Embed Size (px)
Citation preview

Organisation de la variabilitd enzymatique des populations marocaines de luzerne (Medicago sativa L.) : structures gdniques et gdnotypiques
A. BIROUK Institut agronomique et vke'rinaire Hassan II, Rabat, Maroc
Y. DATTEE' Institut national de la recherche agronomique, Universite' de Paris-Sud, Amklioration des plantes, Unite' associe'e n () 115,
9 1405 Orsay CEDEX, France
~ d i t e u r de correspondance : R. S. Singh
R e ~ u le 25 fkvrier 1988
Accept6 le 26 septembre 1988
BIROUK, A., et D A T T ~ E , Y. 1989. Organisation de la variabilitk enzymatique des populations marocaines de luzeme (Medicago sativa L.) : structures gkniques et gknotypiques. Genome, 32 : 120- 128.
Des analyses klectrophorktiques de cinq loci enzymatiques ont kt6 effectukes pour ktudier les structures gkniques et gkno- typiques de 20 populations traditionnelles de luzeme (Medicago sativa L.) du Maroc. Cinq origines gkographiques diffkrente: ont kt6 considkrkes : Ziz, Driia et Dadks (trois oasis sahariennes) ainsi que Demnate et des oasis de r6gions montagneuses. A titre comparatif, trois cultivars (African, Moapa, Europe) et quatre populations (une du Soudan, deux d'Espagne et une de Provence) d'origine ktrangkre ont kt6 inclus dans l'analyse. Les structures gkniques ktudikes sur les 27 populations ou cultivars permettent une bonne discrimination parmi les populations et les origines. Le r6le des allkles de faible frkquence est important dans la discrimination et, en se basant sur la diversit6 allklique, on obtient une meilleure discrimination qu'avec les structures gknotypiques. Le cultivar flamand Europe est distinct de l'ensemble du matkriel mkditerranken (African, Moapa, Provence, populations marocainks) et se rapproche des deux kcotypes espagnols de Mielga. Les deux cultivars African et Moapa se diffkrencient des populations marocaines. Ces demikres sont rkparties entre deux groupes distincts correspondant probable- ment B deux pools gkniques diffkrents : le premier groupe est constitu6 par les populations de Demnate, le second par les populations issues des oasis de montagne et sahariennes, sans distinction nette en fonction de l'origine gkographique. Les structures gknotypiques, lorsqu'elles sont ktablies sur la base de 30 individus analyses par population ou cultivar, ne permettent pas de discriminer les populations. Une augmentation des effectifs reprksentatifs de quelques populations ou cultivars a permis d'ktudier les indices de fixation et d'kprouver I'hypothkse d'kquilibre panmictique. On constate un dkficit d'individus hktkro- zygotes et un kcart important B la panmixie. Les frkquences d'individus monogkniques et digkniques duplex sont klevkes, mais les individus tktragkniques sont rares.
Mots cle's : luzeme, isozymes, structures gkniques, structures gknotypiques, allkles peu frkquents.
BIROUK, A., and D A T T ~ E , Y. 1989. Organisation de la variabilitk enzymatique des populations marocaines de luzeme (Medicago sativa L.) : structures gkniques et gknotypiques. Genome, 32: 120 - 128.
Five enzymatic loci have been analyzed electrophoretically to study the genic and genotypic structures of 20 populations of alfalfa (Medicago sativa L.) from Morocco. Five different geographical origins were considered: Ziz, Dria and Dadks (three oases in the Sahara), Demnate, and oases from mountainous regions. For comparison, three cultivars (African, Moapa, Europe) and four populations (one from Sudan, two from Spain, and one from Provence) of foreign origin were included in the study. The genic structures of the above 27 populations or cultivars allowed a good discrimination among populations and origins. The alleles with low frequency played a significant role in the discrimination and the discrimination was better when established on the basis of allelic diversity than on genotypic structures. The Flemish cultivar Europe differed from all other mediterranean material (African, Moapa, Provence, Morocco populations) but closely resembled two Spanish ecotypes from Mielga. The cultivars African and Moapa proved to be different from the Morocco populations. The latter could be divided into two different genic pools: one consisted of the Demnate populations, the other grouped populations of the Sahara and mountainous oases with no clear distinction between geographical origins. Genotypic structures based on the analyses of 30 plants from each population or cultivar did not allow to discriminate among populations. To study fixation indices and verify the panmictic equilibrium hypothesis, the number of specimen per population or cultivar had to be increased for a few popula- tions. All the populations or cultivars analyzed showed a lack of heterozygous plants and the hypothesis of a Hardy - Weinberg equilibrium could not be accepted. Frequencies of monogenic individuals and digenic duplexes were high, but tetragenic indi- viduals were infrequent.
Key words: alfalfa, isozymes, genic structures, genotypic structures, infrequent alleles. [Journal translation]
Introduction caractkrisation des populations et des cultivars (Huaman 1984;
Les isozymes fournissent un moyen puissant pour l'ktude Chevalier et Dattke 1983; Yndgaard et Iloskuldsson 1985),
de la variabilitk des structures gknetiques des populations l'identification des duplicata dans les banques de genes (Brown vkgktales, spontankes ou cultivkes, notamment dans le et Clegg 1983; Williams 1986), les stratkgies de prospection domaine des ressources gknktiques. 11s sont utilisks pour la (Brown et Munday 1982; Marshall et Brown 1983) ou l'orga-
nisation des complexes d'espkces (Pernes 1984). 1 . Auteur B qui faire parvenir toute correspondance, B l'adresse Peu d'ktudes de ~ol~molphisme enzy matique ant menkes
suivante : Genktique et amklioration des plantes, Institut national de la chez la luzerne (Medicago sativa L.), compte tenu de sa nature recherche agronomique, 78026 Versailles CEDEX, France. allogame autott5traplo'ide, relativement 2i d'autres espkces de
Printed in Canada i lmprimC au Canada
Gen
ome
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
CA
PE B
RE
TO
N U
NIV
on
05/0
2/13
For
pers
onal
use
onl
y.

