Embed Size (px)
Citation preview

-Entité (I) l’activité enzymatique ( la vitesse d’une réaction enzymatique) :
-En se liant à l’enzyme, un inhibiteur peut :
• empêcher la fixation de S au site actif,
• provoquer une déformation du site actif (notamment à l’état de transition) et rendre
l’enzyme moins active (voire inactive) vs S.
-L’affinité de I pour E est traduite par la constante d’inhibition Ki :
• Ki = cte de dissociation de EI en E + I (unité : M),
• Ki = [I] pour laquelle la moitié des sites enzymatiques est occupée,
• + Ki est petit, + l’affinité de I pour E est grande,
• Inhibiteur « idéal » : forte affinité (Ki petit) pour E et forte sélectivité vs S (Km/Ki
grand).
[EI]
[E][I]
i
i-i
k
kKE + I EI
ki
k-i
Exemple :
phosphoglucose
isomérase (PGI)
µM 200F6P
m K
nM 2005PAH
i K1000im KK
Inhibition de l'activité enzymatique
; unités en Molaire
BCM 1502 Cinétique d’inhibition enzymatique Page 1/29

Conception des inhibiteurs :
-Beaucoup d'inhibiteurs (médicaments) ont été trouvés par hasard !
-Ethnopharmacologie
-Criblage à haut débit (HTS ou High Throughput Screening) de produits naturels ou
synthétiques (chimiothèques…)
-Criblage virtuel (screening « in silico »)
-Conception d’un I vs E ciblée par docking
Nature des inhibiteurs :
-Analogues de S (ou P)
-Analogues de l’ET ou IHE
-Complexants de métaux (EDTA…)
-Structures diverses (généralement trouvés par hasard)
« Mimes » : qui ressemblent à… sans être eux-mêmes transformés
CK2
POMs
Inhibition non-compétitive (IC50 = 3 nM) d’une protéine kinase (CK2) par un composé de type polyoxométallate (POM).
Chem. Biol., 2008, 15, 683-692
© CEA
BCM 1502 Cinétique d’inhibition enzymatique Page 2/29

1.1. Différents types cinétiques d’inhibiteurs d'enzymes
Inhibiteurs réversibles :
-Liaison non-covalente (ou covalente) peu stable de l’inhibiteur à l’enzyme (complexes
EI, ESI).
-L'inhibition est réversible (peut être levée), i.e. l’enzyme n’est pas irréversiblement
inhibée :
• Inhibiteurs compétitifs
• Inhibiteurs incompétitifs (et inhibition par excès de substrat)
• Inhibiteurs non-compétitifs purs
• Inhibiteurs non-compétitifs mixtes
Inhibiteurs irréversibles :
-Liaison covalente (ou non-covalente) stable de l’inhibiteur à l’enzyme (complexe E-I)
-Inhibition ne peut pas être levée (inactivation l’enzyme est irréversiblement inhibée) :
• Marqueurs d'affinité
• Inhibiteurs suicides (ou mécanistiques)
• Cas particulier : inhibiteurs dits à interaction lente et/ou à forte affinité :
inhibition non-covalente mais quasi-irréversible (slow-binding, tight-binding
inhibitors)
BCM 1502 Cinétique d’inhibition enzymatique Page 3/29

1.2. Intérêts et usages des inhibiteurs d’enzymes
Recherche fondamental :
-Etudes cinétiques des mécanismes enzymatiques :
• Mode d'interaction d'un inhibiteur ?
• Stoechiométrie et mécanisme de l'inhibition ?
• Effet de l'inhibiteur sur les paramètres cinétiques Km et Vmax (Kmapp, Vmax
app)
• Efficacité d'un inhibiteur (Ki vs Km)
• Comparaison de l'efficacité de ≠ inhibiteurs sur une cible (comparaison des Ki )
• Comparaison de l'efficacité d’un inhibiteur sur ≠ cibles (comparaison des Ki )
4PEA 0.028 1.7
4PEH 0.029 0.057
Ki (mM) Ki (mM)
Inhibiteur SoRpiA MtRpiB
SoRpiA : Rpi d’épinard, type A MtRpiB : Rpi de Mycobacterium tuberculosis, type B
Exemple : Etudes d’inhibition de ribose-5-
phosphate isomérases (Rpi) de type A et B
R5P (Km) 7.5 2.5
BCM 1502 Cinétique d’inhibition enzymatique Page 4/29

