Septembre 2003
8 Pour les trois autres strates travaillées en 2000, deux ont vu leur contour modifié et une (correspondant à un chenal principal) n’a pas été pris en compte.

34
Pour les trois anciennes zones fermées à la pêche, les densités exprimées en effectifs par 0,25 m² sont en baisse dans deux cas sur trois. Exprimées en biomasse, les densités sont en revanche égales ou supérieures à celles obtenues en 2000. Concernant la fraction exploitable, les résultats sont également très variables d’une strate à une autre. La strate S4 présente une forte augmentation de sa fraction exploitable en effectif et en biomasse entre les deux campagnes.
En comparaison, la strate Z3 qui est une zone nouvellement mise en réserve
est caractérisée par des valeurs et par une évolution de ces dernières proches de celles observées pour les strates S1 et S6. Ces observations ne permettent pas de dégager un effet réserve sur ces paramètres du stock.
4. Discussion
4.1. La méthode de prélèvement
La présence de « vases supérieures » a souvent été observée puisqu’on
estime qu’elle s’est produite pour plus de 75 % des prélèvements. Cette accumulation se produit uniquement en présence de vases et n’est pas observée en présence de sédiments à majorité composés de sables ou de débris coquilliers. Cependant, l’analyse du contenu des 70 « vases supérieures » conservées montre que ces prélèvements présentent une quantité d’individus non négligeable par rapport à celle collectée par le godet de la benne (29 % des effectifs). En revanche, il n'y a pas de différence sur la taille des palourdes prélevées.
Ce phénomène n’a pas été détecté lors des évaluations menées dans le golfe
du Morbihan. Il s’est déjà produit lors de la campagne de 2000 sur le bassin d’Arcachon. Il est possible que le Bassin présente des zones de substrats plus meubles qui entraînent un léger enfoncement de l’armature métallique supportant la benne lors de la dépose. Lorsque la benne est actionnée, les tous premiers centimètres de sédiment, situés en surface, s’accumulent alors au-dessus du godet. Pour étayer ces propos et dans le cas d’une nouvelle évaluation, il conviendra de réaliser préalablement des tests avec des observations in situ. Des modifications techniques pourront être apportées (par exemple en augmentant la surface de contact avec le substrat en vue de limiter l’enfoncement) si ce phénomène est confirmé.
Dans tous les cas, ces prélèvements ne peuvent être inclus dans l’évaluation
de biomasse car cette dernière s’effectue à volume constant. Ils sont très
Septembre 2003

35
irréguliers, de taille variable d’un coup de benne à un autre et, de plus, ils ne peuvent pas être systématiquement récupérés (décrochage lors de la sortie de l’eau de la benne). Il est néanmoins important que le prélèvement soit réalisé depuis la surface du sédiment car les palourdes sont essentiellement localisées dans les 10 premiers centimètres (Quéro et Vayne, 1998). Concernant l’estimation réalisée en 2003, le phénomène a pu entraîner une sous-estimation de la biomasse.
4.2. La relation taille poids frais
L'ensemble des strates pour lesquelles on observe les palourdes les plus
lourdes pour leurs tailles sont des strates composées uniquement de sédiments qualifiés de "slikke" sablo-vaseuses (Manaud, 1997). Ce type de sédiment est caractérisé par une gamme granulométrique très étendue avec une forte proportion de matière organique et le plus souvent recouvert de petites zostères. La zone d'investigation dans le cadre de l'évaluation du stock de palourdes est majoritairement recouverte par ce type de sédiment. Il apparaît comme le plus favorable pour leur développement : une granulométrie adaptée à leur capacité d'enfouissement et une richesse en matière organique pour permettre leur développement.
Des études biométriques plus poussées (notamment sur le coefficient
d’allométrie) en liaison avec les caractéristiques du milieu pourront être menées ultérieurement dans le but de compléter des travaux sur la croissance de la palourde dans le bassin d’Arcachon.
4.3. Les espèces
La palourde japonaise domine très largement puisqu’elle représente
respectivement 98 et 99 % des effectifs et de la biomasse estimés. L’évaluation menée en 2003 sur l’ensemble de l’aire de répartition corrobore ainsi les résultats obtenus en 2000 sur près de 24 km² (Bertignac et al., 2001). La palourde européenne et la palourde japonaise présentent des modes de vie similaires et la compétition s’exerce au détriment de la palourde européenne. La palourde jaune qui reste la troisième espèce en biomasse est très peu présente dans les strates situées en fond de Bassin.
Septembre 2003

36
4.4. Les densités et les effectifs totaux en nombre d’individus et en poids
Les densités en effectifs sont en légère baisse par rapport à l’évaluation
menée en 2000 (tableau 9)9. Elles sont similaires à celles observées sur le golfe du Morbihan depuis 2001 (Péronnet et al., 2003) et sont bien supérieures à celles obtenues sur l’étang de Thau en 1998 (palourde européenne- Anonyme, 1999). En poids, l’augmentation de la densité moyenne observée est à mettre en relation avec l’augmentation très forte du poids et de la longueur moyenne des échantillons collectés entre 2000 et 2003. Cette augmentation pourrait s’expliquer par le fait que les nouvelles strates prospectées présentent des palourdes plus grosses que les strates échantillonnées en 2000. Cependant, cette augmentation en poids et en longueur moyenne touche également les zones communes aux deux campagnes. Deux autres facteurs peuvent expliquer cette situation :
- une diminution du niveau d’exploitation suite aux conséquences de la
marée noire provenant du naufrage du Prestige. Cela concerne non seulement l’interdiction de la pêche des coquillages du 4 au 16 janvier 200310, mais surtout la réquisition des navires par les services de l’Etat dans le cadre des plans Polmar Terre et Mer. Ainsi, de janvier à mars, une trentaine de bateaux ayant une activité « petite pêche »11 et détenteurs de la licence « coques et palourdes » ont participé au nettoyage des zones d’estran polluées pour un total de 317 jours. Cette participation a ainsi pu limiter la disponibilité de professionnels pour la pêche de la palourde, alors que le premier trimestre représente un peu plus de 20 % des captures déclarées sur 2001 et 2002 (source : Ifremer). A cela s’ajoute certainement un contexte morose en terme de consommation des produits de la mer. Ce dernier a pu limiter le nombre de jours de sorties des professionnels. Sur le plan national, le marché de Rungis a enregistré un recul de 19 % de la commercialisation des palourdes entre les mois de février et de mai 2002 et 2003. Sur la même période et pour l’huître, la consommation des ménages français diminue de 8 % (Baranger et al., 2003) ;
- la réalisation de conditions environnementales favorables. Les conditions météorologiques des mois précédents la canicule de l’été ont été caractérisées par des températures de l’air supérieures à la normale. Sur le Bassin, les valeurs habituelles ont été dépassées dès le mois de mars
9 La première évaluation porte sur près de 24 km², la deuxième sur 46 km². Pour les zones communes aux deux campagnes et pouvant être comparées, la tendance est identique sauf pour la strate S4. 10 Arrêtés préfectoraux du 4 et du 16 janvier 2003.
Septembre 2003
11 Il n’a pas été possible de procéder au même décompte pour les navires CPP détenteurs de cette même licence.
37
(Auby, comm. pers.). Ces conditions ont pu induire des blooms phytoplanctoniques précoces et être favorables au développement des palourdes dans la mesure où elles sont restées dans les limites du preferendum thermique de l’espèce12.
Tableau 9.- Comparaison des densités, fractions exploitables, longueurs et poids moyens obtenus pour trois stocks de palourdes
Effectif/
m² Biomasse
(g)/m² Fraction
exploitable (effectif)
Fraction exploitable (biomasse)
Longueur moyenne
(mm)
Poids moyen
(g) Arcachon 2000 Bertignac et al. (2001)
26,17
129
9 %
23 %
25,4
4,9
Arcachon 2003 Présente étude
24,32
174
20 %
34 %
29,8
7,3
Morbihan 2003 D’après Peronnet et al. (2003)
E : 24,84ZP : 26
E : 209
E : 18 %
ZP : 19 %
ZP : 34 %
ZP : 29,9
ZP : 7,6
Etang de Thau 1998* Anonyme (1999)
8,3
78,18
30 %
/
/
/
E : Ensemble du gisement ; ZP : Zone de Pêche * Concerne la palourde européenne
La biomasse totale du stock de palourdes du bassin d’Arcachon est estimée à 8 095 tonnes. Cette valeur est très élevée comparée à celles obtenues pour le golfe du Morbihan et pour l’étang de Thau. En effet, le stock du golfe du Morbihan est estimé à 1 273 tonnes pour l’ensemble du gisement dont 724 tonnes pour les zones de pêche (Péronnet et al., 2003). Sur l’étang de Thau, la dernière estimation remonte à 1998 et s’élève à 320 tonnes (Anonyme, 1999). De plus, les résultats obtenus sur l’aire de répartition supposée (couvrant 46,4 km²) sont élevés en comparaison de ceux fournis lors de l’estimation de 2000 pour une aire échantillonnée de 23,1 km² (2 900 tonnes). Ces résultats ne peuvent être expliqués par le seul décalage des dates de campagne d’évaluation 2000 et 2003 (moins de un mois). Ils indiquent que la majeure partie de la biomasse estimée se situe au nord d’un arc de cercle allant d’Audenge à
Septembre 2003
12 18 à 25°C (Quéro et Vayne, 1998).

