Embed Size (px)
Citation preview

235
I. IntroductIon
L’immunité recouvre les mécanismes de défense d’un organisme vis-à-vis d’une agression, qu’elle provienne de l’environnement (les micro-organismes, par exemple) ou d’une modifi -cation de ses propres constituants (les cellules cancéreuses, par exemple). D’une façon plus générale, l’immunité biologique correspond à l’ensemble des mécanismes développés par un organisme pour se maintenir en bonne santé, immun dans le sens sain.
Le concept de l’immunisation naît avec Jenner en 1756. Des vachères avaient des pustules ressemblant à celles de la variole sur les mains, et lors d’épidémie à la variole, elles étaient préservées, elles étaient immunisées. Puis, en 1890, Pasteur met en évidence l’existence des microbes, et notamment des bactéries. Il cultive des cellules de chiens enragés, puis les chauffe et les injecte à un berger mordu. Celui-ci en sort indemne. Il s’agit du vaccin contre la rage.
L’immunité de l’organisme repose en fait sur sa capacité à reconnaître et à éliminer le non soi, via les réponses immunitaires, tout en respectant le soi.
A. NotiondusoiLe soi d’un individu est défi ni par l’ensemble des molécules synthétisées par son organisme qui résultent de l’expression de son génome. Les molécules du soi sont tolérées par le système immunitaire, c’est-à-dire qu’elles ne déclenchent pas de mécanismes de défense.
Certaines de ces molécules ont un caractère propre à chaque individu et constituent la carte d’identité moléculaire de celui-ci. Il s’agit des molécules du Complexe Majeur d’Histocompa-tibilité (CMH) ou du système HLA pour « Human Leucocyte Antigen », et des marqueurs de surface des hématies dont font partie le système ABO et le système Rhésus.
1. LesmoléculesdusystèmeHLA:«HumanLeucocyteAntigen»Ce sont les marqueurs majeurs du soi biologique ; on parle d’antigènes majeurs. Il s’agit de glycoprotéines membranaires spécifi ques d’un individu. Chez l’Homme, elles sont codées par un ensemble de 6 gènes liés, très polymorphes, portés par la paire de chromosome 6 : les gènes A, B, C, DP, DQ et DR. Cette partie du génome constitue le complexe majeur d’his-tocompatibilité : le CMH. Pour chacun des gènes du CMH, il existe un grand nombre d’allèles tous co-dominants. Ceci étant à l’origine d’un nombre de combinaisons extrêmement élevé. En effet, la probabilité que deux personnes non apparentées possèdent les mêmes mar-queurs HLA est quasi nulle. En revanche, cette probabilité s’élève chez les parents proches, car tous les allèles portés par un chromosome sont transmis en bloc, appelé haplotype, comme s’il s’agissait d’un seul gène.
Ces marqueurs sont également nommés antigènes de transplantation, puisque ce sont eux qui sont impliqués dans l’acceptation ou le rejet des greffes. Ils commandent donc la compatibilité ou l’incompatibilité entre deux tissus lors d’une greffe (voir Transplantation d’organe, p. 263). La grande diversité de ces marqueurs cellulaires explique les précautions à prendre lors de transplantations d’organes ou de greffes de tissus. Il faut donc déterminer
13 Immunologie
BAH2_ch13.indd 235 28/02/2014 11:29

236
Anatomie – physiologie
les marqueurs cellulaires du donneur et du receveur afi n d’établir le niveau de compatibilité entre les deux.
On distingue deux groupes de molécules HLA, on parle de classe :
¾¾ Les molécules HLA de classe I : elles sont présentes à la surface de toutes les cellules nucléées de l’organisme. Elles sont codées par les gènes A, B et C de classe I ;
¾¾ Les molécules HLA de classe II : leur présence est strictement limitée à la membrane des cellules immunitaires. Elles sont codées par les gènes DP, DQ et DR de classe II.
On trouve donc 12 molécules HLA différentes (codées par 6 allèles, ou un haplotype, d’ori-gine paternelle, et par 6 allèles, ou un haplotype, d’origine maternelle) à la surface des cel-lules exprimant à la fois les CMH de classe I et II.
Les molécules HLA interviennent dans les réactions immunitaires spécifi ques en s’associant à des fragments de molécules étrangères à l’organisme (voir Les TCR, p. 245).
2. LesmarqueursdesurfacedeshématiesLes hématies étant des cellules anucléées, elles ne présentent pas à leur surface de molé-cules du système HLA. Leur membrane présente en fait des marqueurs, dit mineurs (en opposition au terme « majeur » du CMH), dont le type est génétiquement déterminé. Ces marqueurs déterminent les groupes sanguins. Il s’agit du système ABO et du système Rh.
� Le système ABOCes marqueurs sont des oligosaccharides reliés à une protéine membranaire de l’hématie. C’est leur partie terminale qui diffère d’un marqueur à l’autre, et qui détermine les groupes sanguins. La mise en place de la partie terminale du marqueur est entreprise par une enzyme spécifi que codée par un gène situé sur le chromosome 9 possédant 3 allèles. Deux d’entre eux, les allèles A et B, co-dominent sur le troisième, l’allèle O. De ce fait, il existe donc 4 groupes sanguins : A, B, AB et O, défi nis par la présence ou l’absence des marqueurs à la surface des globules rouges, mais également par la présence d’anticorps (protéine du système immunitaire détectant et neutralisant le non soi de manière spécifi que) dans le plasma dirigés contre les marqueurs absents. Le tableau suivant illustre la situation pour les 4 groupes sanguins, ainsi que les compatibilités lors des transfusions sanguines :
Groupe sanguin Marqueur Anticorps À qui donner ? De qui recevoir ?
A A Anti-B A et AB A et O
B B Anti-A B et AB B et O
AB A et B Aucun AB A, B, AB et O(receveur universel)
O Aucun Anti A et Anti B A, B, AB et O(donneur universel) O
Ces marqueurs sont également appelés « agglutinogènes », car lors d’un contact entre un marqueur et l’anticorps dirigé contre lui (également nommé agglutinine), il y a agglutination (agrégation, réunion en amas des anticorps et des marqueurs) entraînant la lyse massive des globules rouges.
La Figure 1 illustre la comptabilité et l’incompatibilité entre des groupes sanguins avec deux exemples.
BAH2_ch13.indd 236 28/02/2014 11:29

