Embed Size (px)
Citation preview

Les animaux parasites
Introduction
Une daphnie? Non: un écosystème
Les êtres vivants produisent de la matière et de l'énergie (chaleur, travail...) selon le principe d'allocation d'énergie (ou allocation des ressources).
Les échanges d'énergie sont le moteur du fonctionnement des écosystèmes.
Ces échanges se font selon deux grands types de relations:
• - systèmes prédateur-proie; le prédateur tire son énergie de sa proie; • - systèmes parasite-hôte; le parasite tire son énergie de son hôte.
Pourquoi étudier le parasitisme en Biologie animale?
• Parce qu'il joue un rôle esentiel dans la régulation des populations; • Parce qu'il joue un rôle important dans la structuration des écosystèmes; • Parce qu'il joue un rôle important (souvent méconnu) dans l'évolution des espèces.
Définition du parasitisme.
La définition usuelle du parasitisme n'est pas satisfaisante: "Le parasitisme est un mode de vie dans lequel un organisme, le parasite, utilise un autre organisme, l'hôte, comme habitat et comme nourriture."
D'un point de vue écologique, on peut distinguer les différents types d'interaction entre espèces en fonction d'un critère de bénéfice et de la durée des interactions.
- Un système prédateur-proie est une interaction provisoire;
- Un système parasite-hôte est une interaction durable.
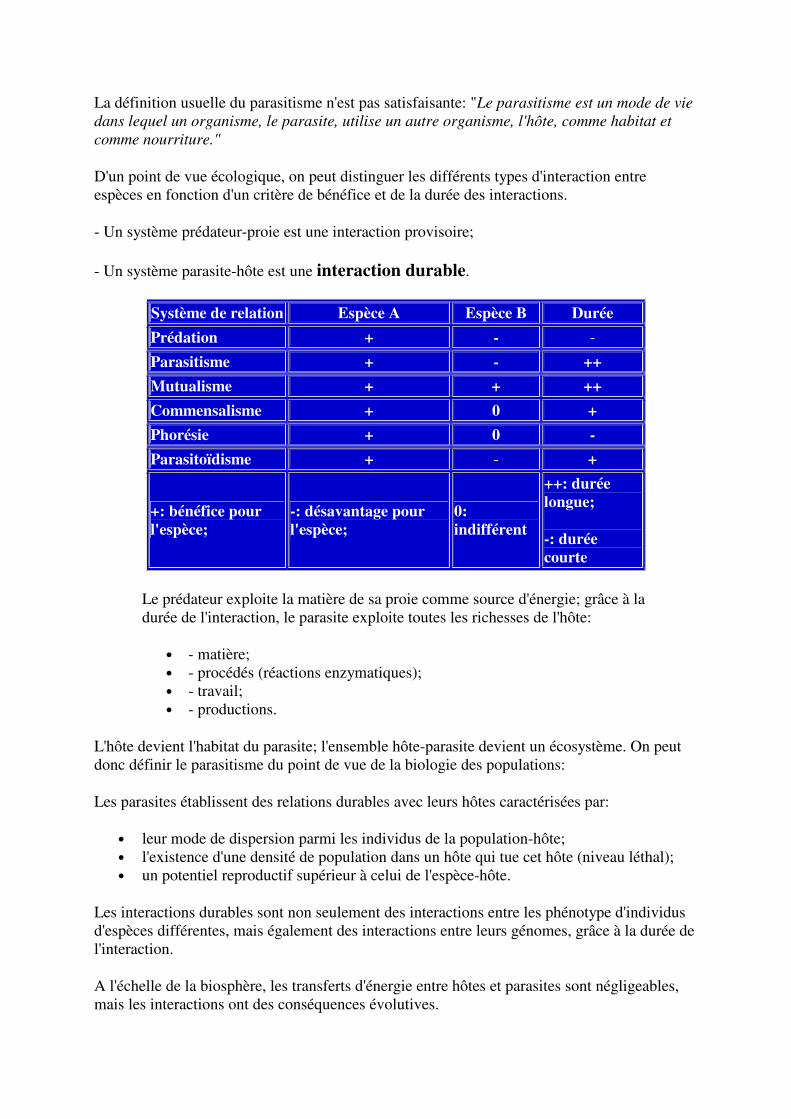
Système de relation Espèce A Espèce B Durée
Prédation + - -
Parasitisme + - ++
Mutualisme + + ++
Commensalisme + 0 +
Phorésie + 0 - Parasitoïdisme + - +
+: bénéfice pour l'espèce;
-: désavantage pour l'espèce;
0: indifférent
++: durée longue;
-: durée courte
Le prédateur exploite la matière de sa proie comme source d'énergie; grâce à la durée de l'interaction, le parasite exploite toutes les richesses de l'hôte:
• - matière; • - procédés (réactions enzymatiques); • - travail; • - productions.
L'hôte devient l'habitat du parasite; l'ensemble hôte-parasite devient un écosystème. On peut donc définir le parasitisme du point de vue de la biologie des populations:
Les parasites établissent des relations durables avec leurs hôtes caractérisées par:
• leur mode de dispersion parmi les individus de la population-hôte; • l'existence d'une densité de population dans un hôte qui tue cet hôte (niveau léthal); • un potentiel reproductif supérieur à celui de l'espèce-hôte.
Les interactions durables sont non seulement des interactions entre les phénotype d'individus d'espèces différentes, mais également des interactions entre leurs génomes, grâce à la durée de l'interaction.
A l'échelle de la biosphère, les transferts d'énergie entre hôtes et parasites sont négligeables, mais les interactions ont des conséquences évolutives.

