Embed Size (px)
Citation preview

Annalesde la
Fondation FyssenN° 27
An
nal
es F
ysse
n N
° 27
20
12
ISSN 0980-157X
fyssen-couv.indd 1 25/02/13 11:48

Annalesde la Fondation
Fyssen2012
pagetitre.indd 1 25/02/13 09:24

Directeur de la publication : Daniel LALLIER
Administration : 194, rue de Rivoli — Paris 1erTél. : 01 42 97 53 16
Secrétariat de rédaction : Julie RUBINe-mail : j.rubinsfondationfyssen.fr
Copyright : Les Annales de la Fondation Fyssen 2012Cette publication ne peut être vendue / Publication not for sale
Tousdroits réservéspour touspays : toute reproduction,diffusionoureprésentation, en toutoupartie,sous quelque forme que ce soit, est interdite sans accord préalable et écrit de la Fondation Fyssen.
All rights reserved for all countries : no part of this publicationmay be reproduced or distributed in any formor by any means without the prior written permission of the Fyssen Foundation.
GRP : fyssen JOB : annales27 DIV : 01⊕mp⊕copyright p. 1 folio : 2 --- 18/2/013 --- 14H33
2

CONSEIL SCIENTIFIQUE AU 01/01/2013
Monsieur Daniel LALLIER
Président
Monsieur Carlo SEVERI (Anthropologie)
Vice-Président, Directeur d’Études à l’E.H.E.S.S., Paris, France
Monsieur Joseph CALL (Primatologie)
Directeur du Centre de Recherche Wolfgang Köhler Primateau Max Planck Institut, Leipzig, Allemagne
Monsieur Laurent COHEN (Neurosciences cognitives)
Professeur à l’Hôpital Salpêtrière, Paris, France
Madame Patrizia D’ETTORRE (Ethologie)
Professeur à l’Université Paris 13, France
Monsieur Vittorio GALLESE (Neurophysiologie)
Professeur à l’Université de Parme, Italie
Madame Sonia GAREL (Neurobiologie)
Directeur de Recherche à l’Institut de Biologie,École Normale Supérieure, Paris, France
Monsieur Patrick HAGGARD (Neuropsychologie cognitive)
Professeur à l’University College de Londres, Grande-Bretagne
Monsieur William F. HANKS (Anthropologie cognitive)
Professeur à l’Université de Californie, Berkeley, Etats-Unis
GRP : fyssen JOB : annales27 DIV : 02⊕mp⊕conseil⊕scientifique p. 1 folio : 3 --- 18/2/013 --- 15H46
3

Monsieur Carl KNAPETT (Technologie et histoire des techniques)
Professeur à l’Université de Toronto, Canada
Monsieur Michael PETRAGLIA (Préhistoire et paléontologie humaine)
Professeur à l’Université d’Oxford, Grande-Bretagne
Madame Nathalie ROUACH (Neurobiologie)
Directeur de Recherche 2, Inserm, Collège de France, Paris, France
COMITÉ DE LECTURE DES ANNALES
DE LA FONDATION FYSSEN NUMÉRO 27
Monsieur Jean CHAVAILLON
Monsieur Michel IMBERT
Monsieur Jacques PERRIAULT
GRP : fyssen JOB : annales27 DIV : 02mp⊕conseil⊕scientifique p. 2 folio : 4 --- 27/2/013 --- 10H32
4

ANTHROPOLOGIE / ETHNOLOGIE
CHARLIER Bernard (Boursier 2010/2011) :Dessiner pour consacrer. Interprétation d’un rituel de consécration d’animaux enMongolie del’ouest ................................................................................................................................ 7
JRAISSATI Yasmine (Boursière 2010/2011) :Catégorisation de la couleur : entre l’universel et le particulier ............................................ 21
THEVENET Corinne (Boursière 2010/2011) :L’invention du cercueil au Néolithique : un changement profond dans la conception desfunérailles .......................................................................................................................... 36
ETHOLOGIE / PSYCHOLOGIE
DELL’ARICCIA Gaia (Boursière 2010/2011) :L’orientation olfactive chez les oiseaux ............................................................................... 49
DI POI Carole (Boursière 2008/2009) :La complainte du bébé phoque en Alaska : une histoire de soutien maternel ........................ 62
NEUROBIOLOGIE
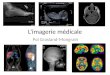
DUBOIS Jessica (Subvention de Recherche 2009) :L’Imagerie par Résonance Magnétique nous révèle quelques mystères du cerveau dunourrisson ......................................................................................................................... 89
GOAILLARD Jean-Marc (Subvention de Recherche 2008) :Robustesse de l’activité neuronale : un autre versant de la plasticité .................................... 103
LEBON Florent (Boursier 2009/2010) :Les processus cognitifs sous jacents de l’imageriemotrice – capacité d’imagerie etmécanismesinhibiteurs -. ...................................................................................................................... 113
GRP : fyssen JOB : annales27Internet DIV : 03mp⊕Sommaire⊕27Internet p. 1 folio : 5 --- 15/5/013 --- 14H20
SOMMAIRE
5

PALÉONTOLOGIE / ARCHÉOLOGIE
KAURIN Jenny (Boursière 2010/2011) :Les fibules : de la pièce unique à la fabrication en série. Innovations et évolution des procédésde fabrication d’un objet du quotidien entre le IIIe s. av. J.-C. et le Ier s. ap. J.-C. .................. 125
ZAZZO Antoine (Subvention de Recherche 2008) :Datation 14C et alimentation : une clé pour appréhender lamobilité des populations côtières dela Péninsule Arabique au Néolithique ................................................................................. 136
BIBLIOTHÈQUE ............................................................................................................. 151
GRP : fyssen JOB : annales27 DIV : 03mp⊕Sommaire⊕27 p. 2 folio : 6 --- 18/2/013 --- 17H33
SOMMAIRE
6

Dessiner pour consacrer. Interprétation d’un rituelde consécration d’animaux en Mongolie de l’ouest*
Bernard CHARLIER
Chercheur affilié au laboratoire d’anthropologie sociale du Collège de France et auMongolia and InnerAsia Studies Unit de Cambridge
RésuméLa consécration de certains animaux est ancienne en Mongolie, Gengis Khan lui-même aurait
initié cette pratique en consacrant des chevaux de course. De nos jours la consécration des animauxa pris différentes formes selon les régions et les ethnies mais le principe reste le même, il s’agit dedédier un animal à une divinité dans l’espérance d’une obtention de bénéfices. La consécrationactualise une relation à l’environnement essentiellement hiérarchique car elle reproduit à l’échellecosmique la relation de dépendance et de protection qu’un éleveur entretient avec son bétail. Parfois,au lieu de consacrer un animal vivant, le propriétaire décide d’en faire un dessin. Cet article a pourbut d’analyser le lien entre une relation singulière à l’environnement, celle de protection, et lastratégie figurative à l’œuvre dans la réalisation de l’image rituelle. L’image n’est pas envisagée icicomme une œuvre esthétique mais comme un système d’actions et de relations complexes.
Mots-clefsMongolie, consécration, fortune, environnement, image rituelle
Drawing to consecrate. Interpretation of a ritualof consecration of animals in west Mongolia*
AbstractThe consecration of animals is ancient inMongolia, Gengis Khan himself would have initiated this
practice by consecrating race horses. Nowadays the consecration of animals takes different formsaccording to the area and the ethnic group but the principle remains the same, an animal is dedicatedto a divinity in the hope of obtaining benefits. The consecration actualizes a hierarchical relationshipto the environment as it reproduces the relationship of dependence and protection the herders havewith their herds at the cosmic level. Sometimes, the owner may decide to make a drawing of aconsecrated animal rather than consecrate a live animal. This article aims at analyzing the linkbetween a particular relationship to the environment, that is, protection, and the figurative strategyat work in the realization of a ritual image. The image is not considered here as an aesthetic work butrather as a system of actions and complex relationships.
KeywordsMongolia, consecration, fortune, environment, ritual image
* Je remercie chaleureusement la Fondation Fyssen quim’a permis de réaliser cette recherche au sein d’un environnementaussi chaleureux que stimulant, le Laboratoire d’anthropologie sociale du Collège de France* I warmly thank the Fyssen Foundation which allowed me to carry out this research in a welcoming and stimulatingenvironment that is the Laboratoire d’anthropologie sociale du Collège de France.
GRP : fyssen JOB : annales27 DIV : 04⊕mp⊕CHARLIER⊕Bernard p. 1 folio : 7 --- 18/2/013 --- 14H36
ANTHROPOLOGIE / ETHNOLOGIE
7

IntroductionEn Mongolie contemporaine, les influences
mêlées du « chamanisme », du « bouddhisme »1,du socialisme et de l’économie de marché appa-rue depuis la finde la domination soviétique dansles années 90, ont donné lieu à une mosaïque deperceptionsde l’environnementnaturel.LesDör-vöd, une minorité ethnique d’éleveurs nomadesvivant principalement dans l’ouest de la Mongo-lie, mêlent à leur propre perception de la naturedes inflexions socialistes selon lesquelles les ani-maux et les ressources naturelles sont exploita-bles grâce à l’intelligence et au travail des hom-mes, et d’autres teintées d’idéologie chrétienneimportées par les nouveaux mouvements mis-sionnaires. Ces influences se retrouvent dansleurs discours religieux, scientifiques, écologi-ques et utilitaristes sur les animaux et l’environ-nement.
Cet article, inspiré des récents travaux deDescola (2010), a pour but d’analyser, parmi lesDörvöd , le lien entre un mode singulier de rela-tionà l’environnement, idéalementperçu commeétant dans un état d’équilibre, et une stratégie defiguration à l’œuvre dans l’élaboration de l’imagerituelle d’un animal consacré. L’analyse ne metpas tant en exergue la dimension esthétique del’artefact que son « agence », une intentionnalitédéléguée entrainant une séquence d’effets (Gell1998).
L’analyse se développe en trois sections. Lapremière est consacrée à la description et l’ana-lysede l’attitudedesDörvödenvers l’exploitationdes ressources naturelles. Dans la deuxième sec-tion, cette attitude estmise en relation avec l’idéede bonne fortune, hishig, et la troisième sectionexamine le lien entre la relation des Dörvöd avecl’environnement et les effets recherchés del’image de l’animal consacré. En conclusion,l’imagerituelle est considéréecommeunsystèmecomplexe d’actions et de relations (Severi 2007,2009 : 11-41).
1. L’environnement : un idéal d’équilibreParmi les Dörvöd, une perception de la nature
liée à la consécration des animaux véhicule l’idéeselon laquelle les ressources naturelles doivent
être utilisées demanière équilibrée. Cet équilibrecorrespond à un ordre moral des choses. Cetteidéologie est exprimée dans une petite légendeselon laquelle le Père Blanc, Tsagaan Aav, lemaître surnaturel du terroir et des animaux sau-vages qui y vivent, ainsi que Bouddha, ont diviséil y a longtemps la nourriture entre tous lesanimaux.Cepartagedes ressources alimentairess’est opéré pour que les hommes et les animauxqui mangeaient la même nourriture, de l’herbe,puissent tous se rassasier équitablement. Unemère humaine et sa fille mangeaient de l’herbejusqu’au jour où il n’y en eu plus assez pour unejument et son poulain. Afin de résoudre ce pro-blème, le Père Blanc et Bouddha décidèrentd’attribuer différentes nourritures aux humainset aux chevaux, de la viande aux premiers et del’herbe aux seconds.
En milieu rural les aînés rappellent souventaux jeunesdenepas couperdesarbresvivants, dene pas creuser de trous, de ne pas uriner ni selaver dans une rivière. Cela engendrerait de lapollution, buzar, et les esprits des lieux, lüs,généralement associés aux sources, enverraientdes maladies en guise de représailles. Ces don-nées font écho à celles qui ont été recueilliesquelques années plus tôt par les membres duprojet Mac Arthur (Humphrey et Sneath 1995a,1995b) initié à Cambridge sur les attitudes deconservation culturelles et environnementalesenAsieCentrale.Auxquestions : coupez-vousdesarbres et des herbes ? Si oui, comment et pour-quoi ? – les personnes interrogées dans différen-tes parties deMongolie ont répondu : «Non, nousne le faisons pas. Selon nos coutumes, nous necoupons pas de plantes ni ne tuons d’animaux sinous n’en n’avons pas besoin ». Une autreréponse était : « Nous ne coupons pas d’herbeverte autrement la nature serait fâchée ». Ouencore :Selon lesvieillespersonnes, dans lepassé il y avaitune coutume selon laquelle on pouvait couperquelques branches d’arbre, mais pas toutes sur lemême arbre. On pensait que la coupe d’un arbreengendrerait la colère des esprits locaux, lüs, etque les coupables souffriraient de maladie oumourraient. (Yenhu 1995 : 12) (c’est nous quitraduisons).La légende de la jument et de son poulain ainsi
que les prohibitions relatives aux usages desressources environnementales véhiculent l’idéeselon laquelle il faut garder l’environnement
1 Pour autant qu’on puisse regrouper sous des termes géné-riques l’importante variété des usages qui s’y apparentent.
GRP : fyssen JOB : annales27 DIV : 04⊕mp⊕CHARLIER⊕Bernard p. 2 folio : 8 --- 18/2/013 --- 14H36
ANTHROPOLOGIE / ETHNOLOGIE
8

naturel dans un état d’équilibre. Cet équilibre estun état de normalité régulé par le maître surna-turel du territoire ainsi que les esprits locaux etimplique de laisser les différentes entités del’environnement exister à leur propre façon.Dans ce contexte, en tant que partie intégrée à unsystème global, les humains peuvent vivre leurvie, ils peuvent utiliser les ressources naturellessi nécessaire, comme les animaux mangent desplantes ou d’autres animaux pour leur survie.Dans le cas des animaux domestiques, une destâches de l’éleveur est de les guider afin d’évitertoute action destructrice. Les personnes qui uti-lisent les ressources de manière excessive, pourleur propre plaisir ou par négligence, risquentd’encourir unepériodedemalchance (Humphrey1993).
2. Induire et conserver la bonne fortuneL’idée de malchance est loin de recouvrir un
terme générique désignant l’avènement d’unhasard malheureux. Il désigne une abstractiondont l’actualisation positive est traduite par leterme de fortune, ou grâce, qui est au cœur durituel de consécration. Des animaux sont consa-crés pour garder la fortune de la famille et il estimpossible d’interpréter le rituel sans prendre encompte l’écologie des relations que la gestion dela fortune implique. La relation de protectionpropre à l’élevage inclut demanière hiérarchiquedeux types de relations dialectiques inhérentes àla gestion de la fortune : la conservation et ladispersion.
Il y a plusieurs types de fortune plus ou moinsdifférenciés et actualisés selon les contextes (huvzaya, az, zol, hishig) mais l’enjeu de la consécra-tion est la protection et la circulation d’une for-tune appelée hishig.2 Cette notion est difficile àdéfinir dans la pratique. Selon Baasanjav, unéleveurDörvöd : «hishig est unepartie dequelquechose qui est bénéfique ». Selon un autre infor-mateur : « Quelqu’un qui a du hishig n’a pasforcément beaucoup d’argent ou beaucoup demoutons, il peut avoir de bons amis et des bons
enfants ». Une signification de hishig est actuali-sée en relation avec un contexte particulier,comme la chasse et l’élevage.
ChasseDans le contextede la chasse, lehishig se réfère
à la proie demandée au maître surnaturel duterritoire, le Père Blanc, ainsi qu’à la portion deviande que le chasseur doit garder pour lui et quis’appelle la hishgiin mah, la viande contenant duhishig. Selon Dorjoo, un éleveur qui pratique lachasse : « Si le chasseur nemangepas cette partieil perd sa fortune. La fortune de la chasse estappelée angyn hishig et c’est comme s’il n’avaitpas chassé du tout ». L’animal entier est consi-déré commedu hishigmais les effets de la fortunesont contenus de manière métonymique dans lapartie appelée hishgiinmah. Ces effets consistenten l’augmentation du potentiel de succès dansles futures activités du chasseur et surtout en lacapacité d’obtenir à nouveau du gibier à lachasse.
Les effets de la fortune sont produits à traversla consommation de la hishgiin mah et le partagedes autres parties dugibier. SelonDorjoo : «Si unchasseur ne partage pas sa proie avec les autreschasseurs, le Père Blanc ne lui donnera plus dugibier car il aura vu qu’il est radin ». Il y a donc uncontrôle contenu dans la mise à mort de l’animaldans la mesure où le chasseur tue à la fois pourlui-même et pour les autres. Après lamise àmort,le chasseur devient l’objet d’exigences de la partdes autres chasseurs et du Père Blanc. Le chas-seur doit partager ce qu’il ne produit pas(Hamayon 1990). Selon Chuluunbat, un autrechasseur : « lorsqu’on chasse dans une régionéloignée, un sanglier par exemple, et que sur leretour, à proximité de l’endroit où on a chassé, oncroise une personne, il faut lui donner une partiedu gibier. Mais cela n’est pas nécessaire si l’oncroise une personne près du camp ». Le partageimmédiat détermine ainsi les résultats futurs dela chasse. Cette pratique est déjà mentionnéedans l’histoire secrète des Mongols (Even et Pop1994 : 257, note 59 ; 292, note 9) mais de nosjours, elle est difficilement applicable car la plu-part des espèces chassées sont protégées. Néan-moins, il y a chez les chasseurs Mongols quiguident les riches chasseurs étrangers sur leurterritoire une réminiscence de cette pratique.Quand un chasseur étranger tue un bouquetin ilprend généralement la peau et la tête, et le reste
2 Ce terme a une histoire complexe et plonge ses ramifica-tions dans l’histoire politique et religieuse de la Mongolie.Pour plus de détails voir Hamayon (1990: 630), Even et Pop(1994: 57,§70, 263,note2),Atwood (2000: 86-139)Ruhlman(2008: 15), Charlier (2010: 35-50).
GRP : fyssen JOB : annales27 DIV : 04⊕mp⊕CHARLIER⊕Bernard p. 3 folio : 9 --- 18/2/013 --- 14H36
ANTHROPOLOGIE / ETHNOLOGIE
9

est jeté. Les chasseurs mongols se partagent laviande et certaines parties sont distribuéesparmi les familles d’éleveurs vivant dans lesenvirons. Comme le dit un chasseur : « tu doispartager ta fortune de chasse, angyn hishig, etavoir bon cœur, saihan setgeltei ».3
Bien que les règles concernant le partage del’animal et que les opinions liées à la localisationde la hishgiinmah peuvent varier d’un chasseur àl’autre, nous voyons que la gestion du hishig estau cœur de deux nécessités dialectiques que sontle partage et la conservation de relations. Dansson étude des Bouriate vivant en Mongolie del’est, Rebecca Empson utilise les idées de sépa-ration et de conservation du hishig pour analyserla façon dont une mère maintient une relationavec ses enfants dans un système de parentépatrilinéaire. La mère garde des cheveux et lescordons ombilicaux de ses enfants cachés dansun coffre. Comme Empson (2007b : 125) le dit :Le cordonombilical est à la fois unepartie actuellede la personne réalisée à travers la séparation etl’expression d’une relation qui, à travers la sépa-ration peut réapparaître : sa conservation estréaliséeà travers ledépart, etpermet lapossibilitéd’un retour. (c’est nous qui traduisons)Le cordon ombilical et les cheveux témoignent
de la relation mère-enfant et c’est la séparationqui permet la conservationde la relation sousuneautre forme. De manière similaire, dans lecontexte de la chasse, le partage de la proiedétermine les relations futures entre le chasseur,le Père Blanc, et les autres chasseurs. La sépara-tion du gibier à travers le partage actualise leseffets de la consommation de la hishgiin mah etvice-versa. Ces effets sont la garantie de chassesultérieures fructueuses. Le partage et la consom-mation appropriés supposent trois types de rela-tion : des relations à l’environnement qui impli-quent la connaissance des lieux où le hishig setrouve, des relations avec d’autres personnes quiimpliquent une politique du partage, et des rela-tions avec soi-même qui impliquent une attitudeéthique. Ces trois types de relation sont présentsdans la gestion du hishig dans un contexte d’éle-vage.
ElevageDurant l’été, une famille avaitdécidédevendre
du bétail afin d’avoir de l’argent en espèce. Avantla vente, la mère de la famille avait gardé un peude laine qu’ils avaient disposé dans un petit sacsupposé contenir le hishig de la famille car laconservation de la fortune garantit son bien-être,ainsi que la prospérité et la croissance du bétail.Selon Empson, cette fortune réside dans le bétailet s’appellemalyn hishig, la fortune du bétail. Elleest perçue comme une entité abstraite et contrai-rement au hishig de la chasse, elle n’a pas de lieuxfixes. Elle est reproduite et transmise de généra-tions en générations à travers le bétail. Les fem-mes qui vendent ou donnent du lait en gardentune petite partie pour elles comme la portion quicontient la fortune accumulée de leurs animaux(Empson 2002 : 138).
L’explication donnée par un informateur Bou-riate à Empson (2002 : 139), afin de clarifier lanotion de hishig, s’applique aussi aux Dörvöd :Si tu veux comprendre ce que signifie la fortune,imagine une construction en briques. Si tu prendsseulement une brique, il se peut que toute laconstruction s’écroule. Alors tu peux essayer detrouver la seule brique que tu peux prendre sansque l’édifice ne s’écroulemais cela risque de pren-dre toute une vie. Au lieu de ça, tu prends desprécautions et à la place d’enlever une briqueentière, tu grattes de lamatièreunpeuà la fois surdifférentes briques.De cettemanière, tu t’assuresd’ en garder toujours un peu afin de maintenir letout car il se peut que la matière recueilliecontienne la fortune. (c’est nous qui traduisons)Garder lehishigdubétail avant la vente est une
façon de retenir sa partie inaliénable, celle quigarantit la prospérité de la famille. En mêmetemps cette pratique permet au bétail vendu dedevenir une marchandise aliénable. Dans lachasse et l’élevage, le hishig ne change pas decontenu sémantique mais de forme. Bien que lafortune de chasse ne soit pas directement pro-duite et ne fasse pas l’objet d’une transmissioncomme la fortune de bétail, cette notion peut êtrequalifiée dans les deux cas de potentiel de crois-sance et de multiplication assurant le bien-être.
Les cinq organes principaux dans les abatscontiennent également du hishig : le foie, lesintestins, les poumons, le coeur et les reins. Ilssont considérés comme les cinq organes les plusimportants dans le mouton, sa meilleure partie,
3 La qualité des intentions exprimée à travers la bonté ducœur est analysée à la fin de la section en relation avec l’idéede mérite.
GRP : fyssen JOB : annales27 DIV : 04⊕mp⊕CHARLIER⊕Bernard p. 4 folio : 10 --- 18/2/013 --- 14H36
ANTHROPOLOGIE / ETHNOLOGIE
10

deej. Selon Tsetsegbal, l’épouse d’un éleveur :« Nous devons offrir les abats à d’autres person-nes comme nous offrons des bonbons et desbiscuits. Quand nous tuons un cheval ou unevache, nous pouvons aussi offrir une vertèbre. »Lesabats sontofferts sansdistinctiondeparenté.Selon Tsetsegbal, après avoir offert les abats, lesvoisins donneront aussi des abats lorsqu’ils tue-ront un mouton. Ainsi, partager son hishig n’estpas seulement un acte de générosité mais aussiune condition nécessaire pour en recevoir. Lepartage assure la production des effets du hishig,la croissance et le bien-être. Il sépare pour conte-nir.
Dans le domainede la chasse et de l’élevage, onpeut observer lamême tension constitutive entrele partage et la conservation. La relation entre lepartage et la conservation de la fortune sembleêtre déterminée par la notion bouddhiste demérite, buyan. Selon Chabros (1992 : 156) :Selon sa signification habituelle de « bonne for-tune » ou de « bénéfice », hishig apparaît fréquem-ment sous la forme composée buyan hishig sansque cela nemodifie son sens. Lemot buyan sembleintensifier la signification plutôt que de la modi-fier, et il apparaît parfois seul en gardant lemêmesens que hishig. (c’est nous qui traduisons)Explicite ou pas, la relation entre hishig et
buyan mérite d’être analysée car ces notionsentretiennent des relations de cause à effet. Lemérite, contrairement au hishig, n’est pas quel-que chose que l’on reçoit, mais quelque chose quel’on produit à travers ses propres actions consi-dérées comme « bonnes » en relation avec unenorme morale. Le terme buyan hishig impliqueune production volontaire de mérites qui estrécompensée par l’obtention de hishig. Le hishigimplique donc des actions volontaires et involon-taires - production et obtention. L’action du par-tage du hishigproduit du mérite qui lui induit unpotentiel de situation favorable ou des transfor-mations silencieuses pour le dire dans les termesde François Julien qui sont les conditions requi-ses pour l’obtention et la conservation du poten-tiel de croissance sous une autre forme (Jul-lien 2009). La relation d’entrelacs entre levolontaire et l’involontaire éclaire la notion demérite car il ne peut y avoir de production demérite sans un ordre moral extérieur qui le cons-titue et exige sa production.
Donner, c’est se constituer en personnemorale, mais l’effet du don, l’obtention de hishig,
reste toujours incertaines. Comme le dit un infor-mateur :Ce n’est pas que nous sommes cent pour centsures que nous allons recevoir du hishig parce quenous avons partagé notre hishig. Néanmoins noussommes plus ou moins sures que nous allons enrecevoir, mais nous ne savons pas quand, ni dequi. Le hishig peut venir de n’importe où, pasautomatiquement de la personne à qui on a donnédu hishig.La même attitude envers le don a été observée
par Laidlaw (2002 : 53) parmi les Jains :Faire un dan (un don gratuit) est méritoire, unacte de punya ou bon karma. En tant que tel il estsupposé, parunprocessus sur lequel personnen’ad’influence, de générer sa propre récompense,bien que personne ne sache quand et de quellemanière la bonne fortune résultante adviendra.(c’est nous qui traduisons)Ainsi le mérite résultant du don désintéressé
localise la personne au centre d’une toile cosmi-que virtuelle et impersonnelle de dons. En tantque point focal, la personne est à la fois le don-neur et le receveur, le commencement et la find’une chaine de causalité qui déploie ses effetsdans un réseau indéterminé dans le temps etl’espace. Le don déclenche le processus d’effec-tuation de la chaine de causalité, et l’espéranced’obtenir du hishig. L’obtention de hishig sanc-tionne la gestion fructueuse d’un équilibre entrel’égoïsme minimal de la conservation etl’altruisme maximal du partage permettant auhishig de déployer ses effets selon une spatialitéet une temporalité anonyme. Le bien être dans cecontexte est un signe du statut moral d’une per-sonne, de sa capacité à gérer la tension entreconservation et dispersion du hishig. Cette idéo-logie est à l’œuvre dans le rituel de consécrationd’un animal.
3. ConsécrationEn Asie centrale comme en Asie du sud (Sté-
panoff 2011 : 175-202, Holler 2002 : 207-226,Heissig 1980, Ferret 2009), le rituel de consécra-tion consiste à laisser vivre un animal d’élevageen renonçant à toute forme d’exploitation. EnMongolie, il s’agit d’un cheval, d’un bouc, d’unbélier ou d’un taureau. Consacrer un animal estune action méritoire, buyantai. L’animal estoffert à une divinité afin d’obtenir sa protectionet, corrélativement, de la bonne fortune, hishig.La consécration, également, sépare pour conte-nir.
GRP : fyssen JOB : annales27 DIV : 04⊕mp⊕CHARLIER⊕Bernard p. 5 folio : 11 --- 18/2/013 --- 14H36
ANTHROPOLOGIE / ETHNOLOGIE
11

Parmi les Dörvöd, l’animal est choisi en fonc-tion de sa couleur. Celle-ci peut varier d’unefamille à l’autremais les couleurs de prédilectionrécurrentes sont le blanc pour le bouc et le bélier,le gris pour le cheval et le noir ou le roux pour letaureau. L’animal doit toujours être jeune etmâle. Il est qualifié de hiimortoi, ce terme désignedans ce contexte son haut potentiel vital.
Lors de la consécration, un lama, prêtre boud-dhiste, est invité à la yourte, celui-ci lit un livrereligieux et brûle du genévrier en faisant troisfois le tour de l’animal dans le sens des aiguillesd’une montre. Le propriétaire fait de même.L’animal consacré est appelé seter, ce terme dési-gne les rubansde soie attachésà sesoreilles. Il estaussi appelé setertei mal, « bétail avec rubans ».L’animal consacré pâture avec les autres ani-maux du troupeau et tous les 9, 19 et 29 ducalendrier lunaire l’éleveur brûle de l’encensautour de l’animal. Un étranger ou une femmenepeuvent toucher l’animal, le hishig de la familles’en irait. Pour les mêmes raisons, ses poils nesont jamais coupés.
Le seter estmarqué comme les autres animauxdu troupeau. Lorsqu’il s’agit d’un cheval, seul lepère et le fils de la famille peuvent le monter et laselle et les étriers sont rangés près de l’auteldomestique de la yourte, du cotémasculin. L’ani-mal est donc étroitement lié à la filiation patrili-néaire car il est transmis de père en fils. Il estcastré. Les testicules sont mangés, ceux du che-val sont accrochés à sa queue et tombent enséchant. Ils ne sont pas utilisés. Parfois ils peu-vent être simplement jetés. La castration rendl’animal unique. Il signifie également le renonce-ment au bénéfice de ses éventuels petits pour letroupeau familial.
Lorsque le seterdevient trop vieux, il est décon-sacré, un autre animal est consacré, et l’ancienseter peut être mangé. Il arrive que le seter soitvolé et dans ce cas la divinité pour laquelle il étaitconsacré retire sa protection et le hishig de lafamille s’en va. Sa fonction est métonymique caril est la partie qui vaut pour le tout. Il est parfoiscomparé à une yourte qui protège les gens qui yhabitent.
Avoir un seter est source d’inquiétude, il nefaut pas le perdre ni se le faire voler sous peine desubir la malchance générée par la divinité offus-quée.C’est la raisonpour laquelle unéleveurpeutdécider de ne pas mettre de rubans à son animal
ou de faire faire un dessin, ou une statuette enbois4 de son animal consacré et de le placer surl’autel domestique dans la partie nord de layourte. Une personne, sans distinction de statut,douée en dessin, peut dessiner l’animal.
Photo de Lars Højer (2004 : 168)
Le propriétaire diminue ou évite ainsi les vols,les pertes et les attaques de loups. L’offrandevivante devient donc dans ce cas une relique.Pour Lars Højer, qui s’est intéressé aux seterparmi les Darhad de Mongolie du nord, il y a desraisons économiques et historiques à cette prati-que. Pendant l’époque socialiste, le bétail étaitcollectivisé et personne n’avait le droit de consa-crer des animaux. Les familles gardaient leurseter secret et à cette fin une personne douée endessin ou en sculpture pouvait réaliser un ani-mal.Ledessinou la statuette était alors consacré.Le dessin comportait sur son dos unmantra, uneprière bouddhiste, choisie en fonction de la divi-nité concernée (Højer 2004 : 165). C’est toujoursle cas de nos jours. L’image rituelle associe doncicônicité et indexicalité (Severi 2009 : 26). Onpeut émettre l’hypothèse selon laquelle la prière,qui n’est pas visible, actualise l’effet dudessin quiest de connecter le propriétaire à un esprit trans-cendant et ainsi de canaliser avantageusement leflux de hishig. L’interaction entre le texte et ledessin génère un effet d’enchâssement entre icô-nicité et indexicalité dans la mesure où la révéla-tion du dessin, localisé au sein d’un champdémonstratif - l’autel domestique - n’impliquepas
4 L’analyse de focalisera uniquement sur l’image rituelle.
GRP : fyssen JOB : annales27 DIV : 04⊕mp⊕CHARLIER⊕Bernard p. 6 folio : 12 --- 18/2/013 --- 14H36
ANTHROPOLOGIE / ETHNOLOGIE
12

seulement un effet visuel mais aussi un effetd’énonciationactualiséuniquementpar le regardd’un spectateur puisque le mantra reste cachéderrière le dessin.
Dans le dessin, on trouve généralement deséléments considérés comme positifs. Le seter estentourédemontagnes et depâtures verdoyantes,et localisé sous des nuages, un soleil et parfoisune lune. Il se trouve devant une table surlaquelle sont posés une ancienne théière, unelampe à huile utilisée pour la propitiation desdieux, et un pot d’encens fumant. De manièreintéressante le cheval vient de la droite vers lagauche et reproduit ainsi le mouvement du soleilqui est aussi celui de l’attraction de la fortune.Selon Batsukh, un lama qui réalise parfois desdessins de seter : « l’important est la présence desélémentsmentionnés ci-dessus sur le dessin. Peuimporte si ledessinestmaladroit ».L’assemblagedes éléments prime donc sur leurs formes icono-graphiques.5
Le dessin semble réunir les éléments dispara-tes nécessaires à la maintenance de l’environne-ment perçu dans sa globalité dans un état d’équi-libre mentionné dans la première section. Onretrouvedans la partie nordde la yourte, la partiela plus respectée, qui est aussi considérée commela plus haute, les traits schématiques idéaux d’unmonde en petit. La terre, les pâtures, le ciel, lesoleil et la lune avec entre eux les montagnes etles nuages ainsi que le seter, mi-animal, mi-of-frande, une offrande qui se trouve prolongée parla prière écrite au dos du dessin. L’image, commel’animal vivant, devient un conteneur, un cataly-seur et un véhicule de hishig, permettant unmouvement centripète vers l’éleveur et doncconnecte différents niveaux cosmologiques ren-dus visibles plutôt que représentés par le dessin.Ce que l’image actualise et donne à voir, ce nesont pas tant les traits iconographiques d’objetsfigurés mais plutôt un ordre de relations entredes éléments constituant l’environnement natu-rel. En d’autres termes, l’image rituelle actualiseune métarelation, c’est-à-dire une relation englo-bant d’autres relations. La relation de protectionenglobe les relations de partage et de conserva-
tionnécessaires à l’inductionduhishig. Elle lie demanière hiérarchique des éléments interdépen-dants localisés à différentes échelles du cosmos.Ledessin selle et révèledemanièremétonymiquedes parties du réseau de liens, ainsi que lesschèmes spatiaux et temporels qui organisentl’environnement et le transforment continuelle-ment. Une caractéristique propre à un mode derelation à l’environnement que Descola nomme« analogisme » (2005 : 280-320). A ce schèmerelationnel correspond une stratégie de figura-tionquidonne«àvoir toutà la foisque l’ensembledesexistantsest fragmentéenunepluralitéd’ins-tances et de déterminations, et qu’il existe tou-jours une voie par laquelle on pourra associercertaines de ces singularités » (2010 : 165).
Conclusions : seter et personne moraleCet article a tenté de montrer la corrélation
qu’il peut y avoir entre un mode singulier derelation à l’environnement et un mode de figura-tion. Comme l’a montré Severi (2007, 2009 :11-41), l’image rituelle n’est pas seulement cons-tituée d’un ensemble de symboles déchiffrablesmais elle est aussi un système d’actions et derelations complexes.
Le dessin pour l’éleveur implique un change-ment de visibilité. Parmi les Dörvöd rencontrés,il n’y apasde seteravecdes rubans.Pour l’éleveurqui décide de faire faire un dessin de son animal,le seter passe de l’anonymat du troupeau à unevisibilité accessible à tout visiteur dans la yourte.Ledessin créedans lepointdevueduvisiteurunevision du propriétaire singulière en récapitulantles relations qui le lient à la divinité et à lui-même.Il récapitule et réplique sous une autre forme larelationdeprotectionqui lie l’éleveurà ladivinitéet corrélativement la relation éthique réflexivegénérée par le mérite. Le dessin actualise maté-riellement la causalité entre constitution de lapersonnemorale et protection. End’autresmots,il agit comme connecteur-catalyseur et, en consé-quence, comme un effet substitué. Comme le ditStrathern (1988 : 182) :un effet substitué ... est souventmontré comme lerésultat transformé de relations magnifiées à tra-vers la réplication. Ces dernières sont donc dou-blement révélées, en tant qu’elles-mêmes et entant qu’elles-mêmes sous une autre forme. (c’estnous qui traduisons)Entrematérialité et immatérialité, visibilité et
invisibilité, le dessin, à la manière d’un artefact,
5 Bien que la réalisation du dessin ne soit pas soumise à descanons esthétiques contraignants et collectivement parta-gés, la personne qui dessine peut avoir des d’exigencesesthétiques personnelles vis-à-vis de sa réalisation.
GRP : fyssen JOB : annales27 DIV : 04⊕mp⊕CHARLIER⊕Bernard p. 7 folio : 13 --- 18/2/013 --- 14H36
ANTHROPOLOGIE / ETHNOLOGIE
13

en même temps selle et révèle des relations à lafois intérieures et extérieures à l’individu. Ilconstitue une biographie visuelle partielle, frag-mentaire et plurielle de l’individu mâle et de sesancêtres, puisque le dessin est transmis de pèreen fils, et la réfracte à la manière d’un prisme.
Pour conclure, la visibilité du seter n’est passeulement soumise à un acte de perception duvisiteur car il a une valeur supplémentaire qui estla connaissance. L’effet visuel du dessin n’est passeulement la capture d’un profil parmi d’autresdu propriétaire mais aussi un acte performatifqui consiste en la réalisation de celui-ci en tantque personne mâle méritoire, buyantai.
Bibliographiee Atwood,C.P. 2004Encyclopedia ofMongolia
and the Mongol Empire.New York: Facts on File.e Chabros, K. 1992Beckoning fortune. A study
of the Mongol dalalya ritual. Wiesbaden: OttoHarrassowitz.
e Charlier, B. 2010 Faces of the Wolf, Faces ofthe Individual. Anthropological Study of Human,Non-Human Relationships in West Mongolia.Ph.D. dissertation (unpublished): University ofCambridge.
e Descola, P. 2005 Par-delà nature et culture.Paris: Gallimard.
e Descola, P. 2010 La fabrique des images.Visions dumonde et formes de la représentation.Paris: Somogy/Musée du quai Branly.
e Empson, R. 2002 Integrating Transforma-tions: a Study of Children andDaughters-In-Law ina New Approach to Mongolian kinship. Ph.D.dissertation (unpublished): University of Cam-bridge.
e Empson, R. 2007b ’Separating and contai-ning people and things in Mongolia’ in ThinkingThrough Things: Theorising Artefacts Ethnogra-phically. A. Henare, M. Holbraad, and S. Wastell(eds.) London: Routledge Press.
e Even, M-D., Pop, R. 1994Histoire secrète desMongols. Paris: Gallimard.
e Ferret, C. 2009 Une civilisation du cheval.Les usages de l’équidé de la steppe à la taïga.Paris:Belin.
e Gell, A. 1998 Art and Agency. An Anthropo-logical Theory. Oxford: Clarendon Press.
e Hamayon, R. 1990. La chasse à l’âme. Paris:Société d’ethnologie.
e Heissig, W. 1980 The Religions of Mongolia.Berkeley: University of California Press.
e Højer, L. 2004 Dangerous Communications:Enmity, Suspense and Integration in Post Socia-list Northern Mongolia. Ph.D. dissertation (unpu-blished): University of Cambridge.
e Holler,D. 2002 ’The ritual of freeing lives’ inReligion and Secular Culture in Tibet. TibetanStudies II. H. Blezer (ed.) Proceedings of theninth seminar of the international associationfor Tibetan studies: Leiden.
e Humphrey, C. et al. 1993 ’Attitudes tonature in Mongolia and Tuva: a preliminaryreport’ Nomadic Peoples 33: 51-61.
e Humphrey, C., Sneath, D. (eds.) 1995a Cul-ture andEnvironment in InnerAsia,Volume1:ThePastoral Economy and the Environment. Cam-bridge: The White Horse Press.
e Humphrey, C., Sneath, D. (eds.) 1995b Cul-ture and Environment in Inner Asia, Volume 2:Society andCulture.Cambridge: TheWhiteHorsePress.
e Jullien, F. 2009 Les transformations silen-cieuses. Chantiers, I. Paris: Grasset.
e Laidlaw, J. 2002 ’A free gift makes nofriends’ in The Question of the Gift. Essays acrossdisciplines. London: Routledge.
e Ruhlman, S. 2008. Une curieuse patisserieen forme de semelle/A Mongolian pastry.,
e Anthropologyof Food, S0, Varia, le numéropermanent, [En ligne], mis en ligne le 15 décem-bre
e 2008. URL: http://aof.revues.org/document5593.html.
e Stépanoff,C. ’Saillances et essences. Le trai-tement cognitif de la singularité chez les éleveursde rennes tou (Sibérie méridionale)’ L’Homme,200:175-202.
e Strathern, M. 1988 The Gender of the Gift.Problems with Women and Problems with Societyin Melanesia. California:University of CaliforniaPress.
e Severi, C. 2007Leprincipe de la chimère.Uneanthropologie de la mémoire. Paris: Rue d’Ulm.
e Severi, C. 2009 ’La parole prêtée. Commentparlent les images?’ Cahiers d’anthropologiesociale, 5: 11-41.
e Yenhu, T. 1995 ’A comparative study of theattitudes of the peoples of pastoral areas of Inner
GRP : fyssen JOB : annales27 DIV : 04⊕mp⊕CHARLIER⊕Bernard p. 8 folio : 14 --- 18/2/013 --- 14H36
ANTHROPOLOGIE / ETHNOLOGIE
14

Asia towards their environments’ in Culture andEnvironment in Inner Asia: 2. C. Humphrey andD. Sneath (eds.) Cambridge: The White HorsePress.
IntroductionIn contemporary Mongolia, the combined
influences of ”shamanism“, ”Buddhism“6, socia-lismand theemergenceof themarket economyatthe end of the Soviet domination in the 90s, gaverise to a mosaic of perceptions of the naturalenvironment. The Dörvöd, who belong to an eth-nic minority of nomadic herders living in westMongolia, have diverse perceptions of natureranging from socialist ideas according to whichanimals as well as the natural environment aredominated and exploited thanks to the humanintelligence and work to other ideas conveyingthe christian division between soul/mind andbody, society and nature. These multiple influen-ces shape their religious, scientific, ecologicaland utilitarian discourses about the animals andthe environment.
This article inspired by Descola’s recent work,aims at analyzing the link between a particularway of interacting with the Dörvöd’s environ-ment, which should ideally remain in a state ofbalance, and thefigurative strategy atwork in therealization of the ritual image of a consecratedanimal. The main focus of the analysis is not somuch the aesthetic dimension of the artefact asits ”agency“, that is, an intentionality attributa-ble to those persons (and things, ...) who/whichare seen as initiating causal sequences of a par-ticular type (Gell 1998).
Thearticledivides into three sections.Thefirstsection describes and analyzes the attitude of theDörvöd towards the exploitation of the naturalresources. In the second section, this attitude isrelated to the idea of good fortune, hishig. Thethird section is devoted to the analysis of the linkbetween the relationship the Dörvöd have withthe environment and the expected effects of theritual image. In the conclusion, the ritual imageis considered as a complex system of actions andrelationships (Severi 2007, 2009: 11-41).
1. Maintaining the environment ideally in astate of balance
Among the Dörvöd, the consecration of ani-mals conveys a perception of the environmentaccording to which the use of the natural resour-cesmust be balanced. Themaintenance of a stateof balance betweenman and nature is based on amoral order. Such an ideology is expressed in alegendaboutTsagaanAav, the supernaturalmas-ter of the land and the wild animals. Buddha wassaid to have divided and given specific food to allthe animals. The sharing out of food began a longtime agowhenmenandanimals still ate the samefood. One day a mother and her daughter wereeating grass next to a mare and its foal. Themother and the daughter were eating faster thanthe horses. They ate the share of the grass reser-ved for the animals. In order to solve this pro-blem, Tsagaan Aav and Buddha decided to giveout different foodstuff to men and horses; meatfor men and grass for horses.
In the countryside, the elders often remind theyouth not to cut live trees, dig holes, urinate orwash in a river as this would generate pollution,buzar, andoffendthespiritsof the land, lüs,whichare generally located at the water sources. Theywould send diseases to take revenge. These dataecho the information collectedby the researchersof theMacArthurProject (Humphrey andSneath1995a, 1995b) on Environmental and CulturalConservation in various regions of Inner Asia.People were asked: ”Do you cut trees and gras-ses? If so, how and why?“ Here are some of thereplies the researches received: ”No, we don’t.According to our customs, we should not cut anyplants orneedlessly kill any animals“, ”Wedonotcut green grasses. If we did, nature would beangry“.
Another answer was:According to old people, in the past it was thecustom that one could cut off a few branches fromthe trees, but not all of them on one limb of a tree.Itwas said that the felling of treeswould anger theGazaryn Luust (dragon-spirit of the earth) andthat people who did so would get sick and maybedie. (Yenhu 1995: 12)The story about the conflicting relationship
between the mare, its foal and the humansconveys the idea that keeping nature in balance,a state of normality which is regulated by thespirit master of the land, means allowing the
6 Shamanism and Buddhism are used as generic termsgathering a great variety of beliefs and practices.
GRP : fyssen JOB : annales27 DIV : 04⊕mp⊕CHARLIER⊕Bernard p. 9 folio : 15 --- 18/2/013 --- 14H36
ANTHROPOLOGY / ETHNOLOGY
15

different entities of the environment to exist intheir own way. In this context, as part of anall-encompassing system, human beings areallowed to live as humans should. They can usenatural resources if necessary, just as animalsmayeatplantsorotheranimals for their survival.It is thought that people who overuse resourcesfor their own pleasure, or through negligence,may incur misfortune as a result. In the case ofdomesticated animals, the herder’s task is toguide them away from destructive activities(Humphrey et al. 1993: 51).
2. Inducing and containing good fortune”Misfortune“ is not a generic term referring to
the occurrence of an unhappy hazard but anabstraction whose positive actualization is trans-lated by the term ”fortune“, or ”grace“, whichis at the heart of the ritual of consecration.Animals are consecrated to keep the fortune ofthe family. It is impossible to interpret the ritualwithout taking into account the ecology of therelationships the management of fortune invol-ves. The relationship of protection peculiar toherding includes two other types of dialecticalrelationships involved in the management of for-tune: containment and dispersion.
There are several types of fortunemore or lessdifferentiated and actualized according to thecontexts (huv zaya, az, zol, hishig) but the stake ofthe consecration is the protection and the circu-lation of a particular type of fortune calledhishig.7 Defining this notion is difficult in prac-tice. For Baasanjav, a Dörvöd herder: ”hishig is apart of something which is beneficial to us“.According to another informant ”Somebody whohas hishig does not automatically have a lot ofmoney or sheep; he/she can have good friendsand good children.“ The hishig must be unders-tood in relation to a particular context such ashunting and herding.
HuntingWhen hunting the hishig refers to the prey
which is asked fromTsagaanAavand to the shareof meat the hunter must keep for himself, the
hishgiinmah (meat containing hishig). Accordingto Dorjoo, a herder who sometimes hunts: ”If thehunter does not eat this part, which is calledangyn hishig [hunting fortune] he loses hishishig. It is as though he had not hunted at all.“Thewhole animal is calledhishigbut the effects offortune are metonymically contained in a parti-cular piece called hishgiin mah. Consuming thispiece of meat increases the hunter’s potential ofsuccess in futureactivitiesandabove thecapacityto obtain game again.
The effects of fortune are produced throughthe consumption of the hishgiin mah and thesharing of the other parts of the game. Accordingto Dorjoo: ”If the hunter does not share theanimal he has hunted, he will not be able to huntanymore, as Tsagaan Aav will have seen that heis mean.“ There is a control embedded in the kill.Thehunterdoesnothuntonly forhimself butalsofor the others. After the kill the hunter becomesthe focus of the expectations of others and ofTsagaanAav.He does not sharewhat he does notproduce (Hamayon 1990). According to Chuluu-nbat, another hunter, ”If we have hunted a wildboar, for instance, far away onhorseback, andwemeet somebody on our way, we should give him/her a part of the animal. The piece of meat mustbe given away if we meet a person near the placewhere we hunted but it is not necessary to sharethe prey if we meet somebody near the encamp-ment.“ As if the immediate sharing was neces-sary to be able to hunt again. Although thispractice is already mentioned in the Secret His-tory of the Mongols (Even and Pop 1994: 257,note 59; 292, note 9), nowadays this practice ismuch less commonasmost species areprotected.However, among the Mongolian hunters whoguide foreign hunters, there is a reminiscence ofthis practice. When a foreign hunter kills a wildsheep, for instance, he takes the skin and thehead, and the rest is thrown away. The body isshared among the Mongolian hunters and someparts of the meat are distributed among theneighbouring yurts. As one guide mentioned:”You have to share your hunting fortune, angyn-hishig, and be good hearted, saihan setgeltei.“8
7 This term which has a complex history originates in thepolitical and religious history of Mongolia. For more detailssee Hamayon (1990: 630), Even et Pop (1994: 57, §70, 263,note2),Atwood (2000: 86-139)Ruhlman (2008: 15),Charlier(2010: 35-50).
8 The quality of intentions as expressed through the kind-ness of the heart is analyzed in relation to the idea of meritat the end of this section.
GRP : fyssen JOB : annales27 DIV : 04⊕mp⊕CHARLIER⊕Bernard p. 10 folio : 16 --- 18/2/013 --- 14H36
ANTHROPOLOGY / ETHNOLOGY
16

Although the rules concerning the sharing ofananimal and theopinions related to the locationof the hishgiin mah vary from one hunter toanother, we can see that the hishig lies at theheart of two dialectical necessities: namely, sha-ring and containing. The separation determinesthe containment of the relation. In her study ofthe Buriat living in the Bachireet district (EastMongolia) Empson uses the ideas of separationand containment of hishig to think about the waya mother maintains a relationship with her childin a patrilineal kinship system.Themother keepssome hair and the umbilical cord of her childhidden in the family chest. As Empson (2007b:125) says:The umbilical cord is both an actual part of aperson achieved through separation and anexpression of a relationship that, through separa-tion, can reappear: its containment is achievedthrough departure, but it remains in order toallow for the possibility of return.The umbilical cord and the hair accounts for
the mother-child relationship. The separationallows the containment of the relationship underanother form. Similarly, in the context of hun-ting, the sharing of the prey determines thefuture relationship between the hunter, TsagaanAav and the other hunters. It can be argued thatthe separation of the game through the sharingactualizes the effects of the consumption of thehishgiinmah, andvice-versa.These effects are theguarantee of successful hunts. The sharing pro-duces effects which are appropriated, containedby the hunter.Hemust share themeat in order tokeep the hishig, which involves three types ofrelationships: firstly, the relationship with theenvironment (he must know where the hishig is),secondly, the relationship with other persons,which involves a politics of sharing, and thirdly,the relationship with oneself, which implies anethical attitude. In connectionwithherding thesethree formsof relationship are alsopresent in themanagement of hishig.
HerdingIn summer a family decided to sell some cattle
so as to have some cash. Before the sale they tooka tuft from the goat’swoolwhich they put in a bagcontaining the hishig of the family. The contain-ment of fortune insures the well-being of thefamily, the prosperity and the growth of thecattle.According toEmpson, that fortune resides
in cattle and is called the fortune of the herd,malyn hishig. It is perceived as an abstract entity,and unlike the hunting hishig, it has no fixedlocation. It is reproduced from generations togenerations through the herd. Therefore womenwho sell or give milk, for instance, keep a little ofit for themselves as the sacred portion whichcontains the accumulative fortune of their ani-mals (Empson 2002: 138).
The explanationgivenbyaBuriat informant toEmpson (2002: 139), in order to clarify thenotionof hishig, also reflects the Dörwöd view:... if you want to understand what fortunemeans,imagine a brick building. If you take just one brickout, the whole building might fall down. You maytry to find a single brick you can take so that thebuilding still remains. Maybe in your whole lifeyou cannot find this brick, so instead you takeprecautions. You decide not to take a whole brickout but just scrape away a little at a time fromdifferentbricks. In thiswayyouensureyoualwayskeep a little back in order to maintain the whole:it may contain the fortune.Keeping the hishig of the cattle before the sale
is a way to retain the inalienable part of it, thepart which guarantees the family’s prosperity.This practice makes cattle an alienable commo-dity. In both contexts, hunting and herding, theconcept of hishig does not change in content butin form. Although the hunting fortune is notdirectly produced and is not transmitted like thefortune of the herd, this notion can be qualified inboth cases as a potential of growth and multipli-cation ensuring the well-being.
The five main organs which are eaten with theinnards are also considered to be endowed withhishig: the liver, the lungs, the intestines, theheart, and the kidneys. They are considered asthe fivemost important organs in the body of thesheep. The innards are considered to be the bestpart, deej. According to Tsetsegbal, a herder’swife: ”We have to offer the innards to otherpeople in the same way as we offer sweets andbiscuits. When we kill a horse or a cow, we canalso offer the vertebrae“. The innards were offe-red to neighbours without any kind of kinshipdistinction. According to Tsetsegbal, if neigh-bours are offered innards of an animal, theyshould give innards themselves when they kill asheep. So sharing one’shishig is not only an act ofgenerosity, but also a necessary condition toreceive hishig. The sharing ensures the produc-
GRP : fyssen JOB : annales27 DIV : 04⊕mp⊕CHARLIER⊕Bernard p. 11 folio : 17 --- 18/2/013 --- 14H36
ANTHROPOLOGY / ETHNOLOGY
17

tion of the effects of hishig, the growth of well-being. It separates in order to contain.
As far as herding and hunting are concerned,we can observe the same constitutive tensionbetween sharing and containing. The rela-tionship between the sharing and the contain-ment of fortune is one ofmerit, buyan. Accordingto Chabros (1992: 156):In its sense of ”good fortune“ or ”benefit“, kesigoccurs frequently in the compound buyan kesig,with the samemeaning. The word buyan seems tointensify the sense rather than modifying it, andoccasionally occurs on its own in the same sense.Explicit or not, the causal relationship
between hishig and buyan is interesting to ana-lyse. The merit, contrary to hishig, is not some-thing you receive, but something you producethrough actions considered to be ’good’ or inkeeping with a moral norm. The term buyanhishig involves a voluntary production of meritwhich is rewarded by obtaining hishig. The hishiginvolves voluntary and non-voluntary actions(producing and obtaining). Sharing one’shishig producesmerit,which inducesapotential ofsituationor silent transformations (Jullien2009)that are necessary conditions to obtain andcontain the potential of growth under anotherform. The interwoven relationships between thevoluntary and the involuntary highlight thenotion ofmerit as there cannot be a production ofmerit without an external moral order whichconstitutes it and demands its production.
To constitute oneself as a moral personimplies giving, but the effect of the gift (obtaininghishig) always remains uncertain. As an infor-mant says:It is not likewe are one hundred per cent sure thatwe are going to receive hishig because we sharedour hishig. Nevertheless, we are quite sure toreceive some, but we do not know when and fromwhom. The hishig can come from anywhere, notautomatically from the person to whom we gavehishig.The sameattitude to the gift has beenobserved
by Laidlaw (2002: 53) among the Jains:Making a dan [free gift] is meritorious, an act ofpunya or good karma.As such, it is expected, by anentirely impersonal process over which no onehas any influence, to bring its own reward,although one cannot know when or in what man-ner the resulting good fortune will come.
So the merit deriving from the disinterestedgift locates thepersonat theheart of a virtual andimpersonal cosmic web of gifts. As a centre, theperson is the giver as well as the receiver, thebeginning and the end of a causality chain thatdeploys its effects into a temporally and spatiallyundetermined network. The gift triggers the pro-cess of effectuation of the causality chain, and thehope for hishig. The obtaining of hishig sanctionsthe successful maintenance of a balance betweentheminimum selfishness of containment and themaximal altruism of the sharing, allowing thehishig to deploy its effects according to an anony-mous space and time. The well-being reflects themoral status of a person, which depends on his/her capacity to manage the tension between thecontainment and the dispersion of hishig. Thisideology is atwork in the ritual of consecration ofan animal.
3. ConsecrationIn Inner and South Asia (Stépanoff 2011: 175-
202, Holler 2002: 207-226, Heissig 1980, Ferret2009), the ritual of consecration consists inrenouncing any form of exploitation of a herdanimal. It must not be killed. In Mongolia, ahorse, a male goat, a male sheep or a bull can beconsecrated. The consecration is a meritorious,buyantai, action. The animal is offered to a divi-nity in order to obtain its protection and, corre-latively, good fortune, hishig. The consecra-tion separates to contain as well.
Among theDörvöd, the animal is chosenaccor-ding to its colour. The colour can vary from afamily to another but the favorite colours are thewhite for the male goat and the male sheep, thegrey for the horse and the black or the red for thebull. The animal must always be a male. It isqualified as hiimortoi, a term that refers to itshigh vital potential.
During the consecration, a lama (Buddhistpriest) is invited in the yurt. He reads a religiousbook, burns juniper and turns around the animalclockwise. Afterwards, the owner burns juniperand turns around the animal in his turn. Theanimal is called seter, meanings that silk ribbonsare attached to its ears. It is also called seterteimal, ”cattle with ribbons“. The consecrated ani-mal pastures onwith the other animals and every9th, 19th and 29th of the lunar calendar theherder burns incense around the animal. A stran-ger or a woman cannot touch it otherwise the
GRP : fyssen JOB : annales27 DIV : 04⊕mp⊕CHARLIER⊕Bernard p. 12 folio : 18 --- 18/2/013 --- 14H36
ANTHROPOLOGY / ETHNOLOGY
18

hishigwouldgoaway. Itshair arenever cut for thesame reason.
The seter is branded like the other animals ofthe herd. When the consecrated animal is ahorse, only the father and the son of the familycanmount it. The saddle and the stirrupsarekeptnear the domestic altar in the male side of theyurt. The animal is thus closely linked to thepatrilineal descent as it is transmitted fromfather to son. It is castrated. Its testicles areeaten, those of the horse are attached to its tailand fall when they are dry. They are not used.Sometimes, they can just be thrown away. Thecastration makes the animal unique and expres-ses thewill to renounce thepossible benefits fromthe offspring of the herd.
When the seter becomes too old it is deconse-crated and another animal is consecrated. Theformer setercanbeeaten. If it is stolenthedivinityto which it was offered withdraws its protectionand the hishig of the family goes away. It has ametonymic function, it is the part that stands forthe whole. The seter is sometimes compared witha yurt that protects the people who live in it.
So, to own a seter may be a source of worries.That is why a herder may decide not to attachribbons to his animal. He will rather make apicture or evenawooden statuette9 of it andplacethe picture or the statuette on the domestic altarin the northern side of the yurt. Anybodywithoutdistinctionof status,who isgifted indrawing, candraw the animal.
Photo by Lars Højer (2004: 168)
In doing so, the owner diminishes or avoid thethefts, the losses and the attacks of wolves. Thelive offering thus becomes a relic. ForLarsHøjer,who paid attention to the seter among theDarhad Mongols in north Mongolia, there areeconomic andhistorical reasons for this practice.During the socialist period, the herds were col-lectivised and people were not allowed to conse-crate an animal. A person could realize a drawingor a statuette of the seter which was kept secret.A mantra, a Buddhist prayer, chosen in relationto a particular divinity was inscribed at the backof the picture (Højer 2004: 165). It is still the casenowadays. The ritual image combines iconicitywith indexicality (Severi 2009: 26).We can arguethat the invisible prayer actualizes the effect ofthe drawing which is the connection between theowner and a transcendent spirit, and correlati-vely, the efficacious channelling of the flow ofhishig. The interaction between the text and thedrawing generates an effect of embedding oficonicity and indexicality. The drawing localizedinside a demonstrative field – the domestic altar– does not only imply a visual effect but also anenunciation act actualized uniquely through thehuman sight as the mantra remains hiddenbehind the drawing.
In the picture, we generally find elements posi-tively considered. The seter is surrounded bymountains and green pastures, and is localizedunder the clouds, the sun and sometimes themoon. It stands in front of a table onwhich an oldtea pot, a butter lamp used to propitiate thedivinities, and a pot of burning incense are loca-ted. Interestingly, the horse comes from the rightto the left and so reproduces the trajectory of thesun, which is also the direction in which thefortune is attracted. According to Batsukh, alama who sometimes realizes drawings of seter:”Themost important is the presence of the abovementioned elements in the picture. Notwithstan-ding the quality of the drawing“. The assemblageof the elements is more important than theiriconographic forms.10
The drawing seems to gather all the disparateelements which are necessary to maintain the
9 The analysis will only focus on the ritual image.
10 Although the realization of the drawing does not have tomeet any constraining and collectively shared aestheticcanons, the drawer may express personal aestheticdemands in his/her own work.
GRP : fyssen JOB : annales27 DIV : 04⊕mp⊕CHARLIER⊕Bernard p. 13 folio : 19 --- 18/2/013 --- 14H36
ANTHROPOLOGY / ETHNOLOGY
19

environment in a global state of balance as men-tioned in the first section. In the northern part ofthe yurt, which is also the ”highest“ and themostrespected one, the simplified ideal traits of aworld in miniature are gathered. The earth, thepastures, the sky, the sun and the moon with,between them, themountains and the clouds, theseter – half animal, half offering – backed up bythewritten prayer behind the picture. So, like thelive animal, the picture becomes a container, acatalyst and a vehicle for hishig allowing a centri-petal move towards the herder and, conse-quently, connects different cosmological levelsmade visible rather than represented by thedrawing. What the picture actualizes and makesvisible are not only the iconographic traits offigured objects but also a particular order of rela-tionships between the elements which constitutethe natural environment. In other words, theritual image actualizes a meta-relationship, thatis, a relationship that encompasses other rela-tionships. The relationship of protection encom-passes the relationship of sharing and contai-ning, which are necessary to the induction ofhishig. It links interdependent elements belon-ging to different cosmological scales. Thedrawing metonymically conceals and revealsparts of the network of links, as well as the tem-poral and spatial schemes which organize theenvironment and continuously transform it.This type of relationship to the environment iscalled ”analogism“ by Descola (2005: 280-320).This relational scheme is correlated to a figura-tive strategy that shows simultaneously that”l’ensemble des existants est fragmenté en unepluralitéd’instancesetdedéterminations,etqu’ilexiste toujours une voie par laquelle on pourraassocier certaines de ces singularités“ (2010:165).
Conclusions: the seter and the moralperson
This article attempted to show the correlationbetween a particular type of relationship to theenvironment and amode of figuration. As Severi
argues (2007, 2009: 11-41), the ritual image isnot a whole of symbols ready to be decipheredbut also a system of actions and complex rela-tionships.
For the herder, the drawing involves a changein visibility. The Dörvöd whom Imet had no seterwith ribbons. For the herder who decides to havehis animal drawn, the seter is no longer an ano-nymous member of the herd. It gets a visibilityaccessible to any visitor in the yurt. The drawingcreates in the point of view of the visitor aparticular perception of the owner by recapitula-ting the relationships that link him to the divinityand to himself. It recapitulates and replicatesunder another form the relationships that linkthe herder and the divinity, and correlatively, thereflexive and ethical relationship generated bythe merit. The drawing actualizes materially thecausality between the constitution of the moralperson and the relationship of protection. Inotherwords, itworks as a connector-catalyst and,as a consequence, as substituted effect. As Stra-thern (1988: 182) says:a substituted effect ... is often staged as the trans-formed outcome of relations magnified throughreplication. These latter are thus doubly revealed,as themselves and as themselves in another form.Betweenmateriality and immateriality, visibi-
lity and invisibility, the drawing conceals andreveals in the same way as an artifact the innerand outer relationships of an individual. It cons-titutes and refracts like a prism a plural fragmen-tary and partial visual biography of the maleindividual and his ancestors as it is transmittedfrom father to son.
To conclude, the visibility of the seter is notdetermined by the perception of a visitor. Vision,in this context, is not only perception, as it has acomplementary value, which is knowledge. Thevisual effect of the drawing is not only the ”cap-ture“ of one profile of the owner among others,but also a performative act, which is the realiza-tion of what he is, a male andmeritorious, buyan-tai, person.
GRP : fyssen JOB : annales27 DIV : 04⊕mp⊕CHARLIER⊕Bernard p. 14 folio : 20 --- 18/2/013 --- 14H36
ANTHROPOLOGY / ETHNOLOGY
20