-Etudes cristallographiques des enzymes (complexes EI cristallisés) :
• Nature et rôle des résidus du site actif
• Mode de fixation du substrat (→ I analogues de S/P)
• Mode de fixation des intermédiaires réactionnels (→ I analogues de IHE/ET)
• Fonctionnement de l’enzyme (mécanisme)
-Etudes de métabolismes (rôle d’une enzyme dans un processus métabolique)
-Génération d’abzymes (anticorps catalytiques) :
• Enzymes artificielles générées par un organisme en réaction à la présence
d’un antigène (inhibiteur analogue de l’ET appelé « haptène »)
PDB 1G98
Biochemistry 2001, 40, 1560-1566
Complexe EI RmPGI-5PAA
5PAA
BCM 1502 Cinétique d’inhibition enzymatique Page 5/29

Applications thérapeutiques :
-Enzymes : cibles de choix
-Utilisation biomédicale des inhibiteurs : bien avant de connaître les enzymes !
-Difficulté : spécificité de l’enzyme inhibée : souvent, les cellules « cibles » et « hôtes »
possèdent les "mêmes" enzymes (→ parasites, cellules cancéreuses…)
-Correction d’un métabolisme défectueux :
• Hypocholestérolémiant (vs excès de cholestérol)
• Antidiabétique (hypoglycémiants)
-Correction d’un "désordre physiologique" :
• Antidépresseurs
• Antiépileptiques
• Antihypertenseurs
• Antihistaminiques (vs effets de l’histamine produite lors de réactions allergiques)
• Antipyrétiques (vs fièvre)
• Analgésiques (vs douleur)
• Antiinflammatoires
BCM 1502 Cinétique d’inhibition enzymatique Page 6/29

Environnement :
-Lutte contre les plantes parasitaires, les insectes et les champignons (xylophages…)
• Herbicides
• Insecticides
• Fongicides
Armes chimiques (!) :
-Neurotoxiques (gaz Sarin, Tabun) : → I irrév. d’une enzyme du SNC
-Vésicants (gaz Moutarde/Hypérite) : → irritants (peau, yeux, muqueuses…)
-Destruction d’un organisme pathogène :
• Antiparasitaires (paludisme, trypanosomiases…)
• Antibactériens
• Antifongiques (vs champignons)
• Anticancéreux
• Antiviraux (antirétroviraux) (vs maturation des virus, interaction virus-cible…)
Pesticides (produits phytosanitaires)
O P
O
F
O P
O
N
Sarin Tabun
BCM 1502 Cinétique d’inhibition enzymatique Page 7/29

1.3. Exemples d’agents thérapeutiques inhibiteurs d’enzymes
http://www.wellesley.edu/Chemistry/Chem101/hiv/HIV-2.html
Infection par le VIH
AZT
ddC
Ritonavir
Trithérapie
-4 cibles dont 2 principales ; d’autres multi-thérapies existent
-résistances vs RT du VIH
(vs résistances)
BCM 1502 Cinétique d’inhibition enzymatique Page 8/29

Ritonavir® Ic Protéase (VIH)
Zidovudine® 3’-azido-2’, 3’-dideoxy-thymidine (AZT)
AZT-TP : Ic Transcriptase Inverse (VIH)
Aciclovir® Aciclovir-TP :
Ic ADN Polymérase (Herpes, VZV)
Antiviraux (antirétroviraux)
O
NH
RR'
S
P O P P
P O P P
AZT AZT
AZT-TP
kinases
X
2’, 3’-dideoxy-cytidine (ddC) ddC-TP :
Ic Transcriptase Inverse (VIH)
P O P P
BCM 1502 Cinétique d’inhibition enzymatique Page 9/29

Antibactériens
Triméthoprim® Ic Dihydrofolate Reductase
Pénicilline Iirr Transpeptidase (paroi cellulaire bactérienne)
Sulfaméthoxazole® Ic Dihydroptéroate Synthase
Autres activités
Kétoconazole® Antifongique Ic C14 a-Demethylase à CytP450 (biosynthèse de l’ergostérol de la paroi cellulaire des champignons)
Méthotrexate® Anticancéreux Ic Dihydrofolate Reductase
Bactrim®
BCM 1502 Cinétique d’inhibition enzymatique Page 10/29

