Embed Size (px)
Citation preview

Impacts sur la qualité de la viande suite à l’alimentation des veaux de grain en période de croissance et de
finition par des sous-produits du Québec
Mémoire
Alex O’Bomsawin-Descôteaux
Maîtrise en sciences animale
Maître ès sciences (M.Sc.)
Québec, Canada
© Alex O’Bomsawin-Descôteaux, 2016

ii

iii
Résumé court
Ce projet visait à évaluer les effets du remplacement partiel ou total du maïs (M) par de
l’orge roulée (B), des drèches sèches de distillerie avec solubles (D) ou du tourteau de
canola (C) sur la qualité de la viande de veau de grain. Huit muscles longissimus dorsi
gauche ont été pris pour chacun des traitements étant assignés aléatoirement : M-
suppléments protéiques (S), BCD, MCD et MSD et donné à volonté. Toutes les rations
étaient isoprotéiques. Des différences significatives entre M et B (P < 0.05) ont été
obtenues pour le pH ultime, la couleur (L*, a* et b*), la force de cisaillement, la longueur
des sarcomères, les pertes à la cuisson, la concentration de malonaldéhyde au jour 7, la
teneur en gras total et des flaveurs indésirables au niveau des analyses sensorielles. La
différence obtenue lors de la force de cisaillement n’était pas perceptible par les panélistes.

iv

v
Table des matières
Résumé court ................................................................................................................................. iii
Table des matières ...........................................................................................................................v
Liste des tableaux .......................................................................................................................... vii
Liste des figures ............................................................................................................................. ix
Remerciements ............................................................................................................................... xi
Avant-propos ............................................................................................................................... xiii
CHAPITRE 1. INTRODUCTION ...................................................................................................1
CHAPITRE 2. REVUE DES TRAVAUX ANTÉRIEURS .............................................................3
2.1. LE MUSCLE ............................................................................................................................3
- Structure générale ................................................................................................................3
- Type de fibres musculaires et fonctionnalités ......................................................................4
- Transformation du muscle en viande ...................................................................................6
2.2. LA QUALITÉ DE LA VIANDE ............................................................................................10
2.2.1. Le pH .....................................................................................................................11
2.2.2. La couleur de la viande ..........................................................................................14
- La myoglobine ..........................................................................................................16
2.2.3. La tendreté .............................................................................................................21
- La longueur des sarcomères .....................................................................................26
2.2.4. Propriétés organoleptiques ....................................................................................30
2.2.5. Analyses sensorielles .............................................................................................36
2.3. ALIMENTATION ..................................................................................................................40
2.3.1. Les céréales .......................................................................................................40
2.3.2. Les sous-produits ..............................................................................................44
CONCLUSION ..............................................................................................................................53
2.5. LISTE DES OUVRAGES CITÉS ..........................................................................................55
CHAPITRE 3. Effect of dietary replacing corn on carcass and meat quality in grain fed
calves ......................................................................................................................61
3.1. ABSTRACT ............................................................................................................................63
3.2. RÉSUMÉ ................................................................................................................................65
3.3. INTRODUCTION ..................................................................................................................67
3.4. MATERIALS and METHODS ..............................................................................................69

vi
3.5. RESULTS and DISCUSSIONS ............................................................................................. 75
3.5.1. Ultimate pH, meat colour and marbling ........................................................... 75
3.5.2. Cooking losses, Warner-Bratzler shear force (WBSF) and Sarcomere
lenght ................................................................................................................ 77
3.5.3. TBA (malonaldehyde concentration) ............................................................... 79
3.5.4. Meat composition ............................................................................................. 80
3.5.5. Sensory analysis ............................................................................................... 81
3.6. CONCLUSION ...................................................................................................................... 83
3.7. ACKNOWLEDGMENTS ...................................................................................................... 85
3.8. REFERENCES ....................................................................................................................... 87

vii
Liste des tableaux
Chapitre 2
Tableau 2.1. Caractéristiques des principaux types de fibres musculaires .....................................5
Tableau 2.2. Composition des muscles Longissimus dorsi, Psoas major et Triceps brachii
capui laterale des carcasses de veaux (moyenne ± écart-type) ..................................6
Tableau 2.3. Valeurs moyennes pour les concentrations plasmatiques avant et après le
transport des veaux ...................................................................................................7
Tableau 2.4. Teneur en glycogène des muscles et du pH selon les manipulations pré-
abattage ......................................................................................................................9
Tableau 2.5. Effets du poids de la carcasse sur la fréquence des coupes sombres de boeuf .........17
Tableau 2.6. Mesures de la qualité de la viande sur 117 longes de porc de chaque classe de
qualité .......................................................................................................................20
Tableau 2.7. Évolution de la couleur selon les manipulations pré-abattage .................................21
Tableau 2.8. Force de cisaillement en fonction du vieillissement du muscle Longissimus
dorsi ......................................................................................................................23
Tableau 2.9. Force de cisaillement et pertes à la cuisson des steaks de bouvillons nourris
avec des rations contenant 0 ou 30% de drèches sèches de maïs avec solubles
en période de croissance et de finition......................................................................24
Tableau 2.10. L’effet de mélanger les groupes de verrats et de cochettes avant le transport
et en stabulation sur deux indicateurs plasmatiques de stress ................................25
Tableau 2.11.Force de cisaillement du muscle Longissimus dorsi de veau avec différentes
méthodes de cuisson .................................................................................................26
Tableau 2.12.Moyennes des longueurs de sarcomère (µm) provenant de différents muscles
de bœuf et selon la méthode de suspension de la carcasse .......................................28
Tableau 2.13. Effets des traitements de refroidissement et du vieillissement sur le Warner-
Bratzler et la longueur des sarcomères chez l’agneau de lait ...................................29
Tableau 2.14. Effet de la quantité de gras dans la viande hachée sur les qualités
organoleptiques .......................................................................................................31
Tableau 2.15. Composition chimique de la viande de veau, % .....................................................32
Tableau 2.16. Capacité de rétention d’eau (CRE) et pertes à la cuisson (PC) selon les
manipulations pré-abattage de jeunes taureaux Holstein Friesian durant le
vieillissement de la viande ........................................................................................33
Tableau 2.17. Pertes à la cuisson et caractéristiques sensorielles des steaks de bouvillons
nourris avec des rations contenant 0 ou 30% de drèches sèches de distillerie
avec solubles en période de croissance et de finition. ...........................................37

viii
Tableau 2.18. Notes distribuées pour la tendreté sensorielle auprès de cinq catégories de
force de cisaillement (FC) ........................................................................................ 38
Tableau 2.19. Notes distribuées pour la tendreté sensorielle auprès de trois catégories de
force de cisaillement (FC) ........................................................................................ 39
Tableau 2.20. Caractéristiques sensorielles selon les manipulations pré-abattage durant le
vieillissement de la viande ....................................................................................... 39
Tableau 2.21. Évaluation des sous-produits .................................................................................. 41
Tableau 2.22. Effets du maïs sur les ratios d’orge dans la ration sur les caractéristiques de la
carcasse des veaux de boucherie nourris aux grains ................................................ 42
Tableau 2.23. Digestibilité des nutriments (%) du maïs entier, de l’orge ronde et de l’orge
roulé ....................................................................................................................... 43
Tableau 2.24. Répartition de l’énergie des veaux nourris avec du maïs entier, de l’orge
ronde et de l’orge roulé ............................................................................................ 43
Tableau 2.25. Comparaison des acides aminés des sous-produits (par rapport aux protéines
du lait) ...................................................................................................................... 45
Tableau 2.26. Fractions de protéines et digestibilité (%) de sept sous-produits de
transformation du maïs ............................................................................................ 47
Tableau 2.27. Effets des suppléments sur les performances des bouvillons Hereford aux
pâturages de blé d’hiver ........................................................................................... 48
Tableau 2.28. Effets des drèches de distillerie sur la couleur objective et les pointages sur
l’apparence subjective de steaks après une simulation d’étalement sur étagère
de 138 heures (Expérience 1 et 2) ............................................................................ 50
Tableau 2.29. Effets des drèches de distillerie sur la force de cisaillement (Expérience 1 et
2) .............................................................................................................................. 51
Tableau 2.30. Effets des drèches de distillerie sur les paramètres sensoriels (Expérience 1 et
2) ......................................................................................................................... 52
Chapitre 3
Table 3.1. Ultimate pH, meat colour and marbling of longissimus dorsi from Holstein
grain-fed calves ........................................................................................................ 91
Table 3.2. Cooking losses, Warner-Bratzler shear force (WBSF) and Sarcomere lenght of
longissimus dorsi from Holstein grain-fed calves ................................................... 92
Table 3.3. Malonaldehyde (TBA) at d0 and d7 from longissimus dorsi Holstein grain-fed
calves ........................................................................................................................ 93
Table 3.4. Meat composition of longissimus dorsi from Holstein grain-fed calves ................. 94
Table 3.5. Sensory analysis of longissimus dorsi from Holstein grain-fed calves ................... 95

ix
Liste des figures
Chapitre 2
Figure 2.1. Structure d’un muscle strié .........................................................................................4
Figure 2.2. Évolution du taux de glycogène musculaire de taurillons après un stress .................8
Figure 2.3. Relations entre le métabolisme énergétique musculaire et les caractéristiques
musculaires déterminant la qualité organoleptique de la viande ..............................10
Figure 2.4. Relation entre le potentiel glycolytique (PG) 48 heures post-mortem et le pH
ultime dans le muscle longissimus dorsi des carcasses de veaux. PG, µmol
d’acide lactique g-1....................................................................................................12
Figure 2.5. Activité de la cathepsine B dans les échantillons de viande de bœuf vieilli à -
1°C jusqu’à 28 jours post mortem à pH ultime bas et élevé ....................................14
Figure 2.6. Les trois formes chimiques de la myoglobine ..........................................................18
Figure 2.7. Évolution du pH musculaire et de la couleur de la viande après l’abattage .............19
Figure 2.8. Force de cisaillement de la viande de bœuf (n=63) dépendamment du pH
ultime durant le vieillissement ..................................................................................22
Figure 2.9. Mouvement des myofilaments d’actine et de myosine lors de la contraction et
de la relaxation des muscles .....................................................................................27
Figure 2.10. Effets du conditionnement sur l’oxydation des gras (TBARS) durant
l’entreposage de la viande de veau ...........................................................................31
Figure 2.11. Effets du conditionnement sur la capacité de rétention d’eau durant
l’entreposage de la viande de veau ...........................................................................34
Figure 2.12. Effets du conditionnement sur les pertes en eau durant l’entreposage de la
viande de veau ..........................................................................................................35
Figure 2.13. Échelle canadienne de persillage de la viande de bœuf............................................37

x

xi
Remerciements
Je tiens à remercier sincèrement l’Union des producteurs agricoles (UPA) et plus
précisément la Fédération des producteurs de bovins du Québec (FPBQ) pour le lancement
de ce projet. Merci également pour l’attribution de la bourse durant mes études. Je remercie
également les producteurs de veau de grain du Québec pour leur implication et leur écoute
lors de la divulgation des résultats. Vous avez permis la création de ce projet et la
continuité afin de permettre l’implantation dans les entreprises.
Je remercie la Ministère de l’agriculture, des pêcheries et de l’alimentation du Québec
(MAPAQ) pour le financement de ce projet et le support constant de mes collègues de
bureau. Je remercie Agriculture et Agroalimentaire Canada (AAC) – Centre de recherche
et de développement sur les aliments (CRDA) pour l’utilisation des locaux, matériaux et la
superbe équipe m’ayant aidé grandement lors des analyses. Claude G., Jacinthe, Nancy et
Claude L., je vous remercie sincèrement pour tout le temps, le travail et le partage de
connaissances que vous m’avez accordé. Merci Claude L. pour toutes les techniques et
façons de travail avec la viande. Merci Claude G. pour le challenge de connaissances et des
réponses aux questions que j’avais à n’importe quel moment. Merci à Jacinthe et Nancy
pour les analyses sensorielles, le temps passé avec vous dans les cuisines m’a fait sentir
comme à la maison. Je remercie également tous ceux et celles que j’ai pu côtoyer. Vous
m’avez fait vivre une expérience mémorable et agréable.
Merci au Centre de développement du porc du Québec (CDPQ) pour toutes les prises de
données et l’aide technique pour le projet. Merci également à Délimax pour les animaux,
les bâtiments pour la recherche et le personnel fortement impliqué durant la phase animale.
Je remercie l’Abattoir de Saint-Germain et Montpak pour l’abattage et la découpe des
veaux respectivement.
Je remercie également Younès Chorfi de la Faculté de médecine vétérinaire dans son appui
lors de la rédaction de l’article scientifique.

xii
Forts remerciements à mon directeur de maîtrise Dany. Merci beaucoup pour le temps
consacré à régulariser la situation avec le comité d’éthique de l’université. Merci pour le
temps donné aux rencontres pour la rédaction, de la présentation aux producteurs et du
support durant mes années d’études. Merci d’avoir cru en moi en m’accordant la chance de
faire ce projet avec toi. Ton expérience et tes connaissances ont été très utiles.
Je remercie ma femme pour sa patience et sa compréhension pour le temps que j’ai mis à
produire ce document et pour tous les moments où j’étais à l’extérieur. Je t’aime mon
amour et merci encore!
Finalement, je tiens spécialement à remercier ma fille Ophélie qui nous apporte beaucoup
depuis sa naissance le 8 janvier 2015. Petite puce, tu nous fais vivre beaucoup de bonheur
et d’émotions depuis que tu t’es pointée la tête. Tu es remplie de surprises, et ce, à tous les
jours. Papa t’aime beaucoup!

xiii
Avant-propos
Ce mémoire contient un chapitre rédigé en anglais sous forme d’article scientifique dont je
suis l’auteur principal. Les coauteurs sont les chercheurs D. Cinq-Mars, C. Gariépy et Y.
Chorfi. L’article ayant pour titre « Effect of dietary replacing corn on carcass and meat
quality in grain fed calves » sera envoyé pour publication dans la revue « Canadian Journal
of Animal Science ».


1
CHAPITRE 1.
INTRODUCTION
La production de veau de grain débute avec un veau ayant une alimentation à base de
lactoremplaceur pour les six premières semaines. Par la suite, l’animal est nourri
exclusivement de maïs-grain et de suppléments protéiques jusqu’à son abattage (Ngapo et
Gariépy. 2006). La viande de veau de grain est reconnue pour être produite au Québec et en
Ontario (Campbell et al, 2013). C’est au Québec que cette production s’est premièrement
implantée à la fin des années 70 (CPAQ, 1999). Le développement de cette activité s’est
produit dans le but de rentabiliser les veaux Holstein des entreprises laitières puisqu’ils sont
aptes à l’engraissement (CPAQ, 1999). Le prix faible des aliments et leurs quantités
disponibles tels que le maïs-grain et la poudre de lait écrémé ont permis également le
lancement de cette production (CPAQ, 1999).
En quinze ans, soit entre 1983 et 1998, la production québécoise de veaux lourds (incluant
le veau de lait et le veau de grain) a connu une augmentation de 402% soit de 43 000 à
173 000 têtes. L’augmentation connue plaçait alors le Québec au premier rang de la
production de veaux lourds canadienne (CPAQ, 1999). En 1998, cette production a généré
25,45 millions de kilogrammes de viande représentant une valeur de 108 millions de
dollars. En 2000, le Québec représentait 62% des abattages canadiens de veaux et l’Ontario
37% (Fornasier, 2002). En 2013, il s’est produit environ 70 000 veaux de grain ayant
générés des sommes de plus de 53 millions de dollars. En date du 1er octobre 2014, le prix
actuel est de 3,075$ par livre soit 6,77$ par kilogramme de viande produite (FPBQ, 2014).
La production québécoise se concentre principalement dans trois grandes régions détenant
plus de 75% du cheptel soient de Saint-Hyacinthe, Centre-du-Québec et Québec (CPAQ,
1999). La concentration dans ces régions est reliée à la proximité et à l’accès aux services
pour l’ensemble des besoins de la production (Fornasier, 2002). Dans la province, la filière

2
veau lourd occupe plus de 4 600 emplois. La production de veau de grain est effectuée par
plus de 160 producteurs (FPBQ, 2014). En 2012, la production de veau de grain québécoise
était intégrée verticalement à un niveau de 47% (Royer et Vézina, 2012).
Au Québec, un programme de certification à la ferme obligatoire encadre les producteurs.
Ces derniers doivent suivre un cahier des charges décrivant les normes et réglementations
de la production telles que la qualité du produit, la régie d’élevage, l’alimentation, la
traçabilité et le bien-être des veaux. Il est important de noter que depuis avril 2004, aucune
hormone de croissance n’est donnée aux veaux suivant lecahier des charges (FPBQ, 2014).
Depuis 2002, la traçabilité permet de suivre le chemin que l’animal parcourt de la ferme
jusqu’aux tablettes. Cela permet d’assurer une surveillance sanitaire tout au long de la vie
de l’animal jusqu’à son abattage (Veau de grain du Québec certifié, 2014). Le Veau de
grain du Québec certifié assure également un produit uniforme pour les consommateurs
(FPBQ, 2014).
La viande de veau de grain se démarque par sa qualité au niveau de sa tendreté et de sa
saveur. C’est ce qui fait d’elle un produit de plus en plus recherché par les consommateurs.
Ces derniers recherchent une viande étant d’une haute qualité. De plus, elle doit être tendre,
juteuse et savoureuse (Leupp et al. 2009). Elle est considérée également comme une viande
extra-maigre (Veau de grain du Québec certifié, 2014).
L’objectif de cette étude est d’évaluer les effets d’un remplacement partiel ou total du maïs
par de l’orge roulée, des drèches sèches de distillerie avec solubles ou du tourteau de canola
sur la qualité de la viande de veaux de grain. Les rations sont comparés à l’alimentation de
base retrouvée dans la production soit maïs-grain et suppléments de protéines.
Le remplacement ou la substitution du maïs pourrait ne pas avoir d’effet sur la qualité de la
viande. Donc, produire une viande avec des paramètres similaires ou même en améliorer la
qualité représente également un élément à vérifier.

3
CHAPITRE 2.
REVUE DES TRAVAUX ANTÉRIEURS
2.1. LE MUSCLE
Avant de subir les processus de transformation, la viande est issue d’un muscle. Ce dernier
est majoritairement composé d’eau et de protéines soit environ 75% et 20% respectivement
(Hocquette et al. 2000). Les teneurs en lipides et en glucides des muscles squelettiques ne
représentent qu’un faible pourcentage soit 1 à 5 % et 1 à 2% respectivement (Hocquette et
al. 2000). Le muscle est également composé par de nombreux éléments importants
correspondant à un rassemblement de milliers de fibres musculaires étant l’unité structurale
du muscle. Elles sont des cellules plurinucléées et composées de faisceaux de myofibrilles
(Hocquette et al. 2000). À l’intérieur de ces myofibrilles se retrouvent les deux types de
myofilaments étant l’actine et la myosine (Gregory, 1998). Nous retrouvons les
mitochondries intermyofibrillaires et subsarcolemmales entre ces myofibrilles et à la
périphérie des cellules musculaires respectivement (Hocquette et al. 2000). Une grande
partie de l’énergie libre (ATP) est produite par ces organites intracellulaires étant
fondamental au muscle (Hocquette et al. 2000). Chacune des fibres musculaires est
entourée par une première couche de tissu conjonctif : l’endomysium. Les fibres sont
regroupées en petits groupes pour former des faisceaux. Ces derniers sont eux, également
recouverts d’une enveloppe plus épaisse de tissu conjonctif, soit le périmysium. C’est dans
cette couche que nous retrouvons des vaisseaux sanguins et des nerfs. Les regroupements
de faisceaux sont entourés d’une dernière couche de tissu conjonctif nommé l’épimysium
(Figure 2.1). Les muscles ne s’attachent pas directement aux os. Pour ce faire, le tendon est
la structure servant d’attache. Il est composé d’une grande quantité de tissu conjonctif
(Lawrie, 1998). Les muscles servent à la thermogénèse (production de chaleur), au
maintien de la posture et de l’activité physique (Hocquette et al. 2000).

4
Figure 2.1. Structure d’un muscle strié
Source : Marieb et Laurendeau, 1993
Dans le muscle, il existe trois types de fibres musculaires étant classées de façon
distinctives. Chacune d’elle participe de façon différente aux activités musculaires. Le
classement se fait principalement par la couleur des fibres, la vitesse de contraction et la
source énergétique consommée (Hocquette et al. 2000; INTERBEV, 2006). Nous
retrouvons donc les fibres βR (ou type I), rouges à contraction lente, αR (ou type IIA),
rouges à contraction rapide, et αW (ou type IIB), blanches à contraction rapide. Selon
l’environnement et l’utilisation des muscles, le type de fibres peut varier seulement dans
cette séquence : βR ↔ αR ↔ αW (I ↔ IIA ↔ IIB). Les fibres βR et αR sont de couleur
rouge puisque leur teneur en myoglobine est élevée et qu’elles ont une vascularisation
importante contrairement aux fibres αW. Pour les fibres βR, la teneur en glycogène est
faible, c’est la raison pour laquelle le métabolisme énergétique de ces fibres est oxydatif.
Plus les muscles ont de fibres βR (ou type I), plus ils contiennent de lipides (Hocquette et
al. 2000). Les fibres βR et αR ont une forte résistance à la fatigue contrairement aux fibres
αW. Les fibres αW sont l’opposé des fibres βR. Leur teneur en myoglobine, la
vascularisation et la teneur en lipides sont faibles. Puisque les fibres αW ont une forte
teneur en glycogène, le métabolisme énergétique de ces fibres est glycolytique. Les fibres

5
αR sont en quelque sorte l’intermédiaire des fibres βR et αW. C’est la raison pour laquelle
leur métabolisme énergétique se situe également à l’intermédiaire des fibres βR et αW, soit
oxydo-glycolytique (Tableau 2.1).
Tableau 2.1 : Caractéristiques des principaux types de fibres musculaires Adapté de
INTERBEV (2006)
Classification βR (type I) αR (type IIA) αW (type IIB)
Couleur Rouge Rouge Blanche
Teneur en myoglobine Forte Forte Faible
Vascularisation Importante Importante Faible
Teneur en lipides Forte Forte Faible
Aire de section Petite Petite Grande
Résistance à la fatigue Forte Forte Faible
Vitesse de contraction Lente Rapide Rapide
Teneur en glycogène Faible Forte Forte
Métabolisme énergétique Oxydatif Oxydo-glycolytique Glycolytique
La composition des muscles n’est pas la même pour l’organisme. Selon le muscle, la
proportion de fibres βR, αR et αW (type I, IIA et IIB), la concentration en pigment et le
potentiel glycolytique peuvent varier (Guignot et al. 1992). Par exemple, le Longissimus
dorsi a significativement plus de fibres blanches (αW ou type IIB), plus de fibres rouges à
contraction rapide (αR ou type IIA) et moins de fibres rouges à contraction lente (βR ou
type I) que le Psoas major. La quantité de fer est également plus élevée chez ce dernier
(Tableau 2.2).

6
Tableau 2.2. Composition des muscles Longissimus dorsi, Psoas major et Triceps brachii
capui laterale des carcasses de veaux (moyenne ± écart-type) Adapté de Guignot et al.
(1992)
LD1 PM1 TB1
Fibres αW (type IIB), % 56 ± 2a 47 ± 1b 52 ± 2a
Fibres αR (type IIA), % 23 ± 2a 16 ± 2bc 18 ± 1ac
Fibres βR (type I), % 21 ± 1a 36 ± 2bc 30 ± 2c
Pigment, µg Fe g-1 4.58 ± 0.2a 6.23 ± 0.2b –
PG, µmol g-1* 151 ± 8 158 ± 6 164 ± 5 a,b,c Moyennes sur une même ligne avec différents exposants sont significativement différentes (P < 0.05). 1 LD = Longissimus dorsi; PM = Psoas major; TB = Triceps brachii capui laterale.
* PG : Potentiel glycolytique
La vitesse de contraction des fibres musculaires dépend du métabolisme énergétique qui
découle de l’isoforme de myosine. La production d’énergie provient principalement de
l’utilisation des sucres et parfois des acides gras (Hocquette et al. 2000; INTERBEV,
2006). Cette énergie est également utilisée pour le développement du muscle tout au long
de la croissance de l’animal (Hocquette et al. 2000). Les surplus de glucose sont stockés
sous forme de glycogène ou de triglycérides pour le glucose et les acides gras (Hocquette et
al. 2000).
Le glucose peut être métabolisé par différentes voies biochimiques dans un muscle au
repos. Il peut s’agir d’une oxydation complète dans les mitochondries, de recyclage sous
forme de lactate ou de conversion en glycogène dans les fibres ou en lipides dans les
adipocytes intramusculaires (Hocquette et al. 2000).
Après l’abattage de l’animal, le muscle continue de fonctionner en puisant dans les réserves
énergétiques qui sont stockées (INTERBEV, 2006). Puisque le l’oxygène est désormais
absente, le métabolisme du glucose ou glycogène est anaérobique. L’activité résiduelle du
muscle résulte en la conversion des réserves de sucres en acide lactique (Hocquette et al.
2000). Cet acide s’accumule dans le muscle pour débuter l’acidification étant un élément
essentiel dans le processus de transformation du muscle en viande (INTERBEV, 2006). Le
taux d’acide lactique produit et la valeur du pH ultime dépendent de la quantité de réserves

7
en glycogène dans les muscles. Ainsi, une diminution de ces réserves en glycogène avant la
mort de l’animal engendre également une diminution de l’acide lactique produit (Scanga et
al. 1998). Ce phénomène résulte en une viande insuffisamment acidifiée (Hocquette et al.
2000; INTERBEV, 2006). Plusieurs facteurs peuvent affecter les réserves de glycogène tel
que les manipulations avant l’abattage, le transport, le stress ou l’activité physique
musculaire avant l’abattage. Ces facteurs font en sorte que la qualité de la viande est
également affectée (Hocquette et al. 2000; Pipek et al. 2003; Önenç et Kaya, 2004;
INTERBEV, 2006).
Après avoir subit un stress, le taux de glycogène dans les muscles diminue de façon
importante pour se retrouver dans la circulation sanguine sous forme de glucose (Tableau
2.3). Lors d’un stress comme le transport ou l’activité physique inhabituelle, le glucose
sanguin augmente afin de permettre au corps de réagir rapidement (Van de Water et al.
2003).
Tableau 2.3. Valeurs moyennes pour les concentrations plasmatiques avant et après le
transport de veaux Adapté de Van de Water et al. (2003)
Avant transport
(moyenne ± É.T.)
Après transport
(moyenne ± É.T.)
P
Glucose (mg/dl) 101 ± 27 932 ± 30.1 *
Cortisol (ng/ml) 6.83 ± 10.84 17.3 ± 15.6 ***
Lactate (mg/dl) 19.0 ± 20.2 52.4 ± 32.9 *** *, *** Différence significative entre les moyennes sur une même ligne (P < 0.05, P < 0.001).
É.T. = Écart-type
Cela demande beaucoup de temps d’attente à l’abattoir afin de rétablir le niveau de
glycogène (plus de 48 heures) même avec une alimentation à volonté (INTERBEV, 2006).
Il est donc préférable de réduire toutes sources de stress afin de préserver le glycogène
musculaire (Figure 2.2).

