Embed Size (px)
Citation preview

Tous droits réservés au Tutorat Associatif Toulousain 1/26
CONCOURS BLANC PACES
SEMESTRE I – 2014-2015
MARAICHERS
CORRECTIONS DETAILLEES
COMPILATION FAITE PAR LE TUTORAT ASSOCIATIF TOULOUSAIN
Avec l’accord préalable de la Scolarité de la Faculté des Sciences
Pharmaceutiques de Toulouse

Tous droits réservés au Tutorat Associatif Toulousain 2/26
SOMMAIRE
UE1 – Chimie ................................................................................................................ 3
UE1 – Biomolécules ...................................................................................................... 6
UE1 – Génome .............................................................................................................. 8
UE2 – Biologie cellulaire, BDR, Embryologie, BDD, Histologie ............................. 11
UE3 – Biophysique ..................................................................................................... 14
UE4 – Biostatistiques .................................................................................................. 20
Remerciements ............................................................................................................ 26

Tous droits réservés au Tutorat Associatif Toulousain 3/26
UE1 – Chimie QCM 1 – CE A. Deux isotopes diffèrent par leur nombre de neutrons (et donc pas leur nombre de masse).
B. Deux isotopes diffèrent par leur nombre de protons.
C. Noyau = protons = charge positive.
D. 7 protons et 8 neutrons.
E. Même élément avec augmentation de A = augmentation du nombre de neutrons.
QCM 2 – C A. 0 ≤ l ≤ n-1.
B. 0 ≤ l ≤ n-1.
D. Une couche électronique peut contenir au maximum 2n2 électrons. E. Une orbitale atomique « 2p » car l = 1.
QCM 3 – ACD B. L’atome de soufre ne possède aucun électron « s » célibataire, les 2 célibataires sont « p ».
C. Attention à ne pas oublier les électrons p remplissant la couche n = 2.
D. Ils en ont 2 célibataires chacun.
E. L’atome de silicium possède une seule orbitale atomique « p » vacante.
QCM 4 – B A. Cette configuration respecte la règle de Klechkowski.
B. C’est le 25Mn C. Cet atome possède 7 électrons de valence.
D. Groupe 7.
E. 4ème période.
QCM 5 – ABCD
D. Be et Sr sont des métaux. E. Be et Sr appartiennent au groupe des alcalino-terreux.
QCM 6 – AB
A. Un électron aura tendance à être sur les couches les plus proches du noyau pour se stabiliser.
B. Utiliser la formule rn = r1 x n² Angstrom C. E3 = -13,6/32 eV D. Un rayonnement appartenant à la série de Paschen correspond au retour de l’électron sur le niveau n = 3 or
ici l’atome d’hydrogène est déjà dans un état correspondant au niveau n = 3.
E. Appartient aux ultraviolets.
QCM 7 – AC B. Orbitale moléculaire anti liante de type π*.
D. L’ordre de liaison de cette molécule est égal à 1 ( (8-6)/2 ). E. Cette molécule est diamagnétique (pas d’électrons célibataires dans le diagramme LCAO).
QCM 8 – ABDE
A. C’est une question de stabilité, plus l’énergie d’un système est faible plus il est stable. La formation d’une
liaison ne se produit généralement que dans ces cas-là. C. Une liaison σ est plus stable qu’une liaison π.

Tous droits réservés au Tutorat Associatif Toulousain 4/26
QCM 9 – CE
A. Attention à bien différencier les figures de répulsions, qui donnent une idée générale, et la géométrie de la
molécule en particulier. Ici, la molécule ayant une géométrie tétraédrique est SiH4. B. Seulement 2 espèces sont linéaires.
C. PCl3. D. Toutes ces molécules ne présentent pas la même figure de répulsion.
E. PCl3.
QCM 10 – BCD A. Cette molécule possède une géométrie pyramidale.
E. Dans cette molécule, l’atome d’azote respecte la règle de l’octet (3 liaisons + 1DNL, au total
mise en commun de 8 électrons).
QCM 11 – BC A. Cette réaction s’accompagne d’une diminution de l’entropie car baisse du nombre de moles gazeuses.
B. ΔrH° < 0 donc exothermique.
C. On passe de 4 d’un côté à 1 de l’autre donc - 3. D. Pour cette réaction effectuée à pression constante, la variation d’énergie interne ΔU = ΔH-ΔnRT (avec Δn
variation du nombre de moles de gaz qui est égale à -3).
E. Une augmentation de la température favorise la réaction dans le sens endothermique, donc ici le sens in-
verse.
QCM 12 – AD B. Mn (+VII), O (-II).
C. H (+I), O (-I) car péroxyde d’hydrogène /!\ Attention cas particulier ! E. Na (+I), H (-I) car hydrure.
QCM 13 – ACD
A. On le sait car il y a une modification des nombres d’oxydation pendant la réaction. B. Cl2 + 2e- = 2Cl- C. La réaction suivante est une réaction d’oxydation (le nombre d’oxydation de (N) augmente : il passe de
(+III) dans NO2- à (+V) dans NO3
-).
D. Relation à savoir ++, c’est elle qui explique aussi notamment pourquoi une réaction d’oxydo-réduction est
toujours spontanée. E. Dans une réaction d’oxydo-réduction, l’oxydant le plus fort réagit sur le réducteur le plus fort.
QCM 14 – ABDE A. Cu est l’élément réducteur car il va subir une oxydation (augmentation de son nombre d’oxydation). C. Cu2+ + 2e- = Cu. Et Mn2+ + 2e- = Mn. Le nombre d’électrons échangés au cours de la réaction spontanée est égale à 2.
D. ΔE° > 0 donc ΔrG° < 0 donc K > 1.
E. Mn est l’élément réducteur car il va subir une oxydation (augmentation de son nombre d’oxydation ; d’une autre façon il suffit de regarder avec les couples : Cu2+ va réagir avec Mn, l’oxydant le plus fort réagis-
sant avec le réducteur le plus fort. Au final ça va donner du Cu et du Mn2+. Le nombre d’oxydation du Mn a
augmenté, passant de 0 a +II pour le Mn2+. Au cours de cette réaction, on peut donc bien dire que le Mn a été
oxydé.

Tous droits réservés au Tutorat Associatif Toulousain 5/26
QCM 15 – D A. Elle est de configuration 2R, 3R, 5Z, 7E.
B. Elle possède 2 fonctions alcools et une fonction acide.
C. Elle en possède 5 : ceux des fonctions alcène, soit 4, plus celui engagé dans la fonction acide carboxylique.
E. C’est l’acide (2R, 3R, 5Z, 7E)-5-fluoro-2,3-dihydroxy-2-méthyl-nona-5,7-diène-oïque.
QCM 16 – ABCE
A. Il présente un plan de symétrie.
B. 2 méthyl d’un côté rends cette molécule achirale (attention, si il y avait eu méthyl et éthyl par exemple, la molécule aurait été chirale : les cumulènes pairs peuvent être chiraux). D. Le composé 3 est thréo.
QCM 17 – BD (Mon conseil lorsque vous vous retrouvez face à ce genre de qcms posant des questions sur ce type de molécules : tout passer dans une même représentation type Fischer, et tout déterminer d’un coup : R, S, diastéréoisomères, … ça permet d’aller beaucoup plus vite ensuite). A. La molécule A est le (3R, 4R)-5-hydroxy-3,4-diméthyl-pentan-2-one.
C. La molécule D n’est pas la représentation de la molécule B en projection de Fischer.
E. Les molécules B et C sont identiques.
QCM 18 – BC Classement global : 1 > 4 > 6 > 2 > 3 > 5.
Rappel : l’eau est plus acide que l’alcool ; un acide minéral type HCl ou HF par exemple sera toujours plus
acide qu’un acide carboxylique ; la proximité d’un groupement présentant un effet inductif attracteur sera plus
importante dans la détermination de l’acidité que le nombre, comme ici entre 4 et 6 où le Cl est plus proche
dans la 4 : le glissement d’électron se fera plus facilement que dans la 6, même s’il y a 2 Cl.
QCM 19 – ABDE A. En effet l’électronégativité diminue la basicité, F est donc moins basique que OH.
B. Il y a groupement conjugué autorisant une mésomérie pour le cycle, ce qui baisse la basicité.
C. CH3-NH2 > NH3 (car les groupements inductifs donneurs augmentent la basicité). D. En effet l’électronégativité diminue la basicité, la molécule portant 3 Cl est donc plus basique que celle
portant 3 F.
E. L’aniline (aminobenzène) présente une mésomérie avec son DNL pouvant se déplacer dans le cycle, tan-
dis que la pyridine a son doublet bloqué, non inclus dans un système conjugué, donc ne pouvant présenter de
mésomérie. La mésomérie diminuant la basicité d’une manière générale, la proposition 5 est bien fausse, ren-
dant l’item vrai (attention aux négations dans ce genre d’items).
QCM 20 – BD A. Neutralité des charges non respectée, N devrait avoir une charge (+). C. On observe le déplacement d’un hydrogène, or la mésomérie ne concerne pas les déplacements d’atomes
(il n’y a pas de système conjugué dans cette molécule).
D. Dans cette molécule l’oxygène a bien un effet mésomère donneur.
E. Le CH2 devrait porter une charge (-) et le Cl une charge (+) sur la figure de gauche car le Cl a un effet mé-
somère donneur et non pas attracteur.

Tous droits réservés au Tutorat Associatif Toulousain 6/26
UE1 – Biomolécules QCM 1 – CD
A. Il s’agit de la forme pyranose d’un aldopentose.
B. Il s’agit de la forme pyranose d’un aldopentose.
E. Elle est, sous la forme furanose, un constituant de l’ADN.
QCM 2 – B
A. Il est constitué d’une molécule de galactose et d’une molécule de glucose.
C. Il peut être clivé par une βD-galactosidase uniquement.
D. Il possède un pouvoir réducteur de la liqueur de Fehling.
E. Il s’agit d’une molécule de lactose.
QCM 3 – ABE
C. La molécule sous forme cyclique (schéma de droite) est un β-D-fructofuranose.
D. Elles n’entrent pas dans la composition de la partie glycanique des glycoprotéines. Le fructose ne fait pas
partie des 8 oses entrants dans la composition des glycoprotéines.
QCM 4 – ADE
B. Seuls les oses simples répondent à la formule Cn(H2O)n. C. Leur caractère anionique et hydrophile rend les tissus à la fois visqueux et élastiques.
QCM 5 – ACDE B. La formation d’acide ascorbique est impossible chez l’Homme.
QCM 6 –AC B. La glycine est l’acide aminé qui comporte le groupement le plus petit, à savoir un hydrogène. D. La radical de la cystéine n’est pas toujours ionisé à pH physiologique. E. La condensation des N-glycanes à une protéine résulte de la condensation d’un ose avec l’asparagine.
QCM 7 – AC B. Il s’agit d’une structure secondaire essentiellement retrouvée aussi bien dans les protéines globulaires que
dans les protéines fibrillaires. D. Chaque tour d’hélice peut contenir plusieurs liaisons hydrogènes. E. Dans les protéines fibrillaires par exemple, elle sera constituée essentiellement d’AA hydrophobes.
QCM 8 – AD B. Pas la majorité car moins de la moitié des acides aminés du peptide sont des acides aminés polaires chargés
au pH physiologique. C. Les structures secondaires sont maintenus uniquement par des liaisons hydrogènes entre le CO et le NH des
acides aminés participant à la liaison peptidique (pas de liaisons hydrogènes impliquant le radical pour la
stabilisation des structures secondaires). E. Aucun des acides aminés de ce peptide ne peut être glycosylé sur son radical (il n’y a ni Ser ni Thr ni Asn).
QCM 9 – C A. La mise en place de la structure tertiaire d’une protéine globulaire nécessite parfois des protéines chape-
ronnes. B. Item ambigü qui ne sera pas reposé de cette manière. D. Elle est stabilisée par des liaisons hydrophobes à l’intérieur des protéines. E. La structure quaternaire du collagène est constituée de sous-unités stabilisées par des pontages covalents
entre lysines.
QCM 10 – ABCE D. La ninhydrine permet le dosage des acides aminés (pas des protéines).