TABLEAU 1. Origine et dknomination du materiel vegetal
Origine gkographique Commentaires
Mielga 1 Mielga 2
Ziz 1 h Ziz 5 Dra 1 hDra5 Dem 1 h Dem 4
Mont 1 h Mont 4
Dad 1 et Dad 2
African
Moapa
Europe
Provence
HCgazi
Mielga, Espagne
VallCe du Ziz VallCe du Dr5a Demnate, Piedmont atlantique
du Haut Atlas Haut Atlas central et Anti-
Atlas (oasis de montagne) Oasis de Dadks
Flandre
Provence
HCgazi, Soudan
Populations collectees au nord de 1'Espagne (Epila et Villamayor) (Delgado 1986)*
Populations marocaines prClevCes dans des exploitations traditionnelles de multiplication de semences (Birouk 1987)
Cultivar produit,aux ~tats-Unis h partir d'une population d'Egypte?.
Cultivar resultant de la selection de neuf clones dans African
Cultivar d'origine flamande (France et nord de 1 ' Europe) *
Population principalement cultivCe dans la region de Provence en France*
VariCtC population* - -
*Fournis parJa Station d'arnelioration des plantes fourragkres, Institut national de la recherche agronornique, Lusignan, France. TV. Marble (University of California), communication personnelle.
m2me importance economique. Le determinisme genetique de (&AMY), les leucine aminopeptidases (LAP) et les peroxydases quatre systkmes enzymatiques a ete elucide ii la suite des (PRX). h i s , en vue de 1'Ctude de la structure gknotypique des popu- travaux de Brunei (1982) et Quires (1982). Des recherches uti- lations, un SOUS-khantillon de 9 populations marocaines, 2 popula-
lisant l'~lectrophor~se effectuees sur la luzeme et les tions Ctrangkres et 2 cultivars a kt6 prClev6 et un total de 60 individus
espkces voisines du genre Medicago (Bingham et yeh 1971; par population ou cultivar a etC analysC pour les m@mes systtmes
Mariani et al. 1978; Damerval 1983; Quiros 1983). Au niveau e n ~ ~ ~ ~ ~ ~ ~ i prottiques ttt obtenus de jeunes feuilles strictement infraspecifique, des travaux d'identification de dtpliCes, broytes dans un tampon phosphate 0,1 M, pH genotypes et de cultivars Ont ete effectuks al- 1972; (Bmnel 1982). Pour les P-amylases et les leucine aminopeptidases, le Deloss 1972; Quiros 1983). support de migration Ctait un gel de polyacrylamide h 8% avec un
Au Maroc, des populations traditionnelles de luzerne sont tampon Tris -borate - EDTA (pH 8,3). Les techniques de rCvClation cultivees depuis de nombreux sikcles dans des aires pedo- climatiques diversifikes.
La prksente etude des ressources gknetiques de luzerne au Maroc vise deux objectifs : ( i ) une approche des structures gkniques et gknotypiques des populations marocaines de luzerne, en comparaison avec des cultivars etrangers; (ii) une classification des luzernes marocaines sur la base des caractkres enzymatiques, et la recherche d'une eventuelle rela- tion entre cette classification et l'origine gkographique.
MatCriel et mCthodes Matelriel velgeltal et analyses enzyrnatiques
Les analyses enzymatiques ont port6 sur 20 populations marocaines provenant d'une prospection dans diffkrentes regions du Maroc (Birouk 1989) et sur 3 cultivars et quatre populations d'origine Ctrangtre servant de tCmoins (tableau 1). Les populations marocaines ont CtC collectCes dans des exploitations agricoles ou la multiplication de semences de luzerne est traditionnellement pratiquCe. Les popula- tions n'ont subi aucune sklection, h part la sClection locale due au milieu. Le materiel d'origine Ctrangkre inclus dans l'analyse h titre comparatif comprenait deux cultivars sClectionnCs aux Etats-Unis et vulgarisCs au Maroc (African et Moapa), deux populations de Mielga, Espagne, et l'une d'HCgazi, Soudan, provenant de prospections, une population de Provence et un cultivar flamand (Europe).
Dans un premier temps, trente individus par population ou cultivar ont CtC analysCs pour trois systkmes enzymatiques : les P-amylases
ont kt6 empruntCes h D. run el (1982) avec une seble modification : le noir solide K (Fast Black K) a CtC utilisC pour la coloration des leucine aminopeptidases. La migration des peroxydases a CtC effec- tuCe en gel discontinu (Ornstein et Davis 1964). La rCvClation a CtC faite selon la mCthode de Graham et al. (1964).
Les rksultats des Ctudes du dCterminisme gCnCtique permettent de conclure h l'existence de six loci : /3-Amy-1, Lap-1, Lap-2, Prx-1, Prx-2 et Prx-3 (Brunel 1982; Quiros 1982).
Les techniques utilisCes ici ont permis de retenir, pour 1'Ctude gCnCtique, cinq loci pluriallCliques avec codominance : /3-Amy-1, Lap-1 , Lap-2, Prx-2 et Prx-3.
Me'thodes d 'analyse de structures gkniques et gknotypiques Analyse des structures geitiques Pour chaque locus, et sur la base du nombre d'allkles quantifiC de 0
h 4 selon I'intensitC des bandes, les frequences allCliques ont CtC calculCes par population ou par origine gkographique.
La variabilitC intrapopulation a CtC apprCciCe par le taux de poly- morphisme, le nombre moyen d'allkles par locus et l'entropie,
n
H = - C pi log, pi i = 1
ou pi reprksente la frkquence du ie allkle et n le nombre d'allkles au locus considCrC. Ce dernier indice, applique h la diversit6 infraspC- cifique par Hutchenson- (1970), donne une estimation quantitative de la variabilitk intragroupe. Par rCfCrence h l'entropie totale, il est aussi possible d'exprimer les parts respectives de l'entropie intrapopu- lation, intra-origine et inteiorigines.
Gen
ome
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
CA
PE B
RE
TO
N U
NIV
on
05/0
2/13
For
pers
onal
use
onl
y.

GENOME, VOL. 32, 1989
8 - S f i q 8 l 8 8 - 1 ; o o o o o o o a
> . d
Par ailleurs, pour n allkles kquifrkquents, H,,, = log2 n et un coef- ficient d'kquitabilitk peut Ctre dkfini par
E = * (Feoli et al. 1984) log2 n
Pour l'analyse de la variabilitk interpopulations, des analyses facto- rielles discriminantes (Dixon 198 1, BMDP 7 M) ont kt6 rkaliskes aux deux niveaux de regroupement, population et cultivar ou origine gko- graphique.
Des distances de Mahalanobis ont kt6 calculkes. Sur la matrice des distances interpopulations, une classification ascendante hikrarchique a kt6 effectuke, le critkre d'aggrkgation des classes ktant la minimisa- tion du moment centrk d'ordre 2.
