Embed Size (px)
Citation preview

Thèse de Doctorat
UNIVERSITÉ PARIS 12 VAL DE MARNE
Spécialité : Sciences et Techniques Environnementales
Présentée par
Fabien THOMAS
Pour obtenir le grade de Docteur de l’Université Paris 12 Val de Marne
Rôle de deux ingénieurs de l’écosystème: le termite Cornitermes sp. et l’annélide Andiodrilus pachoensis sur le fonctionnement du sol dans le
Sud-est amazonien.
Soutenue le 19 Décembre 2006Devant le jury composé de :
• Pr Decaëns Thibaud Rapporteur• DR De Noni Georges Rapporteur• Pr Bruand Ary Examinateur• Pr Lavelle Patrick Examinateur • Pr Garnier-Zarli Evelyne Directrice de thèse
Thèse de DoctoratUNIVERSITÉ PARIS 12 VAL DE MARNE
Spécialité : Sciences et Techniques Environnementales
Présentée par
Fabien THOMAS
Pour obtenir le grade de Docteur de l’Université Paris 12 Val de Marne
Rôle de deux ingénieurs de l’écosystème: le termite Cornitermes sp. et l’annélide Andiodrilus pachoensis sur le fonctionnement du sol dans le
Sud-est amazonien.
Soutenue le 19 Décembre 2006Devant le jury composé de :
• Pr Decaëns Thibaud Rapporteur• DR De Noni Georges Rapporteur• Pr Bruand Ary Examinateur• Pr Lavelle Patrick Examinateur • Pr Garnier-Zarli Evelyne Directrice de thèse

Thèse de Doctorat
UNIVERSITÉ PARIS 12 VAL DE MARNE
Spécialité : Sciences et Techniques Environnementales
Présentée par
Fabien THOMAS
Pour obtenir le grade de Docteur de l’Université Paris 12 Val de Marne
Rôle de deux ingénieurs de l’écosystème: le termite
Cornitermes sp. et l’annélide Andiodrilus pachoensis sur
le fonctionnement du sol dans le Sud-est amazonien.
Soutenue le 19 Décembre 2006
Devant le jury composé de :
• Pr Decaëns Thibaud Rapporteur
• DR De Noni Georges Rapporteur
• Pr Bruand Ary Examinateur
• Pr Lavelle Patrick Examinateur
• Pr Garnier-Zarli Evelyne Directrice de thèse

Remerciements
Remerciements
Ce travail a été réalisé au sein du Laboratoire de Biologie des sols et des eaux
(Université Paris 12) et du département des sciences du sol de l’université fédérale rurale
d’Amazonie (UFRA) au Brésil. Ce travail a été possible grâce d’une part à l’Université Paris
12 Val de Marne qui m’a attribué une bourse de thèse, et d’autre part à l’IRD qui m’a
également aidé financièrement.
Je tiens tout d’abord à remercier Monsieur Paulo Fernando da Silva Martins, directeur
du département des sciences du sol de l’UFRA (Belém) qui m’a chaleureusement accueilli et
qui a mis à ma disposition, pendant toute la durée de mon séjour au Brésil, tous les moyens
dont il disposait pour assurer le bon déroulement de mon travail. Merci également au personnel
du Museum Paraense Emilio Goeldi (Belém) et du Laboratoire socio-agronomique du
Tocantins (LASAT) qui m’a accueilli à Marabá.
Je suis particulièrement sensible à l’honneur que m’ont fait Messieurs Georges De Noni
et M. Thibaud Decaëns pour avoir accepté d’être les rapporteurs de cette thèse. Je suis
également très reconnaissant à Messieurs A. Bruand et P. Lavelle pour avoir bien voulu faire
partie du jury d’examen de la thèse. Je les en remercie sincèrement.
Mme Evelyne Garnier-Zarli, directrice du laboratoire de Biologie des Sols et des Eaux,
était le professeur responsable de ma thèse. Elle a montré beaucoup d’intérêt pour mes
recherches et m’a apporté ses conseils et ses critiques bienveillantes aux moments décisifs de mon
travail. Je tiens à lui exprimer ici ma profonde gratitude.
Ma reconnaissance va aussi à Mr Michel Grimaldi pour la confiance qu’il m’a
accordée et pour sa patience à mon égard durant toutes les étapes de ma thèse. Il a toujours été
disponible pour discuter et partager avec moi sa grande expérience des sols. J’ai beaucoup
appris à son contact.

Remerciements
Je voudrais en particulier remercier Max Sarrazin, technicien de l‘IRD détaché à
l’UFRA, dont la disponibilité, la compétence et l’efficacité m’a été d’une aide précieuse, tant sur
le terrain qu’au laboratoire et qui m’a gentiment accueilli chez lui à Belém.
Nombreuses sont les personnes qui m’ont apporté leur concours et ont contribué à la
réalisation de ce travail :
- Didier Brunet , pour les études NIRS de la matière organique
- Yannick Besnard pour la réalisation des lames minces des structures biogéniques et
Vincent Hallaire pour son aide dans l’étude micromorphologique de ces lames.
-Jean-Pierre Rossi et Jérome Mathieu pour leur contribution dans le traitement
statistique des données.
- Philippe Mora, Thierry Desjardins, Sébastien Barot, Nourreddine Bousshérine, Lise
Dupont, Myriam Harry, Samir Abbad , Corinne Rouland, pour leurs conseils et
encouragements.
A tous, je leur dis merci.
Aux techniciens, Alexandre Livet, Catherine Martin (LBSE), Edouard Miambi,
Annabelle Mellot, Jocelyne Roman(LEST), et à tous mes camarades thésards et ater :
Catherine, Jennifer, Marieke, Manuel, Virginie, avec qui j’ai partagé des « tranches de vie ».
Qu’ils soient tous remerciés.
Je n’oublie pas les agriculteurs de Benfica qui ont accepté que je travaille sur leurs
terres : Sr Melete, Sr Osano, Sr Josué, Sr Cicéro, Sr Léonine…Un grand merci à eux.
Je ne saurais terminer ces quelques mots de remerciement sans parler de mes proches :
mon fidèle ami Déurival, qui m’a beaucoup aidé sur le terrain mais aussi tant apporté
humainement. C’est « um amigo pra todas horas » mes parents pour leur soutien et mon épouse
Neidelane, qui tous les jours est une source de bonheur et de réconfort.

Résumé étendu
Résumé étendu
La perte de biodiversité dans les écosystèmes anthropisés et en particulier dans
les sols, suscite ces dernières années un intérêt croissant. Des mesures, afin de garantir
la conservation de cette diversité biologique, sont maintenant préconisées lors de
grandes conventions internationales.
En Amazonie brésilienne, dans la région de Marabá, la mutation du milieu de la forêt
en pâturage par l’agriculture familiale modifie profondément les propriétés physiques,
chimiques et biologiques des ferralsols argileux.
Les organismes du sol et principalement les invertébrés « ingénieurs » (annélides,
termites et fourmis) jouent un rôle essentiel dans l’évolution et la structuration du sol.
Les conséquences de la déforestation se manifestent par une modification des
populations des organismes du sol. Face au bouleversement du milieu, des espèces
disparaissent ou s’adaptent en modifiant leur activité alors que d’autres colonisent les
nouveaux milieux ouverts. Les organismes « ingénieurs » produisent des structures
biogéniques (déjections d’invertébrés, turricules d’annélides, constructions termitiques,
galeries,…) qui affectent fortement les processus physico-chimiques du sol.
L’objectif de cette étude est de mettre en évidence le rôle particulier de deux
espèces ingénieurs : l’annélide Andiodrilus pachoensis et le termite Cornitermes sp. dans
le fonctionnement du sol et d’établir plus généralement une typologie des structures
biogéniques produites par ces organismes, utilisée comme base à la constitution d’un
indice de qualité des sols, afin de contribuer à une modélisation des sols à but prédictif
voire préventif.
La caractérisation de profils pédologiques (jusqu’à 2 m de profondeur) dans le site
d’étude nous a permis de mettre en évidence que le sol des parcelles choisies est un
ferralsol, encore appelé latosol, un peu moins épais sous pâturage que sous forêt.
Dans les pâturages, l’horizon superficiel (0-10 cm) est légèrement tassé en raison de
la structure localement lamellaire entre les touffes de graminée, et hétérogène à cause
de la distribution en taches de la matière organique et de l’hydromorphie. Sous forêt, un
horizon organique humifère relativement homogène occupe les premiers centimètres du
sol.
La texture des horizons de surface est dans toutes les parcelles argilo-sableuse.
L’altérite apparaît généralement vers 1,5 m de profondeur sous un lit de nodules.
La transformation de la forêt en pâturage par l’agriculture familiale provoque dans le
site d’étude, comme dans de nombreux contextes amazoniens, une évolution des

Résumé étendu
propriétés physiques et chimiques du sol. Il a été observé que la fertilité chimique tend
vers une amélioration mais qu’un tassement du sol s’opère.
Une première étude a été consacrée au termite Cornitermes sp. Les termitières
épigées, de grande taille (jusqu’à 1,5 m de hauteur) dans les pâturages, atteignent une
densité de 38 nids à l’hectare alors qu’aucun de ces édifices n’a été observé sous forêt.
La distribution spatiale des termitières, analysée dans trois pâturages, s’est avérée
hétérogène mais significativement reliée à la profondeur du sol.
Les propriétés physico-chimiques de ces structures ont permis de mesurer leur
stabilité structurale et leur résistance au brûlis annuel des pâturages. La stabilité
structurale de la muraille de termitière déjà très élevée par rapport au sol, s’accroît
lorsque la structure biogénique est brûlée. Le nid est composé de deux parties
distinctes : la paroi périphérique et le feuilleté central. Seul ce dernier constitue une
réserve importante en matière organique (77 %).
En se référant à la granulométrie (54% d’argile et 32% de sables grossiers), au pH
(pH=4) et aux concentrations d’ions échangeables présents dans la muraille ([Al3+] =
0,53 cmolc.kg-1, [Ca2+] = 1,5 molc.kg-1, [Mg2+] = 1,1 cmolc.kg-1, [K+] = 0,3 cmolc.kg-1 ),
il est possible d’estimer la profondeur de prélèvement des matériaux par la caste des
ouvriers termites pour construire la muraille de leur nid entre 10 et 30 cm sous la surface
du sol. Sa concentration en carbone (19 mg.g-1) est du même ordre que celle du sol
témoin jusqu’ à 10 cm de profondeur.
La destruction de ces structures réalisée par les paysans amazoniens suscite ainsi un
effet local immédiatement positif, puis négatif en assurant la destruction de la réserve de
matière organique et une augmentation de stabilité de la muraille. La concentration en
carbone dans la muraille brûlée par rapport à l’état initial est réduit de 23% et le feuilleté
est presque totalement consommé par le feu. Le pH de la muraille augmente (de 4,6 à
5,4) et les concentrations en cations sont plus faibles dans la muraille brûlée qu’à l’état
natif. Seule la concentration en phosphore augmente de l’ordre de 4 fois. Les effets du
feu sur les termitières sont donc de deux ordres. Si l’augmentation du pH et de la
concentration en phosphore disponible dans la muraille tend à favoriser le
développement des végétaux et l’activité microbiologique, le changement de structure et
de constitution (formation d’oxydes de fers) et la diminution des concentrations en
cations et en carbone ont un effet négatif sur la fertilité du matériau.
La transformation du feuilleté en cendres devra être étudiée plus précisément pour
compléter le bilan chimique de ces termitières.

Résumé étendu
Il serait intéressant de préciser la progression de cette espèce dans les pâturages
amazoniens par une étude de sa dispersion à l’aide des méthodes de génétique des
populations.
Une deuxième étude a porté sur l’annélide Andiodrilus pachoensis. Une analyse
comparative des distributions spatiales des turricules de cette espèce a été menée dans 4
pâturages et 4 forêts. Les relations entre la distribution des turricules et les patterns
spatiaux des touffes de la graminée Brachiaria brisantha dans les pâturages ont été
analysées. Les liens entre la distribution des turricules et la quantité de litière sous forêt
ont été étudiés. Dans les deux milieux, tous les turricules et les éléments de la
végétation ont été cartographiés dans des surfaces variables de 25m² à 50m². La
production de turricules a été suivie pendant 2 mois. La densité de turricules s’est avérée
significativement plus faible dans les pâturages (42.103/ha) contre 243.103/ha dans les
forêts, et leur distribution est agrégée aux larges échelles (>15m) ; alors qu’en forêt, les
analyses spatiales ont mis en évidence une grande hétérogénéité entre les parcelles :
distribution aléatoire, agrégée ou inhibée. La densité des turricules de vers ne s’est pas
révélée significativement corrélée à la densité des touffes et leur production est
indépendante de la présence de touffes ou de la quantité de litière dans les forêts.
Dans les forêts et pâturages, les turricules sont composés en moyenne de 94%
d’éléments minéraux et 6% de matière organique. Comme les sols témoins, la
composition granulométrique des turricules est constituée essentiellement d’argile
(environ 45%) et de sables grossiers (environ 30 %).
La densité de motte des turricules est globalement plus faible que celle des sols
témoins. Il s’agit donc de turricules moins denses que le sol témoin et riches en matière
organique. Les turricules d’Andiodrilus pachoensis constituent une grande réserve en
carbone. Sa concentration dans les turricules est en moyenne de 60 mg.g-1 dans les
pâturages, légèrement plus élevée que dans les forêts (53 mg.g-1), significativement plus
forte que dans les sols témoins (0-30 cm). Les turricules dans les deux milieux
présentent un pH acide (4,6 et 5,8 respectivement pour les forêts et pâturages et 4 pour
les sols témoins) et les concentrations en cations sont plus élevées que dans le sol
témoin : sous forêt, [Al3+] = 0,72 cmolc.kg-1, [Ca2+] = 4,7 molc.kg-1, [Mg2+] = 2,2
cmolc.kg-1, [K+] = 0,27 cmolc.kg-1 ; sous pâturage, [Al3+] = 5,1 cmolc.kg-1, [ca2+] = 7,3
molc.kg-1, [Mg2+] = 3,6 cmolc.kg-1, [K+] = 0,75 cmolc.kg-1. Les concentrations en cations
dans les turricules de pâturages sont ainsi plus élevées que dans ceux de forêts.
La distribution en taille des particules, la stabilité structurale et la résistance à
l’érosion sous l’action de la pluie ont été mesurées sur des turricules natifs et des
turricules brûlés selon 2 intensités de pluie. Les turricules sont plus stables que le sol

Résumé étendu
témoin et aucun effet physique (stabilité) et chimique (concentrations en cations) du feu
sur cette structure biogénique n’a été mis en évidence.
Une troisième étude a mesuré les caractéristiques chimiques, obtenues par la
méthode NIRS (Near InfraRed Spectrophotometry), de la matière organique des
biostructures et permis d’établir une typologie d’un ensemble de structures biogéniques
(Turricules d’Andiodrilus pachoensis et d’autres annélides ainsi que les parois de
termitière de Cornitermes et d’autres termitières humivores, déblais et nids de fourmis,
déjections de coléoptères..).
L’état de la matière organique (fraîche ou dégradée) s’est avérée être le premier
facteur qui sépare les différentes structures biogéniques étudiées. L’origine et la nature
de la matière organique ne jouent pas un rôle significatif dans la différenciation des
structures biogéniques.
Deux autres typologies ont été établies à partir des concentrations ioniques et l’étude
de la porosité après analyse micromorphologique de 65 lames minces de biostructures à
deux résolutions (5,88µm et 0,47µm). Les typologies, obtenues de l’analyse des poroïdes
répartis en 3 classes de forme et 5 classes de taille, sont différentes aux deux
résolutions.
Les différences entre les typologies obtenues par les diverses méthodes soulignent
l’importance de considérer un maximum de caractéristiques physico-chimiques afin de
développer des outils méthodologiques visant à séparer les agrégats du sol soumis à la
forte variabilité locale du milieu. Les résultats des typologies n’ont pas mis en évidence
de fortes différences entre les grands genres taxonomiques mais une forte variabilité en
leur sein (variabilité inter espèces).
La modélisation de la dynamique des agrégats biologiques et plus largement des
profils culturaux d’un sol pose ainsi le problème d’une modélisation générale au niveau
taxonomique de l’ordre voire de la famille. L’accumulation de données spécifiques aux
espèces dites « ingénieurs ou clés de voûte » des écosystèmes afin de définir des
groupes fonctionnels, apparaît ainsi comme une étape nécessaire à la modélisation du
fonctionnement du sol.
Mots clés : Andiodrilus pachoensis / Cornitermes sp. / Turricule / Termitière / feuilleté /
xylophage / distribution spatiale / propriétés physiques / cations / stabilité structurale /
Brachiaria brisantha / NIRS / Lames minces / culture sur brûlis / simulation de pluie /
Amazonie

Resumo
Resumo
A privação da biodiversidade dentro dos ecosistemas antropicos, e em particular
no solo, estimula, os ultimos anos, um interesse crescente. Medidas, pra garantir a
conservação biologica, estão agora preconizadas durante grandas convençoes
internacionais. Na amazonia brasileira, no municipio de Marabà, o desmatamento
causado por a agricultura familiar, muda profundamente as propriedades fisicas,
quimicas e biologicas dos latosolos argilosos. Os organismos do sol, e os invertebrados
“engenheiros” (annelidos, cupins é formigas), jogam um papel essencial na evolução e na
estruturacão do solo. As consequências do desmatamento se manifestam por uma
modificação das populações dos organismos do solo. Na frente deste transtormo do meio
ambiente, as espécies desaparecem ou se adaptam, modificando a atividade delas
enquanto outras especies colonizam as novas pastagems. Os organismos “engenheiros”
produzem estructuras biogenicas (dejeçoes dos invertebrados, turriculos dos annelidos,
construçoes dos cupins, galerias...) que afetam os processos fisicos é quimicos do solo.
O objectivo desse estudo tem por objectivo de revelar o papel particular dos dois
espécies engenheiros : o annelido Andiodrilus pachoensis e o cupim Cornitermes sp.
affinis ovatus. no funcionamento do solo e de construir uma tipologia generica das
estruturas biogenicas usada como fundamento na constitução de um indicio de qualidade
do solo, para contribuir na modelisação predicativa do solo.
A caracterisação dos profilos pedologicas (até 2m de profondidade) na area do
estudo nos permiti de evidenciar que o solo das parcelas escolhidas é um ferralsol,
chamado também de latosolo, um poco menos fundo no pasto que na mata.
No pasto, a camada superficial (0-10 cm) esta um pouco comprimido por causa da
estrutura lameliforma entre os pés de Brachiaria brisantha é heterogenea por causa da
distribução heterogena de matéria organica é da hydromorphia. Na mata, a camada
organica homogenia enche os pimeiros centimetros do solo.
A textura das camadas da superfice do solo esta em todas parcelas argila-areiada.
A alterita aparece perto de 1,5 m de profondidade abaixo de uma camada de nodulos
O desmatamento causado por a agricultura familiar desafia na aréa do estudo, como
nos diversos contextos amazonicos , uma evolução das propriedades fisicas é quimicas
do solo. Si a fertilitade quimica consigue melhorar, um compactação do solo aparece.

Resumo
Um primeiro estudo foi realizado com os Cornitermes sp. Os cupinzeiros, de grande
tamanho: até 1.5 m de altura nos pastos, alcançam uma densidade de 38 ninhos por
hectares visto que nenhum destes edifícios foi observado sob a floresta. A distribuição
espacial dos cupinzeiros, analisada em três pastos, provou ser heterogênea e ligada
significativamente à profundidade do solo. As propriedades fisicas e quimicas destas
estruturas fizeram possível medir sua estabilidade estrutural e sua resistência ao
queimada anual dos pastos. A estabilidade estrutural da parede do cupinzeiro é muito
elevado comparada a do solo. Ela aumenta quando a estrutura biogenica é queimada. O
ninho é composto de duas porções distintas: a parede periférica e a “laminada” central.
Somente a “laminada” constitui uma reserva importante da matéria orgânica
(aproximadamente 77%). Ao consultar a granulometrria (argila 54% e areia grosseira
32%), o pH (pH=4) e as concentrações dos íons trocavel na parede ([Al3+] = 0.53
cmolc.kg-1, [Ca2+] = 1.5 molc.kg-1, [Mg2+] = 1.1 cmolc.kg-1, [K+] = 0.3 cmolc.kg-1),
é possível estimar a profundidade de remover dos materiais pelo cupins trabalhadores
para construir a parede de seu ninho entre 10 e 30 cm sob a superfície do solo. Sua
concentração do carbono (19 mg.g-1) é da mesma ordem que aquela do solo até 10 cm
da profundidade. A destruição destas estruturas realizadas pelos agricultures causa
imediatamente um efeito positivo local, mais negativo por causa da destruição da reserva
da matéria orgânica e de um aumento da estabilidade da parede. A concentração do
carbono na parede queimada comparado ao estado inicial é reduzida de 77%. A
“laminada” é consumido quase completamente pelo fogo. O pH da parede aumenta (por
4.6 a 5.4) e as concentrações nos cations são mais baixas na parede queimada do que
no estado fresco. Somente a concentração do phosphorus aumenta cerca 4 vezes. Os
efeitos do fogo nos termitières são assim de dois tipos: Se o aumento no pH e na
concentração do phosphorus disponíveis na parede tender a suportar o desenvolvimento
das plantas e da atividade microbiologica, a mudança da estrutura e do constitução
(formação do óxido do ferro) e a redução das concentrações nos cations e no carbono
têm um efeito negativo na fertilidade do material. A transformação da “laminada” em
cinzas terá que ser estudada mais precisamente para suplementar a avaliação química
destes cupinzeiros. Seria interessante especificar a progressão desta espécie nos pastos
amazonicos por um estudo de sua dispersão usando os métodos dos genetics das
populações.
Um segundo estudo foi desenvolvido no annelido Andiodrilus pachoensis. Um análise
comparativo das distribuições espaciais dos turriculos desta espécie foi realizada em 4
pastos e em 4 florestas. As relações entre a distribuição dos turricules e os testes

Resumo
padrões do espaço dos pés do graminaceous Brachiaria brisantha nos pastos foram
analisadas. As ligações entre a distribuição dos turriculos e a quantidade da liteira sob a
floresta foram estudadas. Nos dois meios, todos os turriculos e os elementos da
vegetação foram feitos um mapa em superfícies variáveis de 25m² ao 50m². A
densidade destes turriculos esta menor nos pastos (42.103/ha) que nas florestas
(243.103/ha). Sua distribuição é agregada nas escalas largas (>15m); visto que na
floresta, as análises do espaço destacaram um grande heterogeneidade entre as partes:
distribuição aleatória, agregada ou regular. A densidade dos turriculos não pareceu
correlacionada com a densidade dos pés de Brachiaria e sua produção é independent da
presença dos pés ou da quantidade da liteira nas florestas. Nas florestas e nos pastos, os
turriculos são compostos em uma média de 94% dos sais minerais e de 6% da matéria
orgânica. Como os solos testemunhos, a textura dos turriculos é composta
primeiramente da argila (aproximadamente 45%) e das areias grosseiras
(aproximadamente 30%). A densidade do agregado dos turriculos é globalmente mais
baixa do que aquela dos solos testemunhos. Nestes turriculos estam compactados e
pobre em matéria orgânica. Os turriculos do Andiodrilus pachoensis nao constituem uma
reserva do carbono. Sua concentração nos turriculos está na média 60 mg.g-1 nos
pastos, ligeiramente mais altamente do que nas florestas (53 mg.g-1), significativamente
mais fortes do que nos solos testemunhos (0-30 cm). Os turriculos nos dois meios
apresentam um pH ácido (4.6 e 5.8 respectivamente para as florestas e os pastos e 4
para as terras de referencia) e as concentrações nos cations são mais elevadas do que na
terra testemunha: sob a floresta, [Al3+] = 0.72 cmolc.kg-1, [Ca2+] = 4.7 molc.kg-1,
[Mg2+] = 2.2 cmolc.kg-1, [K+] = 0.27 cmolc.kg-1; sob o pasto, [Al3+] = 5.1 cmolc.kg-
1, [ca2+] = 7.3 molc.kg-1, [Mg2+] = 3.6 cmolc.kg-1, [K+] = 0.75 cmolc.kg-1. As
concentrações nos cations nos turriculos dos pastos são assim mais elevadas do que
naquelas da floresta. A distribuição em tamanho das partículas, a estabilidade estrutural
e a resistência à erosão sob a ação da chuva foram medidas em turriculos frescos e em
turriculos queimados de acordo com 2 intensidades da chuva. Os turriculos são mais
estáveis do que a terra testemunha e nenhum efeito físico (estabilidade) e o produto
químico (concentrações nos cations) do fogo nesta estrutura biogenica foram destacados.
Um terceiro estudo mediu as características químicas, obteve por método NIRS (perto
infravermelho Spectrophotometry), da matéria orgânica dos biostructuras e as licenças
para estabelecer um typologia das estruturas biogenicas (Andiodrilus pachoensis, outros
turriculos dos annélidos , paredes dos cupinzeiros de Cornitermes, outras estruturas e
cupinzeiros de cupims humivores, ninhos das formigas, dejeçoes dos coleopteros). O
estado da matéria orgânica (fresca ou degradada) provou ser o primeiro fator que separa

Resumo
as várias estruturas biogenicas estudadas. A origem e a natureza da matéria orgânica
não fazem parte significativa na diferenciação das estruturas biogenicas. Dois outras
tipologias eram estabelecidas a partir das concentrações quimicas das estruturas e o
estudo da porosidade após uma análise micromorphologica de 65 lâminas finas dos
biostructuras com duas definições (5,88µm e 0,47µm). As tipologias obtidas a partir da
análise dos poroïdos divididos em 3 categorias da forma e 5 classes do tamanho, são
diferentes nas duas definições. As diferenças entre os tipologias obtidos pelos vários
métodos forçam a importância para considerar um máximo de características fisico-
quimicas a fim de desenvolver as ferramentas metodologicas que visam separando os
agregados do solo sujeitado ao variabilidade local forte. Os resultados dos tipologias não
destacaram fortes diferenças entre os grandes tipos taxonomicos mas um importante
variabilidade em seu centro (variabilidade inter-especies). Modelisação da dinâmica dos
agregados biológicos do solo posa assim o problema da modelisação generalo no nível
taxonomico do ordem e mesmo da família. A acumulação do dados específicos sobre
espécies conhecidas como “coordenadores ou chaves” dos ecosystemos a fim de definir
grupos funcionais, parece assim um estágio necessário a modelar o funcionamento do
solo.
Palavras chave : Andiodrilus pachoensis / Cornitermes sp. / turriculos / cupinzeiro /
laminada / xilofagia / distribução espacial / propriedades fisicas / cations / stabilidade
estructural / Brachiaria brisantha / NIRS / lamina fina / pastos quemados / simulaçao de
chuva / Amazonia

Table des Matières
1
Table des matières._________________________

Table des Matières
2
INTRODUCTION GENERALE .......................................................................................... 7
I. INTRODUCTION....................................................................................................... 10
I.1 LA FORET AMAZONIENNE BRESILIENNE. ............................................................. 11 I.1.1 Hétérogénéité et biodiversité de la forêt amazonienne brésilienne. ....................... 11 I.1.2 La déforestation............................................................................................ 13 I.1.3 Conséquences du déboisement et utilisation des sols en Amazonie. ...................... 16 I.1.4 Impact du feu sur le sol des cultures sur brûlis. ................................................. 17
I.1.4.a Modifications des propriétés physiques et minéralogiques du sol des
cultures sur brûlis.............................................................................................. 17 I.1.4.b Modifications des propriétés chimiques et de la matière organique du sol
dans les cultures sur brûlis. ................................................................................ 18 I.1.4.c Modifications des propriétés biologiques du sol dans les cultures sur brûlis. ..... 19
I.2 LES INVERTEBRES ET LE FONCTIONNEMENT DU SOL ........................................... 20 I.2.1 Classification de la faune du sol....................................................................... 20 I.2.2 Le modèle hiérarchique, les ingénieurs de l’écosystème et leurs structures
biogéniques. ........................................................................................................ 22 I.2.3 Formation d’un sol ou pédogénèse................................................................... 27 I.2.4 Processus du sol affectés par l’activité des ingénieurs......................................... 29
I.2.4.a Effets sur la dynamique de la matière organique.......................................... 29 I.2.4.b Effets sur la structure physique. ................................................................ 30
I.3 FERTILITE ET PROPRIETES PHYSICO-CHIMIQUES DU SOL................................... 31 I.4 PROBLEMATIQUE ET OBJECTIFS .......................................................................... 36
II. LE SITE D’ETUDE. ................................................................................................... 38
II.1 CARACTERISATION DU SITE D’ETUDE. ............................................................... 39 II.1.1 Caractérisation générale. .............................................................................. 39 II.1.2 Le climat. ................................................................................................... 41 II.1.3 Mode de gestion et pratiques agricoles............................................................ 42 II.1.4 Les parcelles étudiées................................................................................... 43
II.2 LA MACROFAUNE DU SOL (Mathieu, 2004) ......................................................... 46 II.2.1 Les populations de Macroinvertébrés de « Benfica ».......................................... 46 II.2.2 Choix des structures biogéniques étudiées....................................................... 47
III. LES METHODES D’ANALYSES................................................................................. 48
III.1 ANALYSES STATISTIQUES................................................................................. 49

Table des Matières
3
III.1.1 Quelques définitions (Krebs, 1998)................................................................ 49 III.1.2 Analyses statistiques classiques. ................................................................... 50 III.1.3 Analyses statistiques spatiales. ..................................................................... 50
III.1.3.a Les processus de points. ........................................................................ 50 III.1.2.b Les Fonctions de Ripley.......................................................................... 51 III.1.2.c Les densités locales de 1er ordre.............................................................. 53 III.1.2.d Les Analyses intertypes ....................................................................... 54
III.1.4 Indice de diversité de Shannon ..................................................................... 55 III.2 ANALYSES PHYSIQUES ET CHIMIQUES. ............................................................ 56
III.2.1 Analyses physiques des horizons et des structures biogéniques. ........................ 56 III.2.2 Analyses de la composition chimique des horizons témoins et des structures
biogéniques. ........................................................................................................ 58
IV. SUBSTRAT GEOLOGIQUE ET COUVERTURE PEDOLOGIQUE DU SITE D’ETUDE. ......... 60
IV.1 SUBSTRAT GEOLOGIQUE ET ORGANISATION DE LA COUVERTURE
PEDOLOGIQUE........................................................................................................... 61 IV.1.1 Substrat géologique..................................................................................... 61 IV.1.2 Couverture pédologique ............................................................................... 61
IV.1.3 Description des fosses pédologiques. .......................................................... 63 IV.1.3.a Description des profils pédologiques sous forêt .......................................... 63 IV.1.3.b Description des profils pédologiques sous pâturages................................... 67 IV.1.3.c Autres parcelles .................................................................................... 70
IV.2 CARACTERISTIQUES PHYSICO-CHIMIQUES DES SOLS TEMOINS........................ 71 IV.2.1 Les latosols amazoniens. .............................................................................. 71 IV.2.2 Profils granulométriques des sols.................................................................. 71 IV.2.3 Densité apparente et densité de mottes des sols témoins.................................. 72 IV.2.4 Stabilité structurale des agrégats du sol en forêt et en pâturage. ....................... 73 IV.2.5 Bilan chimique des horizons de surface des sols de forêt et de pâturages. .......... 75
IV.2.5.a Dosage du carbone du sol....................................................................... 75 IV.2.5.b Le pH. ................................................................................................. 77 IV.2.5.c Les cations, la CEC et le taux de saturation. .............................................. 78 IV.2.5.d Effets du brûlis sur les propriétés chimiques du sol..................................... 82
V. ETUDE DES TERMITIERES DU GENRE CORNITERMES. ............................................. 83
V. 1 MATERIEL ET METHODES.................................................................................... 84 V.1.1 Le termite du genre Cornitermes .................................................................... 84 V.1.2 Etude des termitières de Cornitermes sp.......................................................... 85

Table des Matières
4
V.2 RESULTATS.......................................................................................................... 88 V.2.1 Etude de la distribution spatiale des termitières de Cornitermes sp.au niveau
des parcelles........................................................................................................ 88 V.2.1.a Densité des termitières de Cornitermes sp. ................................................ 88 V.2.1.b Fréquence en classes de taille des termitières de Cornitermes sp................... 89 V.2.1.c Distribution spatiale des termitières épigées de Cornitermes sp dans les
pâturages......................................................................................................... 91 V.2.1.d Relation avec le type de sol...................................................................... 93
V.2.2 Etude des caractéristiques physico-chimiques des termitières de
Cornitermes sp..................................................................................................... 95 V.2.2.a Quantification et propriétés du feuilleté interne du nid. ................................ 95 V.2.2.b Propriétés physiques de la muraille des termitières...................................... 98
b1) Granulométrie de la muraille de termitières (annexes 3 et 3bis)..................................................... 98 b2) Densité de motte des agrégats de muraille des termitières épigées. ............................................... 99 b3) Stabilité structurale de la muraille de termitières.......................................................................... 99
V.2.3 Bilan des éléments chimiques séquestrés dans les murailles des termitières de
Cornitermes sp................................................................................................... 102 V.2.3.a Concentration en carbone des murailles de termitières de Cornitermes sp..... 102 V.2.3.b Le pH des murailles de termitières de Cornitermes sp. ............................... 103 V.2.3.c Concentration en ions dans la muraille de termitière. ................................. 103
V.2.4 Simulation de l’impact du feu sur les termitières. ............................................ 107 V.2.4.a Action du feu sur les murailles des termitières de Cornitermes sp. .............. 107
a1) Texture. .................................................................................................................................107 a2) Stabilité structurale des murailles brûlées. .................................................................................109 a3) Dosage du carbone dans les murailles brûlées. ...........................................................................110 a4) pH des murailles brûlées. .........................................................................................................111 a5) Dosage des cations..................................................................................................................112 A6) Dosage du phosphore assimilable..............................................................................................114
V.2.4.b Action du feu sur le feuilleté des termitières de Cornitermes sp. ................. 114 V.3 DISCUSSION...................................................................................................... 116
VI. ETUDE DES TURRICULES DE L’ANNELIDE ANDIODRILUS PACHOENSIS................. 123
VI.1 MATERIEL ET METHODES ................................................................................ 124 VI.1.1 L’espèce Andiodrilus pachoensis. ................................................................. 124 VI.1.2 Cartographie des turricules d’Andiodrilus pachoensis ...................................... 124 VI.1.3 Analyse des propriétés physico-chimiques des turricules ................................. 127 VI.1.4 Expérience de simulation de l’impact du feu sur les turricules d’Andiodrilus
pachoensis. ....................................................................................................... 128 VI.1.4 Expérience de simulation de l’impact de la pluie sur les turricules..................... 129

Table des Matières
5
VI.2 RESULTATS ...................................................................................................... 130 VI.2.1 Etude de la distribution spatiale des turricules d’Andiodrilus pachoensis. ........... 130
VI.2.1.a Densité et masse des turricules de Andiodrilus pachoensis dans les
différents milieux ............................................................................................ 130 VI.2.1.b Production et dégradation des turricules. ................................................ 134 VI.2.1.c Distribution spatiale des turricules d’Andiodrilus pachoensis....................... 137 VI.2.1.d Stabilité dans le temps de la distribution spatiale de la production des
turricules. ...................................................................................................... 141 VI.2.1.e Relation avec les autres structures biogéniques rencontrées. ..................... 143 VI.2.1.f Relation avec la distribution spatiale des arbres dans les forêts................... 144 VI.2.1.g Expérimentation.................................................................................. 145
g1) Sous forêt, relation avec la quantité de litière. ............................................................................145 g2) Sous pâturage, relation avec la présence de Touffes de Brachiaria brisantha. .................................146
VI.2.1.h relation avec la température du sol. ....................................................... 147 VI.2.2 Propriétés physiques des turricules d’Andiodrilus pachoensis ........................... 148
VI.2.2.a Granulométrie (données annexes 18, 18bis et 19) ................................... 148 VI.2.2.b Densité de Motte................................................................................. 150 VI.2.2.c Stabilité structurale ............................................................................. 151
VI.2.3 Bilan Chimique de séquestration des éléments.............................................. 153 VI.2.3.a Le Carbone......................................................................................... 153 VI.2.3.b Le pH ................................................................................................ 156 VI.2.3.c Les cations et le phosphore................................................................... 157
VI.2.4 Dégradation des Turricules. ........................................................................ 163 V.2.4.a Action du feu sur les turricules. .............................................................. 163
a1) Modifications granulométriques (annexe 20)...............................................................................164 a3) Modifications chimiques. ..........................................................................................................169
VI.2.4.b Action de la pluie sur les turricules d’Andiodrilus pachoensis. ..................... 173 VI.3 DISCUSSION. ................................................................................................... 176
VII. MISE AU POINT D’UNE TYPOLOGIE DES STRUCTURES BIOGENIQUES. .................. 184
VII.1 INTRODUCTION.............................................................................................. 185 VII.2 MATERIEL ET METHODES................................................................................ 187
VII.2.1 Etude de la matière organique par la méthode NIRS (Near Infrared
Resonance Spectophotometry). ............................................................................ 187 VII.2.2 Analyse chimique des structures biogéniques ............................................... 188 VII.2.3 Analyses micromorphologiques de Lames minces.......................................... 188
VII.3 RESULTATS..................................................................................................... 190 VII.3.1 Typologie basée sur l’étude de la matière organique par la méthode NIRS........ 190

Table des Matières
6
VI.3.1.b Analyses multivariées des données de chimie. ......................................... 191 VII.3.2 Analyse micromorphologique des lames minces de biostructures..................... 193
VII.3.2.a Description succincte des lames minces correspondant aux différentes
structures biogéniques. .................................................................................... 193 VII.3.2.b Quantification de la porosité des différentes structures biogéniques........... 196
b1) Résolution de 5,88µm par pixel (annexe 21)...............................................................................196 b2) Résolution de 0,47µm par pixel (annexe 22)...............................................................................199 Les images étudiées à cette échelle mesurent 1500 * 1200 pixels.......................................................199
VII.3.2.c Diversité des poroïdes et typologie des biostructures ............................... 201 c1) Diversité des structures biogéniques. .........................................................................................202 C2) Diversité des genres taxonomiques. ..........................................................................................203 c2) Résolution de 0,47µm par pixel .................................................................................................204
VII.4 DISCUSSION .................................................................................................. 206
VIII. DISCUSSION GENERALE ET CONCLUSION .......................................................... 210
ANNEXES ................................................................................................................. 220
REFERENCES BIBLIOGRAPHIQUES ........................................................................... 246
LISTE DES ILLUSTRATIONS....................................................................................... 264
Tableaux.................................................................................................................. 265 Figures .................................................................................................................... 266 Photos ..................................................................................................................... 272 Illustrations............................................................................................................. 272

Introduction. Chapitre I
7
Introduction générale

Introduction. Chapitre I
8
Tout au long de l’histoire et sur tous les continents, les populations humaines ont
vécu et se sont développées en intéraction avec leur environnement en acquérant la
connaissance empirique, voire scientifique depuis quelques années, de la diversité
biologique et écologique. Cette relation avec la nature leur a permis de connaître et de
créer des techniques spécifiques de gestion des divers écosystèmes, utilisant leur savoir
au développement de techniques adéquates à l’exploitation des ressources naturelles. Le
savoir « paysan », transmis au fil des générations n’a cepedant pas toujours permis aux
écosystèmes naturels fertiles de perdurer jusqu’à aujourd’hui.
L’augmentation des populations humaines et l’agriculture intensive font
qu’actuellement, un certain nombre d’écosystèmes sont menacés.
Le terme « écologie » est désormais connu de tous tant il est incessamment sous
les feux des medias et tend à se distinguer comme une mode. Ce terme, créé en 1866
par Ernst Haeckel, définit l’ensemble des études consacrées aux relations entre les êtres
vivants mais aussi entre ceux-ci et le milieu qu’ils habitent. Ce terme a ensuite évolué
pour y intégrer, entre autre, la dimension « adaptative » de Darwin qui fut l’un des
précurseurs de la science des sols avec la publication en 1881 de son ouvrage intitulé
« Rôle des vers de terre dans la formation de la terre végétale ».
Parmi l’ensemble des milieux terrestres colonisés par les êtres vivants, le sol est
sans doute celui qui impose les plus importantes contraintes adaptatives. Nous
appellerons sol, les premiers centimètres de la surface de la croute continentale (jusqu’ à
la roche mère). Ces contraintes concernent tant le déplacement, la respiration due aux
fortes variations d’aération et d’humidité, et la nutrition en raison de la pauvreté des
ressources nutritives qu’il abrite (Lavelle et al., 1995). Cependant des centaines
d’espèces animales ont colonisé les sols. Ces communautés sont parmi les plus
diversifiées de la planète avec un nombre d’espèces estimé de 5 à 80 millions
d’arthropodes entre autres (Giller et al., 1997). Malgré les nombreuses études
biologiques réalisées ces dernières années, le sol est resté par sa complexité un système
peu connu aussi bien en ce qui concerne son fonctionnement qu’en ce qui concerne le
rôle et la diversité des individus qui y vivent.
Très souvent la science du sol s’est limitée à l’étude physico-chimique plus ou
moins dynamique du sol en décrivant des horizons et expliquant leur genèse (pédologie)
ou bien s’est uniquement consacrée à l’étude de la vie dans le sol (microorganismes,
macroinvertébrés) sans pour cela prendre en compte tous les paramètres de l’écotope.
Plus récemment des études se sont intéressées à son fonctionnement afin de mieux

Introduction. Chapitre I
9
définir l’impact des pratiques agricoles. Les sols abritent des processus écologiques
fondamentaux auxquels la participation des invertébrés dans leur régulation est avérée
(Lee and Foster, 1991 ; Chauvel et al., 1999 ; Lavelle et al., 1999). Cette biocénose
constitue l’un des supports d’une agriculture maîtrisée et par conséquent une ressource
naturelle indispensable à la survie de l’humanité.
Ce travail cherche à concilier ces trois approches (écotope, biocénose et
fonctionnement) de la science du sol. Tout en tentant d’apporter des réponses de gestion
durable à l’exploitation des sols, une étude de l’activité de biocénose sur la fertilité du sol
a été entreprise dans les zones pionnières de déforestation en Amazonie.
Cette région, qui constitue la plus grande forêt tropicale de la planète nous a servi
de cadre d’étude. Suite à la déforestation, c'est-à-dire au bouleversement total de
l’écosystème (couvert végétal et zone racinaire du sol) qui les abritait, de nombreuses
espèces de forêts vont s’éteindre ou s’adapter. Ce changement de milieu va également
s’accompagner de la colonisation de nouvelles espèces (Höfer et al., 2001 ; Wanner and
Dunger, 2002 ; Mathieu, 2004). Le changement d’abondance et de diversité des
populations de macroinvertébrés du sol s’accompagne d’une modification de l’activité de
ces populations animales et donc de leur influence sur les paramètres physico-chimiques
du sol. Notre contribution, à cette immense étude, qui consiste à analyser puis
comprendre le fonctionnement des 30 premiers centimètres de la surface terrestre, en
vue d’une éventuelle modélisation qui permettrait d’anticiper les conséquences de la
déforestation, a été ciblée sur deux espèces abondantes de l’écosystème étudié. Nous
avons étudié l’influence du termite Cornitermes sp. et de l’annélide Andiodrilus
pachoensis, à travers les structures biogéniques qu’ils produisent et la dégradation de
ces dernières, sur les propriétés physico-chimiques du sol. Certaines études de notre
groupe de travail ont d’ores et déjà dénombré et étudié la diversité des structures
biogéniques ou encore mis en évidence les caractéristiques physiques particulières de
certaines d’entre elles (Blanchart et al., 1993a ; Decaëns et al., 2001). L’originalité de
cette étude est d’aborder d’une part la distribution spatiale des structures biogéniques
produites par les deux espèces à une grande échelle et d’autre part de comparer les
propriétés physico-chimiques de ces structures pour s’intéresser à leur dégradation et
leur intégration dans le sol. Ce travail, réalisé à différentes échelles, a permis d’apporter
des éléments de compréhension du rôle des deux espèces étudiées sur la variabilité
locale du sol et donc la mise en place de niches écologiques propices ou non au
développement d’autres espèces végétales ou animales.


Introduction. Chapitre I
10
I. Introduction

La forêt amazonienne brésilienne. Chapitre I.1
11
I.1 LA FORET AMAZONIENNE BRESILIENNE.
La forêt amazonienne correspond au stéréotype de la forêt encore vierge. Le mot
suffit à évoquer la chaleur, l’ombre, la luxuriance et les bruissements d’un monde qui
foisonne sous toutes les formes de la vie. Cette forêt appelle au respect mais aussi à
la fascination. C’est à juste titre, sans doute, que le mot désigne dans le langage
courant une contrée aux richesses surabondantes.
Trop souvent, vue d’Europe, l’utilisation du mot “ Amazonie ” se restreint à la forêt
tropicale. Au Brésil, l’Amazonie est une région, un espace convoité, exploité et peuplé qui
s’aménage et s’urbanise sous le nom de région Nord. La région Nord est une des cinq
grandes régions administratives du Brésil, se différenciant du reste du pays. L’Amazonie
n’est pas qu’une forêt et l’Amazonie n’est pas le Brésil. Le bassin amazonien occupe 7
millions de km2 dont presque 60% reviennent au seul Brésil. Mais d’autres états se
partagent le massif forestier amazonien comme le Pérou, la Bolivie, l’Equateur, la
Colombie, le Venezuela, les Guyanes. La forêt appartient donc à plusieurs Etats même si
le Brésil en détient la plus grande part.
Cette région brésilienne trop souvent associée à l’unique image de la forêt est une
région géographique à part entière. Le Brésil possède 3,6 millions de km² de forêt
(réparties entre forêt tropicale et forêt atlantique), ce qui le place à la 3ème place
mondiale des pays contenant la plus dense surface forestière du monde. C’est la plus
grande forêt tropicale de la planète avec une surface trois fois supérieure à celle du
Congo.
I.1.1 Hétérogénéité et biodiversité de la forêt amazonienne brésilienne.
L’homogénéité n’existe pas dans le relief, le climat, le sol et donc la forêt. En
Amazonie brésilienne, il existe trois grandes catégories de forêts : la forêt de Terra
Firme, la forêt de Varzea et la forêt d’Igapo.
- la forêt de Terra Firme (Terre Ferme) : 253 millions d’hectares.
La forêt de Terra Firme est la plus importante en superficie. Elle s’étend entre les
rivières, sur les interfluves.
La forêt de Terra Firme, “ Mata Densa ” est composée d’arbres géants de 40 à 50 mètres
de hauteur, dont la canopée est très dense, d’arbres moyens de 30 à 40 mètres de haut
et d’une strate arborée de 15 à 20 mètres. Peu de graminées au sol mais surtout des

La forêt amazonienne brésilienne. Chapitre I.1
12
mousses, des champignons qui transfèrent directement aux racines les éléments
nutritifs, des lianes (cipos) et des épiphytes. La forêt de Terra Firme est riche en
essences “ dures ” de densité souvent importante. Il existe aussi certaines nuances selon
les facteurs pédologiques, topographiques, climatiques et édaphiques. Il est donc difficile,
tant l’étendue de l’Amazonie est importante de parler d’uniformité de la forêt de « Terre
Ferme ».
- la forêt de Varzea
La forêt de Varzea est située dans les zones inondables. A l’époque des crues, elle est
périodiquement inondée de Décembre à Juin. En général, elle s’étend sur une largeur qui
peut atteindre jusqu'à 80 kilomètres de part et d’autre des rivières. Cette forêt se
compose surtout d’essences dites “ blanches ”, essences de bois tendres à croissance
rapide et d’un éventail de palmiers. Ce fut dans l’histoire de l’Amazonie, la première forêt
exploitée, bien avant la construction des routes pour d’évidentes raisons d’accessibilité.
D’ailleurs, la quasi totalité des bois extraits était exportée par flottage avant l’ouverture
routière.
- la forêt d’Igapo.
Les Igapos sont des terres marécageuses dont les sols sont hydromorphes, toujours
inondés, possédant une végétation dense mais basse, pauvre en qualité et totalement
inextricable. La forêt d’Igapo a en commun avec la forêt de Varzea quelques essences qui
ne sont pas exploitées aujourd’hui du fait des difficultés techniques. Paradoxe de
l’accessibilité, les terres les plus accessibles autrefois sont celles qui le sont le moins
aujourd’hui depuis l’ouverture routière amazonienne voulue par l’Etat.
A ces trois types de forêt s’ajoute une formation secondaire qui remplace la forêt
dense primaire (celle où l’homme n’est pas intervenu), quand celle-ci a été défrichée.
Cette dernière est plus basse avec un sous-bois plus dense. Dans la mesure où les
grands arbres disparaissent, la lumière atteint plus facilement le sol, permettant le
développement d’un sous-sol dense.
Le terme « biodiversité » ou « diversité biologique » est utilisé pour décrire les
variétés de formes de vie dans une région. Le calcul de la biodiversité se fait à travers la
quantité d’écosystèmes, d’espèces vivantes, des patrimoines génétiques et endémiques.
La forêt amazonienne, au moins dans sa partie brésilienne, recèle la plus grande
biodiversité avec de 10% à 20% des 1,5 millions d’espèces animales et végétales

La forêt amazonienne brésilienne. Chapitre I.1
13
référencées (Ministério do Meio Ambiente (MMA), 1998). Ce qui représente 56 000
espèces de plantes dont près de 7000 endémiques, 502 mammifères, 1677 oiseaux, 600
amphibiens, 2657 poissons (Joly and Bicudo, 1999). C’est également le pays ayant la
plus grande richesse endémique (779 vertébrés) (Myers et al., 2000). La classe des
arthropodes est la plus nombreuse sous les tropiques avec environ 800 000 espèces
connues et de 30 à 100 millions estimées. La disparité entre le nombre d’espèces
référencées et ce que nous supposons exister est très importante. Sur un seul arbre en
forêt, 200 espèces d’arthropodes différentes peuvent être rencontrées (Erwin, 1982).
L’Amazonie joue un rôle primordial dans la stabilité environnementale de la planète ; elle
abrite sous forme fixée une centaine de milliards de tonnes de carbone, sa masse
végétale libère près de 7 milliards de tonnes d’eau annuellement dans l’atmosphère par
évapotranspiration et ses fleuves représentent 20% de toute l’eau douce rejetée dans les
océans sur le globe (Capobianco, 2001).
Ce patrimoine environnemental brésilien est arrivé jusqu’à aujourd’hui avec ses
caractéristiques originales relativement bien conservées. Il constitue un grand potentiel
et une réserve pour les progrès biotechnologiques et génétiques. D’autre part le rapport
national brésilien sur la diversité biologique (Ministério do Meio Ambiente (MMA), 1998)
affirme que ce patrimoine a une importance décisive sur le plan économique. Aujourd’hui
l’exploitation de la biodiversité représente près de 5% du PIB du pays avec 4% de
l’exploitation forestière et 1% du secteur recherche alors que l’utilisation du patrimoine
génétique brésilien pourrait représenter jusqu’à 40% du PIB sans compter le marché en
expansion des produits biotechnologiques (Arnt, 2001). Cependant le futur de l’Amazonie
ne semble pas assurer par son importance environnementale et/ou son potentiel
intrinsèque, et les menaces de dégradation avancent à un rythme accéléré.
I.1.2 La déforestation.
En 1978, la forêt amazonienne couvrait près de 4 millions de km². En 1998, il ne
restait plus qu’environ 86 % de cette surface, 517 000 km2 ayant été défrichés en 20
ans (INPE, 2001). Le rythme moyen de la déforestation s’est accéléré dans les années 90
(17 000 km²/an) et on estime aujourd’hui que la superficie totale des forêts
fragmentées, défrichées ou affectées par des phénomènes de bordure représente le tiers
de l’Amazonie brésilienne (Figure 1). Les derniers calculs de l’INPE estiment à près de
47 000 km² la surface de forêt éliminée entre 2001 et 2003. Cependant la déforestation
est cartographiée à l’aide d’images satellitales Landsat et une étude de 1999 a démontré
que les feux et l’exploitation du bois étaient invisibles par Landsat et endommageaient

La forêt amazonienne brésilienne. Chapitre I.1
14
des milliers de kilomètres carré de forêt chaque année (Laurance and Fearnside, 1999).
Le ministère brésilien de l'environnement a annoncé, le 26 août 2005, que la
déforestation en Amazonie avait beaucoup décru sur la période 2004-2005 par rapport
aux années précédentes. Avec 9 100 km2, selon les données satellitales fournies par
l'INPE (Institut national de recherche spatiale), c'est une diminution drastique par rapport
au chiffre très élevé de 2003-2004 (26 130 km2), qui succédait lui-même à deux très
mauvaises années (24 597 km2 pour 2002-2003 et 23 266 km2 pour 2001-2002). Le
dernier chiffre officiel annoncé n'est encore qu'une estimation, qui sera sans doute revue
à la hausse, mais il marque un infléchissement net de la tendance. On n'avait pas connu
un chiffre aussi bas depuis la saison 1990-1991, où 11 000 km2 de forêt avaient été
détruits (Kempf, 2005).
Le processus de déforestation de l’Amazonie brésilienne trouve son origine dans
l’action de l’Etat, qui à partir de 1950 s’est intéressé à l’intégration des zones peu
peuplées et méconnues du nord par la construction de routes (routes Brasília-Belém et
Brasília-Acre dans les années 1950-1960 et 1970, Transamazonienne depuis les années
1970) (Droulers, 1995).
INPE PRODES Digital, 2004.INPE PRODES Digital, 2004.
Légende
Déforestation avant 2002
Déforestation en 2002/2003
INPE PRODES Digital, 2004.INPE PRODES Digital, 2004.
Légende
Déforestation avant 2002
Déforestation en 2002/2003
Légende
Déforestation avant 2002
Déforestation en 2002/2003
Figure 1 : Carte de la déforestation 2002/2003, Forêt amazonienne brésilienne (INPE, 2004).
A la création de ces infrastructures de transport est associée un vaste programme
de colonisation agricole qui a provoqué une multiplication par dix depuis 1960 de la

La forêt amazonienne brésilienne. Chapitre I.1
15
population humaine de la région (Goodman and Hall, 1990). Ne nécessitant qu'un faible
investissement et donnant des retours financiers rapides, l'élevage bovin fut
immédiatement la modalité choisie par la grande majorité des nouvelles exploitations.
Dans les cas de régions «ouvertes» par les grandes exploitations, les grands
propriétaires défrichent rapidement de vastes zones pour assurer la possession de la
terre, créant des exploitations qui ne s'avèrent pas toujours rentables mais dont l'intérêt
principal, outre le prestige social, est la spéculation foncière. Une autre colonisation s’est
mise en place par la politique de réforme agraire (INCRA), des familles de paysans se
sont installées sur des lots d’une centaine d’hectares en 1970 mais plus souvent entre 25
et 60 hectares dans les années 90, mais la différenciation sociale fit rapidement son
apparition, certains réussissant à acheter les lots de leurs voisins et se bâtissant ainsi des
propriétés de taille moyenne. Rares sont les régions où les petits propriétaires
réussissent à se maintenir, et la plupart du temps eux aussi optent pour l'élevage bovin.
Les routes construites ont par ailleurs rendu accessibles ces régions forestières à des
colonisateurs pratiquant l’abattage industriel et développant des industries extractives.
La foresterie brésilienne exploite en grande majorité des aires naturelles et pas de forêt
de plantation. Malgré un certain boycott européen du bois précieux, le bois amazonien
est consommé à 75% au Brésil même (Smeraldi and Verissimo, 1999), où le marché ne
cesse de croître. Un nouveau groupe apparaît ces dernières années : celui des planteurs
de soja. Attirée par la forte productivité végétale amazonienne, cette activité est en
pleine expansion. Entre 1990 et 2002, l'Etat du Mato Grosso a bondi du troisième au
premier rang des producteurs de soja brésilien, multipliant sa production par quatre et
atteignant près de 12,7 millions de tonnes en 2002 (près de 7% de la production
mondiale) (Le Tourneau, 2004).
Face à cette déforestation élevée, et en raison de la place croissante de l'écologie
dans les préoccupations des opinions publiques des grands pays industrialisés, de
nombreuses stratégies de protection de la forêt sont mises en place. Dès le mandat du
président Sarney (1985-1990), un Institut chargé de protéger l'environnement (IBAMA),
et plus particulièrement l’Amazonie, est créé. Sous le mandat du président Collor (1990-
1992) se tient à Rio de Janeiro le fameux «sommet de la terre», et la terre indigène
Yanomami (96 500 km2) est déclarée zone de préservation. Le gouvernement de
Fernando Henrique Cardoso (1994-2002), avec l'appui du programme du G7 élargit la
zone de protection à environ 1 575 000 km2 (soit 31,5% de l'Amazonie). Avec 15% de la
forêt aujourd’hui déboisée, il reste encore de vastes zones, en particulier dans les États
d'Amazonas et du Pará, dont le destin n'est pas défini.

La forêt amazonienne brésilienne. Chapitre I.1
16
Pour remédier à la réduction de la diversité biologique, les autorités ont
également fait un gros effort dans les domaines de la certification des bois et de la
préservation des forêts (PNUE-CEPALC, 2001). Cependant le positionnement de l’Etat
fédéral brésilien est ambigu ; il juxtapose à d’indéniables mesures de protection de la
forêt, des calculs économiques ou politiques, en particulier la redistribution des terres des
grands propriétaires à des milliers de nouveaux petits paysans en Amazonie.
Les restes de la forêt sont par conséquent menacés par l’aménagement des côtes,
l’abattage incontrôlé, l’expansion de l’agriculture avec des filières qui persistent (bois et
élevage bovin), de nouvelles cultures (culture du soja) et la production toujours
croissante de charbon de bois. La dévastation que subit l’Amazonie a de graves
conséquences écologiques, économiques et sociales à court et long terme au niveau
local, régional et mondial.
I.1.3 Conséquences du déboisement et utilisation des sols en Amazonie.
Les études réalisées par la « Food and Agriculture Organization » (FAO) ont
montré que la plupart des changements de paysage dans les zones tropicales se
caractérisent par une transition de la forêt aux systèmes agro-pastoraux (FAO, 1996). En
Amazonie brésilienne, l'agriculture familiale occupe une place importante dans les
activités de subsistance. Ces pratiques agricoles englobent la culture du manioc, du riz et
de la banane, les plantations d'arbres fruitiers et celles de graminées pour le bétail. Ces
pratiques agricoles donnent des rendements modestes pour les familles et
l'environnement subit de nombreux effets néfastes.
La première modification débute juste après la déforestation, même si les sols ne
sont pas immédiatement utilisés. Ces altérations sont liées à la qualité et la quantité de
matière organique déposée au sol et par conséquent à son turnover.
Une dégradation du sol peut aussi s’opérer quel que soit le couvert végétal
(naturel ou cultivé) en raison de l’érosion suscitée par l’écoulement des eaux de pluie
(Pereira, 1977 ; Guerra et al., 1999). Les sols amazoniens sont très chargés en fer et en
mercure. Ces éléments sont rejetés dans l’eau suite à l’érosion et le ravinement. La
déforestation laisse une terre nue exposée aux pluies intenses. L’activité microbiologique
étant très intense, le mercure inorganique se transforme en methylmercure, fortement
toxique et facilement assimilé par les organismes aquatiques, contaminant ainsi toute la
chaîne alimentaire. Les nutriments sont également lessivés et les sols, sans aucune

La forêt amazonienne brésilienne. Chapitre I.1
17
protection, ne sont plus maintenus. Ces processus de « désertification » aboutissent à
des sols acides, stériles, pollués de métaux lourds (Carmouze et al., 2001).
Par ailleurs une forte perte de biodiversité s’observe. Selon les estimations, 300
espèces d’arbres disparaissent par an. A la disparition de ces arbres se rajoute la perte
d’arthropodes, d’épiphytes, d’oiseaux en association symbiotique avec ces arbres. Ainsi
plus de 180 000 arthropodes disparaîtraient par an. De nombreuses espèces s’éteignent
avant même qu’elles soient référencées et étudiées.
La déforestation joue également un rôle sur le climat. Localement la destruction
de la forêt réduit les précipitations. La balance hydrique est altérée. Au niveau planétaire,
la contribution de la déforestation sur l’accroissement de l’effet de serre et les
changements climatiques qui lui sont associés sont aujourd’hui étudiés.
La durabilité des cultures sur brûlis étant réduite (Gehring et al., 2005), des
systèmes alternatifs réduisant la détérioration de la structure du sol et maintenant sa
fertilité afin de conserver une bonne productivité à long terme sont à l’étude (Alegre and
Cassel, 1996).
Dans les processus de conversion de la forêt en nouvelles aires agricoles, les
conséquences sur le sol peuvent être plus sévères lorsque le feu est utilisé. Les effets
sont multiples et fonction de l’intensité et de la durée du feu (Certini, 2005).
I.1.4 Impact du feu sur le sol des cultures sur brûlis.
De très nombreuses études ont mis en évidence l’effet négatif de la culture sur
brûlis sur la fertilité des sols (Alegre et al., 1996 ; Juo and Manu, 1996 ; Lessa et al.,
1996 ; Kotto-Same et al., 1997 ; Tanaka et al., 1998 ; Garcia-Montiel et al., 2000 ;
Batabyal and Lee, 2003 ; Eneji et al., 2003 ; Norgrove et al., 2003 ; Araujo et al., 2004)
(Da S. Martins et al., 1991 ; Bird et al., 2000 ; Ketterings and Bigham, 2000 ; Dezzeo et
al., 2004).
I.1.4.a Modifications des propriétés physiques et minéralogiques du sol des
cultures sur brûlis.
•La perméabilité à l’eau des sols diminue en raison de la formation d’un horizon continu
imperméable de quelques centimètres sous la surface. Cela implique une augmentation
des écoulements superficiels et l’érosion.

La forêt amazonienne brésilienne. Chapitre I.1
18
•La stabilité structurale diminue en raison notamment d’une réduction des complexes
organo-minéraux, comme résultat de la combustion des ciments organiques.
•La densité apparente augmente, la porosité et la perméabilité diminuent en raison de la
destruction des agrégats et du comblement des vides par l’argile dispersée, pouvant
aboutir à la formation d’une croûte (Martinez and Zinck, 2004 ; Mills and Fey, 2004).
•La distribution en taille des particules minérales ne change pas directement, sauf
lorsque l’érosion entraîne de manière sélective la fraction fine et concentre relativement
les plus grosses particules (sables) (Obale-Ebanga et al., 2003).
•La nature minéralogique des sols ne change qu’à des températures supérieures à
500°C.
•La couleur devient plus sombre, en raison de la carbonisation et rougit en raison de la
formation d’oxydes de fer non hydratés.
•le régime de température change temporairement en raison de la disparition de la
couverture végétale d’un part et d’autre part de l’assombrissement du sol.
I.1.4.b Modifications des propriétés chimiques et de la matière organique du sol
dans les cultures sur brûlis.
•La quantité de matière organique diminue immédiatement après le feu mais, à plus long
terme, elle excède généralement le niveau d’avant le feu.
•La qualité de la matière organique change remarquablement avec un enrichissement
relatif de la fraction plus récalcitrante aux attaques biochimiques. Ceci est dû d’une part
au brûlage sélectif des résidus frais (feuilles, petites branches…) et à la néoformation de
résidus aromatiques fortement polymérisés (comme des acides humiques).
•Le pH des sols (non calcaires) augmente, au moins éphémèrement, en raison de la
libération des cations alcalins (Ca, Mg, K, Na) par la matière organique.
•La disponibilité des nutriments augmente souvent remarquablement mais de façon
éphémère. L’azote organique (indisponible, souvent presque égal à l’azote total) se
volatilise en partie et l’autre partie minéralise sous forme d’ammonium, forme utile pour
les organismes vivants. L’ammonium est destiné à être transformé biochimiquement en
nitrate, qui est rapidement lessivé si non assimilé par les cellules. La disponibilité de
l’azote baisse en dessous des niveaux de pré-feu en quelques années. Le phosphore
organique minéralise en orthophosphate et la perte au travers de la volatilisation est
négligeable. Les orthophosphates ne sont pas lessivés mais s’ils ne sont pas rapidement
assimilés, ils précipitent sous forme minérale peu assimilable. Calcium, magnésium et
potassium souvent augmentent remarquablement mais éphémèrement.
•La capacité d’échange diminue proportionnellement à la perte de matière organique.

La forêt amazonienne brésilienne. Chapitre I.1
19
•La saturation en base augmente en conséquence de la libération dominante des bases
de la matière organique brûlée.
I.1.4.c Modifications des propriétés biologiques du sol dans les cultures sur brûlis.
•La biomasse microbienne diminue. Le retour au niveau du pré-feu dépend
principalement de la rapidité des plantes à recoloniser le milieu.
•La composition de la communauté microbienne change comme conséquence de l’effet
sélectif du feu sur quelques groupes de microorganismes et les modifications imposées à
la végétation. En général les champignons diminuent plus que les bactéries.
•La biomasse des invertébrés du sol diminue mais moins que celle des microorganismes
en raison de leur plus grande mobilité.
•La composition des communautés de macrofaune du sol change. Le temps de retour aux
assemblages d’avant feu diffère beaucoup en fonction des différents phylas.

Les invertébrés et le fonctionnement du sol. Chapitre I.2
20
I.2 LES INVERTEBRES ET LE FONCTIONNEMENT DU SOL
I.2.1 Classification de la faune du sol.
Aujourd’hui de très nombreuses études s’intéressent au rôle des communautés
d’invertébrés dans les processus physico-chimiques et biologiques du sol (Lal, 1988 ;
Lobry De Bruyn and Conacher, 1990 ; Kuperman, 1996 ; Lavelle, 1996 ; Fragoso et al.,
1997 ; Brussaard, 1998 ; Decaëns et al., 1999a ; Cragg and Bardgett, 2001 ; Sarr et al.,
2001 ; Folgarait et al., 2003). En dehors de la classification zoologique, les communautés
d’invertébrés du sol peuvent être divisées en trois grands groupes fonctionnels basés sur
la taille :
Moins de 0.2mm
Microfaune Protozoaires, Nématodes, Rotifères
De 0.2 à 4mm Mésofaune Enchytréïdes, Collemboles, Protoures,
Diploures, Acariens
De 4 à 100mm Macrofaune Mollusques, Lombricidés, Myriapodes, Insectes,
Isopodes, Arachnides
Plus de 100mm
Mégafaune Rongeurs, Insectivores terricoles
Tableau 1 :Classement de la faune par taille (critère théorique) (Bachelier, 1978).
Un classement selon les régimes alimentaires montre également une grande
diversité taxonomique et écologique de ces communautés.
La macrofaune du sol est très diversifiée et très sensible à toute modification du
milieu, ses divers composants répondent différemment suivant leur adaptabilité (Lavelle
et al., 1994).

Les invertébrés et le fonctionnement du sol. Chapitre I.2
21
Prédateurs de surface
•Coléoptères : Staphylinidés, Carabidés
•Hyménoptères : Fourmis
•Myriapodes : Chilopodes
•Arachnides : Araignées, pseudoscorpions
Carnivores
Prédateurs endogés •Larves de coléoptères
Feuilles vivantes •Chenilles : Lépidoptères
Phytophages Racines vivantes
•Larves : Diptères (tipules), Coléoptères
(élatéridés Curculionidés), Homoptères
Phytosaprophages •Lombricidés, Enchytréides, Diplopodes,
Isopodes, Larves de Diptères
Xylophages •Larves de diptères
Coprophages •Coléoptères : scarabées
•Larves de Diptères
Saprophages
Nécrophages •Coléoptères
•Larves de Diptères
Polyphages Pas de spécialité •Blattoptères
Tableau 2 : Classement de la faune par régime alimentaire (critère écologique).
Trois groupes principaux de vers de terre peuvent être reconnus :
•Les épigés vivent horizontalement à la surface du sol dans la litière. Ils
consomment les débris végétaux de la litière participant ainsi à leur décomposition. Ce
sont des vers pigmentés et de petite taille. Ils ne sont pas capables de creuser le sol, ils
n’ont par conséquent pas d’effet sur la structure du sol. Leurs turricules s’accumulent à la
surface du sol.
•Les anéciques présents dans des galeries verticales sont également acteurs de la
décomposition de la litière en l’enfouissant dans le sol. Ils modifient la structure du sol.
•Les endogés occupent l’horizon minéral profond du sol et consomment la matière
organique humifiée (Lavelle, 1983). On peut distinguer trois catégories : les
polyhumiques (ingèrent le sol riche en matière organique) ; les mésohumiques (peu
sélectifs quant à la quantité de matière organique) ; les oligohumiques (consomment un
sol pauvre en matière organique). Dans les milieux tropicaux, les populations d’annélides
sont dominées par les vers de types endogés mésohumiques et oligohumiques.

Les invertébrés et le fonctionnement du sol. Chapitre I.2
22
Les termites sont des insectes sociaux appartenant à l’ordre des isoptères. Mieux
adaptés à la sécheresse que les annélides, ils utilisent efficacement le bois et les résidus
végétaux pauvres en nutriments (cellulose). Ces derniers constituent le groupe dominant
sous les tropiques dans les milieux naturels ou les systèmes agricoles dégradés (Decaëns et
al., 1994). Selon les différents régimes alimentaires, quatre groupes peuvent être
distingués : (Grassé, 1984b)
-les xylophages qui vivent plus ou moins uniquement dans le bois ;
-les champignonnistes qui consomment du bois, des feuilles et des herbes et vivent
en symbiose avec des champignons dans des chambres appelées meules à
champignon ;
-les fourrageurs qui consomment des herbes et de la litière ;
-les humivores se nourrissent d’humus et sont parfois géophages.
Les coléoptères, myriapodes, isopodes et de nombreux autres groupes d’arthropodes
sont associés à la présence d’une litière à la surface du sol et sont ainsi plus caractéristiques
des forêts.
I.2.2 Le modèle hiérarchique, les ingénieurs de l’écosystème et leurs structures
biogéniques.
Le fonctionnement du sol est le résultat d’interactions complexes entre les
facteurs physiques, chimiques et biologiques.
Un modèle hiérarchique général (figure 2) a été proposé pour décrire l’important
potentiel de ces facteurs comme déterminant des processus du sol. Les organismes du
sol sont séparés en 4 groupes fonctionnels présentés dans un ordre hiérarchique qui
dépend de leur fonction et de la nature des interactions qu’ils développent avec les
autres organismes (Lavelle, 1996).

Les invertébrés et le fonctionnement du sol. Chapitre I.2
23
Milieu
ClimatCouverture pédologique
Activités anthropiques
Pratiques culturales
Biodiversité
Peuplement végétal
Faune
Microflore
Structures Biogéniques
Fonctionnement du sol
Constitution et structure du sol
Dynamique de la Matière organique
Propriétés hydriques et chimiques
Evolution du sol
Milieu
ClimatCouverture pédologique
Activités anthropiques
Pratiques culturales
Biodiversité
Peuplement végétal
Faune
Microflore
Structures Biogéniques
Fonctionnement du sol
Constitution et structure du sol
Dynamique de la Matière organique
Propriétés hydriques et chimiques
Evolution du sol
Figure 2 : Place de la faune et de ses structures biogéniques dans le fonctionnement et
l’évolution des sols et des agrosystèmes (d’après (Lavelle, 2000)
Ce modèle est basé sur l’idée que les processus du sol sont déterminés par une suite de
facteurs dont l’effet dépend de l’échelle de temps et d’espace à laquelle ils opèrent. Les
facteurs agissant largement contraignent ceux qui agissent plus localement (Lavelle et
al., 1993). Au niveau inférieur de ce modèle se trouvent les processus de régulation
biologiques issus de l’activité de la faune du sol divisée en micro et macrofaune
•Les microorganismes
Tous les invertébrés du sol ont développé des relations avec les microorganismes. Leurs
relations de prédation ou de mutualisme proviennent de la capacité des
microorganismes, contrairement à la macrofaune, de digérer les nutriments contenus
dans le sol. Cependant leur relative immobilité, les oblige à dépendre de la mise à

Les invertébrés et le fonctionnement du sol. Chapitre I.2
24
disposition de ces nutriments par la macrofaune (« sleeping beauty paradox ») (Lavelle,
1997 ; Brown et al., 2000)
• Les microprédateurs
Ce sont de petits invertébrés, protozoaires ou nématodes de la microfaune, ne
fabriquant aucune structure biogénique.
•Les transformateurs de litière
Ils regroupent des arthropodes de la mésofaune (oligochètes, enchytreïdes) et de
la macrofaune. Ces organismes fabriquent de petites structures (boulettes fécales) qui
sont le lieu d’activation de l’activité des microorganismes.
•Les ingénieurs de l’écosystème
Les ingénieurs de l’écosystème sont des organismes qui modifient directement ou
indirectement l’accès aux ressources des autres espèces en structurant physiquement le
milieu où ils vivent (Jones et al., 1994). D’une autre manière ce sont des organismes qui
peuvent avoir un effet déterminant sur l’abondance et la diversité d’autres organismes
(Nested Biodiversity Hypothesis) (Decaëns et al., 1999b ; Jiménez and Decaëns, 2004).
Ils regroupent notamment de gros arthropodes annélides (vers de terre) et des
insectes sociaux isoptères et hyménoptères (termites et fourmis).
Ces organismes ont souvent développé des relations de mutualisme interne avec les
microorganismes du sol (Lavelle et al., 1997). Ils se caractérisent par leur capacité à
fabriquer des grosses structures organo-minérales appelées structures biogéniques qui
constituent des habitats pour d’autres espèces appelées domaines fonctionnels. Ces
domaines peuvent être épigés ou endogés. Ces structures peuvent persister durant des
périodes supérieures à celles de la vie des invertébrés qui les produisent.
L’accumulation de ces structures crée des domaines fonctionnels présentant des
propriétés physiques et de dynamique de la matière organique particulières (Brown et
al., 2000 ; Lavelle, 2002).

Les invertébrés et le fonctionnement du sol. Chapitre I.2
25
Plusieurs types de structures biogéniques peuvent être distingués :
• Les nids:
-Les termitières
Elles sont de formes et de structures très variées. Cinq catégories peuvent être
cependant distinguées (Noirot, 1970; Grassé, 1984a) :
-Les nids intégrés dans le bois
-Les nids arboricoles (sur les troncs ou branches).
Ces deux premiers types de nids sont constitués d’excréments et du contenu de la panse
rectale.
-Les nids hypogés (souterrains) peuvent être concentrés ou diffus avec plusieurs
chambres souterraines reliées par d’innombrables galeries.
-Les nids épigés sont très variés, ils peuvent atteindre 13 m de haut, et leur
diamètre à la base, jusqu'à 30 m avec, dans certains cas, une partie hypogée. Les nids
peuvent être compacts et présentent généralement des structures complexes réalisées à
partir de matériaux sélectionnés mêlés à de la salive. Ces structures sont ordinairement
très stables. La structure se dégrade uniquement après la mort de la colonie.
-Les nids inquilins situés à l’intérieur des nids d’autres espèces.
Certains termites produisent également en surface des placages qui couvrent le sol, la
litière, le bois lorsqu’ils sortent de leur nid.
Les différents modes de construction et de nutrition des termites déterminent leur rôle
dans l’écosystème.
-Les fourmilières
La fourmilière est constituée, en général, d’une multitude de galeries et de
chambres souterraines dont la répartition est très variable selon les colonies. Les
fourmilières sont creusées dans le sable, l’argile ou au centre d’une vieille souche
d’arbre. Pour creuser les galeries, elles utilisent leurs mandibules comme pelles et elles
se servent de leurs pattes antérieures pour ratisser le sol et pour confectionner des
boulettes faisant office de mortier.
La fourmilière compte de nombreux étages ayant chacun une fonction spécifique. On
rencontre donc plusieurs types de chambres comme le dépotoir, le magasin, le grenier, la
salle commune, la pouponnière à œuf, larves et nymphes qui sont situées dans la partie
la plus obscure de la fourmilière. Ceci dit la répartition des chambres est très variable, le
plus souvent la répartition des chambres est due aux conditions de température et
d’humidité de leur milieu (en Europe souvent une forêt de conifères). Les matériaux de

Les invertébrés et le fonctionnement du sol. Chapitre I.2
26
déblai accumulés à l’extérieur de la fourmilière forment un monticule en forme de dôme.
Le dôme régule donc la température ainsi que l’humidité. La profondeur du nid est très
variable, elle est généralement située entre 30 et 40 cm.
Les fourmis arboricoles bâtissent des nids de feuilles au sommet des arbres. D’autres
espèces de fourmis (comme les fourmis Magnans) constituent leur abri à l’aide de leur
propre corps.
• Les vides et galeries
Les vides se manifestent souvent sous forme de galeries. Elles peuvent être
subhorizontales ou subverticales (galeries de termites et vers anéciques) à la surface.
Certains vers rebouchent leur galerie par des turricules (vers endogés géophages). Les
galeries réalisées par les vers sont à section cylindrique et les parois sont recouvertes de
mucus. Les galeries de fourmis sont plus anguleuses, plus fines et généralement non
recouvertes de mucus. Celles de termites sont généralement recouvertes d’un enduit
fécal.
• Les Turricules
Les turricules sont les déjections produites par les vers de terre. Ce sont
habituellement des structures de formes arrondies isolées du sol par un fin cortex d’argile
et de colloïdes organiques (Blanchart et al., 1993a). Les turricules peuvent être divisés
en deux catégories (Lee, 1985) :
- les turricules globulaires sont issus de l’accumulation de boulettes fécales de
taille pluricentimétrique. Ce sont des structures très résistantes fabriquées par
des vers dits « compactants ».
- les turricules granulaires sont issus de l’accumulation de petites et fragiles
boulettes fécales. Ils sont produits par des vers dits « décompactants ».
Les vers « décompactants » sont souvent des vers de taille réduite (en comparaison aux
vers « compactants »). Ils peuvent consommer, entre autres, des turricules globulaires.
Au niveau de la digestion du sol par les vers, l’activité microbienne est fortement
stimulée en quelques heures via le mécanisme de digestion mutualiste. Lors de ce
processus, l’eau et le carbone soluble, sous forme de mucus intestinal produit par les
vers stimule la microflore dormante. Cette stimulation de la microflore augmente la
décomposition des formes stables de matière organique du sol ingéré. Dans les turricules
et les parois de galeries, l’abondance des ressources en nutriments pour la microflore

Les invertébrés et le fonctionnement du sol. Chapitre I.2
27
perpétue cette stimulation lors du « priming effect » (Lavelle et al., 1995 ; Fontaine et
al., 2003), augmentant un court temps le taux de minéralisation et la biodisponibilité des
nutriments des plantes. Lorsque les parois des galeries ou les turricules commencent à
sécher et se stabiliser avec l’âge (jours ou semaine), particulièrement pour les vers
compactants, la décomposition de la matière organique, la minéralisation des nutriments
et l’activité microbienne diminuent atteignant des taux souvent plus faibles que le sol
non ingéré en raison de la protection de la biostructure. Martin, en 1991, a également
montré que les turricules de Milsonia anomala présentaient une minéralisation du
carbone accélérée dans les premiers jours mais fortement ralentie au bout de quelques
mois. Finalement aux échelles annuelles (ou pluriannuelles) et des profils de sol, il
apparaît que la drilosphère peut exercer une importante régulation de l’incorporation de
la matière organique et des taux de turnover des stocks de carbone. (Brown et al.,
2000).
Les ingénieurs de l’écosystème se définissent ainsi comme bioturbateurs, Ils sont
impliqués dans la dynamique de la matière organique et de la structure du sol (Lavelle et al.,
1997).
I.2.3 Formation d’un sol ou pédogénèse.
La formation d’un sol s’effectue en trois phases :
- l’altération de la roche mère
- l’enrichissement en matière organique
- les transferts de matières et formation d’horizons bien différenciés
La première phase permet de libérer de petites particules minérales (sables,
limons, argiles) ayant plus ou moins subi des modifications minéralogiques. C’est
l’altération de la roche mère. L’altération biogéochimique des roches se fait par
l’intermédiaire de l’eau, dans un environnement aérobie ou non, en présence ou non de
gaz carbonique et/ou d’acides organiques. Ces altérations physico-chimiques s’effectuent
de 5 manières : l’hydratation, la dissolution, l’oxydation, la réduction et l’hydrolyse. De
plus, de par leurs métabolismes, certains microorganismes dégradent directement des
minéraux (batteries chimiolithoautrophes). Indirectement les organismes participent
également à l’altération des roches en modifiant le milieu (pH, concentration en oxygène,
potentiel redox). Ces processus physiques, chimiques et biologiques produisent un
matériel minéral auquel sera intégrée la matière organique dans la deuxième phase.

Les invertébrés et le fonctionnement du sol. Chapitre I.2
28
La matière organique correspond aux déchets, sécrétion et à la mort des êtres
vivants. L’intégration de la matière organique dans le sol nécessite par conséquent
l’intervention des organismes édaphiques. Suite à son dépôt au sein du substrat minéral,
la matière organique va subir trois types de transformation :
-la minéralisation : processus physiques, chimiques et biologiques menant à la
transformation des constituants organiques en constituants minéraux.
-l’humification : processus biochimiques de néosynthèse des substances organiques.
-l’assimilation par les organismes
La minéralisation dégrade la matière organique fraiche ou celle synthétisée par
l’humification. La dégradation débute par les composants les plus fragiles : glucides,
protéines, acides aminés, lipides, acides nucléiques. Si la dégradation est totale, elle
aboutit à la libération des cations, des anions, de l’eau et du dioxyde de carbone. Ces
substances sont ensuite libérées dans l’atmosphère ou absorbées par les végétaux et les
microorganismes ou fixées sur le complexe adsorbant ou lixiviées. La minéralisation est
plus ou moins rapide en fonction de l’origine de la matière organique. Les molécules
synthétisées lors de l’huméfaction sont plus stables, la dégradation est donc plus lente.
L’humification est un processus de synthèse de matière organique. Cette synthèse
s’effectue selon trois voies :
-l’humification par héritage : les composés les plus résistants sont incorporés
directement au complexe argilo-humique sans avoir été beaucoup modifiés. Ils
constituent l’humine résiduelle
-l’humification par polycondensation : des composés phénoliques simples, issus des
premières étapes de la minéralisation ou de la litière se polycondensent en molécules
d’assez grande taille : les acides humiques voire l’humine d’insolubilisation.
-l’humification par néosynthèse bactérienne : des molécules organiques provenant de
dégradations enzymatiques sont transformées par des microorganismes en
polysaccharides stables.
Suite à la deuxième phase, la matière organique humifiée et liée à la fraction
minérale fine au sein du complexe argilo humique. Ce stade correspond aux sols peu
évolués constitués d’un seul horizon (couche de sol parallèle à la surface plus ou moins
homogène) organo-minéral.
Des transferts de matière vont alors s’effectuer, permettant la constitution d’autres
horizons.

Les invertébrés et le fonctionnement du sol. Chapitre I.2
29
L’eau est le principal vecteur vertical de transfert de matière, cependant la pente,
la perméabilité des roches sous jacente et les « ingénieurs de l’écosystème » jouent
également un rôle important.
I.2.4 Processus du sol affectés par l’activité des ingénieurs
Le sol est le siège d’un nombre important de services écosystémiques essentiels,
induits par les organismes présents, tels la capacité à recycler la matière organique,
maintenir voire accentuer les propriétés physiques comme la porosité et l’agrégation qui
déterminent l’aptitude du sol à résister à l’érosion, conserver l’eau et laisser circuler l’air
(Daily et al., 1997). L’activité des ingénieurs a un effet significatif sur la dynamique de la
matière organique et l’organisation de la structure du sol.
I.2.4.a Effets sur la dynamique de la matière organique.
Les effets des annélides et des termites sur la matière organique sont très contrastés
en fonction de l’échelle spatio-temporelle considérée.
A l’échelle journalière, la minéralisation de la matière organique est fortement accélérée
au niveau du transit intestinal des vers. Leur digestion a pour résultat la fragmentation
des débris végétaux ingérés et la libération d’une quantité significative d’azote et de
phosphore minéral (Barois et al., 1987) lors du « priming effect » (Lavelle, 1997) qui
stimule l’activité microbiologique. La digestion des termites n’entraîne pas de libération
de nutriments. Ils ont cependant développé d’efficaces mécanismes de fixation de
l’azote.
A l’échelle de plusieurs jours, les vers et les termites affectent la dynamique de la
matière organique au travers des structures biogéniques produites. Si, dans les
turricules, la minéralisation ralentit fortement après quelques jours (Martin, 1991 ;
Lavelle et al., 1992b), chez les termites champignonnistes, le potentiel de minéralisation
de l’azote dans la paroi des chambres à champignons est plus élevé que dans le sol
(Abbadie and Lepage, 1989).
A plus long terme, (échelle de quelques années) l’effet des vers et termites demeure plus
incertain. Certains termites humivores accumulent la matière organique indigeste dans
les parois de la termitière sous forme d’éléments organo-minéraux qui résistent à la
décomposition, mais d’autres termites xylophages participent fortement à la

Les invertébrés et le fonctionnement du sol. Chapitre I.2
30
décomposition du bois ; enfin d’autres champignonnistes ne semblent pas intervenir dans
la conservation de la matière organique (Garnier-Sillam, 1987).
I.2.4.b Effets sur la structure physique.
Le premier effet est celui issu de la sélection des particules ingérées. Les ingénieurs
du sol sélectionnent la taille et la qualité des particules organiques et minérales qu’ils
consomment. Ce phénomène est variable en fonction de la taille de l’ingénieur considéré
et du type de sol (Barois, 1992). Il semble évident que dans les meilleurs cas, les vers
sélectionnent les grosses particules organiques et les fines particules minérales. Les
termites également affectent la texture du sol. L’utilisation de fines particules (argile de 0
à 2µm) est généralement préférée à celle des limons et des sables lors de la construction
des termitières. D’autre part, l’activité des vers et des termites affecte fortement la
rugosité de surface, la porosité et l’agrégation.
Ces paramètres influent significativement sur les propriétés hydrauliques du sol et
plus particulièrement l’infiltration de l’eau et la circulation de l’air. Les galeries des
termites et des vers ne représentent que 1% du volume du sol (Lepage, 1979 ; Lee,
1985), mais elles régulent fortement l’infiltration de l’eau. L’infiltration augmente avec le
pourcentage de sol couvert par des turricules et des placages de termites (Casenave and
Valentin, 1988), de nombreuses études ont également mentionné l’effet bénéfique de la
présence de macropores et de galeries (Lee, 1985 ; Lavelle et al., 1992a). Le type de
structure biogénique joue un rôle primordial ; l’apparition d’une croûte à la surface du sol
provenant de l’accumulation de turricules compacts de Pontoscolex corethrurus a déjà
été mise en évidence en Amazonie brésilienne. Cette croûte aurait des propriétés
d’imperméabilisation du sol (Chauvel et al., 1999).
Les vers et les termites participent donc au maintien dans le sol d’une porosité
relativement élevée mais aussi affectent l’agrégation du sol. Si les vers endogés
favorisent la formation de macroagrégats très stables, d’autres au contraire réduisent ces
structures en turricules granulaires empêchant l’accumulation des turricules compacts
(Blanchart et al., 1999). Ainsi une régulation de l’agrégation du sol s’opère. Par ailleurs
les boulettes fécales de certains termites tropicaux sont des micro agrégats
organominéraux qui forment les parois et parfois recouvrent le sol avoisinant des
termitières (Garnier-Sillam, 1987)
L’effet à long terme des structures sur la pédogenèse reste encore peu étudié.

Fertilité et propriétés physico-chimiques du sol. Chapitre I.3
31
I.3 FERTILITE ET PROPRIETES PHYSICO-CHIMIQUES DU SOL.
La fertilité est la résultante d’effets multiples et indépendants caractéristiques du
milieu géographique, ainsi que de l’action de l’homme (techniques culturales). En
fonction de ces deux groupes de facteurs, la fertilité naturelle, liée au premier groupe, se
distingue de la fertilité acquise reliée au second. Il est cependant plus réaliste de
distinguer les facteurs qui relèvent des conditions naturelles (climat, sol), difficilement
modifiables par les agriculteurs, de ceux, au contraire, sur lesquels l’homme agit
aisément.
Selon Duchauffour (1997), la fertilité d’un sol dépend à la fois de ses caractères
pédogénétiques (propriétés morphologiques intrinsèques issues de la pédogenèse) et de
ses caractères fonctionnels (propriétés physico-chimiques).
Parmi les propriétés physiques, la structure du sol joue un rôle important dans la
fertilité. L’état de la structure se manifeste par sa porosité c'est-à-dire le volume relatif
des vides. Ces vides se divisent en deux catégories : macroporosité et microporosité,
c'est-à-dire la capacité en air et la capacité en eau, assurant aux plantes l’alimentation
en eau et la respiration de ses racines. Si la porosité renseigne sur la capacité hydrique
du sol, en terme de « réservoir », la circulation de l’eau dépend du mode d’arrangement
des pores c'est-à-dire de leur tortuosité et de leur connectivité. Une diminution de la
porosité (compaction) est souvent interprétée comme une dégradation du sol et donc
une réduction de sa fertilité.
La constitution physique du sol (texture, quantité de matière organique, teneur en
éléments minéraux grossiers de taille supérieure à 2mm) et sa composition ionique
influent considérablement sur sa stabilité structurale. La teneur en matière organique
humifiée a de multiples conséquences sur le fonctionnement du sol. C, H, N, O sont les
atomes majeurs qui entrent dans la constitution de la matière organique des sols. Elle
résulte du bilan entre les pertes par minéralisation et les apports résultant de la
dégradation de la matière vivante. Les teneurs en C et N doivent être suffisantes afin de
maintenir la fixation biologique de ces éléments par les plantes.
Le carbone constitue l’élément clé de toutes les substances biogéochimiques et se
trouve à la base de tous les cycles biogéochimiques.
Le cycle global du carbone (Figure 3) est constitué de deux cycles intimement liés :

Fertilité et propriétés physico-chimiques du sol. Chapitre I.3
32
Figure 3 : Cycle du carbone (Ramade, 1994)
- le cycle du carbone organique est régulé par la photosynthèse et la respiration. La photosynthèse utilise l'énergie solaire pour synthétiser la matière organique en fixant
le carbone dans des hydrates de carbone. La base de ces molécules grosses et complexes
dont demeure les éléments C, H et O, mais auxquels viennent se joindre d'autres
éléments en faibles quantités comme l'azote (N), le phosphore (P) et/ou le soufre (S).
La respiration est l'inverse de la photosynthèse: à partir de l'oxygène libre O2, elle
transforme toute matière organique en CO2. Une partie de la matière organique est
oxydée par les animaux ou les plantes elles-mêmes; une autre partie se retrouve dans
les sols terrestres ou les sédiments marins. La décomposition se fait sous l'action de
micro-organismes aérobies ou anaérobies (fermentation), de bactéries et de
champignons.

Fertilité et propriétés physico-chimiques du sol. Chapitre I.3
33
Les processus discutés plus haut (photosynthèse, respiration, fermentation) affectent le
cycle du carbone organique, et en particulier l'équilibre du CO2 atmosphérique, sur une
échelle de temps inférieure au siècle. Sur des échelles de temps beaucoup plus longues,
ce sont les processus de nature géologique qui deviennent les contrôles les plus
importants. Il s'agit de processus tels l'enfouissement des matières organiques dans les
sédiments et roches sédimentaires, leur transformation en combustibles fossiles et leur
altération.
-le cycle du carbone inorganique implique le carbone qui est contenu dans le dioxyde
(CO2) et dans les calcaires (CaCO3).
L'altération chimique des roches continentales convertit le CO2 dissout dans les eaux
météoriques (eaux de pluie et des sols) en HCO3- qui est transporté dans les océans par
les eaux de ruissellement. Les organismes combinent ce HCO3- au Ca2+ pour secréter leur
squelette ou leur coquille de CaCO3. Une partie de ce CaCO3 se dissout dans la colonne
d'eau et sur les fonds océaniques; l'autre partie s'accumule sur les planchers océaniques
et est éventuellement enfouie pour former des roches sédimentaires carbonatées. Ces
dernières sont ramenées à la surface après plusieurs dizaines de millions d'années par
les mouvements tectoniques. Une partie du carbone des roches carbonatées est recyclée
dans les magmas de subduction et retournée à l'atmosphère sous forme de CO2 émis par
les volcans.
La fertilité minérale des sols et l’activité biologique sont régies, par l’intermédiaire du
pH, par l’état du complexe absorbant et ses modifications éventuelles. Le complexe
absorbant se définit comme étant l’ensemble des « colloïdes » (composés humiques et
argiles) dotés de charges négatives susceptibles de retenir les cations sous la forme
échangeable, c'est-à-dire pouvant être remplacé par d’autres cations (Duchaufour,
1997).
Les cations échangeables sont en équilibre avec la solution du sol. Lors d’une
modification de la composition de la solution, des cations du complexe passent en
solution (désorption) et sont remplacés par des cations de la solution (adsorption).
Deux grandes catégories de cations se distinguent :
- les générateurs d’acidité : H+ et AL3+
- les basiques (bases échangeables) : Ca2+, Mg2+, K+, Na+

Fertilité et propriétés physico-chimiques du sol. Chapitre I.3
34
Ainsi l’importance des cations basiques n’est pas seulement due à leur rôle nutritif
mais aussi à leur rôle essentiel dans la neutralisation de l’acididté, le maintien de
l’activité biologique et la structuration du sol. Les cations sont absorbés par la plante
sous forme de sels solubles. Au fur et à mesure que le sol se désature, la diffusion est de
plus en plus lente. Ils sont remplacés dans le sol par les cations échangeables. Le
complexe absorbant est, à son tour, progressivement resaturé en cations basiques par
mobilisation des cations provenant de l’altération minérale ou de la minéralisation de la
matière organique.
Par ailleurs, le phosphore assimilable joue également un rôle essentiel pour le
vivant en participant à la fabrication de l’ADN et de l’ARN. Il intervient fortement dans la
nutrition des végétaux. Sa forme assimilable est Ca(H2PO4)2. Contrairement aux cations
basiques, le phosphore en solution est toujours en faible concentration par rapport au
phosphore absorbé. Les réserves de phosphore sont d’une part minérale et d’autre part
organique (Figure 4).
Figure 4 : Cycle du phosphore (Ramade, 1994)

Fertilité et propriétés physico-chimiques du sol. Chapitre I.3
35
Le phosphore est un élément limitant dans plusieurs écosystèmes terrestres, du fait
qu’il n’y a pas de grand réservoir atmosphérique de phosphore comme c’est le cas pour
le carbone. Sa disponibilité est directement liée à l’altération des roches. L’activité
humaine intervient dans le cycle du phosphore par la fabrication des fertilisants. Ajoutés
aux sols en excès, les phosphates sont drainés vers les écosystèmes aquatiques. Une
addition de phosphore dans ces systèmes agit comme fertilisant et génère des problèmes
d’eutrophisation (forte productivité biologique résultant d’un excès de nutriments).
La plus importante fonction de la matière organique dans le sol est la réserve
d’azote et de nutriments, indispensable aux plantes, (Craswell and Lefroy, 2001) qu’elle
constitue. La matière organique est influencée par le couvert végétal mais aussi par le
matériau géologique ; elle dépend aussi de la quantité et de la qualité de l’argile et des
précipitations (Tognon et al., 1998). Elle augmente l’agrégation du sol, la capacité de
rétention en eau, et la disponibilité en Phosphore, en réduisant sa fixation. La matière
organique diminue aussi la toxicité de l’Aluminium et du Magnésium et le lessivage des
nutriments en accroissant les cations échangeables Ca, Mg et K (Baligar and Fageria,
1997).
Enfin, dans les zones tropicales humides, la matière organique influe également
sur les propriétés physiques du sol telles que la charge électrique en étant largement
responsable des charges négatives et donc de la capacité d’échange cationique (CEC)
(Raij, 1969). La physico-chimie du sol est une composante importante de la fertilité. Les
échanges d’ions (K, Na, Ca, Mg, P …) entre les constituants minéraux ou organiques et la
phase liquide définissent la capacité d’échange. Cette capacité d’échange conditionne la
diffusion des éléments nutritifs au contact des racines et donc la nutrition des végétaux.
Des techniques agricoles, permettant de maintenir une bonne fertilité des sols à
faible coût (sans amendement chimique), constitueraient une solution pour les petits
agriculteurs amazoniens, en quête de milieux productifs sans pour autant accélérer la
déforestation et la dégradation des écosystèmes.

Problématique et objectifs. Chapitre I.4
36
I.4 PROBLEMATIQUE ET OBJECTIFS
L’agriculture familiale est actuellement l’un des principaux responsables de la
transformation des systèmes naturels amazoniens. De petits agriculteurs se délocalisent
successivement vers les fronts pionniers de déforestation après avoir vendu leurs
anciennes terres qu’ils avaient déjà déboisées et transformées en pâturages peu
productifs. La sédentarisation de ces petits agriculteurs sur des territoires est une
condition nécessaire à la réduction de la déforestation et à la conservation de la
biodiversité. Cependant la prise en compte des conditions socio-économiques de ces
agriculteurs est nécessaire pour promouvoir des modes de gestion agricole
économiquement viables et durables tout en permettant de maintenir dans les
agrosystèmes une diversité biologique élevée.
Cette étude s’insère dans le projet de l’UMR 137 dont l’objectif de recherche
annoncé est de mieux comprendre les relations entre la biodiversité du sol, qu’elle soit
végétale, animale ou microbiologique et le fonctionnement de celui-ci afin d’identifier des
pratiques agricoles durables et conservatrices des écosystèmes dégradés des zones
tropicales.
De nombreuses études ont d’ores et déjà mis en évidence les bouleversements
des populations de macroinvertébrés suite à la déforestation (Barros et al., 1998 ;
Carvalho and Vasconcelos, 1999 ; Didham, 2001 ; Andresen, 2003 ; Dunn, 2004 ;
Mathieu, 2004). Les espèces « ingénieurs » font partie des espèces les plus sensibles à
ces modifications de l’écosystème. Par exemple, les feux dans les forêts causent un
déclin des densités des populations des espèces de vers (Bhadauria et al., 2000). Ainsi,
considérant, comme nous l’avons vu, l’importance du rôle des ingénieurs de l’écosystème
dans la structuration du milieu, nous avons étudié deux espèces ingénieurs de
l’écosystème : le termite Cornitermes sp. et l’annélide Andiodrilus pachoensis. Au travers
de l’étude des structures biogéniques de surface produites et de leur dégradation, nous
avons apporté des éléments de compréhension du rôle de ces deux espèces dans le
fonctionnement du sol et ses propriétés physico-chimiques.
Pour se faire nous avons répondu à quatre questions :
1) Quelles sont les particularités des deux structures
biogéniques étudiées ?

Problématique et objectifs. Chapitre I.4
37
Type de structure, densité, masse, distribution spatiale, relations avec le milieu
(topographie, couvert végétal, distribution spatiale de la végétation)
2) Quelles sont leurs propriétés physiques et chimiques ?
Composition, densité de mottes, stabilité structurale, matière organique, pH,
séquestration d’éléments chimiques.
3) Comment se dégradent-elles et s’intègrent-elles au sol ?
Les agents de la dégradation, l’influence des pratiques agricoles, les conséquences
sur le fonctionnement du sol
4) Peut-on réaliser une typologie des structures biogéniques à
partir de leurs caractéristiques chimiques, de leur porosité? Ces
typologies sont elles identiques ?
5) Dans la discussion globale, nous avons replacé cette étude dans
le contexte de la modélisation des sols à but prédictif.

Le Site d’étude. Chapitre II
38
II. Le site d’étude.