Catégorisation de la couleur : entre l’universelet le particulier
Yasmina JRAISSATI
Maître de conférence Département de Philosophie Université Américaine de Beyrouth (AUB)
RésuméLa discussion sur les facteurs sous-tendant la catégorisation de la couleur s’étend sur plus d’un
siècle de recherche, et se polarise aujourd’hui entre deux positions clairement distinguées : l’univer-salisme, prôné par la théorie des termes basiques de couleur, selon laquelle les catégories de couleurse fondent sur des mécanismes perceptifs et cognitifs ; le relativisme linguistique et culturel,cristallisée autour de l’hypothèse Sapir-Whorf, qui soutient que les catégories résultent de l’usage dulangage. Dans la littérature, ces deux possibilités sont traitées comme si elles épuisaient l’espacelogique.Desmécanismes cognitifs et perceptifs, ou du langage et de la culture, seul l’un ou l’autre typede facteur est vu comme une explication possible de la catégorisation de la couleur. Dans ce qui suit,jemontre les limites de cette dichotomie, et, enme basant sur la littérature, suggère qu’une troisièmepossibilité logique ne peut pas être exclue : la catégorisation peut être due à des mécanismesperceptifs, cognitifs et linguistiques, culturels à la fois.
Mots-clésPerception, Catégories de couleur, Relativisme, Universalisme
Color categorization :between the universal and the particular
AbstractThe discussion regarding the factors underlying color categorization is over a century old. Today,
it polarizes between two clearly distinguished stands: Universalism, offered by the basic color termstheory according to which color categories are grounded on perceptual and cognitive mechanisms;linguistic and cultural relativism, that crystallizes around the Sapir-Whorf hypothesis, which arguesthat categories result from language use. In the literature, these two possibilities are treated as if theyexhausted the logical spaceofpossibilities.Furthermore, either cognitive andperceptualmechanismsor language and culture are viewed like a possible explanation of color categorization. Inwhat follows,I show the limits of this dichotomy. Taking the literature as a starting point, I suggest that a thirdlogicalpossibility cannotbeexcluded: categorizationcanbedue toperceptual, cognitive, linguistic andcultural mechanisms, at once.
KeywordsPerception, Color, Categorization, Sapir-Whorf, Relativism, Universalism
1. IntroductionNos catégories sont constitutives de notre
pensée. Nous ne mangeons jamais la mêmepomme deux fois, mais nous sommes en mesurede déterminer que ce fruit, dont la couleur, latexture et la saveur varient, appartient à la
catégorie « pomme » à travers le temps etl’espace. Dans le cas de la couleur, la questionde comment nous parvenons à déterminerl’extension de la catégorie « bleu » par exempleest d’autant plus pressante. Contrairement aucas des fruits, qui sont des entités discrètes,
GRP : fyssen JOB : annales27 DIV : 05mp⊕JRAISSATI⊕Yasmine p. 1 folio : 21 --- 25/2/013 --- 14H18
ANTHROPOLOGIE / ETHNOLOGIE
21

la description physique de la couleur en faitun continuum. Nos impressions de couleurrésultent de l’interaction des longueurs d’ondede la lumière, avec des surfaces ayant des pro-priétés de reflectance spécifiques, avec notreappareil perceptif. Les longueurs d’ondes, lesstimuli ultimes de notre impression de couleur,varient de façon continue. Et pourtant, notrerapport cognitif à la couleur est typiquementdiscontinu. Nous nommons les couleurs desobjets de façon quotidienne, et distinguonsmême certains objets entre eux en fonction deleur couleur. Si la discontinuité de la couleurn’est pas dans le monde, comment expliquernotre rapport discontinu à la couleur ? Laréponse à cette peut suivre deux directions.Puisque la discontinuité de nos catégories decouleur n’est pas dans le monde perçu, elle estdonc issue du sujet percevant. Soit elle résulte denos mécanismes perceptifs et cognitifs, univer-sels, soit elle résulte de notre langue, particu-lière.
La question spécifique du rapport entrelangage et perception remonte au moins audix-neuvième siècle. Pour les chercheurs decette époque, l’absence de terme pour « bleu »,dans le grec antique des écrits homériques(Gladstone, 1858), ou dans le sanskrit deshymnes Veda (Geiger, 1880), comme sonabsence dans la langue des insulaires duTorres Straight, en Nouvelle Guinée (Rivers,1901), indique l’insensibilité de ces populationsau bleu. Dans ce contexte théorique, la langue estle seul moyen d’accéder à l’esprit de l’êtrehumain. Langue et culture sont étroitementliées à l’esprit et au cerveau. Une culturedite primitive, ou qui ne présente pas les mêmesavancée technologique et sophistication que cellede l’homme blanc de l’Europe de l’ouest, estprimitive parce que l’esprit et le cerveaudesquels elle est issue sont primitifs, c’est-à-direqu’ils n’auraient pas atteint leur plein déve-loppement (Gobineau, 1853-1855). A l’arrière-plan de cette approche de l’homme, de sescapacités cérébrales et perceptives et de leurrelation à la culture, est un universalismeEuropéocentriste. L’homme est un être univer-sel, en développement. Toutes les races humai-nes sont vouées à achever le même accomplisse-ment culturel que celui atteint par l’homme
blanc, au bout de la chaîne de l’évolution del’espèce humaine.
C’est en réaction à un tel universalisme que lerelativisme linguistique et culturel voit le jour,soutenu par Franz Boas (Boas, 1938). Son argu-ment est que race et culture sont deux phénomè-nes indépendants. Par conséquent, une culturemoins avancée technologiquement et moinssophistiquée n’indique pas une race elle aussimoins développée. De plus, culture et langagesont des outils au service du besoin d’une popu-lation. Les sociétés vivant dans d’autres environ-nements, avec d’autres structures sociales, ontd’autres besoins. Ainsi, les cultures et les racesne peuvent être ordonnées sur une ligne évolu-tionnaire. Elles sont différentes, et donc incom-mensurables.
Whorf, élève de Boas, ainsi que de Sapiradopte ce relativisme culturel et linguistique, etle développe. Non seulement les langues et lescultures sont différentes et incommensurables,produisant des catégories linguistiques spécifi-ques aux différents besoins des différentes popu-lations, mais la langue façonne à son tour lapensée, et plus généralement les capacités intel-lectuelles, telles que la perception (Whorf, 1956).En cela consiste l’idée centrale de l’hypothèseSapir-Whorf.
Appliquée au cas de la couleur, l’hypothèseSapir-Whorf implique donc deux choses distinc-tes :– La catégorisation de la couleur est déterminéepar le langage– Les catégories de couleur déterminent la per-ception de la couleurEn 1969, Berlin et Kay publient une monogra-
phie qui apporte unnouveau rebondissement à lalittérature sur la catégorisation de la couleur(Berlin & Kay, 1969). En se tournant vers leursprédécesseurs tels Rivers, ils reconnaissent quela catégorisation de la couleur est universelle, etnon pas relative aux langues et cultures. Maiscontrairement à leurs prédécesseurs, Berlin etKay reconnaissent également avec Boas que raceet culture sont deux phénomènes indépendants.Ainsi, bien que certaines populations n’aient pasde terme pour « bleu », cela n’implique pas queleur système perceptif ne soit pas pleinementévolué, ni qu’ils ne perçoivent pas cette couleur,mais plutôt que leur lexique n’est pas aussi
GRP : fyssen JOB : annales27 DIV : 05mp⊕JRAISSATI⊕Yasmine p. 2 folio : 22 --- 25/2/013 --- 14H18
ANTHROPOLOGIE / ETHNOLOGIE
22

évolué que celui d’autres cultures. Berlin et Kayproposent donc que la catégorisation de la cou-leur est bien universelle, dans la mesure où onretrouve les mêmes catégories à travers lescultures. Cependant, ce lexique de la couleurévolue. Et cette évolution du lexique expliquepourquoi à certaines culturesmanquent certainstermes de couleur. Berlin et Kay s’opposent ainsià l’hypothèse Sapir-Whorf et proposent aucontraire :– La catégorisation de la couleur est déterminéepar nos mécanismes perceptifs– Les catégories de couleur ne déterminent pas laperception de la couleurDans les 40 ans qui nous séparent de la
publication de cette monographie, la recherchesur la catégorisation de la couleur a d’abord étédominée par la théorie des termes basiques (Kay,2002 ; Kay, 1975 ; Kay, 1999 ; Kay & Berlin,1997 ; Kay et al., 1991 ; Kay & Kempton, 1984 ;Kay&McDaniel, 1978 ;Kay&Regier, 2006 ;Kay& Regier, 2003 ; Kay & Webster, 2005 ; Kay &Cook, 2005). Dans les dix dernières annéescependant, l’hypothèse Sapir-Whorf de relati-visme linguistique connaît un regain de popula-rité, soutenu par des résultats expérimentauxrobustes concernant la deuxième thèse, celle del’influence des catégories sur la perception de lacouleur (Franklin et al., 2008 ; Franklin et al.,2008 ; Gilbert et al., 2006 ; Kay & Kempton,1984 ; Roberson et al., 2000 ; Roberson et al.,2008 ; Siok et al., 2009 ; Tan et al., 2008 ;Winawer et al., 2007).
Comme en témoigne l’histoire de la recherchesur le rapport entre culture et perception, ledébat est fortement polarisé. Si aujourd’hui uni-versalistes et relativistes convergent quant àl’influence plusieurs fois observée de la culturesur la perception, ils divergent encore sur ce qui,initialement, détermine les catégories. Les rela-tivistes soutiennent que les catégories résultentexclusivement du langage, même s’ils reconnais-sent quelques contraintes perceptuelles minima-les. Les universalistes soutiennent que les caté-gories résultent exclusivement de nosmécanismes perceptifs, même si certainesinfluences extérieures sont admises.
Pourtant à la question de qu’est-ce qui déter-mine la catégorisation, il existe trois réponsesdans l’espace logique, non pas deux : (a) Lesmécanismes perceptifs, (b) la langue et la cul-
ture, (c) les mécanismes perceptifs ainsi que lalangue et la culture. Dans ce qui suit, je voudraisdémontrer que les résultats de la littératureexistante ne peuvent pas exclure cette troisièmevoie.
2. Les limites du relativismeet de l’universalisme
Dans leur étude, Berlin et Kay démontrentl’universalité de la catégorisation de la couleurpar une comparaison interculturelle de lexiques.En se basant sur une dizaine de langues étudiéessur le terrain, et sur des données ethnographi-ques sur plus de soixante-dix langues, la théoriedes termes basiques argue qu’il existe au plusonze termes basiques de couleur (Berlin & Kay,1969). Certains lexiques, moins développés qued’autres, ont moins de termes. Cependant, tousles lexiques au même stade d’évolution, ont lesmêmes catégories de couleur, et par conséquent,des partitionnements similaires de l’espace descouleurs. Une langue de stade 1 comporte 2 caté-gories basiques, « noir » et « blanc ». Une languede stade 2 en comporte 3, « noir », « blanc » et« rouge ». Ainsi, à chaque stade de l’évolution,émerge une nouvelle catégorie dans un ordrespécifique. Après « rouge », émergent « jaune »,« vert », « bleu », puis « marron », « violet »,« orange », « rose » et « gris ».
La thèse universaliste est renforcée par uneétude étendue des lexiques de couleur : le WorldColor Survey (WCS,http ://http ://www.icsi.be-rkeley.edu/wcs/) qui recense aujourd’hui110 langues de pays non industrialisés. A traversces 110 langues, on retrouve les mêmes régula-rités (Cook et al., 2005).Deplus, les points focauxdes catégories, ou meilleur exemple des catégo-ries, sont eux aussi universels. Ils sont pris alorspour structurer les catégories.
Dès le départ, le défi principal de la théorie destermes basiques, au-delà de démontrer l’univer-salité de la catégorisation, a été de l’expliquer :Pourquoi les catégories sont-elles universelles ?Pourquoi émergent-elles dans cet ordre spécifi-que ? Depuis 1969, les défenseurs de l’universa-lisme ont soutenu que les catégories de couleurétaient universelles, parce qu’elles étaient direc-tement fondées sur nos mécanismes perceptifsde bas niveau (Kay & McDaniel, 1978). Plusprécisément, la proposition était que les pointsfocaux, aussi appelés prototypes, étaient fondéssur les couleurs pures de Hering, qu’on considé-
GRP : fyssen JOB : annales27 DIV : 05⊕mp⊕JRAISSATI⊕Yasmine p. 3 folio : 23 --- 18/2/013 --- 14H37
ANTHROPOLOGIE / ETHNOLOGIE
23

rait alors résulter de l’activité des cellules àopposition chromatique dans le corps genouillélatéral, au bout de la fibre optique (Rosch, 1972 ;Rosch, 1972 ; Rosch, 1973 ; Rosch & Olivier,1972 ; Kay & McDaniel, 1978). Ainsi, « noir »,« blanc », « rouge », « jaune », « vert » et « bleu »,sont des couleurs primaires qui émergent gra-duellement dans le lexique, et sont fondées surles réponses neuronales d’un certain type decellules. « Orange », « rose », « violet », « gris »et « marron », seraient des couleurs complexes,résultant du mélange des couleurs primaires.
A partir des années 1990, cette approche estcependant fortement mise en cause. Non seule-ment on observe certaines disparités entre leslangues dans la localisation du point focal(Regier et al., 2007), mais en plus, la théorie dela vision de la couleur sur laquelle s’appuyait lathéorie des termes basiques est révisée : laperception des couleurs pures de Hering ne peutêtre expliquée par l’activité des cellules à oppo-sition chromatique, mais par des mécanismescorticaux pour l’instant non identifiés (Abramov& Gordon, 1997 ; De Valois & De Valois, 1993).En d’autres termes, l’explication de la catégori-sation prônée par les universalistes pendant plusde vingt ans s’effondre.
Ainsi, au début des années 2000, non seule-ment la deuxième thèse universaliste est rejetée,étant donné qu’il semblerait que les catégories decouleurs déterminent la perception de la couleurcomme le suggèrent les nombreux résultats expé-rimentaux cités plus haut,mais en plus, contra lapremière thèse universaliste, rien dans notreconnaissance actuelle du système visuel ne per-met de soutenir que les catégories de couleursont déterminées par nos mécanismes percep-tifs.
Avec l’observation de l’influence du lexiquesur la perception et la modification de la théoriede la vision de la couleur, l’hypothèse Sapir-Whorf connaît un regain important. Le fait quela perception de la couleur puisse être influencéepar le lexique suggère que les catégories decouleur ne sont pas innées, et que par consé-quent, la catégorisation de la couleur ne peut pasêtre expliquée par un fondement simple sur nosmécanismes perceptifs.
La suggestion des relativistes est que les caté-gories de couleur sont déterminées par le lan-
gage. Alors que les universalistes soutiennentque la couleur x est appelée « C » parce qu’elleappartient à la catégorie C, les relativiste sou-tiennent la thèse contraire selon laquelle lacouleur x appartient à la catégorie C parce qu’elleest appelée « C ». C’est parce que certainescouleurs sont nommées « rouge », qu’elles sontcatégorisées ensemble sous « rouge ». Seull’usage des termes de couleur expliquerait pour-quoi nous partitionnons l’espace des couleurs dela façon dont nous le partitionnons. La seulecontrainte perceptuelle, est que nous catégori-sons ensemble des couleurs qui sont similaires.Une catégorie ne peut pas inclure des échan-tillons bleus et des échantillons jaunes sansinclure aussi des échantillons verts qui se trou-vent entre jaune et bleu dans notre espace per-ceptuel. Cependant, ce qui détermine l’extensionde « vert », de « bleu » et de « jaune », ce qui faitque nous traçons la frontière entre « bleu » et« vert » là où nous la traçons, est notre usage dulangage.
Ainsi, comme Boas avant eux, les relativistesou néo-Whorfiens, soutiennent que les catégo-ries de couleur sont fonction des besoins d’ungroupe donné, et ne sont pas fondés sur desmécanismes perceptifs (Davidoff, 2001 ; Davi-doff et al., 1999). Etant donné que les besoins desdifférentes communautés linguistiques et cultu-relles varient souvent, les catégories de couleurvarient elles aussi. Cependant, les données eth-nographies du WCS ne permettent pas deconclure aussi simplement à la variation inter-culturelle dans la catégorisation de la couleur.Admettant que la thèse évolutionnaire soit reje-tée, et qu’une langue qui partitionne l’espace descouleurs en 5 catégories est simplement diffé-rente d’une langue le partitionnant en 4 catégo-ries, et non pas plus développée, comment expli-quer que deux langues différentes, comportanttoutes deux 5 catégories basiques, partitionnentl’espace des couleurs de la même façon ?
Les données duWCS suggèrent que le rejet del’explication universaliste de la catégorisationselon laquelle les catégories de couleurs sontfondées sur les mécanismes perceptifs de façondirecte, n’implique pas que la thèse relativisteselon laquelle les catégories de couleur résultentdu langage, soit vraie. Si tel était le cas, une tellerégularité inter-linguistique n’aurait pas pu êtreobservée.
GRP : fyssen JOB : annales27 DIV : 05⊕mp⊕JRAISSATI⊕Yasmine p. 4 folio : 24 --- 18/2/013 --- 14H37
ANTHROPOLOGIE / ETHNOLOGIE
24

3. Les limites du nouveau modèleuniversaliste
Face à ces nouveaux défis, mais forts desdonnées du WCS, les défenseurs de l’universa-lisme dans la catégorisation de la couleur, pro-posent un nouveau modèle de catégorisation(Regier et al., 2007). Ce n’est pas que les catégo-ries de couleur soient fondées sur des mécanis-mes perceptifs de bas niveau, identifiables. Maisdans notre expérience de la couleur, certainescouleurs sont perceptivement plus saillantes qued’autres (Kay et al., 1997 ; Guest & Laar, 2000 ;Sturges & Whitfield, 1997 ; Boynton & Olson,1987).
Notre espace des couleurs est irrégulier (Jame-son & D’Andrade, 1997). Notre seuil de sensibi-lité aux variations de longueurs d’onde, varie enfonction des régions du spectre. En résultat de lastructure de nos cônes L, M et S (pour « long »,« medium » et « short » respectivement, enréférence à la sensibilité maximale aux lon-gueurs d’ondes longues, moyennes, et courtes),les courbes de sensibilité des cônes L et M sesuperposent partiellement (Churchland, 2007 ;MacAdam, 1942). Cela implique que le systèmevisuel humain est moins sensible dans lesrégions vert-bleu, que dans les régions rouge-jaune-vert. Les seuils de sensibilité sont plusélevés dans les régions vert-bleu, que dans lesrégions rouge-jaune-vert. L’irrégularité de notreespace perceptuel implique que les rapports desimilitude entre les couleurs perçues ne sont pashomogènes à travers le spectre. Dans les régionsoù le système visuel a des seuils de sensibilitéplus bas, de petites variations dans l’input sontperçues comme plus grandes que des variationsdemêmemagnitude dans les régions du systèmevisuel où les seuils de sensibilité sont plus élevés.Ces régions où la sensibilité est accrue sont dites« perceptivement saillantes », dans le contexte dunouveau modèle universaliste.
A elle seule la saillance perceptuelle ne suffitcependant pas à rendre compte de la régularitéde la catégorisation. Ainsi, le modèle universa-liste de catégorisation de la couleur propose, surla base de l’observation de plusieurs types decatégories perceptuelles, l’existence d’un méca-nisme cognitif de catégorisation optimale.
L’observation de catégories perceptuellesdévoile que les catégories regroupent les objetsles plus similaires entre eux, et séparent les
objets les plus dissimilaires entre eux. Etantdonnée l’irrégularité de l’espace perceptif descouleurs, un nombre limité de catégories opti-males sont possibles. Ainsi, l’irrégularité del’espace couplée de notre capacité à optimale-ment catégoriser mènerait à la catégorisationuniverselle de la couleur.
Dans leur étude, Regier et al. testent lemodèleproposé de façon artificielle. Un algorithme decatégorisation est proposé, qui catégoriseraitl’espace de façon optimale, en se basant sur lesrapports irréguliers de similitude à traversl’espace des couleurs. En se servant de cetalgorithme, on produit des systèmes lexicauxartificiels à 3, 4 et 5 catégories. Les systèmeslexicaux obtenus de façon artificielle sont trèssimilaires aux systèmes lexicaux naturels, com-portant 3, 4 et 5 catégories (Regier et al., 2007).Ce résultat est pris pour confirmer l’hypothèse :la régularité de la catégorisation résulte de l’inte-raction entre une capacité de catégoriser opti-malement et l’irrégularité de l’espace perceptif.
Ce modèle universaliste de catégorisationsemble prometteur, mais il demeure limité. Seu-les les six premières catégories de la séquenceévolutionnaire ont été produites par l’algorithmede catégorisation, laissant ouverte la question desi les cinq dernières catégories de la séquenceévolutionnaire seraient elles aussi produites delamême façon. Cette discrimination entre les sixpremières catégories et les cinq dernières n’estpas surprenante. En effet, dans la littérature, lesétudes prônant une approche universaliste de lacatégorisation distinguent souvent les six pre-mières couleurs (blanc, noir, rouge, jaune, vert etbleu) des cinq dernières (orange, rose, gris, violetet marron), arguant que les six premières sontperceptivement plus saillantes (Guest & Laar,2000 ; Sturges & Whitfield, 1997 ; Boynton &Olson, 1987). Selon cette suggestion, il y auraitune différence qualitative entre les deux ensem-bles de couleur.Mais commenous le verrons plusbas, cette discrimination entre les deux ensem-bles de couleur sur la base de la saillance per-ceptive est problématique.
4. Une alternativeDans leur étude de 1969, Berlin et Kay défi-
nissent les termes basiques sur la base de huitcritères, dont les quatre premiers sont les plusimportants (Berlin & Kay, 1969) : (i) ils sontmono-lexemiques ; (ii) leur extension n’est pas
GRP : fyssen JOB : annales27 DIV : 05⊕mp⊕JRAISSATI⊕Yasmine p. 5 folio : 25 --- 18/2/013 --- 14H37
ANTHROPOLOGIE / ETHNOLOGIE
25

incluse dans celle d’un autre terme basique ; (iii)ils ne s’appliquent pas à une classe restreinted’objets ; (iv) ils sont psychologiquementsaillants. De ces quatre critères, seul le dernierest absolument incontestable et incontesté. Deplus, des huit critères proposés, seul celui de lasaillance psychologique est utilisé à travers lesdifférentes études anthropologiques et psycholo-giques (Berlin & Berlin, 1975 ; Hage & Hawkes,1975 ; Hays et al., 1972 ; Pollnac, 1975 ; Maffi,1990 ; MacLaury, 1991 ; Stanlaw, 1997). Eneffet, contra (i), il a été soulevé quedans certaineslangues, aucun terme de couleur n’est mono-lexemique. Dans d’autres cas, un vocabulaire decouleur riche témoigned’un certainniveaud’abs-traction, même si ces termes s’appliquent à uneclasse restreinte d’objets, ce qui s’oppose à (iii).Enfin, le critère (ii) présuppose la notion de’terme basique’ qu’il est justement censé défi-nir (pour une critique de la définition opération-nelle des termes basiques voir : (Hickerson,1971 ; Kuschel & Monberg, 1974 ; Crawford,1982 ; Lucy, 1997 ; Saunders & Van Brakel,1997 ; Lyons, 1999).
En résultat, les termes et catégories basiquesne sont que des termes et catégories de couleurqui sont psychologiquement saillants – c’est-à-dire que ces termes viennent les premiers àl’esprit, et qu’ils ont une référence stable àtravers la communauté linguistique (Berlin &Kay, 1969).
Cependant, prise seule, la saillance psycholo-gique ne suffit pas à garantir l’universalité de lacatégorisation. En effet, la saillance psychologi-que d’un terme et d’une catégorie de couleur peuttrès bien résulter de l’influence de facteurs exter-nes, tel l’usage. Il est tout à fait envisageablequ’un terme non basique de couleur soit psycho-logiquement saillant pour un expert, tel un his-torien de l’art, oumême un artiste. La critique dela définition opérationnelle des termes basiquesde couleur dévoile les limites de cette notion, etsa dimension ad hoc. En effet, si les termesbasiques de couleur sont simplement des termespsychologiquement saillants, et étant donné quela saillance psychologique est une notion gra-duelle, quelle est la limite entre un terme basiqueet un terme non basique ? Comment décider duseuil au-delà duquel un terme n’est plus consi-déré basique ? Pour cette raison, pour les uni-versalistes, la saillance psychologique doit être
accompagnée d’une contrainte perceptive. Plusprécisément, dans la littérature, il est suggéréque la saillance psychologique d’un terme etd’une catégorie est causée par la saillance per-ceptuelle des couleurs qu’elle regroupe (Guest &Laar, 2000 ; Sturges&Whitfield, 1997 ; Boynton& Olson, 1987).
Mais cette suggestion est aussi limitée.Dans lamesure où tous les termes basiques de couleursont également psychologiquement saillants, pardéfinition, les six premiers termes de la séquenceévolutionnaire (« noir », « blanc », « rouge »,« jaune », « vert », « bleu ») autant que les cinqderniers « marron », « orange », « rose »,« violet », « gris »), comment expliquer lasaillance psychologique de ces derniers, qui neseraient pas aussi perceptivement saillants queles premiers ?
Ces dernières considérations soulève aumoins trois questions : Qu’est ce que la saillancepsychologique ? Qu’est ce que la saillance per-ceptuelle ? Si la saillance psychologique de cer-taines couleur est expliquée par leur saillanceperceptuelle exclusivement, comment rendrecompte de la saillance psychologique des cou-leurs qui ne sont pas, ou moins, perceptivementsaillantes ?
A la lumière de la littérature existante, j’aime-rais proposer que le terme « catégorie de cou-leur » est bien caractérisé par sa saillance psy-chologique. Un terme de couleur dénote unecatégorie de couleur si des échantillons de cou-leur appartiennent à une catégorie donnée, defaçon psychologiquement saillante. Dans lamesure où il est plausible que la saillance psy-chologique soit graduelle, alors la notion de« catégorie de couleur » est elle aussi graduelle.Ainsi, une catégorie est dite psychologiquementsaillante, dans la mesure où un terme de couleury réfère de façon cohérente et consensuelle.
Pour ce qui est de la notion de « saillanceperceptuelle », elle est récurrente dans la litté-rature, mais n’est généralement pas définie (voirpar exemple, Rosch, 1973 ; Sturges & Whitfield,1997, et Jameson & Alvarado, 2002 pour unediscussion de cette notion). Seuls Regier et al.pour qui cette notion joue un rôle central ladéfinissent sur la base de la similitude : sontsaillantes les couleurs dans les régions du spec-tre où les rapports de similitudes sont étroits, en
GRP : fyssen JOB : annales27 DIV : 05⊕mp⊕JRAISSATI⊕Yasmine p. 6 folio : 26 --- 18/2/013 --- 14H37
ANTHROPOLOGIE / ETHNOLOGIE
26

raison des seuils bas de sensibilité dans cetterégion (Regier et al., 2007).
Il nous suffira, dans le contexte de cet article,de contraster saillance perceptive et saillancepsychologique, en notant que la saillance percep-tive est plus étroitement liée à l’apparence d’unecouleur, telle qu’elle est expérimentée par unsujet percevant, que la saillance psychologique,qui est elle liée au système conceptuel d’unepersonne donnée. La notion de saillance percep-tive réfère à la qualité perceptive d’une couleurdonnée, par contraste à la saillance psychologi-que d’une catégorie.
Une couleur peut être dite perceptivementsaillante pour au moins trois raisons. (PER1)Une couleur est perceptivement saillante quandelle a des caractéristiques inhérentes qui larendent saillantes. Les rapports étroits de simi-litude, résultant de la structure de notre appareilperceptif, évoqués par Regier et al. entrent danscette catégorie. (PER2) Une couleur est expéri-mentée comme saillante lorsqu’elle est contras-tée avec une autre couleur. Ainsi, pour prendreun cas extrême, une tâche grise sur un fond blancou jaune est saillante. Les expériences de recher-che visuelle (Roberson et al., 2000 ; Gilbert et al.,2006 ; Drivonikou et al., 2007 ; Winawer et al.,2007 ; Franklin et al., 2008 ; Franklin et al.,2008 ; Roberson et al., 2008 ; Siok et al., 2009 ;Tan et al., 2008) et les effets pop-out (D’Zmura,1991) sont liés à cette sorte de contraste. Noterque dans ce cas, contrairement à (PER1), il n’estpas évident que les effets pop out ou la rapiditéde la discrimination résulte de la structure denotre appareil perceptif. (PER3) Enfin, une cou-leur est plus ou moins perceptivement saillantedans notre jugement de combien elle est un bonexemple d’une catégorie donnée. Ainsi, face à unobjet d’une certaine couleur, une personne don-née aura plus ou moins de facilité à juger si cetobjet est bien « rouge »–plutôt que, disons, « rosefoncé ». Une couleur est dite perceptivementsaillante dans ce cas, si elle est facilement jugéecomme étant un bon exemple de « rouge ». Dansle cas de PER3, l’apparence des couleurs et lesmécanismes psychologiques semblent tous deuxjouer un rôle. Il ne s’agit donc pas ici de dire queles mécanismes psychologiques n’ont aucun rôleà jouer dans au moins certains cas de saillanceperceptive, ni de dire que la saillance psycholo-gique est indépendante de notre appareil percep-
tuel. La notion de saillance perceptuelle réfèretant à quelque chose de déterminé par la struc-ture physiologique de notre système visuel(PER1), qu’à quelque chose qui est aussi psycho-logique, dans la mesure où une certaine couleurest jugée perceptivement saillante (PER3, etPER2dansune certainemesure). Cette approchede la saillance perceptive souligne surtout lafaçon dont la couleur apparaît au sujet, et laisseouverte la question débattue de si cette saillanceest innée ou acquise.
Ainsi, une catégorie de couleur est psycholo-giquement saillante à divers degrés. On observeque les couleurs perceptivement saillantes sontgénéralement regroupées dans des catégorieshautement psychologiquement saillantes. Onobserve aussi cependant, que les couleurs per-ceptivementmoins saillantes peuvent égalementêtre regroupées dans des catégories hautementpsychologiquement saillantes (le violet par exem-ple). L’existence d’un lien causal entre saillanceperceptuelle est saillance psychologique est plau-sible dans certains cas, mais il ne peut pas êtregénéralisé à tous les cas. Dans le cas de couleursperceptivement moins saillantes néanmoinsregroupées dans des catégories psychologique-ment saillantes, clairement, des facteurs d’uneautre nature doivent être introduits pour rendrecompte de la catégorisation. Il ne peut être excluque certains de ces facteurs soient externes.
Ce que le cadre théorique esquissé dans cetarticle sur la base de la littérature existante etétablie suggère, c’est que les contraintes sur lacatégorisationpeuvent bien être à la fois internes(ou perceptives) et externes (ou linguistiques etculturelles). Ces différents types d’influences nesont pas nécessairement mutuellement exclusifset peuvent possiblement converger et/ou intera-gir. Une nouvelle ligne de recherche qui prometd’être fructueuse s’attacherait à expliquer com-ment ces différents facteurs convergeraient versdes partitionnements assez similaires del’espace perceptif à travers les langues. Elledétaillerait aussi plus précisément les contrain-tes non perceptuelles intervenant dans la caté-gorisation.
Bibliographiee Abramov, I. & Gordon, J. (1997). In
C. L. Hardin & L. Maffi (Eds.), Physiologicalmechanisms of color vision. Cambridge: Cam-bridge University Press.
GRP : fyssen JOB : annales27 DIV : 05⊕mp⊕JRAISSATI⊕Yasmine p. 7 folio : 27 --- 18/2/013 --- 14H37
ANTHROPOLOGIE / ETHNOLOGIE
27

e Berlin, B. & Berlin, E. A. (1975). Aguarunacolor categories. American Ethnologist, 2, 61-87.
e Berlin, B. & Kay, P. (1969). Basic ColorTerms: their universality and evolution. Califor-nia: University of California Press.
e Boas, F. (1938). TheMindOfPrimitiveMan.The Macmillan Company.
e Boynton, R. M. & Olson, C. X. (1987).Locating basic colors in the OSA space. ColorResearch and Application, 12(2), 94-105.
e Churchland, P. (2007). On the reality (anddiversity) of objective colors : how color-qualiaspace is a map of reflectance-profile space. Phi-losophy of science, 74, 119-149.
e Cook, R. S., Kay, P., &Regier, T. (2005). TheWorldColorSuvery database: history anduse. InH. Cohen & C. Lefebvre (Eds.), Handbook ofcategorisation in the cognitive sciences (pp. 224-242). Amsterdam: Elsevier.
e Crawford, T.D. (1982).Defining ”basic colorterm“. Anthropological Linguistics, 24(3), 338-343.
e D’Zmura, M. (1991). Color in visual search.Vision Research, 31, 951-966.
e Davidoff, J. (2001). Language and percep-tual categorization. TICS, 5(9), 382-387.
e Davidoff, J., Davies, I., & Roberson, D.(1999). Colour categories of a stone-age tribe.Nature, 398, 203-204.
e De Valois, R. & De Valois, K. (1993). Amulti-stage color model. Vision Research, 36,833-836.
e Drivonikou, G. V., Kay, P., Regier, T., Ivry,R. B., Gilbert, A. L., Franklin, A., & Davies, I. R.L. (2007). Further evidence thatWhorfian effectsare stronger in the right visual field than the left.PNAS, 104(3), 1097-1102.
e Franklin, A., Drivonikou, G. V., Clifford, A.,Kay, P., Regier, T., & Davies, I. R. L. (2008).Lateralization of categorical perception of colorchanges with color term acquisition. PNAS, 105,18221-18225.
e Franklin, A., Pitchford, N., Hart, L., Davies,I. R. L., Clausse, S., & Jennings, S. (2008).Salience of primary and secondary colours ininfancy. British Journal of Developmental Psy-chology, 26(4), 471-483.
e Geiger, L. (1880). Contributions to the His-tory of the Development of the Human Race.London: Tubner and Company.
e Gilbert, A., Regier, T., Kay, P., & Ivry, R.(2006). Whorf hypothesis is supported in theright visual field but not the left. PNAS, 103(2),489-494.
e Gladstone, W. E. (1858). Studies on Homerand the homeric age. Oxford: Oxford UniversityPress.
e Gobineau, J. A. (1853-1855). Essai sur l’iné-galité des Races humaines. Edition Pierre Bel-fond (pour l’édition de 1967).
e Guest, S. & van Laar, D. (2000). The struc-ture of colour naming space. Vision Research,40, 723-734.
e Hage, P. & Hawkes, K. (1975). Binumariencolor categories. Ethnology, 14(3), 287-300.
e Hays,D.,Margolis, E., Naroll, R., &Perkins,D. (1972). Color Term Salience. AmericanAnthropologist, 74(5), 1107-1121.
e Hickerson, N. (1971). Review of Basic ColorTerms: Their Universality and Evolution. Inter-national Journal of Amercian Linguistics, 37(4),257-270.
e Jameson, K. A. & Alvarado, N. (2002).Differences in Color Naming and Color Saliencein Vietnamese and English. Color Research andApplication, 28(2), 113-138.
e Kay, P. (2002). Color categories are notarbitrary. Journal of Vision, 2(10), 44.
e Kay, P. (1975). Synchronic variability anddiachronic change in basic color terms. Lan-guage in Society, 4, 257-270.
e Kay, P. & Berlin, B. (1997). Scienceimperialism: There are nontrivial constraints oncolor naming. Behavioral and Brain Sciences,20(2), 196--201.
e Kay, P., Berlin, B., & Merrifield, W. (1991).Biocultural implications of systems of colornaming. Journal of Linguistic Anthropology,1(1), 12-25.
e Kay, P. & Kempton, W. (1984). What is theSapir-Whorf hypothesis ?. American Anthropo-logist, 86, 65-79.
e Kay, P. & McDaniel, C. K. (1978). Thelinguistic significance of the meanings of basiccolor terms.. Language, 54(3), 610-646.
GRP : fyssen JOB : annales27 DIV : 05⊕mp⊕JRAISSATI⊕Yasmine p. 8 folio : 28 --- 18/2/013 --- 14H37
ANTHROPOLOGIE / ETHNOLOGIE
28

e Kay, P. & Regier, T. (2006). Language,thought, and color: Recent developments.Trends in Cognitive Sciences, 10(2), 51-54.
e Kay, P. & Regier, T. (2003). Resolving thequestion of color naming universals. PNAS, 100,9085-9089.
e Kay, P., Regier, T. & Cook, R. (2005). Focalcolors are universal after all. PNAS, 102(23),8386-8391.
e Kuschel, R. & Monberg, T. (1974). ”Wedon’t talk much about colour here“: A study ofcolour semantics on Bellona Island. Man, 9,213-242.
e Lucy, J. (1997). The linguistics of ”color“. InC. L. Hardin & L. Maffi (Eds.), Color categoriesin thought and language (pp. 321-346). Cam-bridge University Press.
e MacAdam, D. L. (1942). Visual sensitivitiesto color differences in daylight. Journal of theOptical Society of America, 32, 247.
e MacLaury, R. (1991). Social and cognitivemotivations of change: Measuring variability incolor semantics. Language, 67(1), 34-62.
e Maffi, L. (1990). Somali color term evolu-tion: grammatical and semantic evidence.Anthropological linguistics, 32(3-4), 316-334.
e Pollnac, R. (1975). Intra-cultural variabilityin the structure of the subjective color lexicon inBuganda. American Ethnologist, 2(1), 89-109.
e Regier, T., Kay, P., & Khetarpal, N. (2007).Color naming reflects optimal partitions of colorspace. PNAS, 104(4), 1436-1441.
e Rivers, W. H. R. (1901). Introduction andVision. In A. C. Haddon (Ed.), Reports on theCambridge Anthropological Expedition to theTorres Straits. London: Cambridge UniversityPress.
e Roberson, D., Davies, I., & Davidoff, J.(2000). Color categories are not universal: repli-cations and new evidence from a Stone-Ageculture. Journal of Experimental Psychology:General, 129(3), 369-398.
e Roberson, D., Pak, H. S., & Hanley, J. R.(2008). Categorical perception of colour in theleft and right visual field is verbally mediated:Evidence from Korean. Cognition, 107, 752-762.
e Rosch, E. (1972). Universals in colornaming and memory. Journal of ExperimentalPsychology, 93, 1-20.
e Rosch, E. (1972). Probabilities, Sampling,and Ethnoraphic method: The case of DaniColour Names. Man, 7(3), 448-466.
e Rosch, E. (1973). Natural categories. Cogni-tive Psychology, 4(3), 328-350.
e Rosch, E. & Olivier, D. C. (1972). The struc-ture of the color space in naming andmemory fortwo languages. Cognitive Psychology, 3, 337-354.
e Saunders, B. & Van Brakel, J. (1997). Arethere non-trivial constraints on colour categori-zation ?. Behavioral and Brain Sciences, 20,167-228.
e Siok, W. T., Kay, P., Wang, S. Y. W., Chan,A. H. D., Chen, L., Luke, K.-K., & Tan, L. H.(2009). Language regions of brain are operativein color perception. PNAS, 106, 8140-8145.
e Sturges, J. & Whitfield, A. (1997). Salientfeatures of Munsell colour space as a function ofmonolexemic naming response latencies. VisionResearch, 37(3), 307-313.
e Tan, A.H. D., Chan, L. H., Khong, P. L., Yip,L. K. C., & Luke, K.-K. (2008). Language affectspatterns of brain activation associated with per-ceptual decision. PNAS, 105(10), 4004-4009.
e Whorf, B.-L. (1956). Language, thought andreality: selected writings. New York: JohnWiley& Sons.
e Winawer, J., Witthoft, N., Frank, M. C.,Wu, L., Wade, A. R., & Boroditsky, L. (2007).Russian blues reveal effects of language on colordiscrimination. PNAS, 104, 7780-7785.
1. IntroductionOur categories are constitutive of our thought
processes. We never eat the same apple twice,and yet we are able to determine whether a givenfruit, the color, texture, and taste of whichslightly varies across time and space, belongs tothe category ”apple“. In the case of color, thequestion of how we are able to determine theextension of the category ”blue“ for example, iseven more critical.
As opposed to fruit, which are discrete enti-ties, color is physically described as a continuum.Our color impressions result from the interac-tion of light wavelengthswith surface reflectanceproperties and our perceptual apparatus. Wave-lengths, the ultimate stimuli of our color impres-sions, vary continuously. Yet, our cognitive rela-tionship to color is typically discontinuous. We
GRP : fyssen JOB : annales27 DIV : 05⊕mp⊕JRAISSATI⊕Yasmine p. 9 folio : 29 --- 18/2/013 --- 14H37
ANTHROPOLOGY / ETHNOLOGY
29

name color of objects on a daily basis, and evendistinguish among certain objects based on theircolor property. If the discontinuity of color is notin the world, how can we account for our discon-tinuous relation to color? There are two direc-tions the answer to this question can take. Giventhat the discontinuity of our color categories isnot in the perceived world, it must then be due tothe perceiving subject. Thus, either discontinuityresults fromour perceptual and cognitivemecha-nisms, or it results from our particular language.
The specific question of the link between lan-guage andperception goes back to thenineteenthcentury at least. For researchers of the time, theabsence of a term for ”blue“ in the ancient Greekused in Homeric writings (Gladstone, 1858), inthe Sanskrit of the Veda hymns (Geiger, 1880),or in the languages of the Torres Straight in NewGuinea (Rivers, 1901), shows the insensitivity ofthese populations to the color blue. In this theo-retical framework, language is the only way onecan access to the human mind. Language andculture are tightly linked to mind and brain. Aso-called ”primitive“ culture, or a culture thatdoes not manifest the same technological deve-lopment and sophistication as the white man ofWestern Europe, is said to be primitive becausethe mind and brain from which it stems arethemselves primitive. In other words, they didnot attain their full development (Gobineau,1853-1855). In the background of this approachto man, to his intellectual and perceptual capa-cities and their relationship to culture, is aEurocentric universalism. Man is a universalbeing in development. All human races arebound to reach the same cultural accomplish-ment than the one reached by white men, at thetop of the evolutionary chain of the humanspecies.
Linguistic relativism, defended by FranzBoas, emerges in reaction to such a universalism(Boas, 1938). Boas’ argument is that race andculture are two independent phenomena. Conse-quently, a technologically less advanced culture,and one that is less sophisticated, does not implya less developed race. Further, culture and lan-guage are mere tools supposed to serve a givenpopulation’s needs. Different societies living indifferent environments, with different socialstructures have different needs. Thus, culturesand races cannot be arranged in an evolutionary
sequence. They are different, and thereforeincommensurable.
Whorf, a student of Boas and of Sapir, adoptsthis cultural and linguistic relativism, pushing itfurther still. Not only are languages and culturesdifferent and incommensurable, producing dif-ferent lexical categories specific to the needs ofdifferent populations, but language in turn sha-pes thought. More generally, language affectsintellectual capacities, among which perception(Whorf, 1956). Such is the essence of the Sapir-Whorf hypothesis.
When applied to the case of color, the Sapir-Whorf hypothesis implies two distinct claims:– Color categorization is determined by language– Color categories determine color perceptionIn 1969, Berlin and Kay publish a monograph
that leads to new a development in the literatureon color categorization (Berlin & Kay, 1969).Following psychologists like Rivers, theycontend that color categorization is universal,and is not relative to culture and language.However, as opposed to their universalist prede-cessors, Berlin and Kay also admit along withBoas that race and culture are two independentphenomena. Thus, although some populationslack a term for ”blue“, this does not entail thattheir perceptual system is not fully evolved, orthat they do not perceive this color. It howevershows that these populations’ color lexicon is notas evolved as that of others’. Berlin and Kaytherefore offer that color categorization is indeeduniversal, to the extent that the same categoriesare found across cultures. However, this colorlexicon evolves. And the evolution of the lexiconaccounts for the discrepancies across cultures.Berlin and Kay therefore oppose the Sapir-Whorf hypothesis and suggest:– Color categorization is determined by our per-ceptual mechanisms– Color categories do not determine color percep-tionIn the forty years following the publication of
this monograph, research on color categoriza-tion was first dominated by the Basic color termstheory (Kay, 2002; Kay, 1975; Kay, 1999; Kay &Berlin, 1997; Kay et al., 1991; Kay & Kempton,1984; Kay & McDaniel, 1978; Kay & Regier,2006; Kay & Regier, 2003; Kay &Webster, 2005;Kay & Cook, 2005). However, in the past decade,the Sapir-Whorf hypothesis of linguistic relati-
GRP : fyssen JOB : annales27 DIV : 05⊕mp⊕JRAISSATI⊕Yasmine p. 10 folio : 30 --- 18/2/013 --- 14H37
ANTHROPOLOGY / ETHNOLOGY
30

vism has regained some ground. Indeed, severalrobust empirical results were published, pertai-ning to the influence of lexical categories on colorperception, or claim 2 (Franklin et al., 2008;Franklin et al., 2008; Gilbert et al., 2006; Kay &Kempton, 1984; Roberson et al., 2000; Robersonet al., 2008; Siok et al., 2009; Tan et al., 2008;Winawer et al., 2007).
As can be seen from the history of the researchon the relationship between culture and percep-tion, the discussion is extremely polarized.Today, proponents of universalism and relati-vism may converge on the repeatedly observedinfluence of culture and language on perception,but they still diverge on what is supposed toinitially determine these categories. Relativistsargue that categories result exclusively fromlanguage use, even if they are willing to ack-nowledge some minimal perceptual constraints.Universalists contend that categories resultexclusively from perceptual mechanisms, even ifthey acknowledge some external influences.
Yet, when it comes to the question of whatdetermines categorization, there are threeanswers in the space of logical possibilities, nottwo: (a) Perceptual mechanisms; (b) languageand culture; (c) perceptual mechanisms alongwith language and culture. In what follows, Iwould like to argue that the results reported inthe literature do not exclude this third possibi-lity.
2. The limitation of relativismand of universalism
In their study, Berlin and Kay show that colorcategorization is universal through a cross-cul-tural comparison of lexicons. The Basic colorterms theory argues that there are atmost elevenbasic color categories, based on a field study ofsome twenty languages, and an analysis of pre-vious ethnographic work on over seventy-eightlanguages (Berlin & Kay, 1969). Some lexiconsare less developed than others and have fewerterms. However, all lexicons that are at the sameevolutionary stage have the same color catego-ries, and consequently, feature the same parti-tioning of the color space. A stage 1 language has2 basic categories, ”black“ and ”white“. A stage2 language has 3 basic categories, ”black“,”white“, and ”red“. At every stage of the lexicon’sevolution, a new category emerges, following a
specific order. After ”red“, emerge ”yellow“,”green“, ”blue“, and finally ”brown“, ”purple“,”orange“, ”pink“ and ”grey“.
The universalist claim is supported further byan extended study of color lexicons, the WorldColor Survey (WCS,http://http://www.icsi.ber-keley.edu/wcs/), which has to this day gathereddata on 110 languages from non industrializedcountries. Across these 110 languages, the sameregularities are found (Cook et al., 2005). Moreo-ver, category focal points, or best examples, arefound to be universal. They are therefore takento structure color categories.
From the start, the Basic color terms theory’smain challenge goes beyond showing that cate-gorization is universal, and consists in accoun-ting for it: Why are color categories universal?Why do they emerge in the lexicons followingthis constrained order? As of 1978, proponentsof universalism have suggested that color cate-gories are universal because they are directlygrounded on low-level perceptual mechanisms(Kay & McDaniel, 1978). More specifically, theirsuggestion was that category foci, also calledprototypes, are grounded on Hering’s uniquehues, then thought to result from the neuronalactivity of the opponent cells in the lateral geni-culate nucleus, at the end of the optic nerve(Rosch, 1972; Rosch, 1972; Rosch, 1973; Rosch&Olivier, 1972; Kay & McDaniel, 1978). Thus,”black“, ”white“, ”red“, ”yellow“, ”green“, and”blue“, are primary colors that emerge graduallyin the lexicon, and are grounded in the neuronalresponses of a type of cell. ”Orange“, ”pink“,”purple“, ”grey“ and ”brown“, are complex cate-gories, resulting from the mixture of the prima-ries.
Starting in the 1990’s, this approach is howe-ver called into question. Not only are discrepan-cies observed across languages when it comes tolocalizing color foci (Regier et al., 2007), but,further, the color vision theory on which reliedthe Basic color terms theory is revised. Theperception ofHering’s unique hues canno longerbe explained by the activity of the opponent cells,but by cortical mechanisms that are yet to beidentified (Abramov & Gordon, 1997; De Valois& De Valois, 1993). In other words, the accountof categorization that was put forward by propo-nents of the universalism for over twenty yearscollapses.
GRP : fyssen JOB : annales27 DIV : 05⊕mp⊕JRAISSATI⊕Yasmine p. 11 folio : 31 --- 18/2/013 --- 14H37
ANTHROPOLOGY / ETHNOLOGY
31