237
Immunologie
marqueur ou agglutinogène A
marqueur ou agglutinogène B
Receveur :individu de groupe sanguin AB
Donneur :individu de groupe sanguin O
hématie hématie
Receveur :individu de groupe sanguin A
transfusion sanguine compatible
transfusion sanguine incompatible car :
Donneur :individu de groupe sanguin B
hématie hématie
hématie agglutination des hématies du donneur⇒ lyse cellulaire
anticorpsou agglutinine anti-B
+
Figure 1 Transfusions sanguines compatibles et incompatibles
� Le système Rhésus ou RhLe système Rhésus est également un système mineur du soi biologique, il se superpose au système ABO. Les sujets porteurs d’un autre marqueur du soi, appelé marqueur D, sont dits Rh positif. Les sujets ne possédant pas ce marqueur sont dits Rh négatif. L’allèle Rh+ domine sur l’allèle Rh–. Les compatibilités entre Rh+ et Rh– reposent sur les mêmes principes que le système ABO.
B. NotiondunonsoiLe non soi est défi ni par l’ensemble des molécules étrangères au soi, c’est-à-dire non codées par notre génome. Ce sont ces éléments du non soi qui déclenchent les réactions immuni-taires.
Les molécules du non soi proviennent :
¾¾ soit de l’environnement, du milieu extérieur : elles sont dans ce cas, portées par les membranes de cellules étrangères à notre organisme (bactéries, parasites, et champi-gnons, par exemple) ou libres et solubles (les toxines, par exemple) ;
¾¾ soit d’une modifi cation du soi, dans ce cas on parle du soi modifi é : il s’agit des cellules cancéreuses ou infectées par un virus.
BAH2_ch13.indd 237 28/02/2014 11:29
238
Anatomie – physiologie
II.orGanesetceLLuLesImmunItaIres
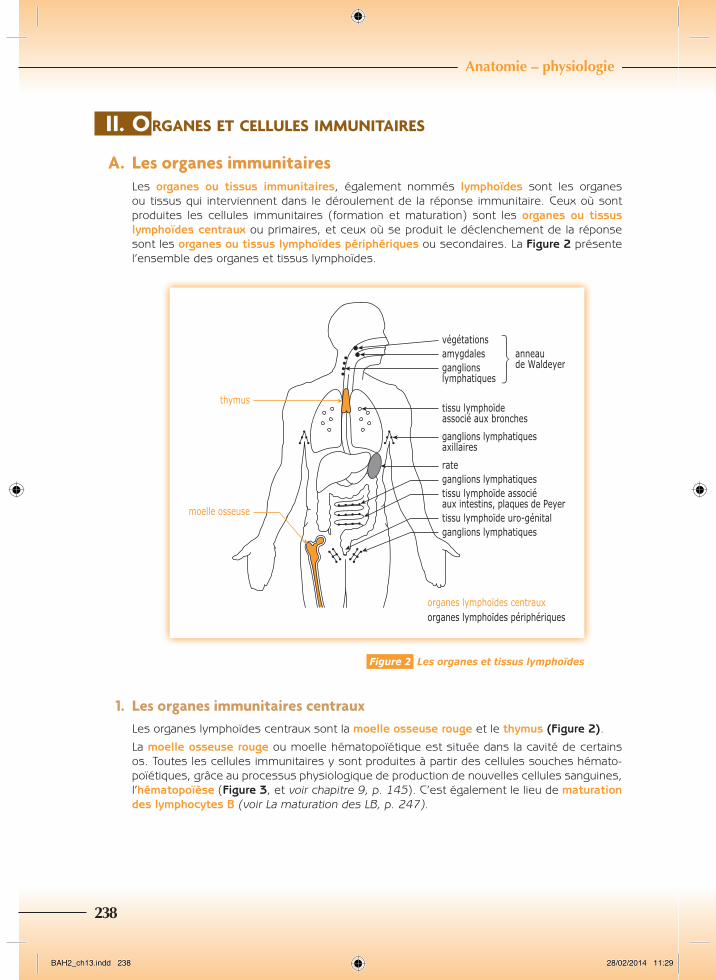
A. LesorganesimmunitairesLes organes ou tissus immunitaires, également nommés lymphoïdes sont les organes ou tissus qui interviennent dans le déroulement de la réponse immunitaire. Ceux où sont produites les cellules immunitaires (formation et maturation) sont les organes ou tissus lymphoïdes centraux ou primaires, et ceux où se produit le déclenchement de la réponse sont les organes ou tissus lymphoïdes périphériques ou secondaires. La Figure 2 présente l’ensemble des organes et tissus lymphoïdes.
thymus
moelle osseuse
végétationsamygdalesganglionslymphatiques
tissu lymphoïdeassocié aux bronches
ganglions lymphatiquesaxillaires
rateganglions lymphatiquestissu lymphoïde associéaux intestins, plaques de Peyertissu lymphoïde uro-génitalganglions lymphatiques
organes lymphoïdes centrauxorganes lymphoïdes périphériques
anneaude Waldeyer
Figure 2 Les organes et tissus lymphoïdes
1. LesorganesimmunitairescentrauxLes organes lymphoïdes centraux sont la moelle osseuse rouge et le thymus (Figure 2).
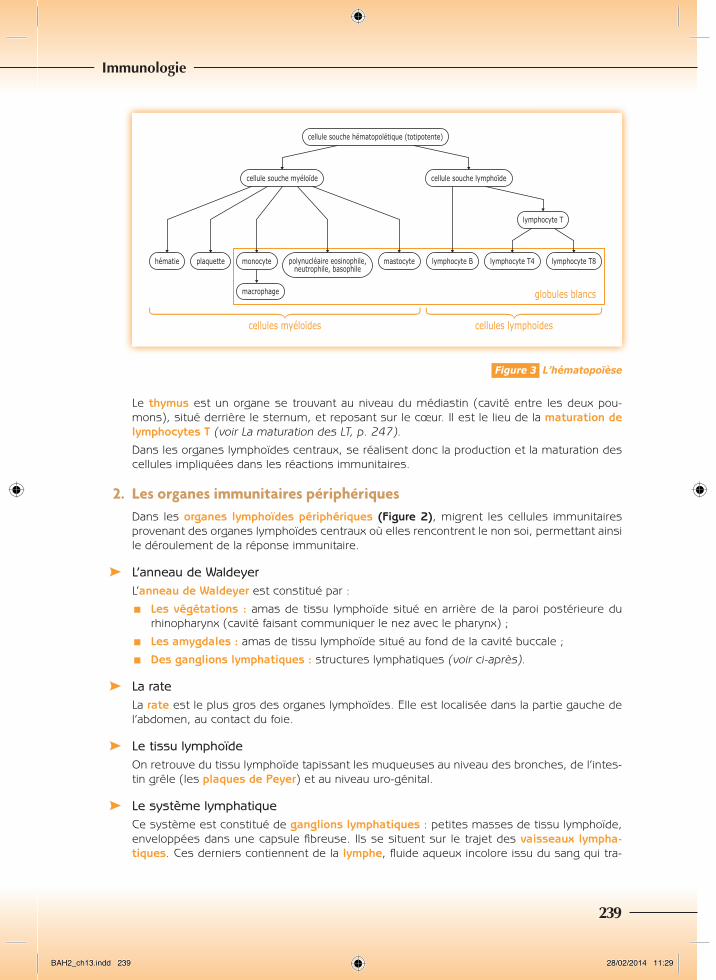
La moelle osseuse rouge ou moelle hématopoïétique est située dans la cavité de certains os. Toutes les cellules immunitaires y sont produites à partir des cellules souches hémato-poïétiques, grâce au processus physiologique de production de nouvelles cellules sanguines, l’hématopoïèse (Figure 3, et voir chapitre 9, p. 145). C’est également le lieu de maturation des lymphocytes B (voir La maturation des LB, p. 247).
BAH2_ch13.indd 238 28/02/2014 11:29
239
Immunologie
cellule souche hématopoïétique (totipotente)
globules blancs
cellules myéloïdes cellules lymphoïdes
cellule souche myéloïde cellule souche lymphoïde
lymphocyte T8lymphocyte T4lymphocyte Bmastocytepolynucléaire eosinophile,neutrophile, basophile
monocyte
macrophage
plaquettehématie
lymphocyte T
Figure 3 L’hématopoïèse
Le thymus est un organe se trouvant au niveau du médiastin (cavité entre les deux pou-mons), situé derrière le sternum, et reposant sur le cœur. Il est le lieu de la maturation de lymphocytes T (voir La maturation des LT, p. 247).
Dans les organes lymphoïdes centraux, se réalisent donc la production et la maturation des cellules impliquées dans les réactions immunitaires.
2. LesorganesimmunitairespériphériquesDans les organes lymphoïdes périphériques (Figure 2), migrent les cellules immunitaires provenant des organes lymphoïdes centraux où elles rencontrent le non soi, permettant ainsi le déroulement de la réponse immunitaire.
� L’anneau de WaldeyerL’anneau de Waldeyer est constitué par :
¾¾ Les végétations : amas de tissu lymphoïde situé en arrière de la paroi postérieure du rhinopharynx (cavité faisant communiquer le nez avec le pharynx) ;
¾¾ Les amygdales : amas de tissu lymphoïde situé au fond de la cavité buccale ;
¾¾ Des ganglions lymphatiques : structures lymphatiques (voir ci-après).
� La rateLa rate est le plus gros des organes lymphoïdes. Elle est localisée dans la partie gauche de l’abdomen, au contact du foie.
� Le tissu lymphoïdeOn retrouve du tissu lymphoïde tapissant les muqueuses au niveau des bronches, de l’intes-tin grêle (les plaques de Peyer) et au niveau uro-génital.
� Le système lymphatiqueCe système est constitué de ganglions lymphatiques : petites masses de tissu lymphoïde, enveloppées dans une capsule fi breuse. Ils se situent sur le trajet des vaisseaux lympha-tiques. Ces derniers contiennent de la lymphe, fl uide aqueux incolore issu du sang qui tra-
BAH2_ch13.indd 239 28/02/2014 11:29
240
Anatomie – physiologie
verse les parois des capillaires sanguins (voir chapitre 10, p. 167). La lymphe passe d’abord dans les espaces intracellulaires des tissus, puis est récupérée par un réseau de vaisseaux lymphatiques parsemé de ganglions.
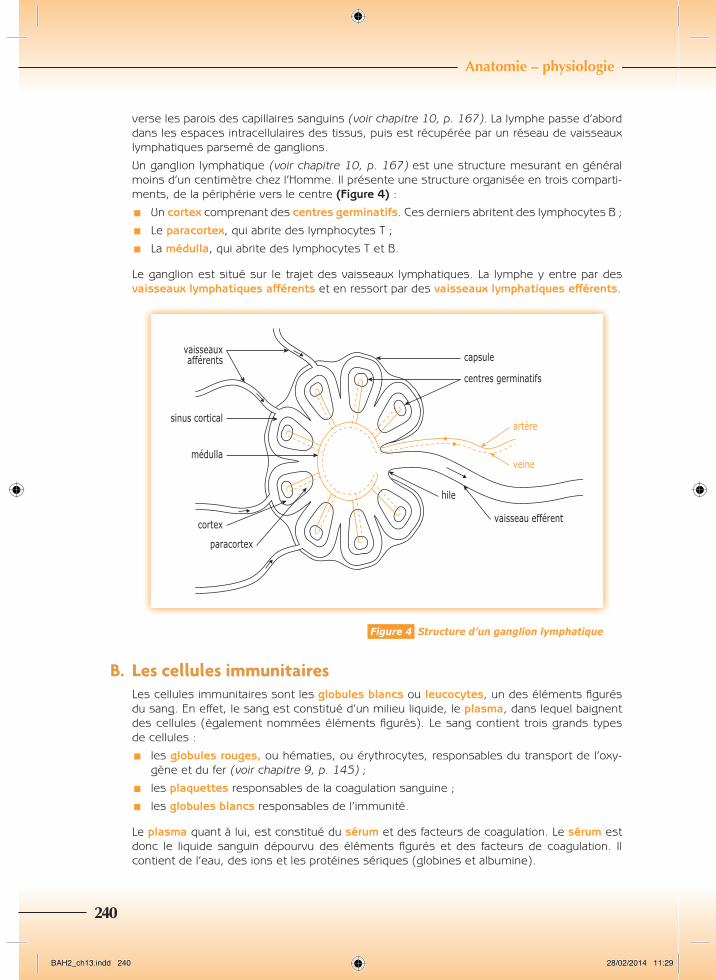
Un ganglion lymphatique (voir chapitre 10, p. 167) est une structure mesurant en général moins d’un centimètre chez l’Homme. Il présente une structure organisée en trois comparti-ments, de la périphérie vers le centre (Figure 4) :
¾¾ Un cortex comprenant des centres germinatifs. Ces derniers abritent des lymphocytes B ;
¾¾ Le paracortex, qui abrite des lymphocytes T ;
¾¾ La médulla, qui abrite des lymphocytes T et B.
Le ganglion est situé sur le trajet des vaisseaux lymphatiques. La lymphe y entre par des vaisseaux lymphatiques afférents et en ressort par des vaisseaux lymphatiques efférents.
vaisseauxafférents capsule
sinus cortical
centres germinatifs
médulla
artère
cortex
veine
paracortex
hile
vaisseau efférent
Figure 4 Structure d’un ganglion lymphatique
B. LescellulesimmunitairesLes cellules immunitaires sont les globules blancs ou leucocytes, un des éléments fi gurés du sang. En effet, le sang est constitué d’un milieu liquide, le plasma, dans lequel baignent des cellules (également nommées éléments fi gurés). Le sang contient trois grands types de cellules :
¾¾ les globules rouges, ou hématies, ou érythrocytes, responsables du transport de l’oxy-gène et du fer (voir chapitre 9, p. 145) ;
¾¾ les plaquettes responsables de la coagulation sanguine ;
¾¾ les globules blancs responsables de l’immunité.
Le plasma quant à lui, est constitué du sérum et des facteurs de coagulation. Le sérum est donc le liquide sanguin dépourvu des éléments fi gurés et des facteurs de coagulation. Il contient de l’eau, des ions et les protéines sériques (globines et albumine).
BAH2_ch13.indd 240 28/02/2014 11:29
241
Immunologie
Les globules blancs sont des cellules nucléées. Elles peuvent se déplacer à contre-courant, franchir la paroi des vaisseaux sanguins (processus de la diapédèse) et pénétrer dans les tissus. Le nombre de globules blancs est d’environ 7 000 par mm3 de sang.
On distingue les leucocytes en 2 lignées cellulaires (Figure 3) :
¾¾ Les cellules myéloïdes : les leucocytes polynucléaires ou granulocytes (neutrophiles, basophiles, éosinophiles), mastocytes, monocytes et macrophages ;
¾¾ Les cellules lymphoïdes : lymphocytes B et T.
Remarque : les globules blancs peuvent également être séparés en 2 familles selon leur aspect cellulaire, et plus particulièrement selon l’aspect de leur noyau. Les leucocytes mono-nucléaires (un noyau ; regroupant les lymphocytes, mastocytes, monocytes et macro-phages) sont à distinguer des polynucléaires (Pris au départ pour plusieurs noyaux. En fait, un seul noyau multilobé. Il s’agit des granulocytes). Les mononucléaires sont également appelés agranulocytes en opposition à la dénomination « granulocytes » des polynucléaires.
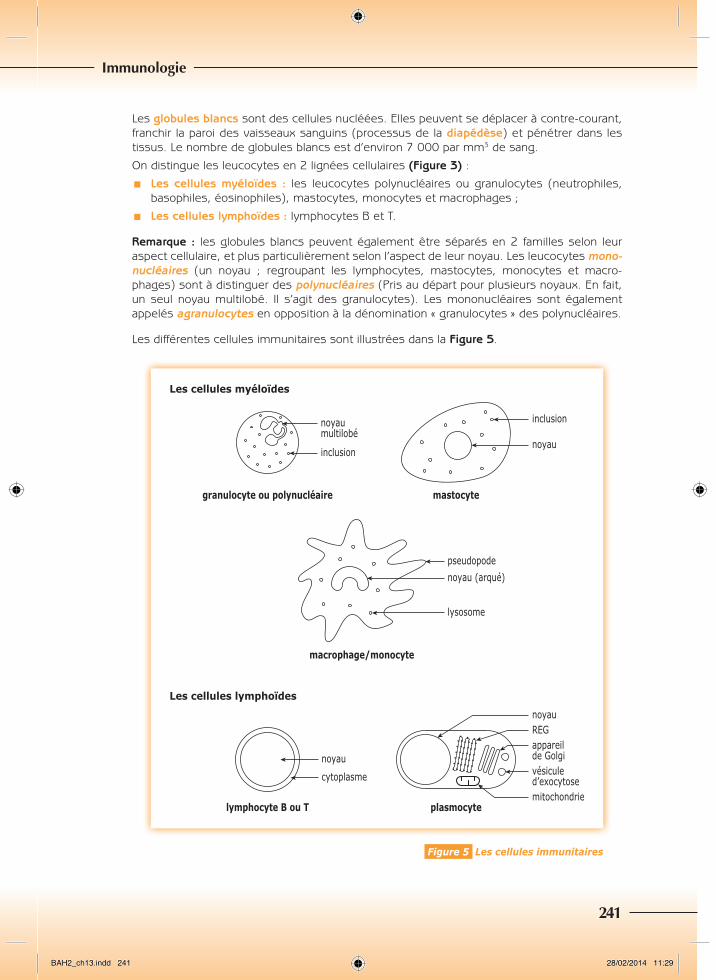
Les différentes cellules immunitaires sont illustrées dans la Figure 5.
Les cellules myéloïdes
Les cellules lymphoïdes
noyau
granulocyte ou polynucléaire mastocyte
inclusion
inclusion
noyaumultilobé
macrophage/monocyte
lysosome
pseudopode
noyau (arqué)
lymphocyte B ou T plasmocyte
vésiculed’exocytosemitochondrie
noyauREGappareilde Golgi
cytoplasme
noyau
Figure 5 Les cellules immunitaires
BAH2_ch13.indd 241 28/02/2014 11:29

