Embed Size (px)
Citation preview

European J. Biochem. 2 (1967) 341-348
La semi-aldkhyde aspartique dkshydrogknase chez Saccharomyces cerevisiae: proprikths et rkgulation
Y. SURDIN
Laboratoire d’Enzymologie du C.N.R.S., Gif-sur-Yvette
(Repu le 31 mai 1967)
The synthesis of threonine and methionine in X. cerevisiae proceeds as follows : aspartic acid + aspartyl-phosphate + aspartic-semi-aldehyde
lr homoserine J \
threonine methionine
isoleucine .1
The aspartic-semi-aldehyde dehydrogenase (ASA dehydrogenase) catalyzes the second reaction. It has been purified from an extract of baker’s yeast and has been obtained with a specific activity 250 times higher than in crude extracts. I ts main properties are : (a) The optimum pH is 8.5 in both directions; (b) Its activity is enhanced 4 or 5 times in both directions by addi- tion of bicarbonate to the reaction mixture. Evidence is presented that the bicarbonate is necessary for the reaction to proceed; (c) I ts molecular weight estimated by filtration on Sephadex G-200 is about 140,000; (d) No feed-back inhibition by threonine or methionine has been found. An inhibition by cysteine and homocysteine (Kt = 1 x M) was found in the reverse reaction. The synthesis of ASA dehydrogenase is regulated by threonine intracellular concentration ; (e) By heating at 60” a t different pH’s it is shown that ASA dehydrogenase exists in two forms. Two mechanisms are proposed: a depolymerisation, or a release of proton.
As in the case of the ~-glyceraldehyde-3-phosphate dehydrogenase, the reaction catalyzed by ASA dehydrogenase can be separated into two parts : the oxidation and the phosphorylation. This mechanism is under investigation.
Chez Xaccharomyces cerevisiae la biosynthese de la threonine et de la methionine s’effectue selon le schema suivant :
Aspartate + Aspartyl-phosphate + Semi-aldehyde aspartique
It Homosbrine J \
Threonine Mhthionine
Isoleucine 4
Abrbviations non usudles. Semi-aldbhyde aspartique, ASA; dibthanolamine, DEA; 2-mercaptobthanol, 2-ME; B-aspartyl-phosphate, BAP; semi-aldbhyde aspartique dbs- hydrogbnase, ASA dbshydrogbnase.
Enzymes. Aspartokinase, ou ATP : ~-aspartate-4-phos- photransfbrase (EC 2.7.2.4); semi-aldbhyde aspartique dbs- hydrogenase, ou L-aspartate-8-semi-aldhhyde : NADP oxydo- rbductase (phosphorylating) (EC 1.2.1.11); homosdrine dbs- hydrogdnase, ou L-homosbrine : NAD oxydorbductase (EC 1.1.1.3) ; glycbralddhyde 3-phosphate dbshydrogdnase, ou ~-glycbraldbhyde-3-phosphate : NAD oxydorbductase (phos- phorylating) (EC 1.2.1.12).
Contrairement aux bactkries, X. cerevisiae ne synthetise pas la lysine B partir du semi-aldbhyde aspartique. Les trois enzymes communs B la bio- synthkse de la thrbonine et de la mbthionine sont : l’aspartokinase, la semi-aldbhyde aspartique des- hydrogenase et l’homos6rine dbshydrogbnase. Les mu- tants auxotrophes pour l’homosbrine ont bt6 analyses biochimiquement et genetiquement [l]. Les gknes contr8lant la synthkse de ces trois enzymes, respecti- vement thr,, thr, et thr,, shgrbgent indbpendemment.
L’Btude de la regulation de cette voie de bio- synthkse a dejh montre que, d’une part, la synthkse et l’activiti? de l’aspartokinase sont contrBl6es par la thrtionine [2] et que, d’autre part, la synthbse de l’homoserine dkshydrogenase est contr816e par la mbthionine et son activite par la thrbonine et la methionine [3].
Le present travail a pour but d’8tudier les pro- pribtbs de la semi-aldhhyde aspartique dbshydro- genase qui occupe la position intermediaire entre ces deux enzymes. On dbfinira les conditions pouvant modifier son activitb et sa synthbse.

342 ASA d6shydrogAnase chez 8. cerevisiae European J. Biochem.
MATGRIEL ET TECHNIQUES Matbiel
Ce travail a &tit effectuh avec la levure de bou- langerie (Springer). On a verifie que 1’ASA des- hydrogenase extraite de la levure de boulangerie a les m6mes propriittes que celle extraite de la souche 4094/B.
Les souches haploides de S. cerevisiae employees au cours de ce travail sont les suivantes: (a) 4094-B provenant de la collection de F. Sherman. Genotype : a, ad,, ur,; (b) J D 144 provenant de la collection de R. K. Mortimer. Genotype: a, ad,, thr,; (c) JE.2 provenant de la collection de R. K. Mortimer. Gi:no- type: a, a$, thr,; (d) T 1-6 provenant de la collec- tion de H. Cherest. Genotype: a, thr,, ad2, ur,.
Prdparation de l’extrait acellulaire La levure pressbe est suspendue dans un poids
itgal de tampon !his-HC1 IO-lM pH 7’2. Cette sus- pension cellulaire est passee dans la presse de French. L’extrait obtenu est centrifuge 8. 8000 x q pendant 15 minutes pour hliminer les debris cellu- laires. I1 est ensuite dialysB 5 heures 8. 4” contre 5 litres de DEA M contenant de 1’EDTA et du 2-ME B la concentration de 10-3M. L’extrait est ensuite maintenu B 55” pendant 6 minutes. Les pro- teines ditnaturees sont eliminees par centrifugation et on constate alors l’absence d’homos8rine dkshydro- ghnase dont l’activitb interfhre, dans l’extrait brut, avec le dosage de 1’ASA-deshydrogitnase. A l’aide d’un extrait d’une souche ne synthetisant pas d‘homos6rine dhshydrogenase (JD 144) on a v6rifi6 que ce chauffage ne modifie pas les principales pro- prihtes de l’enzyme.
