Embed Size (px)
Citation preview

La signalisation cellulaire, un jeu d’interactions protéiques et de modifications post-traductionnelles
I. Introduction et présentation de l’UMR 3 biologie cellulaire
II. Signalisation cellulaire
A- Physiologie de la signalisation B- Médiateurs et récepteurs
1) Médiateurs 2) Récepteurs
C- Mécanismes généraux et acteurs de la signalisation
III. Modifications post traductionnelles A- Modifications stables lors de la biosynthèse
1) Glycosylation 2) Acylation
B- Modifications labiles lors de la biosynthèse 1) Phosphorylation 2) Acétylation
C- Modifications stables lors de la dégradation
IV. Interactions protéiques et méthodes d’études A- Co-immunoprécipitation B- Double hybride chez la levure et pull down assay
1) Double hybride 2) GST pull down assay
C- FRET-BRET 1) FRET 2) FRET vs BRET 3) FRET et FLIM
D- Intégration du signal
UE11 - Parcours 3 Biologie cellulaire et moléculaire
de la signalisation - Cours n°1 24/02/2016
Pr. Eric Clauser [email protected] [email protected]
RT : Colleen Breslin Ségolène Camus RL : Kim Vân Lussiez

Abréviations : RCPG : Récepteur couplé à une protéine G Aa : acide aminé IL : interleukine PM : poids moléculaire Ach : Acétyl choline RE : réticulum endoplasmique IC : intracellulaire CoIP : co immuno précipitation WB : Western Blot GFP : green fluorescent protein GST : glutathion S transférase
Mot du RT : Cours assez long mais quasiment tout avait déjà été vu en P1, donc c’est plutôt facile. Cours relu et corrigé par le prof

I. Introduction et présentation de l’UMR 3 biologie cellulaire But de l’enseignement: Acquérir les bases conceptuelles et méthodes de la recherche actuelle dans le domaine de la communication entre les cellules et comprendre les avancées récentes de la biologie cellulaire . 12 cours donnés par des chercheurs de l’INSERM, du CNRS, ou des universitaires spécialisés dans le domaine . Epreuves : Dates provisoires : 15 juin, 17h-19h30, Modalités : En français, rédactionnel, Etude portant sur un des 12 thèmes vus en cours Questions de cours (25%) et d’analyses de figure (75%) Avant les examens, des annales et leurs corrections seront disponibles sur le Moodle.
II. Signalisation cellulaire
A- Physiologie de la signalisation La signalisation cellulaire consiste en la fixation d’un ligand sur un récepteur entrainant un ensemble d’évènements, pour aboutir à un effet sur la cellule. Les modes de transmissions : Les modes de transmissions de l’information sont : . Endocrine : un médiateur est fabriqué par une cellule ou un organe, puis il est transporté par la circulation sanguine pour aller agir à distance sur son organe cible. C’est le mode de transmission le plus classique. . Paracrine : dans un même tissu, un type cellulaire fabrique un médiateur qui va agir sur une cellule voisine à fonction différente. . Autocrine : Un médiateur est fabriqué par une cellule et agit sur la cellule elle-même. Les médiateurs et récepteurs varient en fonction du mode de transmission utilisé. ➢ Système endocrine Il existe 3 systèmes endocrines : l’axe hypothalamo-hypophysaire, les systèmes de régulation métabolique et les systèmes de maintien de l’équilibre électrolytique. Les médiateurs sont des molécules hydrophobes, des stéroïdes ou des molécules hydrophiles : bioamines, peptides, protéines, glycoprotéines. Ces médiateurs se fixent à des récepteurs qui peuvent être intracellulaires (pour les molécules hydrophobes) ou membranaires : RCPG ou récepteur-enzyme. ➢ Système paracrine : le système nerveux Le système nerveux fonctionne sur un mode paracrine puisque la transmission synaptique se fait entre 2 neurones à proximité via la fente synaptique. Les médiateurs sont ici des neurotransmetteurs, tous hydrophiles. Ce sont des aa (glutamate, glycine) ou des dérivés d’aa (dopamine, noradrénaline, adrénaline dérivent de la tyrosine, la sérotonine dérive du tryptophane, l’histamine de l’histidine, le GABA du glutamate.) Ces derniers se fixent à des récepteurs membranaires RCPG ou récepteur-canaux. ➢ Un exemple de la combinaison des 3 modes de transmission : Le système immunitaire

Le système immunitaire fonctionne sur un mode endocrine, paracrine ou autocrine, avec en plus des communications cellulaires directes. Les médiateurs sont principalement des cytokines (IL 1 à 12, interféron, CSF facteur stimulant les colonies, facteurs de croissance (GH), chémokines). Les cytokines sont soit des peptides, soit des protéines, elles sont donc toutes hydrophiles. Les récepteurs sont des récepteurs membranaires de type enzyme ou RCPG.
B- Médiateurs et récepteurs
1) Les médiateurs Les médiateurs sont des molécules dont les propriétés chimiques déterminent le site de fixation. Ainsi, existe-t-il des médiateurs : ➢ Hydrophobes : Hormones : stéroïdes, thyroïdiennes, cortisol Lipides : isoprénoides (acide rétinoïque), acides gras, écosanoides, phosphoglycérolipides, cannabinoides… Vitamines : vit D3 Ces derniers sont capables de passer la bicouche lipidique de la membrane et de se fixer à des récepteurs intracellulaires. ➢ Hydrophiles : aa et dérivés : neurotransmetteurs (glutamate GABA), hormones (noradrénaline) Peptides : neurotransmetteurs (enképhaline), hormones (hypothalamique, vasoactive, gastrointestinale) Protéines, glycoprotéine, lipoprotéine : hormones (hypophysaire, insuline), cytokine, facteur de croissance ➢ Autres : lumière, odeur (mécanismes plus complexes)
2) Les récepteurs Les récepteurs peuvent être localisés sur la membrane ou à l’intérieur de la cellule et sont constitués d’hélices, de feuillets et de boucles. ➢ Récepteurs intracellulaires :
Il s’agit des récepteurs aux médiateurs hydrophobes. Ils possèdent un domaine de liaison à l’hormone et un à l’ADN qui leur permet de réguler la transcription d’un gène. Leur activation passe souvent par une dimérisation. ➢ Récepteurs membranaires :
L’activation d’un récepteur nécessite, en plus de la fixation du médiateur, un changement de sa conformation. A l’état inactivé, le récepteur est lié à des co-répresseurs qui l’empêchent de se lier à l’ADN ou à des protéines. Quand le médiateur vient se fixer, des coactivateurs se lient aussi au récepteur, qui se détache de ses co-répresseurs. Le récepteur pourra donc se lier à l’ADN ou à des protéines. Il en existe 3 classes : les RCPG, les récepteurs canaux, les récepteurs enzymes.