L'existence de deux génomes côte à côte conduit à considérer le système parasite-hôte comme un super-organisme vivant possédant un super génome, ce qui a 4 conséquences:
1) le génome du parasite peut modifier le phénotype de l'hôte; exemple: les galles des végétaux sont dues à la sélection dans le génome d'insectes de gènes capables de manipuler la croissance des tissus végétaux. Le système hôte-parasite est un système de phénotypes croisés.
2) Dès qu'un système hôte-parasite se forme, il y a une probabilité pour que voisinent des gènes en double, d'où une tendance à la cessation de l'expression de l'un des gènes. La perte est en général subie par le parasite et entraîne des régressions de fonctions pouvant être assurées par le génome de l'hôte: simplification morpho-anatomiques (organes locomoteurs, digestifs, organes des sens); enzymes (déficience du métabolisme des purines chez les protozoaires parasites et les schistosomes. La survie de l'espèce parasite dépend strictement de l'hôte.
3) Le parasite peut apporter dans l'association un gène que l'hôte ne possède pas, mais dont les produits sont avantageux.
C'est l'enchaînement au parasite, rançon du mutualisme, avec coopération (piratage) génique entre les deux partenaires.
S'il y a dépendance physiologique unilatérale, on a affaire au parasitisme; si la dépendance physiologique est bilatérale, c'est le mutualisme. Le mutualisme est une association à "parasitisme réciproque".
4) L'association prolongée entre deux organismes de génomes différents peut favoriser les échanges géniques entre ces organismes.
Comment savoir si un organisme est parasite: le concept d'interaction durable lève toute équivoque.
Exemple:
• le moustique est hématophage, mais il n'y a pas d'interaction durable avec son hôte: ce n'est pas un parasite (l'étude de moustiques peut néammoins se justifier parce qu'ils peuvent être des vecteurs de parasites).
• La tique est un hématophage qui établit une interaction durable avec son hôte: c'est un parasite.