Tous droits réservés au Tutorat Associatif Toulousain 7/26
QCM 11 – D A. L’acide linolénique est un acide gras à 18C comportant 3 doubles liaisons. B. L’acide lignocérique est un acide gras saturé à 24C. C. Le beurre contient également d’autres types d’acides gras (caproïque, caprylique, caprique). E. Les prostanoïdes ont pour précurseur l’acide arachidonique (C20:4) ou l’acide eicosapentaènoïque (C20:5).
QCM 12 – BCE A. Les prostacyclines (à la différence des lipoxines et des leucotriènes) ne possèdent pas de doubles liaisons
conjuguées. D. L’aspirine inhibe la cyclooxygénase et donc la synthèse des prostaglandines (les leucotriènes sont produits
par lipoxygénase).
QCM 13 – ABE C. Les cardiolipides sont amphiphiles mais pas amphotères. D. Les cérébrosides sont bien amphiphiles mais ce sont des sphingolipides.
QCM 14 – AB C. La vitamine K possède un noyau naphtoquinone (c’est la vitamine E qui possède un résidu phytyl linéaire
saturé). D. L’oestrone possède un noyau oestrane à 18C. E. La vitamine D2 dérive de l’ergostérol.
QCM 15 – D A. La liaison de la protéine avec la chaine lipidique est de type thioéther. B. La protéines est géranylgéranylée (20 carbones). C. Le motif isoprénique permet l’ancrage des protéines à la surface interne de la membrane plasmique. E. Voie de synthèse des isoprènes.
QCM 16 – BCD A. Une liaison riche en énergie est une liaison instable. E. La barrière d’activation permet seulement de ralentir une réaction thermodynamiquement favorable mais
pas de la rendre défavorable.
QCM 17 – A B. Une lysine de l’apoenzyme de l’ALAT forme une base de Schiff avec la vitamine B6 activée (phosphate de
pyridoxal). C. Le katal correspond à la transformation d’une mole de substrat par seconde. D. Chaque amino-transférase reconnait deux substrats. E. Les nucléases (endonucléase et exonucléase) font partie de la classe des hydrolases (EC3).
QCM 18 – AD B. Ce coenzyme est un coenzyme de transférase. C. Ce coenzyme est le Coenzyme A. D. Acétate coenzyme A ligase. E. Ce sont les coenzymes nicotiniques réduits qui absorbent la lumière à 340 nm.
QCM 19 – A B. La vitesse maximale est généralement atteinte après quelques secondes. C. La vitesse maximale d’une enzyme Michaelienne est mesurée dans des conditions d’ordre 0. D. La pente de la représentation de Lineweaver-Burk correspond à Km/Vmax. E. Km n’est pas une constante d’affinité.
QCM 20 – BE A. L’inhibiteur compétitif possède une analogie structurale avec le substrat. C. Les effecteurs allostériques se fixent aussi bien sur les enzymes multimériques que sur les enzymes mono-
mériques. D. Les isoenzymes sont des enzymes Michaeliennes (ce sont des enzymes allostériques).

Tous droits réservés au Tutorat Associatif Toulousain 8/26
UE1 – Génome QCM 1 – BD
A. FAUX : Le déplacement des éléments géniques mobiles (transposons) est très rare, ils ne se déplacent pas
plusieurs fois par cycle cellulaire.
B. VRAI : ici le "au moins une copie" est très important car il ne faut pas oublier les chromosomes sexuels
(XY) qui n'ont pas TOUJOURS les gènes en 2 copies (car le Y est plus petit que X). C. FAUX : L’ADN répété correspond à plus de 10% de l’ensemble du matériel génétique humain.
E. FAUX : Les opérons sont absents du génome nucléaire eucaryote (mais attention il en existe dans la mito-
chondrie, faites attention à l'énoncé on parle bien de génome nucléaire eucaryote).
QCM 2 – E
A. FAUX : Il s’agit d’un nucléoside dérivant d’une base purique (attention à ce genre de piège…). B. FAUX : Elle présente 2 cycles aromatiques.
C. FAUX : Elle ne comprend pas de liaisons riches en énergie car c'est un ribonucléoSide donc Sans phosphate. D. FAUX : La guanosine triphosphate (GTP) peut être un substrat de l’ADN polymérase.
/!\ ATTENTION dans l'énoncé on te parle de la guanosine qui est un NUCLEOSIDE (donc aucun phosphate)
et l'ADN polymérase ne prend que sous forme triphosphate (GTP, CTP, TTP, ATP) donc la guanosine ne peut
pas être son substrat.
E. VRAI : Toutes les bases, ainsi que leurs dérivés, nucléosides ou nucléotides absorbent à 260 nm.
Rappel : cette propriété d'absorption est utilisée pour la détection et le dosage des acides nucléiques.
QCM 3 – C
A. FAUX : La télomérase utilise une amorce d’ADN pour la synthèse de l’ADN télomérique.
B. FAUX : Les télomères n’existent pas chez les procaryotes car leur chromosome bactérien est un ADN
double brin circulaire (pas d'extrémités donc pas de télomères). Remarque : l'ADN plasmidique étant aussi de l'ADN double brin circulaire il n'y a donc pas de télomères.
C. VRAI : la télomérase est exprimée dans : les cellules cancéreuses, les cellules souches et durant l'embryo-
génèse/petite enfance. D. FAUX : La télomérase est une ribonucléoprotéine constituée d’une partie protéique et d’ARN entrant dans
la catégorie des « ARN autres ».
E. FAUX : La télomérase est une transcriptase inverse qui agit également dans les cellules souches, cancé-
reuses et pendant la petite enfance.
QCM 4 – AC
A. VRAI : chez les eucaryotes quand on dit "les transposons" on met aussi dans ce "groupe" les rétrotranspo-
sons qui eux augmentent la taille du génome. On peut donc dire que les transposons (le groupe comprenant les
rétrotransposons) augmentent la taille du génome (PIEGE +++). B. FAUX : Les séquences d’insertions (IS) ne codent que pour la transposase.
C. VRAI : si le transposon se déplace et s'intègre au milieu d'un gène, alors ce gène sera inactivé.
D. FAUX : Les rétrotransposons sont des éléments génétiques mobiles qui ne peuvent pas infecter une autre
cellule.
E. FAUX : Chez les procaryotes, les transposons existent également au niveau de l’ADN plasmidique.
QCM 5 – BC
A. FAUX : Cette molécule est un brin d’ADN.
D. FAUX : La liaison indiquée par une flèche est une liaison N-osidique entre le C1’ du sucre et l’azote numéro
9 de la base.
E. Errata (passe faux) : la liaison entre deux nucléotides est une liaison phosphodiester 3’ vers 5’.
QCM 6 – BCD
A. FAUX : Cette molécule n’est pas complémentaire de la séquence d’ADN qui nous est présentée.
B. VRAI : L'ADN polymérase III est bien active lors de la réplication des procaryotes et elle a besoin d'une
amorce d'ARN.
C. VRAI : Après la transcription de l'ADN, l'ARN est sous forme de simple brin dans le noyau.
E. FAUX : L’ADN est plus stable que l’ARN car les bases ont perdu leurs hydroxyles.

Tous droits réservés au Tutorat Associatif Toulousain 9/26
QCM 7 – BCD
A. FAUX : Toutes les caractéristiques héréditaires ne peuvent pas être déterminées par séquençage du génome
nucléaire et mitochondrial (certaines ne dépendant pas de la séquence primaire d’ADN : épigénétique). Donc c'est le "toutes" qui rend l'item FAUX.
E. FAUX : L’ADN mitochondrial est transmis uniquement par la mère chez les eucaryotes supérieurs.
QCM 8 – B
A. FAUX : Cette molécule est un hybride ARN/ADN.
C. Errata : passe vraie. En effet on voit un T (1ère molécule de la B) avec un A (1ère molécule de la A). D. FAUX : Cette molécule a son pic d’absorption à 260 nm grâce aux bases. E. FAUX : Cette molécule est chargée négativement. ADN et ARN sont chargés négativement.
QCM 9 – ACDE
B. FAUX : Elle est complémentaire et antiparallèle. C. VRAI : Pour synthétiser les amorces entre autres.
QCM 10 – CD
A. FAUX : L’initiation de la réplication débute entre autres grâce la reconnaissance de l’origine de réplication
par les protéines dnaA chez les procaryotes. Le promoteur étant pour la transcription.
B. FAUX : L’ARN polymérase (ou primase) synthétise également des amorces d’ARN pour le brin continu.
C. VRAI : Elle va relier l'ADN néosynthétisé de l'ADN polymérase III avec l'ADN néosynthétisé de l'ADN
polymérase I en faisant des liaisons 3'→ 5' phosphodiester.
E. FAUX : Les protéines SSB stabilisent l’ADN simple brin (attention piège !).
QCM 11 – ACDE
B. FAUX : Elles ne permettent pas l’identification d’un excès de sucre lié au diabète dans le sang.
QCM 12 – ABCE
D. FAUX : C’est la dénaturation qui se fait à une température avoisinant les 94°.
QCM 13 – BC
A. FAUX : A est une séquence cis-régulatrice et B est un élément trans-régulateur.
D. FAUX : Ce schéma symbolise la régulation de la transcription.
E. FAUX : Ce gène code pour un ARNm comme en témoigne la présence d’un codon d’initiation de la traduc-
tion (ATG qui deviendra AUG dans l’ARNm) et d’un codon stop (TGA qui deviendra UGA dans l’ARNm).
QCM 14 – ABCD
E. FAUX : Elle permet la transcription des introns et des exons chez les eucaryotes, cependant les introns
seront éliminés lors de l’épissage pour donner un ARNm mature.
QCM 15 – B
A. FAUX : Les ARNr proviennent de la maturation d’un pré ARN par des endonucléases (réalisent des cou-
pures) spécifiques, les exonucléases quant à elles suppriment, mangent des nucléotides. C. FAUX : Les ARNm reçoivent une coiffe (7-méthylguanosine triphosphate) du côté 5’. C’est d’ailleurs la
modification post-transcriptionnelle systématique pour tous les ARNm. D. Errata : passe faux. Tel quel l’item est faux car la queue poly A se trouve en 3’ du pré-ARNm (le reste est
juste). E. FAUX : Les ARNr subissent des modifications post-transcriptionnelles (sauf le 5S). Les ARNr (sauf le 5S)
sont synthétisés sous formes d’un long précurseur puis il va être découpé pour avoir plusieurs petits ARNr
(18S ; 5,8S ; 28S).
QCM 16 – ABE
A. VRAI : En effet la peptidyltransférase nécessite 1 liaison riche en énergie provenant du GTP pour réaliser
une liaison peptidique en 2 acides aminés.
C. FAUX : La séquence Shine Dalgarno n’existe que chez les procaryotes.
D. FAUX : L’activation des acides aminés implique l’hydrolyse d’une molécule d’ATP (pas de molécules de
GTP) en AMP, l’aminoacylARNt synthétase consomme donc 2 liaisons riches en énergie.