Les récepteurs canaux sont en général constitués de plusieurs sous unités (5 le plus souvent). La fixation du ligand à au moins 2 sous-unités engendre un changement de conformation et conditionne leur ouverture (ou leur fermeture) et laisse ainsi passer de façon plus ou moins spécifique des ions.

C’est cette modification des concentrations ioniques qui va provoquer des changements dans la cellule Ex : récepteur de l’Ach à la jonction neuro-musculaire
Les récepteurs enzymes sont généralement constitué d’un domaine extracellulaire, d’un domaine transmembranaire (le plus souvent une hélice alpha avec des AA hydrophobes) et d’un domaine intracellulaire, sur lequel se trouve une enzyme. La fixation du médiateur induit l’activation de l’enzyme souvent liée à la dimérisation du récepteur. Les plus courants sont les récepteurs Tyrosine-kinase (ex : récepteur à insuline, facteur de croissance…) bien qu’il en existe d’autres sortes : tyrosine-phosphatase, sérine kinase, guanylate cyclase…
Les RCPG sont des récepteurs à 7 domaines transmembranaires (7 hélices alpha) formés de 3
modules. Le module récepteur est associé au deuxième module, une protéine G. A l’état inactif, la protéine G est liée au GDP. Lors de la fixation d’un ligand, celle-ci passe à l’état actif liée au GTP. Elle peut alors activer le module effecteur, des enzymes (adénylyl cyclase, phospholipase C…) qui produisent des 2nds messagers.
C- Mécanismes généraux, grands principes et acteurs
Rappelons que les cibles de la signalisation sont l’ouverture/fermeture des canaux ioniques, l’activité des enzymes, des transporteurs, des protéines de structure et la régulation de facteurs de la transcription. Pour que ces changements puissent aboutir, il ne suffit pas que le médiateur se lie à son récepteur. On dénombre 3 modifications essentielles des protéines de signalisation : ➢ des modifications post-traductionnelles rapides, en particulier les cascades de phosphorylation

➢ l’association à un 2nd messager Ex de l’AMPc : la fixation d’un ligand sur un RCPG aboutit à l’activation d’une protéine Gs et donc à celle de l’adénylyl cyclase, augmentant ainsi la quantité d’AMPc. Cette dernière peut se lier à un certain nombre de cibles, et, en particulier à la protéine kinase dépendante de l’AMPc (PKAc). Cette dernière forme un complexe hétérotétramérique avec 2 PKAr. La fixation de l’AMPc va libérer la PKAc qui pourra aller agir sur ses cibles.
➢ la formation d’un complexe multi-protéique. La présence
de protéines dites d’échafaudage est parfois nécessaire pour activer un
complexe multiprotéique.
Pour terminer, tous ces mécanismes peuvent être bien évidemment associés.
III. Modifications post-traductionnelles Deux choses sont essentielles pour la signalisation cellulaire : d’une part les modifications post-traductionnelles (par exemple la phosphorylation permettant l’activation ou l’inhibition des protéines) et un système d’interaction entre protéines.
A- Modifications stables lors de la biosynthèse

Elles sont en générales stables et définitives.
1) Glycosylation
Il s’agit de l’ajout d’un sucre et il existe deux types de glycosylation : - La N-glycosylation : le sucre est lié à un acide aminé par une fonction NH2, cet acide aminé
est toujours une asparagine dans une séquence Asn-X-Ser (Thr ou Cys). Cette N-glycosylation a lieu à l’intérieur des vésicules du RE et du Golgi (maturation dans le Golgi) et ainsi ne concerne que les parties extracellulaires des protéines. Cette glycosylation est prédictible, et a pour fonction : la stabilité des protéines, les interactions, l’adhésion cellulaire.
- L’O-glycosylation : le sucre est associé à une sérine ou thréonine, ces sucres sont plus courts et moins ramifiés que les N-glycosylations. Les serines et thréonines sont aussi des acides aminés qui peuvent être phosphorylés, il existe donc une compétition entre la phosphorylation et la O-glycosylation. 2) Acylation
Cela correspond à l’ajout d’un acide gras sur un acide aminé spécifique (souvent cystéine et lysine). Ils sont ajoutés dans le RE et le Golgi, cet ajout a pour rôle de transformer une protéine cytosolique en protéine membranaire. La protéine peut alors s’ancrer à la membrane plasmique mais aussi à la membrane des mitochondries ce qui permet de localiser les protéines dans les différents compartiments cellulaires.
B- Modifications labiles lors de la signalisation 1) Phosphorylation
Phosphorylation : adjonction d’un groupement phosphate sur un substrat avec un groupement hydroxyle grâce à l’hydrolyse de l’ATP par une kinase. La déphosphorylation se fait grâce à une phosphatase. Il existe des kinases et phosphatases pour les protéines, pour les sucres, pour les lipides… On distingue deux types de protéines kinases et phosphatases : les sérines –thréonines kinases et les tyrosines kinases. La majorité des phosphorylations concerne les sérines et thréonines (99.9%), néanmoins les phosphorylations sur les tyrosines sont très importantes car elles concernent la croissance cellulaire. Exemple des MAP kinases :

Le médiateur active son récepteur et ainsi active son domaine tyrosine-kinase. Ceci permet le recrutement et l’activation de p21ras, elle-même recrute une sérine-tyrosine kinase Raf-1 et l’active. Raf-1 va alors phosphoryler Mek , qui peut ensuite phosphoryler les MAPK ERK1 et ERK2. La phosphorylation de ces deux MAPK induit leur translocation dans le noyau où elles peuvent activer des facteurs de transcription, des enzymes… Exemple de cancers corticosurrénaliens : 90 % des cancers de la corticosurrénale fabriquent massivement un facteur de croissance l’IGF2 et 10% n’en fabriquent pas. On cherche à déterminer une relation entre la production d’IGF2 et la phosphorylation d’ERK et d’Akt.
