Embed Size (px)
Citation preview

UE9-IMMUNOPATHOLOGIE Pr
P.GASQUE
Date : 9/03/2018 Plage horaire : 10h45-12h45 Promo : DFGSM2 2016/2017 Enseignant : Pr GASQUE
Ronéistes : CALPETARD Victor LARRUE Nathan
LE SYSTEME CLEF DE L’IMMUNITE INNEE: LE COMPLEMENT
L. ‘immunité innée A. Immunité Innée et Immunité Adaptative B. Qu'est-ce qu'un pathogène?
1. La reconnaissance sélective (concept 1) 2. Pathogène = microbes + débris dérivés de l'hôte (concept 2)
C. Les mécanismes de reconnaissance 1. Introduction 2. Le rôle du PRR (TLR4) 3- Le rôle du LPS 4- Les différents PRR
D. Un exemple de PRR: Les Toll Like Receptor (TLR)
II. Système de défense contre les virus
III. Système de défense contre les bactéries A. La phagocytose B. Le système du complément
1- Introduction, historique, généralités 2- Les 3 voies d'activation

Introduction
L'objectif de ce cours est de comprendre quel est le rôle de l'immunologie. De manière générale, son rôle essentiel est de tuer les pathogènes.
Le mot «pathogène» devra être pris au sens le plus large possible, ça n'est pas qu'un microbe, ça peut être une cellule cancéreuse ou une cellule apoptotique.
L'immunologie comprend deux grands volets : -l'immunité innée (II) immunité naturelle mise en place dès qu'on sera infecté en contact avec un élément pathogène (phase de minutes à heures) -l'immunité adaptative (IA) ne peut se faire que si l'immunité innée a été instruite. L'objectif étant de produire des anticorps, des cellules T, CD4, CD8 qui vont réagir beaucoup plus tardivement.
En d'autres termes, si votre II n'a pas lieu et que vous attendez l'IA, celle-ci ne pourra pas être efficace. Vous serez déjà décédés de cette infection parce que votre système immunitaire (SI) aura mis trop de temps à réagir contre l'agent infectieux.
L'II nous protège des éléments pathogènes tout en préservant nos cellules normales. Le système du complément, pourtant très cytotoxique, va éliminer les pathogènes mais épargner nos propres cellules.
L’II et l’IA sont extrêmement interconnectés. Sachant que c’est l’II qui doit s’installer dans un 1er temps avant que puisse en tout état de cause s’installer et s’engager l’IA avec la réponse B, T. L’IA est beaucoup plus spécifique que l’II. D’une manière générale, le principal de l’II est d’éliminer les pathogènes, à savoir les éléments du non soi. On peut naturellement penser à microbe de type bactérie, parasites ou encore champignons. On verra que cette notion de pathogène est à élargir à sa complexité notamment incluant les facteurs dérivés de l’hôte. Donc pathogène selon le professeur : illustration de quelque chose pouvant induire la genèse d’une pathologie.
On verra que notre système immunitaire inné est capable de reconnaître les facteurs dérivés de l’hôte tel que la protéine amyloïde, prions … Cette élimination des pathogènes ne peut se faire qu’en préservant nos propres cellules. Si on a une bactérie se baladant dans notre SNC. Il est clair que notre système immunitaire va lutter contre celle-ci mais tout en préservant nos neurones, cela est vrai pour les hépatocytes dans le foie, les myoblastes dans le muscle.
Il faut absolument que ce système de l’II soit suffisamment discriminant pour préserver les éléments du soi (nos propres cellules) et éliminer les pathogènes. Il peut y avoir des échecs à cela, dans le cadre de l’auto immunité, notre système immunitaire peut être dirigé contre nos propres constituants. Ceci est la complexité des pathologies auto immunes.
I. L’immunité innée :
A. Immunité Innée et Immunité Adaptative :
- Avant de rentrer dans les détails de l’II, un élément concernant la séquence lors de l’entrée d’un pathogène dans notre organisme pour la 1ère fois : l’existence d’une dichotomie entre l’II et l’IA (visible en haut de la diapositive), sachant que l’II

sera la 1ère. On ne pourra jamais avoir une IA lymphocytes T, B si notre II ne s’est pas mise en route.