8
Figure 2.2. Évolution du taux de glycogène musculaire de taurillons après un stress
Source : INTERBEV (2006)
Avant l’abattage de l’animal, celui-ci subit un étourdissement visant à le rendre inconscient.
Ainsi, l’animal peut être saigné sans causer de douleur ou de détresse. La technique
d’étourdissement des animaux avant l’abattage a également une influence sur les réserves
de glycogène (Önenç et Kaya, 2004). Les chercheurs ont évalué trois techniques
d’étourdissement avant l’abattage auprès de 30 jeunes taureaux Holstein Friesian, dont le
poids moyen des carcasses était de 266 kg, soit aucun étourdissement, étourdissement
électrique et étourdissement avec un pistolet à percussion. L’étourdissement électrique et
avec le pistolet à percussion sont les deux méthodes dont le taux de glycogène demeure le
plus élevé. Aucune différence significative n’a été observée au niveau du pH24 (Tableau
2.4). Il se peut que les techniques d’étourdissement électrique et avec pistolet à percussion
permettent de réduire le stress des animaux de façon plus importante. Cela pourrait
expliquer le taux de glycogène plus élevé pour ces deux techniques d’étourdissement par
rapport à aucun étourdissement avant l’abattage.

9
Tableau 2.4. Teneur en glycogène des muscles et du pH selon la méthode d’abattage
Adapté de Önenç et Kaya (2004)
Paramètres Sans
étourdissement
Étourdissement
électrique
Étourdissement
par percussion Écart-type P
Glycogène
(mmol/l) 8.84 b 10.12 a 11.25 a 0.51 0.01
pH15 min 6.50 b 6.59 b 6.77 a 0.06 0.01
pH24 h 5.99 5.96 5.75 0.55 0.35 Les moyennes sur une même ligne avec différentes lettres sont significativement différentes (P < 0.05)
Le pH d’un muscle vivant se situe près de la neutralité (7,0) et après abattage, ce dernier
descend à environ 5,5-5,7 pour devenir de la viande (INTERBEV, 2006; Sosin-Bzducha et
al. 2012). Selon l’acidification du muscle, le pH peut se situer entre 5,8 et 6,3 (Guignot et
al. 1994; Pipek et al. 2003; Lomiwes et al. 2014; Vieira et Fernández, 2014).
L’acidification est également importante pour la conservation de la viande et prend
généralement 48 heures pour la viande bovine (INTERBEV, 2006). Les valeurs du tableau
précédent démontrent une diminution insuffisante du pH. Après l’abattage, il se pourrait
que l’étourdissement électrique augmente le taux de glycogène à cause de l’activité des
muscles et du relâchement des cathécolamines dans le sang (Önenç et Kaya, 2004).
Plusieurs modifications de la structure du muscle se produisent lors de l’acidification
(Hocquette et al. 2000; Pipek et al. 2003; INTERBEV, 2006). La vitesse de chute du pH
dépend du type de fibres musculaires et du métabolisme énergétique qu’elles utilisent.
(Hocquette et al. 2000). Le pH post mortem influence la couleur, la capacité de rétention
d’eau, la jutosité et surtout la tendreté de la viande selon l’amplitude et sa vitesse de chute
(Hocquette et al. 2000; INTERBEV, 2006). Différentes colorations seront créées par les
pigments présents dans la viande qui varieront selon leur quantité, l’oxygénation et
l’oxydation produite par le métabolisme (Hocquette et al. 2000; INTERBEV, 2006). La
jutosité est également influencée par la quantité de lipides présente dans le tissu adipeux
intramusculaire. La teneur en phospholipides et triglycérides ainsi que la composition en
acides gras et l’oxydation du gras feront varier la flaveur principalement, mais aussi la
tendreté de la viande (Hocquette et al. 2000). Cette dernière peut également varier selon

10
plusieurs autres paramètres tels que la teneur et la solubilité du collagène et des propriétés
contractiles et métaboliques des fibres musculaires (Figure 2.3).
Métabolisme oxydatif du muscle
Tissu adipeux intramusculaire
Métabolisme glycolytique du muscle
Teneur en
glycogène
Pigments
Teneur,
oxygénation
et oxydation
pH
Amplitude
et vitesse de
chute post
mortem
Collagène
Teneur et
solubilité
Fibres
musculaires
Propriétés
contractiles et
métaboliques
Lipides
Teneurs en
phospholipides
et triglycérides,
composition en
acides gras,
oxydation
Couleur Capacité de
rétention d’eau
Jutosité Tendreté Flaveur
Figure 2.3. Relations entre le métabolisme énergétique musculaire et les caractéristiques
musculaires déterminant la qualité organoleptique de la viande.
Adapté de Hocquette et al. (2000)
2.2. LA QUALITÉ DE LA VIANDE
La qualité de la viande est un concept général regroupant la qualité physico-chimique,
microbiologique et sensoriel. Cette dernière comprend les qualités organoleptiques
perceptibles lors de la consommation telles que la saveur, la jutosité et la tendreté de la
viande mentionnées précédemment. Plusieurs facteurs peuvent influencer la qualité de la
viande de veau. Par exemple, le pH, l’élevage en groupe ou individuel et la durée de

11
transport des animaux ont un effet direct sur la tendreté de la viande donc la qualité de
celle-ci (Campbell et al. 2013).
2.2.1. Le pH
Le pH d’une viande est mesuré à l’aide d’un pH-mètre muni d’une électrode servant à la
prise de donnée. Avant de procéder à une mesure, l’appareil doit être calibré avec des
solutions à pH standards soit à pH 4 et 7 (Campbell et al. 2013). Pour la production de veau
de grain, le pH ultime est obtenu 96 heures après l’abattage de l’animal (Campbell et al.
2013).
Pour la production de veau de grain au Canada, le pH peut varier entre 5,5 et 5,6. Au
Québec, pour une carcasse de veau de 148 kg, la moyenne du pH pour les muscles
longissimus est de 5,57. En Ontario, la moyenne du pH est 5,49 pour une carcasse de veau
de grain de 172 kg (Campbell et al. 2013). Comme mentionné dans la section précédente, le
pH dépend de la quantité de réserves en glycogène dans les muscles (Hocquette et al. 2000;
INTERBEV, 2006). La chute du pH post-mortem est le résultat de la transformation
anaérobie du glycogène en acide lactique (Pipek et al. 2003). Lorsque le potentiel
glycolytique du longissimus dorsi descend en dessous de 150 pmol/g 48 heures post-
mortem, les valeurs du pH ultime restent au-dessus des normales (pH > 6,3) (Guignot et al.
1992). La probabilité d’observer des carcasses à pH élevé est également 3,3 fois supérieure
chez les animaux pesant moins de 305 kg. Cela est probablement causé par les conditions
d’élevage telles que la stabulation en groupe. Les jeunes animaux sont plus excitables ce
qui entraînerait plus de consommation des réserves en glycogène (INTERBEV, 2006). Plus
le potentiel glycolytique est élevé, plus le pH diminue de façon importante (Figure 2.4). Le
potentiel glycolytique représente le niveau des réserves énergétiques du muscle avant la
mort de l’animal. L’estimation de ce potentiel s’effectue en dosant le glycogène musculaire
et des produits de sa dégradation en acide lactique : glucose, glucose-6-phosphate et acide
lactique, selon une formule établie en 1985 par MONIN et SELLIER : [Potentiel

12
glycolytique] = 2 x ([Glycogène] + [glucose] + [Glucose-6-phosphate]) + [acide lactique]
(INTERBEV, 2006).
Figure 2.4. Relation entre le potentiel glycolytique (PG) 48 heures post-mortem et le pH
ultime dans le muscle longissimus dorsi des carcasses de veaux. PG, µmol d’acide lactique
g-1 Adapté de Guignot et al. (1992)
Le pH ultime a une corrélation positive avec la tendreté, la jutosité et la flaveur et une
corrélation négative avec les pertes à la cuisson et les pertes en eau (Guignot et al. 1994;
Pipek et al. 2003; Ngapo et Gariépy, 2006).
Différentes activités protéolytiques responsables de l’attendrissement de la viande se
manifestent avec des valeurs de pH24 variant entre 5,8 et 6,3. L’activité des enzymes
calpaïnes est favorisée pour une viande dont le pH est plus élevé que 6,3 et celle des
enzymes cathepsines pour une viande dont le pH est plus bas que 5,8 (Guignot et al. 1994;
Pipek et al. 2003; Lomiwes et al. 2014; Vieira et Fernández, 2014). Lorsqu’il y a arrêt du
fonctionnement des pompes ioniques après l’abattage et que l’énergie (ATP) contenue dans
les muscles est épuisée. Nous observons une hausse de la concentration de calcium (Ca2+)
sarcoplasmique. Ce calcium est nécessaire pour activer les isoformes µ et m de la calpaïne
pour l’hydrolyse des protéines myofibrillaires. La calpaïne a non seulement la capacité de
dégrader les protéines myofibrillaires, mais également les protéines cytosquelettiques
(Huff-Lonergan et al. 2005; Lomiwes, 2014).

13
Lorsque les deux enzymes protéolytiques (calpaïne et cathepsine) sont plus actives, nous
pouvons observer la production d’une viande plus tendre qu’en dehors de l’échelle de pH
5,8-6,3 (Guignot et al. 1994; Lomiwes et al. 2014). Cela est provoqué par la dégradation
des protéines myofibrillaires à poids moléculaire élevé et bas durant le vieillissement de la
viande par l’activité de la µ-calpaïne et de la cathepsine B (Lomiwes et al. 2014). La vitesse
à laquelle la viande s’attendrit découle également du pH ultime. Toutefois,
l’attendrissement de la viande se produit plus rapidement lorsque le pH ultime est élevé
comparativement à un pH ultime faible. À faible pH ultime, le processus se retrouve
ralentit à cause du stade rigor mortis (rigidité cadavérique). L’activité des deux enzymes est
déclenchée très rapidement suivant la mort de l’animal. De plus, toute valeur de pH en
dehors de la «zone de confort» des enzymes occasionne une limitation dans l’activité de la
protéolyse (Lomiwes et al. 2014).
Pendant que l’activité de l’enzyme µ-calpaïne est plus active à pH ultime élevé (pH 7,5),
celle de la cathepsine B est plutôt faible (entre 5,0 et 6,0). Tout au long du vieillissement de
la viande, plus le pH ultime diminue et la situation inverse se produit. Donc, la µ-calpaïne
diminue son activité pour laisser place à l’activité de la cathepsine B (Lomiwes et al. 2014).
Cette dernière voit son activité pratiquement doublée durant le vieillissement à faible pH,
notamment à pH 4 (Figure 2.5). La cathepsine B est relâchée à l’intérieur du sarcoplasme à
partir des lysosomes lorsque la viande est à pH faible. La cathepsine B n’hydrolyse pas
seulement les protéines myofibrillaires, mais altère également les protéines associées avec
les bandes-A et les disques-Z des protéines myofibrillaires (Lomiwes et al. 2014).
La chute du pH est influencée par l’activité de la glycolyse qui réagit également en
conséquence de la température de refroidissement. Vieira et Fernández (2014) ont fait
diminuer la température de carcasses d’agneaux de trois façons différentes jusqu’à obtenir
2°C. Le refroidissement conventionnel consiste à placer la carcasse d’agneau dans une
chambre à 2°C durant 24 heures. Pour le refroidissement très rapide, la carcasse est placée
3,5 h dans une chambre à -20°C et dans une seconde chambre à 2°C jusqu’à 24 heures. La

14
dernière méthode de refroidissement dite lente consiste à laisser la carcasse durant 7h à
12°C et la transférer également dans une chambre à 2°C jusqu’à 24 heures. Une
température froide réduit l’activité métabolique résiduelle des muscles. Le processus
d’acidification de la carcasse se retrouve également ralentit et cela cause une réduction de
la chute du pH. Le pH ultime est donc influencé par la vitesse et la température de
refroidissement. Puisque le pH a de nombreuses influences sur la qualité de la viande,
plusieurs paramètres peuvent connaître également une modification (Vieira et Fernández,
2014). L’effet du ralentissement de la chute de pH sur la qualité de la viande sera abordé au
cours des autres sections.
Figure 2.5. Activité de la cathepsine B dans les échantillons de viande de bœuf vieilli à -
1°C jusqu’à 28 jours post mortem à pH ultime bas et élevé.
Adapté de Lomiwes et al. (2014)
2.2.2. La couleur de la viande
La valeur de la carcasse de veau se base sur l’un des facteurs les plus importants : la
couleur (Guignot et al. 1992). Il s’agit également du premier point critique que les
consommateurs utilisent pour s’assurer qu’une viande est dite de qualité et soit acceptable
(Abril et al. 2001). La couleur de la viande comporte trois indices de couleurs soit L*, a* et
b* et se mesurent à l’aide d’un colorimètre. L* désigne la luminosité de la viande. Plus
cette valeur est élevée, plus la viande est d’apparence claire. a* représente le spectre variant
de la couleur rouge à vert. Plus la valeur de a* est élevée, plus la viande est rouge.
Act
ivit
é d
e la
ca
thep
sin
e B
(m
U/m
g d
e p
roté
ine)
pH bas
pH élevé
Temps post mortem (jours)

15
Finalement, b* désigne le spectre variant de la couleur jaune à bleu. Plus la valeur de b* est
élevée, plus la viande est jaune (Nade et al. 2012; Campbell et al. 2013). L’angle Hue est
représenté par l’équation Tan-1 (b*/a*) (Campbell et al. 2013). Dès que les indices a* et b*
changent, l’angle Hue change également. La couleur de la viande est influencée par
plusieurs facteurs tels que la myoglobine et son niveau d’oxydation, la structure des
protéines, la valeur du pH ultime, les conditions pré-abattage, l’espèce, le sexe, l’âge, le
poids de l’animal, l’alimentation et le vieillissement de la viande (Guignot et al. 1992;
Abril et al. 2001).
Tel que mentionné dans la section 2.1, l’épuisement des réserves de glycogène pré-abattage
lors d’un stress ou d’un jeûne prolongé conduit à l’obtention d’un pH ultime élevé (Van de
Water et al. 2003). Au niveau de la couleur de la viande, ce phénomène fera une viande dite
DFD (Dark, Firm, Dry) soit Foncée, Dure et Sèche ou connue également sous le nom de
coupe sombre (Scanga et al. 1998; Abril et al. 2001; Pipek et al. 2003; Van de Water et al.
2003). Donc, plus le stress lors d’un transport est long, plus les réserves de glycogène
seront affectées (Van de Water et al. 2003; INTERBEV, 2006). La fréquence des coupes
sombres augmente de façon parallèle lorsque la distance parcourue entre la ferme et
l’abattoir excède 300 km (Jones et al. 1989).
L’acidification du muscle fait en sorte de modifier sa structure musculaire qui réfléchit la
lumière dépendamment du pH produisant ainsi une luminosité différente. Lorsque le pH est
élevé (>6,3), nous nous éloignons du point isoélectrique des protéines dont la structure est
dite ouverte. Les chaînes de protéines se repoussent puisqu’elles sont chargées
électriquement et renferment des molécules d’eau. La lumière pénètre donc en profondeur
dans le muscle et la portion réfléchie est faible, d’où l’obtention d’une viande plus foncée
(Guignot et al. 1994; Abril et al. 2001). De plus, la formation de réseau crée un espace pour
l’accumulation d’eau produisant ainsi une viande plutôt collante (Guignot et al. 1994;
INTERBEV, 2006). Le contraire est observé lorsque le pH est trop faible (<5,8). Nous nous
rapprochons du point isoélectrique des protéines. Elles se resserrent par un effet d’attraction
réciproque puisque leurs charges diminuent. Il s’agit du rétrécissement latéral des
myofibrilles (Ripoll et al. 2013). La structure est donc dite fermée. La lumière pénètre peu

16
dans le muscle et nous pouvons observer une grande quantité réfléchie. Ceci fait en sorte
d’observer une viande plus claire. Toutefois, une structure dite fermée occasionne
également une diminution du pouvoir de rétention d’eau du muscle. Donc, des pertes
importantes d’exsudat se produisent lors de la conservation de la viande (INTERBEV,
2006). Les pertes en eau seraient reliées à la vitesse de chute du pH (Guignot et al. 1994).
Lors du rétrécissement latéral des myofibrilles, l’eau se déverse dans l’espace
extracellulaire des muscles (Huff-Lonergan et al. 2005 et Ripoll et al. 2013). La taille de
ces espaces est influencée par la valeur du pH ultime, donc également les pertes en eau
(Guignot et al. 1994).
La coloration de la viande dépend également de la forme chimique de la myoglobine et de
sa quantité (Guignot et al. 1994; Abril et al. 2001; INTERBEV, 2006). La myoglobine est
un pigment intrinsèque, connue également comme une protéine complexe nommée la
chromoprotéine, et varie selon le muscle, l’espèce, l’âge de l’animal et l’alimentation (Abril
et al. 2001; INTERBEV, 2006). Elle est composée d’une protéine incolore soit la globine et
d’une autre qui lui donne sa coloration étant l’hème. Ce dernier est composé d’un atome de
fer et est responsable de l’intensité de la couleur rouge de la viande (INTERBEV, 2006;
Nikmaram et al. 2011). Cet atome permet donc à la myoglobine de déterminer l’intensité de
la pigmentation du muscle à plus de 90%. (INTERBEV, 2006). Plus une viande est riche en
fer, plus la couleur résultante de cette viande sera d’un rouge vif. La myoglobine a
également un rôle de réserve d’oxygène pour les muscle et de transport de cet oxygène
jusqu’aux structures intracellulaires (INTERBEV, 2006).
La quantité de fer augmente dans les muscles avec le temps pour faire en sorte que la
viande devienne de plus en plus rouge saturé (INTERBEV, 2006). La pigmentation
définitive de la viande bovine dépend de la maturité physiologique de l’animal, notamment
par sa vitesse de croissance et l’objectif du poids de carcasse visé (INTERBEV, 2006).
L’apparition de coupes sombres augmente lorsque le poids de la carcasse est inférieur à 275
kg (Jones et al. 1989). Selon la catégorie de poids de la carcasse, le pourcentage de coupe
sombre chez le bœuf le plus faible se situerait entre 275 et 375 kg (Tableau 2.5). Les

17
coupes sombres sont également associées à une température d’élevage plus élevée (5,4
contre 4,8ºC) (Jones et al. 1989). Cela pourrait s’expliquer par la taille plus petite des
animaux vivants ainsi plus de stress lors du transport que les animaux plus gros (Jones et al.
1989).
Tableau 2.5. Effets du poids de la carcasse sur la fréquence des coupes sombres de boeuf
Adapté de Jones et al. (1989)
Poids carcasse (kg) Nombre de carcasses % carcasses avec coupes
sombres†
150 – 275 48 800 1.04
> 275 – 375 95 024 0.85
> 375 26 710 0.91
†Probabilité Khi-carré P < 0.003.
Le pH a un certain effet sur la myoglobine, donc sur la couleur de la viande (Abril et al.
2001; Pipek et al. 2003). Ce pigment peut se retrouver sous différentes formes dans la
viande crue dépendamment à l’état d’oxydoréduction et d’oxygénation soit sous forme
réduite (Mb), la forme oxymyoglobine (MBO2) et la metmyoglobine (MetMb). Le stade
physico-chimique ou stabilitéde la myoglobine détermine la couleur de la viande
(Hocquette et al. 2000; Abril et al. 2001). L’état de la myoglobine est également en
fonction de la fraîcheur de la viande et de la pression en oxygène (INTERBEV, 2006).
La surface d’une viande de bœuf fraîche telle que nous la voyons est d’un rouge vif. Il
s’agit de la myoglobine oxygénée soit l’oxymyoglobine (MbO2) puisque la pression en
oxygène est importante. La couleur de la viande dépend également de l’épaisseur de la
couche d’oxymyoglobine. En absence d’oxygène, la myoglobine est retrouvée sous une
forme différente pour être désormais d’un rouge plus foncé, voire pourpre. La myoglobine
est donc à l’état réduit et désoxygéné (Mb) puisque la pression en oxygène est nulle. Selon
la présence ou non d’oxygène, la myoglobine peut changer de forme de façon réversible
entre sa forme oxygénée rouge vif à sa forme réduite rouge pourpre. Entre ces deux
couches se trouve une troisième forme étant la metmyoglobine (INTERBEV, 2006). Cette
couche correspond à l’oxydation de l’oxymyoglobine dont la couleur est brune (Hocquette

18
et al. 2000; INTERBEV, 2006). Elle dépend également de plusieurs facteurs comme
l’évolution du pH post mortem, la vitesse de diffusion de l’oxygène et la consommation
d’oxygène par les mitochondries suite à l’abattage (Hocquette et al. 2000; De Palo et al.
2013). La couche de metmyoglobine tend à se développer par oxydation progressive de
l’oxymyoglobine lors de la conservation (Figure 2.6).
Figure 2.6 : Les trois formes chimiques de la myoglobine
Source : INTERBEV (2006)
La couleur de certains muscles du veau peut être liée à la vitesse de chute du pH. Ainsi,
pour un muscle à chute de pH rapide tel que le filet, l’intensité de la couleur dépend plus de
la vitesse de chute du pH que de la teneur en pigment. C’est le contraire pour un muscle à
chute de pH lente (INTERBEV, 2006). La luminosité et la rougeur de la viande diminuent
lorsque la valeur du pH ultime augmente (Guignot et al. 1994). Lorsque le pH ultime est
bas, soit entre 5,2 et 5,5, cela favorise l’autoxydation de la myoglobine oxygénée ou réduite
pour résulter en une teinte plus faible (Guignot et al. 1992; INTERBEV, 2006). Donc, c’est
le cas des viandes PSE (Pale, Soft, Exsudative) soit Pâles, Molles, Exsudatives. Lorsque le
pH ultime est normal, soit entre 5,5 et 5,7, ceci limite la consommation d’oxygène par le
muscle. Ainsi, la myoglobine reste sous sa forme oxygénée rouge vif en surface.
Finalement, lorsque le pH ultime est élevé, soit entre 6,3 et 6,7, la consommation
d’oxygène augmente et la myoglobine demeure à l’état réduit étant rouge sombre (Figure
2.7). Cette viande est donc insuffisamment acidifiée puisque l’acidification s’est arrêtée