Tous droits réservés au Tutorat Associatif Toulousain 10/26
QCM 17 – ACE
B. FAUX : Seule une petite partie de l’ADN eucaryote est traduite en protéines.
C. VRAI : Effectivement on a les ARNm qui sont traduits, les ARNt et les ARNr.
D. FAUX : Les ARNt ne reconnaissent pas les codons stop, la reconnaissance des codons stop se fait par des
facteurs de terminaison.
E. VRAI : La peptidyl transférase réalise la liaison peptidique entre 2 AA et la translocase réalise le déplace-
ment du ribosome d’un codon ainsi l’AA lié se passe du site A vers le site P. Chacune de ces 2 enzymes
nécessitent 1 molécule de GTP.
QCM 18 – AB
C. FAUX : La séquence Shine Dalgarno n’existe que chez les procaryotes.
D. FAUX : Elle peut être régulée par la phosphorylation des facteurs de la traduction. Attention aux énoncés ! E. FAUX : Elle fait intervenir la consommation de 800 liaisons riches en énergie pour la synthèse d’un peptide
de 200 acides aminés : 1 AA ajouté = 4 liaison riche en énergie.
QCM 19 – BE
5’ GTCATTTCACGGTGACTGCGCG ----//----- GAGGTGGCCCTGCATCGTTTAAA 3’ AMORCES B
3’ CAGTAAAGTGCCACTGACGCGC ---//----- CTCCACCGGGACGTAGCAAATTT 5’
A. FAUX : Les amorces sont sur le même brin et dans le même sens.
C. FAUX : Voir item B.
D. FAUX : Regarder le lieu de fixation des amorces, qui ne se fixe pas directement à l’extrémité des brins, ce
qui fait que on 7 nucléotides ne seront pas synthétisés à gauche et 3 nucléotides ne seront pas synthétisés à
droite (attention, les amorces sont bien incluses dans le produit de PCR).
QCM 20 – AD
Attention les réponses données aux items C et E sont dues au fait qu’on ne travaille pas sur un allèle entier
mais seulement sur un exon. B. FAUX : La FISH ne permet d’observer que les macrolésions de l’ADN et non pas les mutations ponctuelles.
C et E comptées vrai en TD année 2019.

Tous droits réservés au Tutorat Associatif Toulousain 11/26
UE2 – Biologie cellulaire, BDR, Embryologie, BDD, Histologie
QCM 1 – ABDE A. VRAI : La mort cellulaire entraîne une augmentation de la perméabilité membranaire ce qui permet au
colorant de rentrer. Les cellules mortes sont colorées en bleu et les cellules vivantes sont non colorées. B. VRAI : Voir A, la perméabilité membranaire au bleu Trypan chez les cellules vivantes est nulle, elles ne
sont alors pas colorées. C. FAUX : La coloration au bleu trypan ne peut pas être réaliser sur des cellules fixées. D. VRAI : C’est un colorant qui permet de distinguer si la cellule est morte ou vivante, c’est donc un colorant
vital.
E. VRAI : la 𝑣𝑖𝑎𝑏𝑖𝑙𝑖𝑡é 𝑐𝑒𝑙𝑙𝑢𝑙𝑎𝑖𝑟𝑒 =135 𝑐𝑒𝑙𝑙𝑢𝑙𝑒𝑠 𝑛𝑜𝑛 𝑐𝑜𝑙𝑜𝑟é𝑒𝑠
150 𝑐𝑒𝑙𝑙𝑢𝑙𝑒𝑠 𝑡𝑜𝑡𝑎𝑙𝑒𝑠 𝑥 100 = 900, donc la viabilité cellulaire est de
90 %. QCM 2 – BC A. FAUX : Sa séquence contient en N-terminal une séquence reconnue par le complexe SRP. B. VRAI : La calréticuline est une protéine résidente du RE, elle comporte donc une séquence KDEL. C. VRAI : en effet, avec la calnexine D. FAUX : La traduction de cette protéine se fait dans le réticulum endoplasmique, elle est cependant initiée
dans le cytosol (comme pour toutes les protéines provenant du génome nucléaire). E. FAUX : Le dolichol participe à la glycosylation des protéines, les lectines chaperonnes (calnexine et cal-
réticuline) ne font que reconnaître les protéines glycosylées (contrôle qualité des protéines). QCM 3 – BDE A. FAUX : Lorsque les tubulines α et β sont seules dans le milieu, il n’y a pas la formation de microtubules,
les microtubules ne peuvent donc pas s’assembler seul in vitro. B. VRAI : Lorsque l’on ajoute uniquement le complexe Gamma TuRC aux tubulines α et β on observe un
début de création de microtubules (en faible nombre, condition 4). C. FAUX : Le protéine 5RAP2 ne favorise pas la nucléation des microtubules en absence de complexe gamma
TuRC (condition 2). D. VRAI : En effet, on observe qu’en présence de la protéine 5RAP2 et de gamma TuRC (condition 5) le
degrès de nucléation des microtubules est augmentée comparé à l’expérience comprenant gamma TuRC seul
(condition 4). E. VRAI : En effet, on observe qu’en présence de la protéine 5RAP2 mutée et de gamma TuRC (condition 6), le degrès de nucléation des microtubules est le même que celle retrouvée dans l’expérience comprenant gamma
TuRC seul (condition 4). QCM 4 – DE A. FAUX : Les GTPases Rab sont impliquées dans le ciblage des vésicules vers les différents compartiments
du système endomembranaire. Rho est impliqué dans la modification du cytosquelette d’actine. B. FAUX : Ces protéines sont activées par la fixation d’un GTP par des protéines régulatrices appelées GEF. C. FAUX : Les ancres GPI permettent l’ancrage à la surface externe de la membrane plasmique, or ces pro-
téines sont liées à la surface interne de la membrane plasmique. D. VRAI : GDI séquestre les GTPase, ainsi elle les rend inactive. E. VRAI : Voir A. QCM 5 – CE A. FAUX : La glucosylcéramide synthase permet au contraire la synthèse du glucosylcéramide et non sa dé-
gradation. B. FAUX : L’accumulation de glucosylcéramide peut être la conséquence d’une diminution de l’activité de la
glucosylcéramidase (hydrolase) dans les lysosomes. C. VRAI : En effet, le glucosylcéramide est bien un sphingolipide, son accumulation provoque une sphingoli-
pidose. D. FAUX : Les ribosomes sont contenus dans le REG ou peuvent être libres dans le cytosol. La fraction enri-
chie en lysosome n’en contient pas après une centrifugation différentielle (les derniers ribosomes libres seront
encore dans le surnageant et se retrouveront dans la toute dernière fraction). E. VRAI : En effet, la LAMP est une glycoprotéine transmembranaire spécifique des lysosomes.

Tous droits réservés au Tutorat Associatif Toulousain 12/26
QCM 6 – BE A. FAUX : Les protéines intégrales sont associées à la membrane plasmique par un domaine transmembranaire
hydrophobe. B. VRAI : Les protéines organisées en hélice alpha sont majoritaires par rapport à celles organisées en feuillet
Bêta. C. FAUX : Les protéines de type 1 ont leur extrémité carboxy-terminale riche en acides aminés basiques. Les protéines de type 2 elles possèdent leur extrémité N-ter riche en acides aminés basiques, ce qui fait que
cette extrémité ne rentre pas dans le translocon et que l’on obtient une protéine avec l’extrémité N-ter cytoso-
lique et C-ter extracellulaire. D. FAUX : Les domaines transmembranaires hydrophobes des protéines intégrales de la membrane plasmique
ne sont pas clivables par la peptidase du signal (signal peptidase) au niveau de la lumière du REG. E. VRAI : Leur synthèse est initiée dans le cytosol et est poursuivie au niveau du translocon du REG. C’est ensuite par le jeu du trafic vésiculaire qu’elles atteindront la membrane plasmique. QCM 7 – BE A. FAUX : Les GLUT (Glucose transporter) sont impliqués dans le transport passif (pas de consommation
d’ATP) du glucose dans le sens de son gradient de concentration. B. VRAI : En effet, ce sont des protéines intégrales de la membrane constituant un pore qui permet le passage
sélectif d’ions du plus concentré vers le plus concentré (donc selon un gradient de concentration). C. FAUX : Le transport des molécules d’eau par les aquaporines n’est pas un phénomène de diffusion simple.
La diffusion simple est un passage simple à travers la membrane plasmique, ici c’est une diffusion facilitée
(car elle implique des protéines transmembranaires). D. FAUX : La pompe sodium/potassium (Na+/K+) est un transporteur primaire permettant de créer un gradient
de concentration de sodium. E. VRAI : Les ions bicarbonates sont des éléments basiques, augmenter la concentration en bicarbonates re-
vient à augmenter la basicité du milieu et donc à augmenter le pH. QCM 8 – ACE A. La membrane pharyngienne est l'équivalent de la bouche primitive.
B. Lors du repliement longitudinal, l’embryon conserve sa symétrie bilatérale indispensable.
D. Les crêtes neurales s’individualisent autour du 23-24ème jour du développement.
QCM 9 – DE A. Les principaux nutriments nécessaires au développement de l’œuf proviennent du milieu extérieur.
B. La réaction pré déciduale de l’endomètre a lieu avec ou sans fécondation.
C. Les jonctions intercellulaires s’établissent au stade 10-16 blastomères.
QCM 10 – ABE A. Ayez en tête le schéma du cours du stade avilleux fruste (permet en le visualisant). B. Idem que pour le A ayez les schémas des 3 stades en tête.
C. Vers le 9ème jour de développement, le syncytiotrophoblaste entourant le cytotrophoblaste se creuse de la-
cunes. Le cytotrophoblaste n'aura jamais de lacunes ! D. Le début de la formation des villosités primaires a lieu au stade avilleux trabéculaire (13ème jour) tandis que
le stade avilleux lacunaire est du 9ème au 12ème jour. E. Il n'y a pas encore de mésenchyme extra embryonnaire au stade avilleux frustre !
QCM 11 – ACE B. La gastrulation correspond à la mise en place de la structure à 3 feuillets lors de la 3ème semaine de déve-
loppement. Le passage d’une structure plane discoïde à une structure cylindrique se fait lors de la 4ème semaine. D. La taille du cœlome extra-embryonnaire diminue au cours du repliement embryonnaire (lors de la 4ème
semaine). Cependant il y a une croissance importante de la cavité amniotique. E. C'est cette migration et différentiation de l'épiblaste par la ligne primitive qui va donner le mésoblaste.

