Embed Size (px)
Citation preview

UE11 – Parcours 3 – Génétique - n°14
22/02/2016Pr S. Romana
RT : Julien THOMASRL : Arnaud KLOKNER
COURS NON RELU PAR LE PROFESSEUR
MITOSE (partie 2)
Plan :
I – MosaïcismeA – Diagnostic pré-implantatoireB – Instabilité chromosomique dans les blastomèresC – RationalisationD – Application au gène PLK4
II – Tumeurs A – Anomalies de nombre de chromosomesB – MVA et cancers

L’objectif de cette partie est d’étudier les applications des checkpoints de la mitose dans deux phénomènes physiologiques ou pathologiques : le mosaïcisme observé lors des premières divisions mitotiques (J0-J4) et l’instabilité chromosomiques dans les tumeurs.
I – Mosaïcisme
A – Diagnostic pré-implantatoire (DPI)
Un couple sur 300 possède un membre porteur d’une translocation chromosomique équilibrée dont la quasi-totalité ne présente aucun symptôme. Cependant, cette translocation peut entraîner de graves déséquilibres génétiques lors de la méiose. Après la fécondation, en fonction de la gravité, elle peut conduire à une stérilité, des avortements spontanés à répétition ou à la naissance d’un enfant présentant des malformations et un retard mental.
Jusqu’à la fin des années 1990, lors de la suspicion d’une maladie génétique dans un couple présentant l’un de ces cas de figure, il n’était possible que d’effectuer un diagnostic prénatal : une ponction de trophoblaste ou de liquide amniotique à 12 semaines indiquait la présence ou l’absence de l’anomalie génétique, et une interruption médicale de grossesse (IMG) était proposée le cas échéant. Par la suite a été mise au point la technique de DPI, encadrée par la loi et ne concernant que les couples susceptibles de transmettre une maladie génétique grave incurable :
1. Des gamètes sont prélevés chez les deux parents,2. Une fécondation in vitro est réalisée pour obtenir une dizaine d’embryons,3. Les divisions se succèdent jusqu’au stade de « morula non compactée » (J3),4. On effectue une biopsie de blastomères sur chaque embryon,5. Les blastomères sont analysés par PCR et séquençage (anomalie génique) ou par
hybridation in situ (anomalie chromosomique).
L’image suivante illustre la recherche d’une maladie récessive liée à l’X entraînant l’élimination des embryons (XY) et l’implantation des (XX) avec des sondes spécifiques des centromères de chaque gonosome :
Dans le cas d’une translocation t(2 ;8) on peut voir la comparaison entre le sang des parents présentant 2 signaux de chaque couleur (en haut) et de deux embryons (en bas) : l’un monosomique 2q (un seul signal jaune, à gauche) et l’autre normal (à droite, la multiplicité des signaux est ici non pertinente et liée au stade de la mitose observée).
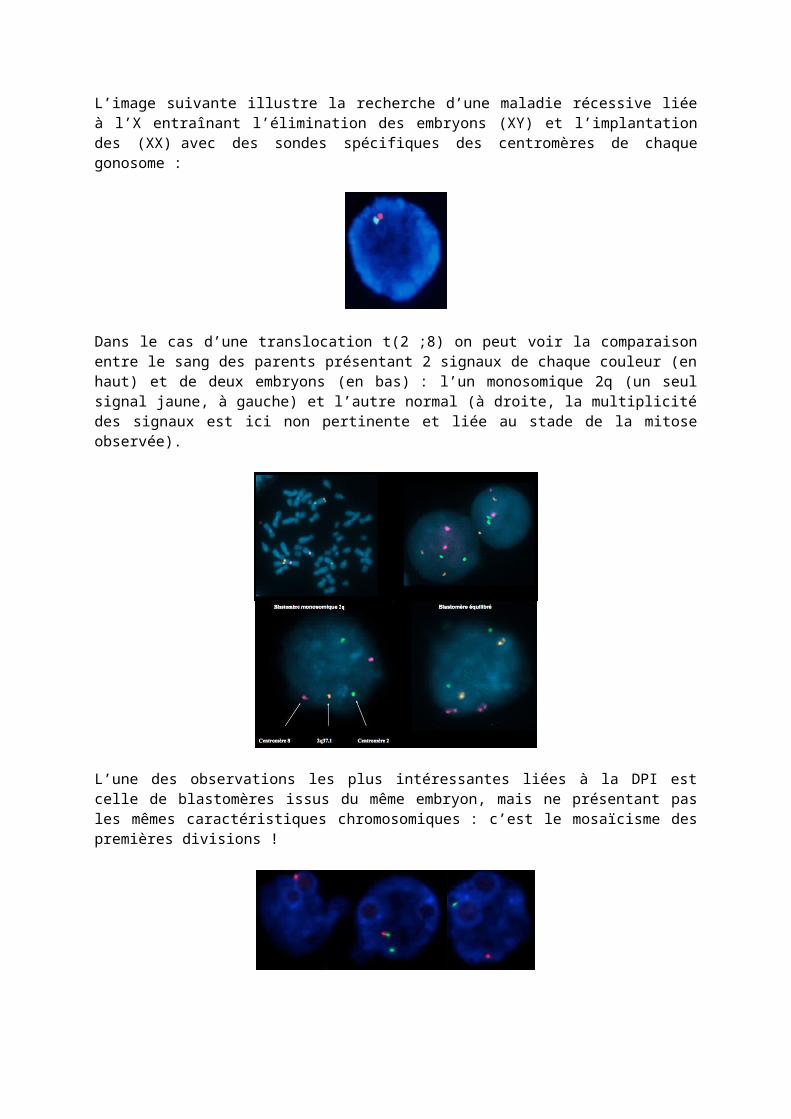