19
alors que le pH est toujours élevé dans le muscle en raison du manque de réserves en
glycogène (INTERBEV, 2006). Une viande est considérée DFD lorsque le pH24 est plus
élevé que 6,2 (Pipek et al. 2003).
Figure 2.7 : Évolution du pH musculaire et de la couleur de la viande après l’abattage
Source : INTERBEV (2006)
Chez le porc, trois catégories de qualité avaient été formulées selon la couleur, la fermeté et
l’exsudation étant PSE pour Pale, Soft, Exsudative (Pâle, Molle, Exsudative); RFN pour
Reddish-pink, Firm, Non-exsudative (Rouge-rose, Ferme, Non-exsudative) et DFD pour
Dark, Firm, Dry (Foncée, Ferme, Sèche). D’autres intermédiares ont été catégorisés pour
ainsi en ajouter deux nouvelles soit : RSE pour Reddish-pink, Soft, Exsudative (Rouge-rose,
Molle, Exsudative) et PFN pour Pale, Firm, Non-exsudative (Pâle, Ferme, Non-exsudative)
(Faucitano et al. 2010). Ces cinq catégories se distinguent par leur pH ultime, leur
luminosité (L*) et leur perte en eau (Tableau 2.6). Les viandes de catégorie PSE sont celles
dont le pH ultime est le plus bas, un L* plus élevé (plus pâle) et ayant des pertes en eau
plus importantes. Les viandes de catégorie DFD sont à l’opposé des viandes de catégorie
PSE. Donc, le pH ultime est le plus élevé, le L* est le plus faible et les pertes en eau sont
les moins importantes. Pour les viandes de catégorie PFN, RSE et RFN, le L* n’est pas
significativement différent et au niveau du pH ultime, PFN est significativement différent

20
des deux autres. RSE est significativement différent que PFN et RFN concernant les pertes
en eau (Fautacino et al. 2010).
Tableau 2.6. Mesures de la qualité de la viande sur 117 longes de porc de chaque classe de
qualité. Adapté de Faucitano et al. (2010)A
Classes de qualitéB pHu L* Perte en eauC
mg
PSE 5.52d 53.41a 125.10a
PFN 5.58cd 52.54ab 45.12c
RSE 5.67bc 46.43b 99.84b
RFN 5.71b 45.92b 33.16cd
DFD 6.21a 40.54c 24.60d
SEMD 0.03 0.47 3.92 A
Moyennes suivies par une lettre différente sur la même ligne sont significativement différentes (P < 0.05). B
PSE (pâle, molle, exsudative); PFN (pâle, ferme, non-exsudative); RSE (rouge, molle, exsudative); RFN (rouge, ferme,
non-exsudative); and DFD (foncé, ferme, sec).
C Perte en eau selon Kaufmann et al. (1986) et les lignes directives du National Pork Board (NPB, 2000).
D SEM, erreur standard de la moyenne.
Le vieillissement de la viande a également un impact sur la couleur de la viande. L’étude de
Campbell et al. (2013) a démontré que chez le veau de grain, le vieillissement de la viande
résulte en une diminution de la valeur de b*. Cependant, pour le Musculus longissimus
thoracis, c’est une augmentation de la valeur de L* et b* de qui est observée et une
diminution de la valeur de a* (Sosin-Bzducha et al. 2012). La protéolyse se poursuit lors du
vieillissement et cela fait en sorte de créer de nouvelles structures dites ouvertes laissant
entrer la lumière plus profondément dans la viande. Donc, plus il y a relâchement au niveau
des protéines avec le temps plus la couleur de la viande deviendra foncée (Önenç et Kaya,
2004; Vieira et Fernández, 2014).
La façon dont l’étourdissement des animaux se produit avant l’abattage peut avoir une
incidence sur la couleur de la viande. Trois jours après l’abattage, les indices de couleurs
L* et b* sont significativement différents pour l’étourdissement électrique avec une viande
plus foncée (Önenç et Kaya, 2004). L’expérience de ces chercheurs a également fait le
portrait sur l’évolution de la couleur de la viande avec le vieillissement. Pour les trois
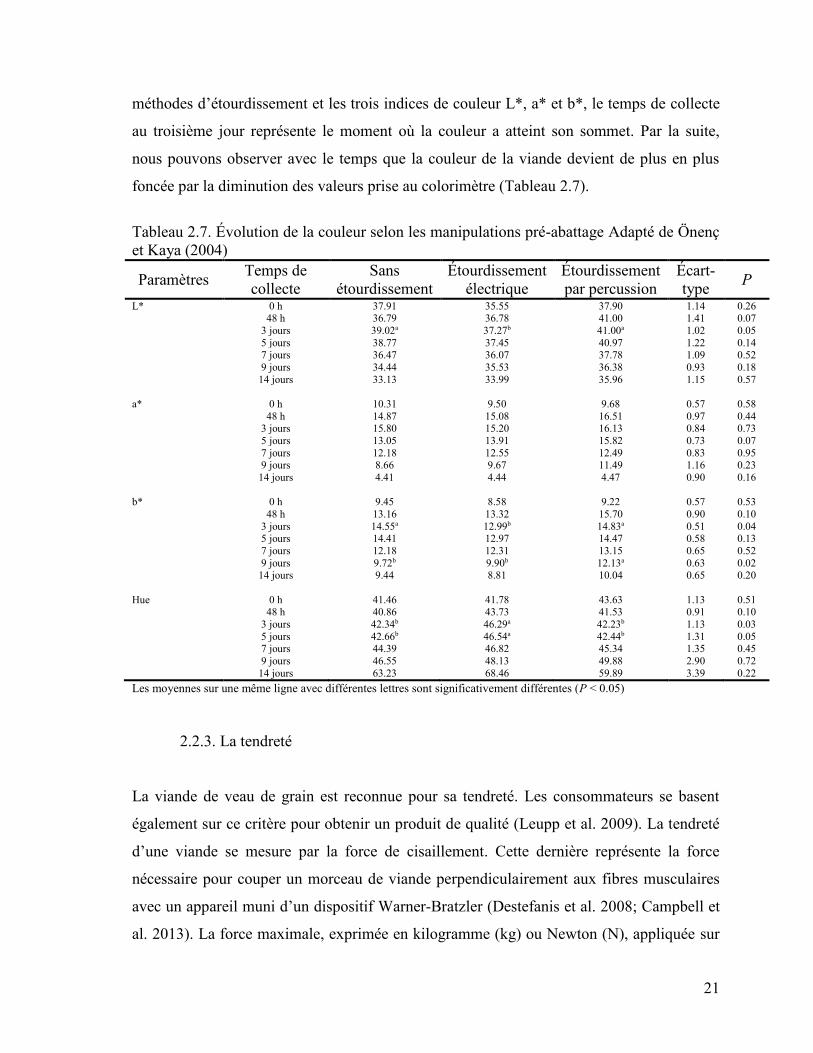
21
méthodes d’étourdissement et les trois indices de couleur L*, a* et b*, le temps de collecte
au troisième jour représente le moment où la couleur a atteint son sommet. Par la suite,
nous pouvons observer avec le temps que la couleur de la viande devient de plus en plus
foncée par la diminution des valeurs prise au colorimètre (Tableau 2.7).
Tableau 2.7. Évolution de la couleur selon les manipulations pré-abattage Adapté de Önenç
et Kaya (2004)
Paramètres Temps de
collecte
Sans
étourdissement
Étourdissement
électrique
Étourdissement
par percussion
Écart-
type P
L* 0 h 37.91 35.55 37.90 1.14 0.26 48 h 36.79 36.78 41.00 1.41 0.07
3 jours 39.02a 37.27b 41.00a 1.02 0.05
5 jours 38.77 37.45 40.97 1.22 0.14 7 jours 36.47 36.07 37.78 1.09 0.52
9 jours 34.44 35.53 36.38 0.93 0.18
14 jours 33.13 33.99 35.96 1.15 0.57
a* 0 h 10.31 9.50 9.68 0.57 0.58
48 h 14.87 15.08 16.51 0.97 0.44 3 jours 15.80 15.20 16.13 0.84 0.73
5 jours 13.05 13.91 15.82 0.73 0.07
7 jours 12.18 12.55 12.49 0.83 0.95 9 jours 8.66 9.67 11.49 1.16 0.23
14 jours 4.41 4.44 4.47 0.90 0.16
b* 0 h 9.45 8.58 9.22 0.57 0.53
48 h 13.16 13.32 15.70 0.90 0.10
3 jours 14.55a 12.99b 14.83a 0.51 0.04 5 jours 14.41 12.97 14.47 0.58 0.13
7 jours 12.18 12.31 13.15 0.65 0.52
9 jours 9.72b 9.90b 12.13a 0.63 0.02
14 jours 9.44 8.81 10.04 0.65 0.20
Hue 0 h 41.46 41.78 43.63 1.13 0.51 48 h 40.86 43.73 41.53 0.91 0.10
3 jours 42.34b 46.29a 42.23b 1.13 0.03
5 jours 42.66b 46.54a 42.44b 1.31 0.05 7 jours 44.39 46.82 45.34 1.35 0.45
9 jours 46.55 48.13 49.88 2.90 0.72
14 jours 63.23 68.46 59.89 3.39 0.22
Les moyennes sur une même ligne avec différentes lettres sont significativement différentes (P < 0.05)
2.2.3. La tendreté
La viande de veau de grain est reconnue pour sa tendreté. Les consommateurs se basent
également sur ce critère pour obtenir un produit de qualité (Leupp et al. 2009). La tendreté
d’une viande se mesure par la force de cisaillement. Cette dernière représente la force
nécessaire pour couper un morceau de viande perpendiculairement aux fibres musculaires
avec un appareil muni d’un dispositif Warner-Bratzler (Destefanis et al. 2008; Campbell et
al. 2013). La force maximale, exprimée en kilogramme (kg) ou Newton (N), appliquée sur

22
la viande pour la couper correspond à la force de cisaillement (Destefanis et al. 2008;
Campbell et al. 2013). Pour la convertir les N en kg, il suffit de diviser les N par
l’accélération gravitationnelle (9,8 m/s2) et l’opération inverse pour convertir les kg en N.
La force de cisaillement, ou la tendreté, dépend de plusieurs paramètres tel que le pH, l’âge,
la race et le sexe de l’animal, le vieillissement de la viande, l’élevage individuel ou en
groupe, l’alimentation, la quantité de collagène, la composition de la viande et la méthode
de cuisson de la viande (Mandell et al. 1997; Hocquette et al. 2000; Pipek et al. 2003;
Nikmaram et al. 2011; Campbell et al. 2013).
Tel que mentionné dans la section 2.2.1, le pH a une incidence sur l’activité des enzymes
protéolytiques (calpaïne et cathepsine) et fait en sorte d’obtenir une viande tendre ou non.
Toute viande dont le pH ultime est inférieur à 5,8 ou plus élevé que 6,3 sera plus tendre
(Guignot et al. 1994; Pipek et al. 2003; Lomiwes et al. 2014). Le vieillissement de la viande
fait en sorte de diminuer la force de cisaillement (Campbell et al. 2013; Lucero-Borja et al.
2014). Lomiwes et al. (2014) ont évalué la force de cisaillement selon différents pH ultimes
et jours de vieillissement de la viande (1, 2, 7 et 28 jours). La tendreté de la viande n’est
pas la même à deux jours ou 28 jours après l’abattage. La viande est également plus tendre
pour celles ayant un pH ultime élevé (Figure 2.8). En effet, plus nous laissons vieillir la
viande, plus elle sera tendre également (Campbell et al. 2013; Lomiwes et al. 2014; Lucero-
Borja et al. 2014).
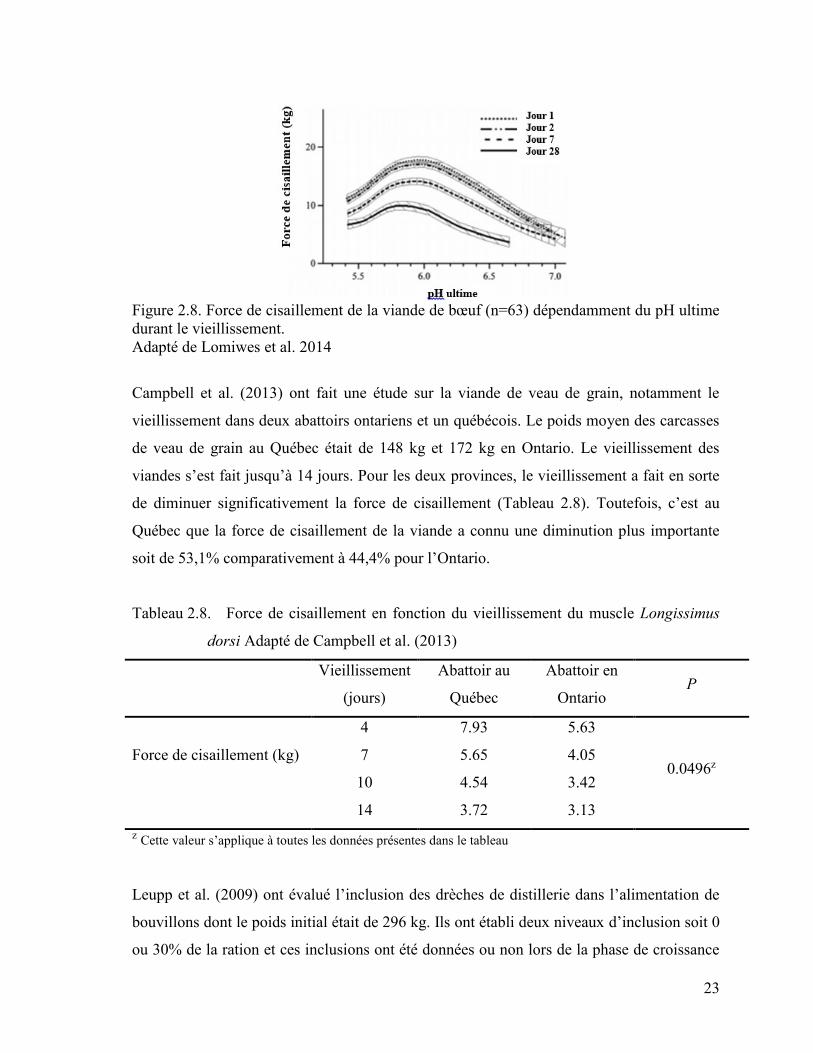
23
Figure 2.8. Force de cisaillement de la viande de bœuf (n=63) dépendamment du pH ultime
durant le vieillissement.
Adapté de Lomiwes et al. 2014
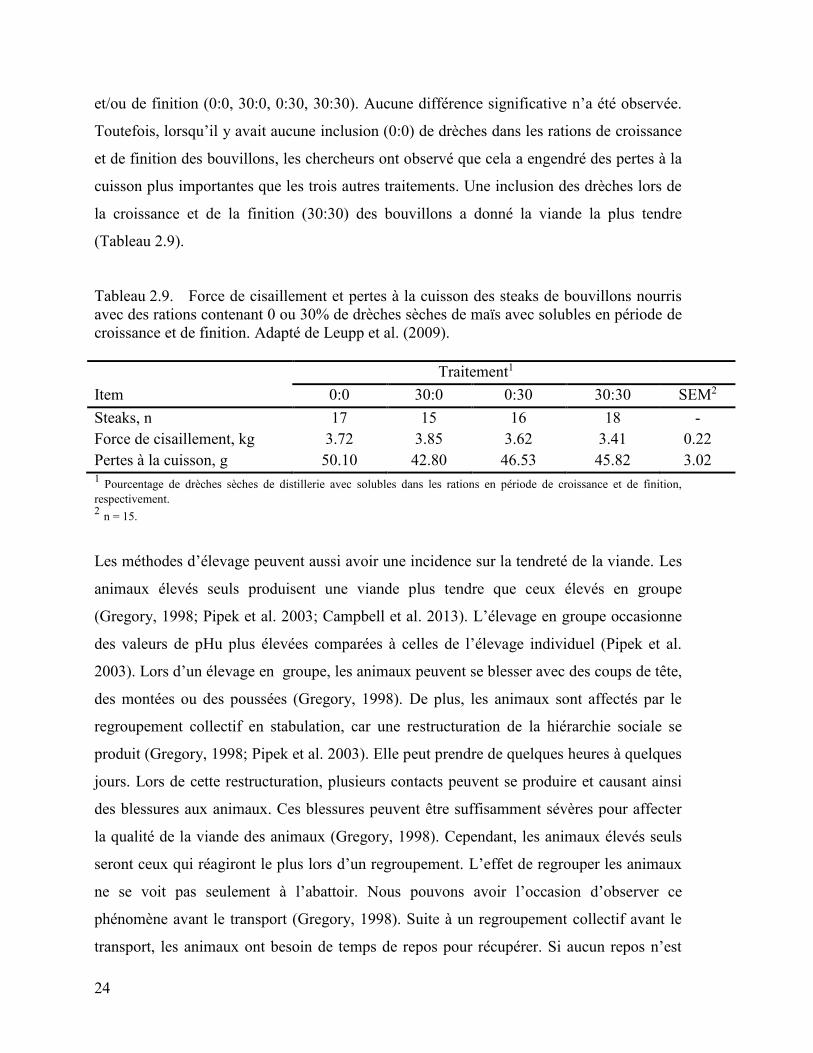
Campbell et al. (2013) ont fait une étude sur la viande de veau de grain, notamment le
vieillissement dans deux abattoirs ontariens et un québécois. Le poids moyen des carcasses
de veau de grain au Québec était de 148 kg et 172 kg en Ontario. Le vieillissement des
viandes s’est fait jusqu’à 14 jours. Pour les deux provinces, le vieillissement a fait en sorte
de diminuer significativement la force de cisaillement (Tableau 2.8). Toutefois, c’est au
Québec que la force de cisaillement de la viande a connu une diminution plus importante
soit de 53,1% comparativement à 44,4% pour l’Ontario.
Tableau 2.8. Force de cisaillement en fonction du vieillissement du muscle Longissimus
dorsi Adapté de Campbell et al. (2013)
Vieillissement
(jours)
Abattoir au
Québec
Abattoir en
Ontario P
4 7.93 5.63
0.0496z Force de cisaillement (kg) 7 5.65 4.05
10 4.54 3.42
14 3.72 3.13
z Cette valeur s’applique à toutes les données présentes dans le tableau
Leupp et al. (2009) ont évalué l’inclusion des drèches de distillerie dans l’alimentation de
bouvillons dont le poids initial était de 296 kg. Ils ont établi deux niveaux d’inclusion soit 0
ou 30% de la ration et ces inclusions ont été données ou non lors de la phase de croissance
24
et/ou de finition (0:0, 30:0, 0:30, 30:30). Aucune différence significative n’a été observée.
Toutefois, lorsqu’il y avait aucune inclusion (0:0) de drèches dans les rations de croissance
et de finition des bouvillons, les chercheurs ont observé que cela a engendré des pertes à la
cuisson plus importantes que les trois autres traitements. Une inclusion des drèches lors de
la croissance et de la finition (30:30) des bouvillons a donné la viande la plus tendre
(Tableau 2.9).
Tableau 2.9. Force de cisaillement et pertes à la cuisson des steaks de bouvillons nourris
avec des rations contenant 0 ou 30% de drèches sèches de maïs avec solubles en période de
croissance et de finition. Adapté de Leupp et al. (2009).
Traitement1
Item 0:0 30:0 0:30 30:30 SEM2
Steaks, n 17 15 16 18 -
Force de cisaillement, kg 3.72 3.85 3.62 3.41 0.22
Pertes à la cuisson, g 50.10 42.80 46.53 45.82 3.02 1
Pourcentage de drèches sèches de distillerie avec solubles dans les rations en période de croissance et de finition,
respectivement. 2 n = 15.
Les méthodes d’élevage peuvent aussi avoir une incidence sur la tendreté de la viande. Les
animaux élevés seuls produisent une viande plus tendre que ceux élevés en groupe
(Gregory, 1998; Pipek et al. 2003; Campbell et al. 2013). L’élevage en groupe occasionne
des valeurs de pHu plus élevées comparées à celles de l’élevage individuel (Pipek et al.
2003). Lors d’un élevage en groupe, les animaux peuvent se blesser avec des coups de tête,
des montées ou des poussées (Gregory, 1998). De plus, les animaux sont affectés par le
regroupement collectif en stabulation, car une restructuration de la hiérarchie sociale se
produit (Gregory, 1998; Pipek et al. 2003). Elle peut prendre de quelques heures à quelques
jours. Lors de cette restructuration, plusieurs contacts peuvent se produire et causant ainsi
des blessures aux animaux. Ces blessures peuvent être suffisamment sévères pour affecter
la qualité de la viande des animaux (Gregory, 1998). Cependant, les animaux élevés seuls
seront ceux qui réagiront le plus lors d’un regroupement. L’effet de regrouper les animaux
ne se voit pas seulement à l’abattoir. Nous pouvons avoir l’occasion d’observer ce
phénomène avant le transport (Gregory, 1998). Suite à un regroupement collectif avant le
transport, les animaux ont besoin de temps de repos pour récupérer. Si aucun repos n’est

25
accordé, une différence au niveau sanguin se produit. En effet, les deux indicateurs
plasmatiques de stress, le cortisol et le lactate, augmentent grandement. Chez le porc, trois
heures de repos après le transport s’avère bénéfique autant pour ceux n’ayant pas été
mélangés avec d’autres que ceux qui l’ont été. Le porc ayant été mélangé dans un autre
groupe et abattu immédiatement à son arrivée à l’abattoir aura des niveaux d’indicateurs
plasmatiques très élevés (Tableau 2.10).
Tableau 2.10. L’effet de mélanger les groupes de verrats et de cochettes avant le transport
et en stabulation sur deux indicateurs plasmatiques de stress Adapté de Gregory. (1998)
Porcs non
mélangés Porcs mélangés
Indicateurs
plasmatiques
Abattage après
3 heures en
stabulation
Abattage
après 3 heures
en stabulation
Abattage à
l’arrivée à
l’abattoir
Erreur standard
de déviation
Cortisol
(µg par 100 ml) 5.5 5.8 14.3 0.7
Lactate
(mg par 100 ml) 54 65 98 7
La méthode de cuisson peut influencer la force de cisaillement de la viande. Lors de la
cuisson, divers phénomènes se manifestent. Une dénaturation des protéines se produit et
cela cause des changements de structure dans la viande. Nous pouvons donc observer la
destruction des membranes cellulaires, le rétrécissement des fibres, de l’agrégation et la
formation d’un gel de protéines myofibrillaires et sarcoplasmiques ainsi qu’un
rétrécissement et une solubilisation du collagène (Nikmaram et al. 2011). La dureté de la
viande dépend également de la méthode de cuisson. Entre une cuisson aux micro-ondes,
une viande rôtie ou braisée, c’est la cuisson par braisage qui créer une viande plus tendre
(Tableau 2.11). La dureté de la viande augmente lors de la cuisson et l’augmentation est
causée par la dénaturation du collagène intramusculaire ou dans la structure des protéines
myofibrillaires. La cuisson solubilise également le tissu conjonctif produisant
l’attendrissement de la viande. Donc, les composantes du muscle comme les protéines
myofibrillaires et les protéines de tissus conjonctifs, le collagène et l’élastine contrôlent la
dureté de la viande (Nikmaram et al. 2011).

26
Les méthodes d’étourdissement peuvent avoir un effet sur la tendreté de la viande. Önenç et
Kaya (2004), tel que mentionné dans la section 2.1, ont évalué trois méthodes
d’étourdissement différentes avant l’abattage des animaux. L’étourdissement électrique a
engendré une viande plus dure (13,83 kg) que les deux autres traitements étant aucun
étourdissement et étourdissement par percussion (9,80 et 9,12 kg respectivement).
Tableau 2.11. Force de cisaillement du muscle Longissimus dorsi de veau avec différentes
méthodes de cuisson Adapté de Nikmaram et al. (2011)
(Moyenne ± SE)
Traitements Force de cisaillement (N)
Crue 16.90 ± 4.87
Micro-ondes 37.00 ± 2.64
Rôtir 37.64 ± 1.16
Braiser 34.02 ± 1.52
La longueur des sarcomères (fibres musculaires) semble avoir une influence sur la tendreté
de la viande. La longueur des sarcomères représente la distance entre deux lignes Z
(Campbell et al. 2013; Ishihara et al. 2013). La longueur des sarcomères s’exprime en
micromètre (µm) (Vieira et Fernández, 2014). Lors d’une contraction musculaire, les
myofilaments d’actine et de myosine glissent entre elles et les lignes Z se rapprochent l’une
de l’autre (Figure 2.9). Ce phénomène occasionne également le rétrécissement des
sarcomères lors du rapprochement de ces lignes Z (Gregory, 1998). Les ramifications des
myofilaments de myosine créent et brisent à plusieurs reprises et entre en contact avec les
myofilaments adjacents d’actine. À chaque fois, un pont d’actomyosine se crée lorsque
l’ATPase de la myosine est activée et celle de l’actine est à zéro. Durant la relaxation, la
myosine doit se recharger d’une molécule d’ATP pour retourner à son état initial. Si l’ATP
n’est pas disponible, les côtés ramifiés des myofilaments de myosine resteront attachés à
l’actine. Cette situation se présente souvent après l’abattage des animaux et c’est la raison
également pourquoi les muscles demeurent contractés (rigor) (Gregory, 1998). Les muscles
demeurent actifs après l’abattage de l’animal et les contractions se poursuivent jusqu’à
épuisement des réserves d’ATP (Gregory, 1998; INTERBEV, 2006).