Thus, in the early 2000’s, based on the obser-ved fact that color categories affect color expe-rience, as suggested by the above mentionedempirical results, the second universalistic claimwas rejected. Further, the first claimaccording towhich color categories are determined by per-ceptual mechanisms is also called into question,as nothing in our actual knowledge of the visualsystem supports it.
With the observation of the influence of lexi-con on perception and the modification of colorvision theory, the Sapir-Whorf hypothesisregains some popularity. The fact that colorperception can be affected by lexicon suggeststhat color categories are not innate, and there-fore, a simple resort to perceptual mechanismscannot explain color categorization.
Relativists suggest that color categories aredetermined by language. Thus, while universa-lists argue that a given color x is called ”C“because it belongs to category C, relativistscontend the opposite view according to which acolor x belongs to category C because it is called”C“. It is because some colors are named ”red“,that they are categorized together under the label”red“. Nothing but our use of color terms canexplain why we partition the color space the waywe do. The unique perceptual constraint is thatonly colors that are similar to each other arecategorized together. One category does notinclude blue and yellow samples if it does not alsoinclude green samples that are found betweenyellow and blue in our perceptual space. Howe-ver, what determines the extension of ”green“,”blue“, and ”yellow“, and makes us draw theboundary between ”blue“ and ”green“ where wedraw it, is our language use.
As Boas before them, the relativists or neo-Whorfians, argue that color categories dependon the needs of a given group, and are notgrounded on perceptual mechanisms (Davidoff,2001; Davidoff et al., 1999). Given that the needsof different linguistic and cultural communitiesvary, color categories also vary. However, theethnographic data of the WCS does not lendsupport to cross-cultural variation. Admittingthat the evolutionary sequence is rejected, andthat a language that partitions the color spaceinto 5 categories is simply different from (andnotmore developed than) a language that partitionsthe color space into 4 categories, how can one
account for the fact that two different languagesthat both have 5 basic categories partition thecolor space in the same way?
TheWCSdata suggest that the rejection of theuniversalistic account of color categorizationaccording towhich color categories are groundedstraightforwardly in perceptual mechanisms,does not entail that the relativist claim, accor-ding to which categories result from languageuse, is true. If that were the case, such a cross-linguistic regularity could not have been obser-ved.
3. The limitations of the new universalisticmodel of categorization
Because of these new challenges, and in lightof the WCS data, proponents of universalismoffer a newmodel of color categorization (Regieret al., 2007). In this model, color categories areno longer based on identified low-level perceptualmechanisms. Rather, in our color experience,some colors are perceptually more salient thanothers (Kay et al., 1997; Guest & Laar, 2000;Sturges & Whitfield, 1997; Boynton & Olson,1987).
Our color space is irregular (Jameson &D’Andrade, 1997). The sensitivity thresholds towavelengths variation vary depending on theperceived areas of the spectrum. Resulting fromthe structure of the L, M and S cones (for ”long“,”medium“ and ”short“ wavelengths respecti-vely), the sensitivity curves of the L and M conespartially overlap (Churchland, 2007; MacAdam,1942). This causes the human visual system to beless sensitive to the green-to-blue area, than tothe red-to-yellow-and-green area. The sensitivitythresholds are higher in the green-to-blue areathan in the red-to-yellow-and-green. The fact thatour perceptual space is irregular means thatperceived similarities between colors are nothomogeneous across the spectrum. In the areaswhere the visual system’s sensitivity thresholdsare low, small input variations are perceived asbeing bigger than variations of the same magni-tude in the areaswhere the sensitivity thresholdsare higher. The areas where sensitivity is highare said to be ’perceptually salient’ in the contextof the new universalistic model.
However, perceptual saliency is not enough toaccount for the observed regularity of colorcategorization. For this reason, the universalis-
GRP : fyssen JOB : annales27 DIV : 05⊕mp⊕JRAISSATI⊕Yasmine p. 12 folio : 32 --- 18/2/013 --- 14H37
ANTHROPOLOGY / ETHNOLOGY
32

tic model of color categorization suggests, on thebasis of other perceptual categories as well, theexistence of a cognitive mechanism of optimalcategorization.
The observation of perceptual categoriesreveals that categories group together items thatare most similar to each other, and separatesitems that are most dissimilar to each other.Given the irregularity of our perceptual space,only a limited number of optimal categories ispossible. Thus, the irregularity of the perceptualspace coupled with our capacity to optimallycategorize would yield universal color categori-zation.
In their paper, Regier et al. test the proposedmodel in an artificial manner. A categorizationalgorithm is offered, which categorizes the spacein an optimal way, on the basis of irregularsimilarities across the color space.With the helpof this algorithm, artificial lexical systems featu-ring 3, 4, and 5 categories are generated. Theartificially obtained lexical systems and naturallanguages that have 3, 4, and 5 categories parti-tion the color space in very similar ways (Regieret al., 2007). This result is taken as a proof of thehypothesis: the regularity of color categorizationis an outcome of the interaction of our capacityto optimally categorize with an irregular percep-tual space.
This universalistic model seems promising,but it remains limited. Only the first six catego-ries of the evolutionary sequence have beengenerated with this categorization algorithm,leaving open the question of whether the 5 lastcategories of the evolutionary sequence wouldalso be generated in the same way. This discri-mination between the first six categories and thefive last ones is not surprising. Indeed, in theliterature, studies adopting a universalisticstand on color categorization often distinguishbetween the first six colors (white, black, red,yellow, green and blue), and the five last colors(orange, pink, grey, purple and brown). It isargued that the first six colors are perceptuallymore salient that the others (Guest &Laar, 2000;Sturges & Whitfield, 1997; Boynton & Olson,1987). According to this line of thought, there isa qualitative difference between the two sets ofcolor. However, as we will see in what follows,this distinction between the two sets of color onthe basis of perceptual saliency is problematic.
4. An alternative
In their 1969 study, Berlin and Kay definebasic color terms on the basis of 8 criteria, thefour first of which are essential (Berlin & Kay,1969). Basic color terms (i) are monolexemic; (ii)their extension is not included in that of anotherbasic color term; (iii) they do not apply to aspecific class of objects; (iv) they are psychologi-cally salient. From these four criteria, only thelast one was never questioned. Also, of the eightcriteria, only psychological saliency is usedacross anthropological and psychological studies(Berlin & Berlin, 1975; Hage & Hawkes, 1975;Hays et al., 1972; Pollnac, 1975; Maffi, 1990;MacLaury, 1991; Stanlaw, 1997).
Indeed, against (i), it was pointed out that insome languages, color terms are not monolexe-mic. In other cases, a rich color lexicon testifiesto a certain level of abstraction, even if theseterms do not apply to a restricted class of objects,as opposed to (iii). Finally, criterion (ii) presup-poses the notion of basic color terms that it isprecisely supposed to define (for a criticism ofthe definition of basic color terms, see: Hicker-son, 1971; Kuschel & Monberg, 1974; Crawford,1982; Lucy, 1997; Saunders & Van Brakel, 1997;Lyons, 1999).
As a result, basic color categories and termsare nothing but psychologically salient terms –that is, terms that come quickly tomind, and thathave a stable reference across the linguisticcommunity (Berlin & Kay, 1969). However,taken on its own, psychological saliency is notenough to guarantee the universality of catego-rization. Indeed, the psychological saliency of aterm and of a category may very well result fromthe influence of external factors, such as use. Itis perfectly imaginable that a non-basic colorterm is psychologically salient to an expert, suchas an art historian or an artist. The criticism ofthe operational definition of basic color termsreveals the limits of this notion, and its ad hocdimension. Indeed, if basic color terms are sim-ply psychologically salient terms, and given thatpsychologically saliency is a gradual notion,where does the limit between a basic term and anon-basic term lie? How can one decide thethreshold beyond which the term is no longerconsidered basic? For this reason, for universa-lists, psychological saliency has to be accompa-
GRP : fyssen JOB : annales27 DIV : 05⊕mp⊕JRAISSATI⊕Yasmine p. 13 folio : 33 --- 18/2/013 --- 14H37
ANTHROPOLOGY / ETHNOLOGY
33

nied by a perceptual constraint. More specifi-cally, it is suggested in the literature that thepsychological saliency of a term and category iscaused by the perceptual saliency of the colorsthey group together (Guest & Laar, 2000; Stur-ges & Whitfield, 1997; Boynton & Olson, 1987).
But this suggestion is also limited. To theextent that all basic color terms are equallypsychologically salient, by definition, the firstsix terms of the evolutionary sequence(”black“, ”white“, ”red“, ”yellow“, ”green“, and”blue“) as much as the last five (”brown“,”orange“, ”pink“, ”purple“, and ”grey“), how canthe last five categories be psychologically salientif they are not as perceptually salient as the firstsix?
Thus, these considerations raise at least threequestions: What is psychological saliency?Whatis perceptual saliency? If the psychologicalsaliency of some colors is explained by percep-tual saliency alone, how can we account for thepsychological saliency of colors that are not, orless, perceptually salient?
In light of the existing literature, I would liketo propose that the notion ”color category“ isindeed characterized by its psychologicalsaliency. A color term denotes a color category ifgiven color samples belong to a given colorcategory in a psychologically salient way. To theextent that it is plausible that psychologicalsaliency is gradual, then the notion of ”colorcategory“ may also be gradual as well. Thus, acategory is said to be psychologically salient tothe extent that a color term refers to a colorcategory in a coherent and consensual way.
As for ”perceptual saliency“, the notion isrecurrent in the literature, but is generally notdefined (see for example, Rosch, 1973; Sturges& Whitfield, 1997, and Jameson & Alvarado,2002 for a recent discussion of this notion).Regier et al. alone, for whom this notion plays akey role, define perceptual saliency on the basisof similarity: colors in the areas of the spectrumthat are perceived as tightly similar in reason oflow sensitivity thresholds, are perceptuallysalient (Regier et al., 2007).
In the context of this paper, it is enough for usto contrast perceptual saliency with psychologi-cal saliency, while noting that perceptualsaliency is more closely related to color appea-
rance as it is experienced by a perceiving subject,than psychological saliency, which is linked to agiven conceptual system. The notion of percep-tual saliency refers to the perceptual quality of agiven color, by contrast to the psychologicalsaliency of a category.
A color can be said to be perceptually salientfor at least three reasons. (PER1) A color isperceptually salient when it has inherent charac-teristics that make it so. The tight similarityrelations, resulting from the structure of ourperceptual apparatus evoked by Regier et al. fithere. (PER2) A color is experienced as salientwhen it is contrasted with another color. Thus,take the extreme case of a grey stain on a yelloworwhite background. Visual search tasks (Rober-son et al., 2000; Gilbert et al., 2006; Drivonikouet al., 2007;Winawer et al., 2007; Franklin et al.,2008; Franklin et al., 2008; Roberson et al., 2008;Siok et al., 2009; Tan et al., 2008) and pop-outeffects (D’Zmura, 1991) are related to this sort ofcontrast. Note that in this case, as opposed to(PER1), it is not obvious that the pop-out effects,or the speed of discrimination results from ourperceptual apparatus. (PER3) Finally, a color ismore or less perceptually salient in our judginghow good an example it is of a given category.Thus, when looking at an object of a given color,a given person will more or less easily judgewhether the given object is ”red“ – instead of say,”dark pink“. In the case of PER3, color appea-rance as well as psychological mechanisms, bothseem to play a role. The point is thus not to saythat psychological mechanisms have no role toplay in at least some cases of perceptual saliency,nor to contend that psychological saliency isindependent from our perceptual apparatus. Thenotion of perceptual saliency refers to the struc-ture of our visual system’s physiology (PER1), asmuch as it refers to something that is alsopsychological, to the extent that a given color isjudged to be perceptually salient (PER3, andperhaps PER2 to some extent). This approach toperceptual saliency mainly stresses the way acolor appears to the subject, and leaves unad-dressed the debated question of its innate oracquired nature.
Thus, a color category is psychologicallysalient at varying degrees. Perceptually salientcolors are generally grouped in categories thatare highly psychologically salient. However,
GRP : fyssen JOB : annales27 DIV : 05⊕mp⊕JRAISSATI⊕Yasmine p. 14 folio : 34 --- 18/2/013 --- 14H37
ANTHROPOLOGY / ETHNOLOGY
34

colors that are less perceptually salient maynevertheless also be grouped in highly psycholo-gically salient categories (take the case of purple,for example). The existence of a causal relationbetween perceptual saliency and psychologicalsaliency is plausible in certain cases, but cannotbe generalized to all cases. In the case of colorsthat are less perceptually salient and neverthe-less grouped in psychologically salient catego-ries, clearly, factors of a different nature need tobe introduced when accounting for them. Itcannot be excluded that some of these factorsmay be external.
The theoretical framework sketched in thispaper suggests that constraints on categoriza-tionmaywell be both internal (or perceptual) andexternal (linguistic and cultural). These differenttypes of influence are not necessarily mutuallyexclusive and can possibly converge and/ orinteract. A new promising line of research wouldconcentrate on explaining how different factorsmight converge towards a similar partitioning ofthe perceptual space, across languages. Such aresearch would also look into more specificallynon-perceptual factors that intervene in catego-rization.
GRP : fyssen JOB : annales27 DIV : 05⊕mp⊕JRAISSATI⊕Yasmine p. 15 folio : 35 --- 18/2/013 --- 14H37
ANTHROPOLOGY / ETHNOLOGY
35

L’invention du cercueil au néolithique :un changement profond dans la conception des funérailles
Corinne THEVENET
Post-doctorante UMR 8215 – Trajectoires, Maison de l’Archéologie et de l’Ethnologie, Nanterre
RésuméPar la reconstitution des funérailles, nous tentons d’appréhender le discours des sociétés préhis-
toriques sur lamort et sur elle-même.Malgré le caractère lacunaire des données archéologiques, nousrestituons une large part des gestes réalisés à l’occasion du décès d’un individu, notamment lesmodalités de dépôt des corps dans les tombes. L’apparition de défunts allongés au sein d’une culturepratiquantmajoritairement le dépôt des corps en position fléchie apparaît a priori commeune ruptureau sein des pratiques funéraires. L’exemple du Néolithique ancien d’Europe centrale et occidentaleillustre en quoi l’apparition d’une nouvelle position des corps signe une modification importante desfunérailles, puisqu’elle traduit ici l’apparitionducercueil.Un tel changement implique inévitablementune modification des funérailles et de leur déroulement.
Mots-clésNéolithique ancien, Europe centrale et occidentale, pratiques funéraires, analyse taphonomique,
position de dépôt, cercueil
The invention of the coffin in the neolithic :a profound change in the concept of funeral rituals
AbstractThrough the reconstruction of burials, we attempt to approach beliefs of the prehistoric societies
on death and on themselves. Regardless insufficient archaeological data, we reconstructed a largerpart of the gestures involved on the death of an individual, notably the types of deposit of the corpsein thegrave.Atafirst glance, theemergenceofburials inextendedpositionwithinaculturedominatedbydeposits in crouchedpositionappears to represent a rupture in the funerarypractices. Theexamplestemming from the Early Neolithic in central and western Europe illustrates how the appearance ofanewpositionof the corpse reveals substantialmodification inburial practices.Moreparticularly, theemergence of the coffin necessarily implicates a change of the burial and the related ritual.
KeywordsEarly Neolithic, Central andWestern Europe, funerary practices, taphonomic analysis, position of
the deposit, coffin
Chaque société se doit d’affronter la mort desesmembres et chacune d’entre elles apporte desréponses qui lui sont propres à travers les funé-railles. Celles-ci sont le support d’un discours quela société tient sur elle-même, non pas tant tellequ’elle est,mais telle qu’elle sepense.Pourappré-hender le discours des sociétés préhistoriques,nous ne disposons que de vestiges nécessaire-ment incomplets, des restes osseux dans une
structure le plus souvent fossoyée et comprenantéventuellement d’autres objets. Ces vestigesconstituent néanmoins le support permettant lareconstitution des gestes réalisés pour le traite-ment des défunts, que ceux-ci conduisent à laconstitution d’une sépulture ou à d’autres traite-ments post-mortem du corps ou du squelette.Bien que l’identification de ces gestes ne soit quepartielle par la naturemême des données archéo-
GRP : fyssen JOB : annales27 DIV : 06⊕mp⊕THEVENET⊕Corinne p. 1 folio : 36 --- 18/2/013 --- 14H39
ANTHROPOLOGIE / ETHNOLOGIE
36

logiques, une large part peut toutefois être abor-dée depuis le choix de l’emplacement de la tombejusqu’aux gestes réalisés après sa fermeture.Grâce à la reconstitution de ces gestes, on peuttenterd’appréhender lediscoursdeshommessurla mort. Il en va ainsi de la préparation et desmodalités de dépôt des corps dans les tombes :déposer le défunt fléchi sur le côté ou allongé surle dos répond à des prescriptions funéraires dif-férentes. Le passage d’une position de dépôt àl’autre au sein d’une même culture signe a prioriune rupture au sein des pratiques funéraires.Que peut dire l’apparition d’une nouvelle posi-tion des corps du déroulement des funéraillesdans ces sociétés ?
Une position rare au sein des pratiquesfunéraires du Néolithique ancien rubané
L’apparition des premières sociétés sédentai-res enEuropeCentrale et occidentale correspondà la culture de la Céramique Linéaire ou Rubané.Ce complexe culturel se développe à partir desplaines hongroises durant la seconde moitié du6e millénaire et atteint les côtes de la Manchedurant le 5e millénaire (Lichardus et al. 1985 ;Strien et Gronenborn 2005). Malgré cette consi-dérable extension géographique (fig. 1), ces pre-mières populations agro-pastorales conserventune identité forte à travers leur culture maté-rielle, notamment l’architecture des maisons etles moyens de subsistance, tout en sachant
Figure 1. Extension maximale de la Céramique Linéaire en Europe tempérée (5500-4900 BC).
GRP : fyssen JOB : annales27 DIV : 06⊕mp⊕THEVENET⊕Corinne p. 2 folio : 37 --- 18/2/013 --- 14H39
ANTHROPOLOGIE / ETHNOLOGIE
37

s’adapter aux nouveaux environnements coloni-sés.
Que ce soit dans les nécropoles ou parmi lessépultures en habitat, l’inhumation primaire estle mode de dépôt largement prédominant.L’interprétation longtemps retenue a été celled’un mode d’inhumation le plus simple qui soit :un défunt déposé dans une fosse ovalaire creuséeaux dimensions d’un corps fléchi sur le côté leplus souvent et immédiatement remblayée après.Or, le cas du Bassin parisien montre que cettesimplicité n’est qu’apparente : les tombes répon-dent à une structuration complexe qui maintientun espace vide autour du corps (Thevenet 2004 ;2010). La présence de défunts en position allon-gée n’a pas modifié la conception d’un moded’inhumation sommaire, tant leur présence ausein des nécropoles rubanées semble rare :N. Nieszery dénombrait ainsi moins de 5 % desdéfunts inhumés dans cette position en EuropeCentrale (Nieszery 1995 : 82). Ils sont pourtantloin d’être anecdotiques enAlsace, où ils peuventconstituer près de la moitié des défunts danscertaines nécropoles (par exemple, 41 des111 défunts à Vendenheim, Boës et al. 2007). Defait, ces défunts en position allongée se concen-trent dans les nécropoles de la plaine du Rhinsupérieur (Alsace, Bade-Wurtemberg) et dansquelques ensembles funéraires de Bavière orien-tale (fig. 1). Dans la plupart de ces nécropoles, àl’exception de certains ensembles funérairesalsaciens, ils représentent une minorité.
Que traduit une telle rupture ?Unanimement considérée comme une excep-
tion à la règle, la position allongée des défunts estgénéralement perçue comme un phénomène tar-dif au sein de la Céramique Linéaire. Si lesmobiliers ne sont pas toujours présents pourattester cette attribution, celle-ci repose implici-tement sur le fait que cette position devient lanorme dans les cultures qui succèdent à la Céra-mique Linéaire dans les régions rhénanes (Hin-kelstein, Großgartach, Rössen).
On l’a mentionné, ce fait n’est pas si anecdo-tique selon les régions. Quant au moment de sonapparition, celui-ci ne fait pas consensus : tardifpour certains (Moddermann 1970, Peschel1992), dès l’étape récente pour d’autres (Jeu-nesse 2006). Il est encore possible que cetteinnovation ne soit pas apparue simultanémentdans toutes les régions de la Céramique Linéaire
ou qu’elle résulte d’une diffusion à l’intérieur ducomplexe culturel. Quoi qu’il en soit, la positionallongée des défunts constitue une rupture parrapport à la norme. La principale hypothèseinterprétative est que ces deux positions descorps traduisent des statuts sociaux différents.N. Nieszery et N. Peschel proposent ainsi un lienentre position allongée et statut social inférieur.ÀDillingen-Steinheim (Bavière), cette hypothèses’appuie sur la relative pauvreté des défuntsinhumés en position allongée (Nieszery 1995 :82). N. Peschel, quant à elle, associe les patholo-gies osseuses observées sur le squelette d’unsujet en position allongée de la nécropole deWittmar à un travail pénible et par extension, àune position sociale inférieure dans la société(Peschel 1992 : 239-240). Enfin, à Vendenheim,Ch. Jeunesse propose que l’opposition entresujets fléchis et sujets allongés exprime pro-bablement un clivage social au sein de la commu-nauté, sans préciser de quelle nature (Jeunesse2006 : 8).
En tout état de cause, ces positions des corps siopposées impliquent des sépultures différentes,ne serait-ce que par la forme et les dimensionsnécessaires de la fosse sépulcrale. Elles peuventaussi impliquer des aménagements différents àl’intérieur des tombes. Ces positions différentespourraient être ainsi la cause ou la conséquenced’une modification des funérailles. Commentappréhender les aménagements de ces sépultu-res lorsque l’on ne dispose que d’un squelettedans une fosse ?
Appréhender les modalités de dépôtdes corps et les dispositifs architecturauxdes tombes : l’apport de l’analysetaphonomique
L’analyse taphonomique du cadavre et de latombe constitue aujourd’hui un acquis méthodo-logique validé par plus de 20 années de recher-ches. Elle se propose de restituer l’agencementoriginel de la sépulture à partir des ossementstels qu’ils sont mis au jour lors de la fouille, encombinant observations ostéologiques et archéo-logiques fines (Duday et Masset 1987 ; Duday etal. 1990 ;Duday 1995, 2009). Les décalages entrel’ordre anatomique tel qu’il est dans le corps et ladisposition des ossements les uns par rapportaux autres dans la tombe permettent de discuterdes modalités de dépôt du corps, ainsi que dumilieu dans lequel le cadavre s’est décomposé.
GRP : fyssen JOB : annales27 DIV : 06⊕mp⊕THEVENET⊕Corinne p. 3 folio : 38 --- 18/2/013 --- 14H39
ANTHROPOLOGIE / ETHNOLOGIE
38

L’observation de l’état des articulations qui liententre eux les ossements du squelette (connexionoudislocation), du typed’articulations (labiles oupersistantes), de la disposition et des déplace-ments des ossements permettent de déduire lanaturede l’espace sépulcral : colmatéouvide.Cesdeux milieux auront en effet des conséquencesdifférentes sur les ossements et le squelette. Parailleurs, la mise en évidence d’effets de parois oud’effets de contrainte sur le squelette participe dela détermination d’éventuels contenants, rigidesou souples. Ces éléments combinés aux donnéesrelatives à la fosse sépulcrale et à la dispositiondu mobilier déposé dans la sépulture autorisentla reconstitution des modalités de dépôts descorps et de l’aménagement des sépultures.
Cette analyse repose sur la connaissance de lachronologie de la décomposition du corps et par-ticulièrement des articulations. Elle nécessitedonc une bonne conservation des ossements etdu squelette pour réaliser les observations ostéo-logiques. Son intérêt pour l’archéologie est depermettre la restitution de l’agencement des élé-ments aujourd’hui disparus et qui constituent lasépulture : le cadavre, les modalités de son dépôtet de son évolution et les aménagements qui l’ontentourés.
Des cercueils au Néolithique ancien :l’exemple de la nécropole rubanée de« Viesenhäuser Hof » (Stuttgart,Bade-Wurtemberg)
La nécropole de « Viesenhäuser Hof » s’étendau nord de la ville actuelle de Stuttgart, sur unebutte de loess et à proximité d’un habitat enpartie contemporain. Elle a connu une très lon-gue durée d’occupation, depuis le Rubané ancienjusqu’au Rubané récent (Kurz 2000 ; Price et al.2003) et comprend 177 sépultures. Parmi celles-ci, on compte 10 sujets en position allongée (6 %.)Nous présentons ici l’une de ces tombes.
La sépulture 65 contient les restes de deuxindividus, un adulte allongé sur le dos et unnouveau-né déposé sur l’épaule droite de l’adulteet dont la position n’est pas restituable (fig. 2). Laformeetlesdimensionsdelasépulturenesontpasdocumentées en surface, seules des traces plussombressontapparuesauniveaudusquelette.Lafossetellequedéfinieparlesfouilleursestovalaireet étroite, mesurant environ 1,70 m de long sur0,40mde large. La profondeur est de 1,40msousleniveaudecirculationactuel.Lesfragmentsd’un
vase décoré ont été découverts dans le rem-plissage de la tombe, 0,20mau-dessus dudéfunt.
L’adulte est orienté la tête au nord. Son crâneest redressé et recouvre l’épaule gauche. Lesmembres supérieurs sont en flexion maximale,l’avant-bras droit sur le bras droit, tandis quel’avant-bras et le bras gauches sont parallèles ; lamain gauche repose sur l’épaule gauche. Lemem-bre supérieur droit recouvre en partie l’hémi-thorax droit. Les membres inférieurs sont enextension et parallèles, pourtant la partie distaledu pied gauche se trouve sous le pied droit. Bien
Figure 2. La sépulture 65 de « Viesenhäuser Hof »,Stuttgart (photo : Landesamt für Denkmalpflege,Baden-Württemberg).
GRP : fyssen JOB : annales27 DIV : 06⊕mp⊕THEVENET⊕Corinne p. 4 folio : 39 --- 18/2/013 --- 14H40
ANTHROPOLOGIE / ETHNOLOGIE
39

que le squelette du sujet adulte témoigne d’unebonne cohérence générale, il n’en présente pasmoins plusieurs distorsions par rapport à l’ordreanatomique, dont certaines de grande ampleur(fig. 3). La plupart sont concentrées sur le côtégauche du corps : la mise à plat accompagnée del’ouverture de l’hémi-thorax, les côtes venantrecouvrir le coude gauche ; la mise à plat de l’oscoxal gauche, entraînant une ouverture du bas-sin ; la dislocation entre le tibia et la fibula gau-ches et surtout la rupture de la cheville due àla rotation médiale de l’ensemble du pied. Surle côté droit du corps, les déplacements sontplus rares (« verticalisation » de la clavicule dont
l’extrémité latérale sortduvolume initialducada-vre, affaissement général du pied), mais l’un estimportant : l’ensemble du membre supérieurdroit, fléchi, amigrédeprès de15 cmendirectiondubassin.Cedéplacement,ainsique l’axeobliquesuivi par une partie des premières vertèbres tho-raciques, est vraisemblablement à mettre sur lecompte de la pression exercée par le corps dunouveau-né déposé sur son épaule.
Ces nombreux déplacements outrepassent levolume initial du cadavre et impliquent unedécompositiondu corps enespace vide.Plusieursparties anatomiques sont néanmoins demeuréesen équilibre après la disparition des contentionsligamentaires et malgré l’espace vide, tandis qued’autres témoignent de contraintes. Le crâne estainsi fortement redressé et les membres supé-rieurs fléchis sont en pendage. La position des osde l’avant-bras gauche est particulièrement ins-table : leur extrémité distale se trouve 6 cm plushaut que leur extrémité proximale, sans que riende pérenne n’ait assuré leur maintien. Une com-pression s’est exercée sur l’hémi-thorax droitdont la fermeture s’est accrue, ainsi que surl’épaule gauche (la clavicule est « verticalisée », lascapula est sur chant et l’humérus est latéralisé).Quant à la position du pied gauche, elle impliqueune forte contrainte car son déplacement estinverse du mouvement anatomique attendu,c’est-à-dire une rotation latérale. Quelque chose anon seulement empêché le mouvement « natu-rel » du pied, mais l’a contrait à s’effondrer ducôté opposé. Par ailleurs, on remarque quel’ensemble des ossements sur le côté droit des-sineuneffetdeparoi rectiligne,depuis lemembresupérieur jusqu’aupied. Il est ànoter que l’impor-tant déplacement du membre supérieur droit arespecté cet alignement.
Les dislocations accompagnées de sorties duvolume initial du corps indiquent donc unedécomposition de l’adulte en espace vide. L’hypo-thèse la plus simple serait une fosse fermée parun couvercle, maintenant ainsi le volume sépul-cral videdurant la décompositionducorps.Néan-moins, les contraintes et les effets de butée sur lesquelette nécessiteraient une fosse particulière-ment étroite, dispositif difficile pour installerl’adulte et le nouveau-né sur son épaule droite.Une seconde hypothèse est la présence d’uneconstruction à l’intérieur de la fosse, assurantnon seulement l’espace vide, mais rendant
Figure 3. Les ruptures articulaires accompagnéesd’une sortie du volume initial du corps indiquentune décomposition du cadavre en espace vide.
GRP : fyssen JOB : annales27 DIV : 06⊕mp⊕THEVENET⊕Corinne p. 5 folio : 40 --- 18/2/013 --- 14H40
ANTHROPOLOGIE / ETHNOLOGIE
40

compte des nombreuses butées sur le squelette.Les parois d’un contenant rigide étroit peuventexpliquer ces différentes caractéristiques (fig. 4).En outre, l’effet de paroi unilatéral suggère untassement de celui-ci, tel qu’il peut se produirelorsque l’on descend le contenant dans la fosse.Le crâne redressé et les poignets situés 0,15 mau-dessus du fond de la fosse impliquent desmontants relativement hauts et plaideraient enfaveur d’un contenant fermé.
Lesobservations réalisées surd’autres tombesbien documentées corroborent ces faits.
Si la présencede cercueil est démontrable, leurnature exacte reste inaccessible en l’absence devestige ligneux : contenant monoxyle ou consti-tué de planches équarries et assemblées... La pré-sence d’outils en pierre permettant le défriche-ment et la coupe d’arbres (herminettes), toutcomme le savoir-faire architectural de ces popu-lations (leurs maisons rectangulaires en bois ettorchispeuvent atteindre40mde long) indiquentqu’elles possèdent les moyens de fabriquer descontenants rigides élaborés. D’un point de vuematérialiste, la fabricationd’uncercueil implique
Figure 4. La décomposition du corps en espace vide, la présence de contraintes et d’un effet de paroi le longdu côté droit du corps permettent de restituer la présence d’un contenant rigide et étroit.
GRP : fyssen JOB : annales27 DIV : 06⊕mp⊕THEVENET⊕Corinne p. 6 folio : 41 --- 18/2/013 --- 14H40
ANTHROPOLOGIE / ETHNOLOGIE
41

un investissement supplémentaire par rapportau fait d’enterrer le défunt sans contenant. Maisl’interprétation en termes hiérarchiques desdéfunts allongés ne permet pas de comprendrecette nouveauté. Si l’invention du cercueil signeune innovation, elle s’inscrit néanmoins dans lacontinuité, celle de préserver le corps du contactde la terre, mais les moyens diffèrent : la fosseseuleassurecette fonction lorsqu’elle contientundéfunt fléchi, mais on a recours à un cercueillorsque le défunt est allongé. Comment s’insèrecette modification au sein des funérailles ? Laposition allongée apparaît la plus appropriée autransport d’un corps. Si l’apparition du cercueilpeutêtremiseenrelationavecdesconsidérationspratiques, elle témoigne dans tous les cas d’unemodification des pratiques pré-sépulcrales. Cesdernières prennent-elles une part accrue durantle déroulement des funérailles ? Ce nouvel usagepourrait en effet traduire une importance plusgrande accordée au transport du défunt et auxprocessions accompagnant ce moment.
Plutôt qu’un simple choix de la position dudéfunt, l’introductionde lapositionallongéedanscertains ensembles funéraires de la CéramiqueLinéaire peut être mise en relation avec l’appari-tion de contenants rigides et mobiles, c’est-à-direde cercueils. Un tel changement implique inévita-blement une modification des funérailles et deleur déroulement. Elle traduit vraisemblable-ment une mutation des représentations socialesde cette culture.
Bibliographiee Boës E., Jeunesse C., Arbogast R.-M.,
LefrancP.,MauvillyM., Schneikert F.&Sidera I.(2007) – Vendenheim « le Haut du Coteau » (Bas-Rhin) : remarques sur l’organisation interned’une nécropole du Néolithique ancien danubienin Besse M. (dir.), Sociétés néolithiques. Des faitsarchéologiquesaux fonctionnements socio-économi-ques, Actes du 27e colloque interrégional sur leNéo-lithique, Neuchâtel 2005. Lausanne, Cahiersd’archéologie romande 108, p. 279-284.
e Duday H. (1995) – Anthropologie de « ter-rain », archéologie de la mort in La Mort, passé,présent, conditionnel, La Roche-sur-Yon, GroupeVendéen d’Études Préhistoriques, p. 33-58.
e Duday H. (2009) – The archaeology of thedead : lectures in archaeothanatology (translatedby A. M. Cipriani and J. Pearce). Oxford, OxbowBooks.
e Duday H. & Masset C., dir. (1987) – Anthro-pologie physique et archéologie. Méthodes d’étudedes sépultures. Actes du colloque de Toulouse,4-6 novembre 1982. Paris, CNRS.
e Duday H., Courtaud P., Crubezy E., SellierP. & Tillier A.-M. (1990) – L’anthropologie de« terrain » : reconnaissance et interprétation desgestes funéraires. Bulletins et Mémoires de laSociété d’Anthropologie de Paris, n. s., p. 29-50.
e Jeunesse C. (2006) – Les traditions funérai-res du néolithique moyen en Europe centraledans le cadre du système funéraire danubien inAlt K., Arbogast R.-M., Jeunesse C. & van Willi-gen S. (éds.), Archéologie funéraire du Néolithi-que danubien, Nouveaux enjeux, nouvelles appro-ches, table ronde de Fribourg-en-Bresgau 2004,Cahiers de l’Association pour la promotion dela Recherche Archéologique en Alsace 20, 2004,p. 3-26.
e Kurz G. (2000) – Gräberfelder und Siedlun-gen beim « Viesenhäuser Hof » (Stuttgart-Mühlhau-sen). (Unpubliziert Manuskript, Stuttgart 2000).
e Lichardus J. et Lichardus-Itten M., avecla coll. de Bailloud G. et Cauvin J. (1985) – LaProtohistoire de l’Europe. Le Néolithique et leChalcolithique. Paris, Presses Universitaires deFrance, Collection Nouvelle Clio n0 1bis.
e Moddermann P. J. R. (1970) – Linearband-keramik aus Elsloo und Stein. Analecta Praehis-torica Leidensia III, 3 volumes.
e Nieszery N. (1995) – LinearbandkeramischeGräberfelder in Bayern. Internationale Archäolo-gie, 16, Espelkamp.
e Peschel Ch. (1992) – Regel und AusnahmeLinearbandkeramische Bestattungssitten in Deuts-chland und angrenzendenGebieten, unter besonde-rer Berücksichtigung der Sonderbestattungen.Internationale Archäologie 9, Verlag MarieL. Leidorf, Buch am Erlbach.
e Price T. D., Wahl J., Knipper C., Burger-Heinrich E., Kurz G. und Bentley R. A. (2003) –Das bandkeramische Gräberfeld vom « Viesen-häuser Hof » bei Stuttgart-Mühlhausen : NeueUntersuchungsergebnisse zum Migrationsver-halten im frühen Neolithikum. Fundberichte ausBaden-Württemberg 27, p. 23-58.
e StrienH.-C., GronenbornD. (2005) –Klima-und Kulturwandel während des mitteleuropäis-chen Altneolithikums (58./57.-51./50. Jahrhun-dert v. Chr.) inGronenborn D. (ed.),Klimaverän-
GRP : fyssen JOB : annales27 DIV : 06⊕mp⊕THEVENET⊕Corinne p. 7 folio : 42 --- 18/2/013 --- 14H40
ANTHROPOLOGIE / ETHNOLOGIE
42

derung und Kulturwandel in neolithischen Gesell-schaftenMitteleuropas, 6700-2200v.Chr. (ClimateVariability and Culture Change in Neolithic Socie-ties of Central Europe, 6700-2200 cal BC), Mainz,p. 131-149.
e Thevenet C. (2004) –Une relecture des prati-ques funéraires du Rubané récent et final du Bas-sinparisien : l’exempledes fosses sépulcralesdansla vallée de l’Aisne. Bulletin de la Société Préhisto-rique Française, tome 101, no 4, p. 815-826.
e Thevenet C. (2010) –Des faits aux gestes, desgestes aux sens ? Pratiques funéraires et sociétédurant le Néolithique ancien du Bassin parisien.Thèse de doctorat, Université Paris1-PanthéonSorbonne, 2 volumes.
Facing the death of its members, each societyprovides specific responses through funeral prac-tices. These latter are the basis of the vision thesociety has on itself, not on its identity but ratheron its beliefs. When approaching prehistoricsocieties, we only dispose of necessarily incom-plete vestigial remains, mostly skeletal elementswithin pit features possibly containing otherobjects. These remains nonetheless are a basisallowing to reconstruct the different stages of thetreatment of the deceased, whether they arelinked to grave construction or other postmortaltreatments of the body or the skeleton. Althoughthe identificationof these gestures is only partial,given the very nature of the archaeological data,a large part, however, can be approached, fromthe choice of the grave location up to the actionsthat occurred after its closing. Thanks to thereconstruction of these actions, men’s beliefs ondeath can be approached. This concerns the pre-paration and the types of deposits of the body inthe grave: deposing the flexed body on one side oron its back reflects different mortuary rites. Thechange from one arrangement of the deposit tothe other within a same culture a priori indicatesa rupture within funerary practices. What doesthe emergence of a new position of the body tellus about the funerary ritual in these societies?
A rare position within the mortuarypractices of the Early Neolithic LinearPottery Culture
The emergence of the first sedentary societiesinCentral andWesternEurope is associatedwiththe Linear Pottery Culture or Danubian culture.This cultural complex develops in the Great
HungarianPlainduring the secondhalf of the6thmillennium and reaches the coast of the Channelduring the 5thmillennium (Lichardus et al. 1985;Strien and Gronenborn 2005). Despite conside-rable geographic expansion (fig. 1), these firstfarmers and stock breeders preserved strongidentity through their material culture, notablyhouse architecture and means of subsistencewhilst adapting to the new environments theycolonized.
In cemeteries as well as in burials within sett-lement context, primary inhumation is the domi-nant type of deposit. A most simple type ofinhumation was a long held interpretation: thedeceased were laid down in an oval-shaped pitdug out in the size of the crouched body. Thecorpse was most often deposited on one side andthe pit immediately filled thereafter. However,the case of theParisBasindemonstrates that thissimplicity is only apparent: the graves revealcomplex structures maintaining a void aroundthe body (Thevenet 2004; 2010). The recognitionof inhumations in extended position has notchanged the concept of a simple type of inhuma-tion as their presence within cemeteries of theLinear Pottery culture seems rare: N. Nieszeryhas registered less than 5 % of the deceasedburied in this position in Central Europe (Nies-zery 1995: 82). However, these cases are not rareat all in Alsace. In this latter region, they mayconstitute nearly half of the inhumations in dis-tinct cemeteries (for example, 41 out of 111 inhu-mations in Vendenheim, Boës et al. 2007). Inhu-mations in extended position indeed concentratein cemeteries of the Upper Rhine plain (Alsace,Baden-Württemberg) and in distinct burial sitesin the eastern part of Bavaria (fig. 1). Except fordistinct funerary assemblages in Alsace, theyrepresent aminority inmost of these cemeteries.
How can such a change be explained?Commonly quoted as an exception from the
rule, the extended position of the deceased isgenerally considered as being a late phenomenonwithin the Linear Pottery Culture. Althoughgrave goods are not always present in order toprecise this attribution, it is implicitly based onthe fact that this position becomes the standardwithin the cultures succeeding to the LinearPotteryCulture in theRhine region (Hinkelstein,Großgartach, Rössen).
GRP : fyssen JOB : annales27 DIV : 06⊕mp⊕THEVENET⊕Corinne p. 8 folio : 43 --- 18/2/013 --- 14H40
ANTHROPOLOGY / ETHNOLOGY
43

Asnoted above, in distinct regions inhumationin extended position is not that anecdotic.Concerning the date of its emergence, there islittle consensus: from the latest phase for certain(Moddermann 1970, Peschel 1992), from the latephase for others (Jeunesse 2006). It is equallypossible that this innovation may have appearedsimultaneously in all the regions occupied bythe Linear Pottery Culture or that it resultedfrom internal spread within this cultural com-plex. Regardless, the extended position of thedeceased marks a rupture compared to the stan-dard. The main interpretation assumes thatthese two positions of the corpse reflect differentsocial status. N. Nieszery and N. Peschel pro-
poseda linkbetweenextendedpositionand lowersocial status. In Dillingen-Steinheim (Bavaria),this hypothesis is corroborated by relativepoverty of grave goods associated with thedeceased buried in extended position (Nieszery1995: 82). N. Peschel puts in relationship bonedisease observed on the skeleton of an individualin extended position in the cemetery of Wittmarwith hard labour and consequently with a lowersocial position within the group (Peschel 1992:239-240). Finally, for Vendenheim, Ch. Jeunesseassumes that the juxtapositionbetweencrouchedand extended positions probably mirrors socialdivision within the group without clarifying itsnature (Jeunesse 2006: 8).
Figure 1. Maximum expansion of the Linear Pottery Culture in central Europe (5500-4900 BC).
GRP : fyssen JOB : annales27 DIV : 06⊕mp⊕THEVENET⊕Corinne p. 9 folio : 44 --- 18/2/013 --- 14H41
ANTHROPOLOGY / ETHNOLOGY
44

In any case, these clearly opposed positions ofthe body necessarily implicate different gravestructureswith regard to the shapeand thesizeofthe sepulchral pit. Equally, they may implicatedifferent arrangements within the graves. Thesedistinct positions may also be the reason or theconsequence from changes in the funerary rite.How can we approach arrangements in thesegraves only observing a skeleton deposited in thepit?
Approaching the deposit modalitiesof the corpses and the architecturalfeatures of the graves: the contributionof taphonomic analysis
Yet taphonomic analysis of the body and thegrave is a methodology validated by more than20yearsof research. It aimsat reconstructing theinitial arrangement of the grave based on thesituation of the bones registered during excava-tion by combining detailed osteological andarchaeological observations (Duday and Masset1987; Duday et al. 1990; Duday 1995, 2009). Thediscrepancies between the «natural» anatomicalorder of the corpse and the arrangements of thebones in the grave allow to analyse the types ofdeposit of the corpse and the environment inwhich the body has decomposed. The analysis ofthe state of the joints connecting the bones of theskeleton (connection or disjunction), of the typesof joints (unstable or persistent), or of the arran-gements and displacements of bones allow todeduce the nature of the sepulchral space: a filledspace or a void. As a matter of fact, the effects ofthese two environments on the bones and theskeleton will be different. Moreover, «walleffects» or «binding effects» recognized on theskeleton indicate the presence of possible contai-ners, made of hard or soft material. These ele-ments, merged with data referring to the gravepit and the arrangement of the grave goods depo-sited in the burial, allow to reconstruct the depo-sit modalities related to the body and the equip-ment in the graves.
This analysis is based on the knowledge of thedecomposition rate of the body and more parti-cularly of the joints. A good preservation of thebones and the skeleton is required in order toallow these osteological observations. Theyare ofinterest for archaeologists in order to recons-truct the arrangement of vanished elements that
constituted the burial: the body, themodalities ofits deposit and its evolution as well as the sur-rounding arrangements.
Coffins in the Early Neolithic: the exampleof the cemetery of the Linear PotteryCulture at ”Viesenhäuser Hof“(Stuttgart, Baden-Württemberg)
The cemetery of ”Viesenhäuser Hof“ expandsnorth of the actual city of Stuttgart, located on aloessic mound, close to a partially contempora-neous settlement. Its occupation spans over a
Figure 2. Burial 65 of «Viesenhäuser Hof», Stutt-gart (photograph: Landesamt für Denkmalpflege,Baden-Württemberg).
GRP : fyssen JOB : annales27 DIV : 06⊕mp⊕THEVENET⊕Corinne p. 10 folio : 45 --- 18/2/013 --- 14H41
ANTHROPOLOGY / ETHNOLOGY
45

very long period of time, from the early to the latephase of the Linear Pottery Culture (Kurz 2000;Price et al. 2003) and it contained 177 burials.Amongst them, 10 individuals are laid down inextended position (6 %.) One of these graves willbe presented here.
Burial 65 contained the remains of two indivi-duals: an adult in extended position on the backand a newborn child deposited on the right shoul-der of the adult and whose position can not bereconstructed (fig. 2). The shape and the dimen-sions of this grave have not been documented.The upper limit, materialized by darker traces,appeared only at the height of the skeleton. Thepit as described by the archaeologists was oval-shaped and narrow, measuring 1,70 m in lengthby 0,40 m in width. It reached a depth of 1,40 mbelow the actual surface. Fragments of a decora-ted vessel have been recovered from the filling ofthe burial, at 0,20 m above the skeleton.
The adult is oriented with the head to thenorth. The skull is uplifted and overlaps with theleft shoulder. The upper limbs are in maximumflexed position, the right forearm lies on the rightarm and the left forearm parallel to the left arm;the left hand lies on the left shoulder. The rightupper limb partially overlapswith the right hemi-thorax. The lower limbs are stretched out andparallel, the distal end of the left foot, however,appears under the right foot. Although the adultskeleton shows general consistency, several dis-tortions from the anatomical order can be obser-ved, some of them being considerable (fig. 3).These distortionsmainly affect the left side of thebody: the flattening accompanied by the openingof the hemi-thorax, the ribs overlapping withthe left elbow; the flattening of the left hipbone leading to the opening of the basin; thedislocation of the left tibia and fibula and moreparticularly thebreakdownof theankledue to themedial rotation of the entire foot. On the rightside of the corpse, displacements are rarer («ver-ticalization» of the clavicle, of which the acromialextremityhasmovedoutside the initial volumeofthe body, collapse of the foot). One, however, issubstantial: the right upper limb has movedabout 15 cm towards the basin. This displace-ment, as well as the oblique axis followed by onepart of the upper thoracic vertebrae, is presuma-bly due to the weight of the newborn child’s bodyon the shoulder.
Thesedisplacementsbeyond the initial volumeof thebody implicate decompositionof the corpsein a void. Nonetheless, a series of anatomicalelements have been maintained in equilibriumafter the breakdown of the ligaments and despitethe void, whilst others attest to constraints. Theskull is stronglyuplifted and thebentupper limbsare inclined. The bones of the left forearm are ina particularly unstable situation: their distalextremity lies 6 cm higher than their proximalextremity, without any object supporting thisposition. Compression of the right hemi-thorax
Figure 3. The ruptures of the joints accompanied bydisplacementsbeyond the initial volumeof thebodyindicate decomposition of the corpse in an emptyspace.
GRP : fyssen JOB : annales27 DIV : 06⊕mp⊕THEVENET⊕Corinne p. 11 folio : 46 --- 18/2/013 --- 14H41
ANTHROPOLOGY / ETHNOLOGY
46

increased its closing. In addition, pressure actedon the left shoulder (the clavicle is «verticalized»;the scapula is turned on edge and the humerusbone on its side). The position of the left footimplicates strong constraint as it moved in theopposite direction compared to the expected ana-tomical movement, i. e. a lateral rotation. Some-thing has not only hindered the «natural» move-ment but has forced the foot to collapse on theopposite side. In addition, it can be stated that alltheboneson the right side showaneffect of lineardelimitation, from the upper limb to the foot. It
has to be noted that the strong displacement ofthe right upper limb followed this alignment.
The dislocaions accompanied by the move-ments of bones beyond the initial volume of thecorpse consequently indicate decomposition ofthe adult in an empty space. The simplest hypo-thesis would be a pit closed by a cover plate,maintaining the sepulchral space void during theprocess of decomposition of the corpse. Nonethe-less, the constraints and thewall effects observedon the skeleton would require a particularly nar-row pit and it would be difficult to lay down an
Figure 4. The decomposition of the corpse in an empty space, the presence of constraints and of a wall effectalong the right side of the bodyallow reconstructing thepresence of anarrowcontainermadeof hardmaterial.
GRP : fyssen JOB : annales27 DIV : 06⊕mp⊕THEVENET⊕Corinne p. 12 folio : 47 --- 18/2/013 --- 14H41
ANTHROPOLOGY / ETHNOLOGY
47

adult and anewborn child onhis shoulder in sucha feature. A second hypothesis is the presence ofan internal construction within the pit, ensuringnot only a void but also explaining thewall effectsobserved on the skeleton. The presence of anarrow container made of hard material mayexplain these different characteristics (fig. 4).Moreover, the unilateral wall effect suggests thatthe body slipped at the moment when the contai-ner was laid down in the pit. The uplifted headand thewrists located 0,15mabove the bottomofthe pit involve the presence of rather high sidesand would be an argument in favour of a closedcontainer.
Observationsmade for other well-documentedgraves corroborate these facts.
Although the presence of a container can bedemonstrated, its true nature remains unclear asorganic features are absent: wooden coffinsfashioned fromhollowed-out logs or crude planks... The presence of stone tools for forest clearingand tree cutting (adzes), as well as the architec-tural skill of these populations (their rectangularhouses constructed with timber and wattle-and-daub can reach 40 m in length) show that theyhad the means to manufacture sophisticatedrigid containers. From the materialistic point ofview, the manufacturing of a coffin involvesadditional investment compared to the burial ofa dead without using a container. However, the
interpretation that the deceased in extendedposition would mirror social hierarchy does notallow to understand this innovation. Althoughthe invention of the coffin is an innovation, it isnonetheless part of a continuum in that it aimedat protecting the corpse from contact with dirt.However, the means of implementation differ:the pit alone ensures this function containing thedead in a crouched position. But when the deadis laid down in an extended position, a coffin isused. How is this change integrated into thefunerary rites? The extended position appears tobe the most appropriate for the transport of thebody. If the emergence of a coffin can be put inrelationship with practical considerations, it tes-tifies in any case to a change of the practicespreceding the burial. Are these latter increasin-gly important within the burial ceremony? Thenew use may indeed mirror greater importanceaccorded to the transport of the dead and toprocessions accompanying this moment.
Rather than a simple choice of the dead’sposition, the introduction of the extended posi-tion in distinct funerary assemblages of theLinear Pottery Culture can be related to theappearance of rigid and mobile wooden contai-ners, i. e. of coffins. Such a change inevitablyimplies a modification of the burial ritual. Itseemingly also highlights a change of the socialrepresentations of this culture.
GRP : fyssen JOB : annales27 DIV : 06⊕mp⊕THEVENET⊕Corinne p. 13 folio : 48 --- 18/2/013 --- 14H41
ANTHROPOLOGY / ETHNOLOGY
48

L’orientation olfactive chez les oiseaux
Gaia DELL’ARICCIA
Chercheur Post-doctoral au Centre d’Ecologie Fonctionnelle et Evolutive à l’UMR 5175 CNRS
RésuméLes pigeons voyageurs sont capables de former des cartes cognitives, sous forme de représentation
neuronale de la distribution hétérogène de molécules odorantes dispersées dans l’atmosphère.Cependant, leur capacité à utiliser des signaux différents (i.e. visuels, magnétiques) en fonction descirconstances, a suscité un débat scientifique quant à l’utilisation de l’olfaction pour l’orientation.L’étude des oiseaux marins de l’ordre de Procellariiformes a apporté de nouvelles et importantesévidences en faveur de la théorie de la navigation olfactive. En effet, de nombreuses espèces de pétrelsutilisent des signaux olfactifs pour localiser les zones d’affouragement au-dessus de la surface del’océan apparemment sans relief. En particulier, ces oiseaux sont capables de détecter et de s’orientervers le sulfure de diméthyle (DMS), une composante volatile présente naturellement dans les océansdumonde entier, au niveau des zones productives d’affouragement. Afin d’étendre la compréhensiondu rôle et de l’utilisation du DMS, j’ai étudié la réponse du Puffin cendré de Méditerranée, à cecomposant. En effet, contrairement aux océans, la Mer Méditerranée est un bassin fermé, où lesdistances entre les côtes peuvent être facilement parcourues par un pétrel en vol dans une journée etoù les émissions de DMS sont faibles. Dans ces conditions, les pétrels peuvent utiliser de repèresvisuels pour construire une carte de navigation à large-échelle, et la sensibilité aux concentrations desurface de DMS peut ne pas être nécessaire pour l’orientation spatiale. Les résultats obtenusmontrent que la réponse auDMS et son utilisation dans l’orientation sont répandues chez les oiseauxmarins, indépendammentde leurshabitudes alimentaires, et dansdifférentmilieumarinsoùd’autresrepères sont également disponibles. La réponse générale à ce composé suggère que le DMS peut êtreimportant, non seulement dans la recherche de nourriture, mais aussi pour d’autres tâches, commepar exemple la navigation globale.
Mots-clésAlimentation, Calonectris diomedea, carte spatiale, DMS, écologie sensorielle, olfaction, orienta-
tion, paysage olfactif, pétrels, Puffin cendré
Olfactory orientation in birdsAbstract
Homing pigeons are able to represent the spatial distribution of atmospheric odours to build anavigationalmap.However, their ability to shift among different cues (i.e. visual,magnetic)made theuse of olfaction for orientation a subject of scientific debate. On the contrary, the investigation ofpetrel seabirds provided new and important evidence supporting olfactory navigation. Many petrelspecies use olfactory cues to locate foraging grounds over the seemingly featureless ocean surface. Inparticular, they are able to detect and orient to dimethylsulfide (DMS), a volatile compound naturallyoccurring in worldwide oceans in correspondence with productive feeding areas. To understand therole and use of DMS in a different ecological setting, I investigated the response to this compound byMediterranean Scopoli’s shearwaters. In contrast to oceans, theMediterranean Sea is a closed basinin which the distances between coasts can be covered easily by a flying petrel within a day, andpresenting low DMS emissions. In this condition, petrels may rely on visual landmarks to build alarge-scale map and the sensing of DMS surface concentrations may not be necessary for spatialorientation. Results show that the response and use of DMS is widespread among different seabirds,independent of feeding habits, and in different marine environments, where other cues are alsoavailable. Such finding suggests that the importance of this compound may be not limited to thelocalization of foraging areas but also be employed for other tasks i.e. global orientation.
GRP : fyssen JOB : annales27 DIV : 07⊕mp⊕DELL⊕ARICCIA⊕Gaia p. 1 folio : 49 --- 18/2/013 --- 14H42
ETHOLOGIE / PSYCHOLOGIE
49

KeywordsCalonectris diomedea, DMS, foraging, olfaction, olfactory landscape, orientation, petrels, Scopoli’s
shearwater, sensory ecology, spatial map
L’orientation olfactive chez les oiseauxParmi les animaux navigateurs, les oiseaux
occupent une place spéciale du fait de leurs capa-cités spatiales exceptionnelles. Ces aptitudes semanifestent notamment dans le contexte de leurretour au nid (Wallraff 2005) et de leurs voyagesde migration, longs de plusieurs milliers de kilo-mètres, qui les ramènent, annéeaprès année, auxmêmes sites de reproduction et d’hivernage (Ber-thold 2001). De nombreuses études qui visent àcomprendre les mécanismes d’orientation chezles oiseaux ont été réalisées en employant lePigeon voyageur (Columba livia) comme espècemodèle. Les pigeons voyageurs sont capables deformer des cartes cognitives, sous forme dereprésentation neuronale de la distribution spa-tiale des stimuli environnementaux. Cette cartede navigation leur permet d’estimer la directionet, peut-être, la distance dupigeonnier depuis dessites éloignés et inconnus. La construction decette carte de navigation reposerait sur leurcapacité à représenter la distribution hétérogènede molécules odorantes dispersées dans l’atmos-phère (Wallraff2005).Cette capacitén’exclut pasque lespigeonspuissentutiliserdes informationssensorielles supplémentaires, fournies par exem-ple par le champ magnétique terrestre (Wilts-chko et Wiltschko 2003). Malheureusement,après plus de 30 ans de recherches (Wallraff2005), l’utilisation de signaux olfactifs reste l’undes sujets le plus débattus dans les études quiportent sur l’orientation chez les pigeons (Wilts-chko et Wiltschko 2003, Wallraff 2004, Wallraff2005, Jorge et al. 2010).
Laméconnaissancede l’implication relativededifférents signaux et repères pour l’orientationest aussi liée au fait que les pigeons volent géné-ralement en survolant la terre, où les repèresvisuels du paysage ont une grande influence surla navigation,même quand ceux-ci sont inconnus(Holland 2003, Lau et al. 2006, Dell’Ariccia et al.2009). De plus, il a été montré que la présence derepères visuels modifie les résultats obtenussuite à des traitements expérimentaux tels quel’anosmie ou le décalage de l’horloge biologique(Bonadonna et al. 2000, Wallraff 2005). Finale-
ment, il apparait de plus en plus évident que lespigeonsvoyageursutilisentunestratégied’orien-tation multimodale et qu’ils sont capables dechanger leur stratégie de navigation selon lescirconstances (Walcott 2005, Wallraff 2005,Dell’Ariccia et al. 2008, Dell’Ariccia et al. 2009).Il apparait donc presque impossible d’isolerl’influence de différents signaux utilisés par cetteespèce et comprendre le rôle exact de chacund’entre eux dans le processus d’orientation. Parconséquent, il parait compromis de pouvoirdémontrer définitivement un mécanismed’orientation olfactive en utilisant cemodèle bio-logique. En revanche, les oiseaux marins, et enparticulier les pétrels, peuvent ouvrir de nouvel-les perspectives. En effet, le milieu marin, à ladifférence du milieu terrestre, est apparemmentsans repères visuels pendant de milliers de kilo-mètres carrés, excluant ainsi cette composantesensorielle du paysage. Par ailleurs, les pétrelssemblent ne pas employer de repères magnéti-ques pour s’orienter (Benhamou et al. 2003,Bonadonna et al. 2003b, Bonadonna et al. 2005).
Les pétrels et l’orientation en milieupélagique
Les pétrels (ordre Procellariiformes) présen-tent unmode de vie pélagique et itinérant à largeéchelle. Ces oiseaux longévifs passent la plupartde leur vie envol sur lameret sont liés àdepetitesîles océaniques uniquement pour la reproduc-tion.Lesairesd’alimentationsontexclusivementpélagiques et doivent être identifiés, ainsi que leszones de reproduction, au milieu d’un vasteenvironnement océanique homogène et mono-tone (Warham 1996). Pendant la période repro-ductive, alors que les zones d’alimentation res-tent pélagiques, les pétrels doivent retournerà la colonie pour relayer le partenaire durantl’incubation de leur unique œuf ou nourrir leurpoussin. Ce mode de vie demande d’une grandeefficacité dans la localisation des zones d’affou-ragement afin de pouvoir assurer une alimenta-tion efficace et le succès reproductif.
Les Procellariiformes ont un système olfactifincroyablement développé d’un point de vue
GRP : fyssen JOB : annales27 DIV : 07⊕mp⊕DELL⊕ARICCIA⊕Gaia p. 2 folio : 50 --- 18/2/013 --- 14H42
ETHOLOGIE / PSYCHOLOGIE
50

neuro-anatomique, par rapport aux autresoiseaux (Fig. 1. Bang 1966). En outre, de plus enplus d’évidences suggèrent que leur sensde l’odo-rat bien développé jouerait un rôle fondamentalpour de nombreux comportements tels que l’ali-mentation (Nevitt et al. 1995), la recherche etl’identification du nid (Bonadonna 2009) et lareconnaissance du partenaire (Bonadonna etNevitt 2004, Bonadonna et al. 2007, Mardon etBonadonna 2009).
L’utilisation de l’odorat chez les oiseauxmarins, en particulier chez les pétrels, a été propo-sée il y a longtemps (Grubb 1972, Hutchison etWenzel 1980, Lequette et al. 1989), mais l’étudedesmécanismespermettantàcesoiseauxd’exploi-ter les composantesnaturellementprésentesdansl’atmosphère pour l’orientation et la navigationest relativement nouvelle. Le principe général surlequel se base la théorie de l’orientation olfactiveest que les pétrels pourraient utiliser des molécu-les odorantes pour construire une carte à largeéchelle comme un paysage olfactif superposé à lasurface de l’océan (Fig. 2). Des changements dansle paysage olfactif pourraient signaler à un pétrelitinérant, sa localisation et/ou la proximité dezones productives où rechercher de la nourriture(Nevitt 2000, Bonadonna et al. 2003a).
Le sulfure de diméthyle (DMS) a été proposécomme candidat parfait pour servir de point de
repère olfactif pour les oiseaux marins qui cher-chent à localiser et exploiter les zones producti-ves (Nevitt et al. 1995, Nevitt 1999, Nevitt etBonadonna 2005b). Le DMS est une composantevolatile présente naturellement dans les océansdu monde entier. Il est dérivé de la décomposi-tionmétabolique du diméthylsulfoniopropionate(DMSP) pendant la dégradation du phytoplanc-ton. La production de DMS est souvent associéeà la présence du zooplancton qui s’alimente duphytoplancton et aux zones à haute productivitéprimaire (Cantin et al. 1996, Simó 2001, Jean etal. 2009). Le DMSP peut être un signal olfactifparticulièrement fort pour les prédateurs du zoo-plancton, et des observations in situ montrentque les bancs de petits poissons s’accumulent à lapériphérie des zones de développement du planc-ton (DeBose et Nevitt 2007, DeBose et al. 2008).Finalement, des concentrations élevés de DMSau niveau local peuvent alerter, de façon oppor-tuniste, les prédateurs d’ordre supérieur, de laprésence d’agrégations de zooplancton et de pré-dateurs du zooplancton, c’est à dire poissons etcalmars qui représentent les principales proiespour de nombreuses espèces de pétrels (del Hoyoetal. 1992,Warham1996,HayetKubanek2002).En effet, la télémétrie par satellite a révélé uneassociation entre les zones d’émissions élevéesde DMS, les agrégations de proies et les airesd’alimentation de différentes espèces de pétrels(Revisé par Nevitt 2000).
Cependant, parmi les différentes espèces depétrels, la réactivité au DMS est très variable,suggérant que la sensibilité olfactive à ce com-posé pourrait être liée à différentes stratégiesécologiques pour la localisation des proies. Pourcertaines espèces (par exemple les pétrels et lesprions), la sensibilité olfactive auDMS fournit unsystème de détection précoce pour se nourrir demanière opportuniste avant que d’autres oiseauxarrivent. Ces détecteurs précoces changent letype des signaux associées aux proies : en effet,en s’alimentant sur le site, ces oiseaux pion-niers fournissent des repères visuels à d’autresoiseaux et induisent le dégagement d’odeurs sup-plémentaires, tels que celles liées à lamacérationdukrill. Ainsi, les espèces àdétection tardive (parexemple les albatros et les pétrels géants) pour-raient employer une stratégie de recherche denourriture multimodale, qui s’appuie sur desindices visuels ainsi que sur une combinaison designaux olfactifs, pour localiser leurs proies (Sil-
Figure 1. Coupe schématique à travers les cham-bres olfactives du Pétrel des neiges; en évidence enrouge l’épithélium olfactif (Modifié de Bang 1966).
GRP : fyssen JOB : annales27 DIV : 07⊕mp⊕DELL⊕ARICCIA⊕Gaia p. 3 folio : 51 --- 18/2/013 --- 14H44
ETHOLOGIE / PSYCHOLOGIE
51

verman et al. 2004, Nevitt et Bonadonna 2005a,Nevitt 2008).
En plus de sa fonction dans le cadre de larecherche de nourriture, le DMS a également étéproposé commerepère pour lanavigation globalesur les océans. Sur la surface des océans, leszones avec les concentrations plus élevées deDMS correspondent aux zones très productives,qui à leur tour sont associées à des caractéristi-ques bathymétriques tels que les zones deremonté d’eau, les montagnes et les plateauxsous-marins (McTaggart et Burton 1992). Le pro-fil de la concentration en surface de DMS pour-rait donc représenter un paysage olfactif naviga-ble pour les oiseaux marins, superposé sur lasurface de l’eau apparemment sans relief (Fig. 2.Nevitt et al. 1995, Nevitt 2000, Bonadonna et al.2003a).
Dans l’ensemble, ces études suggèrent un rôleimportant de signaux biogéniques chez les
pétrels océaniques dans le contexte de la recher-che de nourriture et de la navigation globale.Cependant, jusqu’ici, ces études n’ont exploréque peu d’environnements océaniques et ne sesont intéressées qu’à peu d’espèces. Ainsi, onignore si l’utilisation de signaux olfactifs pourtrouver la nourriture est un phénomène localet/ou spécifique à l’espèce ou une adaptationgénérale des oiseaux de mer ; en particuliers,rienn’est connu en ce qui concerne les pétrels quise reproduisent et se nourrissent dans des mersfermées, où les conditions océanographiquespeuvent être extrêmement différentes et où desrepères visuels pour l’orientation (par exempleles lignes de côte et les îles) sont à proximité.
Le cas de la Mer MéditerranéeAfin de mieux comprendre le rôle et l’utilisa-
tion du DMS dans un contexte écologique diffé-rent, nous avons étudié la réponse à ce compo-sant par le Puffin cendré de Méditerranée
Figure 2. Le paysage olfactif. Les structures bathymétriques tels que les zones de remontéd’eau, lesmontagnes et les plateaux sous-marins correspondent aux zones productives, où les concentrationsde DMS sont les plus élevées, pouvant ainsi servir de point de repère olfactif pour la navigation en mer (DeThivent et Bonadonna, 2003 ©Caroline Picard).
GRP : fyssen JOB : annales27 DIV : 07⊕mp⊕DELL⊕ARICCIA⊕Gaia p. 4 folio : 52 --- 18/2/013 --- 14H49
ETHOLOGIE / PSYCHOLOGIE
52

