Embed Size (px)
Citation preview

171
Troisième partie :
EVALUATION DE LA RESSOURCE ET GESTION DE LA
PECHERIE
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�
�

172
�

173
�
INTRODUCTION Nous avons vu dans la section 3.6. de la première partie de la thèse que l’aménagement de la pêche crevettière à Madagascar est basé essentiellement sur la régulation de l’accès à la pêcherie (zones de pêche et nombre d’unités) par la mise en place d’un système de licences. Les pressions de la part de l’extérieur pour entrer dans la pêcherie sont considérables et la question de la détermination du volume des licences est souvent évoquée. L’évaluation de la ressource peut apporter des éléments de réponse à cette question. Les évaluations passées à l’aide des modèles globaux ont été menées par zone de pêche (zone I à X) laissant penser que celles-ci pouvaient être indépendantes. Par contre celles utilisant les approches analytiques considèrent un stock par espèce (stock de F. indicus, stock de M. monoceros et stock de P. semisulcatus) scindée en deux fractions ; mâles et femelles. Cette approche est surtout liée à l’absence de données suffisamment précises, permettant de ventiler les apports et les structures démographiques par secteur géographique. La deuxième partie de cette thèse avait pour objet d’analyser la variabilité de certains paramètres biologiques nécessaires à l’évaluation des ressources et à la définition des conditions optimales d’exploitation. Cette troisième partie a pour objectif d’effectuer un diagnostic de l’état des stocks de crevettes à partir des méthodes classiques de la dynamique des populations. Plusieurs modèles et jeux de paramètres sont utilisés, afin de comparer les diagnostics obtenus par les différentes méthodes et d’analyser la sensibilité des modèles retenus à l’échelle globale et à l’échelle des grandes zones. Dans un premier temps, une analyse globale est menée à partir des séries de données de capture et effort. Après, nous procédons à une analyse structurale sur une année moyenne où les évaluations par espèce à l’échelle globale et à l’échelle des régions géographiques sont examinées.
8. DIAGNOSTIC PAR AJUSTEMENT DU MODELE GLOBAL
Les stocks de crevettes malgaches sont exploités depuis 1967 et on dispose des séries temporelles de données sur les captures (toutes espèces confondues) et les efforts de la pêche industrielle. Trois séries de données sont prises en compte. Ces séries de données sont de structures et de niveaux d’agrégations différents. Dans un premier temps, nous avons traité séparément ces données et par la suite estimé un indice d’abondance synthétique sur la période 1967 à 2002. On analyse ainsi la réponse de différents stocks à la pression de pêche depuis le début de l’exploitation de la ressource jusqu’à la situation actuelle. Les précautions sur l’utilisation des résultats obtenus comme outil de gestion sont discutées à la fin de ce chapitre.
174
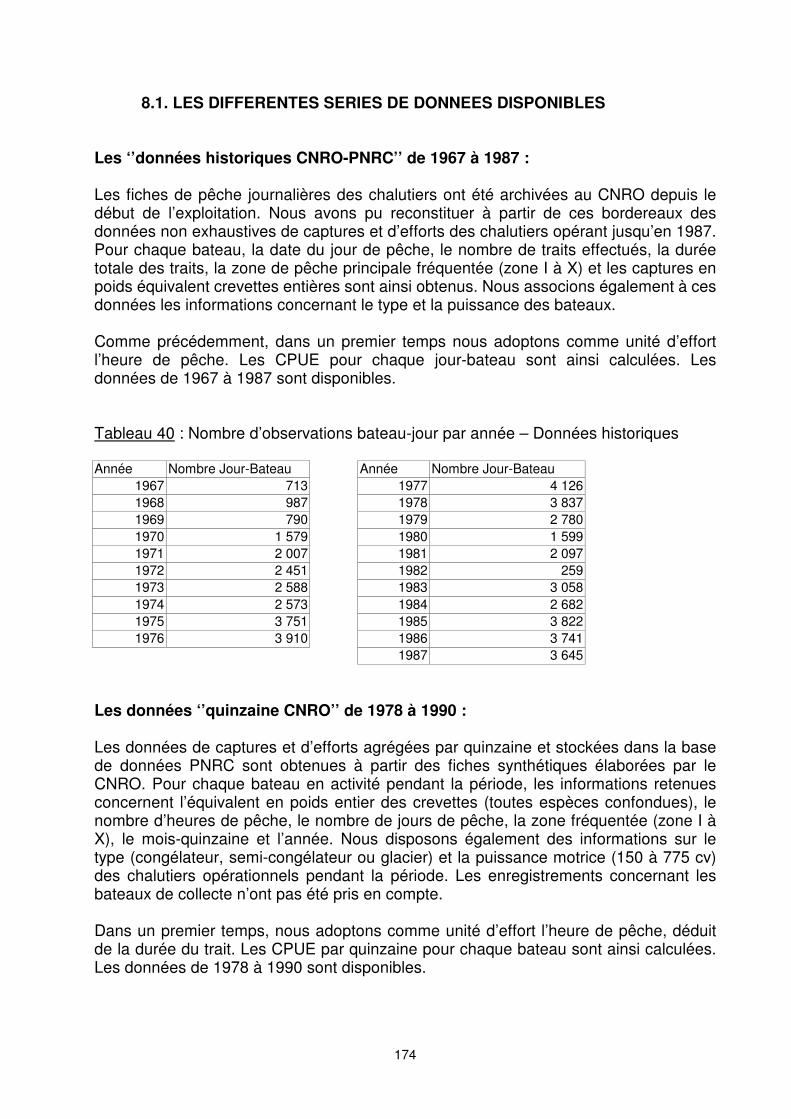
8.1. LES DIFFERENTES SERIES DE DONNEES DISPONIBLES Les ‘’données historiques CNRO-PNRC’’ de 1967 à 1987 : Les fiches de pêche journalières des chalutiers ont été archivées au CNRO depuis le début de l’exploitation. Nous avons pu reconstituer à partir de ces bordereaux des données non exhaustives de captures et d’efforts des chalutiers opérant jusqu’en 1987. Pour chaque bateau, la date du jour de pêche, le nombre de traits effectués, la durée totale des traits, la zone de pêche principale fréquentée (zone I à X) et les captures en poids équivalent crevettes entières sont ainsi obtenus. Nous associons également à ces données les informations concernant le type et la puissance des bateaux. Comme précédemment, dans un premier temps nous adoptons comme unité d’effort l’heure de pêche. Les CPUE pour chaque jour-bateau sont ainsi calculées. Les données de 1967 à 1987 sont disponibles. Tableau 40 : Nombre d’observations bateau-jour par année – Données historiques Année Nombre Jour-Bateau Année Nombre Jour-Bateau
1967 713 1977 4 126 1968 987 1978 3 837 1969 790 1979 2 780 1970 1 579 1980 1 599 1971 2 007 1981 2 097 1972 2 451 1982 259 1973 2 588 1983 3 058 1974 2 573 1984 2 682 1975 3 751 1985 3 822 1976 3 910 1986 3 741
1987 3 645 Les données ‘’quinzaine CNRO’’ de 1978 à 1990 : Les données de captures et d’efforts agrégées par quinzaine et stockées dans la base de données PNRC sont obtenues à partir des fiches synthétiques élaborées par le CNRO. Pour chaque bateau en activité pendant la période, les informations retenues concernent l’équivalent en poids entier des crevettes (toutes espèces confondues), le nombre d’heures de pêche, le nombre de jours de pêche, la zone fréquentée (zone I à X), le mois-quinzaine et l’année. Nous disposons également des informations sur le type (congélateur, semi-congélateur ou glacier) et la puissance motrice (150 à 775 cv) des chalutiers opérationnels pendant la période. Les enregistrements concernant les bateaux de collecte n’ont pas été pris en compte. Dans un premier temps, nous adoptons comme unité d’effort l’heure de pêche, déduit de la durée du trait. Les CPUE par quinzaine pour chaque bateau sont ainsi calculées. Les données de 1978 à 1990 sont disponibles.
175
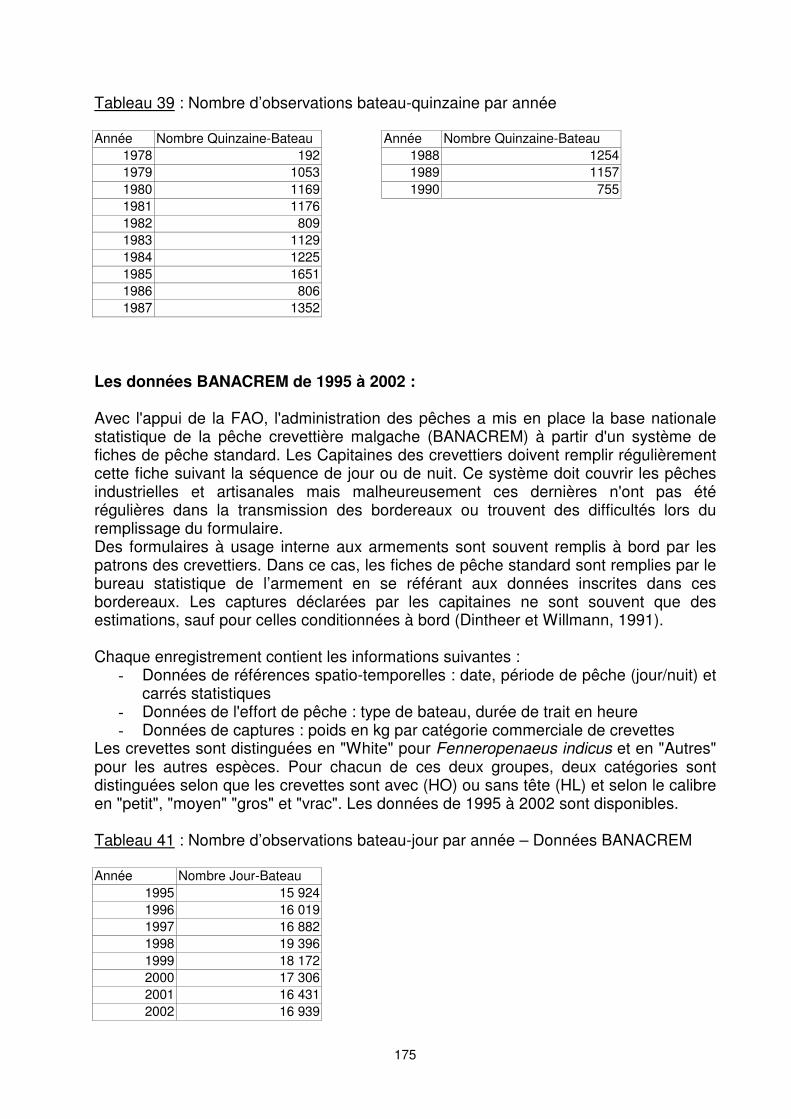
Tableau 39 : Nombre d’observations bateau-quinzaine par année Année Nombre Quinzaine-Bateau Année Nombre Quinzaine-Bateau
1978 192 1988 1254 1979 1053 1989 1157 1980 1169 1990 755 1981 1176 1982 809 1983 1129 1984 1225 1985 1651 1986 806 1987 1352
Les données BANACREM de 1995 à 2002 : Avec l'appui de la FAO, l'administration des pêches a mis en place la base nationale statistique de la pêche crevettière malgache (BANACREM) à partir d'un système de fiches de pêche standard. Les Capitaines des crevettiers doivent remplir régulièrement cette fiche suivant la séquence de jour ou de nuit. Ce système doit couvrir les pêches industrielles et artisanales mais malheureusement ces dernières n'ont pas été régulières dans la transmission des bordereaux ou trouvent des difficultés lors du remplissage du formulaire. Des formulaires à usage interne aux armements sont souvent remplis à bord par les patrons des crevettiers. Dans ce cas, les fiches de pêche standard sont remplies par le bureau statistique de l’armement en se référant aux données inscrites dans ces bordereaux. Les captures déclarées par les capitaines ne sont souvent que des estimations, sauf pour celles conditionnées à bord (Dintheer et Willmann, 1991). Chaque enregistrement contient les informations suivantes : �� Données de références spatio-temporelles : date, période de pêche (jour/nuit) et
carrés statistiques �� Données de l'effort de pêche : type de bateau, durée de trait en heure �� Données de captures : poids en kg par catégorie commerciale de crevettes
Les crevettes sont distinguées en "White" pour Fenneropenaeus indicus et en "Autres" pour les autres espèces. Pour chacun de ces deux groupes, deux catégories sont distinguées selon que les crevettes sont avec (HO) ou sans tête (HL) et selon le calibre en "petit", "moyen" "gros" et "vrac". Les données de 1995 à 2002 sont disponibles. Tableau 41 : Nombre d’observations bateau-jour par année – Données BANACREM Année Nombre Jour-Bateau
1995 15 924 1996 16 019 1997 16 882 1998 19 396 1999 18 172 2000 17 306 2001 16 431 2002 16 939

176
8.2. METHODOLOGIE
8.2.1. L’estimation de l’abondance et des efforts de pêche théorique L’abondance des crevettes est estimée ici à partir de la capture par unité d’effort des données de la pêche commerciale industrielle. Ces CPUE ‘’nominales’’ obtenues sur l’ensemble de la flottille ne sont un bon indice d’abondance que sous l’hypothèse de puissance de pêche constante d’une part et sous l’hypothèse de stock réparti sur une aire de surface constante. Ainsi, on peut formuler l’équation suivante (Gascuel, 1995) :
CPUEn = C / fn = q . B = d . Pg . B où CPUEn : Prise par unité d’effort nominale C : Captures fn : Effort de pêche nominal q : Capturabilité B : Biomasse du stock exploitable d : Disponibilité Pg : Puissance de pêche globale
Laurec (1989) et Gascuel et al. (1997) ont mis en évidence l’intérêt de la méthode d’estimation d’indice d’abondance par modélisation linéaire et analyse de variance. La méthode de Robson (1966) est utilisée pour estimer l’indice d’abondance :
CPUEn = Effet (année) . Effet (flottille) . effet (mois*zone) où Effet (année) : Effet statistique ‘’année’’ interprété comme un indice
d’abondance annuelle Effet(flottille) : Effet statistique ‘’navire’’ interprété comme une puissance
de pêche Effet(mois*zone) : Effet statistique avec interaction ‘’zone de pêche’’ et
‘’saison de pêche’’ interprété comme étant la disponibilité par zone et par mois
Deux facteurs sont utilisés pour caractériser la puissance de pêche : type et puissance motrice du bateau. Le facteur type de navire comprend 3 modalités : chalutier congélateur, chalutier semi-congélateur et chalutier glacier (cf. descriptions § 3.2.1). Les puissances motrices des navires allant de 150 cv à 775 cv sont classées en quatre catégories : �� Classe 150 cv �� Classe 300 cv (contenant 260, 270, 280, 287 et 300 cv) �� Classe 400 cv (contenant 370, 395, 400 et 450 cv) �� Classe 500 cv (contenant 500 et 775 cv)
Les modalités du facteur zone de pêche sont définies par sa situation géographique d’une part et au type de licence d’autre part. Nous distinguons au sein de la zone nord-ouest deux sous-zones, l’une correspondant aux licences de type B et l’autre correspondant aux licences de type C. Ainsi, la zone de pêche est classée en quatre modalités : �� Zone nord ; correspond à la zone où les licences de type A sont autorisées à
pêcher �� Zone nord-ouest correspondant aux licences de type B �� Zone nord-ouest correspondant aux licences de type C �� Zone sud (licence de type C)

177
L’analyse de variance à l’aide de la procédure des modèles linéaires généraux (SAS, PROC GLM) est appliquée aux données des CPUE. La nature multiplicative du modèle et l’examen préalable des résidus du modèle testé nous oriente vers le choix d’une transformation logarithmique de la valeur de la CPUE. Les abondances moyennes par année sont calculées à partir de la fonction LSMEANS de la procédure en appliquant la transformation de Laurent :
IA = exp (LSMEANS + σ 2 / 2 ) -1 De cet indice d’abondance, l’effort théorique correspondant est :
f0 = Y / IA où Y représente la capture totale de la pêche industrielle correspondante
L’effort est exprimé par la suite en multiplicateur de l’effort de la dernière année.
8.2.2. L’ajustement du modèle global et sensibilité à l’évolution des puissances de pêche
Jusqu’à maintenant nous avons assimilé la puissance de pêche globale (Pg), d’une part à la capacité des navires à fréquenter les zones de pêche favorables (effet spatial induit par la puissance motrice du navire), et d’autre part à la capacité de pêche des navires liée à leur taille et leur licence (effet temporel induit par le type de navire, dont dépend notamment la durée de marée). Cette démarche ne prend pas en compte les facteurs technologiques tels que la modification du gréement (changement pêche arrière en pêche à tangon) et l’évolution de la configuration des chaluts (un grand chalut, twin et double-twin), ainsi que l’équipement des navires, notamment l’évolution des appareils de détection (sondeurs) pour le repérage des bancs de crevettes. Par ailleurs, les facteurs sur la capacité de l’équipage à utiliser les moyens dont il dispose ont également une influence sur la mortalité par pêche. Cette capacité s’accroît souvent au cours du temps (Gascuel, 1993). Donc ces facteurs doivent être pris en compte dans les modèles d’estimation de l’abondance. Malheureusement, nous ne pouvons pas estimer ces facteurs qui sont susceptibles d’influencer sur les captures et sur l’effort. Pour tenir compte de cette dérive des puissances de pêche globale, nous étudierons également les résultats issus de l’effort théorique estimé sous l’hypothèse d’un taux d’accroissement des puissances de pêche globales variant entre 0 et 5% par an.
Pgi = Pgi+1 / (1 + α) α : Taux d’accroissement moyen annuel de la puissance de pêche
avec Pgi = 1 pour la dernière année et ainsi : f’0 = Y / IA . Pgi f’0 : effort théorique corrigé
Considérant que les crevettes passent globalement une année dans la phase exploitée, le calcul des efforts à l’équilibre n’est pas nécessaire. Le modèle généralisé de Pella et Tomlinson (1969) est testé :

