Embed Size (px)
Citation preview

C
P
Rda
Rf
0h
ahiers de nutrition et de diététique (2013) 48, 92—97
Disponible en ligne sur
www.sciencedirect.com
HYSIOLOGIE
ôle des récepteurs mu-opioïdes dans la régulation’un axe intestin—cerveau de contrôle de la priselimentaire
ole of mu-opioid receptors in the regulation of a gut—brain axis of control ofood intake
Gilles Mithieuxa,b,c,∗
a Faculté de médecine Laennec Lyon-Est, institut national de la santé et de la recherchemédicale, U 855, 69372 Lyon cedex 08, Franceb Université de Lyon, 69008 Lyon, Francec Université Lyon 1, 69622 Villeurbanne, France
Recu le 10 janvier 2013 ; accepté le 11 fevrier 2013
Disponible sur Internet le 16 mars 2013MOTS CLÉSNéoglucogenèseintestinale ;Protéinesalimentaires ;Prise alimentaire ;Satiété ;Récepteursmu-opioïdes ;Axe intestin—cerveau
Résumé En utilisant des perfusions portales d’agonistes et/ou d’antagonistes des récepteursmu-opioïdes (RMO) chez des rongeurs conscients, nous avons montré que les RMO présentsdans les parois de la veine porte contrôlent un circuit nerveux intestin—cerveau d’induction dela néoglucogenèse intestinale, une fonction contrôlant la sensation de faim. Dans un secondtemps, nous avons montré que les peptides et les protéines promeuvent une induction RMO-dépendante de la néoglucogenèse intestinale. Les peptides n’ont pas d’effet chez les sourisknockout pour les RMO, qui sont également insensibles à l’effet de satiété induit par les régimesprotéinés. De plus, des perfusions portales de modulateurs des RMO n’ont pas d’effet sur la prisealimentaire chez les souris déficientes pour la néoglucogenèse intestinale. Ainsi, la régulationpar les RMO portaux d’un circuit nerveux intestin—cerveau d’induction de la néoglucogenèseintestinale par les peptides est un lien causal dans le phénomène de satiété induit par lesprotéines alimentaires.© 2013 Société francaise de nutrition. Publié par Elsevier Masson SAS. Tous droits réservés.
KEYWORDSIntestinalgluconeogenesis;Dietary proteins;Food intake;Satiety;
Summary Using portal infusions of agonists and/or antagonists of mu-opioid receptors (MORs)in conscious rodents, we have shown that the MORs present in the walls of the portal vein controla gut—brain nervous circuit of induction of intestinal gluconeogenesis, a function control-ling hunger. In a second step, we have shown that peptides and protein digests promote aMOR-dependent induction of intestinal gluconeogenesis. The peptides have no effect in miceknockout for MOR, which are also insensitive to the satiety induced by protein diets. In addition,
∗ Faculté de médecine Laennec Lyon-Est, UMR Inserm U855, 7-11, rue Guillaume-Paradin, 69372 Lyon cedex 08, France.Adresse e-mail : [email protected]
007-9960/$ — see front matter © 2013 Société francaise de nutrition. Publié par Elsevier Masson SAS. Tous droits réservés.ttp://dx.doi.org/10.1016/j.cnd.2013.02.002

int
ulat regutinal
nut
étudié l’effet d’un REP chez le rat après un traitementpériportal par la capsaïcine, une substance qui inactiveles nerfs afférents. Aucune induction de l’activité de laglucose-6 phosphatase (G6Pase) ou de la phosphoénolpyru-vate carboxykinase-C (PEPCK-C), deux enzymes essentiellesde la NGI, n’était observée, contrairement aux animauxtémoins. Pour tester le rôle putatif des RMO dans cetteinduction, nous avons infusé des régulateurs RMO dans laveine porte de rats conscients, comme décrit précédem-ment [11]. Deux agonistes, la �1-7 casomorphine (dérivéede la caséine ß) et le DAMGO ont réduit l’activité G6Paseintestinale (Fig. 1). Au contraire, deux antagonistes RMO,la casoxin C (casoC) provenant de la kappa-caséine et lanaloxone (Nalox), l’ont induite (Fig. 1). Des résultats compa-rables ont été obtenus pour l’expression de la protéinePEPCK-C. La production de glucose intestinale (PGI), quan-tifiée à la fin de la perfusion de Nalox à l’aide d’un traceurmétabolique de glucose (tritié sur le carbone 3), représen-tait 25—30 % de la production de glucose endogène totale(PEG). Au contraire, aucune apparition de glucose n’étaitobservée chez des rats perfusés avec le DAMGO. En accordavec ce qu’on pouvait attendre de l’effet des effecteursRMO sur la néoglucogenèse intestinale, les rats perfu-sés avec un antagoniste RMO (CasoC) dans la veine porte
Rôle des récepteurs mu-opioïdes dans la régulation d’un axe
Mu-opioid receptors;Gut—brain axis
portal infusions of MOR-modgluconeogenesis. Thus, thecircuit of induction of intesinduced by dietary protein.© 2013 Société francaise de
Introduction
Dans le cadre de la compréhension des mécanismes del’homéostasie énergétique, un espace de plus en plus impor-tant de recherche concerne les signaux hormonaux queproduit l’intestin en réponse à l’absorption des nutrimentset qui modulent la faim. Cela concerne notamment les hor-mones gastro-intestinales telles la cholécystokinine (CCK),le glucagon-like peptide 1 ou encore le peptide YY. Il estintéressant de noter que le système nerveux entérique joueun rôle important dans la détection et la transmission dessignaux hormonaux au cerveau [1—3].