(Calonectris diomedea), qui niche sur l’île deLinosa (Îles Pélages, Canal de Sicile, Italie :35o52’ N, 12o52’ E, Fig. 3). Les voyages d’alimen-tations de ce pétrel durent en moyenne 9,4 jourspendant la période d’incubation des œufs et5,5 jours pendant l’élevage de poussins. Pen-dant leurs voyages, ces oiseaux parcourent, enmoyenne, 904 kmpar voyage d’alimentation pen-dant l’incubation et 662 km par voyage pendantl’élevage du poussin, avec une distance moyennepar jour de 135 km (Celada et al. 2009). Parconséquent, la distance entre les côtes méditer-ranéennes peut être facilement traversée dans lajournée par un pétrel en vol et plusieurs fois aucours du même voyage d’alimentation ; les côtespeuvent donc fournir les points de repère néces-sairespour l’orientationdanscebassin fermé.Eneffet, les zones d’affouragement des puffins quinichent à Linosa se situent surtout autour de l’îleou le longde la côte tunisienne (Celadaetal. 2009,Dell’Ariccia et al. 2010). La zone côtière tuni-sienne présente des repères visuels évidents,fournis par la côte elle-même et par les nombreu-ses îles, qui pourraient aider à l’orientation enpermettant aux oiseaux de pointer vers des spotsdenourriture une fois qu’ils sont dans la zone.Enoutre, laMéditerranée présente des émissions deDMS modérées, liées à la faible productivité deses eaux, et qui sont inférieures à celles desOcéans Atlantique et Pacifique (Simó et al. 1997,Simó et Grimalt 1998, Belviso et al. 2003). Leszones où la concentration de DMS est la plusélevée se limitent aux zones côtières, où la pro-ductivité des eaux est majeure (Estrada 1996,
Simó et al. 1997). Dans ces conditions, l’utilisa-tion du DMS comme repère olfactif pourrait êtremoins utile qu’enmilieu océanique et donc l’apti-tude à détecter et à s’orienter vers le DMS pour-rait être réduite ou absente car superflue pourl’orientation, et donc les Puffins cendrés deMédi-terranée pourraient être moins attirés par cecomposé que d’autres pétrels.
Notre expérience s’est déroulée en deux éta-pes : dans un premier temps, nous avons testé lacapacité des Puffins cendrés à détecter et à choi-sir le DMS dans un labyrinthe en Y placé à lacolonie (Fig. 4) ; dans ce test nous avons présen-tés aux oiseaux un choix binaire entre le DMS etune odeur contrôle, en utilisant, en milieu natu-rel et avec des oiseaux sauvages, une méthodeexpérimentale typique des études de laboratoire.Les oiseaux testés ont significativement préféréle bras du labyrinthe qui contenait la solution deDMS, démontrant ainsi que les puffins sont capa-bles de détecter ce composé et ont tendance às’orienter vers celui-ci. Dans un second temps,nous avonsmesuré l’attractivité de ce composanten milieu naturel, en pleine mer. Pour cela nousavons déployé une série de taches d’huile végé-tale, dont la moitié contenait du DMS, et nousavons comparé le recrutement d’oiseaux sur lesdeux types de taches. Les taches parfumées auDMSont systématiquement attiré plus d’oiseauxque les taches contrôles, confirmant que la ten-dance des puffins cendrés à s’orienter vers cecomposé s’observe aussi dans le contexte naturelde recherche de nourriture. Nous avons observéque les taches parfumées au DMS ont égalementattiré d’autres espèces telles que les Océanitestempête de Méditerranée (Hydrobates pelagicusmelitensis). Cette espèce de pétrels est rarementobservéedans laMéditerranée, et presque jamaispendant la journée, en raison de leur abondancetrès faible et des habitudes alimentaires noctur-nes (Warham 1990, del Hoyo et al. 1992, Brooke2004). Leur présence sur les taches contenant duDMS souligne que d’autres espèces de pétrelsméditerranéens peuvent être attirées par cettemolécule. Puffins et Océanites tempête ont volécontre le vent vers les taches parfumées au DMSet ont souvent fait une ou deux boucles avant des’envoler, ce qui suggère soit un intérêt pour latache soit une tentative d’analyser la tache enquête d’une proie. Dans notre expérience, tousles pétrels n’ont été recrutés que par l’odeur caraucun oiseau ne s’est arrêté sur les taches ne
Figure 3. La mer Méditerranée centrale et l’Île deLinosa (flèche) où se situe la colonie de Puffincendré. (Image Google Earth ©2010).
GRP : fyssen JOB : annales27 DIV : 07⊕mp⊕DELL⊕ARICCIA⊕Gaia p. 5 folio : 53 --- 18/2/013 --- 14H42
ETHOLOGIE / PSYCHOLOGIE
53

fournissant donc aucun repère visuel pourd’autres oiseaux présents dans les alentours. Cerésultat suggère que les repères olfactifs, seuls,peuvent être suffisants pour localiser les airesproductives.
Conclusions
Les Puffins cendrés sont capables de détecteret sont attirés par le DMS aussi bien dans descontextes naturels qu’artificiels. La sensibilité auDMS, par conséquent, semble être un trait trèsrépandu chez les oiseaux marins, présent danstoutes les espèces spécifiquement testées à cejour, et dans différents milieux marins, oùd’autres repères sont également disponibles. Laréponse générale à ce composé, indépendantedes habitudes alimentaires des espèces, suggèreque le DMS peut être important, non seulementdans la recherche de nourriture, mais aussi pourd’autres tâches. En particulier, l’étude de l’utili-sation de point de repères olfactifs pour la navi-gation globale mériterait d’être approfondie.
RemerciementsCette étude a été financée par la Fondation
Fyssen, par une bourse du Journal of Experimen-tal Biology et par une bourse Marie Curie[SOMA]. Je voudrais remercier Francesco Bona-donna pour la relecture critique de l’article etAurélie Célérier pour la révision du français,ainsi que les personnes quim’ont aidé pendant letravail de terrain : Aurélie Célérier, MarianneGabirot et Pauline Palmas.
Bibliographiee Bang, B.G. 1966. The olfactory apparatus of
tubenosed seabirds (Procellariiformes). ActaAnatomica 65 :391-415.
e Belviso, S., A. Sciandra, et C. Copin-Monté-gut. 2003. Mesoscale features of surface waterDMSP and DMS concentrations in the AtlanticOceanoffMoroccoand in theMediterraneanSea.Deep-Sea Research Part I 50 :543-555.
e Benhamou, S., F. Bonadonna, et P. Jouven-tin 2003. Successful homing of magnet-carrying
Figure 4. Le labyrinthe en Y placé à la colonie. Le bruit de pas de l’oiseau dans le labyrinthe permettait àl’expérimentateur d’établir le bras choisi. C = bras de choix contenant la solution de DMS et celle contrôle.E = entrée de l’oiseau.
GRP : fyssen JOB : annales27 DIV : 07⊕mp⊕DELL⊕ARICCIA⊕Gaia p. 6 folio : 54 --- 18/2/013 --- 14H42
ETHOLOGIE / PSYCHOLOGIE
54

white-chinned petrels released in the open sea.Animal Behaviour 65 :729-734.
e Berthold, P. 2001. Bird Migration : A Gene-ral Survey. edn 2. Oxford University Press,Oxford.
e Bonadonna, F. 2009. Olfaction in petrels :from homing to self-odor avoidance. Annals ofthe New York Academy of Sciences 1170 :428-433.
e Bonadonna, F., C. Bajzak, S. Benhamou,K. Igloi, P. Jouventin ,H.-P. Lipp, etG.Dell’Omo.2005. Orientation in the wandering albatross :interfering with magnetic perception does notaffect orientation performance. Proc. R. Soc.Lond. B 272 :489-495.
e Bonadonna, F., S. Benhamou, et P. Jouven-tin 2003a. Orientation in ”featureless“ environ-ments : the extreme case of pelagic birds. Pages367-377 in P. Berthold, Gwinner, E. & Sonnens-chein, E., editor. Avian migration. Springer-Ver-lag, Berlin, Heidelberg.
e Bonadonna, F., S. Chamaillé-Jammes,D. Pinaud, et H.Weimerskirch. 2003b. Magneticcues : are they important in Black-browed Alba-tross Diomedea melanophris orientation ? Ibis145 :152-155.
e Bonadonna, F., R. A. Holland, L. Dall’Anto-nia, T. Guilford, et S. Benvenuti. 2000. Trackingclock-shifted homing pigeons from familiarrelease sites. Journal of Experimental Biology203 :207-212.
e Bonadonna, F., E. Miguel, V. Grosbois,P. Jouventin , et J. M. Bessiere. 2007. Individual-specific odour recognition in birds : an endoge-nous olfactory signature on petrels’ feathers ?Journal of Chemical Ecology 33 :1819-1829.
e Bonadonna, F., et G. A. Nevitt. 2004. Par-tner-specific odor recognition in an Antarcticseabird. Science 306 :835.
e Brooke, M. 2004. Albatrosses and Petrelsacross the World. Oxford University Press,Oxford.
e Cantin, G., M. Levasseur, M. Gosselin, etS. Michaud. 1996. Role of zooplankton in themesoscale distribution of surface dimethylsul-fide concentrations in the Gulf of St. Lawrence,Canada. Marine Ecology Progress Series141 :103-117.
e Celada, C., G. Gaibani, J. G. Cecere, L. Cala-brese, et P.Piovani. 2009.Aree importanti per gli
uccelli. Dalla terra al mare. Studio preliminareper l’individuazione delle IBA (Important BirdAreas) in ambiente marino. LIPU, Ministerodell’Ambiente and DPN.
e DeBose, J. L., S. C. Lema, etG.Nevitt. 2008.Dimethylsulfoniopropionateasa foragingcue forreef fishes. Science 319.
e DeBose, J. L., et G. Nevitt. 2007. Investiga-ting the association between pelagic fish anddimethylsulfoniopropionate in a natural coralreef system. Mar. Freshw. Res. 58.
e del Hoyo, J., A. Elliott, et J. Sargatal. 1992.Handbook of the birds of the world. Lynx Edi-cions, Barcelona.
e Dell’Ariccia, G., G. Dell’Omo, et H.-P. Lipp.2009. The influence of experience in navigation :GPS tracking of homing pigeons released overthe sea after directional training. Journal ofExperimental Biology 212 :178-183.
e Dell’Ariccia, G., G. Dell’Omo, B. Massa, etF. Bonadonna. 2010. First GPS-tracking ofCory’s shearwater in the Mediterranean Sea.Italian Journal of Zoology 77 :339-346.
e Dell’Ariccia, G., G. Dell’Omo, D. P. Wolfer,et H.-P. Lipp. 2008. Flock flying improvespigeons’ homing : GPS-track analysis of indivi-dual flyers versus small groups. Animal Beha-viour 76 :1165-1172.
e Estrada,M.1996.Primaryproduction in thenorthwestern Mediterranean. Scientia Marina60 (Suppl 2) :55-64.
e Grubb, T. C. 1972. Smell and foraging inshearwaters and petrels. Nature 237 :404-405.
e Hay,M.E., et J.Kubanek. 2002.Communityand ecosystem level consequences of chemicalcues in the plankton. Journal of Chemical Eco-logy 28 :2001-2016.
e Holland, R. A. 2003. The role of visual land-marks in the avian familiar area map. Journal ofExperimental Biology 206 :1773-1778.
e Hutchison, L. V., et B. M. Wenzel. 1980.Olfactory guidance in foraging by procellarii-forms. Condor 82 :314-319.
e Jean, N., G. Bogé, J.-L. Jamet, D. Jamet, etS. Richard. 2009. Plankton origin of particulatedimethylsulfoniopropionate in a Mediterraneanoligotrophic coastal and shallow ecosystem.Estuarine, Coastal and Shelf Sciences 81 :470-480.
GRP : fyssen JOB : annales27 DIV : 07⊕mp⊕DELL⊕ARICCIA⊕Gaia p. 7 folio : 55 --- 18/2/013 --- 14H42
ETHOLOGIE / PSYCHOLOGIE
55

e Jorge, P. E., P. A. M. Marques, etJ. B. Phillips. 2010. Activational effects ofodours on avian navigation. Proc. R. Soc. Lond.B 277 :45-49.
e Lau, K.-K., S. Roberts, D. Biro, R. Freeman,J. Meade, et T. Guilford. 2006. An edge-detectionapproach to investigating pigeon navigation.Journal of Theoretical Biology 239 :71-78.
e Lequette, B., C. Verheyden, and P. Jouven-tin 1989. Olfaction in suantarctic seabirds :itsphylogenetic and ecological significance. Condor91 :732-735.
e Mardon, J., et F. Bonadonna. 2009. Atypicalhoming or self-odour avoidance ? Blue petrels(Halobaena caerulea) are attracted to theirmate’sodour but avoid their own. Behavioural Ecologyand Sociobiology 63 :537-542.
e McTaggart, A. R., et H. Burton. 1992.Dimethylsulfide concentrations in the surfacewaters of the Australasian Antarctic and Suban-tarctic oceans during an Austral summer. Jour-nal of Geophysical Research 97 :14407-14412.
e Nevitt, G. A. 1999. Foraging by seabirds onan olfactory landscape. American Scientist87 :46-53.
e Nevitt, G. A. 2000. Olfactory foraging byantarctic procellariiform seabirds : Life at highReynolds Numbers. Biological Bulletin 198 :245-253.
e Nevitt, G. A. 2008. Sensory ecology on thehigh seas : theodorworldof theprocellariiformesseabirds. Journal of Experimental Biology211 :1706-1713.
e Nevitt, G. A., et F. Bonadonna. 2005a.Seeing the world through the nose of a bird : newdevelopments in the sensory ecology of procel-lariiform seabirds. Marine Ecology ProgressSeries 287 :292-295.
e Nevitt, G. A., et F. Bonadonna. 2005b. Sen-sitivity to dimethyl sulphide suggests a mecha-nism for olfactory navigation by seabirds. Bio-logy Letters 1 :303-305.
e Nevitt, G. A., R. R. Veit, et P. Kareiva. 1995.Dimethyl sulphide as a foraging cue forAntarcticProcellariiform seabirds. Nature 376 :680-682.
e Silverman, E. D., R. R. Veit, et G. Nevitt.2004. Nearest neighbors as foraging cues : infor-mation transfer in apatchy environment.MarineEcology Progress Series 277 :25-35.
e Simó, R. 2001. Production of atmosphericsulfur by oceanic plankton : biogeochemical, eco-logical and evolutionary links. Trends in Ecologyand Evolution 16 :287-294.
e Simó, R., et J. O. Grimalt. 1998. Spring-summer emissions of Dimethyl Sulphide fromthe North-western Mediterranean Sea. Estua-rine, Coastal and Shelf Sciences 47 :671-677.
e Simó, R., J. O. Grimalt, et J. Albaigés. 1997.Dissolved dimethylsulphide, dimethylsulphonio-propionate and dimethylsulphoxide in westernMediterranean waters. deep-Sea Research PartII 44 :929-950.
e Thivent, V. et Bonadonna, B. 2003 Cesoiseauxqui s’oriententàvuede... nez !LaRecher-che 369 :52-55.
e Walcott, C. 2005. Multi-modal orientationcues in homing pigeons. Integrative and Compa-rative Biology 45 :574-581.
e Wallraff, H. G. 2004. Avian olfactory navi-gation : its empirical foundation and conceptualstate. Animal Behaviour 67 :189-204.
e Wallraff, H. G. 2005. Avian Navigation :Pigeon Homing as a Paradigm. Springer-Verlag.
e Warham, J. 1990. The Petrels their ecologyand breeding system. Harcourt Brace Jovano-vich, London.
e Warham, J. 1996. The behaviour, popula-tion biology and physiology of the Petrels. Har-court Brace & Company, London.
e Wiltschko, R., etW.Wiltschko. 2003. Aviannavigation : from historical to modern concepts.Animal Behaviour 65 :257-272.
Olfactory orientation in birdsBirds hold a special place among animal navi-
gators because of their exceptional spatial abili-ties, which manifest themselves most notably inthe context of their return from migratory jour-neys of thousands of kilometres to the samebreeding andwintering area year after year (Ber-thold 2001), and their homing ability (Wallraff2005). Numerous studies on birds’ orientationwere performed using homing pigeons (Columbalivia) as a model species. A homing pigeons’so-called «navigational map», which enablesthemtoapproximate thedirectionofandperhapsdistance to home from distant and unfamiliarlocations, is based on their ability to represent
GRP : fyssen JOB : annales27 DIV : 07⊕mp⊕DELL⊕ARICCIA⊕Gaia p. 8 folio : 56 --- 18/2/013 --- 14H42
ETHOLOGY / PSYCHOLOGY
56

the heterogeneous distribution of atmosphericodours in space (Wallraff 2005). This does notpreclude the fact that pigeons can use additionalsensory information, such as information on theearth’s magnetic field (Wiltschko and Wiltschko2003). However, after more than 30 years ofstudies (Wallraff 2005), the use of olfactory cuesis still one of the most debated subjects in pigeonorientation (Wiltschko andWiltschko2003,Wal-lraff 2004, Wallraff 2005, Jorge et al. 2010).
The lack of clear answers concerning the res-pective role of alternative orientation cues is alsodue to the fact that pigeons generally fly overland, and landscape features have been shown todeeply influencenavigation, evenwhenunknown(Holland 2003, Lau et al. 2006, Dell’Ariccia et al.2009). In addition, the presence of visual land-marks may modify the result of experimentaltreatments, such as clock-shift or anosmia (Bona-donnaet al. 2000,Wallraff2005).Eventually, it isalways more evident that pigeons use a multimo-dal navigation system and are able to shift fromone navigation strategy to another according tothe circumstances (Walcott 2005,Wallraff 2005,Dell’Ariccia et al. 2008, Dell’Ariccia et al. 2009).It is therefore almost impossible to isolate therelative importance of different cues used fororientation in this species, and, as a conse-quence, to demonstrate definitively an olfactoryorientation mechanism. In this perspective,petrels are probably a better model. Contrary toland, the marine environment where these birdslive is seemingly featureless over thousands ofsquare kilometres, so excluding the landscapecomponent among the possible orientation cues.Moreover, previous studies showed that petrelsappear to not rely on magnetic cues to orient(Benhamou et al. 2003, Bonadonna et al. 2003b,Bonadonna et al. 2005).
Petrels and orientation in a pelagicenvironment
Petrels (order Procellariiformes) are characte-rized by their pelagic and wide-ranging lifestyle.These long living birds spend most of their livesin flight over the sea and are tied to small oceanicislandsonly forbreeding.So, theyhave to localizepatchily distributed foraging areas as well astheir breeding colonies within the vast, seemin-gly featureless oceanic environment (Warham1996). During the breeding season, procellarii-forms are central place foragers (Stephens and
Krebs 1986), i.e. they are temporally constrainedto return to their nest either to relieve a mateor to provision their unique chick, while theforaging grounds remain pelagic. Under theselifestyle constraints, high efficiency in locatingproductive foodsourceswill ensureeffective fora-ging and breeding success.
Procellariiforms have a remarkably developedolfactoryneuroanatomycompared tomanyotherbirds (Fig. 1.Bang1966). There is rising evidencethat their well-developed sense of smell plays animportant function in a number of behavioursranging from locating patchily distributed foodresources in the ocean (Nevitt et al. 1995), tohoming (Bonadonna 2009) and to nest and materecognition (Bonadonna and Nevitt 2004, Bona-donnaet al. 2007,MardonandBonadonna2009).
In seabirds, and particularly petrels, the use ofthe sense of smell to locate food patches wassuggested long ago (Grubb 1972, Hutchison andWenzel 1980, Lequette et al. 1989), but the inves-tigation of how petrels use naturally occurringscented compounds as foraging and navigationcues is relatively new. The basic idea of the olfac-tory orientation theory is that petrels may usestable volatile compounds to build a large-scalemapasanolfactory landscapesuperimposedoverthe water surface (Fig. 2). Changings in the olfac-tory landscape might indicate to a wandering
Figure 1. Diagrammatic section through theolfactory chamber of Snow petrel. The olfactoryepithelium is evidenced in red (Modified fromBang1966).
GRP : fyssen JOB : annales27 DIV : 07⊕mp⊕DELL⊕ARICCIA⊕Gaia p. 9 folio : 57 --- 18/2/013 --- 14H42
ETHOLOGY / PSYCHOLOGY
57

petrel its position and/or the proximity of fora-ging areas (Nevitt 2000,Bonadonnaet al. 2003a).
Dimethylsulfide (DMS), a volatile compoundthat occurs naturally in worldwide oceans, maybe the perfect candidate as olfactory cue forseabirds to locate and exploit productive areas(Nevitt et al. 1995, Nevitt 1999, Nevitt and Bona-donna 2005b). DMS is a by-product of the meta-bolic decomposition of dimethylsulfoniopropio-nate (DMSP) produced during phytoplanktongrazing. Its production is often associated withzooplankton feeding andareaswithhighprimaryproductivity, i.e. high phytoplankton concentra-tion (Cantin et al. 1996, Simó 2001, Jean etal. 2009). DMSP may be a particularly strongsource of olfactory signal for zooplankton preda-tors, and in situ observations indicate that scho-ols of small fish aggregate along the periphery ofplankton blooming areas (DeBose and Nevitt2007, DeBose et al. 2008). Eventually, local ele-vation in DMS may, opportunistically, alert
higher order predators of rapidly accumulatingaggregations of zooplankton and zooplanktonpredators, i.e. fish and squid that are among themain prey for many petrel species (del Hoyo etal. 1992,Warham1996,Hay andKubanek 2002).Indeed, satellite telemetry revealed an associa-tion between areas with high DMS emissions,prey aggregations and foraging grounds of diffe-rent petrel species (Reviewed by Nevitt 2000).
Among petrel species, however, responsive-ness to DMS is quite variable suggesting thatolfactory sensitivity to this compound may belinked to different ecological strategies for preylocation. For some species (e.g. stormpetrels andprions), olfactory sensitivity to DMS provides anearly detection system for opportunistic foragingbefore other birds arrive. These early detectorschange the nature of prey-associated cues: fee-ding at the patch, they provide visual cues toother birds and induce the release of additionalodours, such as those linked to macerated krill.
Figure 2. Olfactory landscape. Productive areas are often associated with bathymetric features, such asupwelling zones and seamountains. Here, DMS emissions are higher and can provide olfactory signposts fornavigation to a wandering seabird (From Thivent and Bonadonna, 2003 ©Caroline Picard).
GRP : fyssen JOB : annales27 DIV : 07⊕mp⊕DELL⊕ARICCIA⊕Gaia p. 10 folio : 58 --- 18/2/013 --- 14H50
ETHOLOGY / PSYCHOLOGY
58

Thus, late detector species (e.g. albatrosses andgiant petrels) may employ amultimodal foragingstrategy that relies on visual cues as well as acombination of olfactory cues to locate prey (Sil-vermanet al. 2004,Nevitt andBonadonna2005a,Nevitt 2008).
In addition to the foraging function, DMS hasbeen also proposed as a signpost for global orien-tation. Over the oceans, highly productive areas,and so higher DMS concentrations, are associa-ted with bathymetric features such as upwellingzones, sea mountains and shelf waters (McTag-gart and Burton 1992). Therefore, the DMSsurface concentration profile could represent anavigable olfactory landscape to seabirds supe-rimposed on the apparently featureless watersurface (Fig. 2. Nevitt et al. 1995, Nevitt 2000,Bonadonna et al. 2003a).
Overall, these studies suggest a strong role ofbiogenic cues in oceanic petrels in a foragingcontext and for global orientation. However, theinvestigation of how petrels use naturally occur-ring scented compounds as foraging and homingcues has so far been limited to few oceanic envi-ronments and few species. Thus, it is not knownwhether the use of olfactory cues to locate food atsea and the nest at the colony is a local and/orspecies-specific phenomenonor a general adapta-tion of seabirds. In particular, nothing is knownregarding petrels breeding and foraging in closeseas, where oceanographic conditions may beextremely different, and visual landmarks are inclose proximity (i.e. coastlines and islands).
The example of the Mediterranean SeaIn the aim to understand the role and use of
DMS in a different ecological setting, we investi-gated the response to this compound byMediter-ranean Scopoli’s shearwaters (Calonectris diome-dea), breeding at the colony on Linosa island(Pelagian Isles, Sicilian Channel, Italy: 35o52’ N,12o52’E,Fig. 3).Here, foraging tripsappear tobeshorter, both in terms of duration (lasting, onaverage, 9.4 days during incubation and 5.5 daysduring chick rearing) and distance travelled.Nonetheless, Scopoli’s shearwaters cover an ave-rage of 904 km per foraging trip during incuba-tion and 662 km per trip during chick rearing,with an average daily distance of 135 km (Celadaet al. 2009). Therefore, the distance betweenMediterranean coasts could easily be covered bya flying petrel within a day and several timeswithin a foraging trip; coasts may provide thenecessary landmarks for orientation in thisclosed basin. Foraging grounds of Scopoli’sshearwaters breeding in Linosa Island aremainly along the Tunisian coast or just offshoreof the island (Celada et al. 2009, Dell’Ariccia etal. 2010). The Tunisian coastal area presentsconspicuous visual landmarks, useful for orien-tation, provided by the coastline itself and by thenumerous islands that could help to point toforaging spots once the birds are in the area.Moreover, this sea presentsmoderateDMSemis-sions, consistent with the low productivity of itsopen waters, and lower surface concentrationsthan both Atlantic and Pacific oceans (Simó etal. 1997, Simó and Grimalt 1998, Belviso etal. 2003). Areas of higher surface concentrationsof DMS are found along coastlines, where pri-mary productivity is higher (Estrada 1996, Simóet al. 1997). In these conditions, the use of DMSas olfactory signpost might be less useful than inthe open ocean, inducing a reduced or absentability to detect this compound and tendency toorient towards it. Therefore, Mediterraneanshearwatersmight be less attracted byDMS thanother petrel species.
In this study, two different experiments werecarried out: first, we tested the ability of Scopoli’sshearwaters to detect and respond to DMS in aY-maze at the colony, a laboratory experimentwith wild birds in their natural environment(Fig. 4). In this test, we presented birds with abinary choice between DMS and a control odour.
Figure 3. The central Mediterranean Sea andLinosa Island (arrow) where the Scopoli’s shearwa-ters’ colony is located. (Image Google Earth ©2010)
GRP : fyssen JOB : annales27 DIV : 07⊕mp⊕DELL⊕ARICCIA⊕Gaia p. 11 folio : 59 --- 18/2/013 --- 14H43
ETHOLOGY / PSYCHOLOGY
59

Tested birds significantly preferred the arm ofthe maze that contained the DMS solution, pro-viding evidence that these shearwaters are ableto smell this compound and have a tendency toorient towards it. Second, we tested the responseto this compound under natural conditions atsea.We compared thenumbers of birds attractedby DMS-scented and non-scented vegetable oilslicks deployed on the water surface. DMS-scen-ted slicks systematically attracted more birdsthan control slicks, confirming that the tendencyof Scopoli’s shearwaters to orient to this com-pound extends to a natural foraging context. Inaddition to the Scopoli’s shearwater, DMS-scen-ted slicks also attracted the MediterraneanStorm Petrel (Hydrobates pelagicus melitensis).These birds are rarely observed in the Medi-terranean, and almost never during daylight,because of their very low abundance and noctur-nal feeding habits (Warham 1990, del Hoyo etal. 1992, Brooke 2004). Their presence at theslicks underlines that other Mediterraneanpetrel species may be attracted by this molecule.Shearwaters and Storm Petrels flew upwind to
theDMS scented slick and oftenmade one or twoloops over it beforeflying away, suggesting eitheran interest in the slick or an attempt to scan theslick for prey. In our experiment, all shearwaterswere recruited only by the smell since no birdstopped at the slick so there were no visual cuesproviding social enhancement to other birds inthe area. This result suggests that olfactory cuesalone may be sufficient to locate productivegrounds.
ConclusionsIn conclusion, Scopoli’s shearwaters are able
to sense and are attracted to DMS both in artifi-cial andnatural contexts. The sensitivity toDMS,therefore, may be a trait that is widespreadamong marine birds, present in all species speci-fically tested todate, and indifferentmarine envi-ronments. Such general response to this com-pound, independent of feeding habits, suggeststhatDMSmaybe important, not only in foraging,but also in other tasks. This idea merits furtherinvestigation, particularly as it relates to the useof olfactory signposts for global navigation.
Figure 4. The Y-maze in the colony. The sounds of the bird walking in the maze allowed the experimenter toeasily assess arm-choice. C = choice arms containing DMS and control solution. E = bird entrance.
GRP : fyssen JOB : annales27 DIV : 07⊕mp⊕DELL⊕ARICCIA⊕Gaia p. 12 folio : 60 --- 18/2/013 --- 14H43
ETHOLOGY / PSYCHOLOGY
60

AcknowledgementsThis study was granted by the Fyssen Founda-
tion, by a Journal of Experimental Biology Travel-ling Fellowship and by a Marie Curie intra-Euro-pean Fellowship [SOMA]. I would like to thank
Francesco Bonadonna for critical reading of themanuscript and Aurélie Célérier for proofreadingthe French version. I am also grateful to peoplewho helpedme during fieldwork: Aurélie Célérier,Marianne Gabirot and Pauline Palmas.
GRP : fyssen JOB : annales27 DIV : 07⊕mp⊕DELL⊕ARICCIA⊕Gaia p. 13 folio : 61 --- 18/2/013 --- 14H43
ETHOLOGY / PSYCHOLOGY
61

La complainte des bébés phoques en Alaska :une histoire de soutien maternel
Carole Di Poi
Chercheur post-doctoral au sein du Groupe Mémoire et Plasticité comportementale (EA4259),Université de Caen Basse-Normandie.
RésuméComprendre l’écologie des pinnipèdes est essentiel pour déterminer les causes des déclins des
populations observés en Alaska. Néanmoins, les études menées nécessitent que les animaux soientcapturés, temporairementmarquésoude façonpermanentetparfoismême,deséchantillonsde tissusprélevés. De telles procédures sont stressantes pour des animaux sauvages, il est donc importantd’évaluer leurs conséquences à court- et long-termes dans le but d’améliorer le succès des activités deconservation et minimiser les effets négatives de la gestion des espèces. L’inquiétude est particuliè-rement grande pour les chiots non sevrés qui sont extrêmement vulnérables pendant la période delactation et qui pourraient être plus sensibles aux perturbations humaines, en particulier s’ils sontséparés de leur mère. Dans cette étude, nous avons mesuré les effets de la présence/séparationmaternelle sur la physiologie du stress et le comportement de jeunes phoques communs (Phocavitulina richardsi) sauvages au cours d’un protocole de capture-maintien en captivité-prélèvement detissus. Les résultats ontmontré que la présence de lamèremodérait significativement l’amplitude dela réponse glucocorticoïde au stresseur de leur chiot ; alors que chez des chiots toujours dépendantsou chez des jeunes sevrés capturés seuls, les concentrations de cortisol sérique étaient significative-ment plus élevées. De plus, les chiots présentaient une agitation et un nombre de vocalisationsimportants dès lors qu’ils étaient séparés de leur mère par rapport à de jeunes phoques seuls depuisune plus longue période de temps. Cette étude est le premier exemple d’amortissement social chez unmammifère marin et apporte de nouveaux éléments quant à l’impact des activités anthropiques surle bien-être des jeunes pinnipèdes. Enfin, ces résultats pourront servir de support dans le choix detechniques de gestion plus judicieuses dans le respect de l’écologie des espèces afin de diminuerefficacement le stress des animaux au cours des procédures scientifiques.
Mots-clésPhoque commun, vocalisations, agitation, stress, glucocorticoïdes, amortissement social.
The lament of harbor seal pups in Alaska:a story of maternal support
AbstractThe monitoring of pinniped life-history is essential to determinate the causes of the Alaskan
population declines. However, these studies require that the animals are captured, temporary orpermanently marked and sometimes tissue samples collected. Since such procedures are potentiallystressful for the animals, it is important to examine their consequences on short- and long-terms inorder to improve the success of conservation activities and minimise the negative effects ofmanagement.A concern is especially raised for pre-weanedpups that are extremely vulnerable duringlactation andmore sensitive to human disturbance especially if they are separated from theirmotherduring procedures. In this study, we studied the effects of maternal presence/separation on stressphysiology and behavior during a capture-handling-restraint protocol in free-ranging Pacific harborseal (Phoca vitulina richardsi) pups. The results showed that the presence of the mother significantlybuffered themagnitudeof theglucorticoid response to the stressor in theoffspringwhereasdependent
GRP : fyssen JOB : annales27 DIV : 08⊕mp⊕DI⊕POI⊕Carole p. 1 folio : 62 --- 18/2/013 --- 16H22
ETHOLOGIE / PSYCHOLOGIE
62

pups alone (caught without their mother) and weaned pups exhibited high levels of serum cortisol.Moreover, pups showed significantly higher levels of agitation and vocalizations as soon as separatedfrom their mother compared to pups separated for a longer period of time. The results give the firstexample of social buffering in marine mammals and bring new insights on the impact of humanprocedures on pinniped pup welfare. Moreover it opens new perspectives to develop judiciousmanagement techniques in respect to the ecology of the species to effectively reduce stress of theanimals during scientific procedure.
KeywordsHarbor seal, vocalizations, agitation, stress, glucocorticoids, social buffering
Le déclin des populations de phoquescommuns en Alaska
Les populations de phoques communs Phocavitulina richardsi ont subi des fluctuations démo-graphiques importantes au cours des trente der-nières années dans le Pacifique Est, et en parti-culier dans le Golfe d’Alaska (Hoover-Miller et al.2011). Selon les scientifiques, le nombre de pho-ques a subitement décliné de 7 à 67 %, selon lesannées et les régions, depuis les derniers recen-sements effectués dans les années 70/80 (revuedansHoover-Miller et al. 2011). Les causes proxi-males de ces fluctuations n’ont jamais été déter-minées. Néanmoins, ces déclins coïncident avecles changements écologiques attribués à l’oscilla-tion décennale du Pacifique (ODP), survenue de1976 à 1977, qui a fait varier la température del’eau dans le bassin de l’océan Pacifique (Hare etMantua 2000). Quoique des conditions de glacesansprécédent soient associéesà l’ODP, les chan-gements climatiques à long termedus au réchauf-fement de la planète ont eux aussi un impact surla fonte des glaces et pourraient expliquer lesdéclins des populations de phoques observéesdans certaines régions. Par exemple, dans lesud-est de l’Alaska, le parc national de GlacierBay et d’autres fjords comme Tracy Arm et Endi-cott Arm sont des sites importants de mise baspour le phoque commun (Mathews et Pendleton2006). De nombreux phoques migrent de façonsaisonnière dans les eaux des fjords pour sereposer, muer, s’accoupler, mettre bas et allaiterleur chiot (figure 1). En effet, la glace dérivantefournit des plateformes flottantes sécuriséespour les mères et leur progéniture qui peuventallaiter sans interruption (Blundell et al. 2011)et apporter les meilleurs soins à leur jeune pen-dant la période estivale (Muelbert et Bowen1993). Les phoques qui vivent sur les glaces
flottantes subissent une perte importante de leurhabitat estival suite à la fonte de la glace de mer.
Les campagnes de recensementet d’échantillonnage des phoques
Afin de faire le bilan démographique à long-terme et de suivre l’état de santé des populationsde phoques communs, des campagnes d’échan-tillonnage et de recensement sont régulièrementorganisées pendant la mue, à la fin de l’été, dansle Golfe d’Alaska. Cependant, relativement peude sites ont été étudiés pendant la saison desnaissances (Hoover-Miller et al. 2011). Plusieursétudes montrent que ces activités de recherchepeuvent entraîner des perturbations (e.g. bruit,capture, maintien en captivité, prélèvements detissus) pouvant être stressantes pour les ani-maux (e.g. Engelhard et al. 2002 ; Petrauskas etal. 2008). Il paraît donc important de connaîtreles conséquences de ces pratiques à la fois sur laphysiologie et le comportement des animaux,mais les publications à ce sujet restent rares.
Figure 1. Une mère phoque commun et son chiotsur une plateforme de glace en Alaska(photos ©C. Di Poi).
GRP : fyssen JOB : annales27 DIV : 08⊕mp⊕DI⊕POI⊕Carole p. 2 folio : 63 --- 18/2/013 --- 16H23
ETHOLOGIE / PSYCHOLOGIE
63

L’inquiétude des scientifiques est d’autant plusgrande pour les mères allaitantes et leur jeune.En effet, la lactation est l’une des périodes la pluscoûteuse en termes de demandes énergétiquesque les femelles expérimentent (e.g. Oftedal1985) ; et la progéniture dépend directement del’investissement et de la condition physique deleur mère pour grandir (e.g. Mellish et al. 1999).C’est donc un moment où les animaux sont par-ticulièrement vulnérables face aux perturbationshumaines.
L’amortissement social sur la réponseau stress
Lorsqu’il est confronté à un stimulus stres-sant, l’organisme va se préparer à fuir, si celaest possible, ou à résister. La réponse au stressest adaptative et universelle chez l’ensembledes Vertébrés. Elle fait intervenir l’axe neuro-endocrinien hypothalamo-hypophyso-surrénales(HHS) qui résulte en une série de cascades hor-monales dont les constituants majeurs sontl’hormone hypothalamique corticotrope (CRH),l’hormone adrénocorticotropine (ACTH) et lesglucocorticoïdes (Sapolsky2002). Les glucocorti-coïdes sont secrétés par les glandes surrénales etlibérés dans le sang ; le cortisol est typiquementchoisi pour évaluer l’état de stress chez lesMammifères, y compris chez les Pinnipèdes (e.g.Myers et al. 2010).
L’amplitude de cette réponse au stress peutêtremodulée par le contexte social. Par exemple,lorsqu’il existe un fort attachement émotionnelentre une mère et sa progéniture, la séparationdes deux membres de la dyade peut provoquerune augmentationde la réponse de l’axeHHSà lasuite d’un événement stressant. Au contraire, laprésence du partenaire social peut amortir leseffets du stress. L’aptitude du partenaire àréduire la réponse au stress, que ce soit au niveauphysiologique et/ou comportemental, est com-munément appelé « amortissement social » ou« social buffering » en anglais (Hennessy 1997 ;DeVries et al. 2003 ;Hennessy et al. 2009). L’effetest plus oumoins important dépendammentde lanature et de la qualité de la relation que la pairepartageait, ainsi que dumoment et de la durée dela séparation. Dans le contexte de la relationmère-jeune, la puissance des liens que la mère etson jeune développe dépend de la structuresociale de l’espèce et du degré de mobilité du
jeune. Selon la théorie de Gubernik (1981), lesliens sont plus forts lorsque le jeune est seul,précoce et très mobile et que la structure socialedu groupe est grégaire, au moins pendant lapériode de reproduction.
La relation mère-chiot chez le phoquecommun
Chez les pinnipèdes, il existe deux stratégiesmaternelles principales (Trillmich 1996) : la stra-tégie du jeûne chez les Phocidés de grande tailleet la stratégie d’approvisionnement chez les Ota-riidés.Chez lesgrandsPhocidés, la femelleallaiteson chiot en continu pendant une période relati-vement courte (de 4 jours à 4 mois) ; elle nes’alimente pas et utilise ses réserves corporellespour couvrir ses propres besoins physiologiquesainsi que la lactation de son petit. Chez les Ota-riidés, la femelle allaite son chiot de manièrediscontinue pendant une période relativementlongue (de 4 mois à 3 ans) ; elle alterne desvoyages en mer en vue de s’alimenter et decourtes visites à terre pour allaiter ce dernier.Contrairement aux grands Phocidés, le phoquecommun a une stratégie d’allaitement intermé-diaire. La femelle n’est pas en mesure de jeunerdu fait de sa petite taille et doit, tous les jours,aller en mer pour s’alimenter pendant la périodede lactation (BonessetBowen1996).Néanmoins,parce que cette période est extrêmement courtechez cette espèce (3 à 6 semaines en moyenne ;Muelbert et Bowen 1993 ; Hoover-Miller 1994),cela implique que la mère et son chiot doiventrester en contact le plus de temps possible. Ilexiste un lien bilatéral entre lamère et son jeune,qui jouent tous les deux un rôle pour garder lacohésion de la paire ; le jeune très précoce ensuivant samère continuellement enmer car il esttrès mobile, et la mère en réduisant le risqued’être séparée de son petit en apportant unesurprotection à ce dernier (Lawson et Renouf1987). Dans ce contexte de fort attachementfiliatif pendant la période de lactation, le phoquecommun est une espèce de choix pour étudier leseffets de l’absence maternelle sur la réponse austress du chiot.
L’objectif de l’étudeLe but de l’étude a été de déterminer si la
réponse au stress du chiot, induite à la suited’une procédure d’échantillonnage (capture/
GRP : fyssen JOB : annales27 DIV : 08⊕mp⊕DI⊕POI⊕Carole p. 3 folio : 64 --- 18/2/013 --- 16H23
ETHOLOGIE / PSYCHOLOGIE
64

maintien/prélèvement), pouvait être moduléepar la présence ou l’absence de la mère chez desphoques communs du Pacifique Phoca vitulinarichardsi sauvages. Trois catégories de chiots ontété étudiées : chiots dépendants capturés avecleur mère mais brièvement séparés pendantl’échantillonnage, chiots dépendants mais cap-turés sans leur mère et des jeunes phoquesrécemment sevrés capturés seuls. La réponseau stress a été évaluée par une approche com-portementale, mesure du nombre de vocalisa-tions et du niveau d’agitation générale, et uneapproche physiologique en mesurant l’activitéde l’axe HHS par la quantification du cortisolsérique.
Expérimentations
Cette étudea été réalisée grâce à l’obtentiondupermis de recherche #358-1757 délivré par laNational Oceanic and Atmospheric Administra-
tion (NOAA)/National Marine Fisheries Service(NMFS) et le permis #09-08 délivré par le Careand Use Committee du Department of Fish andGame’s en Alaska (ADF&G). Toutes les capturesontété réaliséesenévitantdecauserunniveaudeperturbation inacceptable des colonies de pho-ques.
Les deux sites d’échantillonnageCette étude a été menée pendant la saison
estivale des naissances à Endicott Arm et TracyArm, deux fjords situés à environ une soixan-taine de kilomètres au sud de Juneau en Alaska,USA (figure 2).
La procédure de capture des phoquesLes phoques ont été capturés par des filets en
nylon monofilament de (30 m x 3 m) entre 8 :30et 16 :00 (figure 3). Toutes les paires mère-chiotet les jeunes phoques sevrés seuls étaient trans-
Figure 2. Carte des deux sites d’échantillonnage (fourni par ADFG; photos ©C. Di Poi).
GRP : fyssen JOB : annales27 DIV : 08⊕mp⊕DI⊕POI⊕Carole p. 4 folio : 65 --- 18/2/013 --- 16H23
ETHOLOGIE / PSYCHOLOGIE
65

portés dans un bateau servant de laboratoirepuis échantillonnés. Les chiots toujours dépen-dants de leur mère mais capturés seuls étaientéchantillonnés en quelques minutes directementà bord d’un pneumatique de capture et libérés àl’endroit exact où ils avaient été capturés.
Au total, 41 phoques ont été capturés pourcette étude : 5 chiots dépendants et leur mère(Chiots Dépendants Accompagnés), 29 chiotsdépendants capturés sans leur mère (ChiotsDépendants Seuls) et 7 jeunes sevrés capturésseuls (Jeunes Sevrés Seuls). Chez le phoque duPacifique Nord-Est, le sevrage est généralementatteint à l’âge de 1mois après la naissance (Muel-bert et Bowen 1993). Dans notre étude, les jeunesétaient considérés comme « sevrés » si aucunefemelle n’avait été observée aux alentours pen-dant la capture et si leur masse corporelle étaitsupérieure à 25 kg.
Observations comportementalesLes niveaux d’agitation et de vocalisation ont
été quantifiés pour les trois catégories de chiotspendant la 1ère collecte de sang, entre le momentoù lephoqueétaitmaintenuphysiquementà lafindu prélèvement de sang (durée moyenne : 3.9 ±0.3min)dans le butd’évaluer la réponseau stressinitiale des animaux. Pour chaque chiot, le nom-bre de vocalisations parminute a étémesuré et leniveau d’agitation estimé sur une échelle allantde 1 à 3 avec 1 = calme, aucun mouvement duphoque, 2 = agitation modérée et mouvement dela tête du phoque, et 3 = forte agitation, le phoqueétait difficile à maintenir.
Procédure des prélèvements sanguinset dosage du cortisol
Tous les animaux étaient maintenus dans descages individuelles opaques afin d’éviter lescontacts physiques entre eux. Les mères et leurjeune étaient maintenus ensemble. Tous les indi-vidus étaient pesés afin de déterminer le volumede sang à collecter. Les animaux étaient ensuitemaintenus physiquement, sans l’administrationd’anesthésiant, et le sang collecté à partir de laveine vertébrale en position extra-durale. Lesmères et leur chiot ont toujours été échantillon-nés en priorité et enmême temps ; néanmoins ilsétaient brièvement séparés pendant les prélève-ments et réunis après.
Les CDS ont été échantillonné 16 à 213 minu-tes après la capture (temps moyen : 69.2 ±8.9 min) et libérés à l’endroit exact de leur cap-ture. Pour les autres phoques, l’intervalle detemps entre la capture et les prélèvements san-guins variait de 103 à 255 min (temps moyen :164.6 ± 14.0minpour lesCDAet 171.7 ± 19.3minpour les JSS). Enfin, l’heure de la collection dusang oscillait entre 10 :04 à 16 :13 pour les chiotsdépendants seuls, et de 11 :03 à 14 :43 pour lesautres jeunes.
Une fois la prise de sang terminée, tous lesanimaux étaient sexés, pesés et les CDS libéréspour leur permettre de retrouver leur mère. Lesautres phoques étaient replacés dans leur cage(seuls ou avec leur mère) afin qu’ils récupèrentpendant 2.5 heures avant la 2ème collecte desang. Finalement, tous les phoques ont étérelâchés proche de leur lieu de capture ; lespaires mère-chiot ont été relâchées ensemble.Les concentrations de cortisol ont été mesuréesà t0 et t2.5 dans le sérum par dosages radio-immunologiques en phase solide en utilisantl’iode 125 comme marqueur radioactif.
Résultats
Les paramètres pouvant influencerles concentrations de cortisol
Il n’y avait pas d’influence du temps s’écoulantentre la capture et les prélèvements sanguins surles concentrations de cortisol (Analyse de régres-sion linéaire,R2 = 0.0981 ; P = 0.256). Il n’y avaitpas non plus d’influence de l’heure de la journéeà laquelle les prélèvements sanguins avaientété effectués (Analyse de régression linéaire,R2 = 0.0892 ; P = 0.182). De plus, nous n’avons
Figure 3. Positionnement du filet monofilament àEndicott Arm, Alaska et pneumatique de capture(photo ©C. Di Poi).
GRP : fyssen JOB : annales27 DIV : 08⊕mp⊕DI⊕POI⊕Carole p. 5 folio : 66 --- 18/2/013 --- 16H23
ETHOLOGIE / PSYCHOLOGIE
66

pas trouvé d’influence de la masse (Analyse derégression linéaire, R2 = 2.91.10-5, P =0.974) oudu sexe (t-test non apparié, P = 0.930) sur lesniveaux de cortisol.
Les réponses comportementales des chiots
Le nombre de vocalisations ne différait passignificativement entre les groupes de chiots(Kruskal-Wallis, P = 0.056, n = 37 ; figure 4A).Néanmoins, le nombre de vocalisations restaitplus élevé chez les jeunes accompagnés de leurmère depuis la capture mais brièvement séparé-ment pendant l’échantillonnage que chez les jeu-nes capturés seuls, qu’ils soient dépendants ousevrés. De même, le niveau d’agitation ne diffé-rait pas significativement entre les groupes(Kruskal-Wallis, P = 0.0973, n = 37 ; figure 4B) ;même si le niveau d’agitation tendait à augmen-ter pour les chiots récemment séparés de leurmère. Globalement, les niveaux de vocalisations
et d’agitation avaient tendance àdiminuer quandle temps de séparation de la mère augmentait.
Les réponses hormonales des chiotsÀt0, lesniveauxdecortisoldifféraiententre les
groupes (ANOVA, P = 0.005, n = 41 ; figure 5A).Comme attendu, la présence de la mère lorsd’un événement stressant semblait modérer laréponseglucocorticoïdedes jeunes comparéeauxconcentrations mesurées chez les chiots dépen-dants et les jeunes phoques sevrés capturésseuls (Fisher’s PLSD, P = 0.0011 et P = 0.0186respectivement ; figure 5A). À t2.5, les niveaux decortisol ne différaient plus entre les chiots accom-pagnés et les jeunes sevrés (t-test non apparié,P = 0.200 ; figure 5B) ; bien que les jeunes accom-pagnés présentaient toujours une concentrationde cortisol plus faible. L’analyse de variance àmesures répétées n’a pas montré d’évolutionsignificative des concentrations de cortisol dansle temps pour l’ensemble des groupes (ANOVA àmesures répétées, P O 0.05, données non mon-trées).
DiscussionLa notion d’« amortissement social a été défi-
nie chez de nombreuses espèces présentant unfort attachement émotionnel entre la mère et saprogéniture » (e.g. DeVries et al. 2003 ;Hennessyet al. 2009). Selon la nature et la qualité du lienfiliatif, la séparation des deux membres de ladyade peut provoquer une amplification de l’axeHHS au cours d’un événement stressant (e.g.Hennessy 1997) ou au contraire, la présence dupartenaire social peut modérer cette réponse(von Holst 1998).
Les réponses comportementalesLes mesures comportementales telles que les
vocalisations, le repos ou au contraire l’agitationpeuvent être utilisées pour évaluer la réponse austress et/ou la douleur chez les animaux (Lay etal. 1992a,b,c). Immédiatement après la sépara-tion de la mère, la réponse comportementaleinitiale du jeune est caractérisée par l’émissiond’un nombre élevé de vocalisations (e.g. Hen-nessy et Moorman 1989) et par un niveau d’acti-vitéaccru (e.g. chez lesprimatesMinekaetSuomi1978). Après une séparation prolongée, le jeuneprésente une inactivité générale et affiche uneposture avachie caractéristique chez les petitsprimates. Ces deux phases sont respectivementappelées « la phase de protestation » et la « phase
Figure 4. Comparaison du nombre de vocalisations(A) et du niveau d’agitation (B) entre les troisgroupes de jeunes phoques: chiots dépendantscapturés avec leur mère mais séparés pendant lesobservations (CDA), chiots dépendants seuls (CDS)et jeunes sevrés seuls (JSS) à l’aide de tests deKruskal-Wallis.
GRP : fyssen JOB : annales27 DIV : 08⊕mp⊕DI⊕POI⊕Carole p. 6 folio : 67 --- 18/2/013 --- 16H23
ETHOLOGIE / PSYCHOLOGIE
67

de résignation » (Bowlby 1979). Chez le cochond’Inde,Hennessy etMoorman (1989) ontmontréque les jeunes seuls avaient tendance à vocaliserplus que des jeunes accompagnés de leur mèrependant les 30 premières minutes post-sépara-tion (phase de protestation). 60 à 90min après laséparation, le nombre de vocalisations commen-çait à diminuer jusqu’à atteindre lemême niveaude vocalisations que les jeunes accompagnés deleurmère 150-180min après la séparation (phasede résignation). Néanmoins, les mêmes auteursn’ont pas trouvé de différence dans le niveaud’activité locomotrice entre ces deux groupes dejeunes alors qu’une augmentation de l’activitélocomotrice a été observée chez de jeunes pri-mates immédiatement après la séparation(Mineka et Suomi 1978). Dans notre étude,les paires mère-jeune étaient physiquement etvisuellement séparéesquelquesminutesaucoursdes prélèvements sanguins. Pendant cette sépa-ration, les jeunes montraient des niveaux élevésde vocalisations et d’agitation comparés aux jeu-nes capturés seuls et séparés de leurmère depuis
plus longtemps, au moins depuis la capture pourles chiots dépendants. Nos résultats montrentbien la phase de protestation, décrite ci-dessusqui est typiquement observée chez depetitsmam-mifères immédiatement après la séparationmaternelle. En revanche, le faible niveau d’agi-tation et le nombre peu élevé de vocalisationsmesuré chez les chiots non sevrés seuls peu-vent s’expliquer par le fait qu’en moyenne1 heure s’était écoulée entre la capture et lesobservations comportementales (temps moyen :69.2 ± 8.9 min), et que cela coïncide au laps detempsqui s’écoule avant l’entréedans la phasederésignation chez le cobaye avec la baisse signifi-cative du nombre de vocalisations observée 60 à90min après la séparationmaternelle (Hennessyet Moorman 1989).
Les réponses hormonalesLa capture, le maintien en captivité et les
prélèvements de tissus sont autant de facteursstressants pour les animaux sauvages (Engel-hard et al. 2002 ; Petrauskas et al. 2008). De pluschez les rongeurs, les jeunes répondent forte-ment lorsqu’ils sont séparés de la figure mater-nelle et exposés à un événement stressant (e.g.exposition à un nouvel environnement) à traversl’activation de la cascade hormonale de l’axeHHS qui se traduit par l’augmentation desconcentrationsplasmatiques enACTHet cortisolqui atteignent un maximum 90 minutes post-séparation (Hennessy etMoorman1989). Chez lephoque commun, nos résultats montrent que lesjeunes accompagnés de leur mère pendant lacaptureprésentaientune réponsedecortisol séri-que moins élevée que les jeunes capturés seuls,qu’ils soient dépendants ou déjà sevrés ; ten-dance qui se maintient 2.5 heures plus tard. Laprésence de la mère semble donc modérer, aussichez cette espèce, la réponse hormonale de saprogéniture àun stress.Deplus, les jeunes sevrésprésentaient toujours de fortes concentrationsde cortisol sérique, qui auraient peut être pu êtremodérées en présence d’une figure sociale. Eneffet chez certains rongeurs, le lien mère-enfantpersiste au-delà du sevrage (e.g. Newberry etSwanson 2001). Chez le cochon d’Inde, la pré-sence de la mère pendant une exposition brève àun nouvel environnement persiste à réduire leniveau de cortisol de son jeune plusieurs semai-nesau-delàde l’âge typiquedusevrage (Hennessyet al. 1995).
Figure 5. Comparaison des concentrations decortisol sérique à t0 (A) et t2.5 (B) entre les troisgroupes de jeunes phoques CDA, CDS et JSS àl’aided’uneANOVAàun facteur suiviepardes testspost-hoc deFisher.Des lettres différentesmontrentune différence significative entre les groupes alorsque des lettres identiques indiquent qu’il n’y a pasde différence significative.
GRP : fyssen JOB : annales27 DIV : 08⊕mp⊕DI⊕POI⊕Carole p. 7 folio : 68 --- 18/2/013 --- 16H23
ETHOLOGIE / PSYCHOLOGIE
68