Dosage de Z’ASA ddshydroqdnme La reaction catalysee par 1’ASA deshydrogenase
est la suivante : P-aspartyl-phosphate + NADPH + semi-aldit-
hyde aspartique + NADP+ + P, + H+. Le dosage, inspire des conditions decrites par
Black et Wright [4], est effectui: dans les conditions suivantes :
Sens de formation d’ASA. On mesure au spectro- photometre B 340mp la vitesse d’oxydation du NADPH. La concentration finale des rbactifs est la suivante: tampon DEA-HC1, pH 8,5, 10-l M ; BAP, 1,2 x 10-3 M; 2-ME 2 x M; NADPH
Sens de formation de B A P . On mesure la vitesse de reduction du NADP+. La concentration finale des reactifs est la suivante: tampon DEA-HC1 pH 8,5, 10-1 M; %ME, 2 x M; P04HK,, 2 x M;
Dans les deux sens de la reaction, la vitesse de- meure constante pendant les 3 premieres minutes
2,4 x 10-3 M.
ASA, 2 x 10-3 M; NADP+, 1’3 x 10-3 M.
pour une quantite d’enzyme donnant une variation d’absorbance infitrieure B 0’3 par minute.
Une unite enzymatique correspond B la quantite d’enzyme donnant une variation d’absorbance de 0,001 (340 mp et I cm de profondeur) par minute. L’activitb spbcifique est exprimbe en unites enzy- matiques par milligramme de prothines.
Les proteines sont dosees par la mhthode de Lowry et coll. [5], la serum albumine bovine cristal- lisee servant de substance Qtalon.
Gradients de glyckrol Les gradients lineaires de densite de glycerol (5 B
30°/, en volume) sont prepares en milieu acittique 10-l M, prealablement ajuste au pH desire par une solution de potasse. Pour les gradients B pH 3 et 6,5 on a v6rifi6, aprhs la centrifugation que le pH n’a pas varie (ceci permettant de faire tous les gradients dans le m6me milieu). Tous les gradients contiennent du KC1 2 x 10-1 M. Dans les conditions exphrimen- tales employees ici le rapport des distances par- courues par la proteine est Bgal au rapport des con- stantes de s6dimentation) ceci pour deux enzymes de reference : Catalase et alcool deshydrogenase de levure. Les gradients sont centrifuges 14 heures B 0” B 25.000 rev./min dans le rotor SW 39 d’une centrifugeuse Spinco modele L. Environ 20 fractions sont recueillies apres la centrifugation. Un gradient de reference A pH 6’5 contenant de l’alcool deshydro- genase est inclus dans chaque experience. Les r6sul- tats des gradients B pH 3, 4 et 5 sont rapport& au gradient de r6fi:rence en tenant compte des indices de refraction. On dittermine la position des pics d’acti- vit6 [6].
Produits L’ASA est prepare selon la mbthode decrite par
Black et Wright [4]. I1 est titre par une preparation purifiee d’homoskrine dkshydrogenase de levure [3].
Le BAP est prepare selon la methode dbcrite par Black et Wright [7] B partir de l’a’-benzyl-ester-,!I- chlorure de l’acide N-carbobenzoxy-L-aspartique. Ce composi: est prepare B partir de l’acide N-carbo- benzoxy-L-aspartique (m6thode utilisee dans la pr6- paration de l’asparagine) [8].
Le P-aspartyl-phosphate est dose par la methode de mise en evidence des acyl-phosphates par l’hydro- xylamine [9], l’hydroxamate de l’acide aspartique servant de substance &talon.
Le NADP+ et le NADPH sont de Sigma Chemical Company. L’acide N-carbobenzoxy-L-aspartique pro- vient de Mann Research Laboratories ainsi que l’hy- droxamate de l’acide aspartique.
Le sulfate de sodium marque par 35S et le bicar- bonate de sodium marqui: par 14C proviennent du Commissariat a 1’Energie Atomique.
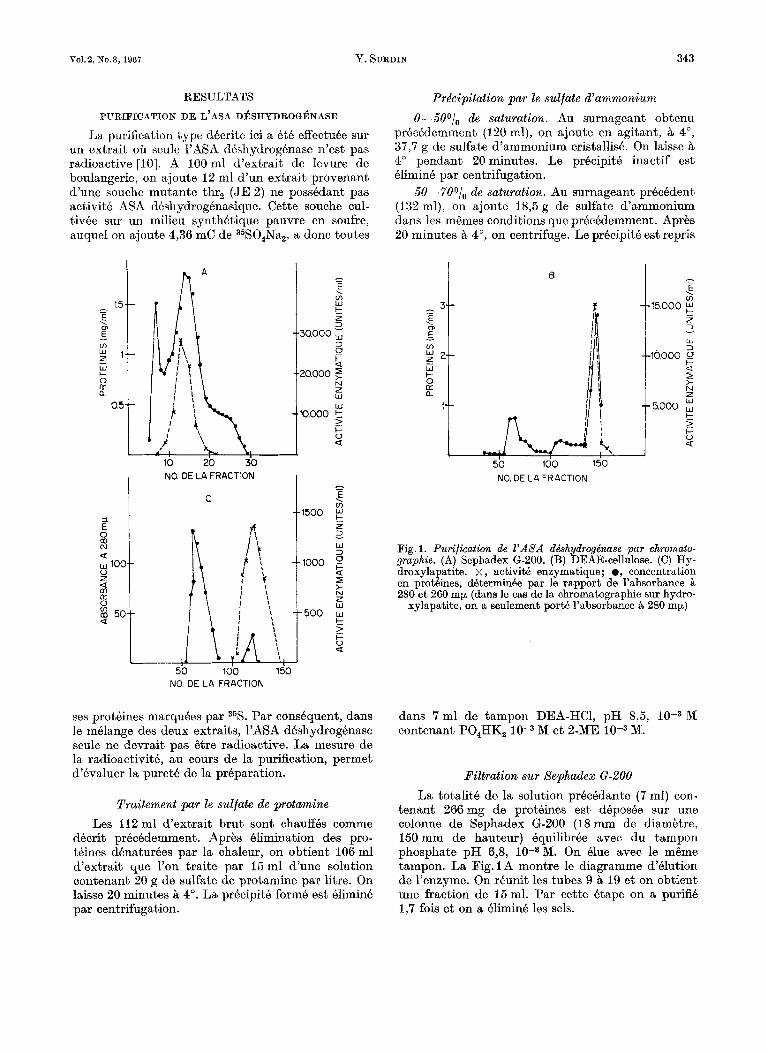
V01.2, n’0.3,1967 Y. SURDIN 343
RESULTATS
PURIFICATION DE L’ASA D~SHYDROGENASE
La purification type decrite ici a A t i t effectuBe sur un extrait oa seule 1’ASA d8shydrog6nase n’est pas radioactive [lo]. A 100 ml d’extrait de levure de boulangerie, on ajoute 12 ml d’un extrait provenant d’une souche mutante thr, (JE 2) ne possedant pas activith ASA dhshydrogknasique. Cette souche eul- tivbe sur un milieu synthetique pauvre en soufre, auquel on ajoute 4,36 mC de 35S0,Na,, a donc toutes
Prdcipitation par le sulfate d’ammonium 0-50°/0 de saturation. Au surnageant obtenu
pritcklemrnent (120 ml), on ajoute en agitant, B 4”, 37,7 g de sulfate d’ammonium cristallis6. On laisse h 4” pendant 20 minutes. Le precipite inactif est Blimini: par centrifugation.