Aspirine Analgégique, antipyrétique, antiinflammatoire Iirr Prostaglandine Synthase (Cyclooxygénase)
Isocarboxazide Antidépresseur Iirr (suicide) Monoamine Oxidase
Allopurinol vs goutte/calculs rénaux Ic Xanthine oxydase (biosynthèse acide urique)
Autres activités (suite)
Captopril Antihypertenseur Ic Enzyme de Conversion de l’Angiotensine (ACE)
Mévinoline Hypocholestérolémiant Ic Hydroxymethylglutaryl-CoA reductase
BCM 1502 Cinétique d’inhibition enzymatique Page 11/29

2. Inhibition réversible : généralités et cinétique
2.1. Inhibition compétitive
Fixation réversible exclusive
Différents modèles de l’inhibition compétitive :
A. Modèle classique : S et I ont le même site de fixation
1.
S et I sont en compétition vs site actif
de part leur analogie de structure.
Site actif de la fructose-1,6-
phosphate aldolase d’H. pylori
BCM 1502 Cinétique d’inhibition enzymatique Page 12/29
FBP
Inh
J Biol Chem. 2011 286, 40219-31

B. Modèles alternatifs : les sites de fixation de S et I sont distincts
2.
L’encombrement stérique de I
empêche la fixation de S
3.
S et I ont un groupe en commun
qui se fixe sur un 3ème site
4.
Les sites de fixation de S et I
se recouvrent
5.
Régulation allostérique :
La fixation de I induit un
changement de conformation de
l’enzyme qui déforme ou
masque le site de fixation de S
(et inversement)
"Enzyme Kinetics " I. Segel (1975), Ed. J. Wiley & Sons, Inc.
Exemples :
-aspartate transcarbamylase
-glutamine synthétase
BCM 1502 Cinétique d’inhibition enzymatique Page 13/29

k1 S
EI
E +
I
k-1
kcat
Ki
E + P
+
ES
• S et I sont en compétition pour leur fixation sur E (inhibition exclusive)
• L’affinité apparente de E : Kmapp
• I peut être déplacé par un excès de S : Vmax n’est pas modifié
[ES]
[E][S]m K
[EI]
[E][I]i K v0 = kcat[ES] [E]T = [E] + [ES] + [EI]
Cinétique de l’inhibition compétitive (rappels) :
BCM 1502 Cinétique d’inhibition enzymatique Page 14/29

Vmax
Vmax/2
Km Kmapp
Vo
[S]
[I] > 0
[S][I]
1
[S]V
m
i
max0
KK
v
avec m
i
m
app [I]1 K
KK
Inhibition compétitive : v0 = f([S])
[S]
[S]V
appmax0
mKvsoit
BCM 1502 Cinétique d’inhibition enzymatique Page 15/29

-1/Km
Km/Vmax
1/[S]
1/Vmax +
+
+
+ +
1/Vo
-1/Km1app
sans inhib.
[I]1
[I]2
[I]3
Km1app/Vmax
Inhibition compétitive : 1/v0 = f(1/[S]) (Lineweaver-Burk)
[S]
1
V
[I]1
V
11
max
m
i
max0
KK
v
[I]3 > [I]2 > [I]1
pente = max
m
i V
[I]1
K
K
oao = 1/Vmax
pente et oao = cte lorsque [I]
oao = ordonnée à l'origine
BCM 1502 Cinétique d’inhibition enzymatique Page 16/29