Les réponses comportementaleset hormonales sont-elles couplées ?
Nos résultats tendent à montrer une dissocia-tion entre la réponse comportementale et laréponse hormonale. De forts niveaux de vocali-sations et d’agitation avaient pu être observéschez les jeunes capturés avec leur mère maisbrièvement séparés pendant les prélèvementsalors qu’une faible réponse glucocorticoïde avaitété mesurée chez ces mêmes individus. Àl’inverse, une inhibition générale des comporte-ments était reliée à de fortes concentrations decortisol chez les autres groupes de jeunes séparésde la figure maternelle depuis bien plus long-temps. Cette dissociation a déjà été mise enévidence chez les primates et les rongeurs (e.g.Hennessy etMoorman1989 ;Wiener et al.1990).Chez ces espèces, le nombre de vocalisationsdiminue graduellement au fur et à mesure que laséparation se prolonge alors que le niveau decortisol s’élève. Cette dissociation se produitenviron 60 à 90 min après la séparation chez lecobaye (Hennessy et Moorman 1989). De nom-breux auteurs se sont interrogés sur le rôle decertaines hormones de l’axe HHS sur la régula-tion des comportements. L’administration péri-phérique de CRH (hormone hypothalamique cor-ticotrope) auneffet suppresseur sur lenombredevocalisations chez le cochon d’inde placé dans unnouvel environnement au coursdes 30premièresminutes après la séparation de la mère (Hen-nessy et al. 1995) et réduit également l’activitélocomotrice immédiatement après la séparation(Hennessy et al. 1995). Des concentrations éle-vées de cortisol observées chez de jeunes indivi-dus à la suite d’un événement stressant dansl’ensemble de ces études, y compris la notre,reflète vraisemblablement une augmentationdesniveaux de CRH en amont et contribueraient à laréduction générale des comportements au coursd’une isolation prolongée. D’un point de vueécologique, l’agitation et l’augmentation du nom-bre de vocalisations qui permettent au chiot et àsa mère de se retrouver requiert une dépenseénergétique importante pour les animaux quisont à un âge critique. De plus, la détresse qu’ilsexpérimentent lorsqu’ils sont seuls pourrait atti-rer les prédateurs. En conséquence, l’inhibitiondes comportements de détresse semble être uneréponse d’évitement des prédateurs efficacesdans la nature.
ConclusionsCette étude montre pour la première fois un
exemple d’amortissement social chez une espècedemammifèresmarins. Notre résultats ontmon-tré à la fois l’amortissement de la réponse phy-siologique au stress chez des chiots en présencede leur mère ainsi que de forts niveaux devocalisations et d’agitation chez ces même jeu-nes après une brève séparation maternelle. Al’inverse, nous avons observé une inhibition com-portementale chez des chiots exposés à une iso-lation prolongée pouvant être régulée au niveauhormonal par la forte réponse glucocorticoïdemesurée. Ce type d’étude est de première impor-tance lorsqu’il s’agit d’espèces en déclin et/ouvulnérables comme le phoque commun. Le suc-cès de la gestion des populations et de la conser-vation des espèces peuvent être améliorées si lesconditions sociales, biologiques et écologiques del’espèce en question sont respectées et si lesperturbations anthropogéniques sont minimi-sées. Nos résultats remettent en questionl’impact et le choix de telles techniques de recher-che pour l’étude des animaux dans leur environ-nement naturel ; et pourraient servir de supportà l’avenir, pour l’utilisation de pratiques plusjudicieuses afin de réduire efficacement l’anxiétédes animaux lorsque cela est possible.
RemerciementsJe tiens tout d’abord à remercier la Fondation
Fyssen qui a financé ce projet de recherche. Jesuis également reconnaissante au Pr. ShannonAtkinson qui m’a accueillie au sein de son labo-ratoire à l’University of Alaska Fairbanks ainsiqu’à mes collaboratrices Gail Blundell (ADF&G)et Ann Hoover-Miller (ASLC). Je remercie égale-ment chaleureusement tous les volontaires pré-sents pour l’observation des phoques et l’assis-tance sur le terrain.
Bibliographiee Blundell G.M., Womble J.N., Pendleton
G.W., Karpovich S.A., Gende S.M., HerremanJ.K. 2011. Use of glacial and terrestrial habitatsby harbor seals in Glacier Bay, Alaska: costsand benefits. Marine Ecology Progress Series429:277-290.
e Boness D.J., Bowen W.D. 1996. The evolu-tion of maternal care in Pinnipeds. New findingsraise questions about the evolution of maternalfeeding strategies. BioScience 46:645-654.
GRP : fyssen JOB : annales27 DIV : 08⊕mp⊕DI⊕POI⊕Carole p. 8 folio : 69 --- 18/2/013 --- 16H23
ETHOLOGIE / PSYCHOLOGIE
69

e Bowlby J. 1979. Themaking and breaking ofaffectional bonds. Tavistock Publications, Lon-don, UK, 184 pp.
e DeVries A.C., Glasper E.R., Detillion C.E.2003.Socialmodulationof stress responses.Phy-siology & Behavior 79:399-407.
e Engelhard G.H., Brasseur S.M.J.H., HallA.J., Burton H.R., Reijnders P.J.H. 2002. Adre-nocortical responsiveness in southern elephantseal mothers and pups during lactation and theeffect of scientific handling. Journal of Compara-tive Physiology B 172:315-328.
e Gubernick D.J. 1981. Parental and infantattachment in mammals. In: Gubernick D.J. andKlopfer P.H. (Ed), Parental care in mammals.Plenum Press Inc., New York, pp.243-305.
e Hare S.R., Mantua N.J. 2000. Empiricalevidence for North Pacific regime shifts in1977 and 1989. Progress in Oceanography47:103-145.
e HennessyM.B. 1997. Hypothalamic-pituita-ry-adrenal responses to brief social separation.Neuroscience and Biobehavioral Reviews21:11-29.
e Hennessy M.B., Kaiser S., Sachser N. 2009.Social buffering of the stress response:Diversity,mechanisms, and functions. Frontiers in Neu-roendocrinology 30:470-482.
e Hennessy M.B., Moorman L. 1989. Factorsinfluencing cortisol and behavioral responses tomaternal separation in guinea pig. BehavioralNeuroscience 103:378-385.
e HennessyM.B., Nigh C.K., SimsM.L., LongS.J. 1995. Plasma cortisol and vocalization res-ponses of postweaning age guinea pigs to mater-nal and sibling separation: evidence for filialattachment after weaning. Developmental Psy-chobiology 28:103-115.
e vonHolst D. 1998. The concept of stress andits relevance for animal behavior. Advances inthe Study of Behavior 27:1-131.
e Hoover-Miller A.A. 1994. Harbor seals(Phoca vitulina): Biology and Management inAlaska. Report to the Marine Mammal Commis-sion.ContractNumberT75134749.Washington,D.C.45 pp.
e Hoover-Miller A. A., Atkinson S., Conlon S.,Prewitt J., Armato P. 2011. Persistent decline inabundance of harbor seals Phoca vitulinarichardsi over three decades in Aialik Bay, an
Alaskan tidewater glacial fjord. Marine EcologyProgress Series 424:259-271.
e Lay D.C.Jr, Friend T.H., Bowers C.L., Gris-som K.K., Jenkins O.C. 1992a. A comparativephysiological and behavioral study of freeze andhot-iron branding using dairy cows. Journal ofAnimal Science 70: 1121-1125.
e Lay D.C.Jr, Friend T.H., Grissom K.K.,Bowers C.L.,MalM.E. 1992b. Effects of freeze orhot-iron branding of angus calves on some phy-siological and behavioral indicators of stress.Ap-plied Animal Behaviour Science 33: 137-147.
e Lay D.C.Jr, Friend T.H., Randel R.D.,Bowers C.L., Grissom K.K., Jenkins O.C. 1992c.Behavioral and physiological effects of freeze- orhot-iron branding on crossbred cattle. Journal ofAnimal Science 70: 330-336.
e Lawson J.W., Renouf D. 1987. Bonding andWeaning in Harbor Seals, Phoca vitulina. Jour-nal of Mammalogy 68:445-449.
e Mathews E.A., Pendleton G.W. 2006. Decli-nes in harbor seal (Phoca vitulina) numbers inGlacier Bay National Park, Alaska, 1992-2002.Marine Mammal Science 22:167-189.
e Mellish J.-A.E., Iverson S.J., Bowen W.D.1999. Individual variation in maternal energyallocation and milk production in grey seals andconsequences for pup growth and weaning cha-racteristics. Physiol. Biochem. Zool. 72: 677–690.
e Mendoza S.P., Smotherman W.P., MinerM.T., Kaplan J., Levine S. 1978. Pituitary-adre-nal response to separation in mother and infantsquirrelmonkeys. Developmental Psychobiology11:169-175.
e Mineka S., Suomi S.J. 1978. Social separa-tion in monkeys. Psychological Bulletin 85:1376-1400.
e MuelbertM.M.,BowenW.D.1993.Durationof lactation and postweaning changes in massand body composition of harbour seal, Phocavitulina, pups. Canadian Journal of Zoology 71:1405-1414.
e Myers M., Litz B., Atkinson S. 2010. Theeffects of age, season and geographic region oncirculating serum cortisol concentrations inthreatened and endangered Steller sea lions(Eumetopias jubatus). General and ComparativeEndocrinology 165:72-77.
GRP : fyssen JOB : annales27 DIV : 08⊕mp⊕DI⊕POI⊕Carole p. 9 folio : 70 --- 18/2/013 --- 16H23
ETHOLOGIE / PSYCHOLOGIE
70

e Newberry R.C., Swanson J.C. 2001. Brea-king social bonds. In: Keeling L.J., GonyouH.W.(Eds), Social Behaviour in Farm Animals. CABIPublishing, Wallingford UK, pp. 307-331.
e Oftedal O.T. 1985. Pregnancy and lactation.In:Hudson R.J. and White R.G. (Eds), Bioener-getics of wild herbivores. CRC Press, Inc., BocaRaton, Fla. pp. 215-238.
e Petrauskas L., Atkinson S., Gulland F., Mel-lish J.A., HorningM. 2008. Monitoring glucocor-ticoid response to rehabilitation and researchprocedures in Californiaand Steller sea lions.Journal of Experimental Zoology 309A:73-82.
e Sapolsky R.M. 2002. Endocrinology of thestress-response. In: Sapolsky, R.M. (Ed), Endo-crinology of the Stress-Response. MassachusettsInstitute of Technology, Cambridge, Massachu-setts, pp.409-450.
e Trillmich F. 1996. Parental investment inPinnnipeds. Advances in the Study of Behavior25:533-577.
Harbor seal population declines in Alaska
Harbor seal (Phoca vitulina richardsi) popula-tionshaveexperienced importantdemographicalfluctuations over the last 30 years in the easternPacific, and particularly in the Gulf of Alaska-(Hoover-Miller et al. 2011). According to scien-tists, the number of harbor seals declined preci-pitously from 7 to 67%, according to the year andthe region, from the time when counts weremade during the 70s/80s (review in Hoover-Miller et al. 2011). The proximal causes of thesefluctuations are still uncertain. Nevertheless,these declines coincided with ecological changesobservedafter the1976 to1977shift in thePacificDecadal Oscillation (PDO), which induced varia-tions of the North Pacific sea surface tempera-ture (Hare and Mantua 2000). Although the alte-rationof sea ice conditionshasbeenassociated tothe PDO shift, the long term climate changesrelated to global warming have also an impact onice melting and might explain harbor seal decli-nes observed in some regions. In SoutheastAlaska, Glacier Bay National Parkand other gla-cial fjords such as Tracy and Endicott Armsare important pupping sites for harbor seal(Mathews and Pendleton 2006). Many sealsmigrate seasonally to tidewater glacial fjordsto rest, molt, breed and nurse pups (figure 1).
Indeed, the ice floes may provide safe floatingplatforms for the lactatingmother seals and theiroffspring who can nurse uninterrupted (Blundellet al. 2011), and better attend to their pup duringthesummernursingperiod (MuelbertandBowen1993). However, harbor seals that use glacial iceexperience loss of summer habitat as tidewaterglaciers retreat onshore.
Harbor seal census and samplingcampaigns
In order to improve the understanding of har-bor seal population dynamics and to examine thehealth andconditionof thesepopulations, censusand sampling campaigns are regularly conductedduring the molt at the end of the summer in theGulf of Alaska. However, relatively few locationshave been studied during pupping season (Hoo-ver-Miller et al. 2011). Several studies haveshown that such research activities, and theirrelated perturbations (e.g. noise, capture, capti-vity handling, tissue sampling), are stressful forthe animals in several species (e.g. Engelhardet al. 2002; Petrauskas et al. 2008). Thus, it isimportant to understand the effects on both phy-siology and behavior; to date, the publications onthe subject are still rare. A concern is periodicallyraised for pups and lactating mothers. Indeed,lactation is among the most expensive energeticdemands female mammals experience (e.g. Ofte-dal 1985) and offspring depend on investmentand physical condition of their mother to grow(e.g. Mellish et al. 1999). Thus, it also is a timewhen animals are more vulnerable to humandisturbances.
Figure 1. Harbor seal mother and her pup on afloating ice plateform inAlaska (photos ©C. Di Poi).
GRP : fyssen JOB : annales27 DIV : 08⊕mp⊕DI⊕POI⊕Carole p. 10 folio : 71 --- 18/2/013 --- 16H23
ETHOLOGY / PSYCHOLOGY
71

Social buffering on stress responseWhen confronted to a stressful stimulus, the
organism will flight, when possible, or fight. Theneuroendocrine stress response is adaptativeand universal in all Vertebrates and involvesthe activation of hypothalamic-pituitary-adrenal(HPA) axis resulting in the elevation of the hor-mone concentrations, specifically corticotrophicreleasing factor (CRF), adrenocortictrophic hor-mone (ACTH) and glucocorticoids (Sapolsky2002). Circulating glucocorticoids, such as corti-sol, are typicallyused to evaluate stress inmarinemammals, including pinnipeds (e.g. Myers etal. 2010). The amplitude of the response can bemodulated by social context.When a strong emo-tional attachment exists between infants andmothers, separation from the other member ofthe dyad can enhance the HPA axis during athreatening situation. On the contrary, the pre-sence of the partner can reduce HPA responsesand hence serves to buffer the individual againstthe stressful stimulus. The ability of a socialpartner to reduce stress responses, both physio-logical and behavioral responses, is commonlyreferred to «social buffering» (e.g. DeVries etal. 2003; Hennessy et al. 2009). The buffering ismore or less important depending upon thenature and the quality of bonds that the dyaddevelops, as well as the time and the duration ofthe separation. In the context of mother-youngrelationship, the strength of bonds during nur-sing that a mother develops with her dependentoffspringdependsupon the social structureof thespecies and the degree of offspring mobility.According to the theory of Gubernick (1981), theprobability of bonding is greater when the youngis single, precocial, and highly mobile and thesocial structure of the group is gregarious, atleast during the reproductive period.
The mother-pup bond in harbor sealIn pinnipeds, there are two maternal strate-
gies often referred to ’’capital breeding’’ in largerPhocidsand ’’incomebreeding» inOtarrids (Trill-mich 1996). In Phocids after a prolonged periodat sea, the mothers follow a fasting strategy andnurse uninterrupted the pups during a relativelyshort lactation period (from 4 days to 4 months)until weaning. They fuel their needs and feedtheir pupentirely out of energy reserves stored inthe body. This maternal strategy corresponds toextreme capital breeding. In contrast, otariids
adopt an income strategy and feed intermittentlyover a prolonged lactation period (from4monthsto 3 years). The mothers alternate between fas-ting ashore while suckling the pups and foragingat sea away from them. Unlike females of thelarger Phocid species, the female harbor sealshave an alternative strategy. As they are smaller,they are unable to store enough energy and fastlong enough to nurse properly their pup; thus,they forage opportunistically at sea during lac-tation (Boness and Bowen 1996). However,because their nursing period is extremely brief(3 to 6 weeks on average; Muelbert and Bowen1993; Hoover-Miller 1994), it requires that themother and her pupmust remain in contact witheach other as much as possible. There is a bilate-ral bond between mother and pup during theshort nursing period with both members havinga role in keeping the pair together; the pup bycontinually following hismother, and themothercan reduce the risk to become separated from hispup by assuming overt attention for it particu-larly while swimming and during periods of dis-turbance (Lawson andRenouf 1987).Within thiscontext of strong attachment during nursing, theharbor seal is a good candidate for studying theeffects of maternal separation on the stress res-ponse of pups.
Objective of the studyThis study aimed to determine whether the
stress response of pups, following a capture-handling-restraint protocol, might be modulatedby the presence or the absence of the mother infree-ranging Pacific harbor seal Phoca vitulinarichardsi. Three categories of pups were studied:dependent pups captured with their mother butbriefly separated during blood sampling, depen-dent pups captured without their mother, andpups, newly weaned, then captured alone. Weassessed the stress response of pups by a beha-vioral approach evaluating the number of vocali-zations and the agitation level, and by a physio-logical approach evaluating theHPAaxis activityby the quantification of serum cortisol.
ExperimentsThis study followed the National Oceanic and
Atmospheric Administration (NOAA)/NationalMarine Fisheries Service (NMFS)Research Per-mit #358-1757, and Care and Use Committeedu Department of Fish and Game’s en Alaska
GRP : fyssen JOB : annales27 DIV : 08⊕mp⊕DI⊕POI⊕Carole p. 11 folio : 72 --- 18/2/013 --- 16H23
ETHOLOGY / PSYCHOLOGY
72

(ADF&G) permit, Protocol #09-08. All captureswere carried out without causing unacceptablelevel of disturbance of the colonies of seals.
The two sampling sitesThe study was carried out during the summer
pupping season in Endicott Arm and Tracy Arm,two glacial fjords about 60 kmSouth of Juneau inSoutheast Alaska, USA (figure 2).
Seal capture procedureSeals were captured with monofilament nets
(30 m x 3 m) during the light phase between8:30 am and 4:00 pm (figure 3). All mother-puppairs and weaned pups alone were transferredto the laboratory boat and then processed. Thedependent pups captured without their motherwere immediately processed in a transport skiffandthenreleasedat theexact locationwhere theywere captured.
During the field effort, 41 animals were captu-red for this study including 5 dependent pupscaptured with their mother (Dependant Pupswith Mothers) but briefly separated from eachother during blood sampling, 29 dependent pupscaptured without their mother (Dependant PupsAlone) and 7 weaned pups captured alone (Wea-ned Pups Alone). Weaning of Northeast Pacificharbor seal is generally regarded as completeabout 1 month after birth (Muelbert and Bowen1993). In our study, pups were considered as«weaned» if no female was sighted with the pupduring capture and if their body mass was above25 kg.
Behavioral observationsThe behavior of the three categories of pups
was monitored during the first blood collection,between the time that the pups were physicallyrestrained to theendof thebloodsampling (mean
Figure 2. Map of the two sampling sites (provided by ADFG; pictures ©C. Di Poi).
GRP : fyssen JOB : annales27 DIV : 08⊕mp⊕DI⊕POI⊕Carole p. 12 folio : 73 --- 18/2/013 --- 16H23
ETHOLOGY / PSYCHOLOGY
73

duration:3.9±0.3min) inorder toevaluate initialstress responsesof theanimals.Foreachpup, thenumber of vocalizations per minute was measu-red and the level of agitation estimated on a scalefrom 1 to 3 with 1 = quiet, no movement; 2 = lowagitation with movement of the seal’s head, and3 = highly agitated, the seal was hard to restrain.
Blood sampling procedure and cortisolquantification
All the other animals were restrained alone ina box or held far away enough from other seals toavoid a physical contact between them. Mothersand pups captured together were handledtogether until blood sampling. Before proces-sing, all animals were weighed in order to deter-mine the weight and the volume of blood thatcould safely be collected. For blood collection,animals were immobilized by physical restraintwithout anaesthesia administration. Blood wascollected fromtheextradural intravertebral vein.Mothers and their pup were always processedfirst and sampled at the same time; however, theywere briefly separated during sampling and thenre-united after it. DPA were sampled in the on-site sampling skiff within 16 - 213 min aftercapture (mean time: 69.2 ± 8.9 min) and thenreleased at the location where they were captu-red. For the other seals, the total interval of timebetween capture and blood collection rangedfrom103 to255min (mean time: 164.6±14.0minforDPMand171.7±19.3min forWPA).The timeatwhichbloodbegan entering the vacutainerwasnoted and ranged between 10:04 am and 4:13 pm
for dependant pups alone and between 11:03 amand 2:43 pm for the other young seals. After thesample collection procedures, sex and bodymasswere determined, and dependant pups alonereleased at the exact location where they werecaptured to allow them to reunite with theirmother. All the other seals were allowed to reco-ver for 2.5 hours in their individual cage (alone orin pairs for mothers and pups) before a secondblood collection trial. Finally all the seals werereleased; the mother-pup pairs were releasedtogether. Cortisol concentrations were measu-red at t0 and t2.5 in serum with 125� solid-phaseradioimmunoassays.
Results
Parameters influencing hormoneconcentrations
Neither the time interval between the captureand blood collection (Linear regression analysis,R2 = 0.0981; P = 0.256), nor the time of bloodcollection during the day (Linear regression ana-lysis, R2 = 0.0892; P = 0.182) was related tocortisol concentrations. Moreover, there was noinfluence of body mass (Linear regression analy-sis, R2 = 2.91.10-5, P = 0.974) or gender of pups(unpaired t-test, t= -0.089,P=0.930)onhormonalconcentrations.
Behavioral responses of pupsThe number of vocalizations did not differ
significantly between the three groups of pups(Kruskal-Wallis, P = 0.056, n = 37; figure 4A).However, the level of vocalization tended to behigher for pups which were accompanied of theirmother but briefly separated during samplingthan for pups alone, either dependent or weanedpups, since capture. Likewise, the level of agita-tion also did not significantly differ betweengroups (Kruskal-Wallis, P = 0.0973, n = 37;figure 4B); but the level of agitation tended to behigher for pups recently separated from theirmother.Overall, vocalization andagitation levelsduring blood sampling decreased as the durationof separation from the mother increased.
Hormonal responses of pupsAt t0, the levels of cortisol differed between
groups (ANOVA, P = 0.005, n = 41; Fig.3A). Asexpected, the presence of the mother since thebeginning of the stressful event (capture-han-dling-restraint) moderated the cortisol response
Figure 3. Positioning of the monofilament net inEndicott Armand capture skiff (picture ©C. Di Poi).
GRP : fyssen JOB : annales27 DIV : 08⊕mp⊕DI⊕POI⊕Carole p. 13 folio : 74 --- 18/2/013 --- 16H23
ETHOLOGY / PSYCHOLOGY
74

of pups compared to dependent pups andweanedpups alone, both exhibiting higher serum cor-tisol levels (Fisher’s PLSD, P = 0.0011 andP = 0.0186 respectively; figure 5A). At t2.5, thelevels of cortisol did not significantly differedbetween pups accompanied by their motherand weaned pups (unpaired t-test, P = 0.200;figure 5B); although accompanied pups still exhi-bited a lower serum cortisol concentration. Theanalysis of variance for repeated measurementsdid not show a significant evolution of cortisolconcentrations over time for any group (repeatedmeasures ANOVA, P O 0.05, data not shown).
Discussion
The notion of «social buffering» has beenwidely demonstrated in many species exhibitingevident mother-infant bonding (e.g. DeVries etal. 2003; Hennessy et al. 2009). Depending uponthe nature and quality of filial attachment, theseparation from the other member of the dyad
can activate the HPA axis during exposure tothreatening conditions (e.g. Hennessy 1997); orat the opposite, the presence of the social partnercan buffer this response (von Holst 1998).
Behavioral responses
Behavioral measures such as vocalizations,resting ormoving around can be used to evaluatestress response and/or pain in animals (Lay etal. 1992a,b,c). Shortly after separation ensues,the initial acute behavioral response of the infantto separation from his mother has been charac-terized by high levels of vocalization (e.g. Hen-nessy andMoorman 1989) and increased activitylevel (e.g. in primates Mineka and Suomi 1978).With prolonged separation, infants of somemon-key species showageneral inactivity andbegin todisplay a characteristic hunched body posture.These two phases are called the «protest» and the«despair» phases respectively (Bowlby 1979). Inguinea pig, Hennessy and Moorman (1989)
Figure 4. Comparison of vocalization (A) andagitation (B) levels of the three harbor seal pupcategories: dependant pups captured with theirmother but newly separated during observations(DPM), dependant pups alone (DPA) and weanedpups alone (WPA) using Kruskal-Wallis tests.
Figure 5. Comparison of serum cortisol concen-trations at t0 (A) and t2.5 (B) between the threegroups of harbor seal pups DPM, DPA and WPAusing one-way ANOVA followed by Fisher’s posthoc tests. Different letters mean significant diffe-rences between groups and same letters indicateno significant difference.
GRP : fyssen JOB : annales27 DIV : 08⊕mp⊕DI⊕POI⊕Carole p. 14 folio : 75 --- 18/2/013 --- 16H23
ETHOLOGY / PSYCHOLOGY
75

showed that separated pups vocalized more thandid pups tested with their mother during theinitial 30 min post-separation only (protestphase) and then decreased from 60-90 min andbecame at the same level of pups with motherafter 150-180 minutes of separation (despairphase). However, they found that locomotor acti-vity did not differ between pups with the motherand pups alone at any point (from 0 to 180 min).Although an increase of the activity level hasalready been observed in young primates shortlyafter separation (Mineka and Suomi 1978). Inour study, mother-pup pairs were physically andvisually separated few minutes during the firstblood collection. During separation, pups exhibi-ted high levels of both vocalizations and agitationcompared to pups already alone since capture.The separation of pups from theirmother causedthe initial protest response observed in smallmammals, typically composed of a high rate ofseparation-induced vocalizations and high agita-ted level as exposed above, compared to pupsseparated from their mother for a longer time.The weakness of behavioral response of unac-companied dependent pups can be explained bythe fact that about one hour elapsed betweencapture and observations (average time: 69.2 ±8.9 min); this period of time was consistent withthe timepassed after the separation in guinea pig(60-90 min) when the infants enter into the des-pair phase and their vocalization response star-ted to decrease (Hennessy and Moorman 1989).
Hormonal responsesCapture, temporary handling in captivity and
tissue sampling represent disturbances to wildanimals (Engelhard et al. 2002; Petrauskas et al.2008). Moreover in rodents, infants experience asignificant activation of the HPA axis when sepa-rated from their mother and exposed to a stres-sful event (e.g. novel environment exposure)that ended with the increase of both plasmaACTH and cortisol reaching a maximum level90 minutes following separation (e.g. Hennessyand Moorman 1989). In harbor seal, our resultsshowed that pups accompanied by their mothersince capture exhibited significantly lower serumcortisol concentrations than pups alone, eitherstill dependent or recentlyweaned; this tendencywas maintained 2.5 hours later. In this species,the presence of the mother from the beginning
of the procedure also seemed to moderate hor-monal response of their offspring to a stressor.Moreover, single weaned harbor seals exhibitedhigh level of serum cortisol compared to pre-weaned pups; which would have been likely buf-fered in presence of a social figure. Indeed insome rodents, the mother-infant relationshiplasts after weaning (e.g. Newberry and Swanson2001). As an example in guinea pig, the presenceof the mother during brief exposure to a novelenvironment continues to reduce the cortisolresponse of their young for several weeks beyondthe typical age of weaning (Hennessy et al. 1995).
Are behavioral and hormonal responsescoupled?
Our results tended to show a dissociationbetween behavioral response and hormonal res-ponse. We observed high levels of vocalizationsand agitation for pups exposed to a brief mater-nal separation during sampling but they showedthe lowest glucocorticoid response; while pupsseparated from their mother from a longer timeexhibited a general inhibition of their behaviorsand higher cortisol concentrations. Severalauthors have previously reported this dissocia-tion in primates and rodents; the number ofvocalizations decreasing as the separation pro-ceeds rather quickly and cortisol level conti-nuing to climb. This dissociation occurs at about60-90 min post-separation in guinea pig (Hen-nessy and Moorman 1989). The possibility thatHPA hormones influence the behavioral res-ponse to separation has received growing atten-tion. Peripheral administration of CRF (cortico-trophin-releasing factor) has a suppressive effecton the level of vocalizations of guinea pig pupsduring the first 30-min period of isolation in anovel environment (e.g. Hennessy et al. 1995)and reduces the locomotor activity immediatelyfollowing separation (e.g. Hennessy et al. 1995).As a consequence, high elevation of plasma cor-tisol observed following a stressful event in thesestudies, including our study, likely reflects anincreased level of peripheral CRH and contribu-tes to the waning of the vocalizations duringprolonged isolation. From a behavioral ecologyperspective, vocalizing and agitation increasingto reunite with the mother require energy expen-diture for pupswhich are at a vulnerable age, andthe distress they experienced when alone may
GRP : fyssen JOB : annales27 DIV : 08⊕mp⊕DI⊕POI⊕Carole p. 15 folio : 76 --- 18/2/013 --- 16H23
ETHOLOGY / PSYCHOLOGY
76

attract predators in the wild. As a consequence,the inhibition of distress responses might berelevant predator avoidance behavior patterns innature.
ConclusionsThe current studyprovides for thefirst timean
example of social buffering in a marine mammalspecies. Our results showed at the same time thebuffering of physiological stress response of pupsaccompanied of their mother as well as highlevels of both vocalizations and agitation whenbriefly separated from them. On the contrary, weobserved a behavioral inhibition in pups expe-rienced a prolonged isolation that might be hor-mone-related by the high glucocorticoid levelsmeasured. The results that stem from the studywill be particularly useful and valuable in decli-ning or vulnerable population species as is the
harbor seal. The success of management andconservation of a species may be improved ifsocial, biological and ecological conditions ofeach species are respected and if anthropogenicstress is minimized. Our results raise questionson the impactand thechoiceof researchpracticesandwill givenew insights to adoptmore judiciousmanagement techniques in order to effectivelyreduce stress of animals when possible.
AcknowledgmentsIwould like to thank theFyssenFoundation for
its financial support. I am also grateful to Pr.ShannonAtkinson forhostingme inher labat theUniversity of Alaska Fairbanks and to my colla-borators Gail Blundell (ADF&G) and Ann Hoo-ver-Miller (ASLC). I also warmly acknowledge allthe volunteers for their help for seal observationand assistant in the field.
GRP : fyssen JOB : annales27 DIV : 08⊕mp⊕DI⊕POI⊕Carole p. 16 folio : 77 --- 18/2/013 --- 16H23
ETHOLOGY / PSYCHOLOGY
77

L’Imagerie par Résonance Magnétique nous révèlequelques mystères du cerveau du nourrisson
Jessica DUBOIS
Chargée de recherche INSERMUnité INSERM-CEAU992 de «Neuroimagerie Cognitive », NeuroSpin
RésuméL’IRM permet aujourd’hui d’explorer le cerveau immature du bébé de façon non-invasive et
d’envisager des corrélations fines entre développement anatomique du cerveau et acquisitionsprécoces des fonctions cognitives. En exemples, sont décrites deux études récentes qui ont évalué leplissement des sillons du cortex chez le nouveau-né prématuré, et l’organisation des faisceaux defibres de substance blanche chez le nourrisson sain. Dans les deux cas, des différences spatio-temporelles de maturation entre régions cérébrales, des asymétries anatomiques précoces entre lesdeux hémisphères, ainsi que des corrélations anatomo-fonctionnelles ont pu être mises en évidence.
Mots-clésDéveloppement cérébral, maturation, sillons du cortex, faisceaux de fibres de substance blanche,
nouveau-né prématuré, nourrisson, IRM, imagerie du tenseur de diffusion (DTI), asymétries inter-hémisphériques, potentiels évoqués visuels (PEV).
Magnetic resonance imaging reveals some mysteriesof the infant’s brain
AbstractNowadays MRI enables to explore the immature brain of babies non-invasively, and to consider
precise correlations between the anatomical development of the brain and the early acquisitions incognitive functions.As examples,wepresent two recent studieswhichevaluated the foldingof corticalsulci in preterm newborns, and the organization of white matter fiber bundles in healthy infants. Forboth studies, we have highlighted spatio-temporal differences in the maturation of cerebral regions,early anatomical asymmetries between the two hemispheres, as well as anatomo-functional correla-tions.
KeywordsBrain development, maturation, cortical sulci, white matter fiber bundles, preterm newborn,
infant, MRI, diffusion tensor imaging (DTI), inter-hemispherical asymmetries, visual evoked poten-tials (VEP).
IntroductionDiverses pathologies cérébrales résultent
d’anomaliesprécocesaucoursdudéveloppementet touchent une proportion importante d’indivi-dus, comme par exemple l’épilepsie, le retardmental et la dyslexie. Le développement du cer-veau peut également être perturbé par des évè-nements précoces, comme une naissance préma-turée, un retard de croissance intra-utérin ou
un accident vasculaire cérébral néonatal. Pourappréhender la complexité de ces pathologies,pour juger de l’impact de telles perturbations, etaussi pour comprendre comment le cerveau del’homme lui a permis de développer des capacitésspécifiques, il est essentiel d’étudier comment lecerveau normal se développe tant au plan anato-mique que fonctionnel. Les mécanismes sous-jacents sont multiples, et les changements céré-
GRP : fyssen JOB : annales27Internet DIV : 09mp⊕DUBOIS⊕Jessica p. 1 folio : 78 --- 14/5/013 --- 15H1
NEUROBIOLOGIE
78

braux sontparticulièrement intensespendant lesdernières semaines de grossesse et les premiersmois post-natals. Le cerveau du bébé n’est pas uncerveau adulte en miniature : sa croissance estinhomogènepuisque certaines régions, telles queles régions sensorielles, se développent précoce-ment et très rapidement, alors que d’autresrégions associatives, telles que les régions fron-tales, ont un développement plus retardé et pro-longé jusqu’à la fin de l’adolescence (Paus etal. 2001). Bien avant l’observation d’une spécia-lisation fonctionnelle hémisphérique, des asymé-tries anatomiques entre hémisphères cérébrauxsontobservées chez le fœtus (Chi et al. 1977b).Enparallèle à cette évolution anatomique, les acqui-sitions psychomotrices et cognitives sont pro-gressivement observées chez le nouveau-né, lenourrisson et l’enfant.
Saisir comment ces acquisitions se déroulenten regard de lamaturation cérébrale requiert desétudes in vivo. Les investigations post mortemsont intrinsèquement limitées car il est rare depouvoir garantir la normalité des cerveaux étu-diés et impossible d’établir des corrélations ana-tomo-fonctionnelles.Mêmesi elles permettent detester certaines hypothèses, les études chez l’ani-mal sont insuffisantes compte tenu de la spécifi-cité du cerveau humain. Les techniques d’ima-gerie non invasives (imagerie par résonancemagnétique IRM, électroencéphalographie EEG,magnétoencéphalographie MEG) présententdans ce contexte un potentiel particulièrementimportant, puisqu’elles permettent la mise enrelation de la maturation des structures cérébra-les et l’émergence d’un nouveau comportementchez le nourrisson sain, ainsi qu’un suivi longi-tudinal des enfants à risque pour évaluer notam-ment la plasticité post-lésionnelle.
Cependant, on se trouve confronté à de nom-breuses difficultés pour faire de l’imagerie chez lebébé. En effet, contrairement au cerveau adultequi se modifie relativement peu de vingt àsoixante ans, le cerveau du nourrisson est encontinuel changement. Les structures sont depetite taille, donc la résolution spatiale doit êtreplus élevée que chez l’adulte, et les tissus sontimmatures, ce qui implique d’adapter l’acqui-sition et le post-traitement des données aux dif-férents âges. Par ailleurs, ces techniques d’ima-gerie sont très sensibles au mouvement,l’acquisition des données doit donc être brève et
sans aucune gêne pour garantir le sommeil ou lacoopération du nourrisson.
Grâce à diverses implémentations méthodolo-giques récentes, nous avons mis en place deuxétudes complémentaires visant à éclairer le déve-loppement anatomique du cerveau humain invivo, et à caractériser de façon quantitative leschangements qui s’y déroulent au cours des pre-miers mois post-natals. La première étude s’estfocalisée sur la formation précoce des sillons ducortex chez des nouveau-nés prématurés imagésen IRM conventionnelle à la naissance, entre sixet huitmois d’âge gestationnel. La seconde études’est intéresséeà l’organisationetà lamaturationultérieures de la substance blanche, évaluées parIRM du tenseur de diffusion (DTI) chez le nour-risson sain de un à quatre mois d’âge post-natal.Chaque étude a bien sûr été réalisée avec l’accorddesparentsetducomitéd’éthique.Plusieursaxesde recherche ont été investigués, notamment lamise en évidence de différences spatio-temporel-les dematuration entre régions cérébrales, d’asy-métries précoces entre hémisphères cérébraux,et de corrélations entre changements anatomi-ques et maturation fonctionnelle.
Formation des sillons du cortexchez le nouveau-né prématuré
Le cortex se développe, s’épaissit et se plisseprincipalement au cours du dernier trimestre degrossesse. En plus de son importante croissanceen taille, lamorphologie du cerveau fœtal devientalors rapidement élaborée avec la formation desgyri et des sillons visibles macroscopiquement.La complexité de ces plis est croissante avecl’âge (Chi et al. 1977a ; Feess-Higgins and Laro-che 1987), et le cerveau adulte présente unegrande variabilité inter-individuelle de sesmotifs morphologiques (Ono et al. 1990). Plu-sieurs hypothèses sont aujourd’hui avancées surles mécanismes de la gyration (facteurs généti-ques, épi-génétiques, mécaniques ou environne-mentaux) (Welker 1990).
Pour explorer in vivo les stades précoces dudéveloppement cortical et établir des normeschronologiques basées sur des marqueurs quan-titatifs du plissement des sillons, des nouveau-nés prématurés, de 6.2 à 8.3 mois d’âge ges-tationnel, ont été étudiés ex utero par IRMconventionnelle et avec de nouveaux outils detraitement d’images dédiés au cerveau immature(Dubois et al. 2008c). A partir d’images pondé-
GRP : fyssen JOB : annales27Internet DIV : 09mp⊕DUBOIS⊕Jessica p. 2 folio : 79 --- 14/5/013 --- 15H1
NEUROBIOLOGIE
79

rées en T2, acquises sur un IRM à 1.5T avec unehaute résolution spatiale (Figure 1a), nous avonspu reconstruire en trois dimensions la surfaceinterne du cortex, à l’interface avec la substanceblanche, sur un intervalle d’âges correspondant
au dernier trimestre de grossesse (Figure 1b).Les sillons se plissent alors de façon spectacu-laire, et un indice de sulcation quantitatif a étémesuré (Figure 1c).
Figure 1Imagerie IRM des sillons du cortex du nouveau-né prématuréa : Images T2 obtenues chez un nouveau-né prématuré de 31 semaines d’âge gestationnel.b : Reconstructions en 3 dimensions de la surface corticale interne de 3 nouveau-nés prématurés d’âges etindices de sulcation croissants. La couleur représente la courbure de la surface. D’après (Dubois et al. 2008c).c : Augmentation de l’indice de sulcation avec l’âge des nouveau-nés. D’après (Dubois et al. 2008c).d : Asymétries inter-hemisphériques. La surface corticale présente des différences de morphologie entre lesdeux hémisphères cérébraux au niveau du sillon temporal supérieur (STS) et de la scissure sylvienne, enantérieur (en regard de la région de Broca) et postérieur (en regard du planum temporale). D’après (Dubois etal. 2010).e : Différences de sulcation en fonction de l’environnement intra-utérin. A âge gestationnel équivalent, lejumeauprésenteun indicedesulcationréduit encomparaisondusingleton (imagesdegauche), selonunretardde maturation harmonieux. A surface corticale équivalente (mais à âge différent), le nouveau-né avec retardde croissance intra-utérin présente un indice de sulcation trop important par rapport au singleton (images dedroite) suggérant un développement disharmonieux. D’après (Dubois et al. 2008d).
GRP : fyssen JOB : annales27Internet DIV : 09mp⊕DUBOIS⊕Jessica p. 3 folio : 80 --- 14/5/013 --- 15H5
NEUROBIOLOGIE
80

En accord avec des atlas postmortem (Chi et al.1977a ; Feess-Higgins and Laroche 1987) et pré-natal par IRM (Garel et al. 2001), l’apparitionprogressive des sillons primaires, secondairespuis tertiaires a été mise en évidence, selon unschéma asynchrone en fonction des régions céré-brales : la surface médiane (sillons calleux et cin-gulaire, scissures pariéto-occipitale et calcarineobservés chez tous les nouveau-nés) se plisseavant la surface latérale, où la différentiationmorphologique des sillons commence dans larégion centrale (sillon central observé chez tous),etprogressedans les lobespariétal (identificationdes sillons post-central, intra-pariétal et pariéto-occipital entre27et31semaines), temporal (iden-tification des sillons temporaux supérieur et infé-rieur entre 27 et 32 semaines, du sillon collatéralentre 29 et 30 semaines) puis frontal (identifica-tion des sillons pré-central, frontaux supérieur etinférieur entre 29 et 30 semaines, identificationdu sillon orbitaire vers 30-31 semaines, et dusillon olfactif à partir de 32 semaines).
Organisation et maturation des faisceauxde fibres de substance blanche dunourrisson
Parallèlement au développement du cortex, lasubstance blanche sous-jacente s’organise etdevient mature. Jusqu’à récemment, l’essentielde nos connaissances sur la myélinisation desfibres, phénomène qui permet d’augmenter lavitesse de conduction de l’influx nerveux (Bau-mann and Pham-Dinh 2001), se basait sur desétudes postmortem (Yakovlev andLecours 1967),laissant de nombreuses questions en suspenscomme les implications fonctionnelles de cettemyélinisation chez le bébé. Depuis quelquesannées, l’IRM conventionnelle permet d’évaluerqualitativement la maturation de la substanceblanche in vivo (Barkovich 2000 ; Paus etal. 2001), en observant les diminutions des pon-dérations T1 et T2 contemporaines respective-ment de la formation de « pré-myéline » par lesoligodendrocytes et de l’enroulement des gainesautour des axones (myélinisation « vraie »).Aujourd’hui, il est possible dequantifier despara-mètres liés à ces mécanismes par imagerie DTI(Neil et al. 2002 ; Huppi and Dubois 2006), carcette technique permet l’exploration non-inva-sive de lamicrostructure tissulaire par lamesuredes mouvements des molécules d’eau (Le Bihanet al. 2001).
Pour étudier l’organisation et quantifier lamaturationde la substanceblanche, etdéfinirdesindices structurels de la maturation fonction-nelle, des nourrissons sains, nés à terme, de 1.3à 4.5 mois d’âge post-natal, ont été imagés avecun protocole DTI dédié au cerveau immature(Figure 2a) sur un IRM à 1.5T (Dubois etal. 2006a). L’hypothèse d’une organisationprécoce de la substance blanche en faisceaux aété validée en reconstruisant par tractographieles principaux faisceaux, même peu myélinisés(Figure 2b), qu’il s’agisse de faisceaux commis-suraux (genou, corps et splénium du corps cal-leux), de projection (faisceaux cortico-spinal etspino-thalamique, radiations optiques et brasantérieur de la capsule interne), limbiques (for-nix et cingulum) ou associatifs (capsule externe,faisceaux unciné, arqué, longitudinaux inférieuret supérieur).
D’importantes variations des indices DTI ontété observées en fonction de l’âge (diminution dela diffusivité moyenne et augmentation de l’ani-sotropie : Figure 2c). Puisque les indicesDTI sontcomplémentaires (Beaulieu 2002), un modèle derelations entre processus de maturation interve-nant au niveau microscopique pendant la myéli-nisation des fibres, et variations des indices DTImesurés à l’échelle du voxel a été proposé pourquantifier les différences de maturation entrefaisceaux de substance blanche (Dubois etal. 2008a). Sur l’intervalle d’âges considéré, lefaisceau cortico-spinal apparaît relativementmature, suivi du faisceau spino-thalamique et dufornix, puis des radiations optiques, des fais-ceaux arqué et longitudinal inférieur, puis dubras antérieur de la capsule interne et du cingu-lum. Ces résultats sont en accord avec les étudesprécédentesdemarquagespost-mortemde lamyé-line (Flechsig1920 ;Yakovlev andLecours1967 ;Brody, Kinney et al. 1987 ; Kinney, Brody et al.1988) et par IRMconventionnelle in vivo (van derKnaap and Valk 1990 ; Paus et al. 2001), qui ontdécrit une progression de la myélinisation desrégions centrales vers la périphérie selon ungradient caudo-rostral, dans les voies sensoriel-les, motrices puis associatives.
Ces deux études IRMont ainsimis en évidencedes différences spatio-temporelles de développe-mententre lesdifférents sillonsducortexet entreles différents faisceaux de substance blanche : àl’instar du développement progressif des capaci-
GRP : fyssen JOB : annales27Internet DIV : 09mp⊕DUBOIS⊕Jessica p. 4 folio : 81 --- 14/5/013 --- 15H5
NEUROBIOLOGIE
81

tés sensori-motrices et cognitives du nourrisson,les régions cérébrales se développent et devien-
nentmatures à des rythmes et sur des intervallesde temps différents.
Figure 2Imagerie DTI de la substance blanche du nourrisson saina : Carte DTI obtenue chez un nourrisson âgé de 7 semaines. Les couleurs codent localement pour la directionprincipale des fibres (rouge : direction droite-gauche ; vert : direction antéro-postérieure ; bleu : directioninféro-supérieure).b : Reconstruction par tractographie de plusieurs faisceaux de fibres chez un nourrisson âgé de 17 semaines.D’après (Dehaene-Lambertz et al. 2006).Abréviations : cc corps calleux ; cg cingulum ; cst faisceau cortico-spinal ; ilf : faisceau longitudinal inférieur ;or radiations optiques ; stt tract spino-thalamique ; unc faisceau unciné.c : Augmentation de l’anisotropie avec l’âge des nourrissons dans différents faisceaux. D’après (Dubois etal. 2008a).d : Asymétries inter-hémisphériques. Une asymétrie de volume du faisceau arqué est observée dans sa partietemporaleen faveurde lagauche (imagedegauche), tandisqu’uneasymétried’organisationmicroscopiqueestobservée dans la par tie pariétale en faveur de la gauche (image du centre). Une asymétrie de maturation dufaisceaucortico-spinal est trouvée dans sa partie inférieure (cp-plic : pédoncules cérébraux – bras postérieur de lacapsule interne) en faveur de la gauche (image de droite). D’après (Dubois et al. 2009).
GRP : fyssen JOB : annales27Internet DIV : 09mp⊕DUBOIS⊕Jessica p. 5 folio : 82 --- 14/5/013 --- 15H6
NEUROBIOLOGIE
82

Asymétries précoces entre hémisphèrescérébraux
En complément de leur complexité anatomi-que, les hémisphères du cerveau adulte sontasymétriques, notamment par le « torque deYakovlev » qui implique les pétalias frontal etoccipital (Toga and Thompson 2003). Certainesdes asymétries structurelles détectées au niveaudes régions péri-sylviennes (gyrus de Heschl,planum temporale et sillon temporal supérieurSTS) ont été décrites au cours du développementprécoce par des études post-mortem de cerveauxde fœtus (Chi et al. 1977b ; Feess-Higgins andLaroche 1987). D’autres asymétries macroscopi-ques évoluent plus tardivement au cours du déve-loppement, comme celle de l’extension posté-rieure de la scissure sylvienne qui progresse del’adolescence à l’âge adulte (Sowell et al. 2002).
Au niveau fonctionnel, la spécialisation hémis-phérique gauche pour le langage est observéeprécocement dans le cerveau du nourrisson(Dehaene-Lambertz et al. 2002 ; Dehaene-Lam-bertz et al. 2006), et la latéralisationde la réponsesomato-sensorielle est également détectée à lanaissance (Erberich et al. 2006). Cependant, lesorigines de ces asymétries anatomiques et fonc-tionnelles, ainsi que leurs relations, restent unsujet à explorer in vivo dans le cerveau en déve-loppement.
Asymétries des sillons des nouveau-nés prématu-rés
Des asymétries précoces ont été démontréesau niveau des surfaces corticales internes desnouveau-nés prématurés, dans trois régions péri-sylviennes du réseau linguistique (Figure 1d) : leSTS, qui se forme plus précocement à droite(Dubois et al. 2008c), et les régions antérieure (enregard de la région de Broca) et postérieure (enregard du planum temporale) de la scissure syl-vienne, régions qui sont plus larges et bombées àgauche (Dubois et al. 2010).
Asymétries des faisceaux de substance blanchedes nourrissons
Trois régions de substance blanche sont éga-lement apparues asymétriques en DTI chez lesnourrissons (Figure 2d) (Dubois et al. 2009) : lesegment temporal du faisceau arqué, plus volu-mineux à gauche, le segment pariétal du faisceauarqué, dont les fibres sont mieux organisées àgauche, et le faisceau cortico-spinal, plus matureàgaucheentre lespédoncules cérébrauxet lebras
postérieur de la capsule interne. Egalement asy-métriques dans le cerveau adulte (Buchel et al.2004 ; Parker et al. 2005), ces faisceaux, qui sontimpliqués l’un dans la perception/production dulangage et l’autre dans la manualité, sont doncasymétriques à un âge où ces deux fonctions sontpeu développées.
Corrélations entre maturation anatomiqueet développement fonctionnel
Desdifférences dedéveloppement anatomiqueentre régions cérébrales et des asymétries inter-hémisphériques précoces ont donc été mise enévidence par ces deux études in vivo. Dans quellemesure les paramètres structurels IRM nousrenseignent-ils également sur le développementpsychomoteur et cognitif des nourrissons ?
Sulcation précoce et suivi longitudinal des nou-veau-nés prématurés
Trois groupes ont été distingués parmi lesnouveau-nés prématurés inclus dans l’étude :1) 27 nouveau-nés sains et issus de grossessesimple ; 2) 12 nouveau-nés sains et issus de gros-sesse gémellaire ; 3) 11 nouveau-nés avec retardde croissance intra-utérin et issus de grossessesimple. Les mesures réalisées à la naissance ontpermis de montrer que l’environnement intra-utérin a un impact majeur sur le développementcérébral (Dubois et al. 2008d). En effet, l’analysequantitative des volumes, surfaces et indices desulcation corticaux a souligné que les jumeauxprésentent un retard dematuration harmonieux,d’environ deux semaines par rapport à leur âgegestationnel, tandis que les nouveau-nés avecretard de croissance intra-utérin présentent unretard disharmonieux, avec des indices de sulca-tion trop importants par rapport à leur faiblesurface corticale (Figure 1e).
De plus, ces mesures corticales à la naissancesont fortement corrélées au développement ulté-rieur des nouveau-nés à l’âge équivalent duterme : les plus grands volumes corticaux et lesmeilleurs scores fonctionnels à terme correspon-dent aux plus grandes surfaces corticales à lanaissance, après correction pour les effets d’âge(Dubois et al. 2008d). A l’avenir, cette métho-dologie devrait permettre de mieux évaluerl’impact, sur le développement du cerveau, dedifférentes stratégies médicales de prise encharge et de soin des nouveau-nés prématurés.
GRP : fyssen JOB : annales27Internet DIV : 09mp⊕DUBOIS⊕Jessica p. 6 folio : 83 --- 14/5/013 --- 15H6
NEUROBIOLOGIE
83

Myélinisation des fibres et vitesse de conductionchez les nourrissons
D’autre part, pour évaluer la valeur fonction-nelle des indices DTI, nous nous sommes focali-sés sur le développement du système visuel.L’EEG, et plus spécifiquement les potentiels évo-qués visuels, sont fréquemment utilisés chez lebébé de par leur facilité d’installation. La latencede la première onde de réponse à un stimulusvisuel (P1) décroit considérablement en fonctionde l’âge, environde250msà lanaissanceà120msvers 4 mois, et ce à cause de divers processusconcomitants (myélinisation des fibres, matura-tion de la rétine et du cortex, etc.) (McCulloch etal. 1999).
A partir de cette latence P1, nous avons estiméla vitesse de conduction de l’influx nerveux dansles voies visuelles, et montré, chez les mêmesnourrissons, que cette vitesse dépend directe-mentdes indicesDTImesurésdans les radiationsoptiques, en plus de l’effet lié à l’âge (Dubois et al.2008b). Cela implique que ces indices structurelssont aussi des marqueurs de la maturation fonc-tionnelle.
ConclusionCaractériser la dynamique du développement
cérébralhumainnormalet lesbasesanatomiquesde la maturation fonctionnelle in vivo pose denombreuses difficultés chez le nourrisson etl’enfant sain, et la mise en œuvre de méthodesd’imagerie et de post-traitements dédiées estnécessaire.Releverde telsdéfisméthodologiqueset proposer de nouveaux marqueurs quantitatifsde la maturation offre l’opportunité d’explorerle cerveau immature sous une perspective nou-velle et transversale, du développement structu-rel du cerveau jusqu’au développement compor-temental du nourrisson, afin de tenter de mieuxcomprendre certaines pathologies du développe-ment et d’aider au diagnostic avec des critèresquantitatifs permettant de prévoir des déficitsfuturs.
RemerciementsCe travail a été financé par la Fondation Fys-
sen, l’Ecole des Neurosciences de Paris, la Fon-dation McDonnell, la Fondation Motrice, l’ANR,le CIBM de Genève et Lausanne, la FondationNationaleSuisse, la Fondation Leenards et leconsortium européen NEOBRAIN. Je remercieparticulièrement L. Hertz-Pannier, G. Dehaene-
Lambertz, D. Le Bihan, J.F. Mangin, P. Hüppi,M. Benders et F. Lazeyras pour m’avoir permisde réaliser ces passionnants travaux.
Bibliographiee Barkovich,A. J. (2000). ”Conceptsofmyelin
andmyelination in neuroradiology.“ AJNRAmJNeuroradiol 21(6): 1099-109.
e Beaulieu, C. (2002). ”The basis of anisotro-pic water diffusion in the nervous system - atechnical review.“ NMR Biomed 15(7-8): 435-55.
e Buchel, C., T.Raedler,M.Sommer,M.Sach,C.Weiller andM. A. Koch (2004). ”Whitematterasymmetry in the human brain: a diffusion ten-sor MRI study.“ Cereb Cortex 14(9): 945-51.
e Chi, J. G., E. C. Dooling and F. H. Gilles(1977a). ”Gyral development of the humanbrain.“ Ann Neurol 1(1): 86-93.
e Chi, J. G., E. C. Dooling and F. H. Gilles(1977b). ”Left-right asymmetries of the temporalspeech areas of the human fetus.“ Arch Neurol34(6): 346-8.
e Dehaene-Lambertz, G., S. Dehaene andL. Hertz-Pannier (2002). ”Functional neuroima-ging of speech perception in infants.“ Science298(5600): 2013-5.
e Dehaene-Lambertz, G., L. Hertz-Pannierand J. Dubois (2006). ”Nature and nurture inlanguage acquisition: anatomical and functionalbrain-imaging studies in infants.“ Trends Neu-rosci 29(7): 367-73.
e Dubois, J., M. Benders, C. Borradori-Tolsa, A. Cachia, F. Lazeyras, R. Ha-Vinh Leuch-ter, S. V. Sizonenko, S. K.Warfield, J. F. Manginand P. S. Huppi (2008d). ”Primary cortical fol-ding in the human newborn: an early marker oflater functional development.“ Brain 131(Pt 8):2028-41.
e Dubois, J., M. Benders, A. Cachia, F. Lazey-ras,R.Ha-VinhLeuchter,S.V.Sizonenko,C.Bor-radori-Tolsa, J. F. Mangin and P. S. Huppi(2008c). ”Mapping the early cortical folding pro-cess in the preterm newborn brain.“ Cereb Cor-tex 18(6): 1444-54.
e Dubois, J., M. Benders, F. Lazeyras, C. Bor-radori-Tolsa, R. H. Leuchter, J. F. Mangin and P.S.Huppi (2010). ”Structural asymmetries of peri-sylvian regions in the preterm newborn.“ Neu-roimage 52(1): 32-42.
GRP : fyssen JOB : annales27Internet DIV : 09mp⊕DUBOIS⊕Jessica p. 7 folio : 84 --- 14/5/013 --- 15H6
NEUROBIOLOGIE
84

e Dubois, J.,G.Dehaene-Lambertz,M.Perrin,J. F. Mangin, Y. Cointepas, E. Duchesnay, D. LeBihan and L. Hertz-Pannier (2008a). ”Asyn-chrony of the early maturation of white matterbundles in healthy infants: quantitative land-marks revealednoninvasively bydiffusion tensorimaging.“ Hum Brain Mapp 29(1): 14-27.
e Dubois, J.,G.Dehaene-Lambertz, C. Soares,Y. Cointepas, D. Le Bihan and L. Hertz-Pannier(2008b). ”Microstructural correlates of infantfunctional development: example of the visualpathways.“ J Neurosci 28(8): 1943-8.
e Dubois, J., L.Hertz-Pannier, A. Cachia, J. F.Mangin, D. Le Bihan and G. Dehaene-Lambertz(2009). ”Structural asymmetries in the infantlanguage and sensori-motor networks.“ CerebCortex 19(2): 414-23.
e Dubois, J., L. Hertz-Pannier, G. Dehaene-Lambertz, Y. Cointepas andD. LeBihan (2006a).”Assessment of the early organization andmatu-ration of infants“ cerebral white matter fiberbundles: a feasibility study using quantitativediffusion tensor imagingand tractography.“Neu-roimage 30(4): 1121-32.
e Erberich, S. G., A. Panigrahy, P. Friedlich,I. Seri, M. D. Nelson and F. Gilles (2006). ”Soma-tosensory lateralization in the newborn brain.“Neuroimage 29(1): 155-61.
e Feess-Higgins, A. and J. C. Laroche (1987).Development of the human foetal brain: an ana-tomical atlas. Inserm-CNRS, Masson.
e Garel, C., E. Chantrel, H. Brisse, M. Elma-leh,D. Luton, J. F.Oury,G. Sebag andM.Hassan(2001). ”Fetal cerebral cortex: normal gestatio-nal landmarks identified using prenatal MR ima-ging.“ AJNR Am J Neuroradiol 22(1): 184-9.
e Huppi, P. S. andJ.Dubois (2006). ”Diffusiontensor imaging of brain development.“ SeminFetal Neonatal Med 11(6): 489-97.
e Le Bihan, D., J. F. Mangin, C. Poupon, C. A.Clark, S. Pappata, N. Molko and H. Chabriat(2001). ”Diffusion tensor imaging: concepts andapplications.“ J Magn Reson Imaging 13(4):534-46.
e McCulloch, D. L., H. Orbach and B. Skarf(1999). ”Maturation of the pattern-reversal VEPin human infants: a theoretical framework.“Vision Res 39(22): 3673-80.
e Neil, J., J. Miller, P. Mukherjee andP. S. Huppi (2002). ”Diffusion tensor imaging of
normal and injured developing human brain - atechnical review.“ NMR Biomed 15(7-8): 543-52.
e Ono, M., S. Kubick and C. Abernathey(1990). Atlas of the cerebral sulci Georg ThiemVerlag.
e Parker,G. J., S. Luzzi,D.C.Alexander,C.A.Wheeler-Kingshott, O. Ciccarelli and M. A. Lam-bon Ralph (2005). ”Lateralization of ventral anddorsal auditory-language pathways in the humanbrain.“ Neuroimage 24(3): 656-66.
e Paus, T., D. L. Collins, A. C. Evans, G. Leo-nard, B. Pike and A. Zijdenbos (2001). ”Matura-tion of whitematter in the human brain: a reviewof magnetic resonance studies.“ Brain Res Bull54(3): 255-66.
e Sowell, E. R., P. M. Thompson, D. Rex,D. Kornsand, K. D. Tessner, T. L. Jernigan andA. W. Toga (2002). ”Mapping sulcal patternasymmetry and local cortical surface graymatterdistribution in vivo: maturation in perisylviancortices.“ Cereb Cortex 12(1): 17-26.
e Toga, A. W. and P. M. Thompson (2003).”Mapping brain asymmetry.“ Nat Rev Neurosci4(1): 37-48.
e van der Knaap, M. S. and J. Valk (1990).”MR imaging of the various stages of normalmyelination during the first year of life.“ Neuro-radiology 31(6): 459-70.
e Welker, W. (1990). ”Why does cerebral cor-tex fissure and fold? A review of determinants ofgyri and sulci. .“ In: Comparative structure andevolution of cerebral cortex. Jones E, Peters Aeds.Plenum,NewYork.Part II,vol8B: 3-136.
e Yakovlev, P. I. and A. R. Lecours (1967).”The myelogenetic cycles of regional maturationin the brain.“ In: Regional development of thebrain in early lifeMinowski A. eds, Blackwell,Oxford: 3-69.
IntroductionSeveral cerebral pathologies result from early
anomalies in brain development and affect animportant number of individuals, for exampleepilepsies, mental retardation and dyslexia.Brain development can also be disturbed early onby abnormal events like a premature birth, anintra-uterine growth restriction or a neonatalstroke. Studying how the healthy brain anatomi-cally develops and becomes functional is impor-tant in order to comprehend the complexity of
GRP : fyssen JOB : annales27Internet DIV : 09mp⊕DUBOIS⊕Jessica p. 8 folio : 85 --- 14/5/013 --- 15H6
NEUROBIOLOGY
85