Dans ce cadre également, la néoglucogenèse intestinale(NGI) est une fonction nouvellement décrite [4—8] qui inter-fère dans le contrôle de l’homéostasie énergétique à l’étatpost-absorptif (pour revue, voir [9]). L’induction de la NGIse traduit par une libération de glucose dans la veine porte,qui collecte le sang de l’intestin entier. Sa détection parun capteur de glucose portal, récemment identifié commeétant le récepteur de glucose SGLT3 [10], et la transmissionde ce signal au cerveau par le système nerveux périphé-rique induisent une diminution de la sensation de faim etune amélioration de la sensibilité à l’insuline de la produc-tion hépatique de glucose. Cela concerne notamment deuxsituations nutritionnelles particulières, les régimes enrichisen protéines (REP) [11—13] et la chirurgie de l’obésité detype by-pass gastrique [14].
Une propriété connue depuis longtemps des protéinesest qu’un certain nombre d’entre elles, en particulierd’intérêt en nutrition humaine, comme les caséines du lait,contiennent des oligopeptides présentant une activité mu-
opioïde in vitro après digestion protéolytique partielle [15].Par ailleurs, une vaste littérature sur le sujet mentionnel’activité mu-opioïde d’oligopeptides variés [16]. Il est éga-lement admis que la modulation des récepteurs mu-opioïdes(RMO) centraux peut interférer avec le contrôle de la prisealimentaire. Les agonistes augmentent la prise alimentaire,tandis que les antagonistes l’inhibent [17]. Fait intéres-sant, les RMO sont exprimés dans l’intestin grêle, où ilscontrôlent la motilité intestinale [18,19]. Les protéines ali-mentaires sont absorbées depuis la lumière intestinale aprèsprotéolyse incomplète et pourraient donc exercer un rôlede signalisation systémique via les résidus protéolytiquesqu’elles libèrent [15]. Cependant, que ces derniers puissentatteindre le cerveau après ingestion orale est exclu en raisonde l’activité protéolytique du foie [20].Cela nous a conduit à tester l’hypothèse d’un rôle desRMO intestinaux dans le contrôle de la sensation de faimpar la NGI.
Les récepteurs mu-opioïdes sont présents dans lesneurones périportaux des rongeurs et de l’homme.
estin—cerveau 93
ors have no effect on food intake in mice deficient for intestinallation by the portal MORs and peptides of a gut—brain neural
gluconeogenesis is a causal link in the phenomenon of satiety
rition. Published by Elsevier Masson SAS. All rights reserved.
Les récepteurs mu-opioïdes de la veineporte régulent la néoglucogenèseintestinale via un circuit intestin—cerveau
Pour tester le rôle du système nerveux gastro-intestinalet/ou périportal dans l’induction de l’expression des gènesde la NGI par les protéines alimentaires, nous avons d’abord
Glc
6Pas
e (m
icro
mol
/min
/g)
Salin Nalox CasoC B1
30
25
20
15
10
5
0-7 DAMGO
*
*
**
Figure 1. Effet des modulateurs mu-opioïdes sur l’expressionde la Glc6Pase intestinale : les modulateurs RMO ont été perfu-sés pendant huit heures dans la veine porte de rats conscients àraison de 8,3 × 10−6 mmol/kg par minute. La Glc6Pase est doséedans un échantillon de jéjunum médian à la fin de la perfusion(moyenne ± SEM, n = 6). *, p < 0,05 (Tuckey’s post hoc test).

9
da
rhPrN1pdnlcpgcacglcmnqldvslcmCsra[
Lom
Qptnlldsapdtm
el
c
(dtoedilpltpoicl
TdPdp
vctTDsmpDcall
4
iminuaient leur prise alimentaire, tandis que ceux infusésvec un agoniste RMO (�1-7) l’augmentaient [16].