50- roo/,, de saturation. Au surnageant pr6cBdent (132 ml), on ajoute 18,5 g de sulfate d’ammonium dans les mkmes conditions que prhchdemment. Aprhs 20 minutes B 4”, on centrifuge. Le pr8cipitB est repris
I 0 20 30
I NO, DE LA FRACTION I
NO. DE L A FRACTION
ses proteines marquees par 35S. Par conshquent, dans le melange des deux extraits, 1’ASA deshydrogknase seule ne devrait pas 6tre radioactive. La mesure de la radioactivite, au cours de la purification, permet d’bvaluer la puretb de la prkparation.
Traitement par b sulfate de protarnine Les 112 ml d’extrait brut sont chauffBs comme
decrit prbcbdemment. Aprbs Blimination des pro- thines denaturbes par la chaleur, on obtient 106 ml d‘extrait que l’on traite par 15 ml d’une solution contenant 20 g de sulfate de protamine par litre. On laisse 20 minutes B 4”. La pr6cipitB form6 est 6liminB par centrifugation.
I
f 3j
15.000 - z 3
w 3
4
N 2
- 16.000 F“
F 5.000
t 2
a t V
NO. DE L A FRACTION
Fig. 1. Purification de 1’ASA dbhydrogdnase par chromato- grapkie. (A) Sephadex 6-200. (B) DEAE-cellulose. (C) Hy- droxylapatite. x , activite enzymatique; e, concentration en protdines, determinke par le rapport de l’absorbance ?i
280 et 260 mp (dans le cas de la chromatographie sur hydro- xylapatite, on it seulement port6 l’absorbanee B 280 mp)
dans 7ml de tampon DEA-HC1, pH 8,5, iO-3M contenant PO,HK, M et 2-ME M.
Filtration sur Sephadex G-200 La totalit6 de la solution prBc6dante (7 ml) con-
tenant 266mg de protBines est dBposBe sur une colonne de Sephadex G-200 (18mm de diambtre, 150mm de hauteur) Bquilibree avec du tampon phosphate pH 6’8, iO-3M. On Blue avec le m6me tampon. La Fig. 1 A montre le diagramme d’hlution de l’enzyme. On reunit les tubes 9 B 19 et on obtient une fraction de 15ml. Par cette &ape on a purifie 1,7 fois et on a BliminB les sels.

344 ASA dCshydrog6nase chez S. cerevisiae European J. Biochern.
Chromatographie sur DEAE-cellulose Une colonne de DEAE-cellulose (diamktre 18 mm,
hauteur 150mm) est equilibree avec du tampon phosphate pH 6,8, 10-3M. On y depose les 15ml (contenant 161 mg de proteines) obtenus aprks Sephadex G-200 e t on lave avec 60ml du m6me tampon. Puis on Blue, par Btapes, avec des volumes Bgaux (60 ml) de tampons de m6me pH mais de molarit6 croissante en phosphate. L’enzyme est Blue B une concentration de 5 x 10-2 M. En rkunissant les tubes 135 B 150, on obtient une fraction de 26ml. La Fig.1B montre le diagramme d’elution. Cette &ape conduit B une purification de 3 fois par rapport B 1’6tape precedente.
Afin de poursuivre la purification par chromato- graphie sur hydroxylapatite, on abaisse la concen- tration en sels de la preparation en filtrant sur une colonne de Sephadex G-25. On blue avec du tampon phosphate pH 6,8, 10-3M. L’activite est alors contenue dans 33 ml.
hydrogBnase au cours de la purification; ces 2 pheno- menes peuvent agir simultanement dans la prepara- tion.
Le rendement de la purification est de loo/,. La teneur en proteines de cette preparation ne per- mettait par. 1’Qtude des proprietes de l’enzyme. Celles- ci ont donc 6th ittudiees sur la preparation obtenue ap rh precipitation au sulfate d’ammonium.
PROPRII~T~S DE L’ASA DI~SHYDROQI~NASE
pH optimum L’optimum d’activite de 1’ASA deshydrogenase
se situe B pH 8,5 dans les deux sens de la reaction. Les tampons utilises sont des tampons tris-HC1, a une concentration finale de 10-l M. L’itchelle de pH examinee est de pH 7’0 B pH 9,4. Les activites sont mesurees dans les deux sens de la reaction en pre- sence de bicarbonate 10-2 M. Cette concentration de bicarbonate ne fait pas varier le pH.