27
Figure 2.9. Mouvement des myofilaments d’actine et de myosine lors de la contraction et
de la relaxation des muscles.
Adapté de Gregory, (1998).
Plusieurs facteurs peuvent influencer la longueur des sarcomères, notamment la cathepsine
B qui altère les protéines associées avec les bandes A et les lignes Z des protéines
myofibrillaires (Lomiwes et al. 2014). Donc, le pH ultime pourrait avoir un impact sur la
longueur des sarcomères puisqu’il joue un rôle dans l’activité de la protéase.
Pour le muscle longissimus provenant d’un boeuf, Ishihara et al. (2013) ont obtenu une
longueur de sarcomère de 1,79 μm tandis que Lucero-Borja et al. (2014) ont obtenu une
moyenne de 2,01 μm tout âge et sexe confondus pour le même type de muscle. England et
al. (2012) ont évalué trois différents types de muscle avec deux méthodes de suspension de
la carcasse de bœuf. Ils ont évalué le Longissimus dorsi, le Psoas major et le muscle
semitendineux avec une suspension normale soit par le tendon d’Achilles et une suspension
par la hanche. Une différence significative concernant la longueur des sarcomères est
observée entre les deux méthodes de suspension de la carcasse pour les trois types de
muscles. Les muscles Longissimus dorsi et le semitendineux ont une longueur des

28
sarcomères plus élevée lorsque la carcasse est suspendue par la hanche. Le muscle Psoas
major a donné un résultat différent des deux autres, car la longueur des sarcomères est
devenue significativement plus petite que la suspension normale (Tableau 2.12). Donc, la
méthode de suspension de la carcasse influence la longueur des sarcomères.
Tableau 2.12. Moyennes des longueurs de sarcomère (µm) provenant de différents muscles
de bœuf et selon la méthode de suspension de la carcasse Adapté de England et al. (2012).
Traitements1
Muscle2 SN SH SEM
LM 1.82 a 2.43 b 0.04
PM 3.64 a 2.06 b 0.04
STN 2.36 a 2.97 b 0.03 a,b Les moyennes avec des exposants différents sur une même ligne sont différents (P < 0,0001). 1 SN = suspension normale (tendon d’Achilles); SH = suspension par la hanche. Les erreurs standards sont pour une
même ligne. 2 LM = muscle Longissimus dorsi; PM = muscle Psoas major; STN = muscle semitendineux.
Le rétrécissement dû au froid est plus connu sous le nom de cold shortening. Ce
phénomène est occasionné par la longueur des sarcomères étant influencée par la
température de refroidissement de la carcasse ainsi que le pH. En dessous de cette mesure,
le cold shortening a pour effet d’augmenter la fermeté de la viande. La force de
cisaillement peut être influencée par la longueur des sarcomères. Il est possible d’observer
un effet de cold shortening si la température interne d’une carcasse diminue en dessous de
10°C et conserve un pH élevé (pH>6.2) 10 heures après l’abattage de l’animal (Vieira et
Fernández, 2014). Le processus de refroidissement s’effectue alors trop rapidement.
Diverses applications ont été développées pour améliorer la tendreté telle que la stimulation
électrique, différents traitements de refroidissement de la carcasse, le vieillissement et la
suspension pelvienne. Afin de prévenir ce phénomène, un ratio temps/température a été
établi. En effet, la température interne de la carcasse ne doit pas diminuer plus de 10°C en
10 heures après l’abattage de l’animal toutes espèces confondues (Bendall, 1973 ;
Campbell et al. 2013).

29
L’étude de Vieira et Fernández (2014) a démontré qu’un refroidissement lent de la carcasse
d’agneaux donnait des longueurs de sarcomères plus élevées que les traitements
conventionnel et très rapide. La force de cisaillement la plus faible a été observée chez le
traitement de refroidissement lent accompagné de la longueur de sarcomère la plus élevée
de façon significative que ce soit avant et après le vieillissement la viande. Le
refroidissement lent est le seul traitement dont la longueur des sarcomères est restée la
même suite au vieillissement (Tableau 2.13). Pour le traitment conventionnel, le
vieillissement de la viande a permis d’obtenir une différence significative pour une
augmentation de la longueur des sarcomères. Pour les traitements conventionnel et très
rapide, le vieillissement de la viande a fait diminuer significativement la force de
cisaillement. Une fois de plus, il est démontré que le vieillissement de la viande pourrait la
rendre plus tendre pour importe le traitement de refroidissement.
Tableau 2.13. Effets des traitements de refroidissement et du vieillissement sur le Warner-
Bratzler et la longueur des sarcomères chez l’agneau de lait.
Conventionnelle Très rapide Lente RSE P
NV V NV V NV V T V T x V
LS1 (µm) 1.5bx 1.6B
y 1.4ax 1.2A
y 1.7c 1.7C 0.10 *** * ns
FC2 (kg) 6.2bx 4.9A
y 7.6bx 5.8B
x 5.4a 4.7A 1.21 *** *** ns
Adapté de Vieira et Fernández, (2014). 1LS = Longueur des sarcomères;
2FC = Force de cisaillement.
NV = viande non vieillie; V = viande vieillie,
C = température de refroidissement; V = vieillissement. a,b moyennes avec différents exposants pour la viande non vieillie représente une différence significative entre les
traitements de refroidissement. A,B moyennes avec différents exposants pour la viande vieillies après cinq jours représente une différence significative
entre les traitements de refroidissement.
y,x moyennes avec différents indices à l’intérieur d’un même traitement de refroidissement représente une différence
significative pour l’effet de vieillissement.
ns : pas de différence significative (p>0.1); *p < 0.05; ***p < 0.001.
Cela implique que peu importe la présence de cold shortening ou non, les sarcomères
subissent par défaut une protéolyse et un attendrissement de la viande (Vieira et Fernández,
2014). Cet attendrissement de la viande, malgré la présence de cold shortening, pourrait
être attribué à un relâchement supplémentaire d’ions calcium du réticulum sarcoplasmique
produit à de basses températures. Ceci fait en sorte d’activer les calpaïnes et engendre un

30
attendrissement de la viande pouvant contourner le problème de cold shortening (Vieira et
Fernández, 2014).
2.2.4. Propriétés organoleptiques
Les propriétés organoleptiques comprennent l’arôme de la viande, la tendreté, la jutosité et
la flaveur (Hocquette et al. 2000 et Nikmaram et al. 2011). Cette dernière est influencée par
plusieurs paramètres dont la composition en acides gras des lipides, l’hydrolyse et
l’oxydation des lipides après l’abattage ainsi que l’oxydation des protéines et les goûts
indésirables (Hocquette et al. 2000). Tous ces facteurs peuvent varier selon divers
paramètres. Par exemple, le taux de gras et la teneur en eau après la cuisson de la viande
peuvent en changer la jutosité (Gregory, 1998). L’épaisseur de gras dorsal doit être
suffisamment élevée pour l’obtention d’une viande moindrement juteuse (Gregory, 1998).
La quantité de gras dépend de l’alimentation soit de sa teneur en énergie, de sa digestibilité,
de son absorption au niveau des intestins, de son métabolisme dans le foie et du transport
des lipides dans le muscle (Sosin-Bzducha et al. 2012). Une alimentation contenant du maïs
aurait tendance à augmenter le gras intramusculaire de la viande comparativement à une
alimentation à base d’orge chez le veau. Cela serait causé par le fait que l’amidon du maïs
est lentement dégradable et augmenterait donc le gras dans la viande (Sosin-Bzducha et al.
2012). Selon l’étude de Leupp et al. (2009), une alimentation contenant des drèches sèches
de distillerie chez le bouvillon, à raison de 30% d’inclusion en période de finition, aurait
tendance à produire une viande plus juteuse et savoureuse. De plus, le gras des animaux a
tendance à être un peu plus jaune lorsqu’ils consomment du maïs par rapport à l’orge
(Boles et al. 2004).
La quantité de gras dans la viande viendrait également jouer sur les paramètres de la
jutosité, la tendreté et la flaveur de la viande (Gregory, 1998 et Hocquette et al. 2000). Plus
nous ajoutons de gras dans la viande hachée, plus cette dernière sera juteuse et plus tendre.
Toutefois, une quantité trop élevée peut diminuer la flaveur de la viande (Tableau 2.14). La
jutosité d’une viande dépend également de la cuisson. Une viande trop cuite deviendra
sèche et ce sera sa capacité en rétention d’eau qui déterminera la jutosité (Gregory, 1998).

31
Tableau 2.14. Effet de la quantité de gras dans la viande hachée sur les qualités
organoleptiques Adapté de Gregory, (1998).
Quantité de gras
9% 20% 28%
Jutosité 7.2a 10.5b 11.3b
Tendreté 9.6a 11.8b 13.1c
Flaveur 9.9a 9.3a 9.0a Un pointage subjectif a été employé : 1 = non appréciée à 15 = très appréciée. a-c
Les lignes sans un exposant commun sont significativement différentes (P = 0.05).
La flaveur de la viande peut être influencée par l’oxydation des lipides. Cette oxydation se
mesure à l’aide du taux de malonaldéhyde présent dans la viande. Le goût de rancidité des
lipides se manifeste de façon plus intense lorsque le taux de TBA est élevé (Nade et al.
2012). Tout au long de la conservation de la viande à 4ºC, la quantité de malonaldéhyde
augmente (Figure 2.10). Pour limiter cette production de malonaldéhyde dans la viande,
une consommation de drèches de distillerie par l’animal pourrait être utile (Nade et al.
2012). En effet, les drèches sont riches en vitamine E (α-tocophérol). Cette vitamine agit
comme antioxydant et pourrait améliorer la conservation de la viande lorsque la ration de
l’animal comprend 100 U.I./veau/jour (Ngapo et Gariépy, 2006; Nade et al. 2012).
Oxydation des gras (TBARS)
Jours
Adapté de De Palo et al. (2013)
Figure 2.10. Effets du conditionnement sur l’oxydation des gras (TBARS) durant
l’entreposage de la viande de veau.
Les lettres différentes sur la même ligne démontrent des différences statistiques pour le même emballage (A–F: P < 0.01;
a–d: P < 0.05). *P < 0.05, **P < 0.01, et ***P < 0.001 représentent les différences statistiques entre les emballages pour
le même jour. MDA = Malondialdéhyde.
Ox
yd
atio
n d
es g
ras
(mg M
DA
/kg)

32
La teneur en eau peut également avoir un impact sur la jutosité de la viande. L’étude de
Sosin-Bzducha et al. (2012) a évalué les caractéristiques de la viande de veau lors d’un
changement d’ingrédients dans la ration. Les veaux étaient de race croisée à prédominance
Holstein âgés entre 10 et 90 jours et séparés en quatre groupes. Chaque de groupe avait une
ration différente, soit une à base d’orge, une à base de maïs-grain, une à 50% d’orge et 50%
d’ensilage de maïs-grain et une à 50% de maïs-grain et 50% d’ensilage de maïs-grain. Les
grains étaient donnés sous forme roulée. Chacune des rations avait la même quantité
d’avoine (18%), de tourteau de soya (21%) et de minéraux. Leur expérience a fait constater
que peu importe l’alimentation du veau, la teneur en eau dans le Musculus longissimus
thoracis se situe autour de 22% (Tableau 2.15).
Tableau 2.15. Composition chimique de la viande de veau, % Adapté de Sosin-Bzducha et
al. (2012).
Groupe
Composition Orge Maïs-grain
50% orge +
50% ensilage de
maïs-grain
50% maïs-grain
+ 50% ensilage
de maïs-grain
Teneur en eau 21.74 22.22 22.08 21.83
La capacité de rétention d’eau peut déterminer également la teneur en eau d’une viande.
Lors de l’étude de Önenç et Kaya (2004), l’étourdissement par percussion a produit plus
d’exsudat, mais pas de façon significative. Cependant, les pertes à la cuisson ont augmenté
avec le vieillissement et elles étaient significativement plus élevées pour les traitements
avec étourdissement (électrique et par percussion) (Tableau 2.16). Cette observation
pourrait être causée par une chute de pH plus rapide. La chute du pH a un rôle significatif
sur la capacité de rétention d’eau ainsi que les pertes en eau à la cuisson (Önenç et Kaya,
2004). Ces deux paramètres sont interreliés, car la capacité de rétention d’eau détermine les
pertes en eau à la cuisson. L’idéal serait d’obtenir une viande dont les pertes à la cuisson
sont faibles afin de ne pas en influencer la jutosité.

33
Tableau 2.16. Capacité de rétention d’eau (CRE) et pertes à la cuisson (PC) selon les
manipulations pré-abattage de jeunes taureaux Holstein Friesian durant le vieillissement de
la viande Adapté de Önenç et Kaya (2004).
Paramètres Vieillissement Sans
étourdissement
Étourdissement
électrique
Étourdissement
par percussion
Écart-
type P
CRE (%) 24 heures 16.32 17.99 15.08 1.78 0.51
7 jours 15.80 15.50 17.80 1.11 0.29
PC (%) 4 jours 22.04 20.01 20.82 1.53 0.78
7 jours 23.01a 26.42a 29.04a 1.74 0.03
14 jours 23.68b 28.12a 30.81a 1.64 0.01
Les moyennes sur une même ligne avec différentes lettres sont significativement différentes (P < 0.05)
Les pertes en eau à la cuisson sont également influencées par les stress, tel qu’un long
transport, avant l’abattage de l’animal (Campbell et al. 2013). Elles ne sont pas influencées
par les traitements de refroidissement de la carcasse. Cependant, les traitements de
température conventionnelle et très rapide démontrent des pertes plus importantes après le
vieillissement de la viande (Vieira et Fernández, 2014). L’étude de Campbell et al. (2013) a
démontré que la moyenne des pertes en eau à la cuisson pour les abattoirs de Québec et
Ontario est respectivement de 21,8% et 19,8% pour la viande de veau de grain.
Ce n’est pas seulement la vitesse de chute du pH et le niveau qu’il atteint qui influencent la
capacité de rétention d’eau de la viande, mais également de la protéolyse et de l’oxydation
des protéines (Huff-Lonergan et al. 2005; De Palo et al. 2013). L’eau est initialement
entreposée dans les cellules incluant les espaces intra et extramyofibrillaires. Après
l’abattage de l’animal, le changement de structure du muscle réduit l’espace disponible
pour l’eau dans les cellules et elle se retrouve donc dans les espaces extramyofibrillaires
(Huff-Lonergan et al. 2005; Ripoll et al. 2013). Il a été démontré par De Palo et al. (2013)
qu’une oxydation élevée des protéines engendre une plus faible capacité de rétention d’eau.
Lorsque de l’oxyation des protéines affecte la capacité de rétention d’eau, il se produit une
conversion de certains acides aminés, notamment l’histidine, en dérivés carbonyl (De Palo
et al. 2013). Ceci fait en sorte de créer des ponts disulfures intra et interprotéines (De Palo

34
et al. 2013). Ces auteurs ont démontré qu’il existe une corrélation négative entre la capacité
de rétention d’eau et l’oxydation des protéines. Donc, lorsque l’oxydation des protéines
augmente, la capacité de rétention d’eau diminue.
Le conditionnement (le type d’emballage) a un impact sur la capacité de rétention d’eau
d’une viande. De Palo et al. (2013) ont évalué notamment la capacité de rétention d’eau et
les pertes en eau en employant deux emballages différents, soit un film Cryovac de 25 µm
d’épaisseur et un film Weegal de 45 µm d’épaisseur pour une viande de veau croisé
Holstein d’un poids vif de 237 kg. Une première différence significative se présente tout au
long de la conservation pour le même emballage. Donc, le vieillissement de la viande a un
impact sur la capacité de rétention d’eau. La seconde différence significative est entre les
deux types d’emballage. À partir du quatrième jour de conservation, le film Weegal permet
d’obtenir une plus grande capacité de rétention d’eau de la viande (Figure 2.11). Donc, un
film plus épais permet réduire l’impact de la conservation sur la capacité de rétention d’eau
grâce à sa perméabilité plus faible à l’oxygène (De Palo et al. 2013). Ceci fait en sorte de
réduire davantage les processus d’altération chimiques et physiques de la viande (De Palo
et al. 2013).
Capacité de rétention d’eau
Jours
Adapté de De Palo et al. (2013)
Figure 2.11. Effets du conditionnement sur la capacité de rétention d’eau durant
l’entreposage de la viande de veau.
Les lettres différentes sur la même ligne démontrent des différences statistiques pour le même emballage (A–F: P < 0.01;
a–d: P < 0.05). *P < 0.05, **P < 0.01, et ***P < 0.001 représentent les différences statistiques entre les emballages pour
le même jour.
Cap
acit
é d
e ré
ten
tion
d’e
au (
%)

35
Nous pouvons observer une étroite relation entre la capacité de rétention d’eau et les pertes
en eau. En effet, plus la capacité de rétention d’eau est faible, plus les pertes en eau sont
importantes (Figure 2.12).
Pertes en eau
Jours
Figure 2.12. Effets du conditionnement sur les pertes en eau durant l’entreposage de la
viande de veau.
Les lettres différentes sur la même ligne démontrent des différences statistiques pour le même emballage (A–F: P < 0.01;
a–d: P < 0.05). *P < 0.05, **P < 0.01, et ***P < 0.001 représentent les différences statistiques entre les emballages pour
le même jour.
Lors de la cuisson de la viande, plusieurs réactions se produisent, notamment la réaction de
Maillard. Cette réaction est fortement contributive pour le goût de la viande, car elle produit
une certaine caramélisation des sucres. Cette caramélisation est produite par la réduction de
sucres se joignant à des acides aminés pour former des N-sucre aminés. Le développement
du goût de la viande dépend de la méthode de cuisson. Une viande rôtie ou grillée aura plus
de saveur qu’une viande mijotée ou cuite au four à micro-ondes. La réaction de Maillard
dépend de la température de cuisson et c’est ce qui explique la raison que les cuissons de
mijotés et aux micro-ondes produisent une viande moins savoureuse. Ceci serait également
causé par un relâchement plus important d’acide inosinique qui augmente la flaveur pour
les viandes rôties ou grillées par exemple. La réaction de Maillard se produit en chauffant
les acides aminés, les peptides, les sucres, les nucléotides et nucléosides de la viande. Les
groupes fonctionnels de la viande comme les protéines, les lipides et les glucides
participent également dans la réaction de Maillard (Gregory, 1998).
Per
tes
en e
au (
%)

36
2.2.5. Analyses sensorielles
Les analyses sensorielles interpellent les perceptions humaines lors de la consommation
d’aliment, notamment la viande (Caine et al. 2003). Lors de la mastication, une multitude
d’interactions sensorielles et physiques se produisent en bouche. La tendreté de la viande
est sans aucun doute le critère le plus important lors de la consommation (Caine et al. 2003;
Destefanis et al. 2008). Les analyses sont faites avec des panélistes entraînés pour
reconnaître les critères de consommation d’un produit (Roeber et al. 2005).
Les différents traitements de refroidissement de la carcasse peuvent avoir un impact sur
quelques éléments perceptibles lors des analyses sensorielles. Les pertes à la cuisson et la
jutosité de la viande ne sont pas influencées par les traitements de refroidissement de la
carcasse (Vieira et Fernández, 2014). Dans l’étude de Vieira et Fernández (2014), les
traitements de refroidissement très rapide et conventionnel ont obtenu des valeurs plus
élevées pour la fermeté, la fibrosité et la masticabilité de la viande comparativement au
refroidissement lent. Toutefois, tel que mentionné dans les sections précédentes, le
vieillissement de la viande vient améliorer favorablement ces trois paramètres lors des
analyses sensorielles.
Selon l’étude de Leupp et al. (2009), l’incorporation de 30% de drèches sèches de distillerie
en finition aurait tendance à produire une viande plus juteuse et savoureuse par rapport à
aucune incorporation (Tableau 2.17). Le persillage (gras intramusculaire) a un effet positif
sur la jutosité et la flaveur de la viande (Gregory, 1998; Leupp et al. 2009). En effet, une
viande sera plus savoureuse et juteuse lorsque le persillage est élevé. Le Canada se base sur
plusieurs critères pour déterminer la classification de la carcasse, dont le persillage (Bœuf
Canada, 2014). Canada A, AA, AAA et Prime (étant le plus élevé) sont classés selon la
présence et l’abondance de persillage dans la viande (Figure 2.13).

37
Tableau 2.17. Pertes à la cuisson et caractéristiques sensorielles des steaks de bouvillons
nourris avec des rations contenant 0 ou 30% de drèches sèches de distillerie avec solubles
en période de croissance et de finition Adapté de Leupp et al. (2009).
Traitements1 Constrates2 P
Item 0:0 30:0 0:30 30:30 SEM2 Croissance Finition
Steaks, n 17 15 16 18 - - - -
Perte à la cuisson, g 50.10 42.80 46.53 45.82 3.02 0.86 0.19 ≥ 0.13
Caractéristiques sensorielles4
Tendreté 6.02 5.81 6.16 6.11 0.16 0.40 0.16 ≥ 0.16
Jutosité 5.84 5.82 5.98 6.03 0.11 0.87 0.10 0.10
Flaveur 6.04 5.74 5.99 6.05 0.08 0.13 0.10 0.10
1 Pourcentage de drèches sèches de distillerie avec solubles dans les rations données en période de croissance et de
finition, respectivement. 2 Croissance = effet principal d’une alimentation avec drèches sèches de distillerie avec solubles durant la période de
croissance (30:0 + 30:30 vs. 0:0 + 0:30); Finition = effet principal d’une alimentation avec drèches sèches de distillerie
avec solubles durant la période de finition (0:30 + 30:30 vs. 0:0 + 30:0). 3 n = 15. 4 Tendreté (8 = extrêmement tendre, 1 = extrêmement dur); jutosité (8 = extrêmement juteux, 1 = extrêmement sec);
flaveur (8 = extrêmement savoureux, 1 = extrêmement fade).
Figure 2.13. Échelle canadienne de persillage de la viande de bœuf.
Source : Bœuf Canada inc. http://www.canadabeef.ca/ca/fr/rt/quality/default.aspx

38
Destefanis et al. (2008) ont comparé la force de cisaillement (Warner-Bratzler) évaluée
mécaniquement et la perception de la tendreté des consommateurs pour la viande de boeuf.
Ils ont créé cinq catégories de tendreté après avoir procédé aux évaluations avec le Warner-
Bratzler. La catégorie 1 correspond à la viande très dure avec une évaluation > 62,59 N; la
catégorie 2 représente la viande dure pour les données entre 52,78 et 62,59 N; la catégorie
3 est pour la viande moyenne dont la tendreté se situe entre 42,87 et 52,68 N; la catégorie
4 représente la viande tendre pour les données entre 42,77 et 32,96 N et la catégorie
5 correspond à la viande très tendre avec une évaluation < 32,96 N. Les résultats de cette
expérience ont démontré que les consommateurs ont de la difficulté à différencier les
catégories 1 et 2 (Dure et Très dure) ainsi que la catégorie 5 de la catégorie 4 (Très tendre
et Tendre) (Tableau 2.18).
Tableau 2.18. Notes distribuées pour la tendreté sensorielle auprès de cinq catégories de
force de cisaillement (FC) Adapté de Destefanis et al. (2008).
FC Classes de
tendreté n Moyenne
de FC
(N)
Moyenne sensorielle
Perception de la tendreté des consommateurs
Catégories Barème (N) 1 2 3 4 5 Très
dure
Dure Moyenne Tendre Très
tendre
% % % % %
1 >62.59 Très dure 92 67.39 2.29A 22.83Bc 39.13B 27.17ABab 7.61Aa 3.26Aa
2 62.59-52.78 Dure 97 58.17 2.53A 18.56Bc 30.93B 35.05Bb 10.31Aa 5.15Aab 3 52.68-42.87 Moyenne 131 48.17 3.11B 0.76Aa 35.11B 26.72ABab 26.72Bb 10.69ABbc
4 42.77-32.96 Tendre 201 37.67 3.42C 6.47Ab 12.44A 29.35ABb 35.82BCb 15.92Bc
5 <32.96 Très tendre 101 28.25 4.16D – – 16.83Aa 50.50Cc 32.67Cd
Valeurs avec des lettres différentes à l’intérieur d’une même colonne sont significativement différentes (a,b = P < 0.05;
A,B = P < 0.01).
Afin d’obtenir de meilleurs résultats, les chercheurs sont passés de cinq catégories à trois,
soit Dure avec > 52,68 N, Moyenne entre 52,68 et 42,87 N et Tendre < 42,87 N. Cette fois-
ci, les consommateurs ont mieux perçu la différence entre les catégories de tendreté. Pour
les catégories Dure et Tendre, la majorité des consommateurs les ont classées correctement.
La tendreté Moyenne a obtenu des résultats répartis dans les trois catégories (Tableau 2.19).
Un changement de 9,8 N (1 kg) peut être perceptible pour des consommateurs aguerris
(Destefanis et al. 2008).