these pathologies and to assess the influence ofdisturbances. It can also help to understand howthe human brain enabled him to elaborate speci-fic cognitive abilities. Brain development reliesonseveralmechanisms, andcerebral changesareparticularly intense during the last weeks ofgestation and the first post-natal months. Thebaby’s brain is not an adult brain in miniature:growth is inhomogeneous, since some regions,like the sensorial regions, develop early on andquickly, whereas other associative regions, likefrontal regions, develop later on and slowly untilthe end of adolescence (Paus et al. 2001). Longbefore that hemispherical functional specializa-tion can be observed, anatomical asymmetriesare present in the foetus cerebral hemispheres(Chi et al. 1977b). Concurrently with this anato-mical evolution of the brain, the newborn andinfant gradually achieve new psycho-motor andcognitive performances.
Understanding how these acquisitions takeplace in relation with brain maturation requiresin vivo studies. Post mortem investigations areintrinsically limited because you cannot be sureof the studied brain normality and you cannotestablishanatomo-functional correlations.Giventhe human brain specificity, animal studiesremain inadequate even if they enable to testparticular hypotheses. In this context, mostpotential is elicited by non-invasive imaging tech-niques (magnetic resonance imaging MRI, elec-troencephalography EEG, magnetoencephalo-graphyMEG) which enable to compare thematu-ration of cerebral structures with the new acqui-sitions by the healthy infant, and to longitudi-nally follow up the development and the brainplasticity in at-risk children.
However, you are faced with many difficultieswhen imaging the baby’s brain. Actually, thisbrain is changing every day, contrarily to theadult brain whose modifications are sparsebetween 20 and 60 years old. Because of thisbrain tissues immaturity, adapting the dataacquisition and post-treatment for each period ofdevelopment is required. Moreover, the baby’scerebral structures are small, so the images spa-tial resolution should be higher. Finally, MRI,EEG and MEG techniques are very sensitive tomotion:dataacquisitionshould remainshort andwithout any discomfort in order to assure thebaby’s sleep or nice cooperation.
Thanks to various methodological implemen-tations, we recently set up two complementarystudies which aimed at highlighting the anatomi-cal development of thehumanbrain in vivo andatquantifying the changes happening during thefirst post-natal months. The first study dealt withthe early folding of cortical sulci in pretermnewborns imaged at birth by conventional MRI,between six and eight months of gestationalage. The second study described the followingorganization and maturation of white matter,using diffusion tensor imaging (DTI) in the heal-thy infant between one and four months ofpost-natal age. For each study, the babies’parents gave written informed consent and theprotocol was approved by the local ethical com-mittee. Several axes of research were investiga-ted. Spatio-temporal differences in the matura-tion of cerebral regions were evaluated, as wellas early asymmetries between cerebral hemis-pheres. Further correlations between anatomi-cal changes and functional maturation wereassessed.
Folding of cortical sulci in the pretermbrain
The brain cortex develops, gets thicker andfolds mainly during the last trimester of gesta-tion. In addition to its important size growth, thefoetal brain morphology becomes quickly elabo-rated with the formation of gyri and sulci visiblemacroscopically. The folds complexity increaseswith age (Chi et al. 1977a; Feess-Higgins andLaroche 1987), and the adult brain shows a highinter-individual variability in its morphologicalpatterns (Ono et al. 1990). Several hypotheseshave been proposed on the mechanisms trigge-ring thegyrificationprocess (genetic, epi-genetic,mecanical or environmental factors) (Welker1990).
In order to explore in vivo the early stages ofcortical development and to define chronologicalstandards based on quantitativemarkers of sulcifolding, we studied preterm newborns (from6.2 to 8.3 months of gestational age GA) ex uterowith conventional MRI and new post-processingtools dedicated to the immature brain (Dubois etal. 2008c). From high-resolution T2-weightedimages acquired on a 1.5T MRI (Figure 1a), wereconstructed in 3 dimensions the inner surfaceof the cortex, at the border with the white matter(Figure 1b). Over this range of ages correspon-
GRP : fyssen JOB : annales27Internet DIV : 09mp⊕DUBOIS⊕Jessica p. 9 folio : 86 --- 14/5/013 --- 15H6
NEUROBIOLOGY
86

ding to the last trimester of pregnancy, the sulcifolding is dramatic, and a sulcation index wasproposed to measure this process quantitatively(Figure 1c).
Theprogressive appearance of primary, secon-dary and tertiary sulci was highlighted, in agree-ment with previous atlases based on post mortemstudies (Chi et al. 1977a; Feess-Higgins andLaro-
Figure 1MRI imaging of cortical sulci in preterm newbornsa: T2-weighted images from a 31week-old preterm newborn.b: 3D reconstructions of the inner cortical surface in 3 preterm newborns with increasing age and sulcationindex. The color codes for the surface curvature. Adapted from (Dubois et al. 2008c).c: Age-related increase in the newborn sulcation index. Adapted from (Dubois et al. 2008c).d: Inter-hemispherical asymmetries. The cortical surface shows morphology differences between the twocerebral hemispheres, at the level of the superior temporal sulcus (STS) and the sylvian fissure, anteriorly (infront of Broca’s region) and posteriorly (at the level of the planum temporale). Adapted from (Dubois etal. 2010).e: Sulcation differences according to the intrauterine environnement. For equivalent gestational age, the twinnewborn shows a reduced sulcation index in comparison with the singleton (left-sided images), according toa harmonious delay in maturation. For equivalent cortical surface (but at different ages), the newborn withintra-uterine growth restriction shows an increased sulcation index in comparison with the singleton(right-sided images), suggesting a disharmonious development. Adapted from (Dubois et al. 2008d).
GRP : fyssen JOB : annales27Internet DIV : 09mp⊕DUBOIS⊕Jessica p. 10 folio : 87 --- 14/5/013 --- 15H7
NEUROBIOLOGY
87

che 1987) and MR prenatal studies (Garel et al.2001). An asynchronous pattern was outlinedaccording to cerebral regions. The sylvian fissurewas folded in all newborns, as well as the medialsurface (callosal and cingulate sulci, parieto-oc-cipital and calcarine scissures). On the lateralsurface, the sulci differentiation began in thecentral region (central sulcus present in all new-borns) and progressed in the parietal lobe (iden-tification of post-central, intra-parietal and parie-to-occipital sulci between 27 and 31w GA), thetemporal lobe (identification of superior and infe-rior temporal sulci between 27 and 32w GA, ofcollateral sulcus between 29 and 30w GA), andfinally in the frontal lobe (identification of pre-central, superior and inferior frontal sulcibetween 29 and 30wGA, of orbital sulcus around30-31w GA, and of olfactive sulcus from 32w GAon).
Organization and maturation of whitematter bundles in the infant brain
At the same time as the cortex develops, thebelow white matter gets organized and mature.Notably, thefibersmyelination is aprocesswhichincreases the conduction speed of the nerveimpulse (Baumann and Pham-Dinh 2001). Untilrecently, most of our knowledge on myelinationwas based on post mortem studies (Yakovlev andLecours 1967), and many questions remainedunanswered, for instance how myelination hasfunctional consequences on the infant’s beha-vior. Since a few decades, conventional MRI hasenabled to assess this maturation in vivo (Barko-vich 2000; Paus et al. 2001), through changes inT1 and T2 weightings related to the «pre-myelin»formation by the oligodendrocytes and to themyelin ensheathment around axons («truemyelination»). Nowadays, quantitative indicesdependant on these processes can be measuredwith DTI imaging (Neil et al. 2002; Huppi andDubois 2006), technique which measures thewater molecules motion and enables to explorethe tissue microstructure non-invasively (LeBihan et al. 2001).
In order to study the organization of whitematter and to quantify its maturation, healthyinfants, bornat term,were imagedbetween1.3 to4.5 months of post-natal age using a 1.5T DTIprotocol dedicated to the immature brain (Figure2a) (Dubois et al. 2006a). We validated the hypo-thesis of an early white matter organization by
reconstructing themajor bundles with tractogra-phy (Figure 2b). Despite a low myelination, wedetected commissural (genu, body and spleniumof the corpus callosum), projection (cortico-spi-nal tract, spino-thalamic tract, optic radiations,anterior limb of the internal capsule), limbic(fornix and cingulum) and associative bundles(external capsule, uncinate, arcuate, superiorand inferior longitudinal fascicles).
Important age-related changes in DTI indiceswere observed (decrease in mean diffusivity,increase in anisotropy: Figure 2c). We aimed atquantifying the differences in maturation acrosswhite matter bundles. Because of the comple-mentarities of DTI indices (Beaulieu 2002), weproposed a model relating maturational proces-ses occurring at the microscopic level and varia-tions in DTI indices measured at the voxel level(Dubois et al. 2008a). Over this range of ages, weobserved from the more mature bundles to theless mature: 1) the cortico-spinal tract, 2) thespino-thalamic tract and the fornix, 3) the opticradiations, the arcuate and inferior longitudinalfascicles, 4) the anterior limb of the internalcapsule and the cingulum. These results were ingood agreement with previous post-mortem stu-diesbasedonahistologicalmarkingof themyelin(Flechsig 1920; Yakovlev and Lecours 1967;Brody, Kinney et al. 1987; Kinney, Brody et al.1988) andwith in vivo studies using conventionalMRI (van der Knaap and Valk 1990; Paus et al.2001). Such studies described the myelinationprogression, from central regions to periphery,according to a caudo-rostral gradient, first in thesensory pathways, second in themotor pathwaysand finally in the associative pathways.
These two MRI studies have highlighted spa-tio-temporal differences in the development ofcortical sulci and between the various white mat-ter bundles. In the manner of the progressiveinfant acquisitions at the sensori-motor andcognitive levels, cerebral regions develop and getmature at different rhythms and over differenttime periods.
Early asymmetries between cerebralhemispheres
In addition to the anatomical complexity, thecerebral hemispheres of the adult brainare asym-metrical, notably through the Yakovlev torquewith frontal and occipital petalias (Toga andThompson 2003). Some of the structural asym-
GRP : fyssen JOB : annales27Internet DIV : 09mp⊕DUBOIS⊕Jessica p. 11 folio : 88 --- 14/5/013 --- 15H7
NEUROBIOLOGY
88

metries observed in peri-sylvian regions (Heschlgyrus, planum temporale, superior temporal sul-cus STS) have been described during early braindevelopment through post-mortem studies of foe-tal brains (Chi et al. 1977b; Feess-Higgins and
Laroche 1987). Other macroscopic asymmetriesevolve later on during development: for instancethe posterior extension of the sylvian fissureprogresses until adolescence and adulthood(Sowell et al. 2002).
Figure 2White matter DTI imaging in healthy infantsa: DTI map acquired in a 7week-old infant. The colors code for the local main direction of thefibers (red: left-right direction; green: anteriorposterior direction; blue: inferior-superior direction).b: Tractography reconstruction of the main fiber bundles in a 17week-old infant. Adapted from (Dehaene-Lambertz et al. 2006).Abbreviations: cc corpus callosum; cg cingulum; cst cortico-spinal tract; ilf: inferior longitudinal fasciculus; oroptic radiations; stt spino-thalamic tract; unc uncinate fasciculus.c: Age-related increase in the anisotropy of infants’ whitematter bundles. Adapted from (Dubois et al. 2008a).d: Inter-hemisphericalasymmetries.Anasymmetry in thearcuate fasciculusvolumeisreported in its temporalpart in favour of the left hemisphere (left-sided image), while an asymmetry in its microscopic organization isobserved in the parietal part of the same side (central image). An asymmetry ofmaturation is described in theinferior part of the cortico-spinal tract (cp-plic: between the cerebral peduncles and the posterior limb of theinternal capsule) in favour of the left hemisphere (right-sided image). Adapted from (Dubois et al. 2009).
GRP : fyssen JOB : annales27Internet DIV : 09mp⊕DUBOIS⊕Jessica p. 12 folio : 89 --- 14/5/013 --- 15H7
NEUROBIOLOGY
89

At the functional level, the left-hemispherespecialization for language processing is obser-ved early on in the infant brain (Dehaene-Lam-bertz et al. 2002; Dehaene-Lambertz et al. 2006),and the lateralization of the somato-sensory res-ponse is also detected at birth (Erberich et al.2006). However, the origins and relationshipsbetween these anatomical and functional asym-metries remain to be explored in vivo in thedeveloping brain.
Asymmetries in the preterm newborn sulciEarly asymmetries were demonstrated on the
inner cortical surfaces of preterm newborns,in three peri-sylvian regions of the languagenetwork (Figure 1d). The STS folded early on inthe right hemisphere (Dubois et al. 2008c). Tworegions of the sylvian fissure were larger on theleft side, in anterior in front ofBroca’s regionandin posterior at the level of the planum temporale(Dubois et al. 2010).
Asymmetries in the infant white matter bundlesThree white matter regions were detected as
asymmetric using DTI imaging in healthy infants(Figure 2d) (Dubois et al. 2009). The arcuate fasci-culus was larger on the left side in its temporalpart. Its left parietal part showed a higher micros-copic organization in comparison with the right.Finally, the cortico-spinal tract was more maturein the left hemispherebetween the cerebral pedun-cles and the posterior limb of the internal capsule.
These two bundles are also asymmetric in theadult brain (Buchel et al. 2004; Parker et al.2005), and they are related to the functionalnetworks of language perception/productionandofmanuality respectively.What is surprisingis that they are asymmetric early on, when thesetwo functions are still little developed.
Correlations between brain anatomicalmaturation and functional development
In these two in vivo studies, we have outlineddifferences between cerebral regions in their ana-tomical development and early inter-hemispheri-cal asymmetries. To which extent the MRI struc-tural indices further provide information on thepsycho-motor and cognitive development of theinfants?
Early sulcation and longitudinal follow-up ofpreterm newborns
Among the preterm newborns, three groupswere distinguished: 1) 27 healthy newborns born
from a single pregnancy; 2) 12 healthy newbornsborn froma twin pregnancy; 3) 11 newbornswithintra-uterine growth restriction (IUGR) and bornfrom a single pregnancy. MR birth measure-ments enabled to demonstrate that intra-uterineenvironment had a major impact on brain deve-lopment (Dubois et al. 2008d). The quantitativeanalysis of cortical volumes, surfaces and sulca-tion indices highlighted that twin newborns pre-sented a harmonious delay in maturation, ofabout two weeks according to their gestationalage. On the contrary, the maturation delay ofIUGR newborns was discordant, with sulcationindices too important in comparisonwith the lowcortical surface (Figure 1e).
Furthermore, these cortical measurements atbirthwere strongly correlatedwith the later new-borns development, at term equivalent age. Thebiggest cortical volumes and the highest functio-nal scoresmatchedwith highest cortical surfacesat birth, after correction for age effects (Dubois etal. 2008d). In the future, this methodology couldhelp to evaluate the impact on brain developmentof various medical strategies for caring pretermnewborns.
Fiber myelination and conduction speed ininfants
Besides, we considered the development of thevisual system in order to evaluate the functionalsignificance of DTI indices. EEG and more par-ticularly visual event-related potentials arewidely used in infants thanks to the easy setting-up. Because of several maturation processes(fiber myelination, retina and cortex develop-ment, etc.), the latency of the first wave of res-ponse to a visual stimulus (P1) decreases drasti-callywith age, fromabout 250msat birth to about120msat 4months of age (McCulloch et al. 1999).
In healthy infants, the conduction speed of thenerve impulse was estimated in the visualpathways from this P1 latency. We showed thatthis speed depended on DTI indices measured intheoptic radiations, besides theage-relatedeffect(Dubois et al. 2008b). This suggests that thesestructural indices also provide information onthe brain functional maturation.
ConclusionStudying thedynamics of normal humanbrain
development and the anatomical bases of func-tional maturation in vivo triggers some difficul-
GRP : fyssen JOB : annales27Internet DIV : 09mp⊕DUBOIS⊕Jessica p. 13 folio : 90 --- 14/5/013 --- 15H7
NEUROBIOLOGY
90

ties in thehealthynewbornand infant. It requiresimplementing dedicated methodologies for ima-ging and post-processing the data. By conside-ring these methodological challenges, new quan-titative markers of maturation can be proposed.Such approach gives the opportunity to explorethe developing brain from a new and transversalperspective, from the brain structural develop-ment to the infant behavioural acquisitions. Itwill help to better understand some developmen-tal pathologies and to improve medical diagnos-tic with quantitative criteria predictive of fol-lowing deficits.
AcknowledgmentsThis work was supported by the Fyssen Fun-
dation, the «Ecole des Neurosciences de Paris»,the McDonnell Fundation, the «FondationMotrice», the ANR French National Agency forResearch, theCIBM imaging center fromGenevaandLausanne, theSwissNationalFundation, theLeenards Fundation and the European consor-tium NEOBRAIN. I particularly thank L. Hertz-Pannier, G. Dehaene-Lambertz, D. Le Bihan,J.F. Mangin, P. Hüppi, M. Benders and F. Lazey-ras, who helped me to lead these fascinatingworks.
GRP : fyssen JOB : annales27Internet DIV : 09mp⊕DUBOIS⊕Jessica p. 14 folio : 91 --- 14/5/013 --- 15H7
NEUROBIOLOGY
91

Robustesse de l’activité neuronale :un autre versant de plasticité
Jean-Marc Goaillard
Chargé de recherche INSERMCR1Unité 1072 « Laboratoire de Neurobiologie des Canaux Ioniques etde la Synapse » à l’Université Aix Marseille
RésuméDe nombreuses études ont démontré que les systèmes biologiques sont particulièrement robustes,
c’est-à-dire capables de maintenir une fonction stable malgré nombre de perturbations externes ouinternes. Bien que cela ait été démontré également pour le système nerveux, notamment en ce quiconcerne la stabilité de l’activité neuronale, les principes moléculaires responsables de cette stabilitésont encoremal connus. Des études récentes suggèrent que les principes de la robustesse de l’activiténeuronale résideraient dans la redondance fonctionnelle entre les nombreux canaux ioniquesexprimés par chaque neurone ainsi que dans la coordination étroite de leurs niveaux d’expression.
Mots-clésRobustesse, Plasticité, Neurones, Canaux ioniques
Robustness of neuronal activity:another facet of plasticity
AbstractA number of studies have demonstrated that biological systems are astonishingly robust in the
sense that they canmaintain a stable functional output in the face of numerous internal and externalperturbations. Although this property has also been described for the nervous system, in particularat the level of neuronal activity, the molecular principles underlying this stability are still largelyunknown. Recent studies suggest that the robustness of neuronal activity may relie on the strongfunctional redundancy between the ion channels expressed by each neuron and on tight co-regulationof their expression levels.
KeywordsRobustness, Plasticity, Neurons, Ion Channels
IntroductionUne des propriétés les plus stupéfiantes des
systèmes biologiques complexes réside dans leurcapacité àmaintenir une fonction stable en dépitdes très nombreuses perturbations internes ouexternes qu’ils subissent constamment. Cettepropriété essentielle à la survie des organismesbiologiques a été nommée « robustesse » biolo-gique. L’omniprésence de la robustesse biolo-gique à tous les niveaux de complexité biologiquea conduit certains auteurs à suggérer qu’elleconstituait une propriété structurelle des systè-
mes biologiques et qu’au cours de l’évolution lessolutions les plus robustes auraient été systéma-tiquement privilégiées (Wagner 2005).
Dans les faits, la robustesse reflète l’existencede solutions très diverses à chaque problèmebiologique. Au niveau du code génétique parexemple, chaque acide aminé est codé par unemultitude de triplets de nucléotides, phénomènequ’onnommedégénérescenceducodegénétique.Et même lorsque les mutations provoquent lasubstitution d’un acide aminé par un autre, lastructure et la fonctionde laprotéineproduite est
GRP : fyssen JOB : annales27Internet DIV : 10mp⊕GOAILLARD⊕JeanMarc p. 1 folio : 92 --- 14/5/013 --- 15H11
NEUROBIOLOGIE
92

rarement modifiée. A un autre niveau, de nom-breuses études ontmontré que le développementde la vulve du vers Caenorhabditis elegans étaitextrêmement robuste à la délétion d’une partiedes cellules progénitrices, et que de multiplesvoies de différenciation cellulaire permettaientd’obtenir un organe fonctionnel. L’existence desolutions multiples repose donc sur une certaineredondance entre les composants d’un systèmebiologique, de telle manière que la perte ou lamodification d’un de ces composants peut êtrecompensée par des composants fonctionnelle-ment proches.
Robustesse du système nerveuxParmi les systèmes biologiques complexes, le
système nerveux, et en particulier le cerveau demammifères, avec ses 1010 neurones intercon-nectés par 1014 synapses, tient une place privilé-giée. Et, sans surprise, notre connaissance crois-sante du fonctionnement du système nerveuxnous révèle que celui-ci est extrêmement robuste.Cette robustesse s’exprime à de nombreuxniveaux et de façon variable. Un des exemples lesplus frappants est la capacité extraordinairedu système nerveux à récupérer (parfois totale-ment) des divers traumatismes qui provoquentdes lésions irréversibles de circuits neuronaux,tels que les accidents vasculaires cérébraux ouencore la présence de tumeurs cérébrales. Dansces cas-là, la robustesse a souvent été assimilée àla plasticité du système nerveux. Mais la robus-tesse du système nerveux s’exprime aussi auniveau de la molécule et du neurone unique, etpas seulement au niveau des réseaux neuronaux.
L’activité électrique des neurones est produitepar l’activation de canaux ioniques qui condui-sent les ions (sodium, potassium, calcium,chlore) à travers la membrane plasmique. Lescanaux dépolarisants (conduisant le sodiumet/ou le calcium) ont tendance à promouvoirl’activité neuronale tandis que les canaux hyper-polarisants (potassium, chlore) tendent à inhiberl’activiténeuronale.Auniveauducorpscellulaireet des prolongements neuronaux (dendrites,axone), le neurone possède une multitude decanaux ioniques aux propriétés variées qui luipermettent de générer de l’activité de façonintrinsèque oude générer de l’activité en réponseaux neurotransmetteurs libérés au niveau dessynapses. Chaque type neuronal exprime un
assortiment particulier de canaux ioniques asso-cié à un type particulier d’activité : les neuronespyramidaux du cortex n’ont pas le même typed’activité ni le même assortiment de canaux ioni-ques que lesmotoneurones de lamoelle épinière.Une question centrale est donc de comprendrecomment un neurone d’un type donné arrive àmaintenir son profil d’activité spécifique durantplusieurs années alors qu’il est potentiellementsoumis à de fortes variations de son environne-ment chimique, du nombre de connexions synap-tiques qu’il reçoit, etc...
Plasticité homéostatique de l’activiténeuronale
Au cours des vingt dernières années, de nom-breuses études ont démontré que les neuronessont capables d’ajuster leurs propriétés électri-ques en fonction de la stimulation électriquequ’ils reçoivent. Ainsi, lorsque les neurones sontprivés d’activité (par un agent pharmacologiquespécifique, ou suite à une privation sensorielle),on observe une augmentation du nombre decanaux ioniques dépolarisants et une diminutiondu nombre de canaux hyperpolarisants, de tellemanière que le niveau global d’activité du neu-rone n’est que peu ou pas altéré. L’inverse seproduit lorsqu’on soumet le neurone à une aug-mentation artificielle d’activité, le nombre decanaux hyperpolarisants et dépolarisants aug-mentant et diminuant respectivement. Cettefaculté des neurones à réguler de façon dynami-que les propriétés ou le nombre de leurs canauxioniques pour maintenir un niveau d’activité sta-ble a été nommée plasticité homéostatique (dugrec Ömoıov, homoios, « similaire » et Ïstcmı,histomi, « immobile ») (Davis 2006). La plasticitéhoméostatique représenteunversantde la robus-tesse (réponse aux perturbations externes) del’activité neuronale, mais malheureusement lesrégulations binaires (augmentation versus dimi-nution) du nombre de canaux dépolarisants ethyperpolarisantsdécritesnepermettentpasvrai-ment de comprendre comment le neurone main-tient un profil d’activité spécifique et précis,maisseulement comment le neurone maintient sonniveau général d’excitabilité. Maintenir un profild’activité précis exige probablement des régula-tions coordonnées très fines des propriétés decanaux ioniques qui partagent une même fonc-tion.
GRP : fyssen JOB : annales27Internet DIV : 10mp⊕GOAILLARD⊕JeanMarc p. 2 folio : 93 --- 14/5/013 --- 15H11
NEUROBIOLOGIE
93

Redondance fonctionnelle entre canauxioniques : manipulation
On sait en effet que chaque neurone exprimebien plus que le nombre minimal de canauxioniques qui lui serait théoriquement nécessairepour générer sonprofil d’activité. Et les expérien-ces sur des souris KO (Knocked Out, souris chezlesquelles l’expression d’un gène a été suppri-mée) démontrent que cette redondance fonction-nelle permet de maintenir un profil d’activitéspécifique même lorsqu’un des canaux ioniquesimpliqués dans la genèse d’activité est supprimé(perturbation interne). Ainsi, les travaux deSwensen et Bean (2005) ont permis de montrerque les neurones de Purkinje du cervelet conti-nuent à générer des bouffées de potentielsd’action même si on supprime un canal sodique(Nav1.6) impliqué dans la genèse de ces bouffées(Figure 1A). Bien que l’application d’une neuro-toxine spécifique de ce canal ionique altère forte-ment l’activité du neurone, la perte chronique ducanal ionique (KO Nav1.6 ou Nav1.6-/-) ne lamodifie que très peu. Ainsi, des mécanismes decompensation (non identifiés dans cette étude)sont mis en jeu qui permettent de pallier la perted’un canal ionique majeur. Une étude similaire apermis demontrer que les neurones pyramidauxdu cortex sont capables de maintenir leur profild’activité même après la suppression d’un canalpotassique (Kv4.2) pourtant considéré commeessentiel dans ces neurones. Dans ce cas, lesauteursontpumontrerque lapertedececanal (etdu courant ionique qu’il engendre) est compen-sée par une augmentation d’amplitude d’autrescourants potassiques (de sorte que leur sommealgébrique est maintenue constante) qui permet-tent au neurone de maintenir son profil d’acti-vité (Nerbonne et al. 2008) (Figure 1B). Cetteétude démontre clairement la redondance fonc-tionnelle entre les différents canaux potassiques(et les courants qu’ils engendrent), et commentcette redondance estmise à profit lors de la perted’un de ces canaux. Cependant, une seule étude apermis de quantifier précisément les modifica-tions de propriétés de canaux ioniques nécessai-res aumaintien d’un profil d’activité stable. Danscette étude réalisée sur le système nerveux de lalangouste, les auteurs ont artificiellement aug-menté l’amplitude d’un courant potassique(porté par le canal ionique Shal) en microinjec-tant l’ARN messager du canal dans un neurone
produisant des bouffées spontanées de potentielsd’action. Malgré cette micro-injection qui provo-que une augmentation importante du courant
Figure 1. Stabilité du profil d’activité desneurones après suppression d’un canalionique. A, les bouffées de potentiels d’actionenregistrées dans des neurones de Purkinje aprèssuppression du canal sodique Nav1.6 (sourisNav1.6 -/-, graphes bleus) sont très similaires àcelles enregistrées chez des souris Nav1.6 +/+(graphes violets). Les traces du haut et du milieumontrent le degré de similarité dans la formegénérale, les histogrammes du bas montrant que lenombre de potentiels d’action générés par boufféeest également très similaire dans des deux cas.B, lepotentiel d’action enregistré dans les neuronespyramidaux du cortex après suppression du canalpotassique Kv4.2 (souris Kv4.2 -/-) présente uneforme identique à celui enregistré chez des sourisKv4.2 +/+. A, d’après Swensen and Bean (2005) ;B, d’après Nerbonne et al. (2008).
GRP : fyssen JOB : annales27Internet DIV : 10mp⊕GOAILLARD⊕JeanMarc p. 3 folio : 94 --- 14/5/013 --- 15H12
NEUROBIOLOGIE
94

potassique, le profil d’activité du neurone nechange pas (MacLean et al. 2005) (Figure 2A).L’augmentation d’amplitude du courant potas-sique (hyperpolarisant) est en fait compenséepar l’augmentation d’un courant dépolarisantnommé IH, tel que l’activité du neurone est ainsistabilisée. C’est ainsi un cas de redondancefonctionnelle sensiblement différent de l’exem-ple cité précédemment (Nerbonne et al. 2008)(Figure 1B), puisqu’ici les deux canaux mis enjeu ont un effet opposé sur le profil d’activité, etque c’est le maintien d’un rapport constant entreles amplitudes de ces deux courants qui permetde maintenir l’activité (Figure 2B).
Dans tous les cas, les trois études citées icidémontrent clairement l’existence d’une redon-dance fonctionnelle entre canaux ioniques dontémerge la robustesse de l’activité neuronale. Ilest aisé d’imaginer que ce type de mécanismen’agit pas seulement en réponse à des mani-pulations expérimentales de l’expression descanaux ioniques (tels que KO ou micro-injectiond’ARNmessager) mais qu’il est également impli-qué dans la réponse à des mutations spontanées(et naturelles) des canaux ioniques entraînantdes altérations fonctionnelles de ces protéines.Bien que la plupart des gènes identifiés dans desformes génétiques d’épilepsie codent pour descanaux ioniques, il y a fort à parier qu’un nombreimportant de mutations de canaux ioniques pas-sent inaperçues parce que la redondance fonc-tionnelle entre canauxpermetdans laplupartdescas une compensation quasi-parfaite de ces alté-rations de fonction. Une étude clinique récente aen effet démontré que les mutations dans lesgènes codant les canaux ioniques sont très nom-breuses, non seulement chez les sujets épilepti-ques, mais également chez les sujets sains, sug-gérantque laplupart desmutationsn’ont quepeud’effet sur le phénotype, et qu’il est du coupimpossible de prédire le risque de développer lapathologie sur la seule base d’un pronostic géné-tique (Klassen et al., 2011).
Redondance fonctionnelle entre canauxioniques : observation
Si la robustesse est une propriété structurelledes systèmes biologiques, les mécanismes qui lasous-tendent devraient être observables au seinde neurones non perturbés par des manipula-tions expérimentales. La redondance fonction-nelle entre les multiples canaux ioniques expri-
més par chaque neurone crée potentiellement unnombre de solutions vertigineux pour générer etmaintenir leur profil d’activité spécifique. Laquestion qui se pose alors est de savoir si lesneurones utilisent effectivement des solutionsvariées, démontrant ainsi que les principes de larobustesse sont à l’œuvre dans des populations« naturelles » de neurones.
Des études réalisées sur le systèmenerveux ducrabe tendent à prouver que des neurones d’unmême type, parfois présentant des profils d’acti-vité quasi-identiques, utilisent des solutions trèsvariées en termes d’expression de canaux ioni-ques qui cependant reposent sur des principesconstants. Ainsi, l’enregistrement électrophysio-logique de trois courants potassiques différentsdans le neurone pylorique latéral (LP en anglais)a révélé des différences importantes dans leuramplitude (3 à 4 fois) d’un neurone LP à unautre, même lorsque ces neurones ont des pro-fils d’activité quasiment indiscernables (Schulz
Figure 2. Compensation de l’effet de la micro-injection d’ARNm du canal Shal par co-variation du courant IH. A, les bouffées depotentiel d’action spontanées générées par leneurone pylorique de la langouste ne sont pasaltérées par la microinjection d’ARN messager ducanal Shal, bien que le courant ionique porté par cecanal (courant IA) soit significativement augmenté(non montré). B, l’augmentation d’amplitude ducourant IA est associée à une augmentationcoordonnée de l’amplitude du courant IH, courantqui a un effet opposé sur l’activité du neurone. Lastabilité de l’activité est due à cette augmentationcompensatoire. D’après MacLean et al. (2003).
GRP : fyssen JOB : annales27Internet DIV : 10mp⊕GOAILLARD⊕JeanMarc p. 4 folio : 95 --- 14/5/013 --- 15H12
NEUROBIOLOGIE
95

et al. 2006) (Figure 3A). Deux explications sontalors possibles face à cette observation : i) lesneurones sont relativement insensibles à ces dif-férences d’amplitude de courants, et ces varia-tions n’ont que peu d’influence sur l’activité duneurone, ou ii) la variabilité des courants mesu-rés est associée à une co-variation de courantsfonctionnellement redondants (complémentai-res ou antagonistes). Les résultats d’expériencesde pharmacologie classiques où l’on bloque l’acti-vité d’un courant ionique montrent que les neu-rones ne sont pas indifférents à des variationsd’amplitude de cet ordre. L’étude de McLean etcollègues citée précédemment (où la variationd’expression du canal Shal entraîne un change-ment coordonné de l’amplitude du courant IH)suggère par contre que certains canaux sont pro-bablement co-régulés pour maintenir un profild’activité stable. Et en effet, une étude ultérieureréalisée sur plusieurs types de neurones du sys-tème nerveux de crabe a permis de confirmercette hypothèse. L’analyse du niveau d’expres-sion de 6 canaux ioniques dans 6 types cellulairesdifférents a permis d’observer un nombre impor-tant de corrélations entre les niveaux d’expres-sion de plusieurs canaux ioniques. L’analyse del’expression des canaux potassiques dans leneurone LP a par exemple démontré que lesniveaux d’expression des canaux potassiquesShal, Shaw, et du canal IH (responsable du cou-rant IH) étaient fortement corrélés (Schulz etal. 2007) (Figure 3B). Ainsi, la variabilitéd’amplitude des courants potassiques pourraitêtre compensée par la variation coordonnéed’autres canaux, par exemple le canal IH quicontrecarrerait les effets du canalShal (MacLeanet al. 2005). Les neurones LP produisent uneactivité stéréotypée non parce qu’ils exprimentun assortiment de canaux ioniques spécifiques àun niveau constant, mais parce que les niveauxd’expressiondecanauxfonctionnellementredon-dantssontrégulésdefaçoncoordonnée.Ainsi, lessolutionsutiliséespar lesneuronesLPpourgéné-rer leur profil d’activité spécifiques sont diversesmais reposent sur des principes constants, enl’occurrence les expressions positivement corré-lées d’au moins 3 canaux ioniques différents(Figure3B).Larobustessereposeraitdoncsur laprésence de canaux fonctionnellement redon-dants ET sur la coordination de leurs niveauxd’expression. Ces expériences démontrent que,même si la robustesse de l’activité ne peut être
miseenévidencedirectementqu’en imposantdesperturbations d’activité à un neurone, ses princi-pes fondamentaux sont à l’œuvre de façon cons-tante dans la vie « normale » du neurone.
Conclusion et discussionLe système nerveux est un organe d’une fasci-
nante complexité, et notre capacité d’adaptation
Figure 3. Variabilité d’amplitude de courantset corrélation entre niveaux d’expression decanaux ioniquesdans lesneuronesde crustacé.A, haut, enregistrements de profils d’activité deneurones LP chez deux crabes (#1 en vert, #2 enorange) montrant la similarité des profils d’activité(superposition des traces, à droite). Bas, enregis-trements de courants potassiques montrant les dif-férences sensibles dans l’amplitude des courants,spécialement pour le courant BK-KCa et le courantIA qui sont respectivement 2 fois et 1.3 fois plusgrands chez les neurone LP#2 (orange). B, graphemontrant la forte corrélation positive dans lesniveaux d’expression des canaux potassiquesShaw et Shal et le canal cationique IH dans lesneuronesLPdedifférents crabes.A, d’aprèsSchulzet al. (2006) ; B, d’après Schulz et al. (2007).
GRP : fyssen JOB : annales27Internet DIV : 10mp⊕GOAILLARD⊕JeanMarc p. 5 folio : 96 --- 14/5/013 --- 15H14
NEUROBIOLOGIE
96

àdes environnements variés et changeants est engrande partie liée à la complexité et à la plasticitéde nos circuits neuronaux. Cependant, à tropmettre l’emphase sur la plasticité du systèmenerveux, nous risquons d’oublier que la plasticitéet la robustesse biologique ne sont pas l’apanagedes seuls systèmes nerveux complexes d’organis-mes pluricellulaires, mais représentent plus pro-bablement des propriétés structurelles commu-nes à tous les systèmes biologiques, de la levureà l’organisme animal le plus complexe. Et il estd’une certaine manière tout à fait étonnant quel’étude des principes de la robustesse de l’activitédes neurones n’ait reçu que peu d’attentionjusqu’à présent, et que les principes de la robus-tesse biologique aient été essentiellement définisà partir d’organismes très simples comme lalevure ou le vers annélideCaenorhabditis elegans.Les résultats présentés dans cet article démon-trent clairement que la fonction neuronalerepose sur des mécanismes robustes (la coordi-nation d’expression de protéines fonctionnelle-ment redondantes) qui sont à la base de l’adap-tation de l’activité des neurones aux mutationsgénétiques ou aux variations de l’environne-ment chimique. Mais ces résultats montrent éga-lement la nécessité d’adopter une approche sys-témique dans l’étude de la fonction neuronale oùde multiples composants (ici des canaux ioni-ques) sont analysés de manière simultanée pourcomprendre les principes émergeant de leursinteractions complexes. Cela implique donc demodifier non seulement nos approches expéri-mentales, notamment en intégrant la modélisa-tion mathématique et informatique, mais égale-ment notre perception de la fonction neuronalepour la considérer comme une propriété émer-geantde la coordinationdemultiples composantsaux propriétés complexes.
e Davis GW. Homeostatic control of neuralactivity: from phenomenology to moleculardesign. Annu Rev Neurosci 29: 307-323, 2006.
e Klassen T, Davis C, Goldma, A, Burgess D,Chen T, Wheeler D, McPherson J, Bourquin T,Lewis L, VillasanaD,MorganM,MuznyD,GibbsR, Noebels J. Exome sequencing of ion channelgenes reveals complex profiles confounding per-sonal risk assessment in epilepsy.Cell, 145: 1036-1048, 2011.
e MacLeanJN,ZhangY,GoeritzML,CaseyR,Oliva R, Guckenheimer J, and Harris-Warrick
RM. Activity-independent coregulation of IA andIh in rhythmically active neurons. JNeurophysiol94: 3601-3617, 2005.
e Nerbonne JM, Gerber BR, Norris A, andBurkhalter A. Electrical remodelling maintainsfiring properties in cortical pyramidal neuronslacking KCND2-encoded A-type K+ currents.J Physiol 586: 1565-1579, 2008.
e Schulz DJ, Goaillard JM, and Marder E.Variable channel expression in identified singleand electrically coupled neurons in different ani-mals. Nat Neurosci 9: 356-362, 2006.
e Schulz DJ, Goaillard JM, and Marder E.Quantitative expression profiling of identifiedneurons reveals cell-specific constraints onhighly variable levels of gene expression. ProcNatl Acad Sci U S A 104: 13187-13191, 2007.
e Swensen AM, and Bean BP. Robustness ofburst firing in dissociated purkinje neurons withacute or long-term reductions in sodium conduc-tance. J Neurosci 25: 3509-3520, 2005.
e Wagner A. Robustness and evolvability inliving systems. Princeton, N.J.; Woodstock: Prin-ceton University Press, 2005, p. 1 v.
IntroductionOne of the most fascinating properties of com-
plexbiological systems is their ability tomaintaina stable function in spite of the numerous inter-nal and external perturbations they are conti-nuously facing. This property, central to the sur-vival of biological organisms, has been namedbiological robustness. Biological robustness hasbeen observed at all levels of biological com-plexity, suggesting that it represents an evolutio-narily-selected structural property of biologicalsystems (Wagner 2005).
Biological robustness reflects the existence ofmultiple solutions toeachbiological problem.Forinstance, at the genetic level, each amino acid isencoded by multiple triplets of nucleotides, aphenomenon named «degeneracy» of the geneticcode. And when random mutations induce thesubstitution of an amino acid for a different one,the structure and function of the encoded proteinare rarely altered. At a different level, numerousstudies have demonstrated that the developmentof the vulva in Caenorhabditis elegans is extre-mely robust to the deletion of several progenitorcells, and that multiple differentiation pathways
GRP : fyssen JOB : annales27Internet DIV : 10mp⊕GOAILLARD⊕JeanMarc p. 6 folio : 97 --- 14/5/013 --- 15H14
NEUROBIOLOGY
97

are leading to a functional organ. Therefore theexistence of multiple solutions relies on somedegree of redundancy between the componentsof a biological system, such that the loss or themodification of one of these components can becompensatedby functionally-overlapping compo-nents.
Robustness in the nervous systemThenervous system, in particular themamma-
lian brain, with its 1010 neurons connected by1014 synapses, is one of the most complex biolo-gical systems. Unsurprisingly, our growingknowledge of its function has revealed that themammalian brain is highly robust to perturba-tions. This robustness can be observed at diffe-rent levels. One of the most striking examples isthe ability of the nervous system to recover(sometimes virtually completely) from variousinjuries irreversibly damaging neuronal circuits,such as strokes or brain tumors. In these cases,robustnesshasoftenbeenmixedwith theconceptof plasticity. But the nervous system robustnessis also visible at the molecular and cellular level,not only at the network level.
The electrical activity of neurons is due to theactivation of ion channel proteins conducing ions(sodium, potassium, calcium, chloride) acrossthe plasma membrane. The depolarizing ionchannels (conducing sodium and/or calcium)tend to promote neuronal activity while hyperpo-larizing ion channels (potassium, chloride) tendto inhibit neuronal activity. Neurons express amultitude of ion channels with different biophy-sical propeties at the level of their cell body andprocesses, which underlie the genesis of electri-cal activity in an autonomous way or in responseto neurotransmitters released at the synapses.
Eachneuronal type express a specific set of ionchannels associated with its specific pattern ofactivity: cortical pyramidal neurons do not dis-play the same pattern of activity or set of ionchannelsasspinal cordmotorneurons.Thereforea crucial question is to understand how neuronsof a given type maintain a stable of pattern ofactivity over years (or decades), while submittedto strong variations of their chemical environ-ment, number of synaptic connections, etc...
Homeostatic plasticity of neuronal activityOver the past twenty years, a number of stu-
dies have demonstrated that neurons are able to
adjust their electrical properties in response tothe electrical stimulation they receive. For ins-tance, when neurons are deprived of activity (byapplication of specific drugs or following sensorydeprivation), the number of depolarizing ionchannels is upregulated while the number ofhyperpolarizing ion channels is downregulated,such that the overall level of activity remainsunaltered. The opposite regulation occurs whenneurons are submitted to artificial increases inactivity, which induce an downregulation and anupregulation of the number of depolarizing andhyperpolarizing ion channels, respectively. Thisability to dynamically regulate the properties orthe number of ion channels to maintain a stablelevel of activity has been named homeostaticplasticity (from the ancient greek Ömoıov,homoios, ”similar“ and Ïstcmı, histomi, «immo-bile») (Davis 2006). Homeostatic plasticity repre-sents one aspect of the robustness of neuronalactivity, i.e. robustness to external perturba-tions. Although the binary regulation of the num-ber of ion channels (increase vs decrease) canexplainhowneuronsmaintain their average levelof activity, it is difficult to understand how aprecise and specific pattern of activity can bemaintained by such basic mechanisms. Maintai-ning a specific activity pattern most likely requi-res precise coordinated regulation of the proper-ties of ion channels involved in a same function.
Functional redundancy between ionchannels: manipulation
It is known that each neuron expresses farmore than the minimal set of ion channels theo-retically required to generate its activity pattern.TheexperimentsperformedonKOmice (knockedout, transgenic mice in which a specific gene hasbeen inactivated) demonstrate that the functio-nal redundancy between ion channels enablesneurons to maintain their pattern of activity inspite of the loss of one of their ion channels(internal perturbation). For instance, Swensenand Bean (2005) have demonstrated that thecerebellum Purkinje neurons are still able togenerateburstsof actionpotentials afterdeletionof a sodium channel (Nav1.6) involved in thegenesis of these bursts of activity (Figure 1A):while the application of a neurotoxin specific forthis ion channel strongly alters the activity of theneuron, the chronic loss of the ion channel
GRP : fyssen JOB : annales27Internet DIV : 10mp⊕GOAILLARD⊕JeanMarc p. 7 folio : 98 --- 14/5/013 --- 15H13
NEUROBIOLOGY
98

(Nav1.6 KO mouse or Nav1.6-/-) only slightlymodifies it.
A similar study has shown that the pyramidalneurons of the cortex are able to maintain theiractivity profile after deletionof apotassiumchan-
nel (Kv4.2) thought to play a critical role in thisneuronal type. In this case, the authors were ableto demonstrate that the loss of theKv4.2 channel(and therefore of the ion current it generates) iscompensated by an increase in the amplitude ofother potassium currents (such that the algebricsum of the total potassium current is maintainedconstant), enabling the neuron to maintain itsprecise activity profile (Nerbonne et al. 2008)(Figure1B).This studyclearlydemonstrates theexistence of functional redundancy between dis-tinct potassium channels (and the currents theygenerate), andhow this redundancy is usedwhenone of the ion channels is lost.
So far however, only one study precisely quan-tified the changes in ion channel properties asso-ciated with the stability of activity patterns. Inthisstudyperformedonthespiny lobsternervoussystem, the authors artificially increased theamplitude of a potassium current (carried by theion channel Shal) bymicroinjecting the ion chan-nel mRNA in a motorneuron spontaneously pro-ducing bursts of action potentials. In spite of thismicroinjection and the associated large increasein the amplitude of the potassium current, thepattern of activity of the motorneuron remainsunchanged (MacLean et al. 2005) (Figure 2A).The increase in amplitude of the potassium cur-rent (hyperpolarizing) is in fact compensated bythe increase in amplitude of a depolarizing cur-rent named IH, such that the activity of the neu-ron is stabilized.
Thus this case of functional redundancy isopposite to the one presented previously (Ner-bonne et al., 2008) (Figure1B), since the two ionchannels involved have an opposite effect onneuronal activity, and the stability of activityrelies on a constant ratio between the amplitudesof these two currents (Figure 2B).
The three above cases clearly demonstrate theexistence of functional redundancy between ionchannels and its role in the emergence of robus-tness of neuronal activity. It is easy to imaginethat this type of mechanisms is not only involvedin the response to experimentalmanipulations ofion channel expression (such as KO or microin-jection of mRNA), but could also be involved inthe response to spontaneous (and natural) muta-tions altering the function of ion channels. Eventhoughmost of the genes associated with geneticforms of epilepsy encode ion channel proteins, it
Figure 1. Stability of the activity pattern ofneurons after deletion of an ion channel. A, thebursts of action potentials recorded in Purkinjeneurons after deletion of the Nav1.6 sodiumchannel (Nav1.6 -/- mice, blue traces) are verysimilar to those recorded in the Nav1.6 +/+ mice(purple traces). The top andmiddle traces show thelevel of similarity in the general shape of the bursts,the histograms (bottom row) demonstrate that thenumber of action potentials during the burst is alsovery similar in both cases. B, the action potentialrecorded in cortical pyramidal neurons afterdeletion of the Kv4.2 potassium channel (Kv4.2 -/-mice) has a shape identical to the action potentialrecorded in Kv4.2 +/+ mice. A, from Swensen andBean (2005); B, from Nerbonne et al. (2008).
GRP : fyssen JOB : annales27Internet DIV : 10mp⊕GOAILLARD⊕JeanMarc p. 8 folio : 99 --- 14/5/013 --- 15H15
NEUROBIOLOGY
99

is likely that a significant number of ion channelmutations have been undetected because inmostcases the functional redundancy between ionchannels provides a virtually perfect compensa-tion of the defect produced by the mutation. Arecent clinical survey indeed demonstrated thatthe mutations in ion channel genes are wides-pread, not only in epileptic patients, but also incontrol subjects, suggesting that most mutationshave no effect on the phenotype, thereforeimplying that the targeted genetic screening isunable to predict the risk to develop the disease(Klassen et al., 2011).
Functional redundancy between ionchannels: observation
If robustness is a structural property of biolo-gical systems, the mechanisms underlying itshould be observable in unperturbed neurons.The functional redundancy between themultipleion channels expressed by each neuron potenti-ally gives rise to a staggeringnumber of solutions
to generate and maintain a specific pattern ofactivity. Therefore, the question arises whetherneurons indeed use variable solutions to gene-rate a same activity pattern, because it woulddemonstrate that neurons in «natural» unpertur-bed populations relie on robust principles togenerate their activity.
Studies performed on the crab nervous systemtend to demonstrate that neurons of the sametype, sometimes presenting virtually identicalactivity patterns, use highly variable solutions interms of channel expression, although constantprinciples aremaintained. For instance, the elec-trophysiological recordings of three distinctpotassium currents in the lateral pyloric (LP)neuron revealed significant differences in theiramplitude (3 to 4-fold) from one LP to another,even when considering neurons with super-imposable activity patterns (Schulz et al. 2006)(Figure 3A). Two explanations can be given inresponsetothisobservation: i)neuronsare insen-sitive to the differences in ampitude of these cur-rents, and these variations do not affect neuronalactivity, or ii) the observed variability is associa-ted with the co-variation of functionally redun-dant ion currents (synergistic or antagonistic).Experiments of pharmacological blockade of ionchannels demonstrate that neurons are sensitiveto changes in current amplitude of lower magni-tude (50%blockade). The study fromMcLeanandcolleagues previously cited (where the variationin expression of the Shal channel triggers a com-pensatory change in the amplitude of the IH cur-rent) however suggests that some ion channelsmight be co-regulated in order to maintain a sta-ble pattern of activity. Indeed, a later study per-formed on several types of neurons of the crabnervous system confirmed this hypothesis. Ana-lyzing the levels of expression of 6 distinct ionchannels in 6 different cell types revealed a largenumber of correlations in the levels of expressionof several ion channels. For instance, the levels ofexpression of theShal,Shaw and IH (responsiblefor the IH current) ion channels were found to bestrongly positively correlated in the LP neuron(Schulz et al. 2007) (Figure 3B). Thus the varia-bility in amplitude of potassium currents in theLP neuron could be compensated by the coordi-nated variations in other currents, for examplethe IH channel, which would counteract theeffects of the Shal channel (MacLean et al. 2005).
Figure 2. Compensation of the effect of Shalchannelmicroinjectionby co-variationof the IHcurrent. A, the spontaneous bursts of actionpotentials generated by the spiny lobster pyloricneuron are not altered after microinjection of ShalmRNA, although the current carried by thischannel (IA current) is significantly increased (notshown). B, the increase in amplitude of IA isassociated with a coordinated increase in theamplitude of IH, which counteracts the effect ofIA on neuronal activity, therefore stabilizing thepyloric neuron activity pattern. From MacLean etal. (2003).
GRP : fyssen JOB : annales27Internet DIV : 10mp⊕GOAILLARD⊕JeanMarc p. 9 folio : 100 --- 14/5/013 --- 15H16
NEUROBIOLOGY
100

Therefore, theLPneuronsproduce a stereotypedpattern of activity not because they express a setof ion channels at a fixed level, but because theexpression levels of functionally redundant ionchannels are coordinately regulated. Thus, the
solutions used by the LP neurons to generatetheir activity profile are diverse but relie on cons-tant principles, i.e. the positively correlatedexpression of at least 3 distinct ion channels(Figure 3B). Robustness of neuronal activitywould therefore relie on the presence of functio-nally overlapping ion channels AND on the coor-dination of their level of expression. These expe-riments demonstrate that, although robustnessof neuronal activitymayonly bedemonstratedbydisturbing the activity of the neuron, its funda-mental principles are expressed during the «nor-mal» life of a neuron.
ConclusionsThe nervous system is a fascinatingly complex
organ, and our ability to adjust to ever changingenvironments ismost likelydue to the complexityand the plasticity of our neuronal networks.Nevertheless,much emphasis has been placed onneuronal plasticity, almost implying that plasti-city and robustness are specific to complexmulticellular organisms, while they most likelyrepresent structural properties common to allbiological systems, from bacteria to humanbeings. It is in fact rather surprising that so littleattention has been paid to the principles of robus-tness of neuronal activity, and that those havebeen defined on very simple organisms such astheyeastor thenematodeCaenorhabditis elegans.The results presented in the current articleclearlydemonstrate thatneuronal function relieson robust mechanisms (coordinated expressionof functionally-overlapping proteins) that consti-tute the adaptive response of neurons to environ-mental and genetic perturbations. But theseresults also point out the need for a system-levelapproach of neuronal function where multiplecomponents (here ion channels) are investigatedsimultaneously in order to decypher the princi-plesarising fromtheir complex interactions.Thisimplies to not only modify our experimentalapproaches, for instance by combining mathe-matical and computer modeling, but also ourview of neuronal function to consider it as aproperty emerging from the tight coordination ofmultiple components with complex non-linearproperties.
Figure 3. Variability in the amplitude ofcurrents and correlation in the expressionlevels of ion channels in crustaceanneurons.A,top row, activity recordings of 2 LP neurons in2different crabs (#1 ingreen, #2 inorange) showingthe similarity in activity profiles (superimposedtraces are shown on the right). Bottom row,recordings of potassium currents showing thedifferences in amplitude, particularly for theBK-KCa and the IA currents, which are 2 and1.3 times larger, respectively, in the LP #2 neuron.B, scatter plot showing the strong positivecorrelation in the levels of expression of thepotassium ion channels Shaw and Shal and thecationic channel IH in the LP neurons fromdifferent crabs.A, fromSchulz et al. (2006);B, fromSchulz et al. (2007).
GRP : fyssen JOB : annales27Internet DIV : 10mp⊕GOAILLARD⊕JeanMarc p. 10 folio : 101 --- 14/5/013 --- 15H16
NEUROBIOLOGY
101