En l’occurrence, cette II se mettant en route dès les premières minutes ou heures d’une infection aura pour principe de reconnaître l’élément pathogène de manière sélective : attention on ne parle pas de spécificité quand il s’agit de l’II (l’immunité spécifique est apportée par nos lymphocytes T, B), le phagocyter (c’est-à-dire l’éliminer de la circulation dans le meilleur des cas), essayer de le lyser (en second cas) et autre chose extrêmement importante : induire la production de facteurs ou molécules :
- soit anti-infectieuses (cas de l’interféron essayant de lutter contre des infections virus),
- soit par production de cytokines pro-inflammatoires qui auront pour objet d’activer nos cellules immunitaires de type macrophages, neutrophiles pour qu’elles infiltrent le site infectieux et qu’elles viennent lutter contre l’agent infectieux,
- soit le complément (plusieurs molécules dont l’objectif est de détruire cet élément pathogène que ce soit une bactérie, un parasite ou un virus).
⇨ L'II est la première à apparaître : les premières heures voire les premiers jours suite à une infection. Elle comprend les phagocytes, les macrophages, les cytokines pro-inflammatoires (TNFα), les interférons anti-virales α et β (les interférons γ concernent l'immunité lymphatique).
Puis il y aura séquentiellement activation du système immunitaire adaptatif avec la capacité de générer spécifiquement des lymphocytes T dirigés contre des protéines virales, bactériennes, contre un constituant parasitaire et générer des Ac mono et polyclonaux dirigés contre cet élément pathogène.
Il n’y a pas de mémoire dans l’II mais dans l’IA. C’est-à-dire que si demain vous avez à nouveau une infection avec ce pathogène, vous n’aurez plus besoin de passer par l’étape II, il y aura une mobilisation rapide de vos LT et de votre panel d’Ac dirigés contre ce pathogène. Il y aura une élimination extrêmement rapide de cet élément pathogène. Le principe de la vaccination est de faciliter cette 1ère étape pour avoir plus rapidement ces LT et ces Ac pour lutter contre cet élément pathogène.
-Inéluctablement, l'IA sera beaucoup plus tardive et ne pourra être établit que si l'II a joué son rôle. Elle comprend les lymphocytes T et les lymphocytes B capables de produire une quantité importante d'anticorps lorsqu'ils se différencient en plasmocytes. (Partie infection approfondie par JJH)
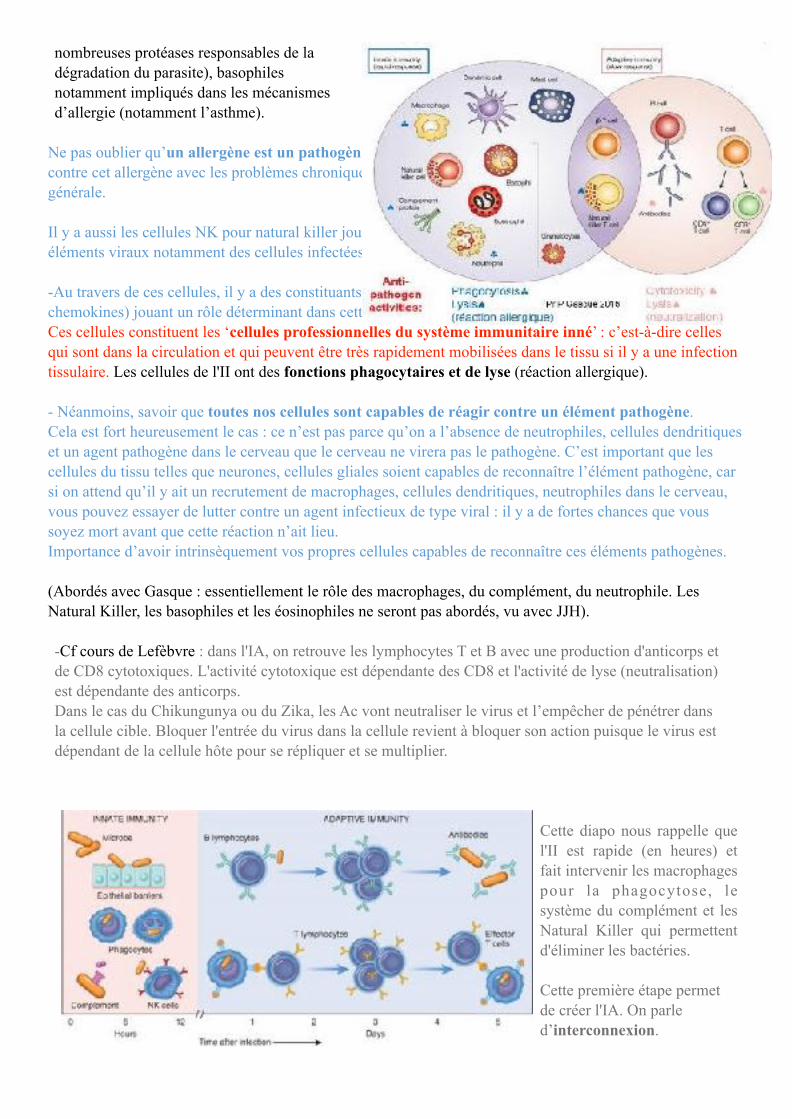
Cells and molecules of immunity Autre représentation de la dichotomie mais cette fois-ci au niveau des cellules impliqués dans la fonction des différents SI : -Quand il s’agit de l’II, on a essentiellement des cellules impliquées dans la reconnaissance de ces éléments pathogènes. D’où la présence de ces cellules quand on parle de l’II : macrophages, neutrophiles, éosinophiles (défense anti parasitaire en allant s’agréger aux parasites et par un mécanisme de dégranulation, libérer de