39
Tableau 2.19. Notes distribuées pour la tendreté sensorielle auprès de trois catégories de
force de cisaillement (FC). Adapté de Destefanis et al. (2008) FC Classes de
tendreté
n Moyenne
de FC
(N)
Moyenne
sensorielle
Perception de la tendreté des consommateurs
Catégories Barème (N) 1 - Dure % 2 – Moyenne % 3 – Tendre %
1 >52.68 Dure 189 62.59 2.41A 55.56C 31.22 13.23A
2 52.68-42.87 Moyenne 131 48.17 3.11B 35.88B 26.72 37.40B
3 <42.87 Tendre 302 45.91 3.67C 12.58A 25.17 62.25C
Valeurs avec des lettres différentes à l’intérieur d’une même colonne sont significativement différentes (A,B = P < 0.01).
L’étude de Önenç et Kaya (2004) démontre que la méthode d’étourdissement des animaux,
et ce tout au long du vieillissement de la viande, a significativement un effet sur toutes les
caractéristiques sensorielles (odeur, flaveur, tendreté et acceptabilité générale). En effet,
l’étourdissement par percussion a obtenu significativement de meilleurs résultats
comparativement à aucun étourdissement et un étourdissement électrique (Tableau 2.20).
Tableau 2.20. Caractéristiques sensorielles selon les manipulations pré-abattage durant le
vieillissement de la viande1 Adapté de Önenç et Kaya (2004)
Paramètres Vieillissement Sans
étourdissement
Étourdissement
électrique
Étourdissement
par percussion
Écart-
type P
Odeur
24 heures 45.10 b 48.40 a 52.10 a 1.94 0.05
4 jours 47.46 b 47.10 b 56.80 a 2.06 0.03
7 jours 49.50 b 50.90 b 55.30 a 1.59 0.04
14 jours 46.90 b 51.80 a 56.00 a 1.74 0.01
Flaveur
24 heures 40.50 c 44.00 b 52.90 a 1.93 0.01
4 jours 45.30 c 46.20 b 55.40 a 2.07 0.01
7 jours 49.80 b 49.10 b 57.30 a 2.03 0.01
14 jours 47.80 c 49.90 b 58.00 a 2.10 0.01
Tendreté
24 heures 36.60 b 43.50 b 54.80 a 1.93 0.01
4 jours 42.50 b 46.40 b 55.60 a 2.93 0.01
7 jours 44.50 b 47.80 b 58.10 a 2.75 0.01
14 jours 45.10 b 50.40 a 57.30 a 2.91 0.02
Acceptabilité
globale
24 heures 41.90 b 46.90 ab 51.40 a 2.31 0.03
4 jours 46.90 b 50.00 ab 55.30 a 1.98 0.02
7 jours 50.50 b 52.90 ab 58.00 a 1.97 0.04
14 jours 50.70 b 51.10 b 58.30 a 2.03 0.02 Les moyennes sur une même ligne avec différentes lettres sont significativement différentes (P < 0.05) 1
Les notes sont données sur une échelle de 10 avec des explications (de 1 = extrêmement mauvais à 10 extrêmement
bon). Dix sessions d’évaluation ont eu lieu. Huit panélistes ont assisté à chaque évaluation. Les valeurs sont le total des
points pour les huit panélistes pour chaque session d’évaluation pour chacune des caractéristiques sensorielles.

40
2.3. ALIMENTATION
Tel que mentionné dans les sections précédentes, l’alimentation a un certain impact sur la
qualité de la viande dépendamment de sa teneur en énergie, en protéines et en lipides.
L’alimentation des animaux élevés pour leur viande peut être composée de céréales (maïs,
orge, avoine), de sous-produits et de suppléments protéiques.
2.3.1. Les céréales
Les céréales procurent une source d’amidon pouvant affecter la qualité de la viande des
veaux au niveau de la composition des acides gras (Sosin-Bzducha et al. 2012). La
digestion de l’orge est faite en très grande partie dans le rumen tandis que celle du maïs-
grain est plus lente. Cela s’explique par le fait qu’il y ait plus d’amidon passant à l’intérieur
du petit intestin avec l’utilisation du maïs-grain (Sosin-Bzducha et al. 2012).
Actuellement, la principale céréale utilisée dans l’alimentation du veau de grain au Québec
est le maïs. Les veaux peuvent être alimentés avec du maïs-grain contenant différentes
teneurs en eau. Plus l’humidité du grain augmente, plus il fermente rapidement dans le
rumen pour produire des acides gras (Sosin-Bzducha et al. 2012). Cela fait en sorte que les
grains de maïs humides sont mieux digérés que les grains secs. Donc, il y a moins de pertes
de source d’amidon dans le gros intestin et finalement dans les fèces (Sosin-Bzducha et al.
2012).
Wright et Lackey (2012) ont comparé le maïs-grain, le tourteau de soya et les drèches de
distillerie sèches et humides. Les drèches sèches de distillerie humides présentent un
avantage au niveau de protéines comparativement au maïs et pourraient être une alternative
au tourteau de soya (Tableau 2.21).

41
Tableau 2.21. Évaluation des sous-produits Adapté de Wright et Lackey (2012)
Aliment % de % d’UNT z % PB y % d’UNT % PB
MS (rapporté à la MS) (de l’aliment)
Maïs-grain 85.0 89.0 9.6 75.7 8.2
Tourteau de soya à 48% 89.0 84.0 53.9 74.8 48.0
Sous-produits de mouture et de transformation
Drèches de distillerie - humides 30.0 84.0 29.5 25.2 8.9
Drèches de distillerie - sèches 92.0 83.7 29.5 77.0 27.1
z UNT = Unités nutritives totales. y PB = Protéines brutes.
Boles et al. (2004) ont démontré qu’une alimentation comprenant de l’orge améliore la
classification et le poids de la carcasse chez le bœuf ainsi que la surface de l’œil de longe
comparativement à l’utilisation du maïs. Les animaux nourris avec de l’orge ont tendance à
produire une viande plus rouge. Cela est possiblement dû à l’augmentation des pigments
hèmes dans les carcasses des animaux (Boles et al. 2004). Même lors de l’utilisation de
sous-produit de l’orge en combinaison avec des drèches de distillerie, la viande des
bouvillons est plus rouge que si nous alimentons avec du gluten de maïs (Roeber et al.
2005). Selon la variété d’orge employée, la viande résultante d’une alimentation composée
de cette céréale a tendance à produire une viande plus pâle (augmentation de l’indice L*)
comparativement au maïs (Boles et al. 2004).
Toutefois, Noon et al. (1998) ont expérimenté divers ratios de maïs et d’orge ronde dans
l’alimentation des veaux de boucherie et ont obtenu des résultats différents. Les ratios de
maïs : orge étaient : 100:0, 75:25, 50:50, 25:75 et 0:100 plus un supplément proétique à
43,6% de protéines brutes. En effet, plus la quantité d’orge augmente dans la ration et plus
le poids de la carcasse diminue. Cette diminution est également observée au niveau du
rendement de la carcasse et du poids de la longe. Après avoir dépassé le ratio 50:50, la
classification des carcasses a tendance à diminuer. Aucune différence significative n’a été
observée pour la couleur (Tableau 2.22).

42
Tableau 2.22. Effets du maïs sur les ratios d’orge dans la ration sur les caractéristiques de la
carcasse des veaux de boucherie nourris aux grains Adapté de Noon et al. (1998).
Rations (% maïs : % orge)
Paramètres 100:0 75:25 50:50 25:75 0:100 SEMz LSy
n 8 8 6 8 7
Poids d’abattage (kg) 260.0 256.5 257.6 252.5 252.5 3.06 NS
Poids carcasse (kg) 163.7 158.8 158.2 152.6 150.5 1.66 L**
Rendement carcasse (%) 63.0 61.9 61.4 60.5 59.7 0.58 L**
Couleurx 42.6 43.0 42.4 40.8 45.7 1.31 NS
Classificationw 2.0 1.9 3.0 3.1 3.0 0.53 LT
A1, # de veaux 0 1 0 0 0
A2, # de veaux 8 7 5 5 5
A3, # de veaux 0 0 0 1 0
B1, # de veaux 0 0 0 0 1
B2, # de veaux 0 0 2 2 1
Longe
Poids (kg) 8.18 7.97 7.74 7.70 7.23 0.19 L**
pH 5.58 5.73 5.73 5.75 5.63 0.10 NS
Couleur Minoltav 45.9 43.7 43.8 41.7 43.6 1.15 LT
z SEM = erreur standard de la moyenne. y LS = niveau significatif, NS = non significatif, L = effet linéaire, **P < 0.01, T = tendance, P < 0.10. x Couleur mesurée à un abattoir par un inspecteur fédéral. w Classification : A1 = 1, A2 = 2, A3 = 3, B1 = 4, A1 = 5, B2 = 6. v Couleur déterminée au laboratoire avec un appareil Minolta D-65 (valeur de L* sur l’échelle de Hunter)
La digestibilité apparente des rations diminuait avec l’augmentation de la proportion
d’orge. La dégradation ruminale de l’amidon pour l’orge est très élevée (90%)
comparativement à celle du maïs (60%). Lorsque les veaux sont nourris avec du maïs, une
augmentation de la quantité d’amidon est acheminée dans le petit intestin. C’est à cet
endroit que se produit la formation de glucose grâce à la digestion enzymatique. Ce glucose
est énergétiquement plus efficace que la métabolisation des acides gras volatils dans le
rumen (Noon et al. 1998). Ainsi, les veaux nourris avec du maïs ont consommé moins de
matière sèche que ceux avec de l’orge puisque le processus de digestion était plus efficace
(Guertin et al. 1995; Noon et al. 1998). De plus, les animaux n’ont pas eu à puiser dans
leurs réserves corporelles pour la gluconéogenèse faisant en sorte de produire une carcasse
plus lourde (Noon et al. 1998). Le contenu en énergie et en amidon est significativement
plus élevé dans le maïs que dans l’orge (Tableau 2.23). L’énergie retenue issue d’une
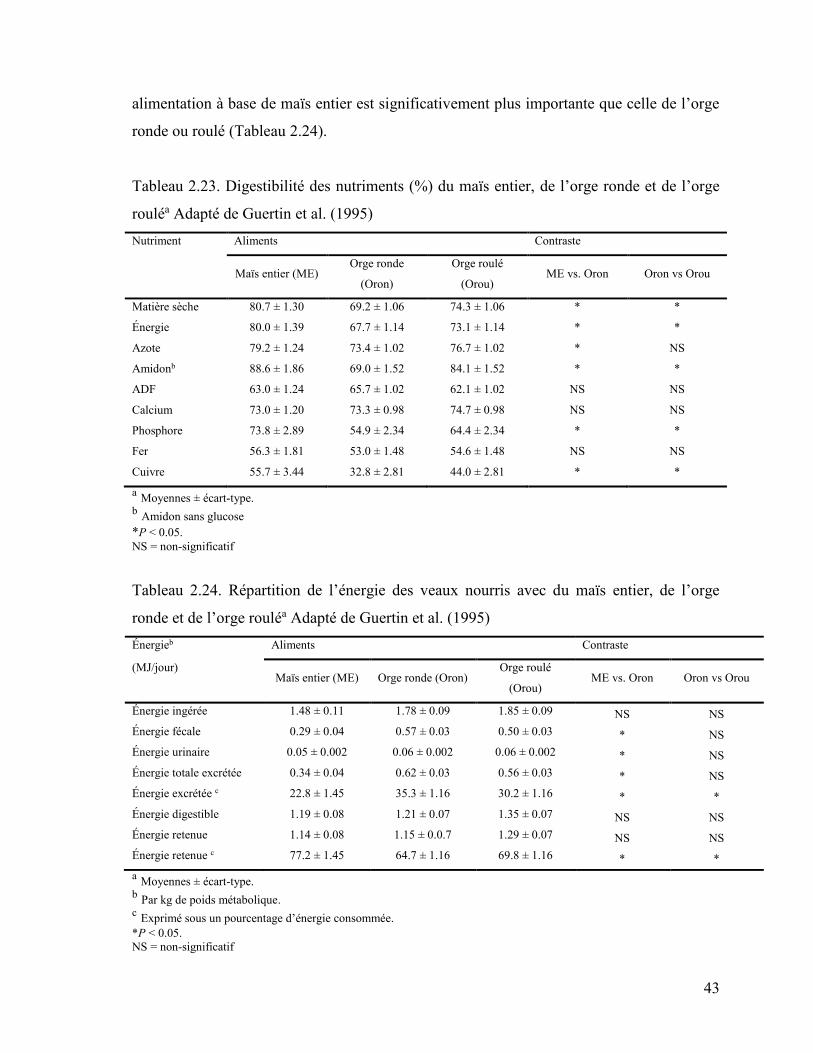
43
alimentation à base de maïs entier est significativement plus importante que celle de l’orge
ronde ou roulé (Tableau 2.24).
Tableau 2.23. Digestibilité des nutriments (%) du maïs entier, de l’orge ronde et de l’orge
rouléa Adapté de Guertin et al. (1995)
Nutriment Aliments Contraste
Maïs entier (ME)
Orge ronde
(Oron)
Orge roulé
(Orou) ME vs. Oron Oron vs Orou
Matière sèche 80.7 ± 1.30 69.2 ± 1.06 74.3 ± 1.06 * *
Énergie 80.0 ± 1.39 67.7 ± 1.14 73.1 ± 1.14 * *
Azote 79.2 ± 1.24 73.4 ± 1.02 76.7 ± 1.02 * NS
Amidonb 88.6 ± 1.86 69.0 ± 1.52 84.1 ± 1.52 * *
ADF 63.0 ± 1.24 65.7 ± 1.02 62.1 ± 1.02 NS NS
Calcium 73.0 ± 1.20 73.3 ± 0.98 74.7 ± 0.98 NS NS
Phosphore 73.8 ± 2.89 54.9 ± 2.34 64.4 ± 2.34 * *
Fer 56.3 ± 1.81 53.0 ± 1.48 54.6 ± 1.48 NS NS
Cuivre 55.7 ± 3.44 32.8 ± 2.81 44.0 ± 2.81 * *
a Moyennes ± écart-type. b Amidon sans glucose
*P < 0.05. NS = non-significatif
Tableau 2.24. Répartition de l’énergie des veaux nourris avec du maïs entier, de l’orge
ronde et de l’orge rouléa Adapté de Guertin et al. (1995)
Énergieb Aliments Contraste
(MJ/jour) Maïs entier (ME) Orge ronde (Oron)
Orge roulé
(Orou) ME vs. Oron Oron vs Orou
Énergie ingérée 1.48 ± 0.11 1.78 ± 0.09 1.85 ± 0.09 NS NS
Énergie fécale 0.29 ± 0.04 0.57 ± 0.03 0.50 ± 0.03 * NS
Énergie urinaire 0.05 ± 0.002 0.06 ± 0.002 0.06 ± 0.002 * NS
Énergie totale excrétée 0.34 ± 0.04 0.62 ± 0.03 0.56 ± 0.03 * NS
Énergie excrétée c 22.8 ± 1.45 35.3 ± 1.16 30.2 ± 1.16 * *
Énergie digestible 1.19 ± 0.08 1.21 ± 0.07 1.35 ± 0.07 NS NS
Énergie retenue 1.14 ± 0.08 1.15 ± 0.0.7 1.29 ± 0.07 NS NS
Énergie retenue c 77.2 ± 1.45 64.7 ± 1.16 69.8 ± 1.16 * *
a Moyennes ± écart-type. b Par kg de poids métabolique. c Exprimé sous un pourcentage d’énergie consommée. *P < 0.05.
NS = non-significatif

44
2.3.2. Les sous-produits
Le tourteau de canola est le sous-produit qui découle de l’extraction de l’huile des graines
de canola (Lardy, 2002). Une étude faite avec deux groupes de génisses laitières a démontré
que l’appétence du tourteau de canola est plus élevée que celle du tourteau de soya. En
effet, en trois minutes, le premier groupe de génisses avec le tourteau de canola avait
consommé 221 g et l’autre groupe avec le tourteau de soya en avait consommé 96 g. Cela
pourrait être à cause de sa teneur élevée en sucrose (Conseil Canadien du Canola, 2014). Il
s’agit d’une consommation 2,3 fois plus grande que le tourteau de soya.
Ce sous-produit est utilisé en production laitière pour son appétence élevée et son contenu
en protéines de haute qualité. Il est riche en histidine, méthionine, cystine et thréonine
(Conseil Canadien du Canola, 2009). Le profil d’acides aminés du tourteau de canola est
bien équilibré entre les acides aminés par rapport à d’autres sous-produits. Le tableau 2.25
démontre la comparaison entre le tourteau de canola, le tourteau de soya et les drèches
sèches de distillerie avec solubles.

45
Tableau 2.25. Comparaison des acides aminés des sous-produits (par rapport aux protéines
du lait)1 Adapté du Conseil Canadien du Canola, 2009.
Sous-produits
Acides aminés Tourteau de canola Tourteau de soya Drèches sèches de
distillerie avec solubles
Arginine 197 225 149
Histidine 138 111 120
Isoleucine 83 89 86
Leucine 82 88 130
Lysine 84 87 37
Méthionine 95 58 87
Phénylalanine 103 116 34
Thréonine 113 98 102
Tryptophane 115 93 77
Valine 88 78 96
1Les chiffres en gras correspond à l’acide aminé limitant du sous-produit.
Ces résultats démontrent qu’il peut y avoir une interaction entre les sous-produits lorsqu’ils
sont incorporés ensemble dans l’alimentation. Par exemple, les drèches sèches de distillerie
sont faibles en lysine et riches en leucine étant le cas inverse pour le tourteau de canola.
Donc, une synergie entre l’utilisation des sous-produits pourrait maximiser les
performances animales.
Une portion des protéines du tourteau de canola semble également être non dégradable au
niveau du rumen (Conseil Canadien du Canola, 2009). Par contre, le taux de protéine non
dégradable au rumen est apparemment similaire même légèrement inférieur à celui du
tourteau de soya (Lardy, 2002).
Le tourteau de canola contient trois fois plus de fibres que le tourteau de soya. C’est ce qui
fait en sorte que sa digestibilité lui est inférieure et par conséquent, la teneur en énergie plus
faible (Lardy, 2002). L’incorporation de 25 % tourteau de canola dans l’alimentation de

46
finition augmente le gain moyen quotidien réduisant ainsi le temps de production (Lardy,
2002).
Les drèches de distillerie sont des sous-produits de la production d’éthanol. Lorsque
l’amidon des grains de maïs est fermenté pour produire de l’éthanol, les résidus sont
environ trois fois plus concentrés que le grain original en protéines, gras et acide gras
insaturé, fibres et en phosphore (Klopfestein et al. 2008; Nade et al. 2012). Elles ont une
teneur élevée en vitamines telles le groupe complexe B, A, D et E (Roeber et al. 2005;
Nade et al. 2012). L’énergie de ce sous-produit provient notamment de son contenu en
fibres résiduelles hautement digestibles. Ces fibres permettent de réduire l’acidose dans les
rations riches en grains (Lackey, 2008). La hausse de la production d’éthanol augmente
également la disponibilité des drèches de distillerie. Cela laisse donc une source
d’alimentation en grande disponibilité pour les animaux. Les drèches de distillerie sont
considérées comme des ingrédients dans les rations de la production bovine (Roeber et al.
2005). Une partie des protéines des drèches de distillerie avec solubles passe dans le rumen
sans être dégradée pour se déverser directement dans le duodénum, et ce, environ 2,6 fois
de plus comparativement au tourteau de soya (Klopfestein et al. 2008).
Kelzer et al. (2010) ont caractérisé les fractions de protéines (A, B1, B2, B3 et C) et ont
évalué les différences de protéines non dégradables au niveau du rumen ainsi que la portion
digestible. Pour ce faire, ils ont évalué sept sous-produits de maïs. Toutes les analyses se
sont produites sur une base de matière sèche. Le pourcentage de protéines brutes pour les
germes était 12,7%, 26,9% pour les drèches sèches de distillerie sans chauffage avant
fermentation (DSDS1), 45,4% pour les drèches sèches de distillerie avec une haute teneur
en protéines (DSDHP), 12,7% pour le son, 30,2% pour les drèches humides de distillerie
avec solubles, 23,1% pour le gluten humide de maïs (GHM) et 26,0% pour les drèches
sèches de distillerie avec chauffage avant la fermentation (DSDS2). Les DSDS2 et DSDHP
sont les deux sous-produits ayant le plus de protéines non digestibles au rumen, mais la
digestibilité totale des protéines se retrouve avec les DSDHP (Tableau 2.26).
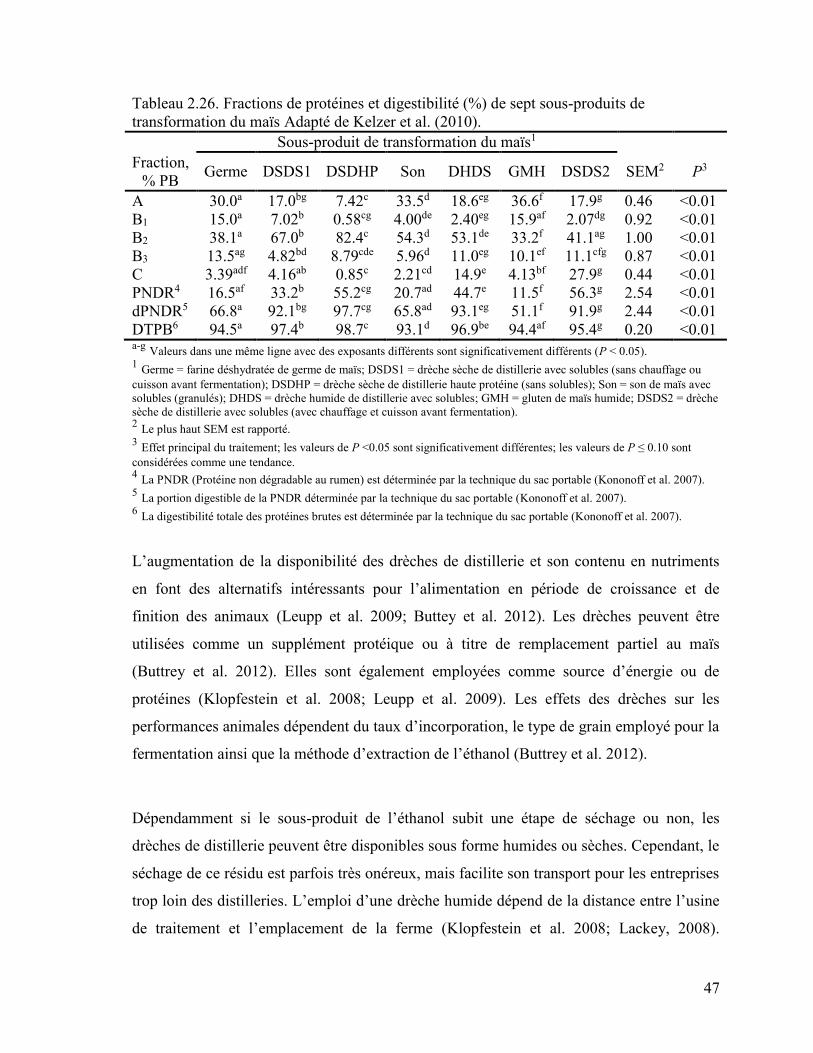
47
Tableau 2.26. Fractions de protéines et digestibilité (%) de sept sous-produits de
transformation du maïs Adapté de Kelzer et al. (2010).
Sous-produit de transformation du maïs1
Fraction,
% PB Germe DSDS1 DSDHP Son DHDS GMH DSDS2 SEM2 P3
A 30.0a 17.0bg 7.42c 33.5d 18.6eg 36.6f 17.9g 0.46 <0.01
B1 15.0a 7.02b 0.58cg 4.00de 2.40eg 15.9af 2.07dg 0.92 <0.01
B2 38.1a 67.0b 82.4c 54.3d 53.1de 33.2f 41.1ag 1.00 <0.01
B3 13.5ag 4.82bd 8.79cde 5.96d 11.0eg 10.1ef 11.1cfg 0.87 <0.01
C 3.39adf 4.16ab 0.85c 2.21cd 14.9e 4.13bf 27.9g 0.44 <0.01
PNDR4 16.5af 33.2b 55.2cg 20.7ad 44.7e 11.5f 56.3g 2.54 <0.01
dPNDR5 66.8a 92.1bg 97.7cg 65.8ad 93.1eg 51.1f 91.9g 2.44 <0.01
DTPB6 94.5a 97.4b 98.7c 93.1d 96.9be 94.4af 95.4g 0.20 <0.01 a-g Valeurs dans une même ligne avec des exposants différents sont significativement différents (P < 0.05). 1 Germe = farine déshydratée de germe de maïs; DSDS1 = drèche sèche de distillerie avec solubles (sans chauffage ou
cuisson avant fermentation); DSDHP = drèche sèche de distillerie haute protéine (sans solubles); Son = son de maïs avec
solubles (granulés); DHDS = drèche humide de distillerie avec solubles; GMH = gluten de maïs humide; DSDS2 = drèche
sèche de distillerie avec solubles (avec chauffage et cuisson avant fermentation). 2 Le plus haut SEM est rapporté. 3 Effet principal du traitement; les valeurs de P <0.05 sont significativement différentes; les valeurs de P ≤ 0.10 sont
considérées comme une tendance. 4 La PNDR (Protéine non dégradable au rumen) est déterminée par la technique du sac portable (Kononoff et al. 2007). 5 La portion digestible de la PNDR déterminée par la technique du sac portable (Kononoff et al. 2007). 6 La digestibilité totale des protéines brutes est déterminée par la technique du sac portable (Kononoff et al. 2007).
L’augmentation de la disponibilité des drèches de distillerie et son contenu en nutriments
en font des alternatifs intéressants pour l’alimentation en période de croissance et de
finition des animaux (Leupp et al. 2009; Buttey et al. 2012). Les drèches peuvent être
utilisées comme un supplément protéique ou à titre de remplacement partiel au maïs
(Buttrey et al. 2012). Elles sont également employées comme source d’énergie ou de
protéines (Klopfestein et al. 2008; Leupp et al. 2009). Les effets des drèches sur les
performances animales dépendent du taux d’incorporation, le type de grain employé pour la
fermentation ainsi que la méthode d’extraction de l’éthanol (Buttrey et al. 2012).
Dépendamment si le sous-produit de l’éthanol subit une étape de séchage ou non, les
drèches de distillerie peuvent être disponibles sous forme humides ou sèches. Cependant, le
séchage de ce résidu est parfois très onéreux, mais facilite son transport pour les entreprises
trop loin des distilleries. L’emploi d’une drèche humide dépend de la distance entre l’usine
de traitement et l’emplacement de la ferme (Klopfestein et al. 2008; Lackey, 2008).

