Embed Size (px)
Citation preview

LES OSTRACODES ET LE PASSAGE PALI OCI NE-] OCENE
DANS LES CORDILLI RES BI TIQUES (COUPE DE CARAVACA, ESPAGNE)
CLAUDE G U E R N E T & EUSTOQUIO M O L I N A
GUERNET C. & MOLINA E. 1997. Les ostracodes et le passage Pal~oc~ne-Eocgne dans les Cordillgres b~tiques (Coupe de Caravaca, Espagne). [Ostracodes across the Paleocene-Eocene boundary in the Betic Cordillera (Caravaca section, Spain)]. [Los ostr4codos y la transicidn Paleoceno-Eoceno en las Cordilleras B6ticas (Corte de Caravaca, Espafia]. GEOBIOS, 30, 1: 31-43. Villeurbanne le 28.02.1997.
Manuscrit d~pos~ le 13.07.1995; accept~ d~finitivement le 05.09.1995.
RESUMt~ - Les associations d'ostracodes du Palfiocgne sup~rieur et de l'Eocgne de Caravaca (Sud de l 'Espagne) sont, au moins en partie, constituges d'esp~ces nouvelles dont Trachyleberidea rnarginata nov. sp., h la fois abondante et caract@istique. De mgme que Cytherella sp. 1 aft .navetensis, Bairdia cf. cymbula, Krithe sp. 1 et Oertliella ? sp. 1 aff. aculeata, T. marginata est prgsente sur toute la hauteur de la coupe. De possibles modifications de l'environne- ment, infralittoral fi gpibathyal, sont sugg@~es par la diminution de l'abondance des carapaces. Les esp~ces pal~oc~nes et ~ocgnes sont partiellement communes et la limite Palgoc~ne-Eoc~ne est donc, pour les ostracodes et en milieu int~alittoral ~ gpibathyal, moins tranchge que dans le NW de l'Europe.
MOTS-CL]~S: OSTRACODES, PALI~OCI~NE, I~OCENE, Tt~THYS, ESPAGNE, CORDILLt~RES Bt~TIQUES.
ABSTRACT - Late Paleocene to early Eocene associations of ostracodes from Caravaca (South of Spain) are, at least partially, constituted of new species of which Trachyleberidea marginata nov. sp. is at the same time abundant and characteristic. Cytherella sp. 1 aff. navetensis, Bairdia cf. cymbula, Krithe sp. 1, Oertliella ? sp. 1 affi aculeata and T. marginata are present throughout the section. Possible modifications of the infralittoral to epibathyal environment (sources of nutriment) are only indicated by the reduced abondance of carapaces. The Paleocene and Eocene species are partially the same, and therefore the Paleocene/Eocene boundary is, for the infralittoral to epibathyal environ- ment, less abrupt than in NW Europe.
KEYWORDS: OSTRACODES, PALEOCENE, EOCENE, TEHYS, SPAIN, BETIC CORDILLERA.
RESUMEN - Las asociaciones de ostrficodos del Paleoceno superior y del Eoceno inferior de Caravaca (Sur de Espafia) estfin, al menos en parte, constituidas de especies nuevas, tal como Trachyleberidea marginata nov. sp. que es a la vez abundante y caracterfstica. Cytherella sp. 1 aff. navetensis, Bairdia cf. cymbula, Krithe sp. 1, Oertliella ? sp. 1 aff. aculeata y T. marginata estfin presentes a lo largo de todo el corte. Las posibles modificaciones del medio ambiente, infralitoral a epibatial, s61o son sugeridas por la disminuci6n del mimero de caparazones. Las especies paleocenas y eocenas son parcialmente comunes, y pot tanto el lfmite Paleoceno/Eoceno es, para los ostr~codos de medio infralitoral a epibatial, menos marcado queen el NW de Europa.
PALABRAS CLAVE: OSTRACODOS, PALEOCENO, EOCENO, TETHYS, ESPAI~A, CORDILLERAS BI~TICAS.
INTRODUCTION
R6f6rence m o n d i a l e p o u r l '~ tude du p a s s a g e M~sozoYque-C~nozo[que, la coupe de C a r a v a c a es t celle d 'une s@ie m a r i n e con t inue du Cr~tac6 s u p @ i e u r EL l 'Eoc~ne moyen . Si tu6e dans la zone Subb~t ique de la Cordil lSre B~t ique (Fig. 1), elle a ~tfi d~cri te p o u r la p r e m i e r e fois p a r D u r a n d Delga & Magn~ (1958) et r66tudi~e p a r Von H i l l e b r a n d t (1974) p o u r les fo ramin i f e re s et p a r
Romein (1979) pou r le n a n n o p l a n c t o n calcaire. Le p a s s a g e Pal~oc~ne-Eocgne a fa i t l 'obje t d ' une ~tude s t r a t i g r aph ique , mic ropa l~on to log ique et g6ochimique p a r Mol ina et al. (1994) e t p a r Canudo et al. (1995). L '~tude des os t racodes du Pal~oc~ne h l 'Eoc~ne in f@ieur es t p r~sen t6e ici pour la p r e m i @ e fois.
Le Pal6oc~ne e t l 'Eoc~ne c o r r e s p o n d e n t h la F o r m a t i o n de J o r q u e r a (Van Veen 1969), c 'est-M

32
L CARAVACA ALICANTj
: a ravaca ~ v 1 K i n 6 5
2 MURCIE ~" o ¢z
' ~F Km 68 N
Cimeti6re .. mm ~ ) Cimeti6r~ ikm
~ Lorca // 1 I I
FIGURE 1 - Situation de la coupe de Caravaca. Location of the Caravaca section.
dire h 225 m de marnes al ternant avec des cal- caires gr6seux. Le Pal6oc6ne est principalement marneux, h intercalations de lits minces de cal- car6nites en sa partie sup6rieure. Le passage l'Eoc~ne se fait au sein de 20 m de marnes et argiles contenant deux niveaux rapproch6s h forte dissolution des carbonates. La base des argiles est gris sombre h noir et contient des organismes ben- thiques qui sugg6rent un 6v6nement anoxique (voir plus loin). L'Eoc6ne inf6rieur est constitu6 de calcaires gris jaune, de calcar6nites, de marnes et d'argiles interstrat i f i6s. Plusieurs horizons grands foraminif6res (nummulites, alv6olines, discocyclines) sont pr6sents dans les calcar6nites. Celles-ci pr6sentent fr6quemment des stratifica- tions obliques, "convolute laminations" et autres structures indiquant un niveau d'6nergie 61ev6. Le sommet de la coupe est principalement mar- neux.
Les marnes pal6oc6nes et 6oc6nes contie.nnent des foraminif~res planctoniques et du nannoplancton calcaire abondants et bien conserv6s. Les petits foraminif~res benthiques sont relativement peu communs tandis que les grands foraminif~res sont pr6sents seulement h certains niveaux. Les zones de foraminiFeres reconnues sent les P3b, P4 (pour le Pal6oc6ne), P6a, P6b, P6c et P7 (pour l'Eoc6ne
inf6rieur) de Berggren & Miller 1988 (Canudo et al. 1995). La zone P5 et la partie inf6rieure de la P6a n'ont pas 6t6 identifi6es du fait de l'existence d'un hiatus s6dimentaire r6v616 par la disparition simultan6e de 7 esp6ces et de l 'apparition subs6- quente de 4 autre esp6ces. Ce hiatus qui pourrait repr6senter 0,7 Ma (Molina et al. 1994) est plac4 2,5 m sous l'intervalle de dissolution qui marque la limite Pal6oc~ne-Eoc6ne d6finie h Alamedflla par la disparition de Igorina laevigata et de Subbotina velascoensis (Arenillas & Molina, sous presse). Cependant, cette limite n'a pas 6t6 d6finie officiel- lement et beaucoup de biostratigraphes la placent
l'extinction de M. velascoensis, soit entre la P6a et la P6b, tandis que d'autres la placent ~ l'extinc- tion de P. pseudomenardii qui d6fmit le sommet de la P4. La limite retenue ici est caract6ris6e par l'ex- tinction massive des petits foraminif~res ben- thiques (Ortiz 1994) et une chute du 613 C, ~v6ne- merits qui correspondraient h une 616vation du niveau de l a m e r et de la temp6rature des eaux profondes avec appauvrissement de celles-ci en oxyg~ne (Molina et al. 1994).