Analyse des structures geizotypiques Les frkquences des individus monogkniques (M), digkniques
simplex (S) , digkniques duplex (D) , trigkniques (T) et tktragkniques (Q) ont kt6 estimkes pour chacun des loci.
Le niveau d'hktkrozygotie des populations a kt6 approchk par l'indice d'hktkrogknkitk,
1 2 5 Ho = -S + -D + -T + Q 2 3 6
lequel a kt6 calculk en considkrant la probabilitk pour chaque gkno- type de produire des gamktes hktkrogknes h la mkiose pour a = 0 (Demarly 1963). Les indices d'hktkrogknkitk des 20 populations et des sept tkmoins pour les cinq loci ont fait l'objet d'une analyse facto- rielle des correspondances.
Pour les populations ou cultivars caractkrisks sur la base de 60 indi- vidus, l'indice de fixation de Wright (1969) a kt6 calculk :
oG He est le taux d'hktkrogknkitk calculk h partir des frkquences all& liques sous l'hypothkse de l'kquilibre de Hardy - Weinberg.
L'indice de fixation F est une apprkciation de la structure gkno- typique h un locus donnk (Jain et Workman 1967; Brown 1979). Cependant, il convient de noter que, chez les tktraplo'ides, les taux d'hktkrogknkitk calculks concement les populations gamktiques, et non pas les populations zygotiques.
Pour cinq populations et un cultivar, un test de la panmixie a kt6 effectuk. Les allkles ont kt6 regroupks en quatre classes au maximum. Les deux premikres classes contiennent chacune un seul allkle (0,3 < frkquence < 0,7), la troisikme et l'kventuelle quatrikme classe contiennent les allkles de frkquence plus faible (0,l < frkquence < 0,3). Les allkles rares sont regroupks avec ceux de la troisikme ou quatrieme classe.
Les comparaisons entre les effectifs observks et les effectifs thko- riques ont port6 sur les cinq classes gknotypiques aprks regroupement des combinaisons allkliques en classes gknotypiques.
Malgrk ces simplifications, et compte tenu des faibles effectifs de certaines classes gknotypiques, il s'avkre impossible, dans certains cas, de satisfaire aux hypothkses nkcessaires h la validitk du test X2.
Analyse des structures geiziques Variabilite' intra-origine et intrapopulation Vingt-huit alleles ont kt6 dknombrks pour l'ensemble des
loci (tableau 2). Tous les allkles ne sont pas kgalement dis- tribuks.
Taux de polymorphisme - Un certain nombre d'allkles (6- Amy-12, Lap-12, L a ~ - 2 ~ , Lap-212, P r ~ - 2 ~ ) sont rares (Marshall et Brown 1975). Leurs frkquences moyennes sont infkrieures B 5%. Une population est considkrke polymorphe en un locus donnk si la frkquence de l'allkle le plus frkquent est infkrieure B 95% (Brown et .Weir 1983). Selon cette dkfinition, tirke d'ktudes sur des populations diploi'des, toutes les populations marocaines et les populations ou cultivars tkmoins sont poly- morphes aux cinq loci. Cependant , gknkralement deux allkles
Gen
ome
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
CA
PE B
RE
TO
N U
NIV
on
05/0
2/13
For
pers
onal
use
onl
y.

BIROUK ET DATTEE 123
TABLEAU 3. Paramktres de variabilite enzymatique : resultats des analyses de
variance sur l'ensemble des loci
Variable F calculk
Nombre moyen d'allkles 7,00** Entropie 14,93** Equitabilite 3,1 I*
*Significatif au seuil de 5 %. **Significatif au seuil de 1 %
sont prkpondkrants chacun des loci et reprksentent eux seuls 80% environ des allkles.
Nombre moyen d9al121es par population ou cultivar et diversite' ge'nique - Les analyses de variance du nombre d'allkles par population ou cultivar et de l'entropie mettent en kvidence des diffkrences hautement significatives entre les origines gkographiques lorsque ces paramktres sont calculks sur l'ensemble des loci (tableau 3). La richesse allklique la plus klevke se trouve au sein des luzernes de Demnate et de Dadks (tableau 4). La population d'Hkgazi et le cultivar Europe ont la plus faible richesse allklique. Le coefficient d'kquitabilitk pennet de classer le cultivar Europe au meme niveau que les populations de Demnate et Mielga. Europe prksente peu d'allkles par locus mais une rkpartition relativement kquilibree des frkquences allkliques .
Discrimination entre les origines ge'ographiques Les individus d'une meme provenance gkographique ont kt6
regroupks sans tenir compte de la structuration en populations ou cultivars.
Lorsque les 10 origines sont incluses dans l'analyse, le pour- centage de classification correcte des individus est de 4 1'4 %. L'origine la mieux classke est le Soudan (57%) suivie de Demnate (51%). Dria est l'origine la moins bien classke (23 %).
Les allkles du locus Lap-2 sont panni les plus discriminants. L'axe 1 isole les populations de Demnate de tous les groupes
et les oppose au cultivar Europe et surtout aux populations de Mielga. Les luzernes de Demnate se distinguent par la prksence de certains allkles rares (Lap-21°, L ~ p - 2 ~ ' et P r ~ - 2 ~ ) et les frkquences relativement faibles de certains alleles en moyenne prkdominants dans les autres origines (0-Amy-13, 0-Amy-15). Les populations de Mielga et le cultivar Europe se distinguent selon l'axe 2, lequel les oppose au reste du matkriel ktudik, en raison de leur richesse en allkles Lap-212 et Prx-2'.
Discrimination parmi les populations et les cultivars Cette analyse est baske sur le regroupement des individus en
27 populations et cultivars. Le pourcentage de classification correcte des individus, en
moyenne de 21 %, varie de 4 % pour la population Dad 1 de Dadks 50% pour celle d'Hkgazi.