38
Arcachon en passant par les chenaux de Moutchalette, de Girouasse et de Mapouchet. La zone Nord, ainsi définie, participe pour près de 60 % de la biomasse totale.
4.5. La fraction exploitable
Elle s’élève à 20 % en effectif13 (proportion d’individus de longueur
supérieure à 35 mm) et à 34 % en biomasse (proportion de la biomasse estimée composée par ces individus de plus de 35 mm). Ces valeurs sont bien supérieures à celles obtenues sur la zone prospectée en 2000 et ces pourcentages se rapprochent de ceux enregistrés pour le golfe du Morbihan. Sur le bassin, la biomasse totale exploitable est élevée puisqu’elle est estimée à 2 730 tonnes début juillet 2003, dont 520 tonnes situées dans les trois zones de réserve actuelles.
Une indication du niveau de recrutement de l’année précédente est fournie
par la proportion d’individus de longueur inférieure à 17 mm (Berthou et al., 1997). Les valeurs obtenues sont faibles (5 % en 2003 sur 46 km², 15 % en 2000 sur 23 km²), mais les histogrammes de fréquence de tailles paraissent particulièrement déséquilibrés entre les individus les plus jeunes et les cohortes suivantes. Cette situation pourrait refléter un sous-échantillonnage de la fraction de la population de longueur inférieure à 17 mm. Considérant la méthode de prélèvement mise en œuvre, elle pourrait provenir du tamisage sur 5 mm (maille carrée) qui occasionnerait des pertes pour les palourdes de longueur voisine de 5 mm. Cette hypothèse, déjà avancée par Bertignac et al. (2001), reste à valider. Dans l’état actuel des connaissances, elle semble plus vraisemblable que le reflet d’un mauvais recrutement, puisque dans ces conditions et d’après les résultats obtenus en 2000 par Bertignac et al. (2001), nous aurions dû observer une diminution du stock en 2002 et en 2003. Pour 2003, cela est contraire aux résultats obtenus lors de la campagne.
De plus, en l’absence de travaux plus poussés sur la croissance de la
palourde dans le Bassin, l’interprétation des histogrammes de taille reste délicate car les données obtenues par Deltreil et al. (1993) amènent à penser que les valeurs obtenues autour de 25 et 37 mm proviennent du chevauchement des valeurs relatives aux cohortes 2001 et 2000. Dans ces conditions, l’appréciation du niveau d’abondance de chacune ne peut être estimée visuellement, ce qui empêche d’avoir une vision dynamique dans le temps de la production du stock. La mise en œuvre d’outils statistiques permettant la
Septembre 2003
13 Cela concerne uniquement la palourde japonaise.

39
séparation de groupes de tailles multinormées couplée à des suivis de croissance in situ permettraient de lever ces difficultés. Pour ces derniers, la méthodologie pourrait s’appuyer sur celle utilisée par de Kergariou et al. (1981) dans le golfe du Morbihan en 1979 et 1980.
Les données déjà disponibles14 montrent que le gain de poids entre 30 et
35 mm est fort et que le respect d’une taille de capture de 35 mm permet de majorer la production en poids pour l’ensemble des exploitants. La capture d’individus de taille inférieure à 35 mm engendre en effet des pertes pondérales importantes au cours de la campagne et se traduirait par un manque à gagner collectif. D’ores et déjà, une première approximation peut être faite, elle devra être modulée par le taux de mortalité.
La relation taille/poids obtenue sur le Bassin (cf. § 3.2.1.) permet d’estimer la perte pondérale subie. En moyenne, une palourde de 30 mm pèse 6,8 g ; une palourde de 35 mm pèse 11,0 g. Dans des conditions d’élevage sur le Bassin, les travaux de Robert et al. (1993) montrent que cet accroissement est obtenu en cinq-six mois. Respecter la taille de capture de 35 mm représenterait un gain pondéral de 60 % dans ce cas. Traduit en terme de valeur, ce gain pourrait être encore supérieur s’il existait un réel écart de prix payé au pêcheur entre les différentes catégories commerciales. Dans la pratique, il semble qu’actuellement ce prix soit indépendant de la taille (de l’ordre de 4,5 à 5 euros au kilo – prix pratiqués en août et septembre 2003). Cependant, le différentiel de prix existe au niveau du mareyeur (il serait de l’ordre de + 3 euros du kilogramme à chaque passage en catégorie supérieure). Dans ces conditions, on peut supposer qu’un meilleur respect du tri permettrait une meilleure valorisation des captures des professionnels et à terme un gain financier pour l’ensemble des exploitants.
4.6. Les anciennes zones de réserve
Concernant les anciennes zones de réserve (S1, S4 et S6) il n’apparaît pas
de différences significatives avec les autres zones pour les paramètres densité et fraction exploitable. De même, les palourdes les plus grosses n’ont pas été observées dans une de ces zones de réserve. Cette situation amène à s’interroger sur « l’effet réserve » réel de ces zones. Elle peut cependant masquer une exploitation des palourdes les plus grosses depuis la réouverture de ces zones à la pêche au 1er janvier 2003 (arrêté préfectoral du 13/12/2002) et la réalisation de la campagne d’évaluation. Des conditions environnementales
Septembre 2003
14 A partir des essais de vénériculture sur le Bassin et par la présente étude.

40
défavorables pour ces strates (nature du substrat, apports en eau douce, courant,…) pourraient également expliquer en partie ces résultats.
Une fraction plus forte des stades les plus jeunes (mode autour de 9 ou 12 mm) a été mise en évidence, indiquant peut-être une plus forte intensité du recrutement. Ces 3 anciennes zones de réserve participent aujourd’hui pour 25 % de la biomasse estimée avec 2 000 tonnes. Les zones de réserves actuelles (Z1, Z2 et Z3) représentent 15 % avec 1 192 tonnes. 5. Conclusion et recommandations
Cette deuxième campagne d’évaluation de la biomasse de palourde a permis d’estimer le stock et ses principales caractéristiques sur l’ensemble de l’aire de répartition supposée de la palourde intra-bassin (46 km²). En 2003, le stock est principalement caractérisé par :
- une très large prépondérance de la palourde japonaise (Ruditapes
philippinarum) puisque cette espèce représente désormais 98 % des effectifs totaux et 99 % de la biomasse totale ;
- une densité moyenne en effectifs par unité d’échantillonnage en léger
repli par rapport à celle enregistrée en 2000, mais compensée par une forte augmentation du poids moyen des individus. Parmi les raisons pouvant expliquer cette situation, une diminution du niveau d’exploitation en début d’année (dû à une baisse de la disponibilité des professionnels suite à la marée noire) et/ou la réalisation de conditions environnementales favorables ont été avancées ;
- une biomasse totale élevée puisqu’elle est estimée à 8 095 tonnes (+/- 1 046 tonnes) début juillet 2003. Pour la palourde, le bassin d’Arcachon se place ainsi au premier rang des zones françaises exploitées bénéficiant d’une évaluation de stock ;
- une biomasse exploitable également élevée car évaluée à 2 730 tonnes au moment de la campagne, dont 2 210 tonnes accessibles puisque situées en dehors des trois zones de réserves. Exprimée en pourcentage de la biomasse totale, cette valeur est cette fois voisine de celles obtenus lors des dernières évaluations menées dans le golfe du Morbihan.
Dans le cadre d'une nouvelle estimation de biomasse sur le bassin d'Arcachon, il conviendra de supprimer le phénomène des « vases
Septembre 2003

41
supérieures » en réalisant préalablement des observations in situ et d’effectuer les adaptations techniques utiles. Ceci afin d'éviter une sous-estimation du stock en place.
L’analyse de la structure en taille reste à approfondir car de nombreuses incertitudes demeurent. Elle montre une faible proportion de palourdes de longueur inférieure à 17 mm et une forte proportion de palourdes de longueur comprise entre 25 et 37 mm correspondant vraisemblablement à deux cohortes distinctes (2000 et 2001). Or, dans le cas d’une espèce à phase exploitable courte (deux à trois ans) présentant de fortes variations inter-annuelles d’abondance, il est nécessaire d’évaluer très régulièrement le stock de palourdes en incluant les pré-recrues afin de limiter les fluctuations de production et tamponner l’effet de la pêche (Lesueur, 2002).
Des travaux sur la simulation de la croissance couplés à des suivis de la croissance in situ permettraient de lever ces difficultés d’interprétation et d’apporter des éléments de réponse sur la dynamique du stock exploité. L’impact de la densité sur la croissance et la survie pourrait de plus être estimé, ce qui permettrait de statuer sur l’intérêt de réensemencer ou non certaines zones.
Parallèlement, il est nécessaire d’améliorer les connaissances concernant la
pêcherie. Actuellement, les informations disponibles à partir des fiches de pêche sont le poids capturé par navire, par marée (ou regroupement de marées) et par espèce à l’intérieur du Bassin, ainsi que la durée de la marée. Même si ces données permettent de fournir une indication sur l’effort de pêche déployé, une information plus précise sur la localisation, mais surtout sur la taille des captures manque fortement. La déclaration des captures ventilées par catégorie de tailles serait utile (voire la mise en place d’un échantillonnage mensuel des captures).
L’ensemble de ces informations est indispensable à la gestion de la pêcherie. En l’état actuel des connaissances, le niveau d’exploitation encadré par l’octroi de 55 licences s’établit à environ 300 tonnes15 pour 2002. Cette valeur reste bien en dessous de l’image instantanée de la biomasse exploitable accessible (hors réserve) estimée à 2 201 tonnes au début du mois de juillet 2003.
Septembre 2003
15 La durée de la marée est généralement comprise entre 4 et 6 heures.