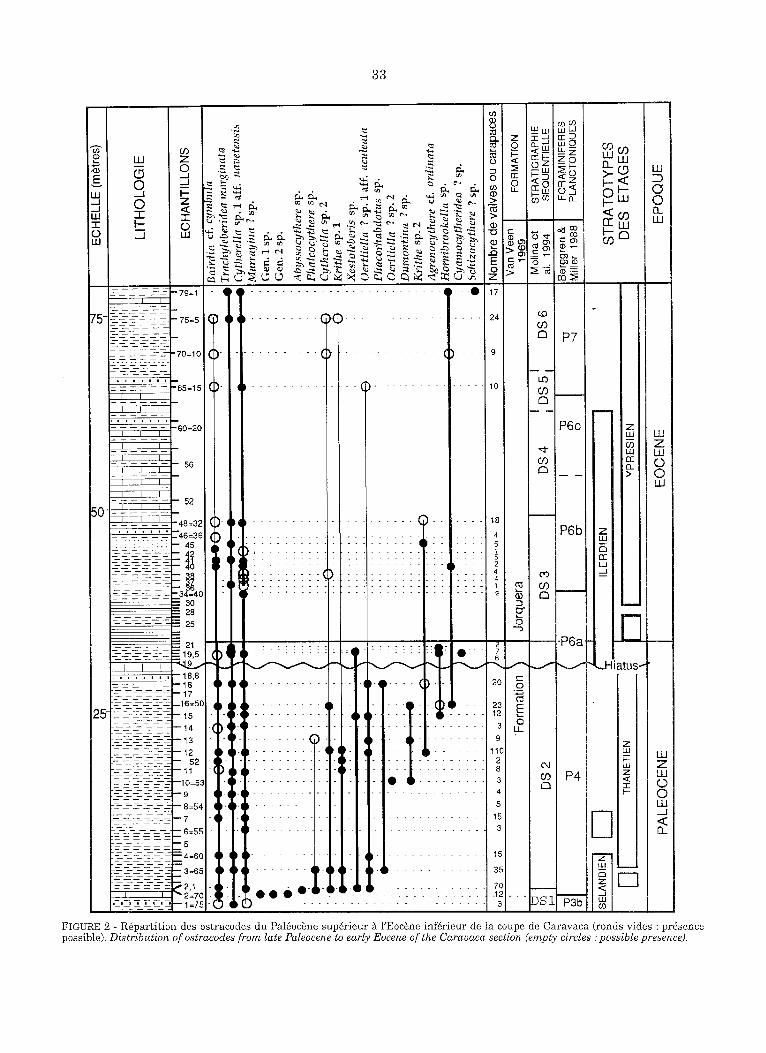
L'~tude pr6sent6e ici porte sur 80 m de s6diments dont 40 m, au voisinage de la limite entre les deux p~riodes, ont fair l'objet d 'un ~chantillonnage serr6 (Fig. 2). Les ostracodes - fr6quents dans le
33
u) cO r,,O e~ LuLU
u_~ '7) 0 3
-'~ _"' z = ~ ~ = ~ ~o ~ ,,, o . . ~ ~ ~ ~ ~o-~,,~_ ___,,,~-~
.~. ~ . ~ ~ ~ ~ - . , ~ ( - . ~ ~ , ~ E >
• _-- . . . . . . 79=1 ~ . . . . . . . . . . . . . . . . . . . . . . . . . . - - - • 17 I I I
Z5 - . - . . . . . . 75=5 i ( 9 ~, ~ 0 0 24 ~ o
. . . . . --- - - ~ P 7
- - - ------7o=~o ( ) - ( ) -( ) . . . . 9
LO " 6 5 : 1 5 ( ) . . . . . . . . . . . . . . . . ( ] ) . . . . . . . . . . . . . . . . . 10 O')
_ L _ L _ % _ !
I I I ~ - - - -
-~o=2o P 6 c z i i i w LLI
Z I i i ~ LLI LLI
L _ L _ ~_ co u: C.) - ~ I i - 5 6 C ' ~ 0.. . . . . . . . > " 0
I , I LU I I I
. . . . . . . 52
50 _ L _ L I I I 4 8 = 3 2 ( I - . ) ~ 0 18
-- - 4 6 = 3 6 ( ) ' , 4 P 6 b Zw
_ o ~ ~ o ' " 3O 28 ( 3 "
. . . . . . ~ 2, ,__ . . . . . . . . . . . . . . . . . . . . . . . I . . . . . . . . ~ . P 6 a - -
- I - - t - ] - ~LE . , , - ~ ~ - . - - ' " - - / ~ ~ - ~ , . . , ~ ' - - . ~ - . ~ " _Hiatus- - -
~.---::.~-z~ ,~ , ~ , ~ . . . . . . . . . . . . . . , , , , . . . . 4 > 1 ~ . . . . . 20 : ~ : - ~ - - ~ - - ~7 ~ "~ :~__---__--_-_-_~-~--16=50 , I ) ! I I . . . . . | ) - - ( I ) l | - - - 23 C::
2 5 : - £ : - 7 _ - - - - 1 5 - -~,-~ ~ - ~ ~2 .'.- - ~ - : = _ - - : - : - - ~ 4 - C , - ~ ,4 , 3 o
. . . . . . . . . . . . . . . . . ~3 '- O [ , ~ • ~ 9 z
:-~-!~-÷-~ ~2 :,-~ '",',:i, ~°~ - 1 - _ ~ . ~ ] : : : : : : i : [ [ [ [ _ ~ _ ~ : : L : : I I I 1 ~- LU
" ~ - - £ ] - - - ~ ~ - - - - I 0 = 5 3 " - - I - - . . . . . . . . . . . . . . . . . • i~ . . . . . . . . . . 3. co P 4 < z LU c::3 -r 0
Z L Z Z Z - 9 ( I - ~ - l - - . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4 ~ 0 . . . . . . . . 8 = 5 4 ( ) ' I H 5 LLI
1 - - - - - - - - - 7 - l , - -I 15 .=:[[ • - - - - - - - - - - - - 6 = 5 5 . . . . . . 4 - . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 . 13_
. . . . . . . . . 5
I - - - - - ~ - - - - - - - - - ? 2,~ '- , ~ - ; . . ~ - • , i , r , ~~ " '~ " 70 " - - - - 1 / I I I ~ 2 = 7 0 I _ . _ ~ - ~ ~ ~ . . . . . . . . . . . . . . . . . . . . . . . . . 12 . . I , ~ , ~ , ~ V _ , ~ , ~ , . - - - 1 = 7 5 , - 0 - ( ) ~... ) 3 DSI I P3b
FIGURE 2 - R@artition des ostracodes du Pal~oc~ne sup~rieur fl l'Eoc~ne inf~rieur de ta coupe de Caravaca (ronds vldes : presence possible). Distribution of ostracodes from late Paleocene to early Eocene of the Caravaca section (empty circles ." possible presence).

34
Pal6oc~ne et rares dans l'Eoc~ne - ont 6t6 pr61ev6s dans les fractions du s6diment de granulom6trie sup6rieure ~ 100 microm~tres.
SYSTl~MATIQUE Les observations syst6matiques portent sur les esp6ces les plus communes ou les plus significatives au point de vue stratigraphique ou 6cologique. Les autres esp6ces, repr6sent6es par un nombre r6duit de valves, sont n6anmoins figur6es; il s'agit de Cyamocytheridea sp., Krithe sp. 1, K. sp. 2, Gem 1 sp., Gen. 2 sp., Gen. 3 sp., Schizocythere sp., Xesto- leberis sp.
Ordre PODOCOPIDA Mfiller, 1894 Sous-ordre PLATYCOPA Sars, 1866
Famille CYTHERELLIDAE Sars, 1866
Genre Cythere l l a JONES, 1849
E s p 6 c e - t y p e - Cytherina ovata ROEMER, 1840.
