Embed Size (px)
Citation preview

Science & Sports, 1 (1986) 1-23 1 �9 Elsevier, Paris
Lactat6mie et exercice musculaire. Signification et analyse critique du concept de ((seuil a&obie-ana6robie
M. RIEU
Laboratoire de Physiologie des Adaptations, UER Cochin, Port Royal, 24, rue du Faubourg-Saint-Jacques, Paris 75014, France
(Refu le 21-1-1986; acceptd le 10-2-1986)
R6sum6 - Les relations existant entre l'intensit6 de la contraction musculaire et la concentration sanguine du lactate ont, depuis le d6but du si~cle, abondamment ~veill6 la curiosit6 des physiologistes.
L'616vation de la lactat6mie pendant l'exercice a ~t6 interpr6t6e comme le reflet de la mise en jeu du m6tabolisme ana&obie qui surviendrait m~me pour des efforts sous-maximaux. Cette accentuation de la glycolyse entraTnerait alors un 6puisement rapide des r6serves en glycog6ne et un arr~t de l'effort. Le niveau de puissance de l'exercice ~t partir duquel interviendraient ces changements m6taboliques est consid6r6 comme un ~ seuil ~ qui conditionne lest< capacit6s d'endurance~ d 'un sujet. La d6termination d 'un tel t< scull ~ a fait l 'objet des pr6occupations de nombreux chercheurs. Comme l'accumulation de lactate dans le sang 6tait pr6sum6e ~tre contemporaine du d6veloppement d'une acidose m6tabolique compens6e sur le plan respiratoire par une hyperventilation relative avec augmentation du rapport VE/VO 2 et rejet accru de CO 2, l'apparition de ces 6v6nements lors de la progression de l'intensit6 de l'effort fut consid~r6e comme refl6tant le d6but de l'intervention du m6tabolisme ana6robie, et la puissance correspondante de l'exercice fut d6finie comme le <~seuil ana6robie~. Plusieurs tests ont ainsi ~t6 d6crits, qui.comprennent une s6quence d'exercices de dur6e 1-4 min d'intensit6 progressivement &oissante et utilisant comme crit~res, soit l'616vation du rapport VE/~rO2 (seuil ventilatoire = S(~/E), soit celle de la lactat6mie au-dessus de sa valeur de repos (seuil lactate = S[LAls). Quel- ques 6tudes montrent une bonne correlation entre les r6sultats de ces tests et les performances r6alis6es lots des courses de fond. Mais d'autres travaux mettent en dvidence des divergences entre les rdsultats des <<seuils ana6robie~ selon qu'ils sont obtenus par la m6thode ventilatoire ou celle du d6but d'augmentation de la lactat6mie. Des questions de fond sont alors pos6es concernant les diverses origines possibles de l 'augmentation de "(/E rapport6e :~ 7 0 2 et les rela- tions existant entre VE et VCO 2 ou encore concernant les variations de la lactat6mie en fonction du lieu de pr61~ve- ment sanguin ou de l'6tat acido-basique de I'organisme.
Une autre approche tente d'objectiver l'intensit6 de l'exercice h laquelle correspondrait une lactat6mie maximale d'6tat stable. Certains auteurs utilisent des tests d'effort par pallet de 3 - 5 m i n e t un niveau fixe de la lactat6mie 6gal b. 4 m m o l . l - ~ pour cette d6termination. L~ encore, les r6sultats sont contradictoires tant dans le domaine de la pr6dic- tion de la performance que dans celui du contr61e physiologique de l'entra~nement.
L'analyse critique du concept de seuil ana6robie r6v~le quelques probl~mes: - d'ordre m6thodologique en soulevant le fair que la mesure de la lactat6mie b. la fin de l'effort ne rend pas compte
de la cin6tique de ce produit du m6tabolisme, surtout lors d'6preuves d'efforts successifs de courte dur6e, qui cumu- lent leurs effets respectifs;
- d'ordre plus fondamental concernant: les rapports entre lactat6mie et ana&obiose qui ne sont pas n6cessairement aussi 6troits qu 'on pouvait ant&ieurement le penser; la r6partition du lactate entre la celluIe musculaire et le milieu extracelIulaire; la signification m~:me de la lactat6mie qui ne repr6sente qu 'un 6quilibre instantan6 entre le d~bit d'apparition et le d6bit de disparition du lactate dans le sang.
En conclusion, trop d'incertitudes subsistent encore pour pouvoir accorder au concept de <<seuil a&obie-ana6robie ~ une signification physiologique certaine.
l ac ta l6mie I exercice / <<seuil a~robie-ana6robie>>

2 M. Rieu
Summary - Lacticemia and muscular exercise. The concept of 'aerobic-anaerobic threshold'. The relationship between the intensity o f muscular contraction and blood lactate concentration has been o f great interest to physiologists.
Increase o f lacticemia upon exercise is interpreted as reflecting anaerobic metabolism, even in the case o f submaxi- real exercise. Such a rise in glycolysis would #tduce a rapid depletion o f glycogen stores and the end o f exercise. These metabolic changes wouM be produced at a certain level o f exercise power considered as a "threshoM" determining the subject "s endurance capacity. AIany scientists have been concerned with the determination o f this "threshold'. Accu- mulation o f lactate in the blood is thought to occur sinudtaneously with metabolic acidosis compensated by relative hyperventilation, with an increase in the ratio I?E/I?O 2 and an increase in CO 2 rejection; therefore, the occurrence o f these phenomena in parallel with an increase in exercise intensity is believed to reflect the onset o f anaerobic meta- bolism, and the corresponding exercise power is defined as 'aerobic threshoM'.
Several tests are described, inchtding a series o f 1 - 4 rain exercises o f increasing intensity in which the increase in the ratio I;'E/ I;'O 2 (ventilatory threshoM = S(I?E)) or the increase in lacticemia above its vahte at rest (lactate thres- hoM = S[LA]s) were used as criteria. Some studies show a good correlation between the results and performance dltring long-distance races. However, other studies have demonstrated differences in vahtes obtained fo r "anaerobic thres- holds" according to the methodology used: by the ventilatory method or f rom onset o f increase in lacticemia. Funda- mental questions then arise concerning the various possible orighTs o f the hwrease hi (ZE/l?02, the relationship between I;'E attd I)CO 2, variations in lacticemia depending on the location o f blood sampling, attd the acid-base con- dition o f the organisnt.
Another approach attempts to determine the level o f exercise that corresponds to a stable level o f maximal lactice- mia. Some attthors use tests with exercises comprising 3 - 5 rain steps and a defined level o f lacticemia o f 4 nunol,1 -I. l tere again, the results do not correlate, either at the level o f predicthlg performance, or at that o f physiological trainhzg.
Critical analysis o f "anaerobic threshoM" raises the following problems: - the question o f methodology, shwe the measurenlent o f lacticemia at the end o f the exercise does not take into
account the kinetics.of this metabolic product, particularly ht tests with successive exercises o f short duration, in which the effects are cunndative;
- furtherfuttdamental questions concerning: the relationship between lacticemia and anaerobiosis, which is not neces- sarily as close as was previously thought; lactate distribution between muscle cells and the extracelhdar medium; the significance o f lacticemia which only represents an htstantaneous balance between the appearance and disappea- rance o f lactate in the blood. Too many t/ncertahtties remain to be able to establish the true physiological significance o f the concept o f "aerobic-
anaerobic threshold'.
lactlcemla / exercise I 'aerobic-anaerobic threshold'
I n t r o d u c t i o n
Depuis que l 'on conna~t l'existence de liens 6troits entre la contraction musculaire et la production d'acide lactique, on a tent6, dans le domaine de la physiologic appliqu6e, d'accorder aux variations de la lactatdmie apparaissant pendant et au d~cours d 'un exercice musculaire, une signification comme tfmoin de l'intervention du mftabolisme ana6robie.
Partant de ces pr~mices, de nombreux tests d 'effor t ont 6t6 mis au point tout au long de ces vingt derni6res anndes. Ils utilisent soit la mesure directe du lactate dans le sang circulant, soit I'enre- gistrement des r6actions compensatrices de l 'orga- nisme ~ l'acidose engendr6e par la production d'acide lactique. Ces tests sont cens6s objectiver la puissance de l'exercice ~ partir de laquelle les besoins 6nerg6tiques n6cessaires ne peuvent plus ~tre
couverts exclusivement par le m6tabolisme a6robie. Ainsi, des protocoles vari6s d'6preuves d'effort ont- ils 6t6 d6crits, dont les m6thodologies ne sont pas toujours sans susciter quelques r6serves darts leurs principes. Par suite, les r6sultats divergents abon- dent dans la litt6rature.
Carte situation pose en outre des questions de fond: de quels facteurs d6pend tr6s exactement la concentration du lactate dans le sang? La produc- tion d'acide lactique par le muscle en activit6 est- elle effectivement synonyme d'ana6robiose?
Ces diff6rents aspects sont pass6s en revue dans les pages suivantes qui tentent, sinon d'apporter une r6ponse d6finitive, tout au moins d'6clairer un d6bat largement nourri au cours des trois ann6es pr6c6dentes par des 6crits scientifiques aussi vigou- reux que contradictoires.

Lactatdmie et exercice musculaire 3
Z o n e de transition a6robie-ana6robie
Historique et probldmatique
A partir des travaux de Claude Bernard et de Pas- teur mettant en 6vidence la transformation du glu- cose en acide lactique et ceux de Du Bois Raymond en 1858 montrant que la contraction musculaire produisait de l'acide lactique, la curiosit6 des physiologistes s'est longuement exerc6e ~ propos de ce produit de la d6gradation du glycog~ne. Sa pr6- sence fur d 'abord consid6r6e comme imm6diate- ment n6cessaire au fonctionnement m6canique du muscle que l 'on d&omposait en une premiere phase t<de contraction>>, survenant en ana6robiose et l 'origine d 'un d6gagement << initial >> de chaleur, et en une deuxi~me phase << de restauration>> a6robie et contemporaine de l 'apparition de la chaleur << retard6e >> (Hill, 1911-1914). A l 'appui de cette id6e venaient les constatations faites par Fletcher et Hopkins (1906) d6montrant que le muscle isol6 pouvait se contracter en absence d'oxyg~ne, pro- duisant alors d 'autant plus d'acide lactique que le travail fourni 6tait plus grand et ceci jusqu'~ une certaine limite h partir de laquelle l'6puisement sur- venait. S ice muscle 6tait plac6 en pr6sence d 'oxy- g~ne, l 'acide lactique form6 disparaissait, cette 61imination 6tant assur6e par sa combustion ou sa transformation en glucose (Meyerhof, 1921). Cette th6orie fut bientft mise en doute (Embden, 1927), l 'acide lactique apparaissant seulement comme un substrat ~nerg~tique. Ult6rieurement, il se r6v~la qu 'un muscle intoxiqu6 par de l 'acide monoiodoa- c6tique et done incapable de former de l 'acide lac- tique conservait n6anmoins la facult6 de se contracter (Lundsgaard, 1931).
En m~me temps, l 'observation que plus l ' inten- sit6 de l'exercice 6tait 61ev6e, plus la concentration du lactate dans le sang 6tait importante et plus la consommation d'oxyg~ne pendant et apr6s l 'effort 6tait grande, conduisit ~ la notion de << dette d 'oxy- g6ne>> (Hill, 1924; Margaria, 1933). Ainsi s 'affirma l'id6e que l'insuffisance d 'apport en oxyg6ne aux muscles en activit6 6tait responsable de l 'accumu- lation du lactate dans le sang (Douglas, 1927). Un peu plus tard, compl6tant ces premi6res donn6es, distinction fut faite entre des exercices de faible intensit6, r6alis6s en a6robiose stricte, ne s 'accom- pagnant pas d 'une augmentation de la lactat6mie et des efforts, toujours sous-maxlmaux mais de puissance plus 61ev6e au cours desquels la concen- tration sanguine du lactate s'accrott (Owles, 1930). Au cours des ann6es 30, d'autres observations con- firm6rent que la concentration sanguine du lactate pouvait augmenter m~me lorsqu'un exercice 6tait
effectu6 avec un niveau de puissance inf6rieur 'r 2 max (Margaria, 1933), mais que, par contre, un effort mod6r6 pouvait r6duire une lactat6mie pr6alablement 61ev6e (Bang, 1936). Enfin, le fait que, pour un exercice d 'une puissance donn6e, le lactate sanguin monte moins chez les sujets entraT- n6s que chez les s6dentaires, 6veillait d6jh l 'atten- tion des chercheurs (Robinson, 1941; Crescitelli, 1944).
Parail~lement, l '6tude des facteurs limitant la performance commenqait ~ ~tre activement entre- prise et la capacit6 de l 'organisme ~ neutraliser les ions H + provenant de l 'acide lactique sembla conditionner le ~t niveau d 'endurance >> des sujets l'exercice (Herbst, 1929).
La d6termination de ce <<niveau d'endurance>> devint alors une pr6occupation ~ laquelle de nom- breux physiologistes pr~t~rent attention tant dans le domaine du travail (Miiller, 1943) que dans celui de l'exercice musculaire.
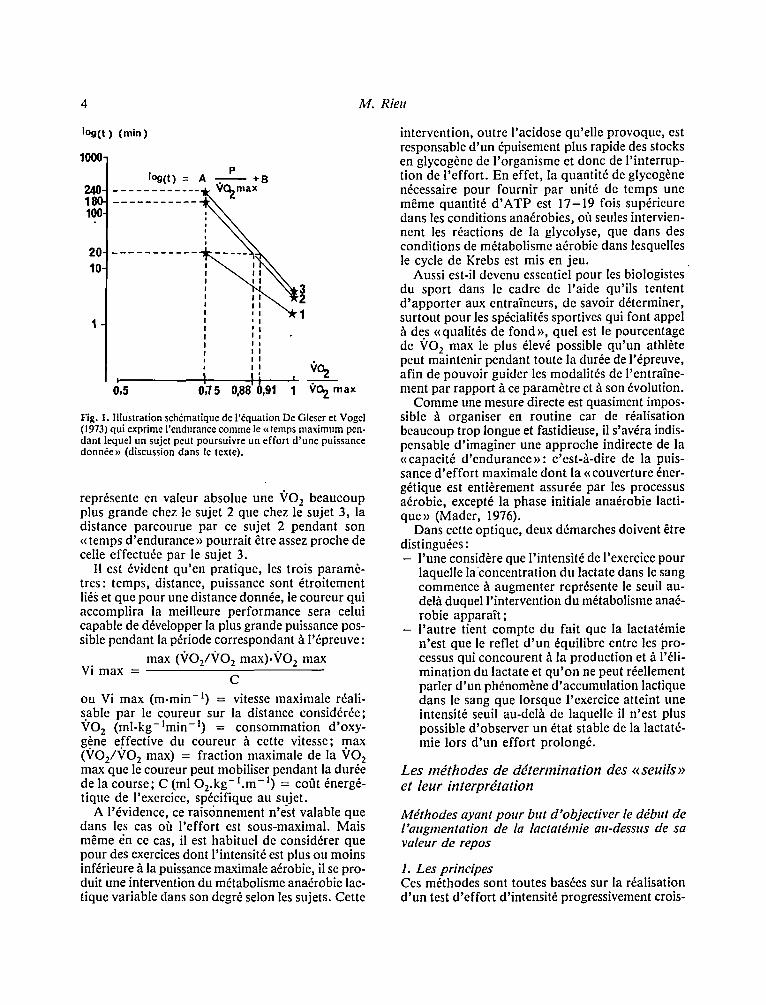
Cette notion a 6t6 largement approfondie depuis ces premiers travaux, l 'endurance pouvant ~tre d6fi- nie comme: << Le temps maximal pendant lequel un sujet est capable de r6aliser un effort d 'une inten- sit6 donn6e.>> La relation temps-puissance de l'exercice rev~t alors une forme exponentielle qui peut s'exprimer par l '6quation suivante (Gleser et Vogel, 1973):
P log(t) = A'~ 'O z max + B
o~ t = la dur6e de l 'effor t , P = la puissance de cet effort .
Les param&res A e t B pouvant ~tre consid6r6s comme l'expression de la << capacit6 d 'endurance >> qui caract6rise un individu. Si P e s t exprim6 en terme de ~rO2~ P /VO 2 max repr6sente la fraction maximale de VO 2 max que, par exemple, un cou- reur peut utiliser pour courir pendant un temps d6fini.