48
Toutefois, les drèches humides ont une durée d’utilisation très limitée entre trois et cinq
jours. Des problèmes de moisissures peuvent donc apparaître en dépassant ce délai (Myer,
2011). Les coûts de transport sont également à considérer dans les calculs car les drèches
humides incluent une grande partie en eau dans le sous-produit. Toutefois, les drèches
humides sont plus efficaces que les sèches (Klopfestein et al. 2008; Lackey, 2008). Afin
d’avoir une utilisation économique de ce sous-produit, il est recommandé de se situer à une
distance maximale de 320 km d’une usine de traitement (Thomas et al, 2010).
Avec l’utilisation des drèches humides de distillerie, il est plus avantageux de les inclure
entre 10 et 20% de l’alimentation pour obtenir des valeurs optimales en période de finition.
Lors de l’emploi de drèches sèches, un apport de 20% semble donner des résultats
similaires (Klopfenstein et al. 2008).
Buttrey et al. (2012) ont évalué l’incorporation de maïs sec roulé et des drèches sèches de
distillerie comme source de supplément pour les bouvillons allant aux pâturages de blé
d’hiver. Trois groupes sont observés soit le groupe témoin sans supplément; le groupe avec
un supplément de maïs sec roulé à raison de 0,5% du poids corporel et le groupe avec des
drèches sèches de distillerie également comme supplément à 0,5% du poids corporel. La
supplémentation de drèches sèches de distillerie a permis d’obtenir un poids final et un gain
de poids significativement plus élevé (Tableau 2.27). Cela peut s’expliquer par le fait que
les drèches de distillerie ont de meilleurs niveaux énergétiques grâce à leur teneur plus
élevée en huiles ou matières grasses et de meilleures teneurs en protéines (Lackey, 2008).
Tableau 2.27. Effets des suppléments sur les performances des bouvillons Hereford aux
pâturages de blé d’hiver Adapté de Buttrey et al. (2012)
CON MSR DSD P
Poids initial, kg 199 198 197 0.79
Poids final, kg 360b 363b 374a 0.04
Gain de poids, kg/j 1.29b 1.31b 1.40a 0.002 1 Traitements expérimentaux : CON = contrôle; MSR = maïs sec roulé; DSD = drèche sèche de distillerie. a,b
Au sein du même rangée, les moyennes sans exposant commun sont significativement différents (P ≤ 0.05).

49
Au niveau de la viande, l’utilisation des drèches sèches de distillerie a une influence sur la
couleur. Le a* de la viande de bouvillons nourris avec 30% de drèches sèches de distillerie
diminue plus rapidement que celle produite sans drèches. Cela pourrait être causé par
l’augmentation de l’oxydation des acides gras non saturés et de la réduction du système
enzymatique contrôlant le taux de metmyoglobine dans la viande (Leupp et al. 2009).
Roeber et al. (2005) ont observé une valeur plus élevé du a* lorsque les bouvillons étaient
nourris avec 12,5%, 25% ou 50% de drèches sèches de distillerie comparativement au
tourteau de soya. De plus, le L* a tendance à augmenter lorsque le taux d’incorporation des
drèches s’élevé autant pour les sèches que les humides (Tableau 2.28).
Roeber et al. (2005) ont également évalué la force de cisaillement et les caractéristiques
sensorielles de la viande des bouvillons nourris avec différents niveaux de drèches de
distillerie sèches et humides comparativement au tourteau de soya. Aucune différence
significative n’a pas été observée lors des deux expériences (Tableau 2.29). Cela signifie
que les drèches de distillerie pourraient remplacer le tourteau de soya à cet égard.

50
Tableau 2.28. Effets des drèches de distillerie sur la couleur objective et les pointages sur
l’apparence subjective de steaks après une simulation d’étalement sur étagère de 138 heures
(Expérience 1 et 2). Adapté de Roeber et al. (2005)
Traitementsa L*b a*b b*b % de steaks «modérément
inacceptable»c
Exp. 1
Tourteau de soya 31.41 5.24z 10.39 c 75.0 x
12,5% DSD 31.35 6.33 xy 11.36 x 64.6 x
25% DSD 32.48 5.70 yz 10.96 yc 72.7 x
50% DSD 32.68 5.69 yz 11.12 xyc 72.9 x
25% DHD 32.36 7.30 x 11.94 x 45.8 y
50% DHD 32.73 6.09 yz 11.49 xy 70.8 x
SEM 0.833 0.452 0.309 0.50
P-value 0.733 <0.05 <0.05 <0.05
Exp. 2
Urée 29.79 y 7.70 x 11.54 40.6 y
Tourteau de soya 31.31 x 7.18 x 11.64 40.6 y
10% DSD 29.77 y 7.17 x 11.60 46.7 y
20% DSD 31.92 x 5.84 yz 11.24 59.4 xy
40% DSD 32.34 x 5.32 z 10.93 71.9 x
10% DHD 30.16 y 7.55 x 11.43 43.8 y
20% DHD 31.28 xy 6.84 xy 11.53 46.9 y
40% DHD 32.38 x 5.17 z 11.19 75.0 x
SEM 0.593 0.414 0.281 0.45
P-value <0.05 <0.05 0.579 <0.05 a
DSD = drèche sèche de distillerie; DHD = drèche humide de distillerie. b L* = valeurs mesurant du noir au blanc (L* élevé indique une couleur plus blanche); a* = valeurs mesurant le spectre
rouge (a* élevé indique une couleur plus rouge); b* = valeurs mesurant le spectre jaune (b* élevé indique une couleur plus
jaune). c Pointage moyen d’un panéliste de 3 ou moins pour un steak «modérément inacceptable» étant considéré au point de
recevoir une réduction pour une vente rapide. x, y, z Au sein de l’expérience et de la colonne, les moyennes de moindre carrés n’ayant pas d’exposant commun sont
différents, P < 0.05.

51
Tableau 2.29. Effets des drèches de distillerie sur la force de cisaillement (Expérience 1 et
2). Adapté de Roeber et al. (2005)
Traitementsa Force de cisaillement, kg
Exp. 1
Tourteau de soya 1.47
12.5% DSD 1.33
25% DSD 1.42
50% DSD 1.65
25% DHD 1.45
50% DHD 1.47
SEM 0.066
P-value 0.200
Exp. 2
Urée 2.47
Tourteau de soya 1.48
10% DSD 1.45
20% DSD 1.46
40% DSD 1.38
10% DHD 1.50
20% DHD 1.40
40% DHD 1.48
SEM 0.072
P-value 0.325 a
DSD = drèche sèche de distillerie; DHD = drèche humide de distillerie.
Concernant les analyses sensorielles, les panélistes ont évalué la tendreté, la jutosité et la
flaveur de la viande. Ils devaient noter sur une échelle de 1 à 9, 9 étant le meilleur, ces trois
critères. Lorsqu’une viande ne les satisfaits pas, des précisions étaient demandées à savoir
lequel des trois critères était la raison de leur choix. Pour l’expérience 1, l’incorporation de
25% de drèches humides de distillerie a obtenu le plus faible pourcentage de panélistes non
satisfaits. Pour l’expérience 2, il s’agit de l’incorporation de 10% de drèches humides de
distillerie qui a connu le plus faible pourcentage de panélistes non satisfaits avec la viande
(Tableau 2.30).

52
Tableau 2.30. Effets des drèches de distillerie sur les paramètres sensoriels (Expérience 1 et 2 Adapté de Roeber et al. (2005).
Traitementsa
Tendretéb
Aime/N’aime
pas
Jutositéb
Aime/N’aime
pas
Flaveurb
Aime/N’aime
pas
% (Nb.) de
panélistes non
satisfaits avec
l’échantillonc
Pourcentage de panélistes non
satisfaits précisément avec :
Tendreté Jutosité Flaveur
Exp. 1
Tourteau de soya 5.51 5.49 5.74 47.4 (45) 60.0 40.0 53.3
12.5% DSD 6.07 5.81 6.31 38.9 (37) 45.9 37.8 40.5
25% DSD 5.62 5.50 5.99 45.3 (43) 62.8 44.2 39.5
50% DSD 5.56 5.44 5.75 49.5 (47) 68.1 51.1 46.8
25% DHD 6.15 6.05 6.12 34.7 (32) 56.3 50.0 50.0
50% DHD 5.47 5.22 5.86 50.5 (48) 52.1 47.9 58.3
SEM 0.30 0.31 0.23 0.60
P-value 0.081 0.083 0.224 0.165
Exp. 2
Urée 6.25 5.79 6.38 34.7 (33) 51.5 33.3 45.5
Tourteau de soya 6.15 5.85 6.29 33.7 (31) 77.4 38.7 58.1
10% DSD 6.31 5.81 5.98 37.9 (36) 50.0 61.1 50.0
20% DSD 6.04 5.75 6.16 36.8 (35) 54.3 34.3 57.1
40% DSD 6.39 5.86 6.14 38.9 (37) 51.4 51.4 48.6
10% DHD 6.35 6.13 6.39 29.5 (27) 48.1 25.9 48.1
20% DHD 6.00 5.68 5.95 41.1 (38) 50.0 55.3 39.5
40% DHD 6.13 5.85 6.05 34.7 (33) 48.5 45.5 42.4
SEM 0.22 0.23 0.22 0.50
P-value 0.852 0.885 0.487 0.785 a
DSD = drèche sèche de distillerie; DHD = drèche humide de distillerie. b Tendreté, jutosité et flavour Aime/N’aime pas : 1 = N’aime pas extrêmement; 9 = Aime extrêmement. c Pourcentage de panélistes ayant indiqué que l’échantillon était, en général, inacceptable.

53
CONCLUSION
La viande est issue d’un muscle. Après l’abattage, l’acidification du muscle dépend
principalement des réserves de glycogène musculaire. La quantité de glycogène est
influencée par plusieurs facteurs, notamment par la teneur en énergie au niveau de
l’alimentation. Cette énergie peut provenir de plusieurs sources, notamment des céréales ou
des sous-produits. Le maïs est souvent présent dans l’alimentation bovine pour la
production de viande. Lors de son remplacement ou d’une substitution par une autre céréale
comme l’orge, il faut prévoir une augmentation de la consommation de matière sèche. En
effet, le maïs contient une énergie retenue significativement plus importante que celle de
l’orge et une meilleure digestibilité. Il en est de même lors de l’introduction de sous-produit
pour remplacement le maïs. Il faut également apporter une attention particulière à toutes
sources pouvant causer un épuisement des réserves de glycogène. Puisque le pH ultime
dépend de ces réserves, plusieurs paramètres de la qualité de la viande peuvent varier.
Donc, le pH est l’élément-clé pour obtenir une qualité de viande optimale.

54

55
LISTE DES OUVRAGES CITÉS
Abril, M., Campo, M.M., Önenç, A., Sañudo, C., Albertí, P., and Negueruela, A.I.
2001. Beef colour evolution as a function of ultimate pH. Meat Sci. 58: 69-78.
Bœuf Canada inc. 2014. http://www.canadabeef.ca/ca/fr/rt/quality/default.aspx. (Page
consultée l2 novembre 2014).
Boles, J. A., Bowman, J. G., Surber, L. M. M.and Boss, D. L. 2004. Effects of barley
variety fed to steers on carcass characteristics and color of meat. J. Anim. Sci. 82: 2087-
2091.
Buttrey, E. K., McCollum III, F. T., Jenkins, K. H., Patterson, J. M., Clark, B. E.,
Luebbe, M. K., Lawrence, T. E. and MacDonald, J. C. 2012. Use of dried distillers
grains throughout a beef production system : Effects on stocker and finishing performance,
carcass characteristics, and fatty acid composition of beef. J. Anim. Sci. 90: 2381-2393.
Caine, W.R., Aalhus, J.L., Best, D.R., Dugan, M.E.R., and Jeremiah, L.E. 2003.
Relationship of texture profile analysis and Warner-Bratzler shear force with sensory
characteristics of beef rib steaks. Meat Sci. 64: 333–339.
Campbell, C. P., Haley, J., Swanson, K. C. and Mandell, I. B. 2013. Packing plant
differences in meat quality for grain-fed veal. Can. J. Anim. Sci. 93: 205-215.
Conseil Canadien du Canola. 2014. http://fr.canolacouncil.org. (Page consultée le 25 avril
2014).
Conseil Canadien du Canola. 2009. Canola meal feed industry guide, 4e Édition.
http://www.canolacouncil.org/media/503589/canola_guide_english_2009_small.pdf. p. 48.
(Page consultée le 25 avril 2014)
CPAQ. 1999. Guide veau lourd. Conseil des productions animales du Québec. 271 pages.
De Palo, P., Maggiolino, A., Centoducati, P., and Tateo, A. 2013. Effects of two
different packaging materials on veal calf meat quality and shelf life. J. Anim. Sci. 91:
2920-2930.
Destefanis, G., Brugiapaglia, A., Barge, M.T. and Dal Molin, E. 2008. Relationship
between beef consumer tenderness perception and Warner–Bratzler shear force. Meat Sci.
78: 153–156.
England, E. M., Fisher, K. D., Wells, S. J., Mohrhauser, D. A., Gerrard D. E. and
Weaver, A. D. 2012. Postmortem titin proteolysis is influenced by sarcomere length in
bovine muscle. J. Anim. Sci. 90: 989-995.
Faucitano, L., Ielo, M.C., Ster, C., Lo Fiego, D.P., Methot, S., and Saucier L. 2010. Shelf life of pork from five different quality classes. Meat Sci. 84: 466-469.

56
Fornasier, A. 2002. Portrait de l’industrie du veau québécois. Colloque sur le veau lourd.
Centre de référence en agriculture et agroalimentaire du Québec. 18 pages.
FPBQ. 2014. Veau de grain. Fédération des producteurs de bovins du Québec.
www.bovins.qc.ca. (Page consultée le 8 mars 2014 et le 1er octobre 2014).
Gregory, N. G. 1998. Animal welfare and meat science. CABI Publishing. Wallingford,
Royaume-Uni.
Guertin, G., Lachance, B., Pelletier, G., St-Laurent, G.J., Roy, G.L. and Petitclerc, D.
1995. Effects of photoperiod and feeding whole maize, whole barley, or rolled barley on
growth performance and diet digestibility in veal calves. Livest. Prod. Sci. 44: 27-36.
Guignot, F., Quilichini, Y., Renerre, M., Lacourta, A. and Monina, G. 1992.
Relationships between muscle type and some traits influencing: veal colour. J. Sci. Food
Agric. 58: 523-529.
Guignot, F., Touraille, C., Ouali, A., Renerre, M. and Monin, G. 1994. Relationships
between post-mortem pH changes and some traits of sensory quality in veal. Meat Sci. 37:
315-325.
Hocquette, J.-F., Ortigues-Marty, I., Damon, M., Herpin. P and Geay. Y. 2000.
Métabolisme énergétique des muscles squelettiques chez les animaux producteurs de
viande. INRA Prod. Anim. 13: 185-200.
Huff-Lonergan, E., and Lonergan, S.M. 2005. Mechanisms of water-holding capacity of
meat: The role of postmortem biochemical and structural changes. Meat Sci. 71: 194–204.
INTERBEV. 2006. Le point sur la couleur de la viande bovine.
http://www.agrireseau.qc.ca/bovinsboucherie/documents/couleur_viande_bovine1.pdf. 113
pages (Document consulté le 17 août 2014).
Ishihara, Y., Moreira, R., De Souza, G., Salviano, A. and Madruga, M. 2013. Study of
the Warner-Bratzler shear force, sensory analysis and sarcomere length as indicators of the
tenderness of sun-dried beef. Molecules. 18: 9432-9440.
Jones, S. D. M. and Tong, A. K. W. 1989. Factors influencing the commercial incidence
of dark cutting beef. Can. J. Anim. Sci. 69: 649-654.
Kelzer, J.M., Kononoff, P.J., Tedeschi, L.O., Jenkins, T.C., Karges, K. and Gibson,
M.L. 2010. Evaluation of protein fractionation and ruminal and intestinal digestibility of
corn milling co-products. J. Dairy Sci. 93: 2803–2815.
Kinman, L. A., Hilton,G. G., Richards, C. J., Morgan, J. B., Krehbiel,C. R., Hicks, R.
B., Dillwith, J. W., and VanOverbeke, D. L. 2011. Impact of feeding various amounts of

57
wet and dry distillers grains to yearling steers on palatability, fatty acid profile, and retail
case life of longissimus muscle. J. Anim. Sci. 89: 179–184.
Klopfenstein, T. J., G. E. Erickson and V. R. Bremer. 2008. Board-invited review : Use
of distillers by-products in the beef cattle feeding industry. J. Anim. Sci. 86: 1223-1231.
Lackey, R. 2008. Le boeuf virtuel du MAAARO – Gros gluten de maïs ou drèches de
distillerie – Que donner? Février. Vol. 7 No. 17. 10 pages
Lardy, G. P. 2002. Canola meal in beef cattle diets. Animal and Range Sciences
Department. North Dakota State University. 6 pages.
Lawrie, R. A. 1998. Lawrie's meat science. 6e édition. Woodhead publishing limited,
Cambridge, Royaume-Unis.
Leupp, J. L., Lardy, G. P., Bauer, M. L., Karges, K. K., Gibson, M. L., Caton J. S.
and. Maddock. R. J 2009. Effects of distillers dried grains with solubles on growing and
finishing steer intake, performance, carcass characteristics, and steak color and sensory
attributes. J. Anim. Sci. 87: 4118-4124.
Lomiwes, D., Farouk, M.M., Wu, G. and Young, O.A. 2014. The development of meat
tenderness is likely to be compartmentalised by ultimate pH. Meat Sci. 96: 646–651.
Lucero-Borja, J., Pouzo, L. B., dela Torre, M.S., Langman, L., Carduza, F., Corva, P.
M. Santini, F. J. and Pavan, E. 2014. Slaughter weight, sex and age effects on beef shear
force and tenderness. Livest. Sci. 163: 140-149.
Mandell, I. B., Gullett, E. A., Wilton, J. W., Allen, O. B. and Osborne, V. R. 1997. Effects of diet, breed and slaughter endpoint on growth performance, carcass composition
and beef quality traits in Limousin and Charolais steers. Can. J. Anim. Sci. 77: 23-32.
Marieb, E. N. et Laurendeau, G. 1993. Anatomie et physiologie humaine. Éditions du
renouveau pédagogique, Québec, QC, Canada.
McKenna, D. R., Roebert, D. L., Bates, P. K., Schmidt, T. B., Hale, D. S., Griffin, D.
B., Savell, J. W., Brooks, J. C., Morgan, J. B., Montgomery, T. H., Belk, K. E. and
Smith, G. C. 2002. National Beef Quality Audit-2000: survey of targeted cattle and carcass
characteristics related to quality, quantity, and value of fed steers and heifers. J. Anim. Sci.
80: 1212-1222.
Myer, R.O. 2003. Alternative feeds for beef cattle. Institute of Food and Agricultural
Sciences. University of Florida. AN128. 5 pages.
Nade, T., Uchida, K., Omori, K., Matsubayashi, K. and Kimura, N. 2012. Effects of
feeding dried distillers grains with solubles (DDGS) on meat quality at the late stage of the
fattening period of Holstein steers. Anim. Sci. J. 83: 310–317.

58
Ngapo, T. M. and Gariépy, C. 2006. Factors affecting the meat quality of veal. J. Sci.
Food Agric. 86: 1412-1431.
Nikmaram, P., Yarmand, M. S., Emamjomeh Z. and Darehabi, H. K. 2011. The Effect
of Cooking Methods on Textural and Microstructure Properties of Veal Muscle
(Longissimus dorsi) ISSN: 1992-6197. 6: 201-207.
Noon, C. D., Seoane, J. R. and Scott, S. L. 1998. The use of corn and barley diets for veal
calves : Effects on performance, diet digestibility and carcass quality. Can. J. Anim. Sci.
78: 351-358.
Önenç, A. and Kaya, A. 2004. The effects of electrical stunning and percussive captive
bolt stunning on meat quality of cattle processed by Turkish slaughter procedures. Meat
Sci. 66: 809–815.
Pipek, P., Haberl, A. and Jeleníková, J. 2003. Influence of slaughterhouse handling on
the quality of beef carcasses. Czech J. Anim. Sci. 9: 371-378.
Ripoll, G., Albertí, P., Casasús I., and Blanco, M. 2013. Instrumental meat quality of
veal calves reared under three management systems and color evolution of meat stored in
three packaging systems. Meat Sci. 93: 336–343.
Roeber, D. L., Gill, R. K. and DiCostanzo, A. 2005. Meat quality responses to feeding
distiller's grains to finishing Holstein steers. J. Anim. Sci. 83: 2455-2460.
Royer, A. et Vézina, F. 2012. Intégration verticale et contractualisation en agriculture.
http://personnel.fsaa.ulaval.ca/fileadmin/fichiers/fichiersPersonnel/AnnieRoyer/Rapport_fi
nal_-_Integration.pdf. 58 pages. (Document consulté le 15 mai 2015).
Scanga, J. A., Belk, K. E., Tatum, J. D., Grandin, T., and Smith G. C. 1998. Factors
Contributing to the Incidence of Dark Cutting Beef. J. Anim. Sci. 76: 2040–2047.
Sosin-Bzducha, E.M., Strzetelski J.A. and Skrzyński, G. 2012. The effect of starch
source on physicochemical and sensory quality of veal meat. J. Anim. Feed Sci. 21: 436–
446.
Suarez-Mena, F. X., Hill, T. M., Heinrichs, A. J. H., Bateman II, G., Aldrich, J. M.
and Schlotterbeck, R. L. 2011. Effects of including corn distillers dried grains with
solubles in dairy calf feeds. American Dairy Science Association. J. Dairy Sci. 94: 3037–
3044.
Thomas, M., Hersom, M., Thrift, T., and Yelich, J. 2010. Wet brewer’s grains for beef
cattle. Institute of Food and Agricultural Sciences. University of Florida. AN241. 3 pages.
Vander Pol, K. J., Luebbe, M. K., Crawford, G. I., Erickson, G. E. and Klopfenstein,
T. J. 2009. Performance and digestibility characteristics of finishing diets containing

59
distillers grains, composites of corn processing coproducts, or supplemental corn oil. J.
Anim. Sci. 87: 639-652.
Van de Water, G., Verjans, F. and Geers, R. 2003. The effect of short distance transport
under commercial conditions on the physiology of slaughter calves; pH and colour profiles
of veal
Livest. Prod. Sci. 82: 171–179.
Veau de grain du Québec certifié – Premier de classe. 2014. www.veaudegrain.com.
(Page consultée le 8 mars 2014).
Vieira, C. and Fernández, A. M. 2014. Effet of ageing time on suckling lamb meat
quality resulting from different chilling regimes. Meat Sci. 96: 682-687.
Wright, T. and Lackey, R. 2012. Comparaison des valeurs nutritives d'aliments pour
ruminants. http://omafra.gov.on.ca/french/livestock/dairy/facts/03-006.htm. (Consulté le 13
septembre 2014).

60

61
CHAPITRE 3.
Effect of dietary replacing corn on carcass and meat quality in grain fed
calves
A. O’Bomsawin-Descôteaux1, C. Gariépy2, Y. Chorfi 3 and D. Cinq-Mars
1,4
1Département des Sciences Animales, Université Laval, 2425 rue de l’agriculture Québec,
QC, G1V 0A6, Canada.
2Centre de recherche et développement sur les aliments, Agriculture et Agroalimentaire
Canada, 3600, boulevard Casavant Ouest, Saint-Hyacinthe, QC, J2S 8E3, Canada.
3Département de Biomédecine Vétérinaire, Faculté de médecine vétérinaire - Université de
Montréal, 3200 Sicotte St., P.O. Box 5000, Saint-Hyacinthe, QC, J2S 7C6, Canada.
4Corresponding author:
Dany Cinq-Mars, tel: 418-656-2131 # 11362, [email protected]
Short title: O’Bomsawin-Descôteaux et al.- Grain-fed calves’ meat quality with by-
products
Abbreviations :
BW : Body weight, TEP : 1,1,3,3-tetraethoxypropan; HCL : Chloridric acid; TBA :
Thiobarbituric acid; TCA : Trichloroacetic acid; BHT : Butylated hydroxytoluene, HCW :
Hot carcass weight, DFD : Dark, Firm, Dry.