Les antagonistes mu-opioïdes perfusés dans la veineporte des rongeurs induisent la néoglucogenèseintestinale.
Un circuit nerveux intestin—cerveau est impliquédans cette régulation.
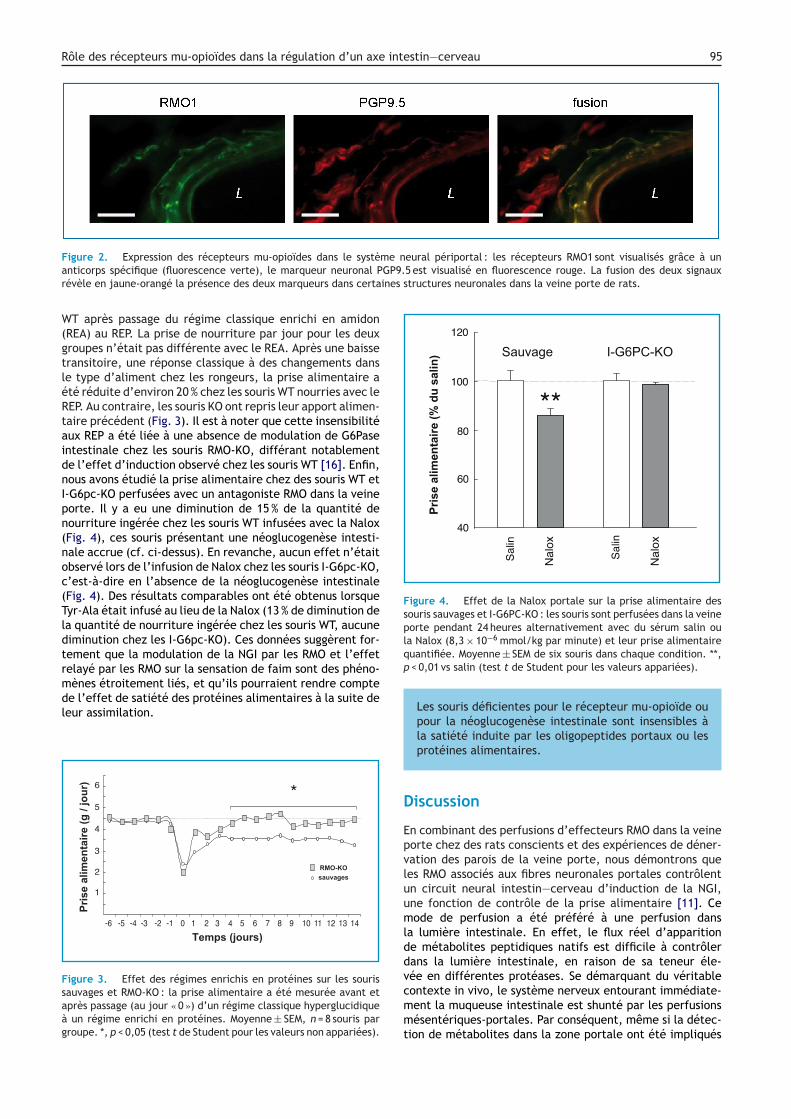
Nous avons utilisé trois méthodes pour préciser leôle joué par les RMO portaux. Les études immuno-istochimiques combinant les RMO et le marqueur neuronalGP9.5 ont révélé la présence des RMO dans les fibres neu-onales dans les parois de la veine porte de rat (Fig. 2).ous avons observé une co-localisation comparable de MOR-
et PGP9.5 dans les branches portales irriguant les espacesortes du foie humain. La présence des RMO dans les paroise la veine porte a été confirmée par western blot. Ensuite,ous avons étudié l’effet de la �1-7 ou de la CasoC sur’expression des gènes de la néoglucogenèse intestinalehez des rats traités par la capsaïcine. Il n’y avait alorslus d’effet sur les enzymes de la néoglucogenèse, sug-érant fortement que le système neuronal périportal estrucial pour la transmission du signal RMO. Enfin, nousvons évalué l’effet de �1-7 et Nalox sur les régions duerveau impliquées dans la réception des signaux d’origineastro-entérique : le complexe vagal dorsal (CVD), qui este principal récepteur du nerf vague, et le noyau parabra-hial (NPB), qui recoit principalement les afférences de laoelle épinière (pour revue : [1]). Par immuno-histochimie,
ous avons étudié l’expression de la protéine C-FOS, un mar-ueur bien connu de l’activation neuronale. La perfusion de’antagoniste Nalox a induit l’expression de C-FOS aussi bienans le CVD que le NPB, ce qui a bien montré que les voiesagale et spinale étaient impliquées dans la transmission duignal RMO au cerveau. La même activation de C-FOS avaitieu dans les principales régions hypothalamiques, qui sontonnectées au CVD et au NPB et qui contrôlent la prise ali-entaire (noyaux arqué et paraventriculaire notamment).