La reprksentation des populations et des cultivars sur le premier plan met en kvidence certains regroupements qui corroborent les tendances observkes dans l'analyse des ori- gines gkographiques (fig. 1) : (i) toutes les populations de Demnate constituent un groupe distinct du reste du matkriel; (ii) les populations de Mielga sont trks rapprochkes l'une de l'autre et se regroupent avec le cultivar Europe; (iii) les popu- lations de Provence et d'Hkgazi font partie d'un groupe central, lequel contient la majoritk des populations marocaines d'oasis et de montagne, sans une structuration nette en fonc-
TABLEAU 4. Estimation des paramktres de variabilite des populations ou cultivars regroup& selon l'origine geographique
Nombre d'allkles Origine par population
geographique ou cultivar Entropie ~ ~ u i t a b i l i t e
Maroc Ziz Dria Demnate Montagne Dadks
Mielga, Espagne Etats-Unis Flandre Provence Hegazi, Soudan
NOTA : Les valeurs suivies de la rn&rne lettre ne sont pas significativernent diffkrentes au seuil a = 5 % par le test de Duncan.
tion de leur origine gkographique. La population Dra 5, origi- naire d'une oasis isolke semble se dktacher du groupe. L'isolement de cette population est lik a la raretk de l'allkle P r ~ - 3 ~ et l'abondance des bandes Lap-212 et 0-Amy-1 I.
Le dendrogramme construit a partir des distances de Maha- lanobis interpopulations et cultivars (fig. 2) rksume les infor- mations apportkes par les analyses factorielles discriminantes.
La proximite des populations de Demnate est a nouveau observke. Le groupe central rkunit la majoritk des populations marocaines originaires des oasis et de montagne.
Les populations de Dria sont les plus disperskes dans le groupe central.
Structures geizotypiques des populations Description des structures ghotypiques des 2 7populations
ou cultivars Le tableau 5 montre la disparitk des distributions de frk-
quences gknotypiques selon les loci. Cependant, pour la plu- part des loci, les individus digkniques duplex sont les plus frkquents, suivis des monogkniques. Par contre, les tktra- gkniques sont rares .
La corrklation entre l'entropie et l'indice d'hktkrogknkitk est hautement significative pour chaque locus a l'exception du locus 0-Amy-' (tableau 6).
Une analyse factorielle des correspondances a kt6 effectuke sur les coefficients d'hktkrogknkitk calculks a chacun des loci. La projection des variables ainsi que des populations et des cultivars sur le premier plan ne met-pas en ividence d'associa- tions prkfkrentielles entre les niveaux d'hktkrozygotie des loci ktudiks. Par ailleurs, ni les structures gknotypiques ni l'hktkro- zygotie ne pennettent de caractkriser des populations ou culti- vars ou des origines. L'ktude des structures gknotypiques a alors kt6 poursuivie sur les seules populations ou cultivars reprksentks par 60 individus.
Indices de fixation et tests de la panmixie La moyenne des indices de fixation sur l'ensemble des loci
indique que les populations prksentent un excks d'environ 15 % de gamktes homogknes par rapport a une distribution conforme a l'kquilibre panmictique (tableau 7). I1 existe peu de varia- tions selon les loci. Aux locus Lap-I, Lap-2 et Prx-3, les popu- lations prksentent un fok excks de gamktes homozygotes et s'kcartent beaucoup de la panmixie. La distribution la plus pro- che de la panmixie parait rkaliske au locus Prx-2.
Gen
ome
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
CA
PE B
RE
TO
N U
NIV
on
05/0
2/13
For
pers
onal
use
onl
y.
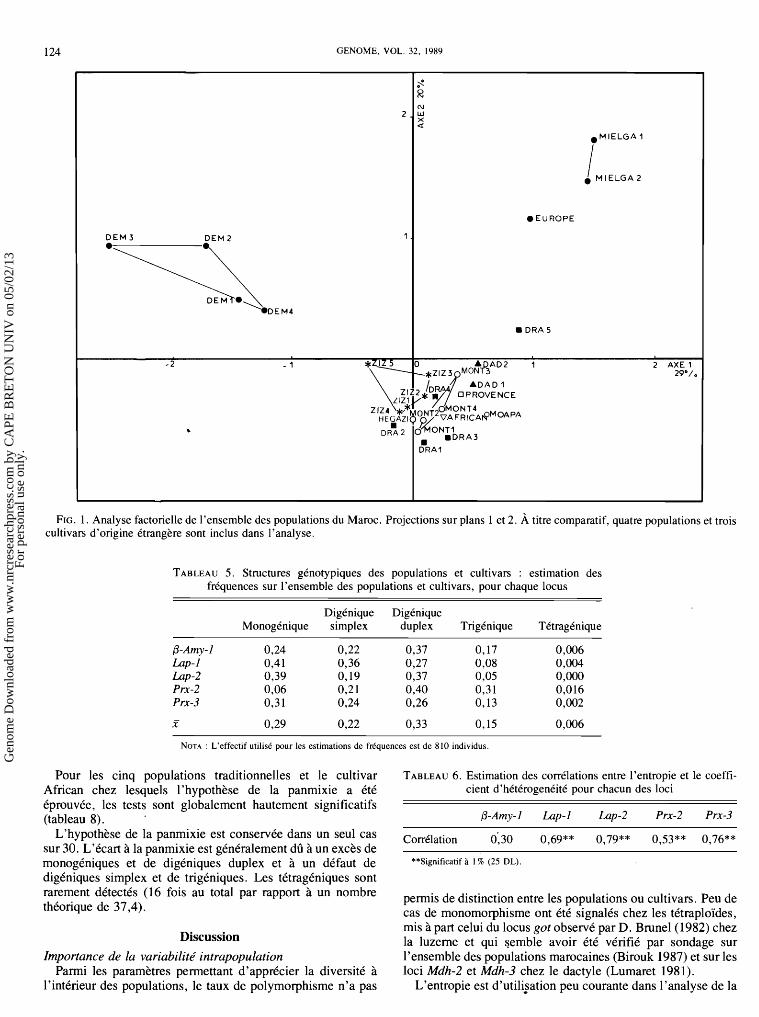
124 GENOME, VOL. 32, 1989
e M I E L G A 1
I M I ELGA 2
FIG. 1. Analyse factorielle de l'ensemble des populations du Maroc. Projections sur plans 1 et 2. A titre comparatif, quatre populations et trois cultivars d'origine Ctrangkre sont inclus dans l'analyse.
2 ,
D E M 3 DEM 2 1 .
@\\ '@DEM~
- 2 - 1
TABLEAU 5. Structures gknotypiques des populations et cultivars : estimation des frkquences sur l'ensemble des populations et cultivars, pour chaque locus
DigCnjque DigCnjque MonogCnique simplex duplex TrigCnique TCtragCnique
s R N
w X
a
DRA 5
-
2 AXE1
NOTA : L'effectif utilisi pour les estimations de fkquences est de 810 individus.