42
6. Bibliographie ANONYME, 1999. Gestion d’une ressource naturelle exploitée. Cas de la palourde (Ruditapes decussatus) dans la lagune de Thau : Diagnostic pluridisciplinaire. Rapport du programme Palourde (Comité Régional des Pêches Maritimes et des Elevages Marins du Languedoc-Roussillon). 103 pages + annexes. AUBY I., 1993. Évolution de la richesse biologique du bassin d’Arcachon. Rapport Société Scientifique d’Arcachon. Laboratoire d’Océanographie Biologique, Ifremer. Contrat SSA n°91 5 527 019, 222 pages + annexes. BARANGER L., BIGOT J.-F., LE BIHAN V. et Y. PERRAUDEAU. – Note de conjoncture socio-économique cultures marines n°15. Observatoire des pêches et des cultures marines du golfe de Gascogne. Aglia, Len Corrail - IMA, 4 pages. BERTHOU P., MAHE J.C., HUET J. et P. NOEL, 1997. Évaluation du Stock de Palourdes du golfe du Morbihan en 1997. Rapport interne Ifremer, 17 pages. BERTIGNAC M., AUBY I., FOUCARD J., MARTIN S., DE MONTAUDOUIN X. et P.-G. SAURIAU, 2001. Évaluation du stock de palourdes du Bassin d’Arcachon. Rapport Interne Ifremer DRV/RH/RST/2001-05, 35 pages. BOBO J.-P., 1996. La contamination des coquillages fouisseurs du bassin d’Arcachon : conséquences sur l’exploitation des gisements. École Nationale des Travaux Agricoles de Bordeaux. 43p. + annexes. COCHRAN W.G. 1977. Sampling Techniques (Third edition) John Wiley & Sons, 428p. FLASSCH J.-P. (red), 1988. La Palourde :dossier d'élevage /IFREMER. Paris. Service de valorisation des Recherches, Ifremer Paris. Direction ressources vivantes, 108 pages. KALYAGINA E. E., 1994. Distribution and population structure of commercial bivalves Ruditapes philippinarum and Mya arenaria in Bousse Lagoon (Southern Sakhalin). Russian Journal of MArine Biology, Vol .20, No 3 :164-168.
Septembre 2003

43
LARUELLE F., 1999.- Phénologie et déterminisme de la reproduction chez Ruditapes decussatus et Ruditapes philippinarum en Bretagne. Thèse 3ème cycle, Univ. Brest. LESUEUR M., 2002. Contribution à l’évaluation des interactions entre usages halieutiques : le cas du gisement classé de Sarzeau (golfe du Morbihan). Mémoire DAA ENSAR, 59 pages. LE TREUT Y., 1986. La Palourde. Anatomie – Biologie – Élevage – Pêche – Consommation – Inspection Sanitaire. Thèse en Doctorat Vétérinaire, École Nationale Vétérinaire de Nantes, 162 pages. MAITRE-ALLAIN T., 1983. Croissance de deux espèces de palourdes dans l'étang de Thau (Herault, France) en fonction des facteurs hydrologiques. Rapp. Comm. Intern de la Mer Med., 28 (6) : 165-166. MAITRE-ALLAIN T., 1992. Suivi d’un ensemencement de palourdes (Ruditapes decussatus et Ruditapes philippinarum) dans l’étang de Thau (Hérault). Rapport CLPM Sète/ ISTPM Sète, non paginé. MANAUD F., 1997. Étude intégrée du bassin d'Arcachon. Synthèse 1997. Rapport Ifremer Arcachon, 123p. POPOVSKY J., GALLET F. et F. PAUTRIZEL, 1999. État des suivis et des évaluations des stocks de coquillages en élevage et des gisements naturels dans les bassins conchylicoles de la façade Aglia. Rapport Observatoire des pêches et des cultures marines du golfe de Gascogne, 91 pages. QUERO J.C. et J.-J. VAYNE, 1998. Les fruits de la mer et plantes marines des pêches françaises. Algues, plantes marines éponges, coraux, coquillages, crustacés, oursins,…Eds Delachaux et Niestlé, Les Encyclopédies du Naturaliste, 256 pages. ROBERT R. et J.-P. DELTREIL, 1990. Élevage de la Palourde japonaise Ruditapes philippinarum dans le bassin d'Arcachon, bilan des dix dernières années et perspectives de développement, Rapport interne Ifremer, RIDRV-90.40-RA/Arcachon, 21p. ROBERT R, TRUT G. et J.-L. LABORDE, 1993. Growth, reproduction and gross biochemical composition of the Manila clam Ruditapes philippinarum in the Bay of Arcachon, France. Marine Biology 116, 291-299.
Septembre 2003

44
TAMURA T., 1970. in Le Treut (1986). TRENKEL V. et I. PERONNET, 2003. Analyse et modification du plan d’échantillonnage utilisé pour l’estimation d’abondance des palourdes dans le golfe du Morbihan. 5 pages. PERONNET I., DIMEET J. et M. SALAUN, 2003. Évaluation de la biomasse de palourdes dans le golfe du Morbihan. 8 pages. SCHERRER B., 1984. Biostatistique. Gaëtan Morin Editeur, 850 pages.
Septembre 2003

45
7. Annexes
Septembre 2003
46
Annexe 1 : carte des 14 strates retenues pour la campagne 2003.
N
N
N
N
N
NN
N
Arès
Lanton
Audenge
Le Teich
Arcachon
Gujan MestrasLa Teste de Buch
Andernos les bains
B
A
D
C
F
S4
E
S6
Z1
S1
H Z2
Z3
G
N
Répartition initiale
Strates
Septembre 2003

47
Annexe 2 : photos de la campagne d’échantillonnage 2003 (Copyright : Ifremer).
Photo 1 : vue de la benne « Hamon »
Photo 2 : "vase supérieure" située au dessus du godet
Septembre 2003

48
Photo 3 : dépôt du prélèvement pour tamisage
Photo 4 : rinçage du prélèvement
Septembre 2003

49
Annexe 3 : échantillonnage stratifié à deux niveaux pour l’évaluation directe des mollusques bivalves (in Berthou et al. 1997) et échantillonnage stratifié à un niveau.
Pour les strates avec deux prélèvements par station, l’échantillonnage est stratifié et comporte deux niveaux ; les strates sont spatiales, les unités secondaires sont de taille constante.
Taux d’échantillonnage
Unités primaires : les unités primaires correspondent aux stations ou points géographiques tirés au hasard dans la strate. Pour une strate h donnée, le taux d’échantillonnage à ce niveau, désigné par f1h est :
h
hh S
Rnf
²1
⋅⋅=
π (1)
avec : nh = nombre de points dans la strate
Sh = Surface totale de la strate
R = Rayon d’activité du bateau de prélèvement autour des coordonnées du point moyen demandé (« point théorique »). On suppose, en fait, que le navire travaille dans un cercle autour d’un point. Ce rayon est une fonction croissante de la longueur du bateau et de la fiabilité des moyens de navigation mis en œuvre (ici GPS) ; le rôle du patron (aptitude de navigation, adaptation aux appareils, etc.) doit également intervenir, mais il est difficile à quantifier. En l’absence d’éléments précis, la valeur de ce rayon était fixée arbitrairement à 25 mètres. En 2003, nous avons ramené ce rayon à 12 mètres car il correspond à la longueur du navire utilisé pour la campagne.
Unités secondaires : les unités secondaires correspondent aux coups de benne Hamon réalisés par point. Dans la strate h, le taux d’échantillonnage à ce niveau, désigné par f2 est égal à :
22 4 Rkf⋅⋅
=π
(2)
avec : k = nombre de coups de benne par point (ou station), fixé ici à deux.
R = rayon d’activité
Septembre 2003

50
Nombre d’échantillons par niveau
Premier niveau : soit Nh le nombre total de points réalisables dans la strate. On doit avoir :
h
h
h
hh N
nS
Rnf =⋅⋅
=2
1π et donc 2R
SN hh ⋅=π
(3)
La probabilité d’intersection entre deux unités d’échantillonnage a été considérée comme nulle.
Deuxième niveau : le nombre potentiel d’unités d’échantillonnage est : 24 R⋅⋅π
Estimateurs
Pour une strate h donnée, l’estimateur de l’effectif moyen ou de la
biomasse moyenne sur 0.25 m2 représenté par est : =
hx
h
n
i
k
jijh
h
n
iih
h nk
x
n
xx
hh
⋅==∑∑∑= ==
=1 11 (4)
xijh est l’effectif capturé par coup de benne j, réalisé sur le point i de la strate h.
Pour le calcul d’une estimation de la variance associée, on fera appel aux notions d’espérance et de variance conditionnelles. La formulation générale du problème (Cochran, 1977) donne (E désignant l’espérance mathématique, V la variance et un estimateur) : θ̂
( ) ( )( ) ( )( )θθθ ˆˆˆ2121 VEEV +=V (5)
La variance est constituée de deux termes. Le premier se rapporte à la variance entre les unités primaires (variance inter-unité), le second se réfère aux unités secondaires au sein des unités primaires (variance intra-unité).
Dans notre cas, le premier terme à droite de l’équation (5) est :
h
h
h
hh n
SS
RnxE21
2
21 1 ⋅
⋅⋅−=
= πV (6)
Septembre 2003