Ainsi, si l 'on examine la Figure I, on note que pour courir pendant 20 min, le sujet 1 pourra au mieux utiliser 7507o de sa VO 2 max alors que les sujets 2 et 3 pourront respectivement mobiliser 8807o et 9107o de leur VO 2 max. A l'inverse, si tous les sujets courent h une allure correspondant pour cha- cun d 'entre eux ~ l'utilisation de 7507o de leur ~'O 2 max, les sujets l , 2, 3 seront respectivement obli- g6s d ' interrompre leur effort au bout de 20 min, 180 min, 240 min. Si leur 7 0 2 max et leur cofit 6nerg6tique sont identiques, leur vitesse sera sem- blable et les distances parcourues seront e.n accord avec cette hi6rarchie. Par contre, si les VO 2 max diff6rent, la x~itesse de course sera variable selon les sujets: ainsi dans l 'hypoth6se o/1 7507o ~'O 2 max
4 M. Riett
Iog(t ) (min)
1000'
240- 180" 100-
20- 10-
P log(t) = A ~ . +B
. . . . . . . . . . .
! I ! ,, ! !
, ,
0,5 0,75 0,88 0,91 1
a V O ~ m a x
Fig. 1. Illustration sch6matique de l'~quation De Gleser et Vogel (1973) qui exprime I'endurance comme le t< temps maximum pen- dant lequel un sujet peut poursuivre un effort d'une puissance donn6e>~ (discussion dans le texte).
repr6sente en valeur absolue une ~/O 2 beaucoup plus grande chez le sujet 2 que chez le sujet 3, la distance parcourue par ce sujet 2 pendant son ~< temps d'endurance >~ pourrait &re assez proche de celle effectu6e par le sujet 3.
I1 est 6vident qu'en pratique, les trois param6- trcs: temps, distance, puissance sont 6troitement li6s et que pour une distance donn6e, le coureur qui accomplira la meilleure performance sera celui capable de d6velopper la plus grande puissance pos- sible pendant la p6riode correspondant h l'6preuve:
max (~/O2/VO 2 max).VO z max Vi max =
C
ou Vi max (m.min- l ) = vitesse maximale r6ali- sable par le coureur sur la distance consid6r6e; ~rO 2 (ml .kg - lmin - l ) = consommation d 'oxy- g.~ne effective du coureur fi cette vitesse; max (VO2/~"O 2 max) = fraction maximale de la ~/O 2 max que le coureur peut mobiliser pendant la dur6e de la course; C (ml O2 .kg - l .m- i ) = cofit 6nerg6- tique de l'exercice, sp6cifique au sujet.
A l'6vidence, ce raisonnement n'est valable que dans les cas off l 'cffort est sous-maximal. Mais m~me ~n ce cas, il est habituel de consid6rer que pour des exercices dont l'intensit6 est plus ou moins inf6rieure h la puissance maximale a6robie, il se pro- duit une intervention du m6tabolisme ana6robie lac- tique variable dans son degr6 seIon les sujets. Cette
intervention, outre I'acidose qu'elle provoque, est responsable d 'un 6puisement plus rapide des stocks en glycog6ne de l 'organisme et donc de l ' interrup- tion de l 'effort. En effet, la quantit6 de glycog~ne n6cessaire pour fournir par unit6 de temps une mfime quantit6 d 'A TP est 17-19 fois supfrieure dans les conditions ana6robies, off seules intervien- nent les r6actions de la glycolyse, que dans des conditions de m6tabolisme a6robie dans lesquelles le cycle de Krebs est mis en jeu.
Aussi est-il devenu essentiel pour les biologistes du sport dans le cadre de l'aide qu'ils tentent d 'apporter aux entra~neurs, de savoir d6terminer, surtout pour les sp6cialitfs sportives qui font appel h des <<qualit6s de fond>), quel est le pourcentage de ~tO 2 max le plus 6lev6 possible qu 'un athlete peut maintenir pendant toute la dur6e de l'6preuve, afin de pouvoir guider les modalit6s de I'entra~ne- ment par rapport fi ce param6tre et fi son ~volution.
Comme une mesure directe est quasiment impos- sible :~ organiser en routine car de r6alisation beaucoup trop longue et fastidieuse, il s'av6ra indis- pensable d'imaginer une approche indirecte de la <<capacit6 d 'endurance~: c'est-fi-dire de la puis- sance d 'effort maximale dont la ~ couverture 6ner- g6tique est enti6rement assur6e par les processus a6robie, except6 la phase initiale ana6robie lacti- q u e , (Mader, 1976).
Dans cette optique, deux d6marches doivent ~tre distingu6es : - l 'une consid~re que l'intensit6 de l'exercice pour
laquelle la concentration du lactate dans le sang commence fi augmenter repr6sente le seuil au- del~ duquel l'intervention du m6tabolisme ana6- robie apparaSt ;
- l 'autre tient compte du fait que la lactat6mie n'est que le reflet d 'un 6quilibre entre les pro- cessus qui concourent :~ la production et :~ l'61i- mination du lactate et qu 'on ne peut r6ellement parler d 'un ph6nom~ne d'accumulation lactique dans le sang que lorsque l'exercice atteint une intensit6 seuil au-delfi de laqueile il n'est plus possible d'observer un 6tat stable de la lactat6- mie lors d 'un effort prolong6.
Les mdthodes de ddtermination des <<seuils)~ et leur interprdtation
Mdthodes ayant pour but d'objectiver le ddbut de l'augmentation de la lactatdmie att-dessus de sa valeur de repos
1. Les principes Ces m6thodes sont toutes bas6es sur la r6alisation d 'un test d 'effort d'intensit6 progressivement crois-
Lactatdmie et exercice musculaire 5
sante menant :~ l'6puisement. Deux protocoles de recueil de donnEes sont couramment utilisfs pour dfterminer le scull d'intensit6 de l 'effort/~ partir duquel le m6tabolisme ana6robie est pr6sum6 inter- venir: l 'un correspond h une analyse directe par dosage du lactate sanguin, nous l 'appellerons le seuil lactate (S[LA]s); I'autre, plus indirecte, pro- c~de b. 1'6tude des 6changes gazeux respiratoires, nous l'appellerons le seuil ventilatoire (S(VE)).
Nous allons darts ce dernier chapitre porter plus particuli~rement notre attention sur ce dernier, compte tenu de l'int6r~t que pr6sentent ses inter- pr6tations physiologiques.
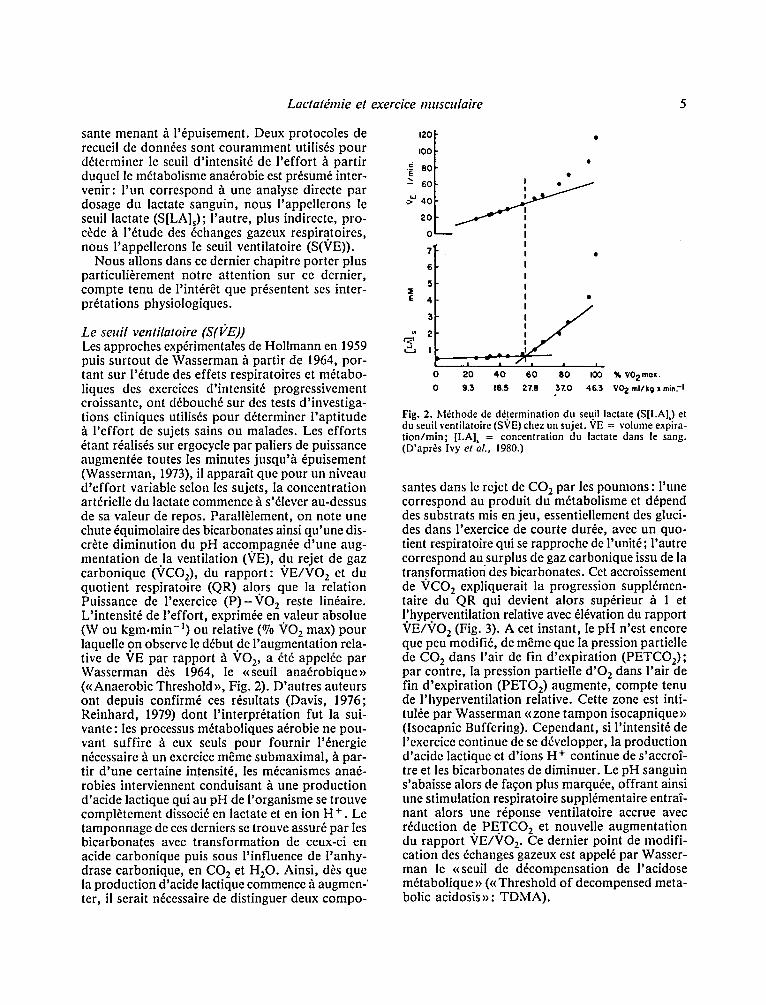
Le seuil ventilatoire (S(I;'E)) Les approches expfrimentales de Hollmann en 1959 puis surtout de Wasserman h partir de 1964, por- tant sur l '6tude des effets respiratoires ct m6tabo- liques des exercices d'intensit6 progressivement croissante, ont d6bouch6 sur des tests d'investiga- tions cliniques utilis6s pour d&erminer l 'aptitude :~ l 'effort de sujets sains ou malades. Les efforts 6tant r6alis6s sur ergocycle par paliers de puissance augment6e toutes les minutes jusqu'.h ~puisement (Wasserman, 1973), il apparaTt que pour un niveau d 'ef for t variable selon les sujets, la concentration artfrielle du lactate commence h s'61ever au-dessus de sa valeur de repos. Parall~lement, on note une chute 6quimolaire des bicarbonates ainsi qu'une dis- cr~te diminution du pH accompagn6e d 'une aug- mentation de la ventilation (VE), du rejet de gaz carbonique (VCO2), du rapport : VE/VO 2 et du quotient respiratoire (QR) alors que la relation Puissance de l'exercice ( P ) - V O 2 reste lin6aire. L'intensit6 de l 'effort, exprim~e en valeur absolue (W ou kgm.min- l) ou relative (~ ~'O 2 max) pour laquelle 9 n observe le d6but de l 'augmentation rela- tive de VE par rapport b. VO 2, a 6t6 appelfe par Wasserman d6s 1964, le r ana6robique , (r Threshold>>, Fig. 2). D'autres auteurs ont depuis confirms ces r6sultats (Davis, 1976; Reinhard, 1979) dont l ' interpr&ation fut la sui- vante: les processus m6taboliques a6robie ne pou- vant suffire ~ eux seuls pour fournir l'6nergie nfcessaire :~ un exercice m~me submaximal, h par- tir d 'une certaine intensit6, les m6canismes anaf- robies interviennent conduisant h une production d'acide lactique qui au pH de l'organisme se trouve compl~tement dissoci6 en lactate et en ion H +. Le tamponnage de ces derniers se trouve assur6 par les bicarbonates avec transformation de ceux-ci en acide carbonique puis sous l 'influence de l 'anhy- drase carbonique, en CO 2 et H20. Ainsi, d6s que la production d'acide lactique commence b. augmen-' ter, il serait n6cessaire de distinguer deux compo-
IZO
I 00
�9 =- so E
- 60
.)u 40
2O
o
7'
6
5 :S E 4 �9
2
i I
o 20 4o 60 ao Ioo "/. VOz~o.. 0 9.3 18.5 :)7.8 37.0 46.3 VO 2 ml/kg x rain. -I
Fig. 2. M6thode de d&ermination du seuil lactate (S[LA]) et du seuil ventilatoire (SVE) chez un sujet. VE = volume expira- tion/min; [LA], = concentration du lactate dans le sang. (D'apr~s Ivy et ak, 1980.)
santes dans le rejet de C O 2 par les poumons : l 'une correspond au produit du m6tabolisme et d6pend des substrats mis en jeu, essentiellement des gluci- des dans l'exercice de courte dur~e, avec un quo- tient respiratoire qui se rapproche de i'unit6; l'autre correspond au surplus de gaz carbonique issu de la transformation des bicarbonates. Cet accroissement de VCO 2 expliquerait la progression supplfmen- taire du QR qui devient alors sup6rieur h l et l~hyperventilation relative avec 616vation du rapport VE/VO 2 (Fig. 3). A cet instant, le pH n'est encore que peu modifi6, de m~me que la pression partielle de CO 2 dans l'air de fin d'expiration (PETCOz); par contre, la pression partielle d 'O 2 dans l 'air de fin d'expiration (PETO 2) augmente, compte tenu de l 'hyperventilation relative. Cette zone est inti- tul6e par Wasserman << zone tampon isocapnique >> (Isocapnic Buffering). Cependant, si l'intensit6 de I'exercice continue de se d6velopper, la production d'acide lactique et d'ions H + continue de s'accro~- tre et les bicarbonates de diminuer. Le pH sanguin s'abaisse alors de fagon plus marquee, offrant ainsi une stimulation respiratoire suppl6mentaire entra2- nant alors une r6ponse ventilatoire accrue avec r6duction de. PE.TCO 2 et nouvelle augmentation du rapport VE/VO 2. Ce dernier point de modifi- cation des 6changes gazeux est appel6 par Wasser- man le <<seuil de d6compensation de l 'acidose m6tabolique >> (<< Threshold of decompensed meta- bolic acidosis>>: TDMA).
6 M. Riett
Q.R.
1,3
1.2
I , I
1
0.9
oK_~/
80
70
6 0
50
40
30,
20'
tO
1... . . . . .t..-.--'I""
VCO 2 (ml.kg - t . m i n -~)
~'o 6'o A ~'o ~b ~o ~ r
70,
60,
50'
40,
30,
20'
10,
~ ~" /o 6'o
[LA]= (mmol. I - l )
10
8
6
4 ;
2, 1
- - / / sb ~b
~tO:, (ml . kg - I . rain -1 )
7b 8'0 ob 10"0 % VO~m~x
7b ~b ~'o ,do ~ ~o~,~
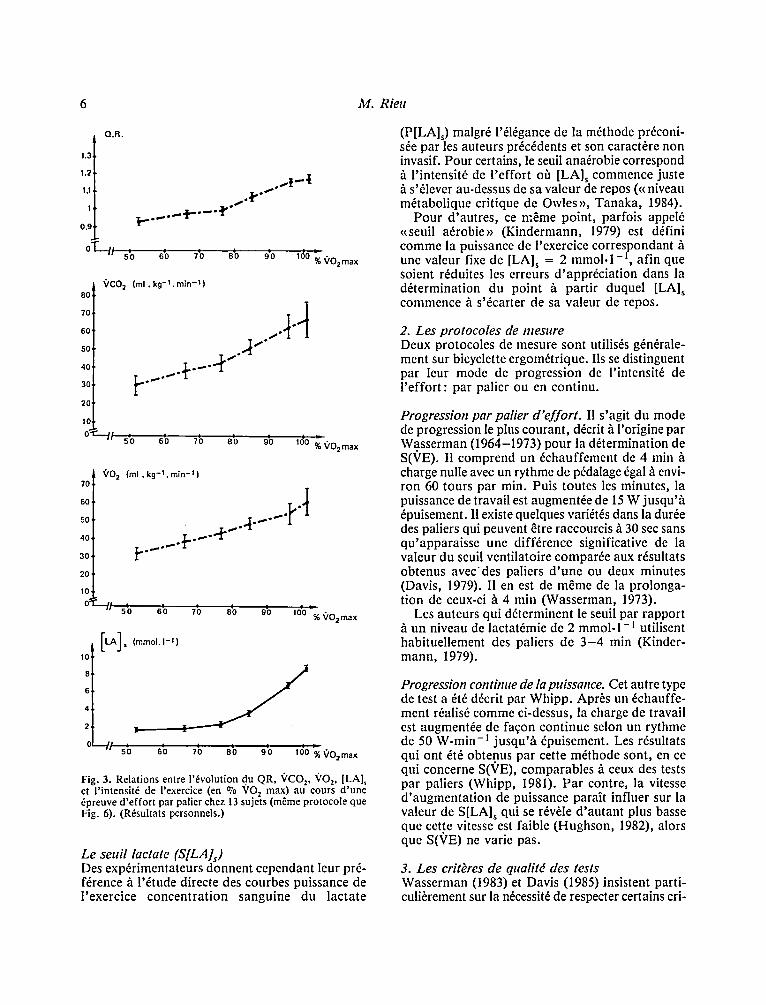
Fig. 3. Relations entre 1'6volution du QR, 'r z, VO 2, [LA], et l ' intensit6 de l'exercice (en o/0 VO 2 max) au eours d 'une 6preuve d 'effor t par palier chez 13 sujets (mfime protocole que Fig. 6). (R6sultats personnels.)