242
Anatomie – physiologie
1. Lescellulesdelalignéemyéloïde
� Les granulocytes ou polynucléairesLes granulocytes ou polynucléaires représentent 65 à 70 % des leucocytes. Ce sont des cellules dont le diamètre est compris entre 9 et 12 µm. Ils ont un noyau multilobé et un cytoplasme granuleux, riche en inclusions (Figure 5). Leur durée de vie est courte, environ deux jours. On distingue trois types de granulocytes :
¾¾ Les granulocytes éosinophiles : ils sont impliqués dans la lutte contre les parasites et se trouvent essentiellement dans les muqueuses ;
¾¾ Les granulocytes basophiles : ils sont impliqués dans la réaction inflammatoire et les phénomènes allergiques (voir La réaction inflammatoire, p. 248 et Les allergies, p. 255, respectivement). On les retrouve essentiellement dans le sang et les muqueuses ;
¾¾ Les granulocytes neutrophiles sont des phagocytes (voir La phagocytose, p. 249).
La différence de dénomination est basée sur leur affinité à absorber des colorants cytolo-giques à base d’éosine, neutres ou basiques. Cette différence repose en fait, sur le contenu de leurs inclusions.
� Les mastocytesLes mastocytes sont des cellules arrondies, à noyau rond central, dont le cytoplasme est granuleux, car, riche en inclusions (Figure 5). Leur diamètre est compris entre 10 et 15 µm. Leur rôle est semblable à celui des granulocytes basophiles, mais contrairement à ces der-niers, ils ne s’observent que dans les tissus.
� Les monocytes et macrophagesLes macrophages dérivent des monocytes (Figure 3). Ces cellules sont caractérisées par leur grande taille, leur diamètre est compris entre 15 et 20 µm. Elles possèdent un noyau arqué (en forme de rein, réniforme, ou de « fer à cheval »), et de nombreux lysosomes, riches en enzymes permettant la digestion d’éléments du non soi (Figure 5). Lorsqu’elles sont activées par la présence d’un élément du non soi, elles développent des prolongements cytoplasmiques, les pseudopodes, leur permettant de « capturer » les éléments du non soi. Comme les granulocytes neutrophiles, ces cellules sont des phagocytes. Elles sont égale-ment des cellules présentatrices d’antigène, des CPA, c’est-à-dire qu’après avoir dégradé l’élément du non soi, elles présentent à leur surface des fragments de celui-ci, qui seront reconnus par les cellules immunitaires intervenant dans les réponses immunitaires spéci-fiques (voir L’immunité spécifique, p. 251). Elles ont une durée de vie relativement longue, pouvant aller jusqu’à plusieurs mois, voire plusieurs années.
2. LescellulesdelalignéelymphoïdeIl s’agit des lymphocytes (Figure 3). Ces cellules sont de petite taille, leur diamètre est com-pris entre 6 et 15 µm. Elles possèdent un noyau arrondi et volumineux entouré d’un fin film cytoplasmique (Figure 5). Les lymphocytes sont présents dans le sang, la lymphe, et tous les tissus et organes lymphoïdes. Ils représentent 25 à 35 % des leucocytes. On distingue 2 grands types de lymphocytes qui diffèrent par leurs marqueurs membranaires :
¾¾ Les lymphocytes T (LT) : ils naissent dans la moelle osseuse, et maturent (c’est-à-dire, deviennent capable de reconnaître les molécules du soi et du non soi) dans le thymus (d’où T comme Thymus). On en distingue 2 types : les lymphocytes T4 (LT4) et les lymphocytes T8 (LT8). Les LT4 participent aux réactions immunitaires spécifiques en tant qu’auxiliaires (LTa) ou helpers (LTh). Ils sont indispensables pour induire les réactions immunitaires spécifiques (rôle pivot dans la réponse immunitaire spécifique), mais n’ont
BAH2_ch13.indd 242 28/02/2014 11:29

243
Immunologie
pas de rôle dans la destruction du non soi. Ils portent à leur surface des récepteurs membranaires T (« T Cell Receptor »), et des marqueurs membranaires CD4. Les LT8 en se différenciant donnent des LT cytotoxiques (LTc). Ce sont les effecteurs de la réaction immunitaire spécifi que à médiation cellulaire (voir La réponse immunitaire à médiation cellulaire, p. 253). Ils portent à leur surface des récepteurs T et des marqueurs membra-naires CD8. Ils permettent la destruction, la cytolyse, des cellules infectées, des cellules cancéreuses ou des cellules greffées étrangères. Ceci, car ils possèdent dans leur cyto-plasme des vésicules remplies de perforine, une enzyme lytique de nature protéique ;
¾¾ Les lymphocytes B (LB) : ces cellules naissent et maturent dans la moelle osseuse rouge (d’où B comme « Bone marrow » = moelle osseuse). Ils possèdent à leur surface des anticorps membranaires (voir Les anticorps, ci-après). En se différenciant, ils donnent les plasmocytes (Figure 5), cellules capables de synthétiser et de sécréter des anticorps (voir ci-après). Ce sont les effecteurs de la réaction immunitaire spécifi que à médiation humorale (voir La réponse immunitaire à médiation humorale, p. 252). Les plasmocytes possèdent toutes les structures caractéristiques d’une cellule productrice et sécrétrice de protéines (REG, appareil de Golgi et vésicules de sécrétion/d’exocytose).
C. Lesrécepteursmembranairesetl’immunocompétence
1. LesanticorpsLa capacité qu’ont les LB à reconnaître un élément du non soi est due à la présence de pro-téines membranaires que l’on appelle anticorps membranaires. Il s’agit de protéines qui ont une structure de base comprenant 4 chaînes polypeptidiques : 2 chaînes légères (L) sem-blables, et 2 chaînes lourdes (H) semblables, reliées entre elles par des ponts disulfures. De par l’arrangement dans l’espace de ses chaînes, la molécule a la forme d’un Y, on parle donc de la structure en Y (Figure 6).
chaîne légère (L)chaîne lourde (H)
fragment Fab
fragment Fc
site de fixationà l’antigèneou paratope
site de liaisonet d’activationdu complément
pont disulfure
régions constantes
région incluse dans la membraneplasmique des LB (anticorps membranaire)ou se liant aux récepteurs des phagocytes(anticorps circulant ou immunoglobuline)
régions variables
Figure 6 Structure en Y d’un anticorps
BAH2_ch13.indd 243 28/02/2014 11:29

244
Anatomie – physiologie
Chaque chaîne possède une partie constance et une partie variable. L’ensemble des parties variables d’une chaîne L et de la chaîne H associée forme un fragment Fab. Un anticorps comprend donc 2 fragments F
ab. L’ensemble des parties constantes des chaînes lourdes et
légères forme un fragment Fc. Un anticorps possède donc 1 fragment Fc.
Un anticorps possède 2 sites de liaison avec l’antigène (élément du non soi, voir L’antigène, p. 251) également nommés paratopes, et ces 2 sites sont identiques. Ils permettent la spécifi cité de reconnaissance de l’antigène. Plus précisément, le paratope reconnaît un épitope, c’est-à-dire une petite partie de l’antigène.
Les chaînes lourdes possèdent une région incluse dans la membrane plasmique d’un LB lorsqu’il s’agit d’un anticorps membranaire (récepteurs membranaires des LB), et une région reconnue par des récepteurs membranaires des phagocytes lorsqu’il s’agit d’un anticorps circulant (anticorps produits par les plasmocytes).
La capacité des lymphocytes B à reconnaître les antigènes lors de la réponse immunitaire spécifi que à médiation humorale (voir La réponse immunitaire à médiation humorale, p. 252) est donc due à la présence d’anticorps membranaires à leur surface. Tous les anticorps membranaires qui se trouvent à la surface d’un lymphocyte B (environ 105) sont identiques, c’est-à-dire qu’ils reconnaissent tous le même antigène. On estime qu’il existe dans l’orga-nisme 1012 lymphocytes B qui se répartissent en 107 clones différant les uns des autres par leurs récepteurs membranaires. Il est à noter que les LB reconnaissent directement les antigènes, qu’ils soient solubles et circulant dans le milieu intérieur, ou qu’ils soient des antigènes membranaires des bactéries, par exemple.
Une fois différencié en plasmocyte, un LB devient sécréteur d’anticorps circulants, égale-ment appelés immunoglobulines (Ig), qui vont aller se fi xer sur les antigènes. Cette fi xation permet la formation d’un complexe antigène-anticorps ou complexe immun. La destruction ultérieure de l’antigène va ensuite différer suivant la classe de l’anticorps.
En effet, il existe plusieurs classes d’anticorps qui diffèrent par leur structure spatiale, et leurs effets dans les réponses immunitaires (Figure 7) :
IgG, IgD et IgE(monomère)
IgM(pentamère)
IgA(dimère)
Figure 7 Structures spatiales
des différentes immunoglobulines
¾¾ La classe des IgM (structure pentamérique, dix sites de reconnaissance). Elles sont pro-duites de manière précoce lors d’une réponse immunitaire primaire (réponse due à un premier contact avec un antigène), elles possèdent un site d’activation du complément, et ne peuvent traverser la barrière placentaire à cause de leur masse importante. On les trouve sur les membranes des LB et sécrétées par les plasmocytes.
BAH2_ch13.indd 244 28/02/2014 11:29

245
Immunologie
¾¾ La classe des IgG (structure monomérique, deux sites de reconnaissance). Elles sont produites plus tardivement que les IgM lors de la réponse immunitaire primaire et sont intégralement responsables de la réponse secondaire (réponse due à un deuxième contact avec le même antigène). Elles possèdent un site d’activation du complément et traversent la barrière placentaire (elles assurent donc l’immunité de l’enfant, le temps que son système immunitaire se mette en place). Principaux anticorps secrétés par les plasmocytes.
¾¾ La classe des IgA (structure dimérique, quatre sites de reconnaissance). Elles sont pré-sentes dans les différentes sécrétions (salives, larmes, lait maternel) et assurent l’immu-nité locale.
¾¾ La classe des IgE (structure monomérique, deux sites de reconnaissance). Elles se fi xent par leur fragment constant (F
c) sur les polynucléaires basophiles et mastocytes, et sont
responsables des réactions allergiques.
¾¾ La classe des IgD (structure monomérique, deux sites de reconnaissance). On les trouve sur les membranes des LB (en quantité infi me, moins de 1 %).
2. LesTCRLa capacité qu’ont les LT à reconnaître un élément du non soi est due à la présence de pro-téines membranaires, différentes des anticorps, que l’on appelle les TCR pour « T Cell Recep-tor » (en parallèle, les anticorps membranaires des LB sont parfois appelés BCR pour « B Cell Receptor »). Il s’agit de protéines qui ont une structure de base comprenant 2 chaînes polypeptidiques dont l’association ne constitue qu’un seul site de reconnaissance de l’anti-gène, donc un seul paratope, contrairement à l’anticorps (Figure 8). De plus, ces récepteurs sont uniquement membranaires. Chaque chaîne possède une région variable et une région constante, un domaine transmembranaire et un court domaine intracytoplasmique. Les deux chaînes sont reliées par un pont disulfure.
site de reconnaissance de l’antigène (paratope)
membrane plasmique
chaîne polypeptidique
pont disulfure
cytoplasme du LT
régions constantes
régions variables
Figure 8 Structure d’un TCR
Contrairement aux anticorps, les TCR ne reconnaissent pas directement les antigènes. Les épitopes doivent être présentés en même temps que des protéines du CMH à la surface des CPA (d’où cette appellation). De plus, les marqueurs CD (4 et 8) interviennent en tant que co-récepteurs dans cette reconnaissance. Cette dernière diffère selon le type de LT :
¾¾ Les TCR des LT4 : ils reconnaissent les épitopes associés au CMH de classe II, et le co-récepteur CD4 reconnaît le CMH II ;
¾¾ Les TCR des LT8 et des LTc : ils reconnaissent les épitopes associés au CMH de classe I, et le co-récepteur CD8 reconnaît le CMH I.
BAH2_ch13.indd 245 28/02/2014 11:29

246
Anatomie – physiologie
Cette double reconnaissance des TCR est schématisée dans la Figure 9.
TCR CD4épitope
CMH II
CD8
LT4
CPA
TCRépitope
CMH I
LT8
CPA
Figure 9 La double reconnaissance du TCR
La capacité des lymphocytes T à reconnaître les antigènes lors de la réponse immunitaire spécifi que à médiation cellulaire (voir La réponse immunitaire à médiation cellulaire, p. 253) est donc due à la présence des TCR à leur surface. Tous TCR qui se trouvent à la surface d’un lymphocyte T (environ 105) sont identiques, c’est-à-dire qu’ils reconnaissent tous le même antigène. On estime qu’il existe dans l’organisme 1012 lymphocytes T qui se répartissent en 107 clones différant les uns des autres par leurs récepteurs membranaires.
3. L’immunocompétenceetlerépertoireimmunologiqueUn lymphocyte est immunocompétent lorsqu’il exprime à sa surface des récepteurs mem-branaires spécifi ques d’un antigène donné, c’est-à-dire des anticorps membranaires pour un LB, et des TCR pour un LT.
Le système immunitaire doit identifi er de très nombreux antigènes différents, dont on a estimé le nombre théorique à 100 millions de milliards. Chez un adulte, la population des lymphocytes est estimée à 1012 dont 107 clones différents, chaque clone fabriquant un seul type de récepteurs membranaires.
Cette infi nie diversité des récepteurs s’explique par des mécanismes génétiques à l’origine de leur synthèse. En fait, les gènes qui codent pour les régions variables de ces récepteurs existent sous forme « morcelée » dans les chromosomes. Ils sont répartis en trois familles, et dans chaque famille, il existe de 4 à 100 allèles différents. Au cours de la synthèse d’une région variable, un membre de chaque famille s’associe aux autres pour former un « grand gène », et ce, de manière aléatoire. Ce réarrangement génétique, totalement aléatoire, per-met ainsi la formation d’un très grand nombre de récepteurs différents.
Puisque ces récepteurs membranaires sont produits au hasard des réarrangements géné-tiques, il existe un risque pour que ceux-ci reconnaissent les molécules du soi comme du non soi. Une telle reconnaissance aurait pour conséquence une autodestruction des cellules de l’individu par son propre système immunitaire (cas des maladies auto-immunes, voir Les maladies auto-immunes, p. 256). Ces lymphocytes possédant des récepteurs qui recon-naissent le soi comme du non soi sont nommés autoréactifs, et sont éliminés lors de la maturation des lymphocytes.
BAH2_ch13.indd 246 28/02/2014 11:29