51
En considérant que l’échantillonnage au deuxième niveau est aléatoire simple, on obtient :
k
S
nRkxV
hn
iih
hh
∑=
=
⋅⋅
⋅⋅−=
1
22
2221
41
π (7)
E1 de l’équation (5) se présente comme suit :
22
1
22
1
22
1 hh
N
iih
h
n
iih
SN
s
n
sE
hh
==
∑∑== (8)
donc :
h
hh nk
SR
kxVE⋅
⋅
⋅⋅−=
= 2
2221 4
1π
(9)
Finalement, la variance de l’estimateur est égale à :
h
h
h
h
h
hh nk
SR
knS
SRnxV
⋅⋅
⋅⋅−+⋅
⋅⋅−=
= 2
22
21
2
411
ππ (10)
Pour estimer cette variance à partir de l’échantillon existant, il est nécessaire de disposer d’une estimation non biaisée de et . Cochran (1977) montre qu’un estimateur non biaisé de est donné par :
21hS 2
2hS21hS
k
sR
ks hh
22
221 4
1 ⋅
⋅⋅−−
π (11)
avec :
11
2
21 −
−
=∑=
=
h
n
ihih
h n
xxs
h
(12)
Septembre 2003

52
( )
( )11 1
2
22 −⋅
−=∑∑= =
kn
xxs
h
n
i
k
jihh
h
h
(13)
Finalement, l’expression de la variance de l’estimateur s’écrit :
h
hé
h
h
h
h
h
hh nk
sR
kS
Rnns
SRnxV
⋅⋅
⋅⋅−⋅
⋅⋅+⋅
⋅⋅−=
= 2
222
12
411
πππ
(14)
L’estimateur effectif (ou biomasse) total(e) sur la strate h, désigné par Th est donné par :
(15) =
⋅⋅= hhh xST 4
et la variance correspondante est égale à :
V (16) ( )
⋅⋅=
=
hhh xVST 216
Pour les strates avec un prélèvement par station, le taux d’échantillonnage est simple (échantillonnage structuré à un seul niveau) et conduit à de nombreuses simplifications. Avec les notations présentées précédemment, l’estimateur de l’effectif moyen ou de la biomasse moyenne sur 0.25 m² est alors :
h
n
ii
hn
xx
h
∑== 1 (17)
xi = effectif ou biomasse par coup de benne
nh = nombre de points dans la strate h
Septembre 2003