Tableau 1. Purification de I’ASA dbhydrogdnase de tevuve
Etape de purification Enzyme totales Activite specifique Radioactivite Proteinea
unites coupslmin coups/rnin mg proteines unit6 enzymatique mg protOiues unites mg
Extrait brut 318.000 1.050 298 255 I,I x 104 2,5 x 104
Chromatographie sur DEAE-cellulose 236.000 36,4 6 500 10 6,7 x 104 Chromatographie sup hydroxylapatite 33.800 0,37a 92.000 1
SO,(NH& 50-1Oo/0 327.000 266 1210 175 Chromatographie sur Sephadex G-200 334.000 161 2 090 33 6,8 x lo4
1,3 x 10-a
Valeurs approchees, la determination des proteines dans ce eas &ant imprhcise.
Chromatographie sur hydroxylapatite Ces 33 ml (contenant 36,4 mg de proteines) sont
deposBs sur un colonne d’hydroxylapatite [ I l l (dia- metre 18 mm, hauteur 150 mm) prealablement 6quilibrBe avec du tampon phosphate pK 6,8,
M. Apres lavage avec 60 nil du m6me tampon, l’enzyme est clue par du tampon phosphate pH 63, 5 x M. La Fig. 1 C montre le profil de 1’Blution.
Le Tableau I resume la purification. A ce stade, la teneur en proteines est trop faible
pour permettre une determination prBcise. La radio- activitB specifique (nombre d’impulsions par minute et par unit6 d’enzyme) montre que la purification totale est de 255 fois par rapport SL la teneur en enzyme de l’extrait brut.
Le nombre d’impulsions par minute et par milli- gramme de proteines reste b peu prks constant au cows de la purification; ceci peut indiquer, ou bien, gue, m6me A la fin de la purification, la proportion d’ASA dBshydrog6nase reste tres faible par rapport aux protBines contaminantes, ou bien, que la souche thr, utilisee synthetise une protBine inactive enzy- matiquement mais se comportant comme 1’ASA d6s-
Action du bicarbonate L’activiti: de 1’ASA deshydrogknase est augmentee
de 4 B 5 fois par addition de bicarbonate dans le melange rhactionnel. L’Btude de ce phenomkne a mis en relief les faits suivants:
a) en Bliminant partiellement le bicarbonate des solutions utilisees pour le dosage (distillation sous vide de la diBthanolamine, decarbonatation de l’eau, preparation des rBactifs en atmosphkre d’azote), on perd 75O/, de l’activitk obtenue avec les solutions prbparees sans precautions speciales. L’activite est retrouvee entikrement en ajoutant du bicarbonate au melange ainsi decarbonath. D’autre part B pH 7, oh les solutions contiennent peu de gaz carbonique dissous, l’enzyme perd goo/, de son activite si on supprime le bicarbonate du melange rhactionnel.
b) le bicarbonate augmente la vitesse de la r6ac- tion dans les deux sens. La chromatographie des produits de la reaction, effectuee en presence de bicarbonate marque par W , n’a pas permis de deceler l’incorporation de 14C0,HNa dans un des produits de la reaction. Ces deux faits rendent improbable la participation du bicarbonate en tant gue substrat.

Vo1.2, N0.3,1967 Y. SURDIN 345
Tableau 2. Constantes cine’tiques de Z’ASA de’shydroginase Les concentrations de bicarbonate ajoutkes sont les suivantes: 2 x &!I B pH 7 et 10-1 &I Q pH 8,s ( K , ASA, PO,HK,, arsk- niate, NADP+) 5 x M Q pH 8,5 ( K , BAP et PU’RDPH). Le pH ne varie pas par addition du bicarbonate. Les variations de concentration des substrats sont les suivantes: B pH 7, pour I’ASA do M; pour le PO,HK, de 10-4 M Q W Q 4 x 10-2 M; pour le NADP+ de 4,3 x M; pour le BAP de 2 x 10-5 M Q 6 x M Q 1,8 x M; pourle PO,HK, de 5 x I f ; pour le NADP+ de 4,3 x 1cI Q 4,3 x 10-3M; pour le BAP M. n eat la pente de la droite obtenue en portant en abscisse log [S] e t en ordonnke log w/( V,,, - w), [S] Btant la concentration du substrat, w la vitesse pour cliaque
concentration en substrat e t V,’,,, la vitessc ?I concentration saturante de substrat
M B p H 7 et
M; pour l’arskniate de 5 x M 8. 2 x
M Q 1,7 x M; pour le NADPH de 1,2 x M A 2,4 x M. A p H 8,5: pour 1’ASA de 2,2 X
M Q lo-, M; pour l’arskniste de 5 x M ?I 4 x M Q 1,3 x M; pour le NADPH de 1,2 x lop5 M A 4,8 x
K , n Substrat variable
PH 7 pH 8,5 PH 7 pH 8,6
ASA 1,2 x 10-4 M ASA + C0,HNa 3,3 x 10-4 M P04HK, s,o x 10-3 M PO,HK, + C0,HNa 8.0 x 10-3 M Argkniaie 6 ; ~ x 10-3 M Arskniate + C0,HNa 6,2 x 10-3 M NADP+ 3,3 x 10-4 M NADP+ + C0,HNa 3,3 x 10-4 $1 BAP 2,7 x 10-4 1c1 BAP + C0,HNa 2,9 x 10-4 M NADPH 3,o x 10-5 M NADPH + C0,HNa 3,0 x 10-5 M
i ,3 x 10-3 M i ,3 x 10-3 M 7,o x 10-4 M 5,o x 10-3 JC 1,s x 10-3 hi 9,o x 10-3 M 5,4 x 10-4 M 5,4 x 10-4 M
i ,6 x 10-3 M 1,2 x 10-4 iw 1,2 x 10-4 M
8,l x M
c) 30 clones reverses provenant de differentes souches mutantes thr, ont &ti: examinbs. Dam tous les cas 1’ASA dbshydrogbnase est activbe par le bicarbonate m6me chez les mutants reverses in- complets.
d) les constantes cinetiques de la reaction (Ta- bleau 2 ) dbterminees au pH optimum de la reaction (pH 8,5) et A pH 7 , montrent que l’addition de bicarbonate ne modifie que le K , du phosphate ii pH 8,5.