Tous droits réservés au Tutorat Associatif Toulousain 13/26
QCM 12 – AD A. Le somatopleure intra-embryonnaire ne se forme que plus tard vers fin 20ème jour-début 21ème jour. B. Et dans le pédicule aussi, mais pas dans la somatopleure (piège). C. Les somites ne commencent à se former qu’au 21ème jour (4ème semaine). E. Les bords latéraux de la gouttière neurale se divisent longitudinalement pour former les crêtes neurales.
QCM 13 – A B. Elle peut se faire entre le 6ème et le 10ème jour après l’ovulation. C. L’œuf fécondé est implanté au stade 4 cellules, or à ce stade le trophoblaste primitif n’est pas encore formé.
De plus, le syncytiotrophoblaste se forme après le trophoblaste primitif.
D. Est considéré comme un accouchement à terme.
E. En moyenne 3 œufs fécondés sont déposés au niveau du pôle supérieur de la cavité utérine.
QCM 14 – B A. Se produit plus tôt lors de la spermiogenèse.
C. Les nucléosomes se dissocient de l’ADN nucléaire permettant la formation de cassures simple brin.
D. Faux car l’item se rapporte à la spermatocytogenèse et non à la spermiogenèse. E. Les spermatozoïdes ne seront matures qu’une fois parvenus dans l’épididyme.
QCM 15 – AC B. Des stéréocils sont retrouvés au niveau du pôle apical des cellules prismatiques de l’épididyme.
D. D’une part l’épithélium des alvéoles des poumons est squameux, d’autre part celui-ci ne porte pas de cils
vibratiles.
E. Le transport paracellulaire n’implique pas le passage de la membrane plasmique de la cellule (le passage se
fait entre les cellules).
QCM 16 – ADE B. Une glande exocrine simple présente toujours une partie excrétrice unique.
C. Les glandes multilobées déversent leurs sécrétions dans des canaux intralobulaires, puis dans des canaux
interlobulaires puis, enfin, dans un canal collecteur.

Tous droits réservés au Tutorat Associatif Toulousain 14/26
UE3 – Biophysique
1 : ADE 2 : BCE 3 : DE 4 : ABCDE 5 : ACD
6 : ACE 7 : ABE 8 : B 9 : B 10 : AC
11 : AB 12 : ACDE 13 : BCDE 14 : AD 15 : BCD
QCM 1 – ADE
On rappelle que :
− Une vitesse correspond à une longueur parcourue en un certain temps (longueur / temps), et est donc
de dimensions L.T-1.
− Une accélération correspond à une vitesse divisée par un temps, et est donc de dimensions L.T-2.
− Une force correspond à une masse multipliée par une accélération, et est donc de dimensions M.L.T-
2.
A. Vrai. Une pression correspond à une force (M.L.T-2) appliquée sur une surface (L2). Ainsi, Force / Surface
nous donne comme dimensions M.L-1.T-2.
B. Faux. Un volume correspond à trois longueurs (les trois dimensions de l'espace, longueur, largeur &
hauteur) multipliées et non divisées entre elles → L3.
C. Faux & D: Vrai. Puisque PV = nRT, on a R = PV/nT.
• PV → M.L-1.T-2.L3 = M.L2.T-2 d'une part ;
• nT → N.Θ d'autre part.
Ainsi, [R] = M.L2.T-2.N-1.Θ-1.
E. Vrai. M.L2.T-2 correspond aux dimensions d'une énergie (Force * Longueur), pouvant être exprimée en
joules (J). Les dimensions N-1.Θ-1 ont respectivement pour unités la mole (mol) et le degré Kelvin (K), d'où
J.mol-1.K-1.
QCM 2 – BCE
On a une vitesse v, une masse m, une hauteur (donc longueur) h, une constante sans dimensions k, et une
accélération g. On a donc [v] = L.T-1 ; [m] = M ; [h] = L ; [k] = ∅ ; [g] = L.T-2.
A. Faux. v étant une vitesse, elle a pour dimensions L.T-1.
B. Vrai. Une vitesse n'implique pas de masse, ainsi la dimension M correspondant à la masse est absente ; on
dit donc qu'elle est de puissance 0, donnant M0 = 1.
C. Vrai. La hauteur h ne donne que la dimension de la longueur L. Or, une vitesse a une dimension de longueur
de puissance 1, et l'accélération elle aussi apporte une dimension de longueur (à la puissance 1) : par
conséquent, la hauteur h confie la moitié de la dimension L, donc à la puissance ½ → L1/2. Il en est de même
pour l'accélération.
D. Faux. Comme dit précédemment, l'accélération confie la seconde moitié de la dimension L de la vitesse, il
faudrait donc que g soit à la puissance ½. Cela nous permet d'obtenir comme dimensions L1/2.T-2/2 = L1/2.T-1.
De plus, cela coïncide également parfaitement au niveau de la dimension du temps : g1/2 offre le T-1 de la
vitesse.
L'énoncé place g à la puissance δ et non à la puissance γ, peut-être une erreur d'impression ou de rédaction de l'énoncé. Ainsi, on obtient bien au final v = k.m0.h1/2.g1/2 → M0.L1/2.L1/2.T-1 = L.T-1.
E. Vrai. Une formule (ici une égalité) est homogène si les dimensions sont respectées. Or (cf. C & D) on vient
de prouver que h et g doivent être à la puissance ½ pour correspondre aux dimensions d'une vitesse.
En rappelant que la puissance ½ correspond à la racine carrée, et que k est sans dimensions, cette formule est
parfaitement homogène.

Tous droits réservés au Tutorat Associatif Toulousain 15/26
QCM 3 – DE
Un équivalent correspond au nombre de charges apportées par une espèce en solution, c'est-à-dire à la molarité
multipliée par la charge (en valeur absolue) et le nombre de fois que l'ion concerné apparaît en se dissociant.
On calcule les équivalents avec la formule : Eq/L = m x k x Z avec m la molarité, k le nombre de fois que l'ion
apparaît après dissociation, et Z son nombre de charges en valeur absolue.
L'osmolarité correspond à la molarité m multipliée par le coefficient d'ionisation i, de formule i = 1 + α (p-1)
avec α le coefficient de dissociation et p le nombre de particules créées lors de la dissociation.
A. Faux. Le sodium Na+ ne provient que du Na2SO4.
Ce dernier a une concentration de 1,42 g/L et une masse molaire de 142 g/mol → pour trouver la molarité en
mol/L, on calcule donc 1,42 / 142 = 10-2 = 0,01 mol/L de Na2SO4 en solution.
Le Na2SO4 se dissocie en 2 Na+ (k = 2 et Z = 1) + SO42-. On a donc 0,01 * 2 * 1 = 0,02 Eq/L de sodium Na+
en solution.
B. Faux. Même raisonnement que pour l'item A, mais en se concentrant sur le SO42-: on a donc 0,01 * 1 * 2 =
0,02 Eq/L de SO42-.
C. Faux. La molarité du Na2SO4 est donc de 0,01 mol/L (cf. A).
Le glucose a pour molarité 0,9 / 180 = 9.10-1 / 18.101 = ½ . 10-2 = 0,5.10-2 = 0,005 mol/L.
Pour un même volume, on a donc 0,01 moles de Na2SO4 et 0,005 moles de glucose, soit deux nombres de
moles différents.
D. Vrai. Le Na2SO4 a une molarité de 0,01 mol/L, un coefficient de dissociation α = 0,9, et donne 3 particules
en se dissociant (2Na+ + SO42-). Ainsi, l'osmolarité du Na2SO4 vaut m' = i x m = [1 + 0,9(3 – 1)] x 0,01 = 2,8
x 0,01 = 0,028 osmol/L.
E. Vrai. L'osmolarité totale de la solution correspond à la somme de celle du glucose et de celle du Na2SO4.
Le glucose ne se dissocie pas en solution, donc α = 0 et i = 1.
Ainsi, l'osmolarité du glucose vaut m' = 1 x m = m = 0,005 mol/L (cf. C).
L'osmolarité totale de la solution vaut donc 0,028 + 0,005 = 0,033 osmol/L.
QCM 4 – ABCDE
A. Vrai. Le sodium Na+ n'est initialement présent que dans la solution A de NaCl.
Le NaCl a une molarité de 0,01 mol/L dans la solution A. On mélange 900 mL (soit 0,9 L) de la solution A
avec la solution B : on apporte donc 0,01 x 0,9 = 0,009 mol de NaCl dans le mélange. Le mélange total formant
1 L (900 mL + 100 mL), on obtient 0,009/1 = 0,009 mol/L de NaCl dans le mélange.
NaCl → 1 Na+ + 1 Cl-, il y a 0,009 x 1 x 1 = 0,009 Eq/L de sodium dans ce mélange.
B. Vrai. Le calcium Ca2+ n'est initialement présent que dans la solution B de CaCl2.
Le CaCl2 a une molarité de 0,005 mol/L dans la solution B. On mélange 100 mL (soit 0,1 L) de la solution B
avec la solution A : on apporte donc 0,005 x 0,1 = 0,0005 mol de CaCl2 dans le mélange. Le mélange total
formant 1 L, on obtient 0,0005/1 = 0,0005 mol/L de CaCl2 dans le mélange.
CaCl2 → 1 Ca2+ + 2 Cl-. On a donc 0,0005 x 1 x 2 = 0,001 Eq/L de Ca dans le mélange.
C. Vrai. Le chlore Cl- intervient dans les deux solutions A et B. Il faut donc prendre en compte les molarités
et équivalents des deux espèces distinctes.
Cf. A & B: on a donc 0,009 mol/L de NaCl et 0,0005 mol/L de CaCl2 dans le mélange. NaCl → 1 Na+ + 1 Cl- et CaCl2 → 1 Ca2+ + 2 Cl-.
• Grâce au NaCl, on a 0,009 x 1 x 1 = 0,009 Eq/L de chlore ;
• Grâce au CaCl2, on a 0,0005 x 2 x 1 = 0,001 Eq/L de chlore.
On a ainsi, au total, 0,009 + 0,001 = 0,01 Eq/L de chlore Cl- dans le mélange.
D. Vrai. Le NaCl a pour osmolarité m' = i x m = [1 + 0,9(2 – 1)] x 0,009 = 1,9 x 0,009 = 0,0171 osmol/L.
Le CaCl2 a pour osmolarité m' = [1 + 0,8(3 – 1)] x 0,0005 = 2,6 x 0,0005 = 0,0013 osmol/L.
Ainsi, la solution a pour osmolarité totale 0,0171 + 0,0013 = 0,0184 osmol/L = 18,4,10-3 osmol/L.
E. Vrai. Cf. D: puisqu'on est en solution aqueuse, on considère que 1L = 1kg (car la masse volumique vaut
donc 1). Dans ce cas, on a bien 18,4.10-3 osmol/kg.