178
Yi / f0i = (a . fi + b)1/(m - 1) avec Yi : Captures totales de l’année I f0i : effort théorique de l’année i
a, b et m sont des paramètres du modèle Comme les valeurs des m ajustées est proche de 1, nous avons utilisé par la suite le modèle exponentiel de Fox (1970) :
Yi / fi = exp(a . fi + b) Les données antérieures à l’année 1971 ne sont pas prises en compte dans l’ajustement des modèles, compte tenu de leur faible fiabilité. L’ajustement des modèles est faite par la méthode des moindres carrés en utilisant le SOLVEUR sous EXCEL. Les valeurs maximales de Yi et les niveaux d’efforts correspondant sont calculés par les formules suivantes :
MSY = -1/a . exp(b - 1) fMSY = - 1/a
8.3. RESULTATS
8.3.1. L’ajustement des modèles d’analyse de variance des rendements horaires
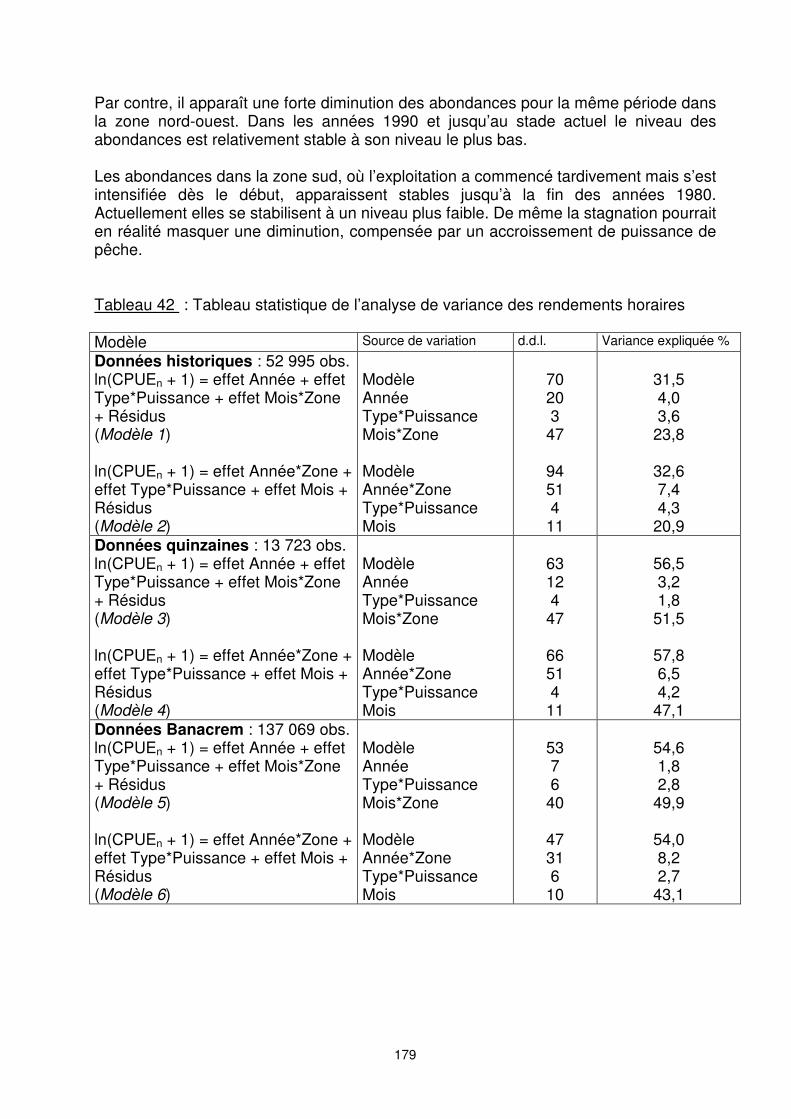
Les modèles de CPUE retenus sont tous hautement significatifs (tableau 42). Les variances expliquées par les modèles à partir des données historiques sont relativement faibles, mais les observations utilisées sont assez nombreuses (52 995 traits). Elles sont de 31,5% pour le modèle 1 et de 32,6% pour le modèle 2. Pour les autres modèles, les variances expliquées sont toutes supérieures à 50% de la variabilité totale. Les modèles 1, 3 et 5 sont utilisés pour estimer les indices d’abondance globale (toutes zones confondues). Les modèles 2, 4 et 6, avec effet d’interaction an*zone sont utilisés pour estimer les indices d’abondances par zone.
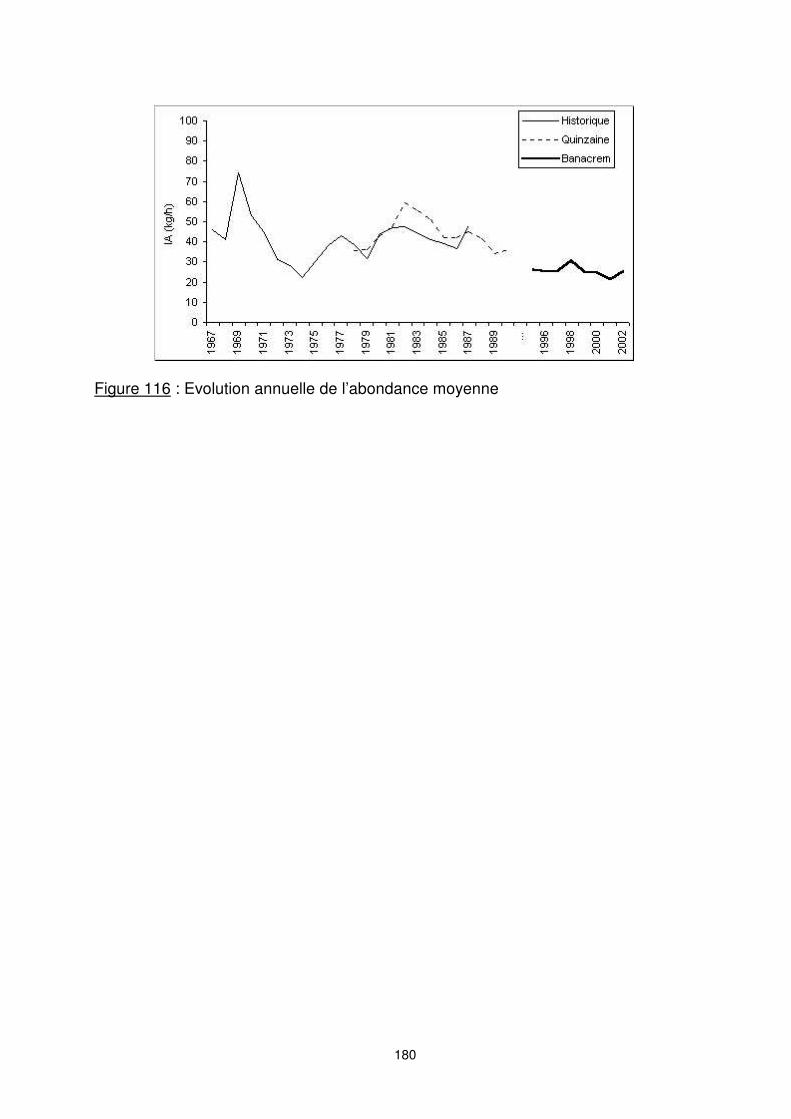
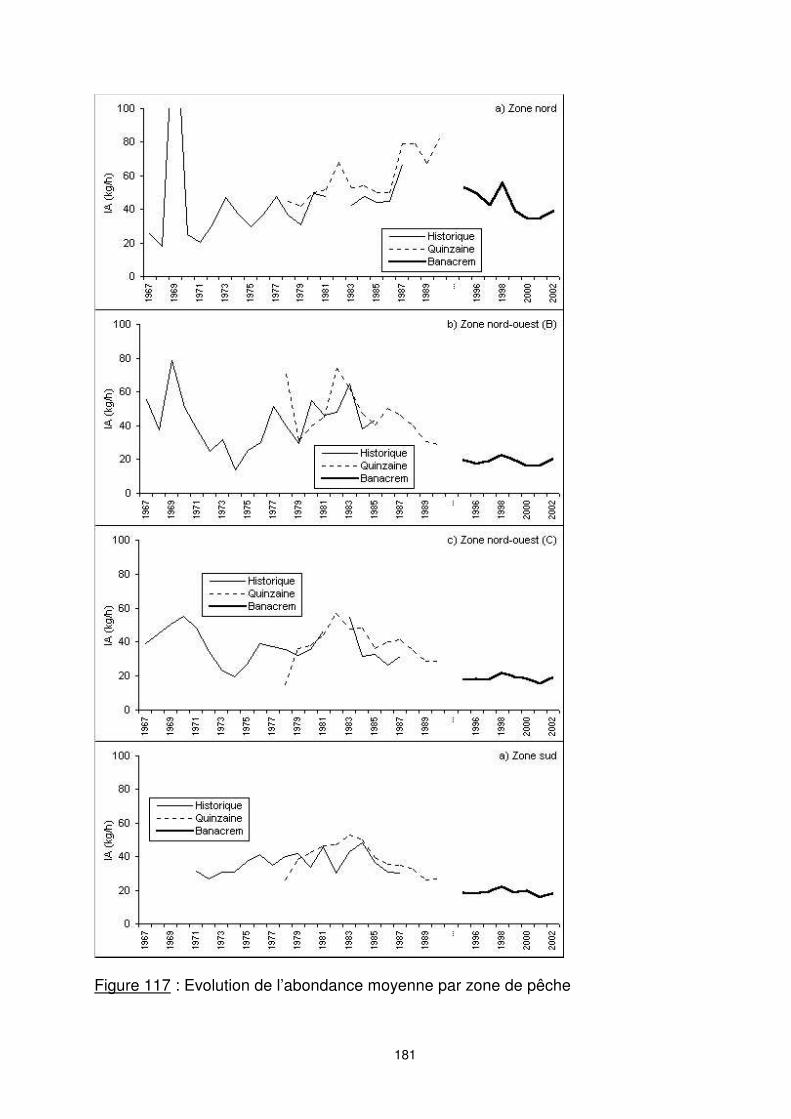
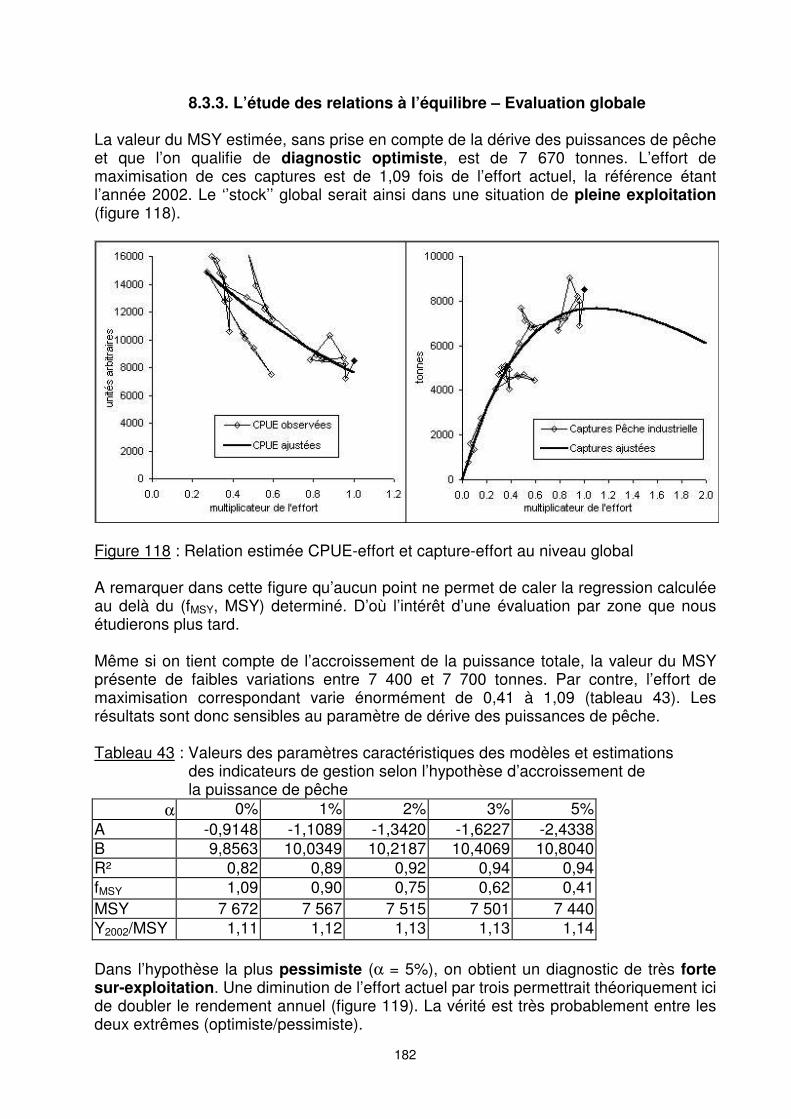
8.3.2. L’évolution annuelle d’abondance L’évolution de l’indice d’abondance (figure 116) montre une forte diminution globale de la biomasse sur la période considérée. Les CPUE nominales passent ainsi de plus de 40 kg/h et jusqu’à 75 kg/h dans les années soixante à moins de 30 kg/h dans les années quatre vingt dix. L’examen de l’évolution de l’indice d’abondance par zone (figure 117) permet de mettre en évidence l’importance et l’évolution tendancielle de l’abondance de.chaque ‘’stock’’. La pêche crevettière malgache a commencé depuis l’année 1967 dans les zones nord. A l’exception des quatre premières années, les indices d’abondances apparaissent croissants jusqu’au début des années 1980. Jusqu’au début des années 1990, les abondances dans la zone nord semblent encore augmenter. Cette augmentation apparente peut cependant être liée à l’accroissement des puissances de pêche, et ne pas traduire un accroissement de la biomasse réelle.
179
Par contre, il apparaît une forte diminution des abondances pour la même période dans la zone nord-ouest. Dans les années 1990 et jusqu’au stade actuel le niveau des abondances est relativement stable à son niveau le plus bas. Les abondances dans la zone sud, où l’exploitation a commencé tardivement mais s’est intensifiée dès le début, apparaissent stables jusqu’à la fin des années 1980. Actuellement elles se stabilisent à un niveau plus faible. De même la stagnation pourrait en réalité masquer une diminution, compensée par un accroissement de puissance de pêche. Tableau 42 : Tableau statistique de l’analyse de variance des rendements horaires Modèle Source de variation d.d.l. Variance expliquée %
Données historiques : 52 995 obs. ln(CPUEn + 1) = effet Année + effet Type*Puissance + effet Mois*Zone + Résidus (Modèle 1) ln(CPUEn + 1) = effet Année*Zone + effet Type*Puissance + effet Mois + Résidus (Modèle 2)
Modèle Année Type*Puissance Mois*Zone Modèle Année*Zone Type*Puissance Mois
70 20 3
47
94 51 4
11
31,5 4,0 3,6
23,8
32,6 7,4 4,3
20,9 Données quinzaines : 13 723 obs. ln(CPUEn + 1) = effet Année + effet Type*Puissance + effet Mois*Zone + Résidus (Modèle 3) ln(CPUEn + 1) = effet Année*Zone + effet Type*Puissance + effet Mois + Résidus (Modèle 4)
Modèle Année Type*Puissance Mois*Zone Modèle Année*Zone Type*Puissance Mois
63 12 4
47
66 51 4
11
56,5 3,2 1,8
51,5
57,8 6,5 4,2
47,1 Données Banacrem : 137 069 obs. ln(CPUEn + 1) = effet Année + effet Type*Puissance + effet Mois*Zone + Résidus (Modèle 5) ln(CPUEn + 1) = effet Année*Zone + effet Type*Puissance + effet Mois + Résidus (Modèle 6)
Modèle Année Type*Puissance Mois*Zone Modèle Année*Zone Type*Puissance Mois
53 7 6
40
47 31 6
10
54,6 1,8 2,8
49,9
54,0 8,2 2,7
43,1
180
Figure 116 : Evolution annuelle de l’abondance moyenne
181
Figure 117 : Evolution de l’abondance moyenne par zone de pêche
182
8.3.3. L’étude des relations à l’équilibre – Evaluation globale La valeur du MSY estimée, sans prise en compte de la dérive des puissances de pêche et que l’on qualifie de diagnostic optimiste, est de 7 670 tonnes. L’effort de maximisation de ces captures est de 1,09 fois de l’effort actuel, la référence étant l’année 2002. Le ‘’stock’’ global serait ainsi dans une situation de pleine exploitation (figure 118).
Figure 118 : Relation estimée CPUE-effort et capture-effort au niveau global A remarquer dans cette figure qu’aucun point ne permet de caler la regression calculée au delà du (fMSY, MSY) determiné. D’où l’intérêt d’une évaluation par zone que nous étudierons plus tard. Même si on tient compte de l’accroissement de la puissance totale, la valeur du MSY présente de faibles variations entre 7 400 et 7 700 tonnes. Par contre, l’effort de maximisation correspondant varie énormément de 0,41 à 1,09 (tableau 43). Les résultats sont donc sensibles au paramètre de dérive des puissances de pêche. Tableau 43 : Valeurs des paramètres caractéristiques des modèles et estimations
des indicateurs de gestion selon l’hypothèse d’accroissement de la puissance de pêche
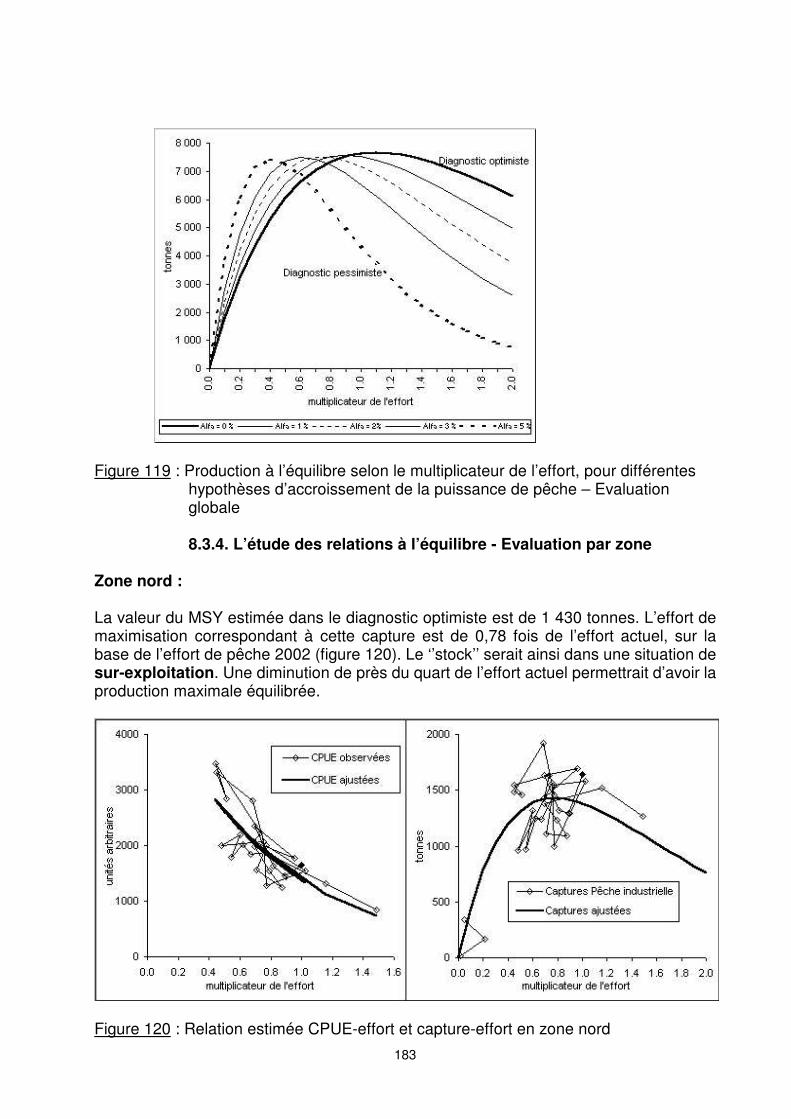
α 0% 1% 2% 3% 5% A -0,9148 -1,1089 -1,3420 -1,6227 -2,4338 B 9,8563 10,0349 10,2187 10,4069 10,8040 R² 0,82 0,89 0,92 0,94 0,94 fMSY 1,09 0,90 0,75 0,62 0,41 MSY 7 672 7 567 7 515 7 501 7 440 Y2002/MSY 1,11 1,12 1,13 1,13 1,14 Dans l’hypothèse la plus pessimiste (α = 5%), on obtient un diagnostic de très forte sur-exploitation. Une diminution de l’effort actuel par trois permettrait théoriquement ici de doubler le rendement annuel (figure 119). La vérité est très probablement entre les deux extrêmes (optimiste/pessimiste).
183
Figure 119 : Production à l’équilibre selon le multiplicateur de l’effort, pour différentes
hypothèses d’accroissement de la puissance de pêche – Evaluation globale
8.3.4. L’étude des relations à l’équilibre - Evaluation par zone
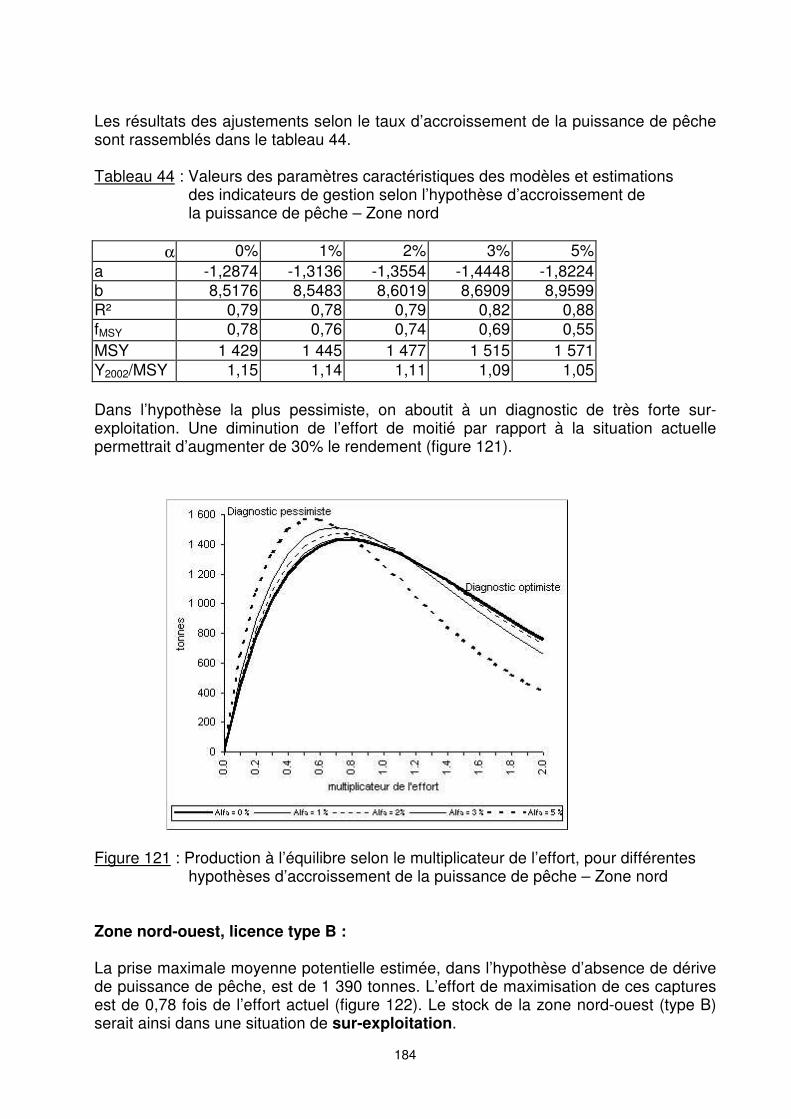
Zone nord : La valeur du MSY estimée dans le diagnostic optimiste est de 1 430 tonnes. L’effort de maximisation correspondant à cette capture est de 0,78 fois de l’effort actuel, sur la base de l’effort de pêche 2002 (figure 120). Le ‘’stock’’ serait ainsi dans une situation de sur-exploitation. Une diminution de près du quart de l’effort actuel permettrait d’avoir la production maximale équilibrée.
Figure 120 : Relation estimée CPUE-effort et capture-effort en zone nord
184
Les résultats des ajustements selon le taux d’accroissement de la puissance de pêche sont rassemblés dans le tableau 44. Tableau 44 : Valeurs des paramètres caractéristiques des modèles et estimations
des indicateurs de gestion selon l’hypothèse d’accroissement de la puissance de pêche – Zone nord
α 0% 1% 2% 3% 5%
a -1,2874 -1,3136 -1,3554 -1,4448 -1,8224 b 8,5176 8,5483 8,6019 8,6909 8,9599 R² 0,79 0,78 0,79 0,82 0,88 fMSY 0,78 0,76 0,74 0,69 0,55 MSY 1 429 1 445 1 477 1 515 1 571 Y2002/MSY 1,15 1,14 1,11 1,09 1,05 Dans l’hypothèse la plus pessimiste, on aboutit à un diagnostic de très forte sur-exploitation. Une diminution de l’effort de moitié par rapport à la situation actuelle permettrait d’augmenter de 30% le rendement (figure 121).
Figure 121 : Production à l’équilibre selon le multiplicateur de l’effort, pour différentes
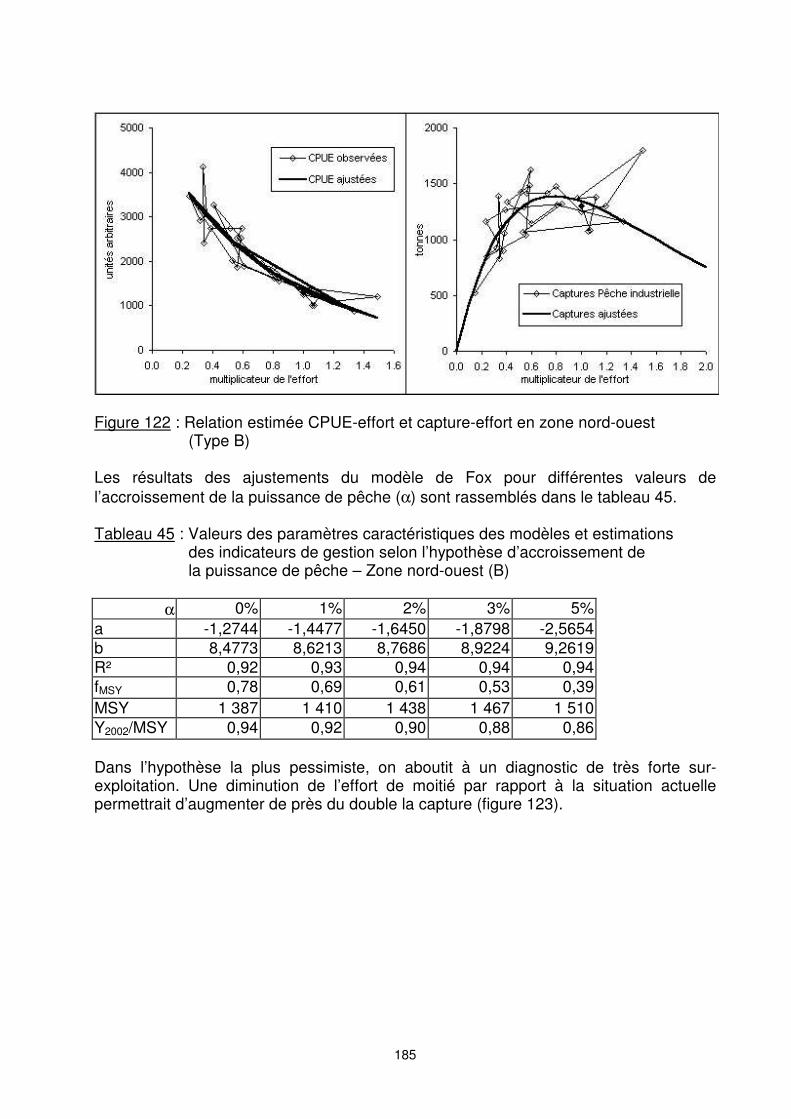
hypothèses d’accroissement de la puissance de pêche – Zone nord Zone nord-ouest, licence type B : La prise maximale moyenne potentielle estimée, dans l’hypothèse d’absence de dérive de puissance de pêche, est de 1 390 tonnes. L’effort de maximisation de ces captures est de 0,78 fois de l’effort actuel (figure 122). Le stock de la zone nord-ouest (type B) serait ainsi dans une situation de sur-exploitation.
