Embed Size (px)
Citation preview

(Laboratoire de Microbiologie, Universitd de Louvain, HdverK'-Iez-Louvain, Belgique).
E T U D E D U M E T A B O L I S M E D ' A C I D E S A M I N E S
C H E Z A S P E R G E L L U S O R Y Z A E
IV. INFLUENCE DU CHLORURE DE CALCIUM SUR LE MI~TABOLISME DES ACIDES AMINI~S 1)
par
PAUL SIMONART.et KWANG YIJ CHOW (Regu le 25 Juillet 1953).
I1 a 6t6 montr6 que le calcium active et s tabi l ise lesprot~inases bact~riennes (1, 2, 3, 4) et que cet ~l~ment est n~cessaire all d~velop- pement de bact~ries dans un milieu qui ne cont ient que des pro- t6ines comme seule source d 'azote (5).
Dans le domaine des moisissures CHOW (6) a mis en 6vidence l ' influence favorable du calcium sur le d~veloppement d'Aspergillus oryzae cultiv6 sur hydro lysa t de cas6ine, ainsi que sur l 'acti~dt~ prot~olyt ique de la prot~inase de cet te moisissure. D ' au t r e par t , SAKAGUCHI (7) a observ~ une augmenta t ion de teneur en azote d)Asp. oryzae en presence de carbonate calcique.
Se basan t sur ces observat ions et faisant suite ~ une ~tude des acides amin6s pr6sents ~ l '6 ta t l ibre dans le myc~lium d'Asp, oryzae (8, 9), il a pa ru int~ressant de rr si le calcium exer~ait une influence sur le m6tabol isme des acides amin6s de ce microchampig- non. I1 en r6sulte qu 'en effet une diff6rence sensible de composi t ion d 'acides amines l ibres s 'observe chez cette moisissure suivant qu'el le est cultiv~e en presence o u c n absence de calcium.
I. I~I~,THODES ]~XPERIMENTALES.
I ) La souche de moisissure et le milieu de culture ut i l i s& dans
1) Part I, II, II1: Antonie van Leeuwenhoek 19, 121, 245, 1953; 20, 174, 1954.

l~tude du m6tabolisme d'acides aminds chez Asp..oryzae. IV. 211
cette ~tude sont ceux qui ont servi aux'recherches pr6c~dentes (8, 9): Asp. oryzae w.f. et une solution g 3 % de bacto-casamino acids (Difco). Mais p lu t6 t que de se faire pa r la mdthode sur flux continu de milieu, les cul tures se font ici en er lenmeyers de 100 ml contenant 20 ml de l iquide nut r i t i f que l 'on ensemence au fil de plat ine au moyen d 'une cul ture sur mofit-agar, vieille de trois jours. L 'er len- meyer ensemenc~ est main tenu ~ 25 ~ C., habi tue l lement pendan t cinq jours.
2) Pour l ' analyse des acides amin6s libres, on retire le myc61ium de l 'e r lenmeyer , et apr~s trois lavages k l ' eau on en ex t ra i t les sub- stances solubles pa r 6bulli t ion de deux minutes dans 20 ml d 'eau. On concentre cet ex t ra i t sous" pression r6duite ~ :k 45 ~ C. et le sou- met ~t un examen chromatographique selon les m6thodes appl i - qu6es pour les 6tudes pr6c6dentes. Cependant , dans cette recherche- ci, il est ut i le de d6barrasser ]a solution ~ chromatographie r des sels ainsi que des substances acides et neutres qu'el le peut contenir. C'est pourquoi il est fai t usage de la facon suivante d 'une colonne de 20 • 1 cm de r4sine ionique "Zeo- Karb" . On lave la colonne avec ! 00 ml de NaOH 2 N, puis b, l ' eau jusqu'~t neut ra l i t6 de l'61uat, ensuite ave*: 100 ml d'HC1 2 N, enfin ~t l 'eau jusqu'~t ce que l 'd lua t a i t un p H = 5. On verse alors sur la colonne 0,5 ~t 1 ml de la solut ion /t 6tudier, on la laisse s ' fcouler doucement , puis on 3' a joute deux lois 1 ml d 'eau, enfin on 61ue/t l ' eau et recueille environ 20 gouttes pa r minute jusqu'b~ ce que le p H de l'61uat a t te igne 5. Apr6s cela on 61ue les acides aminfs ~t I 'NH4OH 1,5 N, recuei l lant deux frac- t ions d'41uat, la premi6re de 100 ~t 150 ml et la seconde de 50 ml; on les concentre sous pression r6duite et les analyse chromatographi - quement. On ne soumet cependant la seconde fraction ~ l ' ana lyse que si des acides amin6s basiques sont pr6sents.