Tous droits réservés au Tutorat Associatif Toulousain 16/26
QCM 5 – ACD
A. Vrai. Il y a 10 g de gaz A pour une masse molaire de 100 g/mol. Il y a donc 10/100 = 0,1 mole de gaz A.
B. Faux. Il y a 2 g de gaz B pour une masse molaire de 20 g/mol. Il y a donc 2/20 = 0,1 mole de gaz B.
C. Vrai. Il y a donc autant de matière pour le gaz A que pour le gaz B avec 0,1 mole de chaque gaz. Cela
signifie que chacun possède la moitié de la quantité de matière totale : autrement dit, chaque gaz a une fraction
molaire de 0,5 (pour un total de 1).
En termes de calculs, on a un total de 0,2 mole de gaz. Chacun des deux a donc une fraction molaire de 0,1/0,2
= 0,5 (sans unité !).
D. Vrai. À 0°C, la loi des gaz parfaits PV = nRT se simplifie par PV = 22,4n (RT = 22,4) avec P en atm, V
en L et n en mol.
On a n = 0,2 mole de matière totale et une pression totale P = 1 atm.
On peut calculer V = 22,4n/P = 22,4 x 0,2 / 1 = 4,48 L.
E. Faux. Tout autre paramètre que P et T restant inchangé, une augmentation de la température (on passe de
0°C à 37°C, soit de 273,15K à 310,15K) doit nécessairement entraîner une augmentation de la pression pour
équilibrer l'égalité (selon PV = nRT, le terme de droite augmentant, celui de gauche doit lui aussi augmenter).
Autrement dit, une augmentation de la température va augmenter l'agitation thermique, augmenter les chocs
des molécules de gaz contre les parois du récipient : la pression va donc augmenter.
QCM 6 – ACE
Il existe 6 types (ou saveurs) de quarks, particules élémentaires de la matière, leur association formant des
hadrons. Parmi ces quarks, les quarks up (de charge +2/3) et down (de charge -1/3) vont notamment s'associer
pour former les nucléons du noyau atomique : les protons et neutrons (qui sont également des baryons, car
composés de 3 quarks chacun).
• Le proton, de charge +1, est formé de 2 quarks up et 1 quark down (+2/3 + 2/3 – 1/3 = +4/3 – 1/3 = +3/3 = +1) ;
• Le neutron, de charge nulle, est formé de 2 quarks down et 1 quark up (-1/3 – 1/3 + 2/3 = -2/3 + 2/3 = 0).
A. Vrai. Le nombre de masse A est de 12 : il y a donc 12 nucléons (comprenant protons et neutrons).
Puisque tous les nucléons sont des baryons à 3 quarks, le noyau du carbone-12 est donc composé de 12 x 3 =
36 quarks.
B. Faux. Le numéro atomique Z est de 6 et le nombre de masse de 12 : il y a donc 6 protons, chacun comprenant
2 quarks up (et 1 down), mais également 12 – 6 = 6 neutrons, chacun comprenant 1 quark up (et 2 down) !
Le noyau du carbone-12 est donc composé de (6 x 2) + (6 x 1) = 18 quarks up.
C. Vrai ? Cet item est compté vrai mais pose problème : les leptons constituent une catégorie de particules fondamentales légères, comprenant notamment les électrons, les neutrinos et leurs antiparticules respectives (entre autres). En ne prenant en compte que les électrons (les seuls qui sont toujours présents), il est faux de dire que le noyau en possède ; par contre, l'atome de carbone-12 en possède bien 6, mais dans le cortège électronique... D. Faux. Les neutrons sont composés de quarks up ou down qui ne sont pas de charge nulle ; la charge globale
nulle du neutron est due à une compensation entre ces différentes charges ! Les 6 saveurs de quarks sont toutes de charge non-nulle : la moitié de charge +2/3, l'autre moitié de charge -1/3. E. Vrai. L'unité de masse atomique est officiellement définie de telle sorte que 1 uma vaut précisément 1/12ème
de la masse d'un atome de carbone-12. Ainsi, la masse d'un atome de carbone-12 vaut très précisément 12 uma.
Mis en équation, cela donne 1 uma = 1/12 x 12C → 12C = 1 uma / (1/12) = 12 uma.
C'est une particularité de cet élément car étant la référence pour l'uma, d'autres atomes n'auront pas forcément la même masse en uma que leur nombre de masse !
QCM 7 – ABE
L'objectif est de vérifier la conservation des nombres de masse A et des numéros atomiques Z entre les réactifs
et les produits.
A. Vrai. A = 98 + 1 = 99
Z = 42 + 0 = 42 →
A' = 0 + 99 = 99
Z' = 0 + 42 = 42 Neutron n: A = 1 & Z = 0
Proton p: A = 1 & Z = 1
Deutérium d: A = 2 & Z = 1
Particule α: A = 4 & Z = 2
Rayon γ: A = 0 & Z = 0
B. Vrai. A = 98 + 2 = 100
Z = 42 + 1 = 43 →
A' = 1 + 99 = 100
Z' = 1 + 42 = 43
C. Faux. A = 99 + 4 = 103
Z = 44 + 2 = 46 →
A' = 2 + 99 = 101
Z' = 2 + 42 = 44

Tous droits réservés au Tutorat Associatif Toulousain 17/26
D. Faux. A = 235 + 1 = 236
Z = 92 + 0 = 92 →
A' = 3 + 99 + 133 = 235
Z' = 0 + 42 + 50 = 92
E. Vrai. A = 235 + 1 = 236
Z = 92 + 0 = 92 →
A' = 5 + 99 + 132 = 236
Z' = 0 + 42 + 50 = 92
QCM 8 – B
Le molybdène-99 possède une période radioactive non-nulle : celui-ci va donc bien subir une désintégration
radioactive.
On rappelle qu'une désintégration radioactive spontanée aboutit forcément à une diminution de la masse et à
une augmentation du défaut de masse.
A. Faux. Une émission β+ permet d'obtenir l'élément de numéro atomique Z – 1 : ici, on passerait du molyb-
dène-99 au niobium-99. Or, le niobium est de masse supérieure, et ne peut donc pas être le fils du molybdène
suite à une désintégration radioactive : ce dernier n'est donc pas émetteur β+.
B. Vrai. Une émission β- permet d'obtenir l'élément de numéro atomique Z + 1 : ici, on passerait du molybdène-
99 au technétium-99 (ou 99m). Puisque le molybdène-99 subit bien une désintégration radioactive (car de
période non-nulle), c'est donc la seule possibilité.
C. Faux. C'est le noyau fils (donc le technétium-99m) qui serait émetteur de photons γ afin de se désexciter,
pas le noyau père.
D. Faux. Toujours selon les masses, le technétium-99m devrait donc passer à l'élément Z + 1 (le ruthénium-99
étant de masse inférieure, le molybdène-99 de masse supérieure) par une transformation β-, pas par une capture
électronique (synonyme de conversion externe). Ne pas confondre conversion externe et conversion interne !
E. Faux. Le technétium-99m, afin de désexciter son noyau, émet des photons et non des électrons γ !
QCM 9 – B
A. Faux. On est dans le cas d'une transformation β-, ce sont donc des antineutrinos qui sont émis avec les
électrons.
B. Vrai. Dans le cas de particules β-, l'énergie moyenne s'obtient par la formule Emoy = Emax/3.
Ainsi, l'énergie maximale vaut Emoy x 3 = 0,4 x 3 = 1,2 MeV.
C. Faux. Le seuil de 1,022 MeV ne s'applique qu'aux transformations β+.
D. Faux. On est toujours dans le cas d'une transformation β-, le seuil de 1,022 MeV ne s'applique donc pas. Dans le cas d'une transformation β+, le seuil de 1,022 MeV serait déjà pris en compte dans l'énergie moyenne ou maximale donnée. E. Faux. L'annihilation résultant en l'émission en sens opposés de deux photons γ de 511 keV n'est systématique
que dans le cas de la transformation β+, où le positon va inévitablement s'arrêter après avoir interagi avec la
matière, et subir l'annihilation par rencontre avec un électron libre du milieu.
QCM 10 – AC
A. Vrai. Dans le cas où, en enceinte close, un élément père et un élément fils sont en contact et que la période
du père est supérieure à la période du fils, on obtient ce que l'on appelle un équilibre du régime.
B. Faux. En raison de l'équilibre de régime, l'activité du fils (le technétium-99m) est très proche de celle du
père (molybdène-99) mais pas exactement égale à celle-ci.
C. Vrai. On rappelle que l'on est dans un équilibre du régime : les activités et périodes apparentes du fils sont
très proches de celles du père. Ainsi, pour le père comme pour le fils, la période est de 66h : cela signifie que
toutes les 66h, l'activité du père comme du fils est divisée par 2, soit décroît d'un facteur 0,5.
D. Faux. 3,3h représentent 1/20ème de l'activité apparente du technétium-99m.
En prenant la formule A(t) = A0.e-λ.t, une diminution par un facteur compris entre 0,92 et 0,94 (environ 0,93)
signifie que e-λ.t = 0,93.
ex peut être approché par (1 – x) lorsque x se rapproche de 0. Ainsi, puisque 1 – x = 0,93, on a x qui est proche
de 0,07. Cela signifie que -λ.t = 0,07, soit -ln(2) / 10 = -0,7 / 10.
Puisque λ = ln(2) / T1/2, il faut donc que t vaille (1/10)T1/2 pour que -λ.t = ln(2) / T1/2 x (1/10)T1/2 = ln(2) / 10.
Autrement dit, le facteur de décroissance de l'activité n'est compris entre 0,92 et 0,94 que si le temps écoulé
correspond à environ 1/10ème de la période, pas 1/20ème.
E. Faux. Bien que les activités et périodes soient en apparence proches, les quantités et nombres d'atomes de
chaque élément père ou fils sont différents. Le rapport s'écrit Np / Nf ≈ Tp / Tf avec Tp et Tf les périodes réelles
des éléments père et fils.
Ainsi, on a Np / Nf ≈ 66 / 6 ≈ 11, soit Np = 11.Nf. Il y a donc 11 fois plus de noyaux pères que de noyaux fils
(donc 11 fois plus de molybdène-99 que de technétium-99m), et non l'inverse !