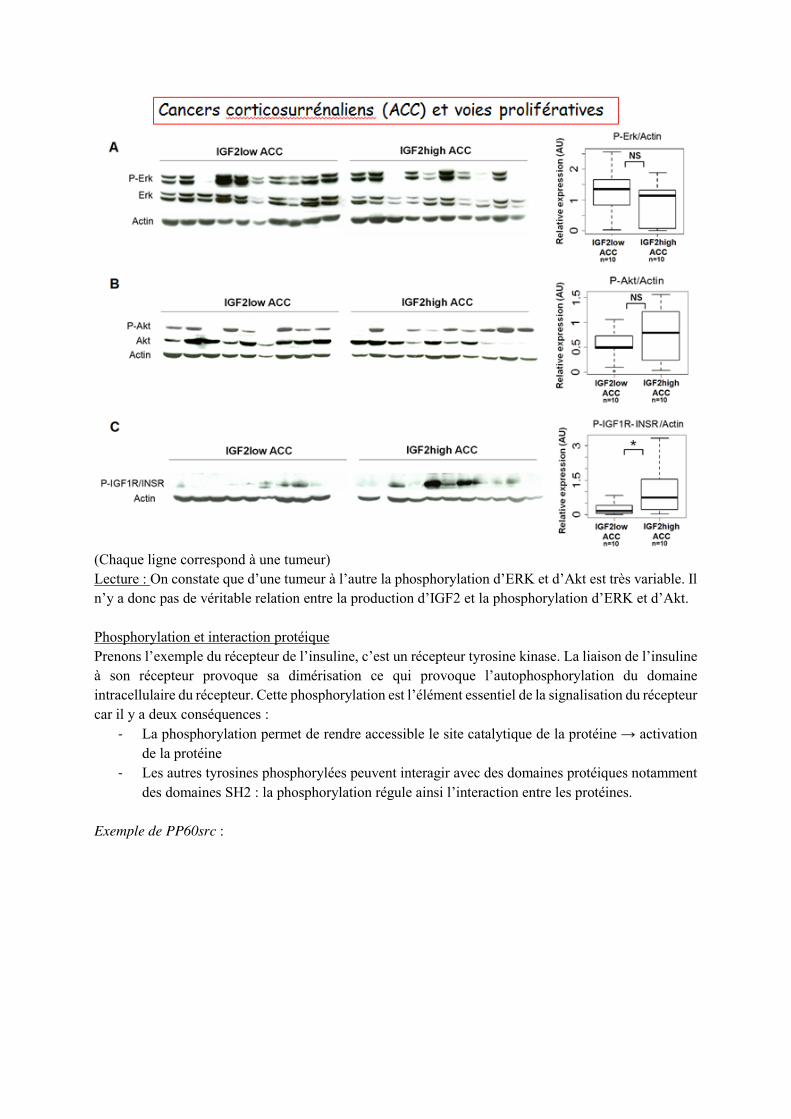
(Chaque ligne correspond à une tumeur) Lecture : On constate que d’une tumeur à l’autre la phosphorylation d’ERK et d’Akt est très variable. Il n’y a donc pas de véritable relation entre la production d’IGF2 et la phosphorylation d’ERK et d’Akt. Phosphorylation et interaction protéique Prenons l’exemple du récepteur de l’insuline, c’est un récepteur tyrosine kinase. La liaison de l’insuline à son récepteur provoque sa dimérisation ce qui provoque l’autophosphorylation du domaine intracellulaire du récepteur. Cette phosphorylation est l’élément essentiel de la signalisation du récepteur car il y a deux conséquences :
- La phosphorylation permet de rendre accessible le site catalytique de la protéine → activation de la protéine
- Les autres tyrosines phosphorylées peuvent interagir avec des domaines protéiques notamment des domaines SH2 : la phosphorylation régule ainsi l’interaction entre les protéines.
Exemple de PP60src :
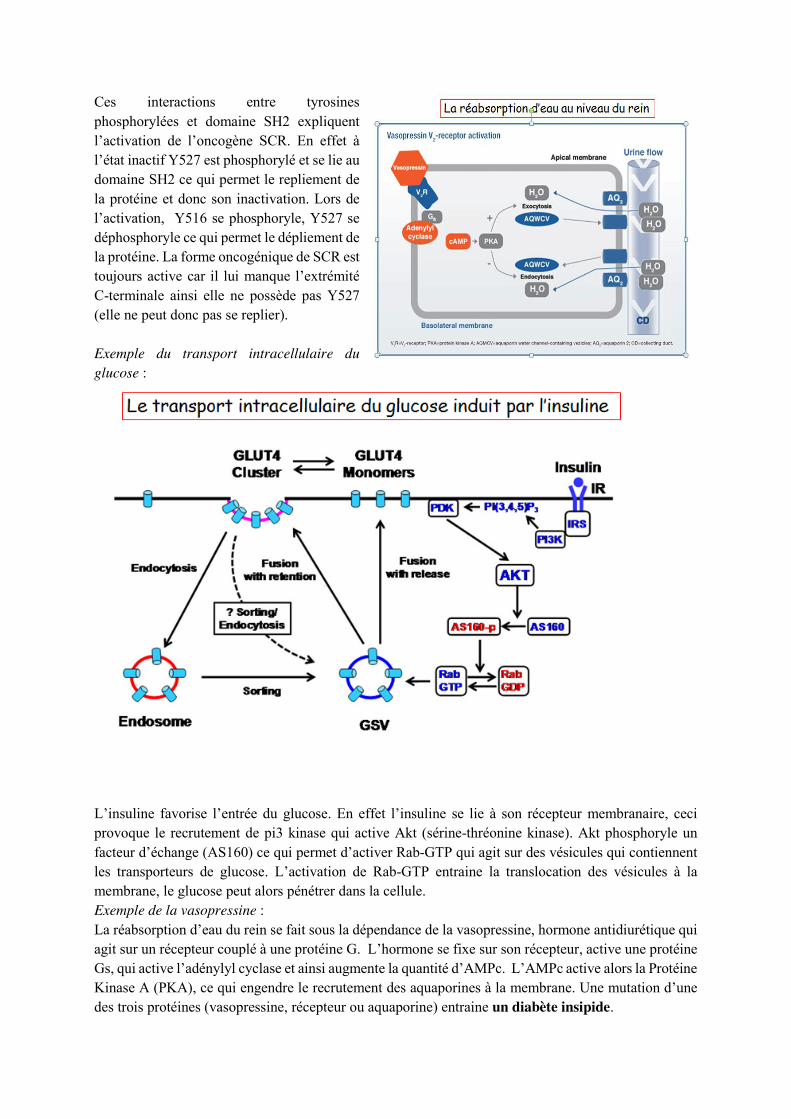
Ces interactions entre tyrosines phosphorylées et domaine SH2 expliquent l’activation de l’oncogène SCR. En effet à l’état inactif Y527 est phosphorylé et se lie au domaine SH2 ce qui permet le repliement de la protéine et donc son inactivation. Lors de l’activation, Y516 se phosphoryle, Y527 se déphosphoryle ce qui permet le dépliement de la protéine. La forme oncogénique de SCR est toujours active car il lui manque l’extrémité C-terminale ainsi elle ne possède pas Y527 (elle ne peut donc pas se replier). Exemple du transport intracellulaire du glucose :
L’insuline favorise l’entrée du glucose. En effet l’insuline se lie à son récepteur membranaire, ceci provoque le recrutement de pi3 kinase qui active Akt (sérine-thréonine kinase). Akt phosphoryle un facteur d’échange (AS160) ce qui permet d’activer Rab-GTP qui agit sur des vésicules qui contiennent les transporteurs de glucose. L’activation de Rab-GTP entraine la translocation des vésicules à la membrane, le glucose peut alors pénétrer dans la cellule. Exemple de la vasopressine : La réabsorption d’eau du rein se fait sous la dépendance de la vasopressine, hormone antidiurétique qui agit sur un récepteur couplé à une protéine G. L’hormone se fixe sur son récepteur, active une protéine Gs, qui active l’adénylyl cyclase et ainsi augmente la quantité d’AMPc. L’AMPc active alors la Protéine Kinase A (PKA), ce qui engendre le recrutement des aquaporines à la membrane. Une mutation d’une des trois protéines (vasopressine, récepteur ou aquaporine) entraine un diabète insipide.