2 Q 0 / o
DRAI
Pour les cinq populations traditionnelles et le cultivar TABLEAU 6. Estimation des corrClations entre l'entropie et le coeffi- African chez lesquels l'hypothkse de la panmixie a kt6 cient d'hCtCrogenCitC pour chacun des loci kprouvke, les tests sont globalement hautement significatifs (tableau 8). 0- my-1 Lap-1 Lap-2 Prx-2 Prx-3
~ ' h ~ ~ o t h k s e de la panmixie est conservke dans un seul cas CorrClation 0,30 0,69** 0,79** 0,53** 0,76** sur 30. L'kcart B la panmixie est gknkralement dfi B un excks de monogkniques et de digkniques duplex et B un dkfaut de **Significatif i I % (25 DL).
digkniques simplex et de trigkniques. Les tktragkniques sont rarement dktectks (I6 fois par rapport un nombre permis de distinction entre les populations ou cultivars. Peu de thkorique de 37'4). cas de monomorphisme ont kt6 signalks chez les tktraploi'des,
mis B part celui du locus got observk par D. Brunel(1982) chez Discussion la luzerne et qui sernble avoir kt6 vkrifik par sondage sur
Importance de la variabilite' intrapopulation l'ensemble des populations marocaines (Birouk 1987) et sur les Parmi les paramktres permettant d'appkcier la diversitk B loci Mdh-2 et Mdh-3 chez le dactyle (Lumaret 1981).
l'intkrieur des populatiork, le taux de polymorphisme n'a pas L'entropie est d7utili?ation peu courante dans l'analyse de la
Gen
ome
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
CA
PE B
RE
TO
N U
NIV
on
05/0
2/13
For
pers
onal
use
onl
y.

MIELGA1 MIELGA2 EUROPE M OA PA
AFRICAN MONT4 DRA 3 Z l Z l ZIZ 2 PROVENCE DAD 1 DRA4 2123 2124 DRA2 Z IZ5 MONT3 D y A 2 HEGAZI MONT2 MONTl
D R A I D F A 5 DEM 1 DEM2 DEM4
DEM3
FIG. 2. Dendrogramme CtaMi sur la base de la distance de Mahalanobis de l'ensemble des populations du Maroc. Quatre populations et trois cultivars d'origine Ctrangkre sont inclus A titre de tkmoins.
TABLEAU 7 . Estimation des indices d'hCtCrogCnCitC (H,) et des taux de fixation ( F ) aux diffkrents loci
Ziz 4 Ziz 5 Dra 1 Dra 5 Dem 3 Dem 4 Mont 2 Mont 4 Dad 2 African Europe Provence HCgazi
variabilite intrapopulation (Hutchenson 1970; Lewontin 1972; Lumaret 1981). Chez les diploides, le paramktre le plus couramment utilise est la diversitk gknique de Nei (1972), dont l'kquivalent, chez ,les tktraploi'des, avait kt6 dkfini par Guy (1972) comme ktant la richesse allklique.
La richesse allklique exprime la probabilitk qu'un individu tirk au hasard soit hktkrogknique. Elle maximise le r6le des allkles prkponderants et met en kvidence les modifications importantes de frkquences entre les populations. La participa- tion des allkles rares, mais qui peuvent jouer un r6le discri- minant important, est par contre minimiske. C'est au vu de l'importance des allkles rares dans les rksultats expkrimentaux que le choix a kt6 port6 sur l'entropie. Sa signification stricte- ment gknktique est cependant moins bien dkfinie que celle de la diversitk gknktique (Brown et Weir 1983).
Sur la base de l'entrovie. la variabilitk avparait surtout d'ori-
gine intrapopulation puisque la diversite interorigine ne reprksente que 8,6 % de la diversitk totale alors que la diversitk intrapopulation ou intracultivar contribue pour 89 % .
Une apprkciation plus pondkrke de la variabilitk intrapopula- tion est foumie par les distances de Mahalanobis puisque le rapport de la moyenne des distances intrapopulations B la moyenne de toutes les distances inter- et intra-populations est de 69%.
La part klevke de la variabilitk intrapopulation n'est pas suprenante, compte tenu de l'ktat polyploi'de de la luzeme et de son mode de reproduction allogame. Elle explique les faibles frkquences de classification correcte observkes au niveau des populations ou cultivars. Le fait que, malgrk cette variabilitk, les origines Demnate et Mielga soient nettement discriminkes renforce la diffkrence entre ces origines gkographiques et les autres.
Gen
ome
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
CA
PE B
RE
TO
N U
NIV
on
05/0
2/13
For
pers
onal
use
onl
y.

GENOME, VOL. 32, 1989
TABLEAU 8. Structures gknotypiques des populations du Maroc, valeurs des X 2 du test de la panmixie
m chant ill on P-Amy-1 Lap-1 Lap-2 Prx-2 Prx-3
Ziz 5 24,1** 58,9** 27,8** 1'7 50,4** Dra 5 15,9** 67,7** 10,3** 6,3* 31,5** Dem 3 5,7** 29,3** 29,3** 37,6** 25,1** Mont 4 7,6** 23,7** 14,3** 19,1** 20,3** Dad 2 28,8** 32,0** 14,6** 17,8** 74,6** TCmoin (African) 47,3** 23,3** 25,6** 9,9** 19,3 * *
Par ailleurs, l'ktat polyploi'de peut expliquer la persistance des allkles rares. La lenteur de l'kvolution des frkquences gkni- ques dans les populations tktraploi'des a en effet kt6 dkmontrke (Demarly 1963). La possibilitk de dktection des allkles de faible frkquence est en relation avec l'effectif de plantes ana- lyskes. D'aprks les travaux de Bray (1983)' sous l'hypothkse de panmixie et avec un risque d'erreur de premikre espkce de 5 %, la frkquence minimale de l'allkle rare dktectable sur un kchantillon de 30 individus serait de 2'5 % . Ce calcul concorde bien avec les frkquences minimales dktectkes par population.