Les cellules de l’immunité sont capables de reconnaitre une cellule infecté contrairement aux virus ceux qui leur permet de phagocyter et lyser cette cellule.
nombreuses protéases responsables de la dégradation du parasite), basophiles notamment impliqués dans les mécanismes d’allergie (notamment l’asthme).
Ne pas oublier qu’un allergène est un pathogène. Il y aura donc une capacité de ces cellules à réagir contre cet allergène avec les problèmes chroniques de l’asthme, de la réaction allergique de manière générale.
Il y a aussi les cellules NK pour natural killer jouant un rôle déterminant dans la reconnaissance des éléments viraux notamment des cellules infectées par des virus ou encore des cellules cancéreuses. -Au travers de ces cellules, il y a des constituants moléculaires (complément, interférons, cytokines, chemokines) jouant un rôle déterminant dans cette II. Ces cellules constituent les ‘cellules professionnelles du système immunitaire inné’ : c’est-à-dire celles qui sont dans la circulation et qui peuvent être très rapidement mobilisées dans le tissu si il y a une infection tissulaire. Les cellules de l'II ont des fonctions phagocytaires et de lyse (réaction allergique).
- Néanmoins, savoir que toutes nos cellules sont capables de réagir contre un élément pathogène. Cela est fort heureusement le cas : ce n’est pas parce qu’on a l’absence de neutrophiles, cellules dendritiques et un agent pathogène dans le cerveau que le cerveau ne virera pas le pathogène. C’est important que les cellules du tissu telles que neurones, cellules gliales soient capables de reconnaître l’élément pathogène, car si on attend qu’il y ait un recrutement de macrophages, cellules dendritiques, neutrophiles dans le cerveau, vous pouvez essayer de lutter contre un agent infectieux de type viral : il y a de fortes chances que vous soyez mort avant que cette réaction n’ait lieu. Importance d’avoir intrinsèquement vos propres cellules capables de reconnaître ces éléments pathogènes.
(Abordés avec Gasque : essentiellement le rôle des macrophages, du complément, du neutrophile. Les Natural Killer, les basophiles et les éosinophiles ne seront pas abordés, vu avec JJH).
-Cf cours de Lefèbvre : dans l'IA, on retrouve les lymphocytes T et B avec une production d'anticorps et de CD8 cytotoxiques. L'activité cytotoxique est dépendante des CD8 et l'activité de lyse (neutralisation) est dépendante des anticorps. Dans le cas du Chikungunya ou du Zika, les Ac vont neutraliser le virus et l’empêcher de pénétrer dans la cellule cible. Bloquer l'entrée du virus dans la cellule revient à bloquer son action puisque le virus est dépendant de la cellule hôte pour se répliquer et se multiplier.
Cette diapo nous rappelle que l'II est rapide (en heures) et fait intervenir les macrophages pour la phagocytose, le système du complément et les Natural Killer qui permettent d'éliminer les bactéries.
Cette première étape permet de créer l'IA. On parle d’interconnexion.