185
Figure 122 : Relation estimée CPUE-effort et capture-effort en zone nord-ouest
(Type B) Les résultats des ajustements du modèle de Fox pour différentes valeurs de l’accroissement de la puissance de pêche (α) sont rassemblés dans le tableau 45. Tableau 45 : Valeurs des paramètres caractéristiques des modèles et estimations
des indicateurs de gestion selon l’hypothèse d’accroissement de la puissance de pêche – Zone nord-ouest (B)
α 0% 1% 2% 3% 5%
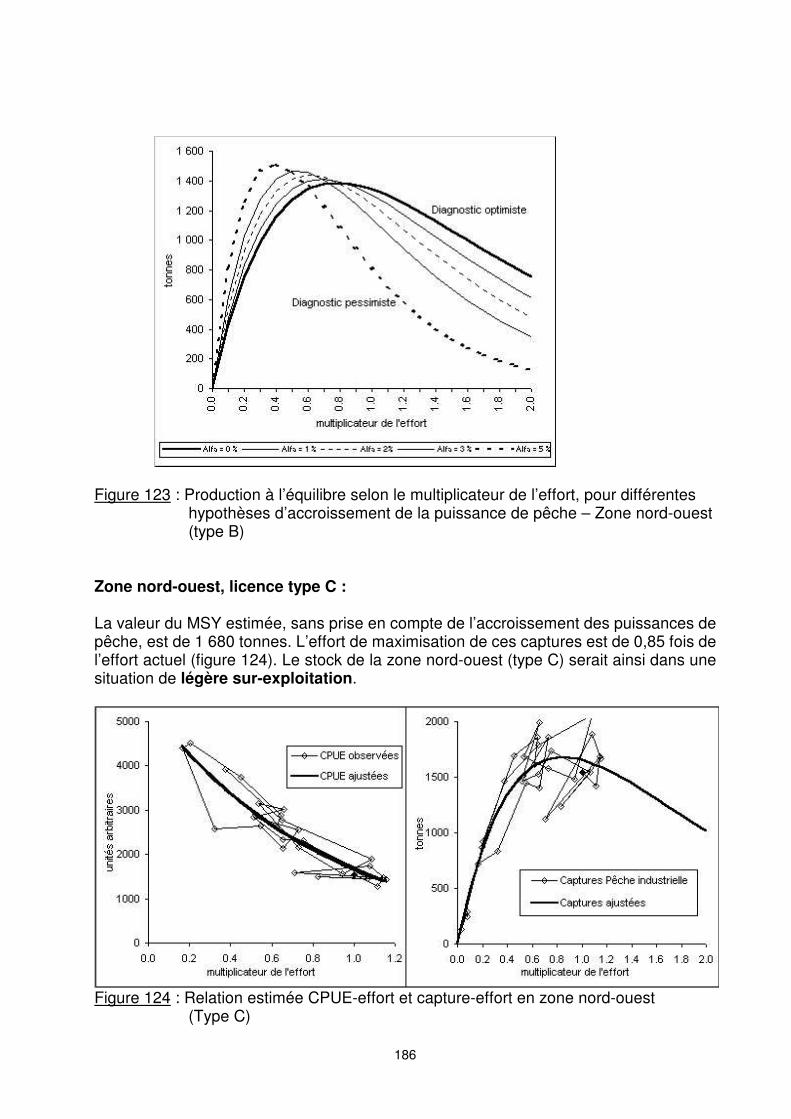
a -1,2744 -1,4477 -1,6450 -1,8798 -2,5654 b 8,4773 8,6213 8,7686 8,9224 9,2619 R² 0,92 0,93 0,94 0,94 0,94 fMSY 0,78 0,69 0,61 0,53 0,39 MSY 1 387 1 410 1 438 1 467 1 510 Y2002/MSY 0,94 0,92 0,90 0,88 0,86 Dans l’hypothèse la plus pessimiste, on aboutit à un diagnostic de très forte sur-exploitation. Une diminution de l’effort de moitié par rapport à la situation actuelle permettrait d’augmenter de près du double la capture (figure 123).
186
Figure 123 : Production à l’équilibre selon le multiplicateur de l’effort, pour différentes
hypothèses d’accroissement de la puissance de pêche – Zone nord-ouest (type B)
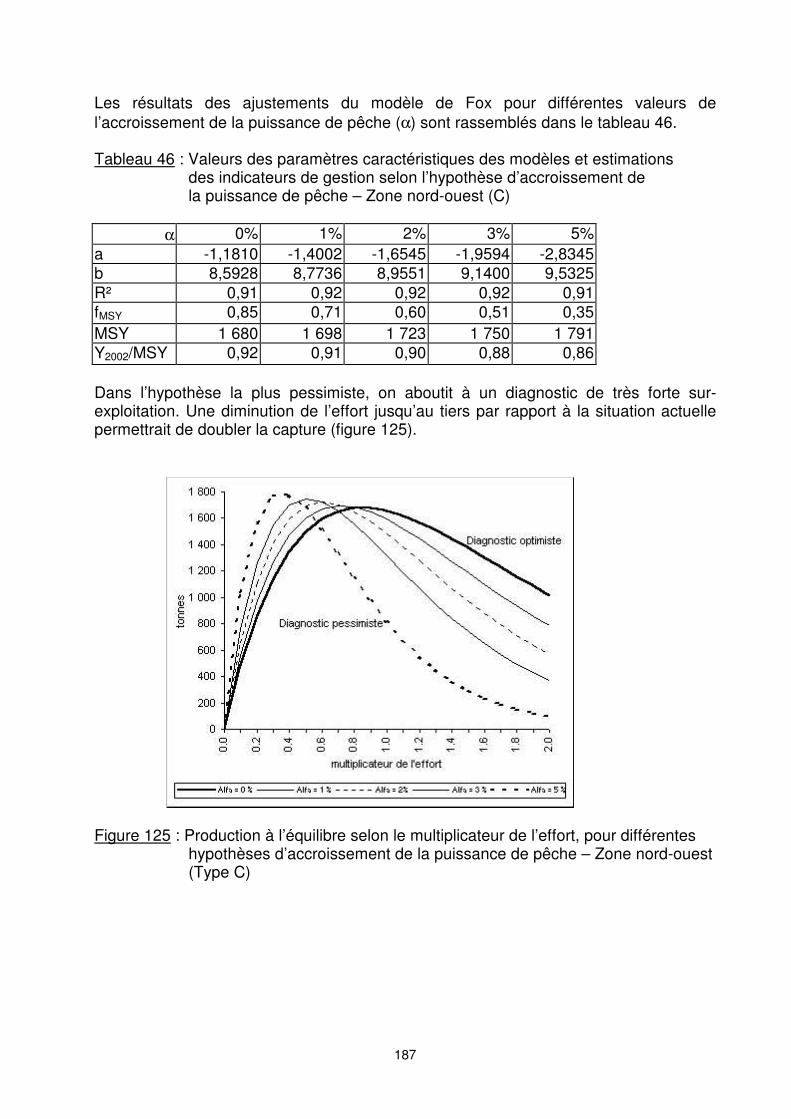
Zone nord-ouest, licence type C : La valeur du MSY estimée, sans prise en compte de l’accroissement des puissances de pêche, est de 1 680 tonnes. L’effort de maximisation de ces captures est de 0,85 fois de l’effort actuel (figure 124). Le stock de la zone nord-ouest (type C) serait ainsi dans une situation de légère sur-exploitation.
Figure 124 : Relation estimée CPUE-effort et capture-effort en zone nord-ouest (Type C)
187
Les résultats des ajustements du modèle de Fox pour différentes valeurs de l’accroissement de la puissance de pêche (α) sont rassemblés dans le tableau 46. Tableau 46 : Valeurs des paramètres caractéristiques des modèles et estimations
des indicateurs de gestion selon l’hypothèse d’accroissement de la puissance de pêche – Zone nord-ouest (C)
α 0% 1% 2% 3% 5%
a -1,1810 -1,4002 -1,6545 -1,9594 -2,8345 b 8,5928 8,7736 8,9551 9,1400 9,5325 R² 0,91 0,92 0,92 0,92 0,91 fMSY 0,85 0,71 0,60 0,51 0,35 MSY 1 680 1 698 1 723 1 750 1 791 Y2002/MSY 0,92 0,91 0,90 0,88 0,86 Dans l’hypothèse la plus pessimiste, on aboutit à un diagnostic de très forte sur-exploitation. Une diminution de l’effort jusqu’au tiers par rapport à la situation actuelle permettrait de doubler la capture (figure 125).
Figure 125 : Production à l’équilibre selon le multiplicateur de l’effort, pour différentes
hypothèses d’accroissement de la puissance de pêche – Zone nord-ouest (Type C)
188
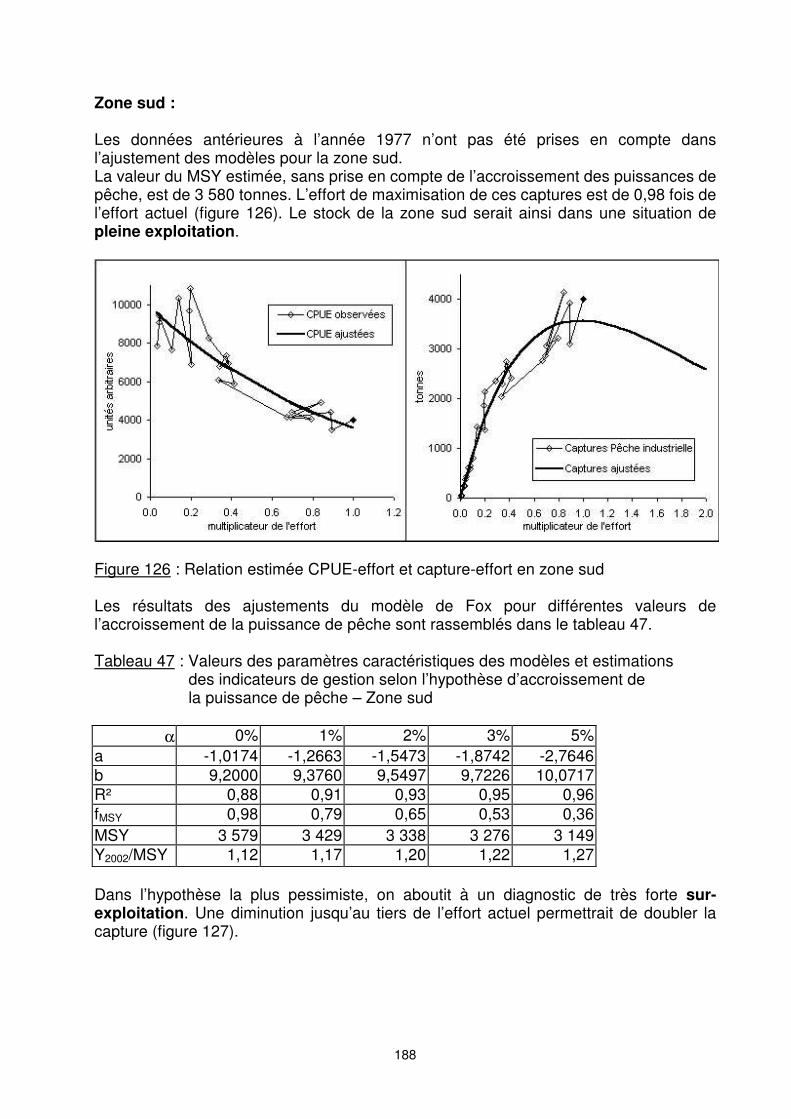
Zone sud : Les données antérieures à l’année 1977 n’ont pas été prises en compte dans l’ajustement des modèles pour la zone sud. La valeur du MSY estimée, sans prise en compte de l’accroissement des puissances de pêche, est de 3 580 tonnes. L’effort de maximisation de ces captures est de 0,98 fois de l’effort actuel (figure 126). Le stock de la zone sud serait ainsi dans une situation de pleine exploitation.
Figure 126 : Relation estimée CPUE-effort et capture-effort en zone sud Les résultats des ajustements du modèle de Fox pour différentes valeurs de l’accroissement de la puissance de pêche sont rassemblés dans le tableau 47. Tableau 47 : Valeurs des paramètres caractéristiques des modèles et estimations
des indicateurs de gestion selon l’hypothèse d’accroissement de la puissance de pêche – Zone sud
α 0% 1% 2% 3% 5%
a -1,0174 -1,2663 -1,5473 -1,8742 -2,7646 b 9,2000 9,3760 9,5497 9,7226 10,0717 R² 0,88 0,91 0,93 0,95 0,96 fMSY 0,98 0,79 0,65 0,53 0,36 MSY 3 579 3 429 3 338 3 276 3 149 Y2002/MSY 1,12 1,17 1,20 1,22 1,27 Dans l’hypothèse la plus pessimiste, on aboutit à un diagnostic de très forte sur-exploitation. Une diminution jusqu’au tiers de l’effort actuel permettrait de doubler la capture (figure 127).
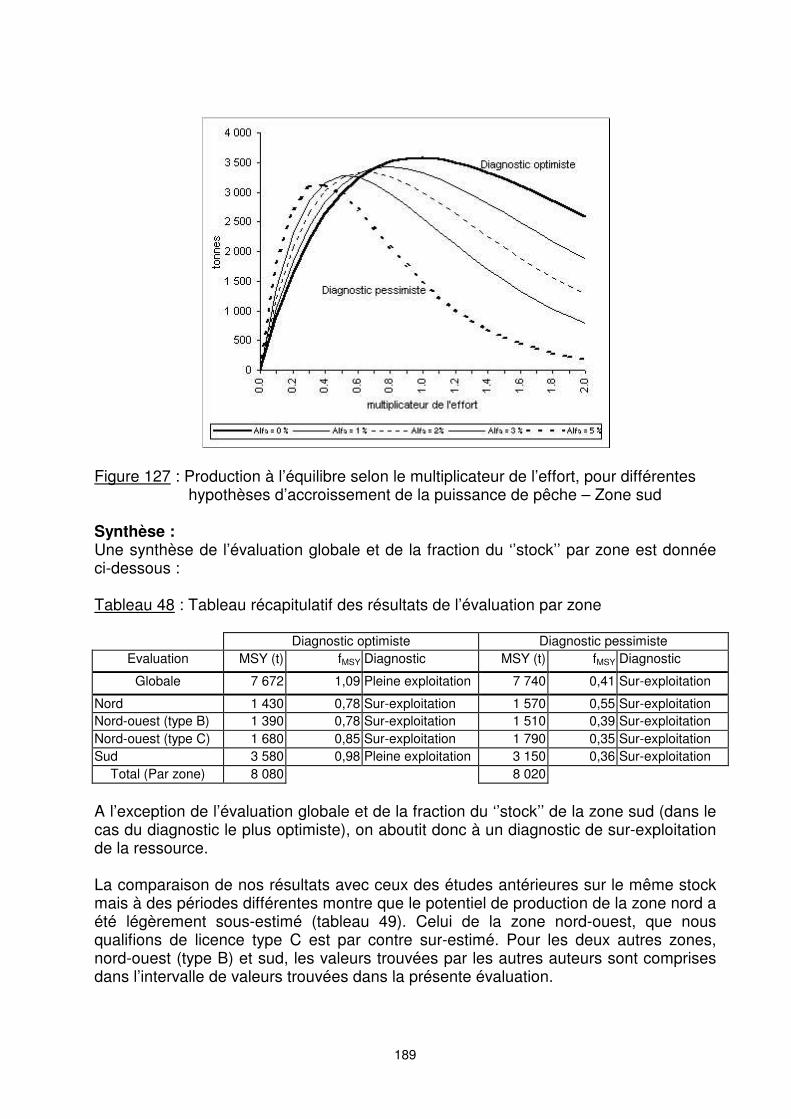
189
Figure 127 : Production à l’équilibre selon le multiplicateur de l’effort, pour différentes
hypothèses d’accroissement de la puissance de pêche – Zone sud Synthèse : Une synthèse de l’évaluation globale et de la fraction du ‘’stock’’ par zone est donnée ci-dessous : Tableau 48 : Tableau récapitulatif des résultats de l’évaluation par zone
Diagnostic optimiste Diagnostic pessimiste Evaluation MSY (t) fMSY Diagnostic MSY (t) fMSY Diagnostic
Globale 7 672 1,09 Pleine exploitation 7 740 0,41 Sur-exploitation
Nord 1 430 0,78 Sur-exploitation 1 570 0,55 Sur-exploitation Nord-ouest (type B) 1 390 0,78 Sur-exploitation 1 510 0,39 Sur-exploitation Nord-ouest (type C) 1 680 0,85 Sur-exploitation 1 790 0,35 Sur-exploitation Sud 3 580 0,98 Pleine exploitation 3 150 0,36 Sur-exploitation
Total (Par zone) 8 080 8 020 A l’exception de l’évaluation globale et de la fraction du ‘’stock’’ de la zone sud (dans le cas du diagnostic le plus optimiste), on aboutit donc à un diagnostic de sur-exploitation de la ressource. La comparaison de nos résultats avec ceux des études antérieures sur le même stock mais à des périodes différentes montre que le potentiel de production de la zone nord a été légèrement sous-estimé (tableau 49). Celui de la zone nord-ouest, que nous qualifions de licence type C est par contre sur-estimé. Pour les deux autres zones, nord-ouest (type B) et sud, les valeurs trouvées par les autres auteurs sont comprises dans l’intervalle de valeurs trouvées dans la présente évaluation.
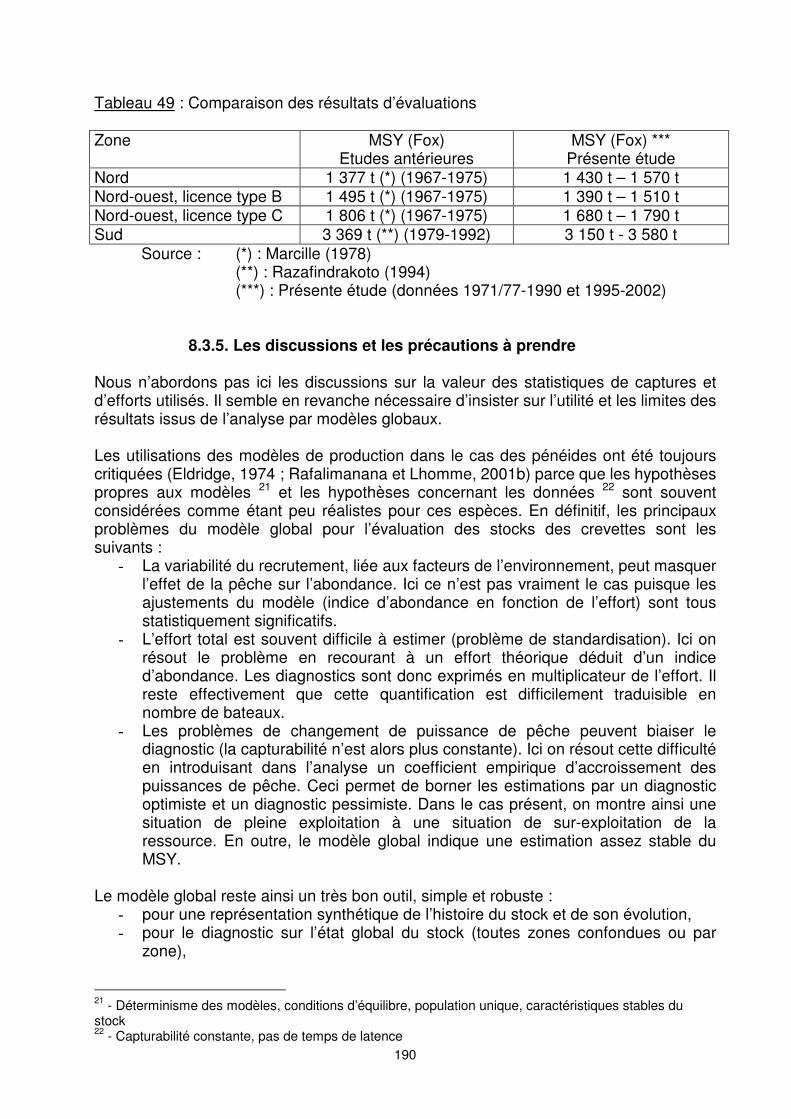
190
Tableau 49 : Comparaison des résultats d’évaluations Zone MSY (Fox)
Etudes antérieures MSY (Fox) ***
Présente étude Nord 1 377 t (*) (1967-1975) 1 430 t – 1 570 t Nord-ouest, licence type B 1 495 t (*) (1967-1975) 1 390 t – 1 510 t Nord-ouest, licence type C 1 806 t (*) (1967-1975) 1 680 t – 1 790 t Sud 3 369 t (**) (1979-1992) 3 150 t - 3 580 t
Source : (*) : Marcille (1978) (**) : Razafindrakoto (1994) (***) : Présente étude (données 1971/77-1990 et 1995-2002)
8.3.5. Les discussions et les précautions à prendre Nous n’abordons pas ici les discussions sur la valeur des statistiques de captures et d’efforts utilisés. Il semble en revanche nécessaire d’insister sur l’utilité et les limites des résultats issus de l’analyse par modèles globaux. Les utilisations des modèles de production dans le cas des pénéides ont été toujours critiquées (Eldridge, 1974 ; Rafalimanana et Lhomme, 2001b) parce que les hypothèses propres aux modèles 21 et les hypothèses concernant les données 22 sont souvent considérées comme étant peu réalistes pour ces espèces. En définitif, les principaux problèmes du modèle global pour l’évaluation des stocks des crevettes sont les suivants : �� La variabilité du recrutement, liée aux facteurs de l’environnement, peut masquer
l’effet de la pêche sur l’abondance. Ici ce n’est pas vraiment le cas puisque les ajustements du modèle (indice d’abondance en fonction de l’effort) sont tous statistiquement significatifs.
�� L’effort total est souvent difficile à estimer (problème de standardisation). Ici on résout le problème en recourant à un effort théorique déduit d’un indice d’abondance. Les diagnostics sont donc exprimés en multiplicateur de l’effort. Il reste effectivement que cette quantification est difficilement traduisible en nombre de bateaux.
�� Les problèmes de changement de puissance de pêche peuvent biaiser le diagnostic (la capturabilité n’est alors plus constante). Ici on résout cette difficulté en introduisant dans l’analyse un coefficient empirique d’accroissement des puissances de pêche. Ceci permet de borner les estimations par un diagnostic optimiste et un diagnostic pessimiste. Dans le cas présent, on montre ainsi une situation de pleine exploitation à une situation de sur-exploitation de la ressource. En outre, le modèle global indique une estimation assez stable du MSY.
Le modèle global reste ainsi un très bon outil, simple et robuste : �� pour une représentation synthétique de l’histoire du stock et de son évolution, �� pour le diagnostic sur l’état global du stock (toutes zones confondues ou par
zone),
�������������������������������������������������21 - Déterminisme des modèles, conditions d’équilibre, population unique, caractéristiques stables du stock 22 - Capturabilité constante, pas de temps de latence