I. Étude d'un exemple: la sacculine
Sacculina carcini, parasite du crabe Carcinus moenas.
L'adulte forme un sac qui fait saillie entre l'abdomen et le céphalothorax du crabe adulte, attaché par un pédoncule. En fait, une partie essentielle du parasite est interne; il est représenté par des "racines" qui envahissent et se nourrissent à partir de tous les tissus.
Les Rhizocéphales sont des parasites d'autres crustacés exclusivement. Ils appartiennent à la classe des Cirripédes (balanes) à cause de la présence de larves cypris. Ces larves recherchent un substrat convenable pour y effectuer une métamorphose de fixation. Dans le groupe-frère des Thoraciques (Balanes), la métamorphose aboutit à la formation d'un adulte filtreur, microphage.
Le sac externe contient le ganglion nerveux, une cavité incubatrice, les ovaires, testicules (en fait des mâles transformés). L'ouverture cloacale permet la sortie des larves. L'adulte ne ressemble pas à un crustacé, mais les stades larvaires permettent de la classer parmi les Cirripèdes .
Le cycle de la sacculine peut être schématisé ainsi.

La sacculine externe
La sacculine est donc gonochorique (sexes séparés; différences morphologiques entre cypris mâle et femelle) et présente l'apparence d'un pseudohermaphrodisme. Les oeufs sont incubés dans la cavité palléale et les larves sont expulsées par l'orifice cloacal (or. de la cavité palléale). La plupart des organes ont disparus, sauf deux ganglions nerveux.

Les larves cypris présentent une segmentation typique des Crustacés, mis à part l'absence d'abdomen. Cette segmentation est contrôlée par l'activité de gènes homéotiques communs à l'ensemble des Métazoaires triblastiques. Il est à noter que, chez la sacculine, le gène abd A qui contrôle le développement de l'abdomen des autres crustacés est absent. D'autres gènes (engrailed) sont exprimés dans la région intermédiaire entre la furca et le thorax (vestige de l'abdomen). Le gène caudal s'exprime dans toutes les régions, sauf l'abdomen vestigial, ce qui pourrait être corrélé avec le développement défectueux de l'abdomen.
Les larves cypris sont sexuées, la larve mâle présente des différences avec la cypris femelle.

Différents stades larvaires de sacculine.
Larve cypris femelle
Formation de la larve kentrogone
Larve cypris mâle avec trichogone

Larve Trichogone récoltée dans la cavité palléale d'une femelle vierge.