II. RI~SULTATS ET DISCUSSION.
1. Influence du CaC12 sur la croissance d'Asp, oryzae.
Asp: oryzae se dfiveloppe tr~s mddiocrement sur 3 % de casamino acids. Mais l ' add i t ion d 'une quant i t~ convenable de calcium favorise tr~s fort le d~veloppement de la rnoisissure ainsi que le montre le tab leau 1. Celui-ci donne le poids du myc~l ium form~ en 5 jours 25 ~ C. sur 20 ml de milieu, en er lenmeyer de I00 ml, et il montre qu 'une concentra t ion M/50 de CaCI 2 donne une pellicule muc6dienne

212 Paul Simonart et Kwang Yii Chow,
qui p~se au delg de six Iois plus que celle qui est form~e en absence de ce sel. D ' au t r e par t , ce tab leau indique aussi le p H de la solut ion de mdtabol isme 5. la fin de la cul ture de cinq iours, et fai t ressor t i r que non seulement le p H le plus bas correspond au d6ve loppement le meilleur, mais que d 'une cul ture k l ' au t re il varie en sens inverse du poids du myc{lium.
TABLEAU [.
Poids du myc41ium sScl% h 105 ~ C. pendant 12 heures
pH de la solution de m6tabolisme
3% bacto-casamino acids; pH = 6.3
0.020
8.2
+M/IO0 CaCL,
0.035 g
7.9
+M/50 CaCI~
0.124 g
6.5
+M/20 CaCl 2
0.i01 g
7.0
-F M/10 CaCI 2
0.084 g
7.1
2. Influence du chlorure de calcium sur la teneur en aoides amin4s de la solution de m6tabolisme.
Le chromatogramme du mil ieu de cul ture tel quel (3 % bac to - casamino acids (Difco)) montre les spots correspondants aux sub- stances suivantes: acide aspar t ique, acide g lu tamique, s~rine, gly- cine, thr6onine, alanine, hist idine, lysine, proline, arginine, ph6nyl- alanine et des acides amin6s du groupe val ine et leucine.
Si l 'on chromatographie le mfime mil ieu de culture addi t iom% de chlorure de calcium, ne fut-ce qu'& la concentrat ion de M/100, on y d~cfile les m~mes acides amin6s qu 'en absence de ce sel, sauf pour les acides aminfis basiques hyst idine, lysfl~e et arginine qu ' i l ne renferme pas.
Ceci s 'expl ique par l ' insolubi l isat ion de ces substances sous forme de sels calciques. En effet, lots de l ' add i t ion de chlorure de calcium se forme un prficipit6 qui fut analys6 comme suit: apr~s f i l t rat ion et lavages r@~t~s 5. l ' eau on le dissout dans H2SO 4 0.5 N; on ~limine par f i l t rat ion le sulfate de calcium; on passe le f i l t rat 5. t r a - vers une colonne de "Zeo K a r b " , et recueille l '~luat ammoniaca l ; la chromatographie de celui-ci montre la pr6sence de lysine h c6t6 de pet i tes quanti t~s d 'arginine et d 'h is t idine.