2) Acétylation
La chromatine est l’enroulement d’ADN autour des histones, l’ensemble étant compacté en une fibre très dense. L’acétylation des histones entraine la décompaction de la chromatine ce qui favorise la transcription de l’ADN. Cette acétylation se fait sur des lysines par des HAT et la déacétylation par des AJDAC. Il existe également des méthylations, des monoubiquitinations et des phosphorylations des histones qui régulent la compaction de la chromatine.
C- Modification lors de la dégradation
L’ubiquitine est une petite protéine exprimée dans toutes les cellules de l’organisme, et elle possède beaucoup de lysines avec lesquelles elle se lie aux protéines. Il existe 3 formes d’ubiquitination : -la monoubiquitnation et la multiubiquitination (ajout d’ubiquitines indépendantes) : fonction dans la signalisation cellulaire comme l’internalisation cellulaire, la régulation des facteurs de transcription…
-la polyubiquitination : les ubiquitines forment une chaine et il en faut au moins 4 pour que la protéine soit dégradée.
Le système d’ubiquitination implique 3 enzymes : - E1 est une enzyme d’activation - E2 est une enzyme de conjugaison - E3 est une ubiquitine-ligase (lie la protéine à dégrader)
E1 en présence d’ATP lie l’ubiquitine et l’active, la transfère à E2 qui se lie à E3, et cela permet le transfert de l’ubiquitine sur la protéine à dégrader. Il existe deux classes essentielles d’E3 :
- HECT - Ring ligase :
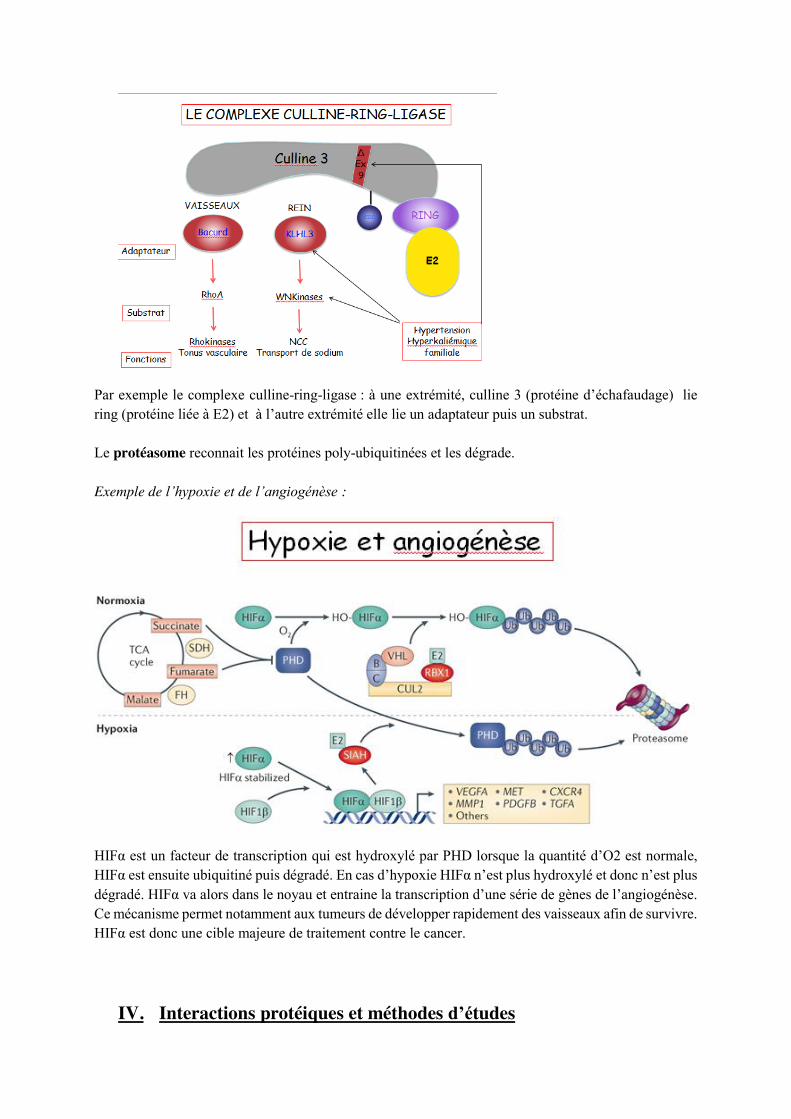
Par exemple le complexe culline-ring-ligase : à une extrémité, culline 3 (protéine d’échafaudage) lie ring (protéine liée à E2) et à l’autre extrémité elle lie un adaptateur puis un substrat. Le protéasome reconnait les protéines poly-ubiquitinées et les dégrade. Exemple de l’hypoxie et de l’angiogénèse :
HIFα est un facteur de transcription qui est hydroxylé par PHD lorsque la quantité d’O2 est normale, HIFα est ensuite ubiquitiné puis dégradé. En cas d’hypoxie HIFα n’est plus hydroxylé et donc n’est plus dégradé. HIFα va alors dans le noyau et entraine la transcription d’une série de gènes de l’angiogénèse. Ce mécanisme permet notamment aux tumeurs de développer rapidement des vaisseaux afin de survivre. HIFα est donc une cible majeure de traitement contre le cancer.