Sacculine interne
La larve vermigon migre le long du tube digestif du crabe; elle forme un système de ramifications qui entourent les organes et prélèvent les molécules
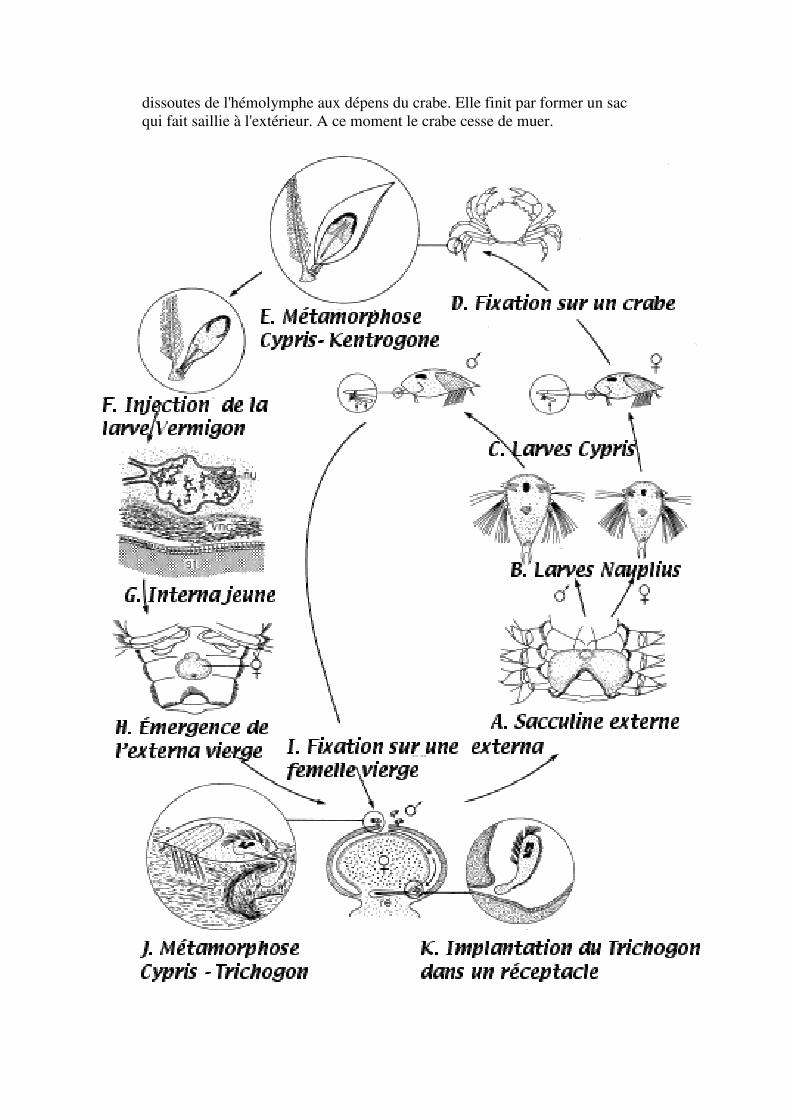
dissoutes de l'hémolymphe aux dépens du crabe. Elle finit par former un sac qui fait saillie à l'extérieur. A ce moment le crabe cesse de muer.
Conclusion
Plusieurs enseignements peuvent être tirés de l'étude de la sacculine:
• Le stade parasite (ici l'adulte) est profondément modifié par rapport à un Crustacé libre.
• Ces modifications concernent: o les structures de la vie végétative et de la vie de relation (A. digestif,
locomoteur, etc...) qui sont dégradées. o les structures liées à la reproduction (appareil et modalités de la
reproduction) qui sont développées. • Konrad Lorenz a créé le terme de sacculinisation pour décrire la réduction de
complexité (évolution rétrograde) de nombreux parasites en comparaison avec les formes libres.
• Cette dégradation ne se retrouve pas chez les stades libres des parasites, en particulier les stades infestants, particulièrement les structures sensorielles.
• Le parasite peut avoir un effet sur le phénotype de l'hôte.
Effets de la sacculine sur l'hôte.
Des modifications physiologiques plus ou moins considérables se produisent. La sacculine prélève sa nourriture sur le crabe, ce qui a pour effet, même lorsqu'elle est interne, de ralentir la croissance et le rythme des mues de l'hôte. Il n'y a pas à proprement parler de castration parasitaire (les racines ne pénètrent pas dans les gonades), mais elles sont privées de ressources énergétiques.
Chez la femelle, il n'y a plus de vitellogenèse ni de ponte; réduction des ovaires.
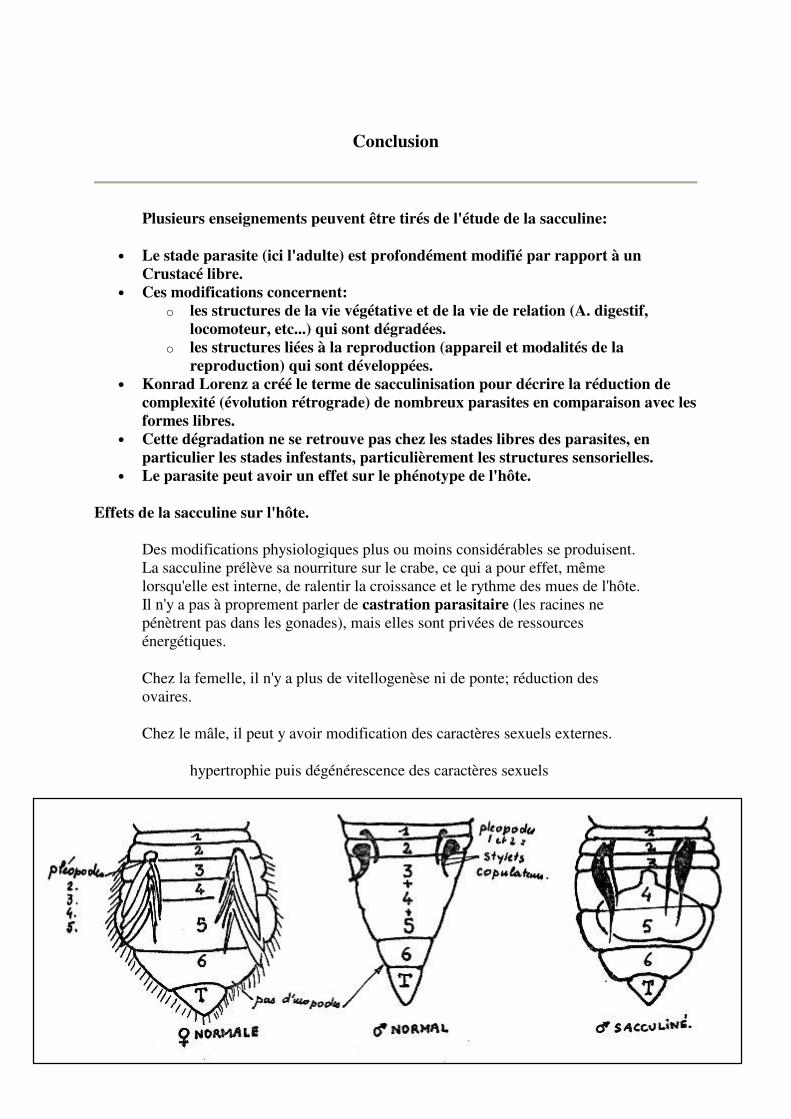
Chez le mâle, il peut y avoir modification des caractères sexuels externes.
hypertrophie puis dégénérescence des caractères sexuels