191
�� pour la médiation des acteurs externes à la pêcherie (en mettant en évidence les gains ou pertes potentiels de toute modification de l’effort de pêche, ainsi que l’incertitude existante sur ces estimations).
Il reste que le modèle global et l’analyse présentée ici constitue une approche assez rudimentaire : �� C’est une analyse de la ressource toutes espèces confondues, ce qui peut
masquer des différences importantes entre espèces. �� Il n’y a pas de prise en compte des changements du diagramme d’exploitation,
liés notamment aux réglementations sur le maillage et les fermetures. Donc le modèle global n’est pas un outil de gestion de ces aspects
�� Le modèle prend en compte seulement la phase marine (pêche industrielle), donc il n’y a pas d’analyse des interactions entre pêcheries et par conséquent il n’y a pas de gestion pluri-metiers.
Ces limites justifient l’analyse de la ressource par le modèle structural, objet du chapitre suivant.

192
9. DIAGNOSTICS PAR ANALYSE DE PSEUDOCOHORTE ET SIMULATIONS DE PRODUCTION
Les espèces principalement exploitées à Madagascar, et les crevettes pénéides tropicales côtières en général, se caractérisent par un cycle vital court puisque leur durée de vie n’excède pas 18 mois. La pêche au cours de la campagne crevettière peut être considérée comme l’exploitation d’une cohorte recrutée au début de l’année. La croissance étant très rapide, l’unité de temps utilisée sera donc le mois. La pêcherie crevettière malgache est une pêcherie multispécifique/multiflottilles. L’ensemble des données de capture des trois segments de la pêche (industrielle, artisanale et traditionnelle) est utilisé pour l’analyse de cohorte. Il est donc nécessaire de connaître la capture totale réalisée pour chacune des espèces et pour chacun des segments de la pêcherie. Les pêcheries sont ici modélisées en utilisant le modèle de pseudocohorte de Jones (1984) basé sur l’analyse des structures en taille 23. Les rendements par recrue sont estimés par le modèle de Thompson et Bell fondé sur la longueur.
9.1. CHOIX DE LA METHODE ET RAPPEL SUR LE PRINCIPE DE L’ANALYSE
Analyse en pseudocohorte : Pour les crustacés en général, du fait de la croissance par mue et des difficultés de déterminer les âges (absence de pièces dures permettant la lecture directe), on est souvent obligé de recourir à une méthode fondée sur la longueur. Compte tenu que la ressource peut être considérée comme annuelle, l’analyse de pseudocohorte fondée sur la longueur est utilisée pour évaluer l’état des stocks. On admet ainsi que l’évolution de l’effectif de l’ensemble des classes de longueur capturée au cours d’une année reflète l’évolution de l’effectif d’une seule cohorte pendant toute sa durée d’existence. La méthode, dérivée de l’analyse de cohorte de Jones (1984), a été appliquée par Dintheer et al. (1996 ; 1998) et Moguedet et al. 24 (2001) sur les données de la pêcherie crevettière malgache de 1994 et 1998. Marcille (1978) avait utilisé le modèle de rendement par recrue de Beverton et Holt (1957) et le modèle exponentiel de Ricker 25 (1958) pour la modélisation analytique de la dynamique des populations de crevettes pénéides exploitées à Madagascar. �������������������������������������������������23 L’Analyse de cohorte est aussi appelé analyse séquentielle de population (ou en anglais VPA pour ‘’Virtual Population Analysis’’). Ce sont des modèles décrivant l’évolution historique d’une population par la reconstruction de l’évolution des cohortes constituant cette population. 24 VPA par le programme JONYFEC (Programme IFREMER, Laurec, Mesnil et Beucher) et simulation par le programme VERDEJON (Programme IFREMER, C . De Verdelhan) : Ce dernier est un programme intégré d’analyse des pseudocohortes de Jones (permettant également la relecture de fichier JONYFEC) et de simulation plurispécifique et plurimétiers permettant ainsi de réaliser des prévisions de captures pour toute modification (segment de la pêche, variations de l’effort de pêche, …) 25 Programme FRG 708 (Lawrance E. Gales, 1963 , cité par Marcille, 1978)

193
L’approche fait appel à l’équation de croissance de Von Bertalanffy pour convertir la longueur en âge :
t(L) = t0 – 1/K . ln (1 – L/L�) ou en classe d’âge :
dt = t (L2) – t (L1) = 1/K . ln [ (L� - L1) / (L� – L2) ] L’équation de l’analyse de cohorte, d’après l’approximation de Pope (1972) en considérant la capture durant une période de temps quelconque ��, est la suivante :
N(t) = [ N(t + dt) . exp(M/2 . dt) + C(t , t+dt) ] . exp(M/2 . dt) C(t, t+dt) = N(t) . F/Z . [ 1 – exp(-Z . dt) ]
N(t) : Nombre de survivant atteignant l’âge t dt : Intervalle de temps M :Mortalité naturelle C(t, t+dt) : Equation de capture fondée sur l’âge F : Mortalité par pêche Z : Mortalité totale ( Z = F + M)
Comme les classes de longueur peuvent être transformées en intervalles d’âge par l’équation inverse de Von Bertalanffy, l’équation de Pope fondée sur l’âge peut convertir en une version fondée sur la longueur, et seul le terme exp(M/2 . dt)�doit être changé����
H(L1 , L2 ) = exp(M/2 . dt) = [(L� - L1) / (L� – L2) ]M / (2 . K)
N(L1) = [ N(L2) . H(L1,L2) + C(L1,L2) ] . H(L1,L2)
C(L1,L2) = N(L1) . F / Z . [ 1 – exp(-Z dt) ] N(L1) : Nombre de crevettes atteignant la longueur L1 N(L2) : Nombre de crevettes atteignant la longueur L2 H(L1,L2) : L’inverse de la fraction de N(L1) qui survit aux morts naturelles entre t(L1) et
t(L1)+ dt/2 C(L1,L2) : Nombre de crevettes capturées de longueurs comprises entre L1 et L2 Connaissant les captures C(L1,L2), la mortalité naturelle M, N(L) et les mortalités par pêche F, ces équations permettent d’estimer les effectifs de la cohorte. La méthode suppose un système à paramètres constants. Elle est très sensible aux valeurs des paramètres de croissance K et L� et à la valeur du paramètre de mortalité M. L’approximation de la VPA est valable pour les valeurs de F.dt allant jusqu’à 1,2 et de M. dt allant jusqu’à 0,3 (Pope, 1972).

194
Modèle de rendement par recrue : Le modèle de Thompson et Bell fondé sur la longueur emprunte ses données d’entrée à une analyse de cohorte de Jones fondée sur la longueur. L’ensemble de formule pour cette analyse est le suivant : - Mortalité totale : Zi = M + mf . Fi où mf : multiplicateur de l’effort - Equation de survie : N(Li+1) = N(Li) [ 1/Hi – mf . Fi / Zi ] / [Hi – mf.Fi / Zi ]
où Hi = [(L� - Li) / (L� - Li+1) ] M / 2K = Ni . exp (-mf . F + M) . dt
- Equations de capture :
en effectif Ci = [ N(Li) – N(Li+1)] . mf . Fi / Zi en poids Yi = Ci . a . [ (Li + Li+1) / 2 ] b
où a et b sont les constantes de la relation longueur-poids Les données de sortie de chaque analyse de cohorte sont constituées de : �� [ Nsi ] les effectifs moyens de la population �� [ Fsi ] les mortalités globales par pêche pour chacune des espèces.
Chacun de ces F revenant à une classe de longueur provient de la mortalité par pêche combinée des différents engins de pêche (flottilles). Ainsi, les F totaux correspondant à chacune des classes de longueur peuvent être redistribués entre les différentes flottilles au moyen de l’équation de ventilation suivante :
Fg = FTot . Cg / CTot Les effectifs capturés par classe de taille sont ainsi ventilés au prorata des : [ Csgi ] ou nombre de crevettes de la classe de longueur i de l’espèce s capturées par la flottille g au cours de la période considérée.
9.2. DONNEES ET TRAITEMENTS PREALABLES Débarquements par segment de flotte et par grande zone : Sur la moyenne de production enregistrée de 2000 à 2002, les apports en équivalents crevettes entières, toutes espèces confondues, sur la côte ouest malgache sont de ������ tonnes environ. L’estimation de la répartition par zone et par segment de pêche de cette quantité débarquée est donnée dans le tableau suivant :
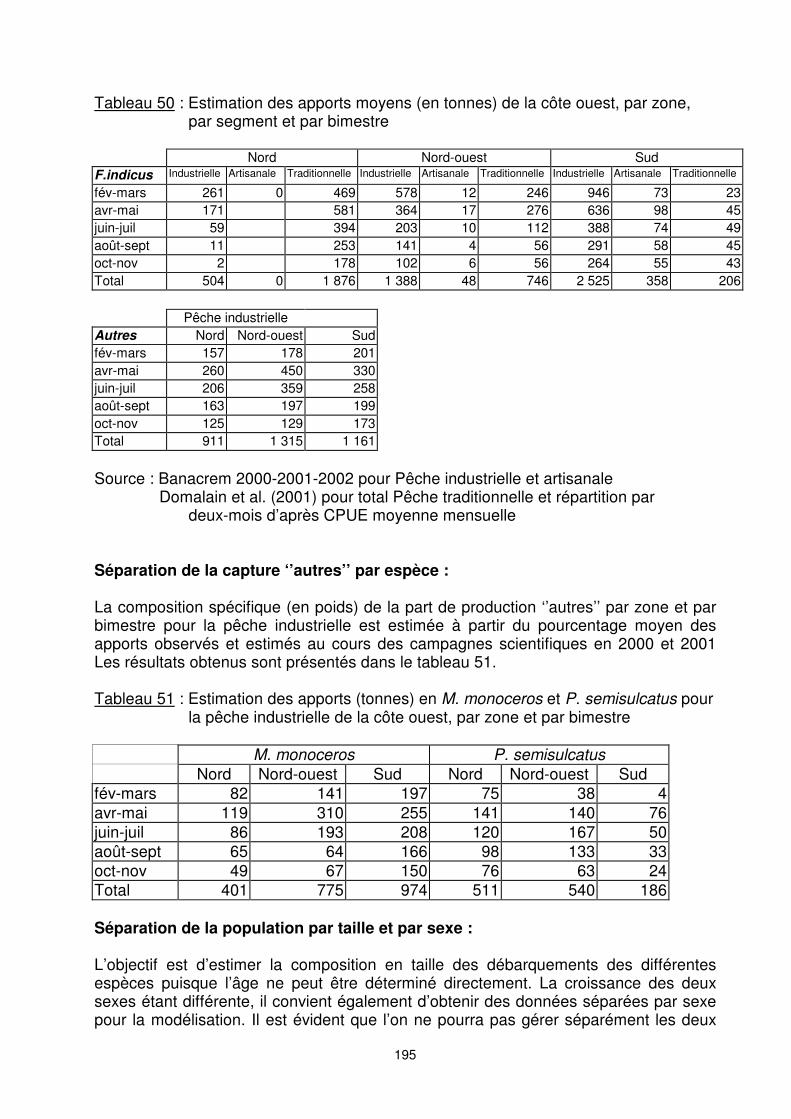
195
Tableau 50 : Estimation des apports moyens (en tonnes) de la côte ouest, par zone, par segment et par bimestre
Nord Nord-ouest Sud F.indicus Industrielle Artisanale Traditionnelle Industrielle Artisanale Traditionnelle Industrielle Artisanale Traditionnelle
fév-mars 261 0 469 578 12 246 946 73 23 avr-mai 171 581 364 17 276 636 98 45 juin-juil 59 394 203 10 112 388 74 49 août-sept 11 253 141 4 56 291 58 45 oct-nov 2 178 102 6 56 264 55 43 Total 504 0 1 876 1 388 48 746 2 525 358 206
Pêche industrielle Autres Nord Nord-ouest Sud fév-mars 157 178 201 avr-mai 260 450 330 juin-juil 206 359 258 août-sept 163 197 199 oct-nov 125 129 173 Total 911 1 315 1 161 Source : Banacrem 2000-2001-2002 pour Pêche industrielle et artisanale Domalain et al. (2001) pour total Pêche traditionnelle et répartition par
deux-mois d’après CPUE moyenne mensuelle Séparation de la capture ‘’autres’’ par espèce : La composition spécifique (en poids) de la part de production ‘’autres’’ par zone et par bimestre pour la pêche industrielle est estimée à partir du pourcentage moyen des apports observés et estimés au cours des campagnes scientifiques en 2000 et 2001 Les résultats obtenus sont présentés dans le tableau 51. Tableau 51 : Estimation des apports (tonnes) en M. monoceros et P. semisulcatus pour
la pêche industrielle de la côte ouest, par zone et par bimestre
M. monoceros P. semisulcatus Nord Nord-ouest Sud Nord Nord-ouest Sud
fév-mars 82 141 197 75 38 4 avr-mai 119 310 255 141 140 76 juin-juil 86 193 208 120 167 50 août-sept 65 64 166 98 133 33 oct-nov 49 67 150 76 63 24 Total 401 775 974 511 540 186 Séparation de la population par taille et par sexe : L’objectif est d’estimer la composition en taille des débarquements des différentes espèces puisque l’âge ne peut être déterminé directement. La croissance des deux sexes étant différente, il convient également d’obtenir des données séparées par sexe pour la modélisation. Il est évident que l’on ne pourra pas gérer séparément les deux