~tude du n%tabolisme d'acides anain4s chez Asp. oryzae. IV. 213
La composition qualitative en acides amin6s de ces milieux de culture, avec ou sans chlorure de calcium ne se modifie gu~re lors du d6veloppement d'Asp, oryzae pendant cinq jours. La seule dif- f6rence perceptible est que l 'on trouve parfois de l 'ornithine, de la glutamine et de la lysine dans la solution de mdtabolisme calcique.
3. Influence du chlorure de calcium sur la teneur en acides aminfis libres du myc~lium.
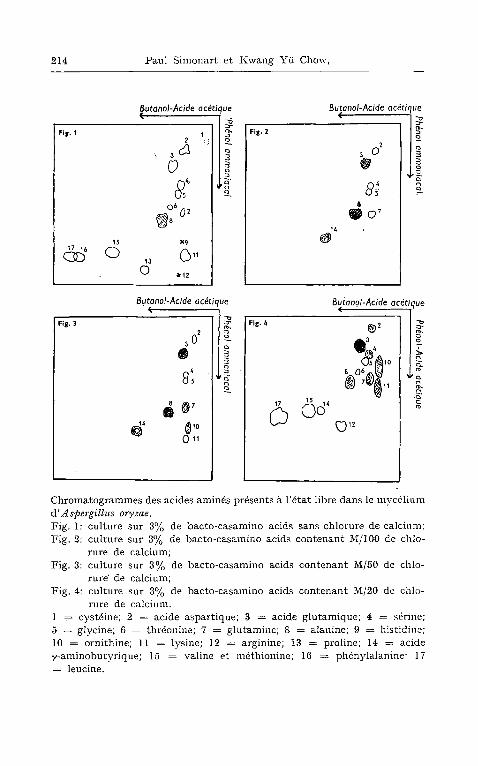
Dans le myc~lium forms sur le milieu exempt de chlorure de cal- cium oll retrouve chromatographiquement (fig. 1) toutes les sub- stances aminfies du bacto-casamino acids, ~ c6td de traces de glu- tamine et de cystfiine ou de cystine.
Lorsque la moisissure se dfiveloppe sur 3 % de bacto-casamino acids contenant du chlorure de calcium 5~ la concentration M/100 on ne d6c~le chromatographiquement (fig. 2) dans son myc~lium que l 'acide aspartique, l 'acide glutamique, la sfirine, la glycine et l 'ala- nine, en tant que constituants initialement prfisents dans le milieu de culture. Mais en plus de ces cinq acides amin6s oll trouve encore en quantit~ apprficiable l 'acide 7-aminobutyrique et la glutamine.
L 'absence de proline, de valine et de leucine peut s 'expliquer soit par une inhibition d 'absorpt ion de ces acides par ]e calcium, soit pa r leur d6composition plus rapide en pr6sence de cet ~l~ment. Les seules donn6es qualitatives dont on dispose ici ne permet tent pas de dire 5. quel ph6nom~ne on a affaire. Des ddterminations quanti- ta t ives trancheraient la question.
Quant on cultive la moisissure sur un milieu plus riche en calcium telles des concentrations en chlorure de calcium de M/50, de M/20 et de lVI/10, la teneur en substances amin6es se distingue de celle qui vient d 'etre vue pour la moisissure cultivfie avcc moins de chlorure de calcium. En effet (fig. 3 et 4) on y ddc~le de l 'ornithine, de la lysine et des traces d'arginine qu 'on ne trouve pas en absence de calcium ni h la concentration M/100 de chlorure de calcium. En outre, ~ la concentration M/20 on ne trouve plus que des traces d 'acide 7-aminobutyrique, tandis qu'h la concentration M/10 cet acide 'ne peut plus fitre mis en fividence. Enfin, il semble que la plus forte concentration en ornithine corresponde k la concentration M/20.
Ces 1-~sultats appellent quelques observations suppl~mentaires en ce qui concerne la formation d'acide 7-aminobutyrique et celle d'acides aminfis basiques.