externes.
Chez Pachygrapsus marmoratus, transformation des testicules en ovaires.
Chez Carcinus moenas, réapparition de la segmentation abdominale, forme élargie rappelant l'abdomen de la femelle; dégénérescence des appendices copulateurs; apparition des pléopodes.
"Castration parasitaire" Féminisation des caractères sexuels
Le passage de la vie libre à la vie parasite.
Introduction.
Présence de parasites dans tous les groupes majeurs d'invertébrés, sauf les échinodermes. Certains groupes sont entièrement parasites comme les Trématodes, les Cestodes ou les Sporozoaires.
Vous devez connaître les parasites étudiés en cours de Biologie animale du DEUG B1:
• Trypanosome • Plasmodium • Douve • Schistosome • Ténia
Les ancêtres des organismes qui vivent en association avec d'autres organismes étaient libres.
1. Les facteurs qui permettent le passage de la vie libre à la vie parasite.

Habitat?
Milieu plus stable? Homéostasie des caractères physico-chimiques, mais existence des défenses de l'organisme: milieu pouvant être mortel.
Abri contre les prédateurs?
Pas entièrement vrai pour les ectoparasites:
• Ectoparasites de poissons ----> poissons nettoyeurs • Ectoparasites de Mammifères ---> oiseaux nettoyeurs
Pas entièrement vrai pour les endoparasites:
• existence d'hyperparasites, toutefois rares • soumis aux prédateurs des hôtes
mais le coût doit être supportable car l'espèce-proie avait survécu; de plus, le parasite peut tirer avantage de la consommation en devenant parasite du prédateur; le parasite est doublement gagnant.
Mobilité?
Le milieu animal est également mobile. Le parasite adulte étant fixe, la mobilité de l'hôte augmente la capacité de dispersion des formes de reproduction. Avantage de la vie parasitaire.
Énergie?