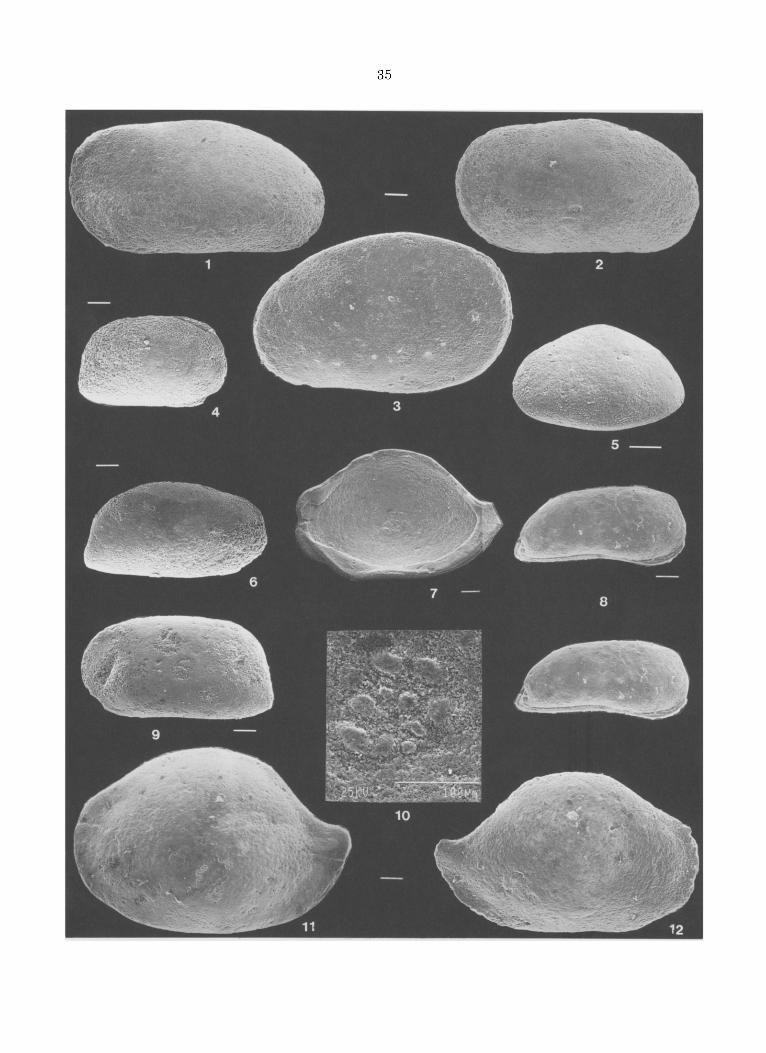
Cytherella sp.1 aff. navetensis BOLD, 1960 Fig. 3.1-3
A Caravaca, le Pal6oc6ne et l'Eec6ne inf6rieur contiennent deux esp6ces attribuables au genre Cytherella. C. sp. 1 appartient au groupe de C. lata BRADY, 1 8 8 0 , actuelle et bathyale, de C consueta DELTEL, 1963, de l'Eoc6ne d'Aquitaine, et de C. nave- tensis. Ces esp6ces poss6dent des valves tr~s dis- sym6triques: valve droite ~ bord dorsal tr~s arrondi plongeant vers l'avant pour se raccorder insensible- ment au bord ant6rieur, valve gauche ~ bord dorsal subhorizontal sauf en son extr6mit6 post6rieure off il plonge obliquement. Cependant le bord dorsal ant6rieur des valves droites de C. lata est nettement moins inclin6 que celui des valves droites de ]a Cytherella de Caravaca tandis que le bord dorsal des valves droites de C consueta est plus r6guli6re- ment arrondi. C'est, par ses contours, de C. naveten- sis que l'esp6ce de Caravaca semble la plus proche. Toutefois les rapports hauteur/longueur sont de 0,66 et 0,62 pour les valves droites de Caravaca sup- pos6es respectivement femelles et m~les, longues de 1,15 mm alors qu'ils sont de 0,61 et 0,55 pour les types de C. navetensis BOLD, 1960, longs de 1,5 ram, diff6rences peut-~tre li6es au caract6re juv6nile des valves recueillies ~ Caravaca.
Cytherella sp. 2, allong6e, ~ bords dorsal et ventral subparall~les, est repr6sent6e par de rares valves en m6diocre 6tat de conservation et de petite taille, peut-~tre larvaires. Elle n'est pas figur6e dans ce travail.
Sous-ordre PODOCOPA Sars, 1866 Superfamille BAIRDIACEA Sars, 1866
Famille BAIRDIIDAE Sars, 1888
Genre B a i r d i a McCoY, 1844 (=Paranes idea MADDOCKS, 1969)
E s p 6 c e t y p e - Bairdia curtus Mc CoY, 1844
Bairdia cf. cyrnbula DELTEL, 1964 Fig. 3.7,10-12.
Les contours et les dimensions des valves des Bairdia juv6niles recueillies ~ Caravaca, aussi bien dans le Pal6oc~ne que dans l'Eoc6ne, sere- blent 8tre ceux de Bairdia cymbula DELTEL, 1964 et notamment de l'holotype. Cependant, les figu- rations originales des types sont anciennes et d'antre part, dans les 6chantillons de Caravaca, la seule valve pr6sum6e adulte, longue de 1,5 ram, est m6diocrement conserv6e: l 'at tr ibution sp6ci- fique est donc incertaine. B. cymbula fur d6crite Peyrehorade (coupe de Lespontes) , dans le Lut6tien du SW de la France (Deltel 1964, p. 139- 140, fig. 21-22, pl. 2 et Deltel 1961 pour la locali- sation g6ographique). Elle est cit6e par son auteur dans le Lut6tien et le "L6dien" ou le "Bartonien' .
Superfamille CYTHERACEA Baird, 1850 Famil]e TRACHYLEBERIDIDAE sensu Sylvester-
Bradley, 1948
Genre T r a c h y l e b e r i d e a BOWEN, 1953
E s p 6 c e - t y p e - Cythereis prestwichiana JONES • SHERBORN, 1887
Les caract6ristiques g6n6riques de T prestwichia- na sont les suivantes (d'apr~s Bowen 1953, 1963; Hazel 1965; Moos 1967; Colin & Babinot 1979 et des observations personnelles de topotypes) : - carapace de longueur g6n6ralement 6gale ou inf6rieure ~ 0,8 ram, aplatie, subtr iangulaire en rue lat6rale, ~ extr6mit6 post6rieure plus ou moins anguleuse ; - bourrelet ant6romarginal se prolongeant ventra-
FIGURE 3 - 1-3. Cytherella sp. 1 aff. navetensis, respect ivement valves gauches male et femelle (Caravaca 12) et valve droite femel- le (Caravaca 2,1). 4,6,9. Krithe sp. 1, respect ivement valve dreite larvaire (femelle?) et valves droite et gauche suppos6es adultes (Car. 12). 8. Krithe sp. 2, carapace en r ue lat6rale droite (Car. 12). 5. Cyamocytheridea ? sp., valve gauche (Car. 19,5). 7,10-12. Bairdia cf. cymbula, respect ivement valve droite immature en r u e in terne (Car. 54) et ses empreintes musculaires, valves gauche (Car. 1,2) et droite (Car. 12). Barre = 100 microm~tres. 1-3. Respectively male and female left valves and female right valve. 4,6,9. Respectively larval right valve (female?) and presumed mature right and left valves. 8. Carapace, right lateral view. 5. Left valve. 7,10-12. Respectively larval right valve (inside view) and its central muscle scar, left and right valves.
35

36
lement en une c6te plus ou moins marquee et se relevant b rusquement avant l'extr~mit6 post~- rieure; c6te m~diodorsale plus ou moins sinueuse, s ' aba i ssan t subver t i ca lement vers l 'arri~re; absence de c6te m~diane ne t tement diff~renci~e ;
tubercule oculaire faiblement d~velopp~ ou absent ; - surface grossi~rement r~ticul~e avec tendance la c~lation, tubercule subcentral ~ l 'emplacement des empreintes de muscles adducteurs et pores normaux simples (parfois pores-conuli), notam- ment aux noeuds r~ticulaires ; - ~pines marginales ant~rieures et post~rieures irr~guli~rement d~velopp~es ; - caract~res internes comme Trachyleberis sauf que l 'empreinte frontale est d~doubI~e et que la dent cardinale post~rieure semble ~tre lob~e; les quatre empreintes de muscles adducteurs sont interconnect~es.