Inhibition compétitive (Lineweaver-Burk)
représentation secondaire : pente = f([I])
-Ki [I]
•
•
•
•
pour pente = 0, [I] = -Ki pentei =
max
m
i
i
V
[I]1
K
K
BCM 1502 Cinétique d’inhibition enzymatique Page 17/29
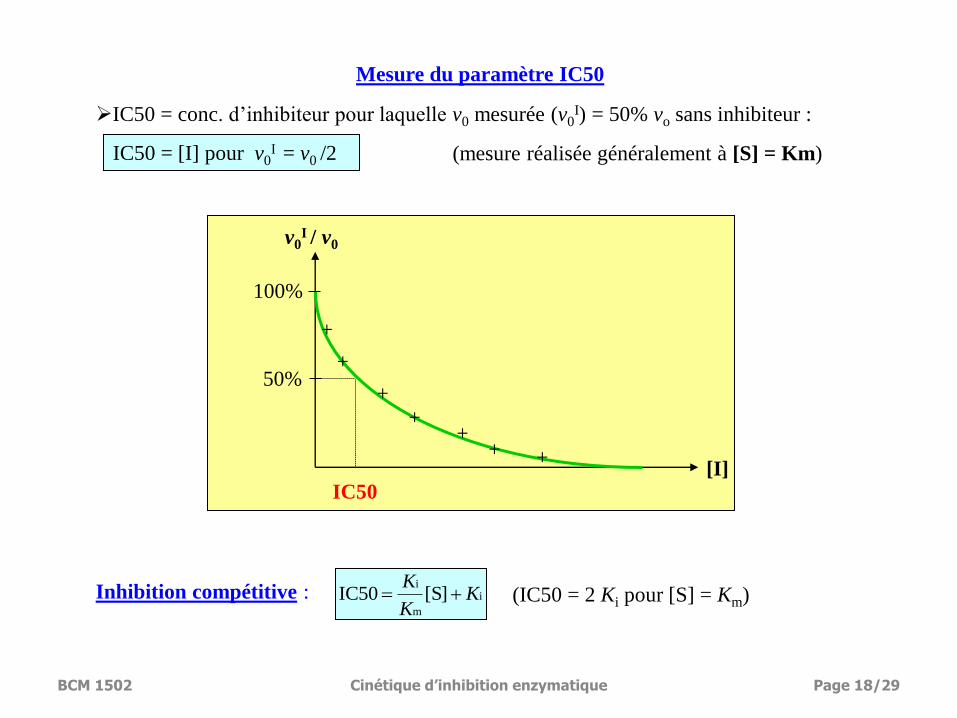
50%
100%
v0I / v0
[I] IC50
Mesure du paramètre IC50
IC50 = [I] pour v0I = v0 /2
IC50 = conc. d’inhibiteur pour laquelle v0 mesurée (v0I) = 50% vo sans inhibiteur :
(mesure réalisée généralement à [S] = Km)
+
+
+
+
+ +
+
Inhibition compétitive : (IC50 = 2 Ki pour [S] = Km) i
m
i[S]IC50 K
K
K
BCM 1502 Cinétique d’inhibition enzymatique Page 18/29

