Embed Size (px)
Citation preview

This article was downloaded by: [University of Aberdeen]On: 05 October 2014, At: 22:47Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH,UK
Acta Botanica Gallica: BotanyLettersPublication details, including instructions forauthors and subscription information:http://www.tandfonline.com/loi/tabg20
Tendances évolutives de lastructure exinique dans lesgenres Ulex, Stauracanthuset Genista (Genisteae,Papilionoideae: Leguminosae)Cristina Pardo a , Paloma Cubas a & Pilar SánchezTestillano ba Departamento de Biología vegetal II, Facultad deFarmacia , Universidad Complutense , E-28040 ,Madridb Centro de Investigaciones Biológicas , CSIC ,Velázquez 144, E-28014 , MadridPublished online: 27 Apr 2013.
To cite this article: Cristina Pardo , Paloma Cubas & Pilar Sánchez Testillano (1994)Tendances évolutives de la structure exinique dans les genres Ulex, Stauracanthuset Genista (Genisteae, Papilionoideae: Leguminosae), Acta Botanica Gallica: BotanyLetters, 141:2, 195-205, DOI: 10.1080/12538078.1994.10515151
To link to this article: http://dx.doi.org/10.1080/12538078.1994.10515151
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all theinformation (the “Content”) contained in the publications on our platform.However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness,or suitability for any purpose of the Content. Any opinions and viewsexpressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of theContent should not be relied upon and should be independently verified withprimary sources of information. Taylor and Francis shall not be liable for any

losses, actions, claims, proceedings, demands, costs, expenses, damages,and other liabilities whatsoever or howsoever caused arising directly orindirectly in connection with, in relation to or arising out of the use of theContent.
This article may be used for research, teaching, and private study purposes.Any substantial or systematic reproduction, redistribution, reselling, loan,sub-licensing, systematic supply, or distribution in any form to anyone isexpressly forbidden. Terms & Conditions of access and use can be found athttp://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
Uni
vers
ity o
f A
berd
een]
at 2
2:47
05
Oct
ober
201
4

Acta bot. CaUica, 1994, 141 (2), 195-205.
Tendances evolutives de Ia structure exinique dans les genres Ulex, Stauracanthus et Genista (Genisteae, Papilionoideae : Leguminosae)
par Cristina Pardo (1), Paloma Cubas (I) et Pilar Sanchez Testillano (2)
(')Departamento de Biologia vegetal II, Facultad de Farmacia, Uuiversidad Complutense, E-28040 Madrid ( 2) Centro de Investigaciones Biologicas, CSIC, Velazquez 144, E-28014 Madrid
Resume.- La tribu des Genisteae presente une morphologie pollinique trils homogilne qui contrasts avec une trils grande variabilite de Ia macro-morpho· Iogie et des nombres chromosomiques, qui correspondent A dilferents niveaux de plo"idie. Cependant, dans Ia parol pollinique, quatre types structuraux (trois chez Ulex et Stauracanthus et quatre chez Genista) sent reconnus et corrales avec le niveau de plo"idie : le type I, avec une structure exinique compacta et un inlratectum peu developpe, inclut presque tous les taxons diplo"ides ; le type II A inlratectum compose A Ia lois de grains et de columelles chez les taxons et populations tetraplo"ides ; le type Ill A lnlratectum columellaire const~ue partois par empilement de grains, present dans les taxons et populations diplo"ides, tetraplo'ides et hexaplo"ides ; et 1e type IV dont toutes les couches exiniques sent constituees de grains, observe seulement chez les taxons octoplo"ides.
Summary.- The pollen morphology in the Genisteae is very homogeneous desp~e the great range of variability displayed by this tribe in macromorphology, chromosome numbers and ploidy levels. However, the wall structure of pollen grains of U/ex, Stauracanthus and Genista shows important differences which allow recognition of four types of exine structure. These types closely correlate with the ploidy levels. Type I, found in most of the diploids, Is characterized by a compact structure of the pollen wall, and an lnlratectum illdeveloped and mainly composed of granules. Type II, characterized by an inlratectum composed of both granules and columellae. This structure is commonly found in pollen grains of tetraploids. Type Ill, showing an inlratectum composed of well developed columellae which are often formed of pillared granules. Pollen grains of diploids, tetraploids and hexaploids show this type of exine structure. In type IV, all the pollen wall layers are formed of densely packed granules which often form columellae. This unusual structure has been only observed in octoploids.
Key word3 : exine structure - Ulex - Stauracanthus - Genista
© Soci8t8 botanique de France 1994. ISSN 1253-8078.
Dow
nloa
ded
by [
Uni
vers
ity o
f A
berd
een]
at 2
2:47
05
Oct
ober
201
4

196 ACTA BOTANICA GALLICA
INTRODUCTION
La tribu d~s Cenisteae prcscnte un pollen pluti)t homogcne, tricolporo'ide, a symctrie rildiale, isopolaire, sphcro'idal en vue mcridicnnc, variant de suboblatc a subprobtc, ct subcirculaire, lcgcrcmcnt lobe ou subtriangulaire en vue polaire. L'ectoapcrture de type colpe, subterminalc ou tcrminale, est constituce par un pli de l'cndexine fortement cpaissie : l'endoaperture est irreguliere, de type poro'ide. L'ornementation varie de perforce a reticulce-pcrforce (Prados et al., 1985). La structure du sporodermc (Ferguson et Skvarla, 1981 ; Missct et al., 1982 ; Cubas et Pardo, 1992) est constituec par une endexine mince ct continue, une foot layer assez epaissc, un infratectum grenu ou columeUaire et un tectum epais avec de rares perforations (Pl. I, A; Pl. II, A).