Caractérisation du site d’étude. Chapitre II.1
39
II.1 CARACTERISATION DU SITE D’ETUDE.
II.1.1 Caractérisation générale.
Notre étude a été réalisée au Brésil, à Benfica, petite communauté de petits et
moyens agriculteurs située dans la municipalité de Itupiranga à 100 km à l’Ouest de
Marabá et à 70 km de la route Transamazonica (Transamazonienne) dans l’état du Parà
(5°16’S ; 49°50’W). Benfica est un Projeto de Assentamento, 1998 (littéralement
« Projet de lotissement») de l’INCRA (Institut National de la Colonisation et de la
Réforme Agraire), (Figure 5).
Cette localité comprend 183 lots répartis sur 124 km² avec une population
estimée aujourd’hui à un millier d’habitants. Le choix de la localité a été effectué par des
chercheurs IRD de l’UMR 137 en collaboration avec les partenaires brésiliens de l’UR64
en Amazonie, de par sa représentativité en terme de milieu biophysique et
principalement pédologique mais aussi de par l’intérêt de travailler au sein d’une
communauté installée relativement récemment. L’avantage d’une telle communauté est
double : a) la forêt occupe encore une partie importante de l’espace, elle constitue le
milieu de référence de notre étude ; b) les lots des agriculteurs étant utilisés depuis peu,
il y est facile de rencontrer différentes phases de l’évolution du paysage.
Le territoire de la communauté d’agriculteurs peut se diviser en deux ensembles
en fonction de son histoire (époque d’occupation et moyen d’acquisition) :
- L’ensemble I correspond à une fazenda peu explorée (« Fazenda Santa
Isabel ») occupée à partir de 1994 par des familles en quête de terre. Elle regroupe 134
familles qui exploitent des lots d’environ 50 hectares (parfois 2 ou 3 par famille). Elle
était composée en 2002 en moyenne de 37% de forêt primaire, 15% de forêt
secondaire ; 7% de cultures destinées à l’alimentation de la famille et 41% de pâturage
(Boukari et Elodie, 2002). Notre étude a été réalisée dans l’ensemble I.
- L’ensemble II correspond à une fazenda voisine de celle de Santa Isabel achetée
entre 1990 et 1992 par deux « petits » fazendeiros. Elle présente plus de pâturages et
moins de forêt que dans l’ensemble I.
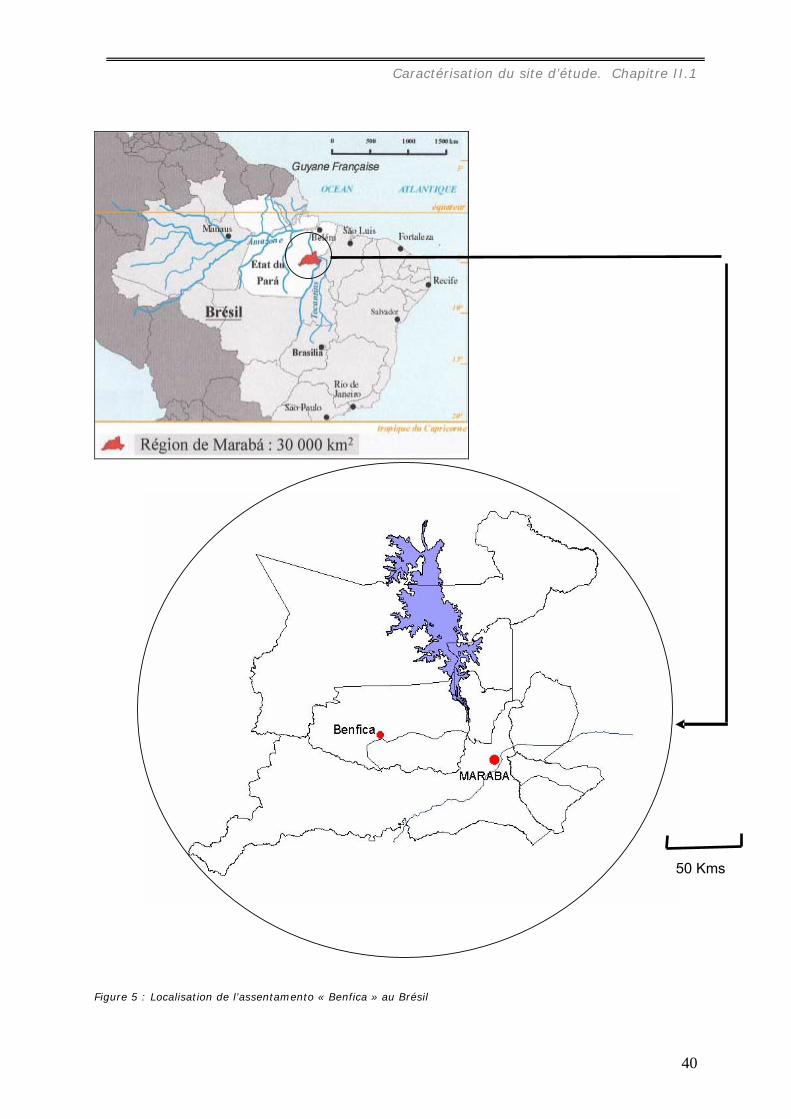
Caractérisation du site d’étude. Chapitre II.1
40
Figure 5 : Localisation de l’assentamento « Benfica » au Brésil
50 Kms
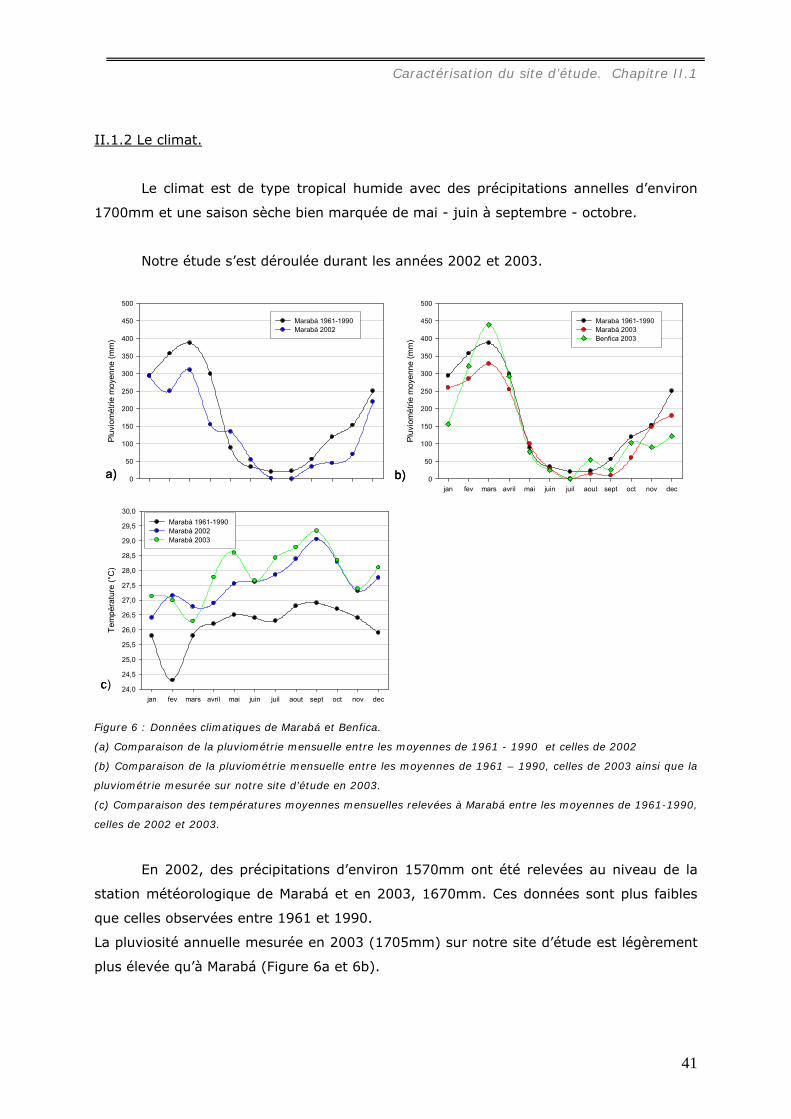
Caractérisation du site d’étude. Chapitre II.1
41
II.1.2 Le climat.
Le climat est de type tropical humide avec des précipitations annelles d’environ
1700mm et une saison sèche bien marquée de mai - juin à septembre - octobre.
Notre étude s’est déroulée durant les années 2002 et 2003.
jan fev mars avril mai juin juil aout sept oct nov dec
Pluv
iom
étrie
moy
enne
(mm
)
0
50
100
150
200
250
300
350
400
450
500
Marabà 1961-1990 Marabà 2002
jan fev mars avril mai juin juil aout sept oct nov dec
Pluv
iom
étrie
moy
enne
(mm
)
0
50
100
150
200
250
300
350
400
450
500
Marabà 1961-1990 Marabà 2003 Benfica 2003
jan fev mars avril mai juin juil aout sept oct nov dec
Tem
péra
ture
(°C
)
24,0
24,5
25,0
25,5
26,0
26,5
27,0
27,5
28,0
28,5
29,0
29,5
30,0Marabà 1961-1990 Marabà 2002 Marabà 2003
a) b)
c)
jan fev mars avril mai juin juil aout sept oct nov dec
Pluv
iom
étrie
moy
enne
(mm
)
0
50
100
150
200
250
300
350
400
450
500
Marabà 1961-1990 Marabà 2002
jan fev mars avril mai juin juil aout sept oct nov dec
Pluv
iom
étrie
moy
enne
(mm
)
0
50
100
150
200
250
300
350
400
450
500
Marabà 1961-1990 Marabà 2003 Benfica 2003
jan fev mars avril mai juin juil aout sept oct nov dec
Tem
péra
ture
(°C
)
24,0
24,5
25,0
25,5
26,0
26,5
27,0
27,5
28,0
28,5
29,0
29,5
30,0Marabà 1961-1990 Marabà 2002 Marabà 2003
a) b)
c)jan fev mars avril mai juin juil aout sept oct nov dec
Tem
péra
ture
(°C
)
24,0
24,5
25,0
25,5
26,0
26,5
27,0
27,5
28,0
28,5
29,0
29,5
30,0Marabà 1961-1990 Marabà 2002 Marabà 2003
a) b)
c)
Figure 6 : Données climatiques de Marabá et Benfica.
(a) Comparaison de la pluviométrie mensuelle entre les moyennes de 1961 - 1990 et celles de 2002
(b) Comparaison de la pluviométrie mensuelle entre les moyennes de 1961 – 1990, celles de 2003 ainsi que la
pluviométrie mesurée sur notre site d’étude en 2003.
(c) Comparaison des températures moyennes mensuelles relevées à Marabá entre les moyennes de 1961-1990,
celles de 2002 et 2003.
En 2002, des précipitations d’environ 1570mm ont été relevées au niveau de la
station météorologique de Marabá et en 2003, 1670mm. Ces données sont plus faibles
que celles observées entre 1961 et 1990.
La pluviosité annuelle mesurée en 2003 (1705mm) sur notre site d’étude est légèrement
plus élevée qu’à Marabá (Figure 6a et 6b).

Caractérisation du site d’étude. Chapitre II.1
42
L’étude des températures a mis en évidence une augmentation significative des
températures en 2002 et 2003 par rapport aux moyennes obtenues entre 1961 et 1990
(figure 4c). La température moyenne mensuelle maximale atteinte est de plus de 37°C
en septembre 2002 et mai 2003, tandis que la température minimale fut d’environ 21°C
en février 2002 et 20°C en octobre 2003. (Données d’après INMET, 2006).
II.1.3 Mode de gestion et pratiques agricoles
L’agriculture familiale est actuellement largement responsable de la
transformation de la forêt amazonienne brésilienne. Depuis les années 70, dans la région
de Marabá, de nombreuses familles de petits agriculteurs venant de toutes les régions du
Brésil, se sont installées sur ces terres nouvelles des fronts pionniers de déforestation,
profitant de nouvelles voies d’accès telle la Transamazonica. Continuant un cycle
traditionnel, ils vendent les terres qu’ils défrichent et sur lesquelles ils installent des
pâturages à de grands propriétaires de fazendas spécialisés dans l’élevage bovin
(Albadalejo and Tulet, 1996 ; Reynal et al., 1997).
Les modes de gestion des petits agriculteurs de la zone d’étude sont légèrement
diversifiés même si la grande majorité se destine à l’élevage bovin ou du moins à mettre
en place un pâturage puis le revendre aux voisins qui désirent développer leur élevage
(Biri Kassoum and Maître d'Hôtel, 2002). Ainsi malgré les recommandations
gouvernementales incitant à la sédentarisation des agriculteurs, une dynamique rapide
de transformation des forêts en pâturages est maintenue.
Deux systèmes agro-pastoraux coexistent à Benfica : un système agricole après
brûlis et un système d’élevage extensif de bovins. Ces deux systèmes sont souvent
utilisés de manière complémentaire.
La valorisation du lot commence par la déforestation (Photo 1) qui s’effectue en
trois étapes. Dans un premier temps, un déboisement du sous-bois est effectué ; s’ensuit
la coupe proprement dite des arbres de diamètre supérieur à 10 cm. Puis le chablis est
brûlé après une vingtaine de jours de séchage juste avant la période des pluies. Cette
dernière étape permet de libérer des nutriments dans le sol à travers les cendres et
d’éliminer une partie des semences restantes enfouies. Le riz pluvial de cycle court est la
culture annuelle pratiquée après le brûlis de la forêt primaire ou secondaire. Ceci
constitue la production agricole essentielle de consommation et de vente à « Benfica ».
Suivent les cultures de manioc, de maïs et de pois. Les cultures annuelles ne reçoivent

Caractérisation du site d’étude. Chapitre II.1
43
pas d’intrants et sont réalisées sur un sol non travaillé et sans traitement phytosanitaire.
Quelques semaines après la semence du riz, les végétaux natifs sont ôtés par bêchage.
L’autre système, d’élevage bovin, est également amplement répandu à
« Benfica ». Ce système agricole passe par la création de pâturages. Cependant, pour
certains exploitants, la mise en pâturage constitue également un moyen d’occuper le lot
et de valoriser la terre. En 2003, grâce aux crédits du PRONAF, toutes les familles étaient
sur le point de posséder au moins 5 têtes et 25% en détenait de 10 à 50. Dans le but
d’améliorer la qualité des pâturages (lutte contre les feux involontaires, les plantes
invasives…), les agriculteurs ont développé de réels plans de gestion. Ils utilisent
différentes espèces de graminées fourragères adaptées à la variabilité du milieu physique
et les plantent de différentes manières en fonction de l’historique des parcelles et des
conditions financières. Plusieurs interventions ensuite telle la coupe manuelle des
adventices et les feux volontaires (Photo 2) sont pratiquées pour conserver une relative
qualité du pâturage. Ainsi les pratiques utilisées par un agriculteur dépendent de ses
connaissances techniques mais aussi de sa situation socio-économique.
II.1.4 Les parcelles étudiées
Les parcelles que nous avons choisies d’étudier sont essentiellement situées dans
les lots de 5 petits agriculteurs caractéristiques de « Benfica » de par le mode de gestion
de leur lot. Un lot représente le terrain alloué par l’INCRA à un agriculteur pour faire de
l’élevage ou cultiver le sol. Un lot est composé d’un pourcentage variable de forêt.
Dans 3 lots une forêt et un pâturage ont été choisis. Deux pâturages, une rizière, une
forêt et une jachère ont également été choisis dans d’autres lots à proximité. Ainsi notre
étude s’est portée sur 11 parcelles et s’est principalement concentrée sur la comparaison
de deux écosystèmes : forêt et pâturages (Tableau 3). Afin de vérifier la ressemblance
de nos répétitions de pâturage et de forêt, le couvert végétal a été examiné. Aucune
différence entre les pâturages n’a été observée ; il s’agit de pâturages de Brachiaria
brisantha âgés de 5 à 6 ans. La disposition des touffes est très irrégulière laissant entre
chaque pied, des espaces de sol nu « inter touffe » (Photo 3).
La forêt F01 a révélé une densité en arbres et troncs de diamètre supérieur à 10 cm,
plus faible (0,34 / m²) que dans les autres forêts (0,82 / m²). Considérant que les gros
arbres avaient été coupés, et que l’intensité des activités humaines dans cette forêt est
importante, cette forêt a été dans cette étude considérée comme une forêt dégradée
(Figure 7).

Caractérisation du site d’étude. Chapitre II.1
44
Agriculteur
Parcelle Pâturage Forêt Jachère Rizière
Sr Amiute x
Sr Cicéro x
Sr Francisco x
Sr Josuel x
Sr Léonine x x
Sr Mélète x x
Sr Osano x x x
Tableau 3 : Répartition des parcelles étudiées en fonction du couvert végétal et de l’exploitant.
Photo 1 : Déforestation (2003)
Photo 2 : Feu de maintien d’un pâturage (2003)

Caractérisation du site d’étude. Chapitre II.1
45
Photo 3 : Disposition clairsemée des touffes de Brachiaria brisantha
Légende
Forêt
Pâturage
Jachère
Rizière
Bas fond
Parcelles étudiées
Légende
Forêt
Pâturage
Jachère
Rizière
Bas fond
Parcelles étudiées
Figure 7 : Disposition des parcelles étudiées dans « l’assentamento » de Benfica en 2002 d’après (Mathieu,
2004).
Le substrat géologique, l’organisation de la couverture pédologique et les
caractéristiques physiques et chimiques des sols du d’étude seront détaillés dans un
prochain chapitre (chapitre IV).
N
500m

La macrofaune du sol. Chapitre II.2
46
II.2 LA MACROFAUNE DU SOL (Mathieu, 2004)
II.2.1 Les populations de Macroinvertébrés de « Benfica ».
L’étude au niveau du paysage des communautés d’invertébrés n’a pas mise en
évidence un patron initial d’organisation spatiale entre les parcelles. La composition des
communautés est plus influencée par le type d’occupation du sol que par la faune des
parcelles voisines.
La densité de la faune est la plus forte sous forêt puis elle diminue
progressivement des jachères aux pâturages puis dans les rizières. La richesse spécifique
est également plus élevée dans les milieux fermés type forêt. Cette tendance est très
forte pour la faune de litière (araignée, diplopodes…). Dès la première culture de riz
après brûlis, la richesse spécifique s’effondre. Les pâturages apparaissent alors plus
favorables à la recolonisation du sol. Peu d’espèces sont abondantes dans tous les
milieux donc généralistes.
De très nombreuses espèces sont indicatrices des milieux fermés (spécificité +
fidélité) et peu des milieux ouverts. Sans doute la majorité des espèces rencontrées en
pâturages sont des espèces forestières tentant de coloniser le nouveau milieu. Les
règnes des coléoptères, fourmis et araignées sont les plus diversifiés. Les groupes les
plus abondants sont les termites (266 ind.m-2), fourmis (192 ind.m-2) et vers de terre
(52 ind.m-2). Les termites et fourmis sont plus diversifiés en forêt que dans les autres
milieux sans doute en raison des nombreuses espèces de termites ayant développées des
relations symbiotiques avec la végétation.
La recolonisation des pâturages ne peut être effectuée que par des espèces de
termites qui peuvent se développer sans présence d’arbres. Une espèce de Cornitermes
sp. s’est bien adaptée aux pâturages. La diversité des vers est également très forte avec
près de 15 espèces collectées. La densité moyenne de ce groupe varie peu. Les
pâturages se caractérisent cependant par l’abondance d’une espèce de ver endogé du
genre Andiodrilus pachoensis. En forêt une espèce anécique et une épigée se rencontrent
préférentiellement.
Dans les différents pâturages étudiés, un effet significatif du type de sol a été
observé. La densité et la richesse spécifique des invertébrés sont supérieures dans les
ferralsols profonds de haut de pente.

La macrofaune du sol. Chapitre II.2
47
Enfin la distribution de la faune n’est pas uniforme au sein des pâturages : la
richesse et la diversité spécifique des invertébrés sont plus élevées sous les troncs des
arbres morts et auprès des touffes d’herbacé que sous le sol nu des entre touffes. Ils
existent donc un effet de microhabitat. (Mathieu, 2004)
II.2.2 Choix des structures biogéniques étudiées.
Une observation visuelle préalable des structures a été réalisée dans les différents
milieux. De nombreuses structures, de tailles et de formes différentes coexistent. Nous
avons répertorié 16 structures biogéniques de surface réparties quelle que soit la
couverture végétale et/ou le type de sol. Ce chiffre ne reflète pas la diversité des
structures biogéniques. Il est très probable que les structures biogéniques déposées et
observables à la surface du sol ne représentent qu’un faible pourcentage de l’ensemble
des structures biogéniques produites. Il est ainsi difficile d’appréhender l’ensemble des
structures biogéniques produites par la faune du sol qui jouent un rôle significatif dans
son fonctionnement global. Notre étude portant sur l’influence des structures
biogéniques sur le fonctionnement du sol, et considérant les résultats obtenus par l’étude
des populations des macroinvertébrés (densité et diversité), nous avons formulé
l’hypothèse que les structures biogéniques de surface, les plus nombreuses, jouaient un
rôle prépondérant.
Notre choix s’est porté sur les deux structures majoritaires:
- Termitières de Cornitermes sp.
- Turricules de Andiodrilus pachoensis

Les méthodes d’analyses. Chapitre III
48
III. Les Méthodes d’Analyses
Dans ce chapitre, les analyses statistiques employées et les analyses physiques
et chimiques réalisées en laboratoire sont détaillées. Les protocoles expérimentaux
particuliers aux deux espèces étudiées seront développés dans les prochains chapitres.

Les analyses statistiques. Chapitre III.1
49
III.1 ANALYSES STATISTIQUES.
III.1.1 Quelques définitions (Krebs, 1998).
Les termes «distribution» et «population» sont utilisés à la fois par les statisticiens
et les écologues, mais avec des significations différentes.
Une distribution est en statistique, une distribution de fréquences alors qu’en
écologie, cela correspond à la répartition d’individus dans l’espace géographique.
Une population est en statistique, l’ensemble des individus étudiés et en écologie,
l’ensemble des individus appartenant à une même espèce et occupant un même
écosystème.
En écologie, il existe trois types de distributions spatiales d’éléments ponctuels dans un
espace géographique : aléatoire, agrégée, ou régulière (Figure 8). Il est possible de
décrire différents degrés d’agrégation.
Une question simple nous renseigne sur le type de distribution spatiale : étant
donné la localisation d’un individu, quelle est la probabilité qu’un autre individu se trouve
à proximité ? Il y a trois possibilités :
1. probabilité non affectée → distribution aléatoire
2. probabilité augmentée → distribution agrégée
3. probabilité réduite → distribution uniforme
Figure 8 : Trois types possibles de distribution spatiale d’organismes dans une population

Les analyses statistiques. Chapitre III.1
50
III.1.2 Analyses statistiques classiques.
Des tests statistiques classiques paramétriques ou non (test T, test de
kruskal et Wallis, test de Mann et Witney, anova,…) et des analyses multivariées (ex :
ACP) ont été appliqués aux données (Scherrer, 1984 ; Legendre and Legendre, 1998).
III.1.3 Analyses statistiques spatiales.
La structure spatiale d’un écosystème c’est-à-dire la manière dont ses
composantes sont organisées dans l’espace, joue un rôle primordial dans son
fonctionnement. La réalité écologique présente de nombreuses sources d’hétérogénéité
qui peuvent influer sur le résultat de l’analyse spatiale. Le concept d’analyse spatiale est
complexe en raison de l’utilisation des deux dimensions du milieu considéré comme un
plan horizontal.
III.1.3.a Les processus de points.
Un processus de points P est un processus aléatoire c’est-à-dire un objet
mathématique similaire à une variable aléatoire dont les réalisations sont des patrons de
points. Les propriétés d’un tel processus définissent des contraintes sur ses réalisations
(densité, voisinage, structure…). Un patron de points peut être hétérogène (a),
anisotropique (une direction privilégiée) (b) ou encore homogène isotropique (patron de
poisson ou encore aléatoire) (c) (Figure 9)
De nombreuses méthodes d’analyse de la structure spatiale peuvent être
rencontrées dans la littérature (Ripley, 1981 ; Diggle, 1983 ; Cressie, 1993). Chaque
méthode nécessite une forme de données particulière : nombre d’individus par cadrat,
distance entre les points, distance au plus proche voisin ou encore cartographie complète
de la zone étudiée. Cette dernière méthode a été choisie parce qu’elle a l’avantage de
décrire la structure spatiale à plusieurs échelles simultanément.

Les analyses statistiques. Chapitre III.1
51
Figure 9 : Exemple d’un patron hétérogène (a) (deux régions de densité différentes) ; un patron anisotropique
(b) (une direction privilégiée) ; un patron homogène isotropique (c) (patron de poisson)
III.1.2.b Les Fonctions de Ripley
• Fonction K(r)
La fonction K(r) de (Ripley, 1977, 1981) permet l’analyse de la structure spatiale
des semis de points homogènes, c’est-à-dire dont on suppose (on peut le vérifier si on
dispose de plusieurs réalisations du processus) que la densité est constante. Cependant
dans de nombreux cas, cette structure est inconnue et les fonctions sont estimées à
partir d’une seule réalisation.
Forme originale :
Cette statistique compte le nombre de points d’un même groupe à l’intérieur d’une
certaine distance à partir de chaque point d’un groupe. On peut donc calculer le K de
Ripley pour plusieurs distances, et donc plusieurs échelles spatiales (Figure 10). La
valeur observée du K de Ripley doit être comparée à une certaine valeur théorique
d’absence de structure spatiale c'est-à-dire que l’on doit comparer le nombre trouvé à
chaque distance au nombre attendu sous une supposition de distribution spatiale de
poisson (homogène).
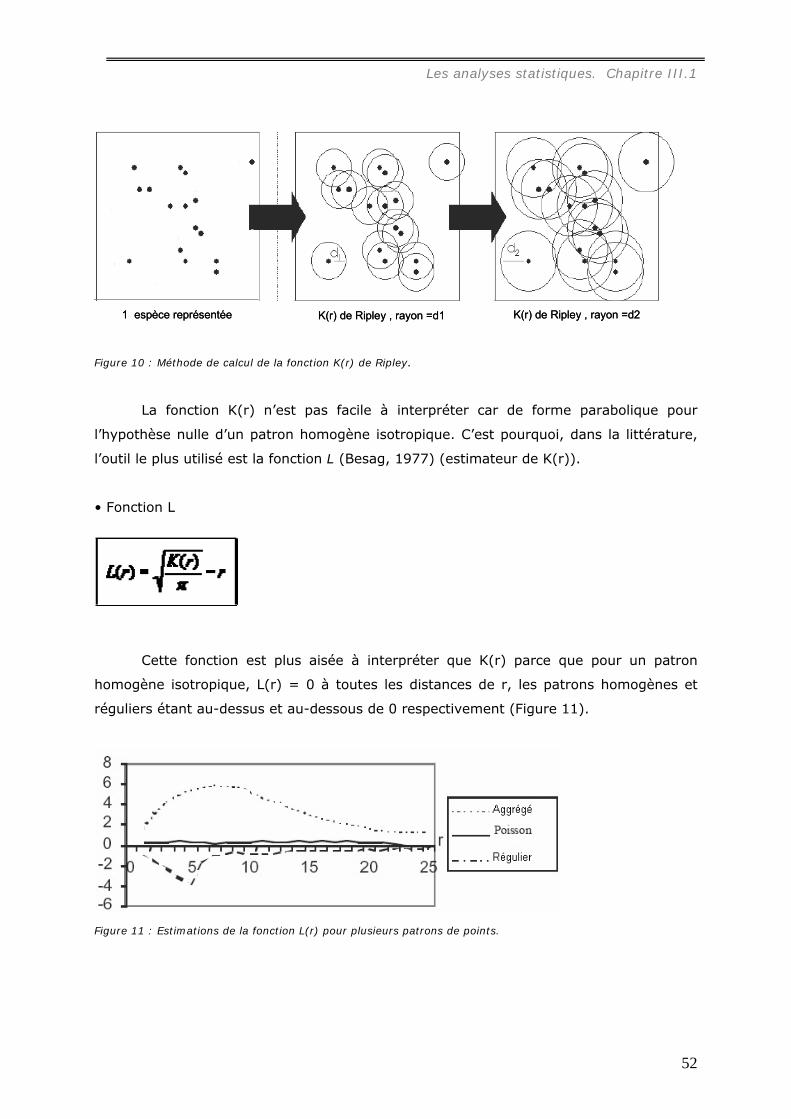
Les analyses statistiques. Chapitre III.1
52
1 espèce représentée K(r) de Ripley , rayon =d1 K(r) de Ripley , rayon =d21 espèce représentée K(r) de Ripley , rayon =d1 K(r) de Ripley , rayon =d2
Figure 10 : Méthode de calcul de la fonction K(r) de Ripley.
La fonction K(r) n’est pas facile à interpréter car de forme parabolique pour
l’hypothèse nulle d’un patron homogène isotropique. C’est pourquoi, dans la littérature,
l’outil le plus utilisé est la fonction L (Besag, 1977) (estimateur de K(r)).
• Fonction L
Cette fonction est plus aisée à interpréter que K(r) parce que pour un patron
homogène isotropique, L(r) = 0 à toutes les distances de r, les patrons homogènes et
réguliers étant au-dessus et au-dessous de 0 respectivement (Figure 11).
Figure 11 : Estimations de la fonction L(r) pour plusieurs patrons de points.
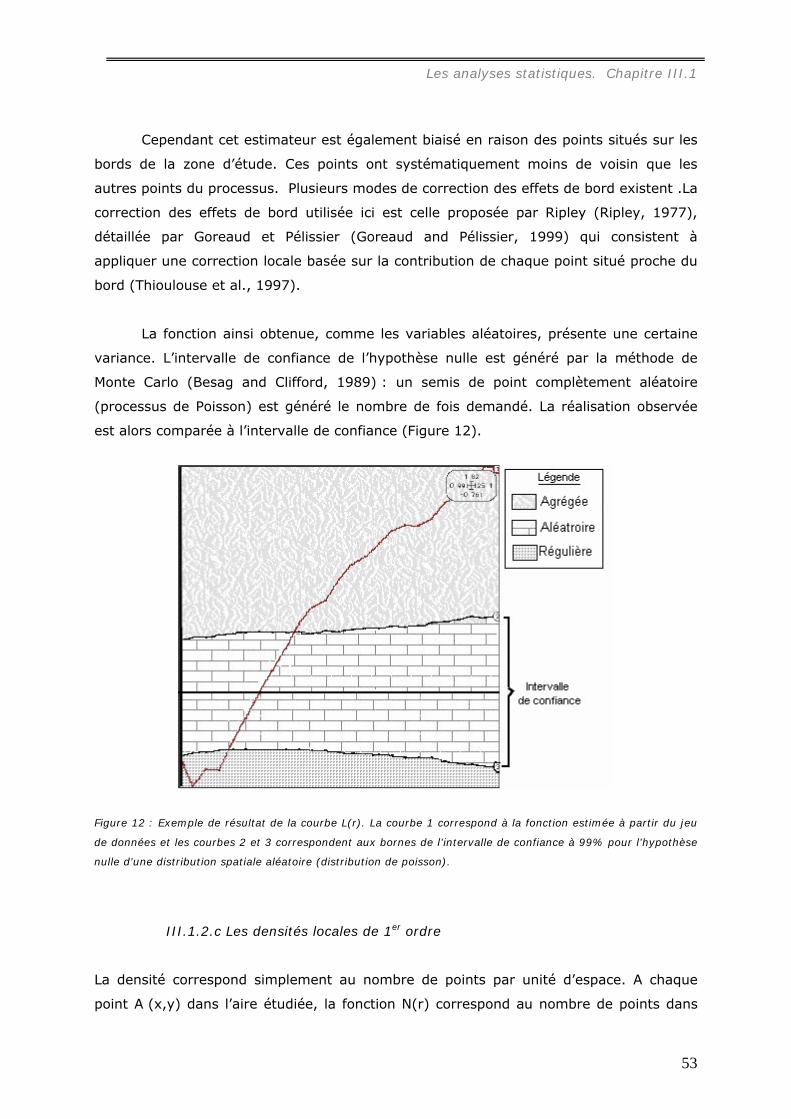
Les analyses statistiques. Chapitre III.1
53
Cependant cet estimateur est également biaisé en raison des points situés sur les
bords de la zone d’étude. Ces points ont systématiquement moins de voisin que les
autres points du processus. Plusieurs modes de correction des effets de bord existent .La
correction des effets de bord utilisée ici est celle proposée par Ripley (Ripley, 1977),
détaillée par Goreaud et Pélissier (Goreaud and Pélissier, 1999) qui consistent à
appliquer une correction locale basée sur la contribution de chaque point situé proche du
bord (Thioulouse et al., 1997).
La fonction ainsi obtenue, comme les variables aléatoires, présente une certaine
variance. L’intervalle de confiance de l’hypothèse nulle est généré par la méthode de
Monte Carlo (Besag and Clifford, 1989) : un semis de point complètement aléatoire
(processus de Poisson) est généré le nombre de fois demandé. La réalisation observée
est alors comparée à l’intervalle de confiance (Figure 12).
Figure 12 : Exemple de résultat de la courbe L(r). La courbe 1 correspond à la fonction estimée à partir du jeu
de données et les courbes 2 et 3 correspondent aux bornes de l’intervalle de confiance à 99% pour l’hypothèse
nulle d’une distribution spatiale aléatoire (distribution de poisson).
III.1.2.c Les densités locales de 1er ordre
La densité correspond simplement au nombre de points par unité d’espace. A chaque
point A (x,y) dans l’aire étudiée, la fonction N(r) correspond au nombre de points dans

Les analyses statistiques. Chapitre III.1
54
un disque de rayon r centré sur A. La fonction de densité locale est alors déduite par :
NA(r) = NA(r) / πr2
La densité locale de 1er ordre peut alors être homogène ou hétérogène.
III.1.2.d Les Analyses intertypes
La fonction K12(r) intertype (Lotwick and Silverman, 1982) est définie comme la
fonction K(r) de Ripley mais pour un patron de points bivariables.
L’interprétation est aussi plus aisée pour la fonction associée L12(r) (Stoyan et al.,
1987). Deux hypothèses peuvent être testées par cette méthode :
•la distribution spatiale de la variable étudiée, en fonction de la seconde, est aléatoire
(distribution de poisson)
•l’indépendance des deux variables.
L’estimateur de la fonction intertype n’est pas biaisé. On représente comme pour
la fonction L(r) de Ripley, la fonction intertype avec son intervalle de confiance
(figure 12).

Les analyses statistiques. Chapitre III.1
55
Figure 13 : Exemple de résultat de la courbe L12(r). La courbe 1 correspond à la fonction estimée pour le jeu de
données initial. Les courbes 2 et 3 correspondent aux bornes de l’intervalle de confiance à 99% de l’hypothèse
nulle de la distribution spatiale aléatoire de l’objet étudié en fonction de la position des points d’une autre
variable qui reste inchangée.
III.1.4 Indice de diversité de Shannon
La diversité indique la manière dont les éléments sont répartis entre les
différentes catégories représentées dans un échantillon. Il s’agit d’un paramètre de
dispersion. Plusieurs indices de diversité ont été proposés mais le plus utilisé est celui de
Shannon (Legendre and Legendre, 1984 ; Scherrer, 1984).
L’indice de Shannon s’écrit : H’ = - Σ ((Ni / N) * log (Ni / N))
i allant de 1 à S, le nombre d’espèces, et log étant de base 2.
Les logiciels statistiques utilisés sont :
-SigmaStat,
-ADE-4 (Thioulouse et al., 1997)
-Module Spatstat (Baddeley and Turner, 2005) utilisé dans R (R Development Core
Team, 2005).

Substrat géologique et couverture pédologique du site d’étude. Chapitre IV
56
III.2 ANALYSES PHYSIQUES ET CHIMIQUES.
III.2.1 Analyses physiques des horizons et des structures biogéniques.
- La texture
La texture influe sur les propriétés physiques d’un sol (perméabilité, aération,
tassement…). Cette connaissance indique la tendance d’un sol. Par exemple, lorsque les
sables grossiers dominent, les sols sont généralement filtrants (forte porosité et absence
de colloïdes), si des sables fins et des limons accompagnent les sables grossiers, le sol se
tasse facilement, il devient battant. Enfin si une forte concentration en argile et en
humus accompagne les sables et les limons, le sol acquiert une structure fragmentée
(Soltner, 1994).
La texture des structures biogéniques et de celle des horizons témoins ont été
comparées (méthode de sédimentation). Ces analyses permettent de classer les
constituants de l’objet d’étude d’après leur grosseur en mesurant le pourcentage de
chaque fraction.
Nous avons utilisé l’horizon témoin (0-30cm) et des structures biogéniques prélevés
dans 3 forêts et 3 pâturages.
-La porosité
Afin d’étudier plus précisément la structure des structures biogéniques, nous avons
évalué leur porosité (volume des vides) et celle du sol. L’examen de la porosité donne
une mesure indirecte de la structure.
1-Densité apparente
Dans un premier temps, nous avons choisi d’étudier la densité apparente (méthode
des cylindres). Il s’agit d’une mesure globale de l’état de compaction du sol. La densité
apparente prend en compte la densité réelle des constituants de l’échantillon et la
manière dont ceux-ci sont organisés. Ainsi l’importance relative des vides entre les
constituants, c'est-à-dire la porosité est (directement) prise en compte dans cette
mesure. La porosité totale à l’échelle de l’horizon est estimée sur le terrain à partir des
mesures de densité apparente.
2-Densité de mottes
L’objectif est de mesurer la densité apparente des agrégats élémentaires qui
correspondent davantage à l’échelle des structures biogéniques. Nous avons donc, dans

Substrat géologique et couverture pédologique du site d’étude. Chapitre IV
57
un second temps, mesuré la densité de mottes ou des agrégats élémentaires (méthode
au pétrole). Le principe de la méthode repose sur la mesure du volume apparent de
l’échantillon, et le pétrole comme liquide d’immersion. Une motte sèche de masse connue
est immergée dans le pétrole, jusqu’à ce que celui-ci remplisse l’espace poral et s’y
maintienne. La différence de masse permet de connaître la quantité de pétrole retenu et
d’en déduire le volume poral.
Cette mesure a été effectuée sur des agrégats obtenus par destruction manuelle des
structures biogéniques et issus des horizons 0-30 cm de profondeur des sols témoins de
2 forêts et 4 pâturages.
-Stabilité structurale
La structure d’un échantillon de sol est l’arrangement spatial des particules
minérales et leur éventuelle liaison par la matière organique, des oxydes ou hydroxydes
de fer et/ou d’aluminium à un moment donné. La stabilité structurale est la solidité de
cet état, sa résistance vis-à-vis de l’action dégradante des facteurs externes, à celle de
l’eau en particulier. La stabilité structurale joue un rôle important dans la fertilité parce
qu’elle influe sur la durabilité des propriétés physiques (aération, circulation de l’eau),
chimiques (échanges chimiques) et biologiques (activité microbiologique) (Mathieu and
Pieltain, 1998).
Plusieurs méthodes ont été mises au point pour mesurer la stabilité structurale. La
méthode de Le Bissonnais et Souder (Le Bissonnais and Souder, 1995) a été choisie.
Cette méthodologie est largement inspirée du test classique de Hénin (Hénin et al.,
1958). Elle est constituée de plusieurs tests :
● Test de désagrégation dans l’eau : L’humectation rapide piège de l’air dans
l’agglomérat qui subit alors une compression. Lorsque la pression est suffisante
l’agglomérat se rompt. C’est un phénomène d’explosion.
● Test de capillarité : L’humectation lente repose sur le fait que l’eau disperse les ciments
argileux et humiques. Il s’agit d’un phénomène de gonflement-dispersion.
● Test de désagrégation dans l’alcool (désagrégation mécanique après prétraitement
dans un liquide miscible à l’eau). Un prétraitement avec un liquide miscible à l’eau
augmente la résistance des particules (Hénin, 1938)
● De manière à essayer de détruire la stabilité des structures biogéniques, nous avons
ajouté un test de désagrégation dans le kérosène. Il s’agit d’une désagrégation
mécanique après prétraitement dans le kérosène liquide non miscible à l’eau. Ce

Substrat géologique et couverture pédologique du site d’étude. Chapitre IV
58
prétraitement est fortement destructeur sauf dans le cas où la matière organique est
abondante. Le kérosène est un mélange d'hydrocarbures contenant des alcanes (CnH2n+2)
de formule chimique allant de C10H22 à C14H30 .Le kérosène est également employé par
les paysans amazoniens pour brûler les termitières.
III.2.2 Analyses de la composition chimique des horizons témoins et des structures
biogéniques.
-Dosage de la matière organique
La matière organique est une source importante d’éléments nutritifs pour les
plantes et la connaissance de sa teneur totale dans le sol renseigne sur sa potentialité
fertilisante. Elle joue également un rôle important dans l’agrégation. La quantité de
matière organique est estimée à partir de la teneur en carbone organique totale dosée
auquel nous appliquons un facteur 2. Le dosage du carbone a été réalisé selon la
méthode Walkley et Black pour le feuilleté de termitière de Cornitermes. Cette méthode
est basée sur l’oxydation du carbone organique par le bichromate de potassium
(K2CR2O7) en milieu acide sulfurique. D’autres dosages de carbone ont été répétés au
moins trois fois dans le sol témoin (0-20 cm) de trois forêts, trois pâturages, et des
structures biogéniques. Ces dosages du carbone ont été réalisés par un auto-analyseur
CHN.
-Dosage de l’acidité d’échange
L’acidité des sols est définie par la concentration en ions H+. L’acidité effective ou
acidité réelle (pH dans H2O) a été mesurée. Elle correspond à la concentration des ions
H+ libres de la solution du sol. Les mesures ont été effectuées au moyen d’un pH-mètre
selon la norme internationale (Afnor, 1994). Trois répétitions d’échantillons de sols
témoins prélevés jusqu’à 25 cm de profondeur dans 2 pâturages et 3 forêts et
d’échantillons de structures biogéniques ont été analysés.
Dans les sols acides, les cations H+ et AL3+ deviennent les principaux cations de la
garniture ionique du complexe absorbant. Ils constituent l’acidité d’échange ou acidité de
réserve. Une comparaison des concentrations en cations Al3+ entres le sol témoin (0-
25cm) (2 pâturages et 3 forêts) et les structures biogéniques a été entreprise (3
répétitions pour chaque site et chaque structure). Ces dosages ont été réalisés par
spectrophotométrie de masse.

Substrat géologique et couverture pédologique du site d’étude. Chapitre IV
59
-Dosage du complexe sorbant
Une comparaison de l’état du complexe absorbant entre les sols témoins et les
structures biogéniques a été effectuée. Les concentrations en cations basiques Ca2+,
Mg2+, K+ ont été dosées par spectrophotométrie de masse. Les mesures ont été répétées
3 fois dans les sols témoin (0-25cm) de 2 forêts, 3 pâturages, et les structures
biogéniques.
Trois valeurs ont enfin été définies pour comparer les sols de forêt, ceux de
pâturage et les structures biogéniques étudiées :
- La capacité d’échange cationique (CEC) correspond à la quantité de cations
adsorbable par le complexe absorbant du sol. Elle provient ainsi des colloïdes qui portent
des charges négatives tels que les minéraux argileux, la matière organique et la silice
colloïdale. L’argile des sols du site d’étude est une kaolinite (Grimaldi, M. communication
personnelle). Le déficit de charge est pratiquement exclusivement dû aux charges
variables. Ces charges proviennent de la rupture du réseau cristallin en bordure des
minéraux argileux.
- La somme des cations échangeables (S). S sera représentée ici par la somme des
concentrations en Ca2+, Mg2+ et K+. Les ions Na+, classiquement inclus dans S, n’ont pas
dans cette étude été dosés. Dans les latossols du site d’étude, leur concentration est
faible, inférieure à 0,1 cmolc.kg-1 et sont négligeables (Grimaldi, M. non publié).
- Le taux de saturation en cations (V). V= S/T% ou T représente la quantité totale de
cations qu’un sol peut adsorber. La saturation en bases désigne la proportion des sites
d’échange de cations du sol qui sont occupés par les différents cations (hydrogène,
calcium, magnésium, potassium). Les surfaces des minéraux et de la matière organique
du sol ont des charges négatives qui attirent et fixent les cations à charge positive. Les
cations qui ont une charge positive (hydrogène, potassium, sodium) se fixent sur un site
à charge négative. Les cations qui possèdent deux charges positives (calcium,
magnésium) occupent deux sites.
-Dosage du Phosphore assimilable
Le phosphore est un élément constitutif essentiel des tissus vivants. Il joue un
rôle important dans la synthèse et le métabolisme des glucides. Son rôle est fondamental
dans la photosynthèse. Les analyses de concentration en phosphore dans les sols ont
pour objectif d’estimer la quantité de phosphore disponible pour les plantes. Nous avons
étudié la concentration en phosphore présente dans les sols témoins de 2 pâturages et
de 3 forêts et dans les structures biogéniques. (extrait avec une solution d’extraction
Mehlich I : HCl 0,05mol L-1 + H2SO4 0,0125mol L-1)

Substrat géologique et couverture pédologique du site d’étude. Chapitre IV
60
IV. Substrat géologique et couverture pédologique du
site d’étude.
Dans ce chapitre, les sols du site d’étude sont présentés au travers de la
description du substrat géologique et de plusieurs fosses pédologiques (3 sous forêts et 3
sous pâturages). L’examen de la texture, de la structure et des propriétés chimiques des
horizons montre les effets de la déforestation sur les sols du site d’étude.

Caractéristiques physico-chimiques des sols témoins. Chapitre IV.2
61
IV.1 SUBSTRAT GEOLOGIQUE ET ORGANISATION DE LA COUVERTURE
PEDOLOGIQUE.
IV.1.1 Substrat géologique
Le paysage de la zone d’étude est constitué d’un ensemble de collines allongées
de taille kilométrique à replats sommitaux et réseaux hydrographiques encaissés. Le
dénivelé est d’environ 50 m (Photo 4).
Ce paysage est développé sur un monzogranite provenant du bouclier brésilien
(Dall’Agnol, non publié)
IV.1.2 Couverture pédologique
La partie sommitale des collines présente un sol classique amazonien ; il s’agit de
ferralsols (WRBSR, 1994) argileux ou Latossolos vermelho amarelo selon la classification
brésilienne des sols (Do Prado, 1996).
Ces sols peuvent présenter une épaisseur variable de 1 m à plus de 3 m. Descendant
le long de la topographie, les ferralsols se transforment en cambisols au niveau du replat
puis en gleysol dans les bas fonds (toposéquence type annexe 1, Grimaldi M.).
Les ferralsols sont issus de l’altération intense de la roche mère. Ils sont caractérisés
par une fraction argileuse dominante peu active (kaolinite) et des oxydes de fer et
d’aluminium (hématite, goethite, gibbsite). Ce sont des sols dont la morphologie, les
propriétés physico-chimiques et minérales sont globalement constantes. Leur texture
peut être variable, plus ou moins argileuse, avec un éventuel horizon plus sableux en
surface. Ils sont généralement poreux, très profonds et peu fertiles avec une
augmentation de la fraction argileuse en profondeur. Ils représentent 40% de la surface
du Brésil (Coelho et al., 2002).
Les structures biogéniques présentent des tailles différentes, de quelques centimètres
pour les turricules à plus de 100 cm de diamètre pour les termitières ; nous avons, par
conséquent, pris en compte pour certaines structures l’échelle de la parcelle et donc le
type de sol alors que pour d’autres, l’étude a été réalisée dans la partie sommitale
(ferralsol) de chaque parcelle. Dans ces zones, le sol y est plus profond et par
conséquent plus propice à l’activité de la macrofaune du sol. De plus, suivant la
technique de déboisement des agriculteurs, les zones de forêt se rencontrent davantage

Caractéristiques physico-chimiques des sols témoins. Chapitre IV.2
62
dans ces parties « hautes », permettant ainsi une meilleure comparaison entre les
pâturages et les forêts.
Photo 4 : Benfica, Modelé du paysage, perspective à partir du pâturage du Sr Osano.
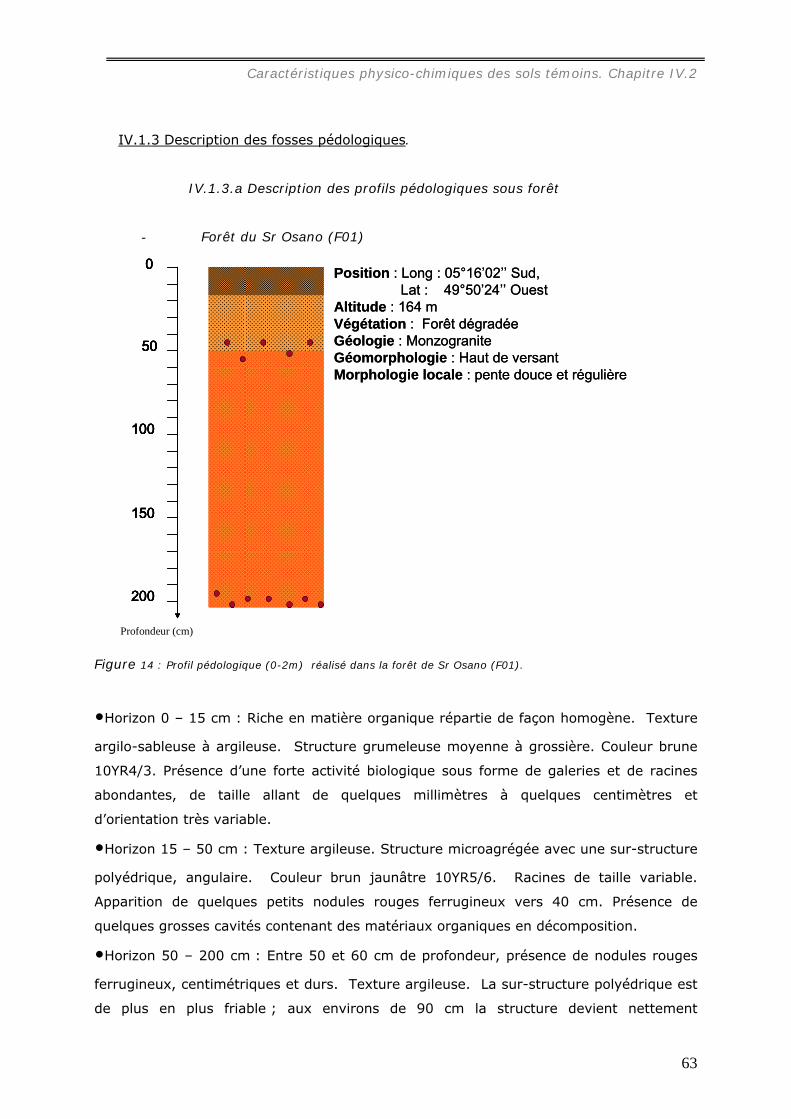
Caractéristiques physico-chimiques des sols témoins. Chapitre IV.2
63
IV.1.3 Description des fosses pédologiques.
IV.1.3.a Description des profils pédologiques sous forêt
- Forêt du Sr Osano (F01)
0
50
100
150
200
Profondeur (cm)
Position : Long : 05°16’02’’ Sud, Lat : 49°50’24’’ Ouest
Altitude : 164 mVégétation : Forêt dégradéeGéologie : MonzograniteGéomorphologie : Haut de versantMorphologie locale : pente douce et régulière
0
50
100
150
200
Profondeur (cm)
0
50
100
150
200
Profondeur (cm)
0
50
100
150
200
0
50
100
150
200
Profondeur (cm)
Position : Long : 05°16’02’’ Sud, Lat : 49°50’24’’ Ouest
Altitude : 164 mVégétation : Forêt dégradéeGéologie : MonzograniteGéomorphologie : Haut de versantMorphologie locale : pente douce et régulière
Figure 14 : Profil pédologique (0-2m) réalisé dans la forêt de Sr Osano (F01).
•Horizon 0 – 15 cm : Riche en matière organique répartie de façon homogène. Texture
argilo-sableuse à argileuse. Structure grumeleuse moyenne à grossière. Couleur brune
10YR4/3. Présence d’une forte activité biologique sous forme de galeries et de racines
abondantes, de taille allant de quelques millimètres à quelques centimètres et
d’orientation très variable.
•Horizon 15 – 50 cm : Texture argileuse. Structure microagrégée avec une sur-structure
polyédrique, angulaire. Couleur brun jaunâtre 10YR5/6. Racines de taille variable.
Apparition de quelques petits nodules rouges ferrugineux vers 40 cm. Présence de
quelques grosses cavités contenant des matériaux organiques en décomposition.
•Horizon 50 – 200 cm : Entre 50 et 60 cm de profondeur, présence de nodules rouges
ferrugineux, centimétriques et durs. Texture argileuse. La sur-structure polyédrique est
de plus en plus friable ; aux environs de 90 cm la structure devient nettement
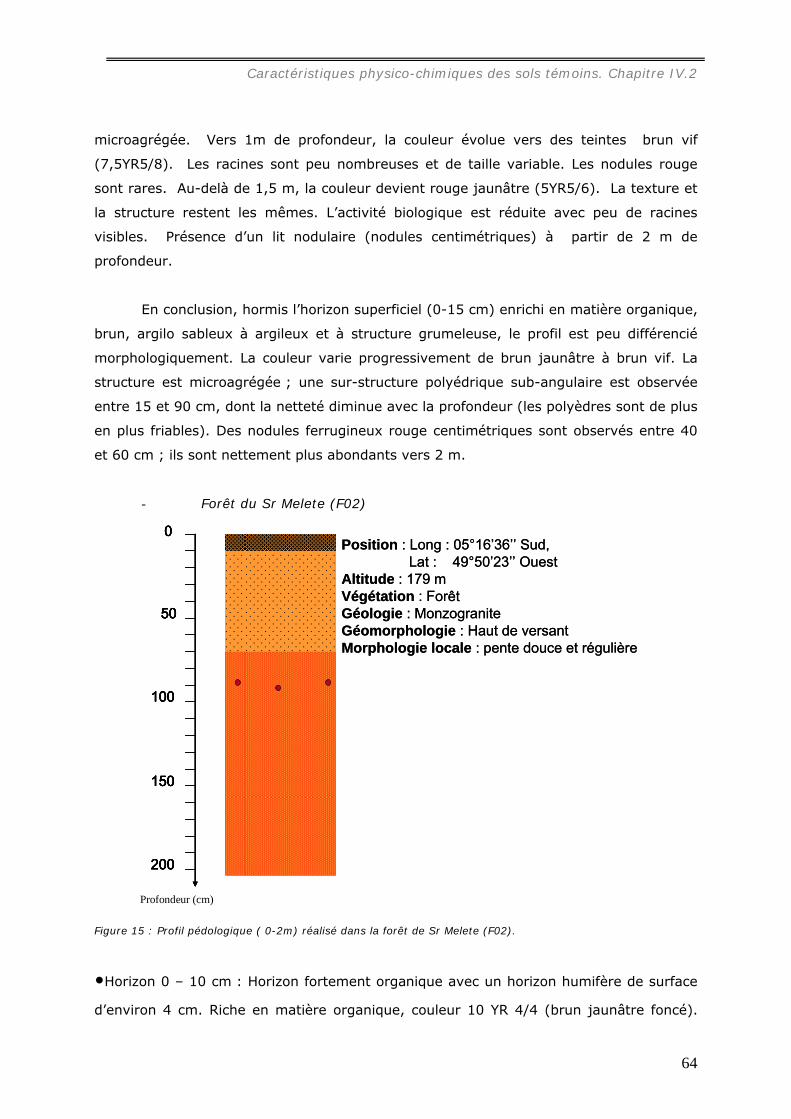
Caractéristiques physico-chimiques des sols témoins. Chapitre IV.2
64
microagrégée. Vers 1m de profondeur, la couleur évolue vers des teintes brun vif
(7,5YR5/8). Les racines sont peu nombreuses et de taille variable. Les nodules rouge
sont rares. Au-delà de 1,5 m, la couleur devient rouge jaunâtre (5YR5/6). La texture et
la structure restent les mêmes. L’activité biologique est réduite avec peu de racines
visibles. Présence d’un lit nodulaire (nodules centimétriques) à partir de 2 m de
profondeur.
En conclusion, hormis l’horizon superficiel (0-15 cm) enrichi en matière organique,
brun, argilo sableux à argileux et à structure grumeleuse, le profil est peu différencié
morphologiquement. La couleur varie progressivement de brun jaunâtre à brun vif. La
structure est microagrégée ; une sur-structure polyédrique sub-angulaire est observée
entre 15 et 90 cm, dont la netteté diminue avec la profondeur (les polyèdres sont de plus
en plus friables). Des nodules ferrugineux rouge centimétriques sont observés entre 40
et 60 cm ; ils sont nettement plus abondants vers 2 m.
- Forêt du Sr Melete (F02)
0
50
100
150
200
Profondeur (cm)
Position : Long : 05°16’36’’ Sud, Lat : 49°50’23’’ Ouest
Altitude : 179 mVégétation : ForêtGéologie : MonzograniteGéomorphologie : Haut de versantMorphologie locale : pente douce et régulière
0
50
100
150
200
Profondeur (cm)
0
50
100
150
200
Profondeur (cm)
0
50
100
150
200
0
50
100
150
200
Profondeur (cm)
Position : Long : 05°16’36’’ Sud, Lat : 49°50’23’’ Ouest
Altitude : 179 mVégétation : ForêtGéologie : MonzograniteGéomorphologie : Haut de versantMorphologie locale : pente douce et régulière
Figure 15 : Profil pédologique ( 0-2m) réalisé dans la forêt de Sr Melete (F02).
•Horizon 0 – 10 cm : Horizon fortement organique avec un horizon humifère de surface
d’environ 4 cm. Riche en matière organique, couleur 10 YR 4/4 (brun jaunâtre foncé).
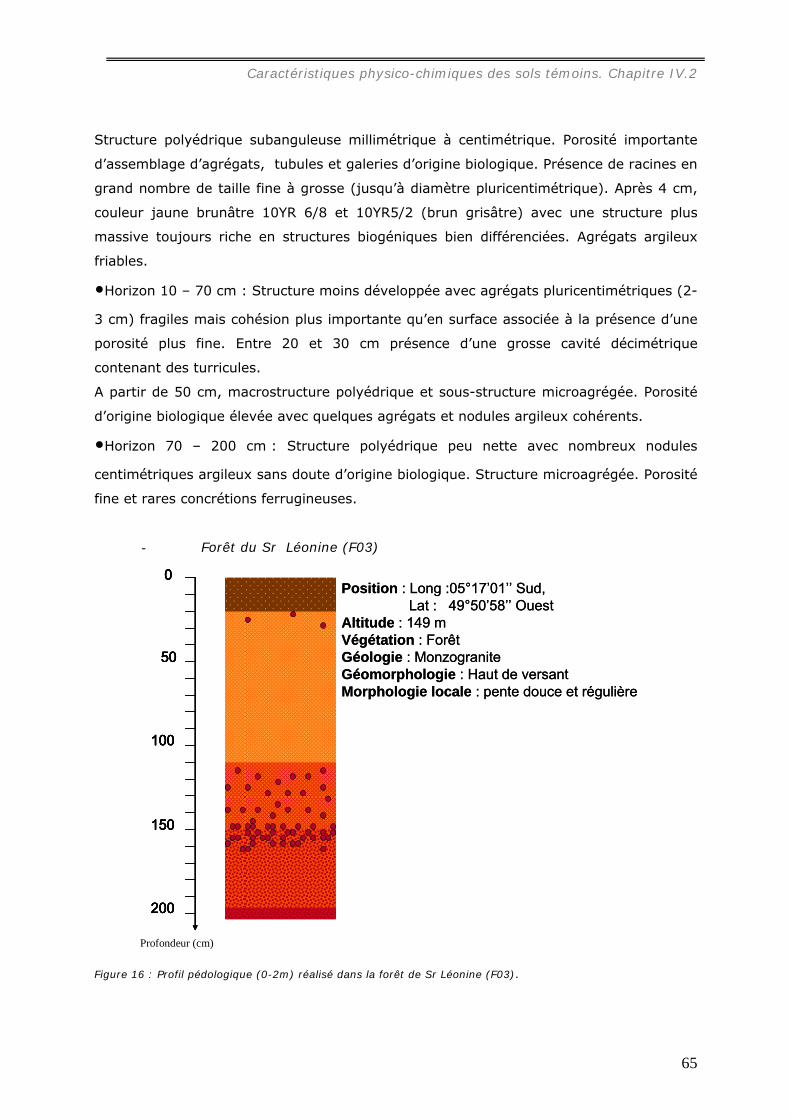
Caractéristiques physico-chimiques des sols témoins. Chapitre IV.2
65
Structure polyédrique subanguleuse millimétrique à centimétrique. Porosité importante
d’assemblage d’agrégats, tubules et galeries d’origine biologique. Présence de racines en
grand nombre de taille fine à grosse (jusqu’à diamètre pluricentimétrique). Après 4 cm,
couleur jaune brunâtre 10YR 6/8 et 10YR5/2 (brun grisâtre) avec une structure plus
massive toujours riche en structures biogéniques bien différenciées. Agrégats argileux
friables.
•Horizon 10 – 70 cm : Structure moins développée avec agrégats pluricentimétriques (2-
3 cm) fragiles mais cohésion plus importante qu’en surface associée à la présence d’une
porosité plus fine. Entre 20 et 30 cm présence d’une grosse cavité décimétrique
contenant des turricules.
A partir de 50 cm, macrostructure polyédrique et sous-structure microagrégée. Porosité
d’origine biologique élevée avec quelques agrégats et nodules argileux cohérents.
•Horizon 70 – 200 cm : Structure polyédrique peu nette avec nombreux nodules
centimétriques argileux sans doute d’origine biologique. Structure microagrégée. Porosité
fine et rares concrétions ferrugineuses.
- Forêt du Sr Léonine (F03)
0
50
100
150
200
Profondeur (cm)
Position : Long :05°17’01’’ Sud, Lat : 49°50’58’’ Ouest
Altitude : 149 mVégétation : ForêtGéologie : MonzograniteGéomorphologie : Haut de versantMorphologie locale : pente douce et régulière
0
50
100
150
200
Profondeur (cm)
0
50
100
150
200
Profondeur (cm)
0
50
100
150
200
0
50
100
150
200
Profondeur (cm)
Position : Long :05°17’01’’ Sud, Lat : 49°50’58’’ Ouest
Altitude : 149 mVégétation : ForêtGéologie : MonzograniteGéomorphologie : Haut de versantMorphologie locale : pente douce et régulière
Figure 16 : Profil pédologique (0-2m) réalisé dans la forêt de Sr Léonine (F03).

Caractéristiques physico-chimiques des sols témoins. Chapitre IV.2
66
•Horizon 0 – 20 cm : Horizon humifère jusqu’à 5 cm, riche en matière organique, de
couleur noire 10 YR 3/2 (brun grisâtre très sombre) à structure peu nette et forte
porosité. Puis à partir de 10 cm structure polyédrique subangulaire centimétrique à
pluricentimétrique. Mât racinaire important avec racines fines à moyennes. Texture argilo
sableuse, présence de quelques pores millimétriques et couleur plus jaunâtre 10 YR 5/4
(brun jaunâtre).
•Horizon 20 – 110 cm : Même texture, structure polyédrique peu nette, friable avec à 30
cm, apparition d’une sous structure microagrégée. Présence de quelques concrétions
ferrugineuses centimétriques. Activité biologique peu importante, porosité faible.
Quelques grosses racines. Couleur jaunâtre 10 YR 5/6 (brun jaunâtre). A partir de 70 cm
quelques pores tubulaires et couleur légèrement plus rouge 7,5 YR 5/8 (brun vif) vers
100 cm de profondeur.
•Horizon 110 – 140 cm : Horizon avec une structure fragmentaire peu nette, peu de
racines, quelques graviers centimétriques et à partir de 120 cm de nombreuses
concrétions ferrugineuses. Peu d’activité biologique remarquable. Même couleur brun vif
7,5 YR 5/8.
•Horizon 140 – 200 cm : horizon de transition vers l’altérite avec de très nombreuses
concrétions ferrugineuses de taille centimétrique à pluricentimétrique. Couleur rouge
jaunâtre 5 YR 5/8 à 5 YR 5/6. Entre les concrétions, structure microagrégée. Pas
d’activité biologique. A 200 cm, large zone de couleur rouge 2,5 YR 5/8.

Caractéristiques physico-chimiques des sols témoins. Chapitre IV.2
67
IV.1.3.b Description des profils pédologiques sous pâturages
- Pâturage Osano (P01)
Profondeur (cm)
0
50
100
150
200
Position : Long : 05°16’10’’ Sud, Lat : 49°50’24’’ Ouest
Altitude : 161 mVégétation : Pâturage de Brachiaria brisanthaGéologie : MonzograniteGéomorphologie : Haut de versantMorphologie locale : pente douce et régulière
Profondeur (cm)
0
50
100
150
200
Profondeur (cm)
0
50
100
150
200
Profondeur (cm)
0
50
100
150
200
0
50
100
150
200
0
50
100
150
200
Position : Long : 05°16’10’’ Sud, Lat : 49°50’24’’ Ouest
Altitude : 161 mVégétation : Pâturage de Brachiaria brisanthaGéologie : MonzograniteGéomorphologie : Haut de versantMorphologie locale : pente douce et régulière
Figure 17 : Profil pédologique (0-2m) réalisé dans le pâturage de Sr Osano (P01).
•Horizon 0 – 10 cm : Horizon riche en matière organique disposée en tâches. Entre de
nombreuses traces millimétriques d’hydromorphie, présence de quelques nodules et de
fragments de charbon de bois. Texture argilo-sableuse à argileuse.
Structure polyédrique subangulaire grossière (2-3cm), parfois lamellaire et compacte.
Couleur brun jaunâtre (10YR5/6). Nombreuses traces d’activité biologique (turricules,
galeries parfois comblées, agrégats de fourmis) et nombreuses racines filamenteuses
verticales.
•Horizon 10 – 50/ 60 cm : Matière organique assez abondante et bien dégradée. Texture
argileuse. Structure polyédrique sub-angulaire centimétrique. Couleur 10YR6/8.
Nombreuses racines verticales réparties préférentiellement sous les touffes de
graminées. Activité biologique réduite (quelques galeries millimétriques horizontales).
•Horizon 50/ 60 – 140 cm : Faible présence de matière organique. Présence de nodules
quartzeux centimétriques. Texture argileuse. Structure polyédrique sub-angulaire
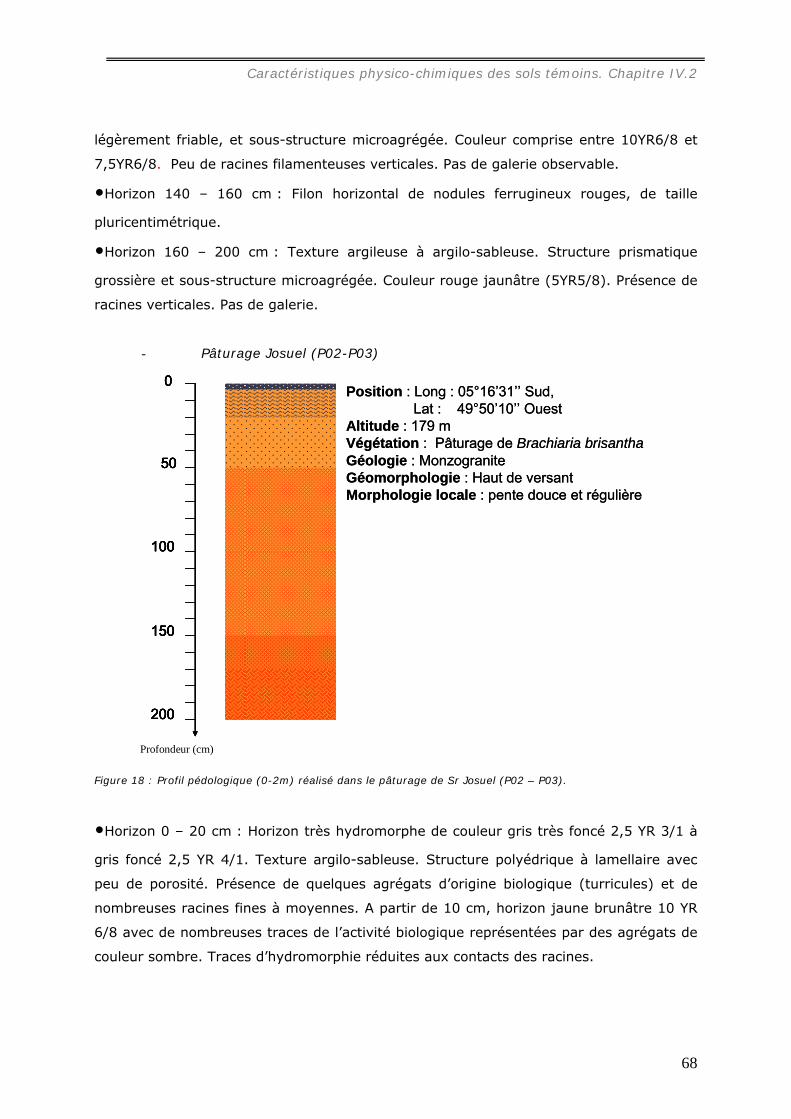
Caractéristiques physico-chimiques des sols témoins. Chapitre IV.2
68
légèrement friable, et sous-structure microagrégée. Couleur comprise entre 10YR6/8 et
7,5YR6/8. Peu de racines filamenteuses verticales. Pas de galerie observable.
•Horizon 140 – 160 cm : Filon horizontal de nodules ferrugineux rouges, de taille
pluricentimétrique.
•Horizon 160 – 200 cm : Texture argileuse à argilo-sableuse. Structure prismatique
grossière et sous-structure microagrégée. Couleur rouge jaunâtre (5YR5/8). Présence de
racines verticales. Pas de galerie.
- Pâturage Josuel (P02-P03)
0
50
100
150
200
Profondeur (cm)
Position : Long : 05°16’31’’ Sud, Lat : 49°50’10’’ Ouest
Altitude : 179 mVégétation : Pâturage de Brachiaria brisanthaGéologie : MonzograniteGéomorphologie : Haut de versantMorphologie locale : pente douce et régulière
0
50
100
150
200
Profondeur (cm)
0
50
100
150
200
Profondeur (cm)
0
50
100
150
200
0
50
100
150
200
Profondeur (cm)
Position : Long : 05°16’31’’ Sud, Lat : 49°50’10’’ Ouest
Altitude : 179 mVégétation : Pâturage de Brachiaria brisanthaGéologie : MonzograniteGéomorphologie : Haut de versantMorphologie locale : pente douce et régulière
Figure 18 : Profil pédologique (0-2m) réalisé dans le pâturage de Sr Josuel (P02 – P03).
•Horizon 0 – 20 cm : Horizon très hydromorphe de couleur gris très foncé 2,5 YR 3/1 à
gris foncé 2,5 YR 4/1. Texture argilo-sableuse. Structure polyédrique à lamellaire avec
peu de porosité. Présence de quelques agrégats d’origine biologique (turricules) et de
nombreuses racines fines à moyennes. A partir de 10 cm, horizon jaune brunâtre 10 YR
6/8 avec de nombreuses traces de l’activité biologique représentées par des agrégats de
couleur sombre. Traces d’hydromorphie réduites aux contacts des racines.
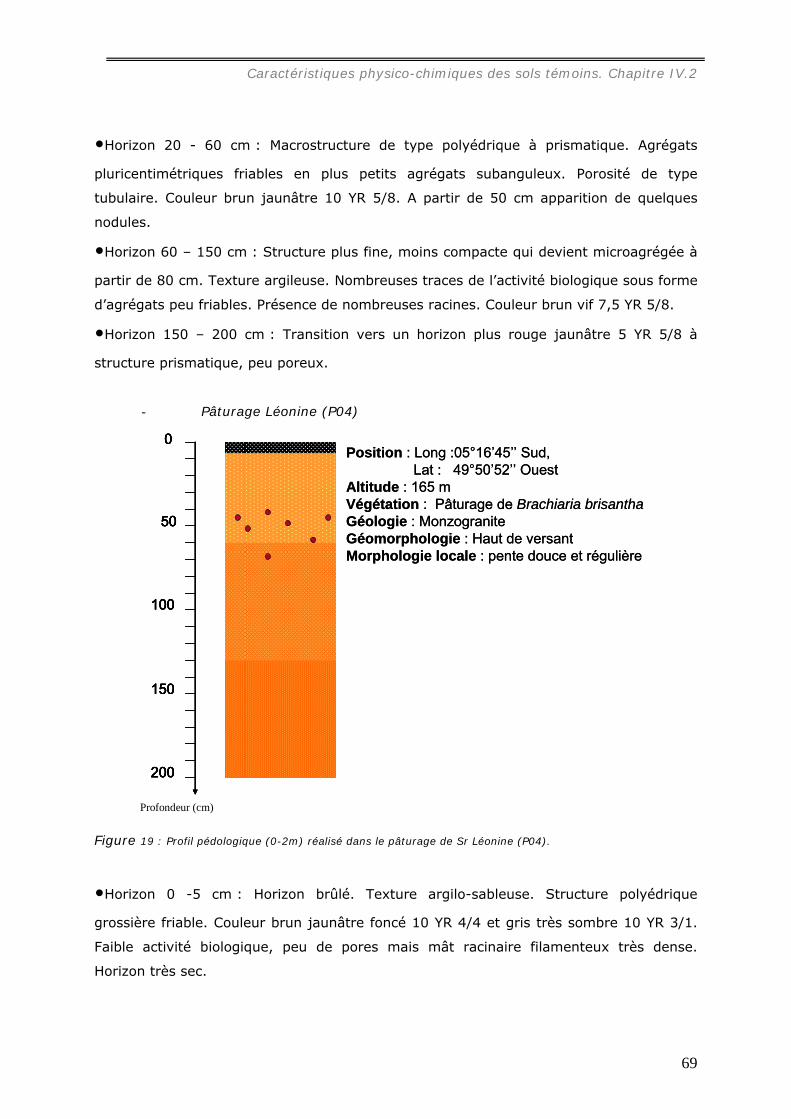
Caractéristiques physico-chimiques des sols témoins. Chapitre IV.2
69
•Horizon 20 - 60 cm : Macrostructure de type polyédrique à prismatique. Agrégats
pluricentimétriques friables en plus petits agrégats subanguleux. Porosité de type
tubulaire. Couleur brun jaunâtre 10 YR 5/8. A partir de 50 cm apparition de quelques
nodules.
•Horizon 60 – 150 cm : Structure plus fine, moins compacte qui devient microagrégée à
partir de 80 cm. Texture argileuse. Nombreuses traces de l’activité biologique sous forme
d’agrégats peu friables. Présence de nombreuses racines. Couleur brun vif 7,5 YR 5/8.
•Horizon 150 – 200 cm : Transition vers un horizon plus rouge jaunâtre 5 YR 5/8 à
structure prismatique, peu poreux.
- Pâturage Léonine (P04)
0
50
100
150
200
Profondeur (cm)
Position : Long :05°16’45’’ Sud, Lat : 49°50’52’’ Ouest
Altitude : 165 mVégétation : Pâturage de Brachiaria brisanthaGéologie : MonzograniteGéomorphologie : Haut de versantMorphologie locale : pente douce et régulière
0
50
100
150
200
Profondeur (cm)
0
50
100
150
200
Profondeur (cm)
0
50
100
150
200
0
50
100
150
200
Profondeur (cm)
Position : Long :05°16’45’’ Sud, Lat : 49°50’52’’ Ouest
Altitude : 165 mVégétation : Pâturage de Brachiaria brisanthaGéologie : MonzograniteGéomorphologie : Haut de versantMorphologie locale : pente douce et régulière
Figure 19 : Profil pédologique (0-2m) réalisé dans le pâturage de Sr Léonine (P04).
•Horizon 0 -5 cm : Horizon brûlé. Texture argilo-sableuse. Structure polyédrique
grossière friable. Couleur brun jaunâtre foncé 10 YR 4/4 et gris très sombre 10 YR 3/1.
Faible activité biologique, peu de pores mais mât racinaire filamenteux très dense.
Horizon très sec.

Caractéristiques physico-chimiques des sols témoins. Chapitre IV.2
70
•Horizon 5 - 60 cm : Texture argilo-sableuse à argileuse et même structure que l’horizon
supérieur. Couleur plus claire, brun jaunâtre 10 YR 5/6. Activité biologique faible mais
présence de quelques agrégats à partir de 10 cm, de quelques racines de diamètre
centimétrique à partir de 20 cm et pores centimétrique à 35 cm. Apparition de quelques
concrétions ferrugineuses millimétriques à 40 cm, structure polyédrique plus grossière
avec une sous-structure microaggrégée.
•Horizon 60 -130cm : même texture et même structure. Couleur entre brun jaunâtre 10
YR 6/8 et brun vif 7,5 YR 6/8. Rares concrétions, porosité fine, peu de racines. Horizon
plus frais et plus friable.
Horizon 130 – 200 cm : Couleur jaune rougeâtre 7,5 YR 6/8 avec quelques filons rouge
clairs 2,5 YR 7/8. Structure polyédrique plus massive et moins friable. Peu d’activité
biologique et peu de porosité. Pas de concrétions.
IV.1.3.c Autres parcelles
D’autres parcelles ont été étudiées : Forêt du Sr Cicéro (F04) ; Jachère du Sr Osano
(J01) ; Pâturages du Sr Cicéro (P05) et du Sr Francisco (P06) ; Rizière du Sr Josuel
(R01). Des sondages à la tarrière y ont été réalisés afin de s’assurer que les sols choisis
étaient bien des ferralsols profonds d’au moins 1,2m d’épaisseur au dessus de l’altérite
ou d’une transition vers l’altérite.
En conclusion, le sol des parcelles choisies est un latosol un peu moins
profond sous pâturage que sous forêt. Dans les pâturages, l’horizon superficiel (0-
10 cm) est légèrement compact en raison de la structure localement lamellaire
entre les touffes de graminée, et hétérogène à cause de la distribution en taches de
la matière organique et de l’hydromorphie, alors, que sous forêt, un horizon
organique humifère relativement homogène occupe les premiers cm du sol. La
texture est dans toutes les parcelles argilo-sableuse en surface et la structure du
sol, polyédrique sub-angulaire, à sous-structure microagrégée à partir de 60 cm.
Dans les deux milieux, vers 1,5 m de profondeur, parfois sous un « lit » de nodules
rouges, apparaît un horizon de transition vers l’altérite, rouge jaunâtre, à structure
prismatique grossière et sous-structure microagrégée, de couleur dominante brun
vif (7,5YR5/6).

Caractéristiques physico-chimiques des sols témoins. Chapitre IV.2
71
IV.2 CARACTERISTIQUES PHYSICO-CHIMIQUES DES SOLS TEMOINS.
Dans l’objectif de comprendre les propriétés physico-chimiques des structures
biogéniques étudiées, une caractérisation des sols, choisis comme « témoin », a tout
d’abord été réalisée. Cette étude a porté dans un premier temps sur la comparaison de
profils granulométriques et des propriétés physiques (compaction, stabilité) puis dans un
second temps, un bilan chimique des sols témoins en forêts et dans les pâturages a été
effectué.
IV.2.1 Les latosols amazoniens.
L’état d’un latosol amazonien sous pâturage de 5 à 7 ans, de faible productivité,
se caractérise par une faible quantité de matière organique par rapport aux sols sous
forêt (Corazza et al., 1999 ; Corazza et al., 2000). La concentration en ions phosphate
disponible pour les plantes est très faible (Vitrac, 2001 ; Corazza, 2002 ; Brunet, 2004).
La structure du sol a été généralement modifiée après la déforestation : une perte de
porosité superficielle s’observe jusqu’à environ 50 cm de profondeur (Fregonezi et al.,
2001 ; Balbino et al., 2002) et un développement de volumes compacts et de croutes de
surface limitant le volume des racines s’opère. Dans le même temps, les propriétés
hydrauliques ne paraissent pas complètement altérées, le desquament et l’érosion
lamellaire sont en Amazonie des phénomènes localisés. La diversité et la biomasse des
invertébrés sont fortement altérées en comparaison avec les communautés observées
sous forêt (Bobillier, 1999 ; Pasini et al., 2000).
IV.2.2 Profils granulométriques des sols.
Les données granulométriques sont présentées pour des profils réalisés jusqu’à
2,20 m de profondeur. (Données en annexe 2 pour les profils de forêts : parcelles F01
(forêt dégradée) et F02 et annexes 3 et 3 bis pour les profils sous pâturages : parcelles
P01, P03 et P04).
Les profils granulométriques obtenus mettent en évidence dans les trois milieux
(forêt, forêt dégradée et pâturage) une forte variabilité des teneurs en argile et en sable
grossier en fonction de la profondeur. La teneur en argile augmente jusqu’à 1m de
profondeur (30% à 40% en surface et 60% à 75% à 1m de profondeur) puis diminue
(40% à 50% au-delà de 2m de profondeur). La teneur en sable grossier plus forte en
surface (30% à 50%) diminue en profondeur (10 à 35% au-delà de 2m). Le profil

Caractéristiques physico-chimiques des sols témoins. Chapitre IV.2
72
d’argile est caractéristique des ferralsols. La classe des limons fins augmente à partir de
150cm de profondeur (7,8%); elle représente jusqu’à 20% à 210cm de profondeur dans
le pâturage P01 et P03.
La texture des sols du site d’étude est sablo-argileuse. Les profils
granulométriques ne différent pas entre les forêts et les pâturages.
IV.2.3 Densité apparente et densité de mottes des sols témoins.
Si La densité apparente permet d’estimer l’état de tassement d’un horizon de sol,
la densité de motte permet d’estimer celui des agrégats élémentaires dont la taille est
plus proche de celle des structures biogéniques tel que les turricules d’annélides.
Selon Grimaldi M. (non publié), les données de densité apparente (méthode des
cylindres) des différents milieux mettent en évidence l’intensité de la compaction du sol
résultant de la déforestation et la profondeur affectée par ce phénomène. Une
augmentation significative de la densité apparente dans les 30 premiers centimètres de
profondeur a été observée dans les pâturages. L’augmentation de densité sous pâturage
est plus forte dans les 5 premiers centimètres. La moyenne des densités varie de
1,1 g.cm-3 sous forêt à 1,45 g.cm-3 sous pâturage de Brachiaria brisantha de 5-6 ans,
soit une diminution de 13% de la porosité du sol. La variabilité spatiale des moyennes de
densité apparente du sol est généralement élevée quelque soit la couverture végétale et
plus particulièrement dans les 5 premiers centimètres des sols de pâturage en raison de
la distribution spatiale des touffes de graminées.
Les valeurs mesurées de densité de motte (méthode au pétrole) n’ont pas permis de
différencier significativement les agrégats prélevés dans les 30 premiers centimètres des
sols de forêts (1,50 ± 0,13) de ceux prélevés dans les sols de pâturage (1,57 ± 0,08). La
densité de motte observée dans la forêt dégradée est significativement plus faible que
dans les autres milieux (1,32 ± 0,16).
La réduction de porosité observée dans les sols de pâturages (environ 13%
dans les 5 premiers centimètres) ne s’observe pas au niveau de la porosité des
agrégats (les densités de motte ne diffèrent pas dans les agrégats de pâturages et
ceux de forêts.) Ce tassement en surface correspond donc à une diminution de
macroporosité sans doute consécutive à la déforestation et aux pratiques agricoles.
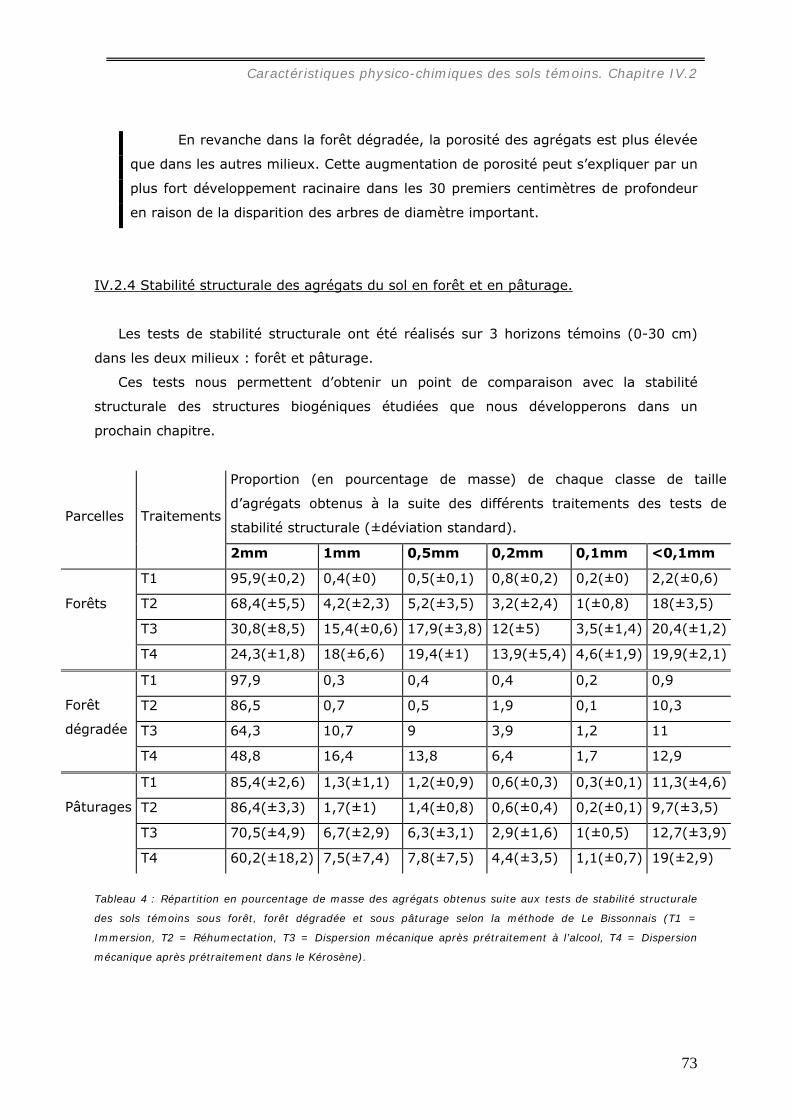
Caractéristiques physico-chimiques des sols témoins. Chapitre IV.2
73
En revanche dans la forêt dégradée, la porosité des agrégats est plus élevée
que dans les autres milieux. Cette augmentation de porosité peut s’expliquer par un
plus fort développement racinaire dans les 30 premiers centimètres de profondeur
en raison de la disparition des arbres de diamètre important.
IV.2.4 Stabilité structurale des agrégats du sol en forêt et en pâturage.
Les tests de stabilité structurale ont été réalisés sur 3 horizons témoins (0-30 cm)
dans les deux milieux : forêt et pâturage.
Ces tests nous permettent d’obtenir un point de comparaison avec la stabilité
structurale des structures biogéniques étudiées que nous développerons dans un
prochain chapitre.
Proportion (en pourcentage de masse) de chaque classe de taille
d’agrégats obtenus à la suite des différents traitements des tests de
stabilité structurale (±déviation standard). Parcelles Traitements
2mm 1mm 0,5mm 0,2mm 0,1mm <0,1mm
T1 95,9(±0,2) 0,4(±0) 0,5(±0,1) 0,8(±0,2) 0,2(±0) 2,2(±0,6)
T2 68,4(±5,5) 4,2(±2,3) 5,2(±3,5) 3,2(±2,4) 1(±0,8) 18(±3,5)
T3 30,8(±8,5) 15,4(±0,6) 17,9(±3,8) 12(±5) 3,5(±1,4) 20,4(±1,2)
Forêts
T4 24,3(±1,8) 18(±6,6) 19,4(±1) 13,9(±5,4) 4,6(±1,9) 19,9(±2,1)
T1 97,9 0,3 0,4 0,4 0,2 0,9
T2 86,5 0,7 0,5 1,9 0,1 10,3
T3 64,3 10,7 9 3,9 1,2 11
Forêt
dégradée
T4 48,8 16,4 13,8 6,4 1,7 12,9
T1 85,4(±2,6) 1,3(±1,1) 1,2(±0,9) 0,6(±0,3) 0,3(±0,1) 11,3(±4,6)
T2 86,4(±3,3) 1,7(±1) 1,4(±0,8) 0,6(±0,4) 0,2(±0,1) 9,7(±3,5)
T3 70,5(±4,9) 6,7(±2,9) 6,3(±3,1) 2,9(±1,6) 1(±0,5) 12,7(±3,9)
Pâturages
T4 60,2(±18,2) 7,5(±7,4) 7,8(±7,5) 4,4(±3,5) 1,1(±0,7) 19(±2,9)
Tableau 4 : Répartition en pourcentage de masse des agrégats obtenus suite aux tests de stabilité structurale
des sols témoins sous forêt, forêt dégradée et sous pâturage selon la méthode de Le Bissonnais (T1 =
Immersion, T2 = Réhumectation, T3 = Dispersion mécanique après prétraitement à l’alcool, T4 = Dispersion
mécanique après prétraitement dans le Kérosène).