onfirmant le rôle des nerfs périphériques dans la transmis-ion du signal, aucune activation ne se produisait chez lesats dont la veine porte avait été traitée par la capsaïcineu moment de la chirurgie pour l’implantation du cathéter16].es digestats de protéines et lesligopeptides sont des antagonistesu-opioïdes
uand les rats sont nourris précédemment avec un REPendant trois jours, une augmentation marquée de la quan-ité des protéines G6Pase et PEPCK-C se produisait, commeous l’avions montré précédemment [11]. Chez ces rats,a perfusion d’agonistes RMO dans la veine porte diminuait’expression des deux protéines. Au contraire, aucun effet’induction supplémentaire n’avait lieu lors de la perfu-ion d’antagonistes RMO. La même tendance était observéevec un REP contenant un mélange de caséines du lait (quiouvait éventuellement libérer de la �1-7 ou de la CasoCans le sang), ou avec un REP à partir de protéines végé-ales (qui ne contenait pas ces peptides). Cela suggérait unécanisme commun d’induction par les antagonistes RMO
pCc
Lnd
PdrrlsltwsDpesé
G. Mithieux
t les protéines alimentaires de toute origine pour induire’expression des gènes de la NGI.
Pour documenter l’hypothèse ci-dessus, nous avonsombiné plusieurs approches.
Tout d’abord, nous avons infusé un digestat protéolytiquepeptones) ou des oligopeptides choisis (di- ou tri-peptides)ans la veine mésentérique portale des rats. Les struc-ures suivantes ont été choisies pour représenter les di-u tri-peptides : avec ou sans une chaîne latérale (Tyr-Alat Gly-Gly, respectivement), ou avec une charge électriqueans la chaîne latérale (Phe-Pro-Arg). Dans tous les cas, unenduction marquée de la G6Pase a été observée à la fois danse jéjunum et dans l’iléon, une partie de l’intestin d’activitélus faible de la G6Pase à l’état basal, mais inductible danse cas des REP [11]. En outre, des études de dilution deraceur glucose ont révélé que la NGI prend place après laerfusion portale de Tyr-Ala ou de peptones. Enfin, commebservé précédemment pour les antagonistes RMO, aucunenduction de la G6Pase par les oligopeptides n’a eu lieuhez les rats dont la zone périportale a été dénervée para capsaïcine.
Deuxièmement, chez les rats perfusés avec le di-peptideyr-Ala, il y a eu une multiplication par trois de l’expressione C-FOS dans le DVC, et dans l’hypothalamus, le NPB et leAG, tout comme chez le rat perfusé avec la Nalox (cf. ci-essus). De plus, aucune augmentation de C-FOS ne prenaitlace après l’inactivation des afférences nerveuses portales.
Troisièmement, nous avons testé si les oligopeptides pou-aient se comporter comme des antagonistes RMO dans uneulture de neuroblastomes exprimant de manière consti-utive les RMO [21,22]. Ainsi, nous avons montré queyr-Ala et Gly-Gly entrent en compétition pour la liaison deAMGO tritié, puis nous avons étudié l’effet d’oligopeptidesur le couplage des RMO à l’adénylate cyclase dans lesêmes cellules. Le RMO est couplé à l’adénylate cyclasear l’intermédiaire d’une protéine G inhibitrice. Ainsi, leAMGO (agoniste) diminuait fortement la teneur en AMPcellulaire. Au contraire, la CasoC (antagoniste) et la Naloxugmentaient considérablement la teneur en AMPc cellu-aire. Il est à noter que tous les oligopeptides augmentaienta teneur en AMPc et empêchaient la diminution induite
ar le DAMGO dans des expériences de co-incubation [16].ela suggère fortement que les oligopeptides se comportentomme des antagonistes RMO in vitro.’induction mu-opioïde-dépendante de laéoglucogenèse intestinale est causaleans l’effet de satiété des protéines
our démontrer le rôle causal des RMO dans l’inductione la NGI par les protéines en cours d’absorption et sonôle dans la satiété associée, nous avons réalisé des expé-iences de perfusion portale sur des souris knockout poure gène codant les RMO (RMO-KO) et sur des souris pré-entant une suppression de la G6Pase spécifiquement dans’intestin (I-G6pc-KO, la G6PC étant la sous-unité cataly-ique de la G6Pase). Lorsqu’ils sont perfusés chez les sourisild-type (WT), les di- et tri-peptides et les peptones indui-
aient l’activité G6Pase dans le jéjunum. Au contraire, leAMGO supprimait l’activité G6Pase et cela était inverséar les peptones dans des expériences de co-perfusion. Ilst à noter qu’aucun de ces effecteurs n’induisait d’effetur la NGI chez des souris RMO-KO [16]. En outre, nous avonstudié la prise alimentaire des souris RMO-KO et des souris
intestin—cerveau 95
me neural périportal : les récepteurs RMO1 sont visualisés grâce à unGP9.5 est visualisé en fluorescence rouge. La fusion des deux signauxines structures neuronales dans la veine porte de rats.