Les cinbtiques pour tous les substrats semblent repondre ii la loi de Henri-Wchaelis. Dans tous les cas on n’observe pas de coopbrativitb entre les molecules de substrat, en presence ou en absence d’un exc&s de bicarbonate [12]. Differentes hypo- th&ses possibles sur le mode d’action du bicarbonate seront examinees clans la discussion.
POIDS MOL~CULAIRE
Le poids moleculaire de 1’ASA cl8shydroghase a b tb estimb par filtration sur gel de Sephadex G-200 [13,14]. La colonne utilisee (diambtre 25 mm et hauteur 400mm) est bquilibrbe avec du tampon phosphate pH 6 3 , lop3 M, contenant du KCl 10-1 M. Ce tampon sert aussi $ l’blution. Les enzymes de reference sont les suivants: cytochrome c coeur de cheval (Sigma Chem. Co.: M = 12.400); peroxydase de raifort (Sigma Chem. Co: M = 40.000); alcool deshydrogenase de levure (Worthington : M = 126.000) ; glutamique deshydrogenase de foie (Sigma Chem. Co.).
Trois experiences indbpendantes donnent pour 1’ASA dbshydrogenase les poids molbculaires sui- vants: 139.000; 140.000; 140.000 (Fig.2).
ALCOOL DESHYDROGENASE
3 100- 0 DESHYDROGENASE >
c
POIDS MOLECULAIRE Fig.2. Ddternzination de la inasse moldculaire de 1’AXA dks- hydrogdnase par filtration sur gel de Xephadex (2-200. Les points reprhsentent les positions des pies d’activitk de chaque rnarquenr utilisir. La fli-che indique la position du pic d’activiti.
de 1’ASA dkshydrogenasc
INHIBITION
Les acides amin& terininaux de la chaine mbta- bolique, threonine et mbthionine, seuls ou en com- binaison, ne sont pas inhibiteurs. Parmi tous les acides aminits essay&, seules la cystbine et l’homo- cystbine inhibent l’activith de 1’ASA dbshydrogknase de fagon importante. Dans le sens AXA-t BAP l’inhibition est de type compbtitif vis-A-vis du sub- strat aminb et non-compbtitif vis-$-vis du phosphate et du NADP (le Km pour I’ASA varie de 1,3 x M a 4,6 x lop3 M en presence de cystitine ou d’homo- cysteine 10-2M, tandis yue la V,,, ne varie pas. Dans le cas du phosphate et du NADP+ les Km
346 ASA d6shydrogAnase ohez 8 . cerevisiae European J. Biochem.
restent inchanges en presence de cette concentra- tion d’inhibiteur, seules les V,,, varient). D’autre part, la concentration de cysteine et d’homocysteine necessaire pour obtenir 50°/, d’inhibition est de
I1 faut remarquer que cette inhibition, observbe en presence de 2-ME B la concentration de 2 x M, ne peut pas &re aishment expliquee par une reduc- tion de ponts disulfure qui serait necessaire ti l’acti- vite de l’enzyme puisque ce dernier n’est actif qu’en milieu r&ducteur, (2-ME ou glutathion). De plus ces inhibitions sont competitives vis-A-vis du substrat aminB (ASA) ce qui semble indiquer une inter- ference particulibre avec l’accessibilite du centre actif de l’enzyme pour le substrat aminh. La signification
M.
t 1
physiologique de cette inhibition n’apparait pas clairement car, d’une part, dans le sens biosynthb- tique BAP + ASA, l’homocyst6ine et la cysteine inhibent trks peu et d’autre part, les formes D de ces deux acides amin& sont egalement inhibitrices.
INACTIVATION THERMIQUE
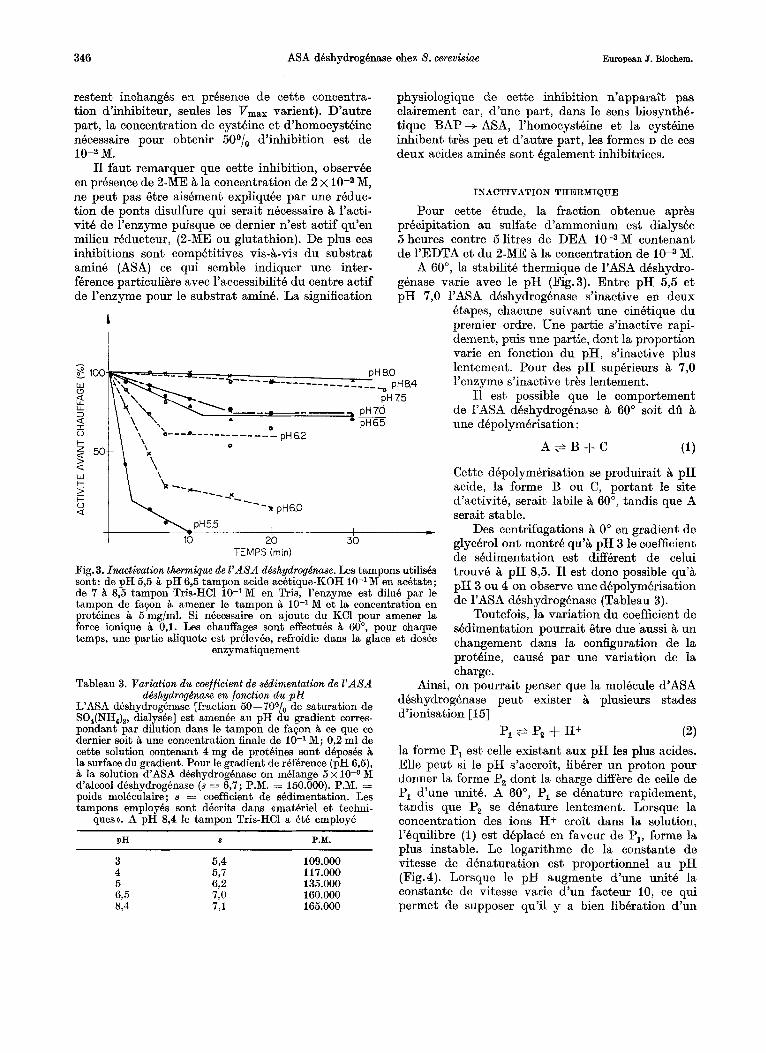
Pour cette etude, la fraction obtenue aprks precipitation au sulfate d’ammonium est dialysee 5 heures contre 5litres de DEA 10-3M contenant de 1’EDTA et du 2-ME t i la concentration de M.