Caractéristiques physico-chimiques des sols témoins. Chapitre IV.2
74
La stabilité des horizons (0-30 cm) des forêts ne se différencie pas
significativement de celle des forêts dégradées qui cependant paraissent plus difficile à
déstructurer (Tableau 4). La structure des horizons 0-30 cm des pâturages est
globalement plus stable que celle des forêts (dégradées ou non). La proportion
d’agglomérats de pâturages non modifiés dont la taille est supérieure à 2mm est restée
significativement plus importante que celle de forêts lors des tests d’immersion ou de
dispersion mécanique après pré-traitement dans l’alcool et le kérosène (de 60% à 86%
d’agrégats non modifiés dont la taille est supérieure à 2mm).
Les sols de surface des forêts (dégradées ou non) se sont avérés plus sensibles à
la réhumectation qu’à l’immersion (25% à 30% de déstructuration). Ces sols sont ainsi
plus sensibles à la dissolution des ciments argileux et humiques qu’aux phénomènes de
pression dans les pores. La dispersion mécanique quel que soit le prétraitement
déstructure fortement les agglomérats de forêt (jusqu’à 70% de déstructuration). Donc
ces horizons se caractérisent par des agrégats fragiles.
Dans les pâturages, l’immersion et la réhumectation ne dégradent pas fortement
la structure de l’horizon 0-30 cm (environ 25% de dégradation). La dispersion mécanique
après pré-traitement dans l’alcool et le kérosène ont permis de mieux dégrader la
structure physique des sols de surface (60% à 70% d’agrégats non modifiés dont la taille
est supérieure à 2mm) (Tableau 4).
Ces résultats confirment que ces sols sont très stables. Cette stabilité s’explique
sans doute par le tassement observé dans cet horizon (diminution de la porosité) et par
la culture sur brûlis qui chaque année engendre la formation de pseudosables par
déshydratation des argiles (pectisation). Ces pseudosables sont des agrégats très
stables.
La dégradation des sols ici observée s’est répartie dans toutes les classes. La
proportion des particules de taille inférieure à 1mm est la plus élevée (de 10% à 15%
dans les forêts et de 13% à 19% dans les pâturages)
La stabilité structurale des horizons de surface (0-30 cm) des forêts ne
diffère pas significativement de celle des forêts dégradées bien que l’horizon de ces
dernières paraît globalement plus stable.
Dans les pâturages la stabilité des horizons est significativement plus
importante que celle des forêts (dégradées ou non) en raison sans doute du

Caractéristiques physico-chimiques des sols témoins. Chapitre IV.2
75
tassement de cet horizon (diminution de la porosité). Ces horizons de pâturages
restent plus sensibles à la dispersion mécanique.
La végétation et la litière sous forêt assure une protection de la surface du
sol or dans les pâturages, la distribution clairsemée des touffes de Brachiaria br.
laissent des zones de sol nu (Chapitre II). De plus les racines qui assurent un
maintien du sol dans les forêts se distribuent dans les pâturages de manière
hétérogène. Les horizons de surface des pâturages sont donc plus sensibles à
l’érosion dans l’espace intertouffe que ceux des forêts.
IV.2.5 Bilan chimique des horizons de surface des sols de forêt et de pâturages.
La croissance et le développement des végétaux nécessitent de l'eau, de la lumière,
du carbone, de l’oxygène et des éléments minéraux. L’air fournit l’oxygène et le gaz
carbonique, source de carbone, que la plante fixe grâce à la photosynthèse. Les éléments
nutritifs minéraux proviennent, en grande partie de la matière organique du sol et ils
sont impliqués dans toutes les fonctions métaboliques dans la plante. Ce sont
principalement les éléments: azote (N), phosphore (P), potassium (K), calcium (Ca) et
magnésium (Mg). Ces éléments se trouvent dans la solution du sol et sont piégés dans le
complexe argilo-humique sous forme d’ions.
Dans l’objectif de mettre en évidence l’effet de la déforestation sur les propriétés
chimiques du sol, un dosage du carbone, des principaux cations (Al3+, Ca2+, Mg2+, K+) et
du phosphore assimilable a été réalisé. Le pH, la CEC et le taux de saturation en base ont
été également mesurés afin de compléter le bilan chimique des sols des deux milieux.
IV.2.5.a Dosage du carbone du sol.
Les forêts jouent un rôle majeur dans le stockage du carbone. Le dioxyde de carbone
de l’air est prélevé de l’atmosphère et utilisé pour produire la matière organique. Lors de
la déforestation, une partie du carbone est restitué au sol.
Le dosage du carbone permet d’évaluer de manière approximative la quantité globale
de matière organique présente dans les sols du site d’étude.
La concentration en carbone a été dosée dans les 30 premiers centimètres sous la
surface dans les trois milieux (2 forêts, 1 forêt dégradée et 2 pâturages).
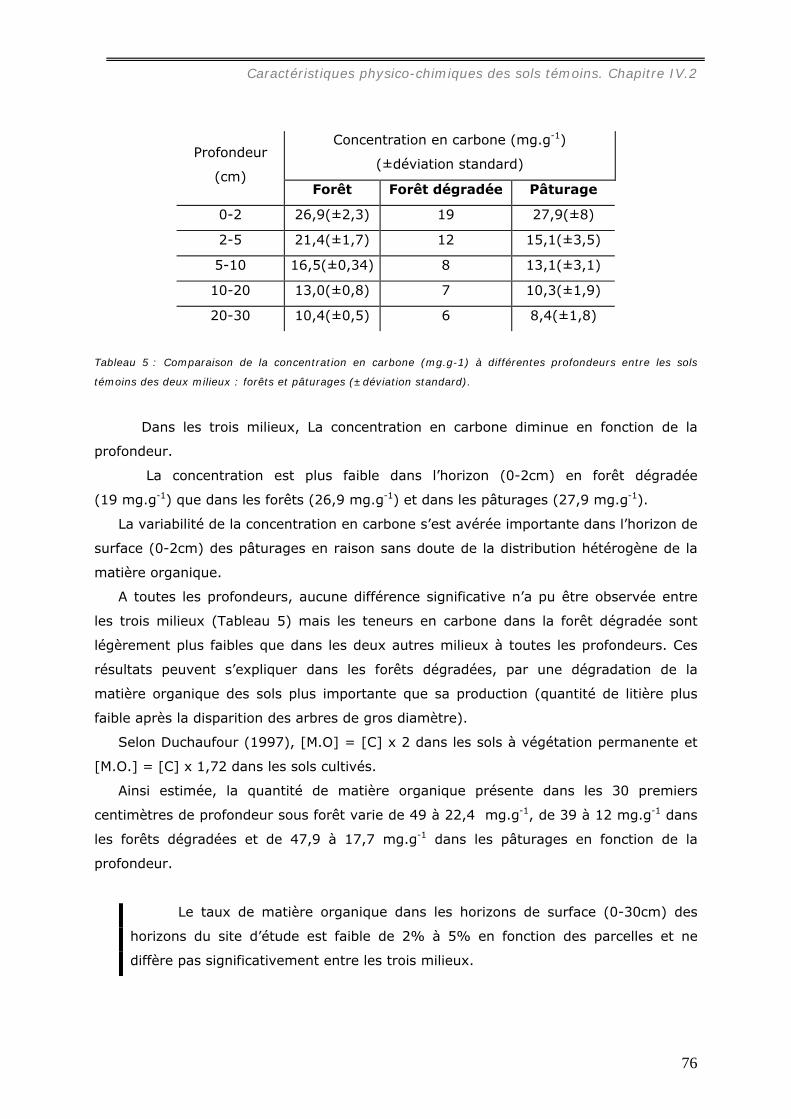
Caractéristiques physico-chimiques des sols témoins. Chapitre IV.2
76
Concentration en carbone (mg.g-1)
(±déviation standard) Profondeur
(cm) Forêt Forêt dégradée Pâturage
0-2 26,9(±2,3) 19 27,9(±8)
2-5 21,4(±1,7) 12 15,1(±3,5)
5-10 16,5(±0,34) 8 13,1(±3,1)
10-20 13,0(±0,8) 7 10,3(±1,9)
20-30 10,4(±0,5) 6 8,4(±1,8)
Tableau 5 : Comparaison de la concentration en carbone (mg.g-1) à différentes profondeurs entre les sols
témoins des deux milieux : forêts et pâturages (±déviation standard).
Dans les trois milieux, La concentration en carbone diminue en fonction de la
profondeur.
La concentration est plus faible dans l’horizon (0-2cm) en forêt dégradée
(19 mg.g-1) que dans les forêts (26,9 mg.g-1) et dans les pâturages (27,9 mg.g-1).
La variabilité de la concentration en carbone s’est avérée importante dans l’horizon de
surface (0-2cm) des pâturages en raison sans doute de la distribution hétérogène de la
matière organique.
A toutes les profondeurs, aucune différence significative n’a pu être observée entre
les trois milieux (Tableau 5) mais les teneurs en carbone dans la forêt dégradée sont
légèrement plus faibles que dans les deux autres milieux à toutes les profondeurs. Ces
résultats peuvent s’expliquer dans les forêts dégradées, par une dégradation de la
matière organique des sols plus importante que sa production (quantité de litière plus
faible après la disparition des arbres de gros diamètre).
Selon Duchaufour (1997), [M.O] = [C] x 2 dans les sols à végétation permanente et
[M.O.] = [C] x 1,72 dans les sols cultivés.
Ainsi estimée, la quantité de matière organique présente dans les 30 premiers
centimètres de profondeur sous forêt varie de 49 à 22,4 mg.g-1, de 39 à 12 mg.g-1 dans
les forêts dégradées et de 47,9 à 17,7 mg.g-1 dans les pâturages en fonction de la
profondeur.
Le taux de matière organique dans les horizons de surface (0-30cm) des
horizons du site d’étude est faible de 2% à 5% en fonction des parcelles et ne
diffère pas significativement entre les trois milieux.
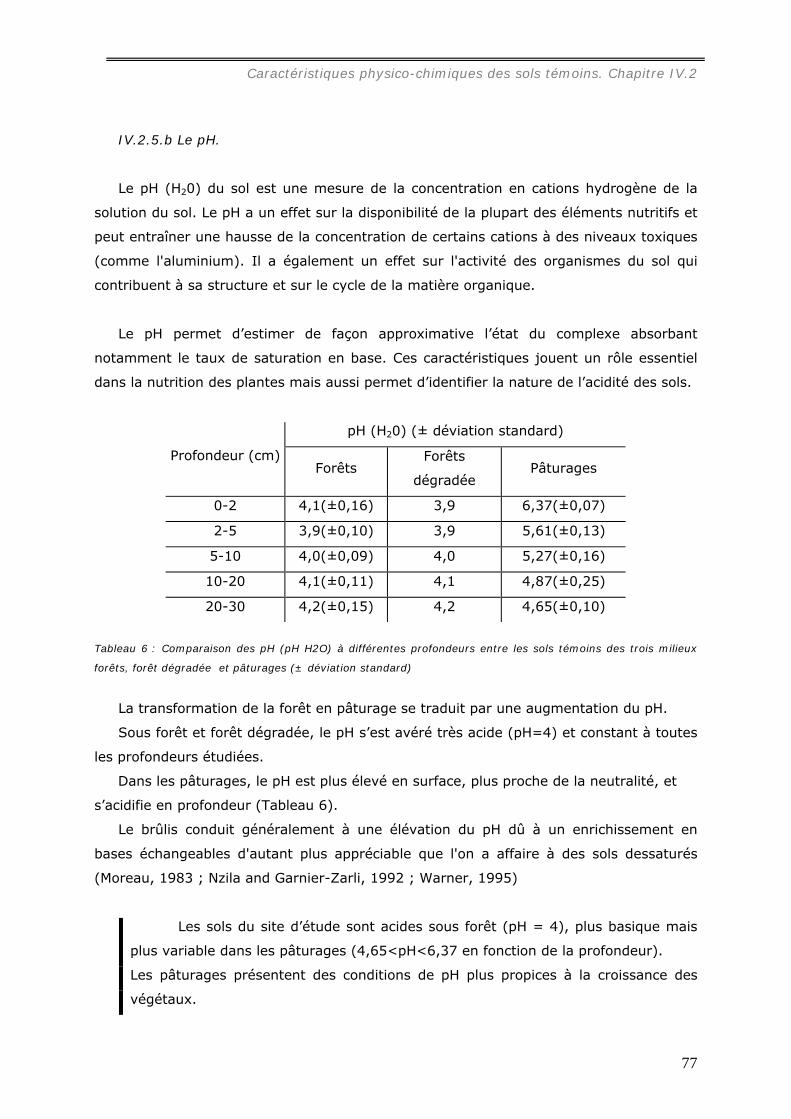
Caractéristiques physico-chimiques des sols témoins. Chapitre IV.2
77
IV.2.5.b Le pH.
Le pH (H20) du sol est une mesure de la concentration en cations hydrogène de la
solution du sol. Le pH a un effet sur la disponibilité de la plupart des éléments nutritifs et
peut entraîner une hausse de la concentration de certains cations à des niveaux toxiques
(comme l'aluminium). Il a également un effet sur l'activité des organismes du sol qui
contribuent à sa structure et sur le cycle de la matière organique.
Le pH permet d’estimer de façon approximative l’état du complexe absorbant
notamment le taux de saturation en base. Ces caractéristiques jouent un rôle essentiel
dans la nutrition des plantes mais aussi permet d’identifier la nature de l’acidité des sols.
pH (H20) (± déviation standard)
Profondeur (cm) Forêts
Forêts
dégradée Pâturages
0-2 4,1(±0,16) 3,9 6,37(±0,07)
2-5 3,9(±0,10) 3,9 5,61(±0,13)
5-10 4,0(±0,09) 4,0 5,27(±0,16)
10-20 4,1(±0,11) 4,1 4,87(±0,25)
20-30 4,2(±0,15) 4,2 4,65(±0,10)
Tableau 6 : Comparaison des pH (pH H2O) à différentes profondeurs entre les sols témoins des trois milieux
forêts, forêt dégradée et pâturages (± déviation standard)
La transformation de la forêt en pâturage se traduit par une augmentation du pH.
Sous forêt et forêt dégradée, le pH s’est avéré très acide (pH=4) et constant à toutes
les profondeurs étudiées.
Dans les pâturages, le pH est plus élevé en surface, plus proche de la neutralité, et
s’acidifie en profondeur (Tableau 6).
Le brûlis conduit généralement à une élévation du pH dû à un enrichissement en
bases échangeables d'autant plus appréciable que l'on a affaire à des sols dessaturés
(Moreau, 1983 ; Nzila and Garnier-Zarli, 1992 ; Warner, 1995)
Les sols du site d’étude sont acides sous forêt (pH = 4), plus basique mais
plus variable dans les pâturages (4,65<pH<6,37 en fonction de la profondeur).
Les pâturages présentent des conditions de pH plus propices à la croissance des
végétaux.
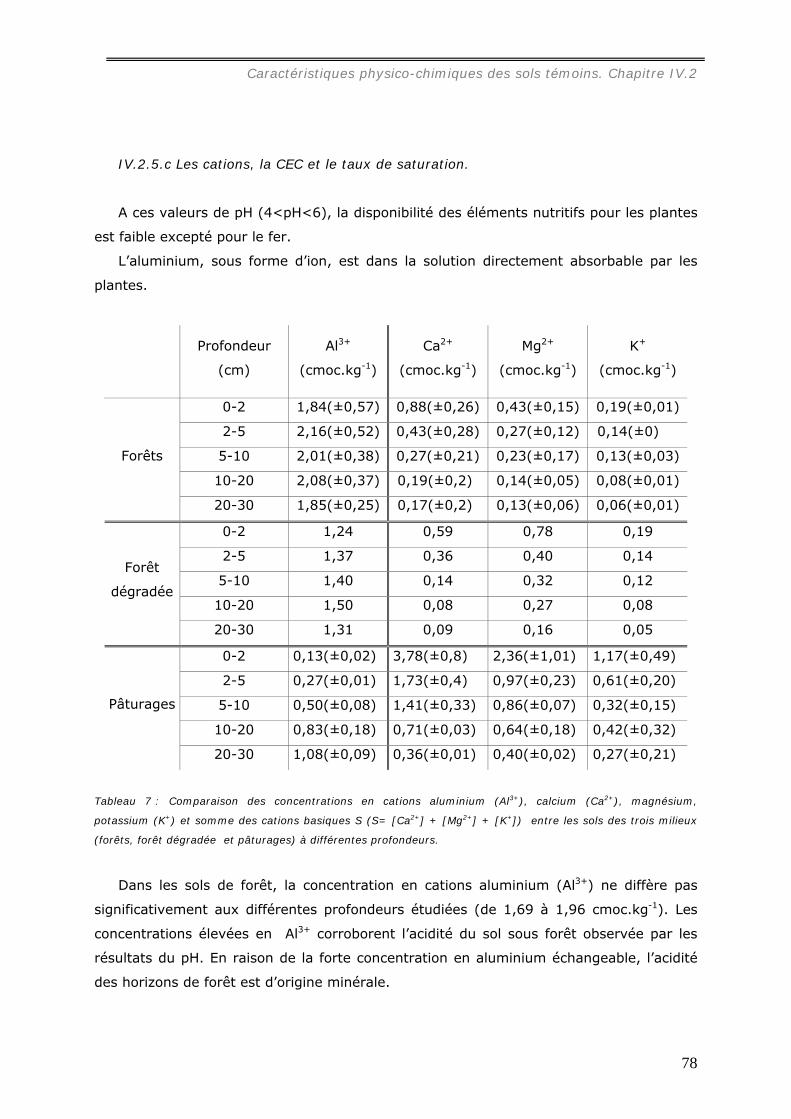
Caractéristiques physico-chimiques des sols témoins. Chapitre IV.2
78
IV.2.5.c Les cations, la CEC et le taux de saturation.
A ces valeurs de pH (4<pH<6), la disponibilité des éléments nutritifs pour les plantes
est faible excepté pour le fer.
L’aluminium, sous forme d’ion, est dans la solution directement absorbable par les
plantes.
Profondeur
(cm)
Al3+
(cmoc.kg-1)
Ca2+
(cmoc.kg-1)
Mg2+
(cmoc.kg-1)
K+
(cmoc.kg-1)
0-2 1,84(±0,57) 0,88(±0,26) 0,43(±0,15) 0,19(±0,01)
2-5 2,16(±0,52) 0,43(±0,28) 0,27(±0,12) 0,14(±0)
5-10 2,01(±0,38) 0,27(±0,21) 0,23(±0,17) 0,13(±0,03)
10-20 2,08(±0,37) 0,19(±0,2) 0,14(±0,05) 0,08(±0,01)
Forêts
20-30 1,85(±0,25) 0,17(±0,2) 0,13(±0,06) 0,06(±0,01)
0-2 1,24 0,59 0,78 0,19
2-5 1,37 0,36 0,40 0,14
5-10 1,40 0,14 0,32 0,12
10-20 1,50 0,08 0,27 0,08
Forêt
dégradée
20-30 1,31 0,09 0,16 0,05
0-2 0,13(±0,02) 3,78(±0,8) 2,36(±1,01) 1,17(±0,49)
2-5 0,27(±0,01) 1,73(±0,4) 0,97(±0,23) 0,61(±0,20)
5-10 0,50(±0,08) 1,41(±0,33) 0,86(±0,07) 0,32(±0,15)
10-20 0,83(±0,18) 0,71(±0,03) 0,64(±0,18) 0,42(±0,32)
Pâturages
20-30 1,08(±0,09) 0,36(±0,01) 0,40(±0,02) 0,27(±0,21)
Tableau 7 : Comparaison des concentrations en cations aluminium (Al3+), calcium (Ca2+), magnésium,
potassium (K+) et somme des cations basiques S (S= [Ca2+] + [Mg2+] + [K+]) entre les sols des trois milieux
(forêts, forêt dégradée et pâturages) à différentes profondeurs.
Dans les sols de forêt, la concentration en cations aluminium (Al3+) ne diffère pas
significativement aux différentes profondeurs étudiées (de 1,69 à 1,96 cmoc.kg-1). Les
concentrations élevées en Al3+ corroborent l’acidité du sol sous forêt observée par les
résultats du pH. En raison de la forte concentration en aluminium échangeable, l’acidité
des horizons de forêt est d’origine minérale.
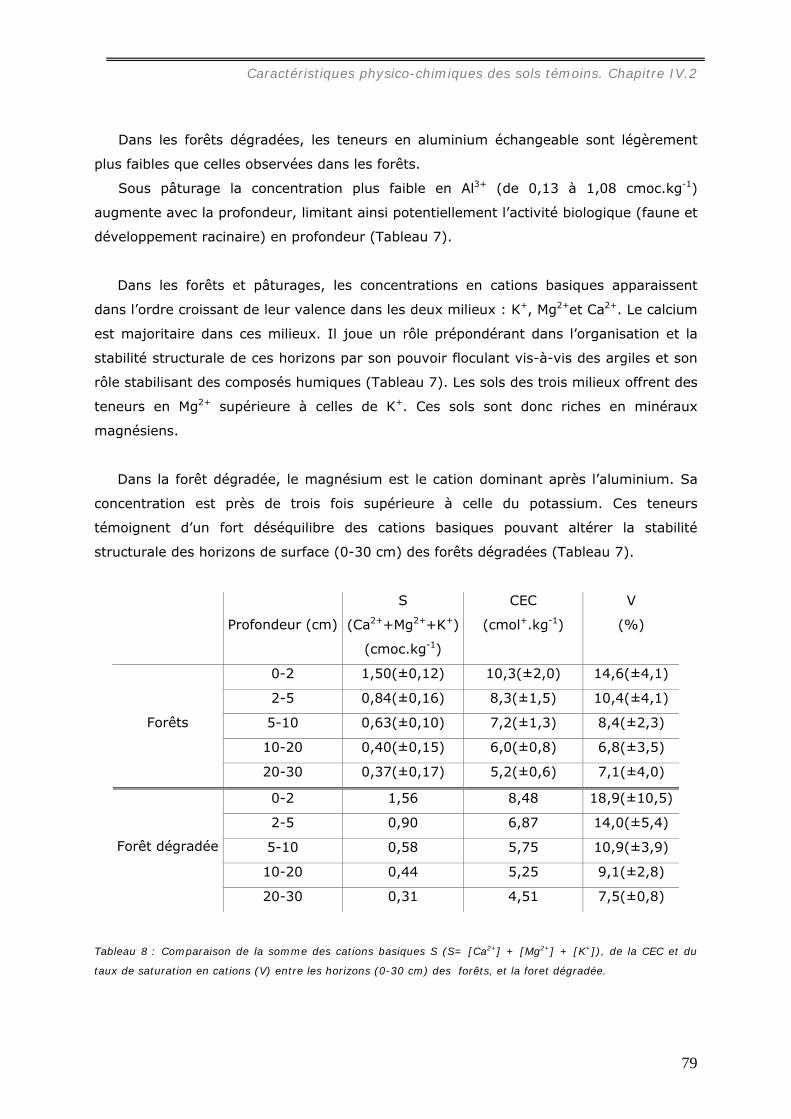
Caractéristiques physico-chimiques des sols témoins. Chapitre IV.2
79
Dans les forêts dégradées, les teneurs en aluminium échangeable sont légèrement
plus faibles que celles observées dans les forêts.
Sous pâturage la concentration plus faible en Al3+ (de 0,13 à 1,08 cmoc.kg-1)
augmente avec la profondeur, limitant ainsi potentiellement l’activité biologique (faune et
développement racinaire) en profondeur (Tableau 7).
Dans les forêts et pâturages, les concentrations en cations basiques apparaissent
dans l’ordre croissant de leur valence dans les deux milieux : K+, Mg2+et Ca2+. Le calcium
est majoritaire dans ces milieux. Il joue un rôle prépondérant dans l’organisation et la
stabilité structurale de ces horizons par son pouvoir floculant vis-à-vis des argiles et son
rôle stabilisant des composés humiques (Tableau 7). Les sols des trois milieux offrent des
teneurs en Mg2+ supérieure à celles de K+. Ces sols sont donc riches en minéraux
magnésiens.
Dans la forêt dégradée, le magnésium est le cation dominant après l’aluminium. Sa
concentration est près de trois fois supérieure à celle du potassium. Ces teneurs
témoignent d’un fort déséquilibre des cations basiques pouvant altérer la stabilité
structurale des horizons de surface (0-30 cm) des forêts dégradées (Tableau 7).
Profondeur (cm)
S
(Ca2++Mg2++K+)
(cmoc.kg-1)
CEC
(cmol+.kg-1)
V
(%)
0-2 1,50(±0,12) 10,3(±2,0) 14,6(±4,1)
2-5 0,84(±0,16) 8,3(±1,5) 10,4(±4,1)
5-10 0,63(±0,10) 7,2(±1,3) 8,4(±2,3)
10-20 0,40(±0,15) 6,0(±0,8) 6,8(±3,5)
Forêts
20-30 0,37(±0,17) 5,2(±0,6) 7,1(±4,0)
0-2 1,56 8,48 18,9(±10,5)
2-5 0,90 6,87 14,0(±5,4)
5-10 0,58 5,75 10,9(±3,9)
10-20 0,44 5,25 9,1(±2,8)
Forêt dégradée
20-30 0,31 4,51 7,5(±0,8)
Tableau 8 : Comparaison de la somme des cations basiques S (S= [Ca2+] + [Mg2+] + [K+]), de la CEC et du
taux de saturation en cations (V) entre les horizons (0-30 cm) des forêts, et la foret dégradée.

Caractéristiques physico-chimiques des sols témoins. Chapitre IV.2
80
Profondeur (cm)
S
(Ca2++Mg2++K+)
(cmoc.kg-1)
CEC
(cmol+.kg-1)
V
(%)
2-5 3,31(±0,43) 6,6(±1,0) 48,3(±7)
5-10 2,6(±0,25) 6,0(±0,9) 32,3(±16,1)
10-20 1,77(±0,48) 5,3(±0,2) 30(±15,1)
Pâturages
20-30 1,04(±0,22) 4,5(±0,3) 20,1(±9,9)
Tableau 9 : Comparaison de la somme des cations basiques S (S= [Ca2+] + [Mg2+] + [K+]), de la CEC et du
taux de saturation en cations (V) entre les horizons (0-30 cm) des pâturages.
Dans les sols des pâturages, la somme des cations varie de 7,3 cmoc.kg-1 à
1,04 cmoc.kg-1 en fonction de la profondeur (0-30 cm) (Tableau 9); dans celles des
forêts, les valeurs sont comprises entre 1,50 cmoc.kg-1 et 0,37 cmoc.kg-1 et ne diffèrent
pas de celles observées dans les forêts dégradées (de 1,56 à 0,31 cmoc.kg-1) (Tableau
8). Comme dans d’autres contextes de déforestation, la concentration en cations est
significativement plus importante dans les pâturages que dans les forêts (dégradées ou
non) à toutes les profondeurs étudiées (Tableau 8 et 9). L’alcanisation des horizons 0-30
cm des pâturages peut s’expliquer par le brûlis qui concentre les cations basiques dans
les cendres mais aussi par la remontée de cations alcalins ou alcalino-terreux des
horizons profonds aux horizons de surface par les cycles biologiques.
La capacité d’échange cationique diminue avec la profondeur dans les trois milieux.
Aucune différence significative n’a été observée entre les sols de pâturages et ceux des
forêts et de la forêt dégradée.
Dans les horizons de surface (0-2 cm), la CEC de la forêt dégradée est plus faible
(8,48) que celles observées dans les pâturages (10,2 ± 1,7) et les forêts (10,3 ± 2,0).
Dans les horizons plus profonds (jusqu’à 30 cm) la CEC est légèrement plus élevée dans
la forêt que dans les deux autres milieux (Tableaux 8 et 9).
Le taux de saturation en bases est faible dans les sols des forêts (de 15% en surface
à 7% à 30 cm de profondeur). Les horizons (0-30 cm) de la forêt dégradée sont
légèrement plus saturés (V de 19% à 7,5%). Les valences du complexe absorbant sont
donc essentiellement occupés dans ces deux milieux par les ions hydrogène et
aluminium.
Dans les horizons de surface 0-2 cm des pâturages, plus de 71% du complexe est
saturé (Tableau 8).

Caractéristiques physico-chimiques des sols témoins. Chapitre IV.2
81
Le rapport [Ca2+]/[Mg2+]= 1,6 dans cet horizon (Tableau 7). Ce rapport indique que
le matériau est magnésique (Baize and Girard, 1995). A 30 cm de profondeur le taux de
saturation est de l’ordre de 20% ; le complexe est alors déssaturé. La variabilité du taux
de saturation est importante à partir de 5 cm de profondeur et diminue en profondeur
dans les pâturages (Tableau 8).
Dans la solution du sol, le phosphore dissous est présent sous forme
d’orthophosphates H2PO4- et/ou HPO4
2-. Ces phosphates peuvent être absorbés par les
plantes mais surtout fixés sur les colloïdes du sol. Généralement dans les latosols
dessaturés acides, une grande partie du phosphore lié à l’alumine précipite de façon
irréversible (Mathieu and Pieltain, 2003).
P (mg.kg-1) Profondeur
(cm) Forêt Forêt dégradée Pâturage
0-2 8,38(±1,71) 5,84 6,95(±7,33)
2-5 5,41(±1,18) 3,87 2,45(±2,53)
5-10 3,53(±0,95) 2,55 1,20(±0,47)
10-20 1,88(±0,47) 1,64 0,65(±0,44)
20-30 0,86(±0,33) 0,94 0,28(±0,36)
Tableau 10 : Comparaison des concentrations en phosphore assimilable (P) entre les horizons des trois milieux
(forêts, forêt dégradée et pâturages) à différentes profondeurs (0-30 cm).
La concentration en phosphore en surface est de 8,38 mg.kg-1 dans les forêts et 6,95
dans les pâturages. Ces teneurs sont supérieures à celle observée dans la forêt dégradée
5,84 mg.kg-1.
Cette concentration diminue avec la profondeur : sous forêt, la concentration est
d’environ 0,86 mg.kg-1 à 25 cm de profondeur, 0,94 mg.kg-1 dans la forêt dégradée et
0,28 mg.kg-1 dans les pâturages.
A toutes les profondeurs, ces concentrations ne diffèrent pas significativement entre
les trois milieux. La variabilité de la concentration en phosphore est bien plus importante
dans les pâturages (Tableau 10).
Le pH et les concentrations cationiques ne diffèrent pas significativement
entre les forêts et les forêts dégradées. Les concentrations élevées en Al3+ des
forêts (dégradées ou non) corroborent l’acidification du sol sous forêt observée par
les résultats du pH. Le taux de saturation en bases est faible (15% en surface). Les

Caractéristiques physico-chimiques des sols témoins. Chapitre IV.2
82
valences sont donc occupés dans ces deux milieux par les ions hydrogène et
aluminium.
Dans les pâturages, la concentration en cations est significativement plus
importante que dans les forêts à toutes les profondeurs étudiées. Dans l’horizon de
surface 0-2 cm, 71% du complexe absorbant est saturé, le matériau est
magnésique. A 30 cm de profondeur le taux de saturation est de l’ordre de 20%, le
complexe est relativement déssaturé.
Aucune différence significative de concentration en phosphore n’a été
observée entre les sols des forêts et ceux des pâturages.
IV.2.5.d Effets du brûlis sur les propriétés chimiques du sol
Le Feu stimule la décomposition biologique et rend bioassimilables des éléments nutritifs
essentiels comme l’azote (N), le soufre (S) et le phosphore (P). Le feu augmente
également la disponibilité des cations échangeables comme le calcium (Ca), le
magnésium (Mg) et le potassium (K) en transformant la matière végétale en cendres.
Ces apports nutritifs peuvent engendrer un foisonnement d’éléments nutritifs. Une
augmentation des cations entraîne une élévation du pH de la couverture morte, laquelle
peut être considérée comme bénéfique puisqu’elle fait augmenter le pouvoir tampon des
sols qui, à son tour, empêche les changements radicaux de l’acidité, laquelle peut nuire à
la croissance des végétaux. Cependant des éléments tels que le carbone, l'azote, le
soufre, et une partie du phosphore sont perdus par volatilisation ou dans les cendres
emportées par le vent.
En conclusion, la transformation de la forêt en pâturage par l’agriculture
familiale provoque dans notre site d’étude, comme dans de nombreux contextes
amazoniens, une évolution des propriétés physiques et chimiques du sol. Alors que
les pâturages mettent en évidence une fertilité chimique qui tend vers une
amélioration, une compaction du sol dont l’intensité et l’étendue est variable d’un
pâturage à l’autre, s’observe simultanément. La compaction de ces sols s’explique
par les pratiques agricoles tel le déboisement mécanique ou encore le surpâturage.
Les sols de pâturage apparaissent également plus sensibles à l’érosion que
ceux sous forêts et par conséquent pourraient être sujet à une dégradation
physique plus rapide suivant le degré d’anthropisation.


Etude des termitières du genre Cornitermes. Chapitre IV.2
83
V. Etude des termitières du genre Cornitermes.
Les termitières épigées de Cornitermes sp. représentent dans les pâturages, les
structures biogéniques les plus remarquables. De très grande taille (pouvant atteindre
1,5 m de hauteur), ces édifices sont en interactions avec le sol. Ce chapitre traite des
interactions sol-termitosphère (écosystème termitière).
Dans un premier temps, après avoir mis en évidence le rôle de la déforestation
sur l’activité des termites Cornitermes, les effets des termitières sur les sols ont été
abordés d’une part, par l’étude de la densité et de la distribution spatiale des édifices et
d’autre part, par la description des propriétés physiques et chimiques de ces structures
biogéniques.
Dans un second temps, afin de mieux comprendre la dégradation des termitières
de Cornitermes sp, les effets des pratiques agricoles (brûlis) ont été analysés.

Termitières du genre Cornitermes, matériel et méthodes. Chapitre V.1
84
V. 1 MATERIEL ET METHODES
V.1.1 Le termite du genre Cornitermes
Les termites sont des insectes sociaux de l’embranchement des isoptères. Le
Brésil recèle une des plus fortes diversités de termites avec près de 300 espèces connues
(Constantino and Acioli, 2006).
Le termite sur lequel nous avons travaillé, du genre Cornitermes fait partie de la
famille des Termitidae. Il fut décrit par Wasmann en 1897. Ce genre est typique des pays
d’Amérique du sud (Brésil, Paraguay, Bolivie, Pérou, Guyane) et d’Amérique centrale., Il
aurait comme ancêtre le genre Syntermes (Grassé, 1984c), mais des études de
génétique moléculaire devront confirmer cette affiliation. Le termite du genre
Cornitermes étudié est un termite xylophage (Amelung et al., 2002).
Ce genre néotropical construit des nids de très grande taille, caractéristiques des
vastes espaces libres du Brésil. Très répandu dans les lieux herbeux et les anciennes
plantations, il apprécie les terres contenant une forte proportion d’argile.
Il parcourt un cycle de développement qui le fait passer d’une situation hypogée
à un état presque entièrement épigé, tandis que la structure de son nid subit des
remaniements constants.
Le nid hypogé est de forme ovoïde reposant sur son pôle postérieur. Sa paroi ou
muraille est continue. Les communications avec l’extérieur se font par des galeries qui
partent de la région inférieure de la termitière. Les parois des chambres sont revêtues
d’un enduit noir, d’origine stercorale (fécès), durci avec le temps. La zone centrale est
presque sphérique avec une structure alvéolaire : le feuilleté, constitué de fécès
agglomérés. Le nid souterrain s’accroît surtout par sa partie supérieure en refoulant vers
le haut la terre qui le surmonte formant ainsi un dôme terreux.
Le nid épigé correspond à l’état terminal même s’il continue à grandir. Le dôme
terreux du stade précédent est intégré dans la muraille du nid (Grassé, 1984a).
Au sommet du nid, la paroi est percée d’orifices de ventilation pouvant faire penser à
l’espèce Cornitermes weberi décrite par Gonçalves et d'Araujo en 1962 (illustration1) ou
Cornitermes ovatus (Emerson, 1954 ; Imbrozio Barbosa et al., 2005). L’espèce ici
étudiée est sans doute affiliée à Cornitermes ovatus. Une étude phylogénétique serait
nécessaire pour déterminer précisément l’affiliation de ce termite. Par conséquent dans la
suite du travail, nous garderons le terme de Cornitermes sp pour le désigner.

Termitières du genre Cornitermes, matériel et méthodes. Chapitre V.1
85
Dans notre site d’étude, les termitières sont épigées, construites la plupart du
temps en incluant un tronc mort ou de grosses branches de bois brûlé. Ces édifices sont
généralement de grande taille: jusqu’à 1,30 m de la base souterraine au sommet sur
environ 1m de diamètre.
La muraille d’au moins 20 cm d’épaisseur est fabriquée à partir d’éléments
minéraux sous forme de boulettes prélevées dans le sol et mélangées à la salive. Au
centre de larges galeries et cavités parcourent la périphérie d’une grande chambre
remplie d’un feuilleté brunâtre composé essentiellement de matière organique d’origine
stercorale (illustration1).
Soldats Cornitermes sp.
Muraille
Zone feuilletée
1 m
1 m
Soldats Cornitermes sp.
Muraille
Zone feuilletée
1 m
1 m
Illustration 1 : Termitières et termites soldats du genre Cornitermes sp.
V.1.2 Etude des termitières de Cornitermes sp.
Trois études ont été menées sur les termitières de Cornitermes sp :

Termitières du genre Cornitermes, matériel et méthodes. Chapitre V.1
86
1) Cartographie des nids de Cornitermes sp.
La cartographie des termitières permet tout en informant sur la densité et la
distribution spatiale des édifices, de mettre en évidence l’influence, au niveau parcellaire,
du type de sol sur les nids de Cornitermes sp.
Dans trois pâturages âgés de 5 à 6 ans (P01, P02, P05), toutes les termitières ont
été référencées géographiquement à l’aide d’un GPS. Plusieurs points de la circonférence
de chaque parcelle ont été callés par plusieurs relevés GPS. Malgré les erreurs induites
par ce système (1 à 5m selon notice du constructeur), la taille des parcelles (de 2,8 ha à
16,7 ha) et le calage des bordures nous assurent une précision suffisante pour étudier la
distribution spatiale des termitières.
Les détails des traitements statistiques (méthode de Ripley) ont été développés
dans le chapitre III.1.
2) Analyses des caractéristiques physiques et chimiques des termitières de
Cornitermes sp.
Cette étude est divisée en deux parties :
- analyses physico-chimiques du feuilleté interne
- analyses physico-chimiques de la muraille externe ou périphérique.
L’influence du sol au niveau local sur les termitières a également été étudiée par
la comparaison des propriétés physico-chimiques de la muraille et du feuilleté des nids
avec celles du sol témoin. Cette étude a porté sur la texture, la densité de motte, la
stabilité structurale, le pH, les concentrations en carbone, en cations (aluminium,
calcium, magnésium, potassium) et en phosphore assimilable.
La description détaillée des techniques utilisées pour réaliser les analyses physico-
chimiques a été présentée au chapitre III.2.

Termitières du genre Cornitermes, matériel et méthodes. Chapitre V.1
87
3) Expérience de simulation de l’impact du feu sur les termitières.
Afin d’essayer de contrôler l’expansion de cette espèce et de limiter ainsi la
surface de pâturage « perdue » par leur implantation, les paysans brûlent les termitières.
En raison du coût financier et de l’effort nécessaire, cette pratique est occasionnelle.
Tous les ans cependant, la culture sur brûlis, pratiquée dans les pâturages du site
d’étude, implique un contact annuel entre le feu et les termitières.
Une analyse de l’impact du feu sur cette structure biogénique a été entreprise.
Pour se faire une expérience a été réalisée dans trois pâturages :
Trois termitières vivantes de Cornitermes sp. ont été choisies dans ces 3
pâturages (P01, P02, P04). Elles ont été cassées et brûlées avec du bois et du kérosène,
selon la méthode utilisée par les agriculteurs locaux. Des prélèvements de muraille et de
feuilleté ont été effectués avant et après calcination (illustration 2).
Illustration 2: Expérience de brûlage des termitières.
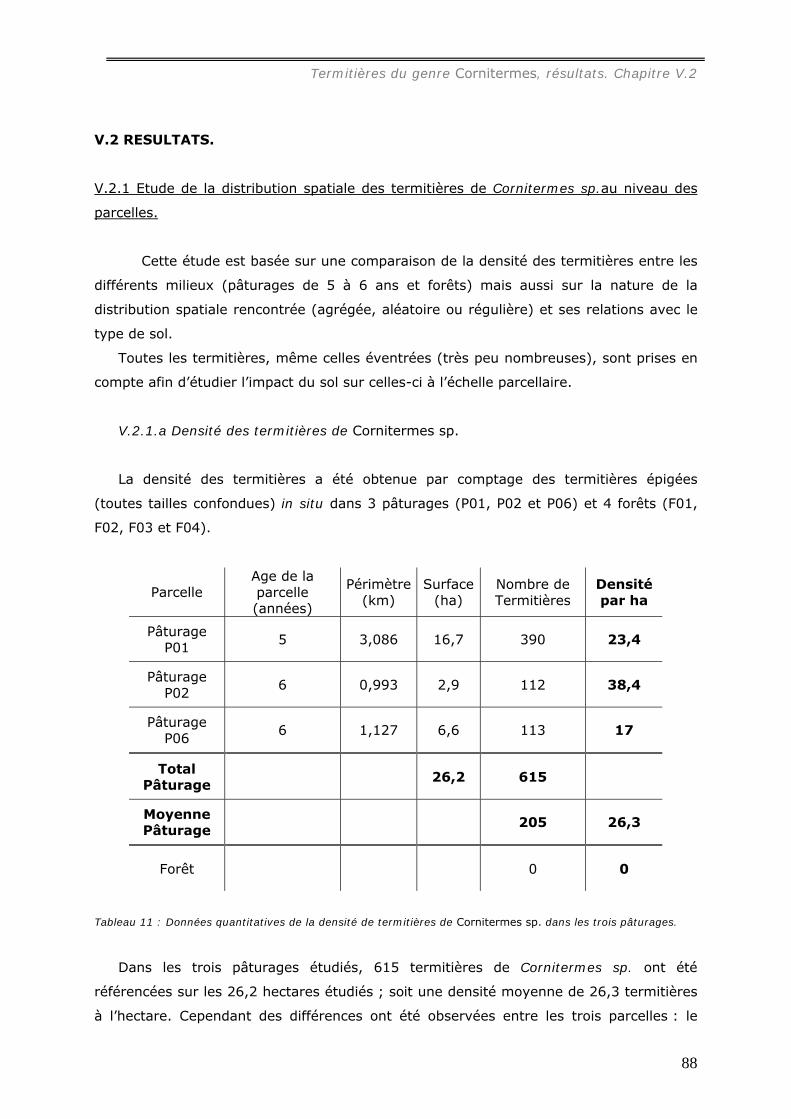
Termitières du genre Cornitermes, résultats. Chapitre V.2
88
V.2 RESULTATS.
V.2.1 Etude de la distribution spatiale des termitières de Cornitermes sp.au niveau des
parcelles.
Cette étude est basée sur une comparaison de la densité des termitières entre les
différents milieux (pâturages de 5 à 6 ans et forêts) mais aussi sur la nature de la
distribution spatiale rencontrée (agrégée, aléatoire ou régulière) et ses relations avec le
type de sol.
Toutes les termitières, même celles éventrées (très peu nombreuses), sont prises en
compte afin d’étudier l’impact du sol sur celles-ci à l’échelle parcellaire.
V.2.1.a Densité des termitières de Cornitermes sp.
La densité des termitières a été obtenue par comptage des termitières épigées
(toutes tailles confondues) in situ dans 3 pâturages (P01, P02 et P06) et 4 forêts (F01,
F02, F03 et F04).
Parcelle Age de la parcelle (années)
Périmètre (km)
Surface (ha)
Nombre de Termitières
Densité par ha
Pâturage P01
5 3,086 16,7 390 23,4
Pâturage P02
6 0,993 2,9 112 38,4
Pâturage P06
6 1,127 6,6 113 17
Total Pâturage
26,2 615
Moyenne Pâturage
205 26,3
Forêt 0 0
Tableau 11 : Données quantitatives de la densité de termitières de Cornitermes sp. dans les trois pâturages.
Dans les trois pâturages étudiés, 615 termitières de Cornitermes sp. ont été
référencées sur les 26,2 hectares étudiés ; soit une densité moyenne de 26,3 termitières
à l’hectare. Cependant des différences ont été observées entre les trois parcelles : le

Termitières du genre Cornitermes, résultats. Chapitre V.2
89
pâturage P02, de surface plus petite (2,9 ha), présente une densité de 38,4 termitières à
l’hectare alors que dans le pâturage P06 (6,6 ha) la densité des nids n’est que de 17 à
l’hectare (Tableau 11, Figure 20).
Dans l’ensemble des forêts étudiées, représentant une surface de prospection
supérieure à 30 ha, aucune termitière épigée de Cornitermes sp. n’a pu être observée.
P01
P02 P05
P01
P02 P05
Figure 20 : Disposition des termitières dans les 3 pâturages (P01, P02, P06). A chaque carré correspond une
termitière de taille et d’âge non précisés.
La distance minimale entre deux termitières épigées dans les pâturages est de 2,6 m.
V.2.1.b Fréquence en classes de taille des termitières de Cornitermes sp.
Les termitières épigées des pâturages ont été classées en trois classes de hauteur :
-Classe des termitières de grande taille (hauteur > 1m)
-Classe des termitières de taille moyenne (60cm < hauteur < 1m)
-Classe des termitières de petite taille (10 cm < hauteur < 60 cm)
Nous n’avons pas observé de termitières dont la hauteur était inférieure à 10 cm.
La fréquence des 3 classes de termitières, (grande, moyenne et petite) basée sur
la hauteur de la termitière, a été observée dans deux parcelles (P02 et P06).
P06
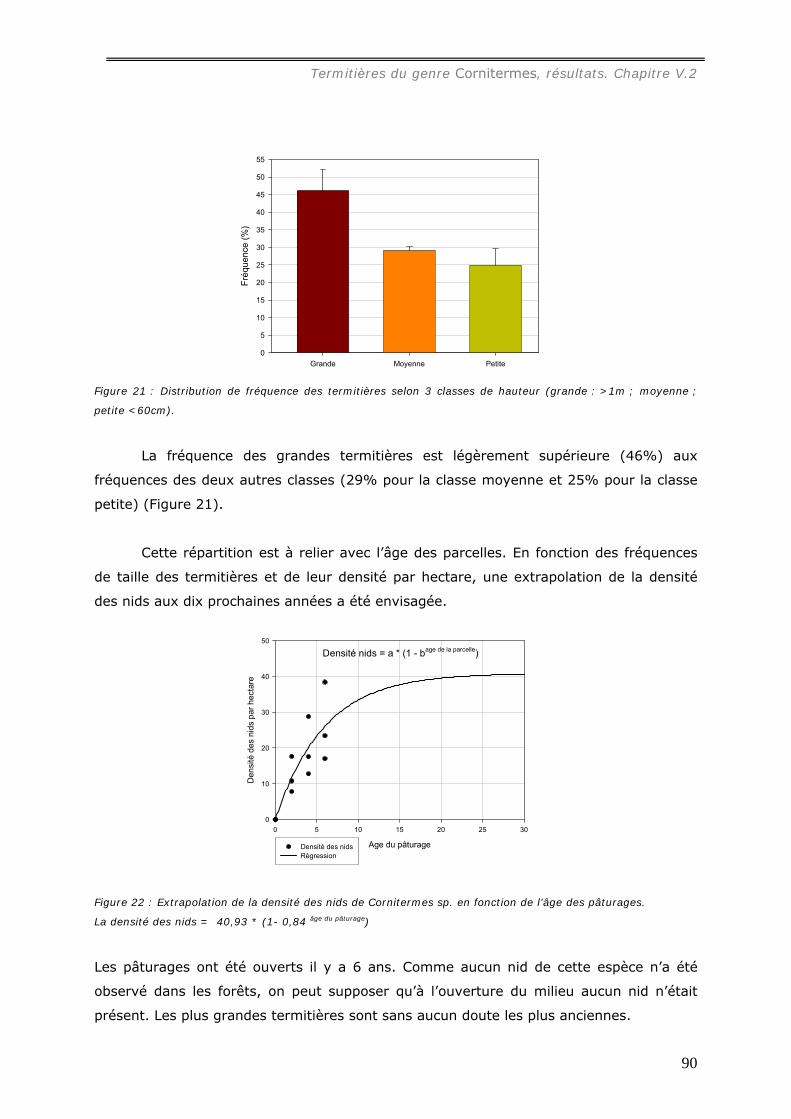
Termitières du genre Cornitermes, résultats. Chapitre V.2
90
Grande Moyenne Petite
Fréq
uenc
e (%
)
0
5
10
15
20
25
30
35
40
45
50
55
Figure 21 : Distribution de fréquence des termitières selon 3 classes de hauteur (grande : >1m ; moyenne ;
petite <60cm).
La fréquence des grandes termitières est légèrement supérieure (46%) aux
fréquences des deux autres classes (29% pour la classe moyenne et 25% pour la classe
petite) (Figure 21).
Cette répartition est à relier avec l’âge des parcelles. En fonction des fréquences
de taille des termitières et de leur densité par hectare, une extrapolation de la densité
des nids aux dix prochaines années a été envisagée.
Densité nids = a * (1 - bage de la parcelle)
Age du pâturage
0 5 10 15 20 25 30
Den
sité
des
nid
s pa
r hec
tare
0
10
20
30
40
50
Densité des nidsRégression
Figure 22 : Extrapolation de la densité des nids de Cornitermes sp. en fonction de l’âge des pâturages.
La densité des nids = 40,93 * (1- 0,84 âge du pâturage)
Les pâturages ont été ouverts il y a 6 ans. Comme aucun nid de cette espèce n’a été
observé dans les forêts, on peut supposer qu’à l’ouverture du milieu aucun nid n’était
présent. Les plus grandes termitières sont sans aucun doute les plus anciennes.
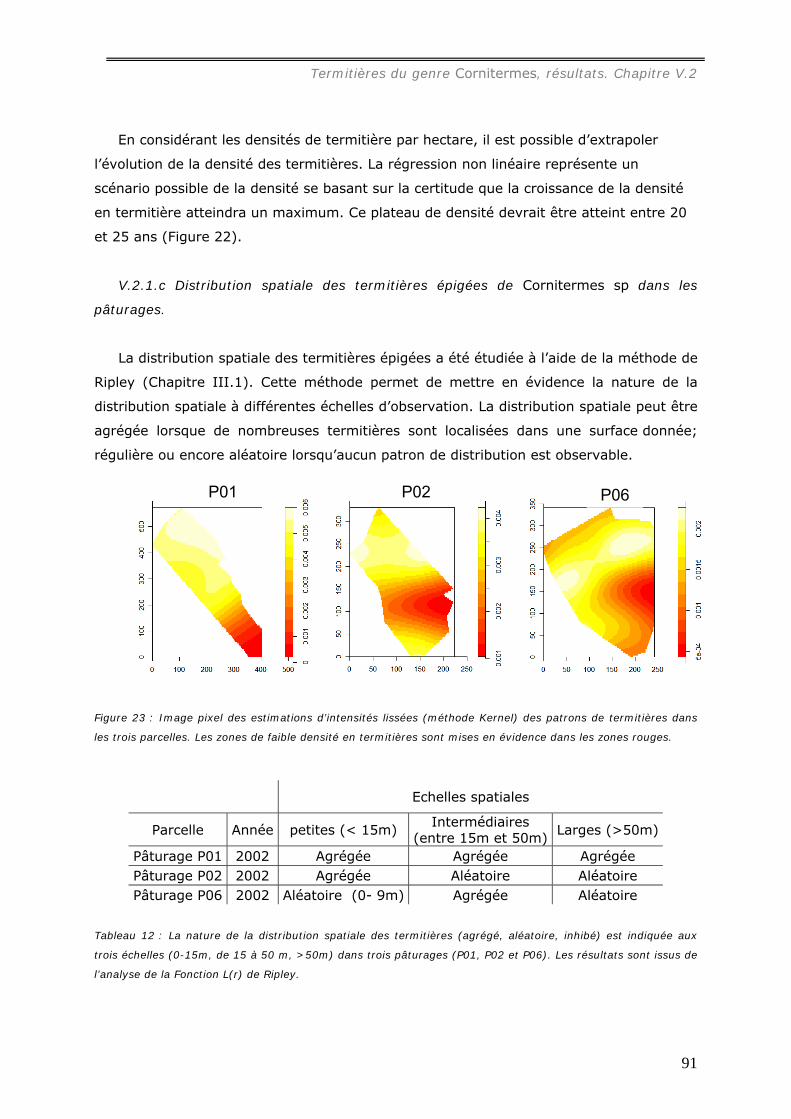
Termitières du genre Cornitermes, résultats. Chapitre V.2
91
En considérant les densités de termitière par hectare, il est possible d’extrapoler
l’évolution de la densité des termitières. La régression non linéaire représente un
scénario possible de la densité se basant sur la certitude que la croissance de la densité
en termitière atteindra un maximum. Ce plateau de densité devrait être atteint entre 20
et 25 ans (Figure 22).
V.2.1.c Distribution spatiale des termitières épigées de Cornitermes sp dans les
pâturages.
La distribution spatiale des termitières épigées a été étudiée à l’aide de la méthode de
Ripley (Chapitre III.1). Cette méthode permet de mettre en évidence la nature de la
distribution spatiale à différentes échelles d’observation. La distribution spatiale peut être
agrégée lorsque de nombreuses termitières sont localisées dans une surface donnée;
régulière ou encore aléatoire lorsqu’aucun patron de distribution est observable.
P06P02P01 P06P06P02P02P01P01
Figure 23 : Image pixel des estimations d’intensités lissées (méthode Kernel) des patrons de termitières dans
les trois parcelles. Les zones de faible densité en termitières sont mises en évidence dans les zones rouges.
Tableau 12 : La nature de la distribution spatiale des termitières (agrégé, aléatoire, inhibé) est indiquée aux
trois échelles (0-15m, de 15 à 50 m, >50m) dans trois pâturages (P01, P02 et P06). Les résultats sont issus de
l’analyse de la Fonction L(r) de Ripley.
Echelles spatiales
Parcelle Année petites (< 15m) Intermédiaires
(entre 15m et 50m) Larges (>50m)
Pâturage P01 2002 Agrégée Agrégée Agrégée Pâturage P02 2002 Agrégée Aléatoire Aléatoire Pâturage P06 2002 Aléatoire (0- 9m) Agrégée Aléatoire
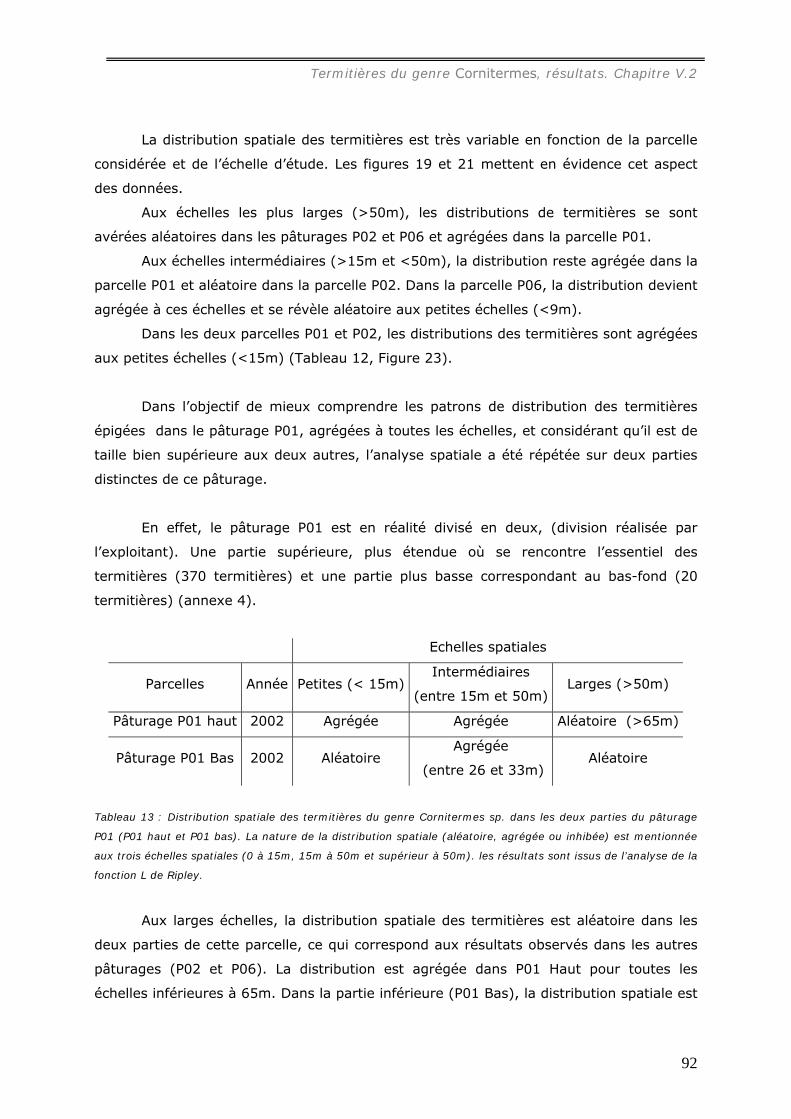
Termitières du genre Cornitermes, résultats. Chapitre V.2
92
La distribution spatiale des termitières est très variable en fonction de la parcelle
considérée et de l’échelle d’étude. Les figures 19 et 21 mettent en évidence cet aspect
des données.
Aux échelles les plus larges (>50m), les distributions de termitières se sont
avérées aléatoires dans les pâturages P02 et P06 et agrégées dans la parcelle P01.
Aux échelles intermédiaires (>15m et <50m), la distribution reste agrégée dans la
parcelle P01 et aléatoire dans la parcelle P02. Dans la parcelle P06, la distribution devient
agrégée à ces échelles et se révèle aléatoire aux petites échelles (<9m).
Dans les deux parcelles P01 et P02, les distributions des termitières sont agrégées
aux petites échelles (<15m) (Tableau 12, Figure 23).
Dans l’objectif de mieux comprendre les patrons de distribution des termitières
épigées dans le pâturage P01, agrégées à toutes les échelles, et considérant qu’il est de
taille bien supérieure aux deux autres, l’analyse spatiale a été répétée sur deux parties
distinctes de ce pâturage.
En effet, le pâturage P01 est en réalité divisé en deux, (division réalisée par
l’exploitant). Une partie supérieure, plus étendue où se rencontre l’essentiel des
termitières (370 termitières) et une partie plus basse correspondant au bas-fond (20
termitières) (annexe 4).
Tableau 13 : Distribution spatiale des termitières du genre Cornitermes sp. dans les deux parties du pâturage
P01 (P01 haut et P01 bas). La nature de la distribution spatiale (aléatoire, agrégée ou inhibée) est mentionnée
aux trois échelles spatiales (0 à 15m, 15m à 50m et supérieur à 50m). les résultats sont issus de l’analyse de la
fonction L de Ripley.
Aux larges échelles, la distribution spatiale des termitières est aléatoire dans les
deux parties de cette parcelle, ce qui correspond aux résultats observés dans les autres
pâturages (P02 et P06). La distribution est agrégée dans P01 Haut pour toutes les
échelles inférieures à 65m. Dans la partie inférieure (P01 Bas), la distribution spatiale est
Echelles spatiales
Parcelles Année Petites (< 15m) Intermédiaires
(entre 15m et 50m) Larges (>50m)
Pâturage P01 haut 2002 Agrégée Agrégée Aléatoire (>65m)
Pâturage P01 Bas 2002 Aléatoire Agrégée
(entre 26 et 33m) Aléatoire

Termitières du genre Cornitermes, résultats. Chapitre V.2
93
apparue agrégée aux échelles comprises entre 26 m et 33 m mais le faible nombre de
termitières dans cette partie incite à considérer ce résultat avec circonspection (Tableau
13).
La densité des termitières épigées de Cornitermes sp, toute taille confondue,
est de 26,3 nids par hectare dans les pâturages.
Aucune termitière épigée de cette espèce n’a pu être observée dans les
forêts (surface de prospection supérieure à 30 ha).
L’extrapolation des densités de nids met en évidence que dans les pâturages
de 6 ans les Cornitermes sont dans une phase d’expansion. Le maximum de densité
devrait être atteint au bout de 20 à 25 ans.
Aux échelles d’observation supérieures à 50 m, la distribution spatiale des
termitières épigées de Cornitermes sp. s’est avérée systématiquement aléatoire
dans tous les pâturages étudiés. A ces échelles, la distribution spatiale des
termitières ne présentent pas de patron de distribution significatif.
Aux échelles intermédiaires (comprises entre 15m et 50m), la distribution
spatiale des termitières épigées est variable en fonction de la parcelle considérée et
de l’échelle d’étude.
Aux petites échelles (<15m), dans toutes les parcelles (excepté P06), la
distribution spatiale des termitières est agrégée. Cette agrégation s’explique sans
doute par l’essaimage des sexués ailés autour des nids mères.
V.2.1.d Relation avec le type de sol.
La relation entre les densités de termitières dans les trois parcelles et le sol est
illustrée par la comparaison des cartes de densité lissée (méthode Kernel) de termitières
avec une représentation tridimensionnelle de l’altitude des parcelles.
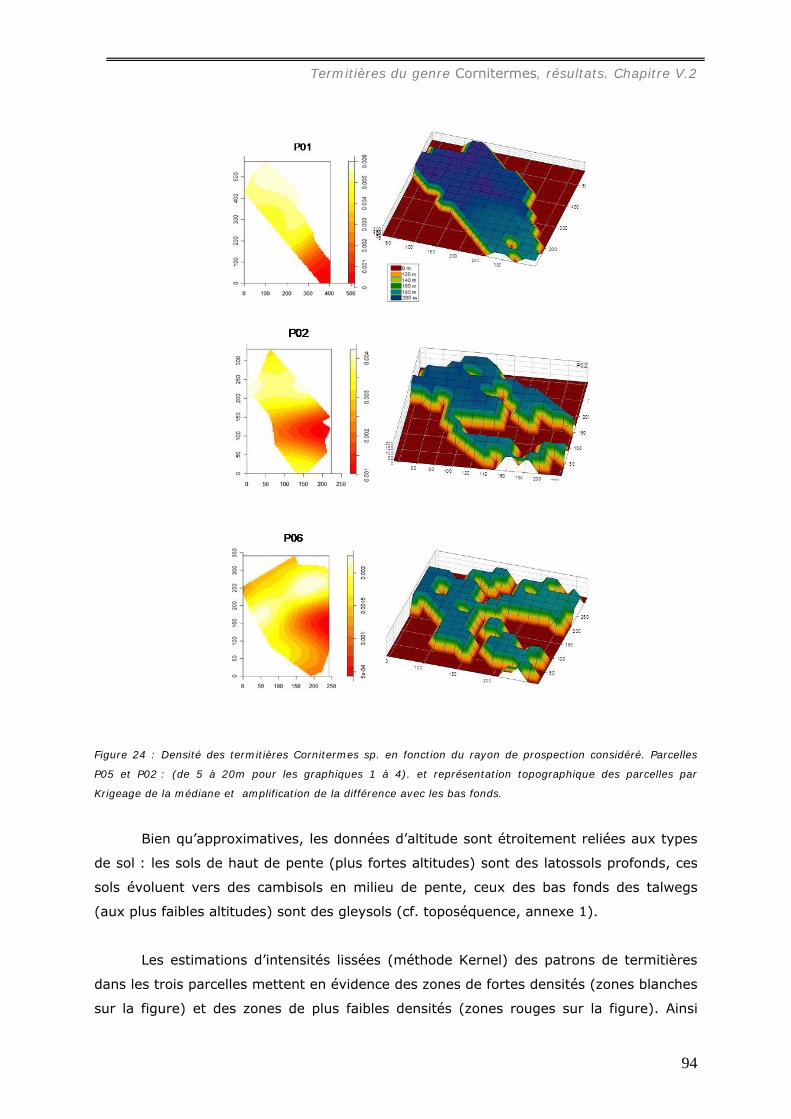
Termitières du genre Cornitermes, résultats. Chapitre V.2
94
Figure 24 : Densité des termitières Cornitermes sp. en fonction du rayon de prospection considéré. Parcelles
P05 et P02 : (de 5 à 20m pour les graphiques 1 à 4). et représentation topographique des parcelles par
Krigeage de la médiane et amplification de la différence avec les bas fonds.
Bien qu’approximatives, les données d’altitude sont étroitement reliées aux types
de sol : les sols de haut de pente (plus fortes altitudes) sont des latossols profonds, ces
sols évoluent vers des cambisols en milieu de pente, ceux des bas fonds des talwegs
(aux plus faibles altitudes) sont des gleysols (cf. toposéquence, annexe 1).
Les estimations d’intensités lissées (méthode Kernel) des patrons de termitières
dans les trois parcelles mettent en évidence des zones de fortes densités (zones blanches
sur la figure) et des zones de plus faibles densités (zones rouges sur la figure). Ainsi

Termitières du genre Cornitermes, résultats. Chapitre V.2
95
dans les 3 pâturages, les taches de fortes densités se situent systématiquement dans des
zones correspondant aux latosols profonds dans le haut de la toposéquence (annexe 1)
alors que les zones peu ou pas occupées par les termitières se situent dans les gleysols
caractéristiques des zones basses humides du site d’étude (Figure 24).
La distribution spatiale des termitières est reliée à l’altitude. La toposéquence
(annexe 1) mettant en évidence la relation entre l’altitude et le type de sol, nous
permet de déduire que les termites s’installent préférentiellement dans les zones
peu humides, et fertiles des latosols profonds (préférés également par les
agriculteurs pour y établir leurs cultures) que dans les gleysols humides des bas-
fonds.
Aucune termitière n’a été observée dans les zones de bas fond caractérisées
par les gleysols.
Entre ces deux types de sol, la densité des nids est variable.
V.2.2 Etude des caractéristiques physico-chimiques des termitières de Cornitermes sp.
La comparaison des propriétés physiques et chimiques des termitières avec celles
des sols témoins du site d’étude a permis de mettre en évidence les interactions locales
entre les termitières et le sol environnant.
Considérant les densités et les distributions spatiales des nids observés
précédemment, une extrapolation des stocks en matière organique et en ions au niveau
parcellaire a été réalisée.
Les termitières sont constituées de deux parties :
• le feuilleté interne
• la muraille ou paroi externe
V.2.2.a Quantification et propriétés du feuilleté interne du nid.
Cette étude a consisté à mettre en relation les classes des termitières (classe de
hauteur) et la masse humide de feuilleté (matière fraîche) présent dans la cavité
centrale de celles-ci.
Pour rappel, la classe des grandes termitières représente les nids dont la hauteur
est supérieure à 1 m, la classe des termitières moyennes ont une hauteur comprise entre
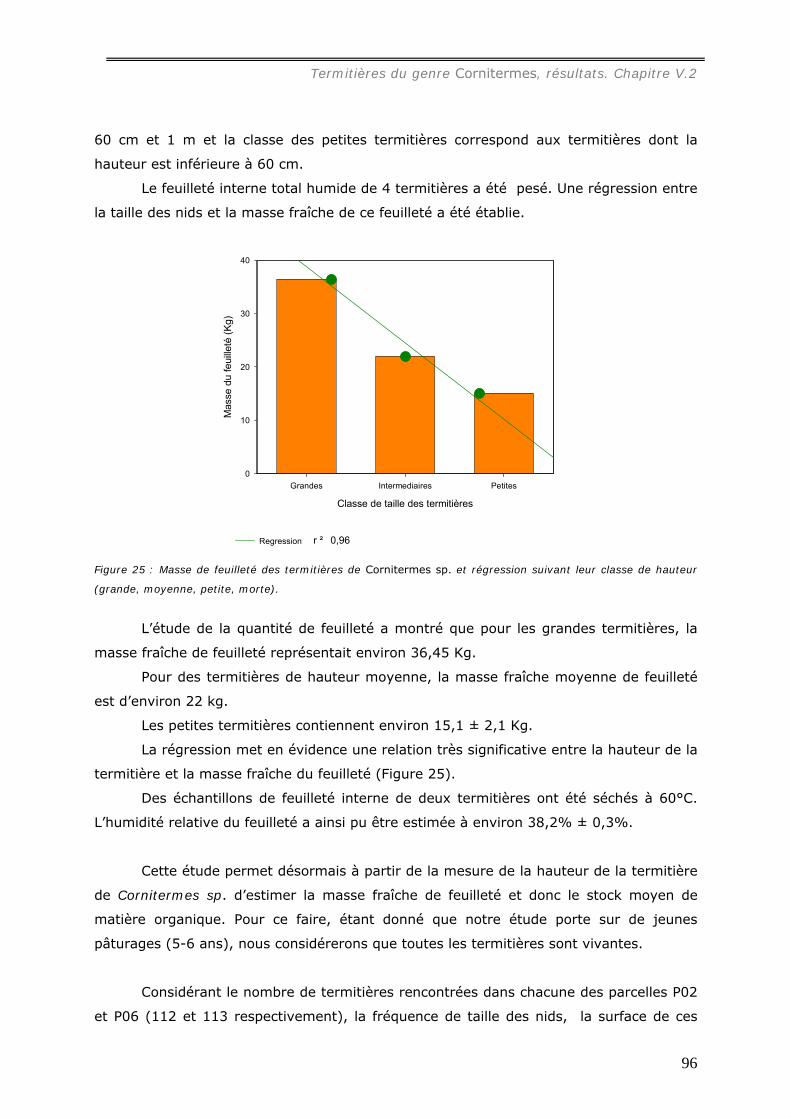
Termitières du genre Cornitermes, résultats. Chapitre V.2
96
60 cm et 1 m et la classe des petites termitières correspond aux termitières dont la
hauteur est inférieure à 60 cm.
Le feuilleté interne total humide de 4 termitières a été pesé. Une régression entre
la taille des nids et la masse fraîche de ce feuilleté a été établie.
Classe de taille des termitières
Grandes Intermediaires Petites
Mas
se d
u fe
uille
té (K
g)
0
10
20
30
40
r ² 0,96Regression
Figure 25 : Masse de feuilleté des termitières de Cornitermes sp. et régression suivant leur classe de hauteur
(grande, moyenne, petite, morte).
L’étude de la quantité de feuilleté a montré que pour les grandes termitières, la
masse fraîche de feuilleté représentait environ 36,45 Kg.
Pour des termitières de hauteur moyenne, la masse fraîche moyenne de feuilleté
est d’environ 22 kg.
Les petites termitières contiennent environ 15,1 ± 2,1 Kg.
La régression met en évidence une relation très significative entre la hauteur de la
termitière et la masse fraîche du feuilleté (Figure 25).
Des échantillons de feuilleté interne de deux termitières ont été séchés à 60°C.
L’humidité relative du feuilleté a ainsi pu être estimée à environ 38,2% ± 0,3%.
Cette étude permet désormais à partir de la mesure de la hauteur de la termitière
de Cornitermes sp. d’estimer la masse fraîche de feuilleté et donc le stock moyen de
matière organique. Pour ce faire, étant donné que notre étude porte sur de jeunes
pâturages (5-6 ans), nous considérerons que toutes les termitières sont vivantes.
Considérant le nombre de termitières rencontrées dans chacune des parcelles P02
et P06 (112 et 113 respectivement), la fréquence de taille des nids, la surface de ces

Termitières du genre Cornitermes, résultats. Chapitre V.2
97
parcelles (P02 : 2,9 ha et P06 : 6,6 ha), la masse fraîche moyenne de feuilleté par
parcelle a été estimée en réalisant le calcul suivant :
(46% de gros nids x 113 nids x 36,45 kg par nid) + (29% de nids intermédiaires x 113
nids x 22 kg par nid) + (25% de petits nids x 113 nids x 15,1 kg par nid) = 3.04 T
L’humidité relative étant de 38%, les 3,04 T de feuilleté frais par parcelle
représentent une masse sèche d’environ 1,9 T dans chaque parcelle.
Ainsi dans le pâturage P02, de 2,9 ha, le stock moyen de feuilleté interne sec estimé est
de 655 kg.ha-1 alors que dans P06 (6,6 ha), il est d’environ 288 kg.ha-1.
Le même calcul a aussi été appliqué pour la parcelle P01 totale :
(46% de gros nids x 390 nids x 36,45 kg par nid) + (29% de nids intermédiaires x 390
nids x 22 kg par nid) + (25% de petits nids x 390 nids x 15,1 kg par nid) = 10,5T de
feuilleté humide soit 6,5T de feuilleté sec.
La surface du pâturage P01 est de 16,7 ha donc le stock de feuilleté sec dans ce
pâturage est d’environ 390 kg.ha-1.
Le feuilleté sec des termitières de Cornitermes sp est composé en moyenne de
38,4 % de carbone (méthode Walkley et Black) soit environ 77% de matière organique
et donc par différence de 23% de matière minérale.
Le stock de matière organique piégé au sein des termitières sous forme de
feuilleté varie donc de 221 kg.ha-1 à 503 kg.ha-1 en fonction de la densité en termitière et
la surface du pâturage.
Le feuilleté est hydrophobe. Il est par ailleurs très friable sous pression manuelle.
La masse fraîche de feuilleté interne est étroitement corrélée à la hauteur
des termitières épigées de Cornitermes sp. dans les pâturages. L’humidité relative
du feuilleté a pu être estimée à environ 38,2% ± 0,3%.
La masse de feuilleté interne sec au sein des nids épigés de Cornitermes sp.
dans les pâturages est compris entre 288 kg.ha-1 et 655 kg.ha-1.
Le feuilleté sec des termitières épigées de Cornitermes sp est composé en
moyenne de 38,4 % de carbone soit environ 77% de matière organique et donc
23% d’éléments minéraux.
Le stock de matière organique piégé au sein du feuilleté interne des
termitières épigées de Cornitermes sp. dans les pâturages a été estimé variant de
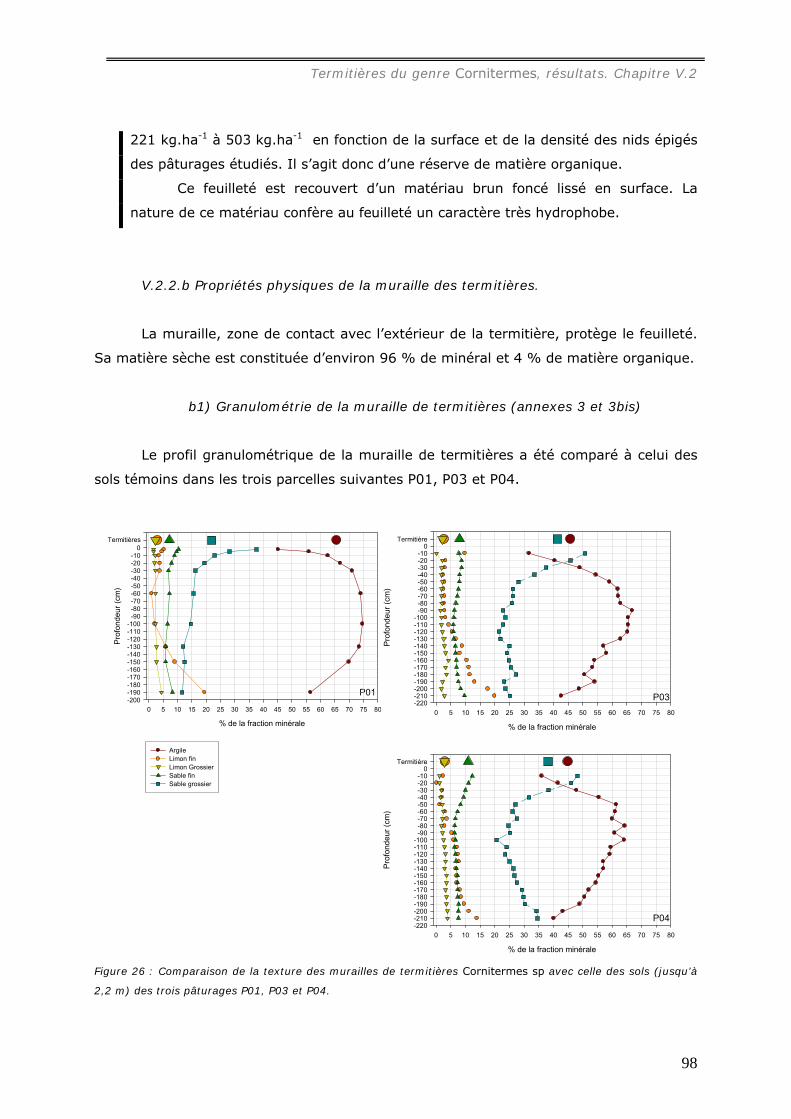
Termitières du genre Cornitermes, résultats. Chapitre V.2
98
221 kg.ha-1 à 503 kg.ha-1 en fonction de la surface et de la densité des nids épigés
des pâturages étudiés. Il s’agit donc d’une réserve de matière organique.
Ce feuilleté est recouvert d’un matériau brun foncé lissé en surface. La
nature de ce matériau confère au feuilleté un caractère très hydrophobe.
V.2.2.b Propriétés physiques de la muraille des termitières.
La muraille, zone de contact avec l’extérieur de la termitière, protège le feuilleté.
Sa matière sèche est constituée d’environ 96 % de minéral et 4 % de matière organique.
b1) Granulométrie de la muraille de termitières (annexes 3 et 3bis)
Le profil granulométrique de la muraille de termitières a été comparé à celui des
sols témoins dans les trois parcelles suivantes P01, P03 et P04.
P01
% de la fraction minérale
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80
Pro
fond
eur (
cm)
-200-190-180-170-160-150-140-130-120-110-100
-90-80-70-60-50-40-30-20-10
01020
ArgileLimon finLimon GrossierSable finSable grossier
Termitières
P03
% de la fraction minérale
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80
Prof
onde
ur (c
m)
-220-210-200-190-180-170-160-150-140-130-120-110-100
-90-80-70-60-50-40-30-20-10
0Termitière
P04
% de la fraction minérale
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80
Pro
fond
eur (
cm)
-220-210-200-190-180-170-160-150-140-130-120-110-100
-90-80-70-60-50-40-30-20-10
0Termitière
P01
% de la fraction minérale
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80
Pro
fond
eur (
cm)
-200-190-180-170-160-150-140-130-120-110-100
-90-80-70-60-50-40-30-20-10
01020
ArgileLimon finLimon GrossierSable finSable grossier
Termitières
P03
% de la fraction minérale
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80
Prof
onde
ur (c
m)
-220-210-200-190-180-170-160-150-140-130-120-110-100
-90-80-70-60-50-40-30-20-10
0Termitière
P04
% de la fraction minérale
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80
Pro
fond
eur (
cm)
-220-210-200-190-180-170-160-150-140-130-120-110-100
-90-80-70-60-50-40-30-20-10
0Termitière
Figure 26 : Comparaison de la texture des murailles de termitières Cornitermes sp avec celle des sols (jusqu’à
2,2 m) des trois pâturages P01, P03 et P04.

Termitières du genre Cornitermes, résultats. Chapitre V.2
99
Dans les trois parcelles étudiées, la texture des horizons de surface est argilo-
sableuse. Les murailles de termitières se caractérisent par une composition moyenne de
54% d’argile, 3% de limons fins, 2,3% de limons grossiers, 8,4% de sable fin et 32,3%
de sable grossier ; soit 57% d’éléments fins et 43% d’éléments grossiers. Ainsi ces
murailles sont essentiellement constituées d’un assemblage d’argile et de sables
grossiers lui conférant une texture argilo-sableuse (Figure 26).
La comparaison avec les profils granulométriques des sols témoins de ces trois
pâturages montre que l’horizon du sol 10-30 cm est celui qui se rapproche le plus des
murailles de termitières (Figure 26).
La texture des murailles de termitières est argilo-sableuse (diagramme des
textures), identique à celle des sols témoins.
Il apparaît donc que les matériaux de construction servant à fabriquer la
muraille des nids sont prélevés entre 10 cm et 30 cm de profondeur dans le sol.
b2) Densité de motte des agrégats de muraille des termitières épigées.
La mesure de la densité de mottes permet d’estimer l’état de compaction des
agrégats en ne prenant pas en compte la macroporosité inter-mottes des horizons des
sols. Ainsi cette étude a permis de comparer des agglomérats des murailles de tailles
pluricentimétriques à des mottes pluricentimétriques issues des horizons témoins
prélevées dans les 20 premiers centimètres.
La densité de motte des murailles des termitières (1,52 ± 0,21) ne diffère pas
significativement de celle du sol témoin (1,57 ± 0,08).
Les murailles des termitières, tout comme les horizons témoins des sols
des pâturages (0-30 cm), sont d’apparence dense et peu macroporeuse. Les valeurs
de densité apparente de mottes sont caractéristiques des structures compactes.
b3) Stabilité structurale de la muraille de termitières
L’étude de la stabilité structurale met en évidence la résistance de la
structure physique des agrégats soumis à divers processus physiques tel l’immersion
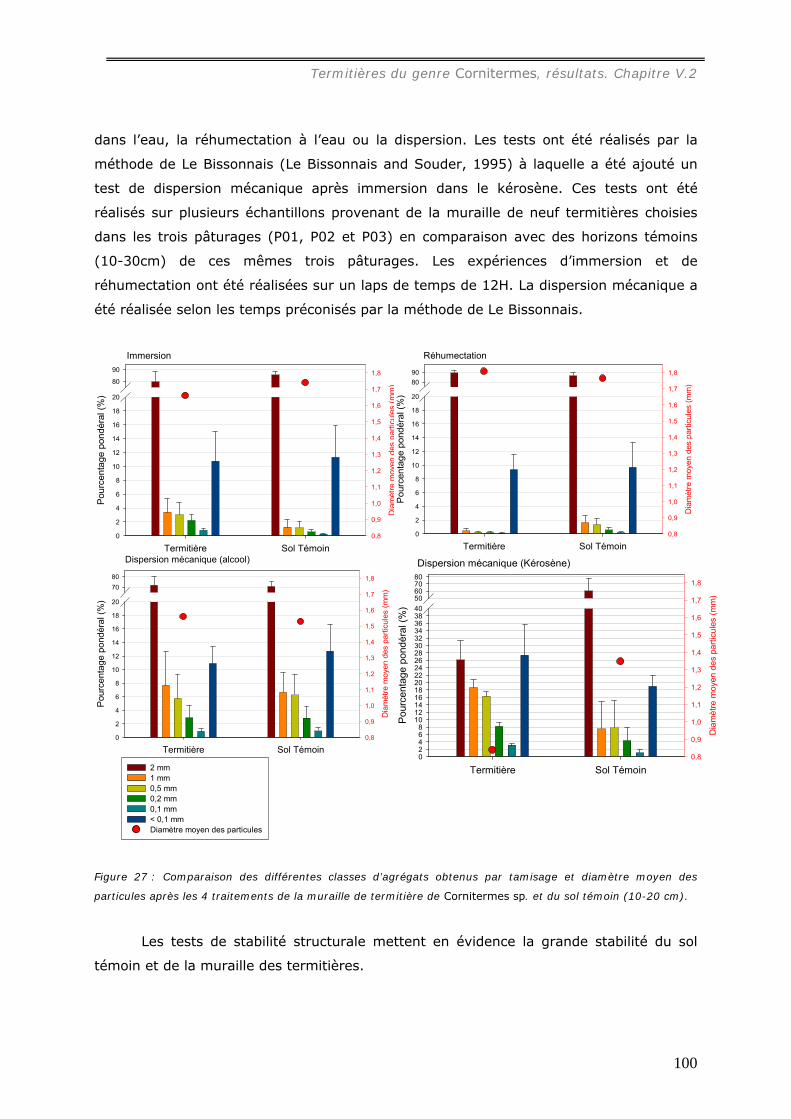
Termitières du genre Cornitermes, résultats. Chapitre V.2
100
dans l’eau, la réhumectation à l’eau ou la dispersion. Les tests ont été réalisés par la
méthode de Le Bissonnais (Le Bissonnais and Souder, 1995) à laquelle a été ajouté un
test de dispersion mécanique après immersion dans le kérosène. Ces tests ont été
réalisés sur plusieurs échantillons provenant de la muraille de neuf termitières choisies
dans les trois pâturages (P01, P02 et P03) en comparaison avec des horizons témoins
(10-30cm) de ces mêmes trois pâturages. Les expériences d’immersion et de
réhumectation ont été réalisées sur un laps de temps de 12H. La dispersion mécanique a
été réalisée selon les temps préconisés par la méthode de Le Bissonnais.
Immersion
Termitière Sol Témoin
Pou
rcen
tage
pon
déra
l (%
)
0
2
4
6
8
10
12
14
16
18
20
80
90
Dia
mèt
re m
oyen
des
par
ticul
es (m
m)
0,8
0,9
1,0
1,1
1,2
1,3
1,4
1,5
1,6
1,7
1,8
Réhumectation
Termitière Sol Témoin
Pou
rcen
tage
pon
déra
l (%
)
0
2
4
6
8
10
12
14
16
18
20
8090
Dia
mèt
re m
oyen
des
par
ticul
es (m
m)
0,8
0,9
1,0
1,1
1,2
1,3
1,4
1,5
1,6
1,7
1,8
Dispersion mécanique (alcool)
Termitière Sol Témoin
Pou
rcen
tage
pon
déra
l (%
)
0
2
4
6
8
10
12
14
16
18
20
7080
Dia
mèt
re m
oyen
des
par
ticul
es (m
m)
0,8
0,9
1,0
1,1
1,2
1,3
1,4
1,5
1,6
1,7
1,8
Dispersion mécanique (Kérosène)
Termitière Sol Témoin
Pou
rcen
tage
pon
déra
l (%
)
02468
1012141618202224262830323436384050607080
Dia
mèt
re m
oyen
des
par
ticul
es (m
m)
0,8
0,9
1,0
1,1
1,2
1,3
1,4
1,5
1,6
1,7
1,8
2 mm 1 mm 0,5 mm 0,2 mm 0,1 mm < 0,1 mm Diamètre moyen des particules
Immersion
Termitière Sol Témoin
Pou
rcen
tage
pon
déra
l (%
)
0
2
4
6
8
10
12
14
16
18
20
80
90
Dia
mèt
re m
oyen
des
par
ticul
es (m
m)
0,8
0,9
1,0
1,1
1,2
1,3
1,4
1,5
1,6
1,7
1,8
Réhumectation
Termitière Sol Témoin
Pou
rcen
tage
pon
déra
l (%
)
0
2
4
6
8
10
12
14
16
18
20
8090
Dia
mèt
re m
oyen
des
par
ticul
es (m
m)
0,8
0,9
1,0
1,1
1,2
1,3
1,4
1,5
1,6
1,7
1,8
Dispersion mécanique (alcool)
Termitière Sol Témoin
Pou
rcen
tage
pon
déra
l (%
)
0
2
4
6
8
10
12
14
16
18
20
7080
Dia
mèt
re m
oyen
des
par
ticul
es (m
m)
0,8
0,9
1,0
1,1
1,2
1,3
1,4
1,5
1,6
1,7
1,8
Dispersion mécanique (Kérosène)
Termitière Sol Témoin
Pou
rcen
tage
pon
déra
l (%
)
02468
1012141618202224262830323436384050607080
Dia
mèt
re m
oyen
des
par
ticul
es (m
m)
0,8
0,9
1,0
1,1
1,2
1,3
1,4
1,5
1,6
1,7
1,8
2 mm 1 mm 0,5 mm 0,2 mm 0,1 mm < 0,1 mm Diamètre moyen des particules
Figure 27 : Comparaison des différentes classes d’agrégats obtenus par tamisage et diamètre moyen des
particules après les 4 traitements de la muraille de termitière de Cornitermes sp. et du sol témoin (10-20 cm).
Les tests de stabilité structurale mettent en évidence la grande stabilité du sol
témoin et de la muraille des termitières.

Termitières du genre Cornitermes, résultats. Chapitre V.2
101
Les tests classiques d’immersion, de réhumectation, et de dispersion mécanique
dans l’alcool n’ont pas permis de dégrader la structure des murailles et des sols témoins
(de 70% à 90% de la structure sont conservés). Seules de fines particules < 1mm sont
libérées (de 8% à 12%).
Après les tests d’immersion et de réhumectation, le diamètre moyen des
agglomérats des murailles est compris entre 1,65 mm et 1,85 mm. Il ne diffère pas
significativement de ceux des horizons témoins (test t, p<0,001).
Le test de dispersion dans l’alcool met en évidence une diminution du diamètre
moyen des agglomérats des murailles et des témoins (1,55 mm).
Seul le test de dispersion mécanique dans le kérosène a permis de déstructurer
les agrégats des murailles de termitières. Les sols témoins sont restés relativement
stables (60% des agglomérats de la structure initiale sont conservés). La dégradation
des murailles s’est répartie dans toutes les classes avec une proportion supérieure pour
les agrégats conservés et les particules fines (> 2mm : 26% et < 1mm : 28%).
Le diamètre moyen des agglomérats résultant de ce test est de 0,85 ± 0,15 mm
pour les échantillons des murailles de termitières contre 1,35 ± 0,3 mm pour ceux des
sols témoins (Figure 27). Le kérosène a ici détruit les liens entre les agrégats. Ce lien
provenait sans doute de la salive ajoutée par les ouvriers termites lors de la construction
(Grassé, 1984a). Cependant ce résultat est à considérer avec précaution en raison de la
variabilité des valeurs des diamètres moyens des agglomérats.
Les tests de stabilité structurale mettent en évidence la grande stabilité des
sols témoins et des murailles de termitières. Après les tests classiques d’immersion,
de réhumectation et de dispersion mécanique après pré-traitement dans l’alcool,
70% à 90% de la structure initiale des agglomérats des murailles des termitières
épigées et des sols témoins sont conservés (diamètre moyen des agglomérats
compris entre 1,65 mm et 1,85 mm).
Le test de dispersion mécanique après pré-traitement dans le kérosène met
en évidence une perte importante de stabilité des agglomérats des murailles
termitiques (diamètre moyen des agglomérats = 0,85 ± 0,15 mm). Ceci est sans
doute dû à une destruction, par le kérosène, du liant salivaire entre les boulettes de
sol excavées puis remaniées par les termites pour construire leur muraille.
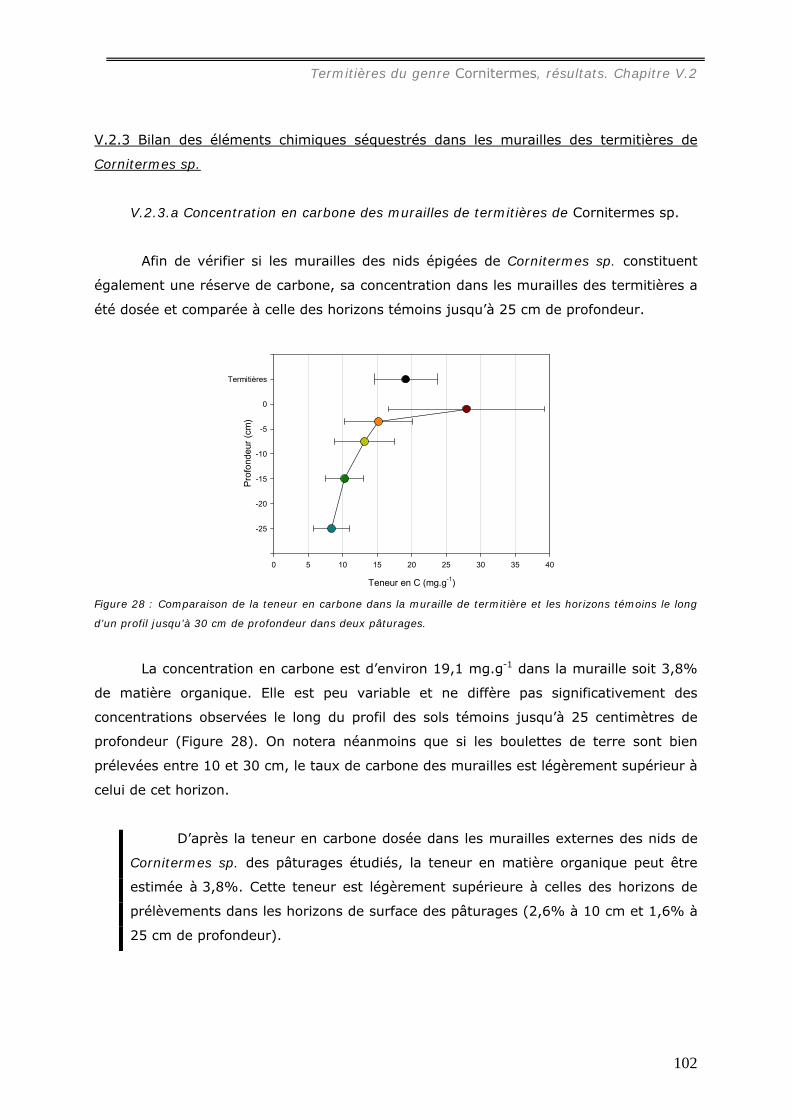
Termitières du genre Cornitermes, résultats. Chapitre V.2
102
V.2.3 Bilan des éléments chimiques séquestrés dans les murailles des termitières de
Cornitermes sp.
V.2.3.a Concentration en carbone des murailles de termitières de Cornitermes sp.
Afin de vérifier si les murailles des nids épigées de Cornitermes sp. constituent
également une réserve de carbone, sa concentration dans les murailles des termitières a
été dosée et comparée à celle des horizons témoins jusqu’à 25 cm de profondeur.
Teneur en C (mg.g-1)
0 5 10 15 20 25 30 35 40
Pro
fond
eur (
cm)
-25
-20
-15
-10
-5
0
Termitières
Figure 28 : Comparaison de la teneur en carbone dans la muraille de termitière et les horizons témoins le long
d’un profil jusqu’à 30 cm de profondeur dans deux pâturages.
La concentration en carbone est d’environ 19,1 mg.g-1 dans la muraille soit 3,8%
de matière organique. Elle est peu variable et ne diffère pas significativement des
concentrations observées le long du profil des sols témoins jusqu’à 25 centimètres de
profondeur (Figure 28). On notera néanmoins que si les boulettes de terre sont bien
prélevées entre 10 et 30 cm, le taux de carbone des murailles est légèrement supérieur à
celui de cet horizon.
D’après la teneur en carbone dosée dans les murailles externes des nids de
Cornitermes sp. des pâturages étudiés, la teneur en matière organique peut être
estimée à 3,8%. Cette teneur est légèrement supérieure à celles des horizons de
prélèvements dans les horizons de surface des pâturages (2,6% à 10 cm et 1,6% à
25 cm de profondeur).
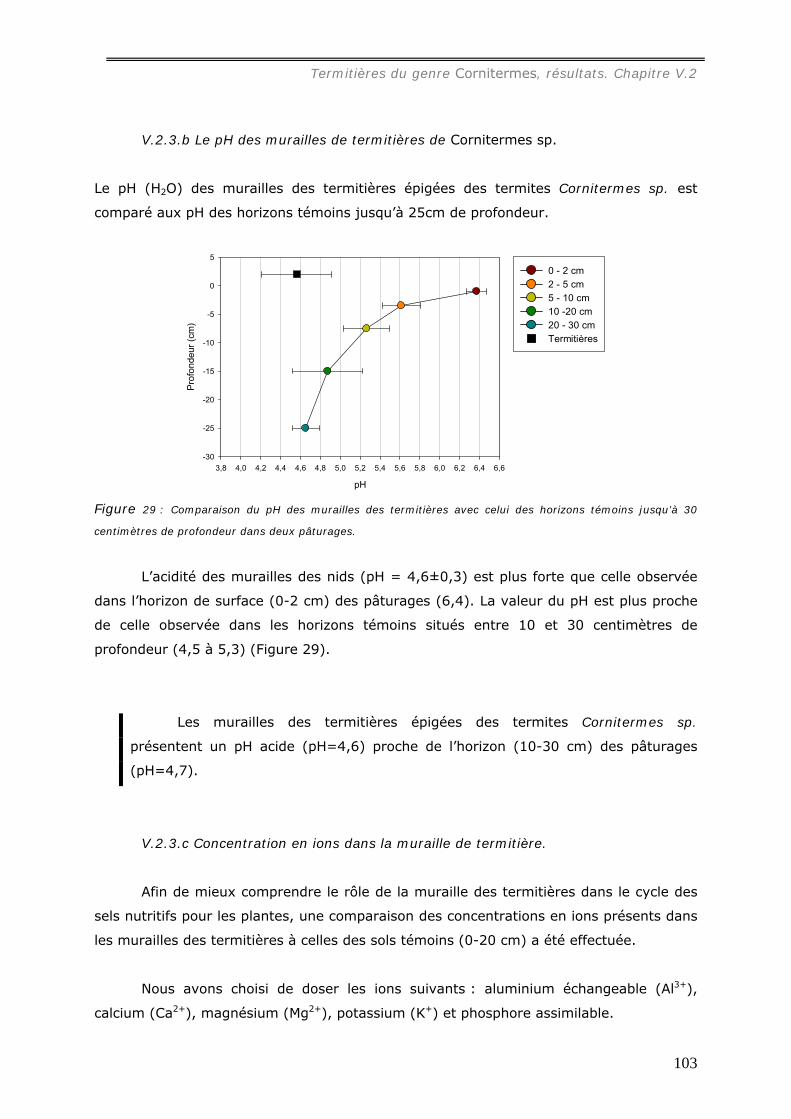
Termitières du genre Cornitermes, résultats. Chapitre V.2
103
V.2.3.b Le pH des murailles de termitières de Cornitermes sp.
Le pH (H2O) des murailles des termitières épigées des termites Cornitermes sp. est
comparé aux pH des horizons témoins jusqu’à 25cm de profondeur.
pH
3,8 4,0 4,2 4,4 4,6 4,8 5,0 5,2 5,4 5,6 5,8 6,0 6,2 6,4 6,6
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
0 - 2 cm2 - 5 cm5 - 10 cm10 -20 cm20 - 30 cmTermitières
pH
3,8 4,0 4,2 4,4 4,6 4,8 5,0 5,2 5,4 5,6 5,8 6,0 6,2 6,4 6,6
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
0 - 2 cm2 - 5 cm5 - 10 cm10 -20 cm20 - 30 cmTermitières
Figure 29 : Comparaison du pH des murailles des termitières avec celui des horizons témoins jusqu’à 30
centimètres de profondeur dans deux pâturages.
L’acidité des murailles des nids (pH = 4,6±0,3) est plus forte que celle observée
dans l’horizon de surface (0-2 cm) des pâturages (6,4). La valeur du pH est plus proche
de celle observée dans les horizons témoins situés entre 10 et 30 centimètres de
profondeur (4,5 à 5,3) (Figure 29).
Les murailles des termitières épigées des termites Cornitermes sp.
présentent un pH acide (pH=4,6) proche de l’horizon (10-30 cm) des pâturages
(pH=4,7).
V.2.3.c Concentration en ions dans la muraille de termitière.
Afin de mieux comprendre le rôle de la muraille des termitières dans le cycle des
sels nutritifs pour les plantes, une comparaison des concentrations en ions présents dans
les murailles des termitières à celles des sols témoins (0-20 cm) a été effectuée.
Nous avons choisi de doser les ions suivants : aluminium échangeable (Al3+),
calcium (Ca2+), magnésium (Mg2+), potassium (K+) et phosphore assimilable.
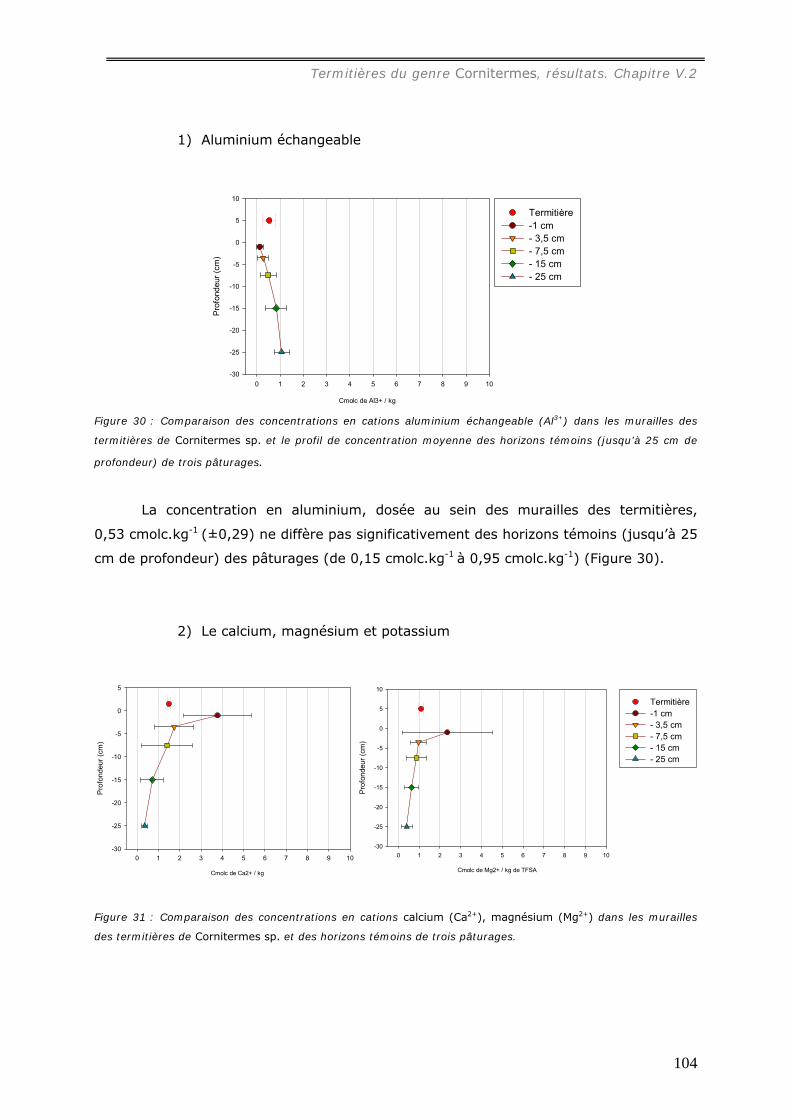
Termitières du genre Cornitermes, résultats. Chapitre V.2
104
1) Aluminium échangeable
-1 cm- 3,5 cm- 7,5 cm- 15 cm- 25 cm
Termitière
Cmolc de Al3+ / kg
0 1 2 3 4 5 6 7 8 9 10
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
10
Figure 30 : Comparaison des concentrations en cations aluminium échangeable (Al3+) dans les murailles des
termitières de Cornitermes sp. et le profil de concentration moyenne des horizons témoins (jusqu’à 25 cm de
profondeur) de trois pâturages.
La concentration en aluminium, dosée au sein des murailles des termitières,
0,53 cmolc.kg-1 (±0,29) ne diffère pas significativement des horizons témoins (jusqu’à 25
cm de profondeur) des pâturages (de 0,15 cmolc.kg-1 à 0,95 cmolc.kg-1) (Figure 30).
2) Le calcium, magnésium et potassium
Cmolc de Mg2+ / kg de TFSA
0 1 2 3 4 5 6 7 8 9 10
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
10
Cmolc de Ca2+ / kg
0 1 2 3 4 5 6 7 8 9 10
Pro
fond
eur (
cm)
-30
-25
-20
-15
-10
-5
0
5
-1 cm- 3,5 cm- 7,5 cm- 15 cm- 25 cm
Termitière
Figure 31 : Comparaison des concentrations en cations calcium (Ca2+), magnésium (Mg2+) dans les murailles
des termitières de Cornitermes sp. et des horizons témoins de trois pâturages.
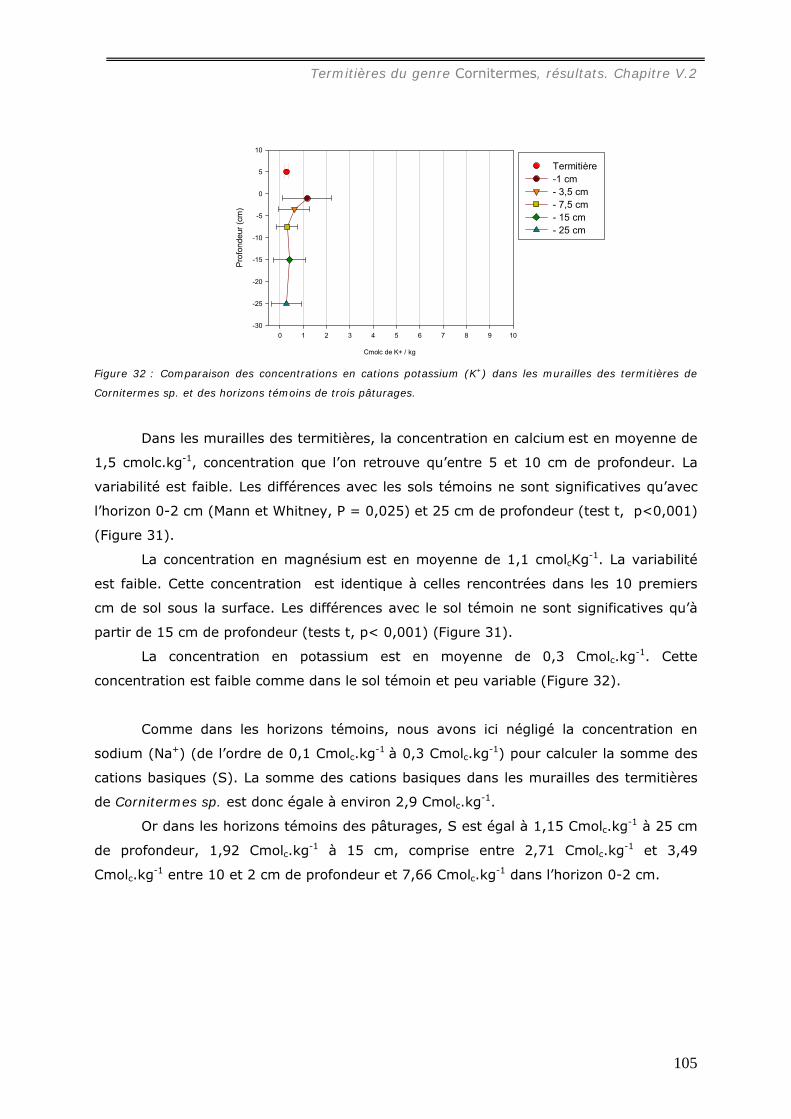
Termitières du genre Cornitermes, résultats. Chapitre V.2
105
Cmolc de K+ / kg
0 1 2 3 4 5 6 7 8 9 10
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
10
-1 cm- 3,5 cm- 7,5 cm- 15 cm- 25 cm
Termitière
Figure 32 : Comparaison des concentrations en cations potassium (K+) dans les murailles des termitières de
Cornitermes sp. et des horizons témoins de trois pâturages.
Dans les murailles des termitières, la concentration en calcium est en moyenne de
1,5 cmolc.kg-1, concentration que l’on retrouve qu’entre 5 et 10 cm de profondeur. La
variabilité est faible. Les différences avec les sols témoins ne sont significatives qu’avec
l’horizon 0-2 cm (Mann et Whitney, P = 0,025) et 25 cm de profondeur (test t, p<0,001)
(Figure 31).
La concentration en magnésium est en moyenne de 1,1 cmolcKg-1. La variabilité
est faible. Cette concentration est identique à celles rencontrées dans les 10 premiers
cm de sol sous la surface. Les différences avec le sol témoin ne sont significatives qu’à
partir de 15 cm de profondeur (tests t, p< 0,001) (Figure 31).
La concentration en potassium est en moyenne de 0,3 Cmolc.kg-1. Cette
concentration est faible comme dans le sol témoin et peu variable (Figure 32).
Comme dans les horizons témoins, nous avons ici négligé la concentration en
sodium (Na+) (de l’ordre de 0,1 Cmolc.kg-1 à 0,3 Cmolc.kg-1) pour calculer la somme des
cations basiques (S). La somme des cations basiques dans les murailles des termitières
de Cornitermes sp. est donc égale à environ 2,9 Cmolc.kg-1.
Or dans les horizons témoins des pâturages, S est égal à 1,15 Cmolc.kg-1 à 25 cm
de profondeur, 1,92 Cmolc.kg-1 à 15 cm, comprise entre 2,71 Cmolc.kg-1 et 3,49
Cmolc.kg-1 entre 10 et 2 cm de profondeur et 7,66 Cmolc.kg-1 dans l’horizon 0-2 cm.
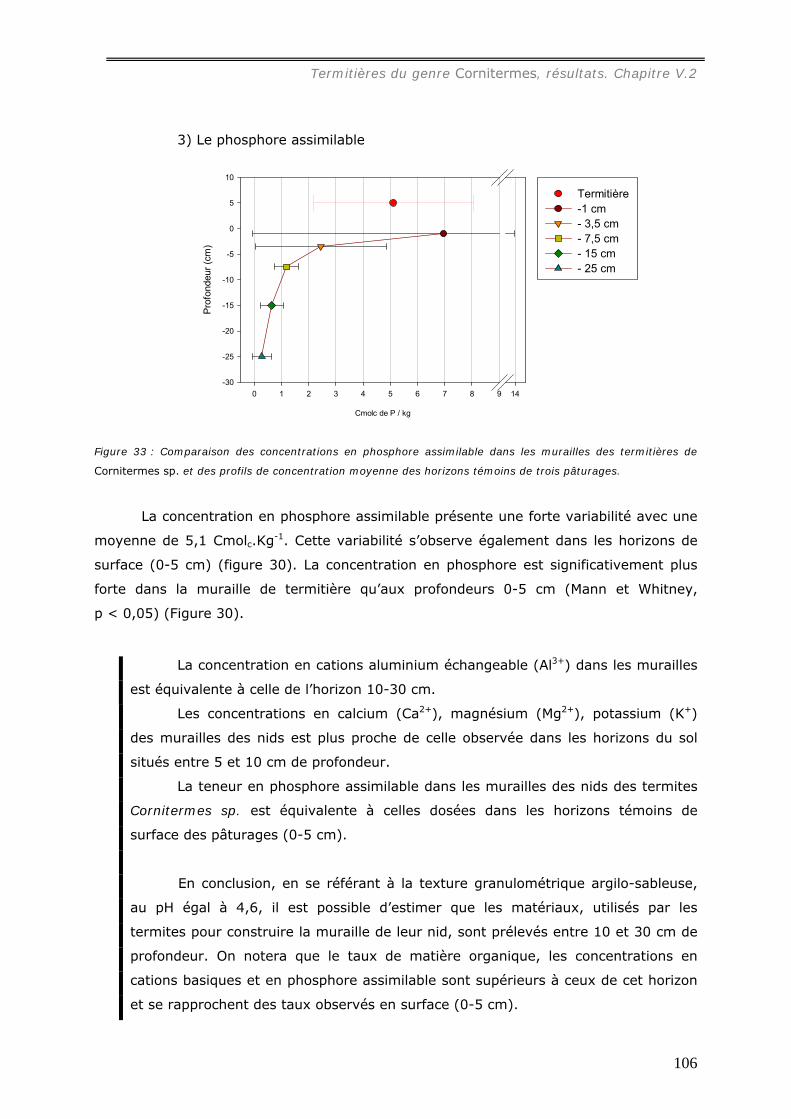
Termitières du genre Cornitermes, résultats. Chapitre V.2
106
3) Le phosphore assimilable
Cmolc de P / kg
0 1 2 3 4 5 6 7 8 9 14
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
10
-1 cm- 3,5 cm- 7,5 cm- 15 cm- 25 cm
Termitière
Figure 33 : Comparaison des concentrations en phosphore assimilable dans les murailles des termitières de
Cornitermes sp. et des profils de concentration moyenne des horizons témoins de trois pâturages.
La concentration en phosphore assimilable présente une forte variabilité avec une
moyenne de 5,1 Cmolc.Kg-1. Cette variabilité s’observe également dans les horizons de
surface (0-5 cm) (figure 30). La concentration en phosphore est significativement plus
forte dans la muraille de termitière qu’aux profondeurs 0-5 cm (Mann et Whitney,
p < 0,05) (Figure 30).
La concentration en cations aluminium échangeable (Al3+) dans les murailles
est équivalente à celle de l’horizon 10-30 cm.
Les concentrations en calcium (Ca2+), magnésium (Mg2+), potassium (K+)
des murailles des nids est plus proche de celle observée dans les horizons du sol
situés entre 5 et 10 cm de profondeur.
La teneur en phosphore assimilable dans les murailles des nids des termites
Cornitermes sp. est équivalente à celles dosées dans les horizons témoins de
surface des pâturages (0-5 cm).
En conclusion, en se référant à la texture granulométrique argilo-sableuse,
au pH égal à 4,6, il est possible d’estimer que les matériaux, utilisés par les
termites pour construire la muraille de leur nid, sont prélevés entre 10 et 30 cm de
profondeur. On notera que le taux de matière organique, les concentrations en
cations basiques et en phosphore assimilable sont supérieurs à ceux de cet horizon
et se rapprochent des taux observés en surface (0-5 cm).