IV. Interactions protéiques et méthodes d’études
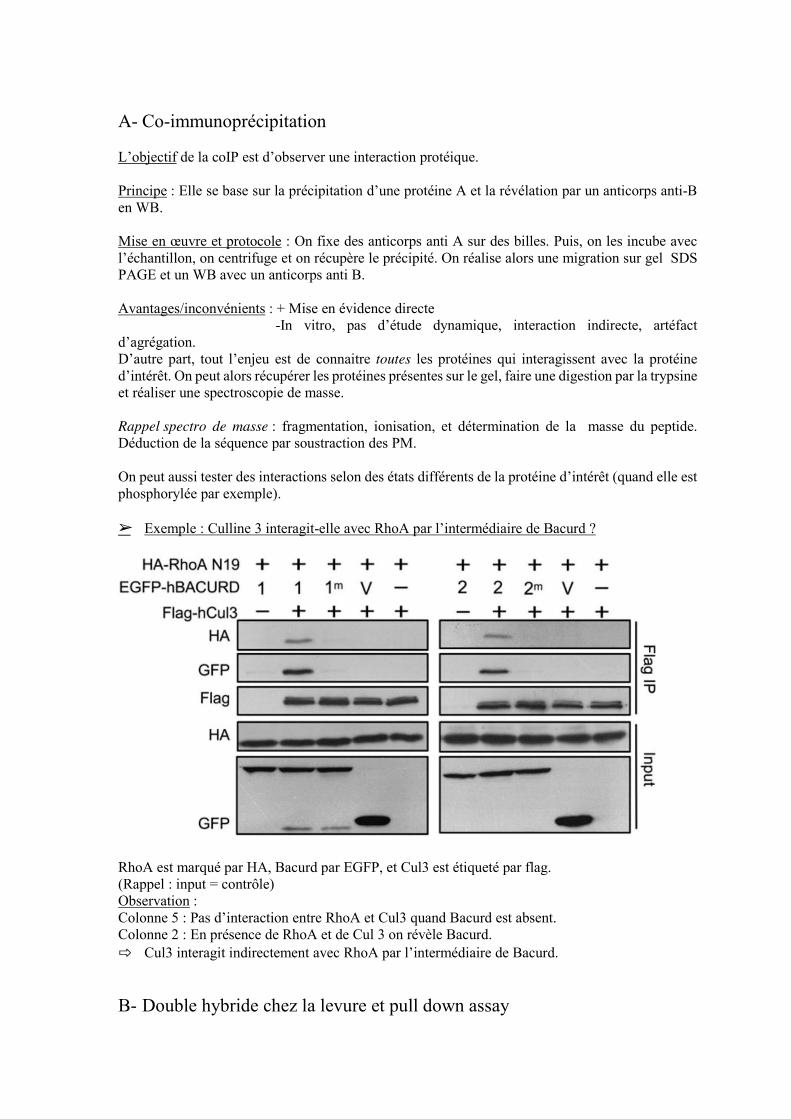
A- Co-immunoprécipitation L’objectif de la coIP est d’observer une interaction protéique. Principe : Elle se base sur la précipitation d’une protéine A et la révélation par un anticorps anti-B en WB. Mise en œuvre et protocole : On fixe des anticorps anti A sur des billes. Puis, on les incube avec l’échantillon, on centrifuge et on récupère le précipité. On réalise alors une migration sur gel SDS PAGE et un WB avec un anticorps anti B. Avantages/inconvénients : + Mise en évidence directe -In vitro, pas d’étude dynamique, interaction indirecte, artéfact d’agrégation. D’autre part, tout l’enjeu est de connaitre toutes les protéines qui interagissent avec la protéine d’intérêt. On peut alors récupérer les protéines présentes sur le gel, faire une digestion par la trypsine et réaliser une spectroscopie de masse. Rappel spectro de masse : fragmentation, ionisation, et détermination de la masse du peptide. Déduction de la séquence par soustraction des PM. On peut aussi tester des interactions selon des états différents de la protéine d’intérêt (quand elle est phosphorylée par exemple). ➢ Exemple : Culline 3 interagit-elle avec RhoA par l’intermédiaire de Bacurd ?
RhoA est marqué par HA, Bacurd par EGFP, et Cul3 est étiqueté par flag. (Rappel : input = contrôle) Observation : Colonne 5 : Pas d’interaction entre RhoA et Cul3 quand Bacurd est absent. Colonne 2 : En présence de RhoA et de Cul 3 on révèle Bacurd. ⇨ Cul3 interagit indirectement avec RhoA par l’intermédiaire de Bacurd.