2.2. Inhibition incompétitive
Un inhibiteur incompétitif (uncompétitif) ne se fixe que sur le complexe ES (le site de fixation de I est induit par celle de S) : L’affinité de E pour S (induit par la formation de ESI) : Kmapp E apparaît moins active (à cause de ESI, inactif) : Vmaxapp
kcat
x
E + P
S
E
ES
ESI
KS
Ki
I
I
S
S
[ES]
[E][S]m K
Si [S] >> Km :
[E]T = [ES] + [ESI]
Vmaxapp = kcat([E]T - [ESI])
(< Vmax)
[ESI]
[ES][I]i K v0 = kcat[ES]
[E]T = [E] + [ES] + [ESI]
BCM 1502 Cinétique d’inhibition enzymatique Page 19/29
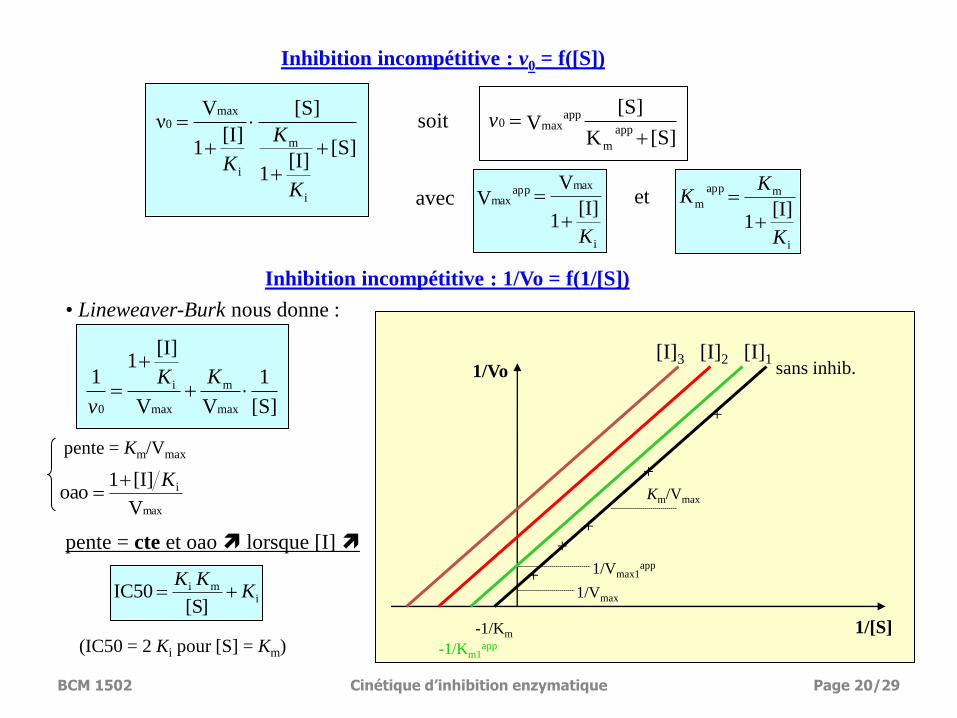
[S][I]
1
[S]
[I]1
Vν
i
m
i
max0
K
K
K
i
max
maxapp
[I]1
VV
K
[S]K
[S]V
m
appmaxapp
0
v
Inhibition incompétitive : v0 = f([S])
avec et
i
mm
app
[I]1
K
KK
• Lineweaver-Burk nous donne :
[S]
1
VV
[I]1
1
max
m
max
i
0
KK
v
pente = cte et oao lorsque [I]
max
i
V
[I]1oao
K
-1/Km
Km/Vmax
1/[S]
1/Vmax
+
+
+
+ +
1/Vo
-1/Km1app
sans inhib. [I]1 [I]2 [I]3
1/Vmax1app
(IC50 = 2 Ki pour [S] = Km)
im i
[S]IC50 K
KK
Inhibition incompétitive : 1/Vo = f(1/[S])
soit
pente = Km/Vmax
BCM 1502 Cinétique d’inhibition enzymatique Page 20/29

2.3. Inhibition non-compétitive pure
• Un inhibiteur non-compétitif peut se fixer sur E et sur ES (mais n’est pas en compétition avec S pour sa fixation à l’enzyme). Il ne peut être déplacé par augmentation de [S]
• E apparaît moins active (à cause de ESI, inactif) : Vmaxapp
• L’affinité de E (et EI) pour S n’est pas modifiée : Km n’est pas modifié (I nc pure)
[ESI]
[EI][S]
[ES]
[E][S]m K
[ESI]
[ES][I]
[EI]
[E][I]i K
v0 = kcat[ES]
[E]T = [E] + [ES] + [EI] + [ESI]
k1
k-1
kcat
Ki
E + P
Ki
k1
k-1
x BCM 1502 Cinétique d’inhibition enzymatique Page 21/29
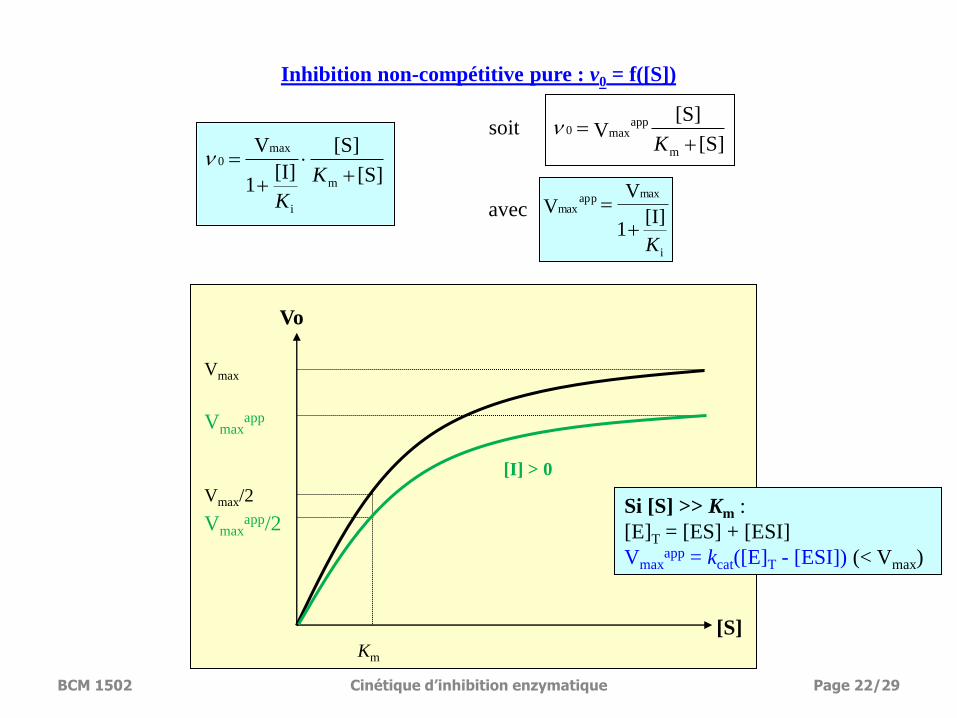
Vmax
Vmax/2
Km
Vo
[S]
[I] > 0
[S]
[S]
[I]1
V
m
i
max0
K
K
avec
i
max
maxapp
[I]1
VV
K
[S]
[S]V
m
maxapp
0
Ksoit
Inhibition non-compétitive pure : v0 = f([S])
Vmaxapp
Vmaxapp/2
Si [S] >> Km :
[E]T = [ES] + [ESI]
Vmaxapp = kcat([E]T - [ESI]) (< Vmax)
BCM 1502 Cinétique d’inhibition enzymatique Page 22/29