Sauvage I-G6PC-KO
alin
)
Rôle des récepteurs mu-opioïdes dans la régulation d’un axe
Figure 2. Expression des récepteurs mu-opioïdes dans le systèanticorps spécifique (fluorescence verte), le marqueur neuronal Prévèle en jaune-orangé la présence des deux marqueurs dans certa
WT après passage du régime classique enrichi en amidon(REA) au REP. La prise de nourriture par jour pour les deuxgroupes n’était pas différente avec le REA. Après une baissetransitoire, une réponse classique à des changements dans
le type d’aliment chez les rongeurs, la prise alimentaire aété réduite d’environ 20 % chez les souris WT nourries avec leREP. Au contraire, les souris KO ont repris leur apport alimen-taire précédent (Fig. 3). Il est à noter que cette insensibilitéaux REP a été liée à une absence de modulation de G6Paseintestinale chez les souris RMO-KO, différant notablementde l’effet d’induction observé chez les souris WT [16]. Enfin,nous avons étudié la prise alimentaire chez des souris WT etI-G6pc-KO perfusées avec un antagoniste RMO dans la veineporte. Il y a eu une diminution de 15 % de la quantité denourriture ingérée chez les souris WT infusées avec la Nalox(Fig. 4), ces souris présentant une néoglucogenèse intesti-nale accrue (cf. ci-dessus). En revanche, aucun effet n’étaitobservé lors de l’infusion de Nalox chez les souris I-G6pc-KO,c’est-à-dire en l’absence de la néoglucogenèse intestinale(Fig. 4). Des résultats comparables ont été obtenus lorsqueTyr-Ala était infusé au lieu de la Nalox (13 % de diminution dela quantité de nourriture ingérée chez les souris WT, aucunediminution chez les I-G6pc-KO). Ces données suggèrent for-tement que la modulation de la NGI par les RMO et l’effetrelayé par les RMO sur la sensation de faim sont des phéno-mènes étroitement liés, et qu’ils pourraient rendre comptede l’effet de satiété des protéines alimentaires à la suite deleur assimilation.Pris
e al
imen
taire
(g /
jour
)
Temps (jours)
RMO-KOsauvages
Figure 3. Effet des régimes enrichis en protéines sur les sourissauvages et RMO-KO : la prise alimentaire a été mesurée avant etaprès passage (au jour « 0 ») d’un régime classique hyperglucidiqueà un régime enrichi en protéines. Moyenne ± SEM, n = 8 souris pargroupe. *, p < 0,05 (test t de Student pour les valeurs non appariées).
ire (%
du
s
Prise
alim
enta
Figure 4. Effet de la Nalox portale sur la prise alimentaire dessouris sauvages et I-G6PC-KO : les souris sont perfusées dans la veineporte pendant 24 heures alternativement avec du sérum salin oula Nalox (8,3 × 10−6 mmol/kg par minute) et leur prise alimentairequantifiée. Moyenne ± SEM de six souris dans chaque condition. **,p < 0,01 vs salin (test t de Student pour les valeurs appariées).
Les souris déficientes pour le récepteur mu-opioïde oupour la néoglucogenèse intestinale sont insensibles àla satiété induite par les oligopeptides portaux ou lesprotéines alimentaires.
Discussion
En combinant des perfusions d’effecteurs RMO dans la veineporte chez des rats conscients et des expériences de déner-vation des parois de la veine porte, nous démontrons queles RMO associés aux fibres neuronales portales contrôlentun circuit neural intestin—cerveau d’induction de la NGI,une fonction de contrôle de la prise alimentaire [11]. Cemode de perfusion a été préféré à une perfusion dansla lumière intestinale. En effet, le flux réel d’apparitionde métabolites peptidiques natifs est difficile à contrôlerdans la lumière intestinale, en raison de sa teneur éle-vée en différentes protéases. Se démarquant du véritablecontexte in vivo, le système nerveux entourant immédiate-ment la muqueuse intestinale est shunté par les perfusionsmésentériques-portales. Par conséquent, même si la détec-tion de métabolites dans la zone portale ont été impliqués

9
dêle[
mllet[jtcatàpcmvtRptcppcpdese
stuhamlrddlpdccdapc[dnjedaqpct
miclacctltpFtmslb[cmf[qp[adld(dCàd
D
Lt
6
ans les conditions expérimentales étudiées ici, il ne peuttre exclu qu’une détection RMO-dépendante puisse avoirieu en amont (par exemple dans le système neuronal quintoure la muqueuse de l’intestin ou la couche musculaire)18,23], en dehors de ce contexte expérimental.