A 60°, la stabilite thermique de 1’ASA deshydro- gknase varie avec le pH (Fig.3). Entre pH 5,5 et pH 7,O 1’ASA deshydrogbnase s’inactive en deux
h, pH 5.5 I I 7 +
10 20 30 TEMPS (rnin)
Fig. 3. Inactivation thermique de Z’ASA ddshydrogdnase. Les tampons utilishs sont: de p H 5,5 it pH 6,5 tampon acide acktique-KOH 10-l M en acetate; de 7 % 8’5 tampon Tris-HC1 10-IM en Tris, l’enzyme est dilue par le tampon de fapon amener le tampon & 10-lM et la concentration en proteines it 5mg/ml. Si necessaire on ajoute du KCl pour amener la force ionique it 0,l. Les chauffages sont effectub % 60°, pour chaque temps, une partie aliquote est prblevee, refroidie dans la glace et dosee
enzymatiquement
Tableau 3. Variation du coefficient de sddimentation de 1’ASA dbhydrogdnase en fonction du pH
L’ASA d6shydroghnase [fraction 50-70°/,, de saturation de SO,(NH,),, dialys6el est amenbe au p H du gradient corres- pondant par dilution dans le tampon de fapon & ce que ce dernier soit & une concentration finale de 10-1 M; 0,2 ml de cette solution contenant 4 mg de proteines sont deposes it la surface du gradient. Pour le gradient de reference (pH 6,5), it la solution d’ASA deshydrogenase on melange 5 x 10P M d’alcool deshydrogenase (s = 6,7; P.M. = 150.000). P.M. = poids moleculaire; s = coefficient de sedimentation. Les tampons employes sont decrits dans ccmathiel e t techni-
ques)). A p H 8,4 le tampon Tris-HC1 a 6tB employe
PH 8 P.M.
3 534 109.000 4 597 117.000 5 6 3 135.000 6,5 7 8 160.000 8-4 7 J 165.000
. -
Qtapes, chacune suivant une cinetique du premier ordre. Une partie s’inactive rapi- dement, puis une partie, dont la proportion varie en fonction du pH, s’inactive plus lentement. Pour des pH superieurs 7 , O l’enzyme s’inactive trks lentement.
I1 est possible que le comportement de 1’ASA dhshydrogknase B 60” soit dii B une depolymkrisation :
A + B + C (1)
Cette depolymerisation se produirait B pH acide, la forme B ou C, portant le site d’activith, serait labile B B O O , tandis que A serait stable.
Des centrifugations B 0” en gradient de glychrol ont montrh qu’b pH 3 le coefficient de sedimentation est diffbrent de celui trouve B pH 8,5. I1 est donc possible qu’8 pH 3 ou 4 on observe une depolymbrisation de 1’ASA deshydrogenase (Tableau 3).
Toutefois, la variation du coefficient de sedimentation pourrait Btre due aussi B un changement dans la configuration de la proteine, cause par une variation de la charge. ”
Ainsi, on pourrait penser que la molhcule d’ASA deshydrogenase peut exister B plusieurs stades d’ionisation [15]
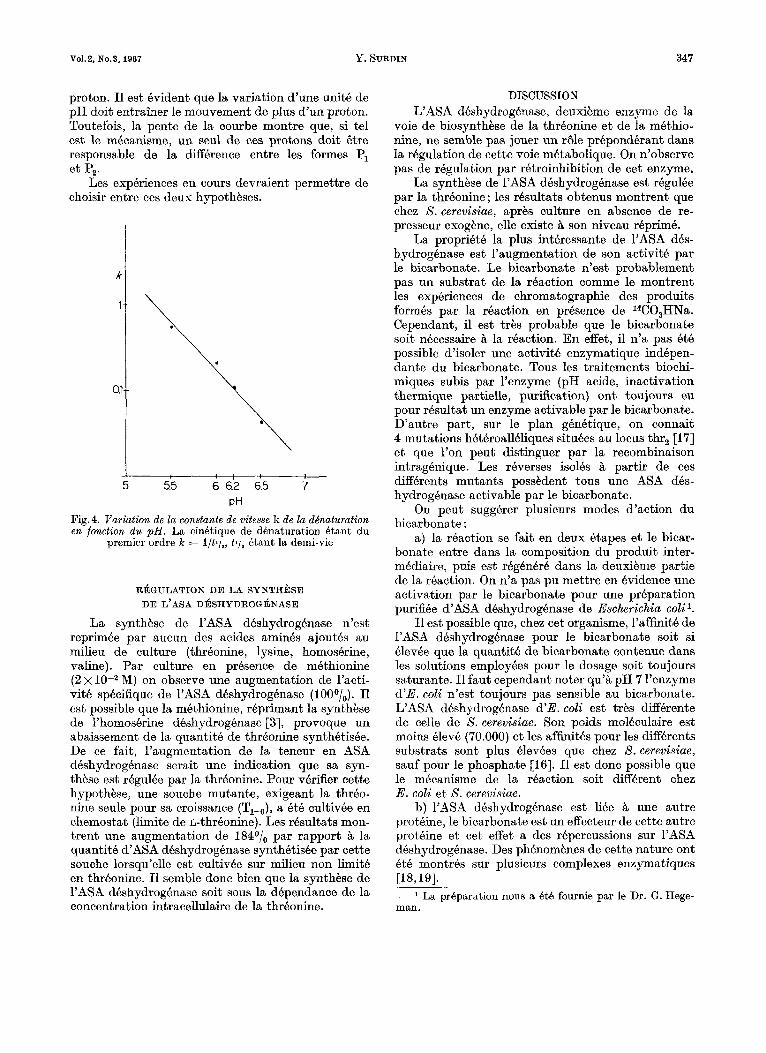
la forme Pl est celle existant aux pH les plus acides. Elle peut si le pH s’accroit, libhrer un proton pour donner la forme Pz dont la charge diffkre de celle de PI d’une unite. A 60”’ Pl se denature rapidement, tandis que Pz se denature lentement. Lorsque la concentration des ions Hf croit dans la solution, 1’8quilibre (1) est d8place en faveur de PI, forme la plus instable. Le logarithme de la constante de vitesse de denaturation est proportionnel au pH (Fig.4). Lorsque le pH augmente d’une unite la constante de vitesse varie d’un facteur 10, ce qui permet de supposer qu’il y a bien liberation d’un
Pi + Ps + H+ (2)
Vol.2, N0.3, 1967 Y. SURDIN 347
proton. I1 est evident que la variation d’une unit6 de pH doit entrainer le mouvement de plus d‘un proton. Toutefois, la pente de la courbe montre que, si tel est le mecanisme, un seul de ces protons doit &re responsable de la difference entre les formes PI et I?,.