2 1 4 P a u l S i m o n a r t e t K w a n g Yt i C h o w ,
FI~. I
ButanoI-Acide ac~tique C IY 2
15 X9
�9 . (5 ~1
0 ~12
ButanoI-Acide ac~tique
3 02 ! 6
8',
Fig, 3
1~' (~10
011
Fig, 2
Fig. 4
17 �9
ButanoI-Acide ac~tique
' li 8'
&
qlPO"
ButanoI-Acide ac~tique , (
1o
6d' ~)12
C h r o m a t o g r a m m e s d e s a c i d e s a m m 6 s p r 6 s e n t s ~ l ' 6 t a t f ib re d a n s le m v c 6 1 i u m
d' A spergillus oryzae. F i g . 1: c u l t u r e s u r 3 % d e b a c t o - c a s a m i n o a c i d s s a n s c h l o r u r e d e c a l c i u m ;
F i g . 2: c u l t u r e s u r 3 % d e b a c t o - c a s a m i n o a c i d s c o n t e n a n t ~ [ / 1 0 0 d e c h l o -
r u r e d e c a l c i u m ; F i g . 3: c u l t u r e s u r 3 % de b a c t o - c a s a m i n o a c i d s c o n t e n a n t M / 5 0 d e c h l o -
rure" d e c a l c i u m ; F i g . 4: c u l t u r e s u r 3 % d e b a c t o - c a s a m i n o a c i d s e o n t e n a n t M / 2 0 d e c h l o -
r u r e de c a l c i u m . 1 .= c y s t 6 i n e ; 2 = a c i d e a s p a r t i q u e ; 3 = a c i d e g l u t a m i q u e ; 4 = s~r ine ; 5 = g l y c i n e ; 6 = t h r ~ o n i n e ; 7 = g l u t a m i n e ; 8 = a l a n i n e ; 9 ~ h i s t i d i u e ;
10 = o r n i t h i n e ; 11 = ly s ine ; 12 = a r g i n i n e ; 13 = p r o l i n e ; 14 = a c i d e
~ , - a m i n o b u t y r i q u e ; 15 = v a l i n e e t m 6 t h i o n i n e ; 16 = p h 6 n y l a l a n i n e ; 17
= l e u c i n e .

]~tude du m6"~abolisme d'acides amin6s chez Asp. oryzae. IV. 215
Pour ce qui est de l 'acide y-aminobutyrique, il est int6ressant de noter que la pr6sence de cet acide dans le myc61ium s'observe d 'une part, lorsque la moisissure est cultiv~e sur milieu contenant M/100 h M/20 de chlorure de calcium et d 'autre part lorsqu'elle est cultiv6e sur "flux cont inu" ~ pH bas (8). L 'ac id i t4 et le calcium semblent donc exercer - - probablement par un m6chanisme diff6rent - - une action analogue sur le m6tabolisme des acides amin6s. Alors que l 'on estimait que la product ion d'acide ) ,-aminobutyrique ne se faisait qu'~ pH bas, l 'on constate ici qu'elle peut avoir lieu ~ pH relative- ment 61ev~ en pr6sence d 'une quantit6 convenable de chlorure de calcium. Des r6sultats analogues ont 4t6 obtenus en pr6sence de carbonate calcique.
Quant aux acides amin6s basiques, ornithine,lysine et arginine que l 'on d4c~le darts le myc61ium d6velopp6 en pr4sence de quantit4s suffisantes de chlorure de calcium, ils ne peuvent provenir que de modifications biochimiques d 'autres acides amin4s, favoris6es par le calcium. En effet, d 'une part, ce milieu de culture ne renferme aucune de ces trois substances en quantitds d6c61ables, d 'autre part, le myc61ium form6 en absence de calcium ne les contient pas da- vantage.