L'homogcneite pollinique de Ia tribu des Cenisteae contraste avec Ia grande diversitc des nombres chromosomiques qu'elle prcsente et encore davantage avec Ia grande variabilite macro-morphologique (Vicioso, 1953, 1962). Pour Saiiudo (1979), les phenomenes d'euplo'idie en sont essentieUement Ia cause et, dans une moindre mcsure, ceux d'aneuplo'idie. L:t proportion des polyplo'ides est tres elcvee par rapport aux diplo'idcs, etant les tctraplo'ides les plus frequents chez les Cenisteae (Saiiudo, 1979).
Deux grands groupes ou alliances sont rcconnus dans Ia tribu des Cenisteae : l'un centre autour de Cytisus et l'autre autour de Cenista (Cytisus and Cenista groups, Gibbs, 1966 ; PolhiU, 1976; llisby, 1981; Cristofolini et Feoli-Chiapella, 1984). Dans ce deuxieme groupe, nous avons concentre notre etude sur Cenista, comme genre central, et sur Ulex ct Stauracanthus considcres comme genres cxtrcmemcnt derives dans Ia lignce evolutive de Genis-
ta (Fcoli-Chiapclla et Cristofolini, 1981 ).
La diversification des genres Ulex et Stauracanthus paralt essentiellcment due a des phenomenes de polyplo'idie : lcs nombres chromosomiques n = 16, 32 et 48 ont ete rapportes chez Ulex, et n = 24, 48 et 72 chez Stauracanthus, c'est-a-dire, les niveaux 2x, 4x et 6x sont presents dans ces genres, pour un nombre de base actuel x = 16 pour Ulex et x = 24 pour Stauracanthus (Cubas, 1986).
La diversification du genre Cenista est beaucoup plus complexe : clle n'est pas seulement due aux phenomenes de polyplo'idie et d'aneuplo'idie mais aussi a une plus grande diversite des nombres de base (il existe une serie presque continue den= 9 jusqu'a n = 60. Saiiudo ( 1979) donne deux nombres de base actuels pour Cenista, x = 9 et x = 12, done il interprete que lcs taxons de Cenista correspondent aux niveaux 2x, 4x, 6x, 8x et lOx en rapport avec ccs deux nombres de base. Par contre, Verlaque (1992), qui donne x = 6 comme nomhre de base primaire pour Cenista, considere que les differents nomhres chromosomiques rapportcs dans ce genre correspondent aux niveaux 3x, 4x, 5x, 6x, 7x, 8x, 12x, 14x, l6x.
MATERIEL ET METHODES
Le materiel pollinique provient de 36 taxons d'Uiex, Stauracanthus et Gem'sta. Les echantillons sont depo· ses dans les herbiers de Ia Faculte de Pharmacie de Madrid (MAF) et du Jardin royal botanique de Madrid (MA). Leur provenance et Ia taxonomie utilisee sont indiquees en annexa. Pour les correlations entre las donnees palynologiques at las nombres chromosomiques (ou niveaux de plo'idie). trois sources d'information ont ate utilisees : les echantillons marques (1) ont ate etudies du point de vue palynologique at caryologique ; les echantillons marques avec (2) ont ate seule· ment etudies palynologiquement, mais ils proviennent de populations proches de celles-ci etudiees caryologiquement par diHerents auteurs ou nous-~mes ; et pour le reste du materiel (3), les nombres chromosomiques ont ate pris dans Ia bibliographie.
Dow
nloa
ded
by [
Uni
vers
ity o
f A
berd
een]
at 2
2:47
05
Oct
ober
201
4

C. PARDO, P. CUBAS ET P. SANCHEZ TESTILLANO 197
Taus las taxons des genres Ulex at Stauracanthus ant ate etudies. Dans Ia cas de Genista, tres riche en taxons, una representation iberique des differents niveaux de plo.idie ainsi que quelques aneuplo"ides ant ate etudies. Pour ce laire, nous las avons groupes salon las nombres de base actuels x = 9 at x = 12 (Salludo, 1979), bien que Verlaque (1992) ail propose comma unique nombre da base x = 6. Ce dernier nombra changerait les niveaux :le plo"idie indiques pour chaque taxon dans cella etude mais nullement les conclusions palynologiques.
Le pollen a ate acetolyse par Ia technique d'Erdtman (1952). Des coupes semi-lines ant ate ellectuees au microtome c\ congelation salon Muller (1973), et des coupes ultra-lines sur du materiel fixe dans Ia glutaraldehyde at post-fixe par Ia tetroxyde d'osmium. Les inclusions ant ate lanes dans l'epon at las coupes colorees par l'acetate d'uranyle 5 %. Las observations ant ate realisees avec un microscope electronique c\ balayage ISI-SX-25 et c\ transmission E.M. Philips 300.