Le seuil lactate (S[LA] s) Des exp6rimentateurs donnent cependant leur prf- f f rence/ l l'6tude directe des courbes puissance de l 'exercice concentrat ion sanguine du lactate
(P[LA]s) malgr6 l'616gance de la mdthode prdconi- s6e par les auteurs pr6c6dents et son caract+re non invasif. Pour certains, le seuil anadrobie correspond
l'intensit6 de l 'effort oil [LA] s commence juste s'61ever au-dessus de sa valeur de repos (<< niveau
m6tabolique critique de Owles>>, Tanaka, 1984). Pour d'autres, ce m~me point, parfois appel6
<<seuil a6robie>> (Kindermann, 1979) est d6fini comme la puissance de l'exercice correspondant/ t une valeur fixe de [LA] s = 2 m m o i . l - l , afin que soient r6duites les erreurs d'appr~ciation dans la d6termination du point /l partir duquel [LA] s commence/ l s'6carter de sa valeur de repos.
2. Les protocoles de mesure Deux protocoles de mesure sont utilis6s g6n6rale- ment sur bicyclette ergom6trique. Ils se distinguent par leur mode de progression de l'intensit6 de l 'effor t : par palier ou en continu.
Progression par palier d'effort. I1 s'agit du mode de progression le plus courant, d6crit/l l'origine par Wasserman (1964-1973) pour la dftermination de S(~rE). II comprend un 6chauffement de 4 min :~ charge nulle avec un rythme de p6dalage ~gal/t envi- ron 60 tours par min. Puis toutes les minutes, la puissance de travail est augment~e de 15 W jusqu'/l 6puisement. I1 existe quelques vari6t6s dans la dur6e des paliers qui peuvent ~tre raccourcis/~ 30 sec sans qu'apparaisse une difffrence significative de la valeur du seuil ventilatoire comparfe aux r6sultats obtenus avecdes paliers d 'une ou deux minutes (Davis, 1979). I1 en est de mfime de la prolonga- tion de ceux-ci/l 4 min (Wasserman, 1973).
Les auteurs qui d6terminent le seuil par rapport /l un niveau de lactat6mie de 2 mmol. 1 - l utilisent habituellement des paliers de 3 - 4 min (Kinder- mann, 1979).
Progression continue de la puissance. Cet autre type de test a ~t6 d6crit par Whipp. Apr6s un 6chauffe- ment r6alis6 comme ci-dessus, la charge de travail est augment6e de fa~on continue selon un rythme de 50 W.min- I jusqu'/l 6puisement. Les r6sultats qui ont 6t6 obtenus par cette m6thode sont, en ce qui concerne S(~'E), comparables/ l ceux des tests par paliers (Whipp, 1981). Par contre, la vitesse d 'augmentation de puissance parait influer sur la valeur de S[LA] s qui se r6v~le d'autant plus basse que cette vitesse est faible (Hughson, 1982), alors que S(~/E) ne varie pas.
3. Les critbres de qualitd des tests Wasserman (1983) et Davis (1985) insistent parti- culi~rement sur la n6cessit6 de respecter certains cri-

Lactatdmie et exercice mttscttlaire 7
t6res si I 'on veut d6tecter la valeur de S(VE) avec pr6cision. En particulier, !I convient de ne pa s se contenter de la mesure de VE et de la simple d6ter- mination de l'intensit6 fi partir de laquelle VE commence h augmenter plus que ne le voudrait l'616vation de la VO 2. Pour conf i rmer cette mesure, il faut y ajouter: l'absence de modification du rapport 9CO2/VE, l 'augmentation du QR et de P E T O 2 sans var ia t ion concomi tan te de PETCO 2.
Si ces r~gles sont observ6es, les critiques de la m~thode li6e aux difficult6s d'appr~ciation de la zone d'inflexion de VE par rapport ~ VO 2, comme le montrent les r6sultats divergents obtenus sur la m~me courbe par quatre observateurs diff6rents (Yeh, 1983), deviennent sans doute moins perti- nentes.
Dans ces conditions, Wasserman et Davis don- nent une valeur pratique h la d&ermination de S(VE) (ou ~tAnaerobic Threshold~ selon leur ter- minologie) arguant en particulier du fait que pour un m~me sujet qui produit un effort, si la puissance d'exercice (P) est inf6rieure :~ S(VE), la VO 2 attein- dra un 6tat stable en 3 min environ, si elle est 6gale ou 16g6rement sup6rieure ~ S(VE) l'6tat stable VO 2 ne sera atteint qu'en 6 - 7 rain, et il ne sera jamais obtenu si P est tr~s sup6rieur fi S(VE), une d&ive continue de 902 apparaissant alors jusqu'/t 9 0 2 max.
Kindermann pour sa part (1979) pense que l 'entrainement au niveau de S[LA] s (2 m m o l . l - l) permet sinon d'accroitre tout au moins de mainte- nir la condition physique dans le domaine de l 'endurance.
4. Les rdsultats Les r6sultats obtenus par les diff6rents auteurs concordent, bien que s'6talant sur une grande 6chelle de valeur en ce qui concerne la d6termina- tion du seuil ventilatoire. Celui-ci se situe entre des puissances de 45 fi 180 W, diminuant avec le vieil- lissement chez une population de condition physi- que et d':~ge (de 40 h 91 ans) tr~s hft&og6ne (Wasserman, 1973). Ou encore en valeur relative, S(VE) repr6sente 64 + 9~ 9 0 2 max (Davis, 1976).
Les tests-retests d6voilent une corr61ation saris- faisante (r = 0,74 et 0,72 respectivement sur ergocy- cle et tapis roulant: Davis 1976; r = 0,91 sur ergocycle" Davis, 1979).
Par ailleurs, la condition physique des sujets sem- ble influer sur la valeur de S(VE). Ainsi, lors d 'une 6tude longitudinale men6e en comparaison avec un groupe t~moin, on observe que l'entra~nement physique de 9 semaines sur ergocycle entraine une
augmentation de VO 2 max de 15o70 et de.S(VE) de 44% (ou de 15o7o si on rapporte S(VE) fi VO 2 max) (Davis, 1979; Yoshida, 1981). Enfin, une excellente corr61ation entre la valeur de S(VE) et le niveau de performance de coureurs de f o n d a souvent 6t6 trouv6e (Farrell, 1979; Kumagai, 1982; Powers, 1984).
5. Les divergences Cependant, un certain nombre de difficult6s sub- sistent, qui ne permettent pas d'accorder fi ces tests une fiabilit6, d6finitive.
Ainsi est-il pour le moins nfcessaire que les conditions d'exdcution des tests soient parfaitement pr6cis6es concernant en. particulier la nature de l'exercice: en effet, S(VE) comme S[LA] S sont abaiss6s lorsque l'exercice est r6alis6 sur ergocycle compar6 au tapis roulant (Koyal, 1976). De m~me, S(VE) dftermin6/l partir d 'un exercice rfalis6 avec les bras est inf6rieur fi celui observ6 lors d 'un effort effectu6 avec les jambes (Davis, 1976; Reybrouck, 1975). Le rythme de p6da]age sur ergocycle influe aussi sur la valeur de S(VE) et de S[LA] s qui se trouvent l 'un et l'autre diminu6s lorsqu'un exercice- test est r6alis6 /~ 90 tours.min -I au lieu de 60 tours.min -I (Hugues, 1982).
Un autre 616ment soul6ve de nombreuses coritro- verses : existe-t-il r6ellement une identitd de nature entre S(k'E) et S[LA] s o u ne s'agit-il que d'ttne co'tncidence conjoncturelle? Cette discussion est essentielle puisque toute la d6finition de S(VE) est bas6e sur l'existence d 'une acidose due fi une aug- mentation de la production d'acide lactique. En la mati+re, les r6sultats sont contradictoires: pour cer- tains, la corr61ation est excellente (Wasserman, 1973; Davis, 1976; Reinhard, 1979; Ivy, 1980; Yo- sfiida, 1981 ; Caiozzo, 1982). Pour d'autres, cette relation est tr6s faible (Hugues, 1982) ou mfime n'existe pas, S.[LA] s se r6v61ant syst~matiquement inf6rieur fi S(VE) (Green, 1983) ou mfime totale- ment al6atoire (Yeh, 1983; Powers,. 1984).
En outre, la dissociation entre S(VE) et S[LA]s peut s'accentuer dans certaines circonstances: ainsi, un r6gime pauvre en glucide ou une d6plftion glyco- g6nique obtenue selon la m6thode de Gollnick (1974) entrainent une .augmentation de S[LA]s et une diminution de S(VE) (Green, 1979; Hugues, 1982; Heigenhauser, 1983). Mieux, chez des sujets atteints d 'un syndrome de MeArdle' s d6fini par une insuffisance de l'activit6 phosphorylasique muscu- laire ayant pour cons6quence une incapacit6 fi pro- duire du lactate, une 6tude (Hagberg, 1982) montre qu'une hyperventilation relative, assimilable fi celle qu! permet chez les sujets normaux de d6terminer S(VE), appara~t pour un niveau d 'effort 6gal fi

8 M. Riett
7 0 - 8 5 % 'v'O 2 max sans 616vation concomitante de la lactatfmie. Par ailleurs, l'entra~nement semble avoir un effet plus marqu6 sur l 'augmentation de S[LA] s que sur celle de S(VE) (Poole, 1984; Brooks, 1985). Enfin, les conditions de passation du test peuvent aussi jouer un r61e: dans le cas des efforts h progression de puissance contii~ue, si la progression est rapide (65,4 W.min- ~), S[LA]~ est sup6rieur ~t S(VE), alors que le contraire est observ6 si la progression est plus lente (8,2 W.min -1) (Hughson, 1982).
6. Les interrogations de f ond Des arguments viennent ~ l 'appui de ces contradic- tions: les premiers se rapportent plus pr6cis6ment ~t la signification de S(VE).
En effet, de nombreux facteurs peuvent &re, en dehors de l'acidose, h l'origine d'une hyperventi- lation relative durant l'exercice, en particulier d 'ordre nerveux ayant trait soit ~ la sensibilit6 des centres respiratoires au CO 2 qui paraR plus grande chez les sprinters que chez les coureurs de fond (Saunders, 1976; Jones, 1982), soit h la nature des sollicitations p6riphfriques qui peuvent varier car, plus l'exercice devient intense, plus le nombre de fibres de type II b qui entrent en jeu grandit (Goll- nick, 1974; Baldwin, 1977; Essen, 1978), pouvant entra~ner une intervention plus importante des r6cepteurs p6riph6riques responsables de la r~gu- lation ventilatoire (Jones, 19.82).
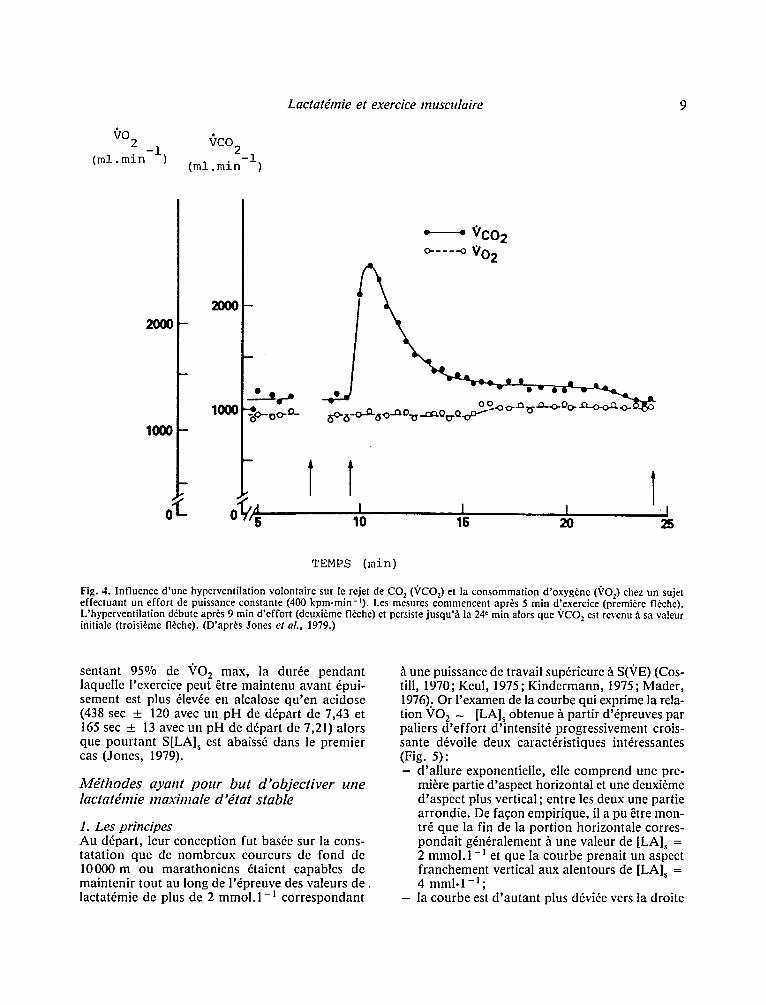
De plus, la liaison entre VCO 2 et (.'E n'est pas simple, la distinction entre lacause et l 'effet res- tant malais6e: ainsi, une hyperventilation volontaire survenant pendant la r6alisation d 'un exercice sous- maximal de puissance constante entraTne une ten- dance ~ la diminution de la pression partielle de CO 2 dans le sang veineux m~16 (PvCO z) et par vole de cons6quence une mobilisation des stocks de CO 2 de l 'organ!sme avec une augmentation sup- pl6mentaire de VCO 2 dont le retour ~ l'6tat stable, l 'hyperventilation persistant, demande plusieurs min (Fig. 4). En outre, si la relation PvCO 2- quantit6 stock6e de CO2 est approximativement lin6aire pour des efforts d'intensit6 mod6r6e, 6tant 6gale ~ 1,83 ___ 0,552 ml CO~.Torr - I PCOz.kg - l , cette capacit6 ~ stocker du CO 2 se r6duit, pour des efforts de puissance 61ev6e, b.: 1,19 + 0,49 ml COz.Torr-~.kg -~ (Jones, 1979). Par ce simple ph6nom~ne, la tendance h excr6ter du CO 2 se trou- verait donc accrue lorsque l'intensit6 de l'exercice s'61~ve (Jones, 1982).
N6anmoins, si l 'on admet qu 'une 6volution vers l'acidose m&abolique appara]t d6s que la puissance de l 'effort augmente, il est 16gitime de penser que celle-ci peut ~tre effectivement ~ l'origine d 'une
hyperventilation qui elle-m~me par la diminution de la PvCO 2 qu'elle entra~ne mobilise le CO 2 dis- sous dans les tissus de l 'organisme. Ainsi durant eette phase, ~tCO 2 pourrai t repr6senter trois composantes: l 'une constitu6e du CO 2 provenant de la combustion des substrats m6taboliques, la seconde du CO 2 provenant de l'utilisation des bicarbonates pour tamponner les ions H § , la troi- si~me du CO 2 provenant de la mobilisation des stocks.
Enfin, S(VE) est difficile it mettre en ~vidence chez les sujets qui pr6sentent une grande variabi- litd ventilatoire comme chez les sujets souffrant d'insuffisance respiratoire ou chez les enfants qui r6v~lent une fr6quence ventilatoire beaucoup plus 61ev6e que l 'adulte pour un niveau d 'effor t donn6 (Jones, 1982; Davis, 1985).
Le second type d'arguments concerne plus par- ticuli~rement la ddtermination de S[LA] s.