196
sexes, mais cette procédure est utile pour avoir une modélisation aussi réaliste que possible de la pêcherie. L’analyse porte sur les deux principales espèces : F. indicus et M. monoceros. La première est typique d’une pêcherie séquentielle et représente la première espèce cible, et la seconde une espèce exploitée pratiquement seule par la pêche industrielle. La structure en taille des captures est extrapolée à partir des données d’échantillonnage 2001 des observateurs à bord pour les cas de la pêche industrielle et de la pêche artisanale. Les échantillons des années 1998 et 1999 sont écartés parce qu’ils ne sont pas représentatifs de la production totale annuelle. En effet, pour ces deux années, les échantillonnages n’ont commencé que vers la fin de la haute saison. Les échantillons utilisés pour le cas de la pêche traditionnelle sont ceux des mensurations réalisées lors des enquêtes PNRC pour les années 1998 et 1999. Aucune autre source de données récentes n’est encore disponible pour ce segment de l’exploitation. Les compositions moyennes sont établies pour chaque espèce, segment d’exploitation, zone et bimestre selon les formules suivantes :
Ni = ni . Ytot. / Wéch. Ni : effectif capturé pour la taille i ni : effectif échantillonné pour la taille i Ytot. : Poids total capturé Wéch. : Poids de l’échantillon
avec : Wéch. = Σ ni . W i W i : Poids individuel pour la taille i Les coefficients d’extrapolation utilisés pour estimer la composition en taille des débarquements sont très variables selon les zone, espèce et segment de la pêcherie (tableau 52). Tableau 52 : Coefficients d’extrapolation utilisés pour la reconstitution de la structrure
démographique des débarquements
Nord Nord-ouest Sud F. indicus M. monoceros F. indicus M. monoceros F. indicus M. monoceros
Industriel 17 937 32 526 31 180 36 374 15 618 38 836 Artisanal 20 2 808 4 898 Traditionnel 4 937 2 145 843 L’obtention d’une structure par taille-sexe de la population pêchée nécessite la connaissance du sex-ratio qui varie énormément en fonction de la taille (tableau 53). Ce faisant, la distinction des femelles et des mâles se justifie du fait de différence de croissance entre les sexes. Nous avons utilisé les valeurs moyennes obtenues par GLM à partir des données Observateurs (cf. § 7.1.2.2. et 7.1.3.2.). Nous considérons que ces valeurs ajustées sont valables pour les trois segments de la pêcherie et les différentes zones de pêche.
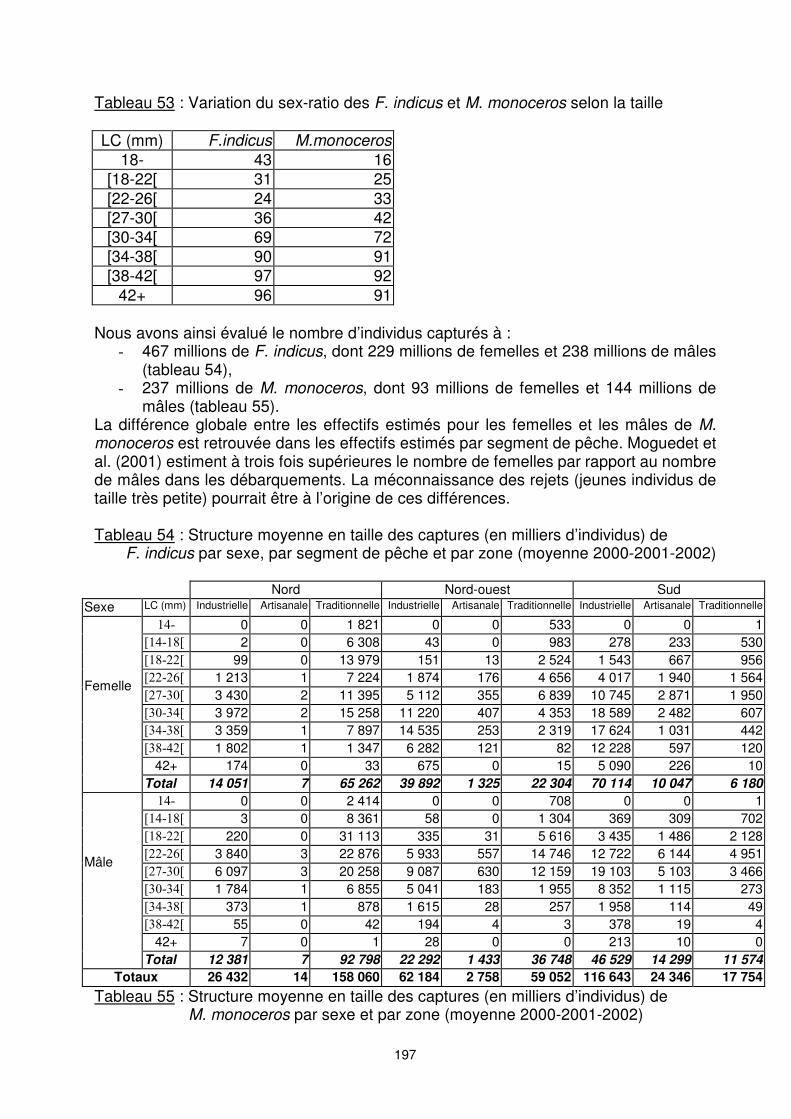
197
Tableau 53 : Variation du sex-ratio des F. indicus et M. monoceros selon la taille LC (mm) F.indicus M.monoceros
18- 43 16 [18-22[ 31 25 [22-26[ 24 33 [27-30[ 36 42 [30-34[ 69 72 [34-38[ 90 91 [38-42[ 97 92
42+ 96 91 Nous avons ainsi évalué le nombre d’individus capturés à : �� 467 millions de F. indicus, dont 229 millions de femelles et 238 millions de mâles
(tableau 54), �� 237 millions de M. monoceros, dont 93 millions de femelles et 144 millions de
mâles (tableau 55). La différence globale entre les effectifs estimés pour les femelles et les mâles de M. monoceros est retrouvée dans les effectifs estimés par segment de pêche. Moguedet et al. (2001) estiment à trois fois supérieures le nombre de femelles par rapport au nombre de mâles dans les débarquements. La méconnaissance des rejets (jeunes individus de taille très petite) pourrait être à l’origine de ces différences. Tableau 54 : Structure moyenne en taille des captures (en milliers d’individus) de F. indicus par sexe, par segment de pêche et par zone (moyenne 2000-2001-2002)
Nord Nord-ouest Sud Sexe LC (mm) Industrielle Artisanale Traditionnelle Industrielle Artisanale Traditionnelle Industrielle Artisanale Traditionnelle
���� 0 0 1 821 0 0 533 0 0 1 ����� 2 0 6 308 43 0 983 278 233 530 ����� 99 0 13 979 151 13 2 524 1 543 667 956 ������ 1 213 1 7 224 1 874 176 4 656 4 017 1 940 1 564 � ���� 3 430 2 11 395 5 112 355 6 839 10 745 2 871 1 950 ������ 3 972 2 15 258 11 220 407 4 353 18 589 2 482 607 ����� 3 359 1 7 897 14 535 253 2 319 17 624 1 031 442 ����� 1 802 1 1 347 6 282 121 82 12 228 597 120
42+ 174 0 33 675 0 15 5 090 226 10
Femelle
Total 14 051 7 65 262 39 892 1 325 22 304 70 114 10 047 6 180 ���� 0 0 2 414 0 0 708 0 0 1
����� 3 0 8 361 58 0 1 304 369 309 702 ����� 220 0 31 113 335 31 5 616 3 435 1 486 2 128 ������ 3 840 3 22 876 5 933 557 14 746 12 722 6 144 4 951 � ���� 6 097 3 20 258 9 087 630 12 159 19 103 5 103 3 466 ������ 1 784 1 6 855 5 041 183 1 955 8 352 1 115 273 ����� 373 1 878 1 615 28 257 1 958 114 49 ����� 55 0 42 194 4 3 378 19 4
42+ 7 0 1 28 0 0 213 10 0
Mâle
Total 12 381 7 92 798 22 292 1 433 36 748 46 529 14 299 11 574 Totaux 26 432 14 158 060 62 184 2 758 59 052 116 643 24 346 17 754
Tableau 55 : Structure moyenne en taille des captures (en milliers d’individus) de M. monoceros par sexe et par zone (moyenne 2000-2001-2002)
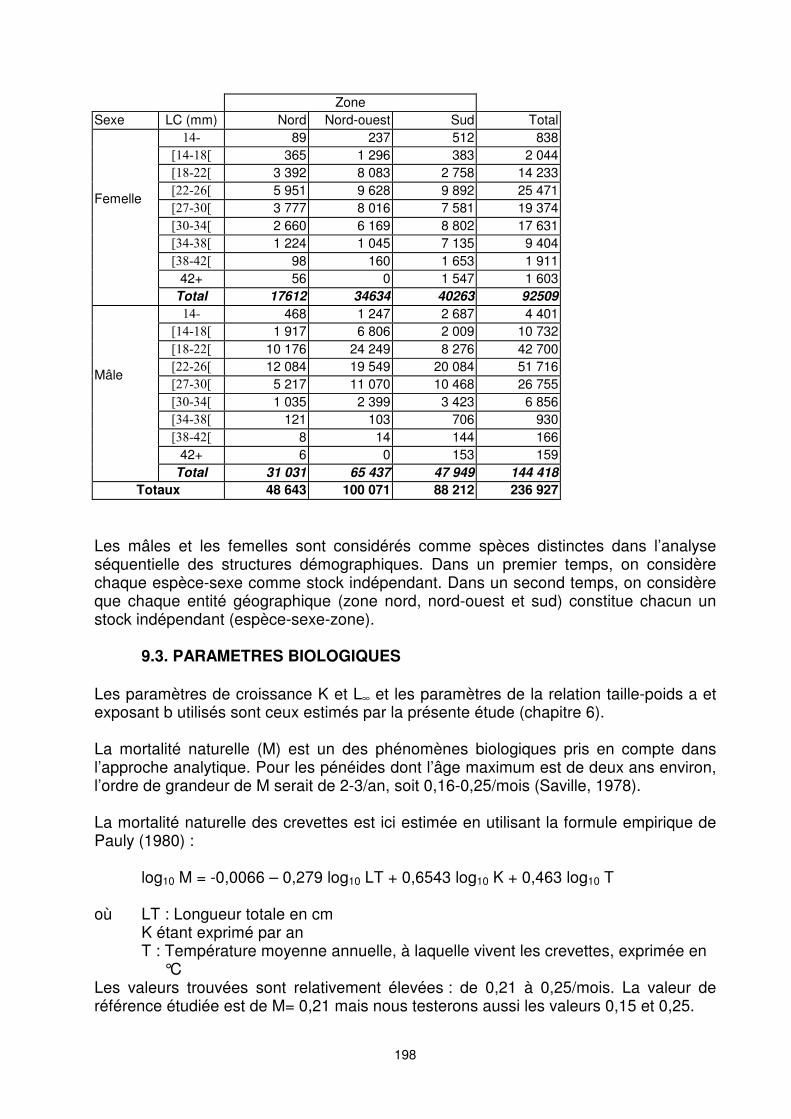
198
Zone Sexe LC (mm) Nord Nord-ouest Sud Total
���� 89 237 512 838 ����� 365 1 296 383 2 044 ����� 3 392 8 083 2 758 14 233 ������ 5 951 9 628 9 892 25 471 � ���� 3 777 8 016 7 581 19 374 ������ 2 660 6 169 8 802 17 631 ����� 1 224 1 045 7 135 9 404 ����� 98 160 1 653 1 911
42+ 56 0 1 547 1 603
Femelle
Total 17612 34634 40263 92509 ���� 468 1 247 2 687 4 401
����� 1 917 6 806 2 009 10 732 ����� 10 176 24 249 8 276 42 700 ������ 12 084 19 549 20 084 51 716 � ���� 5 217 11 070 10 468 26 755 ������ 1 035 2 399 3 423 6 856 ����� 121 103 706 930 ����� 8 14 144 166
42+ 6 0 153 159
Mâle
Total 31 031 65 437 47 949 144 418 Totaux 48 643 100 071 88 212 236 927
Les mâles et les femelles sont considérés comme spèces distinctes dans l’analyse séquentielle des structures démographiques. Dans un premier temps, on considère chaque espèce-sexe comme stock indépendant. Dans un second temps, on considère que chaque entité géographique (zone nord, nord-ouest et sud) constitue chacun un stock indépendant (espèce-sexe-zone).
9.3. PARAMETRES BIOLOGIQUES Les paramètres de croissance K et L∞ et les paramètres de la relation taille-poids a et exposant b utilisés sont ceux estimés par la présente étude (chapitre 6). La mortalité naturelle (M) est un des phénomènes biologiques pris en compte dans l’approche analytique. Pour les pénéides dont l’âge maximum est de deux ans environ, l’ordre de grandeur de M serait de 2-3/an, soit 0,16-0,25/mois (Saville, 1978). La mortalité naturelle des crevettes est ici estimée en utilisant la formule empirique de Pauly (1980) :
log10 M = -0,0066 – 0,279 log10 LT + 0,6543 log10 K + 0,463 log10 T où LT : Longueur totale en cm K étant exprimé par an T : Température moyenne annuelle, à laquelle vivent les crevettes, exprimée en
°C Les valeurs trouvées sont relativement élevées : de 0,21 à 0,25/mois. La valeur de référence étudiée est de M= 0,21 mais nous testerons aussi les valeurs 0,15 et 0,25.