Les experiences en cows devraient permettre de choisir entre ces deux hypothkses.
I 5 55 6 6.2 6.5 7
PH Fig.4. Variation de la constante de vitesse k de la ddnaturation en fonction d u p H . La cinbtique de d6naturation Btant du
premier ordre k = l / t l /a , ti/ , Btant la demi-vie
REGULATION DE LA SYNTH~SE DE L’ASA DESHYDROGENASE
La synthkse de 1’ASA deshydrogenase n’est reprimhe par aucun des acides amines ajoutes au milieu de culture (threonine, lysine, homos&rine, valine). Par culture en presence de methionine (2 x M) on observe une augmentation de l’acti- vite specifique de 1’ASA deshydrogenase ( iOOo/o) . I1 est possible que la mbthionine, reprimant la synthese de l’homosbrine dbshydrogenase [3], provoque un abaissement de la quantiti: de threonine synthhtishe. De ce fait, l’augmentation de la teneur en ASA deshydrogenase serait une indication que sa syn- these est regulee par la threonine. Pour verifier cette hypothkse, une souche mutante, exigeant la threo- nine seule pour sa croissance (T1-,J, a 6th cultivee en chemostat (limite de L-thrbonine). Les resultats mon- trent une augmentation de 1840/, par rapport b la quantite d’ASA deshydrogbnase synthetisee par cette souche lorsqu’elle est cultivhe sur milieu non limit6 en threonine. I1 semble donc bien que la synthkse de 1’ASA deshydrogenase soit sous la dbpendance de la concentration intracellulaire de la thrbonine.
DISCUSSION L’ASA deshydrog&nase, deuxikme enzyme de la
voie de biosynthhe de la threonine et de la mhthio- nine, ne semble pas jouer un r61e preponderant dans la regulation de cette voie metabolique. On n’observe pas de regulation par retroinhibition de cet enzyme.
La synthkse de 1’ASA deshydrogenase est regulee par la threonine ; les resultats obtenus montrent que chez S. cerevisiae, aprks culture en absence de re- presseur exogkne, elle existe b son niveau reprime.
La propriBt6 la plus interessante de 1’ASA des- hydrogenase est l’augmentation de son activite par le bicarbonate. Le bicarbonate n’est probablement pas un substrat de la reaction comme le montrent les experiences de chromatographie des produits form& par la reaction en presence de 1*CO,HNa. Cependant, il est trks probable que le bicarbonate soit necessaire b la reaction. En effet, il n’a pas Btb possible d’isoler une activite enzymatique indepen- dante du bicarbonate. Tous les traitements biochi- miques subis par l’enzyme (pH acide, inactivation thermique partielle, purification) ont toujours eu pour resultat un enzyme activable par le bicarbonate. D’autre part, sur le plan gbnbtique, on connait 4 mutations hBteroall6liques situees au locus thr, [i7] et que l’on peut distinguer par la recombinaison intragbnique. Les reverses isoles b partir de ces Werents mutants posskdent tous une ASA des- hydrogenase activable par le bicarbonate.
On peut suggerer plusieurs modes d’action du bicarbonate :
a) la reaction se fait en deux Btapes et le bicar- bonate entre dans la composition du produit inter- mitdiaire, puis est rBg6nhrA dans la deuxieme partie de la reaction. On n’a pas pu mettre en evidence une activation par le bicarbonate pour une preparation purifiee d’ASA deshydrogbnase de Escherichia coli
I1 est possible que, chez cet organisme, l’affinit6 de I’ASA dbshydrogenase pour le bicarbonate soit si &levee que la quantitb de bicarbonate contenue dans les solutions employees pour le dosage soit toujours saturantc. I1 faut cependant noter qu’b pH 7 l’enzyme d’E. coli n’est toujours pas sensible au bicarbonate. L’ASA deshydrogenase d’E. coli est trks differente de celle de S. cerevisiae. Son poids mol6culaire est moins Bled (70.000) et les affinitks pour les Werents substrats sont plus Blevees que chez 8. cerevisiae, sauf pour le phosphate [16]. I1 est donc possible que le mkcanisme de la reaction soit different chez E. coli et S. cerevisiae.
b) I’ASA deshydrogenase est liBe b une autre proteine, le bicarbonate est un effecteur de cette autre proteine et cet effet a des repercussions sur 1’ASA deshydrogenase. Des phenomknes de cette nature ont 6th montres sur plusieurs complexes enzymatiques [18,19].