Structures ghotypiques des populations et cultivars L'ktude des structures gknotypiques met en kvidence une
importante deviation par rapport a la panmixie, avec un excks d'homozygotes, ou plus prkcisement de la fraction homogkne des gamktes. L'homogknitk des rksultats a un meme locus pour les diffkrentes populations et entre les loci conforte cette affir- mation. Cette situation semble frkquente chez les populations vkgktales spontankes ou cultivkes analyskes pour leur varia- bilitk enzymatique. Brown (1979) a pu dkgager deux faits essentiels, relatifs aux valeurs de l'indice de fixation F. Par rapport aux proportions attendues sous l'hypothkse d'un kqui- libre panmictique, il existe un dkficit gknkralisk d'hktkro- zygotes chez les espkces prkfkrentiellement allogames et, a l'inverse, il y a un excks gknkralisk d'hktkrozygotes chez les espkces prkfkrentiellement autogames. Brown (1979) a pro- posk de dknommer % paradoxe de l'hktkrozygotie . cette situa- tion contradictoire. Diverses hypothkses ont kt6 avanckes pour interprkter ce paradoxe (Brown 1979; Pernes 1984).
Sur un plan thkorique, deux arguments l'un propre aux tktra- ploi'des, l'autre au mode de collecte des populations peuvent &re invoquks.
Chez les tktraploi'des, l'indice de skparation a varie de 0 217 (Demarly 1963). Lorsqu'il n'est pas nul, la fraction homogkne des gamktes est supkrieure sous l'hypothkse pan- mictique. Toutefois, la seule variation de a ne suffirait pas rendre compte des dkficits observks.
Lorsqu'un kchantillon collect6 est composk de plusieurs sous-populations, l'effet Wahlund (1928) se traduit par un biais introduit dans le calcul de l'indice de fixation F. Dans le cas des populations de luzerne cultivke, ce biais pourrait &re dQ a la migration (kchange de semences) ou aux mklanges de rkcoltes d'annkes diffkrentes dans le grenier du paysan. Cependant, toutes les populations marocaines analyskes ici correspondent a la rkcolte d'une meme annke, tout les mklanges de plusieurs annkes ayant kt6 systkmatiquement exclus des analyses (Birouk 1989).
Sur un plan pratique et technique, la prksence d'allkles nuls non dktectks n'est pas a rejeter complktement. Elle pourrait
meme expliquer une partie de l'homozygotie excessive au locus Lap-1 ou Brunel (1979) avait identifik un allkle nul.
Par ailleurs, la variation dktectke par klectrophorkse corres- pond a environ 30% seulement des substitutions de bases, le reste ne conduit pas a une modification de la charge nette de la protkine ou concerne des skquences non traduites comme les introns (Brown et Weir 1983). Des monogkniques pourraient alors correspondre en rkalitk a des gknotypes hktkrozygotes. Cette dernikre hypothkse explicative parait parmi les plus plausibles .
Des klkments liks a la biologie de la reproduction et a la dynamique des populations peuvent aussi intervenir. Chez la luzerne, il existe des diffkrences gknotypiques pour la persis- tance (Hodgkinson et Williams 1983) et il est vraisemblable que des effets de compktition agissent sur la structure gknoty- pique des populations (Assemat 1977; Rotili 1979). Ces facteurs peuvent jouer sur l'kcart a l'kquilibre. I1 faudrait toute- fois supposer qu'ils sont gknktiquement liks aux loci analysks.
La rkgulation gknktique des diffkrentes ktapes de la repro- duction chez la luzerne est complexe (Dattke 1972; Dattke 1977; Guy et Dattke 1982). En supposant l'autofkcondation comme seul klkment explicatif de l'kcart la panmixie, il est possible d'en estimer le taux par la formule de Bennett (1968).
Dans les populations marocaines, les taux d'autofkcondation ainsi estimks seraient de l'ordre de 41 % et remarquablement homogknes selon les populations. Or, les taux moyens d'auto- fkcondation habituellement observks chez cette espkce sont de l'ordre de 5 a 15 %. Par ailleurs, une estimation de 41 % ne serait pas compatible avec les observations agronomiques puis- que, en moyenne, une gknkration d'autofkcondation conduit a une diminution de 30% du rendement en matikre skche. L'autofkcondation ne peut donc a elle seule rendre compte des valeurs observkes de l'indice de fixation.
Les importances relatives des multiples causes possibles et non exclusives de l'kcart a la panmixie ne peuvent pas etre discriminkes.
Si ces rksultats expkrimentaux corroborent bien ceux gknk- ralement obtenus par l'analyse enzymatique de populations vkgktales di- ou' tktra-ploi'des allogames, ils n'en sont pas moins en contradiction avec les klkments thkoriques et expkri- mentaux de l'ktude de l'hktkrosis.
Conclusion Malgrk toutes les limitations knumkrkes, les isozymes
permettent de distinguer certaines populations, et meme des groupes de popu!ations, essentiellement sur la base des struc- tures gkniques. A l'inverse, les structures gknotypiques sont peu discriminantes. La lenteur de l'kvolution des structures
Gen
ome
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
CA
PE B
RE
TO
N U
NIV
on
05/0
2/13
For
pers
onal
use
onl
y.

gknotypiques chez les espkces tktraploides et la richesse allk- lique relativement klevke de l'ensemble du matkriel test6 peu- vent expliquer cette situation.
Le paradoxe de l'hktkrozygotie de Brown (1979) est con- firm6 sur l'ktude des populations de luzerne. I1 est cependant loin de remettre en question l'kvolution des populations tktra- ploides allogames . En effet , les marqueurs enzy matiques ne sont pas reprksentatifs du genome dans son ensemble et ne concernent que des gknes de structures et non pas de rkgu- lation.
Le cultivar Europe, d'origine flamande, est distinct de l'ensemble du matkriel d'origine mkditkrankenne, au sens large. I1 se rapproche nkanmoins des deux kcotypes espagnols de Mielga sur la base de sa composition allklique.
Les deux cultivars African et Moapa, trks apparentks, sont toujours regroupks et distincts d'Europe. Un rksultat similaire a kt6 observe par Quiros (1983) sur la base de la variabilitk de deux systkmes enzymatiques : peroxydase et leucine amino- peptidase. I1 est intkressant de constater que African et Moapa se diffkrencient du matkriel marocain, malgrk leur origine nord-africaine.
Au sein du matkriel marocain, les populations de Demnate se distinguent nettement du reste du matkriel testk, du fait de leur richesse allklique.
Pour les trois systkmes eniymatiques analysks, une grande resemblance caractkrise les populations marocaines origi- naires des oasis et de montagne, bien que ces dernikres prksen- tent, en moyenne, une richesse allklique plus rkduite. Le polymorphisme enzymatique ne semble pas suivre l'organisa- tion de la variabilitk des caractkres agronomiques pour lesquels les populations de montagne sont distinctes, en particulier pour leur dormance hivernale (Birouk et al. 1989).