B. Qu'est-ce qu'un pathogène?
1. La reconnaissance selective :
La question essentielle est la suivante : comment est-ce que les cellules sont capables de reconnaître et d’éliminer un élément pathogène ?
1. Les cellules vont reconnaître l'élément pathogène et vont le cibler sélectivement par l’II. Cette étape clé est la reconnaissance du non- soi: le système immunitaire discrimine nos cellules normales à préserver des cellules cancéreuses/infectées qui se comportent comme un pathogène,
2. Lyser la cible (souvent le cas de la bactérie et du parasite qui sont des gros organismes), 3. Phagocyter (bactéries, cellules infectées par un virus).
Description schéma : Le pathogène exprime un ligand appelé PAMP (Pathogen Associated Molecular Pattern), ex: LPS. La cellule immunitaire (ici le macrophage) reconnaît sélectivement ce ligand via un récepteur appelé PRR (Pattern Recognition Receptor), ex: Toll-like receptor = protéine citruliné, ARN. Lors de la NFS on regarde deux choses: le taux de lymphocytes et neutrophiles. Si on a une augmentation des cellules lymphocytaires = infection bactérienne sinon ça sera une infection virale.
Il est important qu’un macrophage puisse phagocyter une cellule infectée par un virus à la différence de l’avoir lysé. S’il y a lyse d’une cellule infectée par un virus, le virus va se répandre alors qu’en phagocytant une cellule infectée par un virus, la cellule infectée sera totalement phagocytée incluant les virus et ceux-ci ne pourront plus se répandre. C’est pour cela que quand il y a un contexte infectieux de type viral, il est préférable de faciliter la phagocytose plutôt que la lyse de la cellule infectée par les virus.
Le mécanisme mis en jeu est que les cellules jouant un rôle dans l’II, expriment des senseurs ou sensors reconnaissant sélectivement un ligand exprimé par le pathogène. C’est l’interaction entre la cellule immunitaire et le pathogène qui va faciliter la reconnaissance et l’élimination de ce pathogène.
▪ Concept 1 : les PRR reconnaissent sélectivement plusieurs ligands Malheureusement, il y a des pathogènes petits, des découvertes journalières (on n’entendait pas parler des virus chikungunya et Sindbis il y a 10 ans de cela : aujourd’hui ils sont mieux connus, il y a une myriade de ces pathogènes qui circule, tous ne sont pas nécessairement pathogènes mais peuvent acquérir une pathogénicité).
Il est important de comprendre que le système immunitaire soit capable de reconnaître chacun des
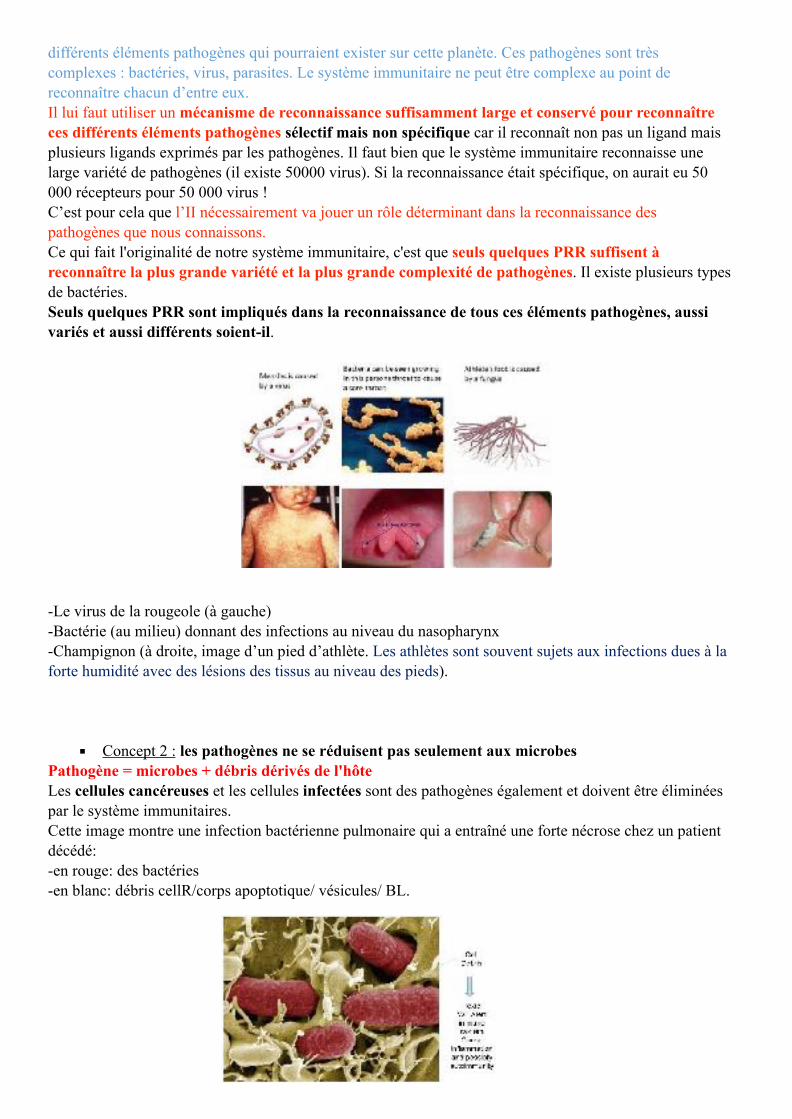