RESULTATS
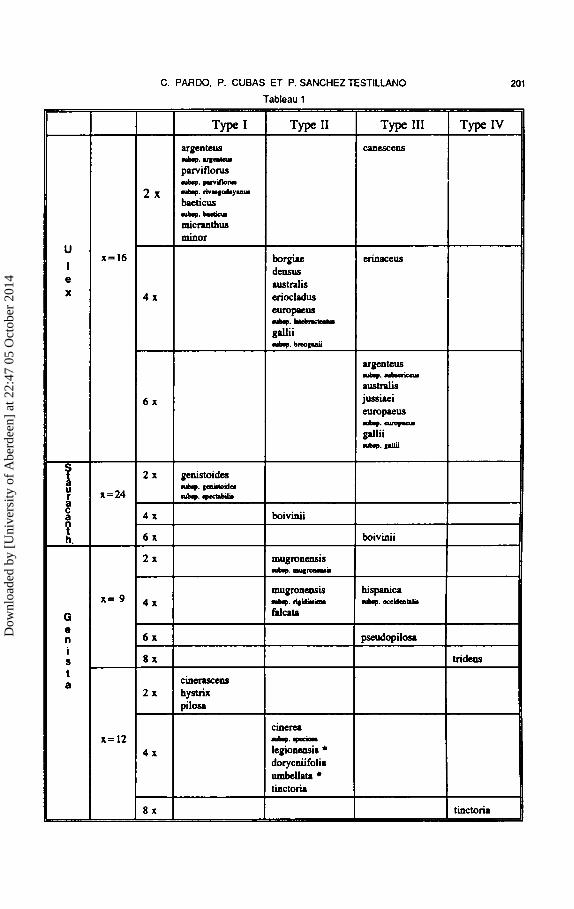
Chez Ulex, trois types structuraux de Ia paroi pollinique extt·a-aperturale ont ete etablis (Cubas et Pardo, 1992; Tableau 1). Le type I est caracterise par une structure compacte, un tectum et une nexine epais par rapport a l'infratectum peu epais et compose de quelques grains individualises partant de Ia nexine et du tectum (e.g. U. parviflorus subsp. rivasgodayanus, Pl. I, B), melanges a des elements ectexiniques radiaux, parfois incomplets. Le type II, presente un infratectum a peu pres de meme epaisseur que Ia nexine et le tectum, compose de grains et de columelles, les grains pouvant etre libres, groupes en amas ou empilf,s formant de courtes columelles pas tres ncttes (e.g. U. australis, Pl. I, C). Le type III est caracterise par un tectum, une nexine et un infratectum bien developpes, cedernier formant Ia couche Ia plus epaisse. Cette couche est composee de columelles parfois constituces de grains empilcs et orientes radialement (e.g. U. australis, Pl. I, D).
Trois types structuraux semblables ont pu etre reconnus chez Stauracanthus (Tableau 1). Le type I est
present dans les deux sous-especcs de St. genistoides (subsp. genistoides, Pl. 1, E, et subsp. spectabilis, Pl. I, F). Les types II et III sont presents chez St. boivinii (Pl. I, Get H).
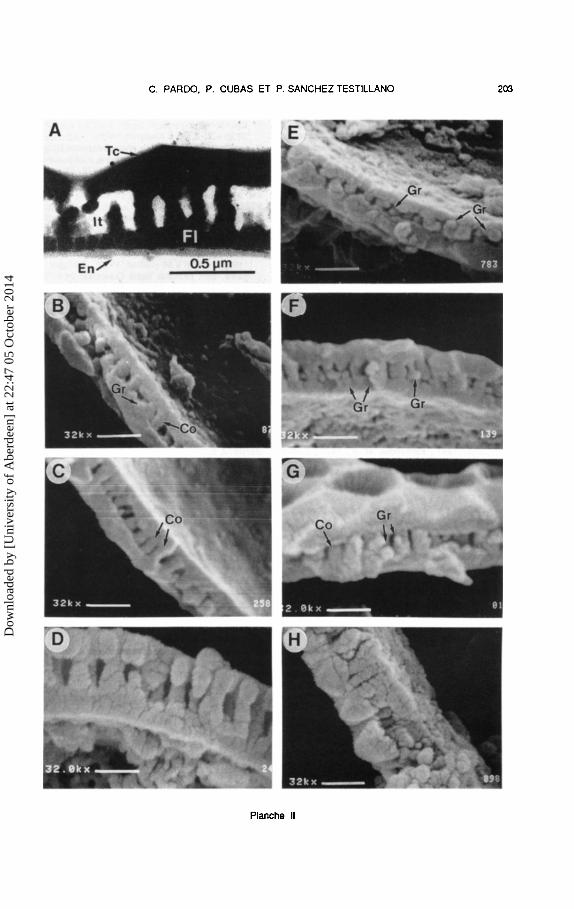
Chez Genista (Tableau 1), on observe une tendance semblable a celle d' Ulex et de Stauracanthus mais avec des legeres differences. La diversification plus importante (un plus grand nombre d'especes et nombres chromosomiques) chez Genista est parallele a celle que l'on observe dans les modeles structuraux de Ia paroi pollinique qui peuvent etre reunis dans 4 types, un de plus que chez Ulex et Stauracanthus. Les types I, II et III (Pl. II, B-G) sont a peu pres semblables a ceux decrits pour Ulex et Stauracanthus. Le type nouveau, type IV, presente une exine tres epaisse par rapport aux autres types du genre Genista ; toutes les couches exiniques sont constituees de grains, ceux de !'infratectum pouvant etre nettement empiles, ce qui donne une apparence columellaire a cette couche (Pl. II, D) ou moins organises (Pl. II, H).
Dans Ia serie de taxons a nombres chromosomiques multiples de x = 9, le type I n'a pas ete trouve. Le type II, a infratectum parfois tres mince, est present chez G. mugronensis subsp. mugronensis, G. mugronensis subsp. rigidissima et G. falcata (Pl. II, B). Le type III est present dans G. hispanica subsp. occidentalis et G. pseudopilosa (Pl. II, C). Le type IV est represente par G. tridens (Pl. II, D).
Dans Ia serie de taxons a nombres chromosomiques multiples de x = 12, les types I, II et IV ont ete rencontres. Le type I, a infratectum grenu et nexine plus mince que le tectum, est present chez G. cinerascens (Pl. II, E), G. hystrix et G. pilosa. Le type II inclut G. cinerea subsp. speciosa, G. legionensis (Pl. II, F), G. dorycniifolia, G. umbellata et G. tinctoria (Pl. II, G). Le type IV est represente dans G. tinctoria (Pl. II, H).
Dow
nloa
ded
by [
Uni
vers
ity o
f A
berd
een]
at 2
2:47
05
Oct
ober
201
4

198 ACTA BOTANICA GALLICA
DISCUSSION
L'etude ultrastructurale de Ia pa,·oi pollinique montre que chez Ulex et Stauracanthus il y a correlation entre lcs types structuraux et les niveaux de plo"idie (Tableau 1). Chez Ulex et Stauracanthus, tous les diploldes appartiennent au type I, sauf Ulex canescens. Le type II est present dans tous les tetraploldes (a !'exception d'U. erinaceus). Le type III est present dans tous les hexaploldes ainsi que chez U. canescens (2x) et U. erinaceus (4x).