53
Pour la variance, seule subsiste la variance entre les unités primaires (variance inter-unités) et le rayon d’action du navire n’intervient plus dans le calcul. Dans ces conditions, la variance est :
( )h
hh
ns
x2
=V (18)
sh = variance relative à la strate h
L’estimateur effectif (ou biomasse) total(e) sur la strate h et la variance correspondante, sont par conséquent :
hhh xS ⋅⋅= 4T (19)
( )
⋅⋅= hhh xVST 216V (20)
Septembre 2003
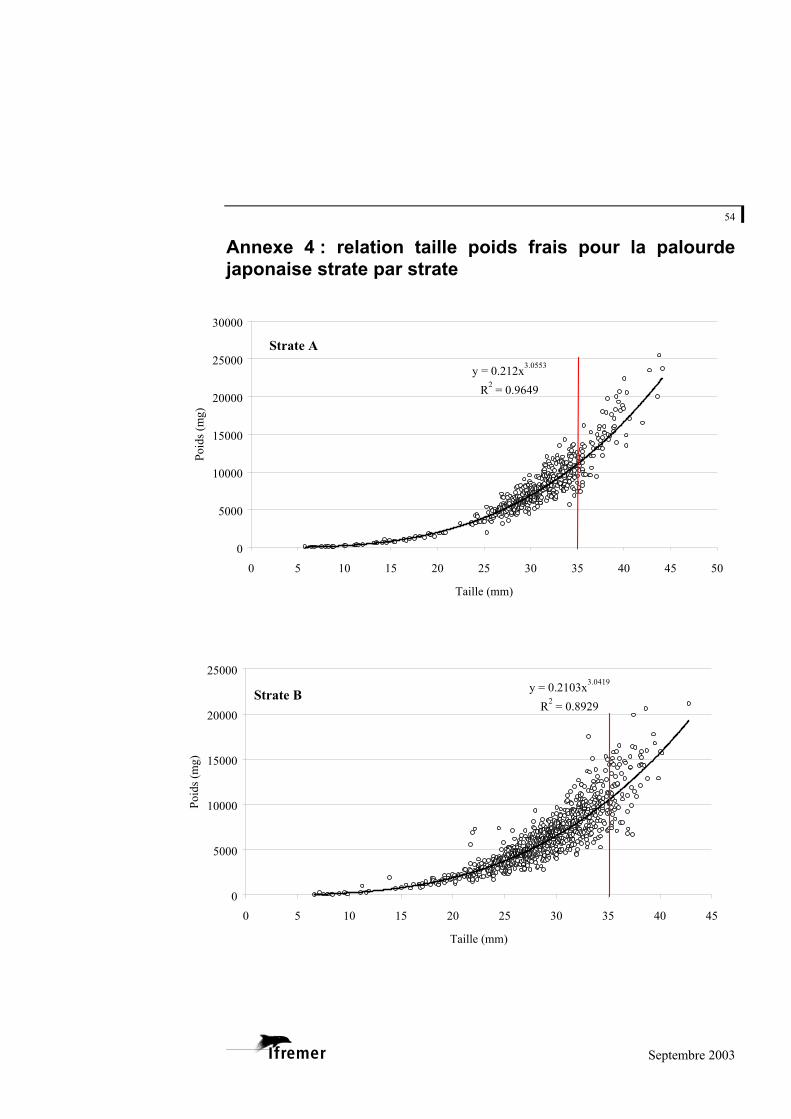
54
Annexe 4 : relation taille poids frais pour la palourde japonaise strate par strate
Strate A
y = 0.212x3.0553
R2 = 0.9649
0
5000
10000
15000
20000
25000
30000
0 5 10 15 20 25 30 35 40 45 50
Taille (mm)
Poid
s (m
g)
Strate B y = 0.2103x3.0419
R2 = 0.8929
5000
10000
15000
20000
25000
0 5 10 15 20 25 30 35 40 45
Taille (mm)
Poid
s (m
g)
0
Septembre 2003
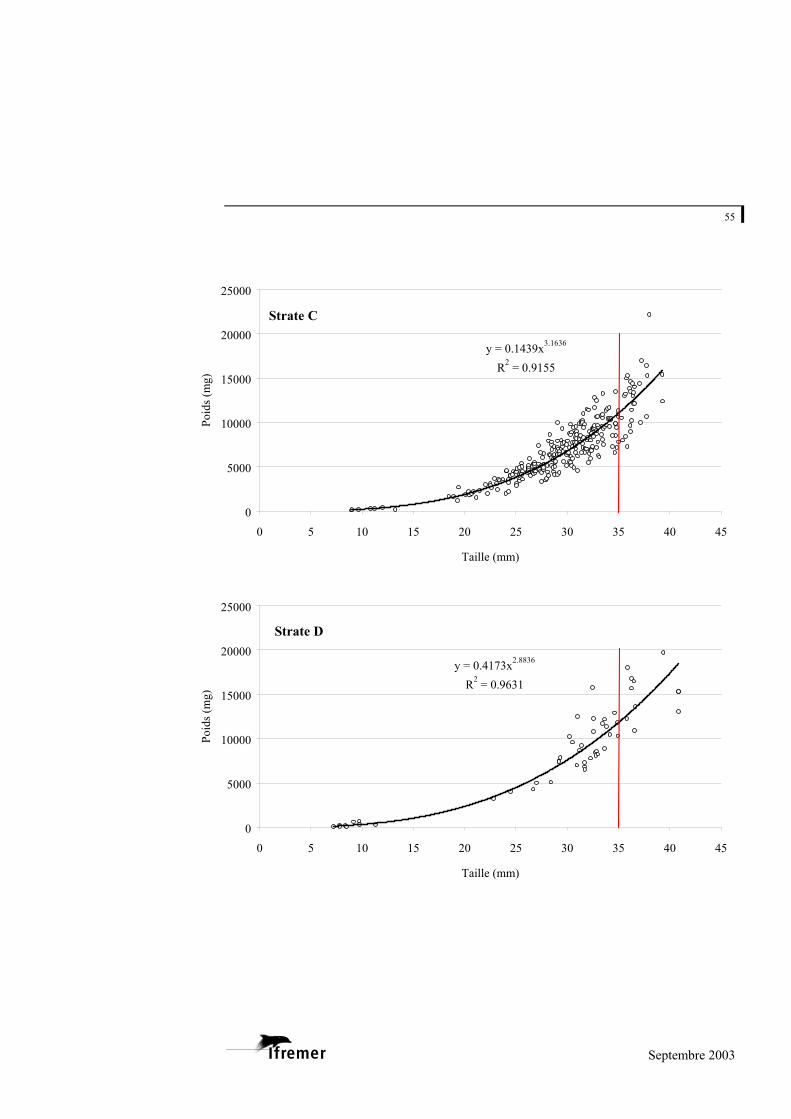
55
Strate C
y = 0.1439x3.1636
R2 = 0.9155
0
5000
10000
15000
20000
25000
0 5 10 15 20 25 30 35 40 45
Taille (mm)
Poid
s (m
g)
Strate D
y = 0.4173x2.8836
R2 = 0.9631
0
5000
10000
15000
20000
25000
0 5 10 15 20 25 30 35 40 45
Taille (mm)
Poid
s (m
g)
Septembre 2003
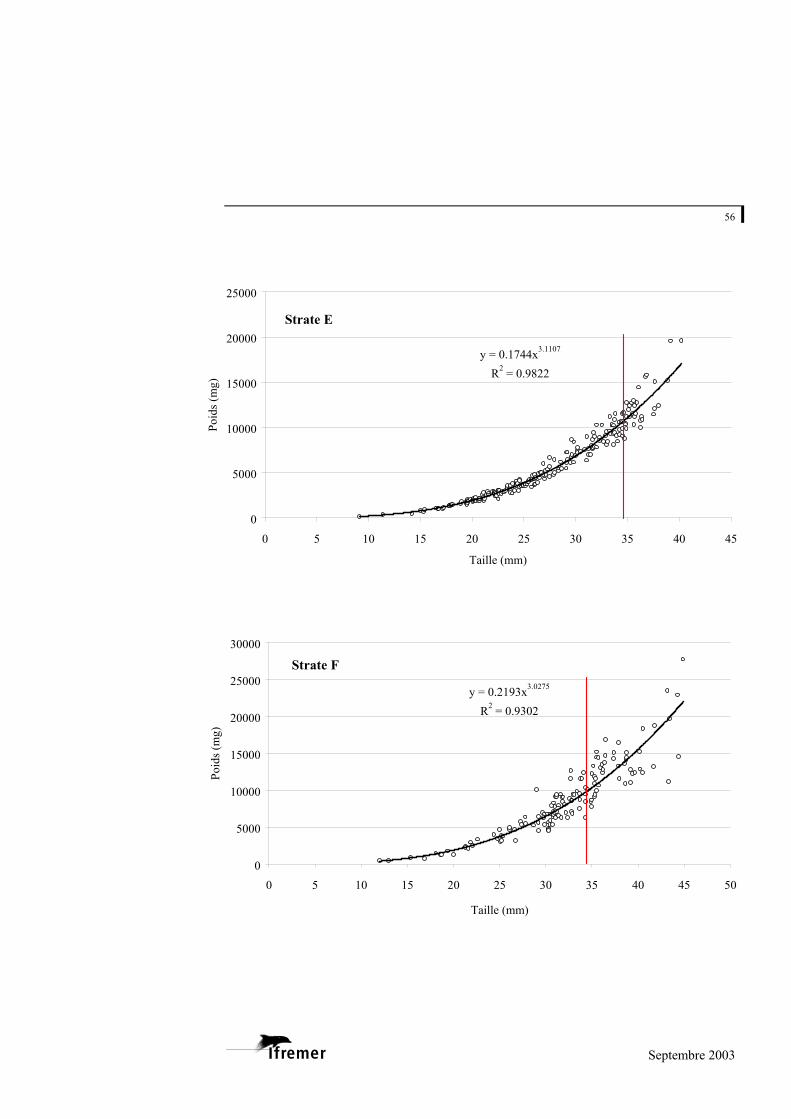
56
Strate E
y = 0.1744x3.1107
R2 = 0.9822
0
5000
10000
15000
20000
25000
0 5 10 15 20 25 30 35 40 45
Taille (mm)
Poid
s (m
g)
30000
25000
20000 15000 10000
Strate F
y = 0.2193x3.0275
R2 = 0.9302
0
5000
0 5 10 15 20 25 30 35 40 45 50
Taille (mm)
Poid
s (m
g)
Septembre 2003
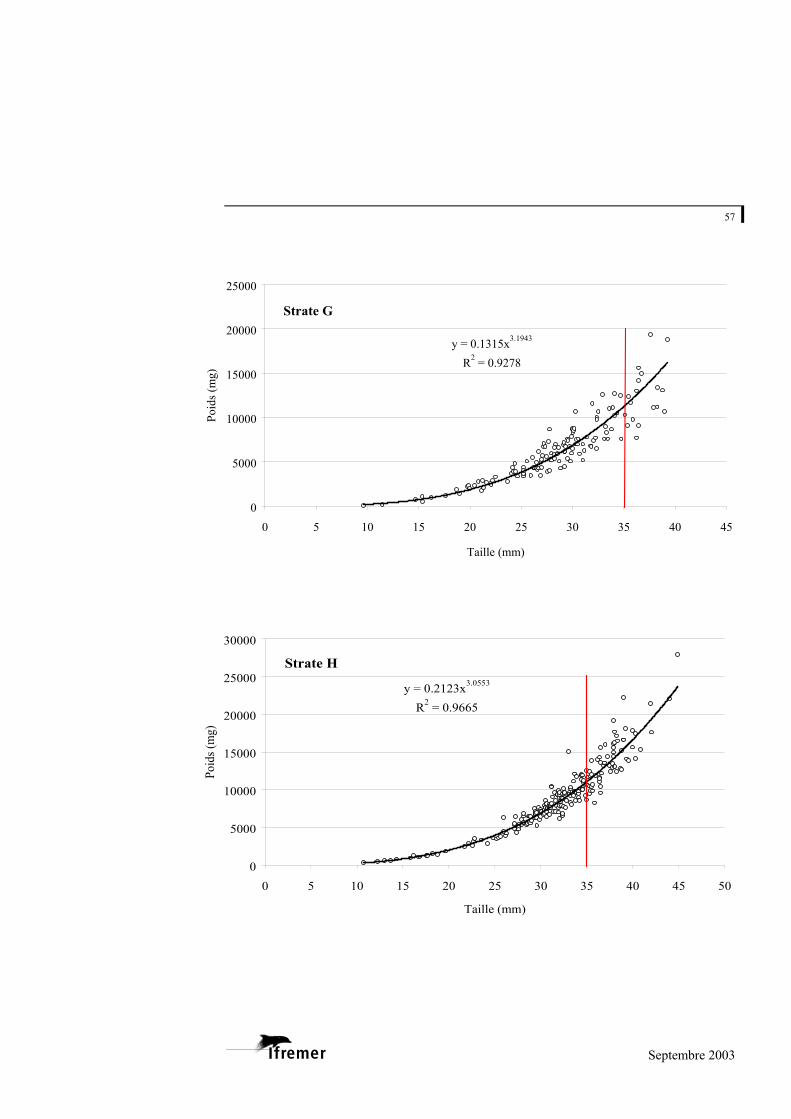
57
Strate G
y = 0.1315x3.1943
R2 = 0.9278
0
5000
10000
15000
20000
25000
0 5 10 15 20 25 30 35 40 45
Taille (mm)
Poid
s (m
g)
500
Strate H
y = 0.2123x3.0553
R2 = 0.9665
0
0
10000
15000
20000
25000
30000
0 5 10 15 20 25 30 35 40 45 50
Taille (mm)
Poid
s (m
g)
Septembre 2003
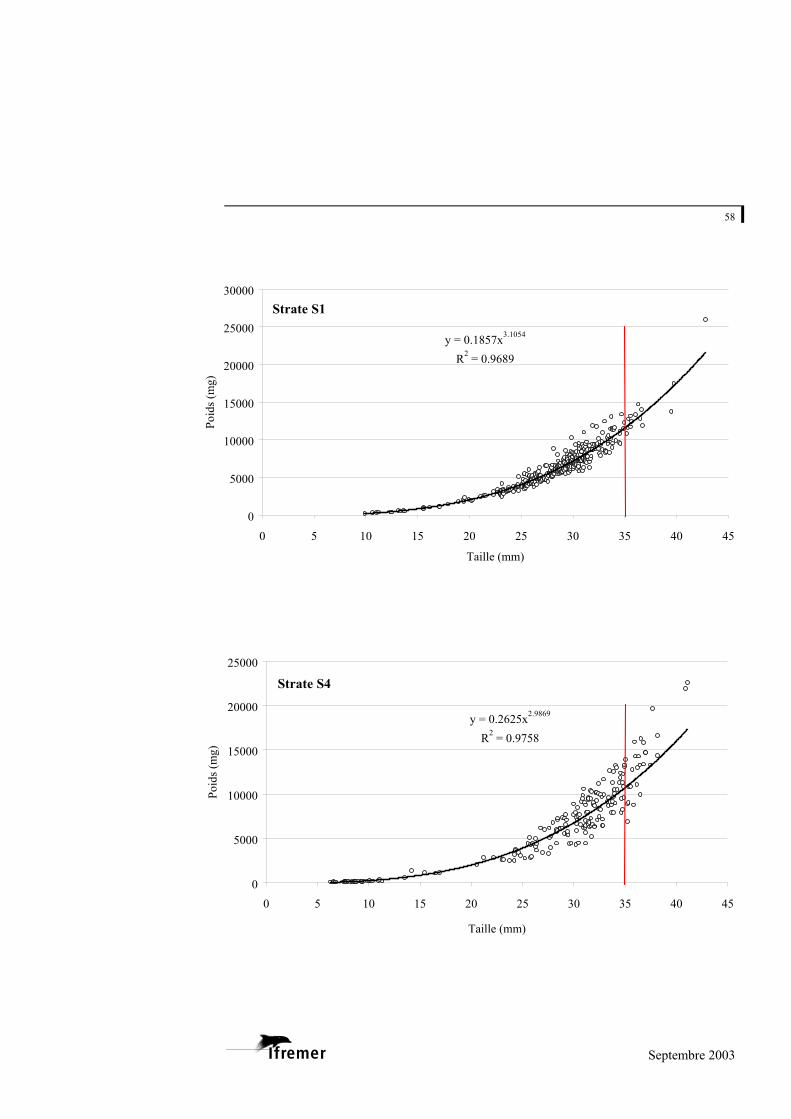
58
Strate S1
y = 0.1857x3.1054
R2 = 0.9689
0
10000
15000
20000
25000
30000
0 5 10 15 20 25 30 35 40 45
Taille (mm)
Poid
s (m
g)
5000
25000
20000 15000 10000
Strate S4
y = 0.2625x2.9869
R2 = 0.9758
0
5000
0 5 10 15 20 25 30 35 40 45
Taille (mm)
Poid
s (m
g)
Septembre 2003
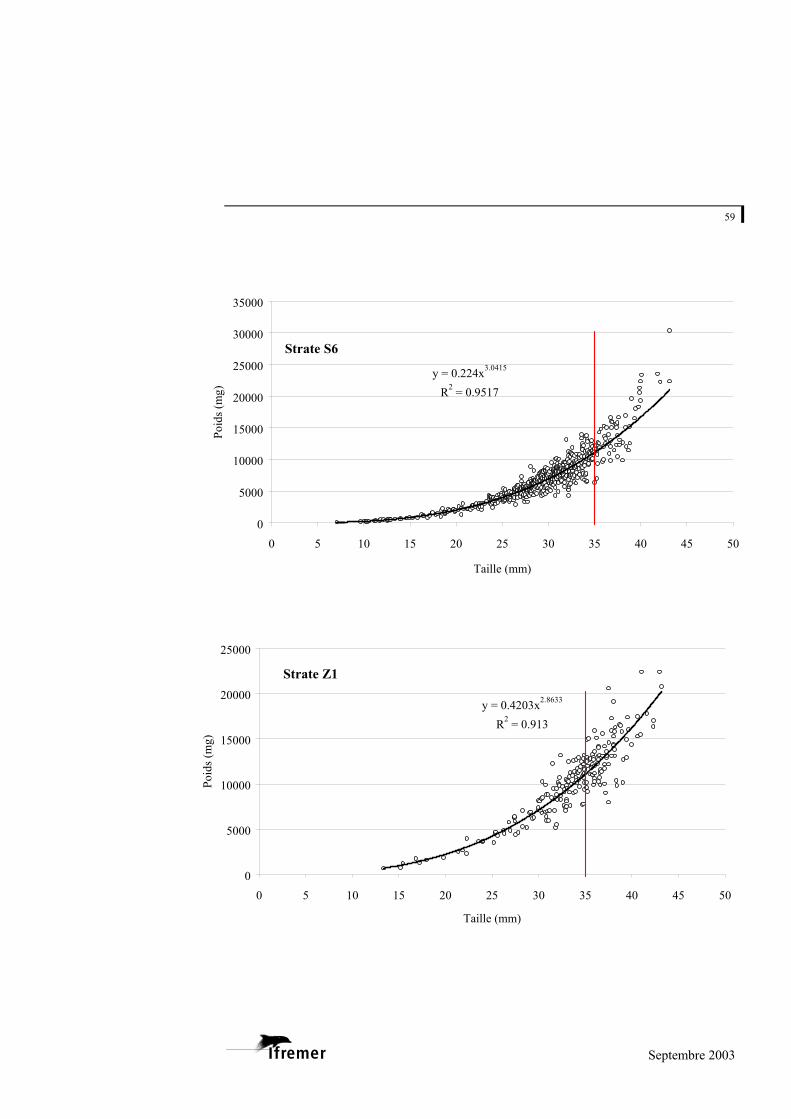