62

63
3.1. ABSTRACT
O’Bomsawin-Descôteaux, A., Gariépy, C., Chorfi, Y. and Cinq-Mars, D. Effect of dietary
replacing corn on carcass and meat quality in grain fed calves. 2015. Can. J. Anim. Sci.
95: 00-00. The aim of this study was to evaluate the effects of partial or total maize (M)
replacement by rolled barley (B), dried distillers grain with soluble (D) or canola meal (C)
on meat quality in grain-fed calves. Eight left longissimus dorsi were utilized from calves
for each diet treatment randomly assigned : M-supplement (S) (36% CP), BCD, MCD and
MSD and fed ad libitum. All diets were isonitrogenous. Ultimate pH (pHu), colour (L*, a*
and b*), Warner-Bratzler shear force (WBSF), sarcomere length, cooking losses,
malonaldehyde concentration at d0 and d7, marbling, meat composition and sensory
analysis were performed. Significant difference between M and B (P < 0.05) were observed
for pHu, colour (L*, a* and b*), WBSF, sarcomere length, cooking losses, malonaldehyde
concentration at d7, total fat content and off-flavor in sensory analysis. The difference
obtained in WBSF was not noticed by panelists. More research is needed before suggesting
maize replacement with barley, canola meal and DDGS in grain fed calves diets.
Key words : Grain-fed veal, colour, sarcomere length, shear force, by-products, meat
quality.

64

65
3.2. RÉSUMÉ
O’Bomsawin-Descôteaux, A., Gariépy, C., Chorfi, Y. et Cinq-Mars, D. L’utilisation de
sous-produits en remplacement du maïs durant la période de croissance et de finition
sur la qualité de la viande de veau de grain. 2015. Can. J. Anim. Sci. 95: 00-00.
L’objectif de cette étude était d’évaluer les effets d’un remplacement partiel ou total du
maïs (M) par de l’orge roulée (B), des drèches sèches de distillerie avec solubles (D) ou du
tourteau de canola (C) sur la qualité de la viande de veaux de grain. Huit muscles
longissimus dorsi gauche ont été pris pour chacun des traitements étant assignés
aléatoirement : M-suppléments protéiques (S), BCD, MCD et MSD et donné à volonté.
Toutes les rations étaient isoprotéiques. Le pH ultime, la couleur (L*, a* et b*), la force de
cisaillement, la longueur des sarcomères, les pertes à la cuisson, la concentration en
malonaldéhyde aux jours 0 et 7, la composition de la viande et des analyses sensorielles ont
été testés au cours de cette étude. Des différences significatives entre M et B (P < 0.05) ont
été obtenues pour le pH ultime, la couleur (L*, a* et b*), la force de cisaillement, la
longueur des sarcomères, les pertes à la cuisson, la concentration de malonaldéhyde au jour
7, la teneur en gras total et des flaveurs indésirables au niveau des analyses sensorielles. La
différence obtenue lors de la force de cisaillement n’était pas perceptible par les panélistes.
Des recherches supplémentaires sont requises avant de suggérer le remplacement du maïs
par l’orge, le tourteau de canola et les drèches sèches de distillerie dans l’alimentation des
veaux de grain.
Mots-clé : Veau de grain, couleur, longueur sarcomère, force de cisaillement, sous-
produits, qualité de viande.

66

67
3.3. INTRODUCTION
Grain-fed veal production starts with a calf fed milk replacer for the first six weeks of its
life. The calf is then gradually fed high level of concentrate based on corn grain, protein
supplement and mineral with very few forage if any (Ngapo and Gariépy 2006).
In past few years, the use of corn grain for ethanol production increased the availability of
distillers dried grain with soluble (DDGS) (Klopfestein et al. 2008) that are used in beef
production partly to replace maize in diet (Roeber et al. 2005; Leupp et al. 2009; Buttrey et
al. 2012). DDGS effects on animal performances depend on the amount of inclusion, the
type of grain used for fermentation and the ethanol extracting method (Buttrey et al. 2012).
Lightness (L*) of Holstein steers carcasses tended to be higher with the inclusion of 20%
DDGS than 10% DDGS but lower for redness (a*) (Roeber et al. 2005). However, Leupp et
al. (2009) showed that feeding 30% DDGS produce a faster decrease of a* than feeding
50% dry rolled corn. The authors suggested that this might be attributable to an increase of
unsaturated fatty acid oxidation and a decrease of enzymatic system which controls
metmyoglobin level in meat. The inclusion of DDGS in maize diets also reduced the shear
force with crossbred and purebred beef steers (Leupp et al. 2009). Noon et al. (1998)
reported that whole barley in replacement of corn significantly decreased carcass quality.
However, these authors did not evaluate the effects of whole barley on meat quality.
Nutrient composition of canola meal depends on the environmental conditions during plant
growth and harvest, bulk density and the processing method for oil extraction (Lardy and
Anderson, 2002). No significant difference was observed in growth performances of 45-
day-old Holstein calves fed canola meal as ingredient in starter rations in replacement of
soybean meal (Lardy and Anderson, 2002).
The objective of this study was to evaluate the effects of partial or total replacement of
maize by rolled barley, DDGS or canola meal on meat quality in grain-fed calves.

68

69
3.4. MATERIALS and METHODS
Animals
Animal and dietary treatments were described by Najid et al. (unpublished). Briefly, 320
clinically healthy 3mo Holstein calves with similar body weight (117±13 kg) were used in
this study. Animals were allotted 10 per pen in 32 pens equipped with automatic waterers.
Pens were randomly assigned to four experimental treatments using 8 pens per treatment.
Relative humidity and temperature were continuously recorded every 5 min. Lighting was
8L: 16D. Animal care procedures followed guidelines of the Canadian Council on Animal
Care (CCAC, 1993) and the protocol was approved by the animal care committee of the
Faculté de médecine vétérinaire of the Université de Montréal.
Diets
During the first 2 weeks, calves received an adaptation diet containing all experimental
ingredients. Four diets consisted of control maize (M) and commercial calf supplement (S)
(Deli Pro, COMAX, St-Hyacinthe, Qc, Ca) diet (MS); M, S and DDGS (MSD) diet; M,
canola meal (C) and DDGS (MCD) diet; and rolled barley (B), CM and DDGS (BCD) diet
fed ad libitum. Calves received 200 g of commercial minerals-vitamins with monensin
sodium at 660 mg kg-1 per day (Prémélange minéralisé veaux lourds, COMAX, St-
Romuald, Qc, Ca). Diets were distributed at 0600, 1030, 1500 and 2030 and were all
isonitrogenous.
Slaughtering
Calves were shipped in mixed group and slaughtered within 60 to 90 minutes of reception.
Calves were slaughtered with percussion stunning without breaking skull at 295 kg of BW
at the Abattoir Saint-Germain (Saint-Germain-de-Grantham, Qc, Ca) which represents 20

70
minutes of transport (22 km). Calves’ carcasses were cooled under the same condition than
beef and were electrically stimulated 2 sec interval during 20 sec.
Samples
Seventy two h after slaughter carcasses were transported to Montpak cutting room (Laval,
Québec) in refrigerated truck. Room temperature remained constant at +4ºC until
processing. Left longissimus dorsi muscle was removed from the carcass as short loin (<
40cm), and then cut in two halves. On one half, colour, ultimate pH (pHu), marbling
measures was performed on fresh meat. Then both halves were vacuum packed and frozen,
-40ºC until analyzed. One half was used for sensory analysis and the other for
physicochemical analysis. A total of 8 short loins per treatment were randomly chosen.
Colour, ultimate pH (pHu) and marbling
Data were recorded 96h post mortem on the samples before packaging. A Minolta CR-300
colorimeter device was used for the colour according to the CIELAB (CIE, 1976) color
system as L* for lightness (100 = white, 0 = black), a* for redness (35 = red, -35 = green)
and b* for yellowness (35 = yellow, -35 = blue) (light source D65; Minolta Camera Co.
Ltd., Osaka, Japan). This unit was calibrated with a CR-A43 white plate. Color has been
taken 5 mn after cutting the short loin in halves.
The pHu was measured with a Beckman Coulter 200 pH-meter (Beckman Instruments,
Inc., CA, USA) with a calibrated glass tip probe (LoT406-M6-DXK-S7/25) Mettler Toledo
(Mettler Toledo Ingold Inc., Bedford, MA, USA) on a polybutyleneterephthalate shaft. The
probe was inserted 25 mm in meat sample.
Marbling has been classified with a sample of the National Pork Producers Council (NPPC,
2000) Official Color and Marbling Standards (P.O. Box 10306. Des Moines. IA. 50306).

71
Cooking losses
Loin samples were thawed at +4ºC the day before analysis. A 10 cm length x 10 cm wide x
6 cm thickness sample of longissimus dorsi was taken and precisely weighted (m0) from
each loin. Then, each sample was placed individually in a polyethylene bag vacuum sealed
with a thermocouple inserted in the middle of the meat. Bags were deposited randomly in a
controlled temperature circulation water tank at 72ºC and cooked until the meat reached
67.5ºC. Bags were then immediately immersed in an ice water bath trough until the internal
meat temperature reach 10ºC. Then samples were refrigerated at +4ºC overnight. Then,
meat samples were sponged and weighted precisely for a second time (mf). Cooking losses
was calculated as the weight difference before and after cooking :
Cooking losses (%) = (m0 – mf)/ m0 x 100
Warner-Bratzler shear force (WBSF)
This analysis was performed on cooked meat samples. Ten 1 cm2 by 10 cm cross sectional
cores were prepared per sample with their lengths paralleled to the fiber axis. The shear
force analysis was conducted by Warner-Bratzler device attached to a TAXT2i Texture
Analyser (Texture Technologies Corp., Scarsdale, NY, USA). Cores were sheared across
the fiber axis with a 20 kg cell at a speed of 1.5 mm/sec and means were calculated with 10
readings.
Sarcomere length
Sarcomere were extracted from 2.5g of raw ground meat from a loin cut and mixed with 25
ml of a sucrose solution 0.25 M. The mix was stirred 25 s at maximum 16 000 rpm with a
Polytron PT 10-35GT (Kinematica, Luzernerstrasse, Lucerne, Suisse) equipped with
grinding Polytron rod PT-DA 12/2EC B154 (12 mm diameter). A drop of solution was
placed on a slide and covered with a slip. Images were taken with a Nikon Eclipse E400
microscope (Nikon Canada instruments Inc., Missisauga, On, Ca) at 40X for 12 myofibrils

72
containing at least 10 sarcomeres. Measures between sarcomeres were taken with Image-
Pro Plus software (Media Cybernetics Inc., Rockville, MD, USA).
Thiobarbituric acid (TBA)
Solution preparations
Malonaldehyde 10-4M standard stock solution was made by mixing 10µl of 1,1,3,3-
tetraethoxypropan (TEP) with 10 ml of HCL 0.1N, heated 5 mn at 95ºC and cooled at room
temperature. Then, 2.4 ml of this stock solution was poured in 100 ml flask and completed
with distilled water. A standard curve was prepared with six different Malonaldehyde
concentrations (0.25; 0.50; 1; 2; 4 and 6 x 10-9M /ml). 3 ml of standard stock solution was
mixed with 3 ml of TBA 20 x 10-3M and heated 30 mn at 100ºC then cooled at room
temperature. 3 ml of the mix were then transferred in a polycarbonate bowl and absorbance
was read at 531 nm as reference.
Sample preparations
Ground meat (5g) was mixed with 20 ml of trichloroacetic acid (TCA) 5% and 50 µl of
butylated hydroxytoluene (BHT) 2.5%. With a Polytron PT 10-35GT equipped with
grinding Polytron rod PT-DA 12/2EC B154 (12 mm diameter), the mix was stirred 1 mn at
maximum 13 500 rpm. Then, 30 ml of TCA 5% was added. The solution was filtered with
a Whatman paper #1. A quantity of 3 ml of the filtrate was mixed with 3 ml of TBA 20 mM
and heated 30 mn at 100ºC, then cooled at room temperature. Then, 3 ml of the mix was
transferred in a glass bowl and sample absorbance was read at 531 nm.
Extraction yield
For extraction yield, 1 ml of standard stock solution 10-4M was mixed with 5 g of ground
meat for a sample, 19 ml of TCA 5% and 50 µl of BHT 2.5%. Then, 3 ml of the mix was
transferred in a glass bowl and absorbance was read at 531 nm. Extraction yield was
calculated with a template from AAFC.

73
Malonaldehyde concentration
For malonaldehyde concentration, the absorbance of 1.5 ml of standard stock solution 10-
4M mixed with 1.5 ml of distilled water was read at 245 nm as reference. All procedures
were made for day 0 and 7 of retail display simulation at +4ºC.
Meat composition (water, total fat and protein content)
Water content
A sample of ground meat was precisely weighted (m0), then lyophilized and weighted a
second time (mf).
Water content (%) = 1-(m0 – mf)/ m0 x 100
Total fat content
Total fat extraction was performed with chloroform/methanol (2:1) solvent, according to
the Soxhlet method using VELP SER 148 (VELP Scientific, Inc., Bohemia, NY, USA). A
quantity of 2g of lyophilized ground meat (m0) was mixed with sand in a cartridge to
increase contact surface with the solvent. Then, a cotton ball was put at the top of the
cartridge and fixed to the device’s adapters. For each aluminum bucket, a tiny amount of
boiling stones was added and weighted precisely (m1). 40 ml of chloroform / methanol
(2:1) solvent was poured in each aluminum bucket. After the extraction, buckets were
placed in a dryer for 30 mn at 100ºC and then cooled in a desiccator at room temperature.
At the end, the aluminum bucket was weighted a second time (m2). Fat weight (mf) was
calculated as follows:
(1) Total fat content (% DM) = 100-(m0 – mf)/ m0 x 100
(2) Total fat content (% AF) = Total fat content (% DM) x Water content (%) /100
Total protein content
Total protein content of meat was performed using a LECO FP428 model (LECO
Corporation, St. Joseph, MI, USA) which doses nitrogen content of the samples. %
nitrogen has been reported in % proteins according these equations:

74
(1) Total protein content (% DM) = % N x 6,25
(2) Total protein content (% AF) = Total protein content (% DM) x Water content (%)
/100
Sensory analysis
A sensory analysis was performed with 10 trained panelists. Steaks were thawed at 4ºC for
24 h prior to cooking, then cooked between plates until the internal temperature reaches
69ºC. Sensory sessions occurred twice a week and lasted 2 wks. The scores were
distributed with a scale of 1 for the lowest score to 10 the highest score given.
Statistical analysis
Meat quality
All statistical analysis except for sensory analysis, were carried out using SAS proc GLM
model (SAS version 9.3, Cary, NC, USA). Significance was declared if P < 0.05 and a
tendency if P < 0.10.
Sensory analysis
Sensory analysis results were carried out using Biosystemes Fizz, version 2.47B
(Biosystemem, Couternon, France) with a Duncan’s test at 5%.

75
3.5. RESULTS and DISCUSSION
3.5.1. Ultimate pH, meat colour and marbling
Treatments had a significant effect on pHu and colour values but not for marbling (Table
3.1). The MS diet had the lowest pHu value in comparison to other treatments. BCD, MCD
and MSD treatments were above pHu 5.7, but MCD and MSD were not significantly
different from MS. Normally, after slaughter muscle pH gradually drops around 5.5-5.7
during the transformation into meat (INTERBEV, 2006; Sosin-Bzducha et al. 2012). Final
meat pH or pHu value depends on the glycogen amount in the muscles pre-slaughter
(Hocquette et al. 2000; INTERBEV, 2006). Many factors may impact the preslaughter
muscle glycogen reserve, such as pre-slaughter handling including sorting, transportation,,
various stress and physical activities (Hocquette et al. 2000; Pipek et al. 2003; Önenç and
Kaya, 2004; INTERBEV, 2006). Najid et al. (unpublisehd) reported a tendency for lower
blood glucose in BCD as compared to other treatments. Blood parameters have shown a
tendency in a decrease of glucose level for all diets but was lower in BCD diet at 96d (see
Najid et al. 2014). Diets were fed ad libitum and were isonitrogenous, but they were not
isoenergetics. Degradation of ruminal starch from barley is higher (90%) than corn (60%),
however starch content of MS was higher than the other treatments (Najid et al.
unpublished). Therefore, it is possible that when calves are fed corn diets, there is an
increase of starch in small intestine which is digested and absorbed as glucose (Noon et al.
1998). Blood glucose is more energy efficient than volatile fatty acids produced in the
rumen (Noon et al. 1998). These authors also showed a significant decrease of dietary
energy values during the growing phase while whole barley gradually replaces corn in grain
fed calves diets. Guertin et al. (1995) reported less retained energy with whole and rolled
barley than with whole maize in dairy calves slaughtered at 222 kg body weight. This may
be because barley has greater fiber content and lower energy concentration than corn (Kim
et al. 2014).
When compared to soybean meal, canola meal has less ruminal bypass protein, total tract
digestibility and digestible energy (Lardy and Anderson, 2002). The hull proportion and

76
fiber content is higher in canola meal than soybean meal and can contribute to the lower
total tract digestibility and digestible energy content in canola meal (Lardy and Anderson,
2002). Therefore, a possible combination of lower blood glucose from barley compared to
corn and higher fiber proportion in canola compared to soybean meal based supplement
may explain higher pHu in BCD compared to MS meat. Džinić et al. (2007) observed a
higher pHu when crushed soybean is replaced by 20% of extruded rape seed in broiler
breast meat, but it was not significantly different.
The MS treatment had the highest colour score in comparison with others treatments.
Higher L* scores means that meat is lighter, higher a* score means that the meat is more
red and higher b* score means that the meat is more yellow (Campbell et al. 2013). On the
contrary, lower colour values scores for BCD (P < 0.05) produced darker meat than MS
diet. Meat from other treatments gave intermediate values. At the cutting room, meat from
BCD was also gummy which tended to be a DFD meat class (Scanga et al. 1998; Abril et
al. 2001; Pipek et al. 2003; Van de Water et al. 2003). In this study, there was a significant
interaction (P < 0.0001) pHu x colour, meaning that pHu significantly influenced meat
color. Muscle acidification changes the muscular structure which reflects light depending
on the pH value and produces a different brightness. When pHu is high, proteins chains
repel themselves since they are electrically charged, which means away from the isoelectric
point, and contained water molecules which give them an open structure. In this situation,
light penetrates deep into the muscle and the reflected light portion is small thereby
obtaining a darker meat (Guignot et al. 1994; Abril et al. 2001). The creation of open
structures generates a water accumulation producing a rather sticky meat (Guignot et al.
1994; INTERBEV, 2006).
Contrary to the present study, Boles et al. (2004) found a lighter meat production while
using four varieties of barley in crossbred steers compared with corn. The increase in
lightness with barley could be associated with antemortem muscle glycogen content and the
relationship with pHu or the rate of pH decline (Boles et al. 2004). High-energy diets can
increase glycogen reserves and decrease pHu compared with roughage-based and low-
energy diets (Boles et al. 2004). Boles et al. (2004) compared different varieties of barley

77
and our study used only one. There may be a difference in energy content depending of
barley variety.
MS diet cooking losses were significantly greater than BCD diet due to the pHu value. In
low pHu, proteins chains attract themselves because the electrical charge descrease. The
muscular structure is closed. Light penetrates poorly into the muscle and a large proportion
of it is reflected. This situation creates a lighter meat. However, this kind of muscular
structure decrease the meat water holding capacity (INTERBEV, 2006). Water losses have
been related to very low pHu (Guignot et al. 1994). When lateral shrinkage of myofibrils
appears, water is transferred into muscles extracellular spaces (Huff-Lonergan et al. 2005;
Ripoll et al. 2013). The size of those spaces is influenced by the pHu (Guignot et al. 1994).
In the present study, using 21% of DDGS gave intermediate values (P < 0.05) for L*, a*
and b* between higher color values for MS and lower values for BCD. According to Leupp
et al. (2009), a* value from steers meat fed 30% of DDGS to substitute dry-rolled corn
decreased faster than those with 0% DDGS inclusion in the diet. This was attributed to the
increase of unsatturated fatty acids oxydation and enzymatic system reduction which
control the metmyoglobin amount in meat (Leupp et al. 2009). In the present study, the
possible effect of metmyoglobin may have play a role, but the combined effect of DDGS
and barley was probably additive. This may explain a production of a darker meat with
BCD diet.
3.5.2. Cooking losses, Warner-Bratzler shear force (WBSF) and Sarcomere length
Treatments had a significant effect on cooking losses, WBSF and sarcomere length (Table
3.2). BCD treatment has generated lower cooking losses, WBSF and sarcomere length
compared with MS (P < 0.05). Cooking losses were significantly higher for MS and MCD
diets compared to BCD diet. MSD diet gave intermediate value. There is almost 2 kg
difference in WBSF between BCD and MS diets. MCD and MSD diets gave intermediate

78
values for WBSF and sarcomere length. All treatments had a sarcomere length values
around 1.80 µm except for BCD with 1.52 µm (Guignot et al. 1994; INTERBEV, 2006).
Enzymatic activity of calpains is favored when pHu is higher than 6.3 and cathepsin
activity when pHu is lower than 5.8 (Guignot et al. 1994; Pipek et al. 2003; Lomiwes et al.
2014; Vieira et Fernández, 2014). When ion pumps stop after slaughter and energy content
in muscle is exhausted, there is an increase of sarcoplasmic calcium concentration. This
calcium is needed to activate μ- and m- calpain isoforms for the myofibrillar proteins
hydrolysis. Calpain not only has the ability to degrade myofibrillar proteins but also the
cytoskeletal proteins (Huff-Lonergan et al. 2005; Lomiwes et al. 2014). It was reported that
the activity of cathepsin double at low pHu compared to high pHu (Lomiwes et al. 2014).
Cathepsin B is released within the sarcoplasm from lysosomes when pHu is low. This
protease does not only hydrolyse myofibrillar proteins, but also alter proteins associated
with A-band and Z-disks of myofibrillar proteins (Lomiwes et al. 2014). In the present
study, pHu of BCD treatment was 6.25 and MS was below 5.8. Therefore, it is possible that
the difference in WBSF may be attributable to these enzyme activities caused by different
pHu between treatments. When both proteolytic enzymes, calpain and cathepsin are more
active, a better tenderization occurs (Guignot et al. 1994; Lomiwes et al. 2014). This may
be attributed to the myofibrillar proteins degradation with high and low molecular weight
during meat aging by the activity of μ-calpain and cathepsin B (Lomiwes et al. 2014). Meat
tenderization seems faster when the pHu is high compared to low pHu. At low pHu, the
process seems to be slower because of rigor mortis. The calpain activity is already
triggered at d0 post mortem due to high pHu (Lomiwes et al. 2014). The cathepsin B
becomes active as the pHu approaches 5.4 (Brewer, 2012). In this study, pHu values tended
to influence WBSF as P = 0.0559. WBSF values may be due to the activity of proteolytic
enzymes, which depend on pHu.
Sarcomere length can be influenced by the calpain which degrades myofibrillar and
cytoskeletal proteins (Lomiwes et al. 2014). pHu could have an impact on sarcomere length
because it has a role in the activity of this protease. This could explain why BCD produced

79
tender meat compared to MS. No significant effect has been observed between WBSF and
sarcomere length (P > 0.10).
Cooking losses are due to the loss of water, molten fat, nitrogenous components and
minerals (Alves de Almeida et al. 2008). According to Campbell et al. (2013), meat
cooking losses of grain-fed calves is around 21.8%. Cooking losses of MS and MCD
treatments were significantly different from BCD. MSD gave an intermediate value. The
diets with by-products (BCD, MCD and MSD) may have an effect on meat cooking losses
and water-holding capacity. Both of these parameters are related to pHu (De Palo et al.
2013). When water-holding capacity is low, there is an increase of cooking losses (De Palo
et al. 2013). In the present study, pHu has significantly influenced cooking losses with P <
0.0001. This is due to the muscular structure variation depending on pHu.
3.5.3. TBA (malonaldehyde concentration)
Treatments had a significant effect on retail display simulation on day 7 (Table 3.3). BCD
treatment developed higher malonaldehyde (P < 0.05) than MS. MCD and MSD diets gave
intermediate values. There was also a significant relationship between malonaldehyde and
pHu (P = 0.0043). Malonaldehyde production is related to lipid peroxidation (Raharjo and
Sofos, 1993) that is one of major causes for quality deterioration in meat products during
refrigerated or frozen storage (Raharjo and Sofos, 1993). It produces a variety of aldehydes
from phospholipids and polyunsaturated fatty acids (Brewer, 2012). After slaughtering, the
antioxidative capacity is no longer active in muscle cells that become overloaded with
prooxidants, peroxidized lipids and oxygen radicals (Raharjo and Sofos, 1993).
Malonaldehyde production and accumulation in meat varies according to the degree of
unsaturation of PUFA, malonaldehyde precursors from non-lipid origin, types of
peroxidation catalysts and malonaldehyde reactivity with other biological materials
(Raharjo and Sofos, 1993). Thiobarbituric acid reacts with malonaldehyde to givean adduct
whose spectrum which is identical with that obtained from malonaldehyde standard
(Raharjo and Sofos, 1993). The reaction rate is function of the concentration of TBA

80
solution, temperature and medium pH (Raharjo and Sofos, 1993). A meat pHu above 5,
may increase malonaldehyde production because the reaction rate is lower. pHu from BCD
diet was the highest this may explained higher score for malonaldehyde production at 7d.
In the present study, calves have received more than 150 I.U./day in all and 21% DDGS
have been used in diets BCD, MCD and MSD treatments (Najid et al. unpublished). In
order to reduce malonaldehyde production during meat storage, DDGS seems to be a good
value in animal diet. Indeed, Nade et al. (2012) compared a substitution of concentrated
feed of corn grain, wheat bran and oil meal by 15% DDGS in 18 mo Holstein steers during
the finishing period. Inclusion of 15% DDGS has significantly reduced malonaldehyde
production after 7d of storage at 5ºC. It has been reported that DDGS has a high amount of
vitamin E (Nade et al. 2012) which acts as a potent antioxidant allowing better meat
conservation when supplemented at 100 I.U./calf/day (Ngapo and Gariépy, 2006).
3.5.4. Meat composition
Treatments had only a significant effect on total fat content. No significant difference was
found for water or total proteins contents (Table 3.4). The fat content of meat from MS
treatment was higher than MCD because corn amount was higher in MS treatment. Sosin-
Bzducha et al. (2012) showed that diets rich in corn grain tended to increase meat
intramuscular fat compared to a barley-based diet in Polish Holstein-Friesian bull calves.
This may be due to the slower ruminal degradability of corn starch and much more starch
passing into small intestine (Noon et al. 1998; Sosin-Bzducha et al. 2012). There is an
intestinal digestion of starch to glucose and increases fat in meat due to glucose absorption
(Noon et al. 1998). The amount of intramuscular fat is influenced by many parameters,
such as energy content of the diet, digestibility, absorption, liver metabolism and lipids
transport in muscles (Sosin-Bzducha et al. 2012). In this study, total fat content may have
been influenced by starch source and level inclusion of grain.