Il est établi que les protéines sont digérées incomplète-ent à l’intérieur de la lumière du tube digestif et qu’elles
ibèrent des fragments d’oligopeptides composés principa-ement de di- ou tri-peptides [24]. Ces oligopeptides sontnsuite transportés dans les entérocytes au moyen d’unransporteur spécifique (PEPT1) de la membrane luminale25,26]. Une fraction subit une protéolyse complémentaireusqu’au stade d’acide aminé pour alimenter la protéosyn-hèse intestinale, connue pour être très intense. Il est moinsonnu qu’une autre fraction est libérée dans le sang portal,vec des acides aminés, par l’intermédiaire de transpor-eurs spécifiques d’oligopeptides et d’acides aminés situés
la membrane basolatérale [27]. Un certain nombre derotéines libèrent des peptides à activité mu-opioïde (voiri-dessus). Toutefois, l’hypothèse que toute protéine ali-entaire peut induire la néoglucogenèse intestinale via une
oie dépendante des RMO implique que les digestats de pro-éines de toutes origines devraient présenter une activitéMO-antagoniste. Cette inférence est fortement soutenuear l’observation que tous les peptides étudiés dans ceravail, que ce soit seuls ou en mélange (peptones), seomportent comme des antagonistes RMO à la fois in vivoour induire la NGI et in vitro dans des expériences de cou-lage à l’adénylate cyclase. L’absence de ces mécanismeshez la souris RMO-KO et leur insensibilité à l’alimentationrotéique, ainsi que l’incapacité des antagonistes RMO etes peptides à moduler la prise alimentaire chez la sourisn absence de NGI, confirment formellement le lien de cau-alité entre l’antagonisation des RMO par les oligopeptidest l’effet de satiété induit par les protéines alimentaires.
Une question intéressante a trait à la complexité de laéquence des événements qui permet aux protéines alimen-aires d’exercer au final leur effet de satiété. En effet,n certain nombre de facteurs connus, par exemple lesormones intestinales, libérées par l’intestin en réponseu repas, sont supposées moduler la consommation ali-
entaire en se liant directement à leurs récepteurs dans’hypothalamus. Il s’agit de mécanismes de mise en placeapide. Dans le cas des protéines alimentaires, une série’événements initie d’abord le programme d’expressiones gènes de la néoglucogenèse dans l’intestin, par’intermédiaire d’un arc réflexe avec le cerveau. Cela estrogressif et se produit au cours de la période postpran-iale. Après cela, le glucose libéré peut induire son actionentrale via les mécanismes de détection portale du glu-ose [10,11]. Il s’agit d’un phénomène basé sur l’inductione gènes et qui peut donc perdurer après l’assimilation desliments. Cela est en accord avec le fait que l’alimentationrotéique diminue la faim à distance du repas précédent,e qui est la définition du phénomène appelé « satiété »28,29]. Cet effet peut se prolonger en termes de contrôlee la faim, c’est-à-dire que l’alimentation protéique dimi-ue la prise alimentaire sur le long terme, pendant plusieursours ou semaines [11,13,30]. Ainsi, des avantages existentn termes de réduction du poids corporel et de la masseu tissu adipeux [11,30]. Au contraire, les rongeurs traitésvant les repas avec la CCK (une hormone de rassasiement,ui est impliquée dans la terminaison des repas) ne réduisentas leur consommation alimentaire globale. En effet, ilsompensent la diminution de la taille des repas en augmen-ant le nombre de repas par jour [31].