Le parasite tire son énergie de son hôte, sans le détruire. Il peut réduire son allocation d'énergie dévolues aux fonctions de recherche de nourriture.
Cumul: La majorité des parasites tirent avantage à la fois de l'habitat, de la mobilité et de la nourriture.
2. Les mécanismes du passage.
Les parasites doivent répondre à quatre caractères essentiels des milieux-hôtes:
• discontinuité dans l'espace (milieux en mosaïque) • discontinuité dans le temps (l'hôte est mortel) • existence de mécanisme immunitaires • évolution del'hôte
a) Discontinuité dans l'espace.
Dans le cas des parasites, les milieux en mosaïque nécessitent des stades spécialisés dans la transmission.
b) Discontinuité dans le temps
Ce n'est pas un apanage des parasites, mais les caractères "mortel" et "fragmenté" conjoints sont à l'origine des cycles parasitaires.
c) Hostilité du milieu
o Chaque organisme (parasite et hôte) est sélectionné sur le seul critère de la transmission de ses gènes.
o Le parasite exploite le milieu et n'a d'autre but que de se reproduire: il est contre-sélectionné si le coût de sa fécondité réduit le succès de sa transmission (effet pathogène).
o De même, l'hôte attaque le parasite, sauf s'il tire avantage de sa présence ou si le coût de la défense est supérieur au coût de la tolérance.
o Les parasites sont obligés de se défendre: sélection de stratégies d'évasion immunitaire.
d) Évolution du milieu
o Modifications anatomiques, physiologiques, du comportement. o Éclatement en populations distinctes ou espèces isolées o Changements de composition de la faune.
3. Répartition taxonomique et parenté des parasites.
Certains groupes taxonomiques sont entièrement constitués de parasites: Sporozoaires, Acanthocéphales.
D'autres sont en majorité libres, mais avec des espèces parasites: Cnidaires, Crustacés.

D'autres sont en majorité parasites: Plathelminthes.
Certaines espèces pratiquent un parasitisme réciproque:
• Les larves d'Unio (glochidium) parasitent les cyprinidés; • La bouvière Rhodeus amarus pond dans les Unio. • Crustacés parasites de crustacés (sacculine) • Poissons parasites de poissons • Oiseaux parasites d'oiseaux (coucous) • Insectes parasites d'insectes.
Le parasitisme est un mode de vie très répandu chez les animaux (comme chez les végétaux). Nous présentons ici une liste des principaux phylums, en indiquant la présence et l'extension du parasitisme.
• en rouge: groupes exclusivement parasites • en vert: groupes comprenant des espèces libres et des espèces parasites • en bleu: groupes sans parasites
Embranchement Classe Exemple
Protozoaires
Microsporidies Nosema apis
Flagellés Trypanosoma gambiense
Rhizopodes Entamoeba histolytica
Sporozoaires Plasmodium falciparum
Ciliés Balantidium coli
Métazoaires
Spongiaires
Cnidaires Cunina
Plathelminthes Turbellariés
Trématodes Fasciola hepatica
Cestodes Taenia solium
Nemathelminthes Ascaris suum
Mollusques Lamellibranches Anodonta cygnea (Paras. larvaire)
Gastéropodes Entoconcha
Céphalopodes
Annélides
Arthropodes Myriapodes
Arachnides Tique
Crustacés Sacculina carcini

Insectes (parasitoïdes)
Echinodermes
Vertébrés
Agnathes Myxine
Chondrichtyens
Osteichtyens (Parasitisme sexuel)
Batraciens
Reptiles
Oiseaux Coucou
Mammifères Homo sapiens (Géoparasitisme)
Chapitre3. Adaptations structurales et physiologiques.
Le parasitisme se traduit par deux catégories d'adaptations: des adaptations négatives, c'est à dire une dégradation de l'organisme parasite par rapport à l'organisme libre (nous en avons vu un certain nombre d'exemples dans le chapitre précédent: perte du tube digestif, perte des structures lies à la mobilité,etc...) et des adaptations positives, c'est à dire toutes les adaptations qui favorisent la vie parasitaire. Ce peut être des adaptations structurales (morphologiques, anatomiques), comportementales ou physiologiques.
A. Exemples d'adaptations structurales.
Les structures observées chez les organismes libres peuvent être profondément m/odifiées chez les parasites, aussi bien sur le plan morphologique qu'ultrastructural; en particulier les structures liées à la fixation du parasite sur l'hôte ou à la pénétration du parasite.
1. Protozoaires parasites.
Ils ont développé diverses sortes d'organelles spécialisés dans la fixation qui adhèrent aux cellules de l'hôte et à la glycocalyx qu'elles sécrétent. Certaines sont basées sur la production de contacts de type desmosomial: exemple de Crithidia fasciculata (Trypanosomatidae).
D'autres protozoaires parasites sont alternativement mobiles et sessiles et ont besoin d'un mécanisme d'attachement réversible: exemple de Giardia muris
Les protozoaires parasites intracellulaires se développent la plupart du temps dans des types cellulaires bien définis, chez des hôtes spécifiques. Les cellules hôtes restent généralement vivantes et fonctionnelles, bien que dans un état altéré. L'entrée de la forme invasive du parasite et son activité intracellulaires doivent être régulés de manière précise, afin d'éviter des dommages non contrôlés de la cellule-hôte. Les phénomènes qui se produisent à l'interface des