Termitières du genre Cornitermes, résultats. Chapitre V.2
107
V.2.4 Simulation de l’impact du feu sur les termitières.
Comme nous l’avons vu précédemment, la stabilité de la muraille externe des nids
de Cornitermes sp. est importante.
Ces nids renferment une importante quantité de matière organique dans leur
feuilleté interne.
Ces nids constituent donc des structures biogéniques stables dans lesquelles est
piégé et protégé un stock de matière organique intéressant pour le maintien voire
l’augmentation de la fertilité chimique des horizons de surface des pâturages.
Ces termitières épigées subissent annuellement des feux lors du brûlis des
pâturages mais aussi lorsque les paysans amazoniens tentent de détruire les colonies.
Des expériences ont donc été réalisées afin de mettre en évidence le rôle
particulier du feu dans la dégradation physico-chimique des termitières.
V.2.4.a Action du feu sur les murailles des termitières de Cornitermes sp.
L’action du feu se traduit par une forte augmentation de la température sur une
durée plus ou moins longue. Cette hausse de la température provoque la destruction de
la matière organique et des populations de microorganismes ; elle modifie ainsi
directement les liaisons chimiques entre les différents constituants organiques et
minéraux mais aussi perturbe les cycles biogéochimiques qui s’y s’opèrent.
a1) Texture.
Les textures des murailles brûlées (3 murailles dans 3 pâturages) sont comparées
à celles des murailles des nids vivants et aux textures des horizons témoins (10-30 cm)
des 3 pâturages P01, P03 et P04. Les tableaux de données sont présentés dans
l’annexe 5.
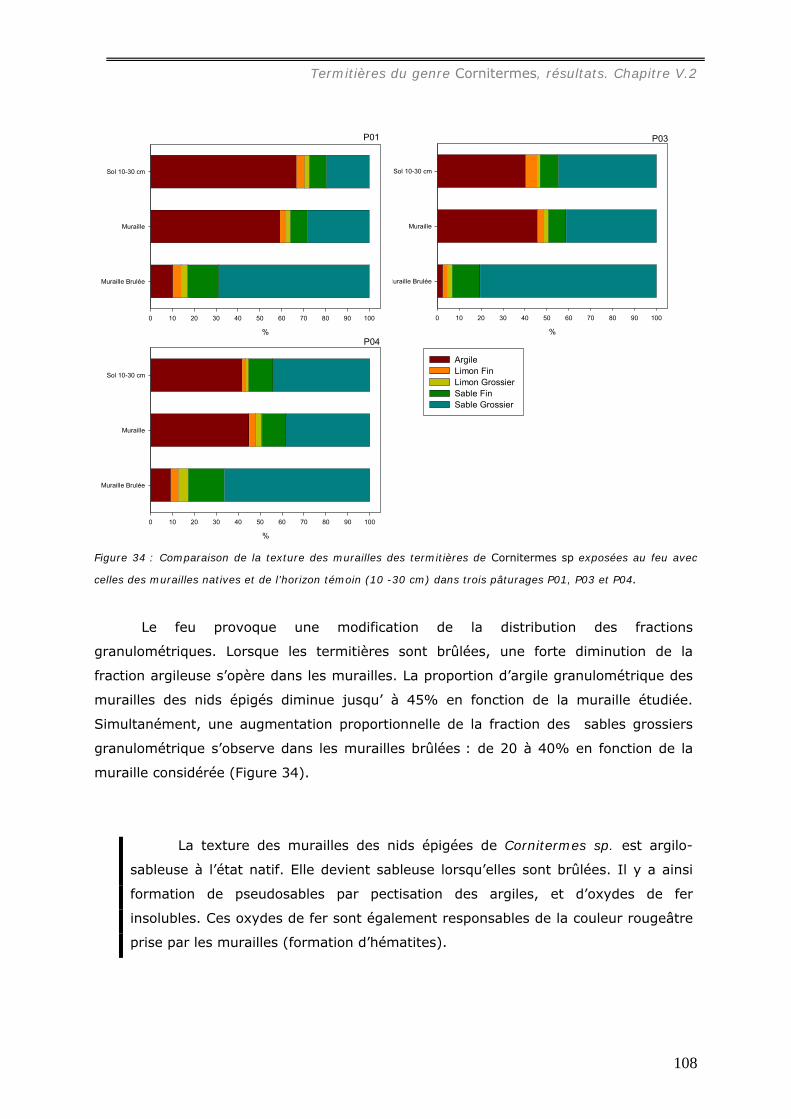
Termitières du genre Cornitermes, résultats. Chapitre V.2
108
P03
%
0 10 20 30 40 50 60 70 80 90 100
Sol 10-30 cm
Muraille
Muraille Brulée
P01
%
0 10 20 30 40 50 60 70 80 90 100
Sol 10-30 cm
Muraille
Muraille Brulée
P04
%
0 10 20 30 40 50 60 70 80 90 100
Sol 10-30 cm
Muraille
Muraille Brulée
ArgileLimon Fin Limon Grossier Sable Fin Sable Grossier
P03
%
0 10 20 30 40 50 60 70 80 90 100
Sol 10-30 cm
Muraille
Muraille Brulée
P01
%
0 10 20 30 40 50 60 70 80 90 100
Sol 10-30 cm
Muraille
Muraille Brulée
P04
%
0 10 20 30 40 50 60 70 80 90 100
Sol 10-30 cm
Muraille
Muraille Brulée
ArgileLimon Fin Limon Grossier Sable Fin Sable Grossier
Figure 34 : Comparaison de la texture des murailles des termitières de Cornitermes sp exposées au feu avec
celles des murailles natives et de l’horizon témoin (10 -30 cm) dans trois pâturages P01, P03 et P04.
Le feu provoque une modification de la distribution des fractions
granulométriques. Lorsque les termitières sont brûlées, une forte diminution de la
fraction argileuse s’opère dans les murailles. La proportion d’argile granulométrique des
murailles des nids épigés diminue jusqu’ à 45% en fonction de la muraille étudiée.
Simultanément, une augmentation proportionnelle de la fraction des sables grossiers
granulométrique s’observe dans les murailles brûlées : de 20 à 40% en fonction de la
muraille considérée (Figure 34).
La texture des murailles des nids épigées de Cornitermes sp. est argilo-
sableuse à l’état natif. Elle devient sableuse lorsqu’elles sont brûlées. Il y a ainsi
formation de pseudosables par pectisation des argiles, et d’oxydes de fer
insolubles. Ces oxydes de fer sont également responsables de la couleur rougeâtre
prise par les murailles (formation d’hématites).
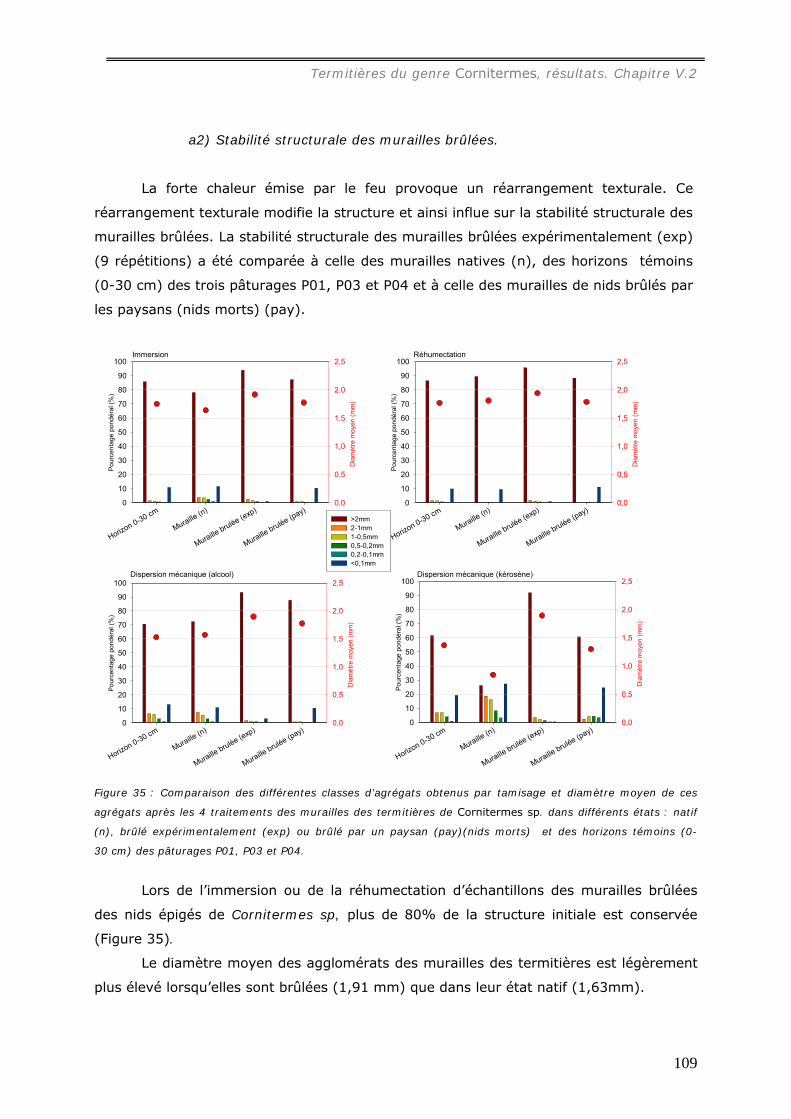
Termitières du genre Cornitermes, résultats. Chapitre V.2
109
a2) Stabilité structurale des murailles brûlées.
La forte chaleur émise par le feu provoque un réarrangement texturale. Ce
réarrangement texturale modifie la structure et ainsi influe sur la stabilité structurale des
murailles brûlées. La stabilité structurale des murailles brûlées expérimentalement (exp)
(9 répétitions) a été comparée à celle des murailles natives (n), des horizons témoins
(0-30 cm) des trois pâturages P01, P03 et P04 et à celle des murailles de nids brûlés par
les paysans (nids morts) (pay).
Immersion
Horizon 0-30 cm
Muraille (n)
Muraille brulée (exp)
Muraille brulée (pay)
Pou
rcen
tage
pon
déra
l (%
)
0
10
20
30
40
50
60
70
80
90
100D
iam
ètre
moy
en (m
m)
0,0
0,5
1,0
1,5
2,0
2,5Réhumectation
Horizon 0-30 cm
Muraille (n)
Muraille brulée (exp)
Muraille brulée (pay)
Pou
rcen
tage
pon
déra
l (%
)
0
10
20
30
40
50
60
70
80
90
100
Dia
mèt
re m
oyen
(mm
)
0,0
0,5
1,0
1,5
2,0
2,5
Horizon 0-30 cm
Muraille (n)
Muraille brulée (exp)
Muraille brulée (pay)
Pou
rcen
tage
pon
déra
l (%
)
0
10
20
30
40
50
60
70
80
90
100
Dia
mèt
re m
oyen
(mm
)
0,0
0,5
1,0
1,5
2,0
2,5Dispersion mécanique (alcool)
Horizon 0-30 cm
Muraille (n)
Muraille brulée (exp)
Muraille brulée (pay)
Pour
cent
age
pond
éral
(%)
0
10
20
30
40
50
60
70
80
90
100
Dia
mèt
re m
oyen
(mm
)
0,0
0,5
1,0
1,5
2,0
2,5Dispersion mécanique (kérosène)
>2mm 2-1mm 1-0,5mm 0,5-0,2mm 0,2-0,1mm <0,1mm
Immersion
Horizon 0-30 cm
Muraille (n)
Muraille brulée (exp)
Muraille brulée (pay)
Pou
rcen
tage
pon
déra
l (%
)
0
10
20
30
40
50
60
70
80
90
100D
iam
ètre
moy
en (m
m)
0,0
0,5
1,0
1,5
2,0
2,5Réhumectation
Horizon 0-30 cm
Muraille (n)
Muraille brulée (exp)
Muraille brulée (pay)
Pou
rcen
tage
pon
déra
l (%
)
0
10
20
30
40
50
60
70
80
90
100
Dia
mèt
re m
oyen
(mm
)
0,0
0,5
1,0
1,5
2,0
2,5
Horizon 0-30 cm
Muraille (n)
Muraille brulée (exp)
Muraille brulée (pay)
Pou
rcen
tage
pon
déra
l (%
)
0
10
20
30
40
50
60
70
80
90
100
Dia
mèt
re m
oyen
(mm
)
0,0
0,5
1,0
1,5
2,0
2,5Dispersion mécanique (alcool)
Horizon 0-30 cm
Muraille (n)
Muraille brulée (exp)
Muraille brulée (pay)
Pour
cent
age
pond
éral
(%)
0
10
20
30
40
50
60
70
80
90
100
Dia
mèt
re m
oyen
(mm
)
0,0
0,5
1,0
1,5
2,0
2,5Dispersion mécanique (kérosène)
>2mm 2-1mm 1-0,5mm 0,5-0,2mm 0,2-0,1mm <0,1mm
Figure 35 : Comparaison des différentes classes d’agrégats obtenus par tamisage et diamètre moyen de ces
agrégats après les 4 traitements des murailles des termitières de Cornitermes sp. dans différents états : natif
(n), brûlé expérimentalement (exp) ou brûlé par un paysan (pay)(nids morts) et des horizons témoins (0-
30 cm) des pâturages P01, P03 et P04.
Lors de l’immersion ou de la réhumectation d’échantillons des murailles brûlées
des nids épigés de Cornitermes sp, plus de 80% de la structure initiale est conservée
(Figure 35).
Le diamètre moyen des agglomérats des murailles des termitières est légèrement
plus élevé lorsqu’elles sont brûlées (1,91 mm) que dans leur état natif (1,63mm).

Termitières du genre Cornitermes, résultats. Chapitre V.2
110
Lorsque les murailles ont été brûlées par un paysan (pay), le diamètre moyen des
agglomérats obtenus est du même ordre que celui des agglomérats natifs (n) (Figure 35).
Pour rappel, les tests de dispersion mécanique dans l’alcool ou dans le kérosène ont
mis en évidence une réduction de la stabilité structurale des murailles natives (n) sans
doute en raison de la destruction du liant salivaire. Lorsque les murailles sont brûlées
expérimentalement (exp), une augmentation significative de la stabilité structurale
s’observe. Cette stabilité ne s’explique pas par le liant salivaire mais sans doute par la
déshydratation des argiles (pectisation) aboutissant à des agglomérats très stables.
Ceci se traduit par une augmentation systématique du diamètre moyen des
agglomérats désolidarisés de la structure. Le diamètre moyen des agglomérats des
murailles augmente de 0,84 mm à 1,89 mm lorsque les murailles sont brûlées
expérimentalement (exp).
Lorsque les murailles sont brûlées par les paysans, leur stabilité structurale diminue
et ne diffère plus significativement de celle des horizons témoins (agglomérats dont la
taille est d’environ 1,29mm) (Figure 35).
L’augmentation de la température induite par le feu a donc tendance à
accroître la stabilité structurale des murailles des nids de Cornitermes sp.
Le pré-traitement dans le Kérosène mettait en évidence l’impact fragile du
liant salivaire dans la stabilité des agglomérats de la muraille fraîche (Chapitre
V.2.2.b2). Le pré-traitement au kérosène ne réduit pas la stabilité de la muraille
brûlée ainsi ce test confirme que l’augmentation de la température a modifié les
liaisons chimiques des constituants à l’origine de la stabilité structurale des
murailles fraîches. Les murailles brûlées se caractérisent par une stabilité de nature
différente de celle des murailles fraîches.
Les résultats obtenus pour les murailles brûlées par les paysans (nids morts)
mettent en évidence la réversibilité de ces modifications.
a3) Dosage du carbone dans les murailles brûlées.
Le feu détruit la matière organique et provoque une forte minéralisation du
carbone. La concentration en carbone dosée dans les murailles brûlées des termitières
épigées est comparée à celle observée dans leur état natif et dans les horizons témoins
(0-10cm) de 3 pâturages P01, P03 et P04.
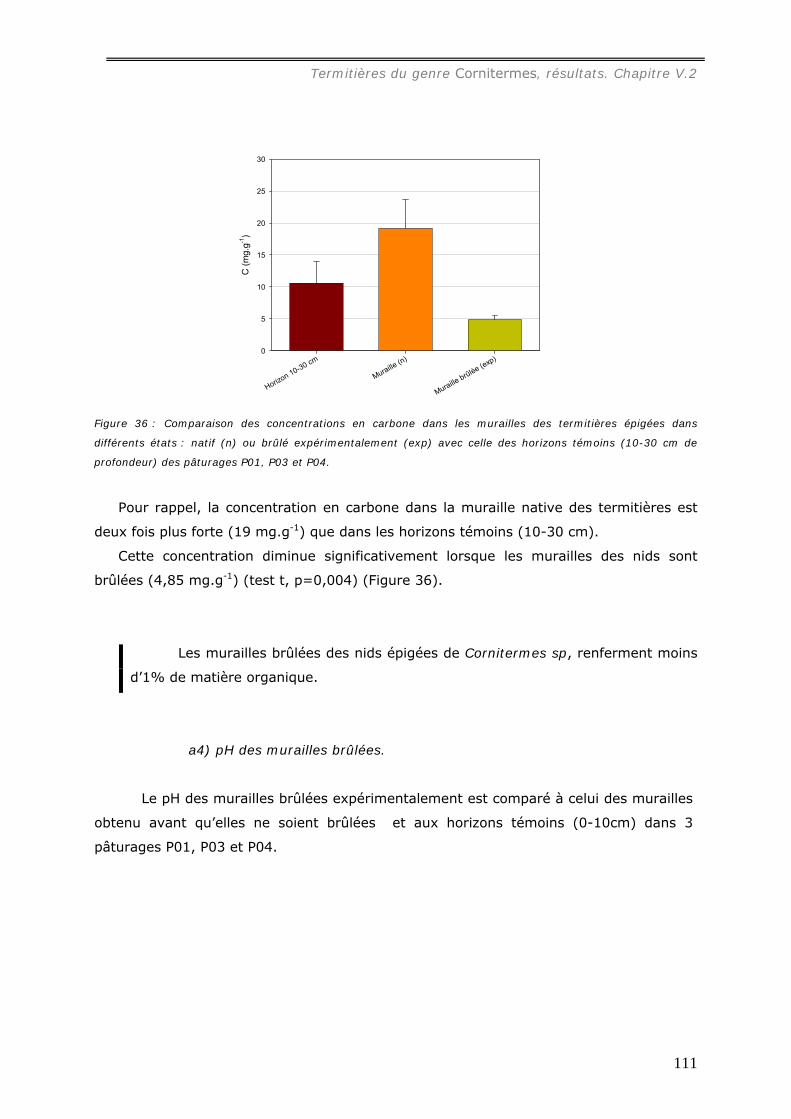
Termitières du genre Cornitermes, résultats. Chapitre V.2
111
Horizon 10-30 cm
Muraille (n)
Muraille brûlée (exp)
C (m
g.g-1
)
0
5
10
15
20
25
30
Figure 36 : Comparaison des concentrations en carbone dans les murailles des termitières épigées dans
différents états : natif (n) ou brûlé expérimentalement (exp) avec celle des horizons témoins (10-30 cm de
profondeur) des pâturages P01, P03 et P04.
Pour rappel, la concentration en carbone dans la muraille native des termitières est
deux fois plus forte (19 mg.g-1) que dans les horizons témoins (10-30 cm).
Cette concentration diminue significativement lorsque les murailles des nids sont
brûlées (4,85 mg.g-1) (test t, p=0,004) (Figure 36).
Les murailles brûlées des nids épigées de Cornitermes sp, renferment moins
d’1% de matière organique.
a4) pH des murailles brûlées.
Le pH des murailles brûlées expérimentalement est comparé à celui des murailles
obtenu avant qu’elles ne soient brûlées et aux horizons témoins (0-10cm) dans 3
pâturages P01, P03 et P04.
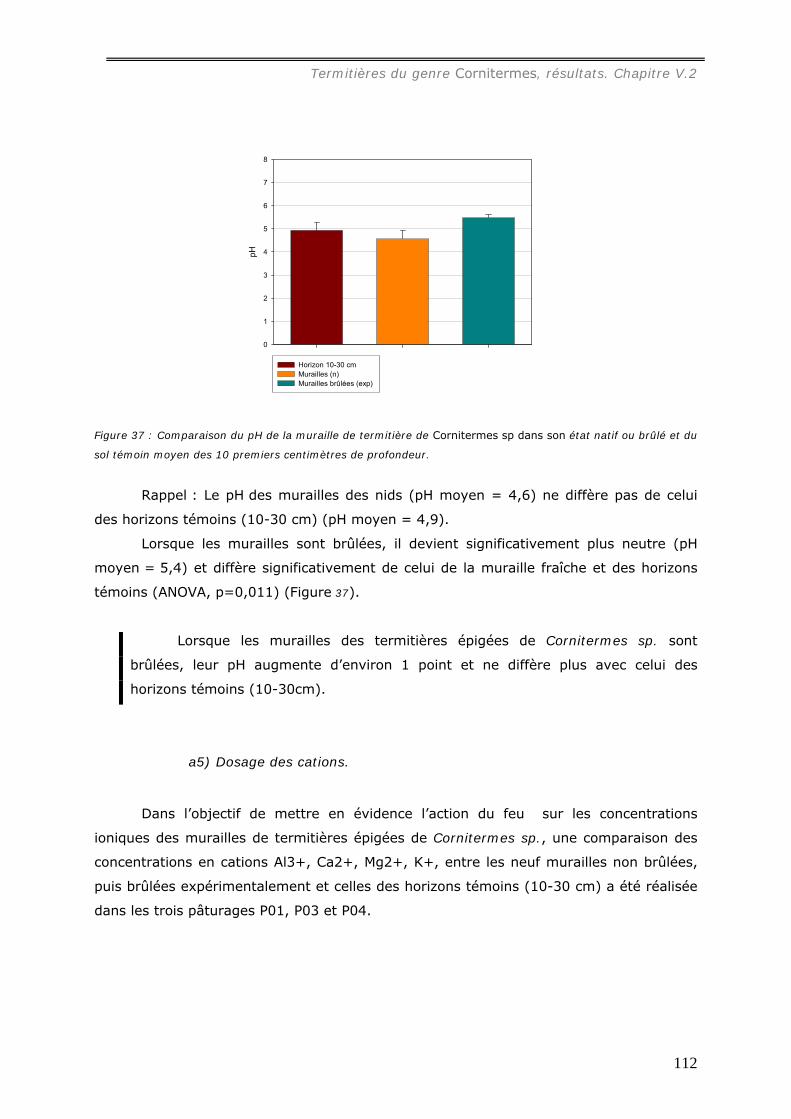
Termitières du genre Cornitermes, résultats. Chapitre V.2
112
pH
0
1
2
3
4
5
6
7
8
Horizon 10-30 cm Murailles (n) Murailles brûlées (exp)
Figure 37 : Comparaison du pH de la muraille de termitière de Cornitermes sp dans son état natif ou brûlé et du
sol témoin moyen des 10 premiers centimètres de profondeur.
Rappel : Le pH des murailles des nids (pH moyen = 4,6) ne diffère pas de celui
des horizons témoins (10-30 cm) (pH moyen = 4,9).
Lorsque les murailles sont brûlées, il devient significativement plus neutre (pH
moyen = 5,4) et diffère significativement de celui de la muraille fraîche et des horizons
témoins (ANOVA, p=0,011) (Figure 37).
Lorsque les murailles des termitières épigées de Cornitermes sp. sont
brûlées, leur pH augmente d’environ 1 point et ne diffère plus avec celui des
horizons témoins (10-30cm).
a5) Dosage des cations.
Dans l’objectif de mettre en évidence l’action du feu sur les concentrations
ioniques des murailles de termitières épigées de Cornitermes sp., une comparaison des
concentrations en cations Al3+, Ca2+, Mg2+, K+, entre les neuf murailles non brûlées,
puis brûlées expérimentalement et celles des horizons témoins (10-30 cm) a été réalisée
dans les trois pâturages P01, P03 et P04.
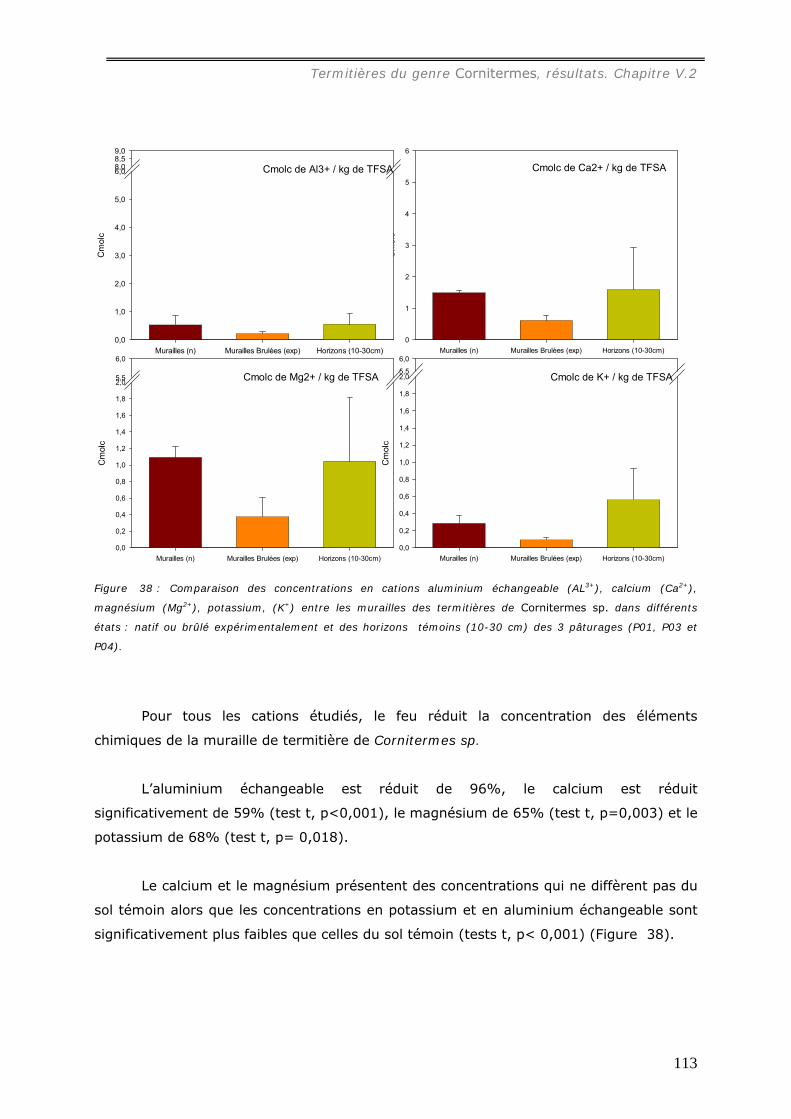
Termitières du genre Cornitermes, résultats. Chapitre V.2
113
Cmolc de Ca2+ / kg de TFSA
Murailles (n) Murailles Brulées (exp) Horizons (10-30cm)
Cm
olc
0
1
2
3
4
5
6
Cmolc de Mg2+ / kg de TFSA
Murailles (n) Murailles Brulées (exp) Horizons (10-30cm)
Cm
olc
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
1,8
2,05,5
6,0
Cmolc de K+ / kg de TFSA
Murailles (n) Murailles Brulées (exp) Horizons (10-30cm)
Cm
olc
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
1,8
2,05,5
6,0
Cmolc de Al3+ / kg de TFSA
Murailles (n) Murailles Brulées (exp) Horizons (10-30cm)
Cm
olc
0,0
1,0
2,0
3,0
4,0
5,0
6,08,08,59,0
Figure 38 : Comparaison des concentrations en cations aluminium échangeable (AL3+), calcium (Ca2+),
magnésium (Mg2+), potassium, (K+) entre les murailles des termitières de Cornitermes sp. dans différents
états : natif ou brûlé expérimentalement et des horizons témoins (10-30 cm) des 3 pâturages (P01, P03 et
P04).
Pour tous les cations étudiés, le feu réduit la concentration des éléments
chimiques de la muraille de termitière de Cornitermes sp.
L’aluminium échangeable est réduit de 96%, le calcium est réduit
significativement de 59% (test t, p<0,001), le magnésium de 65% (test t, p=0,003) et le
potassium de 68% (test t, p= 0,018).
Le calcium et le magnésium présentent des concentrations qui ne diffèrent pas du
sol témoin alors que les concentrations en potassium et en aluminium échangeable sont
significativement plus faibles que celles du sol témoin (tests t, p< 0,001) (Figure 38).
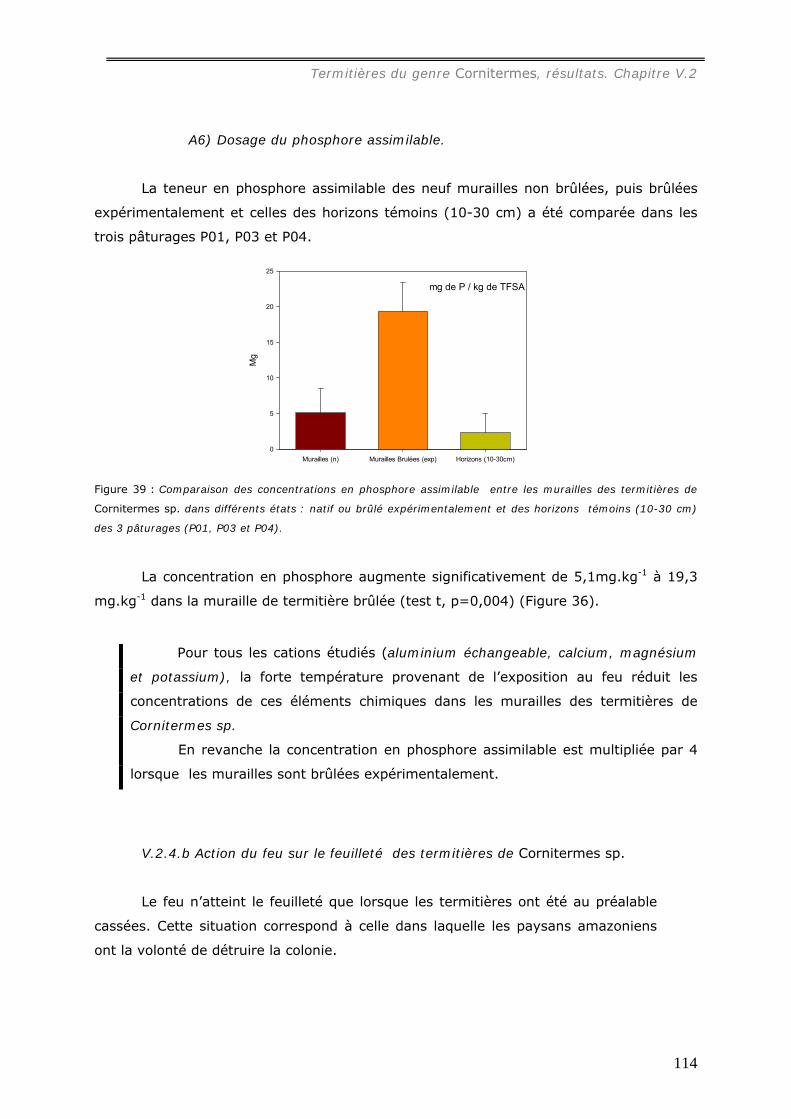
Termitières du genre Cornitermes, résultats. Chapitre V.2
114
A6) Dosage du phosphore assimilable.
La teneur en phosphore assimilable des neuf murailles non brûlées, puis brûlées
expérimentalement et celles des horizons témoins (10-30 cm) a été comparée dans les
trois pâturages P01, P03 et P04.
mg de P / kg de TFSA
Murailles (n) Murailles Brulées (exp) Horizons (10-30cm)
Mg
0
5
10
15
20
25
Figure 39 : Comparaison des concentrations en phosphore assimilable entre les murailles des termitières de
Cornitermes sp. dans différents états : natif ou brûlé expérimentalement et des horizons témoins (10-30 cm)
des 3 pâturages (P01, P03 et P04).
La concentration en phosphore augmente significativement de 5,1mg.kg-1 à 19,3
mg.kg-1 dans la muraille de termitière brûlée (test t, p=0,004) (Figure 36).
Pour tous les cations étudiés (aluminium échangeable, calcium, magnésium
et potassium), la forte température provenant de l’exposition au feu réduit les
concentrations de ces éléments chimiques dans les murailles des termitières de
Cornitermes sp.
En revanche la concentration en phosphore assimilable est multipliée par 4
lorsque les murailles sont brûlées expérimentalement.
V.2.4.b Action du feu sur le feuilleté des termitières de Cornitermes sp.
Le feu n’atteint le feuilleté que lorsque les termitières ont été au préalable
cassées. Cette situation correspond à celle dans laquelle les paysans amazoniens
ont la volonté de détruire la colonie.

Termitières du genre Cornitermes, résultats. Chapitre V.2
115
Les feux, résultant de la pratique de la culture sur brûlis, n’atteignent
généralement pas le centre de la termitière. La muraille joue ici un rôle majeur
dans la protection du feuilleté.
Lorsque le feu atteint le feuilleté, il est généralement totalement consumé
et réduit en cendre. Toute la matière organique est détruite, seule reste la partie
minérale (environ 23% de la matière sèche).

Termitières du genre Cornitermes, discussion. Chapitre V.3
116
V.3 DISCUSSION
L’objectif de cette étude est de mettre en évidence le rôle particulier du termite
xylophage Cornitermes sp. affinis ovatus dans le fonctionnement du sol au travers de
l’étude des interactions termitière-sol.
Ce genre néotropical construit des nids de très grande taille (pouvant atteindre
1,5m de hauteur). Le cycle de développement des nids le fait passer d’une situation
hypogée à un état presque entièrement épigé. Les termitières sont composées de deux
parties distinctes:
-La muraille, composée de boulettes de terre imprégnées de salive, d’origine cibariale,
est percée d’orifices de ventilation au sommet du nid.
-Le feuilleté, correspond aux parois de la chambre intérieure. La structure alvéolaire est
composée de boulettes recouvertes d’un enduit noir d’origine proctodéale (fécès).
De nos résultats, issus des trois études menées sur les termitières de Cornitermes sp :
1) Cartographie des nids de Cornitermes sp.
2) Analyses des caractéristiques physiques et chimiques des termitières
3) Expérience de simulation de l’impact du feu sur les termitières.
Il est apparu que cette espèce est inféodée aux milieux ouverts (pâturages).
Impacts de la déforestation sur le termite Cornitermes sp.
La densité moyenne des termitières épigées rencontrées de Cornitermes sp. dans
les pâturages (surface variant de 2,9 ha à 16,7 ha) est d’environ 26,3 nids par hectare.
Cette densité observée est faible en comparaison de l’étude de Kollar (1932) qui relève
des densités de Cornitermes cumulans fluctuant de 33 nids par hectare à 73 nids par
hectare en fonction du site étudié. (Kollar, 1832). Ces résultats quantitatifs
correspondent à ceux obtenus par Redford (1984) qui a rencontré une moyenne de 55
nids par hectare dans des pâturages de plus de 5 ans. (Redford, 1984 ; Juan Santiago
and Porcel, 2000). D’autres études dans d’autres milieux, tel qu’en en savane africaine,
pour d’autres espèces de termite champignonniste, présentent des densités de
termitières encore nettement plus élevées : plus de 100 nids par hectare pour un termite
cubitermes. (Lepage and Tano, 1986 ; Tano and Lepage, 1990).

Termitières du genre Cornitermes, discussion. Chapitre V.3
117
Plus de 46% des édifices recensés dépassent 1 mètre de hauteur. La surface
occupée par les termitières ne dépasse cependant pas 0,2% de la surface totale des
pâturages les plus envahis.
En considérant les densités de termitière par hectare, et la proportion des trois
classes : 46% de grandes (>1 m), 29% de moyennes (0,6m<hauteur<1 m) et 24% de
petites (<0,6m) ; il est possible d’extrapoler l’évolution de la densité des termitières. La
régression non linéaire obtenue met en évidence que les termites sont dans une phase
d’expansion dans les pâturages de 6 ans et qu’un plateau de densité devrait être atteint
entre 20 et 25 ans.
Dans les forêts, aucune termitière épigée de cette espèce n’a pu être observée.
Cette absence dans les forêts peut s’expliquer par l’incapacité de cette espèce à se
développer dans les milieux fermés (forêt). Le termite Cornitermes serait alors une
espèce colonisatrice qui profiterait de la disparition de la forêt pour coloniser de
nouveaux espaces. Elle pourrait alors être considérée comme une espèce indicatrice des
fronts pionniers de déforestation.
Cette espèce xylophage se développerait en profitant du stock de matière
organique issu des méthodes de déforestation, essentiellement sous forme de racines en
décomposition et de bois brûlé. Ceci expliquerait sa présence dans les nouveaux milieux
ouverts tels que les pâturages mais aussi les rizières. Cette hypothèse sous entend que
la densité de termitière serait fonction du stock de matière organique. Considérant que la
majorité des termitières sont de grande taille (de l’ordre de 45%), il paraît nécessaire de
quantifier précisément la densité des termitières mortes par rapport à la densité des
termitières vivantes dans ces pâturages et de dénombrer le nombre de termitières
présentes dans les autres types de milieux (rizières, jeunes pâturages (1an), vieux
pâturages de plus de 15 ans suite à la déforestation).
Une étude réalisée sur une très grande superficie à l’échelle du paysage avec un
suivi dans le temps permettrait de mettre en évidence l’évolution de cette espèce en
fonction de la progression du front de déforestation.
Autres facteurs limitant l’activité du termite Cornitermes sp.
Aux échelles d’observation supérieures à 15 m, la distribution spatiale des
termitières épigées de Cornitermes sp. est variable en fonction du pâturage étudié. A ces
échelles, la distribution spatiale des termitières ne présente pas de patron de distribution
significatif.

Termitières du genre Cornitermes, discussion. Chapitre V.3
118
Aux petites échelles (<15m), la distribution spatiale des termitières est agrégée.
Cette agrégation locale peut s’expliquer par l’essaimage des ailés autour des nids mères.
La distribution spatiale des termitières s’est révélée reliée à l’altitude. La
toposéquence (annexe 1) mettant en évidence la relation entre l’altitude et le type de
sol, nous permet de déduire que les termites s’installent préférentiellement dans les
zones peu humides, et fertiles des latosols profonds (préférés également par les
agriculteurs pour y établir leurs cultures) plutôt que dans les gleysols humides des bas-
fonds.
Aucune termitière n’a été observée dans les zones de bas fond caractérisées par les
gleysols. Certain auteurs ont déjà mis en évidence les préférences des termites avec les
propriétés de certain sol (Brandão, 1991). A ces zones de sol profond, correspond sous
forêt également, une abondance et une diversité végétale plus élevée que dans les bas-
fonds. Au niveau des sols intermédiaires (latosols peu profonds et cambisols), les
densités de termitières sont sans doute intermédiaires. En effet la réduction de la densité
des termitières est progressive.
Il serait ainsi intéressant de quantifier précisément la densité des nids épigés dans les
cambisols des pâturages amazoniens étudiés. Des régressions multiples pourraient être
établies entre les caractéristiques morphologiques, physiques et biologiques des
termitières et celles des trois types de sol des pâturages afin de déterminer précisément
la ou les propriétés physiques et/ou chimiques des sols qui influencent directement la
mise en place de nouvelles colonies. Mais d’ores et déjà nous pouvons affirmer que le %
d’argile (au moins 40%) et l’humidité du sol sont les paramètres majeurs qui influencent
leur installation.
Caractéristiques physico-chimiques des termitières de Cornitermes sp.
1 - La muraille externe
De texture argilo-sableuse, la muraille présente un léger enrichissement en argile
par rapport aux horizons de surface (0-10). Cette texture permet de supposer que les
matériaux utilisés pour la construction de la muraille ont été puisés entre 10 et 30
centimètres de profondeur. Peu poreuse, la muraille présente une stabilité structurale
supérieure à celle des horizons de référence (de 70% à 90% de la structure est
conservée lors des tests de dispersion) assurant ainsi une bonne protection à l’édifice.
Les agglomérats originaux produits par les termites acquièrent leur stabilité par
l’imprégnation de salive (Jungerius et al., 1999) mais des études ont également mis en
évidence le rôle prépondérant de l’enrichissement en argile dans la stabilité structurale

Termitières du genre Cornitermes, discussion. Chapitre V.3
119
des nids de termites Macrotermes. (Jouquet et al., 2002 ; Jouquet et al., 2004). Donc la
muraille est sans doute constituée de boulettes de terre choisies, prélevées et
régurgitées du cibarium comme chez les termites champignonnistes.
Les murailles ne constituent pas une réserve en matière organique (teneur
estimée de 3,8%).
Les murailles de termitière ont un pH acide (pH = 4,6) plus proche de celui des
horizons (10-30) et sont donc plus acides que les horizons de surface (pH=6,4). Ce
résultat est en contradiction avec ceux obtenus par certains auteurs qui ont observé une
augmentation du pH dans les termitières de Macrotermes par rapport aux horizons de
surface (Stoops, 1964 ; Lepage, 1974). Cette augmentation de pH était expliqué par les
conditions d’humidité et de circulation de l’eau dans la termitière mais une autre étude l’a
également corrélée à une accumulation de carbonate (Lee and Wood, 1971).
De même les concentrations en aluminium échangeable et en bases alcalines et
alcalino-terreuses sont du même ordre que des horizons situés entre 10 et 30 cm. Ces
différents paramètres confirment bien l’horizon de prélèvement de sol par les termites.
Ainsi la muraille de termitière représente une zone agricole où les paramètres de
la fertilité sont faibles : faible porosité, stable, pauvre en matière organique et sans stock
particulier de cations échangeables (concentrations équivalentes à celles du sol).
2 - Le feuilleté interne
Le feuilleté est composé de boulettes proctodéales recouvertes d’un enduit brun
foncé, lissé qui lui confère un caractère hydrophobe.
La masse de feuilleté interne au sein des nids est estimé variant de 288 kg.ha-1 à
655 kg.ha-1 en fonction de la superficie des pâturages et de leur densité respective en
termitières. Très riche en matière organique (77%), le stock de matière organique
protégé au sein des termitières sous forme de feuilleté a été estimé de 222 kg.ha-1 à 504
kg.ha-1. Il s’agit donc d’une réserve de matière organique remaniée tout au long de la vie
de la colonie. Le feuilleté des Cornitermes comme pour celui des Nasutitermes est
d’origine végétale (Amelung et al., 2002).Il est classiquement construit à l’aide de
déjections composées essentiellement de lignine, cette structure renferme un grand
nombre de vide qui lui confère une densité plus faible que celle de l’eau (Korb and
Linsenmair, 2000).
Impact de la calcination sur les termitières de Cornitermes sp. affinis ovatus.
Considérant le diamètre des termitières, la surface recouverte par ces structures
biogéniques est comprise entre 52,4 m² à 95,5 m² en fonction de la densité des

Termitières du genre Cornitermes, discussion. Chapitre V.3
120
termitières. Cette surface au sein des parcelles est inutilisable pour l’agriculteur. Afin
d’éliminer ces colonies géantes de Cornitermes, les agriculteurs percent la structure et
par l’ajout de bois et de carburant, brûlent la structure par l’intérieur (Deurival,
communication personnelle).
La méthode employée par les agriculteurs afin d’éliminer les colonies, même si
celle-ci n’est pas employée de façon systématique, n’est pas une bonne méthode pour
améliorer voire conserver la fertilité du sol. En effet, l’incendie de la structure biogénique
provoque la destruction de la matière organique du feuilleté. Le carbone du feuilleté est
presque totalement minéralisé. Seul demeure la muraille, dont la concentration en
matière organique diminue à moins de 1% après avoir été brûlée. La muraille prend
également une couleur rougeâtre témoignant de la néoformation d’hématites (oxydes de
fer) et de pseudosables par déshydratation des argiles (pectisation) aboutissant à la
constitution de gros agrégats stables dont la taille s’apparente aux sables
granulométriques (Nishita and Haug 1972 ; Giovannini et al., 1988 ; Nzila and Garnier-
Zarli, 1992 ; Mboukou and Lavelle, 1997). Ces processus qui correspondent à ceux
intervenant dans la fabrication de briques expliquent également la légère augmentation
de stabilité observée.
Ainsi si la stabilité structurale de la muraille native provient du liant salivaire entre
les agrégats organominéraux, celle de la muraille brûlée est issue de la pectisation des
argiles en pseudosables.
L’alcalinisation de la muraille brûlée s’accompagne d’une forte réduction des
concentrations en bases alcalines et en aluminium échangeable. L’aluminium
échangeable de la solution est sans doute immobilisé sous forme d’hydroxydes et dans
les argiles.
En revanche, la concentration de phosphore est presque multipliée par 4. Cette
augmentation de la concentration de phosphore est sans doute due à une augmentation
de son assimilabilité. Dans les milieux acides, riches en oxyhydroxydes, le phosphore est
absorbé par le fer au sein de cristaux amorphes. L’augmentation de la température
provoque une réorganisation des hydroxydes de fer et d’aluminium provoquant une
désorption du phosphore qui devient alors assimilable.
Rôle du termite Cornitermes sp. dans le fonctionnement du sol.
L’importance des termites dans l’écosystème tropical n’ est plus à démontrer : ils
constituent souvent la biomasse faunique la plus importante et sont un facteur majeur de
la dynamique du sol (Maldague, 1959 ; Garnier-Sillam, 1987 ; Lal, 1988 ; Lobry De
Bruyn and Conacher, 1990). Comme dans d’autres études sur le termite

Termitières du genre Cornitermes, discussion. Chapitre V.3
121
champignonniste Macrotermes (Jones, 1990 ; Garnier-Sillam and Harry, 1995), le
termite xylophage Cornitermes sp affinis ovatus a un impact physico-chimique sur le sol :
1 - localement (action sur le sol au niveau de la termitière). En sélectionnant leurs
matériaux de construction, une migration d’éléments minéraux s’opère vers la surface
(enrichissement en argile). A ce phénomène s’ajoute l’enrichissement en matière
organique au niveau du feuilleté, qui est piégé de manière durable à l’intérieur de la
colonie.
2 - au niveau parcellaire, par la densité des colonies. La densité en colonie influe
fortement sur le stock de matière organique piégé au sein des différentes structures et
peut, dans une certaine mesure, retarder les processus d’humification des horizons de
surface.
Dans la littérature, l’impact des termites se manifeste différemment sur le cycle
de la matière organique en fonction du groupe trophique étudié.(Lee and Wood, 1971 ;
Lee and Butler, 1977 ; Lafage and Nutting, 1978 ; Garnier-Sillam and Renoux, 1989).
-Les humivores mélangent la matière organique humifiée et les particules minérales du
sol. Ils créent dans leur tube digestif des microagrégats organominéraux stables qui,
incorporés au sol et dans les murailles de la termitière, influencent favorablement le
complexe argilo-humique. Leur action biologique améliore la perméabilité, la stabilité
structurale et en corollaire la fertilité des horizons hémi-organiques. Il participent à la
conversion des colloïdes organiques du sol (Arshad et al., 1988).
-Les champignonnistes, tel que Macrotermes sp. appauvrissent fortement l’horizon
humifère autour de leur nid, par les prélèvements intensifs qu’ils opèrent dans la litière et
la faible restitution de matière organique qu’ils opèrent dans l’écosystème. Ils
déstabilisent leur aire de récolte et limitent les processus d’humification (Garnier-Sillam
and Renoux, 1989).
-Dans le cas du termite xylophage Cornitermes, la sélection des éléments minéraux et
organiques dont il a besoin pour construire les termitières et la séquestration intensive
de la matière organique dans le feuilleté interne lui confèrent un rôle qui s’apparenterait
davantage à celui des champignonnistes. La durée de vie des termitières est mal connue
mais considérant que la durée de vie des termitières d’espèces humivores en Afrique
atteint 25 ans (Nye, 1955 ; Collins, 1981 ; Lepage, 1984 ; Janeau and Valentin, 1987);
les termitières de Cornitermes sp. favorisent certainement une réduction locale de la
fertilité et donc la dégradation des pâturages amazoniens. Les pâturages étudiés ne sont
âgés que de 5 à 6 ans, Il est par conséquent nécessaire de considérer le temps durant
lequel les colonies de termites peuvent agir sur le sol. Il apparaît donc nécessaire
d’étudier l’évolution des populations de Cornitermes sp. au cours du temps (densité dans

Termitières du genre Cornitermes, discussion. Chapitre V.3
122
des pâturages d’au moins 15 ans). Mais d’ores et déjà nous pouvons affirmer que son
impact présente deux effets majeurs :
- un effet mécanique, par la remontée des matériaux argileux de l’horizon 10-30 cm, en
surface, accompagnée d’une aération dans la termitosphère (galeries)
-un effet soustractif de la matière organique, par prélèvement important du bois
(notamment de surface).Les déjections sont séquestrés dans le feuilleté du nid et ne
seront rendues à l’écosystème qu’à la mort de la termitière. Nous pouvons supposer que
cette matière organique sera, à ce moment là, reprise rapidement par les
microorganismes du sol (Garnier-Sillam, 1987).
Cette espèce est considérée dans le site d’étude comme l’une des principales
pestes agricoles (Constantino, 2002). Même si les termites sont souvent considérés
comme néfastes en raison des dommages qu’ils génèrent à l’agriculture, ils contribuent
aux processus d’humification du sol dans les tropiques. Les composes ligneux ingérés
sont soumis, pendant un transit séquentiel, à différents processus de dégradation
chimique (hydrolyse alcaline) et une dégradation microbiologique (fermentation,
respiration anaérobique et minéralisation) (Brauman, 2000).
Pendant la digestion, la matière organique du sol est fortement modifiée en
termes de nature (concentration en matière organique, rapport entre les acides fulviques
et humiques) et organisation (formation de complexes organominéraux avec l’argile
ingéré simultanément). Les déjections sont incluses dans le nid et les galeries qui,
comme un tout, constituent la termitosphère.
Comparée au sol témoin, la matière organique de la termitosphère est plus stable
et protégée de la minéralisation intense qui s’opère sous les tropiques.
Compte tenu de la densité des termites dans les régions tropicales, les
modifications de la matière organique dans le long turnover généré par leur digestion et
le dépôt dans la termitosphère montrent que ces insectes ont un rôle important sur le
cycle de la matière organique.

Etude des turricules de l’annélide Andiodrilus Pachoensis. Chapitre V1
123
VI. Etude des turricules de l’Annélide Andiodrilus
Pachoensis.
L’objectif de cette étude est de mettre en évidence le rôle particulier de l’annélide
ingénieur Andiodrilus pachoensis dans le fonctionnement du sol. Les interactions
turricules-sol sont présentées dans ce chapitre.
• Dans un premier temps, les densités et les distributions spatiales des
turricules dans les forêts et les pâturages ont été étudiés.
• Dans un second temps, l’analyse des propriétés physico-chimiques des
turricules a été effectuée.
• Dans un troisième temps, les effets des turricules sur le sol ont été abordés
par l’étude de leur dégradation. Les effets des pratiques agricoles et des conditions
environnementales sur ces structures biogéniques et plus particulièrement ceux du feu
et de la pluie ont été analysés.

Turricules d’Andiodrilus pachoensis, matériel et méthodes. Chapitre V1.1
124
VI.1 MATERIEL ET METHODES
VI.1.1 L’espèce Andiodrilus pachoensis.
L’espèce Andiodrilus pachoensis fait partie des Annélides anéciques de la famille
des Glossoscolecidae. Il s’agit d’un annélide pigmenté de grande taille.
Le genre Andiodrilus est originaire d’Amérique du Sud et comprend l’immense
majorité des espèces rencontrées en Amazonie avec 35 genres et des centaines
d’espèce. La présence de ce genre dans la région est confirmée par James et Brown
(James and Brown, 2006).
Les turricules étudiés sont constitués par d’une accumulation successive de
déjections de taille centimétrique. Les turricules sont creusés d’un canal central bien
conservé qui persiste au cours du temps. (Tout au long de cette étude, le terme de
turricule désignera cette accumulation de déjections.)
Photo 5 : Andiodrilus pachoensis et turricules.
VI.1.2 Cartographie des turricules d’Andiodrilus pachoensis
La cartographie des turricules a été réalisée dans toutes les parcelles excepté le
pâturage P06 soit 10 parcelles.

Turricules d’Andiodrilus pachoensis, matériel et méthodes. Chapitre V1.1
125
Le travail de cartographie s’est étalé sur 2002 et 2003 durant la saison des pluies
(janvier –juillet).
Ce travail est divisé en 3 axes majeurs :
1. Etude de la distribution spatiale des turricules (2002)
2. Suivi temporel de production de turricules (2002 – 2003)
3. Expériences de calcination in situ et simulateur de pluie en laboratoire (2003)
1. Distribution spatiale des turricules (2002).
Ce travail a été réalisé dans 2 forêts (F01, F04), deux pâturages (P01, P02), 1
jachère (J01) et une rizière (R01).
Pour rappel, 3 milieux ont été comparés : la forêt, la forêt dégradée et le pâturage.
Dans chaque parcelle, une aire de 50 m² (5x10m) a été choisie. La zone d’étude ainsi
définie a été ensuite découpée selon un maillage régulier en carrés de 1m² (1x1m). Dans
chaque carré, toutes les structures biogéniques et principalement les turricules de vers
ont été répertoriées et localisées géographiquement.
Les éléments de la végétation (touffes, arbres, troncs, branches, zone nue…) ont
également été répertoriés.
Toutes les structures biogéniques ont été collectées puis pesées séparément
(illustration 3).
En 2003 une aire de 25m² a également été analysée dans un autre pâturage (P04).
Le détail des analyses statistiques a été présenté précédemment (chapitre III.1).
2. Suivi temporel de production de turricules (2002 – 2003).
Dans cette étude, réalisée en 2003, la cartographie des turricules a été entreprise
dans l’objectif d’observer la production de turricules au cours du temps sur une surface
de 25 m² (aire de 5x5m).
Seules deux aires étudiées en 2002 (la forêt F01 et le pâturage P01) ont été
retrouvées. La végétation dans ces deux zones d’étude était comparable à celle de
l’année précédente. Le suivi temporel a ainsi été réalisé dans ces deux parcelles.
Dans le but de s’assurer des résultats, cette étude a été répétée dans deux forêts
(F02 et F03) et trois autres pâturages (P03, P04 et P05) ayant respectivement un
couvert végétal similaire à la forêt F01 et au pâturage P01 (Tableau 14).
Turricules d’Andiodrilus pachoensis, matériel et méthodes. Chapitre V1.1
126
Parcelles étudiées afin de réaliser la cartographie des
turricules d’Andiodrilus pachoensis
Année 2002 (50 m²) 2003 (25m²)
Couvert végétal Pâturage Forêt Pâturage Forêt
Sr Amiute F04 (30m²)
Sr Cicéro P05
Sr Francisco
Sr Josuel P02 P03
Sr Léonine P04 F03
Sr Mélète F02
Sr Osano P01 F01 P01 F01
Tableau 14 : Localisation (nom du propriétaire de l’exploitation) et type de couvert végétal des diverses
parcelles étudiées pour cartographier les turricules d’Andiodrilus pachoensis.
Dans ces parcelles, selon le même principe que la cartographie réalisée en 2002, tous
les turricules ont été marqués à l’aide de bâtonnets sur lesquels a été inscrite la date
d’observation.
Trois marquages des turricules en 3 mois ont été réalisés avant que tous les
turricules soient répertoriés géographiquement en fonction de leur date d’observation.
Nous avons ainsi obtenu des informations concernant la production des turricules sur
3 mois durant la période des pluies dans toutes les parcelles.
Dans les parcelles P01 et F01, la production en 10 mois a également été calculée (fin
de période des pluies 2002, période sèche 2002-2003 jusqu’au début de la saison des
pluies 2003).
3. Expériences (2003).
Les expérimentations ont été menées dans 2 forêts (F01, F02) et 3 pâturages (P01,
P02 et P04) suivant les premières observations réalisées en 2002.
• Sous forêt, nous avons testé le rôle de la quantité de litière sur la production de
turricules. Dans les deux forêts, tous les turricules sur une aire de 25 m² (5x5m) ont été

Turricules d’Andiodrilus pachoensis, matériel et méthodes. Chapitre V1.1
127
cartographiés. La production a été suivie sur 10m² témoin (2X5m) avec une quantité de
litière non modifiée ; 5m² (5x1m) recouvert d’une quantité réduite de litière et 10m²
(5x2m) sans litière. Ce suivi a été réalisé durant 3 mois (3 observations).
• Sous pâturages, l’effet du couvert des touffes de Brachiaria brisantha sur la
production de turricules a été testé. Dans les 3 pâturages, tous les turricules d’une aire
de 50 m² ont été répertoriés géographiquement. Nous avons comparé la production de
turricules entre une aire de 25 m² (5x5m) témoins avec végétation et une aire de 25m²
(5x5m) où toute la végétation a été retirée le premier jour de l’expérimentation laissant
le sol nu. Cette comparaison a été effectuée durant 3 mois et 3 observations ont été
réalisées tout en laissant repousser les touffes de Brachiaria au cours de
l’expérimentation.
1) Pâturage
2) Forêt1)
2)
1) Pâturage
2) Forêt1)
2)
Illustration 3: Protocole de cartographie et échantillonnage des turricules dans le pâturage et dans la forêt.
VI.1.3 Analyse des propriétés physico-chimiques des turricules
Ces mesures ont été effectuées à partir de 3 turricules prélevés dans chaque parcelle des
deux milieux et comparées avec 3 échantillons témoins issus des 3 forêts (F01, F02 et
F03) et 3 pâturages (P01, P03 et P04).
Le détail des analyses physiques et chimiques a été présenté dans le chapitre III.2

Turricules d’Andiodrilus pachoensis, matériel et méthodes. Chapitre V1.1
128
VI.1.4 Expérience de simulation de l’impact du feu sur les turricules d’Andiodrilus
pachoensis.
Dans une fosse, des turricules d’Andiodrilus pachoensis de forêt (F01, F02 et F03)
et de pâturage (P01, P02 et P04) sont placés sur un gradient de paille sèche avec
présence ou non de bois mort. Ce gradient de paille et bois mort, ainsi mis en place dans
la fosse, provoque en brûlant, un gradient d’intensité et de durée de la période
d’exposition au feu des structures (illustration 3).
Gradient d’intensité et durée du feu
Turricules sur lit de paille sèche
Turricules au sein d’un amas de bois mort
Illustration 4 : Expérience de brûlage des turricules
Ce gradient reflète les conditions naturelles de feu de pâturage avec des zones
plus ou moins couvertes de touffes de végétation et la présence de troncs coupés ou de
souches de bois. Deux états de turricules brûlés sont ainsi observés :
1. Etat noirci : Brûlé +/- (B+/-)
2. Etat rouge brique : Brûlé +/+ (B+/+)

Turricules d’Andiodrilus pachoensis, matériel et méthodes. Chapitre V1.1
129
VI.1.4 Expérience de simulation de l’impact de la pluie sur les turricules
Un simulateur de pluie a été fabriqué (Illustrations 4). Des turricules et du sol
témoin des deux milieux (3 échantillons par type) sont soumis à différents temps
d’exposition de pluies simulées sous deux états (normal et écrasé). Deux hauteurs de
chute des gouttelettes de pluie ont été choisies afin de simuler deux forces de pluie :
•La position haute (F1) correspond à environ 30 cm entre la sortie des gouttelettes et la
structure. Elle simule une faible pluie (quantité importante mais force faible).
•La position basse (F2) correspond à environ 85 cm entre la sortie des gouttelettes et la
structure. Elle simule une forte pluie (quantité plus faible mais force plus élevée).
•L’état écrasé correspond à une structure écrasée entre 2 plaques de fer sous la force de
la main.
Le dispositif expérimental ne nous a permis de simuler qu’une pluie théorique très forte
d’environ 106 ml.min-1.
Méthode d’écrasement
Dispositif Position haute Position basse
Méthode d’écrasement
Dispositif Position haute Position basse
Haut / Bas3Ecrasé
Haut / Bas1 / 3 / 5 Normal
PositionTemps d’exposition
(mn)Etat
Structure
Haut / Bas3Ecrasé
Haut / Bas1 / 3 / 5 Normal
PositionTemps d’exposition
(mn)Etat
Structure
Illustration 5 : Dispositif expérimental de simulation de pluie
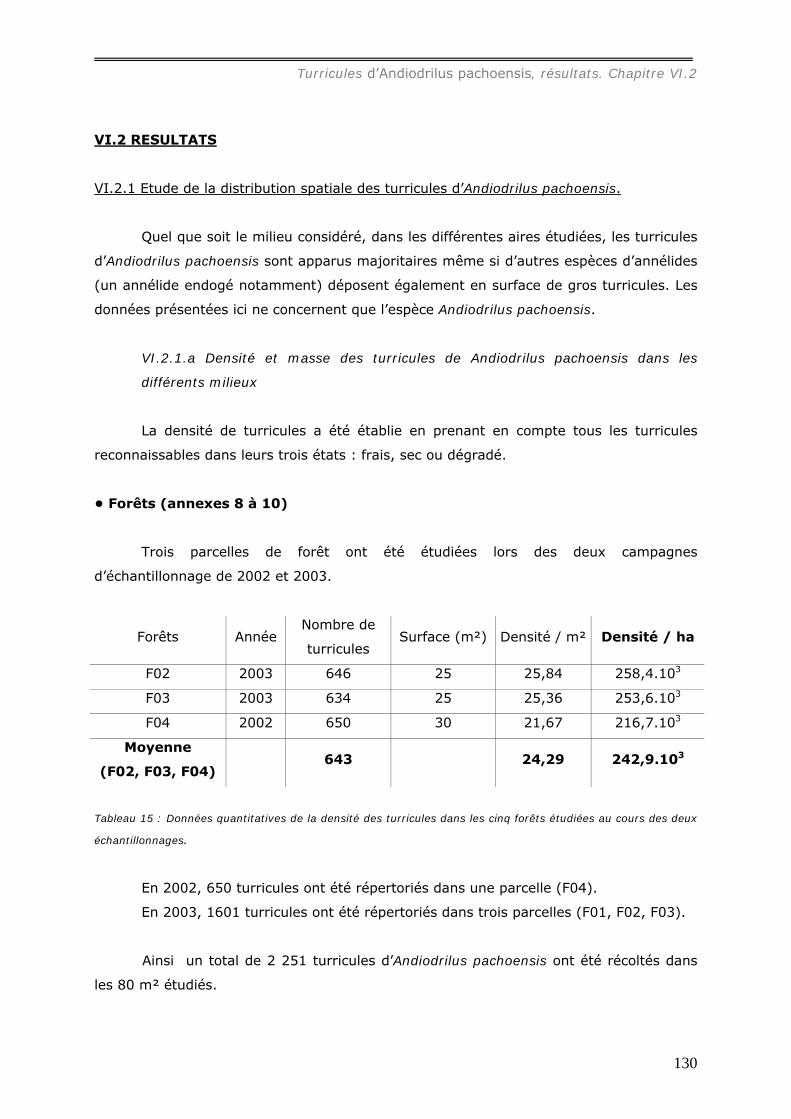
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
130
VI.2 RESULTATS
VI.2.1 Etude de la distribution spatiale des turricules d’Andiodrilus pachoensis.
Quel que soit le milieu considéré, dans les différentes aires étudiées, les turricules
d’Andiodrilus pachoensis sont apparus majoritaires même si d’autres espèces d’annélides
(un annélide endogé notamment) déposent également en surface de gros turricules. Les
données présentées ici ne concernent que l’espèce Andiodrilus pachoensis.
VI.2.1.a Densité et masse des turricules de Andiodrilus pachoensis dans les
différents milieux
La densité de turricules a été établie en prenant en compte tous les turricules
reconnaissables dans leurs trois états : frais, sec ou dégradé.
• Forêts (annexes 8 à 10)
Trois parcelles de forêt ont été étudiées lors des deux campagnes
d’échantillonnage de 2002 et 2003.
Forêts Année Nombre de
turricules Surface (m²) Densité / m² Densité / ha
F02 2003 646 25 25,84 258,4.103
F03 2003 634 25 25,36 253,6.103
F04 2002 650 30 21,67 216,7.103
Moyenne
(F02, F03, F04) 643 24,29 242,9.103
Tableau 15 : Données quantitatives de la densité des turricules dans les cinq forêts étudiées au cours des deux
échantillonnages.
En 2002, 650 turricules ont été répertoriés dans une parcelle (F04).
En 2003, 1601 turricules ont été répertoriés dans trois parcelles (F01, F02, F03).
Ainsi un total de 2 251 turricules d’Andiodrilus pachoensis ont été récoltés dans
les 80 m² étudiés.
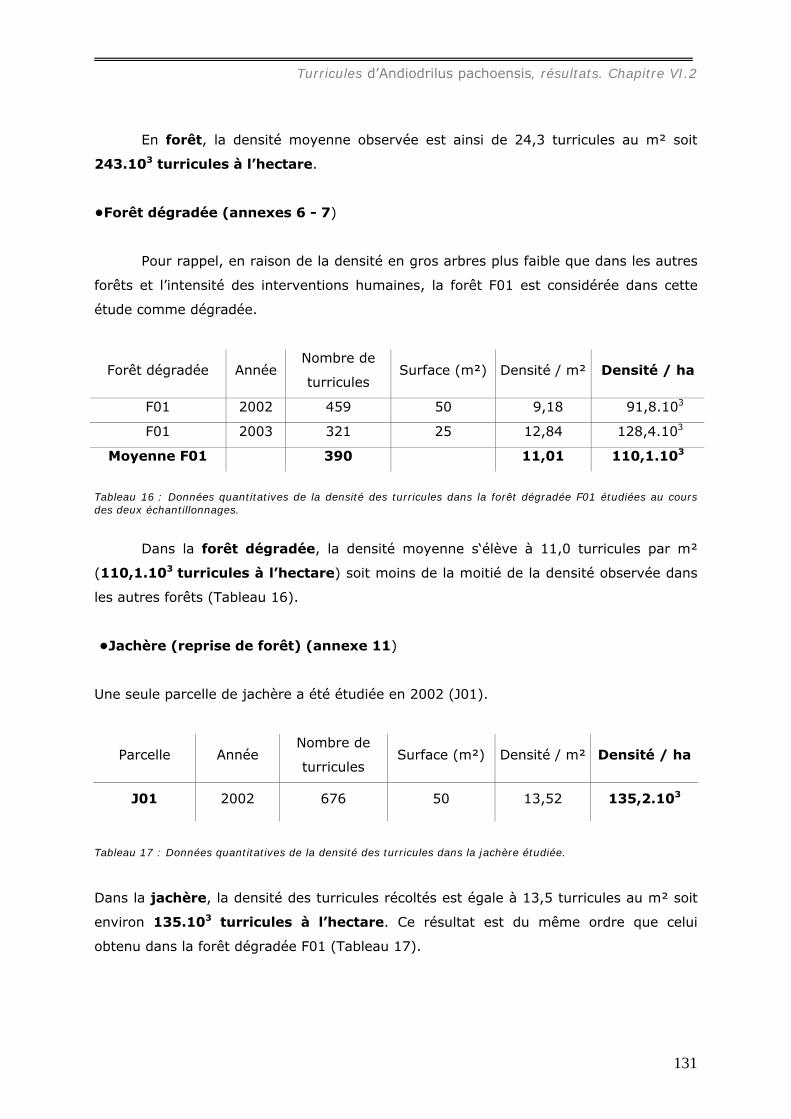
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
131
En forêt, la densité moyenne observée est ainsi de 24,3 turricules au m² soit
243.103 turricules à l’hectare.
•Forêt dégradée (annexes 6 - 7)
Pour rappel, en raison de la densité en gros arbres plus faible que dans les autres
forêts et l’intensité des interventions humaines, la forêt F01 est considérée dans cette
étude comme dégradée.
Forêt dégradée Année Nombre de
turricules Surface (m²) Densité / m² Densité / ha
F01 2002 459 50 9,18 91,8.103
F01 2003 321 25 12,84 128,4.103
Moyenne F01 390 11,01 110,1.103
Tableau 16 : Données quantitatives de la densité des turricules dans la forêt dégradée F01 étudiées au cours des deux échantillonnages.
Dans la forêt dégradée, la densité moyenne s‘élève à 11,0 turricules par m²
(110,1.103 turricules à l’hectare) soit moins de la moitié de la densité observée dans
les autres forêts (Tableau 16).
•Jachère (reprise de forêt) (annexe 11)
Une seule parcelle de jachère a été étudiée en 2002 (J01).
Parcelle Année Nombre de
turricules Surface (m²) Densité / m² Densité / ha
J01 2002 676 50 13,52 135,2.103
Tableau 17 : Données quantitatives de la densité des turricules dans la jachère étudiée.
Dans la jachère, la densité des turricules récoltés est égale à 13,5 turricules au m² soit
environ 135.103 turricules à l’hectare. Ce résultat est du même ordre que celui
obtenu dans la forêt dégradée F01 (Tableau 17).
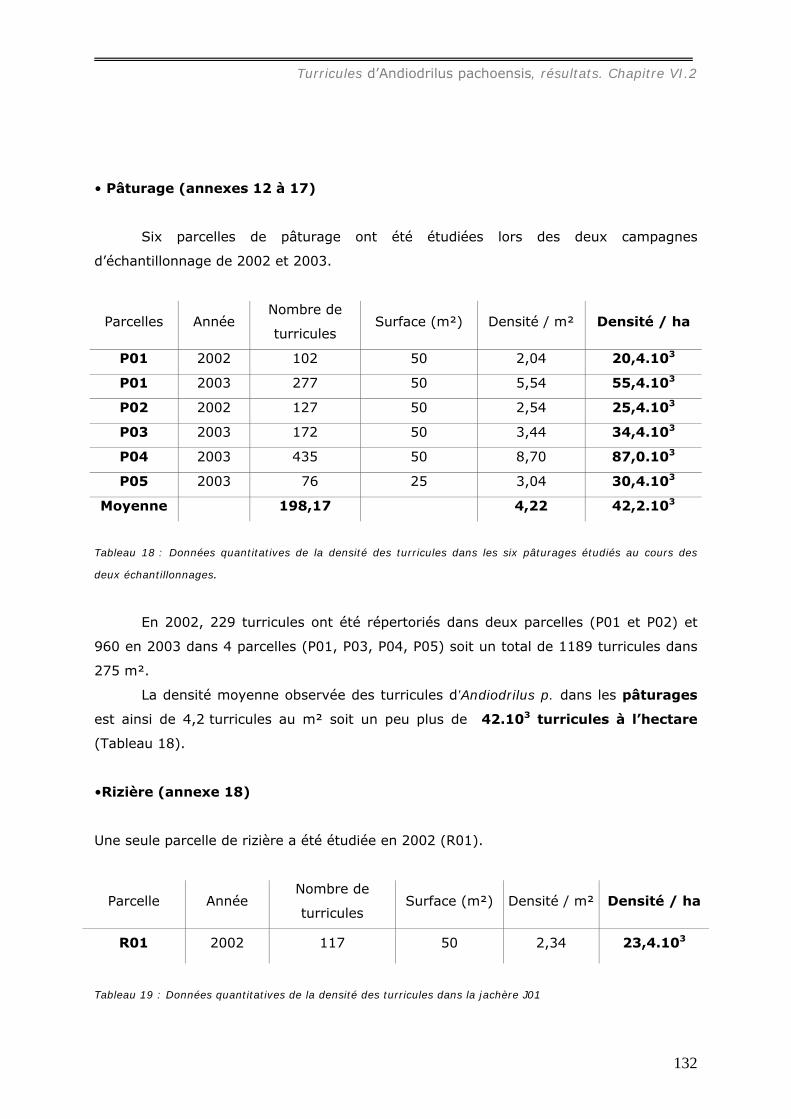
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
132
• Pâturage (annexes 12 à 17)
Six parcelles de pâturage ont été étudiées lors des deux campagnes
d’échantillonnage de 2002 et 2003.
Parcelles Année Nombre de
turricules Surface (m²) Densité / m² Densité / ha
P01 2002 102 50 2,04 20,4.103
P01 2003 277 50 5,54 55,4.103
P02 2002 127 50 2,54 25,4.103
P03 2003 172 50 3,44 34,4.103
P04 2003 435 50 8,70 87,0.103
P05 2003 76 25 3,04 30,4.103
Moyenne 198,17 4,22 42,2.103
Tableau 18 : Données quantitatives de la densité des turricules dans les six pâturages étudiés au cours des
deux échantillonnages.
En 2002, 229 turricules ont été répertoriés dans deux parcelles (P01 et P02) et
960 en 2003 dans 4 parcelles (P01, P03, P04, P05) soit un total de 1189 turricules dans
275 m².
La densité moyenne observée des turricules d’Andiodrilus p. dans les pâturages
est ainsi de 4,2 turricules au m² soit un peu plus de 42.103 turricules à l’hectare
(Tableau 18).
•Rizière (annexe 18)
Une seule parcelle de rizière a été étudiée en 2002 (R01).
Parcelle Année Nombre de
turricules Surface (m²) Densité / m² Densité / ha
R01 2002 117 50 2,34 23,4.103
Tableau 19 : Données quantitatives de la densité des turricules dans la jachère J01
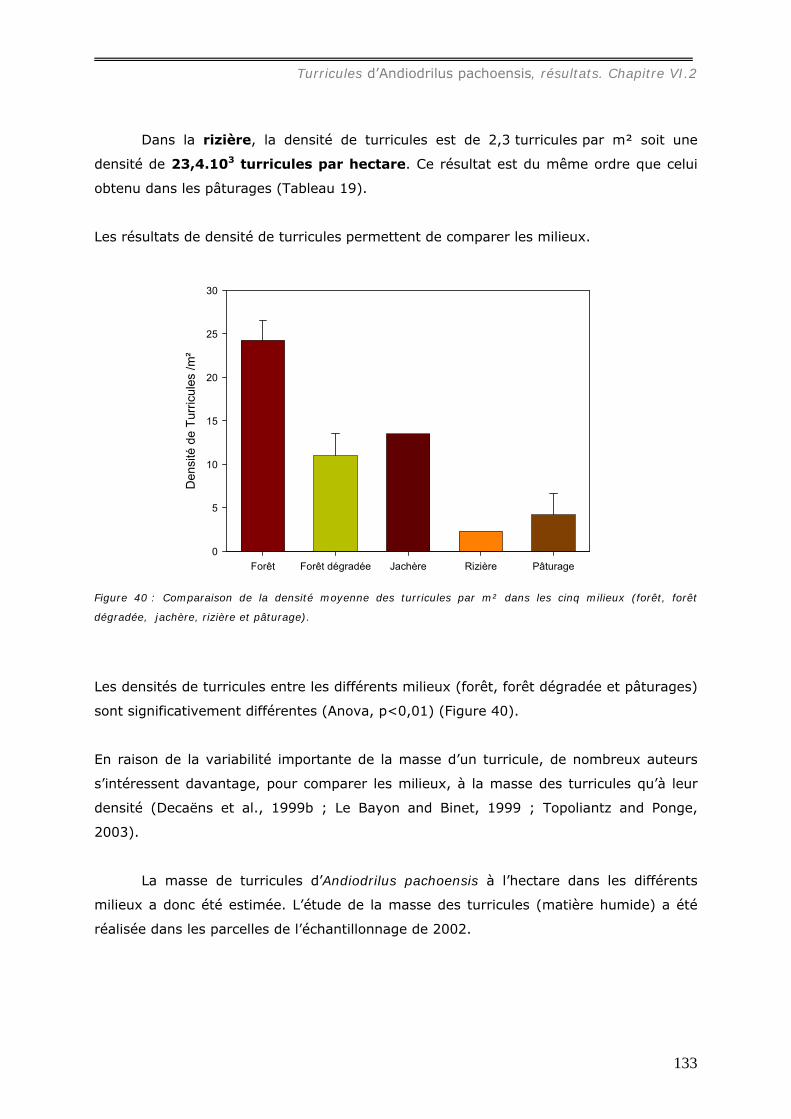
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
133
Dans la rizière, la densité de turricules est de 2,3 turricules par m² soit une
densité de 23,4.103 turricules par hectare. Ce résultat est du même ordre que celui
obtenu dans les pâturages (Tableau 19).
Les résultats de densité de turricules permettent de comparer les milieux.
Forêt Forêt dégradée Jachère Rizière Pâturage
Den
sité
de
Turr
icul
es /m
²
0
5
10
15
20
25
30
Figure 40 : Comparaison de la densité moyenne des turricules par m² dans les cinq milieux (forêt, forêt
dégradée, jachère, rizière et pâturage).
Les densités de turricules entre les différents milieux (forêt, forêt dégradée et pâturages)
sont significativement différentes (Anova, p<0,01) (Figure 40).
En raison de la variabilité importante de la masse d’un turricule, de nombreux auteurs
s’intéressent davantage, pour comparer les milieux, à la masse des turricules qu’à leur
densité (Decaëns et al., 1999b ; Le Bayon and Binet, 1999 ; Topoliantz and Ponge,
2003).
La masse de turricules d’Andiodrilus pachoensis à l’hectare dans les différents
milieux a donc été estimée. L’étude de la masse des turricules (matière humide) a été
réalisée dans les parcelles de l’échantillonnage de 2002.
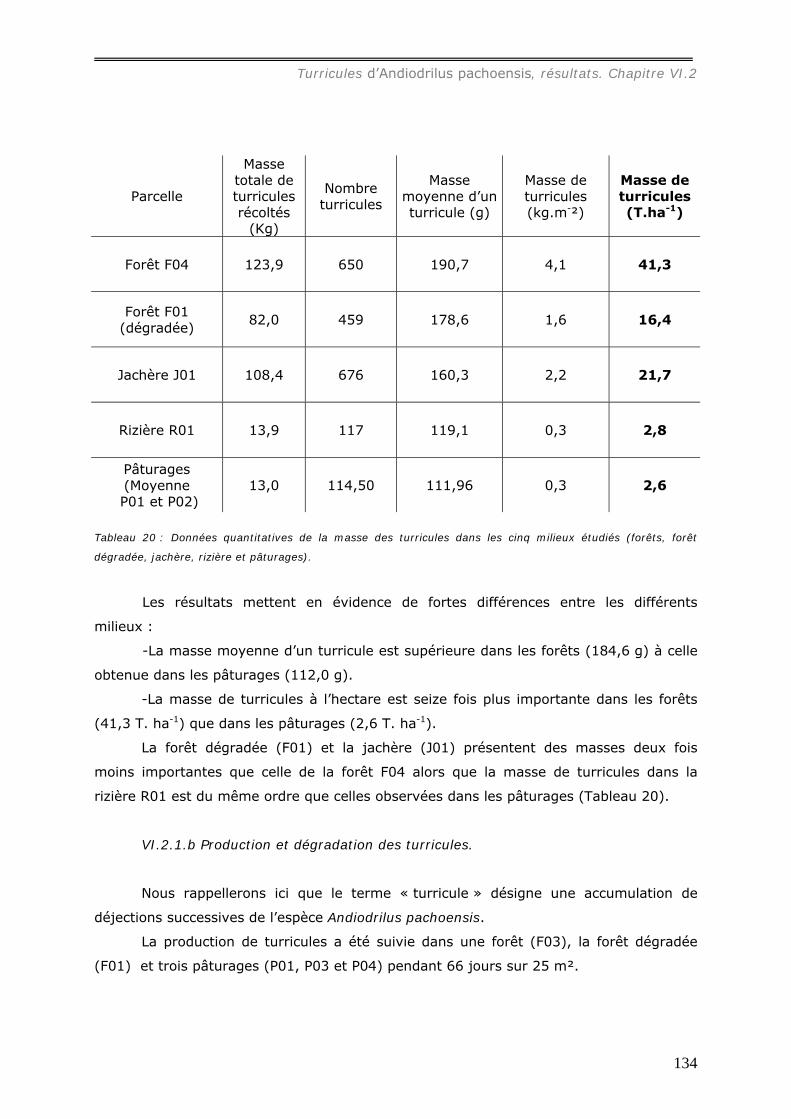
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
134
Parcelle
Masse totale de turricules récoltés
(Kg)
Nombre turricules
Masse moyenne d’un turricule (g)
Masse de turricules (kg.m-²)
Masse de turricules (T.ha-1)
Forêt F04 123,9 650 190,7 4,1 41,3
Forêt F01 (dégradée)
82,0 459 178,6 1,6 16,4
Jachère J01 108,4 676 160,3 2,2 21,7
Rizière R01 13,9 117 119,1 0,3 2,8
Pâturages (Moyenne
P01 et P02) 13,0 114,50 111,96 0,3 2,6
Tableau 20 : Données quantitatives de la masse des turricules dans les cinq milieux étudiés (forêts, forêt
dégradée, jachère, rizière et pâturages).
Les résultats mettent en évidence de fortes différences entre les différents
milieux :
-La masse moyenne d’un turricule est supérieure dans les forêts (184,6 g) à celle
obtenue dans les pâturages (112,0 g).
-La masse de turricules à l’hectare est seize fois plus importante dans les forêts
(41,3 T. ha-1) que dans les pâturages (2,6 T. ha-1).
La forêt dégradée (F01) et la jachère (J01) présentent des masses deux fois
moins importantes que celle de la forêt F04 alors que la masse de turricules dans la
rizière R01 est du même ordre que celles observées dans les pâturages (Tableau 20).
VI.2.1.b Production et dégradation des turricules.
Nous rappellerons ici que le terme « turricule » désigne une accumulation de
déjections successives de l’espèce Andiodrilus pachoensis.
La production de turricules a été suivie dans une forêt (F03), la forêt dégradée
(F01) et trois pâturages (P01, P03 et P04) pendant 66 jours sur 25 m².
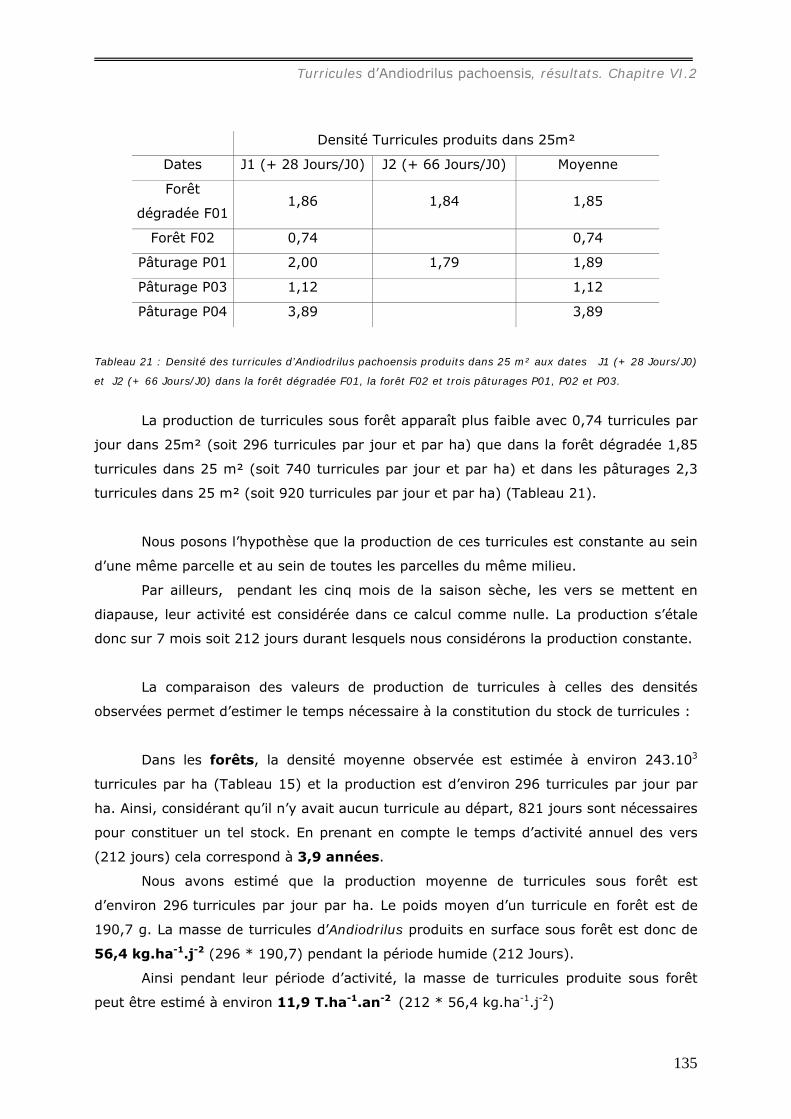
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
135
Densité Turricules produits dans 25m²
Dates J1 (+ 28 Jours/J0) J2 (+ 66 Jours/J0) Moyenne
Forêt
dégradée F01 1,86 1,84 1,85
Forêt F02 0,74 0,74
Pâturage P01 2,00 1,79 1,89
Pâturage P03 1,12 1,12
Pâturage P04 3,89 3,89
Tableau 21 : Densité des turricules d’Andiodrilus pachoensis produits dans 25 m² aux dates J1 (+ 28 Jours/J0)
et J2 (+ 66 Jours/J0) dans la forêt dégradée F01, la forêt F02 et trois pâturages P01, P02 et P03.
La production de turricules sous forêt apparaît plus faible avec 0,74 turricules par
jour dans 25m² (soit 296 turricules par jour et par ha) que dans la forêt dégradée 1,85
turricules dans 25 m² (soit 740 turricules par jour et par ha) et dans les pâturages 2,3
turricules dans 25 m² (soit 920 turricules par jour et par ha) (Tableau 21).
Nous posons l’hypothèse que la production de ces turricules est constante au sein
d’une même parcelle et au sein de toutes les parcelles du même milieu.
Par ailleurs, pendant les cinq mois de la saison sèche, les vers se mettent en
diapause, leur activité est considérée dans ce calcul comme nulle. La production s’étale
donc sur 7 mois soit 212 jours durant lesquels nous considérons la production constante.
La comparaison des valeurs de production de turricules à celles des densités
observées permet d’estimer le temps nécessaire à la constitution du stock de turricules :
Dans les forêts, la densité moyenne observée est estimée à environ 243.103
turricules par ha (Tableau 15) et la production est d’environ 296 turricules par jour par
ha. Ainsi, considérant qu’il n’y avait aucun turricule au départ, 821 jours sont nécessaires
pour constituer un tel stock. En prenant en compte le temps d’activité annuel des vers
(212 jours) cela correspond à 3,9 années.
Nous avons estimé que la production moyenne de turricules sous forêt est
d’environ 296 turricules par jour par ha. Le poids moyen d’un turricule en forêt est de
190,7 g. La masse de turricules d’Andiodrilus produits en surface sous forêt est donc de
56,4 kg.ha-1.j-2 (296 * 190,7) pendant la période humide (212 Jours).
Ainsi pendant leur période d’activité, la masse de turricules produite sous forêt
peut être estimé à environ 11,9 T.ha-1.an-2 (212 * 56,4 kg.ha-1.j-2)

Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
136
Dans la forêt dégradée F01, des calculs identiques sur les densités que ceux
réalisés dans les forêts permettent d’évaluer à 148,8 jours (110,1. 103 turricules.ha-1 /
740 turricules.ha-1), le temps de constitution du stock de turricules soit 70% d’une
année (212 jours). La production moyenne est de 740 turricules à l’hectare. La masse
moyenne d’un turricule dans la forêt dégradée est de 178,6 g. Ainsi la production est de
132,2 kg.ha-1.j-2 (740 * 178,6) soit environ 28 T.ha-1.an-2 (132,2 * 212)
Dans les pâturages, le temps de constitution du stock est évalué à 46 jours
(42,2.103 turricules.ha-1 / 920 turricules.ha-1 par jour), soit 21,7% de l’année. La
production moyenne est d’environ 920 turricules à l’hectare. La masse moyenne d’un
turricule de pâturage étant de 112,0 g, cela représente une masse de 103 kg.ha-1.j-2
(920 * 112,0) pendant la période humide et donc au minimum 21,8 T.ha-1.an-2 (103 *
212).
Ces résultats impliquent que dans les forêts, des turricules ont une durée de vie
supérieure à l’année (3,9 années) alors que dans les forêts dégradées (0,7 année) et les
pâturages (0,21 année) les chiffres sont très en deçà. Ce résultat met également en
évidence la forte dégradation des turricules dans les pâturages.
L’observation de la masse des turricules à un temps t et l’estimation de celle
produite en un an mettent en exergue de fortes différences :
-Sous forêt, la masse de turricules estimée est de 41,3T et la production annuelle
estimée est de 11,9T. De ce fait, la masse observée à un temps t représente 347 % de
la production estimée.
- Sous forêt dégradée, la masse de turricules est estimée à 16,4T et la
production annuelle est estimée à 28 T. Ainsi la masse représente environ 58% de la
production.
-Dans les pâturages elle est de l’ordre de 11,9% de la production (la densité
de turricules observée est de 2,6T et la production annuelle estimée est de 21,8T). La
production annuelle, ainsi estimée, apparaît faible par rapport à la masse de turricules
observées. La différence entre densité et production ne peut s’expliquer que par une
forte dégradation dans un délai relativement court (de l’ordre de 3 à 5 semaines).
Ce résultat est compatible avec celui obtenu lors de l’étude réalisé sur
Martiodrilus c. en Colombie qui évalue à plus de 75% la proportion de turricules cassés
ou désagrégés (Decaëns, 2000)

Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
137
La densité estimée des turricules dans les forêts est d’environ
243.103 turricules par hectare. Dans la forêt dégradée (F01), elle est diminuée de
plus de la moitié (environ 110.103 turricules à l’hectare). Dans les pâturages, la
densité des turricules est fortement réduite mais variable d’un pâturage à l’autre
(42,2.103 turricules ± 22 ,9103 par hectare).
L’évaluation de la masse des turricules a mis en évidence une masse
moyenne d’un turricule supérieure dans les forêts (184,6 g) que dans les
pâturages (112,0 g). Par ailleurs, la masse des turricules à l’hectare est apparue
seize fois plus importante dans les forêts (41,3 T. ha-1) que dans les pâturages (2,6
T. ha-1).
La production de turricules sous forêt est beaucoup plus faible 296 turricules
par jour et par hectare que dans les forêts dégradées (740 turricules par jour et par
ha) et dans les pâturages (920 turricules par jour et par hectare). En comparant ces
résultats à ceux de la densité observée, il apparaît :
1) Dans les forêts, les turricules ont une durée de vie (persistance)
supérieure à l’année alors que dans les forêts dégradées et les pâturages les durées
sont moindres.
2) la forte production dans les forêts dégradées (plus de 2 fois celle de forêt)
met en évidence une stimulation de production de turricules qui peut être reliée à
une augmentation de l’espace disponible suite à une plus forte dégradation.
3) La différence entre densité et production dans les pâturages ne peut
s’expliquer que par une forte dégradation (88% des turricules dégradés ou détruits)
dans un délai relativement court (de l’ordre de 3 à 5 semaines).
Trois facteurs environnementaux peuvent expliquer la dégradation observée dans
les pâturages : la pluie, le feu (culture sur brulis) ou le piétinement bovin. Le rôle de ces
facteurs sera analysé ultérieurement.
Pour comprendre l’effet local sur le sol de la dégradation des turricules,
principalement dans les pâturages, nous avons tout d’abord étudié la distribution spatiale
de ces structures biogéniques.
VI.2.1.c Distribution spatiale des turricules d’Andiodrilus pachoensis.
Compte tenu de la masse importante des turricules d’Andiodrilus pachoensis
récoltés dans le site d’étude et de la forte dégradation qui s’opère dans les pâturages,
nous avons essayé de vérifier si cette incorporation de turricules influencent localement
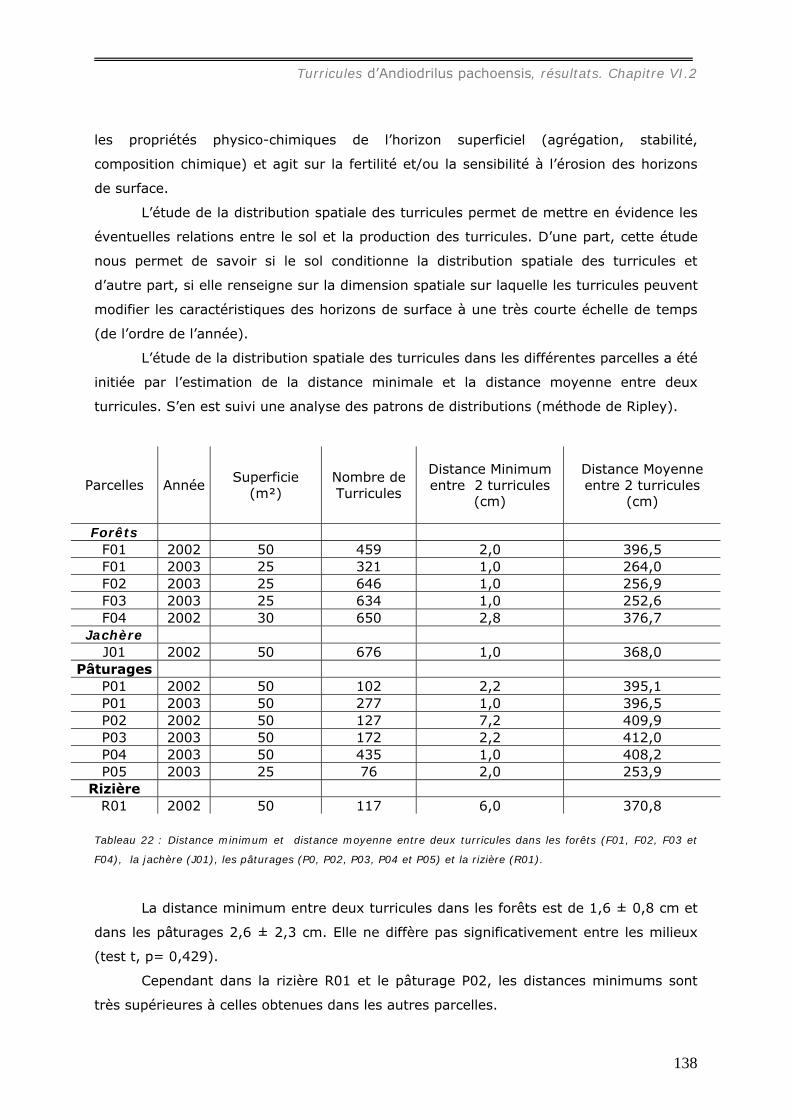
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
138
les propriétés physico-chimiques de l’horizon superficiel (agrégation, stabilité,
composition chimique) et agit sur la fertilité et/ou la sensibilité à l’érosion des horizons
de surface.
L’étude de la distribution spatiale des turricules permet de mettre en évidence les
éventuelles relations entre le sol et la production des turricules. D’une part, cette étude
nous permet de savoir si le sol conditionne la distribution spatiale des turricules et
d’autre part, si elle renseigne sur la dimension spatiale sur laquelle les turricules peuvent
modifier les caractéristiques des horizons de surface à une très courte échelle de temps
(de l’ordre de l’année).
L’étude de la distribution spatiale des turricules dans les différentes parcelles a été
initiée par l’estimation de la distance minimale et la distance moyenne entre deux
turricules. S’en est suivi une analyse des patrons de distributions (méthode de Ripley).
Parcelles Année Superficie
(m²) Nombre de Turricules
Distance Minimum entre 2 turricules
(cm)
Distance Moyenne entre 2 turricules
(cm)
Forêts F01 2002 50 459 2,0 396,5 F01 2003 25 321 1,0 264,0 F02 2003 25 646 1,0 256,9 F03 2003 25 634 1,0 252,6 F04 2002 30 650 2,8 376,7
Jachère J01 2002 50 676 1,0 368,0
Pâturages P01 2002 50 102 2,2 395,1 P01 2003 50 277 1,0 396,5 P02 2002 50 127 7,2 409,9 P03 2003 50 172 2,2 412,0 P04 2003 50 435 1,0 408,2 P05 2003 25 76 2,0 253,9
Rizière R01 2002 50 117 6,0 370,8
Tableau 22 : Distance minimum et distance moyenne entre deux turricules dans les forêts (F01, F02, F03 et
F04), la jachère (J01), les pâturages (P0, P02, P03, P04 et P05) et la rizière (R01).
La distance minimum entre deux turricules dans les forêts est de 1,6 ± 0,8 cm et
dans les pâturages 2,6 ± 2,3 cm. Elle ne diffère pas significativement entre les milieux
(test t, p= 0,429).
Cependant dans la rizière R01 et le pâturage P02, les distances minimums sont
très supérieures à celles obtenues dans les autres parcelles.
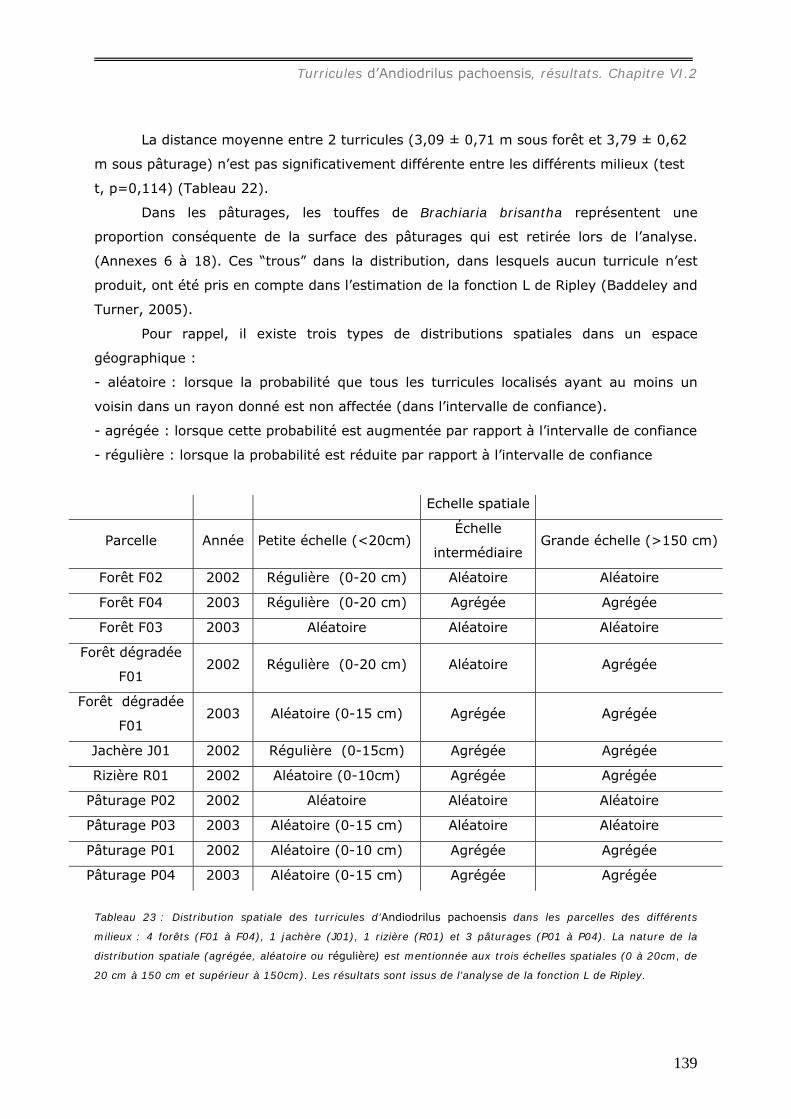
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
139
La distance moyenne entre 2 turricules (3,09 ± 0,71 m sous forêt et 3,79 ± 0,62
m sous pâturage) n’est pas significativement différente entre les différents milieux (test
t, p=0,114) (Tableau 22).
Dans les pâturages, les touffes de Brachiaria brisantha représentent une
proportion conséquente de la surface des pâturages qui est retirée lors de l’analyse.
(Annexes 6 à 18). Ces “trous” dans la distribution, dans lesquels aucun turricule n’est
produit, ont été pris en compte dans l’estimation de la fonction L de Ripley (Baddeley and
Turner, 2005).
Pour rappel, il existe trois types de distributions spatiales dans un espace
géographique :
- aléatoire : lorsque la probabilité que tous les turricules localisés ayant au moins un
voisin dans un rayon donné est non affectée (dans l’intervalle de confiance).
- agrégée : lorsque cette probabilité est augmentée par rapport à l’intervalle de confiance
- régulière : lorsque la probabilité est réduite par rapport à l’intervalle de confiance
Echelle spatiale
Parcelle Année Petite échelle (<20cm) Échelle
intermédiaire Grande échelle (>150 cm)
Forêt F02 2002 Régulière (0-20 cm) Aléatoire Aléatoire
Forêt F04 2003 Régulière (0-20 cm) Agrégée Agrégée
Forêt F03 2003 Aléatoire Aléatoire Aléatoire
Forêt dégradée
F01 2002 Régulière (0-20 cm) Aléatoire Agrégée
Forêt dégradée
F01 2003 Aléatoire (0-15 cm) Agrégée Agrégée
Jachère J01 2002 Régulière (0-15cm) Agrégée Agrégée
Rizière R01 2002 Aléatoire (0-10cm) Agrégée Agrégée
Pâturage P02 2002 Aléatoire Aléatoire Aléatoire
Pâturage P03 2003 Aléatoire (0-15 cm) Aléatoire Aléatoire
Pâturage P01 2002 Aléatoire (0-10 cm) Agrégée Agrégée
Pâturage P04 2003 Aléatoire (0-15 cm) Agrégée Agrégée
Tableau 23 : Distribution spatiale des turricules d’Andiodrilus pachoensis dans les parcelles des différents
milieux : 4 forêts (F01 à F04), 1 jachère (J01), 1 rizière (R01) et 3 pâturages (P01 à P04). La nature de la
distribution spatiale (agrégée, aléatoire ou régulière) est mentionnée aux trois échelles spatiales (0 à 20cm, de
20 cm à 150 cm et supérieur à 150cm). Les résultats sont issus de l’analyse de la fonction L de Ripley.

Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
140
Les patrons spatiaux des turricules sont apparus très variables en fonction des
parcelles et de l’année d’étude (2002 et 2003).
•Forêts
La distribution spatiale des turricules s’est avérée différente dans les cinq
parcelles aux différentes échelles. Les parcelles F02 et F04 ont mis en évidence une
distribution spatiale des turricules inhibée aux petites échelles (0-20cm). Aux échelles
intermédiaires et plus larges, la distribution est aléatoire dans la forêt F02 et agrégée
dans la forêt F04. Dans la forêt dégradée F01, la distribution spatiale de turricules s’est
avérée les deux années d’étude très variable aux échelles petites et intermédiaires alors
qu’aux larges échelles la distribution est agrégée. En 2002, la distribution est inhibée aux
échelles inférieures à 20 cm alors qu’elle est aléatoire en dessous de 15cm en 2003.
Cette différence se retrouve aux échelles intermédiaires où en 2002 la distribution est
aléatoire alors qu’elle est agrégée en 2003.
Dans la parcelle F03 la distribution des turricules est aléatoire quelque soit l’échelle
d’étude considérée (Tableau 23).
• Jachère
Une parcelle a été étudiée lors de l’échantillonnage de 2002. La distribution
spatiale des turricules observés est agrégée aux intermédiaires et grandes échelles
indiquant l’existence de taches de plus forte concentration alors que la distribution est
inhibée aux échelles inférieures à 15cm (Tableau 23).
•Rizière
Cette étude a été effectuée dans une parcelle lors de l’échantillonnage de 2002.
La distribution spatiale s’est révélée aléatoire aux petites échelles (<10 cm) et agrégée
aux autres échelles (Tableau 23).
•Pâturages
La distribution spatiale des turricules dans les pâturages s’est avérée identique,
aléatoire dans les cinq parcelles aux petites échelles. Aux échelles intermédiaires (entre
20cm et 150cm) et aux larges échelles (>150cm), les pâturages P02 et P03 mettent en

Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
141
évidence des distributions aléatoires. Dans les autres pâturages (P01 et P04), les
distributions spatiales sont agrégées aux autres échelles (Tableau 23).
Dans les forêts, les patrons spatiaux se révèlent aléatoires ou réguliers aux
petites échelles (<15 cm). Aux plus grandes échelles, alors que les forêts F02 et
F03 présentent des distributions aléatoires, la forêt dégradée F01 et la forêt F04 se
caractérisent par une distribution agrégée des turricules.
Tous les pâturages analysés ont mis en évidence des patrons aléatoires aux
petites échelles (<15 cm). Aux plus grandes échelles, deux pâturages ont présentés
des patrons aléatoires et les deux autres des patrons agrégés.
Il n’existe ainsi pas de patron général spécifique aux pâturages et/ou aux
forêts. Cette variabilité peut s’expliquer par des sources locales d’hétérogénéité
dans les deux types de milieux tels que le degré de tassement du sol ou encore
dans les pâturages l’intensité du piétinement bovin.
VI.2.1.d Stabilité dans le temps de la distribution spatiale de la production des
turricules.
Cette étude a pour objectif de vérifier l’hypothèse selon laquelle les distributions des
turricules produits aux temps J1 et J2 sont identiques à celle observée au temps J0.
A l’échelle de temps de trois mois, l’emplacement géographique des touffes de
Brachiaria brisantha ne varie pas. Ainsi l’étude de la distribution spatiale de la production
des turricules a été étudiée dans les pâturages sans prendre en compte les distributions
spatiales des touffes.
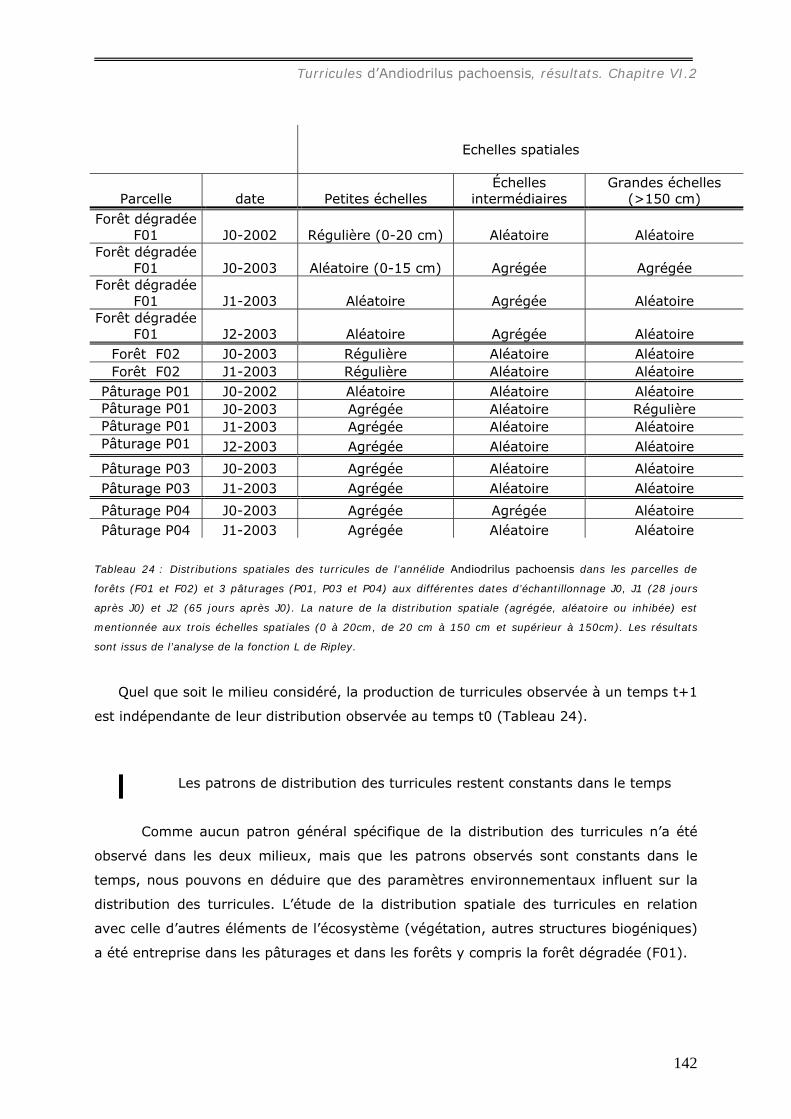
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
142
Tableau 24 : Distributions spatiales des turricules de l’annélide Andiodrilus pachoensis dans les parcelles de
forêts (F01 et F02) et 3 pâturages (P01, P03 et P04) aux différentes dates d’échantillonnage J0, J1 (28 jours
après J0) et J2 (65 jours après J0). La nature de la distribution spatiale (agrégée, aléatoire ou inhibée) est
mentionnée aux trois échelles spatiales (0 à 20cm, de 20 cm à 150 cm et supérieur à 150cm). Les résultats
sont issus de l’analyse de la fonction L de Ripley.
Quel que soit le milieu considéré, la production de turricules observée à un temps t+1
est indépendante de leur distribution observée au temps t0 (Tableau 24).
Les patrons de distribution des turricules restent constants dans le temps
Comme aucun patron général spécifique de la distribution des turricules n’a été
observé dans les deux milieux, mais que les patrons observés sont constants dans le
temps, nous pouvons en déduire que des paramètres environnementaux influent sur la
distribution des turricules. L’étude de la distribution spatiale des turricules en relation
avec celle d’autres éléments de l’écosystème (végétation, autres structures biogéniques)
a été entreprise dans les pâturages et dans les forêts y compris la forêt dégradée (F01).
Echelles spatiales
Parcelle date Petites échelles Échelles
intermédiaires Grandes échelles
(>150 cm)
Forêt dégradée F01 J0-2002 Régulière (0-20 cm) Aléatoire Aléatoire
Forêt dégradée F01 J0-2003 Aléatoire (0-15 cm) Agrégée Agrégée
Forêt dégradée F01 J1-2003 Aléatoire Agrégée Aléatoire
Forêt dégradée F01 J2-2003 Aléatoire Agrégée Aléatoire
Forêt F02 J0-2003 Régulière Aléatoire Aléatoire Forêt F02 J1-2003 Régulière Aléatoire Aléatoire
Pâturage P01 J0-2002 Aléatoire Aléatoire Aléatoire Pâturage P01 J0-2003 Agrégée Aléatoire Régulière Pâturage P01 J1-2003 Agrégée Aléatoire Aléatoire Pâturage P01 J2-2003 Agrégée Aléatoire Aléatoire
Pâturage P03 J0-2003 Agrégée Aléatoire Aléatoire Pâturage P03 J1-2003 Agrégée Aléatoire Aléatoire
Pâturage P04 J0-2003 Agrégée Agrégée Aléatoire Pâturage P04 J1-2003 Agrégée Aléatoire Aléatoire

Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
143
VI.2.1.e Relation avec les autres structures biogéniques rencontrées.
Dans les forêts, comme dans les pâturages, les relations entre la distribution
spatiale des turricules d’Andiodrilus pachoensis et celle des autres structures
biogéniques rencontrées (autres turricules, placages de termite, termitières, fourmilières,
fécès de coléoptères) ont été testées.
Des fonctions intertypes L12(r) ont été calculées entre les positions des turricules
et celles des autres structures biogéniques rencontrées (essentiellement des turricules
d’autres annélides, placages de termites ou fourmilières)
Dans les forêts
Fonction Intertype Turricules / structures
biogéniques
Echelle spatiale
Parcelles Année Petite échelle
(<20cm) Échelle intermédiaire
Grande échelle (>150 cm)
Forêt F02 2002 Aléatoire Aléatoire Aléatoire Forêt F04 2003 Aléatoire Aléatoire Aléatoire Forêt F03 2003 Aléatoire Aléatoire Aléatoire Forêt F01 2002 Aléatoire Aléatoire Aléatoire
Tableau 25 : Distributions spatiales des turricules de l’annélide Andiodrilus pachoensis dans les forêts (F01 à
F04) en fonction de la position des autres structures biogéniques (fonction intertype). La nature de la
distribution spatiale (agrégée, aléatoire ou inhibée) est mentionnée aux trois échelles spatiales (0 à 20cm, de
20 cm à 150 cm et supérieur à 150cm). Les résultats sont issus de l’analyse de la fonction L de Ripley.
Dans toutes les forêts et à toutes les échelles, la distribution des turricules
d’Andiodrilus pachoensis en fonction des autres structures biogéniques est aléatoire
(Tableau 25).
Dans les pâturages
Aucun turricule d’Andiodrilus p. n’a été observé sur ou au sein des termitières de
Cornitermes sp. dans tous les pâturages étudiés; ainsi la présence de ces structures
termitiques inhibe la production des turricules sur la surface occupée par les termitières.
En revanche aucune zone d’inhibition autour de la termitière n’a pu être déterminée.

Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
144
La très faible densité en autres structures biogéniques dans les pâturages ne nous
a pas permis de calculer de fonctions intertypes.
VI.2.1.f Relation avec la distribution spatiale des arbres dans les forêts
Cette étude a pour objectif de comparer la distribution spatiale des turricules en
fonction de la position des arbres ayant une circonférence supérieure à 10 cm.
Les fonctions intertypes L12(r) ont été calculées entre la position des turricules et
celles des centres des arbres dans les quatre forêts.
Fonction Intertype Turricules / arbres
Echelle spatiale
Parcelles Année Petite échelle (<20cm) Échelle
intermédiaire Grande échelle (>150 cm)
Forêt F02 2002 Agrégée Aléatoire Agrégée Forêt F04 2003 Aléatoire Aléatoire Aléatoire Forêt F03 2003 Aléatoire Aléatoire Aléatoire Forêt F01 2002 Agrégée Aléatoire Aléatoire
Tableau 26 : Distributions spatiales des turricules de l’annélide Andiodrilus pachoensis dans les forêts (F01 à
F04) en fonction de la position des arbres (fonction intertype). La nature de la distribution spatiale (agrégée,
aléatoire ou inhibée) est mentionnée aux trois échelles spatiales (0 à 20cm, de 20 cm à 150 cm et supérieur à
150cm). Les résultats sont issus de l’analyse de la fonction L de Ripley.
La distribution spatiale des turricules d’Andiodrilus p. en relation avec le
positionnement des arbres est aléatoire à toutes les échelles dans les forêts F03 et F04.
Dans la forêt F02 une agrégation est mise en évidence aux petites et aux larges échelles
(<20 cm). Dans la forêt dégradée F01, une agrégation entre les turricules et les arbres a
été également mise en évidence aux plus petites échelles (Tableau 26).
Dans les forêts, la distribution spatiale des turricules d’Andiodrilus p. est
indépendante de celle des autres structures biogéniques alors que dans certaines
parcelles, elle est dépendante localement de la position des arbres (agrégation aux
échelles < 20 cm).
Dans les pâturages, la distribution spatiale des turricules est inhibée par la
présence des termitières de Cornitermes. La faible densité en autres structures
biogéniques dans ce milieu ne permet pas de vérifier la dépendance de la
distribution spatiale de ces turricules et de celle des autres structures biogéniques.
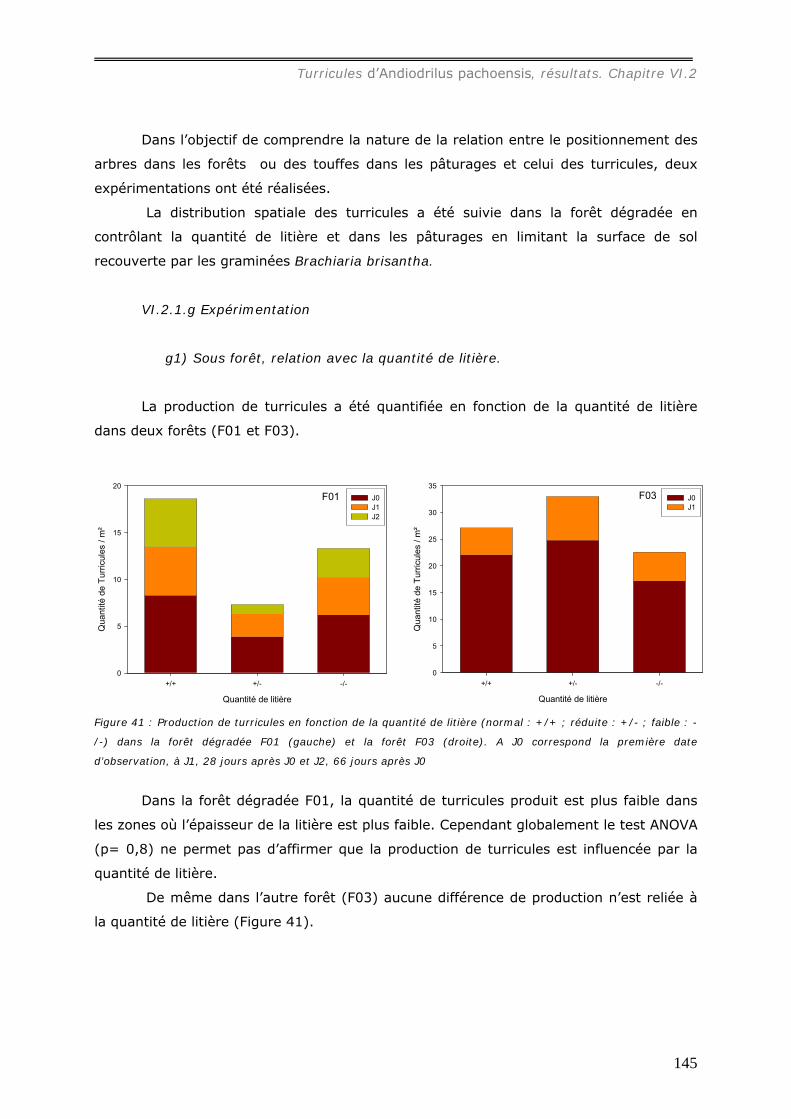
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
145
Dans l’objectif de comprendre la nature de la relation entre le positionnement des
arbres dans les forêts ou des touffes dans les pâturages et celui des turricules, deux
expérimentations ont été réalisées.
La distribution spatiale des turricules a été suivie dans la forêt dégradée en
contrôlant la quantité de litière et dans les pâturages en limitant la surface de sol
recouverte par les graminées Brachiaria brisantha.
VI.2.1.g Expérimentation
g1) Sous forêt, relation avec la quantité de litière.
La production de turricules a été quantifiée en fonction de la quantité de litière
dans deux forêts (F01 et F03).
F01
Quantité de litière
+/+ +/- -/-
Qua
ntité
de
Turr
icul
es /
m²
0
5
10
15
20J0J1J2
F03
Quantité de litière
+/+ +/- -/-
Qua
ntité
de
Turr
icul
es /
m²
0
5
10
15
20
25
30
35J0J1
F01
Quantité de litière
+/+ +/- -/-
Qua
ntité
de
Turr
icul
es /
m²
0
5
10
15
20J0J1J2
F03
Quantité de litière
+/+ +/- -/-
Qua
ntité
de
Turr
icul
es /
m²
0
5
10
15
20
25
30
35J0J1
Figure 41 : Production de turricules en fonction de la quantité de litière (normal : +/+ ; réduite : +/- ; faible : -
/-) dans la forêt dégradée F01 (gauche) et la forêt F03 (droite). A J0 correspond la première date
d’observation, à J1, 28 jours après J0 et J2, 66 jours après J0
Dans la forêt dégradée F01, la quantité de turricules produit est plus faible dans
les zones où l’épaisseur de la litière est plus faible. Cependant globalement le test ANOVA
(p= 0,8) ne permet pas d’affirmer que la production de turricules est influencée par la
quantité de litière.
De même dans l’autre forêt (F03) aucune différence de production n’est reliée à
la quantité de litière (Figure 41).
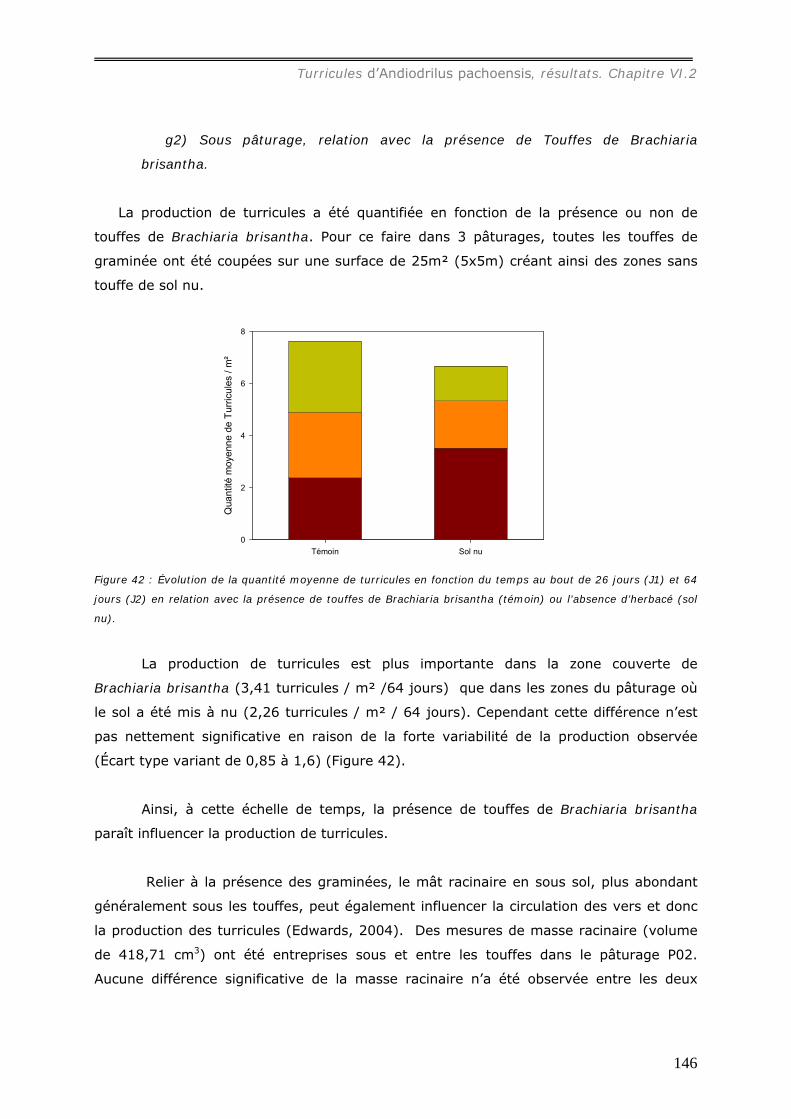
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
146
g2) Sous pâturage, relation avec la présence de Touffes de Brachiaria
brisantha.
La production de turricules a été quantifiée en fonction de la présence ou non de
touffes de Brachiaria brisantha. Pour ce faire dans 3 pâturages, toutes les touffes de
graminée ont été coupées sur une surface de 25m² (5x5m) créant ainsi des zones sans
touffe de sol nu.
Témoin Sol nu
Qua
ntité
moy
enne
de
Turri
cule
s / m
²
0
2
4
6
8
Figure 42 : Évolution de la quantité moyenne de turricules en fonction du temps au bout de 26 jours (J1) et 64
jours (J2) en relation avec la présence de touffes de Brachiaria brisantha (témoin) ou l’absence d’herbacé (sol
nu).
La production de turricules est plus importante dans la zone couverte de
Brachiaria brisantha (3,41 turricules / m² /64 jours) que dans les zones du pâturage où
le sol a été mis à nu (2,26 turricules / m² / 64 jours). Cependant cette différence n’est
pas nettement significative en raison de la forte variabilité de la production observée
(Écart type variant de 0,85 à 1,6) (Figure 42).
Ainsi, à cette échelle de temps, la présence de touffes de Brachiaria brisantha
paraît influencer la production de turricules.
Relier à la présence des graminées, le mât racinaire en sous sol, plus abondant
généralement sous les touffes, peut également influencer la circulation des vers et donc
la production des turricules (Edwards, 2004). Des mesures de masse racinaire (volume
de 418,71 cm3) ont été entreprises sous et entre les touffes dans le pâturage P02.
Aucune différence significative de la masse racinaire n’a été observée entre les deux
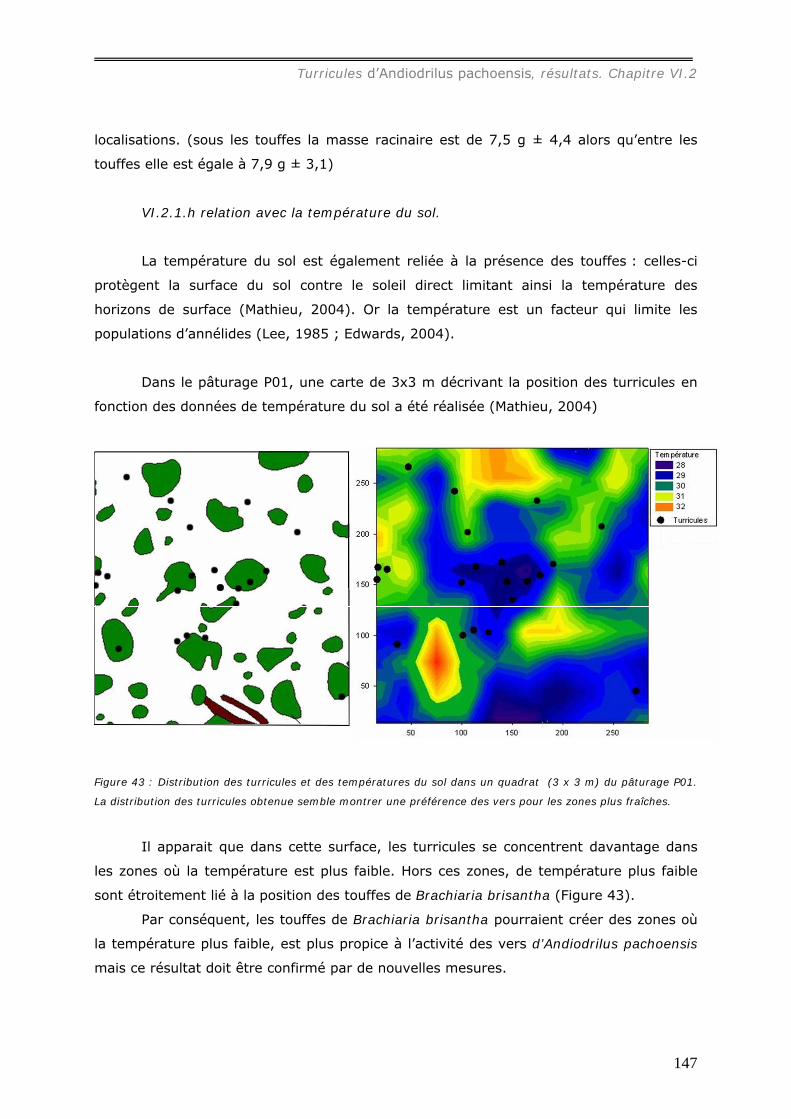
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
147
localisations. (sous les touffes la masse racinaire est de 7,5 g ± 4,4 alors qu’entre les
touffes elle est égale à 7,9 g ± 3,1)
VI.2.1.h relation avec la température du sol.
La température du sol est également reliée à la présence des touffes : celles-ci
protègent la surface du sol contre le soleil direct limitant ainsi la température des
horizons de surface (Mathieu, 2004). Or la température est un facteur qui limite les
populations d’annélides (Lee, 1985 ; Edwards, 2004).
Dans le pâturage P01, une carte de 3x3 m décrivant la position des turricules en
fonction des données de température du sol a été réalisée (Mathieu, 2004)
Figure 43 : Distribution des turricules et des températures du sol dans un quadrat (3 x 3 m) du pâturage P01.
La distribution des turricules obtenue semble montrer une préférence des vers pour les zones plus fraîches.
Il apparait que dans cette surface, les turricules se concentrent davantage dans
les zones où la température est plus faible. Hors ces zones, de température plus faible
sont étroitement lié à la position des touffes de Brachiaria brisantha (Figure 43).
Par conséquent, les touffes de Brachiaria brisantha pourraient créer des zones où
la température plus faible, est plus propice à l’activité des vers d’Andiodrilus pachoensis
mais ce résultat doit être confirmé par de nouvelles mesures.

Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
148
Dans les forêts, la quantité de litière n’est pas apparue comme un facteur
régulant la distribution spatiale des turricules.
Dans les pâturages, l’effet de la présence ou de l’absence de touffes sur la
production de turricules ne s’est pas révélé significatif. Néanmoins, les turricules
semblent cependant se concentrer davantage dans les zones ou la température du
sol est plus basse. Les touffes de Brachiaria brisantha créent des zones, où la
température plus faible est sans doute plus propice à l’activité des vers
d’Andiodrilus pachoensis. Ce dernier résultat devra cependant être vérifié par de
nouvelles mesures.
L’effet local des turricules d’Andiodrilus pachoensis sur leur environnement
immédiat a ensuite été étudié par une approche comparative des propriétés physiques et
chimiques des turricules avec celles des horizons témoins.
Prenant en compte les résultats précédents concernant les densités et les
distributions spatiales observées dans les différents milieux, une extrapolation des stocks
en matière organique au niveau parcellaire a été effectuée.
VI.2.2 Propriétés physiques des turricules d’Andiodrilus pachoensis
Dans la forêt, les turricules sont composés à 94% d’éléments minéraux et 6% de
matière organique.
Dans les pâturages, la proportion de matière organique est très légèrement plus
importante 7% pour 93% de minéral.
VI.2.2.a Granulométrie (données annexes 18, 18bis et 19)
La description des profils granulométriques des sols a été précédemment
développée (Chapitre IV). Les textures des sols témoins mettent en évidence
l’importance de la fraction argileuse et des sables grossiers. Ces textures argilo-
sableuses ne différent pas entre les deux types de milieux (forêt et pâturage).
• Sous forêt.
Les profils granulométriques ont été réalisés à partir de 3 turricules et des sols
témoins dans la forêt dégradée F01 (jusqu’à 220cm) et la forêt F02 (jusqu’à 25cm).
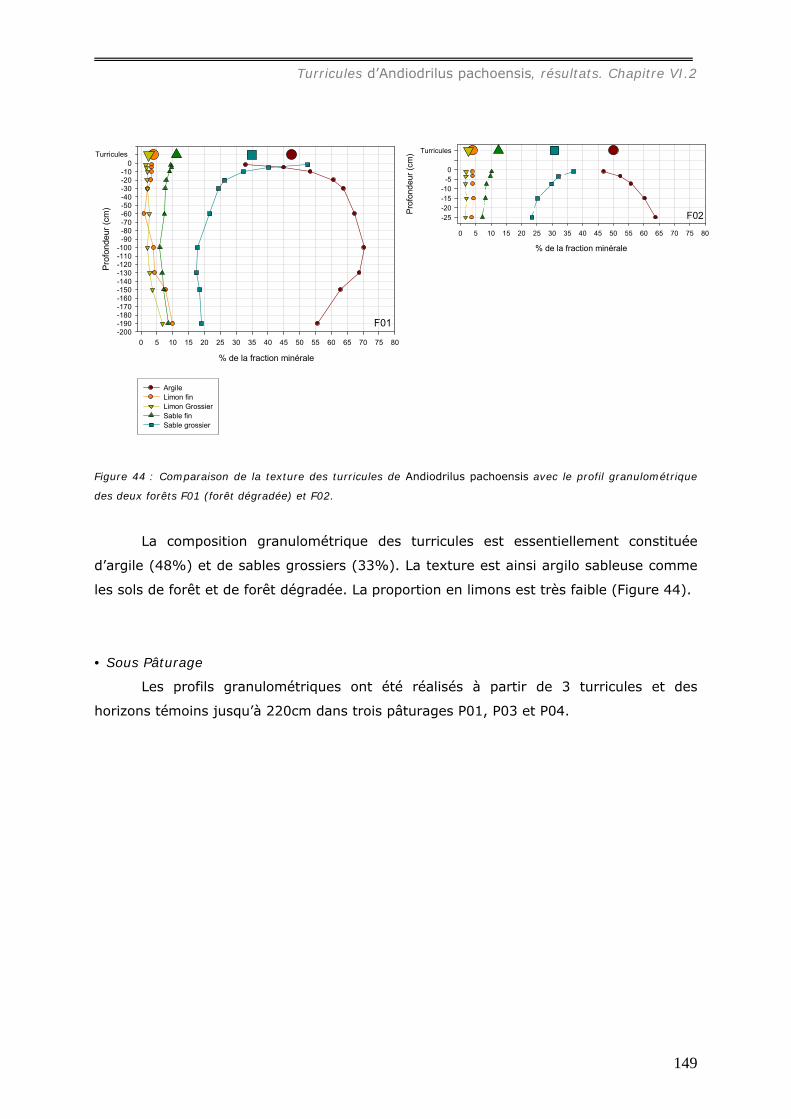
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
149
F01
% de la fraction minérale
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80
Pro
fond
eur (
cm)
-200-190-180-170-160-150-140-130-120-110-100
-90-80-70-60-50-40-30-20-10
01020
ArgileLimon finLimon GrossierSable finSable grossier
Turricules
F02
% de la fraction minérale
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80
Prof
onde
ur (c
m)
-25-20-15-10
-50
Turricules
F01
% de la fraction minérale
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80
Pro
fond
eur (
cm)
-200-190-180-170-160-150-140-130-120-110-100
-90-80-70-60-50-40-30-20-10
01020
ArgileLimon finLimon GrossierSable finSable grossier
Turricules
F02
% de la fraction minérale
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80
Prof
onde
ur (c
m)
-25-20-15-10
-50
Turricules
Figure 44 : Comparaison de la texture des turricules de Andiodrilus pachoensis avec le profil granulométrique
des deux forêts F01 (forêt dégradée) et F02.
La composition granulométrique des turricules est essentiellement constituée
d’argile (48%) et de sables grossiers (33%). La texture est ainsi argilo sableuse comme
les sols de forêt et de forêt dégradée. La proportion en limons est très faible (Figure 44).
• Sous Pâturage
Les profils granulométriques ont été réalisés à partir de 3 turricules et des
horizons témoins jusqu’à 220cm dans trois pâturages P01, P03 et P04.
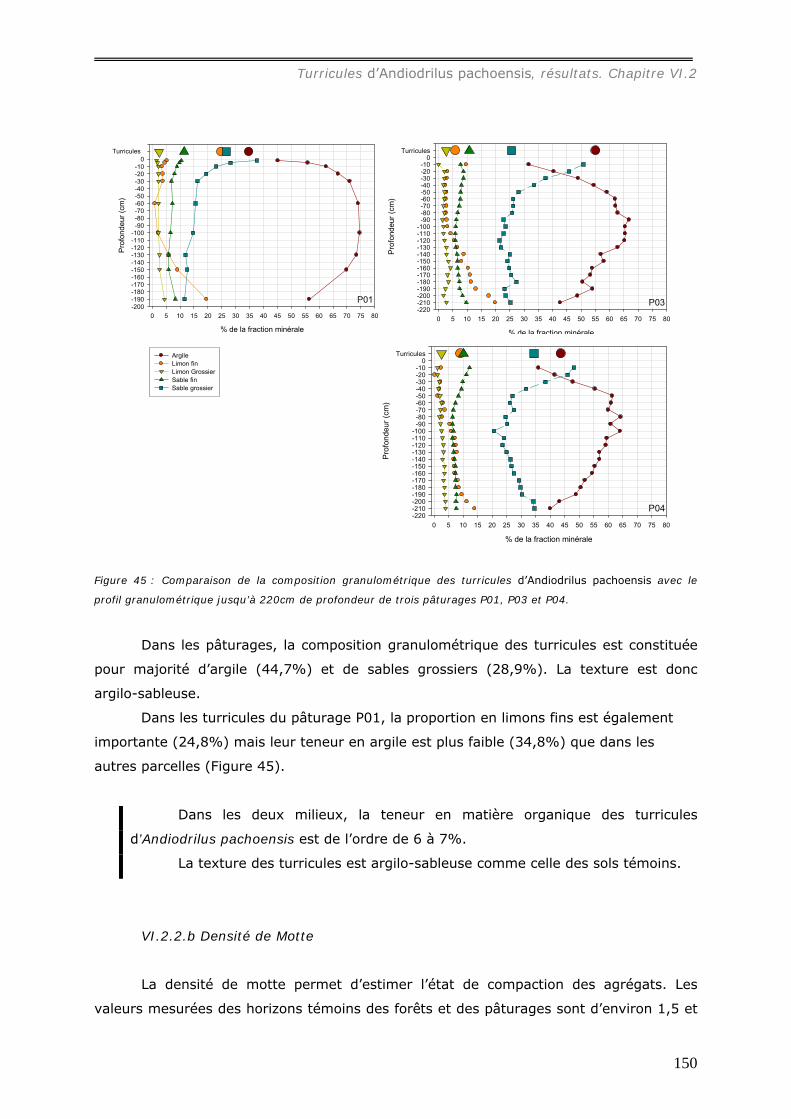
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
150
P01
% de la fraction minérale
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80
Pro
fond
eur (
cm)
-200-190-180-170-160-150-140-130-120-110-100
-90-80-70-60-50-40-30-20-10
01020
ArgileLimon finLimon GrossierSable finSable grossier
Turricules
P03
% de la fraction minérale
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80
Prof
onde
ur (c
m)
-220-210-200-190-180-170-160-150-140-130-120-110-100
-90-80-70-60-50-40-30-20-10
0Turricules
P04
% de la fraction minérale
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80
Prof
onde
ur (c
m)
-220-210-200-190-180-170-160-150-140-130-120-110-100
-90-80-70-60-50-40-30-20-10
0Turricules
P01
% de la fraction minérale
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80
Pro
fond
eur (
cm)
-200-190-180-170-160-150-140-130-120-110-100
-90-80-70-60-50-40-30-20-10
01020
ArgileLimon finLimon GrossierSable finSable grossier
Turricules
P03
% de la fraction minérale
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80
Prof
onde
ur (c
m)
-220-210-200-190-180-170-160-150-140-130-120-110-100
-90-80-70-60-50-40-30-20-10
0Turricules
P04
% de la fraction minérale
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80
Prof
onde
ur (c
m)
-220-210-200-190-180-170-160-150-140-130-120-110-100
-90-80-70-60-50-40-30-20-10
0Turricules
Figure 45 : Comparaison de la composition granulométrique des turricules d’Andiodrilus pachoensis avec le
profil granulométrique jusqu’à 220cm de profondeur de trois pâturages P01, P03 et P04.
Dans les pâturages, la composition granulométrique des turricules est constituée
pour majorité d’argile (44,7%) et de sables grossiers (28,9%). La texture est donc
argilo-sableuse.
Dans les turricules du pâturage P01, la proportion en limons fins est également
importante (24,8%) mais leur teneur en argile est plus faible (34,8%) que dans les
autres parcelles (Figure 45).
Dans les deux milieux, la teneur en matière organique des turricules
d’Andiodrilus pachoensis est de l’ordre de 6 à 7%.
La texture des turricules est argilo-sableuse comme celle des sols témoins.
VI.2.2.b Densité de Motte
La densité de motte permet d’estimer l’état de compaction des agrégats. Les
valeurs mesurées des horizons témoins des forêts et des pâturages sont d’environ 1,5 et

Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
151
différent significativement de celle de la forêt dégradée plus faible (1,32)
(ANOVA, p=0,003).
Dans les forêts, la densité de mottes des turricules (1,44 ± 0,15) ne diffère pas
significativement de celle du sol témoin de forêt (1,56 ± 0,05).
Ce résultat s’observe également dans la forêt dégradée où la densité de motte des
turricules (1,37 ± 0,14) ne diffère par de celle des horizons témoins 0-30cm
(1,32 ± 0,16).
Dans les pâturages, les turricules d’Andiodrilus pachoensis se caractérisent par
une densité de motte de l’ordre de 1,34 ± 0,13. Cette densité est significativement plus
faible que celle observée dans le sol témoin des pâturages compris entre 0 et 30 cm de
profondeur (1,57 ± 0,08) (Mann et Whitney, p< 0,001).
Les déjections d’Andiodrilus p. sont moins denses dans les pâturages que
dans les forêts (dégradées ou non) et diffèrent significativement des horizons
témoins 0-30 cm. Par conséquent les turricules de pâturage sont plus poreux. Les
turricules sont sans doute moins protégés dans les pâturages et la porosité plus
forte peut s’expliquer par une plus grande dégradation de leur structure.
VI.2.2.c Stabilité structurale
Les tests de stabilité structurale permettent de mettre en évidence la résistance
de la structure des agrégats aux agressions physiques du milieu extérieur.
Les expériences d’immersion et de réhumectation ont été réalisées dans un laps
de temps de 12H. La dispersion mécanique a été réalisée selon les temps préconisés par
la méthode de Le Bissonnais (Le Bissonnais and Souder, 1995).
Pour rappel, Les tests de stabilité structurale mettent en évidence la grande
stabilité des horizons témoins 0-30 cm. La stabilité des horizons (0-30 cm) des forêts ne
se différencie pas significativement de celle des forêts dégradées qui cependant
paraissent plus difficiles à déstructurer (chapitre IV.2). Les horizons témoins sous forêt
(toutes forêts confondues) sont cependant plus stables que ceux des pâturages. La
stabilité structurale des turricules de forêt sera comparée à celle des forêts (dégradées
ou non).
La proportion d’agglomérats dont la taille est supérieure à 2mm est restée
significativement plus importante dans les forêts. Les horizons de surface de forêt se sont
avérés plus sensibles à la réhumectation alors que dans les pâturages, l’immersion et la
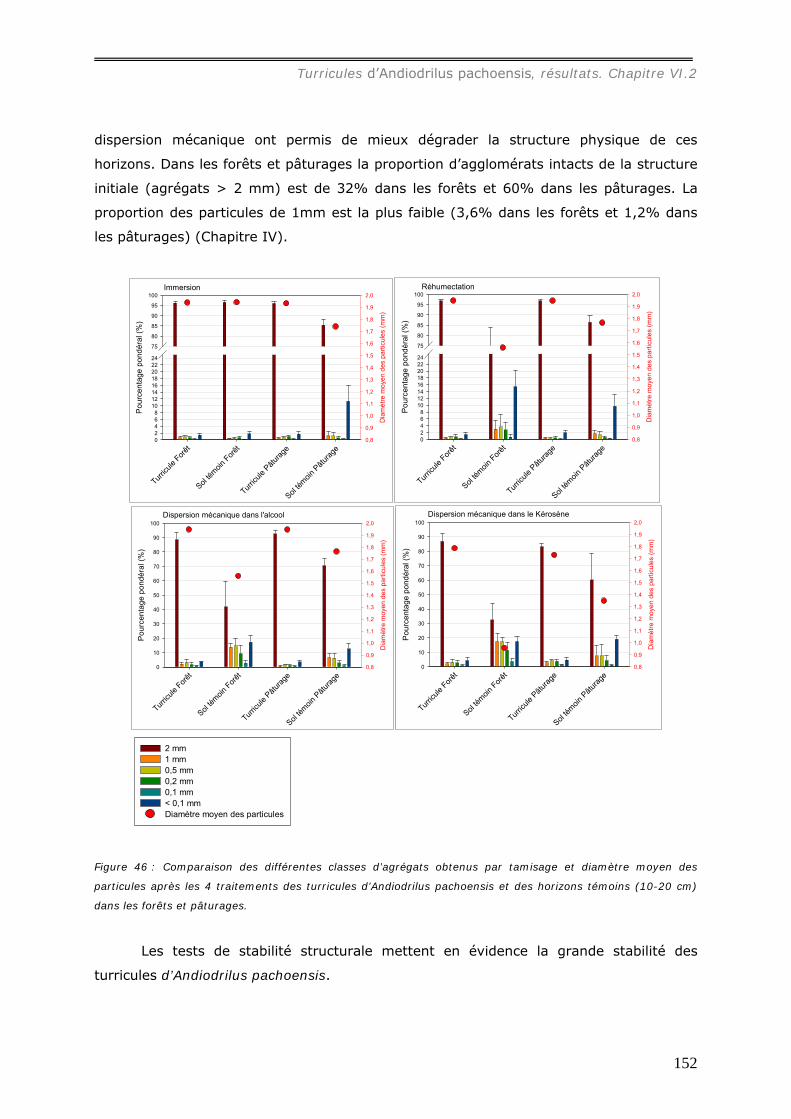
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
152
dispersion mécanique ont permis de mieux dégrader la structure physique de ces
horizons. Dans les forêts et pâturages la proportion d’agglomérats intacts de la structure
initiale (agrégats > 2 mm) est de 32% dans les forêts et 60% dans les pâturages. La
proportion des particules de 1mm est la plus faible (3,6% dans les forêts et 1,2% dans
les pâturages) (Chapitre IV).
Immersion
Turricu
le Forê
t
Sol tém
oin Forê
t
Turricu
le Pâtu
rage
Sol tém
oin Pâtu
rage
Pou
rcen
tage
pon
déra
l (%
)
02468
1012141618202224
75
80
85
90
95
100
Dia
mèt
re m
oyen
des
par
ticul
es (m
m)
0,8
0,9
1,0
1,1
1,2
1,3
1,4
1,5
1,6
1,7
1,8
1,9
2,0Réhumectation
Turricu
le Forê
t
Sol tém
oin Forê
t
Turricu
le Pâtu
rage
Sol tém
oin Pâtu
rage
Pou
rcen
tage
pon
déra
l (%
)
02468
1012141618202224
75
80
85
90
95
100
Dia
mèt
re m
oyen
des
par
ticul
es (m
m)
0,8
0,9
1,0
1,1
1,2
1,3
1,4
1,5
1,6
1,7
1,8
1,9
2,0
Dispersion mécanique dans l'alcool
Turricu
le Forê
t
Sol tém
oin Forê
t
Turricu
le Pâtu
rage
Sol tém
oin P
âturag
e
Pou
rcen
tage
pon
déra
l (%
)
0
10
20
30
40
50
60
70
80
90
100
Dia
mèt
re m
oyen
des
par
ticul
es (m
m)
0,8
0,9
1,0
1,1
1,2
1,3
1,4
1,5
1,6
1,7
1,8
1,9
2,0Dispersion mécanique dans le Kérosène
Turricu
le Forê
t
Sol tém
oin Forê
t
Turricu
le Pâtu
rage
Sol tém
oin P
âturag
e
Pou
rcen
tage
pon
déra
l (%
)
0
10
20
30
40
50
60
70
80
90
100
Dia
mèt
re m
oyen
des
par
ticul
es (m
m)
0,8
0,9
1,0
1,1
1,2
1,3
1,4
1,5
1,6
1,7
1,8
1,9
2,0
2 mm 1 mm 0,5 mm 0,2 mm 0,1 mm < 0,1 mm Diamètre moyen des particules
Immersion
Turricu
le Forê
t
Sol tém
oin Forê
t
Turricu
le Pâtu
rage
Sol tém
oin P
âturag
e
Pou
rcen
tage
pon
déra
l (%
)
02468
1012141618202224
75
80
85
90
95
100
Dia
mèt
re m
oyen
des
par
ticul
es (m
m)
0,8
0,9
1,0
1,1
1,2
1,3
1,4
1,5
1,6
1,7
1,8
1,9
2,0Réhumectation
Turricu
le Forê
t
Sol tém
oin Forê
t
Turricu
le Pâtu
rage
Sol tém
oin P
âturag
e
Pou
rcen
tage
pon
déra
l (%
)
02468
1012141618202224
75
80
85
90
95
100
Dia
mèt
re m
oyen
des
par
ticul
es (m
m)
0,8
0,9
1,0
1,1
1,2
1,3
1,4
1,5
1,6
1,7
1,8
1,9
2,0
Dispersion mécanique dans l'alcool
Turricu
le Forê
t
Sol tém
oin Forê
t
Turricu
le Pâtu
rage
Sol tém
oin P
âturag
e
Pou
rcen
tage
pon
déra
l (%
)
0
10
20
30
40
50
60
70
80
90
100
Dia
mèt
re m
oyen
des
par
ticul
es (m
m)
0,8
0,9
1,0
1,1
1,2
1,3
1,4
1,5
1,6
1,7
1,8
1,9
2,0Dispersion mécanique dans le Kérosène
Turricu
le Forê
t
Sol tém
oin Forê
t
Turricu
le Pâtu
rage
Sol tém
oin P
âturag
e
Pou
rcen
tage
pon
déra
l (%
)
0
10
20
30
40
50
60
70
80
90
100
Dia
mèt
re m
oyen
des
par
ticul
es (m
m)
0,8
0,9
1,0
1,1
1,2
1,3
1,4
1,5
1,6
1,7
1,8
1,9
2,0
Immersion
Turricu
le Forê
t
Sol tém
oin Forê
t
Turricu
le Pâtu
rage
Sol tém
oin P
âturag
e
Pou
rcen
tage
pon
déra
l (%
)
02468
1012141618202224
75
80
85
90
95
100
Dia
mèt
re m
oyen
des
par
ticul
es (m
m)
0,8
0,9
1,0
1,1
1,2
1,3
1,4
1,5
1,6
1,7
1,8
1,9
2,0Réhumectation
Turricu
le Forê
t
Sol tém
oin Forê
t
Turricu
le Pâtu
rage
Sol tém
oin P
âturag
e
Pou
rcen
tage
pon
déra
l (%
)
02468
1012141618202224
75
80
85
90
95
100
Dia
mèt
re m
oyen
des
par
ticul
es (m
m)
0,8
0,9
1,0
1,1
1,2
1,3
1,4
1,5
1,6
1,7
1,8
1,9
2,0
Dispersion mécanique dans l'alcool
Turricu
le Forê
t
Sol tém
oin Forê
t
Turricu
le Pâtu
rage
Sol tém
oin P
âturag
e
Pou
rcen
tage
pon
déra
l (%
)
0
10
20
30
40
50
60
70
80
90
100
Dia
mèt
re m
oyen
des
par
ticul
es (m
m)
0,8
0,9
1,0
1,1
1,2
1,3
1,4
1,5
1,6
1,7
1,8
1,9
2,0Dispersion mécanique dans le Kérosène
Turricu
le Forê
t
Sol tém
oin Forê
t
Turricu
le Pâtu
rage
Sol tém
oin P
âturag
e
Pou
rcen
tage
pon
déra
l (%
)
0
10
20
30
40
50
60
70
80
90
100
Dia
mèt
re m
oyen
des
par
ticul
es (m
m)
0,8
0,9
1,0
1,1
1,2
1,3
1,4
1,5
1,6
1,7
1,8
1,9
2,0
2 mm 1 mm 0,5 mm 0,2 mm 0,1 mm < 0,1 mm Diamètre moyen des particules
Figure 46 : Comparaison des différentes classes d’agrégats obtenus par tamisage et diamètre moyen des
particules après les 4 traitements des turricules d’Andiodrilus pachoensis et des horizons témoins (10-20 cm)
dans les forêts et pâturages.
Les tests de stabilité structurale mettent en évidence la grande stabilité des
turricules d’Andiodrilus pachoensis.

Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
153
Lors de tous les tests, la stabilité structurale du sol témoin s’est avérée plus faible
que celle des turricules. Aucun test n’a permis de dégrader la structure des turricules des
deux milieux (plus de 80% de la structure est conservée) alors que la structure des
horizons témoins est dégradée lors des tests de dispersion mécanique. La proportion de
fines particules (< 1mm) libérées est toujours la plus forte lors de l’immersion ou de la
réhumectation des sols témoin.
Dans les trois premiers tests (immersion, réhumectation et dispersion mécanique
après pré traitement dans l’alcool), le diamètre moyen des particules des turricules est
important (entre 1,9 et 2 mm). Le test de dispersion après prétraitement dans le
kérosène met en évidence une légère diminution du diamètre moyen des particules des
turricules (entre 1,7 et 1,8mm) ; mais qui plus importante que celle des horizons des
pâturages (1,35mm) et des forêts (0,95mm).
La stabilité des turricules sous forêts et pâturages reste ici importante avec de
80% à 90% de la structure conservée et ne diffère pas significativement l’une de l’autre.
(Figure 46).
Les tests de stabilité structurale mettent en évidence la grande stabilité des
échantillons étudiés. Néanmoins, les horizons témoins des pâturages sont plus
sensibles à l’immersion que ceux des forêts. Lors de tous les tests, la stabilité
structurale des turricules des deux milieux s’est avérée plus forte que celle des
horizons témoins. Aucun test n’a permis de dégrader la structure des turricules
(plus de 80% de la structure native est conservée même lors de la dispersion
mécanique après pré traitement dans le kérosène).
La stabilité des turricules reste ici importante avec de 80% à 90% de la
structure conservée.
VI.2.3 Bilan Chimique de séquestration des éléments.
VI.2.3.a Le Carbone
Le dosage du carbone permet d’estimer de manière approximative la quantité
globale de matière organique présent dans les turricules d’Andiodrilus pachoensis. Une
comparaison entre les turricules récoltés sous forêt, sous forêt dégradée et ceux récoltés
dans les pâturages est également entreprise.
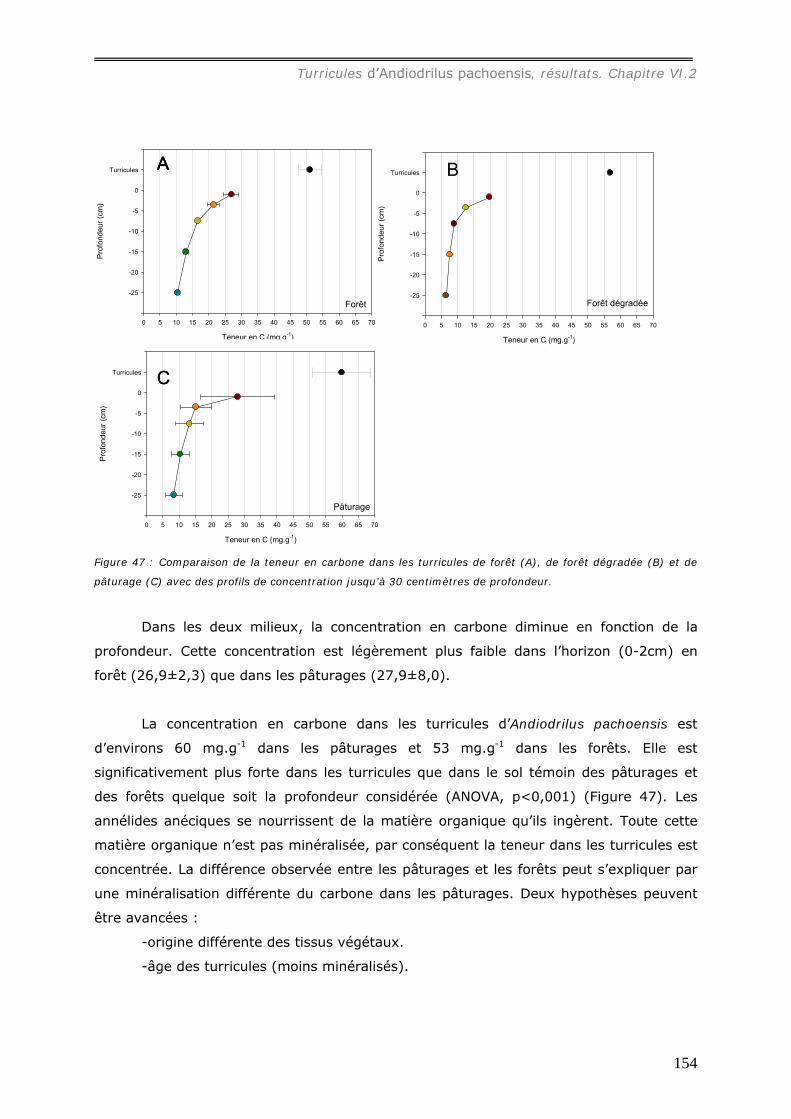
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
154
Forêt
Teneur en C (mg.g-1)
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70
Pro
fond
eur (
cm)
-25
-20
-15
-10
-5
0
Turricules
Pâturage
Teneur en C (mg.g-1)
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70
Pro
fond
eur (
cm)
-25
-20
-15
-10
-5
0
Turricules
Forêt dégradée
Teneur en C (mg.g-1)
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70
Pro
fond
eur (
cm)
-25
-20
-15
-10
-5
0
TurriculesA B
C
Forêt
Teneur en C (mg.g-1)
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70
Pro
fond
eur (
cm)
-25
-20
-15
-10
-5
0
Turricules
Pâturage
Teneur en C (mg.g-1)
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70
Pro
fond
eur (
cm)
-25
-20
-15
-10
-5
0
Turricules
Forêt dégradée
Teneur en C (mg.g-1)
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70
Pro
fond
eur (
cm)
-25
-20
-15
-10
-5
0
TurriculesA B
C
Pâturage
Teneur en C (mg.g-1)
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70
Pro
fond
eur (
cm)
-25
-20
-15
-10
-5
0
Turricules
Forêt dégradée
Teneur en C (mg.g-1)
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70
Pro
fond
eur (
cm)
-25
-20
-15
-10
-5
0
TurriculesA B
C
Forêt dégradée
Teneur en C (mg.g-1)
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70
Pro
fond
eur (
cm)
-25
-20
-15
-10
-5
0
TurriculesA B
C
Figure 47 : Comparaison de la teneur en carbone dans les turricules de forêt (A), de forêt dégradée (B) et de
pâturage (C) avec des profils de concentration jusqu’à 30 centimètres de profondeur.
Dans les deux milieux, la concentration en carbone diminue en fonction de la
profondeur. Cette concentration est légèrement plus faible dans l’horizon (0-2cm) en
forêt (26,9±2,3) que dans les pâturages (27,9±8,0).
La concentration en carbone dans les turricules d’Andiodrilus pachoensis est
d’environs 60 mg.g-1 dans les pâturages et 53 mg.g-1 dans les forêts. Elle est
significativement plus forte dans les turricules que dans le sol témoin des pâturages et
des forêts quelque soit la profondeur considérée (ANOVA, p<0,001) (Figure 47). Les
annélides anéciques se nourrissent de la matière organique qu’ils ingèrent. Toute cette
matière organique n’est pas minéralisée, par conséquent la teneur dans les turricules est
concentrée. La différence observée entre les pâturages et les forêts peut s’expliquer par
une minéralisation différente du carbone dans les pâturages. Deux hypothèses peuvent
être avancées :
-origine différente des tissus végétaux.
-âge des turricules (moins minéralisés).

Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
155
En rapportant les masses moyennes de carbone dans un turricule aux estimations
de la production de turricules dans chaque milieu, on peut évaluer la quantité de carbone
présente au sein des turricules produits. Nous avions vu que la production estimée de
turricules par hectare sous forêt était d’environ 11,9 T, sous forêt dégradée 28 T et sous
pâturages 21,8 T.
Milieux Production de turricules
(T.ha-1.an-2) Masse de Carbone (T.ha-1)
Forêts 11,9 0,63
Forêt dégradée 28,0 1,48
Pâturages 21,8 1,30
Tableau 27 : Evaluation du Carbone stocké dans les turricules d’Andiodrilus pachoensis dans les forêts et les
pâturages en fonction des estimations de masse de turricules dans chaque milieu.
Ainsi calculé, 0,63 T.ha-1 de carbone sont stockées dans les turricules produits dans les
forêts, dans les forêts dégradées cela représente 1,48T et dans les pâturages 1,30 T.ha-1
(Tableau 27).
A partir de ces concentrations en carbone, le stock de matière organique estimé
au sein des turricules produits peut être estimé à 10%- 12% de la matière sèche ; soit
deux fois plus que la valeur de carbone mesurée.
Nous pouvons ainsi estimer que dans les turricules d’Andiodrilus pachoensis
produits sous forêt, 1,26 T.ha-1.an-2 de matière organique sont stockées alors que dans la
forêt dégradée, le stock de matière organique représente presque 3 T.ha-1.an-2 et dans
les pâturages 2,6 T.ha-1.an-2.
La concentration en carbone dans les turricules d’Andiodrilus pachoensis
est d’environs 60 mg.g-1 dans les pâturages et 53 mg.g-1 dans les forêts.
Extrapolée à l’hectare et à l’année, nous pouvons estimer qu’Andiodrilus p.
restitue à l’écosystème 1,26T de matière organique sous forêt, 3T dans les forêts
dégradées et 2,6 dans les pâturages. L’intérêt de ces résultats extrapolés réside
dans le fait qu’ils donnent globalement une idée du taux de matière organique
annuel transitant dans le tube digestif de ce ver. Cette matière organique ayant
subi les attaques enzymatiques lors de la digestion, sera plus rapidement humifiée
et minéralisée dans les sols.
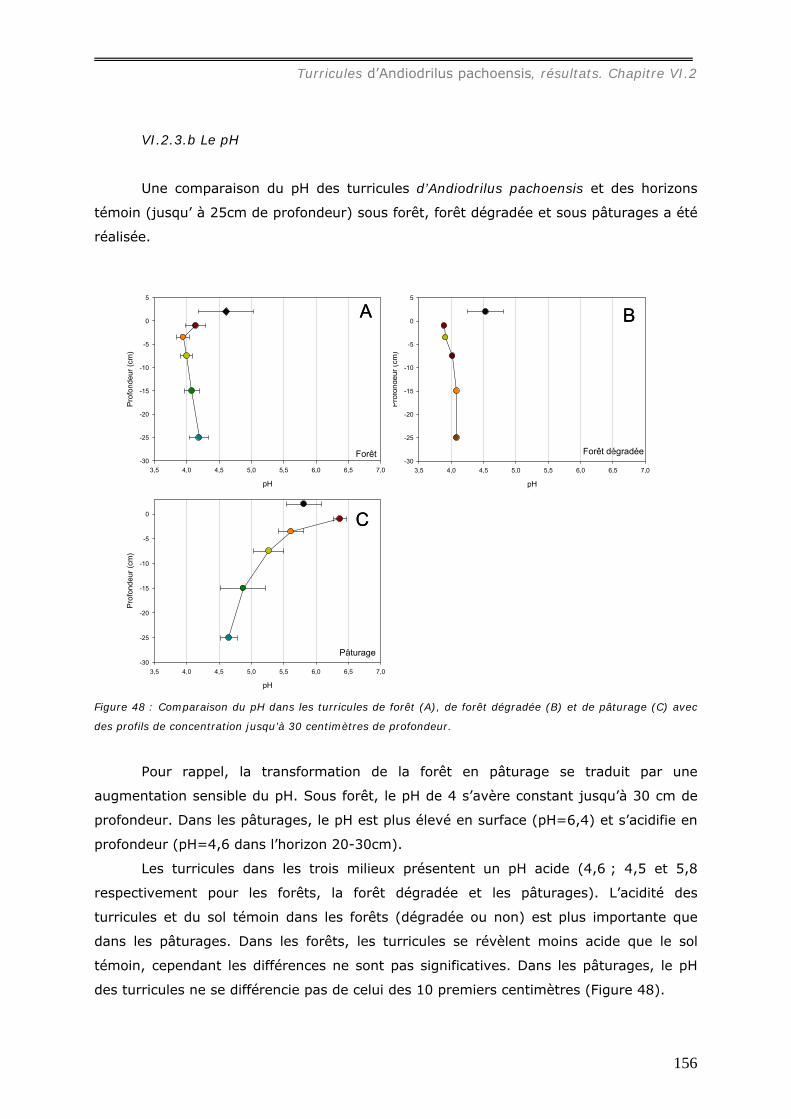
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
156
VI.2.3.b Le pH
Une comparaison du pH des turricules d’Andiodrilus pachoensis et des horizons
témoin (jusqu’ à 25cm de profondeur) sous forêt, forêt dégradée et sous pâturages a été
réalisée.
Forêt dégradée
pH
3,5 4,0 4,5 5,0 5,5 6,0 6,5 7,0
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
Pâturage
pH
3,5 4,0 4,5 5,0 5,5 6,0 6,5 7,0
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
Forêt
pH
3,5 4,0 4,5 5,0 5,5 6,0 6,5 7,0
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
A
C
B
Forêt dégradée
pH
3,5 4,0 4,5 5,0 5,5 6,0 6,5 7,0
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
Pâturage
pH
3,5 4,0 4,5 5,0 5,5 6,0 6,5 7,0
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
Forêt
pH
3,5 4,0 4,5 5,0 5,5 6,0 6,5 7,0
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
Forêt dégradée
pH
3,5 4,0 4,5 5,0 5,5 6,0 6,5 7,0
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
Pâturage
pH
3,5 4,0 4,5 5,0 5,5 6,0 6,5 7,0
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
Forêt
pH
3,5 4,0 4,5 5,0 5,5 6,0 6,5 7,0
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
A
C
BA
C
B
Figure 48 : Comparaison du pH dans les turricules de forêt (A), de forêt dégradée (B) et de pâturage (C) avec
des profils de concentration jusqu’à 30 centimètres de profondeur.
Pour rappel, la transformation de la forêt en pâturage se traduit par une
augmentation sensible du pH. Sous forêt, le pH de 4 s’avère constant jusqu’à 30 cm de
profondeur. Dans les pâturages, le pH est plus élevé en surface (pH=6,4) et s’acidifie en
profondeur (pH=4,6 dans l’horizon 20-30cm).
Les turricules dans les trois milieux présentent un pH acide (4,6 ; 4,5 et 5,8
respectivement pour les forêts, la forêt dégradée et les pâturages). L’acidité des
turricules et du sol témoin dans les forêts (dégradée ou non) est plus importante que
dans les pâturages. Dans les forêts, les turricules se révèlent moins acide que le sol
témoin, cependant les différences ne sont pas significatives. Dans les pâturages, le pH
des turricules ne se différencie pas de celui des 10 premiers centimètres (Figure 48).
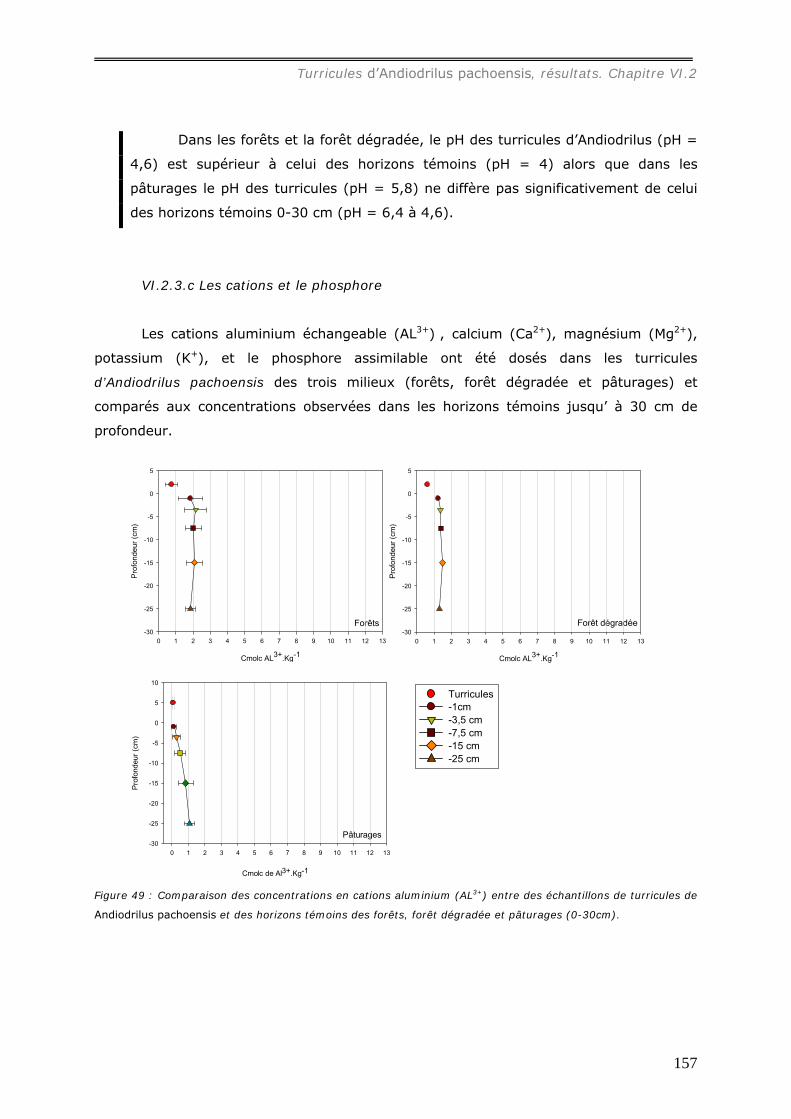
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
157
Dans les forêts et la forêt dégradée, le pH des turricules d’Andiodrilus (pH =
4,6) est supérieur à celui des horizons témoins (pH = 4) alors que dans les
pâturages le pH des turricules (pH = 5,8) ne diffère pas significativement de celui
des horizons témoins 0-30 cm (pH = 6,4 à 4,6).
VI.2.3.c Les cations et le phosphore
Les cations aluminium échangeable (AL3+) , calcium (Ca2+), magnésium (Mg2+),
potassium (K+), et le phosphore assimilable ont été dosés dans les turricules
d’Andiodrilus pachoensis des trois milieux (forêts, forêt dégradée et pâturages) et
comparés aux concentrations observées dans les horizons témoins jusqu’ à 30 cm de
profondeur.
Forêts
Cmolc AL3+.Kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Pro
fond
eur (
cm)
-30
-25
-20
-15
-10
-5
0
5
Forêt dégradée
Cmolc AL3+.Kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
-1cm -3,5 cm -7,5 cm -15 cm -25 cm
Turricules
Pâturages
Cmolc de Al3+.Kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
10
Forêts
Cmolc AL3+.Kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Pro
fond
eur (
cm)
-30
-25
-20
-15
-10
-5
0
5
Forêt dégradée
Cmolc AL3+.Kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
-1cm -3,5 cm -7,5 cm -15 cm -25 cm
Turricules
Pâturages
Cmolc de Al3+.Kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
10
Figure 49 : Comparaison des concentrations en cations aluminium (AL3+) entre des échantillons de turricules de
Andiodrilus pachoensis et des horizons témoins des forêts, forêt dégradée et pâturages (0-30cm).
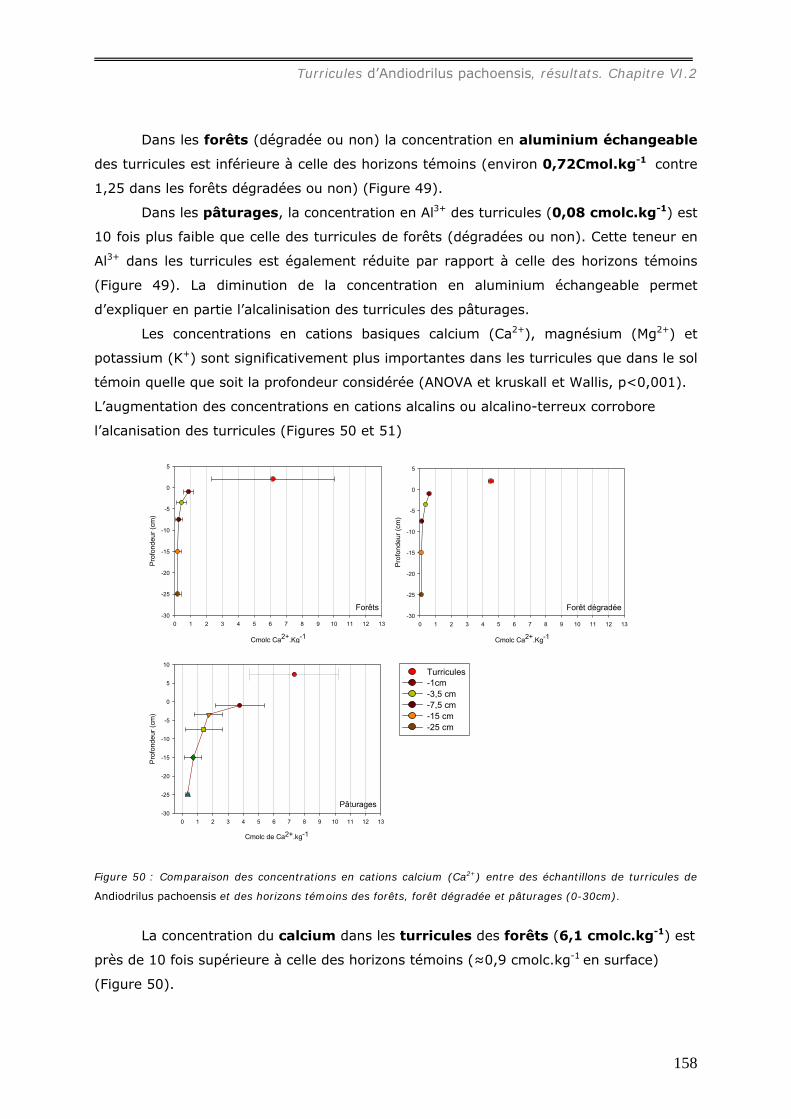
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
158
Dans les forêts (dégradée ou non) la concentration en aluminium échangeable
des turricules est inférieure à celle des horizons témoins (environ 0,72Cmol.kg-1 contre
1,25 dans les forêts dégradées ou non) (Figure 49).
Dans les pâturages, la concentration en Al3+ des turricules (0,08 cmolc.kg-1) est
10 fois plus faible que celle des turricules de forêts (dégradées ou non). Cette teneur en
Al3+ dans les turricules est également réduite par rapport à celle des horizons témoins
(Figure 49). La diminution de la concentration en aluminium échangeable permet
d’expliquer en partie l’alcalinisation des turricules des pâturages.
Les concentrations en cations basiques calcium (Ca2+), magnésium (Mg2+) et
potassium (K+) sont significativement plus importantes dans les turricules que dans le sol
témoin quelle que soit la profondeur considérée (ANOVA et kruskall et Wallis, p<0,001).
L’augmentation des concentrations en cations alcalins ou alcalino-terreux corrobore
l’alcanisation des turricules (Figures 50 et 51)
Forêts
Cmolc Ca2+.Kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
-1cm -3,5 cm -7,5 cm -15 cm -25 cm
Turricules
Forêt dégradée
Cmolc Ca2+.Kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
Pâturages
Cmolc de Ca2+.kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Pro
fond
eur (
cm)
-30
-25
-20
-15
-10
-5
0
5
10
Forêts
Cmolc Ca2+.Kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
-1cm -3,5 cm -7,5 cm -15 cm -25 cm
Turricules
Forêt dégradée
Cmolc Ca2+.Kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
-1cm -3,5 cm -7,5 cm -15 cm -25 cm
Turricules
Forêt dégradée
Cmolc Ca2+.Kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
Pâturages
Cmolc de Ca2+.kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Pro
fond
eur (
cm)
-30
-25
-20
-15
-10
-5
0
5
10
Figure 50 : Comparaison des concentrations en cations calcium (Ca2+) entre des échantillons de turricules de
Andiodrilus pachoensis et des horizons témoins des forêts, forêt dégradée et pâturages (0-30cm).
La concentration du calcium dans les turricules des forêts (6,1 cmolc.kg-1) est
près de 10 fois supérieure à celle des horizons témoins (≈0,9 cmolc.kg-1 en surface)
(Figure 50).
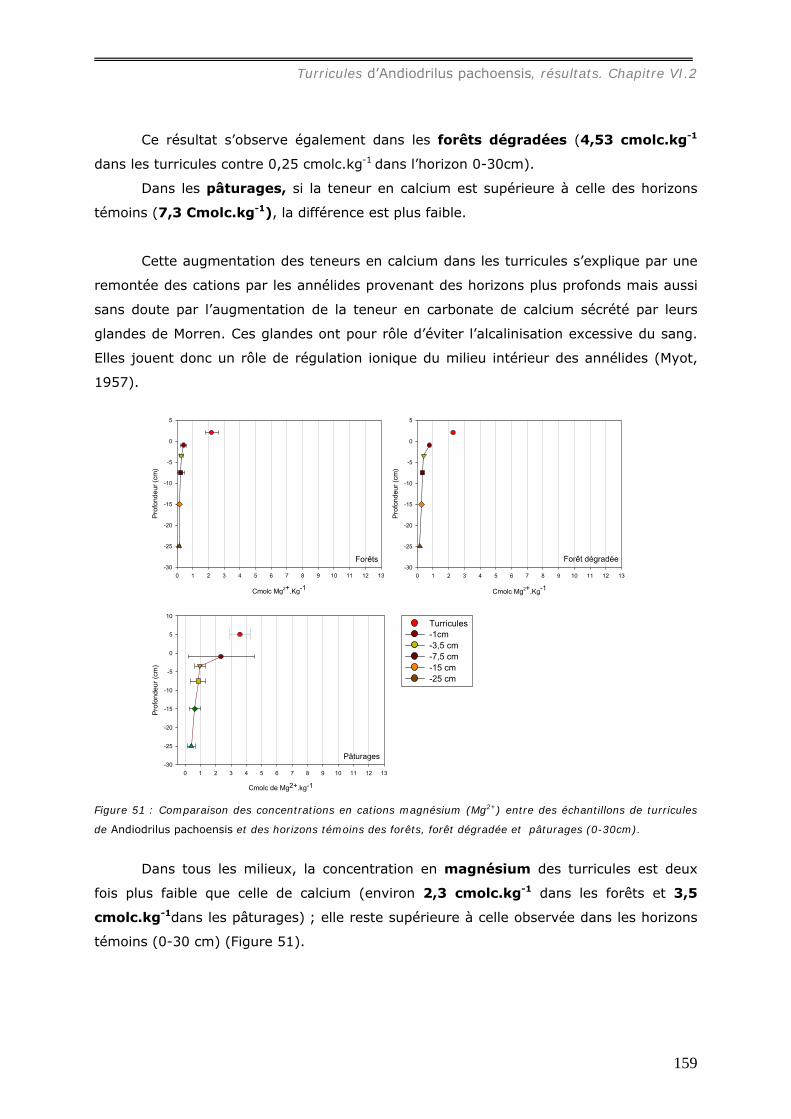
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
159
Ce résultat s’observe également dans les forêts dégradées (4,53 cmolc.kg-1
dans les turricules contre 0,25 cmolc.kg-1 dans l’horizon 0-30cm).
Dans les pâturages, si la teneur en calcium est supérieure à celle des horizons
témoins (7,3 Cmolc.kg-1), la différence est plus faible.
Cette augmentation des teneurs en calcium dans les turricules s’explique par une
remontée des cations par les annélides provenant des horizons plus profonds mais aussi
sans doute par l’augmentation de la teneur en carbonate de calcium sécrété par leurs
glandes de Morren. Ces glandes ont pour rôle d’éviter l’alcalinisation excessive du sang.
Elles jouent donc un rôle de régulation ionique du milieu intérieur des annélides (Myot,
1957).
Forêts
Cmolc Mg2+.Kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
Forêt dégradée
Cmolc Mg2+.Kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
-1cm -3,5 cm -7,5 cm -15 cm -25 cm
Turricules
Pâturages
Cmolc de Mg2+.kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
10
Forêts
Cmolc Mg2+.Kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
Forêt dégradée
Cmolc Mg2+.Kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
-1cm -3,5 cm -7,5 cm -15 cm -25 cm
Turricules
Pâturages
Cmolc de Mg2+.kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
10
Figure 51 : Comparaison des concentrations en cations magnésium (Mg2+) entre des échantillons de turricules
de Andiodrilus pachoensis et des horizons témoins des forêts, forêt dégradée et pâturages (0-30cm).
Dans tous les milieux, la concentration en magnésium des turricules est deux
fois plus faible que celle de calcium (environ 2,3 cmolc.kg-1 dans les forêts et 3,5
cmolc.kg-1dans les pâturages) ; elle reste supérieure à celle observée dans les horizons
témoins (0-30 cm) (Figure 51).
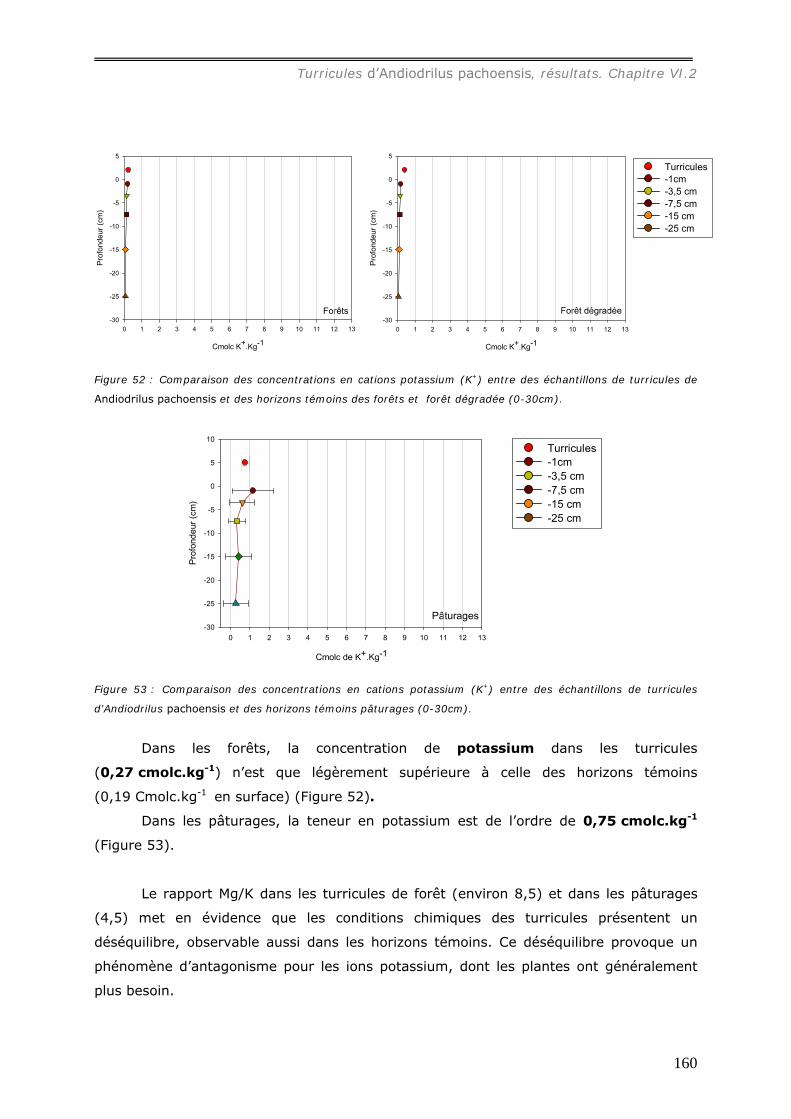
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
160
Forêts
Cmolc K+.Kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
Forêt dégradée
Cmolc K+.Kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
-1cm -3,5 cm -7,5 cm -15 cm -25 cm
Turricules
Forêts
Cmolc K+.Kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
Forêt dégradée
Cmolc K+.Kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
-1cm -3,5 cm -7,5 cm -15 cm -25 cm
Turricules
Figure 52 : Comparaison des concentrations en cations potassium (K+) entre des échantillons de turricules de
Andiodrilus pachoensis et des horizons témoins des forêts et forêt dégradée (0-30cm).
Pâturages
Cmolc de K+.Kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Pro
fond
eur (
cm)
-30
-25
-20
-15
-10
-5
0
5
10
-1cm -3,5 cm -7,5 cm -15 cm -25 cm
Turricules
Pâturages
Cmolc de K+.Kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Pro
fond
eur (
cm)
-30
-25
-20
-15
-10
-5
0
5
10
-1cm -3,5 cm -7,5 cm -15 cm -25 cm
Turricules
Figure 53 : Comparaison des concentrations en cations potassium (K+) entre des échantillons de turricules
d’Andiodrilus pachoensis et des horizons témoins pâturages (0-30cm).
Dans les forêts, la concentration de potassium dans les turricules
(0,27 cmolc.kg-1) n’est que légèrement supérieure à celle des horizons témoins
(0,19 Cmolc.kg-1 en surface) (Figure 52).
Dans les pâturages, la teneur en potassium est de l’ordre de 0,75 cmolc.kg-1
(Figure 53).
Le rapport Mg/K dans les turricules de forêt (environ 8,5) et dans les pâturages
(4,5) met en évidence que les conditions chimiques des turricules présentent un
déséquilibre, observable aussi dans les horizons témoins. Ce déséquilibre provoque un
phénomène d’antagonisme pour les ions potassium, dont les plantes ont généralement
plus besoin.
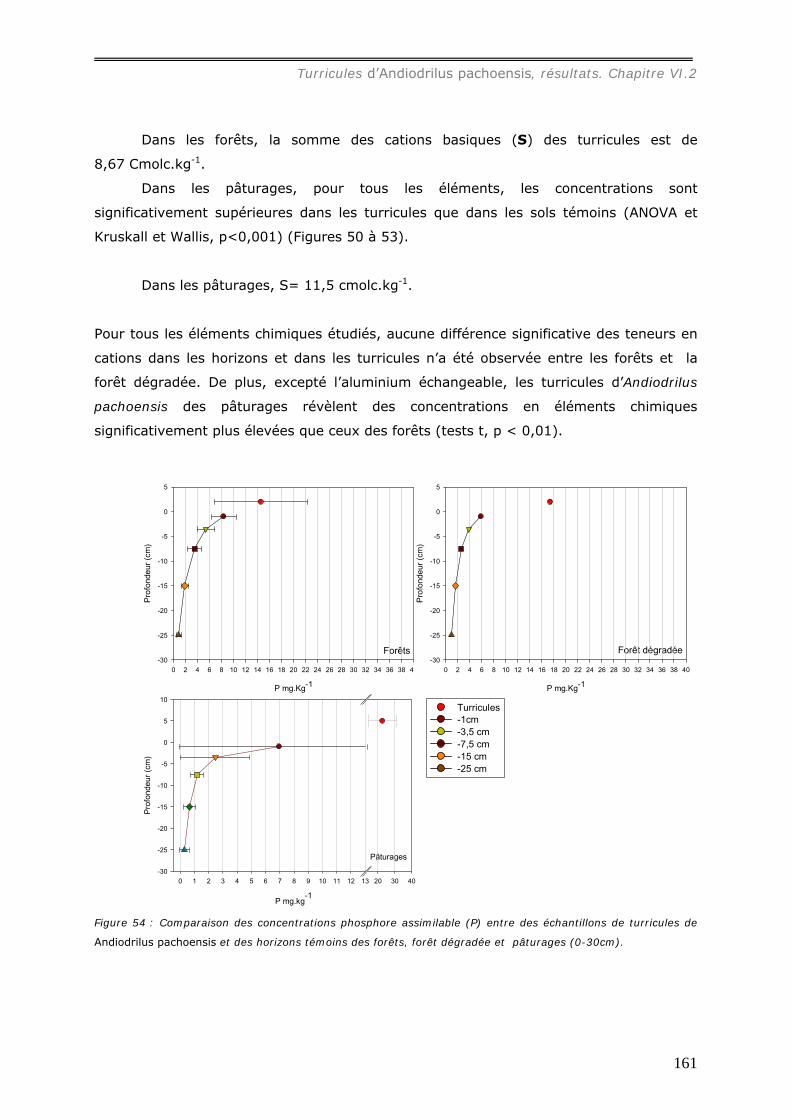
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
161
Dans les forêts, la somme des cations basiques (S) des turricules est de
8,67 Cmolc.kg-1.
Dans les pâturages, pour tous les éléments, les concentrations sont
significativement supérieures dans les turricules que dans les sols témoins (ANOVA et
Kruskall et Wallis, p<0,001) (Figures 50 à 53).
Dans les pâturages, S= 11,5 cmolc.kg-1.
Pour tous les éléments chimiques étudiés, aucune différence significative des teneurs en
cations dans les horizons et dans les turricules n’a été observée entre les forêts et la
forêt dégradée. De plus, excepté l’aluminium échangeable, les turricules d’Andiodrilus
pachoensis des pâturages révèlent des concentrations en éléments chimiques
significativement plus élevées que ceux des forêts (tests t, p < 0,01).
-1cm -3,5 cm -7,5 cm -15 cm -25 cm
Turricules
Pâturages
P mg.kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13 20 30 40
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
10
Forêts
P mg.Kg-1
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34 36 38 40
Pro
fond
eur (
cm)
-30
-25
-20
-15
-10
-5
0
5
Forêt dégradée
P mg.Kg-1
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34 36 38 40
Pro
fond
eur (
cm)
-30
-25
-20
-15
-10
-5
0
5
-1cm -3,5 cm -7,5 cm -15 cm -25 cm
Turricules
Pâturages
P mg.kg-1
0 1 2 3 4 5 6 7 8 9 10 11 12 13 20 30 40
Prof
onde
ur (c
m)
-30
-25
-20
-15
-10
-5
0
5
10
Forêts
P mg.Kg-1
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34 36 38 40
Pro
fond
eur (
cm)
-30
-25
-20
-15
-10
-5
0
5
Forêt dégradée
P mg.Kg-1
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34 36 38 40
Pro
fond
eur (
cm)
-30
-25
-20
-15
-10
-5
0
5
Figure 54 : Comparaison des concentrations phosphore assimilable (P) entre des échantillons de turricules de
Andiodrilus pachoensis et des horizons témoins des forêts, forêt dégradée et pâturages (0-30cm).
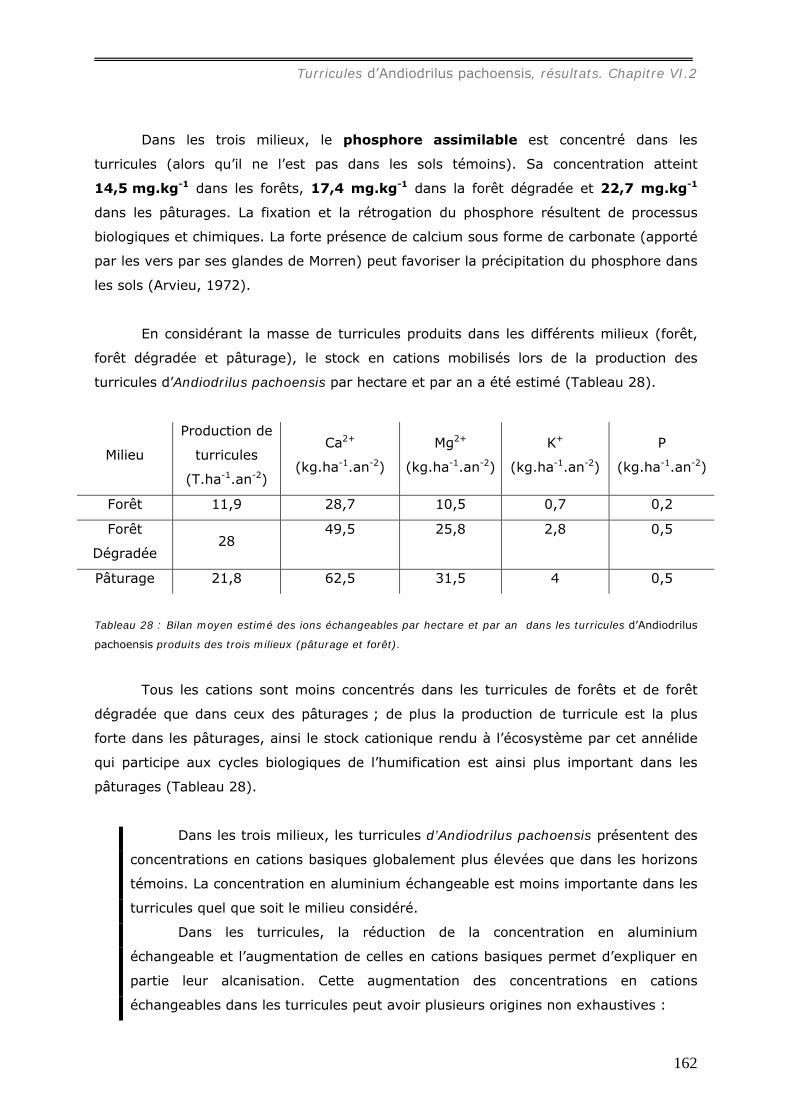
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
162
Dans les trois milieux, le phosphore assimilable est concentré dans les
turricules (alors qu’il ne l’est pas dans les sols témoins). Sa concentration atteint
14,5 mg.kg-1 dans les forêts, 17,4 mg.kg-1 dans la forêt dégradée et 22,7 mg.kg-1
dans les pâturages. La fixation et la rétrogation du phosphore résultent de processus
biologiques et chimiques. La forte présence de calcium sous forme de carbonate (apporté
par les vers par ses glandes de Morren) peut favoriser la précipitation du phosphore dans
les sols (Arvieu, 1972).
En considérant la masse de turricules produits dans les différents milieux (forêt,
forêt dégradée et pâturage), le stock en cations mobilisés lors de la production des
turricules d’Andiodrilus pachoensis par hectare et par an a été estimé (Tableau 28).
Milieu
Production de
turricules
(T.ha-1.an-2)
Ca2+
(kg.ha-1.an-2)
Mg2+
(kg.ha-1.an-2)
K+
(kg.ha-1.an-2)
P
(kg.ha-1.an-2)
Forêt 11,9 28,7 10,5 0,7 0,2
Forêt
Dégradée 28
49,5 25,8 2,8 0,5
Pâturage 21,8 62,5 31,5 4 0,5
Tableau 28 : Bilan moyen estimé des ions échangeables par hectare et par an dans les turricules d’Andiodrilus
pachoensis produits des trois milieux (pâturage et forêt).
Tous les cations sont moins concentrés dans les turricules de forêts et de forêt
dégradée que dans ceux des pâturages ; de plus la production de turricule est la plus
forte dans les pâturages, ainsi le stock cationique rendu à l’écosystème par cet annélide
qui participe aux cycles biologiques de l’humification est ainsi plus important dans les
pâturages (Tableau 28).
Dans les trois milieux, les turricules d’Andiodrilus pachoensis présentent des
concentrations en cations basiques globalement plus élevées que dans les horizons
témoins. La concentration en aluminium échangeable est moins importante dans les
turricules quel que soit le milieu considéré.
Dans les turricules, la réduction de la concentration en aluminium
échangeable et l’augmentation de celles en cations basiques permet d’expliquer en
partie leur alcanisation. Cette augmentation des concentrations en cations
échangeables dans les turricules peut avoir plusieurs origines non exhaustives :

Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
163
1) Remontée de matériau riche de la profondeur du sol
2) Concentration de la matière organique dans les déjections renfermant des sels
nutritifs.
3) Concernant le calcium, sécrétion de carbonate de calcium par les glandes de
Morren.
Les fortes teneurs en phosphore dans les turricules peuvent s’expliquer par
une précipitation plus importante du phosphore en raison des carbonates de calcium
sécrétés par les glandes de Morren des annélides.
Les stocks estimés de cations échangeables contenus dans les turricules qui
participent aux cycles biologiques augmentent du milieu forestier aux pâturages. Ce
résultat est à mettre en relation avec l’impact de la déforestation dans lequel la
forêt dégradée représente un stade intermédiaire.
VI.2.4 Dégradation des Turricules.
Comme nous l’avons déjà souligné, seule une forte dégradation permettrait
d’expliquer les différences observées entre densité et production de turricules dans
les 3 milieux. Dans les pâturages, les turricules subissent annuellement les effets
de la culture sur brûlis et sont moins protégés de la pluie par le couvert végétal
dans les pâturages que dans les forêts.
Nous avons testé deux phénomènes pouvant influer sur la dégradation des
turricules : l’action du feu et de la pluie.
L’effet du feu a été testé selon deux intensités sur les turricules d’Andiodrilus
pachoensis et une expérience de simulation de pluie nous a permis d’analyser
l’action de cette dernière sur la structure physique des turricules.
V.2.4.a Action du feu sur les turricules.
L’augmentation de la température causée par la culture sur brûlis modifie les
caractéristiques texturales des sols.

Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
164
a1) Modifications granulométriques (annexe 20)
Les profils granulométriques ont été réalisés à partir de trois turricules dans
leur état naturel, trois turricules brûlés moyennement (brulé+/-), trois turricules
brulés fortement (brûlé +/+) et du sol témoin de surface pris dans les 30 premiers
centimètres de profondeur.
Afin de simuler l’effet de la culture sur brûlis, ce dispositif a été réalisé pour
des turricules de 3 trois pâturages (P01, P03 et P04).
Par ailleurs, selon le cycle d’utilisation des parcelles (chapitre II.1), la
calcination de la forêt dégradée constitue la troisième étape de la déforestation
dans le cadre de la transformation de la forêt en rizière ou pâturage. Ainsi cette
étude a également été réalisée pour des turricules de forêts (F01, F02 et F03).
Compte tenu que les turricules de forêt dégradée bien que légèrement plus
poreux ne diffèrent pas significativement des turricules de forêt, l’analyse de
l’impact du feu a été réalisée en comparant les turricules de forêts à ceux de
pâturages.
ArgileLimon Fin Limon Grossier Sable Fin Sable Grossier
F01
%
0 10 20 30 40 50 60 70 80 90 100
Sol 0-10 cm
Turricule
Turricule Brulé +/-
Turricule Brulé +/+
F03
%
0 10 20 30 40 50 60 70 80 90 100
Sol 0-10 cm
Tur
Tur Brulé +/-
Tur Brulé +/+
F02
%
0 10 20 30 40 50 60 70 80 90 100
Sol 0-10 cm
Tur
Tur Brulé +/-
Tur Brulé +/+
ArgileLimon Fin Limon Grossier Sable Fin Sable Grossier
F01
%
0 10 20 30 40 50 60 70 80 90 100
Sol 0-10 cm
Turricule
Turricule Brulé +/-
Turricule Brulé +/+
F03
%
0 10 20 30 40 50 60 70 80 90 100
Sol 0-10 cm
Tur
Tur Brulé +/-
Tur Brulé +/+
F02
%
0 10 20 30 40 50 60 70 80 90 100
Sol 0-10 cm
Tur
Tur Brulé +/-
Tur Brulé +/+
Figure 55 : Modification de la composition granulométrique des turricules de Andiodrilus pachoensis suite à
l’exposition plus ou moins forte au feu (2 intensités : +/- et +/+) et comparaison avec celle des horizons
témoins (0 -10 cm) dans trois forêts F01, F02 et F03.
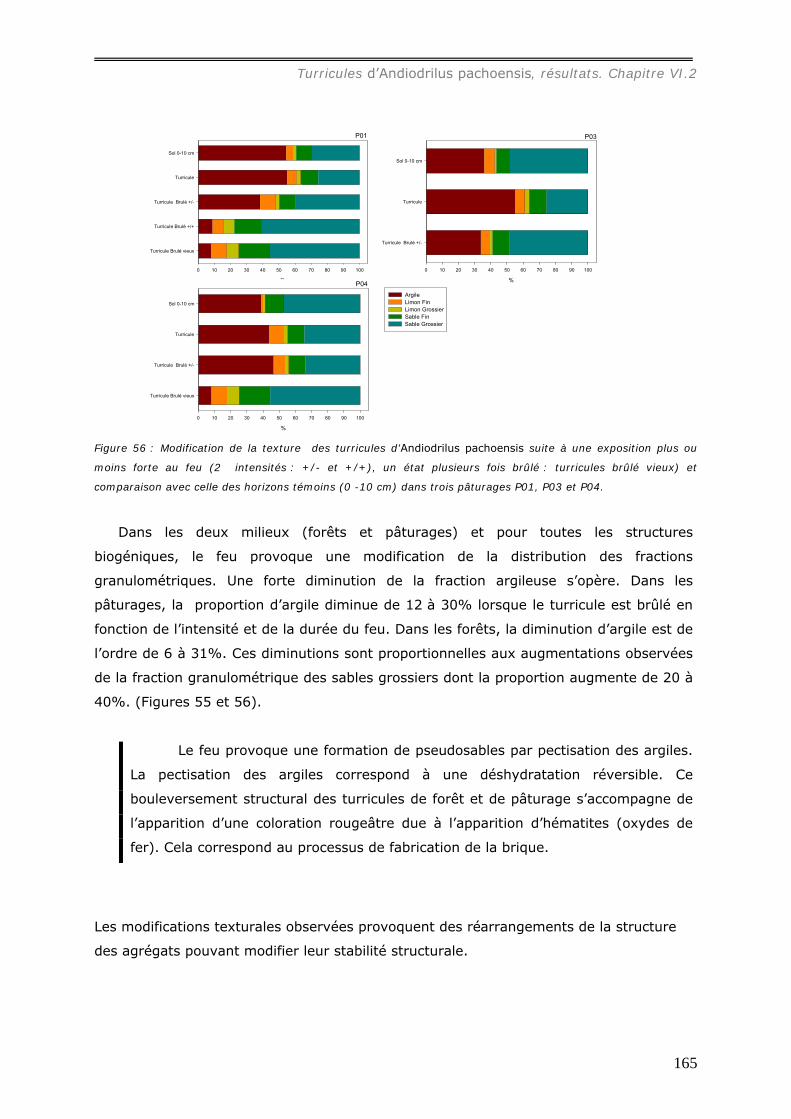
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
165
P03
%
0 10 20 30 40 50 60 70 80 90 100
Sol 0-10 cm
Turricule
Turricule Brulé +/-
P01
%
0 10 20 30 40 50 60 70 80 90 100
Sol 0-10 cm
Turricule
Turricule Brulé +/-
Turricule Brulé +/+
Turricule Brulé vieux
P04
%
0 10 20 30 40 50 60 70 80 90 100
Sol 0-10 cm
Turricule
Turricule Brulé +/-
Turricule Brulé vieux
ArgileLimon Fin Limon Grossier Sable Fin Sable Grossier
P03
%
0 10 20 30 40 50 60 70 80 90 100
Sol 0-10 cm
Turricule
Turricule Brulé +/-
P01
%
0 10 20 30 40 50 60 70 80 90 100
Sol 0-10 cm
Turricule
Turricule Brulé +/-
Turricule Brulé +/+
Turricule Brulé vieux
P04
%
0 10 20 30 40 50 60 70 80 90 100
Sol 0-10 cm
Turricule
Turricule Brulé +/-
Turricule Brulé vieux
ArgileLimon Fin Limon Grossier Sable Fin Sable Grossier
Figure 56 : Modification de la texture des turricules d’Andiodrilus pachoensis suite à une exposition plus ou
moins forte au feu (2 intensités : +/- et +/+), un état plusieurs fois brûlé : turricules brûlé vieux) et
comparaison avec celle des horizons témoins (0 -10 cm) dans trois pâturages P01, P03 et P04.
Dans les deux milieux (forêts et pâturages) et pour toutes les structures
biogéniques, le feu provoque une modification de la distribution des fractions
granulométriques. Une forte diminution de la fraction argileuse s’opère. Dans les
pâturages, la proportion d’argile diminue de 12 à 30% lorsque le turricule est brûlé en
fonction de l’intensité et de la durée du feu. Dans les forêts, la diminution d’argile est de
l’ordre de 6 à 31%. Ces diminutions sont proportionnelles aux augmentations observées
de la fraction granulométrique des sables grossiers dont la proportion augmente de 20 à
40%. (Figures 55 et 56).
Le feu provoque une formation de pseudosables par pectisation des argiles.
La pectisation des argiles correspond à une déshydratation réversible. Ce
bouleversement structural des turricules de forêt et de pâturage s’accompagne de
l’apparition d’une coloration rougeâtre due à l’apparition d’hématites (oxydes de
fer). Cela correspond au processus de fabrication de la brique.
Les modifications texturales observées provoquent des réarrangements de la structure
des agrégats pouvant modifier leur stabilité structurale.
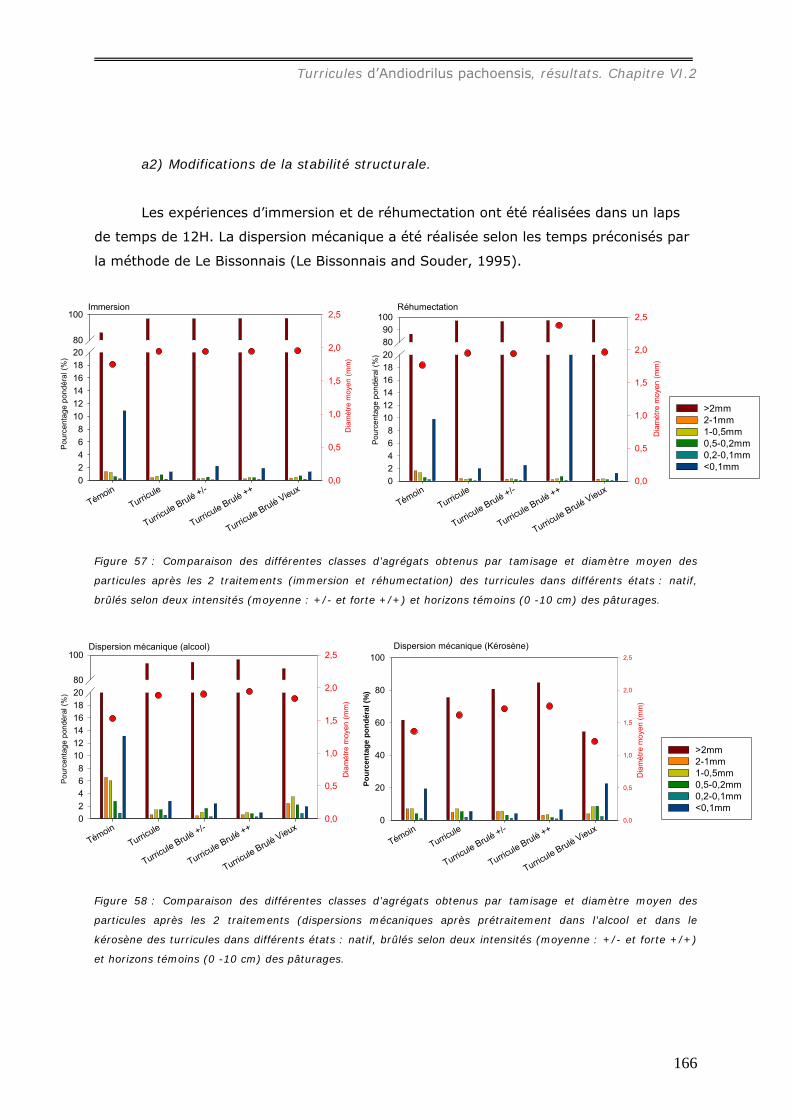
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
166
a2) Modifications de la stabilité structurale.
Les expériences d’immersion et de réhumectation ont été réalisées dans un laps
de temps de 12H. La dispersion mécanique a été réalisée selon les temps préconisés par
la méthode de Le Bissonnais (Le Bissonnais and Souder, 1995).
Immersion
Témoin
Turricule
Turricule Brulé +/-
Turricule Brulé ++
Turricule Brulé Vieux
Pou
rcen
tage
pon
déra
l (%
)
02468
10121416182080
100
Dia
mèt
re m
oyen
(mm
)
0,0
0,5
1,0
1,5
2,0
2,5Réhumectation
Témoin
Turricule
Turricule Brulé +/-
Turricule Brulé ++
Turricule Brulé Vieux
Pour
cent
age
pond
éral
(%)
02468
1012141618208090
100
Dia
mèt
re m
oyen
(mm
)
0,0
0,5
1,0
1,5
2,0
2,5
>2mm 2-1mm 1-0,5mm 0,5-0,2mm 0,2-0,1mm <0,1mm
Immersion
Témoin
Turricule
Turricule Brulé +/-
Turricule Brulé ++
Turricule Brulé Vieux
Pou
rcen
tage
pon
déra
l (%
)
02468
10121416182080
100
Dia
mèt
re m
oyen
(mm
)
0,0
0,5
1,0
1,5
2,0
2,5Réhumectation
Témoin
Turricule
Turricule Brulé +/-
Turricule Brulé ++
Turricule Brulé Vieux
Pour
cent
age
pond
éral
(%)
02468
1012141618208090
100
Dia
mèt
re m
oyen
(mm
)
0,0
0,5
1,0
1,5
2,0
2,5
>2mm 2-1mm 1-0,5mm 0,5-0,2mm 0,2-0,1mm <0,1mm
Figure 57 : Comparaison des différentes classes d’agrégats obtenus par tamisage et diamètre moyen des
particules après les 2 traitements (immersion et réhumectation) des turricules dans différents états : natif,
brûlés selon deux intensités (moyenne : +/- et forte +/+) et horizons témoins (0 -10 cm) des pâturages.
Dispersion mécanique (alcool)
Témoin
Turricule
Turricule Brulé +/-
Turricule Brulé ++
Turricule Brulé Vieux
Pou
rcen
tage
pon
déra
l (%
)
02468
10121416182080
100
Dia
mèt
re m
oyen
(mm
)
0,0
0,5
1,0
1,5
2,0
2,5Dispersion mécanique (Kérosène)
Témoin
Turricule
Turricule Brulé +/-
Turricule Brulé ++
Turricule Brulé Vieux
Pour
cent
age
pond
éral
(%)
0
20
40
60
80
100
Dia
mèt
re m
oyen
(mm
)
0,0
0,5
1,0
1,5
2,0
2,5
>2mm 2-1mm 1-0,5mm 0,5-0,2mm 0,2-0,1mm <0,1mm
Dispersion mécanique (alcool)
Témoin
Turricule
Turricule Brulé +/-
Turricule Brulé ++
Turricule Brulé Vieux
Pou
rcen
tage
pon
déra
l (%
)
02468
10121416182080
100
Dia
mèt
re m
oyen
(mm
)
0,0
0,5
1,0
1,5
2,0
2,5Dispersion mécanique (Kérosène)
Témoin
Turricule
Turricule Brulé +/-
Turricule Brulé ++
Turricule Brulé Vieux
Pour
cent
age
pond
éral
(%)
0
20
40
60
80
100
Dia
mèt
re m
oyen
(mm
)
0,0
0,5
1,0
1,5
2,0
2,5
>2mm 2-1mm 1-0,5mm 0,5-0,2mm 0,2-0,1mm <0,1mm
Figure 58 : Comparaison des différentes classes d’agrégats obtenus par tamisage et diamètre moyen des
particules après les 2 traitements (dispersions mécaniques après prétraitement dans l’alcool et dans le
kérosène des turricules dans différents états : natif, brûlés selon deux intensités (moyenne : +/- et forte +/+)
et horizons témoins (0 -10 cm) des pâturages.

Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
167
Dans les pâturages, le feu ne diminue pas la stabilité structurale des structures
biogéniques. Plus de 80% de la structure initiale est conservée lors de l’immersion ou de
la réhumectation d’un turricule.
L’immersion dans l’eau de la structure biogénique brûlée ne révèle pas de perte de
stabilité. Le diamètre moyen des agrégats d’un turricule est identique avec ou sans
brûlage (1,94 mm) (Figure 57).
Les résultats obtenus par réhumectation sont du même ordre. Seul le diamètre
moyen des turricules brûlés augmente considérablement lorsque le feu dure et
s’intensifie (Figure 57).
Les tests de dispersion mécanique après pré-traitement dans l’alcool ou dans le
kérosène mettent en évidence une augmentation de la stabilité lorsque le turricule est
brûlé. Ceci s’observe notamment par une augmentation systématique du diamètre
moyen des agrégats désolidarisés de la structure. Ce résultat confirme le résultat
précédent de fabrication de brique. Le diamètre moyen des agrégats de turricules croît
de 1,6 mm à 1,74 mm en fonction de l’intensité et de la durée du feu. En vieillissant, la
stabilité structurale des turricules se réduit de nouveau : diamètre moyen des agrégats
de valeur plus faible (1,21mm) que lorsque le turricule est dans leur état naturel
(1,61mm) (Figure 58).
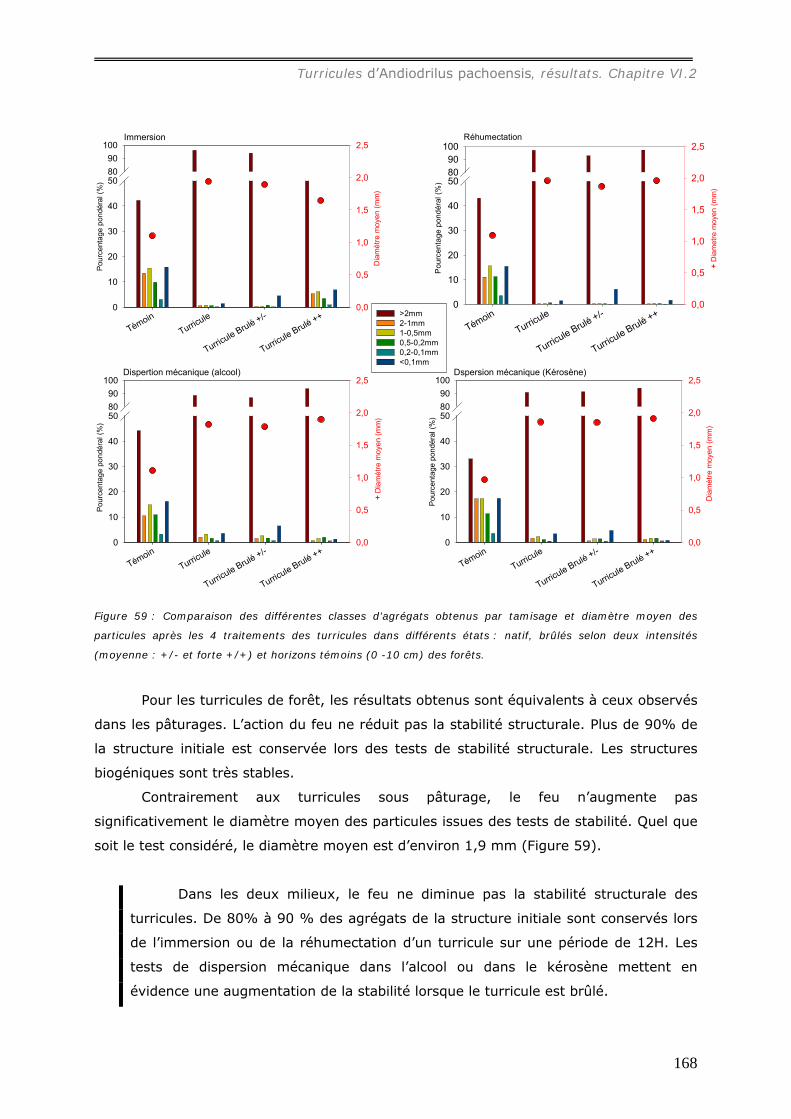
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
168
Témoin
Turricule
Turricule Brulé +/-
Turricule Brulé ++
Pou
rcen
tage
pon
déra
l (%
)
0
10
20
30
40
508090
100
Dia
mèt
re m
oyen
(mm
)
0,0
0,5
1,0
1,5
2,0
2,5Immersion Réhumectation
TémoinTurric
ule
Turricule Brulé +/-
Turricule Brulé ++
Pou
rcen
tage
pon
déra
l (%
)
0
10
20
30
40
508090
100
+ D
iam
etre
moy
en (m
m)
0,0
0,5
1,0
1,5
2,0
2,5
Dispertion mécanique (alcool)
Témoin
Turricule
Turricule Brulé +/-
Turricule Brulé ++
Pour
cent
age
pond
éral
(%)
0
10
20
30
40
508090
100+
Dia
mèt
re m
oyen
(mm
)
0,0
0,5
1,0
1,5
2,0
2,5
TémoinTurric
ule
Turricule Brulé +/-
Turricule Brulé ++
Pour
cent
age
pond
éral
(%)
0
10
20
30
40
508090
100
Dia
mèt
re m
oyen
(mm
)
0,0
0,5
1,0
1,5
2,0
2,5Dspersion mécanique (Kérosène)
>2mm 2-1mm 1-0,5mm 0,5-0,2mm 0,2-0,1mm <0,1mm
Témoin
Turricule
Turricule Brulé +/-
Turricule Brulé ++
Pour
cent
age
pond
éral
(%)
0
10
20
30
40
508090
100
Dia
mèt
re m
oyen
(mm
)
0,0
0,5
1,0
1,5
2,0
2,5Immersion Réhumectation
TémoinTurric
ule
Turricule Brulé +/-
Turricule Brulé ++
Pou
rcen
tage
pon
déra
l (%
)
0
10
20
30
40
508090
100
+ D
iam
etre
moy
en (m
m)
0,0
0,5
1,0
1,5
2,0
2,5
Dispertion mécanique (alcool)
TémoinTurric
ule
Turricule Brulé +/-
Turricule Brulé ++
Pour
cent
age
pond
éral
(%)
0
10
20
30
40
508090
100+
Dia
mèt
re m
oyen
(mm
)
0,0
0,5
1,0
1,5
2,0
2,5
TémoinTurric
ule
Turricule Brulé +/-
Turricule Brulé ++
Pour
cent
age
pond
éral
(%)
0
10
20
30
40
508090
100
Dia
mèt
re m
oyen
(mm
)
0,0
0,5
1,0
1,5
2,0
2,5Dspersion mécanique (Kérosène)
>2mm 2-1mm 1-0,5mm 0,5-0,2mm 0,2-0,1mm <0,1mm
Figure 59 : Comparaison des différentes classes d’agrégats obtenus par tamisage et diamètre moyen des
particules après les 4 traitements des turricules dans différents états : natif, brûlés selon deux intensités
(moyenne : +/- et forte +/+) et horizons témoins (0 -10 cm) des forêts.
Pour les turricules de forêt, les résultats obtenus sont équivalents à ceux observés
dans les pâturages. L’action du feu ne réduit pas la stabilité structurale. Plus de 90% de
la structure initiale est conservée lors des tests de stabilité structurale. Les structures
biogéniques sont très stables.
Contrairement aux turricules sous pâturage, le feu n’augmente pas
significativement le diamètre moyen des particules issues des tests de stabilité. Quel que
soit le test considéré, le diamètre moyen est d’environ 1,9 mm (Figure 59).
Dans les deux milieux, le feu ne diminue pas la stabilité structurale des
turricules. De 80% à 90 % des agrégats de la structure initiale sont conservés lors
de l’immersion ou de la réhumectation d’un turricule sur une période de 12H. Les
tests de dispersion mécanique dans l’alcool ou dans le kérosène mettent en
évidence une augmentation de la stabilité lorsque le turricule est brûlé.
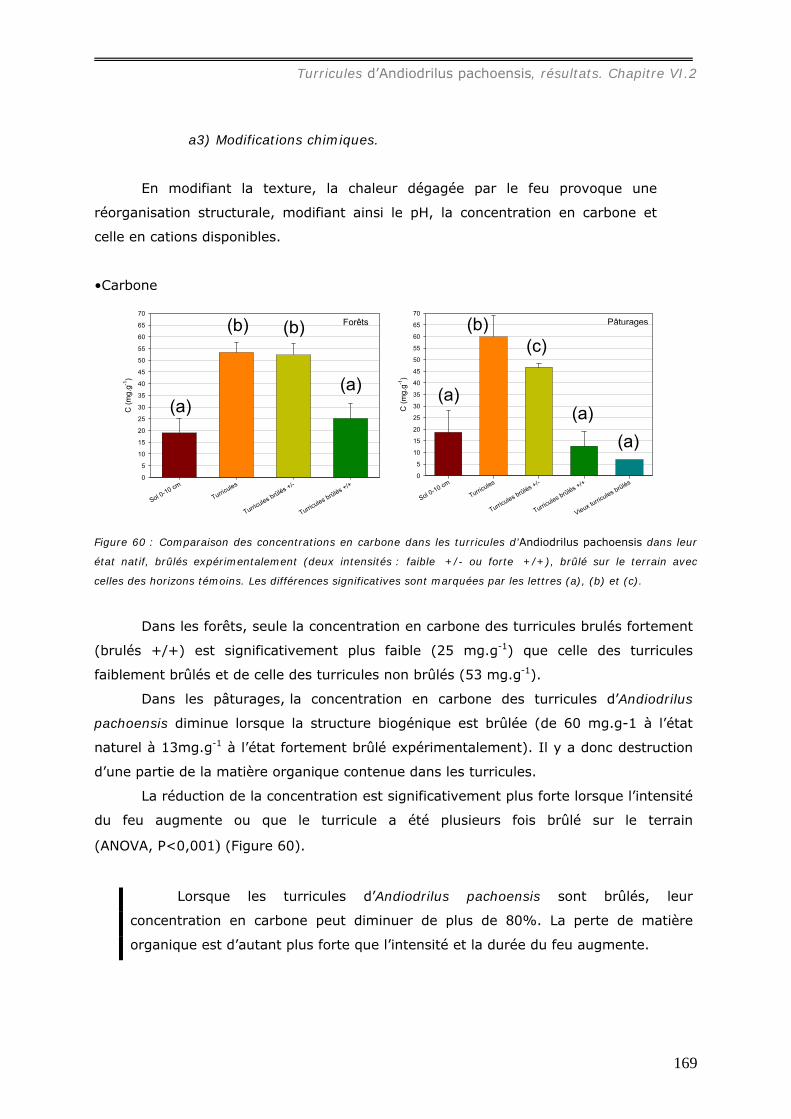
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
169
a3) Modifications chimiques.
En modifiant la texture, la chaleur dégagée par le feu provoque une
réorganisation structurale, modifiant ainsi le pH, la concentration en carbone et
celle en cations disponibles.
•Carbone
Forêts
Sol 0-10 cm
Turricules
Turricules brûlés +/-
Turricules brûlés +/+
C (m
g.g-1
)
0
5
10
15
20
25
30
35
40
45
50
55
60
65
70Pâturages
Sol 0-10 cm
Turricules
Turricules brûlés +/-
Turricules brûlés +/+
Vieux turric
ules brûlés
C (m
g.g-1
)
0
5
10
15
20
25
30
35
40
45
50
55
60
65
70
(a)(a) (a)
(a)(a)
(b) (b) (b)(c)
Forêts
Sol 0-10 cm
Turricules
Turricules brûlés +/-
Turricules brûlés +/+
C (m
g.g-1
)
0
5
10
15
20
25
30
35
40
45
50
55
60
65
70Pâturages
Sol 0-10 cm
Turricules
Turricules brûlés +/-
Turricules brûlés +/+
Vieux turric
ules brûlés
C (m
g.g-1
)
0
5
10
15
20
25
30
35
40
45
50
55
60
65
70Forêts
Sol 0-10 cm
Turricules
Turricules brûlés +/-
Turricules brûlés +/+
C (m
g.g-1
)
0
5
10
15
20
25
30
35
40
45
50
55
60
65
70Pâturages
Sol 0-10 cm
Turricules
Turricules brûlés +/-
Turricules brûlés +/+
Vieux turric
ules brûlés
C (m
g.g-1
)
0
5
10
15
20
25
30
35
40
45
50
55
60
65
70
(a)(a) (a)
(a)(a)
(b) (b) (b)(c)
Figure 60 : Comparaison des concentrations en carbone dans les turricules d’Andiodrilus pachoensis dans leur
état natif, brûlés expérimentalement (deux intensités : faible +/- ou forte +/+), brûlé sur le terrain avec
celles des horizons témoins. Les différences significatives sont marquées par les lettres (a), (b) et (c).
Dans les forêts, seule la concentration en carbone des turricules brulés fortement
(brulés +/+) est significativement plus faible (25 mg.g-1) que celle des turricules
faiblement brûlés et de celle des turricules non brûlés (53 mg.g-1).
Dans les pâturages, la concentration en carbone des turricules d’Andiodrilus
pachoensis diminue lorsque la structure biogénique est brûlée (de 60 mg.g-1 à l’état
naturel à 13mg.g-1 à l’état fortement brûlé expérimentalement). Il y a donc destruction
d’une partie de la matière organique contenue dans les turricules.
La réduction de la concentration est significativement plus forte lorsque l’intensité
du feu augmente ou que le turricule a été plusieurs fois brûlé sur le terrain
(ANOVA, P<0,001) (Figure 60).
Lorsque les turricules d’Andiodrilus pachoensis sont brûlés, leur
concentration en carbone peut diminuer de plus de 80%. La perte de matière
organique est d’autant plus forte que l’intensité et la durée du feu augmente.
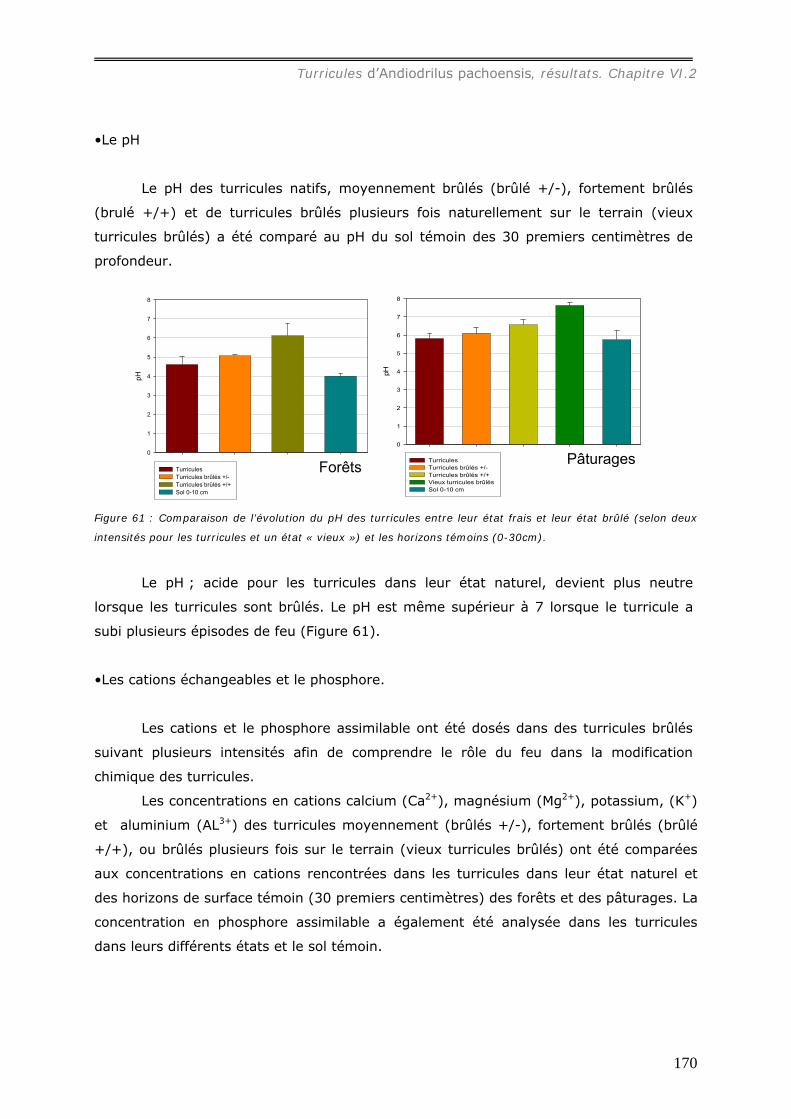
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
170
•Le pH
Le pH des turricules natifs, moyennement brûlés (brûlé +/-), fortement brûlés
(brulé +/+) et de turricules brûlés plusieurs fois naturellement sur le terrain (vieux
turricules brûlés) a été comparé au pH du sol témoin des 30 premiers centimètres de
profondeur.
pH
0
1
2
3
4
5
6
7
8
Turricules Turricules brûlés +/- Turricules brûlés +/+ Vieux turricules brûlés Sol 0-10 cm
Pâturages
pH
0
1
2
3
4
5
6
7
8
Turricules Turricules brûlés +/- Turricules brûlés +/+ Sol 0-10 cm
Forêts
pH
0
1
2
3
4
5
6
7
8
Turricules Turricules brûlés +/- Turricules brûlés +/+ Vieux turricules brûlés Sol 0-10 cm
Pâturages
pH
0
1
2
3
4
5
6
7
8
Turricules Turricules brûlés +/- Turricules brûlés +/+ Sol 0-10 cm
Forêts
Figure 61 : Comparaison de l’évolution du pH des turricules entre leur état frais et leur état brûlé (selon deux
intensités pour les turricules et un état « vieux ») et les horizons témoins (0-30cm).
Le pH ; acide pour les turricules dans leur état naturel, devient plus neutre
lorsque les turricules sont brûlés. Le pH est même supérieur à 7 lorsque le turricule a
subi plusieurs épisodes de feu (Figure 61).
•Les cations échangeables et le phosphore.
Les cations et le phosphore assimilable ont été dosés dans des turricules brûlés
suivant plusieurs intensités afin de comprendre le rôle du feu dans la modification
chimique des turricules.
Les concentrations en cations calcium (Ca2+), magnésium (Mg2+), potassium, (K+)
et aluminium (AL3+) des turricules moyennement (brûlés +/-), fortement brûlés (brûlé
+/+), ou brûlés plusieurs fois sur le terrain (vieux turricules brûlés) ont été comparées
aux concentrations en cations rencontrées dans les turricules dans leur état naturel et
des horizons de surface témoin (30 premiers centimètres) des forêts et des pâturages. La
concentration en phosphore assimilable a également été analysée dans les turricules
dans leurs différents états et le sol témoin.
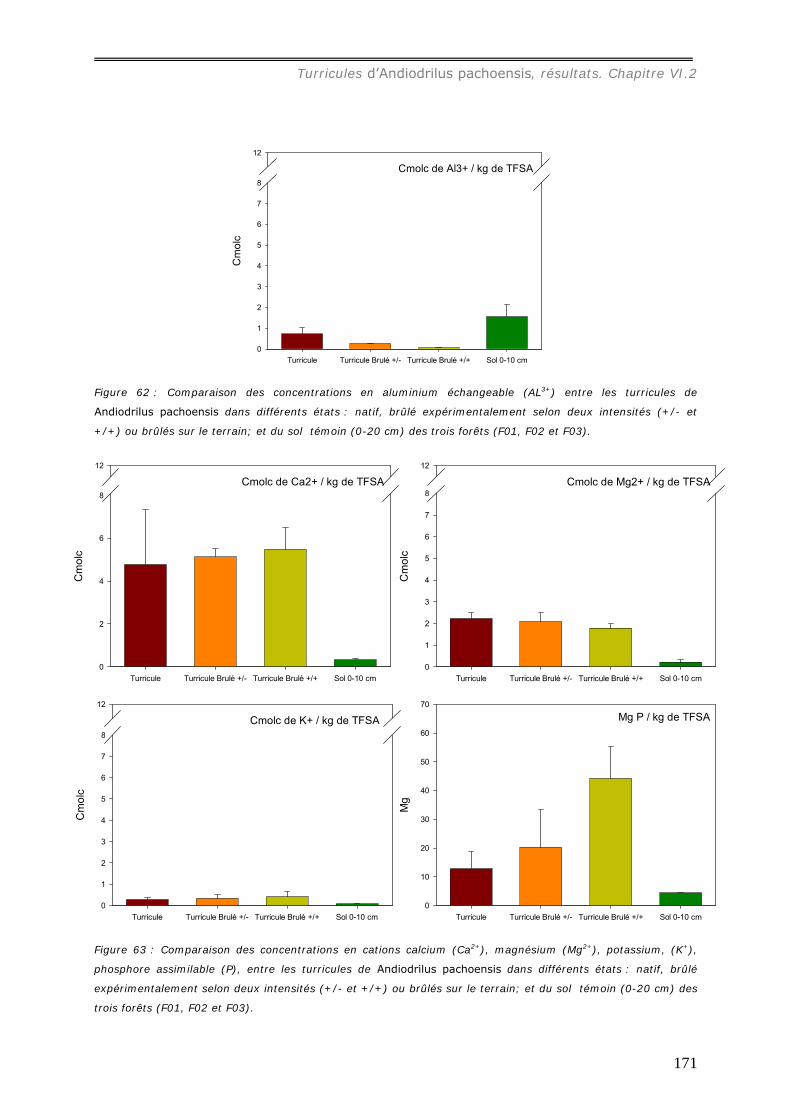
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
171
Cmolc de Al3+ / kg de TFSA
Turricule Turricule Brulé +/- Turricule Brulé +/+ Sol 0-10 cm
Cm
olc
0
1
2
3
4
5
6
7
8
12
Figure 62 : Comparaison des concentrations en aluminium échangeable (AL3+) entre les turricules de
Andiodrilus pachoensis dans différents états : natif, brûlé expérimentalement selon deux intensités (+/- et
+/+) ou brûlés sur le terrain; et du sol témoin (0-20 cm) des trois forêts (F01, F02 et F03).
Cmolc de Ca2+ / kg de TFSA
Turricule Turricule Brulé +/- Turricule Brulé +/+ Sol 0-10 cm
Cm
olc
0
2
4
6
8
12
Cmolc de K+ / kg de TFSA
Turricule Turricule Brulé +/- Turricule Brulé +/+ Sol 0-10 cm
Cm
olc
0
1
2
3
4
5
6
7
8
12
Cmolc de Mg2+ / kg de TFSA
Turricule Turricule Brulé +/- Turricule Brulé +/+ Sol 0-10 cm
Cm
olc
0
1
2
3
4
5
6
7
8
12
Mg P / kg de TFSA
Turricule Turricule Brulé +/- Turricule Brulé +/+ Sol 0-10 cm
Mg
0
10
20
30
40
50
60
70
Figure 63 : Comparaison des concentrations en cations calcium (Ca2+), magnésium (Mg2+), potassium, (K+),
phosphore assimilable (P), entre les turricules de Andiodrilus pachoensis dans différents états : natif, brûlé
expérimentalement selon deux intensités (+/- et +/+) ou brûlés sur le terrain; et du sol témoin (0-20 cm) des
trois forêts (F01, F02 et F03).
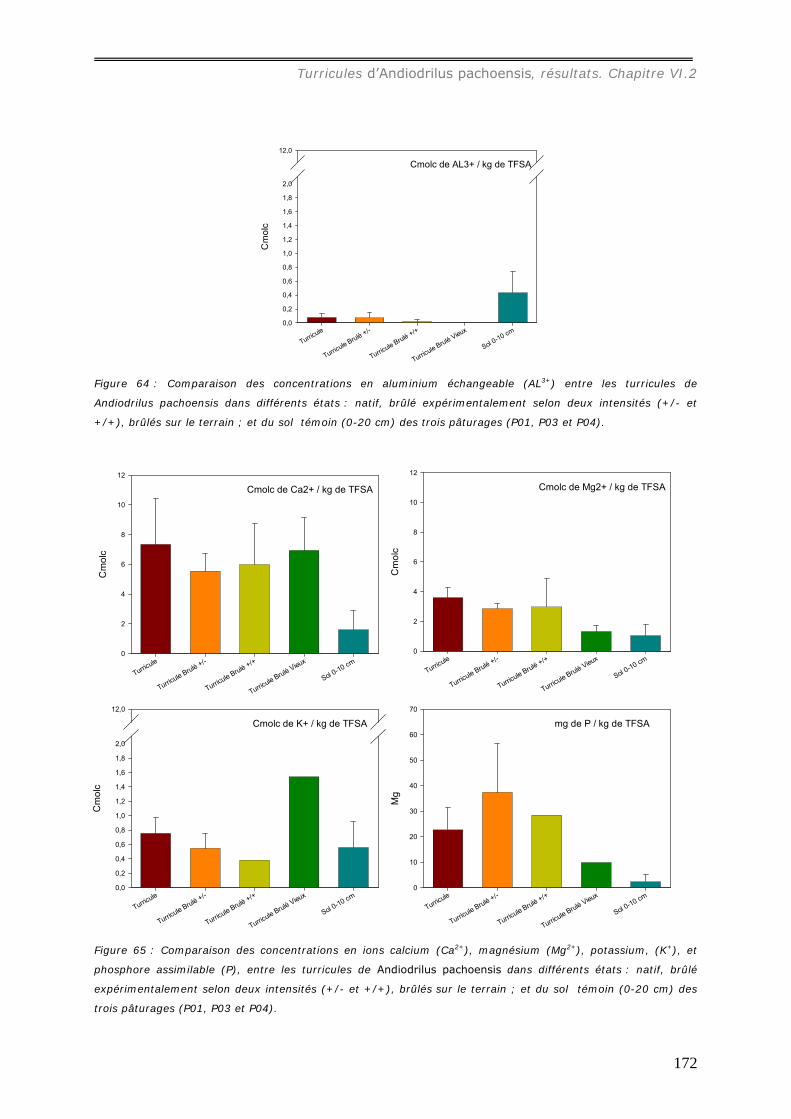
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
172
Figure 64 : Comparaison des concentrations en aluminium échangeable (AL3+) entre les turricules de
Andiodrilus pachoensis dans différents états : natif, brûlé expérimentalement selon deux intensités (+/- et
+/+), brûlés sur le terrain ; et du sol témoin (0-20 cm) des trois pâturages (P01, P03 et P04).
Cmolc de Ca2+ / kg de TFSA
Turricule
Turricule Brulé +/-
Turricule Brulé +/+
Turricule Brulé Vieux
Sol 0-10 cm
Cm
olc
0
2
4
6
8
10
12
Cmolc de Mg2+ / kg de TFSA
Turricule
Turricule Brulé +/-
Turricule Brulé +/+
Turricule Brulé Vieux
Sol 0-10 cm
Cm
olc
0
2
4
6
8
10
12
mg de P / kg de TFSA
Turricule
Turricule Brulé +/-
Turricule Brulé +/+
Turricule Brulé Vieux
Sol 0-10 cm
Mg
0
10
20
30
40
50
60
70
Cmolc de K+ / kg de TFSA
Turricule
Turricule Brulé +/-
Turricule Brulé +/+
Turricule Brulé Vieux
Sol 0-10 cm
Cm
olc
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
1,8
2,0
12,0
Figure 65 : Comparaison des concentrations en ions calcium (Ca2+), magnésium (Mg2+), potassium, (K+), et
phosphore assimilable (P), entre les turricules de Andiodrilus pachoensis dans différents états : natif, brûlé
expérimentalement selon deux intensités (+/- et +/+), brûlés sur le terrain ; et du sol témoin (0-20 cm) des
trois pâturages (P01, P03 et P04).
Cmolc de AL3+ / kg de TFSA
Turricule
Turricule Brulé +/-
Turricule Brulé +/+
Turricule Brulé Vieux
Sol 0-10 cm
Cm
olc
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
1,8
2,0
12,0
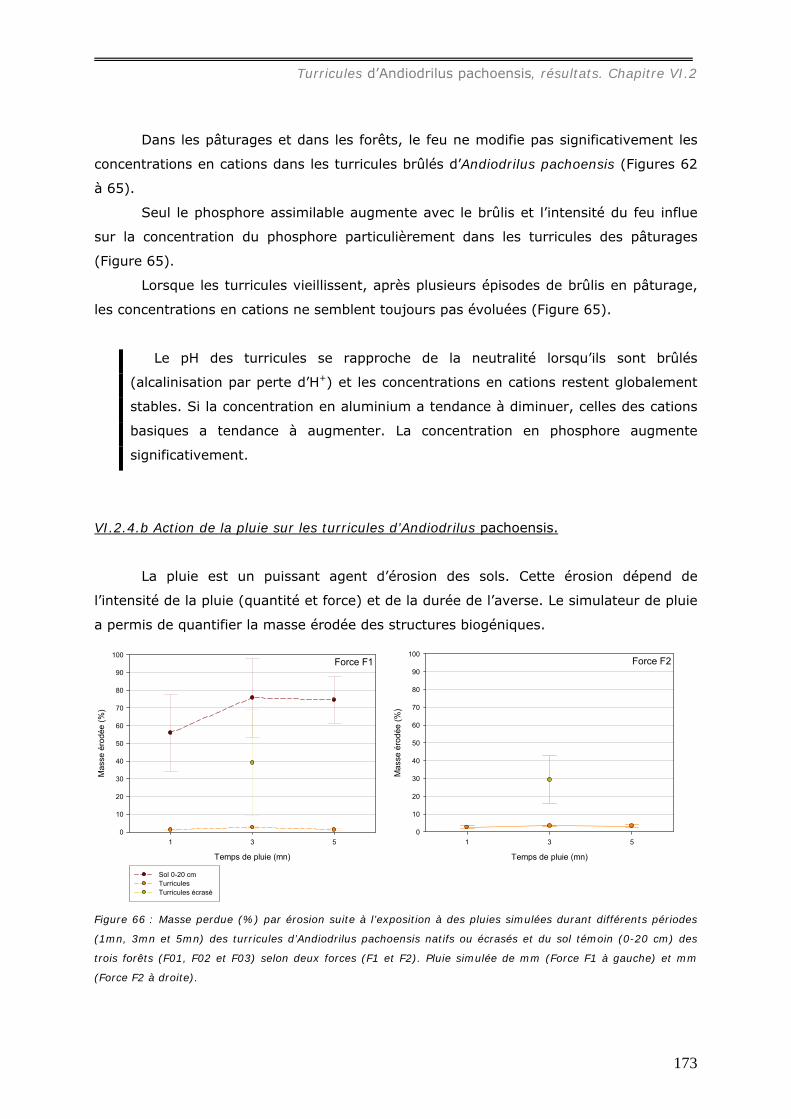
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
173
Dans les pâturages et dans les forêts, le feu ne modifie pas significativement les
concentrations en cations dans les turricules brûlés d’Andiodrilus pachoensis (Figures 62
à 65).
Seul le phosphore assimilable augmente avec le brûlis et l’intensité du feu influe
sur la concentration du phosphore particulièrement dans les turricules des pâturages
(Figure 65).
Lorsque les turricules vieillissent, après plusieurs épisodes de brûlis en pâturage,
les concentrations en cations ne semblent toujours pas évoluées (Figure 65).
Le pH des turricules se rapproche de la neutralité lorsqu’ils sont brûlés
(alcalinisation par perte d’H+) et les concentrations en cations restent globalement
stables. Si la concentration en aluminium a tendance à diminuer, celles des cations
basiques a tendance à augmenter. La concentration en phosphore augmente
significativement.
VI.2.4.b Action de la pluie sur les turricules d’Andiodrilus pachoensis.
La pluie est un puissant agent d’érosion des sols. Cette érosion dépend de
l’intensité de la pluie (quantité et force) et de la durée de l’averse. Le simulateur de pluie
a permis de quantifier la masse érodée des structures biogéniques.
Force F1
Temps de pluie (mn)
1 3 5
Mas
se é
rodé
e (%
)
0
10
20
30
40
50
60
70
80
90
100
Sol 0-20 cm Turricules Turricules écrasé
Force F2
Temps de pluie (mn)
1 3 5
Mas
se é
rodé
e (%
)
0
10
20
30
40
50
60
70
80
90
100Force F1
Temps de pluie (mn)
1 3 5
Mas
se é
rodé
e (%
)
0
10
20
30
40
50
60
70
80
90
100
Sol 0-20 cm Turricules Turricules écrasé
Force F2
Temps de pluie (mn)
1 3 5
Mas
se é
rodé
e (%
)
0
10
20
30
40
50
60
70
80
90
100
Figure 66 : Masse perdue (%) par érosion suite à l’exposition à des pluies simulées durant différents périodes
(1mn, 3mn et 5mn) des turricules d’Andiodrilus pachoensis natifs ou écrasés et du sol témoin (0-20 cm) des
trois forêts (F01, F02 et F03) selon deux forces (F1 et F2). Pluie simulée de mm (Force F1 à gauche) et mm
(Force F2 à droite).
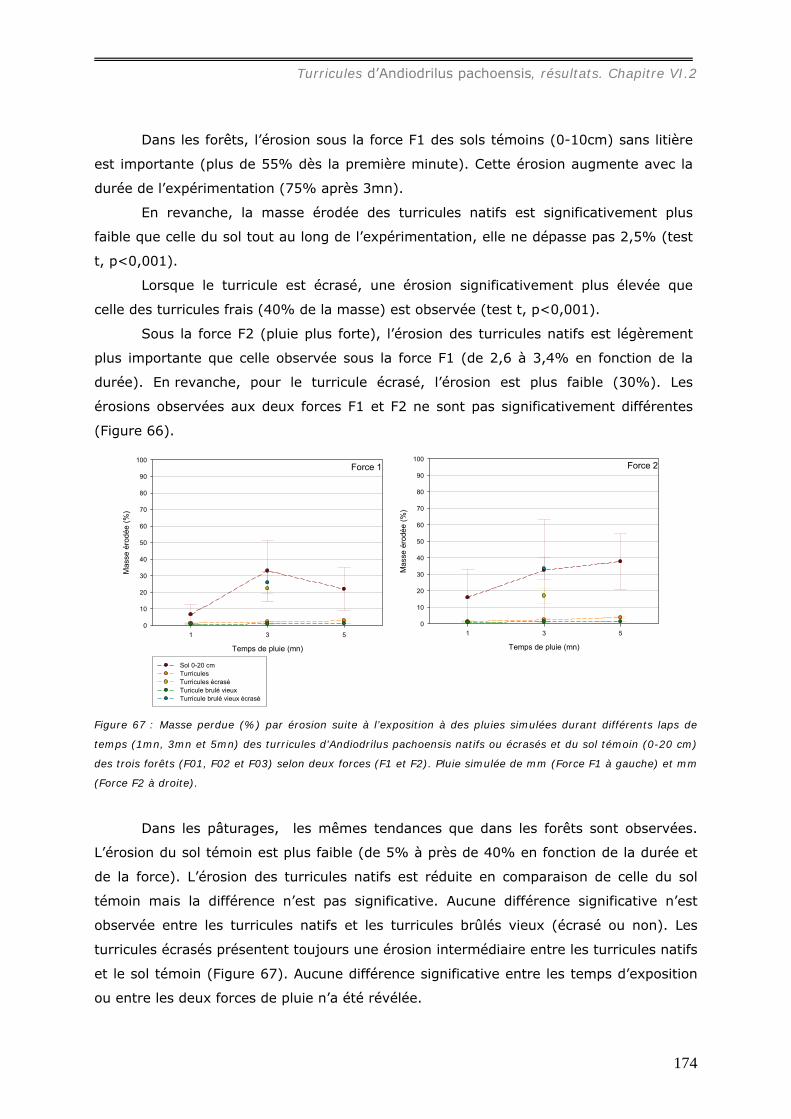
Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
174
Dans les forêts, l’érosion sous la force F1 des sols témoins (0-10cm) sans litière
est importante (plus de 55% dès la première minute). Cette érosion augmente avec la
durée de l’expérimentation (75% après 3mn).
En revanche, la masse érodée des turricules natifs est significativement plus
faible que celle du sol tout au long de l’expérimentation, elle ne dépasse pas 2,5% (test
t, p<0,001).
Lorsque le turricule est écrasé, une érosion significativement plus élevée que
celle des turricules frais (40% de la masse) est observée (test t, p<0,001).
Sous la force F2 (pluie plus forte), l’érosion des turricules natifs est légèrement
plus importante que celle observée sous la force F1 (de 2,6 à 3,4% en fonction de la
durée). En revanche, pour le turricule écrasé, l’érosion est plus faible (30%). Les
érosions observées aux deux forces F1 et F2 ne sont pas significativement différentes
(Figure 66).
Force 1
Temps de pluie (mn)
1 3 5
Mas
se é
rodé
e (%
)
0
10
20
30
40
50
60
70
80
90
100
Sol 0-20 cm Turricules Turricules écrasé Turicule brulé vieux Turricule brulé vieux écrasé
Force 2
Temps de pluie (mn)
1 3 5
Mas
se é
rodé
e (%
)
0
10
20
30
40
50
60
70
80
90
100Force 1
Temps de pluie (mn)
1 3 5
Mas
se é
rodé
e (%
)
0
10
20
30
40
50
60
70
80
90
100
Sol 0-20 cm Turricules Turricules écrasé Turicule brulé vieux Turricule brulé vieux écrasé
Force 2
Temps de pluie (mn)
1 3 5
Mas
se é
rodé
e (%
)
0
10
20
30
40
50
60
70
80
90
100
Figure 67 : Masse perdue (%) par érosion suite à l’exposition à des pluies simulées durant différents laps de
temps (1mn, 3mn et 5mn) des turricules d’Andiodrilus pachoensis natifs ou écrasés et du sol témoin (0-20 cm)
des trois forêts (F01, F02 et F03) selon deux forces (F1 et F2). Pluie simulée de mm (Force F1 à gauche) et mm
(Force F2 à droite).
Dans les pâturages, les mêmes tendances que dans les forêts sont observées.
L’érosion du sol témoin est plus faible (de 5% à près de 40% en fonction de la durée et
de la force). L’érosion des turricules natifs est réduite en comparaison de celle du sol
témoin mais la différence n’est pas significative. Aucune différence significative n’est
observée entre les turricules natifs et les turricules brûlés vieux (écrasé ou non). Les
turricules écrasés présentent toujours une érosion intermédiaire entre les turricules natifs
et le sol témoin (Figure 67). Aucune différence significative entre les temps d’exposition
ou entre les deux forces de pluie n’a été révélée.

Turricules d’Andiodrilus pachoensis, résultats. Chapitre VI.2
175
Sous simulateur de pluie, l’érosion des turricules natifs ou brûlés est presque
nulle (2,5%) alors que celle des sols témoins est très importante (55% en forêt et
40% dans les pâturages). En revanche, des turricules écrasés présentent une
érosion intermédiaire entre le turricule natif et le sol témoin.

Turricules d’Andiodrilus pachoensis, discussion. Chapitre VI
176
VI.3 DISCUSSION.
L’objectif de cette étude est de mettre en évidence le rôle particulier de l’annélide
ingénieur Andiodrilus pachoensis dans le fonctionnement du sol après déforestation au
travers de l’étude des interactions turricules-sol.
Andiodrilus pachoensis est un annélide anécique coloré de la famille des
Glossoscolecidae. Classiquement les vers de terre anéciques vivent dans des galeries
verticales profondes et se nourrissent de la matière organique près de la surface du sol.
De nos résultats est apparu que le changement de milieu, des forêts aux
pâturages, constituait un facteur fondamental influant l’activité de l’annélide Andiodrilus
pachoensis.
Impacts de la déforestation sur l’activité de l’annélide Andiodrilus pachoensis
Une analyse comparative des densités des turricules reconnaissables de cette
espèce a été menée dans les différents milieux (forêts, forêt dégradée et pâturages). Les
densités des turricules comptabilisés à un temps donné (T0) diminuent des forêts
(242,9.103) aux forêts dégradées (110,1.103) puis aux pâturages de 5-6 ans (42,2.103).
Compte tenu du cycle d’utilisation agricole classique des terrains dans le site
d’étude (forêt > forêt dégradée > pâturage), il s’avère que la déforestation provoque une
forte réduction de la densité des turricules ; cette réduction est accompagnée d’une
réduction des densités en individus (Mathieu, 2004). Ainsi Andiodrilus p. s’adapte au
nouvel écosystème pâturage ; il n’est par conséquent pas une espèce indicatrice des
fronts de déforestation puisque après six années de pâturage sa présence prouve qu’il a
réussi à s’adapter aux conditions des pâturages.
Les turricules étudiés sont constitués par une accumulation successive de
déjections de taille centimétrique. Les turricules sont creusés d’un canal central bien
conservé qui persiste au cours du temps. Ainsi la masse de turricule est apparue aussi
comme un bon élément de comparaison des différents milieux.
La masse moyenne d’un turricule s’est révélée supérieure dans les forêts
(184,6 g) à celle observée dans les pâturages (112,0 g). La différence de masse
moyenne d’un turricule peut s’expliquer d’une part, par une période plus longue de
fréquentation de l’annélide dans une même galerie sous forêt et/ou d’autre part par l’âge
des turricules dans les pâturages (plus récents).
La masse totale de turricules a été calculée et leurs productions sur 66 jours
étudiées dans les différents milieux analysés.

Turricules d’Andiodrilus pachoensis, discussion. Chapitre VI
177
La masse des turricules à l’hectare est apparue seize fois plus importante dans les
forêts (41,3 T. ha-1) que dans les pâturages (2,6 T. ha-1) avec une masse de turricules
intermédiaires dans les forêts dégradées (16,4 T. ha-1). Cette masse en surface est
relativement importante en comparaison avec d’autres études effectuées dans des sols
acides (Laverack, 1961 ; Lee, 1985 ; Henrot and Brussaard, 1997 ; Lavelle and Spain,
2001) en raison sans doute de l’espèce d’annélide étudiée. Cependant cette estimation
de la masse ne prend pas en compte les turricules de profondeur sans doute plus
abondants. Aucune relation entre le nombre de turricules et le nombre d’individus
rencontrés dans les 30 premiers centimètres n’a par ailleurs, d’après nos connaissances,
été vérifié pour les annélides en raison de leur écologie et plus particulièrement leur
mobilité verticale et horizontale et en raison de la difficulté d’observer leurs patrons de
distribution (Rossi and Nuutinen, 2004).
Par contre, la production de turricules sous forêt est apparue beaucoup plus faible
(296 turricules par jour et par hectare) que dans les forêts dégradées (740 turricules par
jour et par ha) et dans les pâturages (920 turricules par jour et par hectare). La
production d’Andiodrilus p. dans les pâturages est plus faible que celle observée dans des
pâturages colombiens pour Martiodrilus carimaguensis (1280 turricules par jour et par
hectare ) (Jiménez et al., 1998). L’estimation de la production dans la forêt dégradée est
très élevée. Cette production peut s’expliquer par le fait qu’elle n’a été mesurée que dans
une parcelle où les turricules avaient été l’année précédente retirés. Il est possible que le
fait de retirer les turricules favorise significativement leur production, le déblaiement de
la surface stimulant ainsi leur activité. La mesure de production n’a par ailleurs été
calculée que sur une courte période et ne reflète donc que partiellement la production
mensuelle sur toute la période d’activité des annélides. La production n’est pas à priori
constante. Après la reprise des pluies, suite à une forte production, celle-ci peut se
réduire fortement (Lavelle, 1971).
En comparant ces résultats à ceux de la densité observée, il apparaît :
1) Dans les forêts, les turricules ont une durée de vie nettement supérieure à
l’année (3,9 année) alors que dans les forêts dégradées et les pâturages cette durée de
vie est moindre (149 jours et 48 jours respectivement).
2) la forte production dans la forêt dégradée (plus de 2 fois celle de forêt) met en
évidence une stimulation de production de turricules qui peut être reliée à une
augmentation de l’espace disponible suite à la déforestation. Cette stimulation d’activité

Turricules d’Andiodrilus pachoensis, discussion. Chapitre VI
178
peut aussi s’apparenter à un phénomène de succession secondaire dans lequel certains
processus ou espèces peuvent être stimulés (Horn, 1974).
3) Même si l’assemblage des espèces de vers peut influer sur la production de
turricules (Birang et al., 2003), la différence entre densité et production dans les
pâturages ne peut s’expliquer que par une forte dégradation (88% des turricules
dégradés ou détruits) dans un délai relativement court (de l’ordre de 3 à 5 semaines).
Nos résultats correspondent à ceux obtenus dans d’autres études qui ont montré
que la durée de vie d’un turricule en prairie était variable de quelques jours en milieu
tempéré (Binet and Le Bayon, 1998) à plus d’un an en savane tropicale (Decaëns, 2000).
La déforestation entraine également des modifications des propriétés physico-
chimiques du sol (chapitre IV).
Les propriétés physiques des turricules de forêts et de pâturages diffèrent
faiblement.
De texture granulométrique argilo-sableuse, les turricules de pâturages sont plus
mésoporeux que ceux des forêts. Cette augmentation de la mésoporosité peut
s’expliquer par un état peut-être plus dégradé. Un enrichissement des turricules de
pâturages en limons est observable, ce qui pourrait les rendre plus sensibles à l’érosion.
Un tel résultat a déjà été observé en Côte d’Ivoire (Nooren et al., 1995).
Cependant, la stabilité structurale des turricules ne se différencie pas dans les deux
milieux et s’avèrent très élevée. Plus de 80% à 90% de la structure est conservée lors de
tous les tests de stabilité structurale. Cette stabilité est instantanée et provient du
premier séchage. La stabilité des turricules dépend ainsi sûrement de la composition
originale du sol parent. La force élastique est positivement corrélée avec les teneurs en
argile et en carbonate du sol (Schrader and Zhang, 1997). L’étude de Hindell et al.
(1997) a montré que dans des sols plus humides, le potentiel matriciel
d’approximativement -35kPa et la stabilisation des turricules, sont dus à la cohésion des
particules du sol, au durcissement par l’âge (évaporation d’eau) mais aussi à la
croissance des microorganismes.
Cependant, les débris riches en carbohydrates des tissus végétaux sont également
responsables de la stabilité structurale des turricules d’annélides avec les
mucopolysaccharides dérivés des microorganismes de l’intestin des vers. Du mucilage et
d’autres métabolites se lient aux revêtements des particules minérales et ainsi renforcent
la cohésion des turricules. Les conséquences de cette association de matière organique
fraîche et de la phase minérale serait l’édification d’un pool de carbone protégé
physiquement qui serait libéré parallèlement à la dégradation des turricules (Golchin et

Turricules d’Andiodrilus pachoensis, discussion. Chapitre VI
179
al., 1994 ; Guggenberger et al., 1996). Le passage du sol au travers de l’intestin de
l’annélide brise les micro-agrégats préexistants (Barois et al., 1987) ; les débris
organiques incorporés serviraient alors comme nucleus pour de nouveaux agrégats. Dans
les déjections excrétées, le vieillissement et le séchage faciliteraient le rapprochement et
la liaison des polysaccharides issus des végétaux et des micro-organismes avec d’autres
composés organiques voire humiques associés aux argiles, stabilisant ainsi les nouveaux
micro agrégats.(Shipitalo and Protz, 1989).
En revanche, les turricules des trois milieux diffèrent par leurs propriétés
chimiques.
La concentration en carbone des turricules d’Andiodrilus pachoensis est d’environ
60 mg.g-1 dans les pâturages et 53 mg.g-1 dans les forêts. Le stock de matière
organique produit par an par les vers et piégé dans les turricules est d’environ 1,26 T.ha-
1.an-2 dans les forêts, 3T.ha-1.an-2 dans les forêts dégradées et 2,6 T.ha-1.an-2 dans les
pâturages. Ce stock est important.
Une alcalinisation des turricules de pâturages a été observée en raison de la
minéralisation de la matière organique qui s’opère dans l’appareil digestif des annélides.
Cela se manifeste par une réduction de la concentration en aluminium échangeable et
une augmentation des teneurs en cations basiques notamment du calcium provenant du
carbonate de calcium secrété par les glandes de Morren. La forte teneur en phosphore
assimilable est certainement à relier au taux de calcium, qui permettrait sa précipitation.
Les latosols présentent une forte tendance à retenir le phosphore dans leur phase solide
à cause des hautes teneurs en oxydes et hydroxydes de fer et d'aluminium. Le
phosphore se complexe pour former des oxydes insolubles. (Campello et al., 1994 ;
Fontes and Weeds, 1996). L’augmentation de la concentration en phosphore est sans
doute reliée à la réorganisation des oxydes et une libération du phosphore contenu dans
les argiles qui devient alors assimilable (Jiménez et al., 2003). Cette biodisponibilité du
phosphore crée d’une certaine manière une légère« eutrophisation» des turricules.
Autres facteurs influant l’activité de l’annélide.
L’étude des distributions spatiales des turricules dans les trois milieux (forêt, forêt
dégradée et pâturage) a permis d’analyser le rôle de facteurs locaux sur l’activité de
l’annélide Andiodrilus pachoensis.

Turricules d’Andiodrilus pachoensis, discussion. Chapitre VI
180
Les relations entre la distribution des turricules et les patterns spatiaux des
touffes de la graminée Brachiaria brisantha dans les pâturages ont été analysées. Les
liens entre la distribution des turricules et la quantité de litière sous forêt ont été étudiés.
Aucun patron général spécifique de distribution spatiale des turricules dans les
pâturages et/ou les forêts n’a été mis en évidence. Cette variabilité peut s’expliquer par
des sources locales d’hétérogénéité dans les deux types de milieux tels que le degré de
tassement du sol ou encore dans les pâturages, l’intensité du piétinement bovin. Les
patrons de distribution des turricules restent constants dans le temps
Dans les forêts, la distribution spatiale des turricules d’Andiodrilus p. est
indépendante de celle des autres structures biogéniques mais dépendante localement de
la position des arbres (agrégation aux échelles < 20 cm). Dans les forêts, la quantité de
litière n’est pas apparue comme un facteur régulant la distribution spatiale des turricules.
Néanmoins, des études en microcosmes ont cependant révélé que la qualité de la litière
peut aussi influer sur la quantité de turricules produits de différents Lumbricus (Flegel et
al., 1998).
Dans les pâturages, la distribution spatiale des turricules est inhibée par la
présence des termitières de Cornitermes. La faible densité en autres structures
biogéniques dans ce milieu n’a pas permis de vérifier la dépendance de la distribution
spatiale de ces turricules des autres facteurs. L’effet de la présence ou de l’absence de
touffes sur la production de turricules a présenté une forte variabilité. Les turricules
semblent cependant se concentrer davantage dans les zones où la température du sol est
plus faible. Les touffes de Brachiaria brisantha créent des zones, où la température plus
faible, est sans doute plus propice à l’activité des vers d’Andiodrilus pachoensis.
L’épiderme des vers étant très sensible à la dissécation (Lee, 1985 ; Edwards, 2004),
l’élévation de la température du sol et donc la réduction de l’humidité peut être un
facteur limitant l’activité de cet annélide dans la zone de sol nu.
D’autre part en milieu tempéré, il a été démontré que la distribution spatiale de
Lombricus terrestris peut aussi être corrélée à l’hydromorphie du sol : une tache de forte
biomasse correspondrait à une surface bien drainée et une tache de faible biomasse à
une surface hautement hydromorphique (Cannavacciuolo et al., 1998) mais une telle
relation n’a pas été mise en évidence dans les milieux tropicaux.

Turricules d’Andiodrilus pachoensis, discussion. Chapitre VI
181
Facteurs de dégradation des turricules.
Presque tous les ans, les pâturages sont brûlés. Le feu est également utilisé au
moment de la déforestation. Le feu joue par conséquent un rôle essentiel dans la
coévolution physico-chimique des turricules et des horizons de surface des sols.
Un des principaux effets du feu sur les turricules consiste en une réorganisation des
fractions granulométriques. Dans les deux milieux, la proportion d’argile des turricules
diminue jusqu’ à 31% lorsque les turricules sont fortement brûlés. Cela correspond donc
à une réorganisation de près de 6T d’argile granulométrique. Il y a formation de
pseudosables c'est-à-dire de particules d’argile cuites regroupées par déshydratation
(pectisation) qui forment des agrégats très stables ayant le comportement des sables
(Nzila and Garnier-Zarli, 1992 ; Mboukou and Lavelle, 1997). Les turricules prennent
également une couleur rougeâtre caractéristique de la présence d’hématites (Photo 6A).
Cette réorganisation granulométrique confère aux turricules une stabilité structurale
accrue.
L’autre effet est la calcination de la matière organique qui provoque la minéralisation
de pratiquement tout le carbone contenu dans les turricules lorsque l’intensité et la durée
du feu sont importantes.
Le feu joue un rôle important dans la coévolution des turricules d’Andiodrilus et
des horizons de surface du sol.
De nombreuses études ont mis en évidence que la force de la pluie et le
ruissellement jouaient un rôle primordial dans la destruction des turricules (Nooren et al.,
1995 ; Binet and Le Bayon, 1998 ; Le Bayon and Binet, 1999)
Les expériences de simulation de pluie ont permis de mettre en évidence que
l’érosion des turricules frais est très faible ; elle ne dépasse pas 2,5% de la masse
initiale. Dans les forêts, l’érosion du sol nu est importante (plus de 55% de la masse est
érodée dès la première minute). Dans les pâturages, cette érosion atteint 40% de la
masse initiale. Les sols de nos sites d’études sont fortement sensibles à l’érosion suscitée
par les épisodes pluvieux. Le relief propice à la battance présente les stigmates d’une
forte érosion de surface (Photo 6B).
La pluie n’est pas un facteur prépondérant dans la dégradation physique des
turricules mais son impact est considérable sur les horizons non protégés des pâturages.

Turricules d’Andiodrilus pachoensis, discussion. Chapitre VI
182
Les turricules écrasés présentent toujours une érosion plus forte que celle des
turricules frais mais réduite par rapport aux horizons témoins. Ainsi la stabilité des
turricules pourrait s’expliquer par la présence d’une enveloppe ou cortex qui protège la
structure des agressions extérieures du milieu. Ce cortex a d’ores et déjà été mis en
évidence pour des turricules de Millsonia anomala (Oligochaeta:Megascolecidae) dans des
sols de savane de Côte d’Ivoire (Blanchart et al., 1993b). Cependant, les analyses
micromorphologiques réalisées n’ont pas permis de mettre en évidence une cuticule de
protection autour des turricules d’Andiodrilus pachoensis.
Lorsqu’ils sont écrasés, les turricules perdent leur stabilité et sont facilement
dégradés par la pluie.
Le piétinement du bétail est donc certainement un facteur prépondérant dans la
dégradation des turricules.
Effets des turricules d’Andiodrilus sur le fonctionnement du sol.
L’absence de différence entre les propriétés physico-chimiques des turricules
d’Andiodrilus pachoensis et les horizons de surface (0-30 cm) des pâturages et des forêts
suggèrent, compte tenu de la production estimée des turricules dans les deux milieux,
que les turricules des vers d’Andiodrilus participent fortement à la constitution des
horizons de surface.
Les turricules d’Andiodrilus p. plus poreux que le sol témoin participent peu au
tassement de surface.
Par ailleurs, le stock de matière organique contenu dans les turricules est
considérable, de 1,26 T.ha-1.an-2 à 2,6 T.ha-1.an-2 dans les forêts et pâturages. Les stocks
en cations et en phosphore assimilable sont également importants.
Ces résultats sont par ailleurs déjà largement documentés ; le carbone et les
nutriments sont généralement protégés au sein des turricules ; la matière organique y
est séquestrée et protégée de la minéralisation. (McInerney and Bolger, 2000 ; Mora et
al., 2003); (Bossuyt et al., 2005); (Pulleman et al., 2005). Ces turricules peuvent être
considérés dans ce milieu comme une niche de régénération pour les espèces de plantes
et l’activité des microorganismes (Decaëns et al., 2003).
Dans les pâturages, le piétinement bovin apparaît comme étant le premier facteur de
dégradation des turricules. Suite à l’écrasement, les turricules brisés sous forme de gros

Turricules d’Andiodrilus pachoensis, discussion. Chapitre VI
183
agglomérats, s’intègre aux horizons de surface. L’effet de la pluie et/ou du feu combiné
continue ensuite la dégradation physique des structures aboutissant ainsi à une
désagrégation des agglomérats qui se dissolvent et se fondent progressivement le long
du profil permettant ainsi un retour rapide (3 à 5 semaines) des stocks de matière
organique et de cations au sol. L’intégration au sol des turricules et le brûlis annuel qui
détruit la matière organique permettent d’expliquer l’alcalinisation observée des horizons
de surface des pâturages (ph= 6,4).La couche superficielle compacte entre les touffes de
graminées nous conforte dans notre hypothèse. Cette couche comme dans d’autres
contextes en Amazonie résulte du piétinement du sol par le bétail et de l’action de la
pluie dans ces pâturages. (Grimaldi et al., 1993 ; Chauvel, 1997) d’autant plus que les
risques d’érosion du sol peuvent être augmentés par l’accumulation des turricules de
surface des sols compactés (Binet and Le Bayon, 1998)
Ainsi pour compléter cette étude, des mesures de l’attachement des turricules à la
surface du sol devront être réalisées sur le terrain et le rôle des microorganismes dans la
dégradation des structures biogéniques devra être analysé.
A BA B
Photo 6: A : Vieux turricules brûlés d’Andiodrilus pachoensis en dégradation sous un tronc couché dans un
pâturage. B : Figures d’érosion dans les pâturages de Benfica.
Grimaldi, M.


Mise au point d’une typologie des structures biogéniques. Chapitre VI
184
VII. Mise au point d’une typologie des structures
biogéniques.
Après avoir détaillé les distributions spatiales et les propriétés physico-chimiques
des termitières de Cornitermes sp. et des turricules d’Andiodrilus pachoensis, une étude
comparative des structures biogéniques produites par différentes espèces ingénieurs de
l’écosystème (nids et placages de termites humivores, déblais de fourmis…) a été
entreprise.
Cette étude a pour objectif de mettre en évidence le rôle des espèces ingénieurs
dans le fonctionnement du sol au travers de la séquestration de la matière organique ou
encore de leur influence dans l’agrégation des horizons de surface. Les typologies
obtenues par différentes analyses (NIRS, chimie, micromorphologie) ont pour objectif de
déterminer les meilleurs outils permettant de distinguer les structures biogéniques afin
de pouvoir modéliser le fonctionnement des sols et d’établir un indice de qualité des sols
basé sur les structures biogéniques.

Typologie des structures biogéniques, résultats connus. Chapitre VII.1
185
VII.1 INTRODUCTION
L’analyse spectrale infrarouge de la matière organique contenue dans les
structures biogéniques donne des signatures particulières pour chaque espèce
d’invertébré laissant espérer que les spectres permettraient d’identifier les organismes
responsables de la production d’agrégats dans le sol (Hedde et al., 2005 ; Pelosi, 2005).
Par leur comportement alimentaire, les annélides et les termites, influencent
nettement la matière organique contenue dans leur structure biogénique en y mêlant des
mucus corporels (Lee and Wood, 1971 ; Sleeman and Brewer, 1972 ; Grassé, 1984a ;
Garnier-Sillam et al., 1985 ; Martin et al., 1987).
La dégradation de la matière organique est chimique, via une hydrolyse alcaline
(Rouland-Lefèvre, 2000) et biologique, via l’activité microbienne. Du fait du transit
intestinal, la matière organique est fortement modifiée aboutissant à des molécules qui
marquent les structures biogéniques de façon spécifique.
Deux études ont d’ores et déjà mis en évidence que les structures qui
minéralisent le plus rapidement sont les structures biogéniques et en particulier les
turricules et les nids de termites humivores (Hedde et al., 2005 ; Pelosi, 2005). Une
analyse multivariée montre que la première composante principale de l’analyse peut être
assimilée à un gradient de concentration de matière organique dans les structures
biogéniques. Cette composante permet de rapprocher certains groupes ; tout d’abord, les
structures feuilletés des nids de termites qui sont isolées des autres groupes. Ces
structures se distinguent par leur teneur très importante en carbone et par une forte
minéralisation, ce qui atteste d’une forte labilité du carbone. Ensuite, vient les turricules
d’annélides et les parois produites par les termites humivores. Ces dernières sont moins
riches en matière organique ; ces deux types de structures sont très proches et leurs
taux de carbone et de minéralisation sont quasi-similaires. Enfin, le sol témoin et les
fourmilières présentent un faible taux de carbone; pourtant, celui-ci semble plus modifié
dans les fourmilières que dans le sol témoin car la minéralisation y est plus importante.
Ceci traduit une labilité plus importante de la matière organique, certainement due aux
actions métaboliques du sol par l’organisme ingénieur.
Les signatures NIRS (Near Infrared Spectrophotometry) dépendent également de
la variabilité locale du sol et de la végétation (variation en fonction de la parcelle). En
revanche, s’il n’existe pas d’effet sensible de la nature et de la diversité végétale sur la
signature des turricules frais de vers, cet effet existe sur la signature des classes
micromorphologiques d’agrégats témoins des horizons de surface (Pelosi, 2005).

Typologie des structures biogéniques, résultats connus. Chapitre VII.1
186
Les paramètres qui influencent la signature des structures du sol sont en
interaction, ce qui rend difficile la détermination de l’importance relative de chacun de
ces facteurs dans la signature d’une structure biogénique.
La détermination de l’origine des agrégats d’un sol est une approche difficile sur le
plan de l’expérimentation. Trois typologies ont été testées afin de simplifier les analyses
dans l’objectif de classer ces structures en fonction des services écosystémiques qu’elles
assurent:
-typologie basée sur l’étude de la matière organique (NIRS)
-typologie basée sur une approche chimique (dosage du carbone et des cations)
-typologie basée sur l’estimation de la porosité par micromorphologie
Ces études ont été réalisées afin de mettre en évidence une éventuelle
coévolution des structures biogéniques et des écosystèmes en vue d’identifier un
indicateur physique ou chimique qui permettrait de simplifier la lourde approche
analytique nécessaire à l’identification des structures biogéniques.
Dans un premier temps, une étude de la matière organique par la méthode NIRS
nous a permis d’établir une typologie des structures biogéniques incluant non seulement
les deux structures biogéniques fraîches étudiées dans les chapitres précédents
(termitière de Cornitermes sp. et turricules d’Andiodrilus pachoensis), et ces deux
structures dans leur état brûlé. Cet état brûlé correspond à l’effet de la culture sur brûlis
mais aussi à la destruction des colonies de termite par les paysans. Même si dans les
chapitres précédents, le feu n’est pas apparu comme étant le principal facteur de
dégradation des structures biogéniques, il participe à la modification des propriétés
physico-chimique des structures biogéniques et par conséquent constitue un état
particulier de ces structures.

Typologie des structures biogéniques, matériel et méthodes. Chapitre VII.2
187
VII.2 MATERIEL ET METHODES
VII.2.1 Etude de la matière organique par la méthode NIRS (Near Infrared Resonance
Spectophotometry).
La spectroscopie proche infrarouge (IR) est une spectroscopie d’absorption dont le
principe repose sur l’absorption du rayonnement proche IR par la matière organique.
La méthode s’intéresse aux liaisons chimiques spécifiques du constituant : O-H, N-
H, C-H, etc. Les liaisons chimiques se comportent comme des oscillateurs qui vibrent en
permanence à des fréquences différentes en fonction de leur nature. Une liaison chimique
particulière peut absorber une radiation proche IR dont la fréquence est égale à sa
fréquence de vibration et ainsi passer d’un état fondamental à un état excité. De même,
l’énergie des radiations dont les fréquences sont des multiples de la fréquence
fondamentale peut être absorbée. On parle alors d’harmoniques. Dans la zone du proche
IR, les absorptions ne sont pas dues aux vibrations fondamentales des molécules, mais
aux vibrations harmoniques et aux vibrations de combinaisons. Le spectre d’absorption
recueilli est complexe, les bandes d’absorption de toutes les liaisons chimiques se
chevauchant.
L’interprétation du spectre est donc de fait très difficile. C’est pourquoi des
méthodes de statistiques multivariées sont appliquées afin de comparer les différents
objets entre eux. L’analyse quantitative classique utilise les lois de Beer-Lambert. Cette
analyse nécessite la présence de bandes spécifiques à chaque produit, ce qui limite
fortement son utilisation. Grâce à l’utilisation d’analyses factorielles, le dosage peut être
envisagé sur des mélanges complexes. Lors de la réalisation d’une acquisition par
spectroscopie proche IR, des spectres sont enregistrés. Ils représentent l’absorption
lumineuse en fonction de la longueur d’onde. On obtient ainsi de très nombreuses
valeurs numériques par échantillon. Ces spectres sont par la suite découpés en bandes
de différentes longueur d’onde qui sont traitées comme des variables indépendantes
dans les analyses multivariées (Hruschka, 1990 ; Murray and Williams, 1990 ; Williams,
1990 ; Reeves et al., 1999).

Typologie des structures biogéniques, matériel et méthodes. Chapitre VII.2
188
VII.2.2 Analyse chimique des structures biogéniques
Le détail des analyses chimiques réalisées est détaillé dans le chapitre III.2.
Une analyse multivariée (ACP) a été réalisée a partir des résultats obtenus dans
les chapitres précédents concernant la teneur en carbone, le pH, les concentrations en
cations : aluminium échangeable, calcium, magnésium, potassium et phosphore
assimilable. Les caractéristiques chimiques des murailles de termitière Cornitermes sp
dans leur état frais et brûlé ainsi que celles des turricules d’Andiodrilus p. (frais et brûlés)
ont été comparées à celles des horizons des forêts et des pâturages.
VII.2.3 Analyses micromorphologiques de Lames minces.
L’analyse micromorphologique quantitative du sol a permis de comparer l’état
structural de différents types de structures biogéniques. Soixante cinq échantillons, à
structure conservée, ont été prélevés dans les deux milieux (Forêt, pâturage). Ces
échantillons correspondent à des turricules d’Andiodrilus pachoensis, des murailles et du
feuilleté de termitière Cornitermes sp., mais aussi des déblais de fourmis, des nids et du
placage de termites humivores. Pour les trois dernières catégories (fourmis, nids et
placages de termites humivores) l’étude a été réalisée dans un premier temps au niveau
du groupe taxonomique puis au niveau des différentes espèces.
Afin de limiter les modifications structurales, les structures biogéniques ont été
déshydratées par échange eau-acétone avant d’être imprégnées par une résine
additionnée d’une substance fluorescente permettant de visualiser l’espace poral sous
lumière UV (Hallaire and Curmi, 1994). Une analyse quantitative de l’espace poral
(Ringrose-Voase, 1991 ; Hallaire and Curmi, 1994) a été effectuée par l’analyse d’images
à deux échelles de résolution : 5.88µm/pixel et 0,47µm/pixel. Ces deux échelles
permettent de prendre en compte des pores distincts par leur taille et leur nature :
fissures, tubules et pores d’assemblage d’agrégats à la première ; cavités entre les
particules du squelette à la deuxième.
Le système poral est un réseau tridimensionnel de pores interconnectés ; son
image suivant un plan de coupe supprime ainsi une partie importante des connexions
entre les pores. Les pores individualisés sur l’image sont nommés « poroïde » afin
d’éviter toute confusion (Moran et al., 1988).

Typologie des structures biogéniques, matériel et méthodes. Chapitre VII.2
189
Une typologie des poroïdes à partir de leur taille et de leur forme a été retenue :
● Cinq classes de taille: très petits, petits, moyens, gros et très gros (Tableau 29).
Tableau 29 : Classement des poroïdes en fonction de leur taille aux 2 échelles d’observation.
● Trois classes de forme distincte par l’indice d’allongement I (Tableau 30)
I = P²/4πS où P est le périmètre du poroïde (Coster and Cherman, 1989)
I Type de Poroïdes
I<5 cavités et tubules
5< I <10 fissures
I>10 pores complexes
Tableau 30 : Type de poroïdes en fonction de l’indice de forme.
Echelle d’observation
Très petits petits moyens gros Très gros
5,88 µm/pixel <0,18mm² 0,18 mm² < aire < 0,35mm²
0,35 mm² < aire < 1,8 mm²
1,8 mm² < aire < 18 mm²
>18 mm²
0,47 µm/pixel <0,04mm² 0,04mm² < aire < 0,08 mm²
0,08mm² < aire < 0,38m²
0,38mm² < aire < 3,8mm²
> 3,8mm²

Typologie des structures biogéniques, discussion. Chapitre VII.4
190
VII.3 RESULTATS
VII.3.1 Typologie basée sur l’étude de la matière organique par la méthode NIRS.
La méthode NIRS permet de comparer les différentes structures biogéniques, dans
leur état frais ou brûlé (turricules d’Andiodrilus p., feuilleté et paroi de termitières de
Cornitermes sp)., aux horizons témoins (0 -10 cm) de pâturages et de forêts sur le plan
de la caractérisation de la matière organique.
Figure 68 : Cercle de corrélation (axes 1 et 2) des longueurs d’onde (à gauche) et Projection dans le plan
factoriel des différentes structures biogéniques (à droite) selon les 2 premiers axes (95% de la variabilité
observée). TermP : Paroi de termitière Cornitermes sp. ; TermPBrulé : Paroi de termitière Cornitermes
sp.brûlé ; TermF : Feuilleté de termitière de Cornitermes sp., TurP : Turricules de Pâturages ; TurF : Turricules
d’Andiodrilus de forêt ; TurBrulé : Turricules d’Andiodrilus faiblement brûlé, TurBrulé++ : Turricules
d’Andiodrilus fortement brûlé P : sol témoin de pâturage ; F : sol témoin de forêt.
Les deux premiers facteurs de l’ACP expliquent 95% de l’inertie totale des
variables NIRS (79% et 16 % respectivement). Une analyse discriminante a montré une
séparation significative des différentes structures biogéniques étudiées (P<0.001).
La projection des résultats NIRS des structures biogéniques dans le plan factoriel
des deux premiers facteurs de l’ACP révèle une séparation entre les structures brûlées et
les turricules de pâturage d’un groupe composé des sols témoins, des turricules de forêt
et des deux composants des termitières (feuilleté et muraille) selon l’axe 1. (Figure 61)
P<0,001

Typologie des structures biogéniques, discussion. Chapitre VII.4
191
L’axe 2 sépare quant à lui, le groupe composé des sols témoins et de la muraille
de termitière brûlée et non brûlée d’un autre groupe composé des turricules des deux
milieux, des turricules brûlés et du feuilleté de termitière.
Cependant la projection des longueurs d’onde dans le cercle de corrélation montre
que celles-ci ne se séparent que selon l’axe 2. Les longueurs d’ondes sont donc
fortement corrélées selon l’axe 1. Ainsi selon cet axe les structures biogéniques pauvres
en matière organique s’opposent aux plus riches (Figure 68).
La typologie obtenue par cette méthode sépare deux groupes d’agrégats :
-1- ceux provenant des sols témoins des deux milieux, des turricules frais des forêts
et des structures termitiques (muraille et feuilleté).
-2- ceux issus de toutes les structures biogéniques brûlées et des turricules des
pâturages
Ainsi l’analyse NIRS a permis d’opposer de façon significative les structures
riches en matière organique fraîche de celles dont la matière organique est plus
dégradée. L’état de la matière organique (fraîche ou dégradée) semble ainsi jouer
un rôle prépondérant la séparation des agrégats issus des différentes structures
biogéniques. La deshydratation est un processus qui peut être associée à la
dégradation de la matière organique.
VI.3.1.b Analyses multivariées des données de chimie.
Une analyse multivariée ACP a été réalisée sur les caractéristiques chimiques des
structures afin d’analyser la pertinence de ces caractéristiques dans l’établissement d’une
typologie des agrégats.

Typologie des structures biogéniques, discussion. Chapitre VII.4
192
Figure 69 : Cercle de corrélation (axes 1 et 2) des éléments chimiques et du pH (à gauche) ; Projection dans le
plan factoriel des différentes structures biogéniques (à droite) selon les 2 premiers axes (73% de la variabilité
observée). TermP : Paroi de termitière Cornitermes sp. ; TermF : Feuilleté de termitière de Cornitermes sp.,
TurP : Turricules d’Andiodrilus de Pâturages ; TurF : Turricules d’Andiodrilus de forêt ; P : sol témoin de
pâturage ; F : sol témoin de forêt.
Les deux premiers facteurs de l’ACP expliquent 73% (56% et 17 % respectivement)
de l’inertie totale des variables de chimie (pH, concentration en cations, carbone et
phosphore assimilable).
Une analyses discriminante a montré une séparation significative des différentes
structures biogéniques étudiées (P<0.001) (Figure 69).
La projection des résultats des caractéristiques chimiques des structures
biogéniques dans le plan factoriel des deux premiers facteurs de l’ACP révèle une
séparation entre les différents types de structures biogéniques et les sols témoins.
L’axe 1 (56% de la variabilité) sépare nettement les turricules des pâturages
(brûlés ou non) d’un groupe formé des turricules de forêts, de la muraille des termitières
(fraîche et brûlée) et des sols témoins. Selon le même axe, le cercle de corrélation révèle
une séparation entre le carbone et l’aluminium échangeable des autres cations, pH et
phosphore assimilable.
L’axe 2 (16% de la variabilité) met en évidence un gradient débutant par la muraille
de termitière brûlée puis les turricules brûlés aux deux intensités et la muraille de
termitière fraîche, suivi des turricules de pâturage et de forêt pour se terminer par les
sols témoins. Selon cet axe, le cercle de corrélation sépare le pH et le phosphore
assimilable, des autres cations et du carbone (Figure 69).
P<0,001

Typologie des structures biogéniques, discussion. Chapitre VII.4
193
La typologie des agrégats selon leurs caractéristiques chimiques séparent
les turricules dans leurs deux états (frais ou brûlés) des murailles de termitières
fraîches ou brûlées et des horizons témoins des deux milieux.
L’état de la matière organique et l’acidité (principalement l’acidité de
réserve) des structures constituent ici les facteurs principaux de différenciation des
structures biogéniques.
VII.3.2 Analyse micromorphologique des lames minces de biostructures.
VII.3.2.a Description succincte des lames minces correspondant aux différentes
structures biogéniques.
Des lames minces d’environ 8,5 cm² ont été fabriquées à partir de différentes
structures biogéniques récoltées sur le terrain et conservées de manière à préserver leur
structure physique.
Deux grandes catégories de structures biogéniques peuvent être distinguées :
- les structures non construites (turricules d’annélides)
- les structures construites (structures termitiques ou des fourmis)
Parmi les structures construites se distinguent les structures d’origine cibariale
(muraille de termitière de Cornitermes, placage de termites) des structures proctodéales
(feuilleté de termites Cornitermes, nids des termites humivores). Alors que les structures
cibariales sont des boulettes de terre découpée par les mandibules auxquelles est ajoutée
de la salive, les structures proctodéales (comme les turricules) sont des boulettes
remaniées au sein du tube digestif des invertébrés.

Typologie des structures biogéniques, discussion. Chapitre VII.4
194
Figure 70 : Exemples d’images de lames minces des différentes structures biogéniques : Paroi de termitière
Cornitermes sp. (A) ; Feuilleté de termitière de Cornitermes sp. (B), Nid de termitière humivore (C), Placage de
termitière humivore (D), Turricule d’Andiodrilus pachoensis (E) ; déblais de fourmis (F).
A) La paroi de termitière du genre Cornitermes sp. (Figure 70, A)
La paroi de la termitière est une structure construite d’origine cibariale (mélange
de boulettes de sol et de salive). La couleur de cette structure (10YR 5/6 code Munsel)
est équivalente à celle des horizons 10-30 cm des pâturages. La structure de la paroi de
termitière est uniforme avec présence de fissures plus ou moins fines non orientées,
sécantes qui délimitent partiellement des agrégats. La distribution d’éléments de
fabrication plus larges est liée par des éléments plus petits et le fond matriciel dense.
Cette distribution est caractéristique d’un assemblage porphyrique (Stoops et Jongerius,
1975). La microstructure est microagrégée avec la présence de micro agrégats
organiques dans l’espace intergranulaire. Sur les lames étudiées, quelques quartzs de
tailles plurimillimétriques, des minéraux distribués aléatoirement et quelques concrétions
ferrugineuses plus rares de couleur rouille sont présents. La porosité est constituée de
pores ronds aux bords irréguliers sans doute liée à la construction et des fissures
anguleuses. Aucune cutane n’a été observée.

Typologie des structures biogéniques, discussion. Chapitre VII.4
195
B) Le feuilleté de termitière Cornitermes sp. (Figure 70, B)
Le feuilleté de termitière, d’origine proctodéale, est remanié et de couleur brune
(10YR 4/3). Très dense et uniforme il présente de rares fissures très fines globalement
droites aux bords réguliers. La macroporosité est inexistante à l’échelle de la lame mince.
Aucun large élément n’est observable. La structure est massive. Aucun minéraux et
aucune concrétion n’est observable. A une échelle plus fine, une importante porosité fine
d’assemblage très connectée apparaît. La microstructure est complexe.
C) Parois des nids de termites humivores (Figure 70, C)
La structure de la paroi de termitière humivore est complexe et de couleur brune
(10YR 4/3). Le relief est très prononcé. Elle est constituée de galettes proctodéales
remaniées par l’ajout de minéraux qui ne sont pas passés par le tube digestif.
Des zones présentent une structure proche de celle du feuilleté du type Cornitermes,
d’origine proctodéale. Ces zones sont massives, uniformes avec une porosité fine et une
microstructure complexe. D’autres se caractérisent par des assemblages porphyriques
avec de larges agrégats inclus dans un fond présentant de nombreux éléments plus
petits dans l’espace intergranulaire. Dans ces zones la microstructure est microagrégée.
Sur les lames étudiées, de nombreux quartz distribués aléatoirement sont présents. La
porosité est constituée de quelques fissures irrégulières anguleuses. Aucune cutane n’a
été observée. Des descriptions comparables ont déjà été réalisées sur des termitières du
Congo (Garnier-sillam, 1987).
D) Placage de termites humivores (Figure 70, D)
La structure du placage de termite humivore est proche de celle observée pour le
nid. Sa couleur est brun grisâtre 10YR5/2. Les larges cavités observables proviennent de
la technique de fabrication des lames. En effet les placages de termites sont de formes
concaves et convexes et par conséquent la lame en 2 dimensions présente de larges
cavités. L’assemblage est de type porphyrique avec présence d’un grand nombre de
fissures connectées délimitant des agrégats organo-mineraux. La porosité est complétée
par de nombreux pores complexes. la microporosité est très faible, La microstructure est
massive.
E) Turricule d’Andiodrilus pachoensis (Figure 70, E)
Les turricules d’Andiodrilus pachoensis sont des structures biogéniques non
construites constituées d’une accumulation de déjections ayant la forme de galettes avec
un canal central conservé. Ces fécès sont non remaniés par cette espèce mais peuvent
l’être par d’autres espèces d’invertébrés du sol.

Typologie des structures biogéniques, discussion. Chapitre VII.4
196
Leur structure est caractérisée par la présence d’un très grand nombre de cristaux
de quartz reliés les uns aux autres par une matrice massive. Cette distribution des
éléments rappelle les assemblages porphyriques. La microstructure est complexe
composée de zones granulaires avec des pores complexes connectés et des zones plus
massives. Sur les lames étudiées, quelques pores de forme complexe et de grande taille
sont observables. D’autres minéraux distribués aléatoirement et quelques concrétions
plus rares sont présents. La porosité est constituée de pores complexes aux bords
irréguliers. Aucune cutane n’a été observée.
F) Déblais de fourmis (Figure 70, F)
Les déblais de fourmis sont des structures construites mais dont l’origine est buccale. La
structure des déblais de fourmis est très aérienne, constituée de cristaux de quartz bien
délimités de forme polyédrique en grande quantité. Ces cristaux sont de tailles
plurimillimétriques comparables à ceux observés dans la muraille de termitière de
Cornitermes sp. Ces cristaux sont liés par de petites zones de matrice très uniforme et
massive. La microstructure est massive. La porosité est constituée de pores complexes
très connectés et plutôt de grande taille.
VII.3.2.b Quantification de la porosité des différentes structures biogéniques.
Pour les fourmis et les termites humivores, les résultats obtenus sont des
moyennes de plusieurs images prises sur différentes lames correspondant à des espèces
différentes. L’écart type calculé est un écart type « intra espèce » et « inter espèce ».
Pour les deux espèces étudiées, le ver Andiodrilus et le termite Cornitermes,
l’écart type correspond à des variations entres les images étudiées, donc un écart type
« intra espèce ».
Des exemples d’images binaires (blanc = porosité) obtenues afin de quantifier la
porosité dans les structures biogéniques se situent dans les annexes 14 (résolution de
5,88µm par pixel) et annexe 15 (résolution de 0,47µm par pixel).
A la résolution de 5,88µm par pixel la mésoporosité est quantifiée alors que la
résolution de 0,47µm par pixel permet de mesurer la microporosité.
b1) Résolution de 5,88µm par pixel (annexe 21)
A cette résolution la mésoporosité est prise en compte.

Typologie des structures biogéniques, discussion. Chapitre VII.4
197
Pour rappel, nous avons cinq classes de taille de poroïdes : Très petit (TP), petit
(P), moyen (M), grand (G) et très grand (TG) et 3 types de poroïdes : Cavités (Cav),
Fissures (Fis) ou pores complexes (Comp).
Les résultats comparent la porosité de 7 types de structures biogéniques :
- Muraille de termitière épigée de termites xylophages (Cornitermes sp.) : TermCPa
- Feuilleté de termitière épigée de termites xylophages (Corniterme sp.) : TermCFe
- Paroi de termitière épigée de termites humivores : TermHNd
- Placage de termitière épigée de termites humivores : TermHPl
- Déblai de fourmis : Fourmis
- Turricules frais d’annélides anéciques (Andiodrilus p.) : Tur
- Turricules brûlés d’annélides anéciques (Andiodrilus p.) : Tur Brûlé
Pour chaque type de structures au moins 3 lames ont été examinées et 3 images
ont été analysées sur chacune de ces lames. Les images étudiées à cette échelle
mesurent 1200 * 1100 pixels.
Figure 71 : Porosité des structures biogéniques par analyse micromorphologique à la résolution optique de 5,88
µm par pixel (mésoporosité). Projection dans le pan factoriel des trois types de pores : Cavités (Cav), Fissures
(Fis) et pores complexes (Comp) des 5 tailles : très petit (TP), petit (P), moyen (M), grand (G) et très grand
(TG) et des structures biogéniques de divers invertébrés : Turricule frais d’annélide anécique (Tur), turricule
brûlé d’annélide anécique (Tur Brulé), muraille de nids épigées de termites xylophages TermCPa, feuilleté de
ces termitières de xylophages (TermCFe), Paroi de termitières d’humivores (TermHNd), placage de termites
humivores (TermHPl) et déblais de fourmis (Fourmi). L’axe 1 explique 65% de la variabilité et l’axe 2 : 20%.

Typologie des structures biogéniques, discussion. Chapitre VII.4
198
Quelque soit la structure biogénique étudiée, la porosité totale est inférieure à
25% de la surface étudiée.
Les déblais de fourmis (d’origine buccale) sont à cette échelle d’observation, les
structures biogéniques les plus poreuses mais présentent également une grande
variabilité (25% ± 18% de porosité).
La muraille de termitière de Cornitermes sp. (cibariale) est constituée d’environ
25% de porosité. Ce pourcentage de porosité est néanmoins très variable (25% ± 12%).
La porosité des turricules d’Andiodrilus pachoensis est de l’ordre de 18% ± 13%,
Cette porosité diminue à 11% lorsque le turricule est brûlé.
Les structures biogéniques les moins poreuses correspondent au feuilleté de
termitière Cornitermes sp. (8% ± 2%) et aux nids et placage des termites humivores (de
8% à 10%).
Dans les turricules d’Andiodrilus la porosité est décomposée en pores ronds ou
tubulaires de l’ordre de 35%, de fissures à 20%, et de pores complexes à 45%. Lorsque
le turricule est brûlé, la porosité complexe et les fissures diminuent au profit de la
porosité ronde qui représente alors 50% de la porosité totale.
La paroi de termitière Cornitermes se caractérise par une porosité ronde de 25%,
des fissures à la hauteur de 5% et des pores complexes à 70%.
Dans le feuilleté de termitière Cornitermes, les pores complexes ne représentent
que 5% et les pores ronds près de 80%.
Les nids de termitière humivore présentent une porosité maximale du type rond
(50%) alors que les placages sont constitués de façon équitable de pores ronds (40%) et
de fissures (40%).
Les déblais de fourmis sont constitués en majorité de pores complexes (50%), de
30% de pores ronds et 20% de fissures.
Quelque soit la structure biogénique étudiée, les pores ronds sont répartis de
manière équivalente dans chaque classe de taille. En revanche les pores complexes font
au minimum 1,8mm² (classe de taille 4). Les fissures sont de toutes tailles mais plutôt
supérieures à 0,35mm² sauf dans les termitières de Cornitermes et les turricules
d’Andiodrilus pachoensis brûlés où aucune fissure n’est mesurée au-delà de 18mm².
La projection dans le plan factoriel des structures biogéniques et des différents
types de poroïdes oppose, selon l’axe 1, les poroïdes complexes de moyenne et de très
grande taille, associés aux déblais de fourmis et à la paroi de termitières de Cornitermes
sp. aux autres pores et biostructures. Ces structures sont des structures remaniées. Ainsi

Typologie des structures biogéniques, discussion. Chapitre VII.4
199
le premier facteur de différenciation des structures selon leur porosité serait le
remaniement des boulettes organo-minérales lors de la construction de la structure
(Figure 71).
b2) Résolution de 0,47µm par pixel (annexe 22)
A cette résolution la microporosité a été prise en compte.
Les images étudiées à cette échelle mesurent 1500 * 1200 pixels.
Les mêmes structures biogéniques que dans la précédente analyse ont été
étudiées à cette résolution optique.
Figure 72 : Porosité des structures biogéniques par analyse micromorphologique à la résolution optique de 0,47
µm par pixel (microporosité). Projection dans le pan factoriel des trois types de pores : Cavités (Cav), Fissures
(Fis) et pores complexes (Comp) des 5 tailles : très petit (TP), petit (P), moyen (M), grand (G) et très grand
(TG) et des structures biogéniques de divers invertébrés : Turricule frais d’annélide anécique (Tur), turricule
brûlé d’annélide anécique (Tur Brulé), muraille de nids épigées de termites xylophages TermCPa, feuilleté de
ces termitières de xylophage (TermCFe), Paroi de termitières d’humivore (TermHNd), placage de termites
humivores (TermHPl) et déblais de fourmis (Fourmi). L’axe 1 explique 55% de la variabilité et l’axe 2 : 29%.
A cette résolution, la porosité la plus importante est rencontrée dans les turricules
d’Andiodrilus (21% ± 11% de la surface étudiée). Lorsque le turricule est brûlé, la
porosité est réduite à 15%.