L’une des observations les plus intéressantes liées à la DPI est celle de blastomères issus du même embryon, mais ne présentant pas les mêmes caractéristiques chromosomiques : c’est le mosaïcisme des premières divisions !
B – Instabilité chromosomique dans les blastomères
Sur les images précédentes, la répartition chromosomique n’est pas bonne, mais celle-ci peut procéder d’un désordre total et s’avérer « chaotique » lorsqu’elle varie d’un blastomère à l’autre et ne peut être expliquée par les ségrégations méiotiques classiques :
Des analyses ont alors été menées, prouvant au début des années 2000 qu’à J3, 50% des embryons étaient mosaïques. Après l’élaboration de la méthode de CGH-array sur une cellule, l’équipe de Joris Vermeesch à Louvain a montré sur 23 embryons d’un même couple que ce nombre s’élève en réalité à 90% ! Il s’agissait d’un couple jeune (moins de 35 ans), en bonne santé, ne présentant pas d’avortements à répétitions ou d’anomalies chromosomiques, mais consultant en DPI pour une mutation génique de l’X. L’équipe a étudié la totalité des blastomères de chaque embryon pour les comparer afin de mettre en évidence un éventuel mosaïcisme sur la structure ou le nombre des chromosomes ; seuls deux embryons avaient un contenu de blastomères identiques et équilibrés.
Remarque : le pouvoir reproductif de l’espèce humaine est très faible. Cela peut s’expliquer par le mosaïcisme important lors des premières divisions. Des études ont montré que 30% des embryons ne s’implantent pas et que 30% meurent après implantation.
La méthode de l’étude n’est pas une CGH-array traditionnelle car il faut amplifier toute la cellule et certaines zones ne s’amplifient pas bien. À partir d’un set de référence, ils ont effectué des probabilités d’obtenir une trisomie ou une monosomie pour les différents chromosomes :
Vert : la probabilité est forte d’être en surplus (trisomie) sur toute cette région, Rouge : la probabilité est forte d’être en défaut (monosomie) sur toute cette région, Bleu : la probabilité est forte d’être en quantité normale.

Par exemple sur les figures suivantes, on observe un bras long du chromosome 18 normal dans les deux cas, avec une duplication (à gauche) ou une délétion (à droite) du bras court.
Le résultat final pour chaque embryon se présente sous la forme d’un récapitulatif des différents chromosomes avec les délétions ou duplications observées dans chaque blastomère (chaque bande verticale est un blastomère). Dans l’exemple suivant, on remarque que pour le chromosome 7 certaines bandes présentent une délétion ou une duplication qui se correspondent, manifestement le type et le contre-type d’une anomalie mitotique. En revanche, tous les blastomères ont une trisomie 15 : cette anomalie provient, elle, a priori de la méiose.