-1/Km
Km/Vmax
1/[S]
1/Vmax
+
+
+
+ +
1/Vo
sans inhib.
[I]1
[I]2
[I]3
Km/Vmax1app
Inhibition non-compétitive pure : 1/v0 = f(1/[S]) (Lineweaver-Burk)
[S]
1
V
[I]1
V
[I]1
1
max
m
i
max
i
0
KKK
pente et oao lorsque [I]
1/Vmax1app
[I]3 > [I]2 > [I]1
pente = max
m
i V
[I]1
K
K
max
i
V
[I]1oao
K
IC50 = Ki
BCM 1502 Cinétique d’inhibition enzymatique Page 23/29
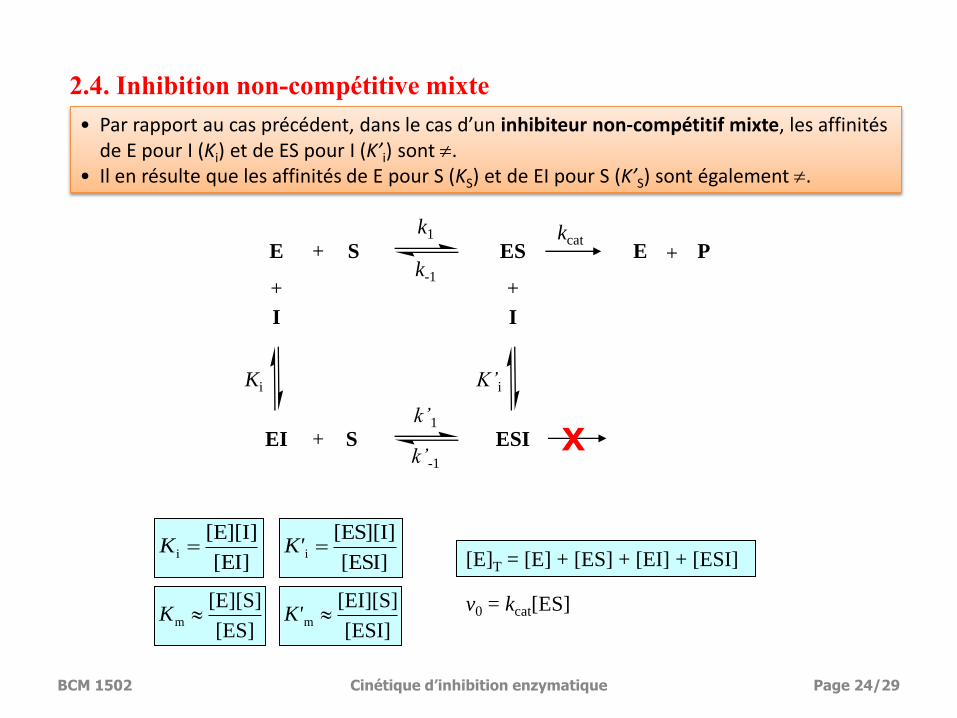
2.4. Inhibition non-compétitive mixte
• Par rapport au cas précédent, dans le cas d’un inhibiteur non-compétitif mixte, les affinités de E pour I (Ki) et de ES pour I (K’i) sont .
• Il en résulte que les affinités de E pour S (KS) et de EI pour S (K’S) sont également .
[ES]
[E][S]m K
[EI]
[E][I]i K
v0 = kcat[ES]
[E]T = [E] + [ES] + [EI] + [ESI] [ESI]
[ES][I]i K'
[ESI]
[EI][S]m K'
ES
ESI
+
+
k1
k-1
kcat
K’i
x
E + P
I
+
Ki
I
EI S
k’1
k’-1
+
E S
BCM 1502 Cinétique d’inhibition enzymatique Page 24/29