B- Double hybride chez la levure et pull down assay
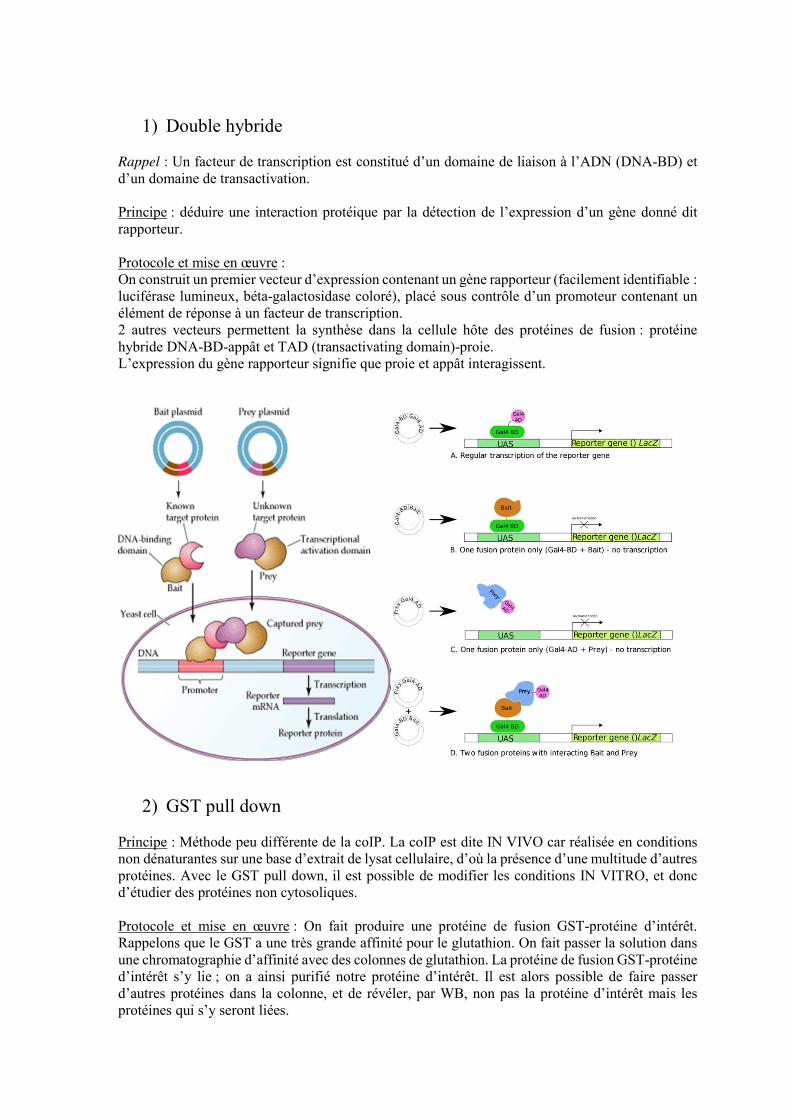
1) Double hybride
Rappel : Un facteur de transcription est constitué d’un domaine de liaison à l’ADN (DNA-BD) et d’un domaine de transactivation. Principe : déduire une interaction protéique par la détection de l’expression d’un gène donné dit rapporteur. Protocole et mise en œuvre : On construit un premier vecteur d’expression contenant un gène rapporteur (facilement identifiable : luciférase lumineux, béta-galactosidase coloré), placé sous contrôle d’un promoteur contenant un élément de réponse à un facteur de transcription. 2 autres vecteurs permettent la synthèse dans la cellule hôte des protéines de fusion : protéine hybride DNA-BD-appât et TAD (transactivating domain)-proie. L’expression du gène rapporteur signifie que proie et appât interagissent.
2) GST pull down Principe : Méthode peu différente de la coIP. La coIP est dite IN VIVO car réalisée en conditions non dénaturantes sur une base d’extrait de lysat cellulaire, d’où la présence d’une multitude d’autres protéines. Avec le GST pull down, il est possible de modifier les conditions IN VITRO, et donc d’étudier des protéines non cytosoliques. Protocole et mise en œuvre : On fait produire une protéine de fusion GST-protéine d’intérêt. Rappelons que le GST a une très grande affinité pour le glutathion. On fait passer la solution dans une chromatographie d’affinité avec des colonnes de glutathion. La protéine de fusion GST-protéine d’intérêt s’y lie ; on a ainsi purifié notre protéine d’intérêt. Il est alors possible de faire passer d’autres protéines dans la colonne, et de révéler, par WB, non pas la protéine d’intérêt mais les protéines qui s’y seront liées.

C- FRET-BRET 1) Le FRET (Fluorescent Resonant Energy Transfer) Principe : Transfert d’énergie non-radiatif entre un fluorophore excité (donneur) et un fluorophore dans son état fondamental (accepteur). Mise en œuvre : On exprime dans une cellule une protéine A marquée par une « étiquette » fluorescente donneuse et une protéine B marquée par un fluorophore accepteur. Si on excite le premier fluorphore à sa longueur d’onde propre d’excitation, il va réémettre de l’énergie à une autre longueur d’onde propre. Si les deux protéines qu’on cherche à tester son assez proches l’une de l’autre, alors l’énergie émise par le premier fluorophore va exciter le second fluorophore, qui va, à son tour, émettre de l’énergie à sa propre longueur d’onde d’émission. On va donc pouvoir détecter l’énergie de ce 2nd fluorophore alors qu’il n’a pas été excité par le manipulateur. Ce transfert d’énergie dépend uniquement de la distance et de la position entre les 2 protéines d’intéret. Distance pour laquelle on a un signal de FRET : 2 à 10 nm. Il s’agit donc d’interactions à l’échelle moléculaire. Avantage/Inconvénients : + dynamique de l’interaction analysable, couplage à la microscopie possible, in vivo. - Les spectres d’émission et d’excitation sont relativement larges, ce qui peut rendre la lecture un peu compliquée (les spectres se chevauchant un petit peu, le fluorophore accepteur peut être excité par la longueur d’onde d’excitation du fluorophore donneur -> bruit de fond). De nombreux progrès ont été réalisés et aujourd’hui on dénombre 75 protéines fluorescentes dans toutes les gammes de couleur.