D’autres études ont repris ces résultats et confirmé que seuls 10 à 20% des embryons de couples jeunes et « normaux » ne présentent pas de mosaïcisme dans les blastomères à J3. Certains couples ont des répartitions plus chaotiques des chromosomes que d’autres.
Mais la première question est : pourquoi les embryons des humains sont-ils aussi déséquilibrés alors que ceux des autres espèces comme la souris ne le sont pas ?
C – Rationalisation
La première réponse serait : les protéines des checkpoints (cf. partie 1) ne fonctionnent pas ! Dans le cas contraire, les cellules auraient été réparées ou seraient entrées en apoptose. Les mécanismes de réparation ne fonctionnent pas non plus. Mais pourquoi ?
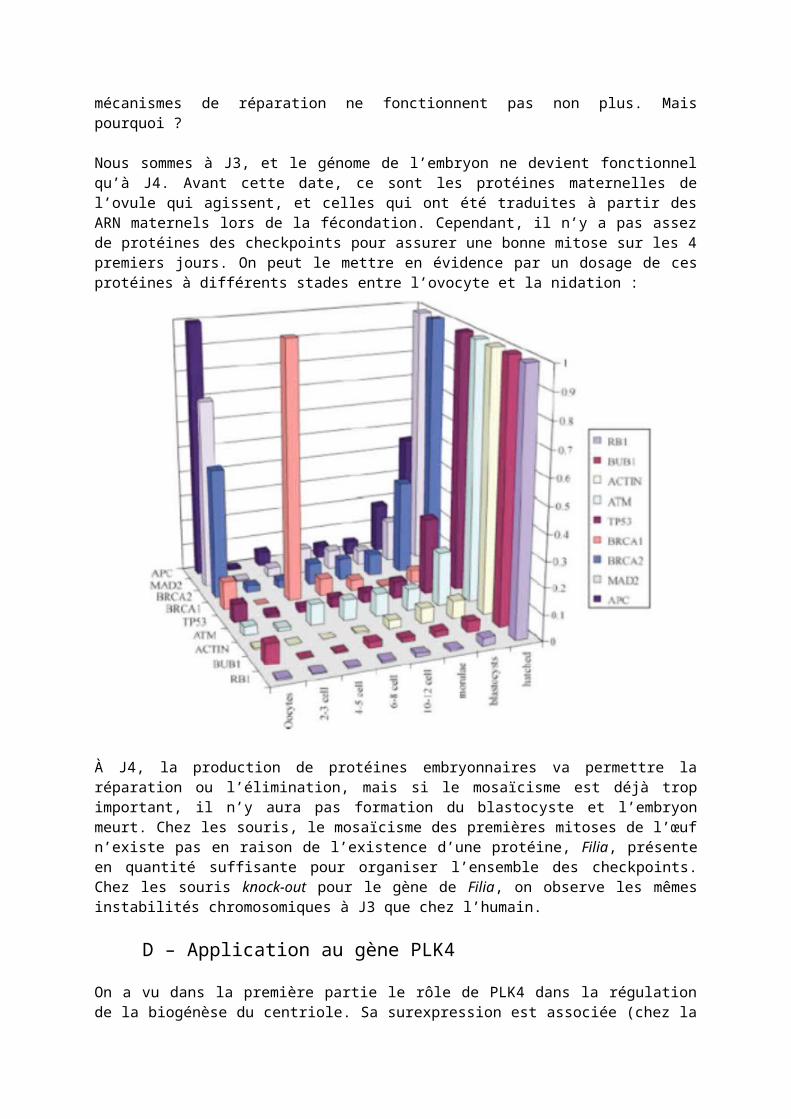
Nous sommes à J3, et le génome de l’embryon ne devient fonctionnel qu’à J4. Avant cette date, ce sont les protéines maternelles de l’ovule qui agissent, et celles qui ont été traduites à partir des ARN maternels lors de la fécondation. Cependant, il n’y a pas assez de protéines des checkpoints pour assurer une bonne mitose sur les 4 premiers jours. On peut le mettre en évidence par un dosage de ces protéines à différents stades entre l’ovocyte et la nidation :

À J4, la production de protéines embryonnaires va permettre la réparation ou l’élimination, mais si le mosaïcisme est déjà trop important, il n’y aura pas formation du blastocyste et l’embryon meurt. Chez les souris, le mosaïcisme des premières mitoses de l’œuf n’existe pas en raison de l’existence d’une protéine, Filia, présente en quantité suffisante pour organiser l’ensemble des checkpoints. Chez les souris knock-out pour le gène de Filia, on observe les mêmes instabilités chromosomiques à J3 que chez l’humain.