Le genre Trachyleberidea est, selon Har tmann et Pur i (1974), synonyme de Recto trachy leber i s RUGGmR% 1952 (=Costa NEWAN% 1928); en fait, il se distingue ne t tement de ce second genre par sa forme triangulaire, la presence d'un tubercule subcentral et l 'absence de c6te m~diane bien d~fi- nie. D u m o n t i n a DEROO, 1966 et Haze l ina Moos, 1966, genres synonymes selon Har tmann et Puri, 1974, sont plus quadrangulaires, avec une extr~- mit~ post~rieure plus arrondie.
Outre T pres twich iana , de l'YprSsien des Bassins de Londres et de Belgique (Keij, 1957), les esp~ces su ivan tes sont ra t tach~es au genre Trachy- leberidea :
T geini tz i (REuss, 1874) du Turonien et du S~nonien de l 'Europe (Babinot 1980) ; - T a landa lusens i s RE:~ENT, 1984 du Turonien d'Espagne (Reyment 1984) ; - T vezerae COLIN, 1973 du Turonien du Bassin d'Aquitaine (France) ; - T arta (DA~OTTE, 1971) du C~nomanien et du Turonien de l 'Europe (Babinot 1980) ; - T sp. du Santonien des Corbi~res (Tambareau et al. 1986, p. 25 et pl. 2, fig, 7) ; - T acuti loba (M~ssoN, 1880) du Campanien et Maestrichtien du Nord de l 'Europe (Deroo 1966; Herrig 1966) ;
T sp. du Campanien sup~rieur du NE de l 'Espagne (Garcia Zarraga & Rodriguez Lazaro 1990). - T. tunis iensis COHN & SAID, 1982, du Campanien-
Maestrichtien du Maghreb (in Donze et al. 1982; Damotte & Fleury 1987) ; - T sp. de l 'Ilerdien des Corbi~res (Tambareau et al. 1986, p. 36) ;
T sp. cf. pres twich iana (T pres tw ich iana in Ducasse et al., 1985) des marnes de Saint-Geours (Ypr~sien?) du bassin d'Aquitaine, cit~e jusque dans l'Oligoc~ne du m~me bassin ; - T posteroacuta (LmNEN~LAUS, 1900) de l'Oligo- c~ne de l'Allemagne du Nord (in Moos, 1966) ; - T blanpiedi (HOWE & LAw, 1936) de l'Oligoc~ne d'Am~rique du Nord (Hazel et a1.1980) ; - T pret iosa LEVINSON, 1974, du Pleistocene circa- littoral ~ bathyal du golfe du Mexique (Leroy & Levinson 1974).
Le genre Trachyleberidea apparai t ra i t donc au Cr~tac~ sup~rieur. I1 semble dispara~tre d'Europe
la fin du Pal~og~ne mais il est actuellement commun "along the Gulf and southeast Atlantic coasts", o~ il est represent6 par T pre t iosa (BENsON in Leroy & Levinson 1974, p. 22). Selon Babinot & Colin (1979) les espSces du genre Trachyleberidea ont migr~ progressivement d'un envi ronnement inf ra l i t tora l (~ hu~tres ou rudistes) au C~nomanien-Turonien ~ des environ- nements beaucoup plus profonds au Tertiaire, avec r~duction ou disparition concomitantes des tubercules oculaires. Cependant, T pres tw ich iana est cit~e par Keij (1957, tabl. 1, p. 27) dans les sables glauconif'eres de l'Ypr6sien du Bassin de Bruxelles, en association avec une ostracofaune caract~re infral i t toral . En r~sum6, le genre Trachy leber idea peut ~tre considSr~ comme caract~ristique, au Pal~og~ne, de milieux oc6a- niques calmes, infralit toraux ~ bathyaux.
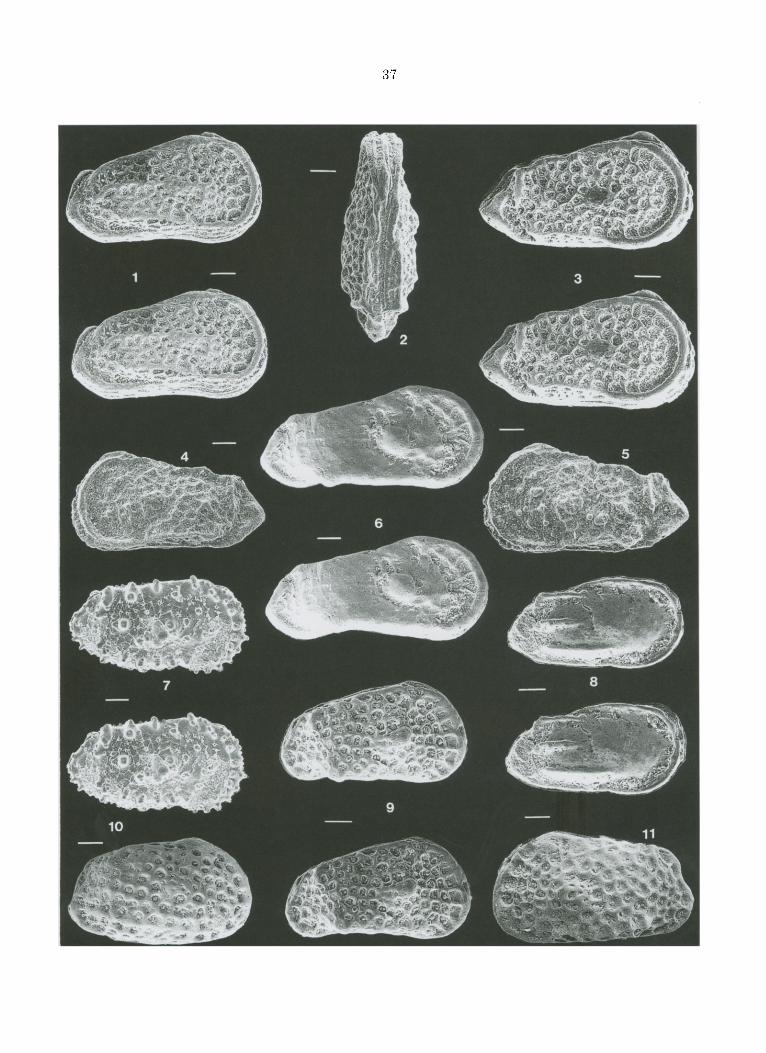
Trachyleberidea marg ina ta nov. sp. Fig. 4.2-5.
D e r i v a t i o n o m i n i s - Allusion au robus te bour re le t qui borde les valves, n o t a m m e n t ~ leur par t ie ant~rieure.
H o l o t y p u s (Fig. 4.2) - Une carapace p r~sum~e femelle n ° P6M1234a (collection labo Micropal. UPMC).
P a r a t y p o i d e s (Fig. 4.4-5) - 4 va lves et une ca rapace n ° P6M1234b.
L o c u s - t y p i c u s - Barranco del Gredero, 4 kin. au Sud de Caravaca de la Cruz, province de Murcie.
S t r a t u m t y p i c u m - Format ion de Jorquera , ~chantil lon 12 at t r ibu~ au Pal~oc~ne sup~r ieur (biozone P4 des foraminif~res planctoniques, Than~tien).
FIGURE 4 - 1. Dumontina ? sp., carapace en vue lat6rale droite (Car. 53). 2-5. Trachyleberidea marginata nov. sp., respec t ivement carapace en vue dorsale (holotype, Car. 12), carapaces en r u e lat~rale droite (Car. 53) et gauche (Car. 12) et valve gauche (Car. 12). 6. Gen. 1 sp., carapace en vue lat~rale droite (Car. 1). 7. Phalcocythere sp., valve droite (Car. 2,1). 8. Phacorhabdo tus sp., carapace en vue latSrale droite (Car. 18). 9. Gen. 3 sp., carapace en vue lat~rale droite (Car. 79). 10-11. Hornibrookella sp., r espec t ivement carapaces en r u e s lat~rale droite et gauche (Car. 1). Barre = 100 microm~tres. 1. Carapace, right lateral view. 2-5. Respectively cara- pace, dorsal view (holotype), carapaces, right and left lateral view, and left valve. 6. Carapace, right lateral view. 7. Right valve. 8. Carapace, right lateral view. 9. Carapace, right lateral view. 10-11. Respectively carapaces, right and left lateral view.