Deux d'entre eux seront ult6rieurement envisa- g6s car ils se rattachent ~ l 'ensemble de la d6mar- che trait6e ici: la lactat6mie n'est en effet que la r6sultante entre les dfbits d 'appari t ion et de dispa- rition du lactate au niveau du sang; la concentra- tion du lactate dans le sang n'est qu 'un reflet
�9 imparfait de celle du lactate dans le muscle. Pour les autres, il s 'agit:
D'une part, de la variation de la lactat6mie en fonction du lieu de prdl~vement des gchantillons sanguins. En effet, il est bien prouv6 que lors d 'un effort maximal, la lactat6mie veineuse est inffrieure h la lactat&fiie art6rielle (Yoshida, 1981) 6tant donn6 la possibilit6 de consommation du lactate pfir les muscles inactifs (Keul, 1967). Encore faut-il que ceux-ci soient effectivement au repos car l 'on sait, par exemple, qu 'un ph6nom6ne de crispation des avant-b]-as sur le guidon appara~t fr6quemment lors d 'un exercice r6alis6 sur ergocycle avec les membres inf6rieurs. Selon que la lactat6mie a 6t6 d6termin6e sur du sang art6riel ou veineux, son augmentation peut ainsi apparaTtre pour des efforts d'intensit6 tr~s faible ou au contraire se retrouver retard6e (Yeh, 1983).
D'autre part, de la variation de la lactat6mie en fonction de l'~tat acidobasique de l'organisme: chez un sujet au repos, une hyperventilation volontaire suffit pour entra~ner une augmentation du lactate sanguin (Huckabee, 1958). De mfime lors d 'un exer- cice de puissance donn6e, l 'acidification ~avant l'effort>) ou ~l'alcalinisation>>, provoqu6e par l 'absorption de comprim6s de C1NH 4 pour la pre- mitre condition ou de CO3HNa pour la seconde, entraTne respectivement une diminution ou une aug- mentation de la lactat6mie art6rielle compar6e aux valeurs t6moins. Cependant pour un effort reprO-
Lactatdmie et exercice musculaire 9
2 -i
(ml.min VCO 2
) (ml .min -I)
1 0 0 0 -
0"
m
m
= _- ~'CO 2 o--- - -o V02
�9 t,_.
1000 "~o-- oo-~
- T I 1 10 16 20 25
TEMPS (min)
Fig. 4. Influence d'une hyperventilation volontaire sur le rejet de CO 2 (VCO2) et la consommation d'oxyg~ne (VO2) chez un sujet effectuant un effort de puissance constante (400 kpm.min-t). Les mesures commencent apr~s 5 min d'exercice (premiere fl~che). L'hyperventilation d~bute apr~s 9 min d'effort (deuxi~me fl~che) et persiste jusqu'/~ la 24' min alors que ~'CO 2 est revenu/l sa valeur initiale (troisi~me fl~che). (D'apr~s Jones et al., 1979.)
sentant 95~ de 9 0 2 max, la dur fe pendant laquelle l 'exercice peut ~tre maintenu avant 6pui- sement est plus 61evfe en alcalose q u ' e n acidose (438 see ___ 120 avec un pH de d6part de 7,43 et 165 sec ___ 13 avec un pH de d6part de 7,21) alors que pour tan t S[LA] s est abaiss6 dans le premier cas (Jones, 1979).
Mdthodes ayant pour but d 'object iver une lactat~mie maximale d'dtat stable
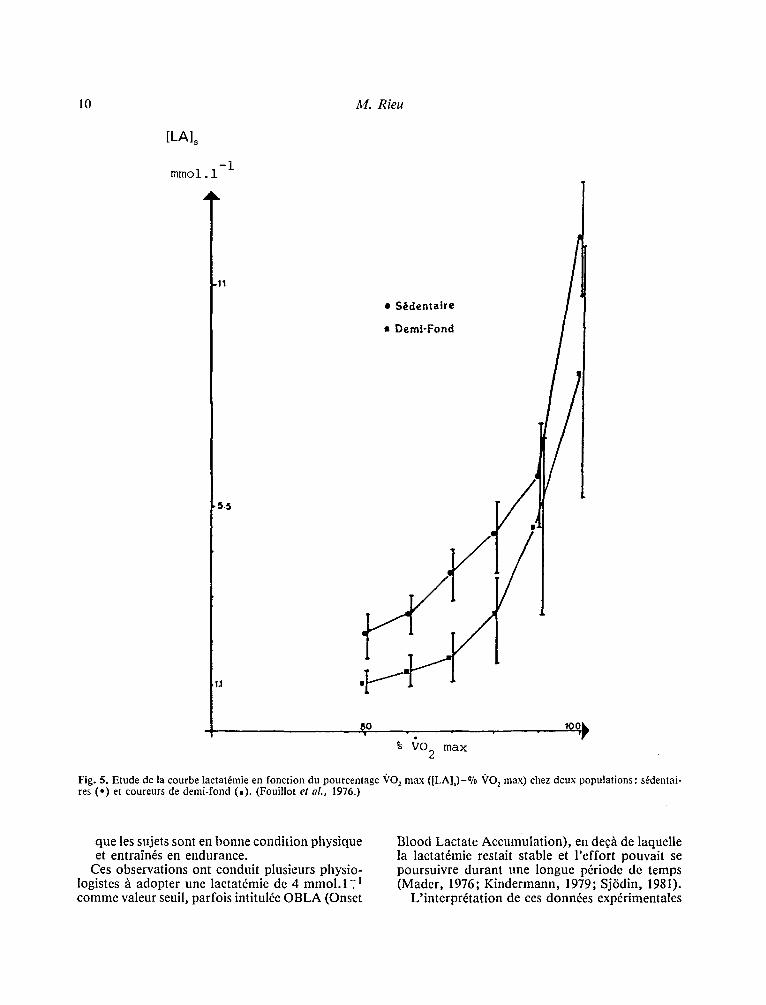
1. Les principes A u d6part , leur concept ion fut bas6e sur la cons- ta ta t ion que de nombreux coureurs de fond de 10000 m ou marathoniens &aient capables de maintenir tout au long de l '6preuve des valeurs de . lactat6mie de plus de 2 mmol . 1- l co r respondan t
/l une puissance de travail sup6rieure ~ S(~'E) (Cos- till, 1970; Keul, 1975; Kindermann, 1975; Mader , 1976): Or l 'examen de la courbe qui exprime la rela- tion VO 2 - [LA] s obtenue/ l partir d '6preuves par paliers d ' e f fo r t d ' intensit6 progressivement crois- sante d6voile deux caract6ristiques int6ressantes (Fig. 5): - d 'a l lure exponentielle, elle comprend une pre-
mi6re partie d 'aspect horizontal et une deuxi6me d ' aspec t plus vertical; entre les deux une patt ie arrondie. De faqon empirique, il a pu ~tre mon- tr6 que la fin de ia por t ion horizontale corres- pondai t g6n6ralement h une valeur de [LA]s = 2 mmol . 1 - ~ et que la courbe prenait un aspect f ranchement vertical aux alentours de [LA] s = 4 mml . l - I "
- la courbe est d ' a u t a n t plus d6vi6e vers la droite
10 M . R ieu
[LA]~
mmol. i -I
.11
.5-5
�9 S ~ d e n t a i t e
�9 Demi-Fond
/ /
(~o 'took �9 b~l,"
% VO 2 max
Fig. 5. Etude de la courbe lactat6mie en fonction du pourcentage ~'O z max ([LA]s)-% ~'O 2 max) chez deux populations: s6dentai- res (,) et coureurs de demi-fond (.). (Fouillot et al., 1976.)
que les sujets sont en bonne condition physique et entrain6s en endurance.
Ces observations ont conduit plusieurs physio- logistes h adopter une lactat6mie de 4 mmol . l _ l comme valeur seuil, parfois intitulfe OBLA (Onset
Blood Lactate Accumulation), en de~h de laquelle la lactat6mie restait stable et l 'effort pouvait se poursuivre durant une longue pfriode de temps (Mader, 1976; Kindermann, 1979; Sj6din, 1981).
L'interpr6tation de ces donn6es exp6rimentales

Lactatdmie et exercice musctdaire 11
tient compte du fait que la lactat6mie est le r6sul- tat d 'une balance entre le d6bit d'apparition du lac- tate dans le sang et son d6bit de disparition. Aussi la lactat6mie peut augmenter transitoirement au d6but de I'exercice pendant la phase de d6ficit en oxyg6ne (Cerretelli, 1977-1979), puis lors de la phase d'6tat stable pour les efforts de faible inten- sit6 se r~duire, r6v61ant ~ ce stade la pr6dominance des processus d'61imination du lactate; pour des exercices de puissance plus 61ev6e, elle peut rester constante t6moignant d 'un 6quilibre respect6 entre apparition et 61imination; pour les efforts intenses, elle augmente tout au long de l'6preuve, montrant ainsi que l'6quilibre pr6c6dent est d6truit.
Le niveau maximum possible de la lactat6mie compatible avec un 6tat stable peut ainsi ~tre consid6r6 comme indiquant le vrai ~seuil ana6ro- bie ~, c'est-~-dire la puissance d'exercice au-delh d e laquelle l'6nergie n6cessaire ne peut plus ~tre four- nie exclusivement par les processus m6taboliques a6robie.
La comparaison d'6preuves d 'effor t par paliers successifs et de cr6neaux d'efforts s6par6s a permis de montrer que, pour un exercice sur tapis roulant de plus de 30 min r6alis6 h la vitesse correspondant au S[LA] s (4 mmol. 1- l) pr6alablement 6tabli lors d 'un test triangulaire, la concentration du lactate dans le sang atteignait cette valeur en environ 5 min, puis restait constante (Kindermann, 1979). C e t auteur proposa une tentative de synth6se des diff6- rentes conceptions en distinguant : - un seuil adrobie correspondant ~t ~l 'anaerobic
threshold~ de Wasserman ddtermin6 soit par la m6thode ventilatoire (S(~/E)), soit par celle du lactate sanguin (S[LA]~) dont le S[LA] s (2 m m o l . l - l ) ;
- un seuil ana6robie correspondant au S[LA]s (4 mmol. 1 - t ) ;
- une zone de transition ~(a6robie-ana6robie~ situ6e entre les deux et assurant le passage d 'un m6tabolisme strictement a6robie h u n m6tabo- lisme dans lequel le suppldment d'dnergie ndces- saire :~ l 'augmentation d'intensit6 de l 'effort est de nature strictement ana6robie.
2. Les protocoles de mesure Les protocoles sont 6tablis de la fa~on suivante: il s'agit d'6preuves r6alis6es sur bicyclette ergom6tri- que ou plus souvent sur tapis roulant. Apr~s un 6chauffement effectu6 dans ce dernier cas ~ une vitesse de 10, voire 12 km.h - I , selon le degr6 d'entra~nement du sujet, les paliers d 'une dur6e de 3 - 5 min se succ~dent entrecoup6s ou non de phase. de repos de 30 sec/~ 1 min. La vitesse est augmen-
t6e entre chaque palier de 1 - 2 km.h- I jusqu'~ la survenue de l'6puisement.
Le lactate sanguin est habituellement dos6 sur des micro6chantillons (50-100 ~1) de sang art6rialis6, pr61ev6 au bout du doigt ou au lobule de l'oreille
la fin de chaque palier. L'6tablissement des courbes vi tesse- [LA]s per-
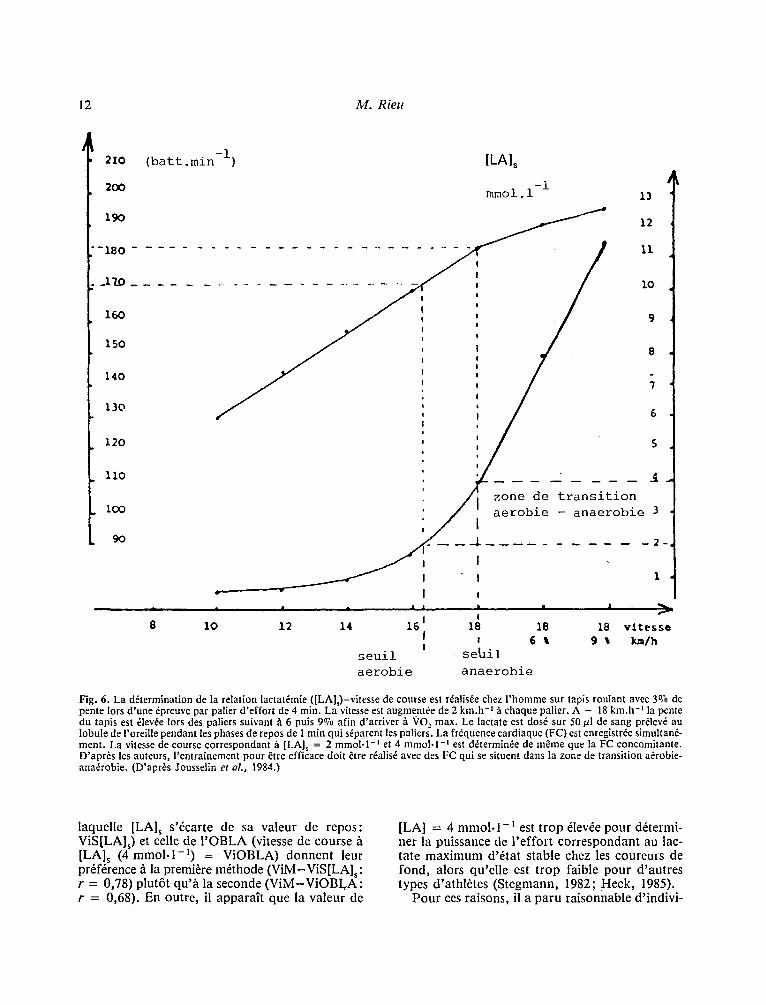
met de d6finir par interpolation la valeur absolue e t /ou relative (en %o VO 2 max) de la vitesse cor- respondant h [LA]s = 4 mmol. l - i . La mesure de la fr6quence cardiaque h ce m~me niveau de vitesse permet en outre d 'avoir une estimation de la puis- sance dfvelopp6e sur la piste d'entra~nement et de sa valeur par rapport au seuil (Fig. 6).
Dans une 6tude r6cente (Heck, 1985), comparai- son a fit6 faite entre d 'une part la vitesse de course sur tapis roulant correspondant h la lactat6mie maximale d'6tat stable (maximal lactate steady state), d6termin6e par des efforts en cr6neaux de 25 min entrecoup6es toutes les 5 min de 30 sec de pause pour le recueil des 6chantillons sanguins, cha- que cr6neau 6tant d'intensit6 croissante, l 'un par rapport h l 'autre et effectu6 ~ distance; d'autre part la vitesse de course correspondant ~ S[LA]s (4 m m o l . l - l ) , pr6alablement d6termin6 par un test par palier de 3 min. La corr61ation est excel- lente (r = 0,979).
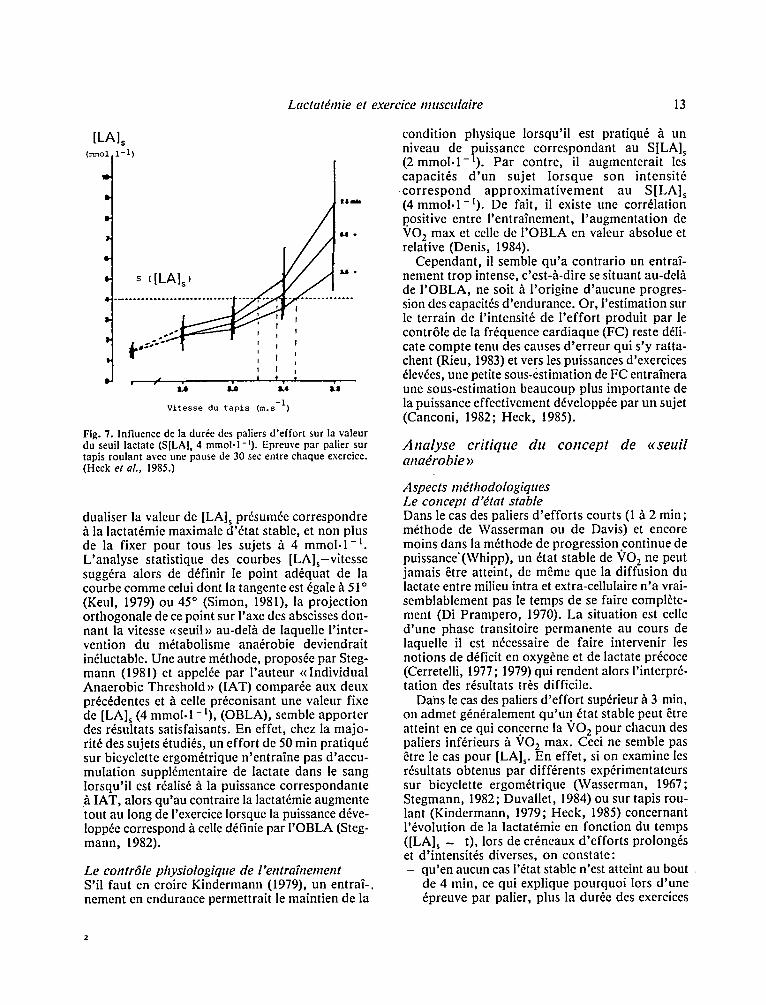
Dans ce mfime travail, les auteurs analysent plu- sieurs points m6thodologiques. Ils montrent en par- ticulier comme auparavant Kindermann (1980) que plus les paliers sont de dur~e 61ev~e (3,5; 5,5; 7,5 min) plus S[LA]s (4 mmol.l - I ) sera bas (Fig. 7), alors que la modification de la dur6e des phases de pauses intercalaires (30 sec; 1 ; 1,5 min) ne joue aucun r61e. Par contre, les caract6ristiques m6ca- niques du tapis roulant influe inconstestablement sur la valeur du seuil. Enfin I'imposition d 'une fai- ble pente ( I - 2 % ) au tapis roulant am61iore les cor- r61ations qui peuvent exister entre les vitesses observ6es au laboratoire et sur le terrain pour une m~me lactat6mie. Par ailleurs, la valeur de S[LA] s (4 mmol .1 - i ) s'est rfv616e ~tre moins 6levee sur bicyclette ergom6trique que sur tapis roulant (Kin- dermann, 1981).