247
Immunologie
� La maturation des LBLes lymphocytes pré-B, formés dans la moelle osseuse rouge par division cellulaire à partir des cellules souches lymphoïdes, vont donc exprimer à leur surface des anticorps membra-naires spécifiques d’un antigène donné. Ils sont donc devenus des LB matures et ont donc acquis leur immunocompétence. Avant la mise en circulation des LB, il faut une dernière étape de sélection qui va consister en l’élimination des Lymphocytes B autoréactifs. Pour cela, les lymphocytes sont mis en contact dans la moelle osseuse avec les marqueurs du soi, et ceux qui vont s’y fixer seront éliminés. Seuls les lymphocytes B ne reconnaissant pas le soi pourront donc devenir circulants et aller coloniser les organes lymphoïdes périphériques.
� La maturation des LTLes lymphocytes pré-T, formés dans la moelle osseuse rouge par division cellulaire à partir des cellules souches lymphoïdes, vont gagner le thymus. Ils vont alors exprimer à leur sur-face des TCR spécifiques d’un antigène donné. Ils deviennent donc immunocompétents. Là encore, il y aura nécessité d’une dernière étape de sélection visant à éliminer les lympho-cytes T autoréactifs. Cette sélection se déroule en 2 étapes : la première a lieu dans la zone périphérique (ou corticale), et la seconde, dans la zone centrale (ou médullaire) du thymus. Dans le cortex thymique, Les LT sont mis en contact avec les marqueurs du CMH, seules ceux s’y fixant seront conservés, et passeront dans la zone médullaire. Dans la médulla du thymus, Les LT sont mis en contact avec la totalité des marqueurs du soi, ceux qui s’y fixent seront éliminés. Seuls les Lymphocytes T reconnaissant le HLA et ne reconnaissant pas le soi passeront donc dans la circulation pour aller coloniser les organes périphériques.
Cette dernière étape de maturation visant à éliminer les lymphocytes autoréactifs permet ainsi d’établir le répertoire immunologique d’un individu. Celui-ci est donc défini par l’en-semble des lymphocytes immunocompétents existants au sein d’un organisme. Autrement dit, il est défini par l’ensemble des récepteurs différents portés par les lymphocytes d’un organisme. On estime à 107 le nombre de récepteurs différents.
III.LesréponsesImmunItaIres
Deux mécanismes sont mis en jeu, l’un est inné, et l’autre est acquis par chaque organisme au cours de sa vie :
¾¾ L’immunité non spécifique (ou innée) : il s’agit des moyens de défense non spécifiques à un germe donné. Ces moyens naturels comprennent les défenses de surface (peau et muqueuses) et la réaction inflammatoire. Cette réponse est génétiquement héritée, opérationnelle dès la naissance, et ne nécessite aucun « apprentissage ». Les modes d’ac-tion sont stéréotypés, sans adaptation particulière aux éléments du non soi concerné ;
¾¾ L’immunité spécifique (ou acquise ou adaptative) : il s’agit des moyens de défense qui sont spécifiques à un germe donné, qui se mettent en place lors de la première rencontre avec celui-ci. Elle permet l’éradication d’un certain nombre d’individus indési-rables de manière spécifique, et d’éviter que ces agressions ne se reproduisent (notion de mémoire immunitaire, voir Vaccination et sérothérapie, p. 260). Elle repose sur les réponses immunitaires spécifiques : la réponse immunitaire à médiation cellulaire (RIMC) et la réponse immunitaire à médiation humorale (RIMH).
BAH2_ch13.indd 247 28/02/2014 11:29

248
Anatomie – physiologie
A. L’immuniténonspécifiqueC’est une réponse de l’espèce, et non de l’individu dans le sens où le déroulement de la réponse fera intervenir les mêmes éléments du système immunitaire chez tous les indivi-dus, et ce, quelle que soit la nature de l’élément étranger visé par cette réponse. C’est la pre-mière barrière de l’immunité. Elle va se mettre en place dès lors qu’un élément étranger va franchir les barrières tissulaires (peau et muqueuses) à l’occasion d’une lésion quelconque.
1. Labarrièrecutanéo-muqueuseLa peau et les muqueuses constituent les frontières naturelles qui nous séparent du monde extérieur. Elles jouent le rôle de barrières tissulaires selon trois mécanismes.
� La peau Un mécanisme physique : la peau est constituée d’un épithélium stratifié avec des cellules kératinisées dans sa couche superficielle. Cette structure lui confère une résistance phy-sique. Elle contient également des cellules pigmentaires, les mélanocytes (la mélanine étant le pigment) qui lui confèrent un rôle de protection contre les radiations, notamment contre les rayons ultraviolets.
Un mécanisme chimique : la peau présente à sa surface un film hydro-lipidique, le sébum, qui contient des acides gras, secrété par les glandes sébacées (ou sudoripares), qui empêche le développement de certains micro-organismes.
Un mécanisme biologique : à la surface de la peau, réside une flore bactérienne dite « rési-dente » ou « commensale » qui permet l’inhibition de la croissance des germes exogènes pathogènes.
� Les muqueusesUn mécanisme physique : les muqueuses produisent du mucus et, selon la muqueuse consi-dérée, certaines variétés de cellules possèdent des ultrastructures (les cils vibratiles des cellules de la muqueuse respiratoire, par exemple) qui permettent l’immobilisation et l’éva-cuation des micro-organismes (toux ou éternuement, par exemple).
Un mécanisme chimique : grâce au pH acide des sucs. Exemple : la muqueuse gastrique et la muqueuse vaginale. De plus, le mucus contient des substances antiseptiques.
Un mécanisme biologique : comme pour la peau, des bactéries dites « commensales » sont présentes sur les muqueuses, et permettent l’inhibition de la croissance de germes exo-gènes pathogènes.
2. LaréactioninflammatoireLa réaction inflammatoire comprend l’ensemble des phénomènes déclenchés par l’effrac-tion de la barrière cutanéo-muqueuse. Cette réaction est en général locale, et comporte plusieurs étapes.
� Une étape vasculaireLes agents infectieux endommagent les tissus, ce qui provoque la libération de substances chimiques, les chimiokines, par des cellules immunitaires qui ont reconnu ces germes. Cette libération va alors provoquer une vasodilatation locale entraînant une augmentation du débit sanguin local et de la perméabilité vasculaire, permettant la sortie de cellules sanguines vers le tissu lésé. Cette vasodilatation est à l’origine de l’apparition de chaleur et de rougeur. De plus, elle provoque la stimulation des terminaisons nerveuses entraînant la douleur.
BAH2_ch13.indd 248 28/02/2014 11:29

249
Immunologie
� Une étape cellulaireLes cellules lésées libèrent des molécules qui vont alors attirer par chimiotactisme les poly-nucléaires neutrophiles se trouvant dans le sang. Ces derniers vont alors traverser la paroi vasculaire par diapédèse. Une fois, dans le tissu lésé, ils phagocytent les éléments du non soi afi n de les éliminer. Lorsque les polynucléaires ne suffi ssent pas pour éliminer le non soi, les macrophages interviennent à leur tour, pour jouer leur rôle de phagocyte. Cette accu-mulation de cellules immunitaires dans le tissu lésé entraîne alors la formation d’un œdème.
� Une étape de réparation Il s’agit de l’étape de cicatrisation. Elle est essentiellement assurée par les fi broblastes qui prolifèrent et permettent la reconstitution du tissu.
La Figure 10 schématise un exemple de réaction infl ammatoire.
écharde lésant le tissu
intrusion de micro-organismes (non- soi)
phagocyte (polynucléaireneutrophile et macrophage)
diapédèse
cellule endothélialede la paroi vasculaire
vasodilatation localehématie
peau
sang
Figure 10 La réaction infl ammatoire
Lorsque les macrophages sont dépassés par l’ampleur de l’agression, et n’arrivent pas à éli-miner l’ensemble des éléments du non soi, ils vont alors activer les réponses immunitaires spécifi ques en devenant des CPA.
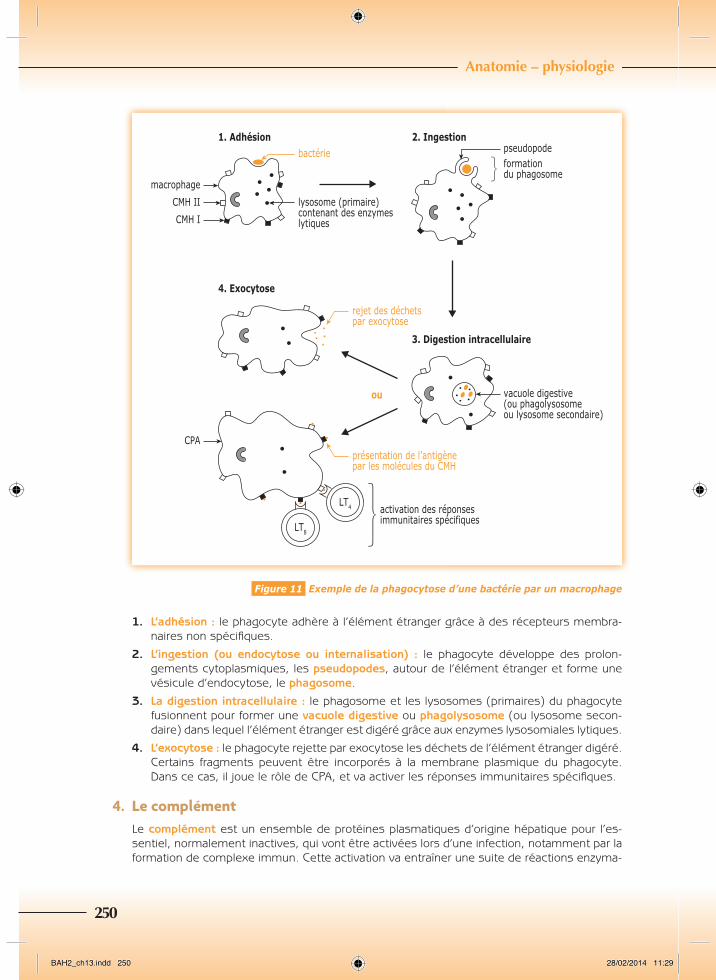
3. LaphagocytoseLa phagocytose est un mécanisme cellulaire qui tend à éliminer les éléments étrangers. Ce mécanisme correspond à l’ingestion et à la digestion des particules étrangères par les cellules phagocytaires, c’est-à-dire soit par les macrophages, soit par les granulocytes neu-trophiles. Elle se déroule en 4 phases (Figure 11).
BAH2_ch13.indd 249 28/02/2014 11:29
250
Anatomie – physiologie
LT8
LT4
1. Adhésion
4. Exocytose
3. Digestion intracellulaire
ou
2. Ingestion
CMH II
macrophage
CPA
CMH I
lysosome (primaire)contenant des enzymeslytiques
pseudopode
vacuole digestive(ou phagolysosomeou lysosome secondaire)
bactérie
rejet des déchetspar exocytose
présentation de l’antigènepar les molécules du CMH
activation des réponsesimmunitaires spécifiques
formationdu phagosome
Figure 11 Exemple de la phagocytose d’une bactérie par un macrophage
1. L’adhésion : le phagocyte adhère à l’élément étranger grâce à des récepteurs membra-naires non spécifi ques.
2. L’ingestion (ou endocytose ou internalisation) : le phagocyte développe des prolon-gements cytoplasmiques, les pseudopodes, autour de l’élément étranger et forme une vésicule d’endocytose, le phagosome.
3. La digestion intracellulaire : le phagosome et les lysosomes (primaires) du phagocyte fusionnent pour former une vacuole digestive ou phagolysosome (ou lysosome secon-daire) dans lequel l’élément étranger est digéré grâce aux enzymes lysosomiales lytiques.
4. L’exocytose : le phagocyte rejette par exocytose les déchets de l’élément étranger digéré. Certains fragments peuvent être incorporés à la membrane plasmique du phagocyte. Dans ce cas, il joue le rôle de CPA, et va activer les réponses immunitaires spécifi ques.
4. LecomplémentLe complément est un ensemble de protéines plasmatiques d’origine hépatique pour l’es-sentiel, normalement inactives, qui vont être activées lors d’une infection, notamment par la formation de complexe immun. Cette activation va entraîner une suite de réactions enzyma-
BAH2_ch13.indd 250 28/02/2014 11:29