81
3.5.5. Sensory analysis
Values for sensory analysis are reported in table 3.5. They were not significantly different
except for Off-flavor which was significantly higher for BCD than MS and MSD
treatments. MCD gave intermediate values. Panelists have observed a livery flavor with
BCD meat. Meat flavor can be influenced by lipids oxidation and measured with
malonaldehyde level (Brewer, 2012; Nade et al. 2012). Rancid fat taste is more intense
when the rate of malonaldehyde is higher (Nade et al. 2012). Humada et al. (2014)
explained that meat rancidity may be detected by consumers when malonaldehyde
exceeded the cut off value of 2 mg kg-1. In the present study, malonaldehyde level in BCD
meat was 0.21 mg kg-1 at d7 of storage. Panelists have been able to detect an off-flavor after
d1 of storage. Malonaldehyde at d7 for MCD meat had a tendency to be higher than MSD
diet for Off-flavor but not significant. The difference in off-flavor between MCD and MSD
may be due to the use of canola meal. Iron content is higher in canola meal (162 mg kg-1)
than soybean meal (83-116 mg kg-1) (Conseil canadien du canola, 2014; Ravindrana et al.
2014). Livery flavor increases as meat iron content increases (Yancey et al. 2006; Brewer,
2012). According to Yancey et al. (2006), livery flavor may be also influenced by
unsaturated fatty acid content in meat. These authors have shown that unsaturated fatty acid
C18:1 is negatively related to livery flavor. Mikulski et al. (2012) demonstrated that C18:1
and polyunsaturated fatty acids significantly increased in turkey meat with high inclusion
of rapeseed meal. Yossinov (2014) obtained the same conclusion in lamb carcass when
rapeseed meal totally replaced sunflower meal. Unsaturated fatty acids have a stronger
relationship with off-flavor than saturated fatty acids (Yancey et al. 2006) and rapeseed
meal significantly lowered levels of saturated fatty acids (Mikulski et al. 2012). Lipid
oxidation is also more susceptible with the increase of unsaturated fatty acids (Mikulski et
al. 2012).
According to Leupp et al. (2009), inclusion of 30% DDGS in finishing state for steers
produced juicier and more flavorful meat compared to 0% inclusion. However no
difference was observed in this study. Juiciness can be influenced by the fat and water
content of meat (Gregory, 1998). In the present study, considering that fat content was

82
significantly different and water content was not, meat juiciness was not significantly
different to panelists. Even although fat content was significantly different, it seems that it
was not detected by panelists to change meat juiciness.
In the present study, the significant difference in WBSF results could not be detected
during sensory analysis. According to Destefanis et al. (2008) a variation of 1 kg in WBSF
can be noticed to trained panelists. The WBSF difference of near 2kg between MS and
BCD in the present study should have been noticed by trained panel. It is likely that this
difference in WBSF would not be observed either by consumers.

83
3.6. CONCLUSION
Partial or total corn replacement by barley and by-products increased meat pHu which
influenced many such as colour, cooking losses, WBSF, sarcomere length, TBA d7, total
fat content and off-flavor in sensory analysis. Introduction of barley and by-products in
claves diets did not change marbling, TBA d0, water and protein content and sensory
analysis except off-flavor. The difference obtained in WBSF was not noticed to panelists.
Substituting corn and protein supplement in grain fed calves for barley, canola or DDGS
increased pHu and lower general meat quality. Therefore, until further research is
performed to address this issue, caution is advised in regard of meat quality.

84

85
3.7. ACKNOWLEDGMENTS
This study was supported by the Volet C «Appui à la réalisation de projets novateurs et
structurants» from the « Programme d’appui financier aux regroupements et aux
associations de producteurs désignés » of the Ministère de l'Agriculture, des Pêcheries et de
l'Alimentation du Québec (MAPAQ). The authors are grateful to the Fédération des
producteurs de bovins du Québec (FPBQ) for supporting this project, to Délimax for all the
help during the experiment, to Julie Chapron for her leadership in the animal trial, to the
Centre de développement du porc du Québec inc. (CDPQ) for the technical help at the
cutting room and to Agriculture and Agri-Food Canada (AAFC) for the help during the
meat quality analysis and the volunteers for the sensory analysis.

86

87
3.8. REFERENCES
Abril, M., Campo, M.M., Önenç, A., Sañudo, C., Albertí, P., and Negueruela, A.I.
2001. Beef colour evolution as a function of ultimate pH. Meat Sci. 58: 69-78.
Alves de Almeida, G. A. Jr., Costa, C., Ribeiro de Carvalho, S. M., Persichetti, P. Jr.
and Panichi, A. 2008. Composição físico-química de carcaças de bezerros holandeses
alimentados após o desaleitamento com silagem de grãos úmidos ou grãos secos de milho
ou sorgo. R. Bras. Zootec. 37:164-170.
Boles, J. A., Bowman, J. G., Surber, L. M. M. and Boss, D. L. 2004. Effects of barley
variety fed to steers on carcass characteristics and color of meat. J. Anim. Sci. 82: 2087-
2091.
Brewer, S. 2012. Handbook of meat, poultry and seafood quality. Second edition. Chapter
15 – Quality measurements in beef.
http://onlinelibrary.wiley.com.acces.bibl.ulaval.ca/book/10.1002/9781118352434 (Site
consulted on 2014, Dec. 9)
Buttrey, E. K., McCollum III, F. T., Jenkins, K. H., Patterson, J. M., Clark, B. E.,
Luebbe, M. K., Lawrence, T. E. and MacDonald, J. C. 2012. Use of dried distillers
grains throughout a beef production system : Effects on stocker and finishing performance,
carcass characteristics, and fatty acid composition of beef. J. Anim. Sci. 90: 2381-2393.
Campbell, C. P., Haley, J., Swanson, K. C. and Mandell, I. B. 2013. Packing plant
differences in meat quality for grain-fed veal. Can. J. Anim. Sci. 93: 205-215.
Canadian Council on Animal Care (CCAC). 1993. Guide to the Care and Use of
Experimental Animals. 2nd ed. EdsOlfert-Cross-McWilliam, ON. Ca.
Conseil Canadien du Canola. 2014. http://fr.canolacouncil.org. (Page consultée le 25 avril
2014).
CIE. 1976. Recommendations on uniform color spaces-color difference equations.
Psychometric Color Terms. Supplement No. 2 to CIE Publication No. 15 (E-1.3.1.) 1978,
1971/(TC-1-3), Commission Internationale de l’Éclairage, Paris, France.
De Palo, P., Maggiolino, A., Centoducati, P., and Tateo, A. 2013. Effects of two
different packaging materials on veal calf meat quality and shelf life. J. Anim. Sci. 91:
2920-2930.
Destefanis, G., Brugiapaglia, A., Barge, M.T. and Dal Molin, E. 2008. Relationship
between beef consumer tenderness perception and Warner–Bratzler shear force. Meat Sci.
78: 153–156.

88
Džinić, N., Petrović, Lj., Tomović, V., Tasić, T. and Filipović, S. 2007. Effect of partial
substitution of standard meal in chicken feed by rape seed on carcass and meat quality.
Biotech. Anim. Husbandry. 23: 323-329.
Fornasier, A. 2002. Portrait de l’industrie du veau québécois. Colloque sur le veau lourd.
Centre de référence en agriculture et agroalimentaire du Québec. 18 pages.
FPBQ. 2014. Veau de grain. Fédération des producteurs de bovins du Québec.
www.bovins.qc.ca. (Page consulted on 2014 Mar. 8 and Oct. 1).
Gregory, N. G. 1998. Animal welfare and meat science. CABI Publishing. Wallingford,
Royaume-Uni.
Guertin, G., Lachance, B., Pelletier, G., St-Laurent, G.J., Roy, G.L. and Petitclerc, D.
1995. Effects of photoperiod and feeding whole maize, whole barley, or rolled barley on
growth performance and diet digestibility in veal calves. Livest. Prod. Sci. 44: 27-36.
Guignot, F., Quilichini, Y., Renerre, M., Lacourta, A. and Monina, G. 1992.
Relationships between muscle type and some traits influencing: veal colour. J. Sci. Food
Agric. 58: 523-529.
Guignot, F., Touraille, C., Ouali, A., Renerre, M. and Monin, G. 1994. Relationships
between post-mortem pH changes and some traits of sensory quality in veal. Meat Sci. 37:
315-325.
Hocquette, J.-F., Ortigues-Marty, I., Damon, M., Herpin. P and Geay. Y. 2000.
Métabolisme énergétique des muscles squelettiques chez les animaux producteurs de
viande. INRA Prod. Anim. 13: 185-200.
Huff-Lonergan, E., and Lonergan, S.M. 2005. Mechanisms of water-holding capacity of
meat: The role of postmortem biochemical and structural changes. Meat Sci. 71: 194–204.
Humada, M.J., Sañudo, C. and Serrano, E. 2014. Chemical composition, vitamin E
content, lipid oxidation, colour and cooking losses in meat from Tudanca bulls finished on
semi-extensive or intensive systems and slaughtered at 12 or 14 months. Meat Sci. 96: 908-
915.
INTERBEV. 2006. Le point sur la couleur de la viande bovine.
http://www.agrireseau.qc.ca/bovinsboucherie/documents/couleur_viande_bovine1.pdf. 113
pages (File consulted, 2014 Aug. 17).
Kim, B. G., Wulf, D. M., Maddock, R. J., Peters, D. N., Pedersen, C., Liu, Y. and
Stein, H. H. 2014. Effects of dietary barley on growth performance, carcass traits and pork
quality of finishing pigs. Rev. Colomb. Cienc. Pecu. 27: 102-113.
Klopfenstein, T. J., G. E. Erickson and V. R. Bremer. 2008. Board-invited review : Use
of distillers by-products in the beef cattle feeding industry. J. Anim. Sci. 86: 1223-1231.

89
Lackey, R. 2008. Le boeuf virtuel du MAAARO – Gros gluten de maïs ou drèches de
distillerie – Que donner? Février. Vol. 7 No. 17
Lardy, G. P. and Anderson, V. 2002. Canola and sunflower meal in beef cattle diets. Vet.
Clin. Food Anim. 18: 327-338.
Leupp, J. L., Lardy, G. P., Bauer, M. L., Karges, K. K., Gibson, M. L., Caton J. S.
and. Maddock. R. J 2009. Effects of distillers dried grains with solubles on growing and
finishing steer intake, performance, carcass characteristics, and steak color and sensory
attributes. J. Anim. Sci. 87: 4118-4124.
Lomiwes, D., Farouk, M.M., Wu, G. and Young, O.A. 2014. The development of meat
tenderness is likely to be compartmentalised by ultimate pH. Meat Sci. 96: 646–651.
Mikulski, D., Jankowski, J., Zdunczyk, Z., Juskiewicz, J. and Slominski, B. A. 2012. The effect of different dietary levels of rapeseed meal on growth performance, carcass
traits, and meat quality in turkeys. Poult. Sci. 91: 215-223.
Nade, T., Uchida, K., Omori, K., Matsubayashi, K. and Kimura, N. 2012. Effects of
feeding dried distillers grains with solubles (DDGS) on meat quality at the late stage of the
fattening period of Holstein steers. Anim. Sci. J. 83: 310–317.
Najid, Y., Couture, Y., Chénier, S., Fournier, A., Cinq-Mars, D. and Chorfi, Y. 2014.
Effects of dietary replacing corn on growth performance, ruminal pH, and blood
biochemistry parameters in grain-fed calves. Unpublished.
NPPC. 2000. Pork Composition and Quality Assessment Procedures. Pork Producers
Council. Des Moines, IA.
Ngapo, T. M. and Gariépy, C. 2006. Factors affecting the meat quality of veal. J. Sci.
Food Agric. 86: 1412-1431.
Noon, C. D., Seoane, J. R. and Scott, S. L. 1998. The use of corn and barley diets for veal
calves : Effects on performance, diet digestibility and carcass quality. Can. J. Anim. Sci.
78: 351-358.
Önenç, A. and Kaya, A. 2004. The effects of electrical stunning and percussive captive
bolt stunning on meat quality of cattle processed by Turkish slaughter procedures. Meat
Sci. 66: 809–815.
Pipek, P., Haberl, A. and Jeleníková, J. 2003. Influence of slaughterhouse handling on
the quality of beef carcasses. Czech J. Anim. Sci. 9: 371-378.
Raharjo, S. and Sofos, J. N. 1993. Methodology for measuring malonaldehyde as a
product of lipid peroxidation in muscle tissues: A review. Meat Sci. 35: 145-169.

90
Ravindrana, V., Abdollahi, M. and Bootwalla, S. 2014. Nutrient analysis, apparent
metabolisable energy and ileal amino acid digestibility of full fat soybean for broilers.
Anim. Feed. Sci. Tech. 197: 233-240.
Ripoll, G., Albertí, P., Casasús I., and Blanco, M. 2013. Instrumental meat quality of
veal calves reared under three management systems and color evolution of meat stored in
three packaging systems. Meat Sci. 93: 336–343.
Roeber, D. L., Gill, R. K. and DiCostanzo, A. 2005. Meat quality responses to feeding
distiller's grains to finishing Holstein steers. J. Anim. Sci. 83: 2455-2460.
Scanga, J. A., Belk, K. E., Tatum, J. D., Grandin, T., and Smith G. C. 1998. Factors
Contributing to the Incidence of Dark Cutting Beef. J. Anim. Sci. 76: 2040–2047.
Sosin-Bzducha, E.M., Strzetelski J.A. and Skrzyński, G. 2012. The effect of starch
source on physicochemical and sensory quality of veal meat. J. Anim. Feed Sci. 21: 436–
446.
Van de Water, G., Verjans, F. and Geers, R. 2003. The effect of short distance transport
under commercial conditions on the physiology of slaughter calves; pH and colour profiles
of veal. Livest. Prod. Sci. 82: 171–179.
Vieira, C. and Fernández, A. M. 2014. Effect of ageing time on suckling lamb meat
quality resulting from different chilling regimes. Meat Sci. 96: 682-687.
Yancey, E.J., Grobbel, J.P., Dikeman, M.E., Smith, J.S., Hachmeister, K.A.,
Chambers IV, E.C., Gadgil, P., Milliken, G.A. and Dressler, E.A. 2006. Effects of total
iron, myoglobin, hemoglobin, and lipid oxidation of uncooked muscles on livery flavor
development and volatiles of cooked beef steaks. Meat Sci. 73: 680–686.
Yossifov, M.R. 2014. Effect of dietary rapeseed meal on fatty acid profile of lamb carcass.
Iran. J. Appl. Anim. Sci. 4: 741-746.

91
Table 3.1. Ultimate pH, meat colour and marbling of longissimus dorsi from Holstein grain-fed calves
Treatmentsz
Parametersy MS BCD MCD MSD
Steaks, n 8 8 9 8
pHu 5.62 ± 0.18B 6.25 ± 0.66 A 5.98 ± 0.52 AB 5.97 ± 0.29 AB
Colourx
L* 39.27 ± 3.27 A 34.40 ± 4.44 B 37.91 ± 5.11 AB 36.44 ± 3.18 AB
a* 14.48 ± 1.75 A 11.14 ± 3.32 B 12.71 ± 3.04 AB 12.35 ± 2.83 AB
b* 8.06 ± 1.51 A 5.70 ± 2.31 B 6.91 ± 2.50 AB 6.20 ± 2.04 AB
Marbling scorew 2.56 ± 0.86 2.38 ± 0.52 2.33 ± 0.79 2.38 ± 0.74
z Treatments : MS = Whole maize + proteins supplements; BCD = Rolled barley + canola meal + DDGS; MCD = Whole maize + canola meal + DDGS; MSD
= Whole maize + proteins supplements + DDGS. y Means with different superscripts within the same row are significantly differents (P < 0.05). x L* : 100 = white, 0 = black; a* : 35 = red, -35 = green and b* : 35 = yellow, -35 = blue. w Marbling score is according to the National Pork Producers Council (NPPC, 2000). Marbling scale is 1 to 5. (1 = extremely small; 5 = extremely abundant).

92
Table 3.2. Cooking losses, Warner-Bratzler shear force (WBSF) and Sarcomere length of longissimus dorsi from Holstein grain-
fed calves
Treatmentsz
Parametersy MS BCD MCD MSD
Steaks, n 8 8 9 8
Cooking losses (%)
23.06 ± 3.00A 15.24 ± 5.40B 20.14 ± 4.20 A
18.96 ± 4.30
AB
WBSFy. kg 4.96 ± 1.60A 3.11 ± 1.72B 3.69 ± 1.72AB 3.84 ± 1.69AB
Sarcomere lenght (µm) 1.82 ± 0.31 A 1.52 ± 0.31 B 1.78 ± 0.28 AB 1.79 ± 0.28 AB
z Treatments : MS = Whole maize + proteins supplements; BCD = Rolled barley + canola meal + DDGS; MCD = Whole maize + canola meal + DDGS; MSD
= Whole maize + proteins supplements + DDGS. y Means with different superscripts within the same row are significantly differents (P < 0.05). x WBSF : Warner-Bratzler shear force; SL : Sarcomere length.
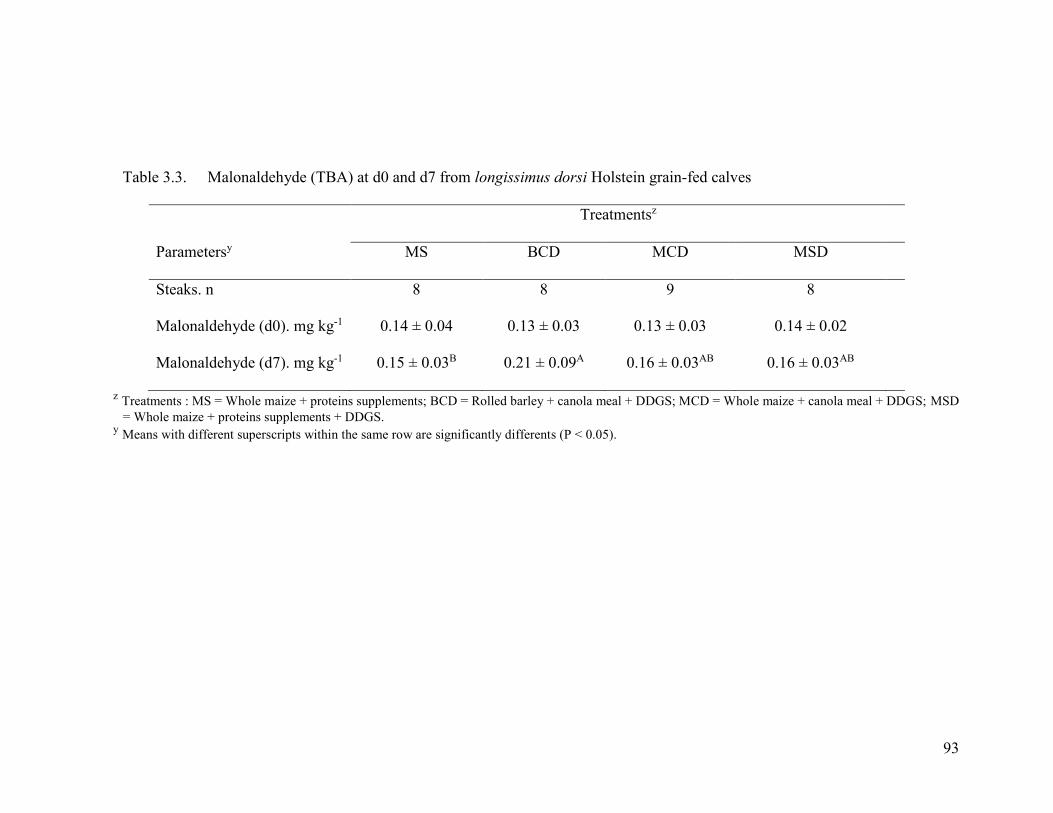
93
Table 3.3. Malonaldehyde (TBA) at d0 and d7 from longissimus dorsi Holstein grain-fed calves
Treatmentsz
Parametersy MS BCD MCD MSD
Steaks. n 8 8 9 8
Malonaldehyde (d0). mg kg-1 0.14 ± 0.04 0.13 ± 0.03 0.13 ± 0.03 0.14 ± 0.02
Malonaldehyde (d7). mg kg-1 0.15 ± 0.03B 0.21 ± 0.09A 0.16 ± 0.03AB 0.16 ± 0.03AB
z Treatments : MS = Whole maize + proteins supplements; BCD = Rolled barley + canola meal + DDGS; MCD = Whole maize + canola meal + DDGS; MSD
= Whole maize + proteins supplements + DDGS. y Means with different superscripts within the same row are significantly differents (P < 0.05).

94
Table 3.4. Meat composition of longissimus dorsi from Holstein grain-fed calves
Treatmentsz
Parametersy MS BCD MCD MSD
Steaks. n 8 8 9 8
Water content. (%) 25.45 ± 0.86 24.98 ± 0.84 24.93 ± 0.83 24.66 ± 0.91
Total fat content. (%) 4.27 ± 0.82A 4.13 ± 0.55AB 3.52 ± 0.63B 3.70 ± 0.80AB
Total protein content. (%) 21.41 ± 0.84 21.37 ± 0.61 21.56 ± 0.66 21.31 ± 0.59
z Treatments : MS = Whole maize + proteins supplements; BCD = Rolled barley + canola meal + DDGS; MCD = Whole maize + canola meal +
DDGS; MSD = Whole maize + proteins supplements + DDGS. y Means with different superscripts within the same row are significantly differents (P < 0.05).

95
Table 3.5. Sensory analysis of longissimus dorsi from Holstein grain-fed calves
Treatmentsz
Parametersy MS BCD MCD MSD
Steaks. n 10 10 10 10
Tendernessx 5.14 ± 1.67 5.13 ± 1.93 4.89 ± 1.86 4.90 ± 2.13
Juiciness 4.26 ± 1.46 4.32 ± 1.56 4.18 ± 1.50 4.06 ± 1.08
Beef flavor 4.89 ± 1.53 4.02 ± 1.70 4.15 ± 1.99 4.49 ± 1.53
Off-flavorw 0.60 ± 0.93B 1.23 ± 1.54A 0.85 ± 1.03AB 0.70 ± 1.18B
z Treatments : MS = Whole maize + proteins supplements; BCD = Rolled barley + canola meal + DDGS; MCD = Whole maize + canola meal +
DDGS; MSD = Whole maize + proteins supplements + DDGS. y
Means with different superscripts within the same row are significantly differents (P < 0.05). x Scores were distributed from 1 to 10 where Tenderness : 1 = extremely tough; 10 = extremely tender; Juiciness : 1 = extremely dry; 10 = extremely
juicy; Beef flavor : 1 = extremely bland; 10 = extremely flavorful; Strange flavor : 1 = nothing; 10 = extremely present. w
Off-flavor regroups all commentaries from panelists as metal, acid, blood and liver taste.





























![Quintette à Cordes en Sol majeur [CNW 59]...Carl Nielsen Udgaven CN 00025 204 arco arco arco 77 pizz. pizz. pizz. 71 II 2 cre scen do cre scen do cre scen do cre scen do 69 6 1 cre](https://img.pdfslide.fr/doc/110x75/60a763b57af8716b955ef719/quintette-cordes-en-sol-majeur-cnw-59-carl-nielsen-udgaven-cn-00025-204.jpg)