3. Les rdsultats Pr~vision de la performance La pr6vision de la performance par la d6termina- tion de I 'OBLA donne des r6sultats contradictoi- res: d'apr~s certains auteurs (Sj6din, 1981), cette mesure rend compte pour 92% des diff6rcntes vites- ses de course observ6es chez les marathoniens (VIM). D'autres (Tanaka, 1984) comparant sur tapis roulant la valeur pr6dictive du S[LA]s (d6finie comme la vitesse de course ~t partir de
12 M. Rieu
210 (batt .rain -I) [LAI~
200 mmol 1 -I �9 13 190 ~ 12
- z s o . . . . . . . . . . . . . . . . . . . . . . . n
_ r m . . . . . . . . . . . . . . . . . . . . . . . . . . . ~ , I 1o
/ I J
i ! 150 8
1 4 0 I i -
1~o : ; , 6 "
no : : s
11o : . _ _ _ _ _ _ . 4
[ ion
I00 i / I aerobie - anaerobie 3
9o - - t - 2-
I I 1
- - I ! I J dl �9 , I .t ,& A |
8 10 12 14 161 18 18 18 v l t e s s e f ~ 6 % 9 % k~lh I
seuil se%il aerobie anaerobie
Fig. 6. La d6termination de la relation lactat6mie ([LA],)-vitesse de course est r~alisde chez l'homme sur tapis roulant avec 30"0 de pente Iors d'une dpreuve par palier d'effort de 4 min. La vitesse est augmentde de 2 km.h -I ~ chaque palier. A - 18 km.h -I la pente du tapis est dlevde lors des paliers suivant h 6 puis 9~ afin d'arriver ~ VO 2 max. Le lactate est dosd sur 50 t~l de sang prdlevd au lobule de l'oreille pendant les phases de repos de 1 rain qui sdparent les paliers. La frdquence cardiaque (FC) est enregistr6e simultand- ment. La vitesse de course correspondant fi [LA], = 2 rnmol-I -~ et 4 mmol. l - t est ddterminde de m~me que la FC concomitante. D'apr6s les auteurs, l'entraTnement pour ~tre efficace dolt &re rdalis6 avec des FC qui se situent dans la zone de transition adrobie- ana6robie. (D'apr~s Jousselin et aL, 1984.)
l a q u e l l e [LA] s s ' dca r t e de sa v a l e u r de r e p o s : ViS[LA]5) et cel le de I ' O B L A (vi tesse de c o u r s e [LA] s (4 m m o l . l - I ) = V i O B L A ) d o n n e n t leur prdf~rence ~ la p remie re m d t h o d e ( V i M - V i S [ L A ] s : r = 0 ,78) p lu t6 t qu ' f i la s e c o n d e ( V i M - V i O B L A : r = 0 ,68) . En ou t r e , il appara~t q u e la v a l e u r de
[LA] = 4 m m o l . l - ] e s t t r o p dlevde p o u r dd te rmi - n e r la p u i s s a n c e de l ' e f f o r t c o r r e s p o n d a n t au lac- t a t e m a x i m u m d ' d t a t s t ab le chez les c o u r e u r s de f o n d , a lors q u ' e l l e est t r o p fa ib le p o u r d ' a u t r e s types d ' a t h l 6 t e s ( S t e g m a n n , 1982; H e c k , 1985).
P o u r ces r a i sons , il a p a r u r a i s o n n a b l e d ' i n d i v i -
Lactatdmie et exercice musctdaire 13
[LA]~ (m~ol l-l)
�9 t o a d
�9 s c [LA]~
I , l
�9 ~, ~ . �9 ~r ~ U L4 & |
Vitesse du tapis (m.s -I)
Fig. 7. Influence de la duroc des paliers d 'ef for t sur la valeur du seuil lactate (S[LA]~ 4 m m o l . 1 - 9 - Epreuve par palier sur tapis roulant avec une pause de 30 sec entre chaque exercice. (Heck et aL, 1985.)
dualiser la valeur de [LA] s prdsumde correspondre b. la lactat6mie maximale d '&at stable, et non plus de la fixer pour t ous l e s sujets b. 4 m m o l . l - I L'analyse statistique des courbes [LA]s-vitesse sugg6ra alors de d6finir le point ad6quat de la courbe comme celui dont la tangente est 6gale b. 51 o (Keul, 1979) ou 45 ~ (Simon, 1981), la projection orthogonale de ce point sur I'axe des abscisses don- nant la vitesse <<seuil>> au-del/l de laquelle l ' inter- vention du mftabolisme ana6robie deviendrait indluctable. Une autre m6thode, proposde par Steg- mann (1981) et appel6e par l 'auteur <<Individual Anaerobic Threshold >> (IAT) compar6e aux deux prdc6dentes e t / l celle pr6conisant une valeur fixe de [LA] s (4 mmol. 1-i) , (OBLA), semble apporter des r6sultats satisfaisants. En effet, chez la majo- rit6 des sujets 6tudids, un effort de 50 min pratiqu6 sur bicyclette ergom6trique n'entraine pas d'accu- mulation suppldmentaire de lactate dans le sang lorsqu'il est r6alis6 h la puissance correspondante :~ IAT, alors qu'au contraire la lactatdmie augmente tout au long de l'exercice lorsque la puissance d6ve- loppde correspond b. celle d6finie par I 'OBLA (Steg- mann, 1982).
Le contr61e physiologique de l'entra~nement S'il faut en croire Kindermann (1979), un entrai-. nement en endurance permettrait le maintien de la
condition physique lorsqu'il est pratiqu~ h un niveau de puissance correspondant au S[LA] s (2 m m o l . l - I ) . Par contre, il augmenterait les capacit~s d 'un sujet lorsque son intensit6
c o r r e s p o n d approx ima t ivemen t au S[LA] s (4 m m o l . l - I ) . De fait, il existe une correlation positive entre l'entraTnement, l 'augmentation de VO 2 max et celle de I 'OBLA en valeur absolue et relative (Denis, 1984).
Cependant, il semble qu'a contrario un entrai- nement trop intense, c'est-~-dire se situant au-delh de I 'OBLA, ne soit .~ l 'origine d'aucune progres- sion des capacit6s d'endurance. Or, l'estimation sur le terrain de l'intensit6 de l 'effort produit par le contr61e de la fr~quence cardiaque (FC) reste d~li- cate compte tenu des causes d 'erreur qui s 'y ratta- chent (Rieu, 1983) et vers les puissances d'exercices filev6es, une petite sous-estimation de FC entrainera une sous-estimation beaucoup plus importante de la puissance effectivement d6velopp6e par un sujet (Canconi, 1982; Heck, 1985).
Analyse critique du concept de <r anadrobie),
Aspects mdthodologiques Le concept d'dtat stable Dans le cas des paliers d 'efforts courts (1 b. 2 min ; m6thode de Wasserman ou de Davis) et encore moins dans la m6thode de progression.continue de puissance'(Whipp), un &at stable de VO 2 ne peut jamais ~tre atteint, de mfime que la diffusion du lactate entre milieu intra et extra-cellulaire n 'a vrai- semblablement pas le temps de se fake compl~te- ment (Di Prampero, 1970). La situation est celle d 'une phase transitoire permanente au cours de laquelle il est n6cessaire de fake intervenir les notions de d~ficit en oxyg~ne et de lactate pr~coce (Cerretelli, 1977; 1979) qui rendent alors l'interpr6- ration des rfisultats tr~s difficile.
Darts le cas des pallets d 'effort sup6rieur ~ 3 rain, on admet g6n~ralement qu 'un ~tat stable peut fitre atteint en ce qui concerne la VO 2 pour chacun des paliers inf6rieurs ~ VO 2 max. Ceci ne semble pas fitre le cas pour [LA] s. En effet, si on examine les r6sultats obtenus par diff6rents exp6rimentateurs sur bicyclette ergom~trique (Wasserman, 1967; Stegmann, 1982; Duvailet, 1984) ou sur tapis rou- lant (Kindermann, 1979; Heck, 1985) concernant l '6volution de la lactat6mie en fonction du temps ([LA] s - t), lors de cr~neaux d 'efforts prolong(:s et d'intensit6s diverses, on constate:
- qu'en aucun cas i'6tat stable n'est atteint au bout de 4 min, ce qui explique pourquoi lots d 'une ~preuve par palier, plus la duroc des exercices

14 M. Rieu
est longue, plus S[LA] s (4 mmol-I - l ) est abaiss6;
- que plus l'exercice est intense, plus l'6tat stable semble long ~t s 'instaurer;
- que l'6tat stable peut appara~tre pour des valeurs tr~s 61ev~es de [LA]s (8 -10 m m o l . l - l ) ;
- qu 'on observe toujours un infldchissement de la courbe de la lactatdmie en fonction du temps, ce qui signifie que, m~me pour des intensit6s proches de VO z max, la puissance de l'exercice restant constante, il y a une tendance vers l'6tat stable, puisqu'au fur / t mesure que la durde de l 'effort se prolonge, l'~quilibre entre les proces- sus qui conditionnent l 'apparition du lactate dans le sang et son 61imination tend/ t se r6ta- blir. Ceci ne devrait pas fitre le cas si [LA]s dtait le reflet de l 'intervention du mdtabolisme ana6- robie qui n'aurait en effet aucune raison de se rdduire, si l 'hypoth~se de base 6tait vraie, l'intensit6 de l 'effort ne variant pas.
La valeur de la lactatdmie enregistrde ~t la 4 r min d 'un effort ne peut donc en aucune mani~re ~tre considdrde comme la valeur correspondant h celle d 'un dtat stable. Peut-elle alors, par des voies indi- rectes qui resteraient ~ 6lucider, correspondre ~ la puissance de l 'effort pour lequel un individu attein- dra sa lactat6mie maximale d'dtat stable, sans pr6- juger de la valeur effective de cette lactatdmie? Rien ne le prouve.
2. Difficultds d'interprdtation de [LA] s Le d o s a g e d e la lactat6mie ~ la fin de chaque exercice lors d 'une ~preuve d 'effor t par palier de 3 - 5 min pose quelques probl~mes d'interpr6tation.
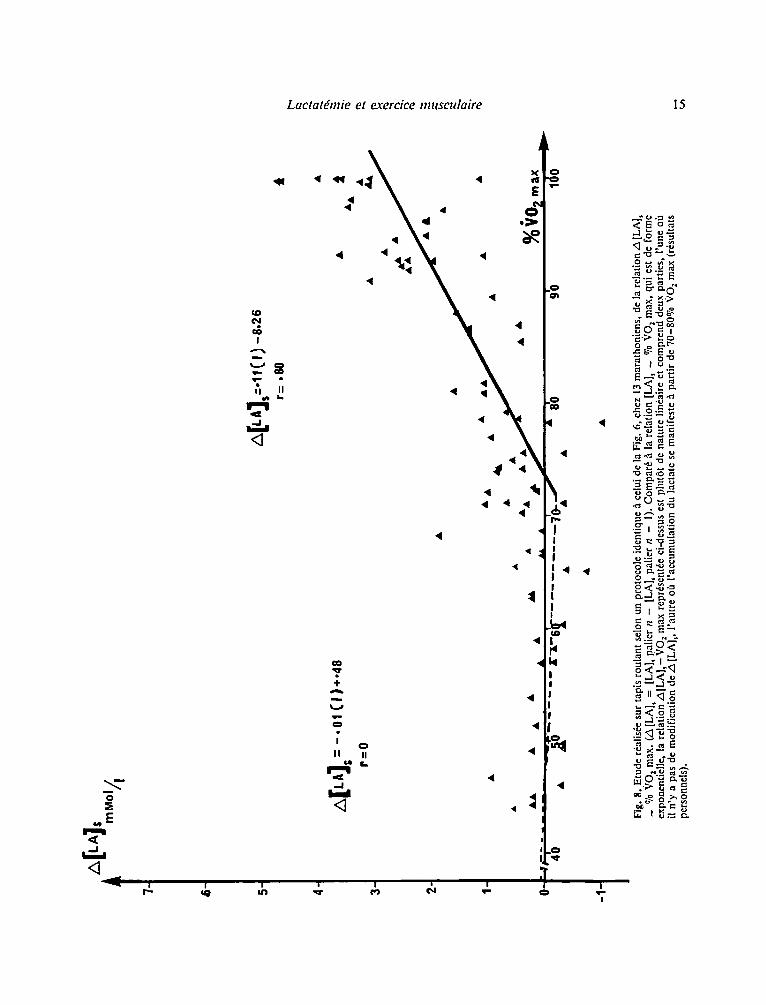
En premier lieu, la relation [LA]~ - VO 2, ~ta- blie lors d 'une 6preuve constitu6e de paliers d 'efforts d'intensit6 croissante, correspond h une cumulation des effets d'exercices successifs dont les r6manences se superposent. Un tel mod61e est dif- ficilement comparable ~ un effort isol6. Aussi peut- on s'interroger sur le sens rdel dans ce cas du para- m~tre [LA] s. Au moins faudrait-il consid6rer les diff6rences de niveau de la lactatdmie:
zS[LAls = [LA] s (palier n)-[LA]s (palier n - l ) ; [LA]s (palier n) = lactat6mie mesur~e h la fin d 'un palier d 'effort n ; [LA] s (palier n - l ) = lactat6mie mesur6e h la fin du palier pr~c6dent. Cette dfmarche, lorsqu'elle est r~alis6e, modifie
l'aspect du phfnom~ne, la relation A [LA]s - VO z pouvant mfime perdre l'allure exponentielle de la relation [LA] s - VO 2 (Fig. 8).
En second lieu, il s'agit d 'une valeur instantande qui ne donne aucune information sur la cin6tique
du lactate sanguin au cours de l 'effort qui vient de s'achever. Ainsi [LA] s peut-il purement et simple- ment repr6senter (Fig. 9): - le reliquat du <<lactate pr6coce>> (LA D) apparu
au d6but de l'exercice consid6r6 et qui igeu! avoir 6t~ consomm6 en totalit6 ou partie par la suite, en particulier.pour des efforts de faible inten- sit6 (~< 50e/0 VO2 max);
- le reflet exact du (LAp); un moment nullemen~ privil6gi6 d 'une phase transitoire lors d'exercices plus intenses qui demandent, nous l 'avons vu, un temps assez long pour que [LA] s parvienne h un 6tat stable.
Enfin on sait qu'apr6s l 'arr& de l'exercice, la lac- tat6mie continue d 'augmenter pendant 3 - 5 rain (Margaria, 1933, 1963; Crescitelli, 1944; Davis, 1971, Di Prampero, 1981). Les mesures effectu6es durant les p6riodes de repos interm6diaires qui, lors des 6preuves par palier, n'exc~dent pas 1 min ne tiennent absolument pas compte de ce ph6nom~ne sans doute Ii6 ~ la lenteur de diffusion du lactate entre les milieux intra et extra-cellulaires.