199
Le Reste (1978) estime à 0,21/mois la mortalité naturelle des F. indicus. Marcille (1978) utilise l’intervalle de valeur [0,15-0,25] pour la modélisation du stock des F. indicus et de [0,10-0,20] pour la modélisation du stock des M. monoceros. Les valeurs utilisées par Moguedet et al. (2001) sont de 0.20/mois pour les F. indicus femelle et de 0,18/mois pour M. monoceros et F. indicus mâle.
9.4. ESTIMATIONS DES MORTALITES PAR PECHE ET DES RECRUTEMENTS INITIAUX
Les résultats des estimations des mortalités par pêche en fonction de la taille, pour différentes hypothèses de mortalités naturelles et de mortalités par pêche terminales sont présentés sur les figures 128 et 129. Nous retrouvons dans toutes les courbes la propriété de la convergence pour différentes valeurs initiales de Ft. Nous avons également vérifié que le choix de M est sans grande importance sur l’estimation du coefficient de mortalité par pêche. Aussi, le choix de M a une influence considérable sur l’estimation des effectifs aux âges et donc du recrutement initial. La sélection du vecteur de mortalité par pêche adopté est effectuée sous la contrainte d’une mortalité constante (ou presque) après le plein recrutement (classe de taille avant le groupe +) et qui donnent des effectifs initiaux comparables entre les deux sexes. Aussi bien pour les F. indicus que pour les M. monoceros, les groupes ‘’+’’, considérés comme la taille terminale de prise en compte de la croissance, sont de [38-42[ pour les femelles. Pour les mâles, ils sont de [34-36] pour les F. indicus et de [26-28] pour les M. monoceros. F. indicus : Pour F. indicus, nous adoptons les résultas issus de la VPA pour un Ft=0,2 et M=0,21 pour les femelles et M=0,15 pour les mâles. Les diagrammes d’exploitation obtenus pour chaque type de pêche sont variables (figure 130). Les mortalités induites par la pêche artisanale sont marginales. La pêche traditionnelle est responsable des prélèvements d’une grande partie des crevettes de petite taille ; jusqu’au [26, 29] mm LC. A partir de cette classe de taille, les prélèvements de la pêche traditionnelle décroient et ceux de la pêche industrielle continuent sa progression.
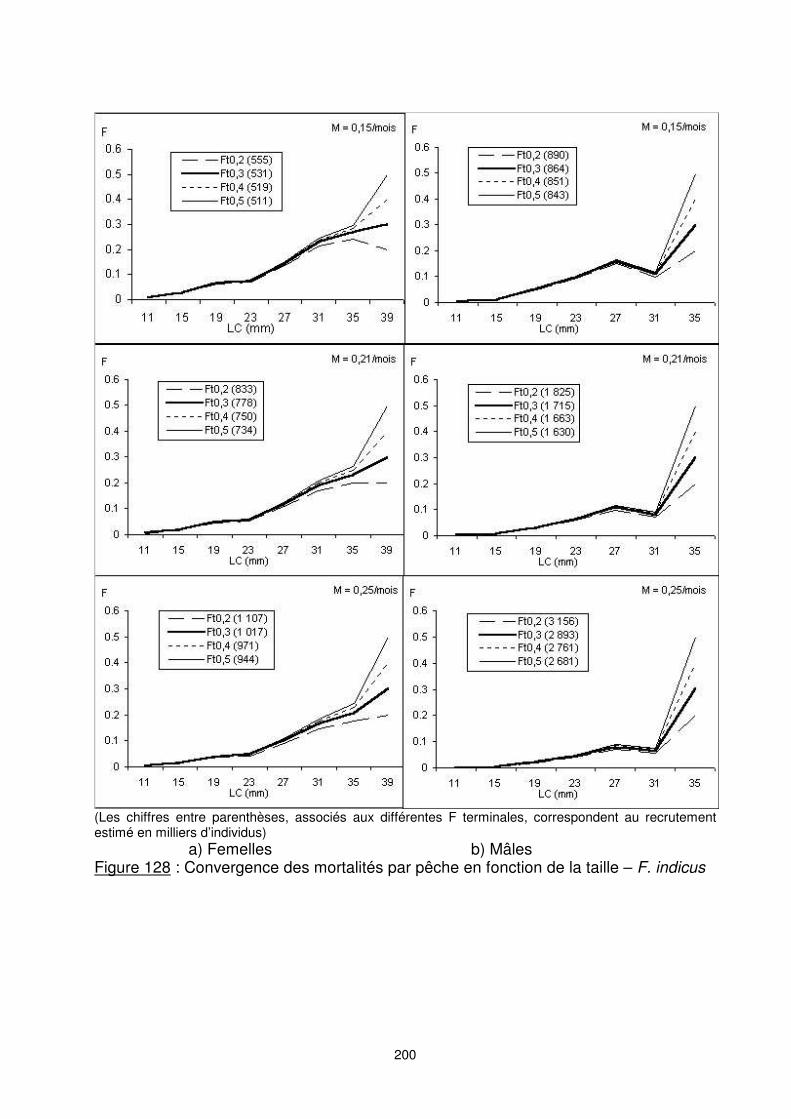
200
(Les chiffres entre parenthèses, associés aux différentes F terminales, correspondent au recrutement estimé en milliers d’individus) a) Femelles b) Mâles Figure 128 : Convergence des mortalités par pêche en fonction de la taille – F. indicus
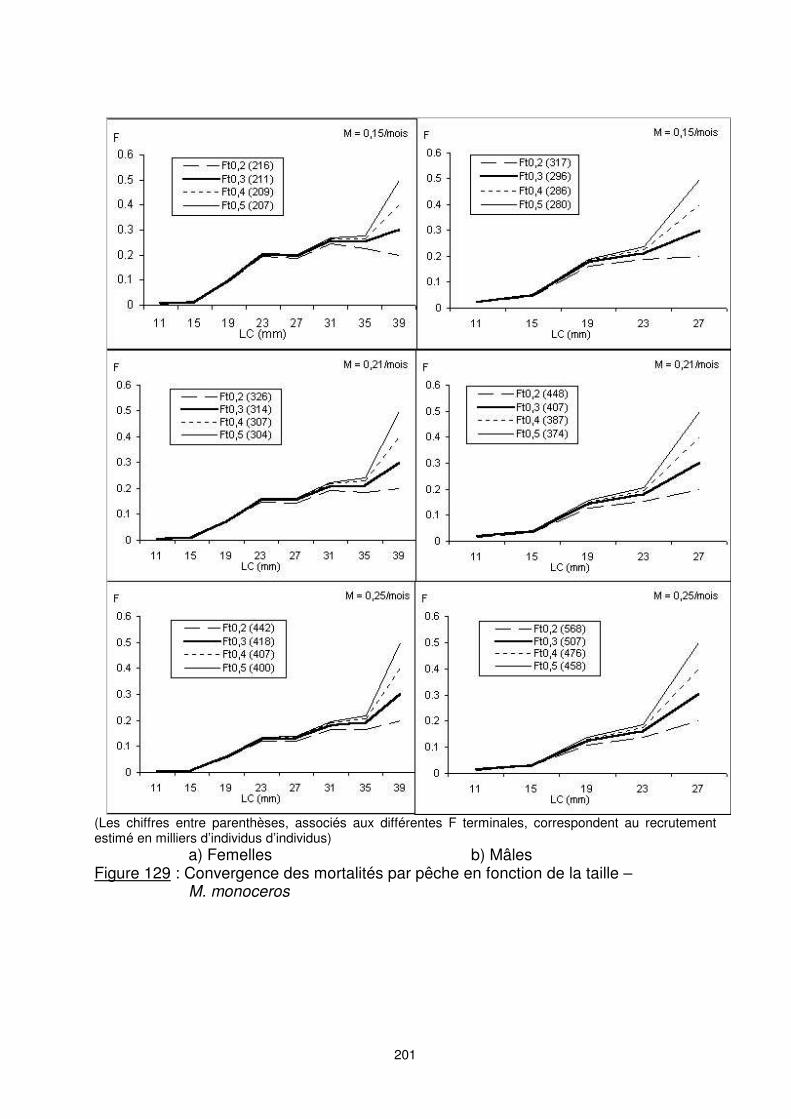
201
(Les chiffres entre parenthèses, associés aux différentes F terminales, correspondent au recrutement estimé en milliers d’individus d’individus) a) Femelles b) Mâles Figure 129 : Convergence des mortalités par pêche en fonction de la taille –
M. monoceros
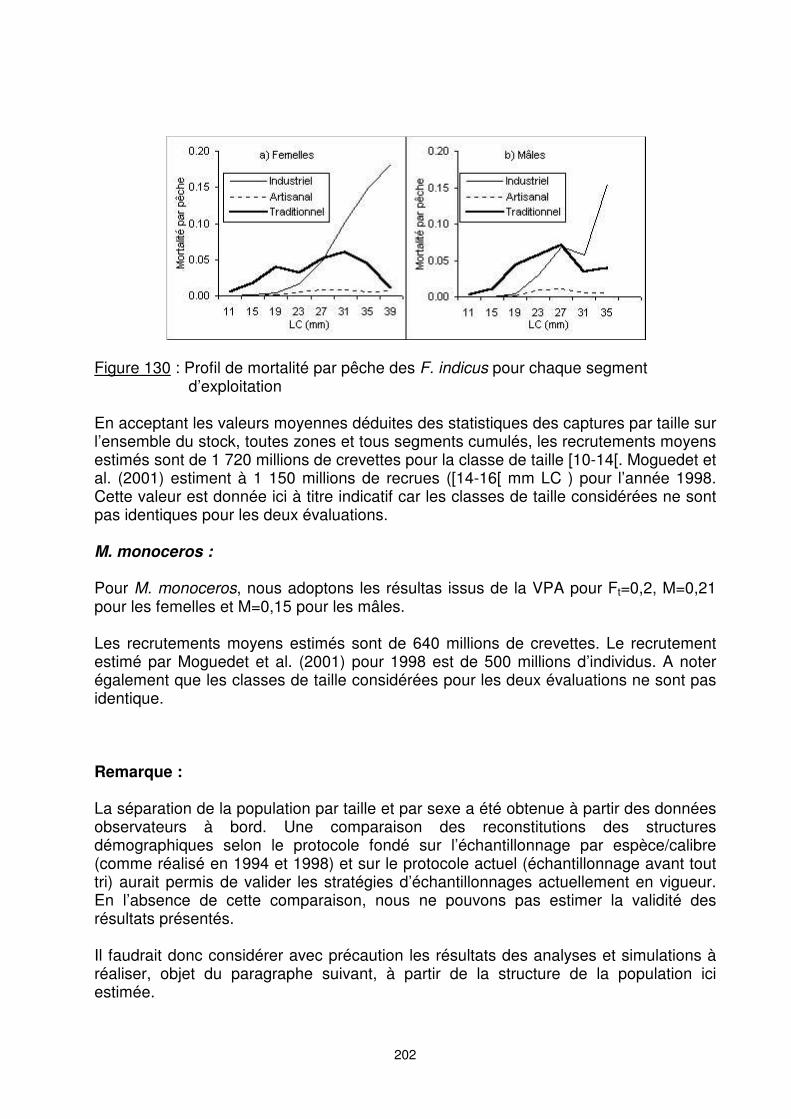
202
Figure 130 : Profil de mortalité par pêche des F. indicus pour chaque segment
d’exploitation En acceptant les valeurs moyennes déduites des statistiques des captures par taille sur l’ensemble du stock, toutes zones et tous segments cumulés, les recrutements moyens estimés sont de 1 720 millions de crevettes pour la classe de taille [10-14[. Moguedet et al. (2001) estiment à 1 150 millions de recrues ([14-16[ mm LC ) pour l’année 1998. Cette valeur est donnée ici à titre indicatif car les classes de taille considérées ne sont pas identiques pour les deux évaluations. M. monoceros : Pour M. monoceros, nous adoptons les résultas issus de la VPA pour Ft=0,2, M=0,21 pour les femelles et M=0,15 pour les mâles. Les recrutements moyens estimés sont de 640 millions de crevettes. Le recrutement estimé par Moguedet et al. (2001) pour 1998 est de 500 millions d’individus. A noter également que les classes de taille considérées pour les deux évaluations ne sont pas identique. Remarque : La séparation de la population par taille et par sexe a été obtenue à partir des données observateurs à bord. Une comparaison des reconstitutions des structures démographiques selon le protocole fondé sur l’échantillonnage par espèce/calibre (comme réalisé en 1994 et 1998) et sur le protocole actuel (échantillonnage avant tout tri) aurait permis de valider les stratégies d’échantillonnages actuellement en vigueur. En l’absence de cette comparaison, nous ne pouvons pas estimer la validité des résultats présentés. Il faudrait donc considérer avec précaution les résultats des analyses et simulations à réaliser, objet du paragraphe suivant, à partir de la structure de la population ici estimée.
203
9.5. DIAGNOSTICS DE RENDEMENT PAR RECRUE ET SIMULATIONS DE PRODUCTION RESULTANT DE CHANGEMENTS DE NIVEAU D’EFFORT
9.5.1. Analyse globale
9.5.1.1. Le cas des F. indicus
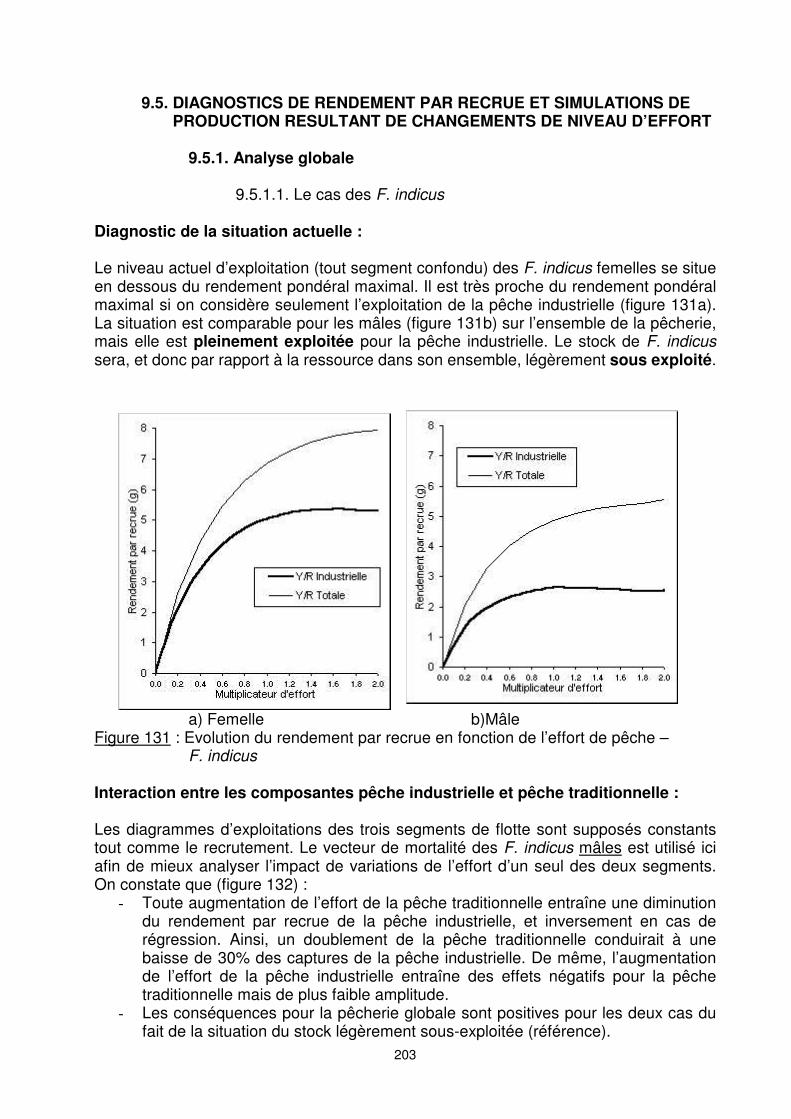
Diagnostic de la situation actuelle : Le niveau actuel d’exploitation (tout segment confondu) des F. indicus femelles se situe en dessous du rendement pondéral maximal. Il est très proche du rendement pondéral maximal si on considère seulement l’exploitation de la pêche industrielle (figure 131a). La situation est comparable pour les mâles (figure 131b) sur l’ensemble de la pêcherie, mais elle est pleinement exploitée pour la pêche industrielle. Le stock de F. indicus sera, et donc par rapport à la ressource dans son ensemble, légèrement sous exploité.
a) Femelle b)Mâle Figure 131 : Evolution du rendement par recrue en fonction de l’effort de pêche –
F. indicus Interaction entre les composantes pêche industrielle et pêche traditionnelle : Les diagrammes d’exploitations des trois segments de flotte sont supposés constants tout comme le recrutement. Le vecteur de mortalité des F. indicus mâles est utilisé ici afin de mieux analyser l’impact de variations de l’effort d’un seul des deux segments. On constate que (figure 132) : �� Toute augmentation de l’effort de la pêche traditionnelle entraîne une diminution
du rendement par recrue de la pêche industrielle, et inversement en cas de régression. Ainsi, un doublement de la pêche traditionnelle conduirait à une baisse de 30% des captures de la pêche industrielle. De même, l’augmentation de l’effort de la pêche industrielle entraîne des effets négatifs pour la pêche traditionnelle mais de plus faible amplitude.
�� Les conséquences pour la pêcherie globale sont positives pour les deux cas du fait de la situation du stock légèrement sous-exploitée (référence).
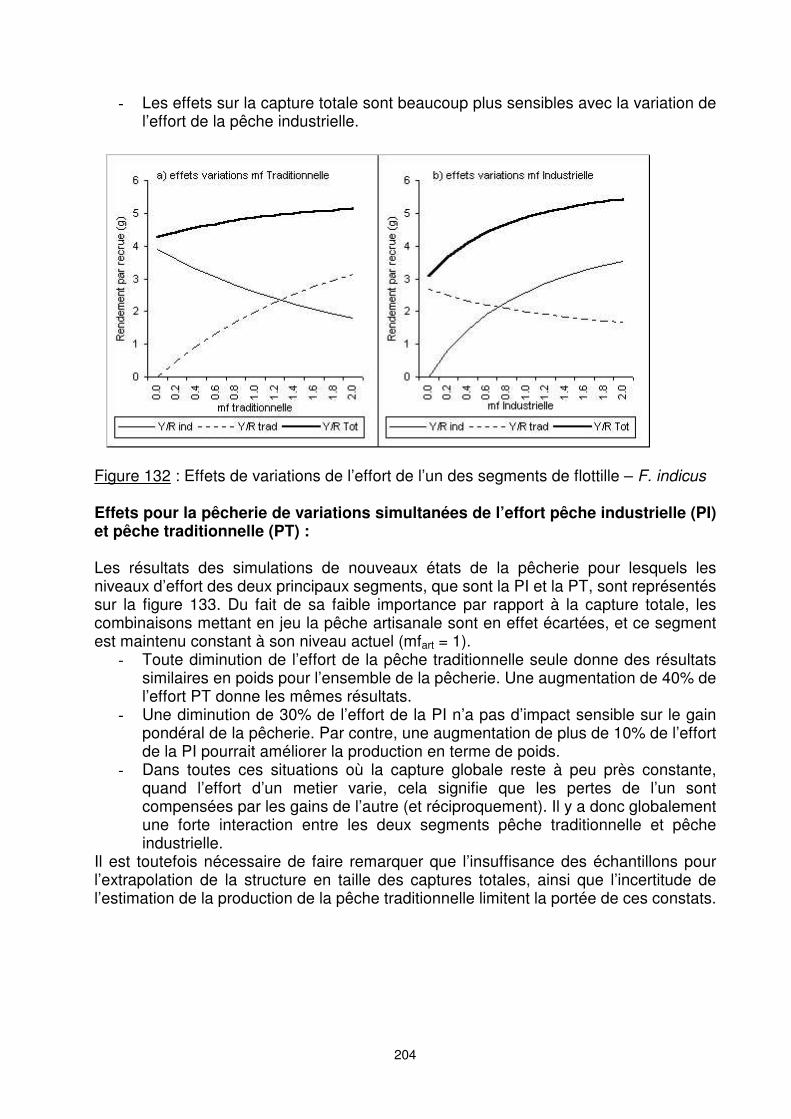
204
�� Les effets sur la capture totale sont beaucoup plus sensibles avec la variation de l’effort de la pêche industrielle.
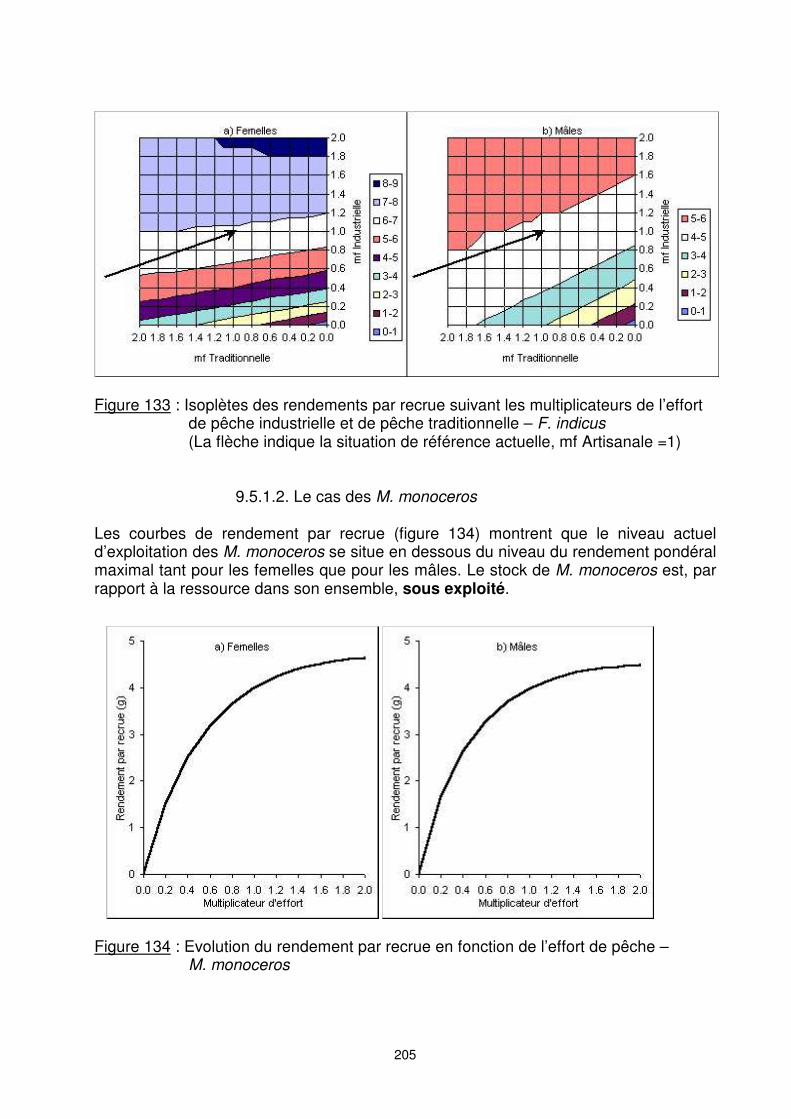
Figure 132 : Effets de variations de l’effort de l’un des segments de flottille – F. indicus Effets pour la pêcherie de variations simultanées de l’effort pêche industrielle (PI) et pêche traditionnelle (PT) : Les résultats des simulations de nouveaux états de la pêcherie pour lesquels les niveaux d’effort des deux principaux segments, que sont la PI et la PT, sont représentés sur la figure 133. Du fait de sa faible importance par rapport à la capture totale, les combinaisons mettant en jeu la pêche artisanale sont en effet écartées, et ce segment est maintenu constant à son niveau actuel (mfart = 1). �� Toute diminution de l’effort de la pêche traditionnelle seule donne des résultats
similaires en poids pour l’ensemble de la pêcherie. Une augmentation de 40% de l’effort PT donne les mêmes résultats.
�� Une diminution de 30% de l’effort de la PI n’a pas d’impact sensible sur le gain pondéral de la pêcherie. Par contre, une augmentation de plus de 10% de l’effort de la PI pourrait améliorer la production en terme de poids.
�� Dans toutes ces situations où la capture globale reste à peu près constante, quand l’effort d’un metier varie, cela signifie que les pertes de l’un sont compensées par les gains de l’autre (et réciproquement). Il y a donc globalement une forte interaction entre les deux segments pêche traditionnelle et pêche industrielle.
Il est toutefois nécessaire de faire remarquer que l’insuffisance des échantillons pour l’extrapolation de la structure en taille des captures totales, ainsi que l’incertitude de l’estimation de la production de la pêche traditionnelle limitent la portée de ces constats.
205
Figure 133 : Isoplètes des rendements par recrue suivant les multiplicateurs de l’effort
de pêche industrielle et de pêche traditionnelle – F. indicus (La flèche indique la situation de référence actuelle, mf Artisanale =1)
9.5.1.2. Le cas des M. monoceros Les courbes de rendement par recrue (figure 134) montrent que le niveau actuel d’exploitation des M. monoceros se situe en dessous du niveau du rendement pondéral maximal tant pour les femelles que pour les mâles. Le stock de M. monoceros est, par rapport à la ressource dans son ensemble, sous exploité.
Figure 134 : Evolution du rendement par recrue en fonction de l’effort de pêche –
M. monoceros