R
Lpc
R
G. Mithieux
En conclusion, les RMO exprimés dans la zoneésentérique portale contrôlent un circuit neuronal
ntestin—cerveau de régulation de la NGI. Cette dernièreontrôle la prise alimentaire [11,13,14]. Jusqu’à présent,e rôle régulateur des RMO dans le contrôle de la priselimentaire avait été documenté dans le système nerveuxentral, par l’intermédiaire de leur implication dans lesircuits dits de « récompense » [32,33]. De plus, les pro-éines alimentaires initient leurs effets de satiété à travers’activité antagoniste �-opioïde de leurs résidus de diges-ion protéolytique, en agissant uniquement sur ce circuitrécédemment insoupconné de neurones intestin—cerveau.ait remarquable, divers antagonistes �-opioïdes adminis-rés par voie orale diminuent la faim chez les humains [17],algré le fait qu’ils n’atteignent pas le cerveau en rai-
on d’un métabolisme hépatique très actif [34]. En outre,’effet de satiété des protéines alimentaires concerne aussiien des espèces animales variées que les êtres humains11,28—30]. Le by-pass gastrique, une opération chirurgi-ale de plus en plus utilisée dans le traitement de l’obésitéorbide, induit une réduction dramatique de la sensation de
aim en quelques jours. Cela est vrai chez les souris obèses14] et chez les humains obèses (pour revue voir [35]). Alorsue la NGI s’est révélée essentielle dans la diminution de larise de nourriture après by-pass gastrique chez la souris14], une NGI a été observée dans des conditions post-bsorptives plusieurs jours après un by-pass gastrique chezes patients obèses non-diabétiques [36,37]. Il faut rappe-er ici que des fibres nerveuses RMO-positives sont présentesans la paroi des ramifications de la veine porte humainecf. ci-dessus). Ces données suggèrent que les mécanismesécouverts ici peuvent également avoir lieu chez l’homme.es nouvelles connaissances pourraient ainsi ouvrir la voie
des approches futures de traitement et/ou de préventiones maladies métaboliques.
éclaration d’intérêts
’auteur déclare ne pas avoir de conflits d’intérêts en rela-ion avec cet article.
emerciements
’auteur remercie les membres de son équipe, qui ontresque tous participé à des titres divers à l’acquisition dees résultats.
éférences
[1] Berthoud HR. Anatomy and function of sensory hepatic nerves.Anat Rec A Discov Mol Cell Evol Biol 2004;280:827—35.
[2] Moran TH. Cholecystokinin and satiety: current perspectives.Nutrition 2000;16:858—65.
[3] Vahl TP, Tauchi M, Durler TS, Elfers EE, Fernandes TM, Bit-ner, et al. Glucagon-like peptide-1 (GLP-1) receptors expressedon nerve terminals in the portal vein mediate the effects ofendogenous GLP-1 on glucose tolerance in rats. Endocrinology2007;148:4965—73.
[4] Croset M, Rajas F, Zitoun C, Hurot JM, Montano S, Mithieux G.Rat small intestine is an insulin-sensitive gluconeogenic organ.Diabetes 2001;50:740—6.
[5] Mithieux G, Rajas F, Gautier-Stein A. A novel role for glucose-6 phosphatase in the small intestine in the control of glucosehomeostasis. J Biol Chem 2004;279:44231—4.

int
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
Rôle des récepteurs mu-opioïdes dans la régulation d’un axe
[6] Mithieux G, Bady I, Gautier A, Croset M, Rajas F, Zitoun C.Induction of control genes in intestinal gluconeogenesis issequential during fasting and maximal in diabetes. Am J PhysiolEndocrinol Metab 2004;286:E370—5.
[7] Rajas F, Croset M, Zitoun C, Montano S, Mithieux G. Induction ofPEPCK gene expression in insulinopenia in rat small intestine.Diabetes 2000;49:1165—8.
[8] Rajas F, Bruni N, Montano S, Zitoun C, Mithieux G. Theglucose-6 phosphatase gene is expressed in human and rat smallintestines: regulation of expression in fasted and diabetic rats.Gastroenterology 1999;117:132—9.
[9] Mithieux G, Magnan C. La néoglucogenèse intestinale : un nou-vel acteur du contrôle de la prise alimentaire. Cah Nutr Diet2006;41:211—5.
10] Delaere F, Duchampt A, Mounien L, Seyer P, Duraffourd C,Zitoun C, et al. The role of sodium-coupled glucose co-transporter 3 in the satiety effect of portal glucose sensing.Mol Metab 2013;2:47—53.
11] Mithieux G, Misery P, Magnan C, Pillot B, Gautier-Stein A, Ber-nard, et al. Portal sensing of intestinal gluconeogenesis is amechanistic link in the diminution of food intake induced bydiet protein. Cell Metab 2005;2:321—9.
12] Pillot B, Soty M, Gautier-Stein A, Zitoun C, Mithieux G. Proteinfeeding promotes redistribution of endogenous glucose produc-tion to the kidney and potentiates its suppression by insulin.Endocrinology 2009;150:616—24.
13] Penhoat A, Mutel E, Amigo-Correig M, Pillot B, Stefanutti
A, Rajas F, et al. Protein-induced satiety is abolished inthe absence of intestinal gluconeogenesis. Physiol Behav2011;105:89—93.14] Troy S, Soty M, Ribeiro L, Laval L, Migrenne S, Fioramonti X,et al. Intestinal gluconeogenesis is a key factor for early meta-bolic changes after gastric bypass but not after gastric lap-bandin mice. Cell Metab 2008;8:201—11.