La preparation now a 6tB fournie par le Dr. G. Hege- man.

348 Y. SURDIN : ASA deshydrogbnase chez 8. cereviske European J. Biochem.
c) le bicarbonate est necessaire B la structure de l’enzyme. En effet, les vitesses maximales sont augmentees par addition de bicarbonate dans tous les cas. Par contre, l’affinite pour le phosphate B pH 8,5 est diminuBe d’un facteur 10 environ.
D’aprBs 1’Qtude cinbtique de 1’ASA dbshydro- gBnase, il semble que le bicarbonate agisse sur le site du phosphate de fapon & augmenter la vitesse maxi- male et b diminuer l’affiniti: pour ce substrat. D’autre part, on peut admettre pour 1’ASA d6s- hydrogenase un mecanisme similaire B celui admis pour la glycBraldBhyde-3-phosphate dBshydrogBnase :
ASA + Enz-SH + NADP+ s aspartyl-S-Enz + NADPH oxydation
phosphorylation.
X i la vitesse de la phosphorylation est infhieure b celle de l’oxydation, la partie limitante de la rBac- tion serait la phosphorylation. Le bicarbonate augmentant la Vm,, pour le phosphate, entrainerait done une augmentation de la Vm,, pour les autres substrats.
A pH 7,0, il semble que la conformation de l’enzyme est telle gue le bicarbonate ne peut plus modifier l’affiniti: pour le phosphate, mais l’effet sur les vitesses subsiste.
Par analogie avec la glycBraldBhyde-3-phosphate dBshydrog6nase [20] un essai a B t B fait pour dissocier effectivement la reaction en deux parties : l’oxyda- tion et la phosphorylation. Des rksultats prhliininaires montrent qu’on peut doser l’oxydation seule en employant un mercaptan comme accepteur d’aycle B la place du phosphate ou de l’arshniate. On a pu montrer que le 2-ME pelt-6tre employe comme accepteur d’acyle. Mais le K , pour le 2-ME dans ces conditions (en absence de phosphate) est tr&s BlevB (environ 10-l M). L’affinitB du complexe aspar- tyl-S-enzyme pour le phosphate est donc plus grande que pour le 2-ME. En prBsence de phosphate et de 2-ME 0,02 M (conditions du dosage) c’est donc bien la phosphorylation qui doit se produire.
aspartyl-S-Enz + PO,= =+ BAP
L’etude de ce mbcanisme est actuellement en cours.
J e remercie Madame Denise Henri e t Mademoiselle Frangoise Eichler pour leur collaboration technique. Que Madame H. de Robichon-Szulmajster qui a dirigb ce travail depuis le debut trouve ici l’expression de ma reconnaissance. Ce travail a b6nkficih de I’aide de la Dblkgation Gknkrale A la Recherche Scientifique et Technique (Convention 61-Fr- 063) e t du Dhpartement de Biologie du Commissariat ti 1’Energie Atomique.
BIBLIOGRAPHIE 1. de Robichon-Szulmajster, H., Surdin, Y., e t Mortimer,
2. de Robichon-Szulmajster, H., et Corrivaux, D., Biochim.
3. Kassarevitch, Y., e t de Robichon-Szulmajster, H., Bio-
4. Black, S., et Wright, N. G., J . Biol. Chem. 213 (1955) 39. 5 . Lowry, 0. H., Rosenbrough, N. J., Farr, A. L., et Ran-
6. Martin, R. G., e t Ames, B. N., J . Biol. Chem. 236 (1961)
7. Black, S., e t Wright, N. G., J. Biol. Chem. 213 (1955) 27. 8. Brown, G. B., Biochemical Preparations, 10 (1963) 10. 9. Lipmann, F., et Tuttle,L. C., J . Biol. Chem. 159 (1945) 21.
10. Truffa-Bachi, P., e t Cohen, G. N., Biochim. Biophys. Acta, 113 (1966) 531.
11. Peterson, E. A., e t Sober, H. A., Dans Methods in En- zymology (edited by s. P. Colowick and N. 0. Kaplan), Academic Press, New York 1962, Vol. V, p. 3.
12. Monod, J., Wyman, J., et Changeux, J. P., J . Mol. Biol. 12 (1965) 88.
13. Iwatsubo, M., e t Curdel, A., Compt. Rend. 256 (1963) 5224.
14. Andrews, P., Biochern. J . 91 (1964) 222. 15. Laidler, K. J., The Chemical Kinetics of Enzyme Action,
Oxford University Press, Oxford 1958, p. 336. 16. Hegeman, G., Communication personnelle. 17. Luzzati, M., Thkse Facult& des Sciences, Paris 1965. 18. Bailin, G., e t Lukton, A,, Biochim. Biophys. Acta, 128
(1966) 317. 19. Patte, J. C., Truffa-Bachi, P., Le Bras, G., et Cohen,
G. N., Biochim. Biophys. Acta, 128 (1966) 426, 440, 450.
20. Wolff, E. C., e t Black, S., Arch. Biochena. Biophys. 8 (1959) 236.
R. K., Genetics, 53 (1966) 609.
Biophys. Acta, 73 (1963) 248.
chim. Biophys. Acta, 73 (1963) 414.
dall, R. J., J . Biol. Chem. 193 (1951) 265.
1372.
Y. Surdin Laboratoire d’Enzymologie du C.N.R.S. 91 Gif-sur-Yvette, France


















![Glucides - 2012 [Mode de compatibilit ]biologie23.e-monsite.com/medias/files/glucides-4.pdf · Un glucide se définit comme un aldéhyde ou une cétone d’un polyalcool Composés](https://img.pdfslide.fr/doc/110x75/5e3798b08c619a00200fad05/glucides-2012-mode-de-compatibilit-biologie23e-un-glucide-se-dfinit-comme.jpg)