251
Immunologie
tiques en cascade qui va aboutir à plusieurs effets en lien avec la neutralisation et la destruc-tion de l’élément étranger. Les deux principaux effets sont :
¾¾ Les protéines du complément peuvent former, par polymérisation, un complexe d’attaque membranaire pouvant aller s’ancrer dans la membrane des éléments étrangers de nature cellulaire. Ceci entraîne la formation de pores membranaires, et donc la destruction de l’élément étranger par choc osmotique (entrée massive d’eau, la cellule va éclater) ;
¾¾ L’opsonisation : certaines protéines du complément fixent l’élément du non soi, et attirent par chimiotactisme les phagocytes. Les opsonines, d’où ce terme d’opsonisation dérive, sont des molécules facilitant la phagocytose. Certaines protéines du complément sont donc des opsonines.
B. L’immunitéspécifiqueEn cas d’échec des barrières de l’immunité non spécifique, les réactions immunitaires spé-cifiques (RIS) seront activées.
C’est une réponse de l’individu et non de l’espèce, dans le sens où le déroulement de la réponse fera intervenir des éléments du système immunitaire différents chez les individus, éléments spécifiques de l’élément étranger visé par cette réponse. Elle va se mettre en place dès lors qu’un élément étranger va réussir à déborder les défenses non spécifiques et passer dans le milieu intérieur.
Les cellules capables d’interagir de manière spécifique avec les déterminants antigéniques sont les lymphocytes. Les lymphocytes B reconnaissent le déterminant de manière isolée grâce à leurs anticorps membranaires. Les lymphocytes T reconnaissent le déterminant anti-génique associé à un marqueur du CMH grâce à leur TCR.
Il existe 2 grands types de réponse spécifique qui différent par la nature des effecteurs qui vont permettre la destruction des éléments étrangers :
¾¾ La RIS à médiation humorale faisant intervenir les LB à l’origine des cellules sécrétrices d’anticorps circulants, les plasmocytes. Elle permet de conserver l’intégrité du milieu extracellulaire ;
¾¾ La RIS à médiation cellulaire faisant intervenir les LT8 à l’origine des LT cytotoxiques. Elle permet de conserver l’intégrité du milieu intracellulaire.
Ces 2 réponses se déroulent en trois étapes successives : la phase d’induction (ou de sélection clonale), la phase d’amplification (ou d’expansion et de différenciation clonale) et la phase effectrice.
1. L’antigèneUn antigène est défini comme toute molécule libre (antigène soluble ou particulaire), ou portée par une cellule (antigène cellulaire ou membranaire), et pouvant être reconnue par le système immunitaire, et par là, induire le développement d’une réponse immunitaire spéci-fique. Il possède donc 2 propriétés fondamentales :
¾¾ L’immunogénicité : possibilité d’induire une réponse immunitaire ;
¾¾ La spécificité antigénique : capacité d’être reconnu par des éléments spécifiques du système immunitaire.
Autrement dit, un antigène est un élément du non soi entraînant des réactions immuni-taires spécifiques.
En fait, l’antigène en tant qu’élément étranger n’est généralement pas reconnu dans son entier. Le système immunitaire va être stimulé par des fragments de nature protéique ou glu-
BAH2_ch13.indd 251 28/02/2014 11:29

252
Anatomie – physiologie
cidique que l’on appelle les déterminants antigéniques, ou épitopes. Un antigène peut donc porter plusieurs déterminants, qu’ils soient identiques ou différents. Il y aura une réponse spécifique dirigée contre chaque déterminant.
2. LaréponseimmunitaireàmédiationhumoraleElle se déroule en plusieurs étapes.
1. La phase d’induction : cette phase a lieu dans les organes lymphoïdes secondaires. Il va y avoir sélection et activation d’un LB et d’un LT4 par le même épitope. La reconnaissance de ce dernier par les LB s’effectue par liaison avec l’anticorps membranaire spécifique. La reconnaissance de l’épitope associé à une molécule du CMH II, présentée par une CPA ou une cellule infectée, par les LT4 s’effectue via le TCR. Ces récepteurs reconnaissent donc des fragments issus de l’antigène exprimés à la surface des cellules infectées ou des CPA et associés aux molécules du CMH. Il s’agit de la double reconnaissance du TCR. Par ailleurs, il existe un co-récepteur exprimé à la surface membranaire des LT4, la glycoprotéine CD4, qui assiste le TCR lors de son interaction avec les protéines CMH II. Il y a donc ici, sélection d’un clone de lymphocyte B, et d’un clone de lymphocyte T4 dans le répertoire immunologique des lymphocytes naïfs, ou vierges (lymphocytes n’ayant jamais été en contact avec l’antigène).
2. La phase d’amplification : cette phase a également lieu dans les organes lymphoïdes secondaires. La reconnaissance de l’épitope par le clone de LB et le clone LT4 a pour effet d’exprimer à leur surface des récepteurs aux interleukines (IL). Les interleukines sont des cytokines, c’est-à-dire des glycoprotéines produites par des cellules immuni-taires qui ont une action intercellulaire (stimulation de la prolifération et différenciation cellulaire) au cours des réponses immunitaires. Grâce à la phase d’induction, les CPA secrètent de l’interleukine 1 qui va alors stimuler la prolifération des LT4 sélectionnés, et leur différenciation en LTa (ou LTh), sécréteur d’interleukines. Ces derniers vont alors secréter les interleukines 2 et 4, qui vont stimuler la prolifération des LB sélectionnés, et les interleukines 4 et 6, qui vont stimuler leur différenciation en plasmocytes, cellules sécrétrices d’anticorps circulants. L’interleukine 2 exerce également une autostimulation sur la prolifération et différenciation des LT4. Lors de la différenciation des LB et des LT4, une petite partie des cellules va se transformer en LB et LT4 mémoire, cellules à durée de vie plus longue qui expriment à leur surface plus de récepteurs membranaires que les lymphocytes naïfs. Ces cellules seront responsables d’une réponse secondaire éventuelle lors d’une rencontre ultérieure avec le même antigène (voir Vaccination et sérothérapie, p. 260).
3. La phase effectrice : cette phase a lieu au niveau des tissus infectés, dans le com-partiment extracellulaire. Les immunoglobulines circulantes libérées dans la circulation sanguine par les plasmocytes vont aller se fixer sur les antigènes permettant leur neutra-lisation. Cette fixation entraîne la formation d’un complexe antigène-anticorps ou com-plexe immun. La destruction ultérieure de l’antigène va différer suivant sa nature. Il peut y avoir phagocytose des complexes immun. Il peut également y avoir activation du complément (par l’intermédiaire du site d’activation présent sur la partie constante des chaînes lourdes des Ig), ce qui entraînera soit la lyse de l’antigène, soit son opsonisation. L’intégrité du milieu extracellulaire est alors préservée.
Un schéma récapitulatif de cette réponse est présenté dans la Figure 12.
BAH2_ch13.indd 252 28/02/2014 11:29

253
Immunologie
bactérie
épitopeTCRCMH IICD4anticorps
membranaire
anticorpscirculants
plasmocyte
Organelymphoïdesecondaire
Sang
Tissuinfecté
Maintien de l’intégrité du milieu extracellulaire
+
+
CPA
IL-2 et 4IL-2
IL-1
IL-4 et 6
Multiplication des LB
LT4 LT4
LB LB
LB LTa LT4m
LBm
1 Phase d’induction : sélection et activation clonale2 Phase d’amplification : multiplication et différenciation cellulaire3 Phase effectrice
3
2
1
complexe immun⇒ neutralisation de l’antigène
Élimination par : – phagocytose – activation du complément
Multiplication et différenciationdes LT4 en LTa et LT4 mémoire
Différenciation en plasmocyteset en LB mémoire
répertoire immunologique(lymphocytes naïfs)
Figure 12 La RIMH
3. LaréponseimmunitaireàmédiationcellulaireElle se déroule en plusieurs étapes :
1. La phase d’induction : cette phase a lieu dans les organes lymphoïdes secondaires. Il va y avoir sélection et activation d’un LT8 et d’un LT4 par le même épitope, mais présenté par des molécules du CMH différentes (CMH I/LT8 et CMH II/LT4). Cette sélection s’effectue donc grâce à la double reconnaissance des TCR. Il existe des co-récepteurs exprimés à la surface membranaire des LT4 et des LT8, respectivement les glycoprotéines CD4 et CD8, qui assistent le TCR lors de son interaction avec les protéines CMH II et CMH I, respective-ment. Il y a donc ici, sélection d’un clone de lymphocyte T8 et d’un clone de lymphocyte T4 dans le répertoire immunologique des lymphocytes naïfs.
2. La phase d’amplifi cation : cette phase a également lieu dans les organes lymphoïdes secondaires. La reconnaissance de l’épitope par le clone de LT8 et le clone de LT4 a pour effet d’exprimer à leur surface des récepteurs aux interleukines. Grâce à la phase d’induction, les CPA secrètent de l’interleukine 1 qui va alors stimuler la prolifération des LT4 sélectionnés, et leur différenciation en LTa (ou LTh), sécréteur d’interleukines. Ces derniers vont alors secréter de l’interleukine 2 qui va stimuler la prolifération et différen-ciation des LT8 sélectionnés en LT cytotoxiques. L’interleukine 2 exerce également une autostimulation sur la prolifération et différenciation des LT4. Lors de la différenciation des
BAH2_ch13.indd 253 28/02/2014 11:29

254
Anatomie – physiologie
LT8 et des LT4, une petite partie des cellules va se transformer en LT mémoire, cellules à durée de vie plus longue qui expriment à leur surface plus de récepteurs membranaires que les lymphocytes naïfs. Ces cellules seront responsables d’une réponse secondaire éventuelle lors d’une rencontre ultérieure avec le même antigène (voir Vaccination et sérothérapie, p. 260).
3. La phase effectrice : les LT cytotoxiques vont alors rejoindre le tissu infecté, et se fi xer sur leurs cellules cibles grâce à leur TCR reconnaissant le même complexe CMH I/épitope que les LT8 sélectionnés. Cette fi xation va entraîner la destruction de la cellule cible (cytolyse) par l’intermédiaire d’un facteur libérer par le LTc, appelé perforine, qui va se polymériser pour former un pore membranaire provoquant la lyse de la cellule par choc osmotique. Le LTc peut également déclencher l’apoptose (mort cellulaire programmée, suicide, baiser de la mort) de sa cellule cible en libérant des facteurs apoptotiques. L’apoptose est un mécanisme d’autodestruction cellulaire programmé génétiquement. Ce mécanisme se caractérise par une fragmentation de l’ADN et un bourgeonnement de la membrane plasmique qui forme des « corps apoptotiques », petites vésicules qui seront ensuite éliminées par les phagocytes. L’intégrité du milieu intracellulaire est alors préservée.
Un schéma récapitulatif de cette réponse est présenté dans la Figure 13.
TCRCD8
CMH IIépitope
CMH I
Organelymphoïdesecondaire
Tissuinfecté
Maintien de l’intégrité du milieu intracellulaire
+
CPA
IL-2
IL-2
IL-1
LT8
TCRCD4
LT4
LTc LT8m+ LTaLT4m
Sang
facteursapoptotiques
H2O 1 Phase d’induction : sélection et activation clonale2 Phase d’amplification : multiplication et différenciation cellulaire3 Phase effectrice
3
2
1
Multiplication et différenciation desLT4 en LTa et LT4 mémoire
Multiplication et différenciation des LT8 en LTc et LT8 mémoire
répertoire immunologique(lymphocytes naïfs)LT4 LT8
perforine(cytolyse)
cellule infectéeou cancéreuse
pore membranaireformé de perforines
LTc
Figure 13 La RIMC
BAH2_ch13.indd 254 28/02/2014 11:29