15] Zioudrou C, Streaty RA, Klee WA. Opioid peptides derived fromfood proteins. The exorphins. J Biol Chem 1979;254:2446—9.
16] Duraffourd C, De Vadder F, Goncalves D, Delaere F, Penhoat A,Brusse B, et al. Mu-opioid receptors and dietary protein sti-mulate a gut-brain neural circuitry limiting food intake. Cell2012;150:377—88.
17] Yeomans MR, Gray RW. Opioid peptides and the controlof human ingestive behaviour. Neurosci Biobehav Rev2002;26:713—28.
18] Hedner T, Cassuto J. Opioids and opioid receptors in peripheraltissues. Scand J Gastroenterol 1987;Suppl. 130:27—46.
19] Sternini C, Patierno S, Selmer IS, Kirchgessner A. The opioidsystem in the gastrointestinal tract. Neurogastroenterol Motil2004;16:3—16.
20] Kreil G, Umbach M, Brantl V, Teschemacher H. Studies onthe enzymatic degradation of beta-casomorphins. Life Sci1983;33(Suppl. 1):137—40.
21] Lorentz M, Hedlund B, Arhem P. Morphine activates calciumchannels in cloned mouse neuroblastoma cell lines. Brain Res1988;445:157—9.
[
[
[
[
[
[
[
estin—cerveau 97
22] Yang JC, Shan J, Ng MBChBa KF, Pang P. Morphine and metha-done have different effects on calcium channel currents inneuroblastoma cells. Brain Res 2000;870:199—203.
23] Sternini C. Receptors and transmission in the brain-gut axis:potential for novel therapies III. �-opioid receptors in the ente-ric nervous system. Am J Physiol Gastrointert Liver Physiol2001;281:G8—15.
24] Daniel C. Molecular and integrative physiology of intestinalpeptide transport. Annu Rev Physiol 2004;66:361—84.
25] Boll M, Markovich D, Weber WM, Korte H, Daniel H, MurerH. Expression cloning of a cDNA from rabbit small intestinerelated to proton-coupled transport of peptides, beta-lactamantibiotics and ACE-inhibitors. Pflugers Arch 1994;429:146—9.
26] Nielsen CU, Brodin B. Di/tri-peptide transporters as drugdelivery targets: regulation of transport under physiologi-cal and patho-physiological conditions. Curr Drug Targets2003;4:373—88.
27] Lee VH. Membrane transporters. Eur J Pharm Sci 2000;Suppl.2:S41—50.
28] Booth DA, Chase A, Campbell AT. Relative effectiveness of pro-tein in the late stages of appetite suppression in man. PhysiolBehav 1970;5:1299—302.
29] Rolls BJ, Hetherington M, Burley VJ. The specificity ofsatiety: the influence of foods of different macronutrientcontent on the development of satiety. Physiol Behav 1988;43:145—53.
30] Jean CGF, Tome D, Larue-Achagiotis C. Wistar rats allo-
wed to self-select macronutrients from wearing to maturitychoose a hith-protein, high-lipid diet. Physiol Behav 2002;76:65—73.31] West DB, Fey D, Woods SC. Cholecystokinin persistently sup-presses meal size but not food intake in free-feeding rats. AmJ Physiol 1984;150:1—22.
32] Glass MJ, Billington CJ, Levine AS. Opioids and foodintake distributed functional neural pathways? Neuropeptides1999;33:360—8.
33] Shin AC, Pistell PJ, Phifer CB, Berthoud HR. Reversiblesuppression of food reward behavior by chronic �-opioidreceptor antagonism in the nucleus accumbens. Neuroscience2010;170:580—8.
34] Reimer K, Hopp M, Zenz M, Maier C, Holzer P, Mikus, et al.Meeting the challenges of opioid-induced constipation in chro-nic pain management — a novel approach. Pharmacology2009;83:10—7.
35] Thaler JP, Cummings DE. Minireview: hormonal and metabolicmechanisms of diabetes remission after gastrointestinal sur-gery. Endocrinology 2009;150:2518—25.
36] Hayes MT, Foo J, Besic V, Tychinskaya Y, Stubbs RS. Is intes-tinal gluconeogenesis a key factor in the early changesin glucose homeostasis following gastric bypass? Obes Surg2011;21:759—62.
37] Mithieux G. Comment about intestinal gluconeogenesis aftergastric bypass in human in relation with the paper by Hayeset al., Obes Surg 2011. Obes Surg 2012;22:1920—2.