2) FRET vs BRET La différence entre ces deux techniques réside dans le fluorophore donneur. Pour le FRET, il s’agit généralement de la GFP ou plus globalement d’une protéine qui a besoin d’être excitée pour fluorescer. Pour le BRET, on utilise souvent la luciférase, protéine autofluorescente (luminophore)

. FRET : forte sensibilité vidéo mais l’intensité du signal de l’accepteur est polluée par la fluorescence du donneur, mais surtout par la longueur d’onde d’excitation. Il y a un bruit élevé et le signal est imprécis. . BRET : grande spécificité du signal car il n’y a pas d’excitation du donneur nécessaire. Toutefois, la sensibilité est faible et l’observation en microscopie est difficile et ne peut se faire que sur de petits extraits cellulaires.
FRET BRET
sensibilité + -
spécificité - + ➢ Exemple : PKA et acrodysostose Cette maladie se caractérise par une résistance multi-hormonale. Elle s’explique par une mutation dans une des sous unités régulatrices de la PKA (voir l’exemple de la signalisation par les 2nds messagers et l’AMPc) On réalise une expérience de BRET pour observer les conséquences sur la dissociation de la PKA suite à la liaison de l’AMPc. On réalise une courbe dose-réponse.
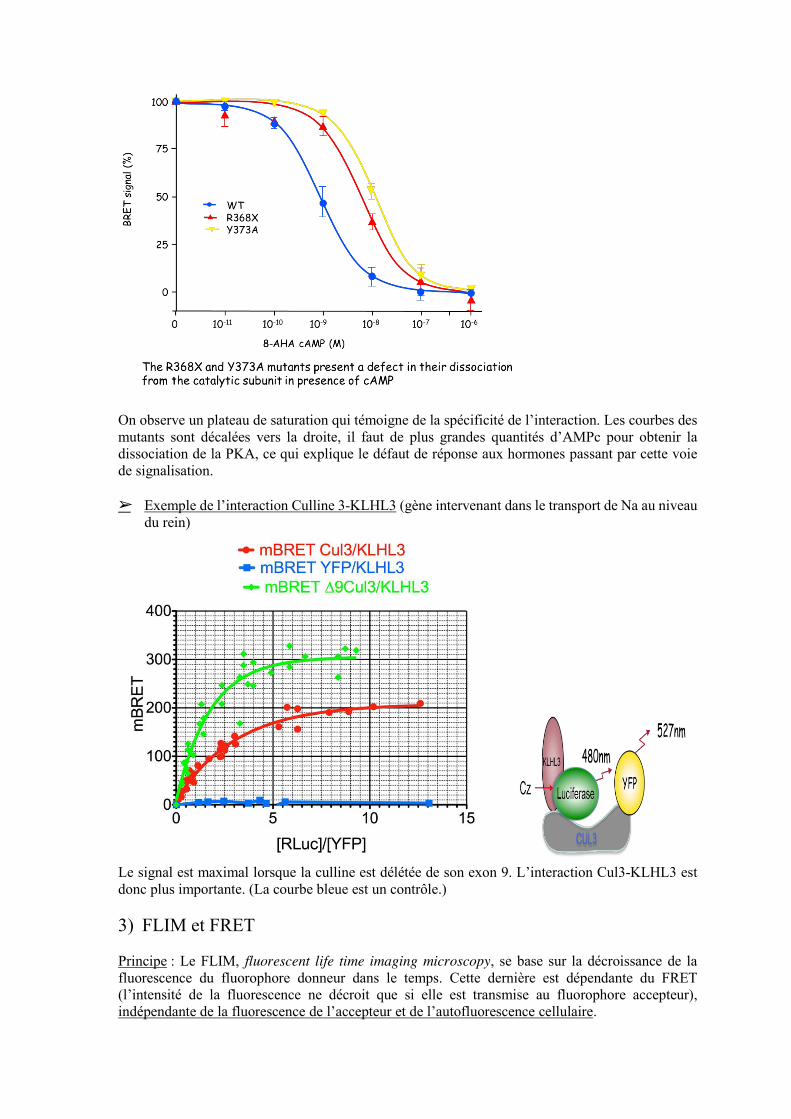
On observe un plateau de saturation qui témoigne de la spécificité de l’interaction. Les courbes des mutants sont décalées vers la droite, il faut de plus grandes quantités d’AMPc pour obtenir la dissociation de la PKA, ce qui explique le défaut de réponse aux hormones passant par cette voie de signalisation. ➢ Exemple de l’interaction Culline 3-KLHL3 (gène intervenant dans le transport de Na au niveau
du rein)
Le signal est maximal lorsque la culline est délétée de son exon 9. L’interaction Cul3-KLHL3 est donc plus importante. (La courbe bleue est un contrôle.)
3) FLIM et FRET
Principe : Le FLIM, fluorescent life time imaging microscopy, se base sur la décroissance de la fluorescence du fluorophore donneur dans le temps. Cette dernière est dépendante du FRET (l’intensité de la fluorescence ne décroit que si elle est transmise au fluorophore accepteur), indépendante de la fluorescence de l’accepteur et de l’autofluorescence cellulaire.

D- Intégration du signal La cellule contient des intégrateurs moléculaires qui orientent la réponse physiologique. D’une façon générale, on ne peut pas dire qu’un facteur A va donner tel ou tel effet sur une cellule, cela dépend de l’intégration et de l’association des différents signaux mis en jeu. L’intégration du signal dépend aussi du type cellulaire concerné.