255
Immunologie
IV.LesdysfonctIonnementsImmunItaIres
A. LesallergiesLes allergies sont des réactions d’hypersensibilité dues à une réponse inadaptée du sys-tème immunitaire vis-à-vis d’un antigène qui devrait, en temps normal, être toléré par celui-ci. On nomme « allergène », une substance qui devrait donc normalement être toléré par l’organisme, mais qui déclenche ou favorise un ensemble de réactions excessives du sys-tème immunitaire de l’organisme suite à un contact, une infection, une ingestion ou une inhalation. On peut distinguer les allergènes selon leur voie d’entrée dans l’organisme.
¾¾ Les allergènes inhalés : ils pénètrent par les voies respiratoires. Par exemple, le pollen, les poils d’animaux, les plumes, et les acariens peuvent être des allergènes inhalés. En général, ils sont responsables d’un asthme (inflammation de la muqueuse bronchique et sécrétion accrue de mucus) allergique, ou d’une rhinite (irritation et inflammation de la muqueuse de la cavité nasale) allergique ;
¾¾ Les allergènes cutanés : ils pénètrent par la voie cutanée (la peau). Par exemple, les piqûres d’insectes, les produits ménagers, et les cosmétiques peuvent être des aller-gènes cutanés. En général, ils sont responsables de dermite allergique (l’urticaire : érup-tion cutanée caractérisée par des rougeurs à contours réguliers, des gonflements et des démangeaisons et l’eczéma : éruption cutanée caractérisée par des rougeurs à contours irréguliers, et des démangeaisons) ;
¾¾ Les allergènes alimentaires : ils pénètrent par la voie digestive. Par exemple, le lait de vache, le blanc d’œuf, les arachides, les fruits de mer, les additifs alimentaires, les conservateurs et certains médicaments sont des allergènes alimentaires. La nature et l’intensité des symptômes varient d’une personne à l’autre. Il peut y avoir des symp-tômes cutanés (démangeaisons, éruptions cutanées, rougeurs, gonflement des lèvres, du visage et des membres), des symptômes respiratoires (respiration sifflante, sensation de gonflement de la gorge, difficulté à respirer, sensation d’étouffement), des symp-tômes digestifs (crampes abdominales, diarrhée, coliques, nausées et vomissements) et des symptômes cardiovasculaires (pâleur, pouls faible, étourdissements et perte de conscience). Tous ces symptômes peuvent se manifester seul ou associés.
L’allergie met en jeu 2 étapes successives :
¾¾ Une étape de sensibilisation, où le système immunitaire va être mis en contact avec l’allergène pour la première fois, et va fabriquer anormalement des éléments spécifiques dirigés contre celui-ci. Cette première étape est le plus souvent totalement inapparente (sans symptômes) ;
¾¾ Une étape de réaction allergique proprement dite, qui se produit lors du second contact avec le même allergène, et voit l’apparition de symptômes spécifiques.
Il existe plusieurs types de réactions d’hypersensibilité dont les plus importantes sont les hypersensibilités immédiates, et retardées.
1. L’hypersensibilitéimmédiateouanaphylaxieCette allergie est une hypersensibilité de la RIMH ; elle résulte d’une production excessive d’IgE.
La sensibilisation : lors du premier contact avec l’allergène, l’organisme déclenche une RIMH. Les LT auxiliaires produits au cours de cette réponse vont stimuler la production d’IgE par des plasmocytes, de manière excessive. Ces immunoglobulines vont aller se fixer sur la membrane des mastocytes (présents dans la peau et les muqueuses) et des granulocytes
BAH2_ch13.indd 255 28/02/2014 11:29

256
Anatomie – physiologie
basophiles (présents dans le sang). Cette phase est le plus souvent progressive, et sans symptômes.
La réaction allergique proprement dite : lors d’un second contact (nommé contact déclen-chant), l’allergène va se fixer sur les IgE portées par les mastocytes et granulocytes baso-philes et induire leur dégranulation. Il y a alors libération de grandes quantités de médiateurs de l’inflammation, dont le principal est l’histamine, qui provoque une vasodilatation locale entraînant une rougeur et un œdème. Cette réaction est relativement rapide puisqu’elle apparaît dans les secondes ou les minutes qui suivent le contact déclenchant.
Le pollen, les acariens, les œufs, les arachides, le lait de vache et certains médicaments peuvent par exemple, induire une hypersensibilité immédiate. Si la réaction est vraiment importante, il peut y avoir choc anaphylactique. Ce dernier est potentiellement mortel, il se traduit en plus des symptômes habituels, par une hypotension, et un œdème de la couche profonde de la peau et des muqueuses entraînant l’œdème de Quincke (œdème du visage et du cou) qui peut rapidement évoluer vers une obstruction des voies respiratoires.
2. L’hypersensibilitéretardéeCette allergie est une hypersensibilité de la RIMC ; elle résulte d’une production excessive de cytokines.
La sensibilisation : lors du premier contact avec l’allergène, l’organisme déclenche une RIMC avec formation de lymphocytes T mémoire.
La réaction allergique proprement dite : lors d’un second contact, l’allergène va stimuler ces lymphocytes mémoire qui vont libérer de manière excessive des interleukines. Celles-ci vont alors exercer un puissant effet chimiotactique sur les mastocytes, et induire leur dégranula-tion entraînant une réaction inflammatoire. Cette réaction est dite retardée car elle apparaît quelques jours après le contact déclenchant.
Certaines infections bactériennes comme la tuberculose, la lèpre et la syphilis et des greffes d’organe peuvent par exemple, induire une hypersensibilité retardée.
B. Lesmaladiesauto-immunesLes maladies auto-immunes sont dues à la rupture de la tolérance au soi. Elles sont dues à une hyperactivité du système immunitaire à l’encontre de substances ou de tissus normale-ment présents et tolérés dans l’organisme.
Leur origine n’est pas encore bien élucidée, mais les substances chimiques introduites depuis 50 ans dans l’environnement (conservateurs, émulsifiants, stabilisateurs alimentaires, pesti-cides, particules volatiles polluantes par exemple) semblent favoriser leur apparition. Divers facteurs sont également reconnus comme favorisant l’apparition et le développement de ces pathologies :
¾¾ Le sexe : les femmes semblent plus exposées ;
¾¾ Des facteurs extérieurs : l’exposition au rayonnement UV, ou certains agents infectieux, par exemple ;
¾¾ Des facteurs génétiques : par exemple, certaines molécules du CMH II, en particulier HLA-DR semblent avoir un lien étroit avec le diabète de type I, qui est une maladie auto-immune. Les individus possédant les allèles codant pour HLA-DR3 et/ou HLA-DR4 montrent une prédisposition génétique au diabète de type 1, c’est-à-dire qu’ils ont plus de risques de développer cette maladie par rapport à des individus qui ne les possèdent pas.
BAH2_ch13.indd 256 28/02/2014 11:29

257
Immunologie
Les maladies auto-immunes sont très différentes dans leur expression clinique, et dans les organes touchés, mais elles ont toutes en commun le même mécanisme : le système immunitaire du malade s’attaque à certains constituants de son propre organisme comme s’il s’agissait de substances étrangères (via les RIMH et RIMC), et produit alors des anticorps dirigés contre ceux-ci, que l’on nomme auto-anticorps. Par contre, elles peuvent être locali-sées à un organe spécifique, ou s’étendre à l’ensemble de l’organisme. Dans cette dernière situation, on parle de maladies systémiques. Voici quelques exemples de maladies auto-immunes localisées :
¾¾ Le diabète de type 1 : il touche le pancréas, et plus particulièrement les cellules bêta des îlots de Langerhans qui produisent l’insuline. On retrouve, chez les personnes atteintes, des auto-anticorps anti-cellules bêta. L’individu atteint souffre alors d’une hyperglycémie chronique ;
¾¾ La sclérose en plaque (SEP) : elle touche la gaine de myéline des fibres nerveuses. On retrouve, chez les personnes atteintes, des auto-anticorps anti-myéline. L’individu atteint souffre de lésions de la gaine de myéline entraînent une déficience musculaire et de la sensibilité, des troubles d’élocution et de la vue.
Voici quelques exemples de maladies auto-immunes systémiques :
¾¾ Le lupus : on retrouve, chez les personnes atteintes, entre autres, des auto-anticorps anti-nucléosome. Les manifestations sont diverses : douleurs articulaires et musculaires, des plaques rouges sur le visage, une hypersensibilité aux rayonnements solaires, un gonflement des jambes, un état dépressif, perte de cheveux, fièvre, fatigue… ;
¾¾ La polyarthrite rhumatoïde : on retrouve, chez les personnes atteintes, des auto-anticorps anti-gammaglobulines. Elle se caractérise par une atteinte articulaire évoluant vers la déformation et la destruction des articulations.
C. LesdéficitsimmunitairesLes déficits immunitaires sont dus à une immunodépression ou à une immunodéficience, c’est-à-dire, respectivement, à une diminution ou à une suppression des capacités du sys-tème immunitaire. On distingue deux types de déficits qui diffèrent par leur origine :
¾¾ Les déficits immunitaires primitifs, ou congénitaux ou innés : ces déficits peuvent tou-cher tous les aspects de la réponse immunitaire. Ils se manifestent chez les nourrissons de plus de 6 mois, c’est-à-dire une fois que l’immunité due aux anticorps maternels ait disparu. Lorsque le déficit est complet, la guérison est possible en faisant une greffe de la moelle osseuse HLA-compatible. Ce sont des maladies rares ;
¾¾ Les déficits immunitaires secondaires, ou acquis : ils touchent aussi bien les enfants que les adultes, et sont beaucoup plus fréquents que les déficits primitifs. Les causes en sont extrêmement variées : des infections bactériennes ou virales, des parasitoses, des intoxications (drogues, médicaments), le vieillissement, un traitement immunosuppres-seur peuvent par exemple causer ce genre de déficit. En général, ces déficits secondaires sont temporaires, ce qui n’est pas le cas pour le SIDA (Syndrome d’ImmunoDéficience Acquise) causé par le VIH (Virus de l’Immunodéficience Humaine).
Le VIH a pour cellules cibles les cellules immunitaires possédant le marqueur membranaire CD4, c’est-à-dire les LT4, les monocytes et les macrophages. Son infection évolue selon trois étapes.
¾¾ La primo-infection : elle correspond à la dissémination du virus dans l’organisme. Elle est le plus souvent asymptomatique. Cependant, dans 30 à 40 % des cas, 2 à 6 semaines après la contamination, elle se manifeste par un syndrome pseudo-grippal avec des signes non spécifiques tels que fièvre, et douleurs musculaires. Cette symptomatolo-
BAH2_ch13.indd 257 28/02/2014 11:29

258
Anatomie – physiologie
gie régresse spontanément au bout d’une à trois semaines. Au cours de cette phase, le virus se multiplie de façon importante, et il en résulte une charge virale plasmatique élevée (la charge virale correspond au nombre de copies d’ARN viral par ml de plasma, et révèle la présence du virus) associée à une diminution du nombre de LT4. L’organisme va alors développer des réponses immunitaires spécifiques (RIMC et RIMH) permettant la production de LTc spécifiques du virus, et d’anticorps neutralisants. L’individu devient alors séropositif pour le VIH, c’est-à-dire que son plasma contient des anticorps anti-VIH.
¾¾ La phase asymptomatique : cette période dure en moyenne 7 à 10 ans en l’absence de traitement. Le virus continue de se multiplier, mais à un niveau inférieur à celui de la primo-infection, grâce aux réponses immunitaires développées. Il n’y a aucun symptôme spécifique associé à ce stade. Preuve d’une réponse immunitaire, les ganglions lympha-tiques sont en général hypertrophiés.
¾¾ La phase SIDA déclaré ou symptomatique : à terme les réponses immunitaires sont dépassées et ne permettent plus le contrôle de la réplication virale. Le taux de LT4 chute considérablement et n’est plus suffisant pour assurer le bon déroulement des réponses immunitaires. Ce stade d’immunodéficience se caractérise par la survenue d’infections opportunistes (pneumocystose, toxoplasmose, infections à mycobactéries, atteintes vis-cérales par le cytomégalovirus) et de proliférations cellulaires (maladie de Kaposi, lym-phomes). Sans traitement, la mort peut survenir en quelques mois.
D. Lesmyélomesetlymphomes
1. LesmyélomesLe myélome ou myélome multiple ou maladie de Kahler concerne environ 1 % des cancers, et 12 % des maladies hématologiques malignes, en France. Il touche en général des indivi-dus âgés d’environ 70 ans.
Cette pathologie est caractérisée par une prolifération anormale et incontrôlée d’un clone de plasmocytes, provoquant une production excessive d’un seul type d’immunoglobuline. L’accumulation de cette population clonale de plasmocytes dans la moelle osseuse provoque une disparition progressive des cellules hématopoïétiques entraînant une anémie, ainsi que des lésions (lacunes, déminéralisation et fractures) et douleurs osseuses. L’atteinte osseuse va alors également entraîner une hypercalcémie due à la déminéralisation. De plus, l’accu-mulation des immunoglobulines va obstruer les vaisseaux sanguins, notamment au niveau rénal, entraînant une insuffisance rénale.
Le traitement consiste en général en une chimiothérapie associée à une radiothérapie. La greffe de moelle osseuse est très rarement préconisée, car elle n’est pas réalisable au-des-sous de 45 ans, pour une allogreffe (voir Transplantation d’organe, p. 263), et au-dessous de 55 ans, pour une autogreffe (voir Transplantation d’organe, p. 263).
2. LeslymphomesLes lymphomes sont caractérisés par une prolifération anormale et incontrôlée des cellules lymphoïdes. Il s’agit donc de cancers du système lymphatique. On distingue le lymphome de Hodgkin des lymphomes non hodgkiniens.
� Le lymphome de HodgkinLa cause de cette pathologie est encore inconnue. En général, ce lymphome touche les individus entre 15 et 40 ans.
BAH2_ch13.indd 258 28/02/2014 11:29