Au niveau infraspecifique, on remarque des differences dans le deg.·e de complexite de Ia structure exinique :
a/ dans des especes ou il existe deux niveaux de ploldie reconnus comme deux taxons infraspecifiques, il y a parallelement deux differents types polliniques (e.g. U. argenteus subsp. argenteus, 2x/type I, et subsp. subsericeus, 6x/type III ; U. europaeus subsp. latebracteatus, 4x/type II, et subsp. europaeus, 6x/type III ; U. gallii subsp. breoganii, 4x/type II, et subsp. gallii, 6x/type III).
b/ dans des especes comme U. australis et St. boivinii, chez lesquellcs nous avons observe des populations qui presentent differents niveaux de plo"idie (4x et 6x), bien qu'elles n'aient pas ete reconnues du point de vue taxonomique, on trouve egalement des types polliniques differents correles avec le niveau de ploldie (type II et III respectivement).
c/ finalement, dans des especes comme U. parviflorus et St. gen~toides, ou un certain degre de diversification macro-morphologique a permis de reconnaitre les sous-especes (Tableau 1), on remarque qu'il n'est pas associe a aucun changement du niveau de ploldie ni de structure exinique.
Chez Genista, Ia correlation niveau de plo"idie/type structural exinique est a peu pres semblable a celle d'Ulex et Stauracanthus. Cette correlation s'ob-
serve de fa~ton identique si l'on part de nombres de base x = 9 et x"' 12 (Sa.iiudo, 1979) ou si l'on accepte le nombre de base x = 6 (Verlaque, 1992). Le type I est present dans tous les diploldes sauf G. mugronensis subsp. mugronensis. Le type II est present dans tous les tetraploldes a !'exception de G. hispanica subsp. occidentalis et aussi de G. mugronensis subsp. mugronensis (2x). Le type III est present dans l'unique hexaplo'ide etudie et egalement chez G. hispanica subsp. occidentalis (4x). Les deux especes octoplo'ides analysees pre-sentent le type IV. ·
Dans le groupe de taxons a nombres chromosomiques multiples de x = 9, on remarque !'absence du type I dans les taxons diplo"ides et l'homogeneite de Ia structure exinique (type II) dans les taxons infraspecifiques a niveaux de ploldie differents (G. mugronensis). Dans les taxons a nombres chromosomiques multiples de x"' 12, on observe d'une part }'absence de taxons iberiques a type structural III parallelement a celle de representants hexaploldes ; d'autre part, les differences du degre de complexite de Ia structure exinique dans les taxons dont les populations presentent differents niveaux de plo"idie (G. tinctoria, 4xltype II et 8x/ type IV).
II faut enfm souligner que chez Genista le taxon diplo'ide a n = 9 presente une structure de Ia paroi pollinique (Type II) plus elaboree que celle des taxons diplo'ides a n = 12 (Type 1). En outre, dans les taxons etudies d'origine aneuplo'idie (G. legionensis et G. umbellata), l'aneuplo"idie ne parait pas se traduire par une variation structurale de }'exine comparee a celle des taxons qui ont le meme niveau de plo"idie.
La tendance de complexite progressive de l'exine chez Ulex, Stauracanthus et Genista se manifeste de telle faliOn qu'une augmentation du nombre chromosomique apparait associee a des variations relatives de l'epaisseur des
Dow
nloa
ded
by [
Uni
vers
ity o
f A
berd
een]
at 2
2:47
05
Oct
ober
201
4

C. PARDO, P. CUBAS ET P. SANCHEZ TESTILLANO 199
differcntcs couches exiniqucs ct a une augmentation de la complexite de Ia couche infratectale. Cette tcndance se manifeste par le passage d'un infratcctum grenu peu organise a un infratcctum compose de grains et de columelles jusqu'a un infratectum columellaire constitue des columelles bien delimitees, lesquelles doivent etre originees par empilement de grains, comme il a ete observe dans d'autres genres et familles (Le Thomas, 1976; Guinet et Ferguson, 1989). Neanmoins, chez Genista, cette tendance se differencie dans les especes octoplo"ides par un type structural tres caracteristique (type IV) dont toutes les
couches exiniqucs sont constituees de grains. L'origine de ce nouveau type rest~ a etablir : il peut etre considere comme le point culminal de la tendance g~ncrale (passage de type III au type IV) ou bien comme une differenciation de cctte tendance avec le passage direct du type II au type IV (ex. G. tinctoria type Il/4x et type IV/8x).
Remerciements.- Nous remercions tout particuliilrerr.ent Mme A. Le Thomas, Directeur du Laboratoire de Phytomorphologie generale et comparee, Museum national d'Histoire naturelle de Paris, pour ses precieuses suggestions et pour Ia revision crnique du manuscrit.
BIBLIOG RA PIIIE
Bisby F.A., 1981.- Genisteae (Adans.) Benth. In: Advances in Legume Systematics. Polhill R.M. & P.H. Raven (Eds.). Kew, 1, 409-425.
Castro D., de, 1941.- Algumas contagens de cromoso· mas no genera Ulex L. (sensu lata). Agron. Lusit., 3,103-113.
Castroviejo S. & E. Valdes-Bermejo, 1983.- Netas sobra los tojos gallegos. Anales Jard. Bot. Madrid, 40 (1), 73-81.
Cristofolini G. & L. Feoll Chiapella, 1984.- Origin and diversification of Genisteae (Fabaceae) : a serosystematic purview. Webbia, 38, 105-122.
Cubas P., 1986.- Numeros cromosomaticos en Ulex L. y Stauracanthus Link ( Genisteae, Papilionaceae). Anales Jard. Bot. Madrid, 43 (2), 217-233.