Typologie des structures biogéniques, discussion. Chapitre VII.4
200
Alors que la paroi de termitière de Cornitermes est caractérisée par une porosité
de 13% ± 10%, le feuilleté présente une porosité plus importante (18% ± 11%).
La porosité des nids de termitière humivore est la plus faible avec 11% ± 9% de
la surface étudiée alors que les placages révèlent une porosité légèrement supérieure à
17% ± 3%.
Une porosité quantitativement équivalente est rencontrée dans les déblais de
fourmis (17% ± 9%).
L’étude de la quantification des poroïdes en fonction de leur type montre que les
pores de type ronds, tubulaires et complexes sont en proportions équivalents dans toutes
les structures biogéniques étudiées de l’ordre de 30% à 50% de la porosité totale pour
chaque type.
La quantité de fissures est légèrement plus basse avec des valeurs comprises
entre 21% et 40%.
La quantité de pores ronds est la plus faible dans les placages de termites
humivores (21%), puis dans les turricules d’Andiodrilus (24%). Ces structures
biogéniques présentent des quantités de pores complexes plus importantes (61% et 50%
respectivement).
La quantification de la porosité des différentes structures biogéniques en fonction
de la taille de ceux-ci au sein de chaque type de pores met en évidence qu’à cette
résolution, aucun pore de taille supérieur à 3,8mm² n’est rencontré quelque soit la
structure biogénique étudiée. Des pores des trois classes de forme sont présents dans
toutes les structures lorsque leur taille est inférieure à 0,04mm². Ces très petits pores
représentent près de 70% de la porosité du feuilleté de termitière de Cornitermes et près
de 65% des nids de termites humivores. Ils ne constituent que 35% de la porosité des
turricules d’Andiodrilus. Cette classe de taille de pore est largement dominée dans
toutes les structures biogéniques par les pores ronds ou tubulaires. Les pores dont la
taille est comprise en 0,04mm² et 0,08mm² sont peu abondants dans toutes les
structures biogéniques de l’ordre de 10 à 20%. Les pores des classes de taille 3 et 4
(pores dont la taille est comprise entre 0,08mm² et 3,8mm²) sont largement dominés
par les fissures et essentiellement les pores de forme complexe. Aucun pore de taille
supérieure à 0,38mm² n’est rencontré à cette résolution dans la paroi des termitières de
Cornitermes sp. et dans les turricules brûlés.
La projection simultanée dans le plan factoriel des structures biogéniques et des
types de poroïdes révèle 3 groupes de structures (Figure 72) :

Typologie des structures biogéniques, discussion. Chapitre VII.4
201
1. Les turricules brûlés d’Andiodrilus p., les murailles des nids de Cornitermes sp.
et les déblais de fourmis. Ces structures sont caractérisées par les cavités
petites moyennes et grandes et les fissures moyennes.
2. Les parois des nids de termites humivores et le feuilleté des termitières de
Cornitermes sp. Les poroïdes caractéristiques de ces structures sont tous de
très petites tailles (les trois formes)
3. Les turricules frais d’Andiodrilus p. et les placages de termites humivores
associés aux fissures et pores complexes de grande taille.
A cette échelle d’analyse, la microporosité est étudiée. Ces trois groupes de
structure biogénique mise en évidence dans cette typologie se distinguent par leur teneur
en matière organique et en minéral. Le groupe 2 est le plus organique et le groupe 1 le
plus minéral.
Ainsi la porosité d’assemblage des constituants organo-minéraux séparent dans
cette analyse les différentes structures biogéniques.
Toutes les structures biogéniques présentent une porosité totale de l’ordre
de 25%. Les structures biogéniques, présentant une forte mésoporosité, sont peu
microporeuses (déblais de fourmis). A l’opposé les structures les moins
mésoporeuses présentent une microporosité plus développée (feuilleté de termitière
de Cornitermes sp.) Les pores de type ronds (cavités) ou complexes sont les plus
abondants dans toutes les structures biogéniques étudiées.
Au niveau de la mésoporosité, le premier facteur de différenciation des
structures biogéniques est le remaniement des boulettes organo-minérales lors de
la construction de la structure.
Au niveau de la microporosité, le principal facteur de différenciation est la
porosité d’assemblage des constituants organo-minéraux.
VII.3.2.c Diversité des poroïdes et typologie des biostructures
La diversité des poroïdes est estimée par le calcul de l’indice de diversité de
Shannon (Chapitre III.1) appliqué aux différentes classes de poroïdes.
Une première partie de l’étude a été consacrée à l’estimation de cet indice de
diversité pour l’ensemble des structures biogéniques analysées. Puis cette étude a été
renouvelée sur les genres taxonomiques.
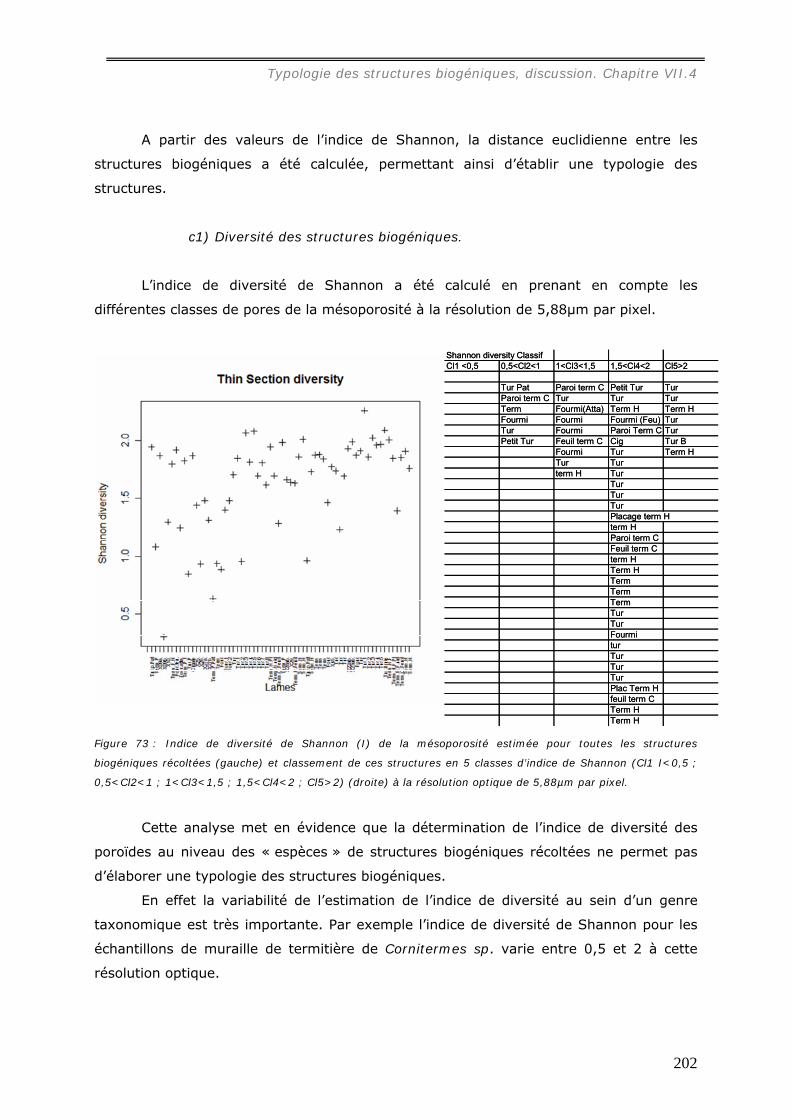
Typologie des structures biogéniques, discussion. Chapitre VII.4
202
A partir des valeurs de l’indice de Shannon, la distance euclidienne entre les
structures biogéniques a été calculée, permettant ainsi d’établir une typologie des
structures.
c1) Diversité des structures biogéniques.
L’indice de diversité de Shannon a été calculé en prenant en compte les
différentes classes de pores de la mésoporosité à la résolution de 5,88µm par pixel.
Shannon diversity ClassifCl1 <0,5 0,5<Cl2<1 1<Cl3<1,5 1,5<Cl4<2 Cl5>2
Tur Pat Paroi term C Petit Tur TurParoi term C Tur Tur TurTerm Fourmi(Atta) Term H Term HFourmi Fourmi Fourmi (Feu) TurTur Fourmi Paroi Term C Tur Petit Tur Feuil term C Cig Tur B
Fourmi Tur Term HTur Turterm H Tur
TurTurTurPlacage term Hterm HParoi term CFeuil term Cterm HTerm HTerm TermTermTurTurFourmiturTurTurTurPlac Term Hfeuil term CTerm HTerm H
Shannon diversity ClassifCl1 <0,5 0,5<Cl2<1 1<Cl3<1,5 1,5<Cl4<2 Cl5>2
Tur Pat Paroi term C Petit Tur TurParoi term C Tur Tur TurTerm Fourmi(Atta) Term H Term HFourmi Fourmi Fourmi (Feu) TurTur Fourmi Paroi Term C Tur Petit Tur Feuil term C Cig Tur B
Fourmi Tur Term HTur Turterm H Tur
TurTurTurPlacage term Hterm HParoi term CFeuil term Cterm HTerm HTerm TermTermTurTurFourmiturTurTurTurPlac Term Hfeuil term CTerm HTerm H
Figure 73 : Indice de diversité de Shannon (I) de la mésoporosité estimée pour toutes les structures
biogéniques récoltées (gauche) et classement de ces structures en 5 classes d’indice de Shannon (Cl1 I<0,5 ;
0,5<Cl2<1 ; 1<Cl3<1,5 ; 1,5<Cl4<2 ; Cl5>2) (droite) à la résolution optique de 5,88µm par pixel.
Cette analyse met en évidence que la détermination de l’indice de diversité des
poroïdes au niveau des « espèces » de structures biogéniques récoltées ne permet pas
d’élaborer une typologie des structures biogéniques.
En effet la variabilité de l’estimation de l’indice de diversité au sein d’un genre
taxonomique est très importante. Par exemple l’indice de diversité de Shannon pour les
échantillons de muraille de termitière de Cornitermes sp. varie entre 0,5 et 2 à cette
résolution optique.
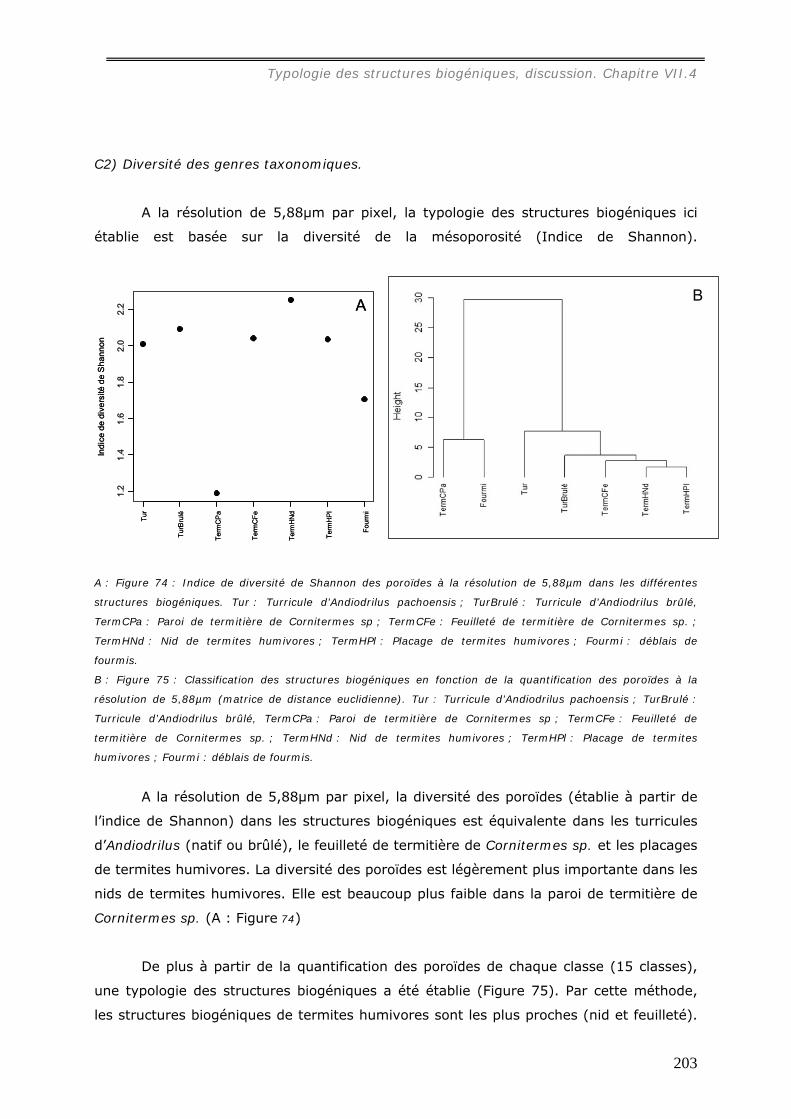
Typologie des structures biogéniques, discussion. Chapitre VII.4
203
C2) Diversité des genres taxonomiques.
A la résolution de 5,88µm par pixel, la typologie des structures biogéniques ici
établie est basée sur la diversité de la mésoporosité (Indice de Shannon).
1 2 3 4 5 6 7
1.2
1.4
1.6
1.8
2.0
2.2
Indi
ce d
e di
vers
ité d
e S
hann
on
Tur
TurB
rulé
Term
CP
a
Term
CFe
Term
HN
d
Term
HP
l
Four
mi
1.2
1.4
1.6
1.8
2.0
2.2 A B
1 2 3 4 5 6 7
1.2
1.4
1.6
1.8
2.0
2.2
Indi
ce d
e di
vers
ité d
e S
hann
on
Tur
TurB
rulé
Term
CP
a
Term
CFe
Term
HN
d
Term
HP
l
Four
mi
1.2
1.4
1.6
1.8
2.0
2.2 A B
A : Figure 74 : Indice de diversité de Shannon des poroïdes à la résolution de 5,88µm dans les différentes
structures biogéniques. Tur : Turricule d’Andiodrilus pachoensis ; TurBrulé : Turricule d’Andiodrilus brûlé,
TermCPa : Paroi de termitière de Cornitermes sp ; TermCFe : Feuilleté de termitière de Cornitermes sp. ;
TermHNd : Nid de termites humivores ; TermHPl : Placage de termites humivores ; Fourmi : déblais de
fourmis.
B : Figure 75 : Classification des structures biogéniques en fonction de la quantification des poroïdes à la
résolution de 5,88µm (matrice de distance euclidienne). Tur : Turricule d’Andiodrilus pachoensis ; TurBrulé :
Turricule d’Andiodrilus brûlé, TermCPa : Paroi de termitière de Cornitermes sp ; TermCFe : Feuilleté de
termitière de Cornitermes sp. ; TermHNd : Nid de termites humivores ; TermHPl : Placage de termites
humivores ; Fourmi : déblais de fourmis.
A la résolution de 5,88µm par pixel, la diversité des poroïdes (établie à partir de
l’indice de Shannon) dans les structures biogéniques est équivalente dans les turricules
d’Andiodrilus (natif ou brûlé), le feuilleté de termitière de Cornitermes sp. et les placages
de termites humivores. La diversité des poroïdes est légèrement plus importante dans les
nids de termites humivores. Elle est beaucoup plus faible dans la paroi de termitière de
Cornitermes sp. (A : Figure 74)
De plus à partir de la quantification des poroïdes de chaque classe (15 classes),
une typologie des structures biogéniques a été établie (Figure 75). Par cette méthode,
les structures biogéniques de termites humivores sont les plus proches (nid et feuilleté).
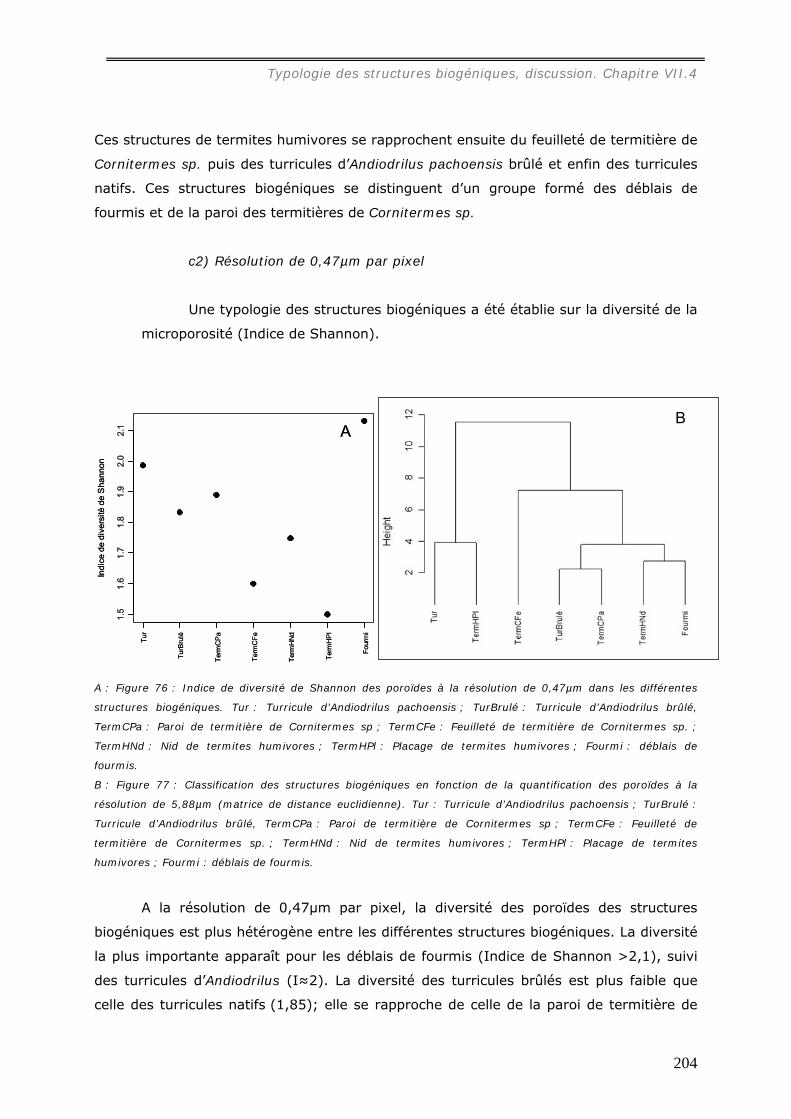
Typologie des structures biogéniques, discussion. Chapitre VII.4
204
Ces structures de termites humivores se rapprochent ensuite du feuilleté de termitière de
Cornitermes sp. puis des turricules d’Andiodrilus pachoensis brûlé et enfin des turricules
natifs. Ces structures biogéniques se distinguent d’un groupe formé des déblais de
fourmis et de la paroi des termitières de Cornitermes sp.
c2) Résolution de 0,47µm par pixel
Une typologie des structures biogéniques a été établie sur la diversité de la
microporosité (Indice de Shannon).
1 2 3 4 5 6 7
1.5
1.6
1.7
1.8
1.9
2.0
2.1
Indi
ce d
e di
vers
ité d
e S
hann
on
Tur
TurB
rulé
Term
CP
a
Term
CFe
Term
HN
d
Term
HP
l
Four
mi
1.5
1.6
1.7
1.8
1.9
2.0
2.1 A
B
1 2 3 4 5 6 7
1.5
1.6
1.7
1.8
1.9
2.0
2.1
Indi
ce d
e di
vers
ité d
e S
hann
on
Tur
TurB
rulé
Term
CP
a
Term
CFe
Term
HN
d
Term
HP
l
Four
mi
1.5
1.6
1.7
1.8
1.9
2.0
2.1 A
B
A : Figure 76 : Indice de diversité de Shannon des poroïdes à la résolution de 0,47µm dans les différentes
structures biogéniques. Tur : Turricule d’Andiodrilus pachoensis ; TurBrulé : Turricule d’Andiodrilus brûlé,
TermCPa : Paroi de termitière de Cornitermes sp ; TermCFe : Feuilleté de termitière de Cornitermes sp. ;
TermHNd : Nid de termites humivores ; TermHPl : Placage de termites humivores ; Fourmi : déblais de
fourmis.
B : Figure 77 : Classification des structures biogéniques en fonction de la quantification des poroïdes à la
résolution de 5,88µm (matrice de distance euclidienne). Tur : Turricule d’Andiodrilus pachoensis ; TurBrulé :
Turricule d’Andiodrilus brûlé, TermCPa : Paroi de termitière de Cornitermes sp ; TermCFe : Feuilleté de
termitière de Cornitermes sp. ; TermHNd : Nid de termites humivores ; TermHPl : Placage de termites
humivores ; Fourmi : déblais de fourmis.
A la résolution de 0,47µm par pixel, la diversité des poroïdes des structures
biogéniques est plus hétérogène entre les différentes structures biogéniques. La diversité
la plus importante apparaît pour les déblais de fourmis (Indice de Shannon >2,1), suivi
des turricules d’Andiodrilus (I≈2). La diversité des turricules brûlés est plus faible que
celle des turricules natifs (1,85); elle se rapproche de celle de la paroi de termitière de

Typologie des structures biogéniques, discussion. Chapitre VII.4
205
Cornitermes sp. qui est légèrement supérieure (1,9). La diversité du feuilleté de
termitière de Cornitermes est plus faible (1,6).
Les structures biogéniques de termites humivores mettent en évidence des
diversités différentes avec le nid dont la diversité d’environ 1,75 est plus importante que
celle du placage (1,5) (A : Figure 76).
A cette résolution, les nids de termites humivores et les déblais de fourmis sont
les plus proches, ainsi que les parois de termitière de Cornitermes et les turricules
d’Andiodrilus brûlés. Ces deux groupes se rapprochent par la suite. Ensuite s’ajoute le
feuilleté de termitière Cornitermes puis un groupe formé des turricules d’Andiodrilus
pachoensis et du placage de termites humivores (Figure 77).
L’étude de la diversité des poroïdes par l’intermédiaire de l’estimation de
l’indice de diversité de Shannon ne permet pas au niveau spécifique d’établir une
typologie des structures biogéniques en raison de la forte variabilité de la porosité
observée pour chaque type de structure biogénique.
Au niveau du genre taxonomique, deux typologies à chacune des résolutions
optiques ont été établies. Ces typologies reflètent les principaux facteurs régulant la
porosité : Le remaniement des boulettes organo-minérales au niveau de la
mésoporosité et l’assemblage organo-minérale au niveau de la microporosité.
Cependant ces typologies sont à considérer avec prudence en raison de la forte
variabilité intra genre observée.

Typologie des structures biogéniques, discussion. Chapitre VII.4
206
VII.4 DISCUSSION
Dans l’objectif de déterminer les meilleurs outils permettant de distinguer les
structures biogéniques et ce afin de modéliser le fonctionnement des sols et d’établir un
indice de qualité des sols basé sur ces biostructures, trois typologies obtenues par
différentes analyses ont été testées : une typologie basée sur la matière organique, une
autre basée sur les caractéristiques chimiques enfin une troisième basée sur l’estimation
de la porosité.
Typologie basée sur l’étude de la matière organique
L’état de la matière organique (fraîche ou dégradée) permet de rapprocher
certaines structures biogéniques :
-Les nids des termites humivores sont isolés et attestent d’une forte labilité du carbone.
-Les turricules d’annélides et les murailles produites par les termites humivores. Ces
dernières sont moins riches en matière organique ; ces deux groupes sont très proches
et leurs taux de carbone et de minéralisation sont quasi-similaires.
-Le sol témoin et les fourmilières présentent un faible taux de carbone.
Les analyses NIRS ont été réalisées sur les turricules d’Andiodrilus pachoensis
frais et brûlés et sur les structures biogéniques du termite Cornitermes sp. (paroi et
feuilleté). Nos résultats confirment que la concentration en matière organique est sans
doute le principal facteur qui sépare les différentes structures biogéniques étudiées.
En effet l’axe 1 du plan factoriel, explique 79% de la variabilité et sépare
clairement un groupe formé des structures biogéniques brûlées (paroi de termitière et
turricules) et les turricules d’Andiodrilus de pâturage d’un autre groupe formé des sols
témoins de pâturage et de forêt, des turricules de forêt natifs et de la paroi de termitière
Cornitermes fraîche. Ceci est confirmé par la projection des longueurs d’onde de l’analyse
NIRS dans le plan factoriel. Toutes les longueurs d’onde sont corrélées selon l’axe 1.
L’axe 2 sépare clairement un groupe formé des structures de termitières (paroi et
feuilleté) dans leur état frais et brûlé et du sol témoin de forêt et de pâturage, d’un autre
groupe formé des turricules d’Andiodrilus pachoensis des deux milieux dans leurs
différents états (brûlés ou non). Cet axe peut être assimilé à la nature de la matière
organique. En effet dans le plan factoriel, les longueurs d’onde se différencient selon
l’axe 2. Un groupe des longueurs d’onde supérieure à 2000nm se différencie des
longueurs d’onde inférieures à 2000nm. A chaque longueur d’onde correspond un type de
liaison carbone et ces liaisons sont responsables de la nature de la matière organique.

Typologie des structures biogéniques, discussion. Chapitre VII.4
207
Ainsi l’analyse NIRS des structures biogéniques permet d’établir une typologie des
structures basée sur la qualité et la quantité de la matière organique.
Typologie basée sur les caractéristiques chimiques
L’analyse multivariée effectuée sur les propriétés chimiques des structures
biogéniques a mis en évidence selon les deux axes une séparation nette des sols
témoins, des structures biogéniques de termites Cornitermes sp. et de celles des
annélides Andiodrilus pachoensis.
Les sols témoins, selon cette analyse, se caractérisent par leur concentration en
carbone et l’aluminium échangeable. Les turricules de pâturage se caractérisent par leur
concentration en potassium, calcium et magnésium. En revanche le pH et le phosphore
assimilable caractérisent les turricules d’Andiodrilus p. brûlés.
Ainsi la minéralisation du carbone et l’acidité d’échange apparaissent dans cette
étude comme étant les deux principaux facteurs permettant d’établir une typologie des
structures biogéniques.
Typologie basée sur l’étude micromorphologique de la porosité
Les typologies des structures biogéniques établies sur la base des résultats
micromorphologiques de la porosité par l’analyse d’image mettent en évidence la
difficulté de caractériser les structures biogéniques par leur porosité.
Cette méthode est généralement utilisée afin de décrire et caractériser la
structure et la porosité des horizons pédologiques (Hallaire and Cointepas, 1993 ;
Hallaire, 1994 ; Hallaire and Curmi, 1994 ; Pérès et al., 1998 ; Jégou et al., 2000).
Les typologies obtenues aux deux résolutions étudiées n’aboutissent pas à la
même organisation des structures biogéniques en raison de la différence des types de
pores rencontrés aux différentes échelles. Aux deux échelles, la variabilité des résultats
au sein d’une espèce est également très forte.
A la plus faible résolution (5,88µm), le principal type de pores est le pore
complexe (porosité de remaniement mécanique) alors qu’à la résolution la plus fine
(0,47µm), le pore caractéristique est le pore rond ou tubulaire (porosité d’assemblage).
Aux deux résolutions les fissures sont peu abondantes en raison de leur origine
fréquemment physique voire mécanique. Or notre étude a déjà mise en évidence la forte
stabilité structurale des structures biogéniques étudiées. Les pores ronds, tubulaires et
complexes correspondent davantage aux pores d’origine biologique.

Typologie des structures biogéniques, discussion. Chapitre VII.4
208
A la résolution 5,88µm, les déblais de fourmis et la paroi de termitière
Cornitermes sont les plus poreux et leur profil poral est comparable en raison sans doute
du mode de fabrication des agrégats de ces structures biogéniques (boulettes buccales).
Ces structures sont des agrégats minéraux additionnés de mucus et façonnés par les
mandibules. Les nids des termites humivores sont également très proches avec une
porosité assez faible (8%). Ils se rapprochent du feuilleté des termitières de Cornitermes.
Ce résultat confirme les observations de terrain et les observations micromorphologiques
mettant en évidence une grande similarité morphologique entre ces structures. Ce
résultat est par ailleurs confirmé par l’existence d’un ancêtre commun du genre
Syntermes (Grassé, 1984c) entre le genre Cornitermes sp. et les termites humivores.
Les turricules d’Andiodrilus pachoensis brûlés présentent ensuite une porosité plus
proche du précédent groupe que des turricules frais. Le feu réduit la porosité en opérant
un réarrangement granulométrique de la biostructure par pectisation des argiles
aboutissant à la formation de pseudosables. Certains pores sont comblés diminuant ainsi
la porosité totale de la structure. L’organisation des différentes structures biologiques ne
se retrouve pas dans l’arbre obtenu à la résolution plus fine même si les arbres des deux
résolutions mettent en évidence des distances comparables entre certaines structures
biogéniques comme par exemple la distance entre la paroi de termitière Cornitermes et
les déblais de fourmis.
L’organisation obtenue à cette seconde résolution trouve plus difficilement
d’explications d’ordre biologique. A cette résolution, l’analyse accède à la microstructure
des structures biogéniques c'est-à-dire à l’organisation des particules. Il est par
conséquent probable que l’origine du matériau permette d’expliquer en partie
l’organisation des biostructures obtenues. La distribution en taille et types de pores ne
permet pas de différencier nettement les structures biogéniques, seuls les turricules
d’Andiodrilus brûlés et la paroi de termitière Cornitermes ne présentent pas de pores
complexes.
Ainsi l’étude réalisée à la première résolution optique apporte de meilleurs
éléments de compréhension pour établir une typologie des structures biogéniques en
fonction de la porosité de leurs agrégats.
Les résultats obtenus mettent en évidence une très forte variabilité de la porosité
interspécifique. Seul, le feuilleté de termites Cornitermes présente une variabilité plus
faible sans doute en raison du matériau d’origine proctodéale qui forme la structure
interne du feuilleté.
La forte variabilité des résultats met en évidence la difficulté d’utiliser la
nomenclature des « genres » taxonomiques afin d’établir une typologie des structures

Typologie des structures biogéniques, discussion. Chapitre VII.4
209
Une étude au niveau des groupes fonctionnels et des espèces clés apparaît par
conséquent plus pertinente.
L’étude de l’ensemble des lames aux deux résolutions montrent que les différents
types de structures biogéniques présentent une quantité et une diversité des poroïdes
variables au sein des groupes de faune considérés en raison sans doute de la variabilité
inter genre mais aussi des variabilités locales des paramètres environnementaux (par
exemple le sol). Il est donc probable que les paramètres tels la porosité ou le taux de
minéralisation soient influencés par le milieu au point qu’une évolution des propriétés des
structures biogéniques lors du changement de milieu soit envisageable. D’autres
analyses ont mis en évidence des co-variabilités des aspects morphologiques des
agrégats et de la faune du sol dans une expérimentation sur le couvert végétal dans les
pâturages brésiliens (Velasquez et al., 2005). Une telle analyse confirme cette hypothèse
de coévolution des structures biogéniques et des écosystèmes.
Conclusions
Notre étude a ainsi révélé trois facteurs importants agissant sur la différenciation des
structures biogéniques :
1. L’état de dégradation et la nature de la matière organique ainsi que ses liens avec
la partie minérale.
2. L’acidité notamment l’acidité d’échange et par conséquent la concentration en
cations basiques.
3. Le remaniement des boulettes organo-minérales d’origine buccale ou métabolique
lors de la fabrication des structures biogéniques.
Ces facteurs constituent des processus fondamentaux dans le fonctionnement du sol.
Il s’agit de processus essentiellement chimiques mais aussi mécaniques qui interviennent
à différentes échelles de temps et d’espace rendant difficile l’estimation de leur impact
dans le fonctionnement global du sol.

Discussion générale. Chapitre VIII
210
VIII. Discussion générale et conclusion

Discussion générale. Chapitre VIII
211
Les organismes du sol et principalement les invertébrés ingénieurs (annélides,
termites) jouent un rôle essentiel dans l’évolution et la structuration des horizons de
surface des sols par l’intermédiaire des processus écosytémiques qu’ils y assurent. Ces
organismes ingénieurs produisent des structures biogéniques (turricules, termitières,) qui
affectent fortement les processus du sol notamment au niveau de la circulation de l’eau
des gaz, et de la matière organique modifiant ainsi les propriétés hydrodynamiques et
par conséquent influent sur les relations sols-plantes-organismes (Grimaldi, 2000).
Aujourd’hui, dans un but prédictif afin de mieux gérer les ressources des
écosystèmes, des études cherchent à modéliser leur fonctionnement et plus
particulièrement celui du sol (Lavelle, 1996, 1997 ; Brussaard, 1998 ; Ekschmitt and
Griffiths, 1998 ; Loreau, 1998 ; Herbert et al., 2003 ; Rossi, 2003).
L’objectif de notre étude était de mettre en évidence l’impact de deux espèces
ingénieurs : le termite xylophage Cornitermes sp. affinis ovatus et l’annélide anécique
Andiodrilus pachoensis, sur les sols.
En Amazonie brésilienne, dans la région de Marabá, la mutation du milieu de la forêt
en pâturages par l’agriculture familiale modifie profondément les propriétés physiques,
chimiques et biologiques des sols argileux. Alors que la fertilité chimique des latosols
s’améliore dans les pâturages soumis à la culture sur brûlis (augmentation du pH et des
concentrations en cations échangeables), une compaction du sol, d’intensité et d’étendue
variable d’un pâturage à l’autre, s’observe. La compaction de ces sols s’explique par les
pratiques agricoles (culture sur brûlis, déboisement mécanique et surtout surpâturage).
En raison de la distribution spatiale clairsemée des touffes de graminées Brachiaria
brisantha, les sols des pâturages apparaissent également plus sensibles à l’érosion dans
l’espace inter-touffes que ceux sous forêts et par conséquent sont sujets à une
dégradation physique plus rapide suivant le degré de la pente.
Les résultats de notre travail permettent de mettre en évidence de multiples
interactions entre les structures biogéniques produites par le termite Cornitermes sp.
(termitières), l’annélide Andiodrilus pachoensis. (turricules), le couvert végétal (forêt ou
pâturage) et le type de sol (latosol, cambisol ou gleysol) (figure 69).

Discussion générale. Chapitre VIII
212
Figure 78 : Schéma des relations entre la couverture végétale, les deux structures biogéniques étudiées (Termitières de Cornitermes sp. et turricules d’Andiodrilus pachoensis) et l’horizon superficiel 0-30 cm des sols du site d’étude.
L’évaluation de la fertilité ou aptitude culturale d’un milieu se fonde sur une
appréciation de son potentiel agricole défini à partir des caractéristiques topographiques,
climatiques et la nature des sols (Pieri, 1989). Elle conduit à établir des « normes »
théoriques d’un maintien ou d’une amélioration de la fertilité du milieu en vue de
l’optimisation de son potentiel cultural.
Concernant les pâturages, le maintien de la fertilité du milieu conditionne le maintien
du couvert végétal herbacé et par voie de conséquence, dans les milieux pentus,
empêche la battance et stabilise la structure.
Ainsi l’action de ces deux espèces ingénieurs du sol porte sur différents paramètres
qui gèrent la fertilité du sol (matière organique, cations basiques, argile).
L’étude biologique du termite Cornitermes sp. a mis en évidence que les termitières
sont absentes des forêts, ce qui signifie que cette espèce de termites est inféodée aux
milieux ouverts. Elle peut être considérée comme indicatrice des fronts de déforestation.
Les densités de termitières observées (23,5 nids par hectare) ne permettent pas de
parler de pullulation mais il faut rappeler que les pâturages étudiés ne sont âgés que de

Discussion générale. Chapitre VIII
213
5 à 6 ans. Le termite Cornitermes est, d’après la courbe de régression, dans une phase
d’expansion.
Les études des distributions spatiales des nids soulignent que le pourcentage d’argile
et le taux d’humidité des sols sont les facteurs majeurs de régulation du développement
des colonies.
L’étude des caractéristiques physico-chimiques des murailles et du feuilleté interne
des termitières a révélé que le rôle de Cornitermes sp. sur le fonctionnement du sol était
triple :
-1- un rôle mécanique local par la remontée de matériaux argileux chargés de cations
primaires (géophagie) pour la construction de la muraille (qui recouvrira les horizons de
surface à la mort de la termitière) et l’aération locale provenant du réseau de galeries
creusées dans le termitosphère. Il s’agit là d’un impact qui peut être jugé positif sur les
paramètres de la fertilité annuelle du milieu.
-2- un rôle soustractif de la matière organique, prélevée dans les horizons de surface
(bois et racines mortes). La matière organique ingurgitée, broyée, transformée et
humifiée par l’arsenal enzymatique du tube digestif des termites est séquestrée sous
forme de déjections et constitue la structure du feuilleté interne au nid. Elle ne sera
rendue à l’écosystème qu’à la mort de la colonie. Cet impact est relativement négatif sur
le fonctionnement annuel du sol.
-3- un ensemencement bactérien et fongique. A la mort du nid, la matière organique
humifiée sera rapidement reprise par les microorganismes.
Considérant le temps de vie d’une colonie, le turnover de la matière organique
orchestré par le termite Cornitermes et rendu à l’écosystème s’effectue donc sur une
durée d’au moins 20 ans.
L’étude de l’annélide anécique Andiodrilus pachoensis révèle que ce ver s’adapte aux
pâturages suite à la déforestation. Il n’est donc pas indicateur des fronts de
déforestation. Si dans les forêts la durée de vie des turricules (accumulation de
déjections) qu’il produit est supérieure à l’année, une stimulation de leur production est
observée dans les milieux découverts. La différence observée entre densité et production
de turricules dans les pâturages ne peut s’expliquer que par une forte dégradation dans
un délai relativement court. Cette forte dégradation est sans aucun doute causée par le
piétinement bovin qui favorise l’intégration d’agglomérats issus des turricules dans les
horizons superficiels des pâturages. La pluie et la calcination (culture sur brûlis)
favorisent ensuite la dissolution des agglomérats et leur intégration le long de la pente.

Discussion générale. Chapitre VIII
214
Les caractéristiques physico-chimiques des turricules permettent d’identifier trois
fonctions majeures de cet annélide sur le sol :
-1- une forte action mécanique (aération par réalisation d’un important réseau de
galeries verticales et brassage des particules du sol remontées en surface sous forme de
déjections
-2- une fonction de dégradation / humification de la matière organique dans le tube
digestif, réexcrétée à la surface du sol.
-3- une activation des microorganismes le long des galeries et enfouissement dans
les horizons profonds.(Lavelle, 1997)
Le temps de vie estimée des turricules (de plus ou moins un an en fonction du milieu)
met en évidence que le turnover de la matière organique réalisé par cet annélide
anécique est rapide et s’effectue à une échelle annuelle dans les milieux ouverts.
La matière organique présente dans le feuilleté des termitières et celle contenue
dans les turricules d’annélides anéciques sont des produits du métabolisme. Ces deux
structures sont composées de déjections qui renferment un taux relativement élevé de
matière organique dégradée. Cette dernière est intimement liée à l’argile ingurgitée
concomitamment (la matière minérale représente 23% du feuilleté et 88% des
turricules). En effet dans le tube digestif de nos deux espèces d’ingénieur (le termite
ouvrier et l’annélide anécique) une succession de décompositions et recombinaisons de la
matière organique s’opère. La matière organique ingurgitée est humifiée sous l’action
mécanique et enzymatique du conduit digestif des termites et des annélides et de leurs
microflores associées. Les transformations aboutissent à la formation de différentes
fractions humifiées et à l’apparition de microagrégats stables d’origine biologique. Nous
pouvons donc supposer que les complexes argilo-humiques ainsi formés dans le tube
digestif de nos deux espèces sont stables et difficilement minéralisables comme l’ont
montré d’autres auteurs. (Garnier-Sillam et al., 1985 ; Barois et al., 1987 ; Garnier-
Sillam, 1987 ; Shipitalo and Protz, 1989 ; Golchin et al., 1994 ; Guggenberger et al.,
1996).
Les deux espèces étudiées présentent donc une action géophagique importante et
une action forte sur la dégradation de la matière organique des litières (ver) et du bois
(termite).
Cependant ces deux structures biogéniques sont de tailles différentes (de l’ordre du
m² pour la termitière et 10cm² pour les turricules) et leur pérennité est différente (20
ans pour la termitière, moins d’un an pour les turricules des pâturages).

Discussion générale. Chapitre VIII
215
Leur impact sur le sol ne s’effectue donc pas aux mêmes échelles spatiales et
temporelles rendant difficile la comparaison des deux structures biogéniques.
Si l’action fertilisante du termite est locale (au niveau de la termitière), celle de
l’annélide s’effectue plus globalement au niveau de la parcelle.
Il assure donc une forme de fertilisation organo-minérale des horizons superficiels.
En raison de ces notions d’échelle, dans ce contexte de déforestation amazonienne, le
rôle du termite Cornitermes sp. affinis ovatus apparaît plus négatif que celui de l’annélide
Andiodrilus pachoensis.
De plus, étant donné que la présence de termitières de Cornitermes parait inhiber la
production des turricules d’Andiodrilus pachoensis, et que la densité des termitières s’est
accentuée sur les 6 années de pâturage, nous pouvons émettre l’hypothèse que ces deux
espèces sont en compétition.
Une prochaine étape dans la dégradation des pâturages amazoniens serait la
disparition de l’annélide anécique et son remplacement au fil du temps par le termite
Cornitermes sp.
Bien que n’agissant pas aux mêmes échelles, les deux structures biogéniques
étudiées sont soumises aux effets des pratiques agricoles (culture sur brûlis, destruction
par le feu). Le principal effet de la calcination des structures biogéniques est une
augmentation de leur stabilité structurale. Sous l’effet des fortes températures, une
pectisation (déshydratation réversible) des particules argileuses s’opèrent. Il en résulte
des agrégats très cohérents qui ne se dispersent pas avec les méthodes classiques, d’où
une augmentation de la fraction des sables grossiers (pseudosables) et une baisse du
taux d’argile granulométrique. Ce processus chimique s’accompagne d’une formation
d’oxyde de fer (hématites) conférant aux structures biogéniques une couleur rougeâtre
en surface lorsqu’elles sont suffisamment longtemps exposées au feu. Les principaux
inconvénients, sur le plan agronomique, de cette pratique est la perte de matière
organique et l’augmentation de stabilité des murailles des termitières et des turricules.
Les avantages sont l’élévation du pH et la libération des sels minéraux (calcium,
magnésium, potassium) sous forme de cendres.
La culture sur brûlis n’affecte pas la partie interne des termitières (en raison du rôle
de la muraille). Le feu n’atteint pas le feuilleté. Cependant la pratique de l’incinération
des nids de termite provoque la destruction du feuilleté et la totalité de la matière
organique est alors minéralisée. Cette pratique n’est donc pas favorable à la conservation
des agrégats argilo-humiques des pâturages.

Discussion générale. Chapitre VIII
216
Ces termites sont affiliées aux Nasutitermes (famille des termiditae) dont le nid
arboricole est uniquement construit en feuilleté. Chez les Cornitermes, la muraille
protège le feuilleté situé au centre de la termitière.
La fabrication de cette muraille peut, peut-être, être assimilée à un stade
intermédiaire d’évolution de cette espèce. Stade plus évolué que celui d’un genre de
termite qui ne construirait pas de muraille mais peut–être moins évolué que celui des
Nasutitermes dont le nid est arboricole. Quoi qu’il en soit, cette espèce inféodée au sol, a
su mettre en place une mesure de défense de la structure fragile (le feuilleté) de son nid.
Ces résultats confirment que les facteurs écologiques sont organisés
hiérarchiquement : nous avons montré que la déforestation, le type de sol, les pratiques
culturales agissent sur les peuplements des deux invertébrés étudiés bouleversant ainsi
les processus écologiques et biologiques qu’ils conditionnent. Le modèle hiérarchique
reflète cette organisation (Lavelle et al., 1993). L’activité de la faune du sol est
déterminée par une suite de facteurs qui agissent largement et la contraignent. Ces
facteurs opèrent à des échelles spatiales variables emboîtées (exemple de la couverture
végétale ou du type de sol) mais aussi à des échelles de temps propre. La réalisation de
ces processus et la production des structures biogéniques s’opèrent à des échelles de
temps très différentes. Les différentes échelles de temps n’étant pas obligatoirement
corrélées dans l’espace, il est ainsi difficile de déterminer précisément les processus
écologiques qui influent sur la production de chacune des structures biogéniques. Ce qui
explique également que la coupe des touffes de Brachiaria brisantha dans les pâturages
n’altère pas significativement la production de turricules. La privation du couvert végétal
pendant une période très réduite ne permet pas de changer suffisamment les conditions
de la surface du sol pour altérer la densité des vers ou leur activité.
Le problème des échelles de perception et de représentation du fonctionnement des
écosystèmes est à l’origine de nombreuses controverses concernant les relations entre la
biodiversité et le fonctionnement des écosystèmes. Aujourd’hui, la théorie acceptée
repose sur le fait que le fonctionnement de l'écosystème serait corrélé avec la diversité
spécifique des organismes à grande échelle, alors que localement le fonctionnement de
l'écosystème serait plutôt lié aux traits des espèces et donc à leur activité spécifique.
Cette théorie de la hiérarchie (Loreau et al., 2001 ; Bond and Chase, 2002) permet de
définir la notion de « groupe fonctionnel ». Les espèces ayant donc des fonctions
écosytémiques équivalentes sont regroupées au sein d’un même groupe fonctionnel. Ces
processus écosytémiques sont réalisés dans les forêts par des organismes qui jouent
partiellement le même rôle dans ces processus écologiques mais ayant des traits

Discussion générale. Chapitre VIII
217
biologiques différents (morphologique, comportementaux). C'est-à-dire à des espèces
d’un même "groupe fonctionnel".
Ainsi les deux espèces ingénieurs étudiées pourraient être classées dans le même
groupe fonctionnel
Les espèces étudiées ont été choisies en se basant sur l’abondance apparente de
leurs structures biogéniques. Ces espèces interviennent de manière prépondérante dans
la bioturbation de l’horizon 0-30cm et nous pouvons affirmer que les horizons superficiels
des sols sont majoritairement constitués des agrégats issus des turricules de cet
annélide.
Les espèces clés dans le fonctionnement du sol sont des espèces dont la présence est
cruciale pour le maintien et l’organisation de leur communauté écologique (Paine, 1969 ;
Scott Mills et al., 1993).
Ainsi, le termite Cornitermes sp. et l’annélide Andiodrilus P. peuvent constituer des
espèces clés pour le fonctionnement du sol mais leurs interactions avec les autres
espèces de la faune du sol doivent être étudiées, en effet les interactions entre les
espèces provenant du changement de diversité spécifique suite au changement de milieu
peuvent influer fortement les processus du sol (Cardinale et al., 2002).
Certain auteurs ont montré que les vers anéciques étaient capables de transformer
rapidement les différents constituants foliaires et racinaires, y compris les plus résistants
comme les pigments bruns (Toutain, 1981 ; Edwards, 2004).
Les termites xylophages sont des organismes extrêmement efficaces vis-à-vis de la
dégradation des composés phénoliques d’origine végétale en particulier les lignines.
Ainsi, les deux ingénieurs étudiés, dans ce milieu, permettent de lever en quelque
sorte le blocage d’ordre biochimique de la dégradation des végétaux (tannage des
protéines des tissus végétaux au moment de leur sénescence ainsi que les noyaux
aromatiques de la lignine). Ils permettent, ainsi, aux autres organismes d’intervenir
ensuite dans la chaîne trophique continue aboutissant à la biodégradation totale des
résidus végétaux.
Notre étude nous a amenés à tenter d’établir une typologie des structures
biogéniques provenant de différentes espèces et groupes taxonomiques en fonction des
différentes propriétés physiques et chimiques de celles-ci. L’analyse de la matière
organique par la méthode NIRS et les mesures de respirométrie (Pelosi, 2005) ont
permis de séparer les différentes structures biogéniques. L’analyse NIRS, dans laquelle
les structures biogéniques brûlées ont été prises en compte, permet de vérifier que le

Discussion générale. Chapitre VIII
218
premier critère de distinction des différentes structures biogéniques est l’état de la
matière organique (fraîche ou dégradée). L’analyse multivariée des caractéristiques
chimiques a permis d’établir une autre typologie distinguant clairement les turricules
d’Andiodrilus pachoensis des parois de termitière de Cornitermes sp. Deux autres
typologies des structures ont été établies par quantification de la porosité par analyse
d’image de lames minces à deux résolutions. La typologie établie à la résolution la plus
large (5,88µm) est apparue comme celle se rapprochant le plus des observations
morphologiques de terrain et des observations micromorphologiques. Cependant la forte
variabilité de la mesure de porosité au sein d’un genre taxonomique met en évidence la
complexité d’utiliser les genres taxonomiques afin d’établir une typologie des structures
et ceci en raison de la forte variabilité des comportements de chaque espèce au sein d’un
« genre » taxonomique.
De ces résultats, il apparaît qu’il est probablement plus important d’étudier les liens
entre les espèces clés, c'est-à-dire de travailler au niveau des groupes fonctionnels qu’au
niveau de la diversité des espèces ou d’un « genre » particulier.
Ainsi, il existe sans doute des relations mécanistiques entre la fonction d’une espèce
dans un écosystème et les fonctions de l’écosystème (Bengtsson, 1998) mais seule
l’accumulation de données spécifiques à chaque espèce permettra d’identifier plus
précisément les « espèces clés » des groupes fonctionnels.
D’autres études ont également mis en évidence le rôle important des
microorganismes présents dans les structures biogéniques (Lavelle, 1997). L’activité
enzymatique peut également servir à établir une typologie des structures biogéniques
(Mora et al., 2005). Ces mesures permettent de classer les structures biogéniques en
séparant clairement des turricules, les placages de termites (Mora et al., 2003).
Des expériences en microcosmes ont permis de mettre en évidence le rôle des
structures biogéniques sur la microflore symbiotique et les conséquences pour les plantes
en zone soudano-sahélienne (Ndiaye, 2003). Une étude de la microflore permettrait de
mettre en évidence le rôle des bactéries et champignons dans la dégradation des
structures biogéniques. L’étude de la microflore associée aux structures biogéniques et
qui devait compléter ce travail, n’a pas pu être effectuée du fait de l’interdiction durant
plusieurs années du gouvernement brésilien de permettre l’exportation de structures
biogéniques non irradiées sur le territoire français.
Notre étude constitue un apport dans l’étude de l’origine et de la dynamique de
l’agrégation dans le sol. L’objectif de ce travail est de développer notamment des outils
méthodologiques visant à séparer les agrégats du sol et établir leur origine afin de

Discussion générale. Chapitre VIII
219
modéliser la dynamique de ces agrégats en vue d’établir un indice de qualité des sols. La
mise au point de tel indice correspond à la nécessité de mettre au point des méthodes de
diagnostic environnemental. La compréhension des processus biologiques et
l’identification des sources de stress des écosystèmes permettront de définir et
promouvoir des plans de gestion durable des écosystèmes naturels.
Notre travail a permis de caractériser particulièrement deux structures biogéniques :
les turricules d’Andiodrilus pachoensis et les termitières de Cornitermes sp. et d’apporter
des éléments à la constitution d’une typologie des structures biogéniques à partir de
leurs propriétés physiques et chimiques. Cette étape a pour objectif de caractériser le
rôle fonctionnel de ces deux espèces dans le fonctionnement du sol.
A terme cela pourrait permettre, en réalisant ce travail sur les « espèces clés » des
groupes fonctionnels, de préparer des recommandations afin de conserver voire de
reconstituer les milieux dans une optique de développement durable.
Notre étude souligne de nouveau que le changement de milieu affecte un grand
nombre de paramètres, en particulier l’activité de la macrofaune, qui modifie directement
et indirectement le fonctionnement du sol.

Annexes
220
Annexes

Annexes
221
Annexe 1
Latossolo
Cambissolo
Gleissolo Gleysol
Cambisol
Latosol
Latossolo
Cambissolo
Gleissolo Gleysol
Cambisol
Latosol
Organisation de la couverture pédologique sur monzogranite (Benfica, Pará)
(Grimaldi M., non publié)
(a) Horizon supérieur de couleur 10YR5/6 (brun jaune), texture argileuse (montante) à
moyennement argileuse (affleurant), avec de la matière organique, compacte;
(b) Horizon de couleur 10YR5/6 (brun jaune), texture argileuse, friable;
(c) Horizon de couleur 7,5YR5/8 (brun), texture argileuse à très argileuse, friable;
(d) Horizon de couleur 7,5YR5/8 (brun), texture argileuse, nodules centimétriques
espars, friable;
(e) Horizon nodulaire de couleur 7,5YR5/8 (brun), texture argileuse, friable;
(f) Horizon de couleur 10YR5/8 (brun jaune), texture argileuse, avec nodules
centimétriques épars, friable;
(g) Horizon d’altération de couleur 5YR5/8 à 5YR5/6 (rouge jaune), texture argileuse
consistance ferme, présence de nodules friables;
(h) Horizon d’altération de couleur 5YR5/8 (rouge jaune), texture argileuse à argilo
limoneuse, avec tâches blanches et jaunes;
(i) Horizon de couleur 7,5YR5/8 (brun) à 7,5YR6/6 (jaune rougeâtre) avec tâches
blanches et jaunes clairs e noirs, texture argilo limoneuse;

Annexes
222
(j) Horizon de couleur 7,5YR5/6 (brun) à 10YR6/8 (jaune brun), avec tâches rougeâtres
et jaune foncé, texture argileuse;
(k) Horizon superficiel de couleur 10YR6/8 (brun jaune) taches de couleur 7,5YR5/6
(brun), texture moyennement argileuse;
(l) Horizon superficiel 10YR3/2 (brun cendre très obscur), texture moyennement
argileuse;
(m) Horizon 2,5Y4/3 (brun olive) com variegados vermelhos, texture argileuse, humide;
(n) Horizon Gley de couleur 2,5Y6/4 (brun jaune clair) à 2,5Y7/2 (gris clair) avec
agrégats (variegado) de couleur grise, blanches, rouge et jaune, texture argileuse,
très humide.
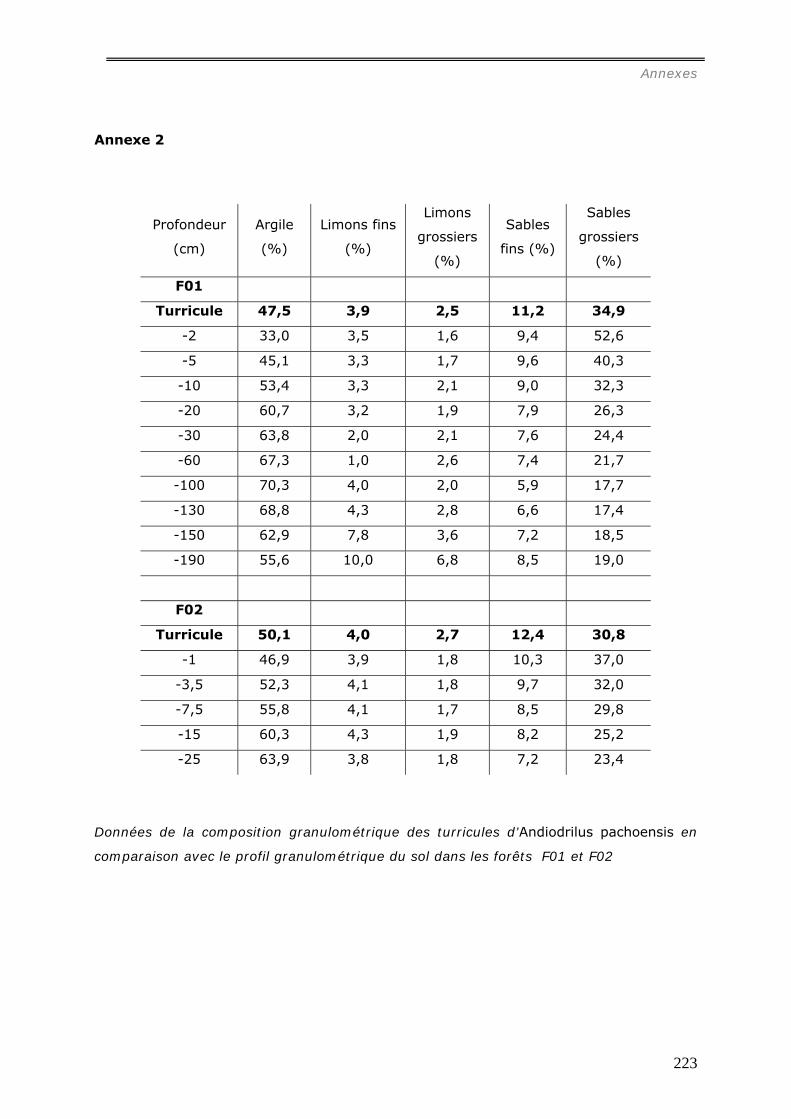
Annexes
223
Annexe 2
Profondeur
(cm)
Argile
(%)
Limons fins
(%)
Limons
grossiers
(%)
Sables
fins (%)
Sables
grossiers
(%)
F01
Turricule 47,5 3,9 2,5 11,2 34,9
-2 33,0 3,5 1,6 9,4 52,6
-5 45,1 3,3 1,7 9,6 40,3
-10 53,4 3,3 2,1 9,0 32,3
-20 60,7 3,2 1,9 7,9 26,3
-30 63,8 2,0 2,1 7,6 24,4
-60 67,3 1,0 2,6 7,4 21,7
-100 70,3 4,0 2,0 5,9 17,7
-130 68,8 4,3 2,8 6,6 17,4
-150 62,9 7,8 3,6 7,2 18,5
-190 55,6 10,0 6,8 8,5 19,0
F02
Turricule 50,1 4,0 2,7 12,4 30,8
-1 46,9 3,9 1,8 10,3 37,0
-3,5 52,3 4,1 1,8 9,7 32,0
-7,5 55,8 4,1 1,7 8,5 29,8
-15 60,3 4,3 1,9 8,2 25,2
-25 63,9 3,8 1,8 7,2 23,4
Données de la composition granulométrique des turricules d’Andiodrilus pachoensis en
comparaison avec le profil granulométrique du sol dans les forêts F01 et F02
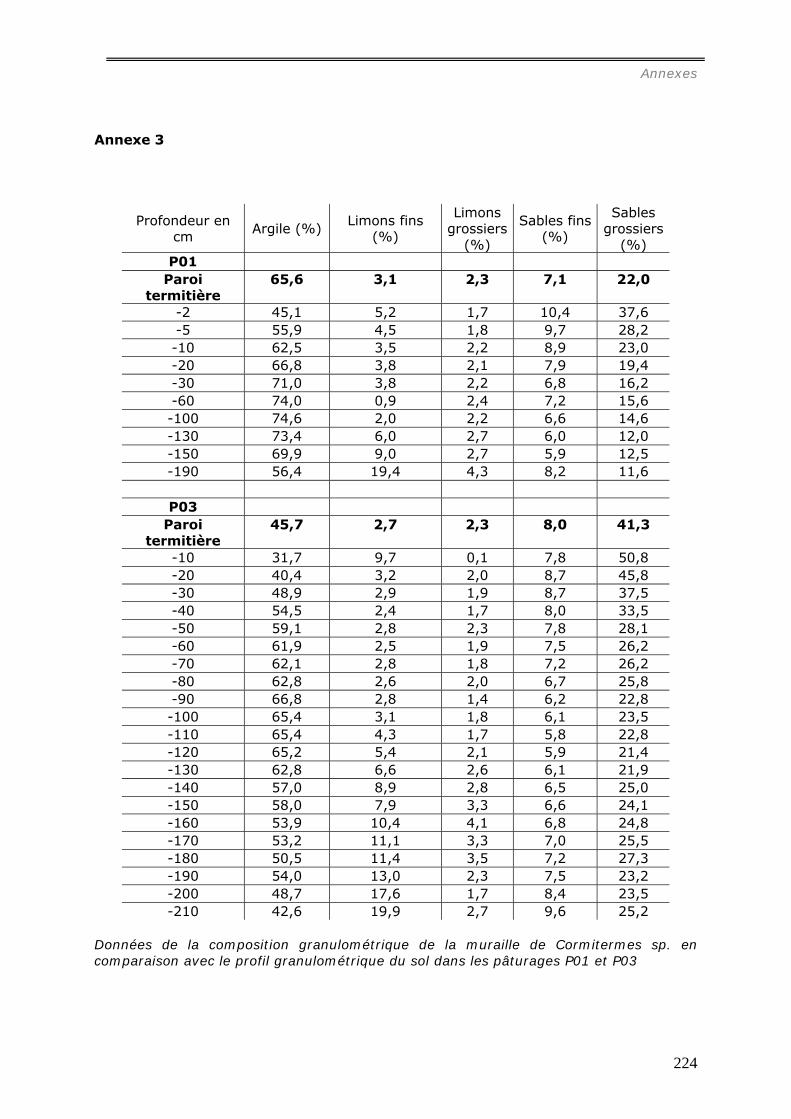
Annexes
224
Annexe 3
Profondeur en cm
Argile (%) Limons fins
(%)
Limons grossiers
(%)
Sables fins (%)
Sables grossiers
(%) P01
Paroi termitière
65,6 3,1 2,3 7,1 22,0
-2 45,1 5,2 1,7 10,4 37,6 -5 55,9 4,5 1,8 9,7 28,2 -10 62,5 3,5 2,2 8,9 23,0 -20 66,8 3,8 2,1 7,9 19,4 -30 71,0 3,8 2,2 6,8 16,2 -60 74,0 0,9 2,4 7,2 15,6 -100 74,6 2,0 2,2 6,6 14,6 -130 73,4 6,0 2,7 6,0 12,0 -150 69,9 9,0 2,7 5,9 12,5 -190 56,4 19,4 4,3 8,2 11,6
P03
Paroi termitière
45,7 2,7 2,3 8,0 41,3
-10 31,7 9,7 0,1 7,8 50,8 -20 40,4 3,2 2,0 8,7 45,8 -30 48,9 2,9 1,9 8,7 37,5 -40 54,5 2,4 1,7 8,0 33,5 -50 59,1 2,8 2,3 7,8 28,1 -60 61,9 2,5 1,9 7,5 26,2 -70 62,1 2,8 1,8 7,2 26,2 -80 62,8 2,6 2,0 6,7 25,8 -90 66,8 2,8 1,4 6,2 22,8 -100 65,4 3,1 1,8 6,1 23,5 -110 65,4 4,3 1,7 5,8 22,8 -120 65,2 5,4 2,1 5,9 21,4 -130 62,8 6,6 2,6 6,1 21,9 -140 57,0 8,9 2,8 6,5 25,0 -150 58,0 7,9 3,3 6,6 24,1 -160 53,9 10,4 4,1 6,8 24,8 -170 53,2 11,1 3,3 7,0 25,5 -180 50,5 11,4 3,5 7,2 27,3 -190 54,0 13,0 2,3 7,5 23,2 -200 48,7 17,6 1,7 8,4 23,5 -210 42,6 19,9 2,7 9,6 25,2
Données de la composition granulométrique de la muraille de Cormitermes sp. en comparaison avec le profil granulométrique du sol dans les pâturages P01 et P03
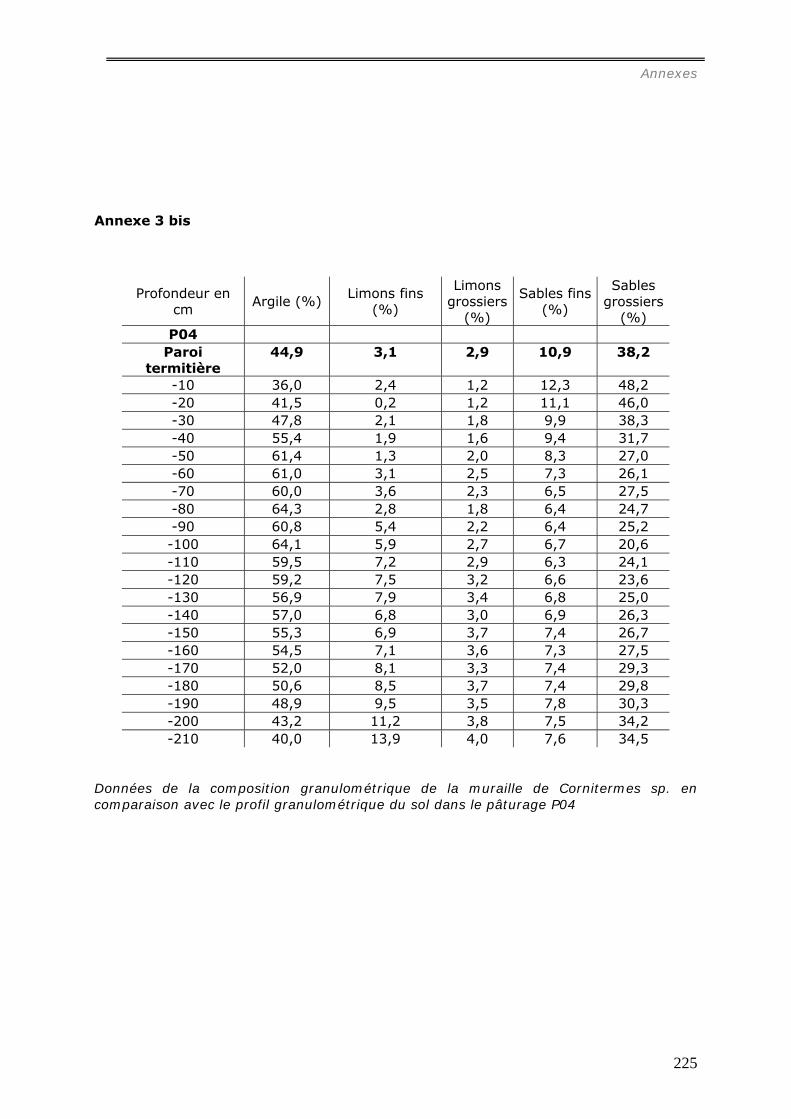
Annexes
225
Annexe 3 bis
Profondeur en cm
Argile (%) Limons fins
(%)
Limons grossiers
(%)
Sables fins (%)
Sables grossiers
(%) P04
Paroi termitière
44,9 3,1 2,9 10,9 38,2
-10 36,0 2,4 1,2 12,3 48,2 -20 41,5 0,2 1,2 11,1 46,0 -30 47,8 2,1 1,8 9,9 38,3 -40 55,4 1,9 1,6 9,4 31,7 -50 61,4 1,3 2,0 8,3 27,0 -60 61,0 3,1 2,5 7,3 26,1 -70 60,0 3,6 2,3 6,5 27,5 -80 64,3 2,8 1,8 6,4 24,7 -90 60,8 5,4 2,2 6,4 25,2 -100 64,1 5,9 2,7 6,7 20,6 -110 59,5 7,2 2,9 6,3 24,1 -120 59,2 7,5 3,2 6,6 23,6 -130 56,9 7,9 3,4 6,8 25,0 -140 57,0 6,8 3,0 6,9 26,3 -150 55,3 6,9 3,7 7,4 26,7 -160 54,5 7,1 3,6 7,3 27,5 -170 52,0 8,1 3,3 7,4 29,3 -180 50,6 8,5 3,7 7,4 29,8 -190 48,9 9,5 3,5 7,8 30,3 -200 43,2 11,2 3,8 7,5 34,2 -210 40,0 13,9 4,0 7,6 34,5
Données de la composition granulométrique de la muraille de Cornitermes sp. en comparaison avec le profil granulométrique du sol dans le pâturage P04
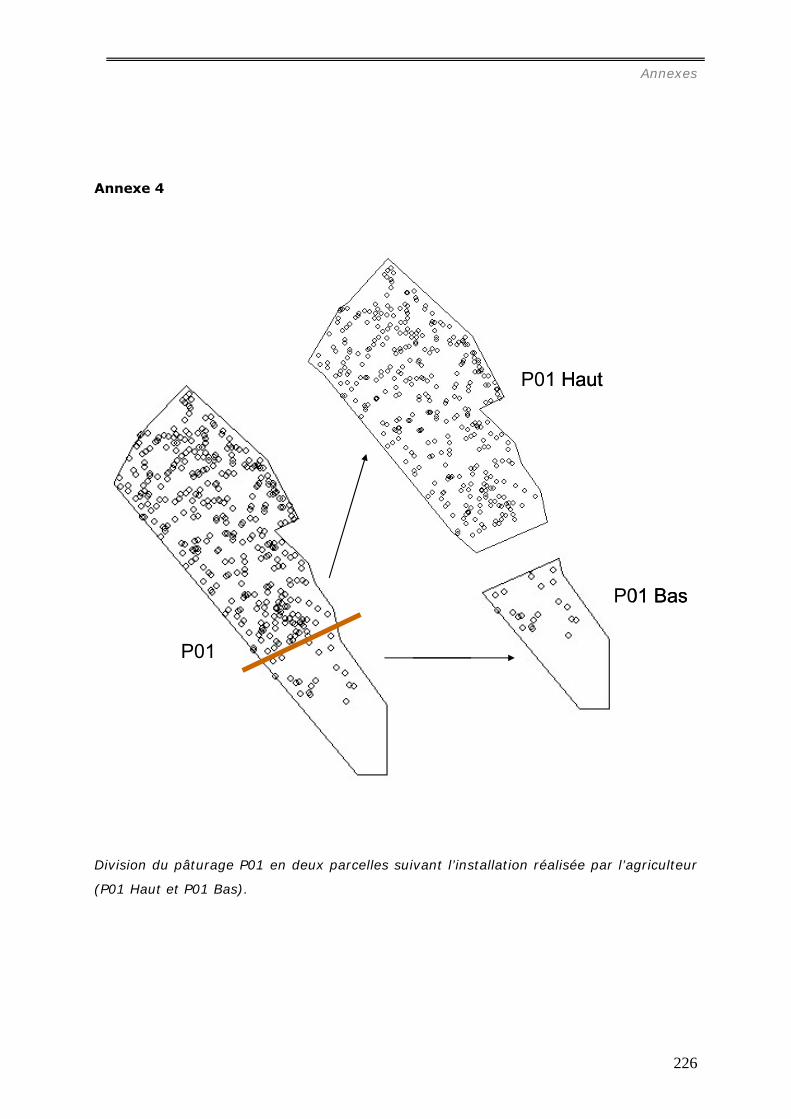
Annexes
226
Annexe 4
P01
P01 Haut
P01 Bas
P01
P01 Haut
P01 Bas
Division du pâturage P01 en deux parcelles suivant l’installation réalisée par l’agriculteur
(P01 Haut et P01 Bas).
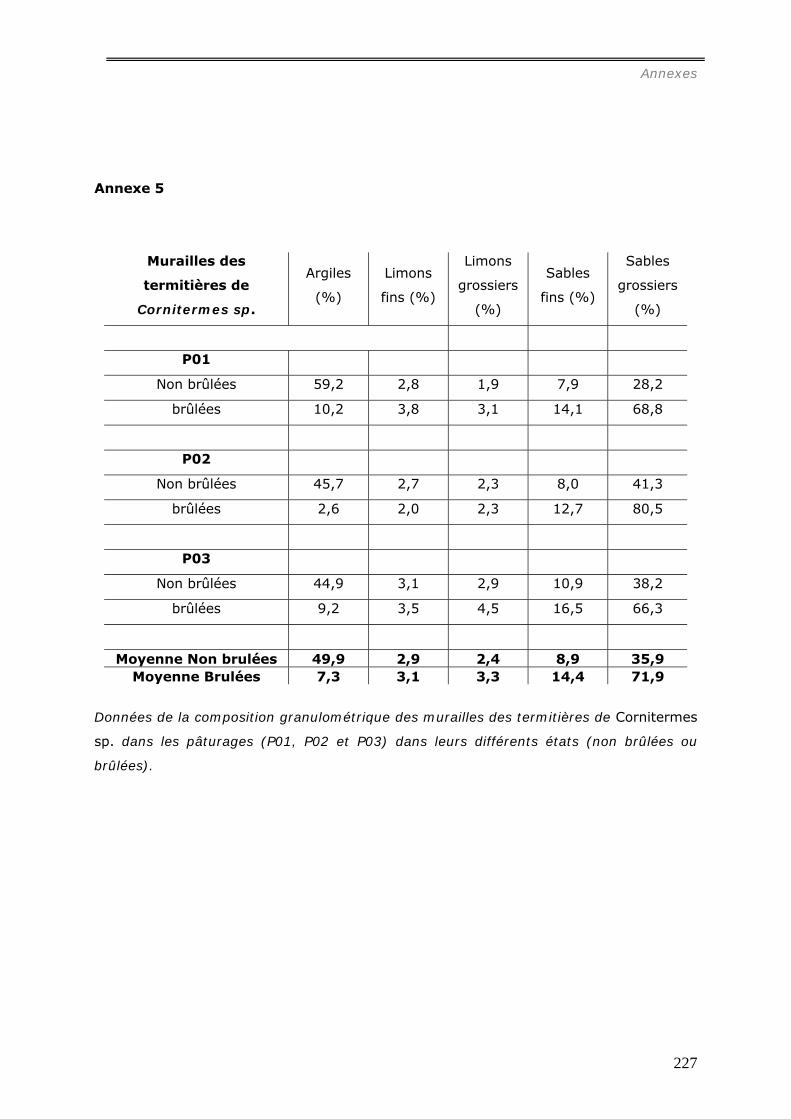
Annexes
227
Annexe 5
Murailles des
termitières de
Cornitermes sp.
Argiles
(%)
Limons
fins (%)
Limons
grossiers
(%)
Sables
fins (%)
Sables
grossiers
(%)
P01
Non brûlées 59,2 2,8 1,9 7,9 28,2
brûlées 10,2 3,8 3,1 14,1 68,8
P02
Non brûlées 45,7 2,7 2,3 8,0 41,3
brûlées 2,6 2,0 2,3 12,7 80,5
P03
Non brûlées 44,9 3,1 2,9 10,9 38,2
brûlées 9,2 3,5 4,5 16,5 66,3
Moyenne Non brulées 49,9 2,9 2,4 8,9 35,9 Moyenne Brulées 7,3 3,1 3,3 14,4 71,9
Données de la composition granulométrique des murailles des termitières de Cornitermes
sp. dans les pâturages (P01, P02 et P03) dans leurs différents états (non brûlées ou
brûlées).
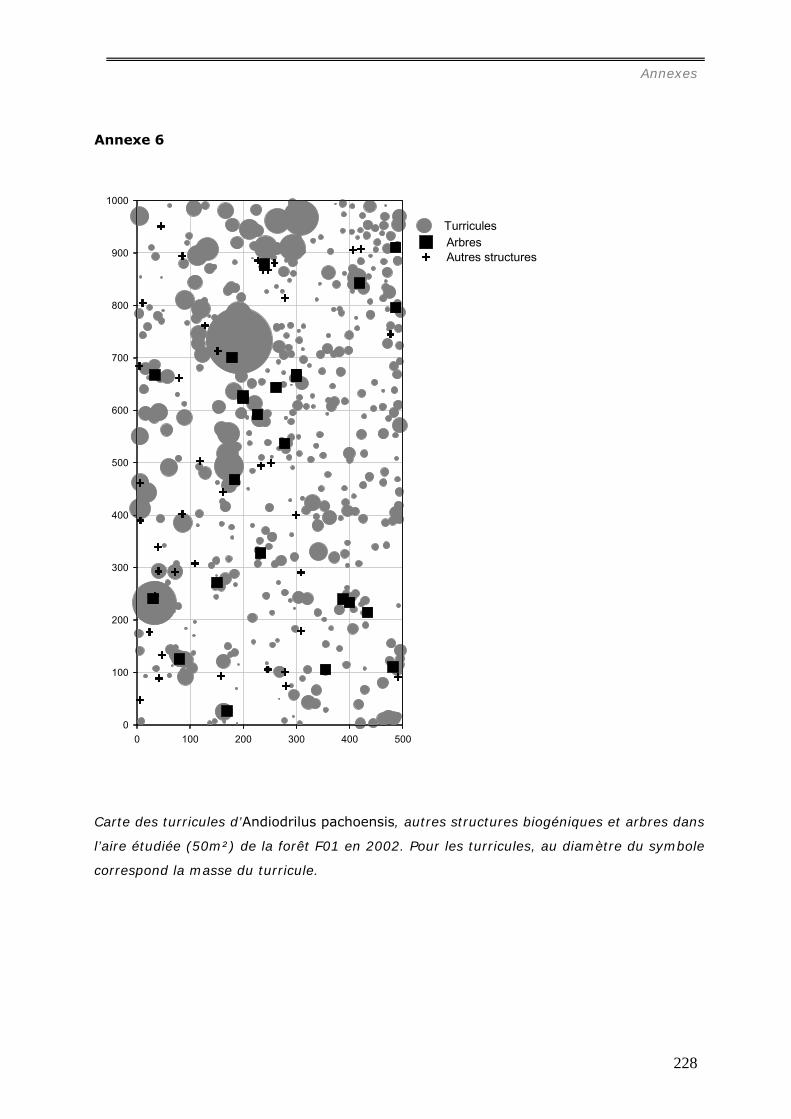
Annexes
228
Annexe 6
0 100 200 300 400 5000
100
200
300
400
500
600
700
800
900
1000
TurriculesArbresAutres structures
Carte des turricules d’Andiodrilus pachoensis, autres structures biogéniques et arbres dans
l’aire étudiée (50m²) de la forêt F01 en 2002. Pour les turricules, au diamètre du symbole
correspond la masse du turricule.

Annexes
229
Annexe 7
0 100 200 300 400 5000
100
200
300
400
500Turricules J0Turricules J1Turricules J2
Carte des turricules d’Andiodrilus pachoensis dans l’aire étudiée (25m²) de la forêt F01
en 2003. Les trois couleurs correspondent aux trois dates d’échantillonnage (J0, J1 et
J2).

Annexes
230
Annexe 8
0 100 200 300 400 5000
100
200
300
400
500Turricules J0Turricules J1Arbres
Carte des turricules d’Andiodrilus pachoensis au jour J0 et J1, ainsi que des troncs dans
l’aire étudiée (25m²) de la forêt F02 en 2003.

Annexes
231
Annexe 9
0 100 200 300 400 5000
100
200
300
400
500Turricules J0Arbres
Carte des turricules d’Andiodrilus pachoensis et des troncs dans l’aire étudiée (25m²) de
la forêt F03 en 2003.
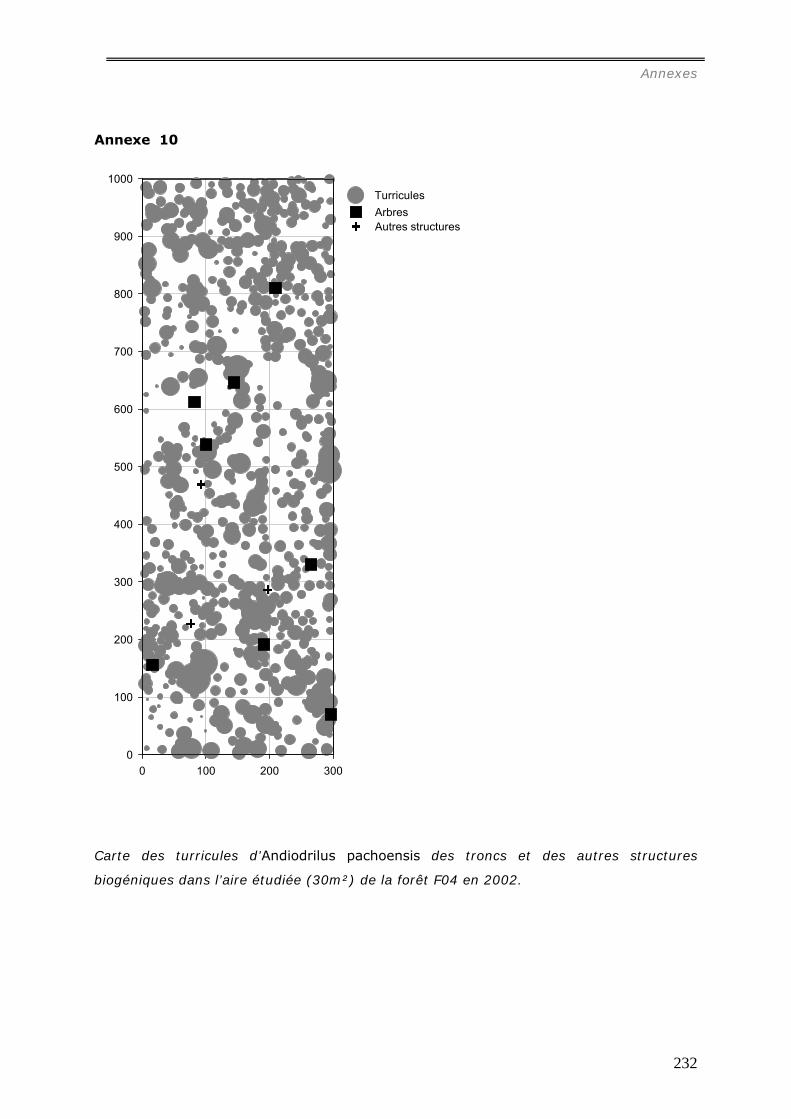
Annexes
232
Annexe 10
0 100 200 3000
100
200
300
400
500
600
700
800
900
1000TurriculesArbresAutres structures
Carte des turricules d’Andiodrilus pachoensis des troncs et des autres structures
biogéniques dans l’aire étudiée (30m²) de la forêt F04 en 2002.
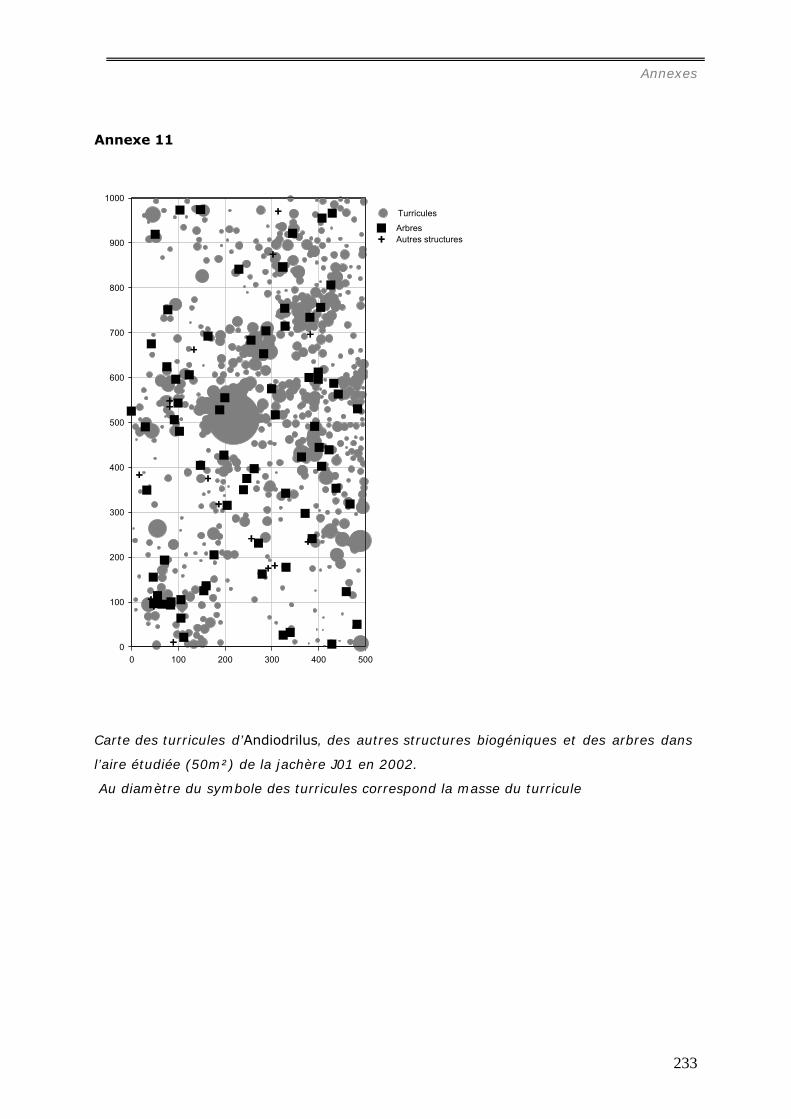
Annexes
233
Annexe 11
0 100 200 300 400 5000
100
200
300
400
500
600
700
800
900
1000
Turricules
ArbresAutres structures
Carte des turricules d’Andiodrilus, des autres structures biogéniques et des arbres dans
l’aire étudiée (50m²) de la jachère J01 en 2002.
Au diamètre du symbole des turricules correspond la masse du turricule
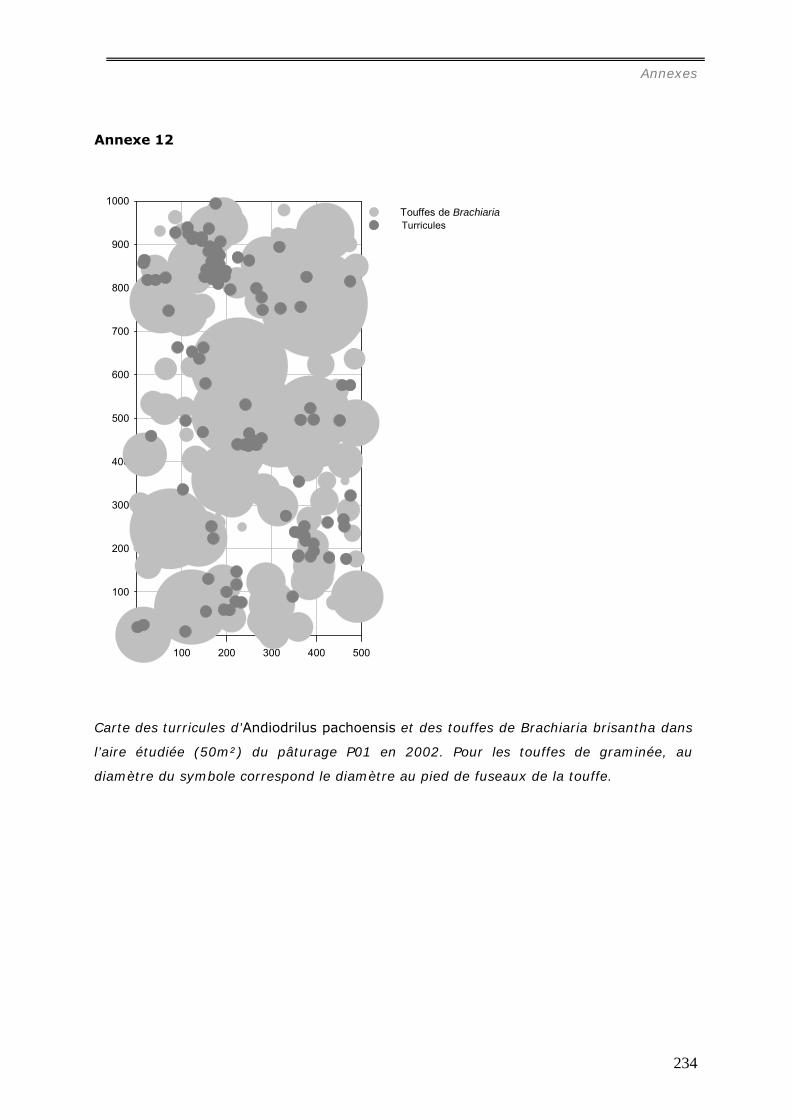
Annexes
234
Annexe 12
Carte des turricules d’Andiodrilus pachoensis et des touffes de Brachiaria brisantha dans
l’aire étudiée (50m²) du pâturage P01 en 2002. Pour les touffes de graminée, au
diamètre du symbole correspond le diamètre au pied de fuseaux de la touffe.
0 100 200 300 400 500
0
100
200
300
400
500
600
700
800
900
1000Touffes de Brachiaria Turricules

Annexes
235
Annexe 13
0 100 200 300 400 5000
100
200
300
400
500
600
700
800
900
1000
Turricules J0Turricules J1Turricules J2
Carte des turricules d’Andiodrilus pachoensis au jour J0, J1 et J2, dans l’aire étudiée
(50m²) du pâturage P01 en 2003.
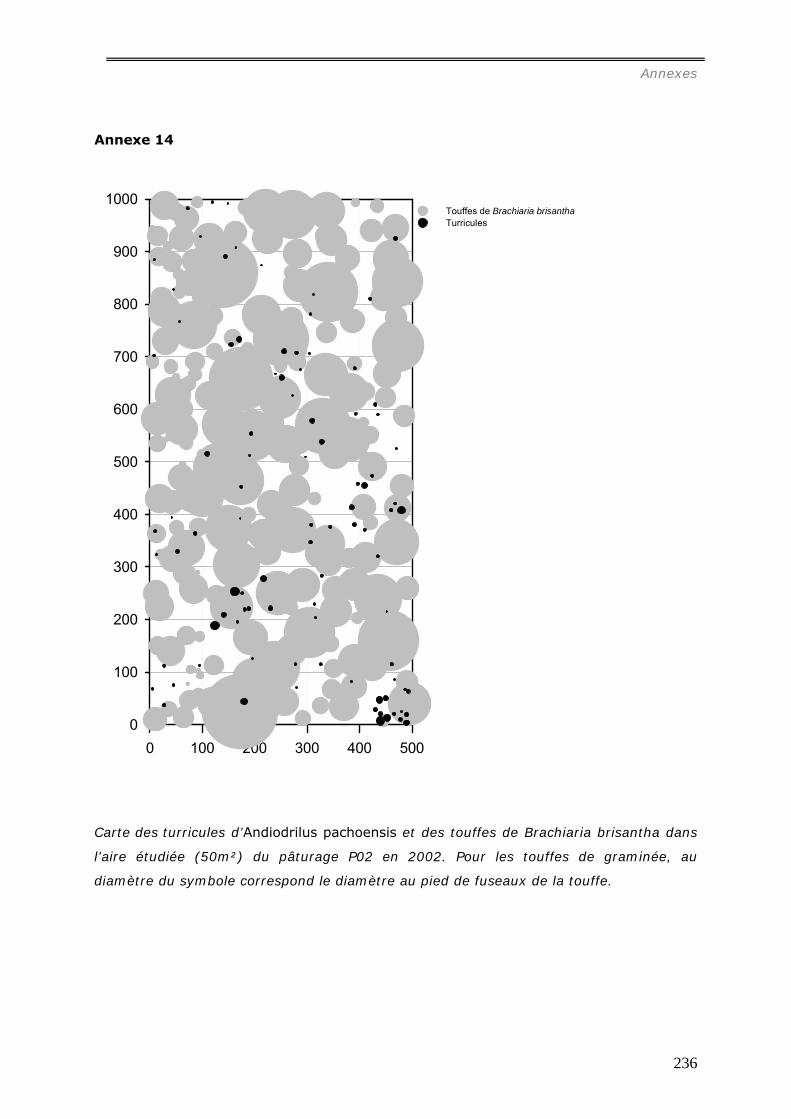
Annexes
236
Annexe 14
0 100 200 300 400 5000
100
200
300
400
500
600
700
800
900
1000Touffes de Brachiaria brisanthaTurricules
Carte des turricules d’Andiodrilus pachoensis et des touffes de Brachiaria brisantha dans
l’aire étudiée (50m²) du pâturage P02 en 2002. Pour les touffes de graminée, au
diamètre du symbole correspond le diamètre au pied de fuseaux de la touffe.

Annexes
237
Annexe 15
0 100 200 300 400 5000
100
200
300
400
500
Touffes
0 100 200 300 400 5000
100
200
300
400
500
600
700
800
900
1000
Turricules J0Turricules J1
Carte des turricules d’Andiodrilus pachoensis au jour J0 et J1 (gauche) dans l’aire étudiée
(50m²) du pâturage P03 en 2003 et carte des turricules et touffes dans les premiers 25
m² (droite).