D – Application au gène PLK4
On a vu dans la première partie le rôle de PLK4 dans la régulation de la biogénèse du centriole. Sa surexpression est associée (chez la drosophile) à une augmentation des aneuploïdies, et une haplo-insuffisance est associée à des anomalies de cytodiérèse. PLK4 est également liée à des lignées de cancers chez la souris.
On a observé dans certains pays des couples qui généraient un nombre important d’embryons déséquilibrés, voire chaotiques. Il a été montré que certains variants du gène PLK4 étaient associés à cette production d’embryons déséquilibrés. C’est une pathologie maternelle, même si la mère n’a aucun symptôme : les anomalies s’expriment lors des checkpoints des premières divisions de l’œuf.
II – Tumeurs
A – Anomalies de nombre de chromosomes
Les premières observations de caryotypes de cellules cancéreuses (années 1960) ont mis en évidence des anomalies dans le nombre de chromosomes. Les génomes sont fortement remaniés, et ces remaniements chromosomiques sont proportionnels à l’avancée du cancer. On observe une anomalie chromosomique toutes les 5 divisions ! Plusieurs cas de figure, dont les mécanismes sont encore mal connus :
1. Un chromosome « part » toutes les 5 divisions : checkpoint mitotique.2. Aneuploïdie touchant un seul chromosome3. Hyperploïdie (23n chromosomes, n>2)4. Hyperdiploïdie (N>46 chromosomes)
On suppose une implication du SAC, équivalent au checkpoint mitotique, qui n’est pas encore prouvée, ou des cohésines qui se sépareraient précocement. On a cependant montré le rôle des centrosomes : une augmentation de leur nombre implique des anomalies de nombre des chromosomes (on retrouve le rôle de la PLK4). Enfin, les cyclines et les gènes des points de cassure sont également impliqués.
Pour étudier le rôle des protéines des checkpoints, on a fait des souris KO pour les gènes impliqués : les embryons obtenus présentaient de telles anomalies qu’ils n’étaient pas viables. Il a alors fallu étudier des hétérozygotes, et même, pour contourner des phénomènes de compensation, des doubles hétérozygotes car les premiers ne présentaient pas d’anomalies évidentes. Le lien de cause à effet n’a pas encore pu être établi avec les observations dans les cancers humains.