Cubas P. & C. Pardo, 1992.· Pollen wall stratification trends in U/ex (Genisteae, Papilionoideae : Leguminosae) in the Iberian Peninsula. Grana, 31, 177-185.
Erdtman G., 1952.- Pollen morphology and plant taxonomy. 1 - Angiosperms. Almqvist & Wiksell, Stock· holm.
Feoli-Chiapella L. & G. Cristofolini, 1981.- Serological contribution to the systematics of U/ex L. (Genisteae-Fabaceae) and allied genera. Nord. J. Bot., 1, 723-729.
Ferguson I.K. & J.J. Skvarla, 1981.· The pollen mor· phology of subfamily Papilionoideae (Leguminosae). In: Advances in Legume Systematics. Polhill R.M. & P.H. Raven (Eds.). Kew, 2, 859-896.
Fernandes A., M.F. Santos & M. Queiros, 1977.- Cytotaxonomie des Spermatophyta du Portugal, IV, suppl. 2. Bol. Soc. Brat., ser. 2. 51, 137·186.
Fernandez Piqueras J. & M. Ruiz Rejon, 1976.· Estudios cariol6gicos sabre Ia flora espanola. Bol. Soc. Brat., ser. 2, 50, 5-13.
Gibbs P.E .. 1966.· A revision of the genus Genista L. Not. R. Bot. Gard. Edinb., 27, 11·99.
Guinet P. & I.K. Ferguson, 1989.· Structure, evolution
and biology of pollen In Leguminosae. In : Advances in Legume Biology. Stirton C.H. & J.L. Zarucchi. Mo. Bot. Gard. Press, StLouis, 77-103.
Le Thomas A. & B. Lugardon, 1976.· De Ia structure grenue ~ Ia structure columellaire dans le pollen des Annonacees. Adansonia, 2 ser., 15, 543-572.
Love A. & E. Kjellqvist, 1974.· Cytotaxonomy of Spanish plants. IV • Dycotiledons : CaesalpiniaceaeAsteraceae. Lagascalia, 4 (2), 153·211.
Missel M.T., J.P. Gourret & A. Huon, 1982.· Le pollen d'Uiex L. (Papilionoideae): morphologie des grains et structure de l'exine. Pollen et Spores. 24, 369-395.
Muller J., 1973.· Pollen morphology of Barringtonia calytrocalyx K. Sch. (Lecythydaceae). Grana, 13, 29-44.
Polhill R.M., 1976.· Genisteae (Adans.) Bentham and related tribes (Leguminosae). Bot. Syst., 1, 143-368.
Prados A., J.L. Ubera & E. Dominguez, 1985.- Estudio palinol6gico de Ia trlbu Genisteae R. Br. en Andalucla Occidental. An. Asoc. Palinol. Leng. Esp., 2, 35-42.
Ruiz Rejon M. & J. Fernandez Piqueras, 1978.· U/ex parvtflorus Pourret. No 72. Lagasca/ia. 8 (1), 120-121.
Santos A.C., 1944.· Algumas contagens de cromosomas nos generos Genista L., e Cytisus L. Bol. Soc. Brot., 19, 519-522.
Sai'ludo A., 1971.· Variabilidad cromos6mica en las Genisteas de Ia flora espanola en relacl6n con su ecologla. I - Numero y comportamiento de los cromosomas durante Ia meiosis. A • Secciones Erinacoides Spach, Spartioides (l.) DC. y Asterospartum Spach del Gen. Genista. Cuad. C. Bioi. Univer. Granada. 1 (1), 5·21.
Sai'ludo A. 1972.· Variabilidad cromos6mica en las Genisteas de Ia flora espanola en relaci6n con su ecologla. B • Secciones Genista L., Spartioides
Dow
nloa
ded
by [
Uni
vers
ity o
f A
berd
een]
at 2
2:47
05
Oct
ober
201
4

200 ACTA BOTANICA GALLICA
Spach, Phillospanum Willk. y Voglera (B. Gaertner, B. Mayer and Seherb) Spach del Gen. Gemsta. Cuad. C. Bioi. Univ. Granada, 1 (2), 43-52.
Sa~udo A., 1973.- Variabilidad cromos6mica de las Genisteas de Ia flora espa~ola en relaci6n con su ecologla. 1 - Numero y comportamiento de los cromosomas durante Ia meiosis. C. Secci6n Cephalospanum del Gen. Genista L. y generos Lygos Adanson, Spanium L., Teline Medicus, Calycotome Link y Argyrolobium Ecklon & Zeyher. Cuad. C. Bioi. Granada. 2 (2), 11 7-120.
Sa~udo A., 1979.- Chromosome variability in the Ge-
nisteae (Adans.) Benth. Webbia, 34, 363-408. Valdes BermeJO E., 1979.- Numeros cromos6micos de
plantas occidentales. 1 - 34. Anales Jard. Bot. Madrid, 36, 373-389.
Verlaque R .. 1992.- Modalites de Ia speciation chez Iss Genisteae. Ades del Simposi Int. de Botanica Pius Font i Ouer, L:eida, 2, 49-68.
Vicioso C., 1953.- Genisteas espa~olas. 1 - GenistaGenistella. Bot. lnst. For. /nv. Exper. Madrid, 67, 1-153.
Vicioso C., 1962.- Revisi6n del genera Ulex en Espana. Bot. lnst. For. lnv. Exper. Madnd, 80, 1-57.
LEGENDES DU TABLEAU ET DES PLANSi1ES
Tableau 1.- Sommaire des correlations entre les types structuraux de Ia paroi pollinique et Iss niveaux de plo'idie chez Ulex, Stauracanthus et Genista. Les taxons aneuploides sont marques d'un asterisque. Nombres de base actuels d'Uiex et Stauracanthus, selon Cubas (1986), et de Genista selon Sa~udo (1979).