Les processus cognitifs sous jacents de l’imagerie motrice– capacité d’imagerie et mécanismes inhibiteurs
Florent LEBON
Maître de Conférences Université de Bourgogne, UFR STAPS INSERM U1093, Cognition, Action etPlasticité Sensimotrice
RésuméL’imagerie motrice est la simulation mentale d’un mouvement sans contraction musculaire
concomitante. La capacité à produire des images mentales vivaces a été mesurée par l’Indiced’Imagerie Motrice. Cet indice combine des mesures psychologiques, comportementales et psycho-physiologiques. L’activation des réseaux neuronaux pendant l’imageriemotrice a ensuite été évaluéepar la technique de stimulation magnétique transcrânienne et corrélée à la capacité d’imagerie. Lesbons imageurs activeraient spécifiquement ces réseaux neuronaux contrairement aux mauvaisimageurs.Enfin, la techniquededouble stimulationmagnétiqueapermisd’estimer l’interactionentreles cortexmoteur primaire et pariétal lors de l’imageriemotrice. Le lobe pariétal inférieur droit seraitainsi impliqué dans une boucle inhibitrice afin de prévenir de toutmouvement,malgré l’activation ducortex moteur primaire.
Mots-clésImagerie motrice, stimulation magnétique transcrânienne, capacité d’imagerie, mécanismes
inhibiteurs.
Cognitive processes underlying motor imagery –imagery ability and inhibitory mechanisms
AbstractMotor imagery is the mental simulation of movement without any motor output. The ability to
produce vivid mental images was measured thanks to the Motor Imagery Index, which combinedpsychological, behavioral andpsychophysiological recordings.Excitability ofneuronalnetworkswerethen assessed by transcranial magnetic stimulation (TMS) and correlated to imagery ability. Goodimagers activated more specifically the neuronal networks compared to poor imagers. Finally,interaction between parietal and primary motor cortices was investigated via image-guided dual-coilTMS during motor imagery. Results showed that right inferior parietal lobe forms part of a distinctinhibitory network that may prevent unwanted movement during imagery tasks.
KeywordsMotor imagery, transcranial magnetic stimulation, imagery ability, inhibitory mechanisms.
IntroductionL’imagerie motrice (IM) est la représentation
mentale d’une action sans production concomi-tante de mouvement (Jeannerod, 1994). Les étu-des d’imagerie par résonance magnétique fonc-tionnelle ont montré que l’exécution motrice, lapréparation d’un mouvement et l’imagerie de cemême geste activaient des régions corticales et
sous corticales similaires. Toutefois certainesaires sont spécifiquement activées lors de l’acti-vité mentale, prouvant que l’IM n’est pas simple-mentunecopiede l’exécutionmotrice impliquantles mêmes structures à un degré inférieur. C’estun phénomène à part entière engageant unréseau neuronal spécifique, qui est notammentimpliqué dans l’inhibition du signal destiné aux
GRP : fyssen JOB : annales27Internet DIV : 11mp⊕LEBON⊕Florent p. 1 folio : 102 --- 14/5/013 --- 15H19
NEUROBIOLOGIE
102

muscles. Toutefois, les réponses du systèmenerveux autonome ne sont, elles, pas inhibées etsont utilisées pour attester la qualité du travailmental (Collet et al., 1999). Par ailleurs, uncertain nombre d’études utilisant la stimula-tionmagnétique transcrânienneontmontré l’évi-dence que l’IM augmentait spécifiquement l’exci-tabilité du réseau neuronal, i.e. le niveau d’acti-vation entre le cortex moteur et les muscles. Unestimulation magnétique focale est appliquée auniveau du cortex moteur, activant les cellulessuperficielles du cerveau. La réponse motrice,communément appelée potentiel moteur évoqué(PME), est alors enregistrée au niveau des mus-cles concernés. L’amplitude du PME reflète ainsile niveau d’excitabilité du réseau neuronal.
En combinant les enregistrements des systè-mes nerveux central et autonome, ce projet avaitpour objectif d’approfondir la compréhensiondes mécanismes de l’IM, et plus généralementdes processus cognitifs. Le but était, dans unpremier temps, d’établir une possible relationentre les systèmes nerveux autonome et centrallors de la simulation mentale du mouvement,puis de définir les processus inhibiteurs de l’IMimpliqués dans la prévention du mouvement.Nous avons fait l’hypothèse que la capacité àformerdes imagesmentales vivacespourrait êtreassociée à une facilitation spécifique de l’excita-bilité neuronale et les mécanismes d’inhibitionpourraient être définis par une interaction entreles cortex moteur primaire et pariétal.
Trois études ont ainsi été réalisées. La pre-mière consistait à évaluer la capacité d’imagerieselon un indicateur combinant des enregistre-ments psychologiques, comportementaux et psy-chophysiologiques.Ladeuxièmeavait pourobjec-tif de mettre en relation l’activation du réseauneuronal évaluéeparstimulation transcrânienneavec laqualitéd’imagerie.Enfin la troisièmes’estattachée à évaluer l’importance de l’interactionentre les cortex moteur primaire et pariétal dansles processus de prévention du mouvement lorsde l’IM.
Expérience 1 : Evaluation de la qualitéd’imagerie
Afin d’évaluer l’habileté à produire des imagesmentales vivaces, plusieurs indicateurs peuventêtre utilisés, tels que des indices subjectifs (ques-tionnaires, estimation personnelle) et objectifs(fréquence cardiaque, activité électrodermale,
chronométrie mentale). Toutefois, la majoritédes études portant sur l’IM n’utilisent qu’un oudeux indices pour évaluer la qualité d’imagerie.Nous avons alors développé un modèle combi-nant desmesures psychologiques, comportemen-tales et psychophysiologiques : l’indice d’ima-gerie motrice, composé de 6 sous-indicateurs(Collet et al., 2011).– Dans un premier temps, le participant répon-dait au questionnaire révisé d’imageriemotrice (Hall & Martin, 1997). Ce test psycho-métrique, validé par la littérature, est composéde 8mouvements qui sont réellement exécutéspuis imaginés. Le participant attribuait unenote comprise entre 1 et 7 après chaque mou-vement imaginé. La note ’1’ représente uneimage floue et ’7’ une représentation mentalenette et précise. Les niveaux de 2 à 6 corres-pondent à des notes intermédiaires. Une noteest obtenue pour l’ensemble des mouvementsimaginés.
– Dans un deuxième temps, le participantmémo-risait une séquence d’opposition du pouce avecles autres doigts. Les 5 autres sous-indicateursétaient alors enregistrés lors de l’exécutionréelle et imaginée de cette séquence :
– Quantification subjective de la qualitéd’imagerie : après avoir imaginé chaqueséquence, le participant attribuait une notebasée sur la même échelle de 1 à 7 que l’indiceprécédent.Unenotemoyenne est obtenuepourl’ensemble des 12 mouvements imaginés.
– Chronométrie mentale : cet indicateur étaitcalculé sur la différence entre les durées dumouvement réel et imaginé, où l’isochroniecorrespondait à une imagerie de bonne qualité.L’équivalence temporelle est en effet un critèrefiable de l’habileté à imaginer. Un bouton pres-soir était placé dans la main gauche du parti-cipant (celle qui n’exécute pas le mouvement).Le participant appuyait sur le bouton de soninitiative afin de marquer le début et la fin dechaque séquence qu’il réalisait (mouvementréellement exécuté et imaginé).
– Conductance cutanée : Le niveau de conduc-tance cutanée enregistré au niveau des doigts(là où l’activité des glandes sudorales est la plusimportante) était ainsi utilisé comme indica-teur physiologique reflétant quantitativementle niveau d’activation durant une tâche. Unediminution de la conductance cutanée est le
GRP : fyssen JOB : annales27Internet DIV : 11mp⊕LEBON⊕Florent p. 2 folio : 103 --- 14/5/013 --- 15H19
NEUROBIOLOGIE
103

reflet d’une baisse du niveau d’activation prou-vant que le participant était trop décontracté.Au contraire, aucune différence avant et aprèsl’essai imaginé montre que le participant étaitimpliqué dans la tâche mentale.
– Durée de perturbation ohmique : cette autremesure de la conductance cutanée était utiliséecomme indicateur du niveau d’attention. Cetindice reflète le temps durant lequel l’informa-tionest traitéeetanalyséeauniveaucentral.Ladurée de perturbation ohmique débute lorsquela courbe de conductance cutanée augmentebrusquement et termine lorsque celle-ciretrouve sa forme initiale. Une durée équiva-lente durant l’IM et l’exécution réelle attested’une bonne qualité d’imagerie.
– Arythmie sinusale respiratoire : ce dernierindicateur est l’amplitudemoyenne de la varia-bilité de la fréquence cardiaque instantanée.Une diminution significative de l’arythmiesinusale respiratoire est typiquement observéelorsque le participant est impliqué dans unetâche cognitive. Cet indice est considérécomme une mesure fiable de l’effort cognitifdurant l’imagerie motrice.L’ensemble des indices étaient ensuite combi-
nés et le score d’imagerie motrice calculé pourclasser selon leur capacité d’imagerie les vingt-trois personnes participant à cette étude. Lescore moyen était de 5.65 (± 0.17), avec un mini-mum de 3.77 et un maximum de 6.96. Les douzeparticipants avec le plus haut score étaient consi-dérés comme bons imageurs, tandis que les onzeautres comme des mauvais imageurs. Les figu-res 1 et 2 représentent les enregistrements desvariables chronométriques et psychophysiologi-ques (indices 3 à 6) de deuxparticipants. Basé surle score d’imagerie motrice, le premier est consi-déré comme un bon imageur et le second commeun mauvais imageur.
Expérience 2 : Excitabilité neuronaleet qualité d’imagerie motrice
Actuellement, la question de savoir com-ment les activations neurophysiologiques sous-jacentes de l’activité mentale reflètent la qualitéde l’imagerie n’a pas encore été résolue. La plu-part des études évaluent la capacité d’IM pardes questionnaires subjectifs. Jusqu’à présent,seuls Guillot et collaborateurs (2008) ont com-biné l’imagerie par résonancemagnétique (IRM)fonctionnelle avec des mesures psychophysiolo-
giques, comportementales et psychologiquespour associer l’activité corticale à la capacitéd’IM. Les bons imageurs activaient de manièreplus importante les régions pariétales et prémo-trices ventrolatérales, connuespour jouerun rôleprépondérant dans la formation des imagesmen-tales. Les mauvais imageurs recrutaient princi-palement le cervelet et les cortex orbitofrontal et
Figure 1 : Indice d’Imagerie Motrice d’un bonimageur.Le score de ce participant était de 6.68 enajoutant le score des 6 sous-indicateurs. A) Enre-gistrement de la conductance cutanée (mS) lorsd’une séquence d’opposition de doigts réellementexécutée et imaginée et d’un temps de repos (tracédu haut) et enregistrement de la durée de ces3 tâches (tracé du bas). Les lignes verticalespointillées représentent la durée de perturbationohmique (DPO) en secondes, indice mesurée à par-tir de la conductance cutanée. La DPO pendantl’imagerie motrice (IM) était similaire à la DPO dumouvement exécuté, montrant un niveau d’atten-tion élevé. Les barres verticales solides repré-sentent le début et lafinde chaque tâche.Lesduréesdes mouvements réellement exécuté et imaginéétaient similaires, montrant une équivalencetemporelle. B) Fréquence cardiaque instantanéependant l’IMet le repos. L’IMa induit une variationde l’arythmie sinusale respiratoire, observée parla diminution de la variabilité de la fréquencecardiaque instantanée. IM = imagerie motrice,mS = micro Siemens, s = seconde.
GRP : fyssen JOB : annales27Internet DIV : 11mp⊕LEBON⊕Florent p. 3 folio : 104 --- 14/5/013 --- 15H19
NEUROBIOLOGIE
104

cingulaire postérieur, impliqués dans l’appren-tissage moteur. Toutefois, aucune étude n’adémontré comment les caractéristiques del’image mentale reflétaient les activations dusystème nerveux, particulièrement au niveau ducortex moteur primaire (M1). Notre hypothèseest que la qualité d’imagerie aurait une influencesur les processus neurophysiologiques évalués
par stimulation magnétique transcrânienne. Laprésente étude s’est attachée à démontrer com-ment les bons et mauvais imageurs activaient demanière préférentielle le réseau neuronal lors del’imagerie d’une tâche simple.
La tâche consistait à taper physiquement oumentalement le pouce de la main droite sur unesurface plane, en rythme avec un métronome(fréquence de 1 Hz). Ce type de mouvementsimple permet de cibler précisément le muscleimpliqué dans la contraction : lemuscle opposantdu pouce. Des stimulations magnétiques trans-crâniennes étaient appliquées au niveau deM1 etplus particulièrement au niveau de la représen-tation corticale du muscle opposant du pouce.Les réponses motrices évoquées étaient alorsenregistrées par électromyographie auniveaudumuscle opposant du pouce mais également dumuscle court abducteur du petit doigt afin d’éva-luer la spécificité spatiale de l’IM. Seule uneaugmentation de la réponse devrait être enregis-trée pour lemuscle impliqué dans lemouvement.De plus, les stimulations étaient déclenchées enrythme avec le métronome ou entre 2 bips afind’évaluer la spécificité temporelle de l’IM. Seuleune augmentation de la réponse devrait êtreenregistrée au moment du bip, comme lors del’exécution réelle où le muscle est activé aumoment du bip et relâché entre 2 bips.
Augmentation de l’excitabilité neuronaleLes résultats confirment ceux des études pré-
cédentes (pour revue, voir Stinear, 2011). Eneffet, l’amplitude de la réponse évoquée lors del’IM était plus importante que celle enregistréeau repos, l’ensemble des participants étantregroupé. Cecimontre une activation plus impor-tante des neurones pyramidaux au sein deM1 lors de l’IM. De plus, cette excitabilité neuro-nale était modulée spatialement et temporelle-ment, c’est-à-dire plus importante pour le muscleimpliqué dans le mouvement (modulation spa-tiale) et lorsque le participant imaginait le mou-vement en rythme avec le métronome (modula-tion temporelle).
Modulation spatiale et temporelle selon lacapacité d’imagerieUne analyse de variance à mesures répétées a
permis de comparer l’amplitude des réponsesévoquées en fonction des groupes d’imageurs.Les résultats montrent que les bons imageursactivaient de façon spécifique les cellules cortica-
Figure 2 : Indice d’Imagerie Motrice d’unmauvais imageur. Le score pour ce participantétait de 4.37 en ajoutant le score des 6 sous-indicateurs. A) Enregistrement de la conductancecutanée (mS) lors d’une séquence d’opposition dedoigts réellement exécutée et imaginée et d’untempsde repos (tracé duhaut) et enregistrement dela durée de ces 3 tâches (tracé du bas). Les lignesverticales pointillées représentent la durée deperturbation ohmique (DPO) en secondes, indicemesuré à partir de la conductance cutanée. La DPOpendant l’imageriemotrice (IM) est plus courte quecelle du mouvement exécuté, montrant une faiblecapacité d’attention. Les barres verticales solidesdans la partie inférieure représentent le début et lafin de chaque tâche. La durée du mouvementimaginé est plus importante que celle du mouve-ment réellement exécuté, montrant une faibleisochronie. B) Fréquence cardiaque instantanéependant l’IM et le repos. Il n’y a pas de diminutionde l’amplitude de la fréquence cardiaque ins-tantanée lors de l’IM, montrant que la tâchementale n’a induit aucune variation de l’arythmiesinusale respiratoire. IM = imagerie motrice,mS = micro Siemens, s = seconde.
GRP : fyssen JOB : annales27Internet DIV : 11mp⊕LEBON⊕Florent p. 4 folio : 105 --- 14/5/013 --- 15H19
NEUROBIOLOGIE
105

les contrôlant le muscle impliqué dans la tâchemotrice tandis que les mauvais imageurs acti-vaient le cortexmoteur demanière plus générale,plusdiffuse.Deplus, l’excitabiliténeuronale étaitplus importante au moment d’imaginer le mou-vement, c’est-à-dire en rythme avec le métro-nome, dans les 2 muscles chez les mauvais ima-geurs. Au contraire, cette différence temporellen’était visible quedans lemuscle impliquédans lemouvement chez les bons imageurs. La modula-tion spatio-temporelle de l’IM serait ainsi asso-ciée à la capacité d’imagerie.
Expérience 3 : Processus inhibiteursde l’imagerie motrice
Les études d’IRM fonctionnelle montrent uneactivation corticale et sous-corticale étendue lorsde l’IM, incluant notamment les aires motricesprimaires et secondaires et pariétales (de Langeet al., 2005). De façon intéressante, les aires del’exécution sont activées alors qu’aucun mouve-ment n’est produit durant l’IM. Un circuit inhi-biteur serait ainsi mis en place pour prévenir detoute contraction musculaire. Nous avons émisl’hypothèse que le cortex pariétal était impliquédans ce circuit inhibiteur car son activation aété systématiquement rapportée pendant l’IM.De plus, des sujets atteints de traumatismes au
niveau du cortex pariétal ont des difficultés pourprédire le temps nécessaire à effectuer un mou-vement (Sirigu et al., 1996) ou pour prévenir detout mouvement lors de l’IM (Schwoebel etal., 2002). Le but de cette troisième étude étaitalors d’établir un lien causal entre le lobe pariétalet M1 par la technique d’une double stimulationmagnétique transcrânienne. La tâche à effectuerréellement ou à imaginer était unmouvement depinçage entre l’index et le pouce de lamain droiteen rythme avec un métronome (fréquence de1Hz).Lemuscleprincipalement étudié était celuide l’index de la main dominante (droite).
Double stimulation magnétique transcrâ-nienne neuronaviguéeUne représentation 3D du cerveau de chaque
participant était créée à partir d’une image ana-tomique obtenue par IRM. Les 2 sites de stimu-lation étaient ainsi précisément déterminés :1) le lobe pariétal inférieur droit dont les coor-données stéréotaxiques étaient déterminées surla base d’études IRM (par exemple, de Lange etal., 2005) et 2) la représentation corticale dumuscle de l’index droit au niveau de M1 gauche.Le positionnement des deux bobines de stimula-tion était alors repéré sur l’image 3D, facilitantune stimulation précise et constante (Figure 4).La technique de double stimulation neuronavi-guée a permis ainsi d’évaluer l’interaction entreces 2 aires corticales lors de l’IM.
Deux types de stimulation étaient délivrés :1) une simple stimulation deM1 gauche et 2) unedouble stimulation correspondant à une stimula-tion au niveau du cortex pariétal envoyée 6 ou12ms avant celle deM1. Le ratio, correspondantà l’amplitude de la réponse évoquée suite à ladouble stimulation par l’amplitude de la réponseévoquée suite à la stimulation simple de M1, apermis de mesurer l’effet du cortex pariétal surM1 lors des tâches de repos et d’IM.
Interaction des cortex moteur primaire etpariétal inférieurLes stimulations consécutives du lobe pariétal
inférieur droit et de M1 gauche montraient unediminution de l’amplitude des réponses lors del’IM, comparativement à une stimulation simple.Au contraire la double stimulation augmentaitl’amplitude des réponses au repos. L’activationdu cortex pariétal inférieur pendant l’IM auraitainsi un effet inhibiteur sur M1 afin d’éviter toutmouvement. Toutefois, cette inhibition n’était
Figure 3 : Somme normalisée des potentielsmoteurs évoqués (PME) lors de l’imageriemotriceen fonctionde laqualitéd’imagerie. Lasomme a été calculée en additionnant l’amplitudemoyenne des PME induits par des stimulationsd’intensité croissante. La somme calculée lorsde l’IM a été normalisée par celle de la conditionde repos. Lorsque les participants étaient grou-pés selon leur capacité d’imagerie, la spécificitéspatio-temporelle n’était visible que chez les bonsimageurs. OP = Opposant du pouce, CAPD = CourtAbducteur du Petit Doigt, ON = stimulationmagnétique en rythme avec le métronome,OFF = stimulation entre les bips du métronome,* = P S 0.05.
GRP : fyssen JOB : annales27Internet DIV : 11mp⊕LEBON⊕Florent p. 5 folio : 106 --- 14/5/013 --- 15H19
NEUROBIOLOGIE
106

visible qu’à un intervalle de stimulation long(12 ms) et non court (6 ms) (Figure 5). Parconséquent, l’interaction entre cortex pariétal etmoteur primaire serait indirecte, impliquant plu-sieurs connexions transsynaptiques du fait de ceplus long intervalle.
Ce résultat explique d’un point de vue neuro-physiologique les mécanismes inhibiteurs impli-qués dans la prévention du mouvement lors del’IM. Toutefois d’autres aires corticales et sous-corticales, tels que le cervelet, semblent fairepartie de ce processus. Des recherches addition-nelles, utilisant la stimulationmagnétique trans-crânienne associée à d’autres techniques d’enre-gistrement et de stimulation, permettraient dedéterminer le réseau neuronal complexe sous-jacent de l’IM.
ConclusionCe projet post-doctoral s’est attaché à appro-
fondir la compréhension des mécanismes neuro-physiologiques sous-jacents de l’IM. Les princi-paux résultats valident l’Indice d’ImagerieMotrice (Collet et al., 2011) et établissent un liendirect entre les systèmes nerveux central et auto-nome. De façon intéressante, le cortex moteur
primaire est à la fois activé et inhibé durant lasimulation mentale du mouvement. Le cortexpariétal inférieur joue, quant à lui, un rôle indi-rect important dans l’inhibition du signal et doncdans la prévention du mouvement. Cette recher-che fondamentale et ses résultats pourraient êtreutilisés pour des applications pratiques. En effet,il est établi que l’IM, intégrée à des programmesde réhabilitation, peut améliorer la performancemotrice après des immobilisations, des trau-matismes corticaux ou encore des blessures péri-phériques. Il serait intéressant de savoir si le
Figure 4 : Technique de double stimulationneuronaviguée. Les représentations 3D du cer-veau d’un participant (haut) étaient créées à partirde son image IRM. Les croix jaunes représententl’orientation des bobines de stimulation magné-tique transcrânienne au niveau du cortex pariétalinférieur droit (haut gauche) et du cortex moteurprimaire gauche (haut droit). Les cibles du basindiquent la position de chaque bobine en direct enfonctiondusitede stimulationet facilitaientdonc lastimulation précise et constante.
Figure 5 : Potentiels moteurs évoqués (PME)conditionnés et non-conditionnés. Les PMEnon-conditionnés (NC) correspondent à la réponseenregistrée au niveau du muscle de l’index de lamaindroite suiteàunesimple stimulationducortexmoteur primaire gauche. Les PME conditionnésreflètent, quant à eux, la réponse enregistrée suiteà une stimulation du lobe pariétal droit suivie d’unestimulation du cortex moteur primaire gauche(double stimulation). Les 2 stimulations sont sépa-réesparunedurée, appelée intervalle interstimulus(IIS), de 6 ou 12 ms. Les tracés gris correspondentaux enregistrements de chaque participant et lestracés noirs auxmoyennes. L’amplitude des répon-ses non-conditionnées était plus importante lors del’imagerie motrice (IM) que lors du repos. Cerésultat montre une augmentation de l’excitabiliténeuronale lors de l’IM. Au contraire, l’amplitude dela réponse était diminuée uniquement lorsque lelobe pariétal était stimulé 12 ms avant le cortexmoteurprimairependant l’IM.Lecortexmoteurestà la fois activé et inhibé pendant la simulationmentale du mouvement et le lobe pariétal joueraitalors un rôle prépondérant dans la prévention dumouvement pendant l’IM.
GRP : fyssen JOB : annales27Internet DIV : 11mp⊕LEBON⊕Florent p. 6 folio : 107 --- 14/5/013 --- 15H19
NEUROBIOLOGIE
107

fait d’améliorer la capacité d’imagerie d’une per-sonne amplifie d’autant plus les effets du travailmental et si l’inhibition du mouvement est unfacteur prépondérant à la performance mentale.
RemerciementsJe voudrais remercier la Fondation Fyssen
et les membres de son comité de m’avoir donnél’opportunité de vivre cette expérience post-doc-torale ; Cathy Stinear et Winston Byblow del’Université d’Auckland (Nouvelle-Zélande) pourleur accueil et leur soutien tout au long de ceprojet ; Christian Collet et Aymeric Guillot del’Université de Lyon pour leurs conseils et lesnombreux échanges ; Martin Lotze de l’Univer-sité de Greifswald (Allemagne) pour sa collabo-ration dans la troisième étude ; et l’équipe duCentre de Recherche sur le Cerveau ainsi quel’équipe du laboratoire de neuroscience du mou-vement de l’Université d’Auckland (NZ).
Bibliographiee Collet C, Dittmar A, & Vernet-Maury E
(1999). Programming or inhibiting action : auto-nomicnervous systemcontrol of anticipation. IntJ Psychophysiol, 32, 261-276.
e Collet C, Guillot A, Lebon F, MacIntyre T& Moran A (2011). Measuring Motor Imageryusing Psychometric, Behavioral and Psychophy-siological Tools. Exerc Sport Sci Rev, 39, 85-92.
e de Lange FP, Hagoort P, & Toni I (2005).Neural topography and content of movementrepresentations. J Cogn Neurosci, 17, 97-112.
e Guillot A, Collet C, Nguyen VA, Malouin F,RichardsC&Doyon J (2008). Functional neuroa-natomical networks associated with expertise inmotor imagery. Neuroimage, 41, 1471-1483.
e Hall C R & Martin KA (1997). Measuringmovement imagery abilities : a revision of themovement imagery questionnaire. J Ment Imag,21, 143-154.
e Jeannerod M (1994). The representingbrain : neural correlates of motor intention andimagery. Behav Brain Sci, 17, 187-245.
e Schwoebel J, Boronat CB, & Branch CoslettH (2002). The man who executed ”imagined“movements : evidence for dissociable compo-nents of the body schema. Brain Cogn,50,1-16.
e Sirigu A, Duhamel JR, Cohen L, Pillon B,Dubois B, & Agid Y (1996). The mental represen-
tation of hand movements after parietal cortexdamage. Science, 273, 1564-1568.
e Stinear CM (2010). Corticospinal facilita-tion during motor imagery. In Guillot & Collet(Eds.),The neurophysiological foundations ofmen-tal and motor imagery. Oxford University Press,pp. 47-61.
IntroductionMotor imagery (MI) is the mental representa-
tion of an actionwithout anymotor output (Jean-nerod, 1994). Research in functional magneticresonance imaging showed thatmotor execution,movement preparation andmotor imagery sharecommon activation of cortical and sub-corticalareas. However, some activation are specific toMI showing that the mental simulation of anaction is not just a copy of motor executioninvolving the same organisation at a lower level.MI engages a specific neuronal network thatinhibits the signal that is sent tomuscles, inorderto prevent from moving. However, responsesfrom the autonomic nervous system are not inhi-bited, justifying the quality of the mental load(Collet et al., 1999). Furthermore, several studiesusing transcranial magnetic stimulation (TMS)showed that MI increased specifically the excita-bility of the neuronal network, i.e. the level ofactivation between the primary motor cortex(M1) and muscles. A focal magnetic stimulationis delivered over M1, activating neurons under-neath and inducing a motor evoked potential(MEP) in the corresponding muscle. The ampli-tude of the MEP reflects the level of neuronalexcitability.
By combining autonomic and central nervoussystem recordings, this project aimed to unders-tand the mechanisms underlying MI, and moregenerally cognitive processes. The purpose wasto determine a relationship between central andautonomic nervous systems while imagining,then to assess the inhibitory processes involvedinmovement prevention duringMI.Wehypothe-sised that one’s ability to form vivid mentalimages would be associated to a specific facilita-tion of the neuronal excitability and that theinteraction between M1 and the parietal cortexwould explain in part the inhibitorymechanisms.
Three experiments were performed to answerthese questions. The first one consisted in asses-sing the imagery ability while combining psycho-
GRP : fyssen JOB : annales27Internet DIV : 11mp⊕LEBON⊕Florent p. 7 folio : 108 --- 14/5/013 --- 15H19
NEUROBIOLOGY
108

logical, behavioural and psychophysiologicalmeasurements. The second one aimed to corre-late imagery ability to neuronal excitability.Finally, the thirdexperimentassessed the level ofinteraction between M1 and the parietal cortexinvolved in movement prevention during M1.
Experiment 1: Imagery ability assessmentTo assess one’s ability to imagine vividly a
movement, several indicators can be used, thatare subjective (questionnaires, self-estimation)and objective (heart rate, electrodermal activity,mental chronometry) measurements. However,most MI studies used just one or two of theseindicators to assess the imagery ability.We com-bined psychological, behavioural and psychophy-siological measurements to obtain the MotorImagery Index (Collet et al., 2011),made of 6 sub-indicators:– First, the participant completed the revisedversion of themovement imagery question-naire (Hall & Martin, 1997). This reliable testis made of 8 movements that are actually per-formed, and then mentally simulated. The par-ticipant rated each imagined movement from1 to 7, where 1 corresponds to a fuzzy imageand 7 to a vivid and clear representation ofthe movement. Intermediate levels from 2 to6wouldcorrespond toamixedevaluationof therepresentation. A score is calculated from the8 imagined movements.
– Then, the participants memorised a thumb-to-finger opposition task. The other 5 sub-indica-tors were then recorded during actual andimagined sequences:
– Self-estimation of imagery quality: aftereach imagined trial, participants estimated thequality of their imagery using the seven-pointscale previously described.
– Mental chronometry: this indicator was cal-culated from the difference between durationsof actual and imagined movements. Temporalequivalence (i.e., isochrony) is a reliablemeasure of imagery quality. The participantused his/her left thumb to depress a pushbutton switch tomark the beginning and end ofeach trial.
– Skin conductance: this sub-indicator is areliable measure of arousal. The signal wasrecorded continuously by two electrodes posi-tioned on the index andmiddle finger of the lefthand (where exocrine sweat glands under the
control of the sympathetic system are mostprominent). A decrease in skin conductancereflects a decrease in arousal, indicating thatthe participant relaxed. On the contrary, nodifference between before and after MI trialshows the participant was involved in thecognitive task.
– Ohmic perturbation duration: this is theduration of the change in skin conductancewhencognitiveactivityproducesasympatheticresponse and it reflects the level of attention.GoodMI is characterised by a similar durationduring imagined and actual task performance.
– Respiratory sinus arrhythmia: this last sub-indicator is the mean amplitude of instanta-neousheart ratevariabilityand is influencedbyparasympathetic activity. A significant reduc-tion of respiratory sinus arrhythmia is typi-cally observed while the participant is involvedin cognitively demanding tasks, and is consi-dered a reliable index of cognitive effort duringMI.The 6 sub-indicators were then combined and
the Motor Imagery Index calculated to rank thetwenty-three persons participating in this studyaccording to their imagery ability. Mean scorewas 5.65 (± 0.17), range 3.77-6.96. The twelveparticipants with the highest score were conside-red as good imagers, while the eleven others aspoor imagers. Figures 1 and 2 represent measu-rements of chronometric and psychophysiologi-cal variables (sub-indicators from 3 to 6) of twoparticipants. Based on theMotor Imagery Index,the first one is considered as a good imager whilethe second one as a poor imager.
Experiment 2: Neuronal excitabilityand motor imagery quality
Understandinghowneurophysiological activa-tion underlying MI mirror imagery qualityremains an open question.Most studies assessedthe ability to imagine vividly a movement withsubjective questionnaires. So far, only Guillotand collaborators (2008) combined functionalmagnetic resonance imaging (MRI) data withpsychophysiological, behavioural and psycholo-gical measurements to correlate cortical activa-tion to imagery capacity. Good imagers activatedto a greater extent the parietal and ventrolateralpremotor regions, which are known to play acritical role in the generation of mental images.By contrast, poor imagers recruited the cerebel-
GRP : fyssen JOB : annales27Internet DIV : 11mp⊕LEBON⊕Florent p. 8 folio : 109 --- 14/5/013 --- 15H19
NEUROBIOLOGY
109

lum, orbito-frontal and posterior cingulate corti-ces, involved in motor learning. However, noresearch showed how characteristics of the men-tal image mirror the activation of the nervoussystem, particularly of M1. The hypothesis wasthat motor imagery quality would influence neu-rophysiological mechanisms assessed by TMS.The present study aimed to demonstrate howgood and poor imagers activated specifically theneuronal network while imagining a simplemovement.
Participants were instructed to imagine orperform an actual thumb-tapping movement in
timewith a 1Hzmetronomewith their dominant(right) hand.The targetmusclewas theopponenspollicis muscle, agonist of the movement. TMSwas delivered over the cortical representation ofthis muscle in M1. Motor evoked potentials werethen recorded in the opponens pollicis muscleand the abductor digiti minimi muscle to assessspatial specificity of MI. An increase of the res-ponse should only be recorded in the target mus-clewhile imagining a simplemovement involvingthis muscle. Furthermore, TMS was triggered intimewith themetronomeor between twobeats inorder to assess temporal specificity of MI. Anincrease of the response should only be recorded
Figure 1:Motor Imagery Indexmeasurementsof a good imager (score 6.68). (A) Skin conduc-tance (mS) and duration (s) of an actual and imagi-ned thumb-to-finger movement, and during rest.The dashed vertical bars in the upper part of thefigure represent the calculation of the ohmic pertur-bationduration (OPD) inseconds.OPDduringMI issimilar to the duration of actual movement, indica-ting good MI performance. The solid vertical barsrepresent the start and end of each task. The dura-tions of the actual and imagined movement aresimilar, showing isochrony. (B) Instantaneousheart rate during MI and rest. MI elicited variationin the respiratory sinus arrhythmia, observed asa decrease in variability of instantaneous heartrate. MI = motor imagery, mS = micro Siemens,s = second.
Figure 2:Motor Imagery Indexmeasurementsof a poor imager (score 4.37). (A) Skinconductance (mS) and duration (s) of an actual andimagined thumb-to-finger movement, and duringrest. The dashed vertical bars in the upper part ofthe figure represent the calculation of the ohmicperturbation duration (OPD) in seconds. OPDduring MI is shorter than during actual condition,indicating poor MI performance. The solid verticalbars represent the start and end of each task. Theduration of the imaginedmovement is greater thanthe duration of the actual task, showing poorisochrony. (B) Instantaneous heart rate during MIand rest condition. There was no decrease in theamplitude between maximal and minimal heartrate, showing that MI did not elicit variation in therespiratory sinus arrhythmia. MI =motor imagery,mS = micro Siemens, s = second
GRP : fyssen JOB : annales27Internet DIV : 11mp⊕LEBON⊕Florent p. 9 folio : 110 --- 14/5/013 --- 15H19
NEUROBIOLOGY
110

in time with the beat, as during actual perfor-mancewhere the targetmuscle is activatedon thebeat and relaxed between two beats.
Increase of neuronal excitabilityThe results confirmed previous research (for
review, see Stinear, 2011). Indeed, amplitude ofmotor evoked potentials during MI was moreimportant than at rest, when all the participantswere pooled. This showed an increase of pyrami-dal neuron activation during MI. Moreover, theincreasewasmore important in the targetmuscle(spatial modulation) and when participants ima-gined themovement in timewith themetronome(temporal modulation).
Spatial and temporalmodulationand ima-gery qualityRepeated measure analysis of variance was
used to compare the response amplitudesbetween and within the participants. Resultsshowed that good imagers activated more speci-fically the cortical cells controlling the thumbmusclewhile poor imagers experienced a generalincrease of M1 during MI, activating both thetarget muscle and the muscle not involved in thetask. Besides, this increase was greater whileparticipants imagined the movement on timewith themetronome, in bothmuscles for the poorimagers but only in the thumb muscle for the
good imagers. Muscle-specific temporal modula-tion of MI was, therefore, correlated to the ima-gery quality.
Experiment 3: Inhibitory processesof motor imagery
Studies in functional MRI showed extensivecortical and sub-cortical activation during MI,includingparticularly theprimaryandsecondarymotor and parietal areas (de Lange et al., 2005).Interestingly,motor execution areaswere activa-ted, although nomovement was produced duringMI. An inhibitory network would therefore pre-vent any muscle contraction. We hypothesisedthat the parietal lobe would be involved in thisnetwork since its activation has been systemati-cally reported while imagining. Besides, patientswith parietal cortex damage encountered difficul-ties to predict the time to perform a movement(Sirigu et al., 1996) or to prevent from movingduring MI (Schwoebel et al., 2002). The purposeof this third study was to establish a causal linkbetween the parietal lobe and M1 with a double-coil TMS. Participants were instructed to ima-gine an index-to-thumb pinching task or to per-form the actual movement in time with a 1 Hzmetronome. The index muscle of the dominant(right) hand was the target muscle.
Neuronavigated dual-coil transcranialmagnetic stimulationA 3D representation of each participant’s
brain was created from an anatomical MRI scan.Two stimulation sites were precisely located:1) the inferior parietal lobule whose stereotaxiccoordinates were based on MRI studies (forexample, de Lange et al., 2005) and 2) the corticalrepresentation of the right index muscle overleft M1. The position of the two coils was thenmarked on the 3D image to reproduce a preciseand constant stimulation over the two sites(Figure 4). The neuronavigated dual-coil TMSwas used to assess any interaction between thesetwo areas during MI.
Two types of stimulation were delivered:1) single stimulation over left M1 and 2) doublestimulation corresponding to one stimulationover the parietal lobe 6 or 12 ms prior to M1 sti-mulation. The ratio,MEP amplitude after doublestimulation over MEP amplitude after single sti-mulation over M1, was used to test any effect ofthe parietal lobe on M1 during MI and at rest.
Figure 3: Normalised sum of motor evokedpotentials (MEP) during motor imageryrelative to imagery ability. The sum wascalculated by adding mean amplitude of MEPevoked by increasing magnetic stimulationintensity. The sum during motor imagery (MI) wasnormalised to the sum at rest. When participantswere pooled relative to their imagery ability, spatialand temporal specificity was only observed in goodimagers. OP = Opponens Pollicis, ADM = AbductorDigiti Minimi, ON = magnetic stimulation in timewith the metronome beep, OFF = stimulationbetween the beeps, *= P S 0.05.
GRP : fyssen JOB : annales27Internet DIV : 11mp⊕LEBON⊕Florent p. 10 folio : 111 --- 14/5/013 --- 15H21
NEUROBIOLOGY
111

Interaction between inferior parietal andprimary motor corticesThe results showed a decrease of MEP ampli-
tude after double stimulation during MI compa-red to single stimulation over M1. On thecontrary, double stimulation increased MEPamplitude at rest.Activation of the inferior parie-tal lobe during MI would therefore inhibit M1 toprevent from moving. However, the interactionbetween the two cortices would be indirect invol-ving trans-synaptic connections, since inhibitionwas only observedwith long (12ms) but not short(6 ms) inter-stimulus interval (Figure 5).
This study explains, fromaneurophysiologicalpoint of view, the inhibitory mechanisms invol-ved in movement prevention during MI. Howe-ver, other cortical and sub-cortical areas, such asthe cerebellum, seem to be part of this process.Additional research, using TMS associated toother recording and stimulation techniques,would help to determine the complex neuronalnetwork underlying MI.
Conclusion
This post-doctoral research project aimed tobetter understand neurophysiological processes
underlying MI. The main results validated theMotor Imagery Index (Collet et al., 2011) andestablished a direct link between central andautonomic nervous systems relative to imageryability. Interestingly, the primarymotor cortex isboth activated and inhibited duringmental simu-lation of movement. The inferior parietal cortexplays an important indirect role in signal inhibi-tion and, therefore, movement prevention whileimagining. This fundamental research and itsresults could be used for practical applications.Indeed, it is well established that MI improvesmotor performance after immobilisation, centraldamage or spinal injury, when integrated intorehabilitation programs. It would be interesting
Figure 4: Neuronavigated dual-coil TMS. 3Drepresentations of the brain (upper part) werecreated from a MRI scan. Yellow crosses show theorientation of the TMS coils over the right inferiorparietal lobe (upper left) and the left primarymotorcortex (upper right). Targets at the bottom showedlive the position of each coil relative to thestimulation site, allowing a precise and constantstimulation.
Figure 5: Conditioned and non-conditionedmotor evoked potentials (MEP). Non-conditioned (NC) MEP are responses recorded inthe index muscle of the right hand and elicited bysingle stimulation over the left primary motorcortex. Conditioned MEP are responses elicitedby a stimulation of the right parietal lobe followedby a stimulation of the left primary motor cortex(double stimulation). The 2 stimuli were deliveredwith an inter-stimulus interval (ISI) of 6 or 12 ms.Grey and black lines represent participants’recordings and mean amplitude, respectively.Amplitude of NC MEP was greater during MIthan at rest, showing that corticomotor excita-bility was increased while imagining. On thecontrary, MEP amplitude was reduced only whenparietal lobe was stimulated 12 ms prior to themotor cortex during MI. Primary motor cortex isboth activated and inhibited during mentalsimulation of movement while the parietal lobewould play an important role in movementprevention during MI.
GRP : fyssen JOB : annales27Internet DIV : 11mp⊕LEBON⊕Florent p. 11 folio : 112 --- 14/5/013 --- 15H21
NEUROBIOLOGY
112

to know whether imagery ability improvement iscorrelated to performance increase and whethermovement inhibition while imagining is predo-minant to mental performance.
AcknowledgementI would like to thank the Fyssen Foundation
and the committee members who gave me theopportunity to carry out this post-doctoral pro-ject; CathyStinear andWinstonByblow from the
University of Auckland (New Zealand) for theirsupport and help throughout this project; Chris-tian Collet and Aymeric Guillot from the Univer-sity of Lyon (France) for their advice and themany exchanges; Martin Lotze from the Univer-sity of Greifswald (Germany) for the collabora-tion in the third experiment; and the teams ofthe Centre for Brain Research and of the Move-ment Neuroscience Laboratory (University ofAuckland, NZ).
GRP : fyssen JOB : annales27Internet DIV : 11mp⊕LEBON⊕Florent p. 12 folio : 113 --- 14/5/013 --- 15H21
NEUROBIOLOGY
113

Les fibules : de la pièce unique à la fabrication en série.Innovations et évolution des procédés de fabrication
d’un objet du quotidien entre le IIIe s. av. et le Ier s. ap. J.-C.
Jenny KAURINChercheur associé, UMR 6298-ARTeHIS
RésuméLes fibules sont des objets emblématiques de la culture celtique et abondent sur les sites archéolo-
giques européens. Ce sont des épingles de sûreté destinées à fixer les plis d’un vêtement, portées à lafois par les hommes et par les femmes, qui témoignent de nombreuses variations de formes, de décorset de matériaux, suivant les évolutions de la mode vestimentaire et des usages locaux. Leur classifi-cation en chaîne opératoire technique permet ici de révéler des savoir-faire distincts, reflétant uneconception différente de l’objet fibule et de sa fabrication. L’analyse des répercussions de ces savoir-faire sur l’organisation du travail permet d’apporter des éléments de compréhension inédits sur laplace occupée par les artisans entre la fin du IIIe s. av. J.-C. et le début du Ier s. ap. J.-C. Autantd’éléments qui permettent de contribuer à une meilleure connaissance de la société celtique.
Mots-clésArchéologie, protohistoire celtique, Europe, fibule, technique de fabrication, savoir-faire, métal,
artisan
Brooches : from unique piece to mass production.Innovations and evolution of manufacturing processes
of an object of the everyday life between the 3rd c.and the 1st c. AD
AbstractBrooches are emblematic objects of the Celtic culture and abound on European archeological sites.
Brooches are safety pins intended to fix tails of a garment and are worn at the same time by men andwomen. They show numerous variations of forms, ornaments andmaterials, following the evolutionsof clothing fashionand localuses.Their classification in technical operatingchainallowshere to revealdifferent know-how, reflecting a conception different from the object brooch and its manufacturing.The analysis of consequences of these know-how on the organization of work allows to bringunpublished elements of understanding on the place occupied by craftsmen between the end of 3rdc. BC and the beginning of 1st c. AD.
KeywordsArchaeology, celtic period, Europe, brooch, technique of manufacturing, know-how, metal, crafts-
man
Lesfibules sont des objets emblématiques de laculture celtique. Ces épingles de sûreté, utiliséespour fixer les plis d’un vêtement ou pour lefermer, sont portées à la fois par les hommes
et par les femmes. Les fibules témoignent denombreuses variations de formes, de décors,de dimensions et de matériaux (fer, alliagescuivreux, plus rarement argent ou or, parfois
GRP : fyssen JOB : annales27Internet DIV : 12mp⊕KAURIN⊕Jenny p. 1 folio : 114 --- 14/5/013 --- 15H24
PALÉONTOLOGIE / ARCHÉOLOGIE
114

associés sur un même objet), répondant aux évo-lutions de la mode vestimentaire et aux usageslocaux. A côté de la vaisselle en céramique, ils’agit d’un des types d’objet les plus abondantssur les sites archéologiques celtiques et gallo-romains, qu’il s’agisse d’habitats, de nécropolesou de lieux de culte. Leur évolution rapide et leurdiffusion sur de vastes espaces géographiques enfont des objets essentiels pour la définition desgroupes culturels et l’établissement des chrono-logies, sur lesquels repose toute la démarchearchéologique en tant que science historique. Oncomprend dès lors pourquoi les fibules comptent,depuis le XIXe s. et la naissance de l’archéologiemoderne, parmi les objets les plus étudiés (parmiles travaux pionniers : Déchelette 1913 ; Rei-necke 1965 ; Viollier 1916 ; parmi les travauxrécents emblématiques : Feugère 1985 ; Geb-hardt 1991, Metzler 1995).
Poser un regard renouvelé sur ces objets n’estdonc pas aisé. Nous nous sommes orientée versune étude technique visant d’une part, à caracté-riser les savoir-faire sous-tendus par leurs procé-dés de fabrication et d’autre part, à mettre enévidence leur évolution en fonction des innova-tions progressivement mises en place. Pour cefaire, nous nous sommes appuyée sur l’approchenovatrice initiée par J.-P. Guillaumet et M. Per-
not à la fin des années 80 (Guillaumet 1993 ;LehöerffetPernot2003) etdontonpeut regretteraujourd’hui qu’elle n’ait pu être plus développée.
De la description technique...La réflexion a été portée à l’échelle de l’Europe
celtique et s’est concentréeplusparticulièrementsur les régionsoccidentales.Lesprincipaux typesde fibules connus pour la période comprise entrela findu IIIe s. av. J.-C. et le début du Ier s. ap. J.-C.ont été soumis à une grille d’analyse technique,les décrivant selon leurs éléments constitutifs(fig. 1). En effet, une fibule est constituée de deux-parties : le corps et le mécanisme de fixation. Lecorpsest lui-mêmecomposéde l’arc (qui supportel’essentiel de la charge ornementale de l’objet) etdu pied (destiné à maintenir la fibule fermée). Lemécanismede fixation comprendun ardillon (quitraverse et maintient les pans du tissu à fixer) etunmécanismequipermetsonarticulation. Ilpeuts’agir soit d’un ressort (l’articulation se fait parpression) soit d’une charnière (l’articulation sefait par rotation). Lemécanisme de fixation est leprincipal point de fragilité de la fibule et en cons-titue la contrainte technique majeure.
Indépendamment de leur variabilité morpho-logique et ornementale (qui se décline en plu-sieurs centaines de types et variantes), plusieurs
Fig. 1 : éléments constitutifs d’une fibule
GRP : fyssen JOB : annales27Internet DIV : 12mp⊕KAURIN⊕Jenny p. 2 folio : 115 --- 14/5/013 --- 15H25
PALÉONTOLOGIE / ARCHÉOLOGIE
115

critères se sont détachés pour leur valeur chro-nologique :– le matériau : fer ou alliage cuivreux ;– la manière dont le pied est mis en forme :attachésur l’arcou faisant corpsavec lui (fig.2aet b) ;
– la nature du mécanisme de fixation : à ressortinclus, à ressort rapporté ou à charnièreincluse (fig. 2c, d et e) ;
– la techniqued’obtentiondesdifférentes partiesconstitutives de la fibule : par martelage et/oufonte, avec ou sans assemblage.
Fig. 2 : différents types de pied et de mécanisme defixation observés sur les fibules du corpus
La séquence chronologique considérée ici estmarquée par quatre innovations majeures. AuIIIe s. av. J.-C., toutes les fibules sont obtenuespar martelage d’un fil de métal et se distinguentpar leur pied attaché sur l’arc, à l’aide d’uneagrafe ou d’une bague de fixation (fig. 3a).
LasecondemoitiéduIIe s. av.J.-C. estmarquéepar l’apparitiond’unenouvelle techniquedemiseen forme du pied : le pied faisant corps avec l’arc.Cette innovationvadepair avec la réintroductiond’un matériau tombé en désuétude depuis ledébut du IIIe s. : les alliages cuivreux (fig. 3b).
L’exploitation des propriétés plastiques de cesalliages conduit, au début du Ier s. av. J.-C., à lamise en place d’un nouveau procédé de fabrica-tion, lui aussi inusité depuis le début du IIIe s. av.J.-C. : la fonte à la cire perdue (fig. 3c).
Il faut attendre la fin du Ier s. av. J.-C. pour quel’on s’affranchisse de la contrainte technique quereprésente le ressort inclus.Le ressort rapporté al’apparenced’un traditionnel ressort inclus,maisfonctionne comme une charnière : par rotation àl’intérieurd’uneboîtecylindriqueetnonparpres-
sion (fig.4a).L’apparitionde la charnière incluse,fonctionnant par rotation autour d’un axe, cons-titue l’aboutissement de ce processus (fig. 4b).
Le développement et la diffusion de ces inno-vations ont deux conséquences principales. Lapremière est une déprise croissante du fer, qui nese travaille à cette époque que par martelage, auprofit de la généralisation des métaux à faibletempérature de fusion, au premier rang desquelsse trouvent les alliages cuivreux. La seconde estun affranchissement progressif des contraintestechniques liées à la fabrication de l’objet fibule,qui requiert au fil du temps un savoir-faire demoins en moins élevé.
... à l’évolution des savoir-faireL’utilisation du fer renvoie à un savoir-faire
métallurgique élevé. La qualité et la solidité de lafibule reposent sur le choix d’un fer à la fois douxet résistant pour autoriser les déformations plas-tiques dues à la mise en forme de l’objet et lespressions fréquentes impliquées par son ouver-ture et sa fermeture dansunusage courant. Cettecontrainte propre au matériau explique sansdoute en grande partie pourquoi une grandepartie des fibules en fer se distingue par un arc deforme simple et un décor très limité (fig. 3a).Toutefois, cette apparente simplicité est à nuan-cer. Dès le début du Ier s. av. J.-C., les artisansforgerons cherchent à donner aux fibules en ferun développement formel analogue à celui desfibules en alliage cuivreux, dont le corps estpourtant obtenu par coulée. Cette productionreste cependant très limitée. Ainsi, qu’il s’agissedes séries importantes de la fin du IIIe et de lapremière moitié du IIe s. av. J.-C. ou bien desproductions plus restreintes que l’on trouve àpartir de la seconde moitié du IIe s. av. J.-C., lafabrication des fibules en fer relève du savoir-faire de l’artisan forgeron, qui travaille les piècesles unes après les autres.
En revanche, l’utilisation des alliages cui-vreux, et d’une manière générale des métaux àfaible températuredefusion, renvoieàdessavoir-fairequi évoluentaufildu temps. Jusqu’à lafinduIIe s. av. J.-C., comme pour les fibules en fer, lafabrication nécessite un savoir-faire métallurgi-que élevé. La qualité et la solidité de la fibulereposent sur un alliage qui associe propriétésplastiqueset résistance.Aussi, lesmodèleset leurdécor restent relativement simples et le travail del’artisan bronzier, qui fabrique les pièces les unes
GRP : fyssen JOB : annales27Internet DIV : 12mp⊕KAURIN⊕Jenny p. 3 folio : 116 --- 14/5/013 --- 15H25
PALÉONTOLOGIE / ARCHÉOLOGIE
116

Fig. 3 : proposition de restitution schématique des différentes étapes de la fabrication des : A) fibules à piedattaché sur l’arc, B) fibules à pied faisant corps avec l’arc, C) fibules dont le corps est obtenu par coulée et lemécanisme de fixation par martelage (en une seule pièce).
GRP : fyssen JOB : annales27Internet DIV : 12mp⊕KAURIN⊕Jenny p. 4 folio : 117 --- 14/5/013 --- 15H25
PALÉONTOLOGIE / ARCHÉOLOGIE
117

après les autres, relève de la petite dinanderie(fig. 3b).
L’introductionde lacouléeaudébutduIer s. av.J.-C. rend nécessaire un nouveau savoir-faire. Lerecours à la technique de la fonte à la cire perdueaccordeuneplace primordiale au travail de sculp-ture sur cire, préalable essentiel à la fabricationdes moules en terre cuite. C’est l’artisan sculp-teur qui va donner aux fibules leurs caractéristi-ques morphologiques. Parallèlement, un savoir-faire métallurgique élevé reste nécessaire demanièreàautoriser tantunecouléedequalitéquela réalisation dumécanisme de fixation, toujoursobtenuparmartelage.Ainsi, lesfibules restent enune seule pièce : le moule donne une fibule ébau-chée où le corps est prolongé par un long fil, dontla mise en forme permettra l’obtention du méca-nisme de fixation (fig. 3c). Aussi, la fabrication sefait toujours pièce par pièce à raison, très vrai-semblablement, d’un moule par fibule, comme lesuggère l’un des raresmoules connus attribuableàcettepériode (Guillaumet1993,pl. 54)et la fortevariabilité des fibules, qui ne semblent pas pré-senter de standardisation.
Ainsi, l’époque augustéenne, dès la fin du Ier s.av. J.-C., connaît une évolution majeure dansl’organisation du travail : la fabrication en série.
Cette évolution conceptuelle dans l’organisationdu travail a été renduepossible par l’introductionde la charnière. La charnière permet en effet dedissocier les parties fonctionnelles de la fibule(corps et mécanisme de fixation) en plusieurspièces, qui seront assemblées mécaniquement.Dès lors, il est possible d’envisager la fabricationsimultanée de plusieurs pièces grâce à l’élabora-tiondemoulesadaptés.Lacharnièresedéclineendeux types : le ressort rapporté (rotation d’unfaux ressort à l’intérieur d’une boîte cylindrique,fig. 4a), abandonné dès lemilieu du Ier s. ap. J.-C.,et la charnière incluse (avec rotation autour d’unaxe fig. 4b), dont l’usage sera généralisé dès lemilieu du Ier s. ap. J.-C. En s’affranchissant de lacontrainte technique majeure que constituait leressort, la charnière induit de fait une perted’importance du savoir-faire métallurgique del’artisan bronzier au profit du sculpteur sur cire.L’alliage n’a plus besoin d’avoir des propriétésspécifiques permettant l’obtention d’un ressort àla foissoupleetrésistantet l’onassisteàunepertede qualité des alliages utilisés, observable à l’œilnu.En revanche, le travail de sculpture sur cire etl’habileté à confectionner des moules autorisantla fabrication simultanée d’un maximum de piè-ces deviennent fondamentaux. L’époque augus-téenne entérine ainsi un basculement essentiel :
Fig. 4 : proposition de restitution schématique des différentes étapes de la fabrication des : A) fibules à ressortrapporté, B) fibules à charnière incluse (en plusieurs pièces).
GRP : fyssen JOB : annales27Internet DIV : 12mp⊕KAURIN⊕Jenny p. 5 folio : 118 --- 14/5/013 --- 15H26
PALÉONTOLOGIE / ARCHÉOLOGIE
118

la fabricationde lafibulene résulteplusdu travailde l’artisan bronzier, mais de l’action croisée dusculpteur, du fondeur et surtout de l’ouvrier spé-cialisé qui assemble mécaniquement ou par bra-sure les différentes pièces constitutives de l’objetet finalise le décor. De fait, on assiste à une stan-dardisation importante des fibules.
L’introduction d’une innovation, quelles quesoient les répercussions qu’elle peut avoir, tantau niveau du développement formel de la fibuleque de l’organisation du travail, ne signifie paspour autant l’abandon des procédés techniquesconnus par ailleurs. Ainsi, les fibules à pied atta-ché sur l’arc perdurent et continuent à être déve-loppées, même après l’invention du pied faisantcorps avec l’arc à la fin du IIe s. av. J.-C. Demême,l’apparitionde la coulée audébut du Ier s. av. J.-C.ne se traduit pas par l’abandon des fibules entiè-rement obtenues par martelage, mais semble enrevanche marquer l’abandon des fibules à piedattaché sur l’arc (qui ne perdurent que dansles régions sud-alpines). Au contraire, la char-nière, qu’elle soit incluse ou à ressort rapporté,s’impose très rapidement. Les possibilités offer-tes par la fabrication en série semblent avoir euraison de tous les procédés impliquant la fabri-cation de pièces uniques. Seules les fibules enfer, peu nombreuses au début du Ier s. ap. J.-C.,maintiennent un savoir-faire hérité de plusieurssiècles. Toutefois, elles disparaîtront rapidementdevant la rentabilité de la fabrication en série defibules coulées et assemblées mécaniquement,qui autorise le recours à des alliages meilleursmarchés et ne nécessite pas un savoir-faire spé-cialisé.
Cette première étude doit maintenant êtrevérifiée par des analyses paléo-métallurgiquesafin d’asseoir les restitutions proposées sur lesprocédés de fabrication. Au-delà, un travail dequantification pourrait permettre de mesurerl’importance respective de l’utilisation de cesdifférentsprocédés techniquesaucoursdu tempset selon les régions du monde celtique, demanière à préciser les modalités de cette évolu-tion. Cette approche, dont nous avons pu évaluerici la pertinence, pourrait se révéler extrême-ment porteuse.
RemerciementsJe tiens en premier lieu à remercier la Fonda-
tion Fyssen qui m’a donné la possibilité de pour-suivre mes recherches à l’Université de Rome la
Sapienza et la professeure P. Piana Agostinettipour son accueil et son aide précieuse. Je remer-cie également les autres institutions qui m’ontaccueillies pour des recherches documentaires :l’Ecole Française de Rome et le centre archéolo-gique européen de Bibracte.
Bibliographiee Déchelette 1913 : Dechelette (J.) – Manuel
d’archéologie préhistorique, celtique et gallo-romaine, Paris, 1913.
e Feugère 1985 : Feugere (M.) – Les fibules enGaule mériodionale de la conquête à la fin duVe siècle après J.-C., (Revue archéologique deNarbonnaise Supplément 12), Paris, 1985.
e Gebhardt 1991 : Gebhardt (R.) – Die Fibelnaus dem oppidum vonManching, Stuttgart, 1991.
e Lehöerff et Pernot 2003 : Dallo scavoall’archeologia sperimentale. La produzione difibule a Bibracte e Autun : le tappe della ricerca ei risultati, dans : Formigli (E.) – Fibulae. Dall’etàdel bronzo all’alto Medioevo tecnica e tipologia,(Atiquarium di Poggio Civitate, comune diMurlo, 4), Firenze, 2003, pp. 173-187.
e Guillaumet 1993 : Guillaumet (J.-P.) – Lesfibules de Bibracte. Technique et typologie, (Centrede recherches sur les techniques gallo-romainesno14), Dijon, 1993.
e Metzler 1995 : Metzler (J.) – Das treverischeOppidum auf dem Titelberg. Zur Kontinuität zwis-chen spätkeltischen und frührömischen Zeit inNord-Gallien, (Dossier d’archéologie du muséenational d’histoire et d’art III), Band 1 et 2,Luxembourg, 1995.
e Reinecke 1965 : Reinecke (P.) – MainzerAufsätze zur Chronologie der Bronze- und Eisen-zeit, Bonn, 1965 (réédition de l’ouvrage paru en1902).
e Viollier 1916 : Viollier (D.) – Les sépulturesduSecondAgeduFer sur le plateau suisse,Genève,1916.
Brooches are emblematic objects of theCeltic culture
These safety pins, used to fix tails of a garmentor to close it, are worn at the same time by menandwomen.Brooches shownumerous variationsof form, ornament, size and material (iron,
GRP : fyssen JOB : annales27Internet DIV : 12mp⊕KAURIN⊕Jenny p. 6 folio : 119 --- 14/5/013 --- 15H26
PALEONTOLOGY / ARCHAEOLOGY
119

cuprous alloy, more rarely silver or gold, someti-mes associated on the same object), answeringthe evolutions of clothing fashion and local uses.Next to pottery, it’s one of the most plentifultypes of object onCeltic andGallo-Romanarcheo-logical sites: settlements, cemeteries or sanctua-ries. Their fast evolution and distribution on vastgeographical spaces make of them essentialobjects for definition of cultural groups and esta-blishment of chronologies, on which all thearchaeological approach as an historical scienceis based on. Because of it, brooches are one ofmost studied objects since the 19th c. and birth ofmodern archaeology (among pioneer works:Déchelette 1913; Reinecke 1965; Viollier 1916;among emblematical works: Feugère 1985; Geb-hardt 1991, Metzler 1995).
Making a new approach is not something easy.We turned to a technical study aiming on onehand, at characterizing know-how from themanufacturing processes and on the other hand,at highlighting evolution of know-how accordingto innovations. To do it, we based on the innova-tive approach introduced by J.-P. Guillaumet andM. Pernot at the end of the 80s (Guillaumet 1993;Lehöerff and Pernot 2003). We can regret todaythis approach was not more developed.
From technical description...Reflection concerns Celtic Europe and is
concentrated more particularly on the westernareas. Main known types of brooches for periodincluded between the end of 3rd c. BC and thebeginning of 1st c. AD were recorded in a tech-nical analysis charts, describing them accordingto their constituent elements (fig. 1). A brooch iscomposed of two parts: the body and the mecha-nism of binding. The body is composed itself ofthe bow (which bears the ornaments) and the foot(intended to maintain the brooch closed). Themechanism of binding associates the pin (whichpierces andmaintains tails of a garment) and themechanism which allows its articulation. It canbe or a spring (articulation is made by pressure)or a hinge (articulation is made by rotation). Themechanism of binding is the main point of fragi-lity of the brooch and constitutes the major tech-nical stress.
Independently of their morphological andornemental variability (brooches decline in seve-ral hundreds of types and variants), several cri-teria have a chronological value:– the material: iron or cuprous alloy;– the way the foot is shaped: attached on the bowor making body with him (fig. 2a and b);
Fig. 1: Constituent elements of a brooch
GRP : fyssen JOB : annales27Internet DIV : 12mp⊕KAURIN⊕Jenny p. 7 folio : 120 --- 14/5/013 --- 15H26
PALEONTOLOGY / ARCHAEOLOGY
120

– the nature of the mechanism of binding: inclu-ded spring, brought back spring or includedhinge (fig. 2c, d and e);
– the technique of manufacturing of variousconstituent parts of the brooch: by hammeringand/or casting, with or without assembly.
Fig. 2: Différent types of foot and mechanism ofbinding
The chronological sequence considered here ismarked by four major innovations. In 3rd c. BC,all brooches are manufactured by hammering ofa metal shaft and distinguish by their foot atta-ched on the bow with a hook or a ring of binding(fig. 3a). The second half of the 2nd c. BC ismarked by the introduction of a new technique ofshaping of the foot: the footmaking bodywith thebow. This innovation go hand in hand with thereintroduction of amaterial gone out of use sincethe beginning of 3rd c.: cuprous alloys (fig. 3b).
Exploitation of plastic properties of thesealloys leads at the beginning of 1st c. BC to thedevelopment of a new manufacturing process:cast in lost wax (fig. 3c). This process was alsouncommon since the 3rd c. BC.
We have to wait for the end of 1st c. BC to freebrooches from the technical stress which is theincluded spring. The brought back spring lookslike a traditional included spring, but works as ahinge: by rotation and not by pressure (fig. 4a).The appearance of included hinge working byrotation on an axis constitutes the outcome ofthis process (fig. 4b).
Development and distribution of these innova-tions have two main consequences. The first oneis decrease of iron (which is worked only byhammering) for the benefit of low smelting pointmetals, in the front row of which are cuprous
alloys. The second one is a progressive emanci-pation of technical stress bound to the manufac-turing of brooches, which require over time a lessand less raised know-how.
... to the evolution of know-howUse of iron refers to a highmetallurgical know-
how. Quality and solidity of brooches are basedon the choice of an at the same time soft andresistant iron to authorize plastic deformationsdue to the shaping of the object and the frequentpressures involved by its opening and lock in acurrent use. This stress characteristic of thematerial explainsdoubtless largelywhyabigpartof iron brooches distinguishes itself by a simpleshape bow and a very limited ornament (fig. 3a).However, this apparent simplicity has to be qua-lified. From the beginning of 1st c. BC, smithcraftsmen try to give to iron brooches a formaldevelopment similar to the development of broo-ches in cuprous alloy, from which the body isobtained by casting. This production remainshowever very limited. Considering importantseries of the end of the 3rd c. and the first half of2nd c. or more restricted productions of thesecond half of 2nd c. BC, manufacturing of ironbrooches belongs to know-how of smith crafts-man, who works brooches one after the other.
On the other hand use of cuprous alloys and ina general way low melting point metals sendsback to know-howwhich evolve over time. Till theend of the 2nd c. BC, as for iron brooches, themanufacturing requires a high metallurgicalknow-how. Quality and solidity of the brooch isbased on an alloy which associates plastic pro-perties and resistance. So, shapes and ornamentremain relatively simple and work of the crafts-man, who makes brooches one after the other,recovers from small brasswork (fig. 3b).
Introduction of casting at the beginning of the1st c. BC makes necessary a new know-how. Useof the technique of cast in lostwax gives an essen-tial part to the work of sculpture on wax, impor-tant prerequisite in themanufacturing ofmouldsin terra-cotta. It’s the sculptor craftsman who isgoing to give to brooches their morphologicalcharacteristics. At the same time, a raisedmetal-lurgical know-how remains necessary to obtainboth a quality casting and the shaping of mecha-nismof binding, still obtainedbyhammering. So,brooches stay in a single piece: the mould gives asketched brooch where the body is extended by a
GRP : fyssen JOB : annales27Internet DIV : 12mp⊕KAURIN⊕Jenny p. 8 folio : 121 --- 14/5/013 --- 15H26
PALEONTOLOGY / ARCHAEOLOGY
121