59
Strate S6
y = 0.224x3.0415
R2 = 0.9517
0
5000
10000
15000
20000
25000
30000
35000
0 5 10 15 20 25 30 35 40 45 50
Taille (mm)
Poid
s (m
g)
Strate Z1
y = 0.4203x2.8633
R2 = 0.913
0
5000
10000
15000
20000
25000
0 5 10 15 20 25 30 35 40 45 50
Taille (mm)
Poid
s (m
g)
Septembre 2003
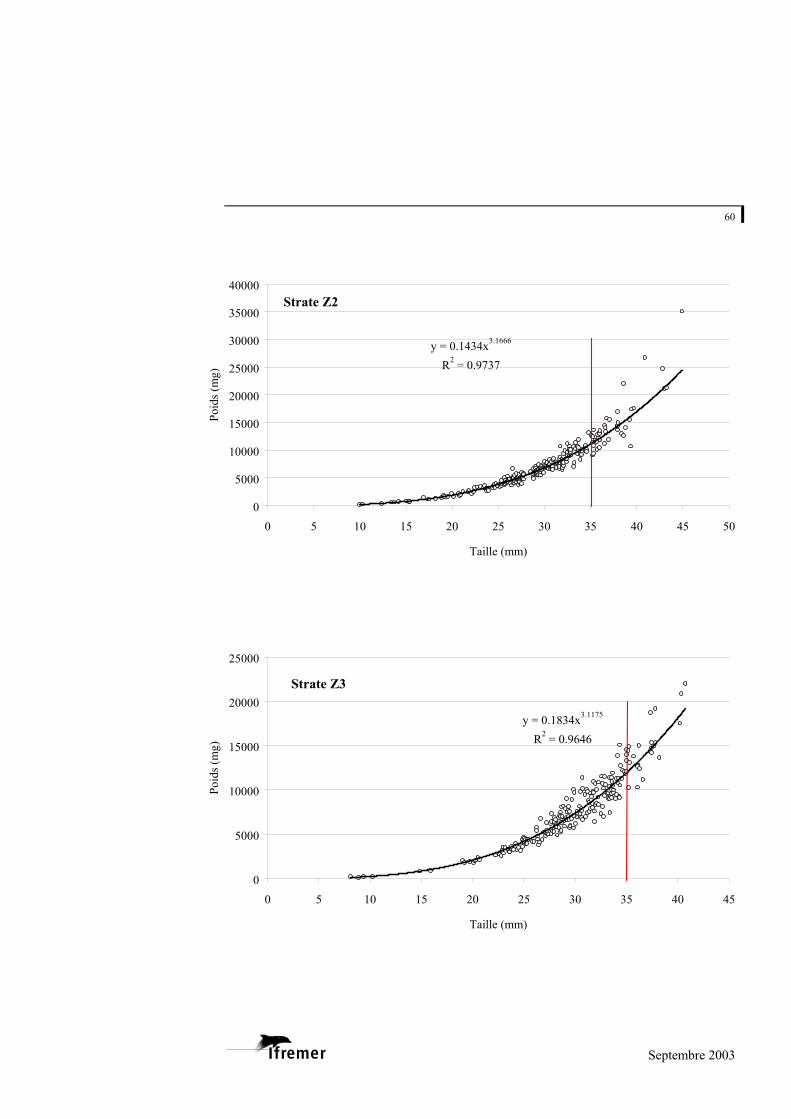
60
Strate Z2
y = 0.1434x3.1666
R2 = 0.9737
0
5000
0 5 10 15 20 25 30 35 40 45 50
Taille (mm)
Poid
s (m
g)
40000
35000 30000
25000 20000 15000
10000
Strate Z3
y = 0.1834x3.1175
R2 = 0.9646
0
5000
10000
15000
20000
25000
0 5 10 15 20 25 30 35 40 45
Taille (mm)
Poid
s (m
g)
Septembre 2003
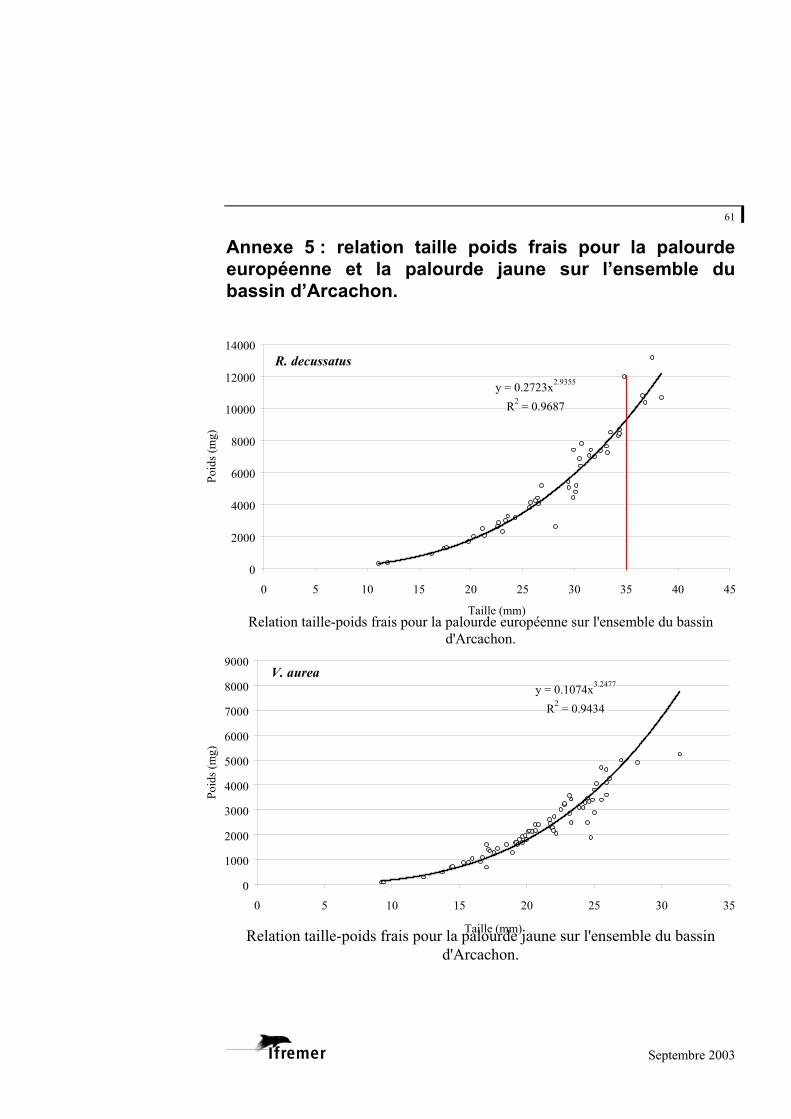
61
Annexe 5 : relation taille poids frais pour la palourde européenne et la palourde jaune sur l’ensemble du bassin d’Arcachon.
R. decu
8000
6000
Relation taille-poids frais pour la palourde européenne sur l'ensemble du bassin
d'Arcachon.
ssatus
y = 0.2723x2.9355
R2 = 0.9687
0
2000
4000
10000
12000
14000
0 5 10 15 20 25 30 35 40 45
Taille (mm)
Poid
s (m
g)
V. aureay = 0.1074x3.2477
R2 = 0.9434
0
1000
2000
3000
4000
5000
6000
7000
8000
9000
0 5 10 15 20 25 30 35
Poid
s (m
g)
Relation taille-poids frais pour la palourde jaune sur l'ensemble du bassin d'Arcachon.
Taille (mm)
Septembre 2003
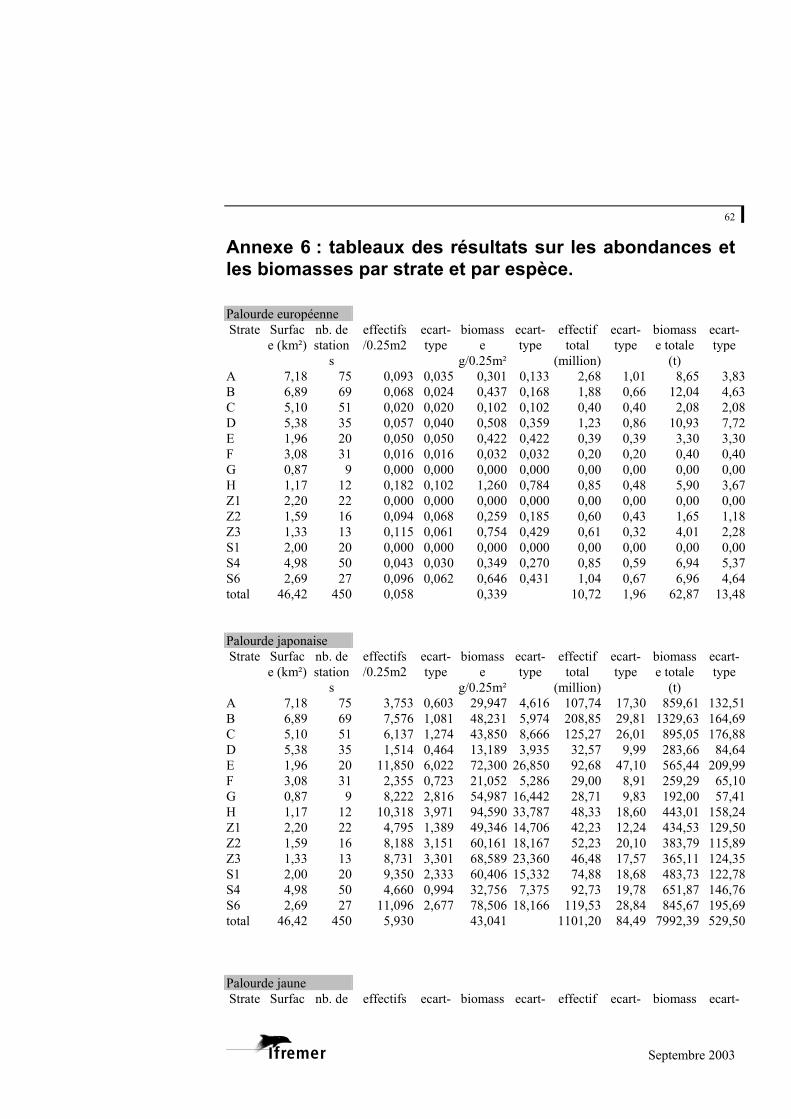
62
Annexe 6 : tableaux des résultats sur les abondances et les biomasses par strate et par espèce.
Palourde européenne Strate Surfac
e (km²) nb. de station
s
effectifs /0.25m2
ecart-type
biomasse
g/0.25m²
ecart-type
effectif total
(million)
ecart-type
biomasse totale
(t)
ecart-type
A 7,18 75 0,093 0,035 0,301 0,133 2,68 1,01 8,65 3,83B 6,89 69 0,068 0,024 0,437 0,168 1,88 0,66 12,04 4,63C 5,10 51 0,020 0,020 0,102 0,102 0,40 0,40 2,08 2,08D 5,38 35 0,057 0,040 0,508 0,359 1,23 0,86 10,93 7,72E 1,96 20 0,050 0,050 0,422 0,422 0,39 0,39 3,30 3,30F 3,08 31 0,016 0,016 0,032 0,032 0,20 0,20 0,40 0,40G 0,87 9 0,000 0,000 0,000 0,000 0,00 0,00 0,00 0,00H 1,17 12 0,182 0,102 1,260 0,784 0,85 0,48 5,90 3,67Z1 2,20 22 0,000 0,000 0,000 0,000 0,00 0,00 0,00 0,00Z2 1,59 16 0,094 0,068 0,259 0,185 0,60 0,43 1,65 1,18Z3 1,33 13 0,115 0,061 0,754 0,429 0,61 0,32 4,01 2,28S1 2,00 20 0,000 0,000 0,000 0,000 0,00 0,00 0,00 0,00S4 4,98 50 0,043 0,030 0,349 0,270 0,85 0,59 6,94 5,37S6 2,69 27 0,096 0,062 0,646 0,431 1,04 0,67 6,96 4,64total 46,42 450 0,058 0,339 10,72 1,96 62,87 13,48 Palourde japonaise Strate Surfac