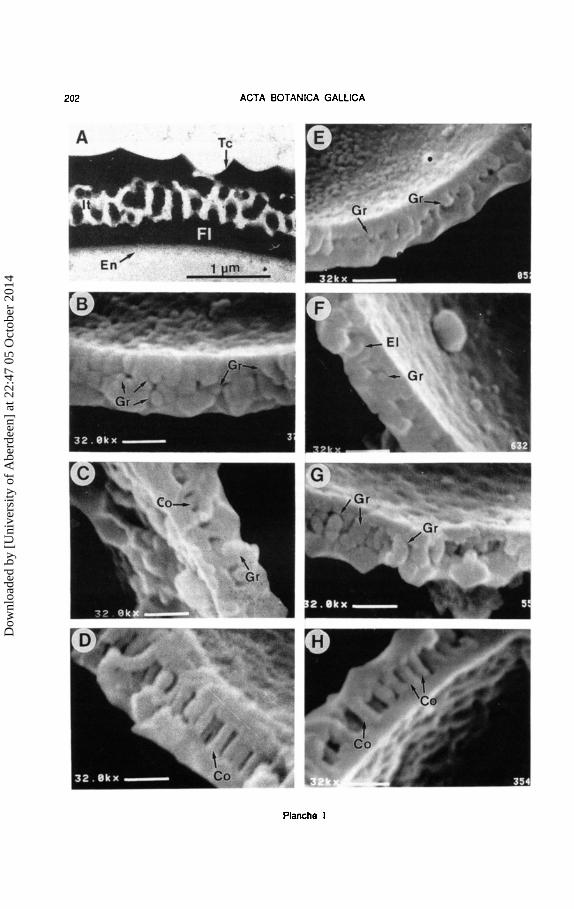
Planche 1.- Structure de Ia paroi exinique chez Ulex et Stauracanthus. A . U. erinaceus, MAF 106562, coupe au MeT montrant le tectum (Tc), l'infratectum (It). Ia foot layer (Ft) et l'endexine (En) mince ; B : U. parviflorus subsp. rivasgodayanus, type I, MAF 108601, grains part ant du tectum et de Ia nexine ; C : U. australis, tetraplo'ide, type II, MAF 68460, columelles courtes et quelques grains isoles ; D : U. australis, hexaploide, type Ill, MAF 106565, columelles longues et nettes ; E : St. genistoides subsp. genistoides, type I, MAF 138024, grains part ant du tectum et de Ia nexine ; F : St. genistoides subsp. spectabilis, type I, MAF 106632, grains isoles et elements radiaux incomplets; G: St. boivinii, tetraploioe, type II, MAF 138023, grains en a mas et empiles ; H: St. boivinii, hexaploide. type Ill, MAF 106621, co1umelles nettes et d'autres constituees par grains empiles et orientes radialement. Co . columella ; El : element radial incomplet; Gr: grain. Echelle = 0.63 ~m saul A.
Planche 11.- Structure de Ia paroi pollinique chez Genista. A: G. florida, MAF 138123, coupe au MeT montrant le tectum (Tc), l'infratectum columellaire (It). Ia foot layer (FI) et l'endexine mince (En) ; B: G. fa/cata. type II. MAF 126508, infratectum a columelles courtes melangees avec gra1ns en amas; C: G. pseudopilosa, type Ill, MA 348632, columelles nettes ; D : G. tridens, type IV, MAF 107263, notez que toutes Iss couches sont constituees de grains ; E : G. cinerascens, type I, MAF 101494, infratectum a grains partant de Ia nexine et isoles ; F : G. legionensis, type II, MA 261867, infratectum a grains empiles et isoles ; G : G. tinctoria, tetraploide, type II, MA 425983, grains melanges avec des columelles pas tres nettes ; H: G. tinctoria, octoploide, type IV, MAF 69164, petits grains fortement empiles dans toutes les couches. Co: columella ; Gr: grain. Echelle = 0.63 ~m saul A.
CAPTIONS OF THE TABLE AND THE FIGURES
Table 1.- Summary of the correlation between structure types of the poll8n wall and ploidy levels in Ulex, Stauracanthus and Genista. Aneuploid taxa are indicated by an asterisk. Base number of Ulex and Stauracanthus, after Cubas (1986), and Genista after Sa~udo (1979).
Plate 1.- Exine structure of pollen grains in U/ex and Stauracanthus. A: U. erinaceus, MAF 106562, TEM section showing the tectum (Tc). the infratectum (It), the foot layer (Ft) and the thin endexine (En) ; B: U. parviflorus subsp. rivasgodayanus, type I, MAF 108601, granules risen from both the tectum and the nexine; C: U. australis, tetraploid, type II, MAF 68460, short columellae and isolated granules ; D : U. australis, hexaploid, type Ill, MAF 106565, long and well-developed columellae ; E : St. genistoides subsp. genistoides, type I, MAF 138024, granules risen from the tectum and the nexine; F: St. genistoides subsp. spectabilis, type I, MAF 106632, isolated granules and incomplete processes radially oriented ; G : St. boivinii, tetraploid, type II, MAF 138023, grouped and piled granules; H: St. boivinii, hexaploid, type Ill, MAF 106621, well-developed columellae composed by granules arranged in radially oriented rows. Co :columella ; El : radial process ; Gr : granule. Scale = 0.63 ~m except in A.
Plate 11.- Exine structure of pollen grains in Genista. A: G. florida, MAF 138123. TEM section showing the tectum (Tc), the columellar infratectum (It), the foot layer (FI) and the thin endexine (En) ; B : G. falcata, type II, MAF 126508, infratectum with short columellae and grouped granules; C: G. pseudopilosa, type Ill, MA 348632, well-developed columellae ; D : G. tridens, type IV, MAF 1 07263, note that all the layers are composed by granules; E :G. cinerascens. type I, MAF 101494, infratectum with granules in the nexine and isolated; F: G. legionensis, type II, MA 261867, infratectum with isolated and piled granules ; G : G. tinctoria, tetraploid, type II, MA 425983, granules interspersed with ill-developed columellae; H: G. tinctoria, octoploid, type IV, MAF 69164, note that all the layers are composed by small granules densely packed. Co :columella ; Gr. :granule. Scale = 0.63 ~m except in A.