Aspects fondamentaux La signification physiologique propre de la lacta- t6mie peut elle-m~me ~tre remise en question et en partieulier, l 'identification trop souvent faite entre augmentation de la concentration du lactate san- guin et d6veloppement d 'une ana6robiose. En outre, le mode de r6partition du lactate entre le muscle et le milieu sanguin est loin d '&re enti6re- ment 6lucid6. Enfin l'analyse de l'6quilibre existant entre les processus responsables d 'une part de l 'apparition du lactate dans le sang et d 'autre part de sa disparition laissefit encore bien des points obscurs.
1. Lactat~mie et ana~robiose Bien que toutes les conditions exp6rimentales ten- dant ~ r6duire l 'apport d'oxyg~ne aux tissus telles que I'an6mie aigu~ (Woodson, I978) ou encore l ' inhalation d 'oxyde de carbone (Pirnay, 1971) entraTnent un abaissement de S[LA] s, on ne saurait pour autant identifier la production du lactate et son apparition dans le sang exclusivement hun d6fi- cit en oxyg6ne.
En effet, au repos les muscles produisent du lac- tate et leur production augmente pour des exerci- ces m~me mod6r6s (Jorfeldt, 1970; Hagenfeldt, 1972), une relation lindaire peut ainsi ~tre 6tablie chez le rat entre la V.O z et le d6bit de renouvelle- ment du lactate (LAr) (Donovan, 1983). Dans aucune de ses conditions, on ne peut mettre en 6vi- dence un d6ficit quelconque en oxyg6ne. D'ailleurs, m~me au cours d'exercices 6puisants, la PO 2 du
Lactatdmie el exercice musculaire 15
<1 d. !
~D
I
M
<]
r �9 er
�9 " N N , ~ " .
" : \ Lo ".\F
4"
I I I II
. - / I= . , . I <]
I &
4 �9
t I I
!
I |
4
4
. 0
I mini 1 ' 0
!
I
q o - 2 O ~ N o - u.} ~
- ~ . >
~o~T "~:.~ ,-. o
�9 ~ ' =_~
"~, ~ ~ ~
o < ,~ .--, ~.~ ~.o
N o ~
~ ,...-," I ~ ~ < .-:<1
N O - . . 3 ~.--~ . .> ~
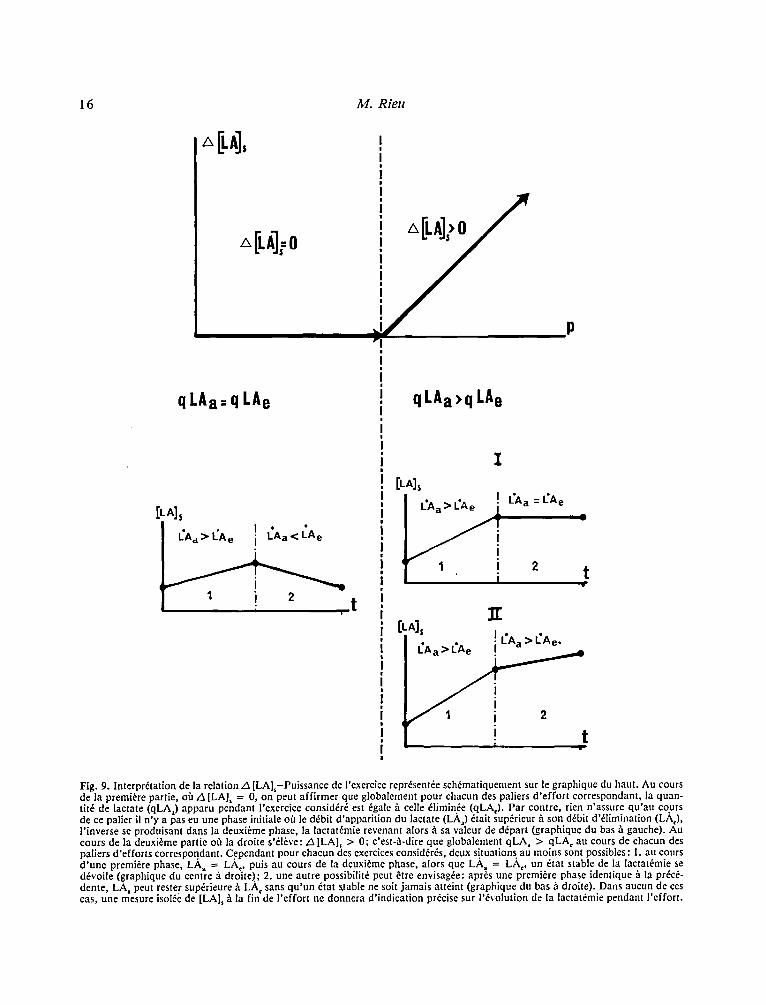
16 M. Rieu
q L A a = q L A e
[LA]s
i ' i ,t
I
i I
I I
I [LA]to /
i / d J " q
i i I
J LA e i qLAa>q i
I I
I i Z |
! [,A], ] ' �9 �9 = t ' A a - - t : A e
!
! ' z t I I �9
i |
i 112 i A]S I L*Aa > L.'Ae ~ i 0,~ > L'A~ i
i
! ~ t |
Fig. 9. Interpretation de la relation .A [LA],-Puissance de I'exercice repr~sent6e sch~matiquement sur le graphique du haut. Au cours de la premiere partie, off z5 [LA], = 0, on peut affirmer que globalement pour chacun des paliers d 'effor t correspondant, la quan- tit6 de lactate (qLA) apparu pendant I'exercice consid~r~ est ,~gale fi celle ~limin6e (qLA,). Par contre, rien n'assure qu'au cours de ce palier il n 'y a pas eu une phase initiale ot~ le d~bit d'apparition du lactate (L./k) ~tait sup6rieur/t son d6bit d'~limination (LA,), l 'inverse se produisant dans la deuxi~me phase, la lactat6mie revenant alors ~ sa valeur de d~part (graphique du b a s h gauche). Au cours de la deuxi~:me partie off la droite s'~l~ve: z5 [LA], > 0; e'est-~.-dire que globalement qLA= > qLA, au cours de chacun des paliers d 'efforts correspondant. C.ependant pour chacun des exercices consid6r6s, deux situations . au moins sont possibles : 1. au cours d 'une premiere phase, LA= = LA,, puis au cours de la deuxi~me phase, alors que LA= = LA o un dtat stable de la lactat~mie se d6voile (graphique du centre/~ droite).; 2. une autre possibilit~ peut etre envisag~e: apr6s une premiere phase identique h la pr~c6- dente, LA, peut rester sup~rieure b. LA, sans qu 'un 6tat stable ne soit jamais atteint (graphique du bas/ t droite). Dans aucun de ces cas, une mesure isol~e de [LA], 1~ la fin de l 'effort ne donnera d'indication precise sur l'~volution de la lactat6mie pendant I 'effort.

Lactatdmie et exercice musculaire 17
sang veineux issu du territoire musculaire en acti- vit6 n'est jamais inf6rieure ~t 15-20 mmHg (Pir- nay, 1971). On doit cependant s 'interroger sur la valeur de cet argument. En effet le muscle n'6tant pas un syst6me homog~ne, on peut sans doute pen- ser que toutes les boucles capillaires ne sont pas ouvertes simultanfment, ce qui entraTnerait un effondrement des r6sistances vasculaires incompa- tibles avec l'6quilibre h6modynamique. Aussi l'exis- tence d 'un shunt fonctionnel art6rio-veineux est-elle vraisemblable. Deux 616ments confortent n6an- moins l'id6e que dans des circonstances normales il n 'y a pas d'insuffisance d 'apport en oxyg6ne au cours de l'exercice m~me intense: d 'une part, l '6tude fluoroscopique du rapport N A D / N A D H intracellulaire dans le muscle isol6 du chien ne r6v~le aucune.limite d'activit6 de la chaTne respiratoire mitochondriale m~me au cours des contractions maximales (Jobsis, 1968); d 'autre part, sur le mus- cle gracilis de chien stimul6 61ectriquement, la ~rO 2 tissulaire estim6e par l '6tude cryomicrospectrosco- pique de la myoglobine ne tombe jamais au-dessous de 2 torr, ce qui est suffisant pour assurer les r6ac- tions oxydatives m~me lors de la stimulation maxi- mum puisqu'~ ce niveau intense de secousses, l'616vation de PO 2 fi 10 torr (valeur sup6rieure h celle du repos) ne modifie pas la VO 2 (Connett, 1984).
En fait, le lactate appara~t plut6t comme un mdtabolite intermddiaire de la glycolyse en dquili- bre selon la loi d'action de masse avec le pyruvate qui tend d 'autant plus fi s 'accumuler que la ddgra- dation du glycog6ne est intense: en effet, la vitesse des rdactions du cycle de Krebs est connue pour ~tre nettement plus faible que celle de la glycolyse alors qu 'au contraire, l'activit6 enzymatique de la LDH est tr6s 61ev~e.
On sait d 'autre part, que les fibres FTb (fibres blanches)/l m6tabolisme glycolytique poss~dent une capacit6 de production de lactate environ 15 fois sup6rieure fi celle des fibres ST (fibres rouges) ~t m6tabolisme oxydatif qui, au contraire, sont capa- bles de capter et d'utiliser le lactate comme subs- trat 6nerg6tique. Aussi, comme lorsque la puissance d 'un exercice augmente, le recrutement des fibres FT s'accentue (Gollnick, 1974; Baldwin, 1977; Essen, 1978), il est 16gitime que la production de lactate s'accroisse dans de grandes proportions sans qu'il soit n6cessaire de faire intervenir une insuffi- sance d 'appor t en oxyg~ne. Aussi comprend-on pourquoi I 'OBLA (Sjfdin, 1981) et S[LA] s (Ivy, 1980) sont positivement corr616s au pourcentage de fibres ST entrant dans la composition du muscle et ~ la capacit6 d'homog~nats de muscles ~ oxyder le pyruvate (Ivy, t980).
D'autres facteurs enfin sont connus pour Etre res- ponsables d 'une augmentation de la lactat6mie et ne peuvent ~tre directement rattach6s fi un d6ficit en oxyg6ne. Ainsi chez l 'homme comme chez le chien, une alcalinisation du sang provoqufe par une hyperventilation volontaire ou une perfusion de bicarbonate, la perfusion d 'une solution de pyru- vate de pH5 (0 ,2-0 ,4mM.min-1) , ou encore celle de 500 ml de glucose ~t 10~ entra~nent une ~16va- tion de [LA] s sans modification du rapport lac- tate/pyruvate (Huckabee, 1958).
Au vu de ces r6sultats, on peut m~me se poser la question de savoir si l 'abaissement de S[LA] s observ6e dans les cas d'an6mie aiguE ou d'intoxi- cation ~t l 'oxyde de carbone (voir plus haut) ne serait pas, pour une partie, la cons6quence indirecte d'une hyperventilation compensatrice entra~nant une alca- lose gazeuse.
De m~me, l 'injection continue pendant 30 min chez l 'homme de 0,15 mg.kg -1. min -1 d'adr6na- line a pour cons6quence un accroissement de [LA] s avec augmentation du rapport lactate/pyruvate (Greene, 1961).
2. Lactate musculaire et lactate sanguin Trop souvent encore, l 'accumulation du lactate dans le sang observ6e lors d'6preuve par palier ou d'exercice intense est considfr6e comme directement li6e fi la production de lactate. Cette assimilation est abusive. En effet, au cours de l'exercice, le lac- tate produit au sein de la cellule musculaire peut: s'accumuler sur place et pour une part subir un pro- cessus d 'oxydat ion; pour une autre part diffuser dans l'espace extracellulaire o6 il peut fitre capt6 et utilis6 par d'autres fibres musculaires voisines fi mftabolisme oxydatif plus prononc6; e t /ou enfin, apparaTtre dans le sang veineux eff6rent du terri- toire musculaire consid6r6. Au cours de la recircu- lation qui suit, le lactate peut alors &re capt6 par de nombreux autres organes tels que les muscles au repos, le myocarde, le foie, etc. Toutes les compa- raisons faites entre la concentration du lactate dans les homog6nats de muscles [LA] met celle du lac- tate sanguin [LA] s montrent qu'fi partir d 'une puissance d'effort sup~rieure ~t 75-80~ VO 2 max, la valeur de [LA] m est sup6rieure ~t celle de [LA] s (Diamant, 1968; Karlsson, 1971; Jorfeldt, 1978). Les r~sultats sont cependant tr~s variables. Ainsi, pour des exercices d'intensit6s modfr~es (30, 50 et 70~ ~'O 2 max) r~alis~s sur ergocycle, Jordfeldt (1978) trouve dans les homog6nats de muscle vaste externe, des concentrations de lactate inf6rieures fi celles du sang artfriel pr6Iev6 simultanfment fi la 4 ' et 12 ~ min. Par contre, Green (1983) au cours d 'un test effectu6 sur ergocycle ~ trois puissances

18 3/1. Rieu
diff6rentes (79~ S(VE); 94o70 S(~/E); 110~ S(VE)) observe des valeurs de [LA] m nettement plus rile- vdes que ceIles de [LA] s mesurdes pr6alablement lors d ' un test par palier, d 'une minute d'intensit6 croissante au niveau S(VE) (Tableau I).
La condition physique semble aussi pouvoir influer sur cette rdpartition du lactate entre milieu intra et extracellulaire: en effet, pour un m~me effor t submaximal, [LA] m est moins dlevd apr~s entrMnement qu 'avant (Karlsson, 1972).
En outre, ~ l 'arr~t de l 'effor t les cindtiques de [LA] m e t de [LA]s ne sont pas superposables: la premi6re diminue brutalement et rapidement, la seconde s'dlevant encore pendant 3 - 5 min (les deux phdnom6nes &ant sans doute li6s) avant de dimi- nuer ensuite parall~lement l 'une et l 'autre de fa~on exponentielle (Diamant, 1968).
Les mdcanismes rdgissant la sortie de lactate hors de la fibre musculaire ne sont pas bien connus. Il semble n6anmoins qu 'h part ir d 'un certain niveau de concentration intracellulaire le phdnom6ne sature, la quantitd de lactate d6versde dans le sang par units de temps h partir des muscles en contrac- tion reste alors constante m~me lorsque [LA]m
continue d 'augmenter (Jorfeldt, 1978) (Fig. 10). Certains facteurs peuvent interfdrer avec ces dchan- ges de lactate soit en les facilitant soit en les contre- carrant . Le pH extracellulaire para~t jouer un r61e important en ce domaine" en effet, dans des expd- riences rdalisdes sur des muscles perfusds de gre- nouilles et stimulds dlectriquement, l 'appari t ion de lactate dans le liquide de perfusion est d 'autant plus importante que le p H de celui-ci est augment6 par un accroissement de la concentration en bicarbo- nate (Mainwood, 1975, Hirche, 1975). Ainsi dans les exercices de puissance dlevde, l 'appari t ion de l 'acidose pourrait en elle-m~me constituer un frein
l 'appari t ion du lactate dans le sang. D 'aut re part , lorsque l 'exercice n ' implique que
de petites masses musculaires, l 'espace de dilution extracellulaire est grand vis-a-vis de celui intracel- lulaire qui est h l 'origine de la production de lac- tate. De plus, le degrd d'activitd des diffdrentes fibres au sein d 'un m~me muscle, voire des diffd- rents muscles au sein d 'un mEme groupe, peut se rdvdler &re indgalement rdparti. Le sang effdrent de ce territoire n 'est alors que le mdlange de sang de provenance et donc de qualitds diverses.
Tableau I. Etude chez l'homme A partir de donn6es de la littdrature de la concentration du lactate dans le sang ([LA L mmol-1 -t) et dans le muscle ([LA] m mmol.kg- ' muscle) selon l'intensitd de l'effort (P) et le temps (t) h partir du ddbut
de l'exercice ofl le prdl~vement et la biopsie musculaire ont dtd effectu6s.
t [LA]ra [LA]s Protocole expdrimental Auteurs P (rain) (mmol.kg-I) (mmol.l-t)
B. Diamant Repos 3,0 + 1,5 1,4 + 0,3 Exercice sur bicyclette ergonomique (1968) Supra max ~puisant 3 menant ~ l'dpuisement en 3 min.