259
Immunologie
Ce type de lymphome est caractérisé par la présence de grandes (environ 50 µm) cellules anormales, les cellules de Reed-Sternberg. Elles sont caractérisées par un noyau volumi-neux multilobé.
Les symptômes initiaux de cette pathologie s’apparentent souvent à ceux d’une grippe : adénopathies (hypertrophie des ganglions lymphatiques) au niveau du cou, des aisselles ou de l’aine, fièvre, fatigue et sueurs nocturnes. Des pertes d’appétit et de poids peuvent également être observées.
Le traitement dépend du stade du lymphome. En effet, on distingue 4 stades dans cette pathologie :
¾¾ Le stade I : le lymphome est confiné à l’intérieur des ganglions lymphatiques ;
¾¾ Le stade II : il s’est propagé dans le système lymphatique, mais n’a pas traversé le diaphragme.
Pour ces deux stades, le traitement préconisé est en général une radiothérapie où seuls les ganglions sont visés par les rayons. Parfois, cette radiothérapie est associée à une chimio-thérapie.
¾¾ Le stade III : il s’est propagé dans tout le système lymphatique ;
¾¾ Le stade IV : il s’est propagé au-delà du système lymphatique. Il y a une atteinte viscé-rale (foie, cœur, poumon, tube digestif, os, système nerveux). Dans ce cas, l’individu peut souffrir d’une pleurésie (inflammation de la plèvre, membrane entourant les pou-mons), de pneumopathies (infections pulmonaires), d’une gastrite (inflammation de la muqueuse gastrique), d’une atteinte hépatique, d’une paraplégie (paralysie des deux membres inférieurs), de névralgies (douleurs suite à l’inflammation d’un nerf sensitif) et d’une splénomégalie (augmentation du volume de la rate).
Pour ces deux stades, le traitement préconisé est en général une chimiothérapie intensive suivie d’une radiothérapie.
� Les lymphomes non hodgkiniensLes lymphomes non hodgkiniens (également nommés lymphosarcomes) sont cinq fois plus fréquents que le lymphome hodgkinien. Il survient en général vers l’âge de 60 ans à 70 ans. Les patients immunodépressifs ou immunodéficients (par exemple, dans le cas du SIDA ou d’une greffe avec traitement immunosuppresseur) sont les plus à risque. Environ 10 % des patients infectés par le VIH souffriront d’un lymphome non hodgkinien.
Il existe plusieurs types de lymphomes non hodgkiniens. On les identifie selon l’apparence des cellules sous le microscope. Parmi eux, on distingue 2 grandes catégories : les lym-phomes indolents et les lymphomes agressifs. Les premiers se développent plus lentement, et engendrent moins de symptômes. Les seconds ont une croissance plus rapide. Les symp-tômes et traitements sont sensiblement les mêmes que pour un lymphome de Hodgkin. Si la chimiothérapie et la radiothérapie ne sont pas suffisantes, on a recourt à une autogreffe (voir Transplantation d’organe, p. 263) de moelle osseuse.
Parmi ces lymphomes, deux formes particulières sont à distinguer : le lymphome de Burkitt et le mycosis fongoïde.
Le lymphome de Burkitt est caractérisé par une prolifération anormale des lymphocytes B, notamment en dehors des ganglions lymphatiques. De ce fait, la pathologie est souvent décelée suite à la découverte de masses abdominales au niveau intestinal. Cette maladie s’observe essentiellement chez les enfants noirs d’Afrique centrale, sous forme endémique, et sous forme sporadique, chez les adultes, en Europe et en Amérique du Nord.
BAH2_ch13.indd 259 28/02/2014 11:29

260
Anatomie – physiologie
Le mycosis fongoïde est caractérisé par une prolifération anormale des lymphocytes T. Il s’agit d’un lymphome non hodgkinien cutané, rare, qui évolue lentement, mais de façon défavorable. Il se manifeste par des plaques rouges, qui peuvent régresser ou devenir per-manentes, s’accentuant et s’infiltrant de manière irrégulière en plusieurs mois ou années. Si un traitement adéquat n’est pas entrepris, de véritables tumeurs apparaissent dont la res-semblance avec des champignons explique le nom de mycosis fongoïde. Parallèlement, les ganglions lymphatiques peuvent être atteints et, dans les formes très évoluées, les organes.
V.LesappLIcatIonscLInIquesetthérapeutIques
A. VaccinationetsérothérapieBien que l’organisme soit doté d’un système immunitaire efficace, les maladies infectieuses restent, encore aujourd’hui, une des premières causes de mortalité dans le monde. Les connaissances sur les mécanismes immunitaires, et notamment sur l’existence d’une mémoire immunitaire, ont été mises à profit par l’Homme pour renforcer ces défenses, grâce à la vaccination et la sérothérapie.
1. LamémoireimmunitaireLa première rencontre avec un antigène induit donc des réponses immunitaires spécifiques, ces réponses sont dites primaires. Comme nous l’avons vu précédemment, des lympho-cytes mémoires, cellules possédant un plus grand nombre de récepteurs membranaires, et une durée de vie plus longue que les lymphocytes naïfs, sont produites lors de cette réponse primaire. La capacité de cette mémoire immunitaire peut être observée grâce à l’étude du taux des anticorps plasmatiques (Figure 14). Lors de cette réponse dite primaire, un délai de plusieurs jours est nécessaire avant que ces anticorps n’apparaissent. Ce délai correspond au temps nécessaire à la reconnaissance de l’antigène, l’activation, la multiplication et la dif-férenciation des LB en plasmocytes sécréteurs. Le taux d’anticorps passe par un maximum puis diminue ensuite pour devenir pratiquement nul au bout de quelques semaines. Cette disparition des anticorps circulants et des plasmocytes qui les sécrètent est due au fait que les plasmocytes sont des cellules à durée de vie courte et que les anticorps sont soumis au catabolisme. Une rencontre ultérieure avec le même antigène induit une réponse immuni-taire dite secondaire qui, dans le cas des anticorps, se traduit par une réponse plus rapide, et plus ample : la quantité d’anticorps produite est beaucoup plus importante, et le taux d’anticorps circulants diminue plus lentement que lors de la réponse primaire. Ainsi, une fois que le système immunitaire a reconnu un antigène, et répondu à ce dernier, il présente une mémoire immunitaire, c’est-à-dire qu’une rencontre ultérieure avec le même antigène induit un état augmenté de réactivité immunitaire.
BAH2_ch13.indd 260 28/02/2014 11:29

261
Immunologie
( anti-antigène 2)
1 2
Taux plasmatique d’anticorps ( anti-antigène 1)
Premier contactavec l’antigène 1
temps (jours)100
Deuxième contact avec l’antigène 1et premier contact avec l’antigène 2
20 30 40 50 60 70 80 90 100
1 Délai 1
2 Délai 2
Figure 14 Le taux d’anticorps plasmatiques
lors des réponses primaire et secondaire
2. LavaccinationCette propriété de mémoire du système immunitaire a donc été mise à profi t par l’Homme lors de la vaccination. Il s’agit d’induire une réponse primaire spécifi que grâce à l’utilisation d’un antigène (normalement pathogène) dénué de son pouvoir pathogène, afi n de faire pro-duire à l’individu vacciné une mémoire immunitaire spécifi que, qui lors d’un contact ultérieur avec le même antigène, mais cette fois-ci pathogène, lui permettra de se défendre effi cace-ment. Cette technique est donc une méthode acquise activement, puisque c’est l’individu, lui-même, qui produit l’immunité. Cette dernière est lente à apparaître, mais elle est durable. L’intérêt est donc préventif.
On distingue quatre types de vaccin :
¾¾ Ceux contenant des microbes (virus ou bactéries) inactivés ou tués. C’est le cas par exemple, pour les vaccins contre la poliomyélite et le choléra ;
¾¾ Ceux contenant des antigènes microbiens. C’est le cas par exemple, pour les vaccins contre la coqueluche, la grippe et l’hépatite B ;
¾¾ Ceux contenant des microbes vivants atténués, c’est-à-dire des microbes voisins du microbe pathogène. C’est le cas par exemple, pour les vaccins contre les oreillons, la rougeole, la rubéole et la varicelle ;
¾¾ Ceux contenant des anatoxines (toxines traitées pour être non pathogènes). C’est le cas par exemple, pour les vaccins contre la diphtérie ou le tétanos.
Dans tous les cas, cette introduction provoque une réaction immunitaire de type primaire pour l’organisme et permet la production de cellules à mémoire spécifi ques. Un vaccin n’est effi cace que s’il contient la plupart des antigènes du micro-organisme. Cette réponse primaire doit donc la plupart du temps être renforcée par un (ou des) rappel(s). À la suite de la vaccination, une exposition de l’organisme au micro-organisme provoque une réponse immunitaire de type secondaire, plus rapide et plus effi cace que la réponse primaire, qui élimine avec succès le micro-organisme pathogène.
BAH2_ch13.indd 261 28/02/2014 11:29

262
Anatomie – physiologie
Pour la population générale, certaines vaccinations sont obligatoires :
¾¾ contre la diphtérie et le tétanos : seule la primo-vaccination avec le premier rappel à 18 mois sont obligatoires ;
¾¾ contre la poliomyélite : la primo-vaccination et les rappels sont obligatoires jusqu’à l’âge de 13 ans ;
¾¾ contre la fièvre jaune : pour toutes les personnes résidant en Guyane.
D’autres vaccinations sont fortement recommandées par les autorités sanitaires :
¾¾ contre la coqueluche,
¾¾ contre la tuberculose (BCG, bacille de Calmette et Guérin, bacille tuberculeux bovin non virulent pour l’Homme),
¾¾ contre l’hépatite B,
¾¾ vaccin ROR contre la rougeole, les oreillons et la rubéole,
¾¾ contre la grippe saisonnière chez les personnes âgées ou souffrant d’une maladie chro-nique.
Malheureusement, certaines maladies infectieuses ne sont pas encore prévenues par la vac-cination. C’est le cas, entre autres, pour le SIDA, l’hépatite C et le paludisme.
3. LasérothérapieIl s’agit de protéger un individu contre une maladie infectieuse donnée, lorsqu’on le sait, ou le suppose contaminé, en lui administrant directement des anticorps fabriqués par un autre organisme (un Homme ou un animal). Elle est donc utilisée à titre curatif. L’immunité est acquise passivement, puisque l’individu ne la produit pas. Elle est immédiate, mais de courte durée (un à trois mois). Cette méthode est notamment utilisée dans le traitement d’hépatites virales ou du tétanos.
B. ImmunothérapieUtilisés en cancérologie, les traitements par immunothérapie n’agissent pas directement sur les cellules cancéreuses. Ils ciblent le système immunitaire, et permettent de le stimuler pour renforcer son action sur les cellules cancéreuses. Il existe deux approches thérapeu-tiques d’immunothérapie :
¾¾ L’immunothérapie passive : elle consiste à administrer au malade des anticorps mono-clonaux, anticorps qui ne reconnaissent qu’un seul type d’épitope. Ils sont destinés à viser une cible moléculaire précise, présente de façon spécifique à la surface des cel-lules cancéreuses. Ces anticorps peuvent également être utilisés pour cibler des facteurs nécessaires à la multiplication des cellules cancéreuses, dont on bloque ainsi la proliféra-tion. Ce type d’immunothérapie est actuellement utilisé dans certains cas de cancers du sein, et pour certains lymphomes ;
¾¾ L’immunothérapie active : notre système immunitaire est capable de détruire les cellules cancéreuses. Malheureusement, cela ne fonctionne pas toujours assez bien pour contrer la formation d’une tumeur, soit parce que ces défenses immunitaires sont affaiblies, soit parce que la tumeur est suffisamment « furtive » pour ne pas être détectée par le système de surveillance de l’organisme. Le principe de l’immunothérapie active est d’améliorer le fonctionnement du système immunitaire en agissant sur l’un ou l’autre de ces deux aspects. L’immunothérapie active se pratique selon deux modalités :
– L’immunothérapie non spécifique : cette approche vise à stimuler l’activité globale du système immunitaire, sans cibler la tumeur particulière d’un malade. On utilise pour cela
BAH2_ch13.indd 262 28/02/2014 11:29