cellules ont donc une importance fondamentale dans l'association hôte-parasite.
Ils pénètrent dans la cellule-hôte par deux méthodes: soit par effraction à travers la membrane plasmique qui se reforme après leur passage, soit en détournant à leur profit les propriétés phagocytaires des cellules-hôtes. Dans ce cas, il y a souvent, mais pas toujours, formation d'une vacuole parasitophore.
Exemple de pénétration par effraction: les Microsporidies.
Exemple de formation de vacuole parasitophore: les Sporozoaires.
2. Les surfaces des Plathelminthes parasites.
Les Plathelminthes comprennent trois groupes dont toutes les espèces sont parasites à l'état adulte.
• les Monogènes: ils ressemblent beaucoup aux Turbellariés libres; ils sont ectoparasites à un seul hôte sur des Vertébrés poïkilothermes.
• Les Trématodes (digènes): la plupart sont endoparasites de Vertébrés; ils ont un cycle indirect avec une phase de reproduction asexuée sur un hôte intermédiaire (Mollusque).
• Les Cestodes: les plus modifiés par le parasitisme; ils n'ont plus de tube digestif; ils vivnet dans le tube digestif des Vertébrés.
(Il existe également des Turbellariés parasites)
Le tégument des Plathelminthes parasites est caractérisé par une organisation syncitiale.
o Structure du tégument
Origne de l'organisation syncitiale.
Exemple du schistosome
3. Les mécanismes de fixation.
Ils représentent l'un des aspects de l'interface hôte parasite auquel on s'est beaucoup intéressé. On a ainsi décrit une cinquantaine de catégories de modes d'attachement.
B. Adaptations physiologiques
1. Adaptations liées à la nutrition
L'étude des échanges nutritionnels dans une association parasitaire est un des aspects qui permet de comprendre ces associations. L'acquisition de la

nourriture a été l'un des facteurs permettant le maintien d'associations qui seraient autrement instables.
Nutrition des Protozoaires
• trypanosomes • amibe dysentérique
Nutrition des Métazoaires
• broutage de surface • hématophagie • ingestion de tissus • ingestion par la paroi du corps
2. Stratégies de reproduction
Une caractéristique liée au parasitisme: l'augmentation du potentiel biotique.
Les parasites utilisent des processus de reproduction leur permettant d'augmenter leur potentiel biotique.
Augmentation du nombre d'oeufs:
• Ascaris: production de 60*106 oeufs par an (1700 fois son poids). • Ténias: T. solium, 80*106 oeufs par an; T. saginata: 10*109 oeufs en 18
ans. Multiplication des appareils reproducteurs: 45000 proglottis chez Polygonoporus (ténia de cétacé, 40 m de long).
• Ankylostome: production de 1000 à 1500 oeufs par jour • Heterodera • Protozoaires: existence de cycles de multiplication à différentes phases
du cycle. • exemple du Plasmodium
Chez les espèces libres, le gonochorisme est le mode de reproduction le plus répandu. Par contre, la rencontre des sexes peut constituer un problème chez les parasites, en raison de leur dispersion, avec quelquefois un seul parasite par hôte. Plusieurs stratégies ont été adoptées par les parasites:
• exagération du dimorphisme sexuel. • hermaphrodisme • parthénogenèse • multiplication asexuée






