Fig. 3: Proposal of schematic restoration of different steps of manufacturing: A) brooches with foot attachedon the bow, B) brooches with foot making body with the bow, C) brooches with body made by casting andmechanism of binding by hammering (in one piece).
GRP : fyssen JOB : annales27Internet DIV : 12mp⊕KAURIN⊕Jenny p. 9 folio : 122 --- 14/5/013 --- 15H27
PALEONTOLOGY / ARCHAEOLOGY
122

long thread of metal (fig. 3c). Shaping of thisthread of metal will allow the obtaining of themechanism of binding. Brooches are still madepiece by piece, using very credibly one mould bybrooch.This issuggestedbyoneof therareknownmoulds attributable to this period (Guillaumet1993, pl. 54) and the strong variability of broo-ches, which present no standardization.
So the augustan period, from the end of 1st c.BC, knows a major evolution in organization ofwork: the mass production. This conceptual evo-lution in organization of work wasmade possibleby introduction of the hinge. The hinge indeedallows to separate functional elements of thebrooch (body andmechanism of binding) in seve-ral pieces, whichwill bemechanically assembled.Therefore it’s possible to envisage simultaneousmanufacturing of several pieces thanks to elabo-ration of adapted moulds. The hinge declines intwo types: the brought back spring (rotation of afalse spring inside a box fig. 4a), abandoned fromthe middle of 1st c. AD, and the included hinge(with rotation on an axis fig. 4b), use ofwhichwillbe generalized from themiddle of 1st c. AD. Freeof the major technical stress which is the spring,the hinge infers de facto an important loss ofmetallurgical know-how of bronze-caster crafts-
man for the benefit of the sculptor on wax. Thealloy doesn’t need any more to have specificproperties allowing the obtaining of a spring atthe same time supple and resistant.We can see aloss of quality of alloys. On the other hand, workof sculpture on wax and skill to make mouldsauthorizing the simultaneousmanufacturingof amaximum of pieces become fundamental. Theaugustanperiod confirmsan essential fall:manu-facturing of brooches doesn’t result any morefrom the only work of bronze-caster craftsman,but of the united action of the sculptor, thecaster and especially the semi-skilled workerwho assembles mechanically or by brasure thevarious essential parts of brooches and finalizesthe ornament. Actually, we attend an importantstandardization of brooches.
Introduction of an innovation, whatever arethe repercussions it can have both at the level offormal development of the brooch and organiza-tion of the work, doesn’t mean for all the surren-der of known technical processes besides. So,brooches with foot attached on the bow continueand keep on being developed, even after theinvention of the footmaking bodywith the bowatthe end of 2nd. c. BC. The introduction of castingat the beginning of 1st c. BC is not translated by
Fig. 4: Proposal of schematic restoration of different steps of manufacturing: A) brooches with brought backspring, B) brooches with included hinge (in several pieces).
GRP : fyssen JOB : annales27Internet DIV : 12mp⊕KAURIN⊕Jenny p. 10 folio : 123 --- 14/5/013 --- 15H27
PALEONTOLOGY / ARCHAEOLOGY
123

the surrenderofbroochescompletelyobtainedbyhammering. But it seems to mark the surrenderof brooches with foot attached on the bow (whichcontinue only in south-alpine area). On thecontrary, the hinge, that it is included or obtai-ning with a brought back spring, stands out veryquickly. Possibilities offered by mass productionseem to have been right of all the processesinvolving manufacturing of unique piece. Onlyironbrooches, littlenumerousat thebeginningof1st c. AD, maintain a know-how inherited fromseveral centuries. However they will quicklydisappear in front of profitability ofmass produc-tion of brooches casted and assembled mechani-cally, which authorize recourse to alloys lessexpensive and don’t require a specialized know-how.
This first study must be now verified by paleo-metallurgical analyses to confirm restorations
proposed onmanufacturing processes. Awork ofquantification could also allow to measure res-pective importance of use of these various tech-nical processes in time and according to thedifferent aeras of theCelticworld, so as to specifythe modalities of this evolution. This approachcould show itself extremely pertinent.
AcknowledgementsI would like to thank, first and foremost, the
Fyssen Fondation, that gaveme the possibility tocarry on with my researches at the University ofRoma la Sapienza and Professor P. Piana Agos-tinetti for her welcome and her invaluable help. Iwould like to thankalso theother instituteswhichwelcome me for my documentary researches:the French School of Roma (Ecole française deRome) and the european archaeological center ofBibracte.
GRP : fyssen JOB : annales27Internet DIV : 12mp⊕KAURIN⊕Jenny p. 11 folio : 124 --- 14/5/013 --- 15H27
PALEONTOLOGY / ARCHAEOLOGY
124

Datation 14C et alimentation : une clé pour appréhenderla mobilité des populations côtières
de la Péninsule Arabiqueau Néolithique
Antoine ZAZZO
Chargé de recherches au CNRS UMR 7209 « Archéozoologie, Archéobotanique : Sociétés, Pratiques etEnvironnements » au Muséum national d’Histoire naturelle
Olivia MUNOZ
Doctorante à l’université Paris 1 Panthéon-Sorbonne-CNRS UMR 7041
RésuméMalgré une position souvent marginale dans les études préhistoriques, les côtes ont été un lieu
d’attractivité primordial pour les installations humaines. Dans la péninsule d’Oman, et principale-ment le long des côtes du golfe d’Oman et de la mer d’Arabie, la présence d’environnements marinstrès productifs, associée à des ressources terrestres relativement limitées, ont conduit à uneexploitation intense des ressources marines comme en témoigne le grand nombre de sites côtiersconnus depuis le VIe millénaire avant notre ère. La question de la mobilité de ces groupes humainsa été soulevée à plusieurs reprises par les chercheurs travaillant dans la région. Notreétude aborde cette question par une approche géochimique originale, faisant intervenir la datationpar la méthode du carbone 14. Cet article en présente le contexte, les méthodes, et les premiersrésultats.
Mots-clésSaisonnalité, Néolithique, Péninsule Arabique, radiocarbon, diet
14C dating and diet : a key to understanding the mobilityof coastal populations of the Arabian Peninsula
during the Neolithic
AbstractDespite an oftenmarginal position in prehistoric studies, the coasts have been a place of attraction
for human settlements. In the Oman peninsula, mainly along the coast of the Gulf of Oman and theArabian Sea, the presence of highly productive marine environments, combined with relativelylimited land resources, have led to intense exploitation of marine resources as evidenced by the largenumber of coastal sites since the sixth millennium BCE. The issue of mobility of human groups hasbeen raised repeatedly by researchers working in the area. Our study addresses this issue by anoriginal geochemical approach, involving the radiocarbon dating method. This article presents thebackground, methods, and first results.
KeywordsSeasonality, Neolithic, Arabic Peninsula, radiocarbon dating, diet
GRP : fyssen JOB : annales27Internet DIV : 13mp⊕ZAZZO⊕Antoine p. 1 folio : 125 --- 14/5/013 --- 15H29
PALÉONTOLOGIE / ARCHÉOLOGIE
125

IntroductionMalgréunepositionsouventmarginaledansles
études préhistoriques, les côtes ont été un lieud’attractivité primordial pour les installationshumaines, l’accroissement de la population et sadispersion, puis sont aussi devenues des zonesdynamiques d’interaction culturelle et de change-ment social (Bailey et Milner 2002 ; Bailey 2004).Ces auteurs évoquent aussi la difficultéd’analyseret classifier la complexification sociale des popu-lations de pêcheurs comme on l’a fait pour leschasseurs-cueilleurs. Dans la péninsule d’Oman,et principalement sur la côte du golfe et de la merd’Oman, la présence d’environnements marinstrès productifs, associée à des ressources terres-tres relativement limitées, ont conduit à uneexploitation intense des ressourcesmarines (Tosi1975 ; 1986). Dans l’Antiquité, plusieurs récitsont décrit les populations côtières de l’Arabiecomme des ichtyophagoi, en raison de leur régimealimentaire tourné vers les ressources marines.
Si la présence humaine est attestée dans larégion depuis au moins la moitié du VIIIe millé-naire par des assemblages lithiques (faciès de« Fasad »), l’érosion, la déflation éolienne, et laremontée du niveaumarin empêchent de connaî-tre les habitats de ces groupes humains (Méryet Charpentier 2009). Néanmoins, à partir duVIe millénaire avant notre ère, un grand nombredesitesarchéologiques identifiés le longdescôtesdu golfe d’Oman et de la mer d’Arabie atteste del’ancienneté de l’exploitation des environne-ments côtiers par les hommes dans la région. Eneffet, sur ces sites, les données archéologiquesmontrent la place prédominante des ressourcesmarines au sein de l’économie de subsistance despopulations côtières dès le VIe millénaire, lachasse et l’élevage jouant un rôle secondaire bienque réel. Cette exploitation des ressources mari-nes ne s’est pas limitée à l’acquisition de nourri-ture, puisque les populations ont employé systé-matiquement lesmatériauxmarins (coquilles, os,perles fines) disponibles pour la productiond’outils et d’objets de prestige, pouvant aussi êtredestinés aux échanges.
A partir de 3200 av. J.-C. environ, les sociétésoccupant la péninsule d’Oman connaissent deprofondes transformations économiques avecl’entrée dans un réseau d’échanges inter-régio-naux, l’adoption de nouvelles techniques (métal-lurgie, poterie), de nouveaux modes de produc-
tiondenourriture (agricultured’oasis, irrigation,intensification de la pêche), des innovations pro-bables dans les techniques de conservation desproduits de l’agriculture et de la pêche (Cleuziou,2002). Ces innovations s’accompagnent demuta-tions sociales, perceptibles notamment à traversdeschangementsdesmodesd’habitat (structurespérennes en brique crue), et des pratiques funé-raires.Cetteévolutionse traduitparuneaugmen-tation de la population et un changement del’occupation et de l’exploitation du territoire.
Du VIe au IVe millénaire avant notre ère, lagrande majorité des sites connus sont situés lelong de la côte, mais on connaît également quel-ques rares sites dans l’intérieur des terres, et leschercheurs s’accordent en général pour qualifierces installations de saisonnières. Les argumentsproposés peuvent être liés à la stratigraphie dessites (présence de couches stériles intercaléesavec des couches d’occupation), la nature desstructures d’habitat (structures temporairesen matériaux légers), les données archéozoo-logiques (courbe d’abattage des animaux domes-tiques, courbe de croissance des coquillages etdes poissons, présence d’espèces dont la migra-tion est saisonnière), les données archéobotani-ques (présence de fruits dont la maturité estsaisonnière), les données anthropologiques etfunéraires (absence de certaines classes d’âgedans les tombes, orientation des corps selon laposition du soleil, présence de tombes secondai-res), et la présence d’objets ou de matériauxexogènes (coquilles marines dans l’intérieur) etles conditions climatiques (température élevéesur la côte en été, associée à une richesse halieu-tique moindre, disponibilité des ressources eneau selon les saisons).
A partir de la fin du IVe millénaire, de nom-breux sites apparaissent dans l’intérieur des ter-res, souvent associés à l’agriculture d’oasis ou àl’extraction de minerai de cuivre et de pierretendre, tandis que sur les côtes, l’occupation sepoursuit sur des sites où les indices d’échangesintra (présence de dattes, de cuivre, et de pierretendre sur la côte, et de coquilles dans l’intérieur)et inter-régionaux (objets importés de laMésopo-tamie, de l’Iran et de l’Indus) sont de plus en plusintenses.
La question de la mobilité de ces groupeshumains a été soulevée à plusieurs reprises parles chercheurs travaillant dans cette région. La
GRP : fyssen JOB : annales27Internet DIV : 13mp⊕ZAZZO⊕Antoine p. 2 folio : 126 --- 14/5/013 --- 15H29
PALÉONTOLOGIE / ARCHÉOLOGIE
126

péninsuled’Omancomporteunegrandediversitéde niches écologiques exploitables par l’homme,depuis le littoral (milieu marin, mangrove),jusqu’aux hauts plateaux de l’intérieur des terresen passant par les zones désertiques et de pié-mont. Soulignons que les conditions climatiquesarides qui prévalent actuellement, caractériséespar une absence des pluies d’été, se sontmises enplace au cours du IVe millénaire avant notre ère.La saisonnalité de la disponibilité des ressourcesdans cesnichesdiverses, associée àunmilieupeufavorable, est à l’origine d’un modèle générald’exploitation saisonnière des ressources, pro-posé sur la based’observations ethnographiques.
Les observations ethnographiques desbédouins sub-actuels en Oman (dans la région duJa’alan), ont mis en évidence une mobilité desgroupes (oud’unepartie), dont l’économiereposesur trois ressources principales : l’existence d’unsystème agricole fondé sur le palmier-dattier,l’élevage et la pêche. Au cours de cycles annuelsou pluri-annuels, les populations se déplacent enfonction de la période de récolte des dattes, ladisponibilité de pâturages, ou la saison la plusfavorable pour la pêche, Les ethnologues ontaussimontré que les surplus éventuellement pro-duits sont échangés au sein d’un réseau régional(Cordes et Scholz 1980 ; Lancaster et Lancaster1992 ; 1996 ; 2002).
Si ce modèle général semble pouvoir s’appli-quer aux populations archéologiques pratiquantl’agriculture d’oasis (à partir de la fin du IVe mil-lénaire avant notre ère), qu’en était-il aux pério-des précédant l’Age du Bronze ancien, quand lasubsistance reposait sur d’autres ressources ali-mentaires ? Les indices archéologiques sontténus, et si à toutes les périodes, la présenced’objets ou denrées exogènes sur les sites attesteparfois de leur déplacement – des dattes, du cui-vre, ou des pierres tendres, provenant de l’inté-rieur sont fréquemment retrouvés dans les sitescôtiers, tandis que des coquillages marins manu-facturéssontretrouvésdans l’intérieurdesterres– il reste à savoir si c’est dans le cadre de réseauxd’échangeset/oudemouvementsdepopulations,et quelles étaient lamesure et lesmodalitésde cesmouvements. Quelle que soit la période concer-née, plusieurs questions restent ouvertes : lesdéplacements concernaient-ils tout ou une partieseulement du groupe ? Étaient-ils réalisés sur labase de cycles saisonniers, annuels, ou pluri-an-
nuels ? La péjoration du climat a-t-elle eu unimpact sur les comportements ? Une sédentaritérelative a-t-elle pu voir le jour dans certainsmilieux favorables ? L’intensification des échan-ges au cours du IIIemillénaire a-t-elle transforméles modes d’occupation du territoire ?
Danscet article, nousprésentons lesprincipauxrésultatsd’uneétudepiloteutilisant lesoutilsde lagéochimie isotopique afin de reconstruire la partde ressources marines entrant dans la diète despopulations côtières omanaises au IVe millénaireavantnotreère.Atravers l’alimentation,notrebutest d’obtenir des informations sur la mobilité desgroupes humains étudiés.
Carbone 14, effet réservoir marinet alimentation
Depuis une trentaine d’année, une littératureabondante a établi les bases de l’écologie isotopi-que et a validé l’utilisation des isotopes stablespour reconstruire différents aspects de l’alimen-tation humaine chez les populations actuelles etpassées (Ambrose 1993). Pour ce qui est de ladétection de ressourcesmarines dans l’alimenta-tion, les traceurs généralement employés sont lesrapports en isotopes stables du carbone (d13C) etsurtout de l’azote (d15N) qui peuvent être mesu-résdanslecollagènedel’os.Enzonetempérée, lesressourcesmarinesontgénéralementdesvaleursd13C et de d15N plus élevées que les ressourcesterrestres, et cette différence se retrouve chez leshumains qui consomment ces deux types de res-sources. Cette caractéristique a par exemple per-mis d’établir l’importance des ressources mari-nes et plus particulièrement du poisson, dansl’alimentation des populations mésolithiquescôtières d’Europe occidentale (Richards et Hed-ges 1999). Il est beaucoupmoins aisé d’appliquerce type d’approchedans la péninsuleArabique oulesplantesàphotosynthèseenC3etC4co-existentet où l’aridité est forte, ce qui peut induire unrecouvrement des valeurs de d13C et de d15N desressources terrestre et marines. De plus, le colla-gène est rapidement dégradé dans les restesarchéologiques du fait des conditions arides et nepeut donc être extrait. C’est pourquoi nous avonseurecoursauneapprocheoriginale impliquant ladatation par la méthode du carbone 14 (14C).
Si les propriétés du 14C comme chronomètresont bien connues des archéologues, celles du 14Ccomme traceur de l’alimentation le sont moins.Cetteutilisationest rendupossiblepar le faitque le
GRP : fyssen JOB : annales27Internet DIV : 13mp⊕ZAZZO⊕Antoine p. 3 folio : 127 --- 14/5/013 --- 15H29
PALÉONTOLOGIE / ARCHÉOLOGIE
127

carbone présent dans le réservoir marin n’est pasà l’équilibre avec le carbone présent dans le réser-voiratmosphérique, lieudeproductionducarbone14. En moyenne, à l’échelle globale, le carbonemarin est environ 400 ans plus vieux que le car-bone atmosphérique : c’est ce qu’on appelle l’effetréservoir marin (ERM). Ce décalage, lié au tempsde résidencemoyen du carbone dans l’océan, peutvarier à l’échelle locale en fonction des spécificitésrégionales de la circulation océanique. Un termecorrectif appelé l’effet réservoir local (noté DR) aété défini pour tenir compte de ces variations. Desvaleurs négatives deDRsontmesurées lorsque lesconnections avec l’océan ouvert sont limitées, oulorsque les apports d’eau douce (à l’équilibre avecle CO2 atmosphérique) sont importants. Al’opposé, des valeurs positives de DR sont mesu-rées dans les régions ou les courants marins fontremonter des eaux froides et anciennes à la sur-face, un phénomène appelé upwelling par lesanglo-saxons. C’est le cas en mer d’Arabie et desvaleurs de + 100 à + 200 ans ont étémesurées pourles périodes récentes le long des cotes omanaises.Ces questions sont bien connues des archéologuestravaillant en milieu côtier car bien souvent lescoquilles marines sont les seuls objets datablesdans les sites d’occupation humaine. La connais-sance de l’effet réservoir est donc cruciale si l’onveut dater le site avec précision.
En quoi le carbone 14 peut-il servir de traceuralimentaire ? Les atomes de carbone qui sontcontenus dans l’alimentation sont métaboliséspar notre organisme et contribuent au renouvel-lement tissulaire. La signature chimique de l’ali-mentation se retrouve ainsi dans nos organes,muscles, os, oudent, validant l’adage selon lequel«onestcequ’onmange».Ainsi,un individuqui senourrit exclusivement de ressources terrestressera à l’équilibre avec le réservoir de 14C del’atmosphère. En d’autres termes, son âge 14Csera identique à son âge biologique à l’erreur demesure près (± 30 ans en général). A l’opposé, unindividu dont l’alimentation serait exclusive-ment constituée de ressources marines sera àl’équilibre avec le réservoir de 14C marin et donccomparativement plus « vieux » (en âge 14C) queson âge biologique. La différence d’âge 14C entredeux individuscontemporainsdépendradoncà lafois de la proportion de ressources marines dansla diète et de l’importance de l’effet réservoir enun lieu donné. En contexte archéologique, si la
dated’inhumationet l’effet réservoir sont connusavec suffisamment de précision, alors la diffé-rence entre l’âge 14Cd’un individu et le temps quis’est écoulé depuis sa mort est fonction de laproportion de ressourcesmarines contenue dansson alimentation.
L’effet réservoir local varie dans l’espace maiségalement dans le temps. Il est donc délicatd’appliquer automatiquement les valeurs moder-nes aux périodes anciennes. Comment procède-t-on pour mesurer cet effet réservoir dans lespériodes anciennes ? L’approche communémentadoptée consiste à comparer l’âge 14Cmesuré surdes paires contemporaines de restes marin(coquille de mollusque, os de vertébré marin) etcontinental (charbon, os de vertébré terrestre).La différence d’âge indique alors l’importance del’effet réservoir marin pour cette période. Toutela difficulté réside dans le fait de sélectionnerdeuxobjets qui soient réellement contemporains,ce qui peut se révéler délicat en contexte archéo-logique. Les tombes sont généralement considé-rées comme susceptibles de fournir un contexteclos a priori qui, s’il renferme des restes terres-tres et marins, offre une bonne chance qu’ilssoient contemporains.
Figure 1 – Location des principaux sites préhis-toriques de la région de Ra’s al-Hamra et Qurum(Mascate, Sultanat d’Oman).
GRP : fyssen JOB : annales27Internet DIV : 13mp⊕ZAZZO⊕Antoine p. 4 folio : 128 --- 14/5/013 --- 15H30
PALÉONTOLOGIE / ARCHÉOLOGIE
128

Le site de Ra’s al-Hamra RH5Situédans laville actuelledeMascate,RH5fait
partie d’un ensemble de sites archéologiquesidentifiés sur un promontoire rocheux, à proxi-mité de la côte et d’une mangrove (Durante etTosi, 1977) (Figure 1). RH5 se caractérise par undépôt anthropique pluristratifié qui témoigned’une occupation quasi ininterrompue depuis lafin du Ve jusqu’au milieu du IVe millénaire avantnotre ère. On y distingue des zones d’habitat etdes traces d’activités (alignements de trous depoteaux, tranchées de fondation, foyers, etc.)(Biagi et al., 1984 ; Biagi et al., 1989 ; Biagi andNisbet, 1999, 2006) ainsi qu’un cimetière où plusde 215 individus sont représentés (Coppa et al.,1986, 1990 ; Salvatori, 1998, 2007). Les défunts,qui portaient parfois des ornements personnels(colliers, bracelets et boucles d’oreille) étaientgénéralement enterrés dans une fosse ovale,reposant en décubitus latéral dans une positionfléchie. Des dépôts de faune ont également étéretrouvés dans certaines tombes (tortue marine,coquillages, poissons). La nouvelle campagne defouilles qui a débuté en 2005 a permis demontrerque le cimetière était plus étendu vers l’ouest queceque l’onpensait alors (Munoz souspresse). Lesnouvelles tombes identifiées ont été fouillées ensuivant les méthodes de « l’anthropologie deterrain »définies parH.Duday (2009) afind’enre-gistrer le maximum d’informations relatives auxprocessus complexes des dépôts durant les céré-monies funéraires. Ce sont ces tombes, apparte-nant aux niveaux les plus récents de la nécropole,qui ont été sélectionnées pour les besoins de cetterecherche.
Estimation de l’effet réservoir marinà RH5
Nous avons tout d’abord cherché à déterminerl’ERM dans trois tombes fouillées en 2008. Danschaque tombe, nous avons daté des restes terres-tres (charbons) et des restes marins et de man-grove (coquilles). Les âgesmesurés sur charbonsau sein d’une même tombe montre une certainevariabilité (jusqu’à 240 ans en âge 14C) qui pour-rait être due à un effet « vieux bois », classiquedans cesmilieux arides, bien qu’un remaniementde charbons anciens présents dans le sol lors ducreusement de la tombe ne puisse pas être tota-lement à écarter. Dans tous les cas, on peutconsidérer que la date d’inhumation est appro-chée aumieux par le charbon le plus récent. Pour
chaque tombe, nous avons ensuite calculé la dif-férence d’âge entre ce charbon et les coquillesmarines et de mangrove. Si trois coquilles indi-quent un ERM supérieur à 700 ans pouvant êtrelié à un effet « vieille coquille » la plupart desrestes marins (n= 18) indiquent un effet réser-voir compris entre 500 et 700 ans. Une analyseselon la méthode des clusters indique deuxfamilles de dates significativement différentesl’unede l’autre et dont lamoyennese situeautourde 508 ± 16 et 668 ± 41 ans, respectivement(Figure 3). Plusieurs arguments suggèrent quecette variabilité pourrait correspondre à un phé-nomène réel. Aujourd’hui présente à l’état derelique, la mangrove de Qurum était beaucoupplus développée au IVe millénaire et devait assu-rer une partie non négligeable des ressourcesalimentaires disponibles sur la côte. Lesmangro-ves se situent à l’interface du continent et del’océan et constituent ainsi une zone de recyclagede la matière organique terrestre. Nous avonsmesuré un effet réservoir plus faible dans descoquillages strictement inféodées à cet environ-nement (comme le gastéropode Terebralia palus-tris), ce qui suggère que la variabilité observéeserait bien d’origine environnementale. Parailleurs, nous avons obtenu deux dates diver-geant de plus de 140 ans sur unmême bivalve, cequi pourrait témoigner d’une variabilité saison-nière de l’effet réservoir, un phénomène observépar ailleurs en Californie, une région égalementmarquée par la présence d’un upwelling côtier(Culleton et al. 2006). Selon notre hypothèse, lamoyenne haute correspondrait donc à l’ERM,tandis que la moyenne basse correspondrait auxvaleurs enregistrées par les organismes vivantsous l’influence directe de la mangrove.
Estimation de la proportion de ressourcesmarines dans la diète
Nous avons ensuite cherché à dater leshumains présents dans ces trois tombes. Il s’agitde trois adultes et d’un enfant. En règle généralele collagène de l’os est utilisé comme support desdatations 14C. Du fait des conditions aridesrégnant dans la région, le collagène de l’os esttotalement dégradé et ne pouvait donc être daté.La seule alternative était de dater la fractionminérale de l’os qui contient suffisamment decarbone (sous forme de carbonate) pour permet-tre une datation 14C. Des travaux menés parnotre équipe ont montré que dans les zones
GRP : fyssen JOB : annales27Internet DIV : 13mp⊕ZAZZO⊕Antoine p. 5 folio : 129 --- 14/5/013 --- 15H30
PALÉONTOLOGIE / ARCHÉOLOGIE
129

arides (Afrique, Péninsule Arabique) le carbo-nate de l’apatite était un support extrêmementfiable desdatations 14Caumoinspour l’Holocène(Zazzo et Saliège 2011). Les résultats montrentque les individus ont un âge 14C compris entre5250 et 5360 BP, c’est-à-dire environ 390 à540 ans plus vieux que les charbons des mêmestombes (Figure 3). Une simple règle de troispermet de calculer que la proportion de ressour-ces marine dans la diète représente 80 à 100 %
environ pour les trois adultes, et 60 à 80 % pourl’individu immature. Les restes humains appar-tenant à 16 individus retrouvés dans d’autrestombes ont pu être datés et confirment ce résul-tat, avec des âges compris entre 5100 et 5450BP.Dans plusieurs de ces tombes, des charbons ontpu être également datés et indiquent une diffé-rence de 400 ans environ avec les humains. Cerésultat indique une consommation très impor-tante de ressources aquatiques de la part de la
Figure 2 – Photo (gauche) and schéma (droite) de la tombe 411, Ra’s al-Hamra RH5, Sultanat d’Oman,montrant les restes fauniques (haut) déposés sur le squelette d’un adulte male en position fléchie et reposantsur le côté droit (bas).
GRP : fyssen JOB : annales27Internet DIV : 13mp⊕ZAZZO⊕Antoine p. 6 folio : 130 --- 14/5/013 --- 15H31
PALÉONTOLOGIE / ARCHÉOLOGIE
130

population, même s’il est pour l’instant difficilededistinguer la part relative des ressourcesmari-nes à proprement parler et de mangrove. Il fauten outre garder à l’esprit que certaines ressour-ces alimentaires sont uniquement trouvées sur lacôte mais possèdent une signature terrestrecomme les mollusques de mangrove Terebraliapalustris qui sont retrouvés en quantité dans lesite de RH5. Le signal marin incorporé dans lestissus des humains doit donc être considérécomme une estimation minimale de la dépen-dance au milieu côtier. Etant donné la place pré-pondérante du signal marin dans l’alimentation,il est difficile d’imaginer que les individus analy-sés s’éloignaient durablement de la côte, sauf àimaginer qu’ils emportaient avec eux ces ressour-ces durant leurs déplacements. Cette hypothèseest plausible, car les récits de voyageurs antiquessuggèrent que les techniques de conservation desdenrées marines (séchage, fumage et salage du
poisson, séchage des coquillages), – par ailleursbien documentées par des travaux ethnographi-ques actuels (ElMahi 2000) – étaient déjàmaîtri-sées il y a longtemps par les populations côtièresomanaises. Cependant, si cette hypothèse étaitvérifiée, elle impliquerait tout demême une fortedépendance vis-à-vis des ressources marines.
Nos résultats vont donc clairement dans lesens d’une occupation permanente et non saison-nière de la côte. Cette conclusion rejoint celleformulée récemment par Biagi et Nisbet (2006)sur la base notamment du réexamen des structu-res d’habitat, du choix des sites retenus par lespopulations du Ve millénaire, du type d’activités(artisanat) réalisées sur ce site et de la présenced’un cimetière pendant au moins 200 ans, quiindique un ancrage territorial transmis sur aumoins dizaine de générations. RH5 est l’un desnombreux amas coquilliers datant du Ve millé-naire identifiés lors de prospections (Cleuziou
Figure 3 – Effet réservoir marin (ERM) enregistré dans les coquilles marines et de mangrove, les poissons etles humains des tombes 405, 408 et 411.
GRP : fyssen JOB : annales27Internet DIV : 13mp⊕ZAZZO⊕Antoine p. 7 folio : 131 --- 14/5/013 --- 15H31
PALÉONTOLOGIE / ARCHÉOLOGIE
131

2005). Parmi ces sites, seuls quatre ensemble desites ont livré des tombes : Ra’s al-Hamra (RH),Wadi Shab (GAS), Ra’s al-Khabbah (KHB) etSuwayh (SWY). Ces sites ont en commun d’êtreinstallés à proximité d’une mangrove ou d’unenvironnement marin particulièrement produc-tif.Dansunmilieuqui s’aridifie, cesniches ontpuattirer des populations qui s’y sont sédentariséesavant que de nouvelles techniques transformentles possibilités d’exploitation du territoire,notamment à travers un réseau d’échange quis’intensifie.
Ce travail ouvre la voie à d’autres études. Siune extension à l’échelle régionale est envisagea-ble pour le Néolithique, l’évolution des pratiquesfunéraires à partir de l’âge du Bronze renddélicate l’utilisation de ce type d’approche pourdes sites plus récents. En effet, les sépulturesindividuelles du Néolithique laissent leurplace aux sépultures collectives de l’époqueHafit et Umm an-Nar. L’intervention répétée del’homme pendant la durée de fonctionnement deces structures réduit fortement les chances detrouver des associations de restes terrestres etmarin contemporains des individus inhumés,condition nécessaire du succès de notre appro-che. Cependant le recours à d’autres approchesgéochimiques, faisant intervenir la mesure durapport isotopique du carbone (d13C) de l’oxy-gène (d18O), et du strontium (87Sr/86Sr) dans lafractionminérale des os et des dents permettrontpeut-être d’aborder l’évolution des pratiques ali-mentaires dans la région.
RemerciementsCe travail a été rendu possible grâce au
soutien financier de la Fondation Fyssen, duCNRS- INSHS et du Ministère Omanais de laCulture. Il a bénéficié de l’aide et des conseils denombreux collègues sur le terrain (L.G. Mar-cucci, G. Seguin, N. Gryspeirt, N. Rolland,M. Tosi) et au laboratoire (P. Béarez, S. Kerneur,C. Martin, C. Pierre, M. Tengberg, J. Ughetto,ainsi que toute l’équipe du LMC14). Cet articleest dédié à la mémoire de S. Cleuziou et J.-F.Saliège.
Bibliographiee Ambrose S. (1993). Isotopic analysis of
paleodiets: Methodological and interpretiveconsiderations. In (M. K. Sandford Ed.) Investi-
gations ofAncientHumanTissue: Chemical Ana-lyses in Anthropology. Langhorne, PA: Gordonand Breach Science Publishers,pp. 59–130.
e Bailey G.N. (2004). World prehistory fromthe margins: the role of coastlines in humanevolution. Journal of InterdisciplinaryStudies inHistory and Archaeology 1 (1): 39–50
e BaileyG.N.,MilnerN.J. (2002).Coastalhun-ters and gatherers and social evolution:marginalor central? Before Farming: the Archaeology ofOld World Hunter-Gatherers 3–4 (1): 1–15.
e Biagi P., Maggi R., Nisbet R. (1989). Exca-vations at the aceramic coastal settlement ofRH5 (Muscat, Sultanate of Oman) 1983-85. In:Frifelt K., Sorensen P. (Eds.), South AsianArchaeology (1985), pp. 1-8
e Biagi P., Nisbet R. (1999). The shell-middensites of RH5 and RH6 -Muscat, Sultanate ofOman. Archaeologia Polona 37, 31-47.
e Biagi P., Nisbet R. (2006). The prehistoricfisher-gatherers of the western coast of the Ara-bian Sea: a case of seasonal sedentarization?World Archaeology 38, 220-258.
e Biagi P., TorkeW., TosiM.,UerpmannH.-P.(1984). Qurum:ACaseStudy ofCoastal Archaeo-logy in Northern Oman. World Archaeology 16,43-61.
e Cleuziou S. (2002). The Early bronze Age ofthe Oman Peninsula: from chronology to thedialectics of trade and State formation. In: Cleu-ziou S., Tosi M., Zarins J. (Eds.), Essays on theLate Prehistory of the Arabian Peninsula, Rome:IsIAO Serie Orientale Roma, pp. 191-236.
e Cleuziou S. (2005). Pourquoi si tard? Nousavons pris un autre chemin. In: Guilaine J. (Ed.)Aux marges des grands foyers du Néolithique.Périphéries débitrices ou créatrices ? Paris,Errance, pp. 123-148.
e Coppa A., Macchiarelli R., Salvatori S., San-tini G. (1986). The Prehistoric Graveyard of Ra’sal-Hamra (RH5): a short preliminary report onthe 1981-1983 excavations. The Journal of OmanStudies 8, 97-102.
e Coppa A., Damadio S.M., Armelagos G.J.,MancinelliD.,VargiuR. (1990).PaleobiologyandPaleopathology. A preliminary study of the pre-historic fishing population of Ra’s al-Hamra5 (Qurum, Sultanate of Oman, 3700-3200 BC).Antropologia Contemporanea 13, 329-336.
GRP : fyssen JOB : annales27Internet DIV : 13mp⊕ZAZZO⊕Antoine p. 8 folio : 132 --- 14/5/013 --- 15H31
PALÉONTOLOGIE / ARCHÉOLOGIE
132

e Cordes R., Scholz F. (1980). Bedouins,wealth, andchange:Astudyof rural developmentin the United Arab Emiratesand the Sultanate ofOman. United NationsUniversity (Tokyo,Japan).
e Culleton B.J., Kennett D.J., Ingram B.L.,Erlandson J.M., Southon J.R. (2006). Intrashellradiocarbon variability in marine molluscs.Radiocarbon 48, 387-400.
e Duday H. (2009). The Archaeology of theDead, London, Oxbow.
e Durante S., Tosi M. (1977). The AceramicShell Middens of Ra’s al-Hamra: A PreliminaryNote. Journal of OmanStudies 3, 137-172.
e El Mahi AT (2000) Traditional fish preser-vation in Oman: the seasonality of a subsistencestrategy.Proceedings of theSeminar forArabianStudies 30:99-113.
e Lancaster W., Lancaster F. (1992). Tribe,Community and theConcept ofAccess toResour-ces: Territorial Behaviour in South-East Ja’alan,in Casimir M. J. et A. Rao (eds), Mobility andTerritory, Social and Spatial Boundaries amongForagers, Fishers, Pastoralists and Peripatetics,New-York, p. 343-363.
e Lancaster F (2002) Trade and stratifiedsociety information from south-east Ja’alan,Oman. In S Cleuziou, M Tosi and J Zarins (eds.):Essays on the Late Prehistory of the ArabianPeninsula. Rome: IsIAO, pp. 247-256.
e LancasterW,andLancasterF (1998)Reflec-tions on the social organization of the ArabianBedu in coastal Oman. The Prehistory of ArabiaandOceania, Proceedings of the XIIIth Congressof the IUPPS, Forli, 8-14 sept. 1996, pp. 141-154.
e Méry S, and Charpentier V (2009) Ritesfunéraires du Néolithique et de l’âge du Bronzeancien en Arabie orientale. In J Guilaine (ed.):Sépultures et sociétés. Du Néolithique à l’His-toire. Paris: Errance, pp. 11-40.
e Munoz O. (sous presse). Funerary practicesat Ra’s al-Hamra RH5: New data from (2005)-2009 excavations. In: Cleuziou S., Tosi M. (Eds.),In the shadow of the ancestors, 2d edition, Mus-cat: Ministry of Heritage & Culture.siteRH-5 (Ra’s al-Hamra, Muscat, Sultanate ofOman). Proceedings of the Seminar for ArabianStudies.
e Richards M.P., Hedges R.E.M. (1999). Sta-ble isotope evidence for similarities in the types
of marine foods used by late Mesolithic humansat sites along the Atlantic coast of Europe. Jour-nal of Archaeological Science 26, 717–722.
e Salvatori S. (1998). Death and ritual in apopulation of coastal food foragers in Oman. In:Afanas’ev G., Cleuziou S., Lukacs J.R., Tosi M.(Eds.), The Prehistory of Arabia and Oceania,Proceedings of the 13th Congress of the IUPPS,Forli, 8-14 Sept. (1996). Forli: ABACO, pp. 205-222.
e Salvatori S. (2007). The Prehistoric Gra-veyard of Ra’s al-Hamra 5, Muscat, Sultanate ofOman. The Journal of OmanStudies, 14.
e Tosi M. (1975). Notes on the distributionand exploitation of natural resources in ancientOman. The Journal of OmanStudies 1: 187–206.
e Tosi M. (1986). The emerging picture ofprehistoric Arabia. Annual Review of Anthropo-logy, 15: 461–90.
e Uerpmann M., Uerpmann H-P., Jasim, S.A.(2000). Stone Age nomadism in SE-Arabia –pa-laeoeconomic considerations on the Neolithicsite of Al-Buhais 18 inthe Emirate of Sharjah,U.A.E. Proceedings of the Seminar for ArabianStudies, 30: 229–34.
e Zazzo A., Saliège J.F. (2011). Radiocarbondating of biological apatites: a review. Palaeogeo-graphy, Palaeoclimatology, Palaeoecology 310,52-61.
IntroductionDespite an often marginal position in prehis-
toric studies, the coasts have been a place ofattraction for human settlements, increasingpopulation and its dispersion, and also becamedynamic zones of cultural interaction and socialchange (Bailey and Milner 2002, Bailey 2004).These authors also mention the difficulty of ana-lyzing and classifying the social complexity offishing communities as it was the case for hunter-gatherers. In the peninsulaof Oman, mainly onthe coast of the Gulf and of the Oman Sea, thepresence of highly productive marine environ-ments, combined with relatively limited landresources, have led to intense exploitation ofmarine resources (Tosi 1975, 1986). In ancienttimes, many stories described the Arabic coastalpopulations as ichtyophagoi, due to theirmarine-oriented diet.
If the human presence is attested in the regionsince at least half of the eighth millennium by
GRP : fyssen JOB : annales27Internet DIV : 13mp⊕ZAZZO⊕Antoine p. 9 folio : 133 --- 14/5/013 --- 15H31
PALÉONTOLOGY / ARCHÉOLOGY
133

lithic assemblages (”Fasad“ facies), erosion,wind erosion, and sea level rise destroyed thehabitat of these human groups (Méry and Char-pentier 2009). However, from the sixth millen-nium BC, a large number of archaeological sitesidentified along the coastline of the Gulfof Omanand the Arabian Sea demonstrates the antiquityof the exploitation of coastal environments byman in the region. Indeed, on these sites,archaeological data show the dominance ofmarine resources in the subsistence economy ofcoastal populations from the sixth millennium,hunting and livestock playing a secondary role,although real. The exploitation of marine resour-ces was not limited to the acquisition of food, aspeople have used systematically marine mate-rials (shells, bones, pearls) for the production oftools and objects of prestige, which could also beintended for trade.
From about 3200 BC, the societies occupyingthe peninsula of Oman underwent profound eco-nomic transformation with the entry into anetwork of inter-regional trade, the adoption ofnew technologies (metallurgy, pottery), newmodes of food production (oasis agriculture, irri-gation, intensification of fishing), and possibleinnovations in techniques for preserving agricul-tural products and fisheries (Cleuziou, 2002).These innovations are accompanied by socialchange, particularly throughperceptible changesin settlement patterns (permanent structures ofmud brick), and funerary practices. This is reflec-ted by an increase in population and a change ofoccupation and land use.
From the sixth to the fourth millennium BCE,the vast majority of known sites is located alongthe coast, with also a few sites in the interior, andresearchers generally agree to describe thesefacilities as seasonal. The arguments put forwardmay be related to the stratigraphy of the sites(presence of sterile layers between layers of occu-pation), the nature of the habitat structures (tem-porary structuresmadeof lightmaterials), zooar-chaeological data (mortality profile of domesticanimals, growth rate of shellfish and fish, thepresence of specieswhosemigration is seasonal),archaeobotanical data (presence of fruit whosematurity is seasonal), and anthropological data(absence of certain age classes in graves, orien-tationof thebody, presence of secondary graves),and the presence of non-local objects ormaterials
(marine shells in the interior) and climatic condi-tions (high temperatures on the coast in thesummer, associated with decreased fish and fres-hwater availability).
From the late fourth millennium, many sitesappear in the interior, often associatedwith oasisagriculture or extraction of copper ore and softstone, while on the coast, the occupation conti-nues on sites where the indices of intra (presencedates, copper and soft stone on the coast and inthe interior shells) and inter-regional (objectsimported from Mesopotamia, Iran and Indus)exchange are more intense.
The issue of mobility of human groups hasbeen raised repeatedly by researchersworking inthis area. TheOmanPeninsula has awide varietyof ecological niches exploited by man from thecoast (marine,mangrove), to the highlands of theinterior through the desert and foothills. Notethat arid climatic conditions currentlyprevailing, characterized by an absence of sum-mer rains, appeared during the fourth millen-niumBCE.Seasonality of resource availability inthese various niches, associated with a less favo-rable environment, form the basis of a generalmodel of seasonal exploitation of resources, pro-posed based on ethnographic observations.
Ethnographic observations of sub-modernBedouin populations in Oman (in the Ja’alanregion) showed a mobility of the groups (or partof the groups), whose economy is based on threemain resources: the existence of an agriculturalsystem based on the date palm, livestock andfishing. During annual or multi-annual cycles,populations move depending on the time of dateharvest, the availability of pastures, or the mostfavorable season for fishing. Ethnologists havealso shown that the surplus products are exchan-gedwithina regionalnetwork (CordesandScholz1980; Lancaster and Lancaster 1992, 1996,2002).
If this general pattern seems to apply toarchaeological populations practicing oasis agri-culture (from the late fourth millennium BCE),does it hold for periods prior to the Bronze Age,when the subsistence economy was based onother food resources? Archaeological evidence isscarce, and if at all periods, the presence ofexogenous food items or sites attests sometimestheir movement - dates, copper or soft stonesfrom the interior are frequently found in coastal
GRP : fyssen JOB : annales27Internet DIV : 13mp⊕ZAZZO⊕Antoine p. 10 folio : 134 --- 14/5/013 --- 15H31
PALÉONTOLOGY / ARCHÉOLOGY
134

sites, while manufactured marine shells arefound in the interior – the question remainswhether this is through exchange networks and/ or population movements, and what was theextent and terms of thesemovements.Whateverthe period, several questions remain open: didthe movements concern all or part of the group?Were they made on a seasonal, annual or multi-annual basis?Did the deterioration of the climatehave an impact on mobility? Could a form ofsedentarity have appeared in more favorableenvironments? Did increased trade during thethird millennium transform patterns of landuse?
In this paper, we present the main results of apilot study using isotope geochemistry to recons-truct the proportion of marine resources in thediet of coastal Oman populations during thefourthmillenniumBCE. Through diet, our aim isto obtain information on the mobility of humangroups studied.
Radiocarbon, diet and marine reservoireffect
For thirty years, an extensive literature hasestablished the basis for isotopic ecology andvalidated the use of stable isotopes to reconstructaspects of the human diet in the past and presentpopulations (Ambrose 1993). Regarding thedetection of marine resources in the diet, thetracers commonly used are stable isotope ratiosof carbon (d13C) and especially nitrogen (d15N)measured in bone collagen. In temperate zones,marine resources typically have d13C and d15Nhigher than the terrestrial resources, and thisdifference is recorded in humans who consumethese types of resources. This characteristic hasallowed, for example to establish the importanceof marine resources, especially fish in the diet ofMesolithic coastal populations of WesternEurope (Richards and Hedges 1999). It is muchmore difficult to apply this approach in theArabian Peninsula where C3 and C4 photosyn-thesis types co-exist and where aridity is strong,which can induce an overlap in d13C and d15Nvalues of terrestrial and marine resources. Inaddition, collagen is rapidly degraded in archaeo-logical remains due to the arid conditions andcannot be extracted. That is why we have used anovel approach involving radiocarbon (14C)dating.
If the properties of 14Cas a dating tool are wellknown to archaeologists, the properties of 14C asa dietary tracer are not. This use is possiblebecause the carbon in themarine reservoir is notin equilibrium with the carbon in the atmosphe-ric reservoir, where radiocarbon is produced. Onaverage, on a global scale, the marine carbon isabout 400 years older than the atmosphericcarbon: this is called the marine reservoir effect(MRE). This shift, related to the mean residencetime of carbon in the ocean can vary locallydepending on the specific regional ocean circu-lation. A correction term called the local reser-voir effect (denoted DR) was defined to reflectthese changes. Negative values of DR are measu-red when the connections with the open oceanare limited, or when freshwater inflows (in equi-librium with atmospheric CO2) are important.In contrast, positive values of DR are measuredin areas where ocean currents bring cold andold water up to the surface, the so-called upwel-ling. This is the case in the Arabian Sea andthe DR values of +100 to +200 years weremeasured for recent periods along the coastofOman. These issues arewell known to archaeo-logists working in coastal areas as marineshells are often the only datable objects in humansite occupation. Estimation of the reservoireffect is crucial if we are to date the site accura-tely.
How can radiocarbon be used as a dietarytracer? The carbon atoms contained in food aremetabolized by the body and contribute to tissueturnover. The chemical signature of the foodthus finds itself in our organs, muscles, bones, orteeth, validating the adage ”you are what youeat.“ Thus, an individual who feeds exclusivelyon terrestrial resources will be in equilibriumwith the atmospheric reservoir of 14C. In otherwords, the 14C age is broadly identical to thebiological age. In contrast, an individual whosediet would consist exclusively of marine resour-ces will be in equilibrium with the marine 14Creservoir and therefore comparatively older (in14C age) that his biological age. Radiocarbon agedifference between contemporary individualsdepends onboth theproportionofmarine resour-ces in the diet and the importance of the reser-voir effect in a given location. In archaeologicalcontexts, if the date of burial and reservoir effectare known with sufficient accuracy, then thedifference between the 14C age of an individual
GRP : fyssen JOB : annales27Internet DIV : 13mp⊕ZAZZO⊕Antoine p. 11 folio : 135 --- 14/5/013 --- 15H31
PALÉONTOLOGY / ARCHÉOLOGY
135

and the time that has elapsed since his death isa function of the proportion of marine resourcescontained in its diet.
The local reservoir effect varies in space butalso in time. It is therefore difficult to applymodern DR values automatically to ancienttimes. How does one measure this reservoireffect in ancient times? The commonly adoptedapproach is to compare 14C age of contemporarymarine (shell mollusk, marine vertebrate bones)and continental (coal, terrestrial vertebratebones) remains. The age difference then indica-tes the importance of the marine reservoir effectfor that period. The difficulty lies in the fact thatfinding two objects that are truly contemporarycan be tricky in archaeological context. Thetombs are generally considered capable of provi-ding such closed contexts which, if they containterrestrial and marine remains, offers a goodchance they are contemporaries.
The site of Ra’s al-Hamra RH5Located in the modern city of Muscat, RH5 is
part of a series of archaeological sites identifiedon a rocky promontory, near the coast and man-grove (Durante andTosi, 1977) (Figure 1).RH5 ischaracterized by a multilayered anthropogenicdeposit indicating an almost continuous occupa-tion from the late fifth to mid-fourth millenniumBCE. It differs fromresidential areas andactivitytraces (rows of postholes, foundation trenches,shelters, etc.). (Biagi et al., 1984; Biagi et al.,1989; Biagi and Nisbet, 1999, 2006) and a ceme-tery where more than 215 individuals are repre-sented (Coppa et al., 1986, 1990, Salvatori, 1998,2007). The deceased, who sometimes wore per-sonal ornaments (necklaces, bracelets andearings) were generally buried in an oval pit, in alateral decubitus flexed position. Faunal depositswere also found in some tombs (turtle, shellfish,fish). The new excavations that began in 2005showed that the cemetery extended westwardmore than if was previously thought (Munoz inpress). New tombs were identified and excavatedusing the methods of the ”field anthropology“defined by Duday (2009) to record as much infor-mation on the complex processes of depositsduring the funeral ceremonies. These are thetombs belonging to the latest levels of the necro-polis,whichwere selected for the purposes of thisresearch.
Estimation of marine reservoir effect atRH5
Wefirst sought to determine theMRE in threetombs excavated in 2008. In each grave, we datedthe terrestrial (charcoal) marine and mangroveremains (shells). Ages measured on charcoalwithin the same tomb shows some variability (upto 240 years in 14C age) which could be due to an”old wood“ effect, typical in arid sites, althoughremobilization of charcoal already present in theground while digging the grave can not be com-pletely ruled out. In any case, we can considerthat the date of burial is best approached by theyoungest charcoal. For each grave, we then cal-culated the difference in age between the char-coal marine and mangrove shells. If three shellsindicate an MRE higher than 700 years that canbe linked to an ”old shell“ effect, most marineremains (n= 18) show a MRE between 500 and700 years. Cluster analysis shows two families ofdates significantly different from each other andwhose average is around 508 ± 16 and 668 ±41 years, respectively (Figure 3). Several argu-ments suggest that this variability may reflect areal phenomenon. Today present as a relic, the
Figure 1 – Location of the main prehistoric sites inthe Ra’s al-Hamra and Qurum region (Muscat,Oman).
GRP : fyssen JOB : annales27Internet DIV : 13mp⊕ZAZZO⊕Antoine p. 12 folio : 136 --- 14/5/013 --- 15H31
PALÉONTOLOGY / ARCHÉOLOGY
136

Qurum mangrove was much more developedduring the fourth millennium and should ensurea significant portion of available food resourceson thecoast.Mangrovesareat the interfaceof thecontinent and the ocean, and thus constitute anareawhere terrestrial organicmatter is recycled.We measured a lower reservoir effect in man-grove shells such as the gastropod Terebraliapalustris, suggesting that the observed variabilitywould be of environmental origin. In addition,we
obtained two dates diverging more than140 years on a single bivalve, which may reflectseasonal variability of the reservoir effect, a phe-nomenonalsodocumented inCalifornia, a regionalso marked by the presence of a coastal upwel-ling (Culleton et al. 2006). According to our hypo-thesis, the high average could thus correspond totheMRE,while the lowaverage could correspondto the values recorded by organisms living underthe direct influence of the mangrove.
Figure 2 – Photograph (left) and schematic (right) of the grave 411, Ra’s al-Hamra RH5, Sultanate of Oman,showing faunal remains (top) deposited on the skeleton of an adult male in a flexed position and based on theright side (bottom).
GRP : fyssen JOB : annales27Internet DIV : 13mp⊕ZAZZO⊕Antoine p. 13 folio : 137 --- 14/5/013 --- 15H32
PALÉONTOLOGY / ARCHÉOLOGY
137

Estimate of the proportion of marineresources in the diet
We then sought to date the humans in thesethree graves. There are three adults and a child.Generally bone collagen is the support of choicefor radiocarbon dating. Because of the aridconditions prevailing in the region, bone colla-gen is totally degraded and could not be dated.The only alternative was to date the mineralfraction which contains enough carbon (in theform of carbonate) to allow for 14C dating. Workcarried out by our team showed that in arid areas(Africa, Arabian Peninsula) carbonate apatitewas a reliable support for 14C dating at least forthe Holocene (Zazzo and Saliège 2011). Theresults show that individuals have a 14C agebetween 5250 and 5360 BP, i.e. 390 to 540 yearsolder than the charcoals of the same graves(Figure 3). A simple rule of thumbs is used tocalculate the proportion of marine resources inthe diet of about 80 to 100% for three adults, and60-80% for immature individual. Sixteen humanremains found in other tombs were also datedand confirm this result, with ages rangingbetween 5100 and 5450 BP. In many of thesetombs, charcoals have been dated and also indi-cate a difference 400 years with the humanremains. This result indicates a very highconsumption of aquatic resources from the popu-lation, although it is currently difficult to distin-guish between the relative importance of marineand mangrove resources. It should also be bornein mind that some food resources are found onlyon the coast but have a terrestrial signature likethemangrove dwellingTerebralia palustriswhichare found in large quantities at RH5. Marinesignal embedded in human tissues should beconsidered a minimum estimate of the depen-dence to coastal environments. Given the promi-nence of the marine signal, it is difficult toimagine that the individuals analyzed perma-nently moved away from the coast, except toimagine that they carried with them theseresources during their travels. This hypothesis isplausible because the stories of ancient travelerssuggest that techniques of marine food preser-vation (drying, smoking and salting fish, shell-fish drying) - also well documented by ethno-graphic observation of modern populations(El Mahi 2000) - had already been mastered
long ago by the Omani coastal populations.However, if this hypothesis were true, it wouldstill imply a strong dependence vis-à-vis themarine resources.
Our results are therefore clearly in favor of apermanent and non-seasonal occupation of thecoast. This conclusion is similar to that recentlyformulated by Biagi and Nisbet (2006) mainly onthe basis of the review of habitat structures, thechoice of sites selected by the people of the fifthmillennium, craft activities and the presence ofa graveyard for at least 200 years, which indica-tes a territorial anchoring transmitted over atleast ten generations. RH5 is one of the manyshell middens dating from the fifth millenniumidentified during surveys (Cleuziou 2005).Among these sites, only four sets of sites haveyielded graves: Ra’s al-Hamra (RH), Wadi Shab(GAS), Ra’s al-Khabbah (KHB) and Suwayh(SWY). These sites have in common to be instal-led near a mangrove or a particularly productivemarine environment. In an environment under-going aridification these niches have been able toattract people who settled down, before newtechniques transform the exploitation potentialof the territory under a growing exchangenetwork.
This work opens the way for further studies.If an extension at the regional level is possible forthe Neolithic, the evolution of burial practicesfrom the Bronze Agemakes it difficult to use thisapproach for more recent sites. Indeed, indivi-dual burials from the Neolithic have been repla-ced by collective burials of the Hafit and Umman-Nar periods. Man-made repeated interven-tions on these structures greatly reduces thechances of finding associations between contem-porary terrestrial and marine remains, a condi-tion necessary for the success of our approach.However, the use of other geochemical, toolsincluding the measurement of the carbon, oxy-gen and strontium (d13C, d18O, 87Sr/86Sr) iso-tope ratio in themineral component of bones andteeth may help addressing the evolution of thedietary practices in the region.
AcknowledgmentsFirst of all we would like to thank the Fyssen
Foundation, CNRS-INSHS, and the OmaniMinistry of Culture for their financial support.Thisworkwas benefited from the help and adviceof many colleagues in the field (LG Marcucci,
GRP : fyssen JOB : annales27Internet DIV : 13mp⊕ZAZZO⊕Antoine p. 14 folio : 138 --- 14/5/013 --- 15H32
PALÉONTOLOGY / ARCHÉOLOGY
138

G. Seguin, N. Gryspeirt, N. Rolland, M. Tosi)and laboratory (P.Béarez, S.Kerneur, C.Martin,C. Pierre, M. Tengberg, Ughetto J. and the
whole team of the LMC14). This article is dedi-cated to the memory of S. Cleuziou and J.-F.Saliège.
Figure 3 – Marine reservoir effect (MRE) recorded in mangrove and marine shells, fish and humans graves405, 408 and 411.
GRP : fyssen JOB : annales27Internet DIV : 13mp⊕ZAZZO⊕Antoine p. 15 folio : 139 --- 14/5/013 --- 15H33
PALÉONTOLOGY / ARCHÉOLOGY
139

BIBLIOTHÈQUE
Nous remercions vivement les généreux donateurs avec lesquels nous échangeons nos Annales :
IIAS/International Institute for Asian Studies Newsletter No61 – Autumn 2012
Max Planck Research The Science Magazine No 3.2012
Société des NeurosciencesLa lettre des NeurosciencesNo 42 – Printemps/Eté 2012No 43 – Automne/Hiver 2012
Académie des Sciences La lettre de l’Académie des Sciences No 29-30
François SIGAUT – CNRS Editions « Comment Homo devint faber » – François SIGAUT
F.-Xavier ALARIO – Ed. Odile JACOB « Toutes les questions que vous vous posez sur votrecerveau » F.-Xavier ALARIO
Universidad de Granada Cuadernos de Prehistoria y arqueologiaNo21 –2011
Nous remercions pour leur collaboration les auteurs soutenus dans leurs travaux de recherche par laFondation Fyssen, nos lecteurs pour leur fidélité, et tous nos correspondants et bibliothèques qui permettentla libre consultation de notre revue.
We thank for their collaboration the authors supported in their researchworks by the Fyssen Foundation, ourreaders for their fidelity, and all our correspondents and libraries which allow the free consultation of ourreview.
GRP : fyssen JOB : annales27Internet DIV : 14mp⊕bibliotheque p. 1 folio : 140 --- 14/5/013 --- 15H39
140