différents éléments pathogènes qui pourraient exister sur cette planète. Ces pathogènes sont très complexes : bactéries, virus, parasites. Le système immunitaire ne peut être complexe au point de reconnaître chacun d’entre eux. Il lui faut utiliser un mécanisme de reconnaissance suffisamment large et conservé pour reconnaître ces différents éléments pathogènes sélectif mais non spécifique car il reconnaît non pas un ligand mais plusieurs ligands exprimés par les pathogènes. Il faut bien que le système immunitaire reconnaisse une large variété de pathogènes (il existe 50000 virus). Si la reconnaissance était spécifique, on aurait eu 50 000 récepteurs pour 50 000 virus ! C’est pour cela que l’II nécessairement va jouer un rôle déterminant dans la reconnaissance des pathogènes que nous connaissons. Ce qui fait l'originalité de notre système immunitaire, c'est que seuls quelques PRR suffisent à reconnaître la plus grande variété et la plus grande complexité de pathogènes. Il existe plusieurs types de bactéries. Seuls quelques PRR sont impliqués dans la reconnaissance de tous ces éléments pathogènes, aussi variés et aussi différents soient-il.
-Le virus de la rougeole (à gauche) -Bactérie (au milieu) donnant des infections au niveau du nasopharynx -Champignon (à droite, image d’un pied d’athlète. Les athlètes sont souvent sujets aux infections dues à la forte humidité avec des lésions des tissus au niveau des pieds).
▪ Concept 2 : les pathogènes ne se réduisent pas seulement aux microbes Pathogène = microbes + débris dérivés de l'hôte Les cellules cancéreuses et les cellules infectées sont des pathogènes également et doivent être éliminées par le système immunitaires. Cette image montre une infection bactérienne pulmonaire qui a entraîné une forte nécrose chez un patient décédé: -en rouge: des bactéries -en blanc: débris cellR/corps apoptotique/ vésicules/ BL.

En même temps que se développent les bactéries, il y a accumulation de facteurs dérivés de l’hôte qui ne sont pas des composés normaux. Ils s’accumulent quand il y a une infection causant des dommages aux tissus et ces facteurs dérivés de l’hôte sont autant pathogéniques que la bactérie car il s’agit de débris et s’il y a trop de débris qui s’accumulent, ils vont également contribuer à la pathologie pulmonaire. Notre système immunitaire pourrait potentiellement réagir contre nos propres cellules, réaction auto-immune car ce sont des composé toxique qui sont pas assez éliminé.
Cela peut être des facteurs dérivés du noyau, des cytosols des cellules, de la membrane des cellules, souvent cela correspond à des corps apoptotiques : c’est-à-dire que la cellule qui est proche d’un virus ou d’une bactérie va subir un mécanisme d’apoptose (mort programmée de la cellule) générant des corps apoptotiques extrêmement toxiques. L’organisme n’est pas capable de les éliminer et ces éléments issus de nos cellules peuvent être à l’origine de maladies auto immunes. Ils peuvent contenir de l’ADN, des phosphatidyl-sérines, de nombreux constituants perçus par notre système immunitaire comme des auto antigènes. L’organisme réagit et créer des auto anticorps à l’origine des pathologies auto immunes.