B – MVA et cancers
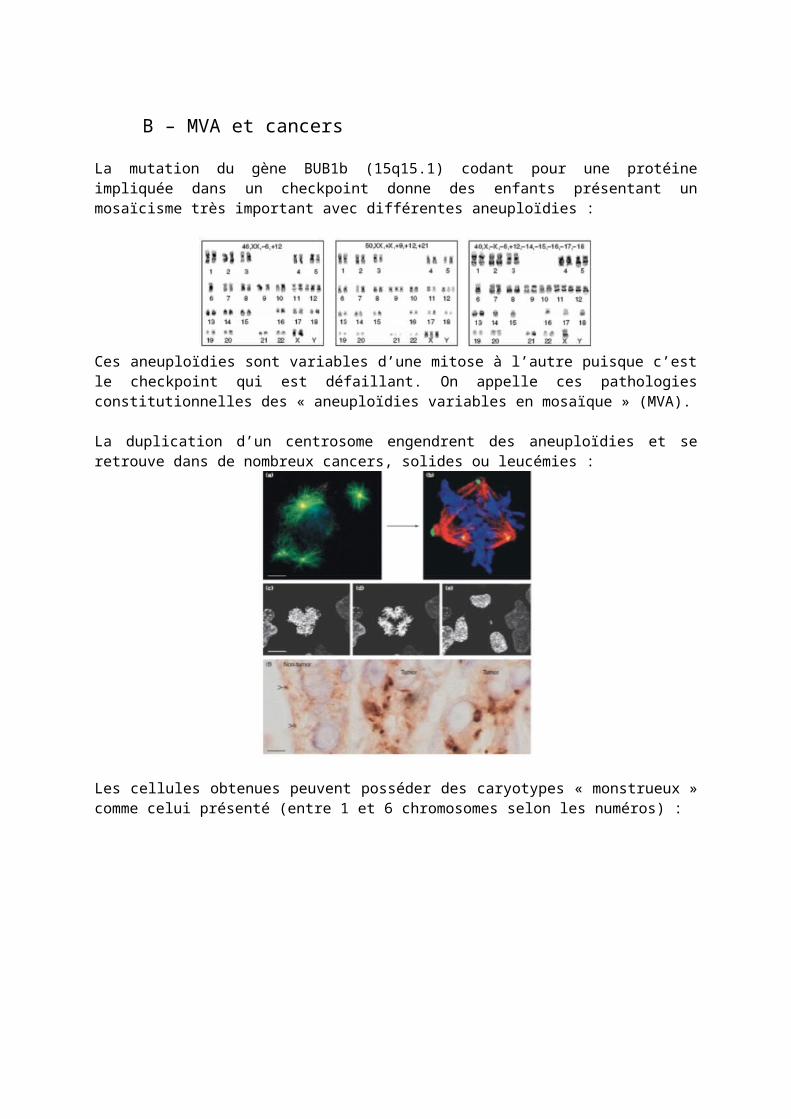
La mutation du gène BUB1b (15q15.1) codant pour une protéine impliquée dans un checkpoint donne des enfants présentant un mosaïcisme très important avec différentes aneuploïdies :
Ces aneuploïdies sont variables d’une mitose à l’autre puisque c’est le checkpoint qui est défaillant. On appelle ces pathologies constitutionnelles des « aneuploïdies variables en mosaïque » (MVA).
La duplication d’un centrosome engendrent des aneuploïdies et se retrouve dans de nombreux cancers, solides ou leucémies :

Les cellules obtenues peuvent posséder des caryotypes « monstrueux » comme celui présenté (entre 1 et 6 chromosomes selon les numéros) :
Les mécanismes impliqués sont très compliqués et ne sont pas encore tous élucidés. On sait qu’ils impliquent de nombreuses erreurs, sur les différents checkpoints, ainsi que sur la biologie du fuseau mitotique.
Abréviations : CGH-array : puce d’hybridation génomique comparativeDPI : diagnostic pré-implantatoireIMG : interruption médicale de grossesseMVA : aneuploïdie variable en mosaïque SAC : spindle assembly checkpoint
Mot du RT
FICHE RECAPITULATIVE
Il est très important de bien comprendre que le phénomène de mosaïcisme est très présent dans les premières étapes du développement de l’embryon (Jusqu’au stade J4) et que cela concerne aussi bien les couples sains que ceux touchés par des anomalies génétiques.
Apparition en 1990 de la technique de diagnostic pré-implantatoire qui permet dans les cas de risques de maladies graves UNIQUEMENT de repérer les embryons malades dès le stade blastocyste au lieu de devoir attendre 12 semaines de grossesse ce qui impliquait ensuite de devoir faire une IMG.
90% des blastocystes au stade J3 sont des mosaïques même chez les couples en bonne santé car il n’y a pas assez de protéine pour assurer les checkpoints avant l’activation du génome embryonnaire à J4. Cela explique les nombreux embryons qui meurent autour de la période implantatoire (avant ou juste après) lorsque l’intégrité génomique ne peut être rétablie.
Le gène PLK4 sert dans la répartition des chromosomes lors de la mitose (action sur les centrioles). Sa mutation amène à la production d’embryons déséquilibrés.
Cancer et remaniement du génome sont très liés à cause des dysfonctionnements du système de partage des chromosomes lors des divisions cellulaires ce qui amène parfois à des caryotypes aberrants.