263
Immunologie
les cytokines (l’interleukine 2, par exemple) qui vont stimuler la prolifération des cellules immunitaires. Ce type d’immunothérapie est actuellement utilisé dans le cas de cancers du rein, les mélanomes et certains lymphomes ;
– L’immunothérapie spécifique : cette stratégie thérapeutique consiste à prélever, à mettre en culture et à manipuler au laboratoire les cellules tumorales ou les cellules immuni-taires du malade avant de les lui réinjecter. Dans le premier cas, on cherche à rendre les cellules tumorales plus immunogènes. Dans le second cas, on cherche à stimuler les cellules immunitaires pour les rendre plus agressives et plus efficaces pour détruire la tumeur. Dans les deux cas, les cellules sont modifiées par génie génétique (voir livre Biologie de l’alimentation humaine, tome I). Jusqu’à présent, cette approche de thérapies cellulaires est encore au stade expérimental.
Actuellement, l’immunothérapie est de plus en plus utilisée. Néanmoins, elle est encore prescrite en complément des autres traitements : chirurgie, chimiothérapie, et radiothérapie.
C. Transplantationd’organeUne greffe ou transplantation est une opération chirurgicale consistant à remplacer un organe malade par un organe sain, appelé greffon et provenant d’un donneur. Ce dernier doit avoir un groupe sanguin compatible avec celui du receveur. La compatibilité CMH entre don-neur et receveur est également nécessaire. Une fois greffé, l’organisme du receveur consi-dère le nouvel organe comme un objet étranger. Le malade devra donc suivre un traitement immunosuppresseur pour éviter le rejet du greffon, c’est-à-dire déclencher des réactions immunitaires visant à éliminer ce greffon. On distingue quatre types de greffes :
¾¾ L’autogreffe : greffe où donneur et receveur sont la même personne. Le taux de réussite de ce type de greffe est maximal, étant donné que les complexes majeurs d’histocompa-tibilité (CMH) du donneur et du receveur sont les mêmes. Aucune réaction immunitaire n’est déclenchée ;
¾¾ L’isogreffe : greffe où le donneur est le vrai jumeau (ou monozygote) du receveur. Le pour-centage de réussite est maximum. En effet, issu d’une même cellule œuf, les jumeaux monozygotes possèdent le même complexe majeur d’histocompatibilité (CMH) ;
¾¾ L’allogreffe : il s’agit des greffes les plus courantes. Ce sont des greffes entre deux indivi-dus distincts. Donneur et receveur possèdent des complexes majeurs d’histocompatibi-lité (CMH) différents, mais le plus proches possible. Dans ces cas, la greffe s’accompagne d’un traitement immunosuppresseur. Plus les CMH sont ressemblants, plus la greffe a des chances de réussite ;
¾¾ La xénogreffe ou hétérogreffe : le donneur est d’une espèce biologique différente de celle du receveur. Le porc est le meilleur animal donneur d’organes pour l’humain.
D. LeshybridomesetlesanticorpsmonoclonauxLes anticorps monoclonaux ont de multiples applications en recherche, en diagnostic, et en clinique humaine, exploitant leur spécificité de reconnaissance. Ils sont par exemple, utilisés dans les tests d’ovulation ou de grossesse, en immunothérapie, ou dans les tests ELISA (voir livre Biologie de l’alimentation humaine, tome I). Afin de les produire, on utilise des hybridomes.
Un hybridome est une cellule qui provient de l’hybridation (fusion) entre un clone de lym-phocyte B normale de mammifères, et une cellule cancéreuse de myélome. L’intérêt est de cumuler les propriétés de ces deux cellules, c’est-à-dire la production spécifique d’anti-corps monoclonaux pour le lymphocyte, et l’immortalité (multiplication infinie) pour la cellule
BAH2_ch13.indd 263 28/02/2014 11:29

264
Anatomie – physiologie
cancéreuse. Les hybridomes donnent donc des cellules immortalisées stables productrices d’anticorps monoclonaux.
E. LasérologieLa sérologie est l’étude du sérum. Elle consiste à évaluer l’immunité vis-à-vis d’une patholo-gie en détectant ou en mesurant la quantité d’anticorps spécifiques de celle-ci. Elle fait appel à des techniques variées comme l’immunofluorescence ou l’ELISA (voir livre Biologie de l’alimentation humaine, tome I). Dans le cadre médical, elle permet donc de poser un séro-diagnostic, c’est-à-dire déterminer le statut sérologique d’un individu vis-à-vis d’une maladie infectieuse. Un individu est dit « séropositif » pour un antigène lorsqu’il présente dans son sérum des anticorps spécifiques de cet antigène, révélant qu’il y a, ou qu’il y a eu, une RIMH spécifique de celui-ci et donc, qu’il est, ou a été, atteint par cette pathologie. Inversement, un individu est dit « séronégatif » pour un antigène lorsqu’il ne présente pas dans son sérum d’anticorps spécifiques de cet antigène, révélant qu’il n’est pas atteint par cette pathologie.
Voici quelques exemples d’utilisation médicale de la sérologie :
¾¾ Dans le diagnostic des maladies auto-immunes : les auto-anticorps sont recherchés ;
¾¾ Dans le diagnostic d’une infection par le VIH : on recherche la présence d’anticorps anti-VIH via un test ELISA en première intention, puis on confirme leur présence avec un wes-tern blot (voir livre Biologie de l’alimentation humaine, tome I) en deuxième intention ;
¾¾ Dans le diagnostic de la toxoplasme et de la rubéole : ces diagnostics concernent plus particulièrement la femme enceinte, car ces 2 maladies infectieuses peuvent causer de graves malformations chez l’enfant. On va donc, chez la femme enceinte, déterminer son statut sérologique vis-à-vis de ces deux infections.
BAH2_ch13.indd 264 28/02/2014 11:29

265
Immunologie
testersesconnaIssances
Exercice1Relever les propositions exactes pour chaque QCM.
Q À propos des marqueurs de surface définissant le soi :
A les molécules HLA de classe I sont uniquement présentes à la surface des hématies.
B les molécules HLA de classe II sont présentes à la surface de toutes les cellules nuclées.
C les molécules HLA sont codées par les gènes A, B, C, DP, DQ et DR.
D le système rhésus est un système mineur du soi biologique.
W À propos des organes lymphoïdes :
A le thymus et la moelle épinière sont des organes lymphoïdes centraux.
B la maturation des lymphocytes T se produit dans le thymus.
C la production des lymphocytes a lieu dans la moelle osseuse rouge.
D l’anneau de Waldeyer, constitué des végétations, des amygdales et des ganglions lym-phatiques, fait partie des organes lymphoïdes centraux.
E À propos des cellules immunitaires :
A on distingue trois types de polynucléaires dont la dénomination est basée sur leur affinité à absorber des colorants histologiques.
B les granulocytes neutrophiles et les mastocytes ont des rôles similaires.
C les monocytes, les macrophages et les polynucléaires éosinophiles sont des cellules phagocytaires, et sont donc riches en lysosomes.
D les lymphocytes représentent entre 25 et 35 % des leucocytes.
R À propos des cellules immunitaires :
A les lymphocytes T4 agissent en tant qu’auxiliaires dans les réponses immunitaires spécifiques.
B les lymphocytes T4 n’ont pas de rôle direct dans la destruction du non soi.
C les lymphocytes T cytotoxiques présentent à leur surface des récepteurs T et des mar-queurs CD8.
D les lymphocytes B présentent à leur surface des anticorps membranaires.
T À propos des récepteurs membranaires :
A les anticorps membranaires des LB possèdent chacun un seul paratope, spécialisé dans la reconnaissance de l’antigène.
B à la surface d’un LB, on peut trouver des immunoglobulines de classe D.
C les récepteurs T ont une double reconnaissance : celle de l’antigène et celle du CMH.
D les récepteurs T des LT4 reconnaissent les molécules HLA de classe I.
BAH2_ch13.indd 265 28/02/2014 11:29

266
Anatomie – physiologie
Y À propos de l’immunocompétence et du répertoire immunologique :
A un macrophage est immunocompétent lorsqu’il présente à sa surface des anticorps spécifiques d’un antigène donné.
B les lymphocytes autoréactifs reconnaissent le non soi comme du soi.
C les lymphocytes B et T acquièrent leur immunocompétence dans la moelle osseuse rouge.
D le répertoire immunologique d’un individu est défini comme l’ensemble des lympho-cytes immunocompétents existants au sein de son organisme.
U À propos des réponses immunitaires :
A la RIMH et la RIMC font partie des moyens de défenses non spécifiques à un germe donné.
B la barrière cutanéo-muqueuse constitue le premier moyen de défenses contre le monde extérieur.
C les flores commensales de la peau et des muqueuses empêchent la croissance des germes pathogènes.
D l’acidité des sucs sécrétés par certaines muqueuses empêche le développement des germes pathogènes.
I À propos des réponses immunitaires :
A la réaction inflammatoire comprend trois étapes : une étape vasculaire, une étape cellulaire et une étape de réparation du tissu lésé.
B lors de la digestion intracellulaire durant la phagocytose, les lysosomes secondaires fusionnent avec le phagosome pour former une vacuole digestive.
C certaines protéines du complément sont des opsonines, c’est-à-dire qu’elles facilitent la phagocytose.
D les phagocytes sont des cellules de l’immunité adaptative.
O À propos des réponses immunitaires :
A la phase effectrice de la RIMC comprend la phagocytose des complexes immuns et, éventuellement l’intervention du complément.
B lors de la phase d’induction de la RIMC, un clone de LT8 et un clone de LT4 sont sélec-tionnés par le même épitope.
C le TCR d’un lymphocyte T4 et le co-récepteur CD4 reconnaissent les molécules HLA de classe II.
D la cytolyse d’une cellule infectée par l’intermédiaire de perforines est un mécanisme de la phase d’amplification de la RIMC.
P À propos des allergies :
A l’hypersensibilité retardée résulte d’une production excessive d’immunoglobulines E.
B l’hypersensibilité immédiate résulte d’une production excessive de cytokines.
C lors d’une hypersensibilité retardée, lors du second contact avec l’allergène, les lym-phocytes mémoires vont libérer des interleukines en quantité excessive.
D lors d’une hypersensibilité immédiate, lors du premier contact avec l’allergène, des immunoglobulines E vont aller se fixer sur la membrane des polynuclaires basophiles et des mastocytes.
BAH2_ch13.indd 266 28/02/2014 11:29

267
Immunologie
{ À propos des déficits immunitaires :
A un déficit immunitaire congénital peut se guérir en faisant une greffe de moelle épi-nière HLA-compatible.
B le sigle SIDA signifie « syndrome d’Immunodéficience acquise ».
C lors de la phase asymptomatique d’une infection par le VIH, les ganglions lympha-tiques sont en général hypertrophiés, montrant la présence de réponses immunitaires.
D lors de la phase SIDA déclaré de l’infection par le VIH, les maladies opportunistes apparaissent.
} À propos des myélomes et des lymphomes :
A la maladie de Kahler se caractérise par une prolifération anormale de macrophages.
B la présence de cellules de Reed-Sternberg caractérise un lymphome non hodgkinien.
C pour un lymphome de Hodgkin au stade II, on préconise en général une radiothérapie seule.
D le lymphome de Burkitt est non hodgkinien, et se caractérise par une prolifération anormale des lymphocytes B.
q À propos de la vaccination :
A la vaccination confère une immunité acquise passivement, qui est lente à apparaître, mais durable.
B la primo-vaccination contre la diphtérie est obligatoire en France.
C la vaccination ROR est obligatoire en France.
D la vaccination contre la fièvre jaune est obligatoire pour toutes les personnes résidant en Guyane.
w À propos de l’immunothérapie :
A l’immunothérapie est utilisée en cancérologie, et permet de renforcer l’action du sys-tème immunitaire sur les cellules cancéreuses.
B l’immunothérapie passive, qui utilise des anticorps monoclonaux, est utilisée dans certains cas de cancers du sein.
C l’immunothérapie active spécifique utilise le génie génétique.
D l’immunothérapie active non spécifique, qui utilise les cytokines, est utilisée dans cer-tains cas de cancers du sein.
Exercice2Définir les phrases suivantes à l’aide d’un mot ou d’une expression.
Q Étude permettant de déterminer le statut sérologique d’un individu vis-à-vis d’une maladie infectieuse.
W Cellule provenant de l’hybridation d’un clone de lymphocyte B normale de mammifères, et d’une cellule cancéreuse de myélome.
E Molécules produites par des hybridomes.
BAH2_ch13.indd 267 28/02/2014 11:29

CORR
IGÉ
268
Anatomie – physiologie
R Greffe entre deux individus de la même espèce, qui ont des CMH différents, mais relative-ment proches.
T Bacille tuberculeux bovin utilisé dans le vaccin contre la tuberculose.
Y Technique curative visant à protéger un individu contre une maladie infectieuse donnée, lorsqu’on le sait, ou le suppose contaminé, en lui administrant directement des anticorps fabriqués par un autre organisme.
U Lymphome non hodgkinien caractérisé par une prolifération anormale de lymphocyte T.
I Stade d’un lymphome de Hodgkin où le lymphome est propagé dans tout le système immu-nitaire.
O Première phase de l’infection par le VIH qui se manifeste par un syndrome pseudo-grippal.
P Maladie auto-immune touchant la gaine de myéline des fi bres nerveuses.
{ Pathologie du système immunitaire due à la rupture de la tolérance au soi.
} Mécanisme d’autodestruction cellulaire programmé génétiquement, et pouvant être déclen-ché dans la cellule cible d’un LTc.
q Cytokines stimulant la différenciation des lymphocytes B en plasmocytes.
w Conséquence de la formation d’un complexe d’attaque membranaire au niveau d’un élé-ment étranger de nature cellulaire.
Exercice3
D’après vos connaissances, remplissez le tableau suivant :
Groupe sanguin Marqueur Anticorps À qui donner ? De qui recevoir ?
A
B
AB
O
Exercice4
Schématisez la structure d’un anticorps.
BAH2_ch13.indd 268 28/02/2014 11:29