Pr61~vement: sang artdrialis6 au bout du doigt. Biopsie: m. quadriceps.
L. Jorfeldt (1978)
Repos 1,1 _+ 0,4 1,0 + 0,2 29 +_ 0,01070 ~/O 2 max 4 0,8 + 0,I 1,6 +_ 0,2
12 1,0 +_ 0,1 1,4 _+ 0,3 51 _+ 0,01o70 ~'O 2 max 4 1,8 + 0,5 2,8 _+ 0,3
12 0,7 _+ 0,3 2,4 + 0,3 71 + 0,01o7"o XrO 2 max 4 3,7 __ 0,4 4,9 _ 0,6
12 2,8 _+ 0,1 5,0 + 0,6 87 _+ 0,01% 4 0 2 max 4 9,3 +_ 1,4 6,7 + 1,0
12 11,8 _+ 2,4 9,0 + 1,5
H.J. Green (1983)
Repos 1,59 ___ 0,98 79~ + 5.,1 S(VE)
(50% VO 2 m.ax) 9 4,49 __ 2,05 1,29 + 0,56 94,3~ _+ 12 S(VE)
(60~ VO, max). 11 6,37 + 2,13 1,70 _ 0,27 107,707o + 4,1 S(VE)
(70o7o VO 2 max) 13 11,3 + 1,76 2,04 + 0,58
Exercices en crdneaux rdalisds ~t distance les uns des autres ~ quatre puissances d'effort diff6rentes sur ergocycle. Prdl6vement: artdriel fdmoral. Biopsie: m. vaste externe.
Trois tests d'efforts par palier de I min augmentds l'un apr6s l'autre de 16,3 W, avec arrfit de chacune des dpreuves h des pourcentages variables de S(VE) d~ter- min6 prdalablement. Prdl6vement: artdrialis6 (veine de la main). Biopsie: m. vaste externe.
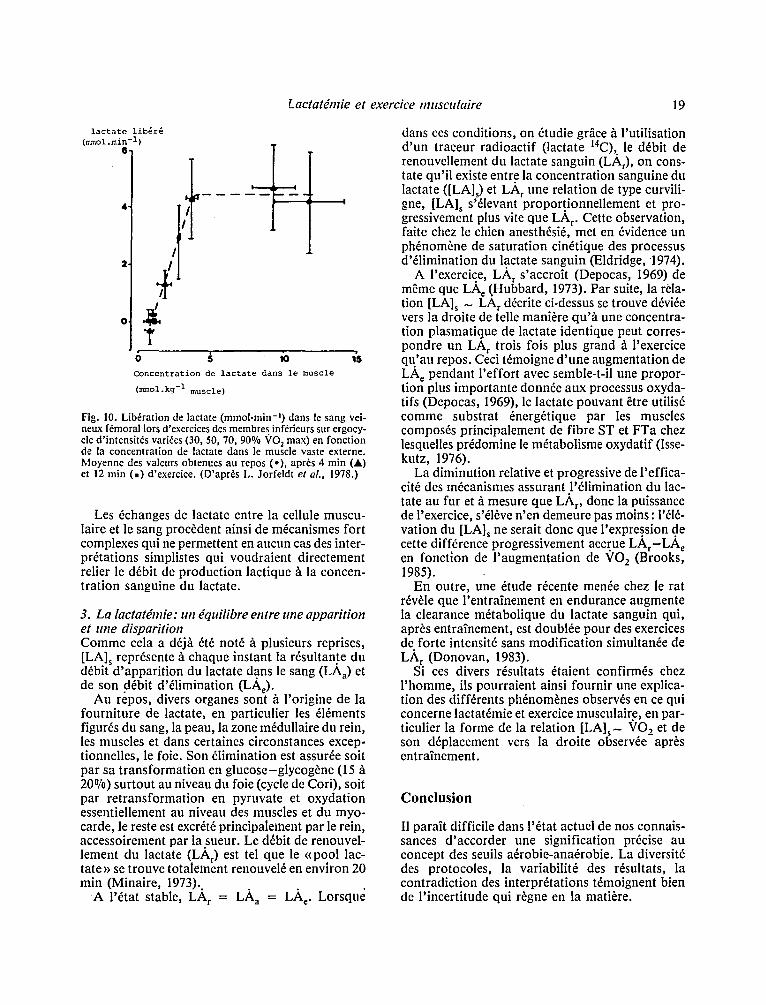
Lactat6mie et exercice mttsculaire 19
lactate lib~r~ (r .rain -I)
Concentration de lactate dans le muscle
(mm~ �9 kg -I muscle)
Fig. 10. Lib6ration de lactate (mmol.min-') dans le sang vei- neux f~moral Iors d'exercices des membres inf~rieurs sur ergocy- cle d'intensit6s vari6es (30, 50, 70, 90% %tO 2 max) en fonction de la concentration de lactate dans le muscle vaste externe. Moyenne des valeurs obtenues au repos (o), apr6s 4 min (A) et 12 rain (m) d'exercice. (D'apr~s L. Jorfeldt et aL, 1978.)
Les 6changes de lactate entre la cellule muscu- laire et le sang proc~dent ainsi de m6canismes fort complexes qui ne permettent en aucun cas des inter- pr6tations simplistes qui voudraient directement relier le d6bit de production lactique/l la concen- tration sanguine du lactate.
3. La laetatdmie: un dquilibre entre une apparition et une disparition Comme cela a d6j~ 6t6 not6 h plusieurs reprises, [LA] s repr6sente h chaque instant la r6sultante du d6bit d'apparition du lactate dans le s a n g (LAa) et de son d6bit d'61imination (LAe).
Au repos, divers organes sont h l 'origine de la fourniture de lactate, en particulier les 616ments figur6s du sang, la peau, la zone m6dullaire du rein, les muscles et dans certaines circonstances excep- tionnelles, le foie. Son 61imination est assur6e soit par sa transformation en glucose-glycog~ne (15/l 20%) surtout au niveau du foie (cycle de Cori), soit par retransformation en pyruvate et oxydation essentiellement au niveau des muscles et du myo- carde, le reste est excr6t6 principalement par le rein, acCessoirement par la sueur. Le d6bit de renouvel- lement du lactate (LAr) est tel que le <<pool lac- tate>> se trouve totalement renouvel6 en environ 20 min (Minaire, 1973)..
A l'6tat stable, LA r = L,A, a = LA e. Lorsque
dans ces conditions, on &udie grace h l'utilisation d 'un traceur radioactif (lactate 14C),. le d6bit de renouvellement du lactate sanguin (LAr) , o n cons- tate qu'il existe entre la concentration sanguine du lactate ([LA]s) et LA rune relation de type curvili- gne, [LA] s s'61evant proportionnellement et pro- gressivement plus vite que L A r. Cette observation, faite chez le chien anesth6si6, met en 6vidence un ph6nom6ne de saturation cin6tique des processus d'61imination du lactate sanguin (Eldridge, 1974).
A l'exercice, LA r s'accroTt (Depocas, 1969) de mfime que LA e (Hubbard, 1973). Par suite, la rela- tion [LA]s - LA r d6crite ci-dessus se trouve d6vi6e vers la droite de telle mani~re qu'h une concentra- tion plasmatique de lactate identique peut corres- pondre un LA r trois fois plus grand it l'exercice qu 'au repos. Ceci t6moigne d 'une augmentation de LA e pendant l 'effort avec semble-t-il une propor- tion plus importante donn6e aux processus oxyda- tifs (Depocas, 1969), le lactate pouvant ~tre utilis6 comme substrat 6nerg6tique par les muscles compos6s principalement de fibre ST et FTa chez lesquelles pr6domine le m6tabolisme oxydatif (Isse- kutz, 1976).
La diminution relative et progressive de l 'effica- cit6 des m6canismes assurant l'61imination du lac- tate au fur et/l mesure q u e L,A~ r, donc la puissance de l'exercice, s'61~ve n'en demeure pas moins: 1'616- vation du [LA] s ne serait donc que l'expression de cette diff6rence progressivement accrue L A r - L A e en fonction de l 'augmentation de ~'O 2 (Brooks, 1985).
En outre, une &ude r6cente men6e chez le rat r6v~le que l'entraTnement en endurance augmente la clearance m6tabolique du lactate sanguin qui, apr~s entrainement, est doubl6e pour des exercices de forte intensit6 sans modification simultan6e de LA r (Donovan, 1983).
Si ces divers r6sultats 6taient confirm6s chez l 'homme, ils pourraient ainsi fournir une explica- tion des diff6rents ph6nom~nes observ6s en ce qui concerne lactat6mie et exercice musculaire, en par- ticulier la forme de la relation [LA] s - VO 2 et de son d6placement vers la droite observ6e apr~s entraTnement.
Conclusion
I1 para]t difficile dans l'6tat actuel de nos connais- sances d 'accorder une signification pr6cise au concept des seuils a~robie-ana6robie. La diversit6 des protocoles, la variabilit6 des r6sultats, la contradiction des interpr6tations t6moignent bien de l 'incertitude qui r6gne en la mati6re.

20 M. Rieu
L'assimilation longtemps faite ent red6vat ion de la concentration du lactate dans le sang et ana6ro- biose est actuellement controvers6e. En effet, l'acc6- 16ration de la production d'acide lactique est in61uctable d~s le moment off apparait un accrois- sement de la glycolyse dont l 'augmentation ne sem- ble pas n6cessairement li6e ~ une insuffisance d ' appor t en oxyg6ne. D'ailleurs, on sait que la rela- tion VO 2 - puissance de l'exercice (P) reste lin6aire pour des valeurs de P tr+s sup6rieures au t<seuil ana6robie>>, ce qui ne devrait pas fitre le cas si l 'on admet que celui-ci marque l 'intensit6 de l ' e f for t h partir de laquelle le m6tabolisme ana6ro- bie dolt obligatoirement intervenir pour rendre pos- sible la r6alisation d 'un exercice de puissance sup6rieure h cette valeur << seuil>>.
De m~me peut-on s ' interroger sur le bien-fond6 des protocoles des tests utilis6s pour la d6termina- tion de ces seuils car est-il justifi6 de se servir de r6sultats obtenus h partir de sdquences d'exercices brefs et r6p6t6s, dont les effets de type << phase tran- sitoire>> se cumulent, pour tirer des conclusions concernant l 'aptitude d 'un sujet h maintenir un effort isol6 et prolong6?
En outre, la mesure d 'un param6tre sanguin (qui n'est lui-m~me que l'expression d 'un 6quilibre entre des d~bits d 'appari t ion et de disparition dans le sang) h intervalle de temps relativement long ne per- met gu6re d 'analyser une cin6tique et reste donc d ' interpr6tation difficile.
II n 'en reste pas moins que la relation lacta- t6mie - puissance de l'exercice se modifie avec l'entra~nement physique. Ce fait exp6rimental pour- rait trouver son explication dans une production moindre d'acide lactique chez le sujet entrain6 pour des efforts de mfime intensit6 avant et apr6s entra]- nement. Mais en ce cas, la consommation d 'oxy- g~ne pendant l 'effort consid6r6 devrait 6tre plus 6lev6e apr6s l 'entrainement qu 'avant , or un tel ph6- nom~ne n ' a jamais fit6 constat6 jusqu'i t ce jour.
Une autre hypoth6se avanc6e dans une p6riode r6cente serait que l 'entra~nement augmente l 'acti- vit6 des m6canismes responsables de l'61imination du lactate sanguin. Mais cette conception ne r6pond que partiel lement/l l 'object ion 6mise ci-dessus.
C 'es t plus vraisemblablement vers l '6tude des m6canismes d'6changes entre la cellule musculaire et le milieu extracellulaire et vers celle des compar- timents de distribution du lactate, comme le sug- g6rent les travaux de Zouloumian, qu' i l faut s 'orienter si l 'on souhaite donner des bases scienti- fiques solides aux d~marches empiriques couram- ment entreprises dans le domaine de la biologic du sport.
Bibliographie
Astrand P.O. (1967) Diet and athletic performance. Fed. Proc. 29, 1772-1777
Baldwin K.M., Campbell P.J. & Cooke D.A. (1977) Glycogen lactate and alanine changes in muscle fiber types during graded exercise. J. Appl. PhysioL 43, 288-291
Bang O. (1936) The lactate content of the blood during and after exercise in man. Skand. Arch. PhysioL 74 (Suppl.)
Bergstrom J., Hermanssen B., Hultman E. & Saltin B. (1967) Diet, muscle glycogen and physical perfor- mance. Acta Physiol. Scand. 71, 140-150
Brooks G.H. (1985) Anaerobic threshold: review of the concept and direction for future research. Med. Sci. Sports. Exercise 17, 22-31
Caiozzo V.J., Davis J.A., Ellis J.F., Azus J.L., Vanda- griff R., Prietto C.A. & MacMaster W.C. (1982) A comparison of gas exchange indices used to detect the anaerobic threshold. J. AppL PhysioL 53, 1184-1189
Canconi F., Ferrari M., Ziglio P.G., Droghetti P. & Codega L. (1982) Determination of the anaerobic threshold by a non invasive field test for runners. J. AppL Physiol. 52, 869-873
Cerretelli P., Shindell D., Pendergast D.P., Di Prampero P.E. & Rennie D.W. (1977) Oxygen uptake transients at onset and offset of arm and leg work. Respir. Physiol. 30, 81-97
Cerretelli P., Pendergast D., Paganelli W.C. & Rennie D.W. (1979) Effects of specific muscle training on VO2 on response and early blood lactate. J. AppL PhysioL 47, 761-769
Clode M., Clark T.J.H. & Campbell E.J.M. (1967) The immediate CO 2 storage capacity of the body during exercise. Clin. Sci. 32, 161-165
Connet R.J., Gayeski T.E.J. & Honig C.R. (1984) Lac- tate accumulation in fully aerobic working dog graci- lis muscle. Am. J. Physiol. 246, HI20-HI28
Costill D.L. (1970) Metabolic responses during distance running. J. Appl. PhysioL 28, 251-255
Crescitelli F. & Taylor C. (1944) The lactate response to exercise and its relationship to physical fitness. Am. J. Physiol. 141,630-640
Davis J.A. (1985) Anaerobic threshold: review of the concept and directions for future research. Med. Sei. Sports. Exercise 17, 6-18
Davies C.T.M. & Crockford G.W. (1971) The kinetic of recovery oxygen intake and blood lactic concentration measured to a baseline of mild steady work. Ergono- mics 14, 721-731
Davis J.A., Vodak P., Wilmore J.H., Vodak J. & Kurtz P. (1976) Anaerobic threshold and maximal aerobic power for three modes of exercise. J. AppL PhysioL 41, 544-550
Davis J.A., Frank R.H., Whipp B.J. & Wasserman K. (1979) Anaerobic threshold alterations caused by endu- rance training in middle aged men. J. Appl. PhysioL 46, 1039-1046

Lactatdmie et exercice musculaire 21
Denis C., Dormois D. & Lacour J.R. (1984) Endurance training, VO 2 max and OBLA: A longitudinal study of two different age groups. Int. J. Sports Med. 5, 167-173
Depocas F., Minaire Y. & Chatonnet J. (1969) Rates of formation and oxydation of lactic acid in dogs at rest and during moderate exercise. Can. J. PhysioL Phar- macoL 47, 603-610
Diamant B., Karlsson J. & Saltin B. (1968) Muscle tis- sue lactate after maximal exercise in man. Acta PhysioL Scand. 72, 383-384
Di Prampero P.E. (1981) Energetics of muscular exer- cise. Rev. PhysioL Biochem. PharmacoL 89, 143-222
Di Prampero P.E., Cerretelli P.E. & Piiper J. (1970) Lac- tic acid formation in gastrocnemius muscle of the dog and its relation to O z debt contraction. Respir. PhysioL 8, 347-353
Donovan C.M. & Brooks G.A. (1983) Endurance trai- ning affects lactate clearance not lactate production. Am. J. PhysioL 244, E83-E92
Douglas C.G. (1927) Respiration and circulation with variations in bodily activity. Lancet I, 213-218, 265-269
Duvallet A., Verdier J.C., Thieulart L., Giraud- Chauveau M. & Rieu M. (1985) Cin&ique de la lacta- t6mie au cours d'exercices sous-maximaux d'intensit6 variable. J. PhysioL (Paris) 80, 2A
Eldridge F.L. (1975) Relationship between turnover and blood concentration in exercising dogs. J. Appl. PhysioL 39, 231-234
Eldridge F.L., T'so L. & Chang H. (1974) Relationship between turnover rate and blood concentration of lac- tate in normal dogs. J. AppL PhysioL 37, 316-320
Embden G., Lehnartz E. & Henschel H. (1927) Der zeit- liche Verlauf der Milchs/iurebildung bei der Muskel- kontraktion. Hoppe Seyler's Z. Physiol. Chem. 65,255
Essen B. (1978) Glycogen depletion of different fibre types in humanskeletal muscle during intermittent and continuous exercise. Acta PhysioL Scand. 103, 446-455
Farrel P.H., Wilmore J.H., Coyle E.F., Billing J.E. & Costill D.L. (1979) Plasma lactate accumulation and distance running performance. Med. Sci. Sports 11, 338-344
Fletcher W.M.. & Hopkins F.G. (1906) Lactic acid in amphibian muscle. J. PhysioL (London) 35,247-309
Fouillot J.P., Duvallet A., Gaillard M.T. & Rieu M. (1976) Etude de la relation entre la lactat6mie et la puis- sance a6robique selon la nature de l'activit6 physique pratiqu~e. J. Physiol. (Paris) 72, 7A
Gleser M.A. & Vogel J.A. (1973) Endurance capacity for prolonged exercise on the bicycle ergometer. J. AppL Physiol. 34, 438-442
Gollnick P.D., Piehl K. & Saltin B. (1974) Selective glyco- gen depletion pattern in human muscle fibres after exercise of varying intensity and a varying pedalling rates../. PhysioL (London) 24, 45-57
Green H., Houston M., Thomson J. & Reid P. (1979) Alterations in ventilation and gas exchange during exercise induced carbohydrate depletion. Can J.' PhysioL PharmacoL 57, 615-618
Green H.Z., Hughson R.L., Worr G. & Ranney D.A. (1983) Anaerobic threshold blood lactate and muscle metabolites in progressive exercise. J. AppL PhysioL 54, 1032-1038
Greene N.M. (1961) Effect of epinephrine on lactate pyruvate and excess lactate production in normal human subjects. J. Lab. Clin. Med. 58, 682.