Annexes
238
Annexe 16
0 100 200 300 400 5000
100
200
300
400
500
600
700
800
900
1000
Turricules J0Turricules J1
Carte des turricules d’Andiodrilus pachoensis au jour J0 et J1 dans l’aire étudiée (50m²)
du pâturage P02b en 2003.
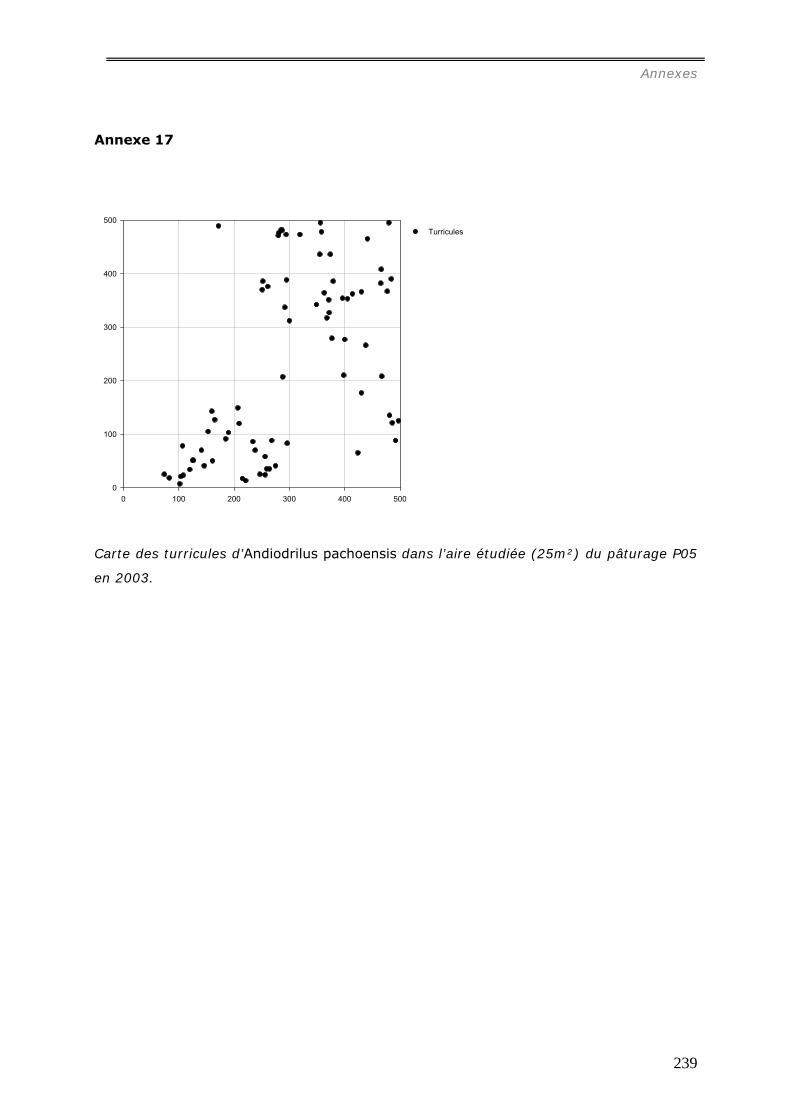
Annexes
239
Annexe 17
0 100 200 300 400 5000
100
200
300
400
500Turricules
Carte des turricules d’Andiodrilus pachoensis dans l’aire étudiée (25m²) du pâturage P05
en 2003.
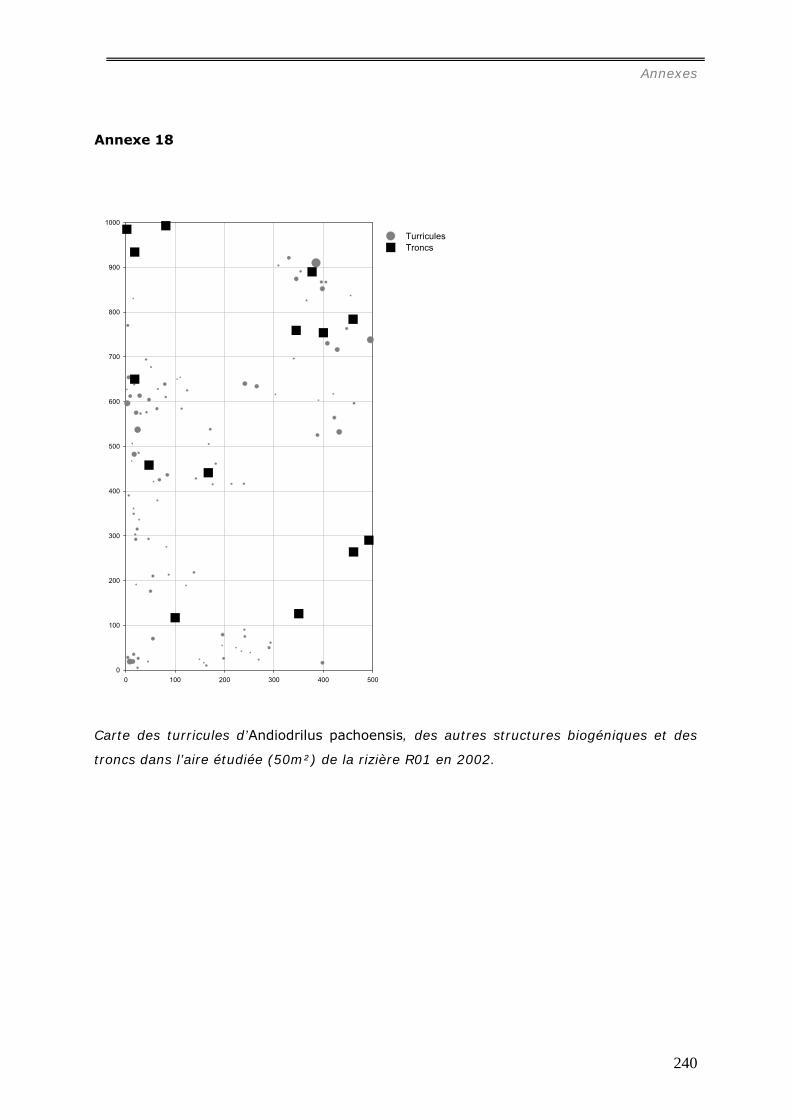
Annexes
240
Annexe 18
0 100 200 300 400 5000
100
200
300
400
500
600
700
800
900
1000
TurriculesTroncs
Carte des turricules d’Andiodrilus pachoensis, des autres structures biogéniques et des
troncs dans l’aire étudiée (50m²) de la rizière R01 en 2002.
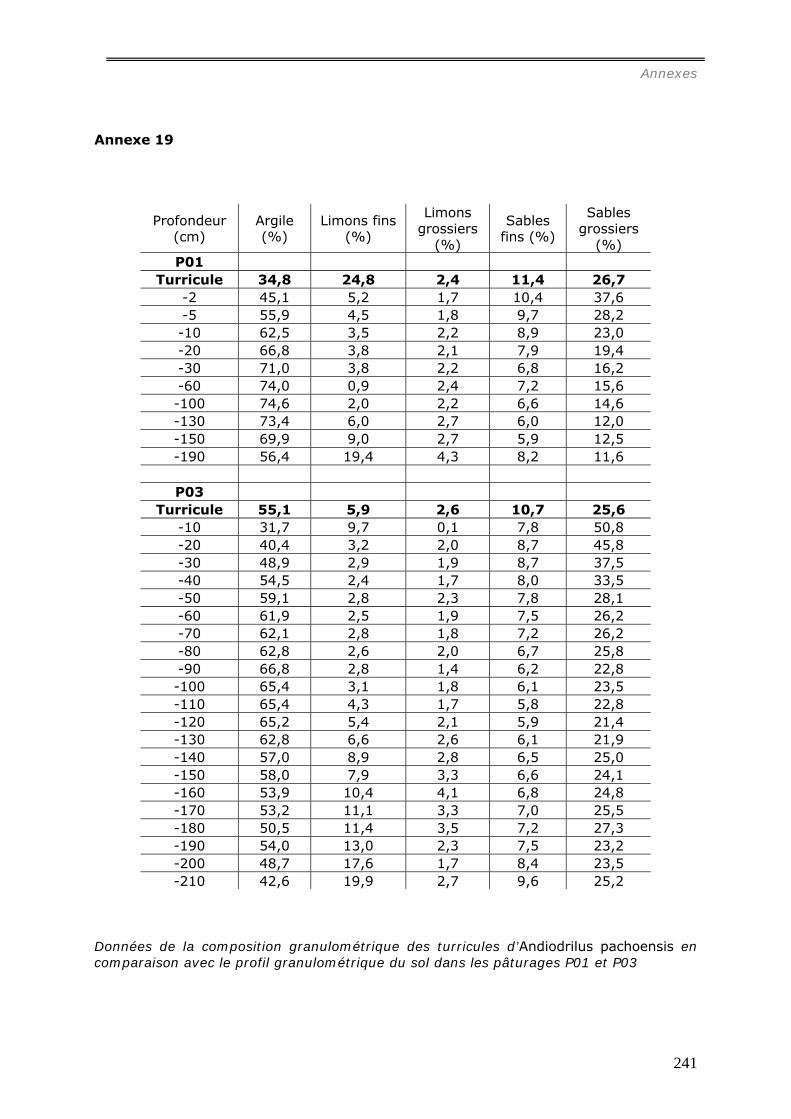
Annexes
241
Annexe 19
Profondeur (cm)
Argile (%)
Limons fins (%)
Limons grossiers
(%)
Sables fins (%)
Sables grossiers
(%) P01
Turricule 34,8 24,8 2,4 11,4 26,7 -2 45,1 5,2 1,7 10,4 37,6 -5 55,9 4,5 1,8 9,7 28,2 -10 62,5 3,5 2,2 8,9 23,0 -20 66,8 3,8 2,1 7,9 19,4 -30 71,0 3,8 2,2 6,8 16,2 -60 74,0 0,9 2,4 7,2 15,6 -100 74,6 2,0 2,2 6,6 14,6 -130 73,4 6,0 2,7 6,0 12,0 -150 69,9 9,0 2,7 5,9 12,5 -190 56,4 19,4 4,3 8,2 11,6
P03
Turricule 55,1 5,9 2,6 10,7 25,6 -10 31,7 9,7 0,1 7,8 50,8 -20 40,4 3,2 2,0 8,7 45,8 -30 48,9 2,9 1,9 8,7 37,5 -40 54,5 2,4 1,7 8,0 33,5 -50 59,1 2,8 2,3 7,8 28,1 -60 61,9 2,5 1,9 7,5 26,2 -70 62,1 2,8 1,8 7,2 26,2 -80 62,8 2,6 2,0 6,7 25,8 -90 66,8 2,8 1,4 6,2 22,8 -100 65,4 3,1 1,8 6,1 23,5 -110 65,4 4,3 1,7 5,8 22,8 -120 65,2 5,4 2,1 5,9 21,4 -130 62,8 6,6 2,6 6,1 21,9 -140 57,0 8,9 2,8 6,5 25,0 -150 58,0 7,9 3,3 6,6 24,1 -160 53,9 10,4 4,1 6,8 24,8 -170 53,2 11,1 3,3 7,0 25,5 -180 50,5 11,4 3,5 7,2 27,3 -190 54,0 13,0 2,3 7,5 23,2 -200 48,7 17,6 1,7 8,4 23,5 -210 42,6 19,9 2,7 9,6 25,2
Données de la composition granulométrique des turricules d’Andiodrilus pachoensis en comparaison avec le profil granulométrique du sol dans les pâturages P01 et P03
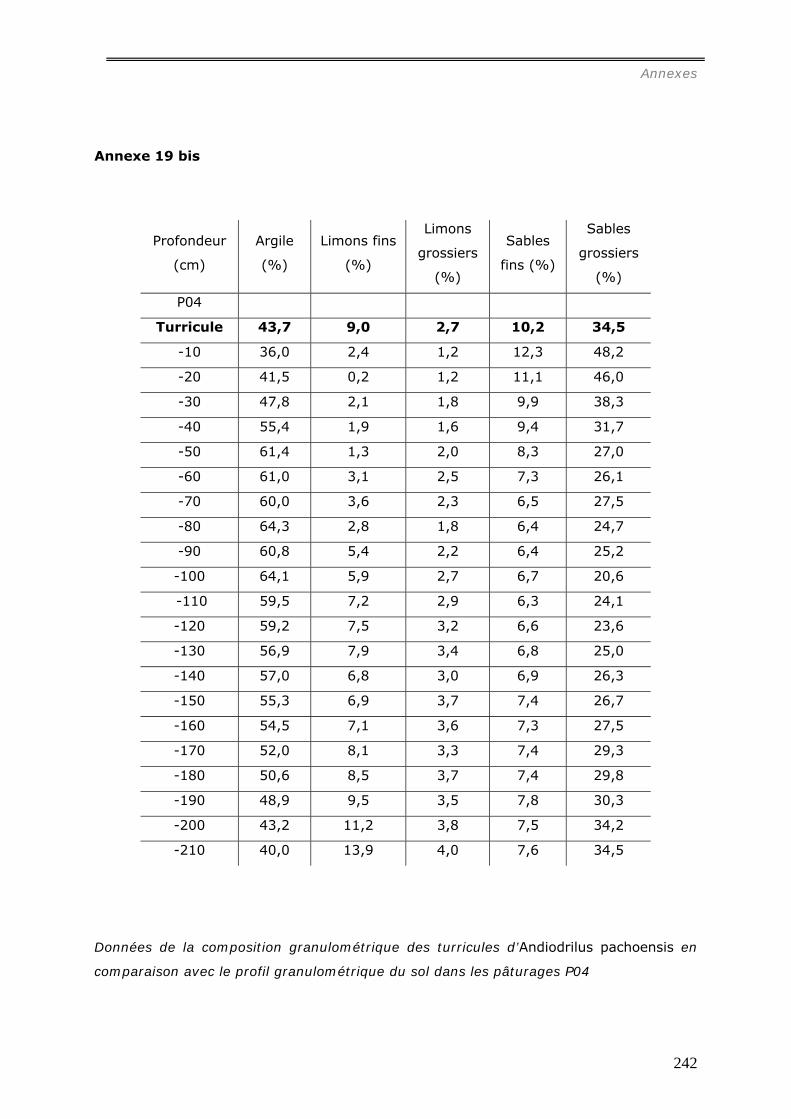
Annexes
242
Annexe 19 bis
Profondeur
(cm)
Argile
(%)
Limons fins
(%)
Limons
grossiers
(%)
Sables
fins (%)
Sables
grossiers
(%)
P04
Turricule 43,7 9,0 2,7 10,2 34,5
-10 36,0 2,4 1,2 12,3 48,2
-20 41,5 0,2 1,2 11,1 46,0
-30 47,8 2,1 1,8 9,9 38,3
-40 55,4 1,9 1,6 9,4 31,7
-50 61,4 1,3 2,0 8,3 27,0
-60 61,0 3,1 2,5 7,3 26,1
-70 60,0 3,6 2,3 6,5 27,5
-80 64,3 2,8 1,8 6,4 24,7
-90 60,8 5,4 2,2 6,4 25,2
-100 64,1 5,9 2,7 6,7 20,6
-110 59,5 7,2 2,9 6,3 24,1
-120 59,2 7,5 3,2 6,6 23,6
-130 56,9 7,9 3,4 6,8 25,0
-140 57,0 6,8 3,0 6,9 26,3
-150 55,3 6,9 3,7 7,4 26,7
-160 54,5 7,1 3,6 7,3 27,5
-170 52,0 8,1 3,3 7,4 29,3
-180 50,6 8,5 3,7 7,4 29,8
-190 48,9 9,5 3,5 7,8 30,3
-200 43,2 11,2 3,8 7,5 34,2
-210 40,0 13,9 4,0 7,6 34,5
Données de la composition granulométrique des turricules d’Andiodrilus pachoensis en
comparaison avec le profil granulométrique du sol dans les pâturages P04
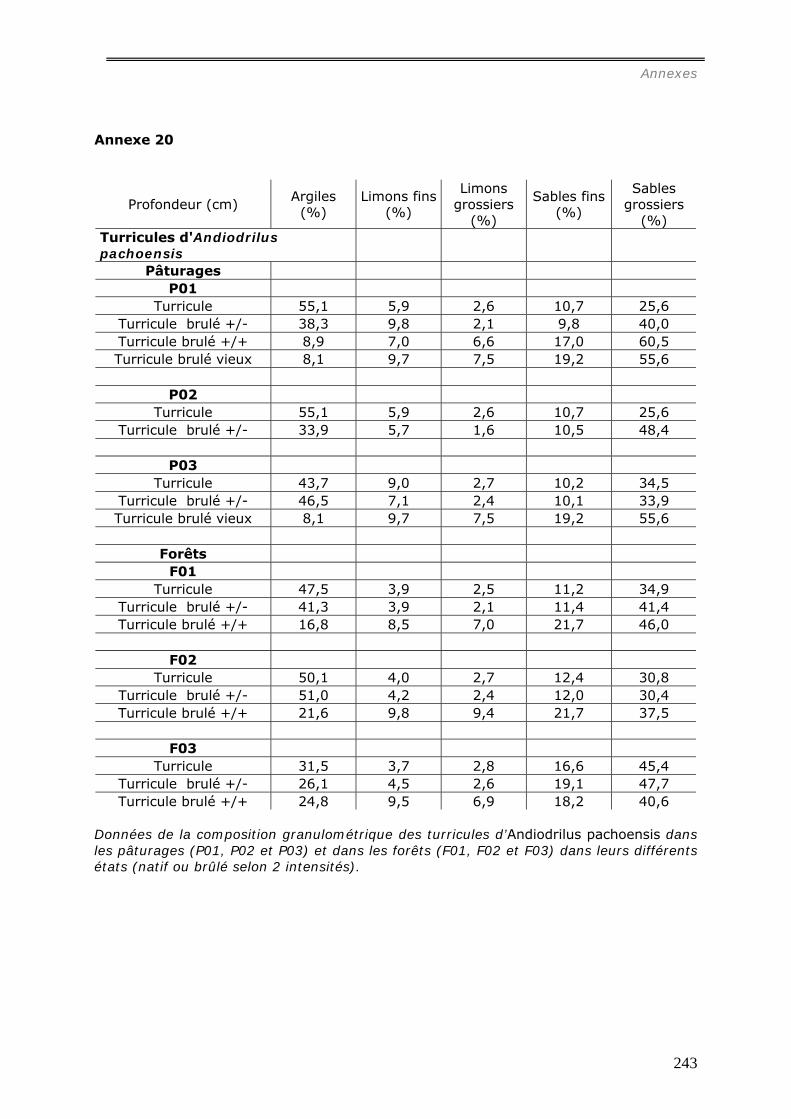
Annexes
243
Annexe 20
Profondeur (cm) Argiles (%)
Limons fins (%)
Limons grossiers
(%)
Sables fins (%)
Sables grossiers
(%) Turricules d'Andiodrilus pachoensis
Pâturages P01
Turricule 55,1 5,9 2,6 10,7 25,6 Turricule brulé +/- 38,3 9,8 2,1 9,8 40,0 Turricule brulé +/+ 8,9 7,0 6,6 17,0 60,5 Turricule brulé vieux 8,1 9,7 7,5 19,2 55,6
P02
Turricule 55,1 5,9 2,6 10,7 25,6 Turricule brulé +/- 33,9 5,7 1,6 10,5 48,4
P03
Turricule 43,7 9,0 2,7 10,2 34,5 Turricule brulé +/- 46,5 7,1 2,4 10,1 33,9 Turricule brulé vieux 8,1 9,7 7,5 19,2 55,6
Forêts
F01 Turricule 47,5 3,9 2,5 11,2 34,9
Turricule brulé +/- 41,3 3,9 2,1 11,4 41,4 Turricule brulé +/+ 16,8 8,5 7,0 21,7 46,0
F02
Turricule 50,1 4,0 2,7 12,4 30,8 Turricule brulé +/- 51,0 4,2 2,4 12,0 30,4 Turricule brulé +/+ 21,6 9,8 9,4 21,7 37,5
F03
Turricule 31,5 3,7 2,8 16,6 45,4 Turricule brulé +/- 26,1 4,5 2,6 19,1 47,7 Turricule brulé +/+ 24,8 9,5 6,9 18,2 40,6
Données de la composition granulométrique des turricules d’Andiodrilus pachoensis dans les pâturages (P01, P02 et P03) et dans les forêts (F01, F02 et F03) dans leurs différents états (natif ou brûlé selon 2 intensités).
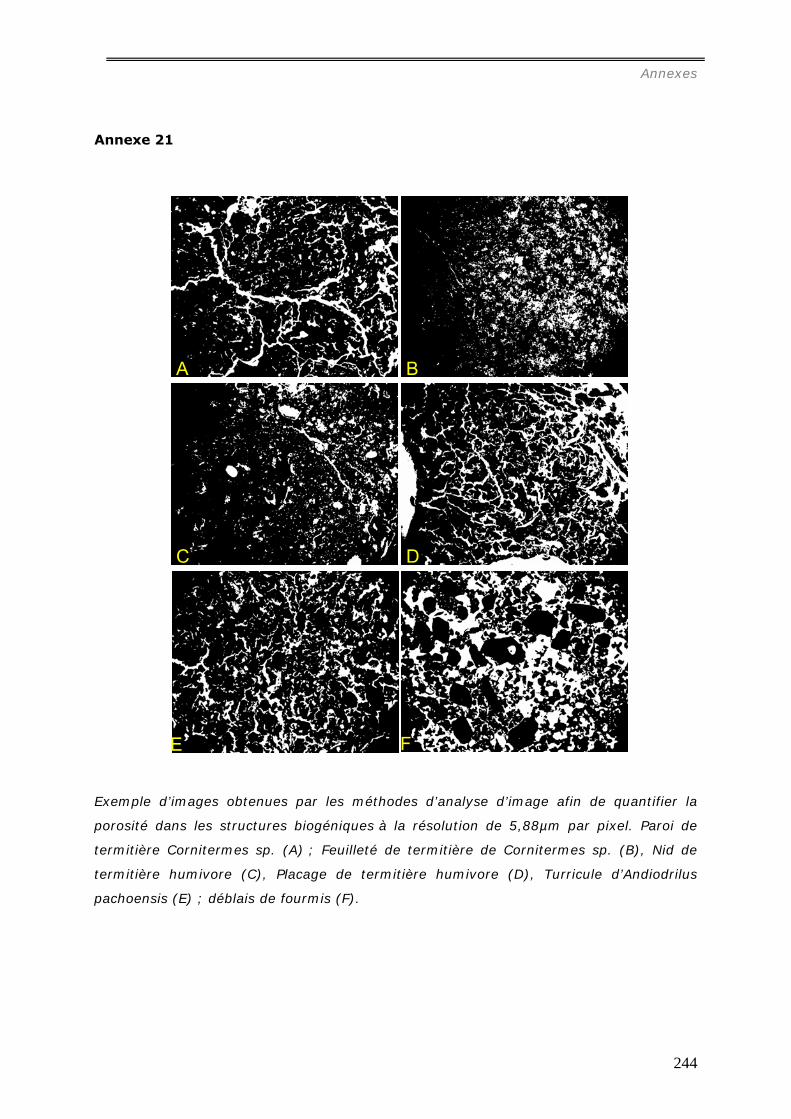
Annexes
244
Annexe 21
Exemple d’images obtenues par les méthodes d’analyse d’image afin de quantifier la
porosité dans les structures biogéniques à la résolution de 5,88µm par pixel. Paroi de
termitière Cornitermes sp. (A) ; Feuilleté de termitière de Cornitermes sp. (B), Nid de
termitière humivore (C), Placage de termitière humivore (D), Turricule d’Andiodrilus
pachoensis (E) ; déblais de fourmis (F).
A B
C
FE
D
A B
C
FE
D

Références bibliographiques.
245
Annexe 22
Exemple d’images obtenues par les méthodes d’analyse d’image afin de quantifier la
porosité dans les structures biogéniques à la résolution de 5,88µm par pixel. Paroi de
termitière Cornitermes sp. (A) ; Feuilleté de termitière de Cornitermes sp. (B), Nid de
termitière humivore (C), Placage de termitière humivore (D), Turricule d’Andiodrilus
pachoensis (E) ; déblais de fourmis (F).
A
C D
B
F E
A
C D
B
F E

Références bibliographiques.
246
Références bibliographiques

Références bibliographiques.
247
Abbadie, L., Lepage, M., 1989. The role of subterranean fungus comb chambers
(Isoptera, Macrotermitinæ) in soil nitrogen cycling in a preforest savanna (Côte
d'Ivoire). Soil Biology and Biochemistry 21, 1067-1071 pp.
Afnor. 1994 Qualité du sol - Détermination du pH.
Albadalejo, C., Tulet, J.-C., 1996. Les Fronts pionniers de l’Amazonie brésilienne : la
formation de nouveaux territoires. L’Harmattan, Paris, 358 pp.
Alegre, J. C., Cassel, D. K., 1996. Dynamics of soil physical properties under alternative
systems to slash-and-burn. Agriculture, Ecosystems and Environment 58, 39-48
pp.
Alegre, J. C., Pashanasi, B., Lavelle, P., 1996. Dynamics of soil physical properties in a
low input agricultural system inoculated with the earthworm Pontoscolex
corethrurus in the amazon region of Peru. SSSA Journal 60, 1522-1529 pp.
Amelung, W., Martius, C., Bandeira, A. G., Garcia, M. V. B., Zech, W., 2002. Lignin
characteristics and density fractions of termite nests in an Amazonian rain forest--
indicators of termite feeding guilds? Soil Biology and Biochemistry 34, 367-372
pp.
Andresen, E., 2003. Effect of forest fragmentation on dung beetle communities and
functional consequences for plant regeneration. Ecography 26, 87-97 pp.
Araujo, E. A., Lani, J. L., Amaral, E. F., Guerra, A., 2004. Land use and physical and
chemical properties of a distrophic Yellow Argisol in the western Amazon region.
Revista Brasileira De Ciencia Do Solo 28, 307-315 pp.
Arnt, R. 2001 Tesouro verde.52-64 pp.
Arshad, M. A., Schnitzer, M., Preston, C. N., 1988. Characterization of humic acids fron
termite mounds and surronding soils, Kenya. Geoderma 42, 213-225 pp.
Arvieu, J. C. 1972. Etude cinétique des réactions entre orthophosphates solubles et
CaCO3. Propriétés physico-chimiques des apatites phospho-calciques formées.
INRA - Faculté des Sciences de Nice,151 pp.
Bachelier, G., 1978. La faune des sols son écologie et son action. ORSTOM, Paris, 391
pp.
Baddeley, A., Turner, R., 2005. Spatstat: an R package for analyzing spatial point
patterns. Journal of Statistical Software 12, 1-42 pp.
Baize, D., Girard, M.-C., 1995. Référentiel pédologique, Paris, 332 pp.
Balbino, L. C., Bruand, A., Brossard, M., Grimaldi, M., Hajnos, M., Guimaraes, M. F.,
2002. Changes in porosity and microaggrégation in clayey Ferrasols of the
Brazilian Cerrado on clearing for pasture. European Journal of Soil Science 53,
219-230 pp.
Baligar, V. C., Fageria, N. K., 1997. Nutrient use efficiency in acid soils : nutrient
management and plant use efficiency. in Moniz, A. C., Furlani, A. M. C., Schaeffer,

Références bibliographiques.
248
R. E., Fageria, N. K., Rosolem, C. A., Cantarella, H., (eds). Plant-Soil Interactions
at low pH. Brasilian Soil Sciance Society, Campinas, SP, Viçosa, MG, Brazil, 75-96
pp.
Barois, I., 1992. Mucus production and microbial activity in the gut of two species of
Amynthas (Megascolecidae) from cold and warm tropical climates. Soil Biology
and Biochemistry 24, 1507-1510 pp.
Barois, I., Verdier, B., Kaiser, P., Mariotti, A., Rangel, P., Lavelle, P., 1987. Influence of
the tropical earthworm Pontoscolex corethrurus (Glossoscolecidæ) on the fixation
and mineralization of nitrogen. in Bonvicini, A. M., Omodeo, P., (eds). On
Earthworms. Mucchi, Bologna, Italy, 151-158 pp.
Barros, E., Grimaldi, M., Desjardins, T., Sarrazin, M., Chauvel, A., Lavelle, P. 1998
Conversion of forest into pastures in Amazonia : effects on soil macrofaunal
diversity and soil water dynamics. 8 pp.
Batabyal, A. A., Lee, D. M., 2003. Aspects of land use in slash and burn agriculture.
Applied Economics Letters 10, 821-824 pp.
Bengtsson, J., 1998. Which species? What kind of diversity? Which ecosystem function?
Some problems in studies of relations between biodiversity and ecosystem
function. Applied Soil Ecology 10, 191-199 pp.
Besag, J. E., 1977. Comments on Ripley's paper. Journal of the Royal Statistical Society
series B 39, 193-195 pp.
Besag, J. E., Clifford, P., 1989. Generalized Monte Carlo Significance Tests. Biometrika
76, 633-642 pp.
Bhadauria, T., Ramakrishnan, P. S., Srivastava, K. N., 2000. Diversity and distribution of
endemic and exotic earthworms in natural and regenerating ecosystems in the
central Himalayas, India. Soil Biology and Biochemistry 32, 2045-2054 pp.
Binet, F., Le Bayon, R. C., 1998. Space-time dynamics in situ of earthworm casts under
temperate cultivated soils. Soil Biology and Biochemistry 31, 85-93 pp.
Birang, M. A., Hauser, S., Brussaard, L., Norgrove, L., 2003. Earthworm surface casting
activity on slash-and-burn cropped land and in undisturbed Chromolaena odorata
and young forest fallow in southern Cameroon. Pedobiologia 47, 811-818 pp.
Bird, M. I., Veenendaal, E. M., Moyo, C., Lloyd, J., Frost, P., 2000. Effect of fire and soil
texture on soil carbon in a sub-humid savanna,Matopos, Zimbabwe. Geoderma
94, 71–90 pp.
Biri Kassoum, B., Maître d'Hôtel, E. 2002 Contribution à l’étude de la durabilité de
l’agriculture familiale en zone de front pionnier : exemple de la communauté de
Benfica en Amazonie Orientale Brésilienne.84 pp.
Blanchart, E., Albrecht, A., Alegre, J., Duboisset, A., Gilot, C., Pashanasi, B., Lavelle, P.,
Brussaard, L., 1999. Effects of earthworms on soil structure and Physical

Références bibliographiques.
249
Properties. in Lavelle, P., Brussaard, L., Hendrix, P. F., (eds). Earthworm
Management in tropical agrosystems. CAB international, Wallingford, UK, 149-
172 pp.
Blanchart, E., Bruand, A., Lavelle, P., 1993a. The physical structure of casts of Millsonia
anomala (Oligochaeta: Megascolecidae) in shrub savanna soils (Cote d'Ivoire).
Geoderma 56, 119-132 pp.
Blanchart, E., Bruand, A., Lavelle, P., 1993b. The physical structure of casts of Millsonia
anomala (Oligochaeta:Megascolecidae) in shrub savanna soils (Côte d'Ivoire).
Geoderma 56, 119-132 pp.
Bobillier, B. 1999. Premiers inventaires de la macrofaune d’invertébrés du sol sous
pâturages de faible productivité du Cerrado (Centre-Ouest du Brésil). mém. de
D.E.S.S 59+ 21 anexos, Université Paris-Val de Marne: IRD: Embrapa Cerrados,
Paris.
Bond, E. M., Chase, J. M., 2002. Biodiversity and ecosystem functioning at local and
regional spatial scales. Ecology Letters 5, 467-470 pp.
Bossuyt, H., Six, J., Hendrix, P. F., 2005. Protection of soil carbon by microaggregates
within earthworm casts. Soil Biology and Biochemistry 37, 251-258 pp.
Brandão, D., 1991. Relações espaciais de duas espécies de Syntermes (Isoptera,
Termitidae) nos cerrados da região de Brasilia. Revista Brasileira De Entomologia
35, 745-754 pp.
Brauman, A., 2000. Effect of gut transit and mound deposit on soil organic matter
transformations in the soil feeding termite: A review. European Journal of Soil
Biology 36, 117-125 pp.
Brown, G. G., Barois, I., Lavelle, P., 2000. Regulation of soil organic matter dynamics
and microbial activity in the drilosphere and the role of interactions with other
edaphic functional domains. European Journal of Soil Biology 36, 177-198 pp.
Brunet, D. 2004. As propriedades físicas e a hidrodinâmica de um Latossolo argiloso sob
pastagens no Cerrado: resultados do experimento da fazenda Rio de Janeiro
(GO). Relatório de síntese 30, Embrapa Cerrados: IRD, Planaltina, DF.
Brussaard, L., 1998. Soil fauna, guilds, functional groups and ecosystem processes.
Applied Soil Ecology 9, 123-135 pp.
Campello, M. R., Novais, R. F., Fernandez, I. E. R., Fontes, M. P. F., Barros, N. F., 1994.
Avaliação da reversibilidade de fosforo não labil em solos com diférentes
caracteristicas. Revista Brasileira De Ciencia Do Solo 18, 157-165 pp.
Cannavacciuolo, M., Bellido, A., Cluzeau, D., Gascuel, C., Trehen, P., 1998. A
geostatistical approach to the study of earthworm distribution in grassland.
Applied Soil Ecology 9, 345-349 pp.

Références bibliographiques.
250
Capobianco, J. P. R., 2001. Introduçao. in Socioambiental, E. L. I., (eds). Biodiversidade
na Amazônia brasileira, Brasilia / Sao Paulo,
Cardinale, B. J., Palmer, M. A., Collins, S. L., 2002. Species diversity enhances
ecosystem functioning through interspecific facilitation. Nature 415, 426-429 pp.
Carmouze, J.-P., Lucotte, M., Boudou, A., 2001. Le mercure en Amazonie : rôle de
l'homme et de l'environnement, risques sanitaires, Paris, 494 pp.
Carvalho, K. S., Vasconcelos, H. L., 1999. Forest fragmentation in central Amazonia and
its effects on litter-dwelling ants. Biological Conservation 91, 151-157 pp.
Casenave, A., Valentin, C., 1988. Les états de surface de la zone sahélienne : influence
sur l'infiltration, Paris, 200 pp.
Certini, G., 2005. Effects of fire on properties of forest soils: a review. Oecologia 143, 1-
10 pp.
Chauvel, A., 1997. Mise en valeur de la forêt et modifications écologiques. in Théry, H.,
(eds). Environnement et Développment en Amazonie Brésilienne. Belin, Paris, 42-
75 pp.
Chauvel, A., Grimaldi, M., Barros, E., Blanchart, E., Desjardins, T., Sarrazin, M., Lavelle,
P., 1999. Pasture damage by an Amazonian earthworm. Nature 398, 32-33 pp.
Coelho, M. R., Santos, H. G. d., Silva, E. F., Aglio, M. L. D., 2002. O Recurso Natural
Solo. in Manzatto, C. V., Freitas Junior, E., Peres, J. R. R., (eds). Uso agricola dos
solos brasileiros. Embrapa Solos, Rio de Janeiro, 1-11 pp.
Collins, N. M., 1981. The role of termites in the decomposition of wood and leaf litter in
the Southern Guinea of Nigeria. Oecologia 51, 389-399 pp.
Constantino, R., 2002. The pest termites of South America: taxonomy, distribution and
status. Journal of Applied Entomology 126, 355-365 pp.
Constantino, R., Acioli, A. N. S., 2006. Termite Diversity in Brazil (Insecta: Isoptera). in
Moreira, F. M. S., Siqueira, J. O., Brussaard, L., (eds). Soil Biodiversity and Other
Brazilian Ecosystems. Cabi Publishing, London, 117-128 pp.
Corazza, E. J. 2002. Biodisponibilidade do íon fosfato em solos sob pastagens cultivadas
na região do Cerrado. Piracicaba. Universidade de São Paulo-ESALQ, São
Paulo,127 pp.
Corazza, E. J., Brossard, M., Dias, C. V. 2000 Soil chemical characteristics under low
productivity pastures: a contribution to a regional approach in the Cerrado region.
Corazza, E. J., Brossard, M., Oliveira, A., 1999. Variabilidade química das camadas
superficiais de solos do município de Unaí (MG). in CONGRESSO BRASILEIRO DE
CIENCIA DO SOLO. SBCS - Embrapa Cerrados, Brasília, DF.Planaltina,
Coster, M., Cherman, J. L., 1989. Précis d’analyse d’images. Presses du CNRS, Paris,

Références bibliographiques.
251
Cragg, R. G., Bardgett, R. D., 2001. How changes in soil faunal diversity and composition
within a trophic group influence decomposition processes. Soil Biology and
Biochemistry 33, 2073-2081 pp.
Craswell, E. T., Lefroy, R. D. B., 2001. The role and function of organic matter in tropical
soils. in Martius, C., Tiessen, H., Vlek, P. L. G., (eds). Managing organic matter in
tropical soils : Scope and Limitations. . Kluwer, Dordrecht, The Netherlands, 7-18
pp.
Cressie, N. A., 1993. Statistics for spatial data. John Wiley & Sons, New York,
Da S. Martins, P. F., Cerri, C. C., Volkoff, B., Andreux, F., Chauvel, A., 1991.
Consequences of clearing and tillage on the soil of a natural Amazonian
ecosystem. Forest Ecology and Management 38, 273-282 pp.
Daily, G. C., Alexander, S., Ehrlich, P. R., 1997. Ecosystem services: benefits supplied to
human societies by natural ecosystems. Issues in Ecology 2, 18 pp.
Decaëns, T., 2000. Degradation dynamics of surface earthworm casts in grasslands of
the eastern plains of Colombia. Biology and Fertility of Soils 32, 149-156 pp.
Decaëns, T., Galvis, J. H., Amezquita, E., 2001. Properties of the structures created by
ecological engineers at the soil surface of a Colombian savanna. Comptes Rendus
de l'Academie des Sciences - Series III - Sciences de la Vie 324, 465-478 pp.
Decaëns, T., Jiménez, J. J., Lavelle, P., 1999a. Effect of exclusion of the anecic
earthworm Martiodrilus carimaguensis Jiménez and Moreno on soil properties and
plant growth in grasslands of the eastern plains of Colombia. Pedobiologia 43,
835-841 pp.
Decaëns, T., Lavelle, P., J.J., J., Escobar, G., Rippstein, G., 1994. Impact of land
management on soil macrofauna in the Oriental Llanos of Colombia. European
Journal of Soil Biology 30, 157-168 pp.
Decaëns, T., Mariani, L., Betancourt, N., Jiménez, J. J., 2003. Seed dispersion by surface
casting activities of earthworms in Colombian grasslands. Acta Oecologica 24,
175-185 pp.
Decaëns, T., Mariani, L., Lavelle, P., 1999b. Soil surface macrofaunal communities
associated with earthworm casts in grassland of the eastern plains of Colombia.
Applied Soil Ecology 13, 87-100 pp.
Dezzeo, N., Noemi Chacon, Sanojab, E., Picon, G., 2004. Changes in soil properties and
vegetation characteristics
along a forest-savanna gradient in southern Venezuela. Forest Ecology and Management
200, 183–193 pp.
Didham, R. K. P. i., editors. . , , 2001. The Influence of Edge Effects and Forest
Fragmentation on Leaf Litter Invertebrates in Central Amazonia. in Bierregaard, R.
O., Gascon, C., Lovejoy, T. E., Mesquita, C. G., (eds). Lessons From

Références bibliographiques.
252
Amazonia.The ecology and conservation of a fragmented forest. Yale University
Press, New Haven & London, 55-70 pp.
Diggle, P. J., 1983. Statistical analysis of spatial point patterns. Academic Press., London,
148 pp.
Do Prado, H., 1996. Manual de classificaçao de solos do Brasil, 3ème 'edition', Sao Paolo,
Brasil, 194 pp.
Droulers, M., 1995. L'Amazonie. Nathan, Paris, 190 pp.
Duchaufour, P., 1997. Abrégé de Pédologie : sol, végétation, environnement, 5ième
'edition', Paris, 291 pp.
Dunn, R. R., 2004. Managing the tropical landscape: a comparison of the effects of
logging and forest conversion to agriculture on ants, birds, and lepidoptera. Forest
Ecology and Management 191, 215-224 pp.
Edwards, C. A., 2004. Earthworm Ecology, 2nd Ed 'edition'. CRC Press, London, 3-12 pp.
Ekschmitt, K., Griffiths, B. S., 1998. Soil biodiversity and its implications for ecosystem
functioning in a heterogeneous and variable environment. Applied Soil Ecology 10,
201-215 pp.
Emerson, A. E., 1954. Nouvelles espèces de termites décrites au cours des années 1952
et 1953 Insectes Sociaux 1,
Eneji, A. E., Agboola, A., Aiyelari, E. A., Honna, T., Yamamoto, S., Irshad, M., Endo, T.,
2003. Soil physical and micronutrient changes following clearing of a tropical
rainforest. Journal of Forest Research 8, 215-219 pp.
Erwin, T. L., 1982. Tropical Forests : Their Richness in Coleoptera and other Arthropod
Species. Coleopterists Bulletin 36, 74-75 pp.
FAO, 1996. Forest Ressources Assessment 1990 : Survey of Tropical Forest Cover and
Study of Change Processes. FAO Forestry paper 120,
Flegel, M., Schrader, S., Zhang, H., 1998. Influence of food quality on the physical and
chemical properties of detritivorous earthworm casts. Applied Soil Ecology 9, 263-
269 pp.
Folgarait, P. J., Thomas, F., Desjardins, T., Grimaldi, M., Tayasu, I., Curmi, P., Lavelle,
P., 2003. Soil properties and the macrofauna community in abandoned irrigated
rice fields of northeastern Argentina. Biology and Fertility of Soils 38, 349-357 pp.
Fontaine, S., Mariotti, A., Abbadie, L., 2003. The priming effect of organic matter: a
question of microbial competition? Soil Biology and Biochemistry 35, 837-843 pp.
Fontes, M. P. F., Weeds, B., 1996. Phosphate adsorption by clays from brazilian oxysols :
relationship with specific surface area and mineralogy. Geoderma 72, 37-51 pp.
Fragoso, C., Brown, G. G., Patron, J. C., Blanchart, E., Lavelle, P., Pashanasi, B.,
Senapati, B., Kumar, T., 1997. Agricultural intensification, soil biodiversity and

Références bibliographiques.
253
agroecosystem function in the tropics: the role of earthworms. Applied Soil
Ecology 6, 17-35 pp.
Fregonezi, G. A. d. F., Brossard , M., Guimarães, M. F., Medina, C. C., 2001.
Transformações morfológicas e físicas de um latossolo argiloso sob pastagens.
Revista Brasileira De Ciencia Do Solo 25, 1017-1027 pp.
Garcia-Montiel, D. C., Neill, C., Melillo, J., Thomas, S., Steudler, P. A., Cerri, C. C., 2000.
Soil phosphorus transformations following forest clearing for pasture in the
Brazilian Amazon. Soil Science Society Of America Journal 64, 1792-1804 pp.
Garnier-Sillam, E. 1987. Biologie et rôle des termites dans les processus d'humification
des sols forestiers tropicaux du Congo. Thèse d'état. Paris V-de Marne,276 pp.
Garnier-Sillam, E., Harry, M., 1995. Distribution of humic compounds in mounds of some
soil-feeding termite species of tropical rainforests: Its influence on soil structure
stability. Insectes Sociaux 42, 167-185 pp.
Garnier-Sillam, E., Renoux, J., 1989. Les composés humiques des termitières de
Thoracotermes macrothorax (humivore) et de Macrotermes mülleri
(champignonniste). Soil Biology and Biochemistry 21, 499-505 pp.
Garnier-Sillam, E., Villemin, G., Toutain, F., Renoux, J., 1985. Formation de micro-
agrègats organo-minéraux dans les fèces de termites. Comptes Rendus de
l'Academie des Sciences, Paris, Série 3 301, 213-218 pp.
Gehring, C., Denich, M., Vlek, P. L. G., 2005. Resilience of secondary forest regrowth
after slash-and-burn agriculture in central Amazonia. Journal of Tropical Ecology
21, 519-527 pp.
Giller, K. E., Beare, M. H., Lavelle, P., Izac, A. M. N., 1997. Agricultural intensification,
soil biodiversity and agroecosystem function. Applied Soil Ecology 6, 3-16 pp.
Giovannini, G., Lucchesi, S., Giachetti, M., 1988. Effect of heating on some physical and
chemical parameters related to soil aggregation and erodibility. . Soil Science and
Plant Nutrition 146, 255-261 pp.
Golchin, A., Oades, J. M., Skjemstad, J. O., P., C., 1994. Soil structure and carbon
cycling. Australian Journal of soil Research 32, 1043- 1068 pp.
Gonçalves, C. R., d'Araujo, S., 1962. Observaçoes sôbre Isopteros no Brasil. arquivos do
museum nacional 52, 193-208 pp.
Goodman, G., Hall, A., 1990. The Future of Amazonia : Destruction or Sustainable
Development ? in colombien, M. G., (eds). El Comercio Ilegal de Especes, Londres
(R.-U.),
Goreaud, F., Pélissier, R., 1999. On explicit formulas of edge effect correction for Ripley’s
K-function. Journal of Vegetation Science 10, 433-438 pp.
Grassé, P.-P., 1984a. Termitologia : nid, construction, Masson 'edition'. Fondation
Singer-Polignac, Paris, 613 pp.

Références bibliographiques.
254
Grassé, P.-P., 1984b. Termitologia: anatomie, physiologie, reproduction, Masson
'edition'. Fondation Singer-Polignac, Paris, 676 pp.
Grassé, P.-P., 1984c. Termitologia: comportement, écologie, évolution, systematique,
Masson 'edition'. Fondation Singer-Polignac, Paris, 715 pp.
Grimaldi, M., 2000. Effet des structures biogéniques sur le fonctionnement du système
sol-plante. Comptes rendus de l'Académie des Sciences série III - Sciences de la
vie - Life Sciences 8, 137-146 pp.
Grimaldi, M., Sarrazin, M., Chauvel, A., Luizao, F., 1993. Effets de la déforestation et des
cultures sur la structure des sols argileux d'Amazonie brésilienne. Cahiers
Agriculture 2, 36-47 pp.
Guerra, A. J. T., Silva, A. S., Botelho, R. G. M., 1999. Erosao e Conservaçao dos Solos :
conceitos, temas, e aplicaçoes, Rio de Janeiro,
Guggenberger, G., Thomas, R. J., Zech, W., 1996. Soil organic matter within earthworm
casts of an anecic-endogeic tropical pasture community, Colombia. Applied Soil
Ecology 3, 263-274 pp.
Hallaire, V., 1994. Description of microcrack orientation in a clayey soil using image
analysis. Development in Soil Science 22, 549-557 pp.
Hallaire, V., Cointepas, J. P., 1993. Caractérisation de la macroporosité d'un sol de
verger par analyse d'image. Agronomie 13, 155-164 pp.
Hallaire, V., Curmi, P., 1994. Image analysis of pore space morphology in soil section in
relation to water movement. Development in Soil Science 22, 559-567 pp.
Hedde, M., Lavelle, P., Joffre, R., Jiménez, J. J., Decaëns, T., 2005. Specific functional
signature in soil macro-invertebrates biostructures. Functional Ecology 19, 785
pp.
Hénin, S. 1938. Etude physico-chimique de la stabilité structurale des terres., Paris
Hénin, S., Monnier, G., Combeau, A., 1958. Méthode pour l'étude de la stabilité
structurale des sols. Annales agronomiques 9, 73-92 pp.
Henrot, J., Brussaard, L., 1997. Abundance, casting activity, and cast quality of
earthworms in an acid Ultisol under alley-cropping in the humid tropics. Applied
Soil Ecology 6, 169-179 pp.
Herbert, D. A., Williams, M., Rastetter, E. B., 2003. A model analysis of N and P
limitation on carbon accumulation in Amazonian secondary forest after alternate
land-use abandonment. Biogeochemistry 65, 121-150 pp.
Hindell, R. P., McKenzie, B. M., Tisdall, J. M., 1997. Influence of drying and ageing on the
stabilization of earthworm (Lumbricidae) casts. Biology and Fertility of Soils 25,
27-35 pp.
Höfer, H., Hanagarth, W., Garcia, M., Martius, C., Franklin, E., Römbke, J., Beck, L.,
2001. Structure and function of soil fauna communities in Amazonian

Références bibliographiques.
255
anthropogenic and natural ecosystems. European Journal of Soil Biology 37, 229-
235 pp.
Horn, H. S., 1974. The Ecology of Secondary Succession Annual Review of Ecology and
Systematics 5, 25-37 pp.
Hruschka, W. R., 1990. Data analysis: Wavelength Selection Methods. in Williams, P.,
Norris, K., (eds). Near Infrared Technology in the Agricultural and Food
Industries. American Association of Cereal Chemists, Inc, St Paul, Minnesota,
USA, 35-53 pp.
Imbrozio Barbosa, R., Pereira do Nascimento, S., Atlântico Figueiredo de Amorim, P.,
Ferreira da Silva, R., 2005. Notas sobre a composição arbóreo-arbustiva de uma
fisionomia das savanas de Roraima, Amazônia Brasileira
Acta Botanicos Brasileiros 19, 323-329 pp.
INMET, I. N. d. M. 2006 Dados meterologicos.
INPE. 2001 Projeto PRODES.
INPE. 2004 Amazonia : Deforestation.
James, S. W., Brown, G. G., 2006. Earthworm Ecology and Diversity in Brazil. in Moreira,
F. M. S., Siqueira, J. O., Brussaard, L., (eds). Soil Biodiversity and Other Brazilian
Ecosystems. Cabi Publishing, London, 56-116 pp.
Janeau, J. L., Valentin, C., 1987. Relation entre les termitières Trinervitremes sp. et la
surface du sol : réorganisation, ruissellement et érosion. Revue d'Ecologie et de
Biologie du Sol 24, 637-647 pp.
Jégou, D., Cluzeau, D., Hallaire, V., Balesdent, J., Trehen, P., 2000. Burrowing activity of
the earthworms Lumbricus terrestris and Aporrectodea giardi and consequences
on C transfers in soil. European Journal of Soil Biology 36, 27-34 pp.
Jiménez, J. J., Cepeda, A., Decaëns, T., Oberson, A., Friesen, D. K., 2003. Phosphorus
fractions and dynamics in surface earthworm casts under native and improved
grasslands in a Colombian savanna Oxisol. Soil Biology and Biochemistry 35, 715-
727 pp.
Jiménez, J. J., Decaëns, T., 2004. The impact of soil organisms on soil functioning under
neotropical pastures: a case study of a tropical anecic earthworm species.
Agriculture, Ecosystems and Environment 103, 329–342 pp.
Jiménez, J. J., Moreno, A. G., Lavelle, P., Decaëns, T., 1998. Population dynamics and
adaptive strategies of Martiodrilus carimaguensis (Oligochaeta, Glossoscolecidae),
a native species from the well-drained savannas of Colombia. Applied Soil Ecology
9, 153-160 pp.
Joly, C. A., Bicudo, C. E. M., 1999. Biodiversidade do estado de São Paulo: síntese do
conhecimento ao final do século XX: invertebrados terrestres., Fundação de
Amparo à Pesquisa do Estado de São Paulo 'edition', São Paulo, 279 pp.

Références bibliographiques.
256
Jones, C. G., Lawton, J. H., Shachak, M., 1994. Organisms as ecosystem engineers.
Oikos 69, 373-386 pp.
Jones, J. A., 1990. Termites, soil fertility and carbon cycling in dry tropical africa : a
hypothesis. Journal of Tropical Ecology 6, 291-305 pp.
Jouquet, P., Mamou, L., Lepage, M., Velde, B., 2002. Effect of termites on clay minerals
in tropical soils : fungus-growing termites as weathering agents. European Journal
of Soil Science 53, 521-527 pp.
Jouquet, P., Tessier, D., Lepage, M., 2004. The soil structural stability of termite nests:
role of clays in Macrotermes bellicosus (Isoptera, Macrotermitinae) mound soils.
European Journal of Soil Biology 40, 23-29 pp.
Juan Santiago, T., Porcel, E. 2000 Estudios Preliminares sobre Densidad y Estimación de
Poblaciones de Nidos de Cornitermes cumulans (Kollar, 1832) (Isoptera,
Termitidae, Nasutitermitinae) de diferentes tamaños en Mburucuyá (Dto.
Mburucuyá, Corrientes).4 pp.
Jungerius, P. D., Van den Ancker, J. A. M., Mucher, H. J., 1999. The contribution of
termites to the microgranular structure of soils on the Uasin Gishu Plateau, Kenya.
Catena 34, 349-363 pp.
Juo, A. S. R., Manu, A., 1996. Chemicals dynamics in slash and burn agriculture.
Agriculture, Ecosystems & Environment 58, 49-60 pp.
Kempf, H., 2005. Le Brésil annonce une baisse record de la déforestation en Amazonie.
Le monde 30.08.2005,
Ketterings, Q. M., Bigham, J. M., 2000. Soil Color as an Indicator of Slash-and-Burn Fire
Severity and Soil Fertility in Sumatra, Indonesia. Soil Science Society of America
Journal 64, 1826-1833 pp.
Korb, J., Linsenmair, K. E., 2000. Thermoregulation of termite mounds: what role does
ambient
temperature and metabolism of the colony play? Insectes Sociaux 47, 357–363 pp.
Kotto-Same, J., Woomer, P. L., Moukam, A., Zapfack, L., 1997. Carbon dynamics in slash
and burn argriculture and land-use alternatives of the humid forest zone in
Cameroon. Agric. Ecos. Envir. 65, 245-256 pp.
Krebs, C. J., 1998. Ecological methodology, 2nd 'edition'. Addison Wesley Longman,
Menlo Park, 620 pp.
Kuperman, R. G., 1996. Relationships between soil properties and community structure
of soil macroinvertebrates in oak-hickory forests along an acidic deposition
gradient. Applied Soil Ecology 4, 125-137 pp.
Lafage, J. P., Nutting, W. L., 1978. Nutrient dynamics of termites. in Brian, M. V., (eds).
Production and ecology of ants and termites. Cambridge University Press, 165-
232 pp.

Références bibliographiques.
257
Lal, R., 1988. Effects of macrofauna on soil properties in tropical ecosystems. Agriculture,
Ecosystems and Environment 24, 101-116 pp.
Laurance, W. F., Fearnside, P. M., 1999. Amazon burning. Tree 14, 457 pp.
Lavelle, P. 1971. Etude démographique et dynamique des populations de Millsonia
anomala (Acanthodrilidae-Oligochètes). Doctorat 3ème cycle. Faculté des Sciences
de Paris, Paris,88 pp.
Lavelle, P., 1983. The structure of earthworm communities. in Satchell, J. E., (eds).
Earthworm Ecology: from Darwin to Vermiculture. . Chapman & Hall, London, UK,
449-466 pp.
Lavelle, P., 1996. Diversity of Soil Fauna and Ecosystem Function. Biology International
33, 3-16 pp.
Lavelle, P., 1997. Faunal activities and soil processes : adaptive strategies that
determine ecosystem function. Advances in Ecological Research 27, 93-132 pp.
Lavelle, P., 2000. Macro-Invertébrés du sol: des intestins de la terre aux organismes
ingénieurs. Comptes Rendus de l'Academie d' Agriculture France 86, 121-127 pp.
Lavelle, P., 2002. Functional domains in soils. Ecological Research 17, 441–450 pp.
Lavelle, P., Bignell, D., Lepage, M., Wolters, V., Roger, P., Ineson, P., Heal, O. W.,
Dhillion, S., 1997. Soil function in a changing world: the role of invertebrate
ecosystem engineers. Eur J Soil Biol 33, 159-193 pp.
Lavelle, P., Blanchart, E., Martin, A., Martin, S., Spain, A., Toutain, F., Barois, I.,
Schaefer, R., 1993. A hierarchical model for decomposition in terristrial
ecosystems : application to soils of the humid tropics. Biotropica 25, 130-150 pp.
Lavelle, P., Brussaard, L., Hendrix, P. F., 1999. Earthworm Management in Tropical
Agroecosystems. CAB International,, New York, 300 pp.
Lavelle, P., Dangerfield, J. M., Fragoso, C., Eschenbrenner, V., Lopez-Hernandez, D.,
Pashanasi, B., Brussaard, L., 1994. The relationship between soil Macrofauna and
tropical soil fertility. in Swift, M. J., Woomer, P., (eds). Tropical Soil Biology and
Fertility. John Wiley-Sayce, New York, 137-169 pp.
Lavelle, P., Lattaud, C., Trigo, D., Barois, I., 1995. Mutualism and biodiversity in soils. in
H.P. Collins, G. P. R. e. M. J. K. e., (eds). The significance and regulation of soil
biodiversity. Kluwer Academic Publisher, 23-33 pp.
Lavelle, P., Melendez, G., Pashanasi, B., Schaefer, R., 1992a. Nitrogen mineralization and
reorganization in casts of the geophogous tropical earthworm pontoscolex
corethurus (glossoscolecidae). Biology and Fertility of Soils 14, 49-53 pp.
Lavelle, P., Spain, A. V., 2001. Soil Ecology. Kluwer Scientific Publications, Amsterdam,
Lavelle, P., Spain, A. V., Blanchart, E., Martin, A., Martin, S., 1992b. The impact of soil
fauna on the properties of soils in the humid tropics. in Lal, R., Sanchez, P. A.,

Références bibliographiques.
258
(eds). Myths and Science of Soils of the Tropics. SSSA Special Publication,
Madison, Wisconsin, 157-185 pp.
Laverack, M. S., 1961. Tactile and chemical perceptions in earthworms. II. Responses to
acid pH solutions. Comparative Biochemistry and Physiology 2, 22-34 pp.
Le Bayon, R. C., Binet, F., 1999. Rainfall effects on erosion of earthworm casts and
phosphorus transfers by water runoff. Biology and Fertility of Soils 30, 7-13 pp.
Le Bissonnais, Y., Souder, C., 1995. Mesurer la stabilité structurale des sols pour évaluer
leur sensibilité à la battance et à l'érosion. Etude et Gestion des Sols 2, 43-55 pp.
Le Tourneau, F.-M., 2004. Jusqu'au bout de la forêt? Causes et mécanismes de la
déforestation en Amazonie brésilienne. M@ppemonde 75,
Lee, K. E., 1985. Eartworms, their ecology and relationships with soils and land use. in.
Academic Press, Sydney, 411 pp.
Lee, K. E., Butler, J. H. A., 1977. Termites, soil organic matter decomposition and
nutrient cycling. Ecological Bulletin 25, 544-548 pp.
Lee, K. E., Foster, R. C., 1991. Soil fauna and soil structure. Australian Journal of soil
Research 29, 745-775 pp.
Lee, K. E., Wood, T. G., 1971. Physical and chemical effects on soils of some Australian
termites, and their pedological significance. Pedobiologia 11, 376-409 pp.
Legendre, P., Legendre, L., 1984. Ecologie numérique : Le traitement multiple des
données écologiques, 2nd 'edition'. Presse de l'université du Quebec, Quebec, 260
pp.
Legendre, P., Legendre, L., 1998. Numerical Ecology. Elsevier, Amsterdam,
Boston,London, New York, San Fransico, Singapore, Sydney,Tokyo.,
Lepage, M. 1974. Les termites d'une savanne sahélienne (Ferlo septentrional, Sénégal):
peuplements, populations, consommation, rôle dans l'écosystème. Sciences,
Paris,344 pp.
Lepage, M., 1979. La récolte en strate herbacée de Macrotermes aff. subhyalinus
(Isoptera: Macrotermitinae) dans un écosystème semi-aride (Kajiado-Kenya).
Cahier de Recherche UIEIS société française- Lausanne 7-8 sept.,
Lepage, M., 1984. Distribution, density and evolution of macrotermes bellicious nests
(Isoptera : Macrotermitinae) in the North-East of Ivory Coast. J. Anim. Ecol. 53,
107-117 pp.
Lepage, M., Tano, Y., 1986. Les termitières épigées d'un bassin versant en zone
soudanienne : premiers résultats obtenus. Actes des colloques insectes sociaux 3,
133-142 pp.
Lessa, A. S. N., Anderson, D. W., Moir, J. O., 1996. Fine root mineralization soil organic
matter and exchangeable cation dynamics in slash and burn agriculture in the
semi-arid northeast of Brazil. Agri. Ecosys. Envir. 59, 191-202 pp.

Références bibliographiques.
259
Lobry De Bruyn, L. A., Conacher, A. J., 1990. The role of termites and ants in soil
modification : A review. Australian Journal of soil Research 28, 55-93 pp.
Loreau, M., 1998. Biodiversity and ecosystem functioning: a mechanistic model. Proc.
Natl. Acad. Sci. USA 95, 5632-5636 pp.
Loreau, M., Naeem, S., Inchausti, P., Bengtsson, J., Grime, J. P., Hector, A., Hooper, D.
U., Huston, M. A., Raffaelli, D., Scmid, B., Tilman, D., Wardle, D. A., 2001.
Biodiversity and Ecosystem Functioning: Current Knowledge and Future
Challenges. Science 294, 804-808 pp.
Lotwick, H. W., Silverman, B. W., 1982. Methods for analysing spatial processes of
several types of points. Journal of the Royal Statistical Society series B 44, 403-
413 pp.
Maldague, M. E., 1959. Analyse des sols et des matériaux des termitières du Congo
belge. Insectes Sociaux 6, 343-359 pp.
Martin, A., 1991. Short- and long-term effects of the endogeic earthworm Millsonia
anomala (Omodeo) (Megascoleidae, Oligochaeta) of tropical savannas, on soil
organic matter. Biology and Fertility of Soils 11, 234-238 pp.
Martin, A., Cortez, J., Barois, I., Lavelle, P., 1987. Les mucus intestinaux de Ver de
Terre, moteur de leurs interactions avec la microflore. Rev. Ecol. Biol. Sol 24,
549-558 pp.
Martinez, L. J., Zinck, J. A., 2004. Temporal variation of soil compaction and deterioration
of soil quality in pasture areas of Colombian Amazonia. Soil & Tillage Research 75,
3-17 pp.
Mathieu, C., Pieltain, F., 1998. Analyse physique des sols. Méthodes choisies, Londres,
275 pp.
Mathieu, C., Pieltain, F., 2003. Analyse chimique des sols. Méthodes choisies, Londres,
387 pp.
Mathieu, J. 2004. Etude de la macrofaune du sol dans une zone de déforestation en
Amazonie du Sud-Est, au Bresil, dans le contexte de l'agriculture familiale. Paris 6
Jussieu, Paris,237 pp.
Mboukou, I., Lavelle, P. 1997. Les macroinvertébrés du sol dans différents systèmes
d'agriculture au Congo : cas particuluier de deux systèmes traditionnels
(ecobuage et brûlis) dans la vallée du Niari. Université de Paris 06, Paris,171 pp.
McInerney, M., Bolger, T., 2000. Temperature, wetting cycles and soil texture effects on
carbon and nitrogen dynamics in stabilized earthworm casts. Soil Biology and
Biochemistry 32, 335-349 pp.
Mills, A. J., Fey, M. V., 2004. Frequent fires intensify soil crusting: physicochemical
feedback in the pedoderm of long-term burn experiments in South Africa.
Geoderma 121, 45–64 pp.

Références bibliographiques.
260
Ministério do Meio Ambiente (MMA). 1998. National Report to the Biological Convention
on Biological Diversity. 283, Ministery of Environment, Brazil.
Mora, P., Miambi, E., Jimenez, J. J., Decaens, T., Rouland, C., 2005. Functional
complement of biogenic structures produced by earthworms, termites and ants in
the neotropical savannas. Soil Biology and Biochemistry 37, 1043-1048 pp.
Mora, P., Seugé, C., Chotte, J.-L., Rouland, C., 2003. Physico-chemical typology of the
biogenic structures of termites and earthworms: a comparative analysis. Biology
and Fertility of Soils 37, 245-249 pp.
Moran, C. J., Koppi, A. J., Murphy, B. W., McBratney, A. B., 1988. Comparison of the
macropore structure of a sandy loam surface soil horizon subjected to two tillage
treatments. Soil Use Management 4, 96-102 pp.
Moreau, R., 1983. Evolution des sols sous différents modes de mise en culture, en Côte
d'Ivoire forestière et préforestière Cahiers - ORSTOM. Série pédologie 20, 311-
325 pp.
Murray, I., Williams, P. C., 1990. Chemical principles of Near Infrared Technology. in
Williams, P., Norris, K., (eds). Near-Infrared technology in the Agricultural and
food industries. American Association of Cereal Chemists, Inc, St. Paul, Minnesota,
USA, 17-34 pp.
Myers, N., Mittermeier, R. A., Mittermeier, C. G., Da Fonseca, G. A. B., Kent, J., 2000.
Biodiversity hotspots for conservation priorities. Nature 403, 853-858 pp.
Myot, C., 1957. Etudes de la glande Morren chez deux oligochètes lombricidés. Archives
de zoologie expérimentale et générale 94, 61 - 87 pp.
Ndiaye, D. 2003. Impact des structures biogéniques dans l’activité et la diversité des
communautés microbiennes liées au cycle de l’azote en zone soudano-sahélienne.
Ecologie. Université Pierre et Marie Curie
Paris,216 pp.
Nishita, H., Haug , R. M., 1972. Some physical and chemical characteristics of heated
soil. Soil Science 113, 422-430 pp.
Nooren, C. A. M., Van Breemen, N., Stoorvogel, J. J., Jongmans, A. G., 1995. The role of
earthworms in the formation of sandy surface soils in a tropical forest in Ivory
Coast. Geoderma 65, 135-148 pp.
Norgrove, L., Nkem, J. N., Hauser, S., 2003. Separating the agronomic effects of burning
and biomass removal in a typical slash and burn rotation system in southern
Cameroon. Biological Agriculture & Horticulture 21, 367-381 pp.
Nye, P. H., 1955. Some soil-forming processes in the humid tropics IV. The action of the
soil fauna. Journal of Soil Science 6, 73-83 pp.

Références bibliographiques.
261
Nzila, J. d. D., Garnier-Zarli, E. 1992. Study of the structural and physico-chemical
transformations in a ferralitic acid soil in the Niari Valley (Congo) submitted to the
practice of burn-beating. Université de Paris 12, Créteil, FRANCE,190 pp.
Obale-Ebanga, F., Sevink, J., de Groot, W., Nolte, C., 2003. Myths of slash and burn on
physical degradation of savannah soils: impacts on Vertisols in north Cameroon.
Soil Use and Management 19, 83-86 pp.
Paine, R. T., 1969. A note on trophic complexity and species diversity. American
Naturalist 100, 91-93 pp.
Pasini, A., Fonseca, I. C. B., Brossard, M., Guimarães, M. F. 2000 Soil macrofauna under
pastures in the Cerrado of Uberlândia-MG, Brazil.
Pelosi, C. 2005. Caractérisation des agrégats produits par les invertébrés du sol dans
l’Est Amazonien. . 38, Université Paris 6 / IRD, Paris.
Pereira, W. 1977. Avaliaçao da erosividade das chuvas em diferentes locais do Estado de
Minas Gerais. Universidade Federal de Viçosa, Viçosa, Minas Gerais, Brasil
Pérès, G., Cluzeau, D., Curmi, P., Hallaire, V., 1998. Earthworm activity and soil
structure changes due to organic enrichments in vineyard systems. Biology and
Fertility of Soils 27, 417-424 pp.
Pieri, C., 1989. Fertilité des terres de savanne. CIRAD et Ministère de la Coopération et
du Développement, Montpellier, 444 pp.
PNUE-CEPALC. 2001. The Sustainability of Development in Latin America and the
Caribbean : challenges and opportunities., Santiago (Chili).
Pulleman, M. M., Six, J., Uyl, A., Marinissen, J. C. Y., Jongmans, A. G., 2005. Earthworms
and management affect organic matter incorporation and microaggregate
formation in agricultural soils. Applied Soil Ecology 29, 1-15 pp.
R Development Core Team, 2005. R : A language and environment for statistical
computing. .http://www.R-project.org.
Raij, B. v., 1969. Capacidade de troca de fraçoes organica e minerais dos solos.
Bragantia 28, 85-112 pp.
Ramade, F., 1994. Elément d'Ecologie : écologie fondamentale, Paris, 579 pp.
Redford, K. H., 1984. The termitaria of Cornitermes cumulans (Isoptera, Termitidae) and
their role in determining a potential keystone species. Biotropica 16, 112-119 pp.
Reeves, J. B., McCarty, G. W., Meisinger, J. J., 1999. Near-Infrared reflectance
spectroscopy for the analysis of agricultural soil. Journal of Near Infrared
Spectroscopy 7, 179-193 pp.
Reynal, V. d., Muchagata, M. G., Topall, O., Hebette, J., 1997. Des paysans en
Amazonie: agriculture familiale de développement du front pionnier amazonien. in
Théry, H., (eds). Environnement et Développment en Amazonie Brésilienne. Belin,
Paris, 76-123 pp.

Références bibliographiques.
262
Ringrose-Voase, 1991. Micromorphology of soil structure : Description, quantification,
application. Australian Journal of Soil Science 29, 777-813 pp.
Ripley, B. D., 1977. Modelling Spatial Patterns. Journal of the Royal Statistical Society
series B 39, 172-212 pp.
Ripley, B. D., 1981. Spatial statistics. John Wiley & Sons., New York, 255 pp.
Rossi, J. P., 2003. The spatiotemporal pattern of a tropical earthworm species
assemblage and its relationship with soil structure. Pedobiologia 47, 497-503 pp.
Rossi, J. P., Nuutinen, V., 2004. The effect of sampling unit size on the perception of the
spatial pattern of earthworm (Lumbricus terrestris L.) middens. Applied Soil
Ecology 27, 189–196 pp.
Rouland-Lefèvre, C., 2000. Symbiosis with fungi. in Abe, T., Bignell, D., Higashi, M.,
(eds). Termites: evolution, sociality, symbioses, ecology. Kluwer Academic
publishers, Dordrecht, 289–306 pp.
Sarr, M., Agbogba, C., Russell-Smith, A., Masse, D., 2001. Effects of soil faunal activity
and woody shrubs on water infiltration rates in a semi-arid fallow of Senegal.
Applied Soil Ecology 16, 283-290 pp.
Scherrer, B., 1984. Biostatistiques, Montréal, 850 pp.
Schrader, S., Zhang, H., 1997. Earthworm casting: Stabilization or destabilization of soil
structure? Soil Biology and Biochemistry 29, 469-475 pp.
Scott Mills, L., Soule, M. E., Doak, D. F., 1993. The Keystone-Species Concept in Ecology
and Conservation Biosciences 43, 219-224 pp.
Shipitalo, M. J., Protz, R., 1989. Chemistry and micromorphology of aggregation in
earthworm casts. Geoderma 45, 357-374 pp.
Sleeman, J. R., Brewer, R., 1972. Microstructures of some Australian termite nests.
Pedobiologia 12, 347-373 pp.
Smeraldi, R., Verissimo, J. A. d. O., 1999. Acertando o alvo: consumo de madeira no
mercado interno brasileiro e promoção da certificação florestal. : Amigos da Terra,
Programa Amazônia, São Paulo, Piracicaba: IMAFLORA; Belém: IMAZON.,
Soltner, D., 1994. Les bases de la production végétale, Phytotechnie Générale 'edition'.
Le clos Lorelle, Angers, 467 pp.
Stoops, G., 1964. Application of some pedological methods to the analysis of termite
mounds. in Bouillon, A., (eds). Etudes sur le termite africain. Masson, Paris, 379-
388 pp.
Stoyan, D., Kendall, W. S., Mecke, J., 1987. Stochastic geometry and its applications.,
New-York, 345 pp.
Tanaka, S., Funakawa, S., Kaewkhongkha, T., Yonebayashi, K., 1998. Labile pools of
organic matter and microbial biomass in the surface soils under shifting cultivation
in northern Thailand. Soil Science and Plant Nutrition 44, 527-537 pp.

Références bibliographiques.
263
Tano, Y., Lepage, M., 1990. Les termites: dynamique des nids épigés et interactions avec
les composantes du milieu. in Orstom, C., (eds). Etudes et Thèses, 105-116 pp.
Thioulouse, J., Chessel, D., Dolédec, S., Olivier, J. M., 1997. ADE-4: a multivariate
analysis and graphical display software. Statistics and Computing 7, 75-83 pp.
Tognon, A. A., Demattê, J. L. I., Demattê, J. A. M., 1998. Tear e distribuçao da materia
norganica em Latossolos das regioes da floresta amzônica e dos cerrados de Brasil
central. Scientia Agricola 55, 343-354 pp.
Topoliantz, S., Ponge, J.-F., 2003. Burrowing activity of the geophagous earthworm
Pontoscolex corethrurus (Oligochaeta: Glossoscolecidae) in the presence of
charcoal. Applied Soil Ecology 23, 267-271 pp.
Toutain, F., 1981. Activite biologique des sols, modalités et lithodépendance Biology and
Fertility of Soils 3, 31-38 pp.
Velasquez, E., Lavelle, P., Barrios, E., Joffre, R., Reversat, F., 2005. Evaluating soil
quality in tropical agroecosystems of Colombia using NIRS. Soil Biology and
Biochemistry 37, 889-898 pp.
Vitrac, M. 2001. Analyse à court terme et indicateurs des dégradations de pâturages
cultivés de la savane brésilienne. Mémoire de fin d’études 75, ISTOM: IRD:
Embrapa Cerrados, Planaltina, DF.
Wanner, M., Dunger, W., 2002. Primary immigration and succession of soil organisms on
reclaimed opencast coal mining areas in eastern Germany. European Journal of
Soil Biology 38, 137-143 pp.
Warner, K., 1995. Agriculteurs itinérants : Connaissances techniques locales et gestion
des ressources naturelles en zone tropicale humide. FAO, 81 pp.
Williams, P. C., 1990. Variables affecting near infrared reflectance spectroscopic analysis.
in Williams, P., Norris, K., (eds). Near-Infrared technology in the Agricultural and
food industries. American Association of Cereal Chemists, Inc, St. Paul, Minnesota,
USA, 143-167 pp.
WRBSR, 1994. World Reference Base for Soil Resources, Wageningen / Rome, 161 pp.

Liste des illustrations.
264
Liste des illustrations.

Liste des illustrations.
265
Tableaux
• Tableau 1 :Classement de la faune par taille (critère théorique) (Bachelier, 1978). ..... 20 • Tableau 2 : Classement de la faune par régime alimentaire (critère écologique). ........ 21 • Tableau 3 : Répartition des parcelles étudiées en fonction du couvert végétal et
de l’exploitant. .................................................................................................. 44 • Tableau 4 : Répartition en pourcentage de masse des agrégats obtenus suite aux
tests de stabilité structurale des sols témoins sous forêt, forêt dégradée et sous pâturage selon la méthode de Le Bissonnais (T1 = Immersion, T2 = Réhumectation, T3 = Dispersion mécanique après prétraitement à l’alcool, T4 = Dispersion mécanique après prétraitement dans le Kérosène). ................................. 73
• Tableau 5 : Comparaison de la concentration en carbone (mg.g-1) à différentes profondeurs entre les sols témoins des deux milieux : forêts et pâturages (±déviation standard). ....................................................................................... 76
• Tableau 6 : Comparaison des pH (pH H2O) à différentes profondeurs entre les sols témoins des trois milieux forêts, forêt dégradée et pâturages (± déviation standard) ......................................................................................................... 77
• Tableau 7 : Comparaison des concentrations en cations aluminium (Al3+), calcium (Ca2+), magnésium, potassium (K+) et somme des cations basiques S (S= [Ca2+] + [Mg2+] + [K+]) entre les sols des trois milieux (forêts, forêt dégradée et pâturages) à différentes profondeurs.................................................................... 78
• Tableau 8 : Comparaison de la somme des cations basiques S (S= [Ca2+] + [Mg2+] + [K+]), de la CEC et du taux de saturation en cations (V) entre les horizons (0-30 cm) des les forêts, et la foret dégradée. .......................................... 79
• Tableau 9 : Comparaison de la somme des cations basiques S (S= [Ca2+] + [Mg2+] + [K+]), de la CEC et du taux de saturation en cations (V) entre les horizons (0-30 cm) des pâturages. ...................................................................... 80
• Tableau 10 : Comparaison des concentrations en phosphore assimilable (P) entre les horizons des trois milieux (forêts, forêt dégradée et pâturages) à différentes profondeurs (0-30 cm). ...................................................................................... 81
• Tableau 11 : Données quantitatives de la densité de termitières de Cornitermes sp. dans les trois pâturages. ............................................................................... 88
• Tableau 12 : La nature de la distribution spatiale des termitières (agrégé, aléatoire, inhibé) est indiquée aux trois échelles (0-15m, de 15 à 50 m, >50m) dans trois pâturages (P01, P02 et P06). Les résultats sont issus de l’analyse de la Fonction L(r) de Ripley. ...................................................................................... 91
• Tableau 13 : Distribution spatiale des termitières du genre Cornitermes sp. dans les deux parties du pâturage P01 (P01 haut et P01 bas). La nature de la distribution spatiale (aléatoire, agrégée ou inhibée) est mentionnée aux trois échelles spatiales (0 à 15m, 15m à 50m et supérieur à 50m). les résultats sont issus de l’analyse de la fonction L de Ripley........................................................... 92
• Tableau 14 : Localisation (nom du propriétaire de l’exploitation) et type de couvert végétal des diverses parcelles étudiées pour cartographier les turricules d’Andiodrilus pachoensis................................................................................... 126
• Tableau 15 : Données quantitatives de la densité des turricules dans les cinq forêts étudiées au cours des deux échantillonnages. ............................................. 130
• Tableau 16 : Données quantitatives de la densité des turricules dans la forêt dégradée F01 étudiées au cours des deux échantillonnages. .................................. 131
• Tableau 17 : Données quantitatives de la densité des turricules dans la jachère étudiée. ......................................................................................................... 131
• Tableau 18 : Données quantitatives de la densité des turricules dans les six pâturages étudiés au cours des deux échantillonnages.......................................... 132

Liste des illustrations.
266
• Tableau 19 : Données quantitatives de la densité des turricules dans la jachère J01................................................................................................................ 132
• Tableau 20 : Données quantitatives de la masse des turricules dans les cinq milieux étudiés (forêts, forêt dégradée, jachère, rizière et pâturages). .................... 134
• Tableau 21 : Densité des turricules d’Andiodrilus pachoensis produits dans 25 m² aux dates J1 (+ 28 Jours/J0) et J2 (+ 66 Jours/J0) dans la forêt dégradée F01, la forêt F02 et trois pâturages P01, P02 et P03. ................................................... 135
• Tableau 22 : Distance minimum et distance moyenne entre deux turricules dans les forêts (F01, F02, F03 et F04), la jachère (J01), les pâturages (P0, P02, P03, P04 et P05) et la rizière (R01). .......................................................................... 138
• Tableau 23 : Distribution spatiale des turricules d’Andiodrilus pachoensis dans les parcelles des différents milieux : 4 forêts (F01 à F04), 1 jachère (J01), 1 rizière (R01) et 3 pâturages (P01 à P04). La nature de la distribution spatiale (agrégée, aléatoire ou régulière) est mentionnée aux trois échelles spatiales (0 à 20cm, de 20 cm à 150 cm et supérieur à 150cm). Les résultats sont issus de l’analyse de la fonction L de Ripley. ........................................................................................ 139
• Tableau 24 : Distributions spatiales des turricules de l’annélide Andiodrilus pachoensis dans les parcelles de forêts (F01 et F02) et 3 pâturages (P01, P03 et P04) aux différentes dates d’échantillonnage J0, J1 (28 jours après J0) et J2 (65 jours après J0). La nature de la distribution spatiale (agrégée, aléatoire ou inhibée) est mentionnée aux trois échelles spatiales (0 à 20cm, de 20 cm à 150 cm et supérieur à 150cm). Les résultats sont issus de l’analyse de la fonction L de Ripley........................................................................................................ 142
• Tableau 25 : Distributions spatiales des turricules de l’annélide Andiodrilus pachoensis dans les forêts (F01 à F04) en fonction de la position des autres structures biogéniques (fonction intertype). La nature de la distribution spatiale (agrégée, aléatoire ou inhibée) est mentionnée aux trois échelles spatiales (0 à 20cm, de 20 cm à 150 cm et supérieur à 150cm). Les résultats sont issus de l’analyse de la fonction L de Ripley..................................................................... 143
• Tableau 26 : Distributions spatiales des turricules de l’annélide Andiodrilus pachoensis dans les forêts (F01 à F04) en fonction de la position des arbres (fonction intertype). La nature de la distribution spatiale (agrégée, aléatoire ou inhibée) est mentionnée aux trois échelles spatiales (0 à 20cm, de 20 cm à 150 cm et supérieur à 150cm). Les résultats sont issus de l’analyse de la fonction L de Ripley........................................................................................................ 144
• Tableau 27 : Evaluation du Carbone stocké dans les turricules d’Andiodrilus pachoensis dans les forêts et les pâturages en fonction des estimations de masse de turricules dans chaque milieu........................................................................ 155
• Tableau 28 : Bilan moyen estimée des ions échangeables par hectare et par an dans les turricules d’Andiodrilus pachoensis produits des trois milieux (pâturage et forêt). ........................................................................................................ 162
• Tableau 29 : Classement des poroïdes en fonction de leur taille aux 2 échelles d’observation. ................................................................................................. 189
• Tableau 30 : Type de poroïdes en fonction de l’indice de forme. ............................. 189
Figures
• Figure 1 : Carte de la déforestation 2002/2003, Forêt amazonienne brésilienne (INPE, 2004). ................................................................................................... 14
• Figure 2 : Place de la faune et de ses structures biogéniques dans le fonctionnement et l’évolution des sols et des agrosystèmes (d’après (Lavelle, 2000) .............................................................................................................. 23
• Figure 3 : Cycle du carbone (Ramade, 1994)......................................................... 32

Liste des illustrations.
267
• Figure 4 : Cycle du phosphore (Ramade, 1994) ..................................................... 34 • Figure 5 : Localisation de l’assentamento « Benfica » au Brésil................................. 40 • Figure 6 : Données climatiques de Marabá et Benfica.............................................. 41 • Figure 7 : Disposition des parcelles étudiées dans « l’assentamento » de Benfica
en 2002 d’après (Mathieu, 2004). ....................................................................... 45 • Figure 8 : Trois types possibles de distribution spatiale d’organismes dans une
population ........................................................................................................ 49 • Figure 9 : Exemple d’un patron hétérogène (a) (deux régions de densité
différentes) ; un patron anisotropique (b) (une direction privilégiée) ; un patron homogène isotropique (c) (patron de poisson)....................................................... 51
• Figure 10 : Méthode de calcul de la fonction K(r) de Ripley...................................... 52 • Figure 11 : Estimations de la fonction L(r) pour plusieurs patrons de points. .............. 52 • Figure 12 : Exemple de résultat de la courbe L(r). La courbe 1 correspond à la
fonction estimée à partir du jeu de données et les courbes 2 et 3 correspondent aux bornes de l’intervalle de confiance à 99% pour l’hypothèse nulle d’une distribution spatiale aléatoire (distribution de poisson)............................................ 53
• Figure 13 : Exemple de résultat de la courbe L12(r). La courbe 1 correspond à la fonction estimée pour le jeu de données initial. Les courbes 2 et 3 correspondent aux bornes de l’intervalle de confiance à 99% de l’hypothèse nulle de la distribution spatiale aléatoire de l’objet étudié en fonction de la position des points d’une autre variable qui reste inchangée...................................................... 55
• Figure 14 : Profil pédologique (0-2m) réalisé dans la forêt de Sr Osano (F01). .......... 63 • Figure 15 : Profil pédologique ( 0-2m) réalisé dans la forêt de Sr Melete (F02). .......... 64 • Figure 16 : Profil pédologique (0-2m) réalisé dans la forêt de Sr Léonine (F03). ......... 65 • Figure 17 : Profil pédologique (0-2m) réalisé dans le pâturage de Sr Osano (P01). ..... 67 • Figure 18 : Profil pédologique (0-2m) réalisé dans le pâturage de Sr Josuel (P02
– P03). ............................................................................................................ 68 • Figure 19 : Profil pédologique (0-2m) réalisé dans le pâturage de Sr Léonine
(P04)............................................................................................................... 69 • Figure 20 : Disposition des termitières dans les 3 pâturages (P01, P02, P06). A
chaque carré correspond une termitière de taille et d’âge non précisés...................... 89 • Figure 21 : Distribution de fréquence des termitières selon 3 classes de hauteur
(grande : >1m ; moyenne ; petite <60cm). .......................................................... 90 • Figure 22 : Extrapolation de la densité des nids de Cornitermes sp. en fonction de
l’âge des pâturages. .......................................................................................... 90 • Figure 23 : Image pixel des estimations d’intensités lissées (méthode Kernel) des
patrons de termitières dans les trois parcelles. Les zones de faible densité en termitières sont mises en évidence dans les zones rouges. ...................................... 91
• Figure 24 : Densité des termitières Cornitermes sp. en fonction du rayon de prospection considéré. Parcelles P05 et P02 : (de 5 à 20m pour les graphiques 1 à 4). et représentation topographique des parcelles par Krigeage de la médiane et amplification de la différence avec les bas fonds................................................ 94
• Figure 25 : Masse de feuilleté des termitières de Cornitermes sp. et régression suivant leur classe de hauteur (grande, moyenne, petite, morte). ............................ 96
• Figure 26 : Comparaison de la texture des murailles de termitière Cornitermes sp avec celle des sols (jusqu’à 2,2 m) des trois pâturages P01, P03 et P04. ................... 98
• Figure 27 : Comparaison des différentes classes d’agrégats obtenus par tamisage et diamètre moyen des particules après les 4 traitements de la muraille de termitière de Cornitermes sp. et du sol témoin (10-20 cm).................................... 100
• Figure 28 : Comparaison de la teneur en carbone dans la muraille de termitière et les horizons témoins le long d’un profil jusqu’à 30 cm de profondeur dans deux pâturages....................................................................................................... 102

Liste des illustrations.
268
• Figure 29 : Comparaison du pH des murailles des termitières avec celui des horizons témoins jusqu’à 30 centimètre de profondeur dans deux pâturages............ 103
• Figure 30 : Comparaison des concentrations en cations aluminium échangeable (Al3+) dans les murailles des termitières de Cornitermes sp. et le profil de concentration moyenne des horizons témoins (jusqu’à 25 cm de profondeur) de trois pâturages................................................................................................ 104
• Figure 31 : Comparaison des concentrations en cations calcium (Ca2+), magnésium (Mg2+) dans les murailles des termitières de Cornitermes sp. et des horizons témoins de trois pâturages. .................................................................. 104
• Figure 32 : Comparaison des concentrations en cations potassium (K+) dans les murailles des termitières de Cornitermes sp. et des horizons témoins de trois pâturages....................................................................................................... 105
• Figure 33 : Comparaison des concentrations en phosphore assimilable dans les murailles des termitières de Cornitermes sp. et des profils de concentration moyenne des horizons témoins de trois pâturages................................................ 106
• Figure 34 : Comparaison de la texture des murailles des termitières de Cornitermes sp exposées au feu avec celles des murailles natives et de l’horizon témoin (10 -30 cm) dans trois pâturages P01, P03 et P04. .................................... 108
• Figure 35 : Comparaison des différentes classes d’agrégats obtenus par tamisage et diamètre moyen de ces agrégats après les 4 traitements des murailles des termitières de Cornitermes sp. dans différents états : natif (n), brûlés expérimentalement (exp) ou brûlée par un paysan (pay)(nids morts) et des horizons témoins (0-30 cm) des pâturages P01, P03 et P04................................... 109
• Figure 36 : Comparaison des concentrations en carbone dans les murailles des termitières épigées dans différents états : natif (n) ou brûlé expérimentalement (exp) avec celle des horizons témoins (10-30 cm de profondeur) des pâturages P01, P03 et P04. ............................................................................................. 111
• Figure 37 : Comparaison du pH de la muraille de termitière de Cornitermes sp dans son état natif ou brûlée et du sol témoin moyen des 10 premiers centimètres de profondeur................................................................................ 112
• Figure 38 : Comparaison des concentrations en cations aluminium échangeable (AL3+), calcium (Ca2+), magnésium (Mg2+), potassium, (K+) entre les murailles des termitières de Cornitermes sp. dans différents états : natif ou brûlé expérimentalement et des horizons témoins (10-30 cm) des 3 pâturages (P01, P03 et P04). ................................................................................................... 113
• Figure 39 : Comparaison des concentrations en phosphore assimilable entre les murailles des termitières de Cornitermes sp. dans différents états : natif ou brûlé expérimentalement et des horizons témoins (10-30 cm) des 3 pâturages (P01, P03 et P04). ................................................................................................... 114
• Figure 40 : Comparaison de la densité moyenne des turricules par m² dans les cinq milieux (forêt, forêt dégradée, jachère, rizière et pâturage). .......................... 133
• Figure 41 : Production de turricules en fonction de la quantité de litière (normal : +/+ ; réduite : +/- ; faible : -/-) dans la forêt dégradée F01 (gauche) et la forêt F03 (droite). A J0 correspond la première date d’observation, à J1, 28 jours après J0 et J2, 66 jours après J0 ................................................................................ 145
• Figure 42 : Évolution de la quantité moyenne de turricules en fonction du temps au bout de 26 jours (J1) et 64 jours (J2) en relation avec la présence de touffes de Brachiaria brisantha (témoin) ou l’absence d’herbacé (sol nu)............................ 146
• Figure 43 : Distribution des turricules et des températures du sol dans un quadrat (3 x 3 m) du pâturage P01. La distribution des turricules obtenue semble montrer une préférence des vers pour les zones plus fraîches. ........................................... 147
• Figure 44 : Comparaison de la texture des turricules de Andiodrilus pachoensis avec le profil granulométrique des deux forêts F01 (forêt dégradée) et F02. ............ 149

Liste des illustrations.
269
• Figure 45 : Comparaison de la composition granulométrique des turricules d’Andiodrilus pachoensis avec le profil granulométrique jusqu’à 220cm de profondeur de trois pâturages P01, P03 et P04. ................................................... 150
• Figure 46 : Comparaison des différentes classes d’agrégats obtenus par tamisage et diamètre moyen des particules après les 4 traitements des turricules d’Andiodrilus pachoensis et des horizons témoins (10-20 cm) dans les forêts et pâturages....................................................................................................... 152
• Figure 47 : Comparaison de la teneur en carbone dans les turricules de forêt (A), de forêt dégradée (B) et de pâturage (C) avec des profils de concentration jusqu’à 30 centimètres de profondeur. ............................................................... 154
• Figure 48 : Comparaison du pH dans les turricules de forêt (A), de forêt dégradée (B) et de pâturage (C) avec des profils de concentration jusqu’à 30 centimètres de profondeur. ................................................................................................ 156
• Figure 49 : Comparaison des concentrations en cations aluminium (AL3+) entre des échantillons de turricules de Andiodrilus pachoensis et des horizons témoins des forêts, forêts dégradée et pâturages (0-30cm)............................................... 157
• Figure 50 : Comparaison des concentrations en cations calcium (Ca2+) entre des échantillons de turricules de Andiodrilus pachoensis et des horizons témoins des forêts, forêt dégradée et pâturages (0-30cm). .................................................... 158
• Figure 51 : Comparaison des concentrations en cations magnésium (Mg2+) entre des échantillons de turricules de Andiodrilus pachoensis et des horizons témoins des forêts, forêt dégradée et pâturages (0-30cm). .............................................. 159
• Figure 52 : Comparaison des concentrations en cations potassium (K+) entre des échantillons de turricules de Andiodrilus pachoensis et des horizons témoins des forêts et forêt dégradée (0-30cm)..................................................................... 160
• Figure 53 : Comparaison des concentrations en cations potassium (K+) entre des échantillons de turricules d’Andiodrilus pachoensis et des horizons témoins pâturages (0-30cm)......................................................................................... 160
• Figure 54 : Comparaison des concentrations phosphore assimilable (P) entre des échantillons de turricules de Andiodrilus pachoensis et des horizons témoins des forêts, forêt dégradée et pâturages (0-30cm). .................................................... 161
• Figure 55 : Modification de la composition granulométrique des turricules de Andiodrilus pachoensis suite à l’exposition plus ou moins forte au feu (2 intensités : +/- et +/+) et comparaison avec celle des horizons témoins (0 -10 cm) dans trois forêts F01, F02 et F03. ............................................................... 164
• Figure 56 : Modification de la texture des turricules d’Andiodrilus pachoensis suite à une exposition plus ou moins forte au feu (2 intensités : +/- et +/+), un état plusieurs fois brûlé : turricules brûlé vieux) et comparaison avec celle des horizons témoins (0 -10 cm) dans trois pâturages P01, P03 et P04. ........................ 165
• Figure 57 : Comparaison des différentes classes d’agrégats obtenus par tamisage et diamètre moyen des particules après les 2 traitements (immersion et réhumectation) des turricules dans différents états : natif, brûlés selon deux intensités (moyenne : +/- et forte +/+) et horizons témoins (0 -10 cm) des pâturages....................................................................................................... 166
• Figure 58 : Comparaison des différentes classes d’agrégats obtenus par tamisage et diamètre moyen des particules après les 2 traitements (dispersions mécaniques après prétraitement dans l’alcool et dans le kérosène des turricules dans différents états : natif, brûlés selon deux intensités (moyenne : +/- et forte +/+) et horizons témoins (0 -10 cm) des pâturages. ............................................ 166
• Figure 59 : Comparaison des différentes classes d’agrégats obtenus par tamisage et diamètre moyen des particules après les 4 traitements des turricules dans différents états : natif, brûlés selon deux intensités (moyenne : +/- et forte +/+) et horizons témoins (0 -10 cm) des forêts........................................................... 168

Liste des illustrations.
270
• Figure 60 : Comparaison des concentrations en carbone dans les turricules d’Andiodrilus pachoensis dans leur état natif, brûlés expérimentalement (deux intensités : faible +/- ou forte +/+), brûlé sur le terrain avec celles des horizons témoins. Les différences significatives sont marqués par les lettres (a), (b) et (c). .... 169
• Figure 61 : Comparaison de l’évolution du pH des turricules entre leur état frais et leur état brûlé (selon deux intensités pour les turricules et un état « vieux ») et les horizons témoins (0-30cm). ..................................................................... 170
• Figure 62 : Comparaison des concentrations en aluminium échangeable (AL3+) entre les turricules de Andiodrilus pachoensis dans différents états : natif, brûlé expérimentalement selon deux intensités (+/- et +/+) ou brûlés sur le terrain; et du sol témoin (0-20 cm) des trois forêts (F01, F02 et F03). .................................. 171
• Figure 63 : Comparaison des concentrations en cations calcium (Ca2+), magnésium (Mg2+), potassium, (K+), phosphore assimilable (P), entre les turricules de Andiodrilus pachoensis dans différents états : natif, brûlé expérimentalement selon deux intensités (+/- et +/+) ou brûlés sur le terrain; et du sol témoin (0-20 cm) des trois forêts (F01, F02 et F03). .................................. 171
• Figure 64 : Comparaison des concentrations en aluminium échangeable (AL3+) entre les turricules de Andiodrilus pachoensis dans différents états : natif, brûlé expérimentalement selon deux intensités (+/- et +/+), brûlés sur le terrain ; et du sol témoin (0-20 cm) des trois pâturages (P01, P03 et P04). ............................ 172
• Figure 65 : Comparaison des concentrations en ions calcium (Ca2+), magnésium (Mg2+), potassium, (K+), et phosphore assimilable (P), entre les turricules de Andiodrilus pachoensis dans différents états : natif, brûlé expérimentalement selon deux intensités (+/- et +/+), brûlés sur le terrain ; et du sol témoin (0-20 cm) des trois pâturages (P01, P03 et P04). ......................................................... 172
• Figure 66 : Masse perdue (%) par érosion suite à l’exposition à des pluies simulées durant différents périodes (1mn, 3mn et 5mn) des turricules d’Andiodrilus pachoensis natifs ou écrasés et du sol témoin (0-20 cm) des trois forêts (F01, F02 et F03) selon deux forces (F1 et F2). Pluie simulée de mm (Force F1 à gauche) et mm (Force F2 à droite).............................................................. 173
• Figure 67 : Masse perdue (%) par érosion suite à l’exposition à des pluies simulées durant différents laps de temps (1mn, 3mn et 5mn) des turricules d’Andiodrilus pachoensis natifs ou écrasés et du sol témoin (0-20 cm) des trois forêts (F01, F02 et F03) selon deux forces (F1 et F2). Pluie simulée de mm (Force F1 à gauche) et mm (Force F2 à droite).............................................................. 174
• Figure 68 : Cercle de corrélation (axes 1 et 2) des longueurs d’onde (à gauche) et Projection dans le plan factoriel des différentes structures biogéniques (à droite) selon les 2 premiers axes (95% de la variabilité observée). TermP : Paroi de termitière Cornitermes sp. ; TermPBrulé : Paroi de termitière Cornitermes sp.brûlé ; TermF : Feuilleté de termitière de Cornitermes sp., TurP : Turricules de Pâturages ; TurF : Turricules d’Andiodrilus de forêt ; TurBrulé : Turricules d’Andiodrilus faiblement brûlé, TurBrulé++ : Turricules d’Andiodrilus fortement brûlé P : sol témoin de pâturage ; F : sol témoin de forêt...................................... 190
• Figure 69 : Cercle de corrélation (axes 1 et 2) des éléments chimiques et du pH (à gauche) ; Projection dans le plan factoriel des différentes structures biogéniques (à droite) selon les 2 premiers axes (73% de la variabilité observée). TermP : Paroi de termitière Cornitermes sp. ; TermF : Feuilleté de termitière de Cornitermes sp., TurP : Turricules d’Andiodrilus de Pâturages ; TurF : Turricules d’Andiodrilus de forêt ; P : sol témoin de pâturage ; F : sol témoin de forêt. ............ 192
• Figure 70 : Exemples d’images de lames minces des différentes structures biogéniques : Paroi de termitière Cornitermes sp. (A) ; Feuilleté de termitière de Cornitermes sp. (B), Nid de termitière humivore (C), Placage de termitière humivore (D), Turricule d’Andiodrilus pachoensis (E) ; déblais de fourmis (F)........... 194
• Figure 71 : Porosité des structures biogéniques par analyse micromorphologique à la résolution optique de 5,88 µm par pixel (mésoporosité). Projection dans le

Liste des illustrations.
271
pan factoriel des trois types de pores : Cavités (Cav), Fissures (Fis) et pores complexes (Comp) des 5 tailles : très petit (TP), petit (P), moyen (M), grand (G) et très grand (TG) et des structures biogéniques de divers invertébrés : Turricule frais d’annélide anécique (Tur), turricule brûlé d’annélide anécique (Tur Brulé), muraille de nids épigées de termites xylophages TermCPa, feuilleté de ces termitières de xylophage (TermCFe), Paroi de termitières d’humivore (TermHNd), placage de termites humivores (TermHPl) et déblais de fourmi (Fourmi). L’axe 1 explique 65% de la variabilité et l’axe 2 : 20%. .................................................. 197
• Figure 72 : Porosité des structures biogéniques par analyse micromorphologique à la résolution optique de 0,47 µm par pixel (microporosité). Projection dans le pan factoriel des trois types de pores : Cavités (Cav), Fissures (Fis) et pores complexes (Comp) des 5 tailles : très petit (TP), petit (P), moyen (M), grand (G) et très grand (TG) et des structures biogéniques de divers invertébrés : Turricule frais d’annélide anécique (Tur), turricule brûlé d’annélide anécique (Tur Brulé), muraille de nids épigées de termites xylophages TermCPa, feuilleté de ces termitières de xylophage (TermCFe), Paroi de termitières d’humivore (TermHNd), placage de termites humivores (TermHPl) et déblais de fourmi (Fourmi). L’axe 1 explique 55% de la variabilité et l’axe 2 : 29%. .................................................. 199
• Figure 73 : Indice de diversité de Shannon (I) de la mésoporosité estimé pour toutes les structures biogéniques récoltés (gauche) et classement de ces structures en 5 classes d’indice de Shannon (Cl1 I<0,5 ; 0,5<Cl2<1 ; 1<Cl3<1,5 ; 1,5<Cl4<2 ; Cl5>2) (droite) à la résolution optique de 5,88µm par pixel. ............................................................................................................. 202
• A : Figure 74 : Indice de diversité de Shannon des poroïdes à la résolution de 5,88µm dans les différentes structures biogéniques. Tur : Turricule d’Andiodrilus pachoensis ; TurBrulé : Turricule d’Andiodrilus brûlé, TermCPa : Paroi de termitière de Cornitermes sp ; TermCFe : Feuilleté de termitière de Cornitermes sp. ; TermHNd : Nid de termites humivores ; TermHPl : Placage de termites humivores ; Fourmi : déblais de fourmis. ............................................................ 203
• B : Figure 75 : Classification des structures biogéniques en fonction de la quantification des poroïdes à la résolution de 5,88µm (matrice de distance euclidienne). Tur : Turricule d’Andiodrilus pachoensis ; TurBrulé : Turricule d’Andiodrilus brûlé, TermCPa : Paroi de termitière de Cornitermes sp ; TermCFe : Feuilleté de termitière de Cornitermes sp. ; TermHNd : Nid de termites humivores ; TermHPl : Placage de termites humivores ; Fourmi : déblais de fourmis. ......................................................................................................... 203
• A : Figure 76 : Indice de diversité de Shannon des poroïdes à la résolution de 0,47µm dans les différentes structures biogéniques. Tur : Turricule d’Andiodrilus pachoensis ; TurBrulé : Turricule d’Andiodrilus brûlé, TermCPa : Paroi de termitière de Cornitermes sp ; TermCFe : Feuilleté de termitière de Cornitermes sp. ; TermHNd : Nid de termites humivores ; TermHPl : Placage de termites humivores ; Fourmi : déblais de fourmis. ............................................................ 204
• B : Figure 77 : Classification des structures biogéniques en fonction de la quantification des poroïdes à la résolution de 5,88µm (matrice de distance euclidienne). Tur : Turricule d’Andiodrilus pachoensis ; TurBrulé : Turricule d’Andiodrilus brûlé, TermCPa : Paroi de termitière de Cornitermes sp ; TermCFe : Feuilleté de termitière de Cornitermes sp. ; TermHNd : Nid de termites humivores ; TermHPl : Placage de termites humivores ; Fourmi : déblais de fourmis. ......................................................................................................... 204
• Figure 78 : Schéma des relations entre la couverture végétale, les deux structures biogéniques étudiées (Termitières de Cornitermes sp. et turricules d’Andiodrilus pachoensis) et l’horizon superficiel 0-30 cm des sols du site d’étude. ... 212

Liste des illustrations.
272
Photos
• Photo 1 : Déforestation (2003)............................................................................ 44 • Photo 2 : Feu de maintien d’un pâturage (2003) .................................................... 44 • Photo 3 : Disposition clairsemée des touffes de Brachiaria brisantha ......................... 45 • Photo 4 : Benfica, Modelé du paysage, perspective à partir du pâturage du Sr
Osano.............................................................................................................. 62 • Photo 5 : Turricules d’Andiodrilus pachoensis....................................................... 124 • Photo 6: A : Vieux turricules brûlés d’Andiodrilus pachoensis en dégradation sous
un tronc couché dans un pâturage. B : Figures d’érosion dans les pâturages de Benfica. ......................................................................................................... 183
Illustrations
• Illustration 1 : Termitières et termites soldats du genre Cornitermes sp. ................... 85 • Illustration 2: Expérience de brûlage des termitières. ............................................. 87 • Illustration 3: Protocole de cartographie et échantillonnage des turricules dans le
pâturage et dans la forêt. ................................................................................. 127 • Illustration 4 : Expérience de brûlage des turricules ............................................. 128 • Illustration 5 : Dispositif expérimental de simulation de pluie................................. 129

RESUME
En Amazonie, la transformation de la forêt en pâturage par l’agriculture familiale modifie les propriétés physico-chimiques des sols argileux et se manifeste aussi par une mutation des populations d’invertébrés qualifiés d’ingénieur du sol et donc des structures biogéniques de surface produites.
L’objectif de cette étude est de mettre en évidence le rôle de deux espèces ingénieurs du Nord-est brésilien: l’annélide Andiodrilus pachoensis et le termite Cornitermes sp. dans le fonctionnement du sol et d’établir une typologie globale des structures biogéniques produites par les organismes ingénieurs.
Les Latosols des parcelles sont caractérisés par un horizon de surface argilo-sableux. Dans les forêts, l’horizon 0-10 cm est organique humifère relativement homogène alors que dans les pâturages, soumis à la culture sur brûlis, il est tassé et hétérogène en raison de la structure localement lamellaire entre les touffes de Brachiaria brisantha ainsi que de la distribution en taches de la matière organique et de l’hydromorphie.
Les termitières épigées dans les pâturages atteignent une densité de 38 nids/ha alors qu’aucun édifice n’a été observé sous forêt. La distribution spatiale des termitières est hétérogène mais reliée au type de sol. Les propriétés physico-chimiques de ces structures termitiques mettent en évidence la forte stabilité structurale et la résistance au brûlis annuel des termitières. Leur destruction par les paysans, suscite un effet local immédiatement positif, puis négatif en assurant la destruction de la réserve de matière organique du feuilleté interne et une formation de pseudo-sables (pectisation des argiles) et d’hématites assurant une augmentation de le stabilité structurale de la muraille.
La densité des turricules d’Andiodrilus p. est apparue plus faible dans les pâturages (42/ha) que dans les forêts (176/ha). Leurs distributions spatiales y sont agrégées aux larges échelles (>15m) ; alors qu’en forêt, elles sont très hétérogènes entre les parcelles. La texture des turricules est argilo-sableuse et leur stabilité structurale est supérieure à celle des horizons témoins. Malgré la pectisation des argiles et la formation d’hématite, aucun effet de la culture sur brûlis sur la stabilité structurale ou les concentrations en cations des turricules n’a été révélé.
Différentes typologies des structures biogéniques ont été établies à partir de l’étude de leur matière organique (NIRS), ou encore de leur porosité par analyses micromorphologiques. Les différentes typologies obtenues soulignent l’importance d’accumuler des données spécifiques à chaque espèce clés des sols afin de définir des groupes fonctionnels pertinents en vue de modéliser le fonctionnement du sol.
Mots clés : Andiodrilus pachoensis / Cornitermes sp. / Turricule / Termitière / feuilleté / xylophage / distribution spatiale / propriétés physiques et chimiques / cations / stabilité structurale / Brachiaria brisantha / NIRS / Lames minces / culture sur brûlis / simulation de pluie / Amazonie
ABSTRACT In Amazonia, transformation of forest to pasture by family agriculture alters physical and chemical properties of
clayey soil and is also reflected by changes in soil engineers populations, , and thus in their own-making biogenic structures.
The aim of this study was to determine the impacts on soil functions of two soil engineer species of the Brazilian north-east Amazonia: the earthworm Andiodrilus pachoensis and the termite Cornitermes sp. as well as to establish a global typology of biogenic structure produced by the engineers.
Study fields were characterised by latosols with a clayey-sandy superficial layer. In forest, the 0-10 cm layer was organic and homogeneous whereas in slash and burned pastures it was compact and heterogeneous because of locally lamellar structure between Brachiaria brisantha clumps and the spot distribution of organic matter and hydromorphy.
In pastures, Cornitermes mounds density was of 38 nests per ha while in forest, no mound was found. Their spatial distribution was heterogeneous but significantly linked to the type of soil. Physical and chemical properties of these structures showed a great structural stability and resistance to annual slash and burned culture. The farming destruction of termite mounds creates an immediate positive effect but, in a second time, a negative effect, when organic matter stocked in the internal foliated part has been totally burnt and structural stability of the mounds wall has been increased by pseudo-sand and hematite making.
Andiodrilus pachoensis casts density was significantly lower in pastures and their distribution was clumped at the large scale (> 15 cm) whereas in forest spatial analyses showed that the cast distribution varied highly between plots: from random to clumped or inhibited. Earthworm casts density was not significantly correlated with tuft density and their production was independent to B. brisantha tufts presence in pastures or litter quantity in forests. Texture of casts was clayey sandy like soil superficial layer and its structural stability was greater than the surface soil one. When they were burnt, casts took a reddish colour, evidence of clay pectisation and hematite making but their structural stability and cations concentration did no change.
Various biogenic structures typologies were established from organic matter study (NIRS) or porosity estimation (micromorphologic analysis). Results underline the necessity to gather specifics data on soil keystone species in order to define significant functional group for modelling of global soil functions.
Key words: Andiodrilus pachoensis / Cornitermes sp. / casts / termite mounds/ foliated / spatial distribution / physical and chemical properties / cations / structural stability / Brachiaria brisantha / NIRS / thin sections / slash and burn / rain simulation / Amazonia





