Tous droits réservés au Tutorat Associatif Toulousain 18/26
QCM 11 – AB
A. Vrai. Contrairement aux désintégrations β, les électrons de conversion interne (suite à une désexcitation
nucléaire) ne peuvent prendre que des niveaux précis d'énergie. Ils sont donc émis selon un spectre de raies.
B. Vrai & C Faux. Le parcours moyen des électrons dans de l'eau est donné par la formule Rmoy (cm) = E(MeV)
/ 2ρ (la masse volumique ρ valant 1 dans l'eau).
On a donc un parcours moyen de (0,140 – 0,021) / 2 = 0,119 / 2 ≈ 0,06 cm.
Le parcours maximal est ensuite donné par Rmax = Rmoy x 1,5, soit 0,06 x 1,5 = 0,09 cm dans l'eau.
D. Faux. Le parcours dans l'eau est significativement faible : si une telle épaisseur d'eau arrête tous les élec-
trons, le parcours maximal dans les tissus organiques sera très proche voire plus faible (en rappelant que les tissus organiques sont composés à majorité d'eau). En bref, le parcours maximal des électrons est trop faible
pour qu'ils soient détectés hors du patient.
E. Faux. Ne pas pouvoir détecter des électrons hors du patient ne signifie pas que l'élément est inutile en
médecine nucléaire : au contraire, c'est un critère de choix afin de réaliser des radiothérapies internes (on veut
que les électrons n'irradient que la cible interne et pas le milieu externe au patient, comme sa famille, ses amis ou le personnel médical).
QCM 12 – ACDE
A. Vrai. L'énergie moyenne d'ionisation dans l'eau est de 32 eV. Le nombre d'ionisations réalisés par un flux
d'électrons sera donc de n = Ec / Eionisation = 119 keV / 32 eV = 119.103 / 32 ≈ 3,7.103.
B. Faux. La densité linéique moyenne se calcule par la formule DLImoy = n / Rmoy. On a 3,7.103 ionisations et
un parcours moyen de 0,06 cm, donc une DLImoy = 3,7.103 / 0,06 ≈ 6,2.104 ionisations.cm-1.
C. Vrai. Même raisonnement, en sachant que 6,2.104 ionisations.cm-1 équivalent à 6,2.103 ionisations.mm-1.
Puisque 1 mm = 103 μm, on a donc 6,2 ionisations.μm-1.
D. Vrai. Le transfert d'énergie linéique moyen est donné par la formule TELmoy = DLImoy * Eionisation. On a donc
un TEL moyen de 6,2.104 * 32 ≈ 200.104 eV.cm-1 = 2.106 eV.cm-1 = 2 MeV.cm-1.
E. Vrai. De la même manière mais en convertissant le résultant, on obtient 2.10-1 MeV.mm-1, soit 200 keV.mm-
1.
QCM 13 – BCDE
A. Faux & B. Vrai. Les photons (rayons) X proviennent du cortège électronique et de ralentissements d'élec-
trons, mais pas du noyau. Les photons γ proviennent eux du noyau.
C. Vrai. Les rayonnements ionisants le sont soit directement s'ils sont chargés électriquement (comme l'élec-
tron ou le positon), soit indirectement s'ils ne sont pas chargés électriquement (comme les photons ou les
neutrons).
D. Vrai. Selon le diagramme d'Evans, à partir de 0,1 MeV (soit 100 keV), pour des milieux de faible numéro
atomique Z comme l'eau, l'effet Compton devient prépondérant (mais coexiste toujours avec l'effet photoélec-
trique !). L'effet de matérialisation est ici inenvisageable car les énergies sont inférieures à 1,022 MeV. E. Vrai. Les rayonnements indirectement ionisants auront des interactions dites stochastiques, probabilistes. Il
n'y a donc aucune certitude quant à leur arrêt : on ne peut que les atténuer. L'atténuation de ces rayonnements
indirectement ionisants, aussi bien chez les photons que les neutrons par exemple, suit une loi de décroissance
exponentielle (on approchera mais n'atteindra jamais exactement un flux nul).
QCM 14 – AD
A. Vrai. Une chambre d'ionisation permet en effet de mesurer une dose produite par des rayonnements ioni-
sants.
B. Faux. Le rendement désigne le rapport des photons détectés sur les photons atteignant le détecteur.
R = Ndétectés / Natteignant le détecteur. Ainsi, un rendement de 1 ou 100% signifie uniquement que tous les photons
atteignant le détecteur sont détectés, pas que tous les photons émis par la source sont détectés.
C. Faux. Le nombre de photons subit, même dans l'air, une atténuation exponentielle. La diminution ne sera
donc pas véritablement inversement proportionnelle, mais dépendante de la distance selon une loi exponen-
tielle.
D. Vrai. Les détecteurs à scintillation peuvent en effet être utilisés comme spectromètres.
E. Faux. En spectrométrie gamma, on observera bien un pic photoélectrique situé à l'énergie initiale des pho-
tons (ici 140 keV), mais on observera ensuite un diffusé Compton (et non un pic) à des énergies inférieures au
pic (l'effet photoélectrique correspondant à un transfert total de l'énergie du photon, tandis que l'effet Compton admet plusieurs transferts d'énergie, inférieurs à l'énergie totale du photon).

Tous droits réservés au Tutorat Associatif Toulousain 19/26
QCM 15 – BCD
A. Faux. Cf. 13 C. Les neutrons n'étant pas des particules chargées, ce sont donc des rayonnements indirecte-
ment ionisants.
B. Vrai. Les probabilités d'interaction des neutrons avec d'autres éléments de la matière que les noyaux sont
très faibles voire carrément nulles. On considère donc qu'ils n'interagissent qu'avec les noyaux atomiques.
C. Vrai. Lors de cette diffusion élastique, les neutrons confèrent une partie de leur énergie cinétique aux noyaux
d'hydrogène qu'ils rencontrent, ils seront donc ralentis.
D. Vrai. Cf. 13 E. Puisque non chargés, les neutrons auront des interactions probabilistes et non déterministes
avec la matière : ils ne seront donc qu'atténués selon une loi exponentielle.
E. Faux. Les neutrons interagissent préférentiellement, lors des diffusions élastiques, avec des élément légers
(des noyaux dont le diamètre et la masse sont proches de ceux du neutron isolé, le noyau d'hydrogène n'étant composé que d'un proton). On veut éviter les phénomènes d'absorption de neutrons (entraînant des réactions
nucléaires potentiellement en chaîne), donc on privilégie la diffusion élastique au moyen d'éléments légers.

Tous droits réservés au Tutorat Associatif Toulousain 20/26
UE4 – Biostatistiques QCM 1 – DE
A. FAUX. La fonction cosinus est paire.
B. FAUX. La fonction cosinus est décroissante sur [0;𝜋
2] donc sur [0;
𝜋
3], elle est décroissante.
C. FAUX. Attention cos est inférieur ou égal à 0 entre 𝜋
2 𝑒𝑡 𝜋! Car 𝑐𝑜𝑠(𝜋) = 0.
D. VRAI. Soit on le voit directement en connaissant la courbe de la fonction inverse, et en transposant à cette
fonction, soit on fait la dérivée de 𝑔(𝑥): 𝑔′(𝑥) = −2𝑥
(𝑥^2+1)^2
Or,−2𝑥 est négatif pour tout 𝑥 supérieur à 0 (donc aussi sur [0;1]). Et le dénominateur est un carré qui ne n’annule pas donc il est toujours strictement supérieur à 0 et positif.
Donc la dérivée est négative sur [0;1], donc 𝑔(𝑥) est décroissante sur cet intervalle. E. VRAI. Cette fonction est une fonction composée, c’est la composée de :
- 𝑔(𝑥) × 𝜋 qui est décroissante sur [0;1] (multiplier une fonction par une constante positive ne change
pas ses variations)
- par la fonction 𝑐𝑜𝑠 qui est décroissante sur [0;1] aussi (car 1 est compris entre 0 et 𝜋
2).
- La composée de deux fonctions décroissantes donne une fonction croissante (“-” par “-” fait “+”).
MAIS attention ici cette fonction composée est multipliée par une constante négative −5 qui inverse donc les
variations (se souvenir “-” par “+” fait “-”). Donc 𝑓(𝑥) est décroissante sur [0;1].
QCM 2 – BD
A. FAUX. 𝐷1 a une pente moins raide que 𝐷2 donc le coefficient directeur de 𝐷1est inférieur à celui de 𝐷2.
Donc c’est l’inverse : 𝐷1→ équation a 𝐷2→ équation b B. VRAI. Il suffit de regarder sur le graphique : 𝑦 = 3 correspond au point de l’axe des ordonnées où les
tangentes se croisent. Donc si on regarde entre -1 et 1 la courbe de 𝑓(𝑥) ne dépasse jamais 3. C. FAUX. Non il semble que 𝑙𝑖𝑚
𝑥 → +−∞= 0.
D. VRAI. En effet, la courbe de 𝑓(𝑥) “monte” vers l’infini. E. FAUX. Non il semble que 𝑓(𝑥) n’ait qu’un seul minima en 𝑥 = 1.
QCM 3 – BD
A. FAUX. Cette fonction est définie sur IR*+ car 𝑙𝑛(𝑥) n’est pas défini en 0 !
B. VRAI. Cela signifie que lorsque S tend vers 0, cette fonction tend vers +/- ∞. En effet : 𝑙𝑖𝑚𝑥 → 0
𝑙𝑛(𝑆
𝑆9) = −∞
d’où : 𝑙𝑖𝑚𝑥 → 0
−1
𝑘𝑇𝑒𝑙𝑛(
𝑆
𝑆0) = +∞
augmente. Comme il y a un signe "-" devant 1
𝑘𝑇𝐸, si C. FAUX. Si S augmente, alors
ln (𝑆
𝑆0) augmente, alors −
1
𝑘𝑇𝐸ln (
𝑆
𝑆0) diminue, donc C(S) diminue.
D. VRAI. En effet, on peut considérer que S0 et C sont inversement proportionnelles. E. FAUX. Si la concentration en agent de contraste est constante et que Te augmente, il faut que S0 diminue
pour compenser la diminution !
QCM 4 – ABCD
A. VRAI. 𝑓(0 + 𝑥) = 𝑐𝑜𝑠(0) + 𝑐𝑜𝑠′(0). 𝑥 + 𝑜(𝑥) = 1 − 𝑠𝑖𝑛(0). 𝑥 + 𝑜(𝑥) = 1 + 𝑜(𝑥) B. VRAI. C’est une FI : donc pour la résoudre, on fait un DL en 0 du dénominateur :
𝑓(0 + 𝑥) = 0 + 1
2√1 − 0× 𝑥 + 𝑜(𝑥) =
1
2. 𝑥 + 𝑜(𝑥)
C. VRAI. La fonction racine carrée est négligeable devant les polynômes en +∞. D. VRAI. Oui, à apprendre par cœur ! E. FAUX. Attention, on aurait tendance à simplifier cette limite en gardant seulement les exponentielles de
plus haut degré comme on le fait avec les polynômes, mais on ne peut pas car ce ne sont pas des polynômes ! Dans ce cas, quand 𝑥 tend vers −∞, les exponentielles tendent vers 0, il reste donc 3 au numérateur et 1 au
dénominateur. Donc la limite en −∞ est 3.