206
Une augmentation de l’effort de la pêche industrielle pourrait améliorer les gains pondéraux pour ce stock. Un accroissement de 20% de l’effort conduirait par exemple à une hausse de 25% des captures pour les femelles et de 20% pour les mâles. Les mêmes remarques faites pour les F. indicus demeurent toujours appropriées, notamment sur la validité de l’extrapolation des échantillons pris par les observateurs.
9.5.2. Analyse par fraction de ‘’stock’’, l’exemple des F. indicus Le cas des F. indicus présente un intérêt particulier pour l’analyse par fraction de ‘’stock’’ parce que l’importance de chaque segment de pêche est différente selon les zones.
9.5.2.1. Le cas de la zone nord La zone nord est caractérisée par la présence d’une pêche traditionnelle très dynamique où les pêcheurs utilisent des engins de pêche variés : engins non sélectifs (‘’Valakira’’, ‘’Poteau’’, …) et engin sélectif (filet maillant calé sur le fond). Les apports annuels moyens estimés sont de 180 millions de crevettes. Le nombre de mâles estimé est légèrement supérieur à celui des femelles. Pour F. indicus de la zone nord, nous adoptons les résultas issus de la VPA pour un Ft=0,2 et M=0,25 pour les femelles et M=0,15 pour les mâles (figure 135). Toujours, sous réserve d’accepter les valeurs moyennes calculées des statistiques des captures par taille sur l’ensemble de la fraction du ‘’stock’’ de la zone nord et tous segments cumulés, les recrutements moyens estimés sont de 850 millions de crevettes. En tenant compte des vecteurs de mortalités par pêche retenus, la fraction de stock de la zone nord apparaît sous-exploitée sur l’ensemble de la pêcherie. Un gain pondéral significatif pourrait attendre d’un accroissement de l’effort actuel. Par ailleurs, une situation de pleine exploitation est constatée pour la composante pêche industrielle (figure 136). (Les chiffres entre parenthèses, associés aux différentes F terminales, correspondent au recrutement estimé en milliers d’individus)
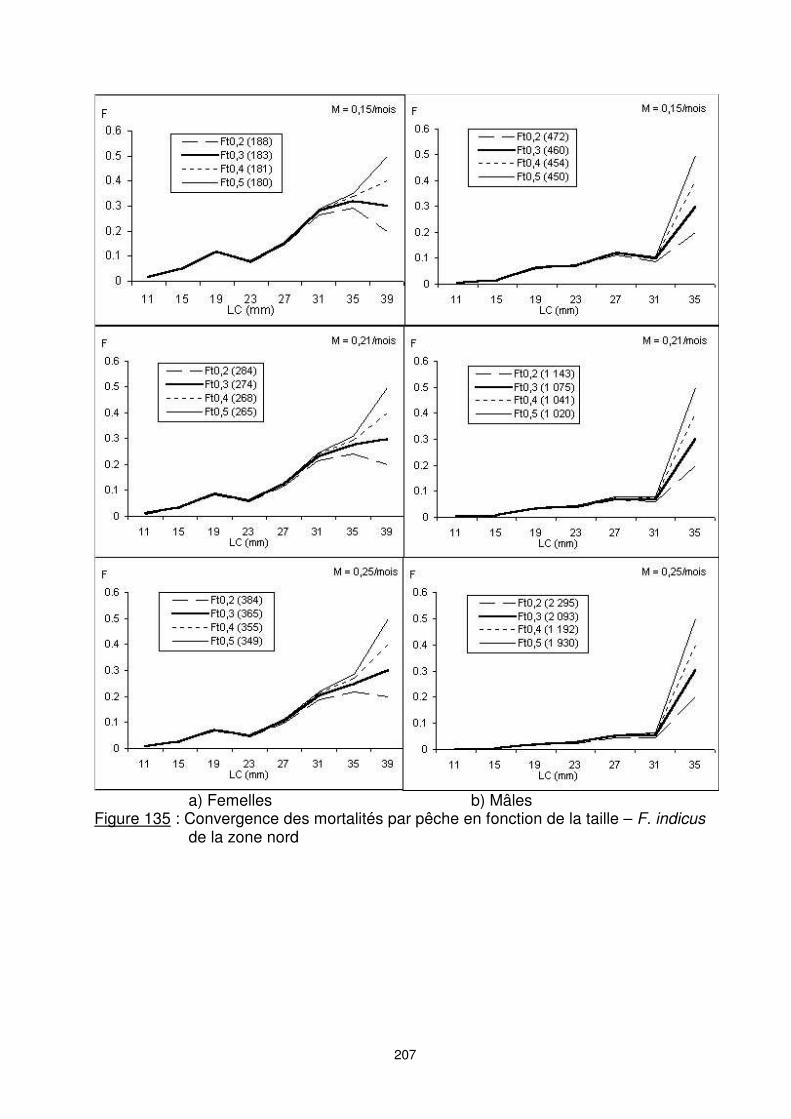
207
a) Femelles b) Mâles Figure 135 : Convergence des mortalités par pêche en fonction de la taille – F. indicus
de la zone nord
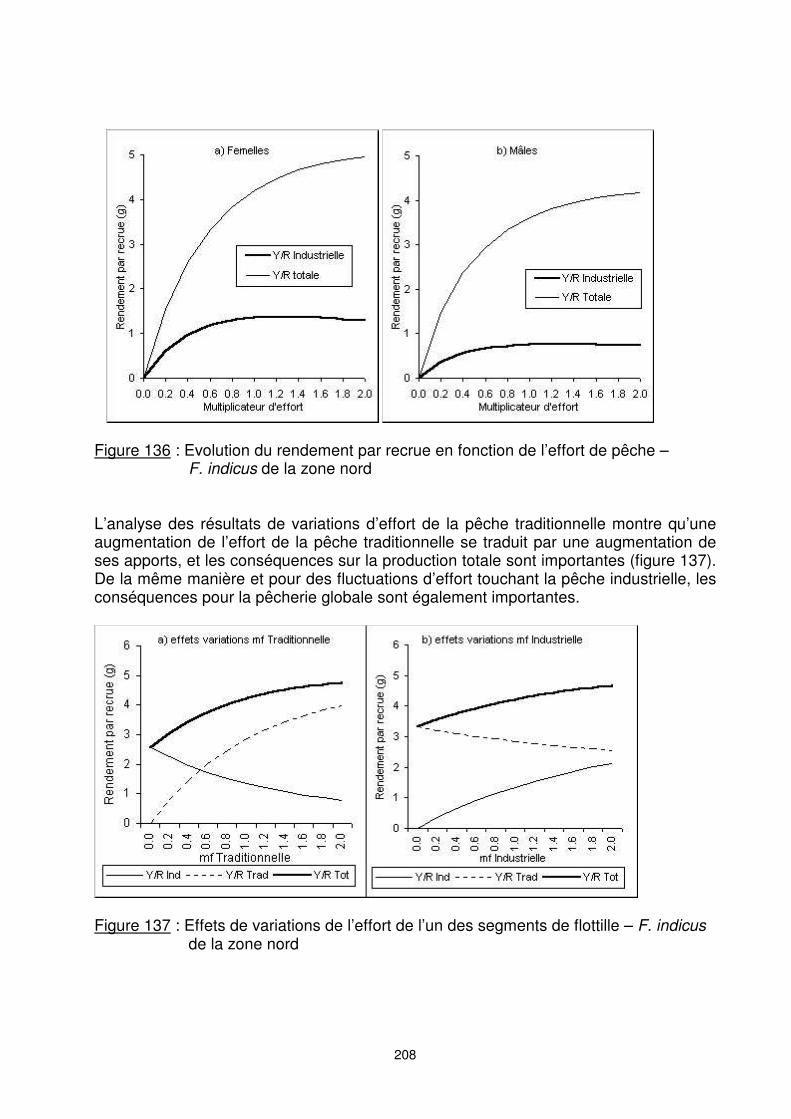
208
Figure 136 : Evolution du rendement par recrue en fonction de l’effort de pêche –
F. indicus de la zone nord L’analyse des résultats de variations d’effort de la pêche traditionnelle montre qu’une augmentation de l’effort de la pêche traditionnelle se traduit par une augmentation de ses apports, et les conséquences sur la production totale sont importantes (figure 137). De la même manière et pour des fluctuations d’effort touchant la pêche industrielle, les conséquences pour la pêcherie globale sont également importantes.
Figure 137 : Effets de variations de l’effort de l’un des segments de flottille – F. indicus
de la zone nord

209
Ce phénomène de substitution peut s’expliquer par l’interaction directe et opposée entre pêche industrielle et pêche traditionnelle car elles travaillent pratiquement sur les mêmes tailles de crevettes.
9.5.2.2. Le cas de la zone nord-ouest La zone nord-ouest est caractérisée par la dominance des activités de la pêche industrielle. Les pêches artisanale et traditionnelle existent mais très localisées et leurs apports sont relativement faibles. L’effectif des apports globaux en F. indicus pour la zone nord-ouest est estimé à 120 millions d’individus. Les vecteurs de mortalités par pêche issus de la VPA pour un Ft=0,2 et M=0,21 pour les femelles et M=0,15 pour les mâles (figure 138) sont retenus pour l’analyse du rendement par recrue. Toujours, sous réserve d’accepter les valeurs moyennes calculées des statistiques des captures par taille sur l’ensemble de la fraction du ‘’stock’’ de la zone nord-ouest et tous segments cumulés, les recrutements moyens estimés sont de 750 millions de crevettes. Considérant les vecteurs de mortalités par pêche retenus, la fraction de stock de la zone nord-ouest apparaît sous-exploitée sur l’ensemble de la pêcherie. Un gain pondéral significatif pourrait attendre d’un accroissement de l’effort actuel. De même, une situation de légère sous-exploitation est constatée pour la composante pêche industrielle (figure 139). Une augmentation de l’effort de la pêche traditionnelle se traduit par une augmentation presque linéaire de ses apports, et en conséquence une diminution, également linéaire du rendement de la pêche industrielle. La résultante de ces variations se traduit par une légère augmentation du rendement par recrue global (figure 140). L’augmentation de l’effort de la pêche industrielle se traduit par une augmentation notable de son rendement, et en conséquence une augmentation du rendement de la production totale. L’effet des variations de l’effort de la pêche industrielle sur le rendement de la pêche traditionnelle est insignifiant. Ces observations sont représentatives d’une pêcherie séquentielle car les résultats de la pêche industrielle dépendent directement de l’activité des pêcheurs traditionnels, alors que la réciproque n’est pas vraie. Pour le profil de capture de la pêche traditionnelle, Domalain et Rasoanandrasana (2001) ont noté que : �� La majeure partie des captures sur l’année de la région de la baie de Narindra
est attribuée aux ‘’Kaokobe’’, et se compose de crevettes de tailles comprises entre les classes [20-25[ mm LC et [30-35[ mm LC
�� Celles de la région de Boeny et la région de Soalala sont attribuées aux ‘’Kopiko’’. Les crevettes capturées sont toutes de tailles inférieures à [25-30[ mm LC.
Les même engins utilisés dans la région de la baie d’Ambaro (zone nord) capturent des crevettes encore plus petites que les précédentes, mais les captures relativement importantes des ‘’Periky’’ qui sont des engins sélectifs masquent le phénomène séquentiel de la pêcherie.
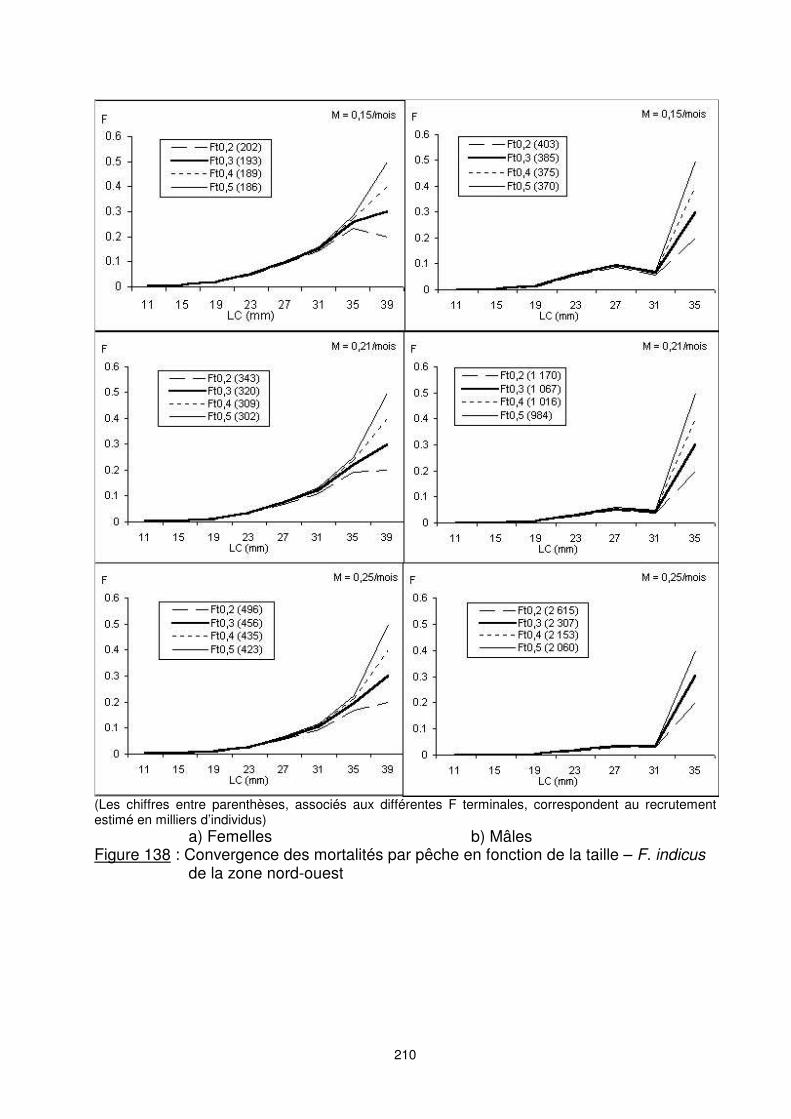
210
(Les chiffres entre parenthèses, associés aux différentes F terminales, correspondent au recrutement estimé en milliers d’individus) a) Femelles b) Mâles Figure 138 : Convergence des mortalités par pêche en fonction de la taille – F. indicus
de la zone nord-ouest
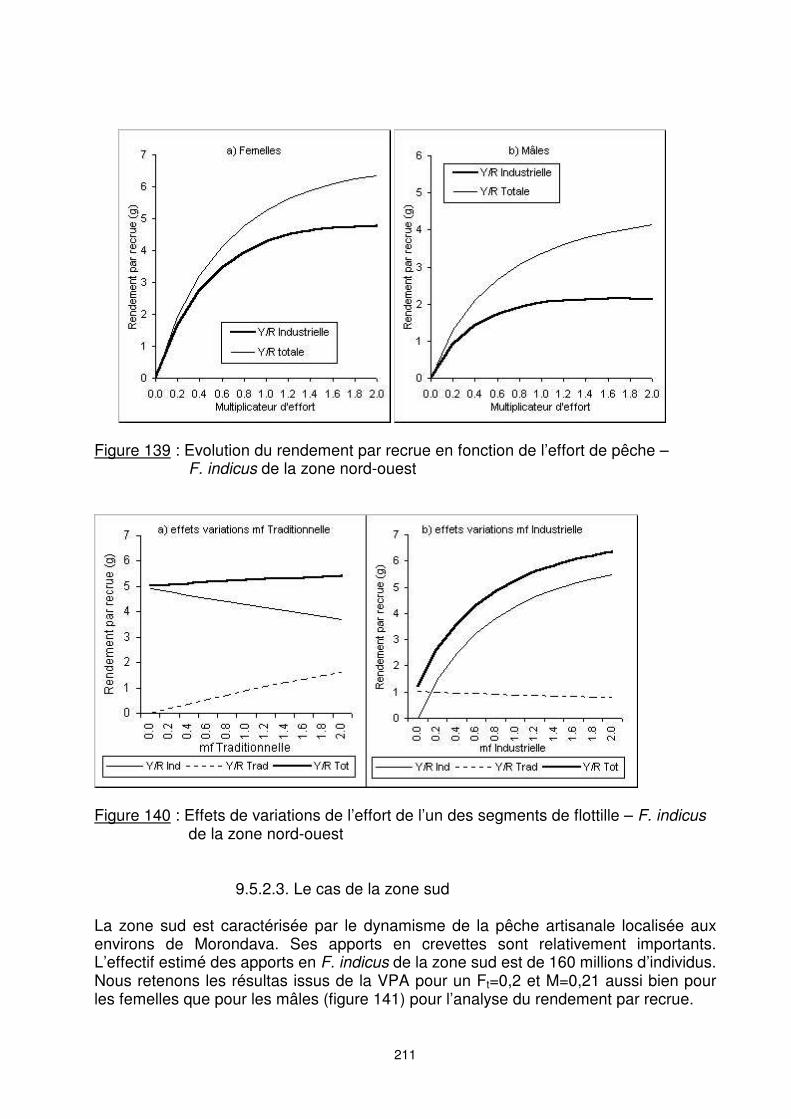
211
Figure 139 : Evolution du rendement par recrue en fonction de l’effort de pêche –
F. indicus de la zone nord-ouest
Figure 140 : Effets de variations de l’effort de l’un des segments de flottille – F. indicus
de la zone nord-ouest
9.5.2.3. Le cas de la zone sud La zone sud est caractérisée par le dynamisme de la pêche artisanale localisée aux environs de Morondava. Ses apports en crevettes sont relativement importants. L’effectif estimé des apports en F. indicus de la zone sud est de 160 millions d’individus. Nous retenons les résultas issus de la VPA pour un Ft=0,2 et M=0,21 aussi bien pour les femelles que pour les mâles (figure 141) pour l’analyse du rendement par recrue.
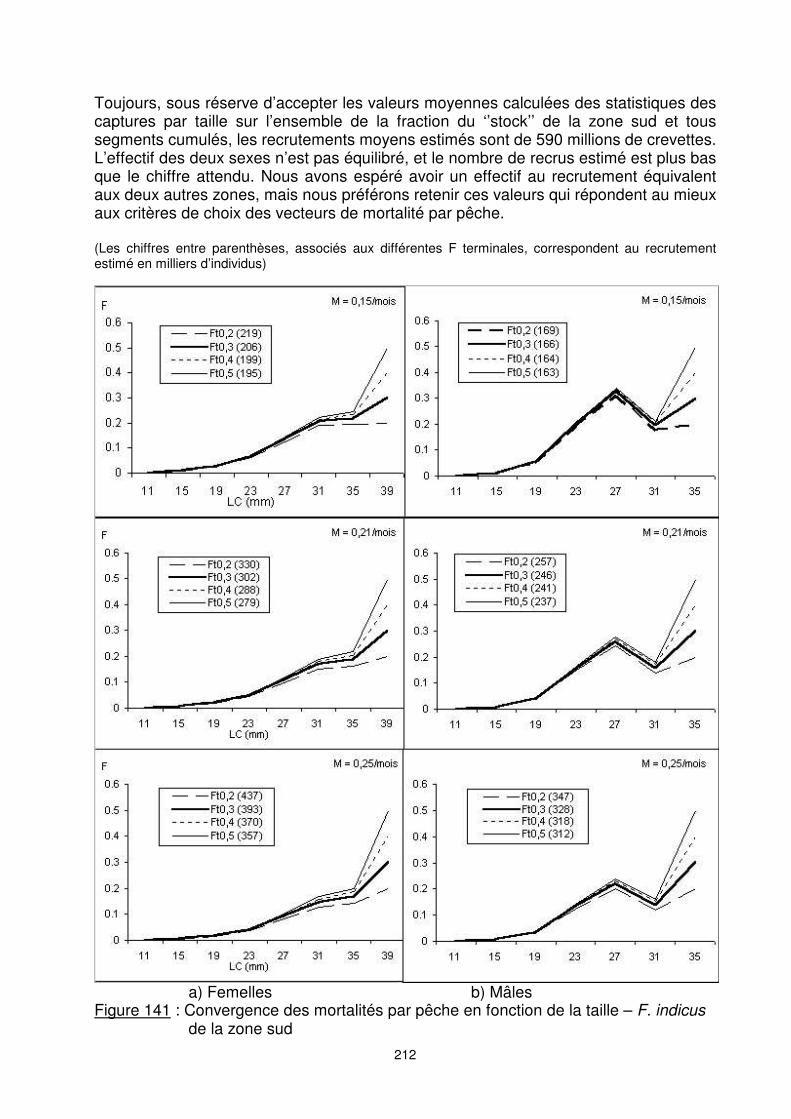
212
Toujours, sous réserve d’accepter les valeurs moyennes calculées des statistiques des captures par taille sur l’ensemble de la fraction du ‘’stock’’ de la zone sud et tous segments cumulés, les recrutements moyens estimés sont de 590 millions de crevettes. L’effectif des deux sexes n’est pas équilibré, et le nombre de recrus estimé est plus bas que le chiffre attendu. Nous avons espéré avoir un effectif au recrutement équivalent aux deux autres zones, mais nous préférons retenir ces valeurs qui répondent au mieux aux critères de choix des vecteurs de mortalité par pêche. (Les chiffres entre parenthèses, associés aux différentes F terminales, correspondent au recrutement estimé en milliers d’individus)
a) Femelles b) Mâles Figure 141 : Convergence des mortalités par pêche en fonction de la taille – F. indicus
de la zone sud
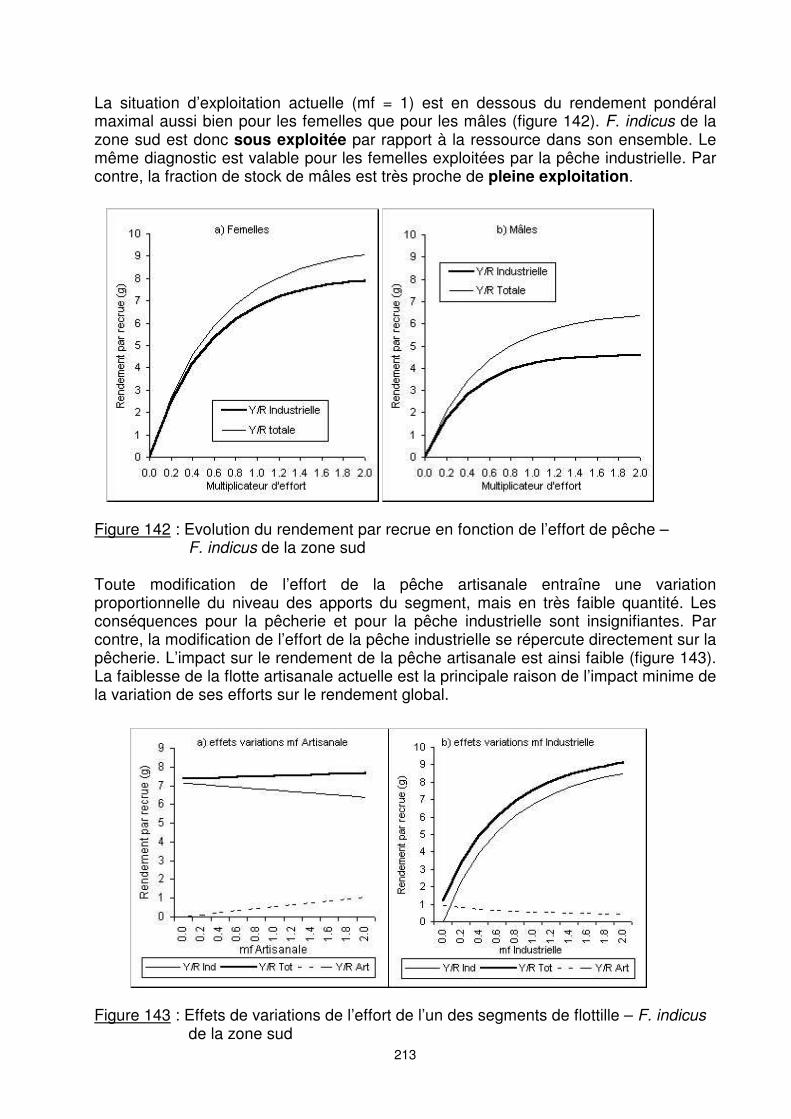
213
La situation d’exploitation actuelle (mf = 1) est en dessous du rendement pondéral maximal aussi bien pour les femelles que pour les mâles (figure 142). F. indicus de la zone sud est donc sous exploitée par rapport à la ressource dans son ensemble. Le même diagnostic est valable pour les femelles exploitées par la pêche industrielle. Par contre, la fraction de stock de mâles est très proche de pleine exploitation.
Figure 142 : Evolution du rendement par recrue en fonction de l’effort de pêche –
F. indicus de la zone sud Toute modification de l’effort de la pêche artisanale entraîne une variation proportionnelle du niveau des apports du segment, mais en très faible quantité. Les conséquences pour la pêcherie et pour la pêche industrielle sont insignifiantes. Par contre, la modification de l’effort de la pêche industrielle se répercute directement sur la pêcherie. L’impact sur le rendement de la pêche artisanale est ainsi faible (figure 143). La faiblesse de la flotte artisanale actuelle est la principale raison de l’impact minime de la variation de ses efforts sur le rendement global.
Figure 143 : Effets de variations de l’effort de l’un des segments de flottille – F. indicus
de la zone sud
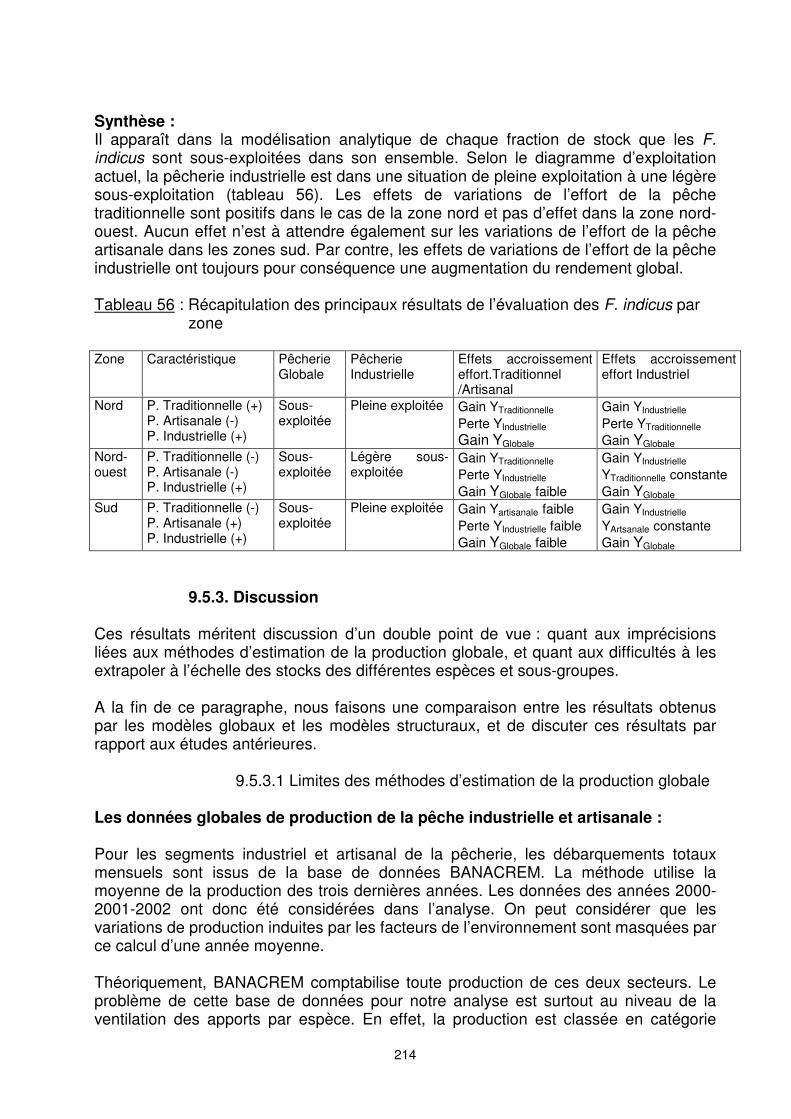
214
Synthèse : Il apparaît dans la modélisation analytique de chaque fraction de stock que les F. indicus sont sous-exploitées dans son ensemble. Selon le diagramme d’exploitation actuel, la pêcherie industrielle est dans une situation de pleine exploitation à une légère sous-exploitation (tableau 56). Les effets de variations de l’effort de la pêche traditionnelle sont positifs dans le cas de la zone nord et pas d’effet dans la zone nord-ouest. Aucun effet n’est à attendre également sur les variations de l’effort de la pêche artisanale dans les zones sud. Par contre, les effets de variations de l’effort de la pêche industrielle ont toujours pour conséquence une augmentation du rendement global. Tableau 56 : Récapitulation des principaux résultats de l’évaluation des F. indicus par
zone Zone Caractéristique Pêcherie
Globale Pêcherie Industrielle
Effets accroissement effort.Traditionnel /Artisanal
Effets accroissement effort Industriel
Nord P. Traditionnelle (+) P. Artisanale (-) P. Industrielle (+)
Sous-exploitée
Pleine exploitée Gain YTraditionnelle Perte YIndustrielle Gain YGlobale
Gain YIndustrielle Perte YTraditionnelle Gain YGlobale
Nord-ouest
P. Traditionnelle (-) P. Artisanale (-) P. Industrielle (+)
Sous-exploitée
Légère sous-exploitée
Gain YTraditionnelle Perte YIndustrielle Gain YGlobale faible
Gain YIndustrielle YTraditionnelle constante Gain YGlobale
Sud P. Traditionnelle (-) P. Artisanale (+) P. Industrielle (+)
Sous-exploitée
Pleine exploitée Gain Yartisanale faible Perte YIndustrielle faible Gain YGlobale faible
Gain YIndustrielle YArtsanale constante Gain YGlobale
9.5.3. Discussion Ces résultats méritent discussion d’un double point de vue : quant aux imprécisions liées aux méthodes d’estimation de la production globale, et quant aux difficultés à les extrapoler à l’échelle des stocks des différentes espèces et sous-groupes. A la fin de ce paragraphe, nous faisons une comparaison entre les résultats obtenus par les modèles globaux et les modèles structuraux, et de discuter ces résultats par rapport aux études antérieures.
9.5.3.1 Limites des méthodes d’estimation de la production globale Les données globales de production de la pêche industrielle et artisanale : Pour les segments industriel et artisanal de la pêcherie, les débarquements totaux mensuels sont issus de la base de données BANACREM. La méthode utilise la moyenne de la production des trois dernières années. Les données des années 2000-2001-2002 ont donc été considérées dans l’analyse. On peut considérer que les variations de production induites par les facteurs de l’environnement sont masquées par ce calcul d’une année moyenne. Théoriquement, BANACREM comptabilise toute production de ces deux secteurs. Le problème de cette base de données pour notre analyse est surtout au niveau de la ventilation des apports par espèce. En effet, la production est classée en catégorie