e (km²) nb. de station
s
effectifs /0.25m2
ecart-type
biomasse
g/0.25m²
ecart-type
effectif total
(million)
ecart-type
biomasse totale
(t)
ecart-type
A 7,18 75 3,753 0,603 29,947 4,616 107,74 17,30 859,61 132,51B 6,89 69 7,576 1,081 48,231 5,974 208,85 29,81 1329,63 164,69C 5,10 51 6,137 1,274 43,850 8,666 125,27 26,01 895,05 176,88D 5,38 35 1,514 0,464 13,189 3,935 32,57 9,99 283,66 84,64E 1,96 20 11,850 6,022 72,300 26,850 92,68 47,10 565,44 209,99F 3,08 31 2,355 0,723 21,052 5,286 29,00 8,91 259,29 65,10G 0,87 9 8,222 2,816 54,987 16,442 28,71 9,83 192,00 57,41H 1,17 12 10,318 3,971 94,590 33,787 48,33 18,60 443,01 158,24Z1 2,20 22 4,795 1,389 49,346 14,706 42,23 12,24 434,53 129,50Z2 1,59 16 8,188 3,151 60,161 18,167 52,23 20,10 383,79 115,89Z3 1,33 13 8,731 3,301 68,589 23,360 46,48 17,57 365,11 124,35S1 2,00 20 9,350 2,333 60,406 15,332 74,88 18,68 483,73 122,78S4 4,98 50 4,660 0,994 32,756 7,375 92,73 19,78 651,87 146,76S6 2,69 27 11,096 2,677 78,506 18,166 119,53 28,84 845,67 195,69total 46,42 450 5,930 43,041 1101,20 84,49 7992,39 529,50 Palourde jaune Strate Surfac nb. de effectifs ecart- biomass ecart- effectif ecart- biomass ecart-
Septembre 2003
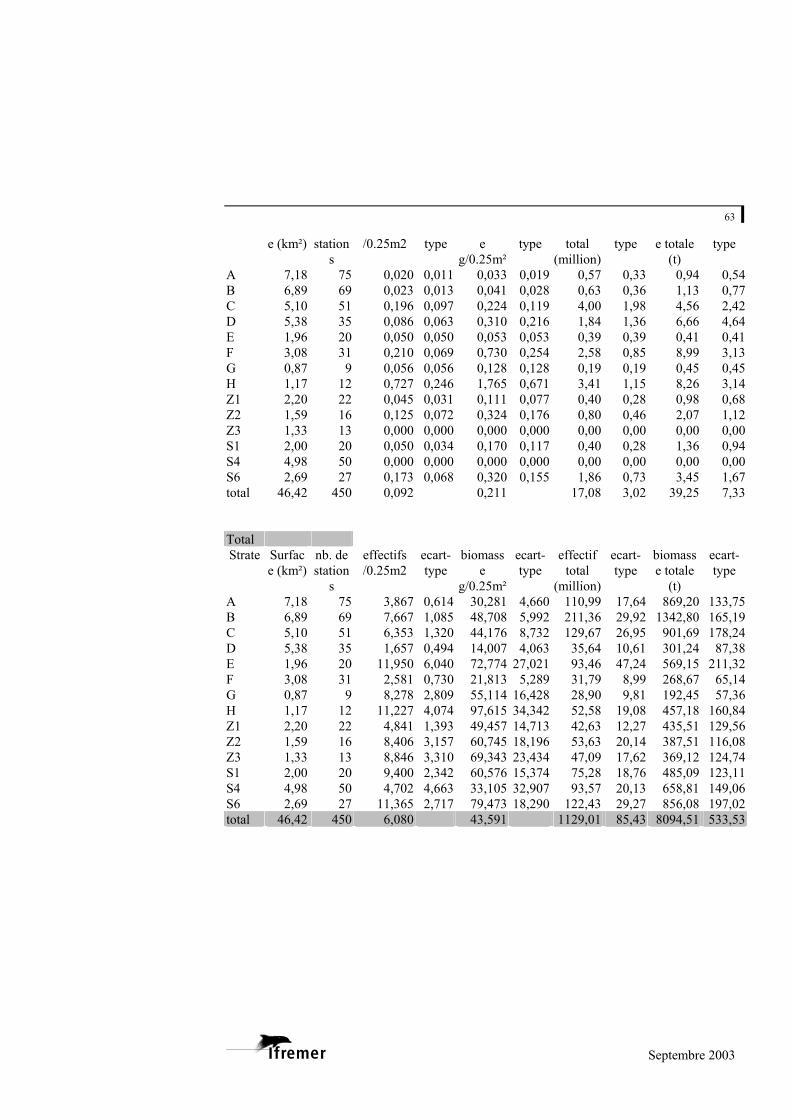
63
e (km²) stations
/0.25m2 type e g/0.25m²
type total (million)
type e totale (t)
type
A 7,18 75 0,020 0,011 0,033 0,019 0,57 0,33 0,94 0,54B 6,89 69 0,023 0,013 0,041 0,028 0,63 0,36 1,13 0,77C 5,10 51 0,196 0,097 0,224 0,119 4,00 1,98 4,56 2,42D 5,38 35 0,086 0,063 0,310 0,216 1,84 1,36 6,66 4,64E 1,96 20 0,050 0,050 0,053 0,053 0,39 0,39 0,41 0,41F 3,08 31 0,210 0,069 0,730 0,254 2,58 0,85 8,99 3,13G 0,87 9 0,056 0,056 0,128 0,128 0,19 0,19 0,45 0,45H 1,17 12 0,727 0,246 1,765 0,671 3,41 1,15 8,26 3,14Z1 2,20 22 0,045 0,031 0,111 0,077 0,40 0,28 0,98 0,68Z2 1,59 16 0,125 0,072 0,324 0,176 0,80 0,46 2,07 1,12Z3 1,33 13 0,000 0,000 0,000 0,000 0,00 0,00 0,00 0,00S1 2,00 20 0,050 0,034 0,170 0,117 0,40 0,28 1,36 0,94S4 4,98 50 0,000 0,000 0,000 0,000 0,00 0,00 0,00 0,00S6 2,69 27 0,173 0,068 0,320 0,155 1,86 0,73 3,45 1,67total 46,42 450 0,092 0,211 17,08 3,02 39,25 7,33 Total Strate Surfac