Tous droits réservés au Tutorat Associatif Toulousain 21/26
QCM 5 – A
A. VRAI. C’est la définition d’un point critique B. FAUX. 𝑑𝑓(𝑥, 𝑦) = (4𝑥𝑦^2 − 2𝑥𝑦 + 3𝑦 ). 𝑑𝑥 + (4𝑥^2𝑦 − 𝑥^2 + 3𝑥 − 15𝑦^2). 𝑑𝑦 Cette différentielle s'annule pour (x ; y) = (0 ; 0) mais pour (x ; y) = (-3 ; 0) on a :
𝑑𝑓 = 0. 𝑑𝑥 + (0 − 9 − 9 + 0). 𝑑𝑦 = 0. 𝑑𝑥 − 18. 𝑑𝑦 la différentielle ne s’annule pas pour ce couple de
valeurs ! C. FAUX. Et impossible car 𝑒𝑥𝑝(𝑥) ne fait jamais 0, donc la différentielle ne s’annulera jamais. D. FAUX. Non c’est impossible car cette fonction n’a aucun point critique, or si une fonction n’a pas de point
critique elle ne peut pas avoir d’extremum (pas de maximum ni de minimum). Un extremum est un point
critique ; mais un point critique n’est pas toujours un extremum. E. FAUX. Cf item D.
QCM 6 – ACE
A. VRAI. Une fonction rationnelle comme celle-ci est définie sur IR si et seulement si son dénominateur ne
s’annule jamais. Pour cela, on peut faire la méthode du discriminant : ∆ = 𝑏^2 − 4𝑎𝑐 = 4 − 8 = −4< 0 donc il n’y a pas de valeur de 𝑥 qui annule le dénominateur. De plus, 𝑥^2 + 2𝑥 + 2 est bien strictement positif : donc 𝑓(𝑥) est définie sur IR. B. FAUX. 𝒍𝒏(𝒙) est négligeable par rapport à 𝑥 en +∞. D. FAUX. On a vu que 𝑙𝑛(𝑥) était négligeable devant 𝑥, et de façon générale devant un polynôme en +∞,
donc :
𝑙𝑖𝑚𝑥 → +∞
𝑙𝑛(2 + 2𝑥 + 𝑥^2)
2 + 2𝑥 + 𝑥^2= 0
E. VRAI. C’est une forme indéterminée (0
0) : attention on ne peut pas utiliser le théorème de l’Hospital pour
une limite finie. Dans ce cas, on va faire un changement de variable :
𝑍 = 2 + 2𝑥 + 𝑥^2
Donc 𝑔(𝑍) = 𝑙𝑛(𝑍)
𝑍−1 : étudier la limite de 𝑔(𝑥) quand 𝑥 tend vers -1 revient à étudier la limite de g(Z) quand
Z tend vers 1. Si on fait le DL du numérateur et dénominateur on trouve :
𝑓(1 + 𝑥) = 0 +1
1× 𝑥 + 𝑜(𝑥) = 𝑥 + 𝑜(𝑥)
Si on fait le DL du dénominateur, on trouve :
ℎ(1 + 𝑥) = 0 + 1 × 𝑥 + 𝑜(𝑥) = 𝑥 + 𝑜(𝑥)
Donc 𝑙𝑖𝑚𝑥 → −1
𝑔(𝑥) = 𝑙𝑖𝑚𝑥 → 1
𝑔(𝑍) = 𝑙𝑖𝑚𝑥 → 1
𝑥 +𝑜(𝑥)
𝑥+𝑜(𝑥)= 1
QCM 7 – B
A. FAUX. On applique (𝑢^𝑛)′ = 𝑛 𝑢′ 𝑢^𝑛−1. Donc : 𝑑𝑀𝐵𝐹 = 230 (0,48 𝑃^−0,52𝑇^0,50 𝐴^0,13. 𝑑𝑃 + 0,50 𝑇^−0,50𝑃^0,48𝐴^−0,13. 𝑑𝑇
− 0,13 𝐴^−1,13𝑃^0,48𝑇^0,50. 𝑑𝐴) 𝑑𝑀𝐵𝐻 = 259 (0,48 𝑃^−0,52𝑇^0,50 𝐴^0,13. 𝑑𝑃 + 0,50 𝑇^−0,50𝑃^0,48𝐴^−0,13. 𝑑𝑇
− 0,13 𝐴^−1,13𝑃^0,48𝑇^0,50. 𝑑𝐴) Attention : ce n’est pas +0,13 A, mais -0,13 A.
B. VRAI. Les incertitudes absolues sont les suivantes : (𝛥𝑀𝐵𝐹)𝑚𝑎𝑥 ∼ 230 (|0,48 𝑃^−0,52𝑇^0,50 𝐴^0,13. 𝛥𝑃| + |0,50 𝑇^−0,50𝑃^0,48𝐴^−0,13. 𝛥𝑇| + |
− 0,13 𝐴^−1,13𝑃^0,48𝑇^0,50. 𝛥𝐴|) (𝛥𝑀𝐵𝐻)𝑚𝑎𝑥 ∼ 259 (|0,48 𝑃^−0,52𝑇^0,50 𝐴^0,13. 𝛥𝑃| + |0,50 𝑇^−0,50𝑃^0,48𝐴^−0,13. 𝛥𝑇| + |
− 0,13 𝐴^−1,13𝑃^0,48𝑇^0,50. 𝛥𝐴|) En effet, si P, T et A sont identiques chez un homme et une femme, la différence de l’incertitude absolue réside
dans le coefficient, or 259 > 230, donc elle sera plus élevée chez un homme que chez une femme.
C. FAUX. Si la taille et l’âge sont connus sans imprécision, 𝛥𝑇et 𝛥𝐴 valent 0. Ainsi, (𝛥𝑀𝐵𝐹)𝑚𝑎𝑥 ∼ 230 (|0,48 𝑃^−0,52𝑇^0,50 𝐴^0,13. 𝛥𝑃|).Si le poids augmente, 𝑃^−0,52diminue, donc
(𝛥𝑀𝐵𝐹)𝑚𝑎𝑥diminue. D. FAUX. Si le poids et la taille sont connus sans imprécision, 𝛥𝑃et 𝛥𝑇 valent 0. Ainsi, (𝛥𝑀𝐵𝐻)𝑚𝑎𝑥 ∼ 259 (| − 0,13 𝐴^−1,13𝑃^0,48𝑇^0,50. 𝛥𝐴|).Si l’âge augmente, 𝐴^−1,13diminue ; et le
signe - de -0,13 A disparaît avec la valeur absolue ; donc (𝛥𝑀𝐵𝐻)𝑚𝑎𝑥diminue.
Tous droits réservés au Tutorat Associatif Toulousain 22/26
E. FAUX. La seule différence entre MBF et MBH est la constante (230 et 259) ; pour les variations et
incertitudes relatives, les constantes disparaissent, donc il n’y aura pas de différences entre hommes et femmes.
QCM 8 – ADE
A. VRAI. On a 10 personnes vaccinées, donc 𝑛 = 10 ; et une allergie pour 100 vaccinations, donc 𝜋 = 0,01. B. FAUX. 𝐸(𝑋) = 𝑛𝜋 = 10 × 0,01 = 0,1. C. FAUX. 𝑉(𝑋) = 𝑛𝜋 (1 − 𝜋) = 0,1 × 0,99 = 0,099. D. VRAI. 𝐸(𝑋) = 𝑛𝜋 = 100 × 0,01 = 1. E. VRAI. Quand 𝑛 tend vers +∞, la loi binomiale converge vers la loi de Poisson. Alors, on a 𝐸(𝑋) = 𝑉(𝑋) =𝜆, le paramètre de la loi de Poisson.
QCM 9 – B
Dans la loi de Poisson, on a : 𝐸(𝑋) = 𝑉(𝑋) = 𝜆.On va donc vérifier que les espérances et les variances valent
la même valeur. A. FAUX. 𝐸(𝑎𝑋) = 𝑎 𝐸(𝑋) → 𝐸(2𝑋) = 2 𝐸(𝑋) = 2 𝜆𝑥
𝑉(𝑎𝑋) = 𝑎^2𝑉(𝑋) → 𝑉(2𝑋) = 2^2𝑉(𝑋) = 4 𝑉(𝑋) = 4 𝜆𝑥 On a 𝐸(2𝑋) ≠ 𝑉(2𝑋). B. VRAI. 𝐸(𝑋 + 𝑌) = 𝐸(𝑋) + 𝐸(𝑌) = 𝜆𝑥 + 𝜆𝑦
𝑉(𝑋 + 𝑌) = 𝑉(𝑋) + 𝑉(𝑌) + 2 𝑐𝑜𝑣(𝑋, 𝑌). Les deux variables étant indépendantes, 𝑐𝑜𝑣(𝑋, 𝑌) = 0. Donc
𝑉(𝑋 + 𝑌) = 𝑉(𝑋) + 𝑉(𝑌) = 𝜆𝑥 + 𝜆𝑦
On a 𝐸(𝑋 + 𝑌) = 𝑉(𝑋 + 𝑌). C. FAUX. 𝐸(𝑋 − 𝑌) = 𝐸(𝑋) − 𝐸(𝑌) = 𝜆𝑥 − 𝜆𝑦
𝑉(𝑋 − 𝑌) = 𝑉(𝑋) + 𝑉(𝑌) − 2 𝑐𝑜𝑣(𝑋, 𝑌). Les deux variables étant indépendantes, 𝑐𝑜𝑣(𝑋, 𝑌) = 0. Donc
𝑉(𝑋 − 𝑌) = 𝑉(𝑋) + 𝑉(𝑌) = 𝜆𝑥 + 𝜆𝑦
On a 𝐸(𝑋 − 𝑌) ≠ 𝑉(𝑋 − 𝑌). D. FAUX. 𝐸(𝑋 + 𝑎) = 𝑎 + 𝐸(𝑋) → 𝐸(𝑋 + 2) = 2 + 𝐸(𝑋) = 2 + 𝜆𝑥
𝑉(𝑋 + 𝑎) = 𝑉(𝑋) → 𝑉(𝑋 + 2) = 𝑉(𝑋) = 𝜆𝑥 On a 𝐸(𝑋 + 𝑎) ≠ 𝑉(𝑋 + 𝑎). E. FAUX. 2 (𝑋 + 𝑌) = 2𝑋 + 2𝑌.
𝐸(𝑎𝑋 + 𝑏𝑌) = 𝑎 𝐸(𝑋) + 𝑏 𝐸(𝑌) → 𝐸(2𝑋 + 2𝑌) = 2 𝐸(𝑋) + 2 𝐸(𝑌) = 2 𝜆𝑥 + 2 𝜆𝑦
𝑉(𝑎𝑋 + 𝑏𝑌) = 𝑎^2𝑉(𝑋) + 𝑏^2𝑉(𝑌) + 2 𝑎𝑏 𝑐𝑜𝑣(𝑋, 𝑌). Les deux variables étant indépendantes, 𝑐𝑜𝑣(𝑋, 𝑌) =0. Donc, 𝑉(2𝑋 + 2𝑌) = 2^2𝑉(𝑋) + 2^2𝑉(𝑌) + 2 × 2 × 2 × 0 = 4 𝑉(𝑋) + 4 𝑉(𝑌) = 4 𝜆𝑥 + 4 𝜆𝑦
On a 𝐸(2 (𝑋 + 𝑌)) ≠ 𝑉(2 (𝑋 + 𝑌)).
QCM 10 – ABD
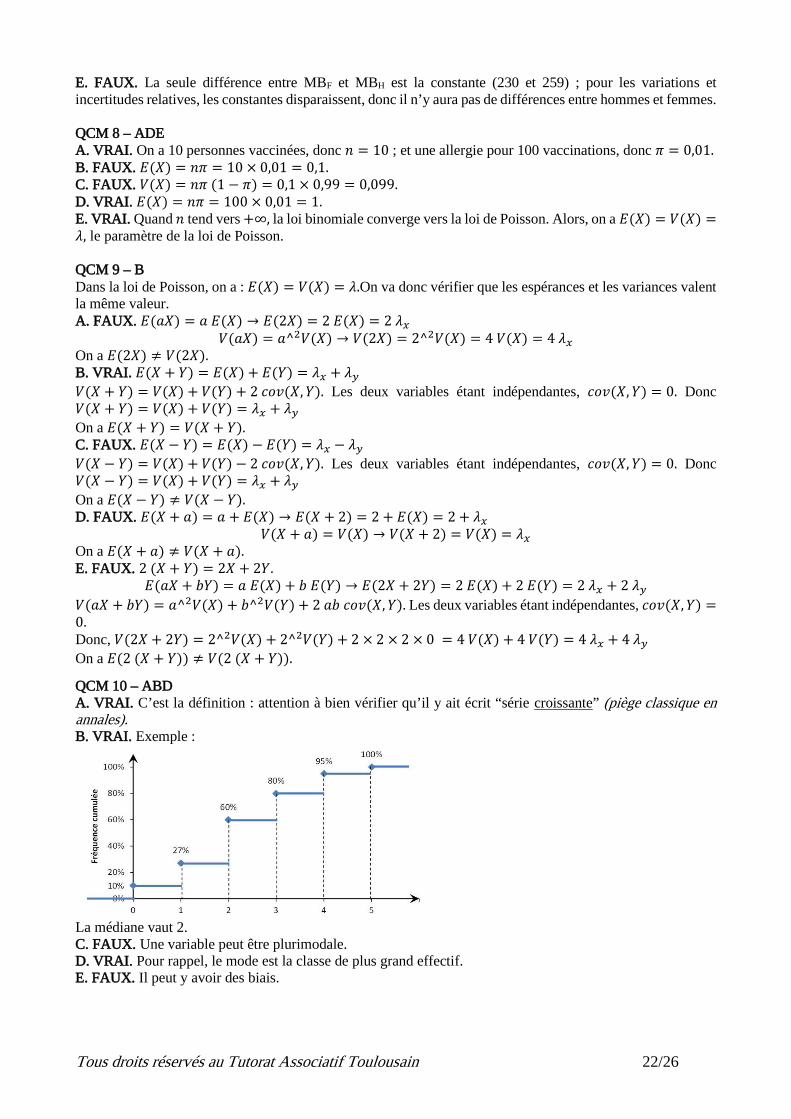
A. VRAI. C’est la définition : attention à bien vérifier qu’il y ait écrit “série croissante” (piège classique en annales). B. VRAI. Exemple :
La médiane vaut 2.
C. FAUX. Une variable peut être plurimodale. D. VRAI. Pour rappel, le mode est la classe de plus grand effectif. E. FAUX. Il peut y avoir des biais.