37

D i a g n o s e - Carapace allong6e subtriangulaire large bourrelet marginal relay6 vers l'arri~re par des cStes dorsale et ventrale; 6pines marginales peu d6velopp6es et c61ation intra-r6ticulaire r6duite.
D i m e n s i o n s holotype, ca rapace p rSsum6e femelle , Iongueur 0,83 ram. (+ ou - 0,01), h a u t e u r : 0,44 (+ ou - 0,02), I a rgeu r 0,31 (+ ou - 0,01) ; paratypes, ca rapace p r 6 s u m 6 e femelle , L: 0,83 (+ ou - 0,01), h: 0,44 (+ ou - 0,02), 1:0,31 (+ ou - 0,02); V.G. p r 6 s u m 6 e femelle , L: 0,84 (+ ou - 0,01), h: 0,42 (+ ou - 0,01); V.D. p r 6 s u m 6 e femelle , L: 0,84 (+ ou - 0,01), h: 0,41 (+ ou - 0,01) ; V.G. p r 6 s u m 6 e male , L: 0,86 (+ ou - 0,01), h: 0,39 (+ ou - 0,02); V.D. p r 6 s u m 6 e ma le , L: 0,85 (+ ou - 0,01), h: 0,42 (+ ou - 0,01).
R a p p o r t s e t d i f f 6 r e n c e s - L'espbce connue la plus proche de T marginata semble 6tre T prest- wichiana qui est plus petite et dent les bourrelets et c6tes sent ne t tement plus 6troits.
D i s t r i b u t i o n - Cette esp~ce est pr6sente sur route la hau teur de la coupe ~tudi~e, du Pal~oc~ne
l'EocSne inf~rieur (zones P3 h P7 pour les fora- miniFeres planctoniques, NP5 et NP12 pour les nannofossiles calcaires).
Genre D u m o n t i n a DEROO, 1966 (=?Hazelina Moos, 1966)
E s p ~ c e - t y p e - Cythere puncturata BOSQUET, 1854
Dumontina? sp. Fig. 4.1.
Trois carapaces suppos6es pal6oc~nes (Fig. 2) poss~dent les caract6ristiques g6n6riques exter- nes du genre Dumontina (DERO0, 1966, p. 144) et no tamment ta petite taille (longueur de l'ordre de 0,8 mm.), le profil g6n6ral 6troit en vue dorsale, le bourrelet ant6romarginal, l 'absence de tubercule subcent ra l , la surface r6ticul6e et costul6e. L'extr6mit6 post6rieure en angle relat ivement obtus et l 'absence de tubercule subcentral permet de dist inguer les esp~ces du genre Dumontina de celles a t t r ibuables au genre Trachyleberidea. Cependant, les caract~res internes des valves n 'ayant pu ~tre observ6s, leur attribution g6n~- rique est incertaine.
ExtSrieurement, la Dumontina ? sp. se distingue des esp~ces connues suppos~es cong~n~riques (in Deroo 1966) par sa forme ~ la lois triangulaire et t rapue (en vue lat~rale), par l 'existence d'une cSte ~troite mais bien individualis~e et d'une r~ticula- tion l~che et irr6guli~re.
38
Genre Agrenocythere BENSON, 1972
E s p ~ c e - t y p e -Agrenocy the re spinosa BENSON, 1972
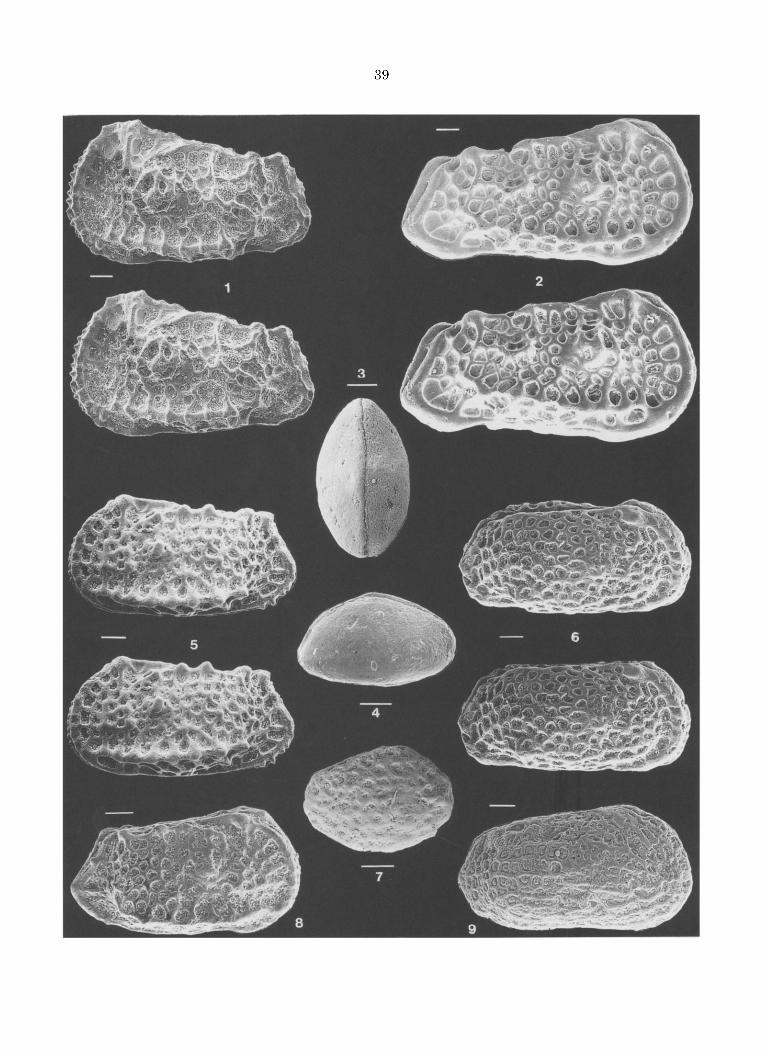
Agrenocythere ? cf. ordinata DELTEL, 1964 Fig. 5.1.
Une valve gauche (Fig. 5.1) et deux d6bris prove- nant d'~chantillons attribu~s au Pal~oc~ne sup~- rieur (Fig. 2) appart iennent ~ l'espSce d~crite par Deltel dans le Lut~tien du Bassin d'Aquitaine (figuration dans Ducasse et al. 1985) ou plus vrai- semblablement ~ une esp~ce voisine ~ bord post~- rodorsal plus rectiligne et ~ c6te dorsale plus r6guli~rement convexe.
A. ordinata est cit~e par Ducasse et al. (1985) dans tout l 'Eoc~ne et l 'OligocSne du Bassin d'Aquitaine. L'un d'entre nous (C. G.) a observ5 des valves identiques aux valves ~oc~nes du Bassin d'Aquitaine dans l 'Ypr6sien du Pr~rif (~chantillon Feinberg BG 23,5 et DA133).
Genre O e r t l i e l l a POKORNY, 1964
E s p ~ c e - t y p e - Cythere reticulata KAFKA, 1866
Oertliella ? sp. 1 aff. aculeata Fig. 5.5, 8.
Oertliella ? sp. 1 se distingue des esp~ces du Pal6oc~ne ou de l'Eocbne du Bassin d'Aquitaine et des Pyr6n6es attr ibu6es h Oertliella aculeata (Carbonnel 1975; Ducasse 1972, Ducasse et al. 1985; Tambareau 1972, etc.) par l 'ensemble des caract~res suivants: bord dorsal peu inclin6, 6pines et pores conuli 6mouss6s (par suite d'une usure?), alv6oles ant6romarginaux sensiblement festonn6s. Elle est repr6sent~e dans le Pal6oc~ne de Caravaca par de rares carapaces en m6diocre 6tat de conservation (Fig. 2).