Deux cas peuvent être distingués :
• Ki > K’i : Kmapp
• Ki < K’i : Kmapp
Dans les deux cas :
• Vmaxapp (à cause de ESI)
[S]α'
α
[S]
α'
V
m
max0
K
avec
[S]
[S]V
m
appmaxapp
0
Kvsoit
α'
VV
max
maxapp et m
app
α'
αKKm
i
[I]1α
K
i
[I]1α'
K'
Inhibition non-compétitive mixte : Vo = f([S])
• Lineweaver-Burk nous donne :
[S]
1
V
α
V
α'1
max
m
max0
K
v
Inhibition non-compétitive mixte : 1/Vo = f(1/[S])
pente = max
m
i V
[I]1
K
K
pente et oao lorsque [I]
max
i
V
[I]1oao
K'
BCM 1502 Cinétique d’inhibition enzymatique Page 25/29

Inhibition non-compétitive mixte : 1/v0 = f(1/[S])
-1/Km
Km/Vmax
1/[S] 1/Vmax
+
+
+
+ +
1/Vo
sans inhib.
[I]1
[I]2
[I]3
a1Km/Vmax
[I]3 > [I]2 > [I]1
a’1/Vmax
m1
1
α'
αK
Ki < K’i
BCM 1502 Cinétique d’inhibition enzymatique Page 26/29

Inhibition non-compétitive mixte : 1/v0 = f(1/[S])
-1/Km
Km/Vmax
1/[S] 1/Vmax
+
+
+
+
+
1/Vo
sans inhib. [I]1
[I]2
[I]3
a1Km/Vmax
[I]3 > [I]2 > [I]1
a’1/Vmax
m1
1
α'
α
K
Ki > K’i
BCM 1502 Cinétique d’inhibition enzymatique Page 27/29

Deux modèles alternatifs d’inhibition non-compétitive (mixte)
kcat
E + P
Ki
KS
K’S
kcat
E + P
Ki K’i
KS
EI peut fixer S (pour donner ESI inactif),
mais ES ne peut pas fixer I
ES peut fixer I (pour donner ESI inactif),
mais EI ne peut pas fixer S
BCM 1502 Cinétique d’inhibition enzymatique Page 28/29

Tableau comparatif des inhibitions réversibles
Finalement, les inhibitions compétitive, incompétitive et non-compétitive pure sont des cas particuliers de l’inhibition non-compétitive mixte :
Inhibition Effet sur Vmax Vmaxapp Effet sur Km Km
app
K’i = compétitive - Vmax aKm
Ki = incompétitive Vmax /a’ Km/a’
Ki = K’i non-compétitive pure Vmax /a - Km
Ki > K’i non-compétitive mixte Vmax /a’ aKm/a’
Ki < K’i non-compétitive mixte Vmax /a’ aKm/a’
i
[I]1α
K
i
[I]1α'
K'
Remerciement à Dr. Laurent Salmon, Institut de Chimie Moléculaire et des Matériaux d’Orsay (ICMMO), Université Paris-Sud 11, 91405 Orsay Cedex, France
BCM 1502 Cinétique d’inhibition enzymatique Page 29/29




























![Synthèse des analogues de l’[azaPhe4]-GHRP-6 comme](https://img.pdfslide.fr/doc/110x75/618c27ade26e05549a04661f/synthse-des-analogues-de-lazaphe4-ghrp-6-comme-.jpg)