Dow
nloa
ded
by [
Uni
vers
ity o
f A
berd
een]
at 2
2:47
05
Oct
ober
201
4
2x
u x=l6
I e X 4x
6x
5 2x 1 u x=24 r a c 4x a ~ 6x h.
2x
x= 9 4x G e n 6x i s Sx t a
2x
x=l2 4x
Sx
C. PARDO, P. CUBAS ET P. SANCHEZ TESTILLANO
Tableau 1
Type I Type II Type III
argenteus canescens ..a.. . .,. ....... parvi florus oubop. porvillo-..a... riv•lodafODoa
baeticus ..a...-.. micranthus minor
borgiae erinaceus densus australis eriocladus europaeus ..a...-gallii ..a... breo1111ii
argenteus ..a... ouboericcuo
australis jussiaei europaeus ....... _ gallii ouloop. lallii
genistoides ............... id .. ..a... ope<tabilio
boivinii
boivinii
mugronensis ouloop.--•
mugrooensis hispanica ......... idiooilllo oubop.occidealalio
falcata
pseudopilosa
cinerascens hystrix pilosa
cinerea ....... opec;.. legionensis • dorycniifolia umbellata • tine tori a
201
Type IV
tridens
tinctoria
Dow
nloa
ded
by [
Uni
vers
ity o
f A
berd
een]
at 2
2:47
05
Oct
ober
201
4
202 ACTA BOTANICA GALLICA
Planche I
Dow
nloa
ded
by [
Uni
vers
ity o
f A
berd
een]
at 2
2:47
05
Oct
ober
201
4
C. PARDO, P. CUBAS ET P. SANCHEZ TESTILLANO 203
Planche II
Dow
nloa
ded
by [
Uni
vers
ity o
f A
berd
een]
at 2
2:47
05
Oct
ober
201
4

204 ACTA BOTANICA GALLICA
ANNEXE: MATERIEL ETUDIE
ULEXL U. argenteus Webb subsp. argenteus : Portugal, Al
garve, Vila do Bispo, 10.3.1978, Andres, BogOnez & Cubas, MAF 106551, (2) 2n = 32 (Cubas, 1986).
U. argenteus Webb subsp. subsericeus (Coutinho) Rothm. : Portugal, Algarve, Faro, cerca del aeropuerto, 26.5.1979, Cubas et a/., MAF 106559, (1) 2n = 96 (Cubas, 1986).
U. australis Clemente : Espana. Cadiz, Chiclana, Pin ares de Ia Dehesilla, 22.9. 1965, Bo~a. MAF 68460 ( 1) 2n = 64, (Cubas, 1986) ; Espana. Hue tva, Donana, Matagorda, 18.3.1979, Guerrero eta/., MAF 106565 (1) 2n = 96, (Valdes-Bermejo, 1979; Cubas, 1986).
U. baeticus Boiss. subsp. baeticus : Espana, MAlaga, Puerto de las Abejas, de Yunquera a El Burgo, 7.4.1979, Andres eta/., MAF 106610, (2) 2n = 32 (Cubas, 1986).
U. borgiae Rivas Martinez : Esparla, Cadiz, Puerto del Cabrito, de Tarifa a Algeciras, 6.4.1977, Moreno et a/., MAF 108581, (2) 2n = 64 (Ruiz RejOn & FernAndez Piqueras, 1978 ; Cubas, 1986).
U. canescens Lange : Espana, Almeria, Cabo de Gata, 27.1.1980, Andres & Cubas, MAF 106689, (1) 2n .. 32 (FernAndez Piqueras & Ruiz RejOn, 1976).
U. densus Webb. : Portugal, Estremadura, rio de Mauro, 22.4.1978, Prada et al., MAF 106577, (2) 2n = 64 (Castro, 1941 ; Fernandes et a/., 1977 ; Cubas, 1986).
U. erinaceus Webb. : Portugal, Algarve, de Vila do Bispo al Cabo, 27.3.1980, CantO & Cubas, MAF 106562, (1) 2n = 64 (Cubas, 1986).
U. eriocladus C. Vicioso : Espana. Badajoz, BarcarrotaJerez de los Caballeros, 4. 11.1977, Prada & Cubas, MAF 106570, (1) 2n = 64 (Cubas, 1986).
U. europaeus L. subsp. europaeus : Espana, Navarra, Azpilicueta, 16.4.1972, Fuertes & Ladera, MAF 134351, (3) 2n = 96.
U. europaeus L. subsp. latebracteatus (Mariz) Rothm. : Espana, Pontevedra, Bayona, 10.9.1977, Cubas, MAF 134350, (1) 2n = 64 (Cubas, 1986).
U. gallii Planchon subsp. gallii: Espana, Vizcaya, Cabo de Machichaco, 27.9.1977, CantO & Cubas, MAF 106646, (1) 2n = 96 (Cubas, 1986).
U. gallii Planchon subsp. breoganii (Castroviejo & Valdes) Rivas Martinez et a/. : Espana. Pontevedra, Carril, 11.9.1977, Cubas, MAF 134353, (2) 2n = 64 (Castroviejo & Valdes-Berrnejo, 1983).
U. jussiaei Webb : Portugal, Beira l~oral, de Coimbra a Ceira, pasado el desvlo a Penacova, 25.4.1978, Prada et aL, MAF 106656, (1) 2n • 96 (Cubas, 1986).
U. micranthus Lange : Portugal, Beira literal, Vendas do Porco, 25.4. 1978, Cubas eta/., MAF 1 06579, (2) 2n = 32 (Cubas, 1986).