215
‘’White’’, ‘’Autres’’ et ‘’Mélange’’. Cette classification n’est pas homogène pour chaque armement. Quelquefois, la catégorie ‘’White’’ est assimilée aux espèces de ‘’crevettes’’, dont F. indicus, M. monoceros et P. semisulcatus, et la catégorie ‘’Autres’’ assimilée au ‘’Camaron’’ ; l’espèce P. monodon. Généralement, la production de la pêche artisanale est classée dans la catégorie ‘’Mélange’’. La ventilation du débarquement par mois et par grande zone ne pose pas problème. Il faut signaler également le cas de rejet de petites crevettes, mentionné dans les fiches des observateurs embarqués à bord des chalutiers commerciaux, mais cette quantité est minime. Deux autres problèmes sont également signalés sur le débarquement total effectif de crevettes : celui de détournement de crevettes évalué à 800 tonnes par an (site Web www.madagascar-contacts/GAPCM) et le débarquement de 7 chalutiers ciblant les poissons depuis l’année 2002. Cette flottille exploite la ressource poissonnière démersale, mais elle opère parfois sur des fonds à crevettes et est donc susceptible d’effectuer des captures (Sandon et al., sous presse). Les données globales de production du segment de la pêche traditionnelle : Les données considérées sont celles de l’enquête sur les captures et les efforts réalisée par le PNRC en 1998 et 1999 et extrapolées sur le recensement actualisé de l’année 1998. Le suivi de la production est réalisé dans les villages de pêche les plus actifs et l’extrapolation sur l’ensemble de la région pourrait être sur-estimée. Par ailleurs, les effectifs de pêche des villages les plus reculés n’ont pas été mis à jour ; ce qui pourrait sous estimer la production totale, car les crevettes autoconsommées sont ignorées. Il semble très difficile d’évaluer la qualité des estimations jusqu’à ce jour. L’utilisation des données de collecte pourrait remédier ce problème, mais celles-ci enregistrent seulement les crevettes de taille marchande. Les estimations de l’autoconsommation et de la commercialisation des crevettes de petite taille réalisées en 1989 (Rafalimanana, 1990) ne sont pas généralisables sur l’ensemble de la côte ouest et méritent une mise à jour. Toutefois, cette valeur de l’estimation de la production de la pêche traditionnelle a un impact capital surtout pour l’évaluation de la fraction de ‘’stock’’ de la zone nord où la production estimée est la plus importante.
9.5.3.2. Portées des extrapolations sur la composition spécifique, la composition en taille et le sex-ratio
Le problème de ventilation par espèce pour les pêches industrielle et artisanale est évoqué précédemment. Nous avons utilisé les échantillons prélevés par les observateurs à bord des chalutiers commerciaux pour estimer le rapport M. monoceros/P. semisulcatus dans les captures, ainsi que la composition en taille par espèce-bimestre. La répartition des captures par classe de taille dans les échantillons traités est extrapolée à l’ensemble de la production industrielle et artisanale. Les taux d’extrapolation sont faibles dans son ensemble et variables selon les zones-mois. Nous ne pouvons pas confronter nos résultats avec les données ‘’processing’’ qui a l’avantage de fournir les informations réelles de débarquement. Cette méthode est utilisée par Dintheer et al. (1998) et Moguedet et al. (2001) pour l’évaluation de cette pêcherie pour les années 1994 et 1998 respectivement. Sandon et al. (sous presse) utilisent les données de traitement à bord d’une société qui possède des navires actifs dans les trois zones (licence type A, B et C).

216
Les sex-ratios utilisés sont ceux issus d’une estimation par analyse de variance des données observateurs. Nous considérons que ces valeurs sont fiables car les facteurs années, mois et zone ont été pris en compte dans les modèles. Seulement, la généralisation pour l’ensemble de la pêcherie pourrait engendrer des biais car seules les données de la pêche chalutière ont été modélisées. Toutefois, la comparaison de ces estimations standardisées avec les données brutes des échantillons de la pêche traditionnelle ne montre pas de différence significative, et nous préférons utiliser l’estimation par analyse de variance. L’échantillon utilisé pour la pêche traditionnelle, répartie sur les principaux sites de débarquement de la côte ouest, présente l’avantage de tenir compte des tailles réelles des captures au débarquement des pirogues et non après tri de la part des collecteurs, contrairement aux données de collecte. Toutefois, à défaut des données récentes, nous avons considéré que ces données des années 1998/1999 restent encore valables sous l’hypothèse que le diagramme d’exploitation ne change pas beaucoup pour la période considérée.
Toutes ces hypothèses de travail impliquent que des données perfectibles ont été introduites dans les analyses, ce qui impose de prendre quelques précautions concernant le diagnostic obtenu. Au-delà de ces considérations sur les données extrapolées, la sélectivité des engins n’a pas été prise en compte dans les analyses de l’impact de la pêche sur la dynamique des populations. Ces facteurs n’apparaient pas possibles dans l’état actuel des connaissances. Une étude sur la sélectivité des ‘’Valakira’’ est en cours (Mémoire DEA, Université de Toliara). Celle des autres engins utilisés par la pêche traditionnelle vient d’être finalisée et présentée au cours de l’atelier de juin 2003 (De Rodellec et Rasoanandrasana, sous presse). L’intégration de ces paramètres dans l’analyse de la population pourrait affiner les résultats et permettre de faire des simulations à moyen terme (changement de maillage et modification des durées de fermeture).
9.5.3.3. Comparaison des résultats obtenus par les modèles globaux
et les modèles structuraux Les résultats des évaluations de stocks obtenus par les modèles globaux et les modèles structuraux sont liés au niveau de la représentativité des données. En reprenant l’exemple des évaluations à l’échelle globale, on notera que le diagnostic de pleine exploitation (optimiste) par les modèles globaux est obtenu, alors que le diagnostic d’une legère sous-exploitation est déduit des modèles structuraux. La différence mérite précisions et quelques commentaires. Dans le cas des modèles globaux, l’évaluation ne porte que sur une partie du stock. Ici, on considère seulement l’exploitation industrielle ; les activités de pêches artisanale et traditionnelle ne sont pas prises en compte. Par ailleurs, les modèles sont appliqués à la capture totale de l’ensemble des espèces. Ainsi, les aspects sur les interactions biologiques et techniques sont ignorés. Aussi, le résultat de l’évaluation est influencé directement sur l’état de la pêcherie de la zone sud, qui représente presque le double du débarquempent mais dont l’effort de pêche n’est pas très intense par rapport aux zones nord.

217
Les résultats par analyses structurales conduites pour chaque espèce sont obtenus à partir des données de captures et d’effort total de chaque type de pêche. Un diagnostic d’une légère sous-exploitation aussi bien pour les F. indicus (segment industriel) que pour les M. monoceros est déduit de l’analyse. La différence entre ces diagnostics et ceux de l’approche globale est vraisemblablement liée à la mauvaise connaissance de la structure de taille des captures de chacune des composantes de la pêcherie. Les résultats de Moguedet et al. (2001) pour l’année 1998 sont les mêmes que la présente évaluation en ce qui concerne les M. monoceros. Par ailleurs, ces auteurs montrent que le stock des F. indicus sur la côte ouest est pleinement exploité, le même résultat est obtenu par Sandon et al. (2003, sous presse) à partir des données de débarquement 2002. Ces résultats sont légèrement differents des ceux issus de la présente étude. Il faut noter que les évaluations réalisées par ces auteurs utilisent les paramètres biologiques de Le Reste et Marcille (1976) et de Coss et al. (1993) pour K et L∞ et ceux de Le Reste et al. (1974) pour les coefficients de relation taille-poids, et donc les résultats ne sont pas directement comparables avec la présente étude. Enfin, il faut remarquer que l’extrapolation utilisée pour la reconstitution des structures démographiques des captures lors de la présente étude est assez théorique par rapport aux deux autres études antérieures. Ces résultats sont donc à considérer avec prudence tant que les structures démographiques des apports extrapolées n’auront pas été validées.

218
CONCLUSION DE LA TROISIEME PARTIE
A partir des modèles de production, on a pu mettre en évidence à l’échelle globale ou à l’échelle des grandes zones géographiques pour le segment industriel que les stocks de crevettes pénéides (toutes espèces confondues) de la côte ouest malgache est actuellement en situation de pleine exploitation à une sur-exploitation selon les hypothèses d’accroissement des puissances de pêche. De l’autre côté avec les modèles stucturaux, en analysant les stocks par espèce-sexe et en tenant compte de l’ensemble des segments d’exploitation (industriel, artisanal et traditionnel), on observe une situation de pleine exploitation à une légère sous-exploitation. Les deux méthodes convergent donc vers un diagnostic de pleine exploitation de la ressource. Chacune des deux méthodes utilisées a ses avantages et ses limites. On a pu estimer des valeurs assez stables et statistiquement acceptables du MSY à partir des modèles globaux. D’autre part, les niveaux de l’effort de pêche de maximisation des captures sont également donnés. Ces modèles restent donc un très bon outil, simple et robuste. Mais les analyses effectuées constituent une approche assez rudimentaire : prise en compte seulement de la phase marine de la ressource et non prise en compte de la spécificité des espèces-sexes. Par contre, les modèles structuraux ont l’avantage de lever ces contraintes, mais la principale contrainte se situe au niveau de la qualité des données, notamment les données pour la reconstitution des structures démographiques. Au travers de ce travail, les paramètres estimés dans la deuxième partie de la thèse ont été utilisés. Nous n’avons pas de problèmes majeurs pour leur utilisation et les résultats obtenus sont cohérents.





