Oertliella ? sp. 2 Fig. 5.2
Cette esp~ce est repr6sent6e dans les 6chantillons than6tiens recueillis h Caravaca par une carapa- ce (male?) longue de 1,3 mm, et par un fragment de valve. Tr~s allong6es, les valves sent orne- ment6es d 'un tubercule subcent ra l 6voquant plut6t celui des Oertliella que celui des Agreno- cythere. Elles sent creus6es de fossettes triangu- laires ~ quadrangulaires ou arrondies
FIGURE 5 - l.Agrenocythere ? cf. ordinata, valve gauche . 2. Oertliella ? sp. 2, carapace, v u e la t~ra le droi te (Car. 53). 3-4. Xestoleberis sp., c a rapace en v u e dorsa le e t la t~ra le droite (Car. 19,5). 5,8. Oertliella sp. 1, c a r apaces en v u e la t6ra le g a u c h e (Car. 12) et droi te (Car. 13). 6. Murrayina ? sp., ca rapace en vue la t~ra le droite (Car. 70). 7. Schizocythere ? sp., ca rapace en r u e la t~ra le droite (Car. 1). 9. Gen. 2 sp., ca rapace en vue la t~ra le droite (Car. 70). B a r r e = 100 microm~tres . 1. Left valve (Car. 20). 2. Carapace, right late- ral view. 3-4. Carapace, dorsal and right lateral view. 5,8. Carapaces, right and left lateral view Car 12 and Car 13 respectively. 6. Carapace, right lateral view. 7. Carapace, right lateral view. 9. Carapace, right lateral view.
39

40
Genre Phalcocythere SIDDIQUI, 1971
E s p ~ c e - t y p e - Cythere horrescens BOSQUET, 1854
Phalcocythere sp. Fig. 4.7
Les valves du Than~tien de Caravaca se distin- guent de celles de la P. horrescens de l'Eoc~ne moyen des Bassins de Paris et d'Aquitaine par l'absence de tubercule oculaire, la non diff~rencia- tion d'une car~ne ventrale, le nombre moins ~lev~ des tubercules ou pores conulis et leur grossi~ret~ relative et enfin par l'existence d'une r~ticulation t~nue. Par leur petite taille, l'existence d'un bour- relet marginal et la relative grande taille de leur tubercules, elles ~voquent les Phalcocythere du Pal~oc~ne d'Afrique (P. modesta APOSTOLESCU, 1963 et P. cultrata APOSTOLESCU in Apostolescu, 1961; Barsot t i 1963; Carbonnel et al. 1990; Carbonnel & Reyment 1993) mais s'en distin- guent ne t tement par l'absence de tubercule ocu- laire, la grossi~ret~ et complexit~ du tubercule musculaire et, en ce qui concerne P. modesta, l'ex- tr6mit6 postSrieure moins triangulaire.
Genre Murrayina PURI, 1954
E s p ~ c e - t y p e - Murrayina howei PuaI, 1954
Murrayina ? sp. Fig. 5.6
Une carapace et une valve pal~oc~nes (Fig. 2) sent a t t r ibutes avec doute au genre Murrayina dent elles poss~dent les valves quadrangulaires et allong6es, la surface r~ticul6e et la charniSre amphidonte. Elles se rapprochent de M. ? monten- sis (Marli~re 1958; Deroo 1966) par ces m~mes caract~res et par la forme de leurs alvSoles ant~- romarginales; elles s'en 51oignent par l'absence de c6te dorsale et d'~paulement postUre-dorsal mar- qu6
Genre Phacorhabdo tus HOWE • LAURENCICH, 1958
E s p ~ c e - t y p e - P. texanus HOWE & LAURENCICH, 1958
Phacorhabdotus sp. Fig. 4.8
Bien que reprSsent~e par seulement 2 carapaces pal~oc~nes (Fig. 2), cette esp~ce est int~ressante au point de vue stratigTaphique - elle appartient ~ un genre exclusivement Cr~tac~ sup~rieur (Coniacien)
OligocSne sup~rieur (Pietrzeniuk, 1965) - et sur- tout au point de vue ~cologique. En effet, les esp6ces de ce genre, g~nSralement d~pourvues de tubercules oculaires et souvent associ~es ~ des
tests de foraminif~res planctoniques sent consid~r~es comme un indicateur de mers "pro- fondes". En fait, ce caract~re profond est relatif, correspondant ~ l'6tage circalittoral des biologistes
par exemple pour l'Oligoc~ne d'Allemagne of~ deux esp~ces de Phacorhabdotus sent d5crites (Pietrzeniuk 1965) - ou ~ la partie sup~rieure de l'~tage bathyal - par exemple pour le Maestrichtien du Kef ~ P. inaequicostata (Donze et al. 1982) ou le Pal~oc~ne du SSn~gal dans lequel 5 esp~ces ont 5t~ d~crites (Apestolescu 1961; Colin 1987).
Au point de vue syst~matique, les carapaces de Caravaca semblent bien d~finies par leur nette convexit~ m~diodorsale et leurs c6tes longitudi- nales au nombre de 3, subparallMes, bien mar- qu~es et de longueur croissante de hau t en bas. Un bourrelet marginal est prSsent sur l 'une des carapaces et absent (us~ ?) sur l 'autre.
Genre Hornibrookel la Moos, 1965
E s p ~ c e - t y p e - Cythere anna LIENENt~.LAUS, 1884
Hornibrookella sp. Fig. 4.10-11
Quatre carapaces plus ou moins bien conservSes et r~parties dans trois 6chantillons d'age pal~ocS- ne ~ ~oc~ne inf~rieur (Fig. 2) sent attribuables au genre Hornibrookella. Leur contour, leur taille (L = 8 mm et 0,75, respectivement, pour les deux carapaces males et femelles les mieux pr~serv~es) et leur ornementation sent tr~s proches de ceux de H. persica TA~BAREAU, 1972 (voir aussi figura- tion dans Ducasse et al. 1985). Cependant, H. per- sica, d6crite dans le Than~tien nord-pyr~n~en, poss~de des c6tes ventrales et dorsales bien diff6- renci~es qui m a n q u e n t sur les valves de Caravaca.
OSTRACODES ET PASSAGE PAL]~OCENE-]~OC]~NE
A Caravaca, le Pal6oc~ne et l'Eoc~ne inf6rieur, tels que d~finis ici, ont livr6 respectivement 367 et 107 valves ou carapaces adultes et juveniles attri- bu6es ~ une vingtaine d'esp~ces. Les ~chantillons, de taille comparable (500 cm 3, approximative- ment), sent in6galement riches, ceux du Pal~oc~ne contenant jusqu'~ 110 valves ou carapaces, soit un peu plus de 13 en moyenne tandis que ceux de l'Eoc~ne en contiennent au maximum 24, soit pros de 5 en moyenne (Fig. 2). Toutes les esp6ces sent nouvelles ou d'attribution incertaine (Bairdia cf. cymbula et Agrenocythere cf. ordinata notam- ment) et la biodiversit~ ~tait faible, au Pal~oc~ne comme ~ l'Eoc~ne. D'une part, en effet, les ~chan-

41
tillons significativement riches en individus sont pauvres en esp6ces (8 a 9 esp6ces darts l'6chan- tillon 2,1 qui contenait 70 individus, 7 ~ 8, dans le 12 qui en contenait 110). D'autre part, consid6r6es globalement les 265 valves ou carapaces pal6oc6nes d6termin6es se r6partissent en 18 esp6ces tandis que les 68 valves ou carapaces 6oc6nes appart iennent a 9 esp6ces. Cette faible biodiversit6 correspond sans doute en partie a un faible hombre de niches 6cologiques disponibles, cons6quenee d'une par t de la relative homo- g6n6it6 des milieux oc6aniques situ6s sous la zone euphotique, d 'autre par t de la sp6cialisation moins grande des esp6ces vivant dans un milieu relat ivement pauvre en nutriments.