Hagberg J.M., Coyle E.F., Carroll J.E., Miller J.M., Martin W.H. & Brooke M.H. (1982) Exercise hyper- ventilation in patients with McArdle's disease. J. AppL PhysioL 52, 991-994
Hagenfeldt L. & Wahren J. (1972) Human foream mus- cle metabolism during exercise VII. FFA uptake and oxydation at different work intensities. Scand. J. Clin. Lab. lnvest. 30, 429-436
Harken A.M. (1976) Hydrogen ion concentrations and oxygen uptake in an isolated canine hindlimb. J. AppL PhysioL 40, I - 5
Heck H., Mader A., Hess G., Mucke S., Muller R. & Hollmann W. (1985) Justification of the 4 mmol.l- lactate threshold, lnt. J. Sports Med. 6, 117-130
Heigenhauser G.J.F., Sutton J.R. & Jones N.L. (1983) Effect of glycogen depletion on the ventilatory res- ponse to exercise. J. AppL PhysioL 54, 470-474
Herbst R. (1929) Der Energieverbrauch bei sportlichen Leistungen. Sportiirztetagung, Frankfurt am Main, 1929; Fischer-Verlag, Jena, 1930
Hermansen L. & Vaage O. (1977) Lactate disappearance and glycogen synthesis in human muscle after maxi- mal exercise. Am. J. PhysioL 223, E422-E429
Hill A.V., Long G.N.H. & Lupton H. (1924) Muscular exercise lactic acid and the supply and utilization of oxygen. Part VII, Muscular exercise and oxygen intake. Proc. R. Soc. London Ser. B. 97, 155-167
Hirche H., Hombach V., Langohr H.D., Wacker U. & Busse J. (1975) Lactic acid permeation rate in working gastrocinemii of dogs during metabolic alkalosis and acidosis. Pflfigers Arch. 356, 209-222
Hollmann W. (1959) The relationship between pH-lactic acid-potassium in the arterial and venous blood, the ventilation (PoW) and pulse frequency during increa- sing spiroergometric work in endurance trained and untrained persons. In: Pan-American Congress for Sports Medicine, Chicago
Hubbard J.L. (1973) The effect of exercise on lactate metabolism. J. PhysioL (London) 231, 1-18
Huckabee W.E. (1958) Relationship of pyruvate and lac- tate during anaerobic metabolism. I. Effects of infu- sion of pyruvation on glucose and of hyperventilation. J. Clin. lnvest. 37, 244-254
Hughes E.F., Turner S.C. & Brooks G.A. (1982) Effects of glycogen depletion and pedaling speed on 'anaero- bic threshold'..L AppL PhysioL 52, 1598-1607
Hughson R.L. & Green H.J. (1982) Blood acid-base and lactate relationships studied by ramp work test. Med. Sci. Sports Exercise 14, 297-302
Issekutz B., William J.R., Shaw A.S. & Issekutz A. (1976) Lactate metabolism in resting and exercising dogs../. AppL PhysioL 40, 312-319
Ivy J.L., Whithers R.T., Handel P.J., Elger D.H. & Cos- till D.L. (1980) Muscle respiratory capacity and fiber

22 M. Rieu
type as determinants of the lactate threshold. J. AppL PhysioL 48, 523-527
Jones N.L. & Jurkowski J.E. (1979) Body carbon dioxide storage capacity in exercise. J. AppL PhysioL 46, 811-815
Jones N.L. & Ehrsam R.E. (1982) The anaerobic thres- hold. Exercise Sports Sci. Rev. 10, 49-83
Jobsis E.F. & Stainbsy W.N. (1968) Oxydation of NADH during contractions of circulated mammelian skeletal muscle. Respir. PhysioL 4, 292-300
Jor feldt L. (1970) Metabolism of L-(+ )-lactate in human skeletal muscle during exercise. Acta PhysioL Scand. SuppL 338, 5-67
Jorfeldt L., Juhlin-Dannfeit A. & Karlsson J. (1978) Lac- tate release in relation to tissue lactate in human ske- letal muscle during exercise. J. AppL PhysioL 44, 350-352
Jousselin E., Handschuh R. & Stephan H. (1984) D~ter- mination de la zone de transition a6robie-ana6robie et utilisation pratique pour l'entra3nement. Cindsiologie 23,301-304
Karlsson J. (1971) Lactate and phosphagen concentra- tions in working muscle of man. Acta. PhysioL Scand. SuppL 358, 1-72
Karlsson J., Nordesjo L.O., Jorfeldt L. & Saltin B. (1972) Muscle lactate ATP and CP levels during exercise after physical training in man. J. AppL PhysioL 33, 199-203
Keul J., Doll J. & Keppler D. (1967) The substrate supply of the human skeletal muscle at rest during and after work. Experimentia 23,974-979
Keul J., Huber G. & Kindermann W. (1975) Unterschied- liche Wirkung des Skilanglaufes und des Skiabfahrts- laufes auf Kreislaug und Stoffwechsel. Sportarzt Sportmed. 26, 49-58
Keul J., Simon G., Berg A., Dickhuth H.H., Goertler I. & Kubel R. (I 979) Bestimmung der individuellen anae- roben Schwelle zur Leistungsberwertung und Trai- ningsgestaltung. Dtsch. Z. Sportmed. 30, 212-218
Kindermann W., Huber G. & Keul J. (1975) Aerobe Energiebereitsellung und Herz frequenz w~ihrend und nach verscheidenen trainings Methoden des Mittel- trecklers. Leitungssport 5, 66-70
Kindermann W., Simon G. & Keul J. (1979) The signifi- ciance of the aerobic-anaerobic transition for the determination of work load intensities during endu- rance training. Fur. J. AppL PhysioL 42, 25-34
Kindermann W., Schramm M. & Keul J. (1980) Aerobic performance diagnostics with different experimental setting. Int. J. Sports Med. 1, 110-114
Koyal S.N., Whipp B.J., Hontsman D., Bray G.A. & Wassermann K. (1976) Ventilatory responses to the metabolic acidosis of treadmill and cycle ergometry. J. AppL PhysioL 40, 864-867
Kumagai S., Tanaka K., Matsura Y., Matsuzaka A., Hirakoba K. & Asano K. (1982) Relationships of the anaerobic threshold with 5 km, 10 km, and 10 mile races. Eur. J. Appl. PhysioL 49, 13-23
La Fontaine T.P., Londeree B.R. & Spath W.K. (1981) The maximal steady state versus selected running events. Med. Sci. Sports Exercise 13, 190-192
Lundsgaard E. (1931) Uber die Energetik der anaeroben Muskelkontraktion. Biochemistry 2, 233-322
Mader A., Liesen H., Heck H., Philippi H., Rost R~, Schurch P. & Hollmann W. (1976) Zur Beurteilung der sportartspezifischen Andauerleistungsf/~higkeit in Laber. Sportarzt Sportmed. 27, 80-88, 109-112
Mainwood G.W. & Worsley-Brown P. (1975) The effects of exti'acellular pH and buffer concentration on the efflux of lactate from frog sartorius muscle. J. Physiol. (London) 250, 1-~22
Margaria R., Edwards H.T. & Dill D.B. (1933) The pos- sible mechanism of contracting and paying the oxy- gen debt and the role of lactic acid in muscular contraction. Am. J. PhysioL 106, 689-714
Margaria R., Cerretelli P., Di Prampero P.E., Massari C. & Torelli G. (1963) Kintics and mechanism of oxygen debit contraction in man. J. AppL PhysioL 18, 371-377
Mazzeo R.S., Brooks G.A., Bundinger T.F. & Shoeller D.A. (1982) Pulse injection, 13C tracer studies of lac- tate metabolism in human during rest and two levels of exercise. Biomed Mass Spectrom. 9, 310-314
Meyerhof O. (1921) Die Energieumwandlung im Mus- kel. V. Milchs~iurebildung und mechanische Arbeit. Pfliigers Arch. Ges. Physiol. 191, 128
Minaire Y. (1973) Origine et destin6e du lactate plasma- tique. J. PhysioL (Paris) 66, 229-257
Mfiller E.A. (1943) Die Pulszahl als Kennzeichen fiir Stof- faustausch und Ermudbarkeit des arbeitenden Mus- kels. Arbeitsphysiologie 12, 271-280
Owles W.H. (1930) Alterations in the lactic acid content of the blood as a result of light exercise and associa- ted changes in the CO z combining power of the blood and in the alveolar CO2 pressure. J. PhysioL (Lon- don) 69, 214-237
Pendergast D., Cerretelli P. & Rennie D.W. (1979) Aero- bic and glycolytic metabolism in arm exercise. J. AppL PhysioL 47, 754-760
Pirnay F., Dujardin J., Deroanne R. & Petit J.M. (1971) Muscular exercise during intoxication by carbon monoxide. J. AppL PhysioL 31, 573-575
Poole D.C. & Gaesser G.A. (1984) Effects of continuous and interval training on lactate threshold and maxi- mal aerobic capacity. Med. Sci. Sports Exercise 16, 183-190
Powers S.K., Dodd S. & Garner R. (1984) Precision of ventilatory and gas exchange alterationsas a predic- tor of the anaerobic threshold. Eur. J. AppL PhysioL 52, 173-177
Reinhard V.P., Muller P.H. & Schmuelling R.M. (1979) Determination of anaerobic threshold by ventilation equivalent in normal individuals. Respiration 38, 36-42
Reybrouck T., Heigenhauser G.J.F. & Faullkner J.A. (1975) Limitation to maximum oxygen uptake during arm, leg and combined arm-leg ergometry. J. AppL Physiol. 38, 774-779
Ribeiro J.P., Fielding R.A., Hugues V., Black A., Bochese M.A. & Knuttgen H.G. (1985) Heart rate

Lactat~mie et exercice musculaire 23
break point may coincide with the anaerobic threshold and not the aerobic threshold. Int. J. Sports Med. 6, 220-224
Rieu M., Fouillot J.P.,_Duvallet A., Handschuh R. & Tekaia F. (1983) Fr6quence Cardiaque et exercice physi- que. Bull. Soc. Med. Paris 11, 65-69
Robinson S. & Harmon P.A. (1941) The lactic acid mechanism and certain properties of the blood in rela- tion to training. Am. J. PhysioL 132, 757-766
Saunders N.A., Leeder S.R. & Rebuck A.S. (1976) Ven- tilatory response to carbon dioxide in young athletes : a family study. Am. Rev. Respir. Dis. 113,497-502
Segal S.S. & Brooks G.A. (1979) Effect of glycogen depletion and work load on postexercise 02 consump- tion and blood lactate. J. Appl. Physiol. 47, 514-521
Simon O,, Berg A., Dickhuth H . H , Simon-Alt A. & Keul J. (1981) Bestimmung der anaeroben Schwelle in Abh/ingigkeit vom Alter und vonder Leutungsffihig- keit. Dtsch. Z. Sportraed. 32, 6-14
Sjodin B. & Jacobs I. (1981) Onset of blood lactate accu- mulation and marathon running performance. Int. J. Sports Recreation, 2, 23-26
Stegmann H., Kindermann W. & Schabel A. (1981) Lac- tate kinetics and individual anaerobic threshold. Int. J. Sports Med. 2, 160-165
Stegmann H. & Kindermann W. (1982) Comparison of prolonged exercise tests at the individual anaerobic threshold and the fixed anaerobic threshold of 4 mmol.1 -~ lactate, lnt. J. Sports Med. 3, 84-89
Suzuki Y. (1979) Mechanical efficiency of fast and slow twitch muscle fibers in man during cycling. J. AppL PhysioL 47, 263-267
Tanaka K. & Matsuura (1984) Marathon performance, anaerobic threshold and onset of blood lactate accu- mulation. J. AppL PhysioL 57, 640-643
Tanaka K., Matsuura Y., Matsuura A., Matsuzaka A., Hirakoba K., Kumagai, Sun S.O. & Asano K. (1984) A longitudinal assessment of anaerobic threshold and
distance running performance. Med. Sci. Sports Exer- cise 16, 278-282
Tanaka H. & Shindo M. (1985) Running velocity at blood lactate threshold of boys aged 6-15 years compared with untrained and trained young males. Int. J. Sports Med. 6, 90-94
Wasserman K. & Mcllroy M.B. (1964) Detecting the threshold of anaerobic metabolism. Am. J. Cardiol. 14, 844-852
Wasserman K., Van Kessel A. & Burton G.G. (1967) Interaction of physiological mechanisms during exer- cise. J. AppL PhysioL 22, 71-85
Wasserman K., Whipp B.J., Koyal S.N. & Beaver W.L. (1973) Anaerobic threshold and respiratory gas exchange during exercise. J. AppL PhysioL 35, 236-243
Wasserman K. & Whipp B.J. (1975) Exercise physiolo- gic in health and disease. Am. Rev. Respir. Dis. 112, 219-249
Whipp B.J., Davis J.H., Torres F. & Wasserman K. (1981) A test to determine parameters of aerobic func- tion during exercise. J. AppL PhysioL 50, 217-221
Woodson R., Wills R.E. & Lenfant C. (1978) Effect of acute and established anemia on 02 transport at rest, submaximal and maximal work. J. Appl. Physiol. 44, 36-43
Yeh M.P., Gardner R.M., Adams T.D., Yanowitz F.G. & Crapo R.O. (1983) 'Anaerobic threshold': problems of determination and validation..L Appl. Physiol. 55, 1178-1186
Yoshida T., Nagata A., Maro M., Takeuchi N. & Sada Y. (1981) The validity of anaerobic threshold determination by a Douglas bag method compared with arterial blood lactate concentration. Eur. J. Appl. PhysioL 46, 423-430
Zouloumian P. & Freund J. (1981) Lactate after exer- cice in man. Mathematical model. Eur. J. AppL PhysioL 46, 135-148









![Livret de l’utilisateur - docs.mesmateriaux.comSOTRALENTZ]Fiche... · Plastepur ®, une gamme complète en pré-traitement anaérobie et en épuration aérobie - soit par infiltration,](https://img.pdfslide.fr/doc/110x75/5ae3d2807f8b9a5d648e7526/livret-de-lutilisateur-docs-sotralentzficheplastepur-une-gamme-complte.jpg)



