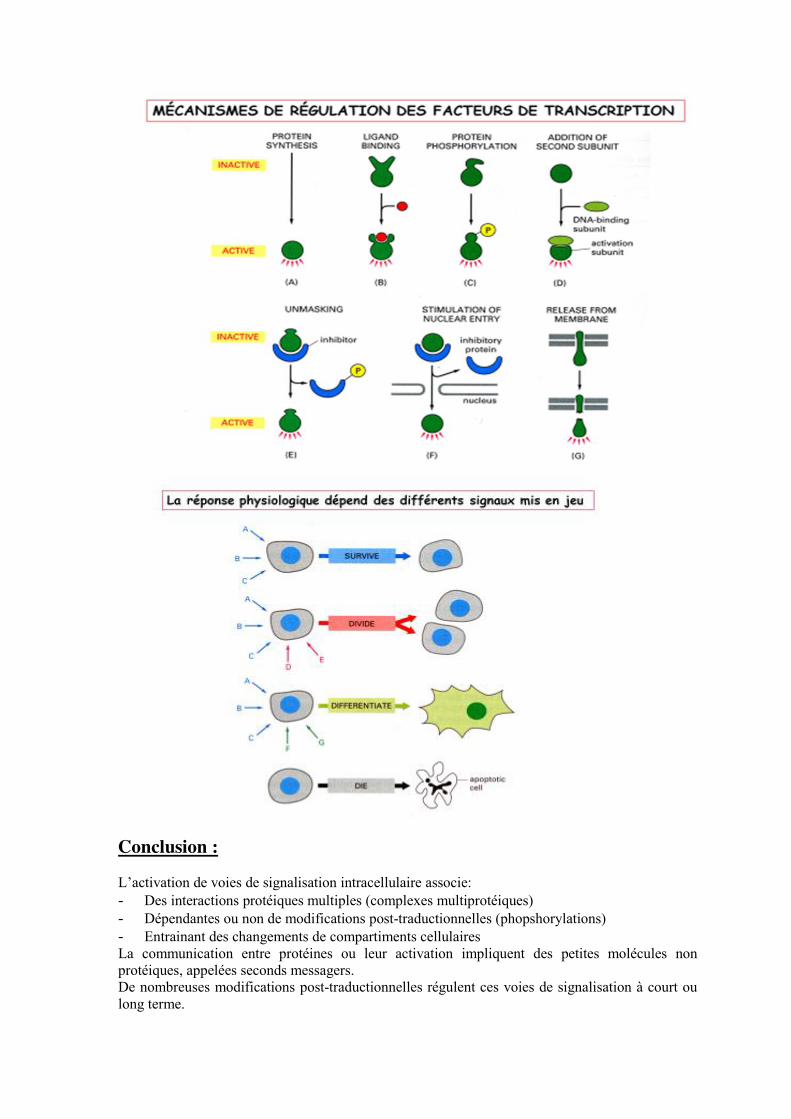
Conclusion : L’activation de voies de signalisation intracellulaire associe: - Des interactions protéiques multiples (complexes multiprotéiques) - Dépendantes ou non de modifications post-traductionnelles (phopshorylations) - Entrainant des changements de compartiments cellulaires La communication entre protéines ou leur activation impliquent des petites molécules non protéiques, appelées seconds messagers. De nombreuses modifications post-traductionnelles régulent ces voies de signalisation à court ou long terme.

FICHE RECAPITULATIVE La signalisation cellulaire repose sur la transmission de l’information d’une cellule à l’autre, par voies - endocrine → (système endocrine) - paracrine → (système nerveux) - autocrine + (système immunitaire) Les médiateurs peuvent être - hydrophiles → récepteurs intracellulaires, régulent la transcription de l’ADN - hydrophobes → récepteurs membranaires (récepteurs canaux, récepteurs enzymes, RCPG) Le signal est détecté grâce à - l’association à un second messager - la formation d’un complexe multi-protéique (protéines d’échafaudage) - des modifications post-traductionnelles L’intégration du signal se fait par une multitude de modifications. Les modifications post-traductionnelles lors de la biosynthèse sont stables et définitives : - La N-glycosylation est l’ajout d’un sucre sur une asparagine (Asn-X-Ser ou Thr ou Cys). Rôle dans la stabilité de la protéine et dans son adhésion à la membrane. - La O-glycosylation est l’ajout d’un sucre sur une sérine ou une thréonine. Rôle dans le contrôle de la phosphorylation (compétition des sucres et des groupements phosphates). - L’acylation est l’ajout d’un acide gras. Rôle dans la localisation membranaire de la protéine. Les modifications post-traductionnelles labiles sont réversibles. Parmi celles-ci : - la phosphorylation, par les sérines-thréonines kinases (99.9%) et les tyrosines kinases (0.01%) - l’acétylation, des histones notamment, qui favorise la transcription de l’ADN. Les modifications post-traductionnelles existent jusque dans le processus de dégradation, par l’ubiquitination des protéines et par l’action du protéasome. Elle se fait grâce à trois enzymes : - E1, enzymes activatrice - E2 enzymes de conjugaison - E3 enzymes ubiquitine-ligase, de deux classes, HECT et culline-ring-ligase Une protéine peut être ubiquitinée sous trois formes : mono- multi- poly- ubiquitinée. Méthodes d’études des interactions protéiques : 1) La co-immunoprécipitation : précipitation de la protéine A, puis révélation de la protéine B. Inconvénients majeurs : pas d’étude dynamique et contraintes de l’in vivo 2) La technique du double-hybride : révélation de l’interaction protéique par la transcription d’un gêne rapporteur. Elle n’est possible que si le DNA-BD et le domaine de transactivation du FT sont proches. Inconvénient majeur : n’étudie que les protéines cytosoliques. 3) Le GST pull down : étude IN VITRO. La purification de la protéine A se fait par chromatographie d’affinité GST-glutathion. Puis passage de la protéine B dans la colonne. Etude par WB. 4) Le FRET : excitation du fluorophore donneur, dont l’émission excitera le fluorophore accepteur seulement s’il est assez proche. Inconvénient majeur : faible spécificité, bruit de fond 5) Le BRET : excitation autonome d’un luminophore (la luciférase), dont l’émission excitera le fluorophore accepteur, seulement s’il est assez proche. Inconvénient majeur : moins sensible que le FRET 6) Le FLIM : étude de la vitesse de décroissance de l’intensité du signal émis par le fluorophore donneur.





