En ce qui concerne les foraminif'eres benthiques, une extinction massive des benthiques au passage Pal6oc6ne-Eoc6ne et une diminution de la biodi- versit6 a 6t6 constat6e fi Caravaca et h Zumaya; elles seraient li6es ~ une hausse brutale (et tran- sitoire) de la temp6rature (Ortiz 1994). Si l'abon- dance des ostracodes diminue aussi du Pal6oc6ne
l'Eoc6ne inf6rieur, l 'extinction de certaines espgces est peut-6tre plus apparente que r6elle. En effet, les esp6ces pal6oc6nes (7 a 10 sur 19, el. Fig. 2) qui n'ont pas 6t6 retrouv6es dans l'Eocgne, ne sont repr6sent6es que par une carapace (Gen. 1 sp., Gen. 2 sp., Abyssocythere sp., Oertliella sp. 2 et Cyamocytheridea ? sp.), voire deux ou trois (Murrayana ? sp., Phalcocythere sp., Xestoleberis sp., Dumontina sp., Agrenocythere cf. ordinata). A priori, leur absence darts les 6chantillons 6oc6nes n'est pas plus significative que leur absence dans la plupart des 6chantillons pal6oc6nes. D'autant plus, a contrario, que les esp6ces repr6sent6es dans le Pal6oc6ne par plusieurs dizaines d'indivi- dus (Cytherella sp. 1 et sp. 3, Bairdia ? aff. cymbu- la et Trachyleberidea marginata) sont pr6sentes aussi dane', l'Eoc6ne inf6rieur. I1 est n6anmoins possible sinon probable, car le sort des ostracodes ne peut 6tre dissoci6 de celui des foraminit'eres benthiques, qu'une partie des esp6ces pal6oc6nes disparaissent r6ellement, notamment a la faveur de l'6v6nement anoxique mis en 6vidence au d6but de l'Eoc6ne, en milieu circalittoral ou 6pibathyal (voir supra): Phalcocythere sp., Dumontina sp. et Oertliella aff. aculeata (cette derni6re peut-~tre repr6sent6e par un d6bris de valves darts un niveau 6oc6ne riche en 616ments remani6s) sont ainsi des esp6ces susceptibles de caract6riser le Pal6oc6ne. Notons cependant, qu'en M6diterran6e orientale, darts des pal6omilieux semblables, Honigstein et Rosenfeld (1995) ne constatent pas de "significant changes" des assemblages d'ostra- codes au passage Pal6oc6ne-Eoc6ne.
Au point de vue 6cologique, l'omnipr6sence de genres ubiquistes tels Cytherella, Bairdia et Krithe, la pr6sence de genres habituels en milieux
marins profonds tels Agrenocythere et Trachy- leberidea et enfin la pr6sence de rares Trachyte- beridid6s pourvus de tubercules oculaires (Mur- rayana ? sp. et Hornibrookella sp.) plaident en faveur d'un milieu situ6 sous la zone euphotique, tr6s faiblement 6clair6, circalittoral ~ 6pibathyal.
CONCLUSION Les ostracodes recueillis appart iennent pour la plupart /~ des esp6ces qui semblent nouvelles - telle Trachyleberidea marginata nov. sp. - et qui ne s'identifient pas fi celles d6crites dans le Than6tien du Kef, en Tunisie, dans des milieux de profondeurs et d'fige suppos6s semblables (Donze et al. 1982), peut-~tre parce que les niveaux 6chantillonn6s ~ Caravaca sont plus anciens on parce qu'ils appart iennent fi une autre province biog6ographique. Ils sont sp6cifiquement et le plus souvent g6n6riquement diff6rents des ostra- codes d6crits dans le Pal6oc6ne ou l'Eoc6ne inf~- rieur de la T6thys occidentale (Bassiouni 1970; Bassiouni & Lfiger 1990; Boukhary & Guernet 1982; Honigstein & Rosenfeld 1995, etc.) ou de l'Afrique nord-occidentale (Carbonnel 1988, 1990, etc.). Enfin les esp6ces pal~oc6nes et 6oc~nes sont au moins en part ie communes et la l imite Pal6oc6ne-Eoc6ne est done, pour les ostracodes et en milieu infralittoral ~ 6pibathyal, moins tran- ch6e que dans le NW de l'Europe o~, en liaison sans doute avec l'existence d'une 6mersion pro- long6e et de l'6pisode lagunaire sparnacien, le renouvellement des esp6ces est total.
R e m e r c i e m e n t s - Cette recherche a ~t6 par t ie l lement financ6e par le projet PS91-0172 de la Direcci6n General de Investigaci6n Cientifica y T6cnica. Nous exprimons aussi notre gratitude aux rapporteurs de notre manuscrit , R. Reyment et Y. Tambareau pour leurs judicieuses critiques.
RI~FI~RENCES APOSTOLESCU V. 1961 - Contribution a l'6tude pal6onto-
logique (ostracodes) et stratigraphique des bassins er6tacgs et tertiaires de l'Afrique Occidentale. Revue de l'Institut Fran~ais du P~trole, 14/7-8: 779-867.
ARENILLAS I. & MOLINA E. 1996 - Bioestratigraffa y evo- lucidn de las asociaciones de foraminfferos planctd- nicos del transito Paleoceno-Eoceno en Alamedilla (Cordilleras B6ticas). Revista Espa~ola de Micro- paleontologfa, 28, 1, sous presse.
BABINOT J.-F. 1980 -Les ostracodes du Cr6tac6 sup6- rieur de Provence. Syst6matique, Biostratigraphie, Pal6o6cologie, Pal6og6ographie. Travaux du Labo- ratoire de Ggologie Historique et de Paldontologie de l'Universitg de Provence, 10, 634 p
BABINOT J.-F. • COLIN J.-P. 1979 - Taxonomic and pateoe- cologic comments on the genus Trachyleberidea

42
BOWEN, 1953. Proceeding of the VII International Symposium on Ostracodes, Serbian Geologogical Society (ed.).: 55-60.
BARSOTTI G. 1963 - Paleocenic ostracods of Libya (Sir te Basin) and the i r wide Afr ican dis t r ibut ion. Revue de l'Institut Fran~ais du Pdtrole, 18/11: 1520-1135.
BASSIOUNI M. 1971 - Os t racoda (Maur i t s in inae und T r a c h y l e b e r i d i n a e ) und ihre B e d e u t u n g ffir die B ios t r a t i g r aph ie des Maas t r i ch t und des Al t te r - terti~ir yon Jo rdan ien . Beihefte zur geologischen Jahrbuchb, 106: 5-52.
BASSIOUNI M. & LUGER P. 1990 - Maas t r i ch t i an to Ear ly Eocene Os t racoda from sou thern Egypt . Paleon- tology, paleoecology and b ios t ra t igraphy. Berliner geowissenschaftliche Abhandlungen, 120/2: 755-928.
BERGGREN W. & MILLER K. 1988 - Paleogene planktonic foraminiferal biostratigraphy and magnetobiochro- nology. Micropaleontology, 34: 362-380.
BENSON R. 1972 - The Brad leya problem, wi th descrip- t ion of two new psychrospher ic ostracode genera, Agrenocythere and Poseidonamicus (Ost racoda: Crustacea) . Smithsonian Contributions to Paleobio- logy, 12, 138 p.
BOLD W., VAN DEN 1960 - Eocene and Oligocene Ostracoda of Trinidad. Micropaleontology, 6/2: 145-196.
BOUF~L~Y M., GUERNET C. & MANSOUR H. 1982 - Ost ra- codes du Ter t ia i re inf~rieur de l 'Egypte. Cahiers de Micropaldontologie, 1: 13-20.
BOWEN R. 1953 - Os t racoda from the London Clay. Proceedings of the Geologists Association, 64/4: 285- 286.
CANUDO J., KELLER G., MOLINA E. & ORTIZ N. 1995 - Planktic foraminiferal turnover and 8C13 isotopes across the Paleocene-Eocene transition at Caravaca and Zumaya , Spain . Palaeogeography, Palaeocli- matology, Palaeoecology, 113: 75-100.
CARBONNEL G. 1975 - Les os t racodes de l ' I l e rd ien (Eocene inf~rieur) du bass in de Tremp (Espagne); s t r a to type et coupes avois inantes . Revista Espa~ola de Micropaleontologfa, 7/1: 37-50.
CARBONNEL G. 1980 - Ost racodes pal~og~nes et n~og~nes du sondage offshore de Cape Timirisi Maur i tan ie . Revue de Micropaldontologie, 31/3: 147-155.
CARBONNEL G. 1990 - Les t ra i t s majeurs de la faune d'os- t r acodes palSog~nes (P3A-P14) dans les bass ins cStiers de l 'Afrique de l 'Ouest et du Golfe de Guin~e. Courier Forschunginstitut Senckenberg, 123: 199-208.
CARBONNEL G., ALZOUMA K. & DIKOUMA M. 1990 - Les ostracodes pal6oc~nes du Niger: taxinomie - un t~moignage de l'existence 6ventuelle de lamer trans- saharienne? Geobios, 23/6: 671-697.