Les facteurs ayant pu intervenir dans la diffkrenciation des populations sont multiples : kventuels effets de fonda- tion, action sklective du milieu, chronologie des introductions et, kventuellement , pratiques culturales traditionnelles des pay sans.
L'individualisation des luzernes de Demnate est probable- ment en relation avec leur origine gknktique, laquelle a kt6 prkservke griice ii la faible ktendue de cette localitk et aux prati- ques des paysans qui prennent un soin particulier ii ne pas mklanger leur semence rkputke avec celles d'autres prove- nances.
L'hypothkse de la diversification des luzernes de montagne et des oasis ii partir d'un meme pool gknique est plausible, compte tenu de certains klkments d'ordre historique et gkogra- phique. Les premikres introductions de luzerne dans les oasis remonteraient, en effet, au 1 er sikcle av. J. -C. (Scofield 1908; Klinkowski 1933; Gunn et al. 1978), et les versants sud des montagnes de 1'Atlas ont servi plusieurs fois de refuge aux habitants des vallkes sahariennes (El Bekri 1068; Berque 1955).
En conclusion on pourrait rkpartir le matkriel marocain en deux pools gkniques diffkrents : celui des luzernes de Demnate, et celui des populations de 1'Atlas sud.
Une telle conclusion demanderait confirmation. Plusieurs questions restent en effet poskes quant ii l'individualisation de certaines populations telles Dra 5, ou meme Dem 3, et ii la position globale de la variabilitk enzymatique du matkriel marocain par rapport ii celle des luzernes de diffkrentes origines, en particulier d'Afrique du Nord.
Les identitks et les dissemblances, mises en kvidence par le polymorphisme enzymatique, si elles ne concordent que
partiellement avec celles de la variabilitk agronomique, appor- tent nkanmoins des klkments nouveaux utiles pour la con- naissance et la conservation des ressources mkditerrankennes.
ASSEMAT, L. 1977. Modkles d'analyse de la concurrence entre populations vCgCtales. Thkse de doctorat, spCcialitC sciences biolo- giques, mention Ccologie, UniversitC des sciences et techniques du Languedoc, Montpellier.
BENNETT, J. H. 1968. Mixed self- and cross-fertilization in a tetra- somic species. Biometries, 24(3) : 485 -500.
BERQUE, J . 1955. Structure sociale du Haut Atlas. Maisonneuve, Paris.
BINGHAM, E. T., et YEH, K. L. 197 1 . Electrophoretic patterns among alfalfa seed from selected varieties, experimental stocks, and species accessions. Crop Sci. 11 : 58 -61.
BIROUK, A. 1987. Les ressources phytogCnCtiques du Maroc : analyse de la variabilitC gCnCtique d'une espkce fourragkre : la luzerne (Medicago sativa L.). Thkse de doctorat en sciences, UniversitC de Paris XI, Paris.
1989. Collecte de populations traditionnelles de luzerne au Maroc : caractCrisation pCdoclimatique des regions prospectCes et recherche de caractkres adaptifs. Plant. Genet. Resources Newsl. Sous presse.
BIROUK, A., DATT~E, Y., SADIKI, M., et ROUMET, P. 1989. ~ v a l u a - tion agronomique des populations de luzerne traditionnelles du Maroc dans deux environnements. Agronomie. Sous presse.
BRAY, R. A. 1983. Strategies ,for gene maintenance. Dans Genetic Resources of forage plants. Editeurs : J. F . McIvor et R. A. Bray. Commonwealth Scientific and Industrial Research Organization, Melbourne. pp. 157- 158.
BROWN, A. H. D. 1979. Enzyme polymorphism in plant populations. Theor. Popul. Biol. 15(1) : 1-42.
BROWN, A. H. D., et CLEGG, M. T. 1983. Isozyme assessment of plant genetic resources. Dans Isozymes current topics in biological and medical research. Vol. 1 1 . Editeurs : M. C. Rattazi, J. G . Scandalios et G. S. Whitt. A. R. Liss Inc., New York. pp. 285 - 295.
BROWN, A. H. D., et MUNDAY, J. 1982. Population genetic structure and optimal sampling of land races of barley from Iran. Genetics, 58 : 85-96.
BROWN, A. H. D., et WEIR, R. S. 1983. Measuring genetic variabil- ity in plant populations. Dans Isozymes in plant genetics and breeding. Editeur : S. D. Tanksley et T. J. Orton. Elsevier, Amsterdam. pp. 219 -239.
BRUNEL, D. 1979. Recherche du determinisme gCnCtique de quelques systkmes enzymatiques chez la luzerne (M. sativa L.) di- et tCtraploide. Thkse de 3" cycle, UniversitC de Paris XI, Paris.
1982. Mise en Cvidence de 4 locus enzymatiques chez la luzerne (Medicago sativa L.) di- et tetra-ploide. Agronomie, 2(2) : 133 - 148.
CHEVALIER, M. H., et DATT~E, Y. 1983. VariabilitC enzymatique chez le mais. Can. J. Genet. Cytol. 26 : 214 -228.
DAMERVAL, C. 1983. Comparaison de six espkces de luzernes annuelles h l'aide de caractkres biomCtriques et enzymatiques. Agronomie, 3(10) : 97 1 -982.
DATT~E, Y. 1972. Analyse quantitative de l'auto- et l'inter-fertilitC chez quelques familles de luzerne. Ann. Amelior. Plant. 22(1) : 5 -21.
1977. Mode de reproduction et maintien de I'hCtCrozygotie chez Medicago sativa L. Physiol. Veg. 15 : 475 -479.
DELGADO, I. E. 1986. Characterization of luzerne (Medicago sativa L.) from Aragon (Spain). Servicio de Investigacion Agraria, Zaragoza.
DELOSS, F. J. 1972. Identification of alfalfa genotypes by electro- phoresis of soluble proteins and selected enzymes. Thkse de Ph.D. University of Missouri, Columbia, MO.
DEMARLY, Y. 1963. GCnCtique des tCtraploi'des et amklioration des plantes. Ann. Amelior. Plant. 13(4) : 307 -400.
DIXON, W. J . 1981. BMDP statistical software. University of Cali-
Gen
ome
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
CA
PE B
RE
TO
N U
NIV
on
05/0
2/13
For
pers
onal
use
onl
y.