Tous droits réservés au Tutorat Associatif Toulousain 23/26
QCM 11 – AE
A. VRAI. Puisqu’il contient toutes les valeurs entre Q1 (25% des valeurs sont inférieures ou égales à Q1) et
Q3 (75% des valeurs sont inférieures ou égales à Q3) : 75 - 25 = 50% des valeurs. B. FAUX. On peut repérer aisément les quartiles. C. FAUX. Plus les valeurs sont dispersées, plus la variance est élevée, donc plus l’écart-type est élevé car 𝜎 =
√𝑉.
D. FAUX. 𝐶𝑉 =𝜎
𝑥: c’est le rapport de l’écart-type sur la moyenne.
E. VRAI. Car il est sans unité.
QCM 12 – C
A. FAUX. C’est une variable qualitative ordinale. B. FAUX. Par un secteur d’angle 90°, car cela correspond à 25% des valeurs, et un cercle complet vaut 360° :
0,25 x 360 = 90°. C. VRAI. “Inférieur au bac” et “master ou plus” sont les deux classes de plus grand effectif (30%). D. FAUX. On fait une moyenne avec des variables quantitatives : on ne fait pas une moyenne sans chiffres. E. FAUX. La médiane et le troisième quartile sont dans la même classe.
QCM 13 – AB
On note G l’événement “avoir la grippe” et V l’événement “être vacciné contre la grippe”. On a 𝑃(𝐺) = 0,3, 𝑃(𝑉) = 0,7, et 𝑃(𝐺|𝑉) = 0,05. A. VRAI. 𝑃(𝑉 ∩ 𝐺) = 𝑃(𝑉) × 𝑃(𝐺|𝑉) = 0,7 × 0,05 = 0,035. B. VRAI. 𝐺et 𝐺 sont deux événements contraires, donc 𝑃(𝐺) = 1 − 𝑃(𝐺) = 1 − 0,3 = 0,7.
C. FAUX. 𝑃(𝐺|𝑉) =𝑃(𝐺∩𝑉)
𝑃(𝑉)=
?
1−0,3. Pas besoin de calculer 𝑃(𝐺 ∩ 𝑉), on voit déjà que c’est une division et
non une multiplication. D. FAUX. 𝑃(𝑉) × 𝑃(𝐺) = 0,3 × 0,7 = 0,21 et 𝑃(𝑉 ∩ 𝐺) = 0,035. Ce sont deux valeurs différentes, donc les
événements ne sont pas indépendants. E. FAUX. Non, puisque 𝑃(𝑉 ∩ 𝐺) ≠ 0.(Cf item A).
QCM 14 – ACDE
On note H l’événement “être hypercholestérolémique” et D l’événement “être diabétique”. On a 𝑃(𝐻) = 0,20
et 𝑃(𝐷) = 0,10. On va se baser sur ce diagramme de Venn pour que ce soit plus clair :
Le rose correspond à 𝐷 ∪ 𝐻, et le bleu correspond à 𝐷 ∩ 𝐻.
A. VRAI. Être diabétique et ne pas être diabétique sont deux événements contraires, donc 𝑃(𝐷) = 1 − 𝑃(𝐷) =1 − 0,10 = 0,90. B. FAUX. Comme D et H sont deux événements indépendants, on calcule : 𝑃(𝐷 ∩ 𝐻) = 𝑃(𝐷) × 𝑃(𝐻) =0,10 × 0,20 = 0,02. On a ainsi :
𝑃(𝐷 ∩ 𝐻) = 1 − 𝑃(𝐷 ∪ 𝐻) = 1 − (𝑃(𝐷) + 𝑃(𝐻) − 𝑃(𝐷 ∩ 𝐻)) = 1 − (0,10 + 0,20 − 0,02) = 0,72. C. VRAI. Comme calculé dans l’item B. D. VRAI. 𝑃(𝐷 ∩ 𝐻) = 𝑃(𝐷 ∪ 𝐻) − 𝑃(𝐷) = 0,28 − 0,1 = 0,18. E. VRAI. Comme calculé dans l’item B.

Tous droits réservés au Tutorat Associatif Toulousain 24/26
QCM 15 – DE
M + M -
T + VP FP
T - FN VN
A. FAUX. Il y a 80 vrais positifs. B. FAUX. La sensibilité correspond à la probabilité d’avoir un test positif sachant qu’on est malade : 𝑆𝑒 =
𝑉𝑃
𝑉𝑃 + 𝐹𝑁=
80
100.
C. FAUX. On ne peut pas calculer la valeur prédictive positive dans la population. C’est la valeur prédictive
de l’échantillon qui vaut 𝑉𝑃𝑃 =𝑉𝑃
𝑉𝑃 + 𝐹𝑃=
80
110.
D. VRAI. La spécificité correspond à la probabilité d’avoir un test négatif sachant qu’on n’est pas malade :
𝑆𝑝𝑒 =𝑉𝑁
𝑉𝑁 + 𝐹𝑃=
70
100.
E. VRAI. Cf item C.
QCM 16 – BC
A. FAUX. La variabilité analytique pré-instrumentale. C. VRAI. Comme on a le même prélèvement, on est censé trouver toujours la même valeur s’il n’y a pas
d’erreur de mesure instrumentale. D. FAUX. Mais cela ne veut pas dire qu’il n’y a pas de variabilité intra-individuelle, on est dans le cas non-
déterminé du tableau récapitulatif. E. FAUX. Cf items C et D : elles ne proviennent que de l’erreur de mesure instrumentale. QCM 17 – AB
A. VRAI. C’est du cours. B. VRAI. Il faut prendre la première partie de l’item comme une affirmation ; donc si c’est difficile de mesurer
la pression artérielle, alors il y a effectivement des risques d’erreur de mesure instrumentale. Pour rappel, en
gros : pré-instrumentale = conditions de mesure ; instrumentale = mesure en elle-même. C. FAUX. Il y a toujours de la variabilité (et quand il y a écrit “inutile” dans un item, il est rarement vrai hein). D. FAUX. On ne prescrit pas un traitement à plusieurs individus, cette précaution ne concerne qu’un individu
à la fois : elle a pour but de réduire la variabilité intra-individuelle. E. FAUX. Il y a toujours de la variabilité, aucun instrument n’est précis à 100%.
QCM 18 – ABCE
A. VRAI. C’est le principe des tests statistiques. B. VRAI. C’est la définition des fluctuations d’échantillonnage. C. VRAI. Encore une fois, c’est le concept. D. FAUX. Non, on définit toujours l’hypothèse nulle et l’hypothèse alternative a priori. E. VRAI. On n’affirme jamais que l’hypothèse nulle est vraie : on se contente de ne pas la rejeter.

Tous droits réservés au Tutorat Associatif Toulousain 25/26
QCM 19 – ACD
C’est cette diapositive du poly qui explique (et celles qui suivent avec les cas unilatéraux)
A. VRAI. Cf diapo. B. FAUX. Elles sont dans la zone de rejet car de fréquence faible. C. VRAI. C’est la définition de la p-value : plus la p-value est faible, moins H0 est crédible. D. VRAI. 𝑧𝛼ou E. FAUX. C’est toujours une égalité.
QCM 20 – D
A. FAUX. Le risque d’erreur de première espèce 𝛼 est la probabilité de rejeter l’hypothèse nulle si l’hypothèse
nulle est vraie. La définition de l’item est celle de la puissance. B. FAUX. Le risque d’erreur de deuxième espèce 𝛽 est la probabilité de ne pas rejeter l’hypothèse nulle si
l’hypothèse alternative est vraie. C. FAUX. Il est généralement fixé a priori à 0,05. D. VRAI.
● 𝑝 < 𝛼 → rejet
● 𝑝 ≥ 𝛼 → non rejet
E. FAUX. Comme 𝑝𝑢𝑖𝑠𝑠𝑎𝑛𝑐𝑒 = 1 − 𝛽, plus le risque d’erreur de deuxième espèce 𝛽 est grand, plus la
puissance est faible.

Tous droits réservés au Tutorat Associatif Toulousain 26/26
Remerciements
Tout particulièrement, merci :
RMs Biomolécules Elian CAUBET & Juliette GAMBARA
RMs Chimie Bruno BERT & Idris MEZERAI
RMs Génome Germain GROS & Ugo VENTURIN
RMs Biologie cellulaire Antonin COLLET GOURC & Clara PUEL
RMs Embryologie/BDD Téo BERTHON & Maialen MURYN
RMs Histologie/BDR Ismaël BARKOUKI & Gaëlle SECHES
RMs Biophysique Manon BLANLUET & Sébastien VILLEY
RMs Biostatistiques Eulalie LIORZOU & Léna TARAZAGA
Ainsi qu’à Clément JEANJEAN sans qui ces corrections n’auraient pas pu exister.
Tendresse, Amour et Tutorat,
Courage




![MAHI thèse Doctorat 2017 - dspace.univ-tlemcen.dzdspace.univ-tlemcen.dz/bitstream/112/10054/1/ETUDE-THEORIQUE-D… · Orbitale Moléculaire ... La réaction de cycloaddition [4+2],](https://img.pdfslide.fr/doc/110x75/5b9d02f309d3f29a298ba9e7/mahi-these-doctorat-2017-orbitale-moleculaire-la-reaction-de-cycloaddition.jpg)
