Comme ces acides amin6s basiques trouvent probablement leur origine dans les r6actions coupl4es de d6samination et d 'amination, cela revient ~ dire que, plut6t que de donner lieu ~ la mise en libert4 d 'ammoniaque, la d4samination de tel ou tel acide amin6 produi- rait - - directement ou indirectement - - des acides bi- ou pluri- amin4s. Dans ces conditions le milieu ne deviendra pas aussi alcalin que si l 'on avait affaire ~ des d6saminations simples. E t il est plausible d 'admet t re que le r61e essentiel du calcium - - et peut ~tre aussi dans une certaine mesure celui de l 'acidit6 - - dans le m6tabo- lisme qui est 4tudi6 ici, se ramfine surtout ~ exercer indirectement une action tampon.
Cette hypoth~se se confirme encore par le f a r que l'acide y-ami- nobutyr ique et la glutamine prennent aussi naissance dans le myc6- lium de la moisissure lorsque le milieu contient une quantit6 con- venable de calcium (ou qu'il a un pH bas). Or, d 'une part, la d4carbo- xylation de l'acide glutamique avec formation d'acide y-aminobuty- rique d6sacidifie le milieu, en supprimant un groupement carboxyle et d 'autre part, l 'amidation de l'acide glutamique bloque aussi un groupe carboxyle en m~me temps qu'elle 6vite la mise en libert4 d 'une mol6cule d'ammoniaclue.

216 Paul S i mona r t et K w a n g yi:~ Chow.
On constate done que par formation d'acides anain6s basiques et d'amide ou encore accessoirement par d6carboxylation, le calcium agit k la fa~on d'un tampon. Mais il n'est pas 6tabli si cet 416ment favorise des ph6nom6nes de transamination ou bien s'il inhibe les d4saminations simples. Cette action tampon permet encore d'ex- pliquer l'influence clu calcium dans d'autres phases du m4tabolisme de la cellule. Mais cette hypoth6se n'exclut nfanmoins pas qu'un r61e particulier puisse ~tre jou6 par l'acide )J-aminobutyrique.
R f s u m 6 .
La teneur en acides amin6s libres du myc41ium est diff~rente selon que l'on cultive Aspergilh~s oryzae sur 3 % de bacto-casamino acids tel quel ou sur ce milieu additionn4 de chlorure de calcium.
Pour des concentrations M/100 de CaC12 on y d6c~le chromato- graphiquement en quantit6 appr6ciable l'acide y-aminobutyrique et la glutamine que l'on ne trouve pas en absence de ce sel. Tandis que pour des concentrations M/50 on y met encore en 4vidence l'or- nithine, la lysine et l'arginine que l'on ne d6c5le pas pour des con- centrations plus faibles de chlorure de calcium.
Ces rfisultats sugg~rent que le calcium d6clenche indirectement une action tampon en favorisant d'une part la formation d'acides amin6s basiques et d'autre part la d6carboxylation de l'acide glu- tamique avec production d'acide ),-aminobutyrique.
I1 est k remarquer que cette derni~re r6action, en prfsence de chlo- rure de calcium, se produit ~ pH 61evG alors qu'il est gfin6ralement admis que cette dfic~boxylation ne se fait qu'~ pH bas.
B i b l i o g r a p h i e .
I. BI.:u,~IEn, J. 1941. Ac ta Biol. Belg. 2, 273, 276. 2. GOnINI, L. et FROMAGEOT, C. 1949. Compt . Rend . 229, 559. 3. GORINI, L. et 17Ir C. 1950. Biochim. Biophys . Ac t a 5, 524. 4. GORINI, L. ] 950. Biochim. Biophys . Ac ta 6, 237. 5. GORINI, L. et ANDRAIN, L. 1951. Biochim. Biophys . Ac ta 6, 477. 6. CHow, K. Y. 1951. E n z y m e s d'dspergilhts et b iochimie de la f e rmen-
t a t i on du Soja. Th6se doer., Univ . Louva in . 7. SAKAGUCHI, K. 1950. J. Agr. Chem. Soc. J a p a n 24, 77. 8. SIMONAI~.T, P. et CHow, I":. Y. 1953. An ton ie v a n Leeuwenhoek 19,
121, 245. 9. SJMO.NAR'r, P. et CHow, I,:. Y. 1954. Anton ie van L eeuwenhoek 20, 174.