128 GENOME, VOL. 32, 1989
fornia Press, Berkeley, CA. pp. 519-537. EL BEKRI. 1068. Description de 1'Afrique septentrionale. (Traduit par
Slane.) Maisonneuve, Paris, 1965. FEOLI, E., LANGONERO, M., et ORLOCI, L. 1984. Information analy-
sis of vegetation data. W. Junk Publishers, La Hague. GRAHAM, R. C., LUNDHOLM, U., et KARNOVSKY, M. J. 1964. Cyto-
chemical demonstration of peroxydase activity with 3-amino- 9-ethylcarbazole. J. Histochem. Cytochem. 13 : 150 - 152.
GUNN, C. R., SKRDLA, W. H., et SPENER, H. C. 1978. Classification of Medicago sativa L. using legume characters and flower colors. U.S. Dep. Agric. Tech. Bull. 1574.
GUY, P. 1972. Structure gCnotypique d'un hybride autotCtraplo'ide multipliC en panmixie. 11. Elements de gCnCralisation. Ann. Amelior. Plant. 22(3) : 239 -26 1.
GUY, P., et DATT~E, Y. 1982. Reproduction patterns of M. sativa. Dans Seed regeneration in cross-pollinated species. Editeurs : E. Porceddu, et G. Jenkins. A. A. Balkema, Rotterdam. pp. 125-141.
HODGKINSON, K. C., et WILLIAMS, 0 . B. 1983. Adaption to graz- ing in forage plants. Dans Genetic resources of forage plants. Editeurs : J. G. McIvor et R. A. Bray. Commonwealth Scientific and Industrial Research Organisation, Melbourne. pp. 85 - 100.
HUAMAN, Z. 1984. The evaluation of potato germplasm at the inter- national potato center. Dans Crop genetic resources: conservation and evaluation. Editeurs : J. H. W. Holden et J. T. Williams. George Allen et Unwin, London. pp. 199 - 201.
HUTCHENSON, K. 1970. A teit for comparing diversities based on the Shannon formulae. J. Theor. Biol. 29 : 25 1 - 254.
JAIN, S. K., et WORKMAN, P. L. 1967. Generalized F-statistics and the theory of inbreeding and selection. Nature (London), 214 : 674 - 678.
KLINKOWSKI, M. 1933. Imperial bureau of plant genetics. Herbage Plants, Bull. 12, Aberystwyth, Wales. (Traduit par G. M. Reserveare. )
LEWONTIN, R. C. 1972. The apportionfnent of human diversity. Duns Evolutionary biology. Vol. 6. Editeurs : T. Dobzhansky, M. M. Hecht et W. C. Steere. Appleton-Century -Crofts, New York, N.Y. pp. 381 -383.
LUMARET, R. 1981. Structure gCnCtique d'un complexe polyploi'de : Da,ctylis glomerata L. (Fam. GraminacCes). Thkse de doctorat d'Etat, UniversitC des sciences et techniques du Languedoc, Mont- pellier.
MARIANI, A,, ARCIONI, S., et VERONESI, F. 1978. Cytological analy-
sis and electrophoretic patterns of seed proteins in M. sativa, M. glutinosa and their hybrids. Genet. Agrar. 32 : 21 -29.
MARSHALL, D. R., et BROWN, A. H. D. 1975. Optimum sampling strategies in genetic conservation. Dans Crop genetic resources for today and tomorrow. Vol. 2. Editeurs : 0 . H. Frankel et J. G. Hawkes. Cambridge University Press, New York, N.Y. pp. 53 -80.
1983. Theory of fofage plant collection. Dans Genetic resources of forage plants. Editeurs : J. G. McIvor et R. A. Bray. Commonwealth Scientific and Industrial Research Oganization, Melbourne. pp. 135 - 148.
MILLER, M. K., SCHONHORST, M. H., et MCDANIEL, R. G. 1972. Identification of hybrids from alfalfa crosses by electrophoresis of single seed proteins. Crop Sci. 12: 535 -537.
NEI, M. 1972. Genetic distances between populations. Am. Nat. 106 : 283 - 292.
ORNSTEIN, L., et DAVIS, B. 1964. Disc electrophoresis. Ann. N.Y. Acad. Sci. 121 : 32 1 - 347.
PERNES, J. 1984. Gestion des ressources gCnCtiques des plantes. Tome 2. Lavoisier, Paris.
QUIROS, C. F. 1982. Tetrasomic segregation for multiple alleles in alfalfa. Genetics, 101 : 1 17 - 127.
1983. Alfalfa, luzerne (Medicagq sativa L.) Dans isozymes in plant genetics and breeding. Part B. Editeurs : S. D. Tanksley et T. J. Orton. Elsevier, Amsterdam. pp. 253 -294.
ROTILI, P. 1979. Contribution h la mise au point d'une mCthode de sClection de la luzerne crenant en compte les effets d'interfkrence entre les individus. I. Etude expCrimentale de la structure de la luzernikre. Ann. Amelior. Plant. 29(4) : 353 - 38 1.
SCOFIELD, C. 1908. The botanical history and classification of alfalfa. USDA Tech. BPI no 131. pp. 11-19.
WAHLUND, S. 1928. Zuzammens et zug von population und Korelationserscheinunge rom standpunkt der vererbungslehree ans betrachtet. Hereditas, 11 : 65 - 106.
WILLIAMS, J. T. 1986. Classification of genetic resources samples and breeders' stocks. Duns Infraspecific classification of cultivated and wild plants. Editeur : B. T. Styles. Clarendon Press, Oxford. pp. 255-261.
WRIGHT, S. 1969. Evolution and the genetics of populations. Univer- sity of Chicago Press, Chicago, IL.
YNDGAARD, F., et ILOSKULDSSON, A. 1985. Electrophoresis: a tool for gene-banks. Plant Genet. Resources Newsl. 63 : 34 -40. G
enom
e D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y C
APE
BR
ET
ON
UN
IV o
n 05
/02/
13Fo
r pe
rson
al u
se o
nly.













![6 Bacterio NouveauxOutils.ppt [Mode de compatibilité]...détection des résistances bactériennes est meilleure par les techniques phénotypiques que génotypiques Seuls les dépistages](https://img.pdfslide.fr/doc/110x75/609e89aea4c5bc7e1e122285/6-bacterio-mode-de-compatibilit-dtection-des-rsistances-bactriennes.jpg)