CARBONNEL G. & REYMENT R. 1993 - L'ornementation chez l'ostracode Phalcocythere du Pal~oe~ne: r~sultat d'une action morphog6ne antagoniste? Lethaia, 26: 81-88.
COLIN J.P. 1987 - E tude sys t6mat ique des ostracodes de la Fo rma t ion des Madele ines (Danien du S~n6gal). Cah iers de Micropaldontologie, 2/3-4: 114-124.
DAMOTTE R. & FLEUR¥ J.-J. 1987 - Ostracodes maas - t r i ch t i ens et pal~oc6nes du Djebel Dyr pros de Tebessa (Alg6rie orientale) . G~ologie m~diterranden- ne, 14/2: 87-107.
DELTEL B. 1961 - Les ostracodes du Pal6og~ne moyen et sup~rieur d'Aquitaine m~ridionale. Th~se de la Facult~ des Sciences de l'Universit~ de Bordeaux, 215 p.
DELTEL B. 1963 - Nouveaux ostracodes de l 'Eoc~ne et de l 'Oligoc~ne de l 'Aqui ta ine m~ridionale. Actes de la Socigtg Linngenne de Bordeaux, 100: 127-211.
DE~OO G. 1966 - Cy the racea (ostracodes) du Maas t r i - cht ien de Maas t r i ch t (Pays-Bas) e t des r~gions voi- sines; r~sul ta t s s t r a t ig raph iques et pal~ontologiques de leur ~tude. Mededelingen van de Geologische Stichting, C, 2-2, 197 p.
DONZE P., COLIN J.-P., DAMOTTE R., OERTLI H., PEYPOUQUET J.-P. & SAID R. 1982 - Les ostracodes du Campanien terminal ~ l'Eoc~ne inf~rieur de la coupe du Kef, Tunisie Nord-Occidentale . Bulletin des Centres de Recherche Exploration-Production Elf-Aquitaine, 6,/2: 273-335.
DUCASSE O., GUERNET CL. ~z TAMBAREAU Y. 1985 - Pal~og~ne. In OERTLI (~d.): A t l a s des ostracodes de France. Mdmoire Elf-Aquitaine, 9: 257-311.
DURAND DELGA M. & MAGNE J. 1958 - Donn~es s t r a t ig ra - p hiques et micropal~ontologiques nouvel les su r le N ummul i t i que de l 'Es t des Cordi l l~res b~t iques (Espagne). Revue de Micropaldontologie, 1/3: 155- 175.
GAliCIA ZARRAGA E. & RODRIGUEZ LAZARO J. 1990 - Late Cretaceous ostracode faunas from the Biscay syncli- norium (Basque Arc, northern Spain). Courier Forschunginstitut Senckenberg, 123: 229-238.
HARTMANN G. • PURI g . 1974 - S u m m a r y of Neonto- logical and Paleontological Class i f icat ion of Ost ra- coda. Mitteilungen aus dem Hamburgischen Zoolo- gischen Museum und Institut, 70: 7-73.
HASKINS C. 1963 - Revision of the ostracode genus Trachyleberidea BOWEN. Micropaleontology, 9/1: 71-74.
HAZEL J. 1965 - Notes on t h e o s t r acode genus Trachyleberidea BOWEN. Journal of Paleontology, 39/3: 501-503.
HAZEL E., MUMMA M. & HUFF W. 1980 - Ostracode bios- tratigraphy of the lower Oligocene (Vicksburgian) of Mississ ippi and Alabama. Transactions, Gulf Coast Association of Geological Societies, 30: 361-401.
HERRIG E. 1966 - Ost racoden aus der weissen Schrei- bkre ide (Unter -Maas t r ich t ) der Insel Rfien. Deutsch. Berichte der Deutschen Gesselschaft fi~r Geologische Wissenschaten, Reihe A, Geologie und Pal~iontologie, 2/4: 693-1024.
HONIGSTEIN A. & ROSENFELD A. 1995 - Palaeocene ostra- cods from s o u t h e r n I s rae l . Revue de Micropa- lgontologie, 38/1: 49-62.
KEIJ A. 1957 - Eocene and Oligocene Os t racoda of Belgium. Mgmoire de l'Institut Royal des Sciences naturelles de Belgique., 136, 210 p.
LEREY D. & LEVINSON S. 1974 - A deep-wa te r Pleis tocene microfossil assemblage from a well in the no r the rn Gulf of Mexico. Micropaleontology, 20/1, 37 p.
LIENENKLAUS E. 1894 - Monographie der Os t racoden des nordwes tdeu t schen Terti~irs. Zeitschrift tier deut- schen Geologischen Gesellschaft., 56: 158-268.
MARLI]~RE R. 1958 - Os t racodes du Mon t i en de Mons et r~su l ta t s de leur 5tude. Mdmoires de la Socidtd

43
Belge de G~ologie, de Pal~ontologie et d~Hydrologie, 5, 53 p.
MOLINA E., CANUDO J., GUERNET C., McDoUGALL K., ORTLZ N., PASCU~ J., PARES J., SAMSO J., SERRA-KmL J. & TOSQUELLA J. 1992 - The s t ra to typ ic I l e rd ian revisi- ted: inte~.-ated s t r a t i g r a p h y across the Paleocene/ Eocene boundary. Revue de Micropal~ontologie, 35/2: 143-156.
MOLINA E., CANUDO J., MA~TINEZ RU~Z F. & ORTIZ N. 1994 Integral~ed s t r a t i g r a p h y across the Paleocene/
Eocene b o u n d a r y a t Ca ravaca , sou the rn Spain . Eclogae geolicae Helvetiae, 87-1: 47-61.
ORTIZ N. 1994 - La extinci6n mas iva de Foraminiferos ben t6n icos b a t i a l e s y ab i sa l e s en el l imi t e Paleoceno/Eoceno. In MOLINA E. (6d.), Extincion y egis t ro fosil. Cuadernos Interdisciplinares, Univ. Zaragoza, 5: 201-218.
PIETRZENIUK E. 1969 - Die Ga t t ung Phacorhabdotus (Ostracoda) im norddeu tschen Pal~iogen. Geologie, 14/9: 1102-1113.
PuRI H. 1954 - Con t r ibu t ion to the s tudy of the Miocene of the F l o r i d a Panhand le . III , Ost racoda. Florida Geological Survey, Geological Bulletin, 36: 217-345.
REYMENT R. 1984 - Uppe r Cretaceous Ost racoda of Nor th Cen t ra l Spain. Bulletin of the Geological Institutions of the University of Uppsala, 10: 67-110.
ROMEIN A. 1979 - Lineages in ear ly Paleocene nanno- p lankton. Utrecht Micropaleontology Bulletin, 22: 18- 22.
SIDDIQUI Q. 1971 - Ea r ly Ter t i a ry Os t racoda of the fami- ly Trachyleber id idae from Wes t -Pak i s tan . Bulletin of the British Museum (Natural history), Geology, suppt . 9, 98 p.
TAMBAREAU Y. 1972 - Than6t ien sup6r ieur et I l e rd ien des Pet i tes Pyr6n6es, du P l a n t a u r e l et des cha~nons audois. Th6se es Sciences, Toulouse, 377 p.
TAMBAREAU Y., BESSIERE G., BILOTTE M., VILLATE J., BABINOT J.-F. & LETHmRS F. 1985 - Journ6e d '6tude des ostracodologistes de langue fran~aise dans les Hautes-Corbi6res (11 Mai 1984). Bulletin de la Soci~t~ d'Etudes Scientifiques de l'Aude, 85: 13-47.
VAN VEEN G. 1969 - Geological inves t iga t ions in the region wes t of Ca ravaca sou theas t e rn Spain. Th~se de Doctorat de l'Universit~ d'Amsterdam, 143 p.
VON HILLEBRANDT A. 1974 B i o e s t r a t i g r a f i a del Paleogeno en el Sures te de Espaf ia (provincias de Murcia y Alicante). Cuadernos de Geologfa, 5: 135- 153.
C. G U E R N E T Universit6 P. et M. Curie
Laboratoire de Micropal~ontologie D6partement de g6ologie s6dimentaire
CNRS URA 1761 - GDR 88 F-75252 Paris Cedex 05
E. MOLINA Departamento de Ciencias de la Tierra (Paleontologia)
Universidad de Zaragoza E-50009 Zaragoza



