U. minor Roth : Portugal, Algarve, Odeceixe a Rojil, 24.5.1979, Cubas, MAF 134354, (2) 2n .. 32 (Castro, 1941 ; Cubas, 1986).
U. parviflorus Pourret subsp. parviflorus : Espana. Jaen, Huelma, 26.1.1980, Andres & Cubas, MAF 106549,
(2) 2n = 32 (Love & Kjellqvist, 1974). U. parviflorus Pourret subsp. rivasgodayanus Cubas :
Espana. MAlaga. Sierra de Almijara, subida al Puerto del Collado a COmpeta, 26.6.1978, Moreno et al .. MAF 108601, (1) 2n = 32 (Cubas, 1986).
STAURACANTHUS LINK St. boivinii (Webb) Samp. : Espana, CAdiz, Los Bar
rios, 7.5.1980, Cubas et al., MAF 106620, (1) 2n = 96 (Cubas, 1986); Espana, MAlaga, P1o. Galis a Jimena de Ia Frontera, Cubas et al., MAF 138023, (1) 2n = 96 (Cubas, 1986) ; Portugal : Algarve, San Tectonic hacia Odeceixe, 24.5.1979, Cubas eta/., LAF 106621, (1) 2n = 144 (Cubas, 1986).
St. genistoides (Brat.) subsp. genistoides: Espana, Huelva, Donana, La Gravija, 18.3.1979, Cubas et a/., MAF 138024, (1) 2n = 48 (Cubas, 1986); Portugal, Estremadura, Peniche, Cabo Carvoeiro, 13.4.1978, Pardo et a/., MAF 138025, (2) 2n = 48 (Cubas, 1986).
St. genistoides subsp. spectabilis (Webb) Rothm. : Portugal, Baixo Alentejo, Sines, 4.5.1979, Cubas eta/., MAF 106632, (1) 2n = 48 (Cubas, 1986); Portugal, Algarve, Sag res, 27 .3.1980, Cubas et a/., MAF 106691, (1) 2n .. 48 (Cubas, 1986).
GENISTA L G. cinerascens Lange : Espana. CAceres, Toril, Vega
del Chiquero, 4.5.1980, D. Belmonte MAF 117011, (3) 2n = 24 ; Espana. Avila, Sierra de MalagOn, Puerto de las Piletas, 12.6.1978, Rivas Goday, MAF 101494, (2) 2n = 24 (Sanudo, 1979).
G. cinerea (Viii.) DC. subsp. speciosa Rivas Martinez et a/. : Espana, Jaen, Monte Javalcruz, 5.4.1977, BolOs eta/., MAF 90978, (2) 2n = 48 (Sanudo, 1972).
G. dorycniifolia Font Quer : Espana, Baleares, lbiza, cerca de Portinatx, 4.5.1980, Rivas Martinez eta/., MAF 422218, (2) 2n = 48 (Santos, 1944).
G. falcata Brat. : Espana, Luge, el Caurel, Ferramulin, 26.5.1981, lzco et a/., MAF 126508, (3) 2n = 36.
G. florida L. : Espana. CAceres, Garganta Ia Olla a Jaraiz de Ia Vera, 16.5.1992, SAnchez Mata, MAF 138123, (3) 2n = 48.
G. hispanica L. subsp. occidentalis Rouy : Espana. Asturias, de Aviles a Grado, 28.3.1970, Rivas Goday & Demetrio, MAF 83447, (3) 2n .. 36.
G. hystrix Lange : Espana, LeOn, Manzanal del Puerto, , Puerto de Manzanal, 20.6.1980, FernAndez Casas, MAF 116062, (3) 2n = 24.
G. legionensis (Pau) Lalnz : Espana, Santander, Pates, Valle de Ia Hermida, 20.5.1970, Sanudo, MA 261867, (1) 2n = 40 (Sanudo, 1972).
G. mugronensis (Viehr.) subsp. mugronensis: Espana. Valencia, de Almansa a Requena, Sanudo, MA 261918, (1) 2n =18 (Sanudo, 1971).
G. mugronensis subsp. rigidissima (Viehr.) FernAndez Casas : Espana. Soria, Arbujuelo, 4.6.1977, Segura, MA 261917, (2) 2n = 36 (Sanudo, 1971 ).
Dow
nloa
ded
by [
Uni
vers
ity o
f A
berd
een]
at 2
2:47
05
Oct
ober
201
4

C. PARDO, P. CUBAS ET P. SANCHEZ TESTillANO 205
G. pilosa l. : Espalla, Guadalajara, Puerto de Ia Quesera, 11.6.1979, Rivas Martinez, MAF 108642, (3) 2n = 24.
G. pseudopilosa Cosson : Espal\a, Albacete, Sierra de Alcaraz. las Espineras del le6n, 28.5.1976, Fernandez Casas, MA 348632, (3) 2n = 72.
G. tinctorial. :Espana, Castell6n, Almenara, 2.6.1984, M. Blasco. MA 425983, (3) 2n = 48. Espai'la, Madrid, Puerto de los leones de Castilla, 6.7.1967, Borja et a/., MAF 69164 (sub. var. e/ata), (3)
2n = 96 ; Espal\a, Santander, Alonos de Carriedo, 20. 7.1969, sanudo, MA 293295 (sub. var. e/ata), (1) 2n = 96 (Sanudo, 1972).
G. tridens (Cav.) DC : Espana, Cadiz, Chiclana, entre Chiclana y Co nil, 17.6.1980, Navarro & Cant6, MAF 107263, (2) 2n" 72 (Sai\udo, 1972).
G. umbellata (l'Herbier) Poiret in lam. : Espal\a, Granada, lobre, 6.4.1985, Molina, MAF 124779, (2) 2n = 42 (Sai\udo, 1973).
Dow
nloa
ded
by [
Uni
vers
ity o
f A
berd
een]
at 2
2:47
05
Oct
ober
201
4