e (km²) nb. de station
s
effectifs /0.25m2
ecart-type
biomasse
g/0.25m²
ecart-type
effectif total
(million)
ecart-type
biomasse totale
(t)
ecart-type
A 7,18 75 3,867 0,614 30,281 4,660 110,99 17,64 869,20 133,75B 6,89 69 7,667 1,085 48,708 5,992 211,36 29,92 1342,80 165,19C 5,10 51 6,353 1,320 44,176 8,732 129,67 26,95 901,69 178,24D 5,38 35 1,657 0,494 14,007 4,063 35,64 10,61 301,24 87,38E 1,96 20 11,950 6,040 72,774 27,021 93,46 47,24 569,15 211,32F 3,08 31 2,581 0,730 21,813 5,289 31,79 8,99 268,67 65,14G 0,87 9 8,278 2,809 55,114 16,428 28,90 9,81 192,45 57,36H 1,17 12 11,227 4,074 97,615 34,342 52,58 19,08 457,18 160,84Z1 2,20 22 4,841 1,393 49,457 14,713 42,63 12,27 435,51 129,56Z2 1,59 16 8,406 3,157 60,745 18,196 53,63 20,14 387,51 116,08Z3 1,33 13 8,846 3,310 69,343 23,434 47,09 17,62 369,12 124,74S1 2,00 20 9,400 2,342 60,576 15,374 75,28 18,76 485,09 123,11S4 4,98 50 4,702 4,663 33,105 32,907 93,57 20,13 658,81 149,06S6 2,69 27 11,365 2,717 79,473 18,290 122,43 29,27 856,08 197,02total 46,42 450 6,080 43,591 1129,01 85,43 8094,51 533,53
Septembre 2003
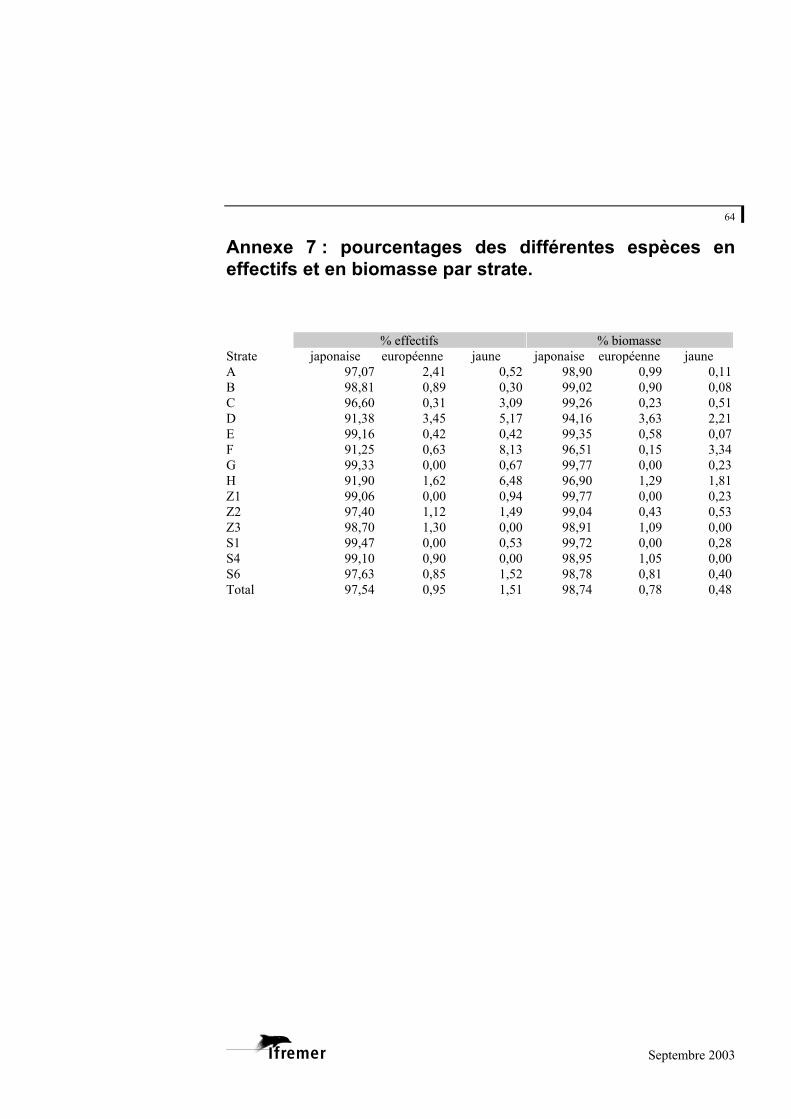
64
Septembre 2003
Annexe 7 : pourcentages des différentes espèces en effectifs et en biomasse par strate.
% effectifs % biomasse
Strate japonaise européenne jaune japonaise européenne jaune A 97,07 2,41 0,52 98,90 0,99 0,11B 98,81 0,89 0,30 99,02 0,90 0,08C 96,60 0,31 3,09 99,26 0,23 0,51D 91,38 3,45 5,17 94,16 3,63 2,21E 99,16 0,42 0,42 99,35 0,58 0,07F 91,25 0,63 8,13 96,51 0,15 3,34G 99,33 0,00 0,67 99,77 0,00 0,23H 91,90 1,62 6,48 96,90 1,29 1,81Z1 99,06 0,00 0,94 99,77 0,00 0,23Z2 97,40 1,12 1,49 99,04 0,43 0,53Z3 98,70 1,30 0,00 98,91 1,09 0,00S1 99,47 0,00 0,53 99,72 0,00 0,28S4 99,10 0,90 0,00 98,95 1,05 0,00S6 97,63 0,85 1,52 98,78 0,81 0,40Total 97,54 0,95 1,51 98,74 0,78 0,48