⇨ Ces débris cellulaires comportent des substances toxiques et se comportent donc comme des pathogènes, au même titre que les bactéries présentes. Ils doivent donc être éliminés.
⇨ Si l'organisme a une déficience dans l'élimination et la reconnaissance de ces débris cellulaires, cela peut aboutir à des maladies auto-immunes : -débris cellulaires riches en ADN dans le lupus = forte nécrose du cerveau du à la mort par apoptose des cellules de l’immunité => il faut éliminer tout les corps étrangers qui n’appartiennent pas au milieu d’origine. -myéline dans la sclérose en plaque.
Dans le cerveau, l'accumulation de myéline doit être éliminée parce qu'elle fait partie des débris cellulaires. Si elle n'est pas éliminée, des Ac ou lymphocytes T vont être auto-réactifs contre la myéline donnant la maladie de la sclérose en plaque.
Role of host-derived cofactors in pathological mechanisms (hence a pathogen)
Cette image montre une infection par un aspergillus chez un patient décédé qui a été atteint du VIH et fortement immunodéficient avec un taux CD4 faible.
La flèche du haut représente l'infection par un aspergillus.
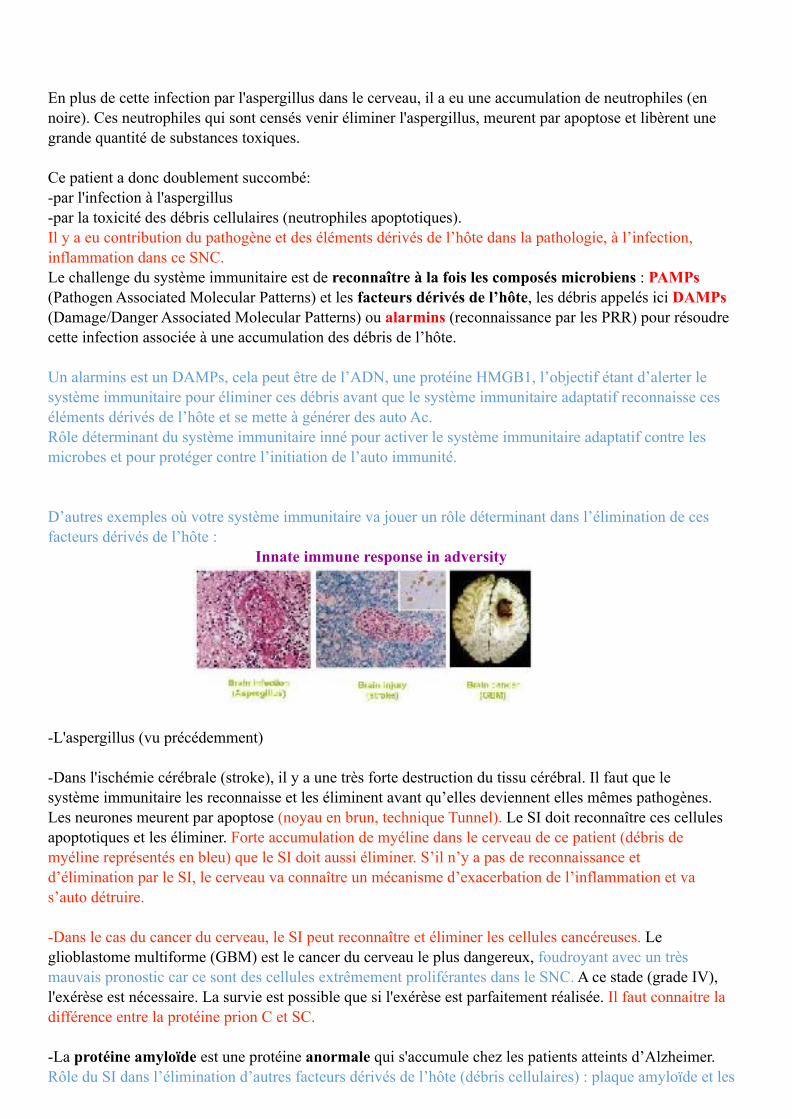
En plus de cette infection par l'aspergillus dans le cerveau, il a eu une accumulation de neutrophiles (en noire). Ces neutrophiles qui sont censés venir éliminer l'aspergillus, meurent par apoptose et libèrent une grande quantité de substances toxiques.
Ce patient a donc doublement succombé: -par l'infection à l'aspergillus -par la toxicité des débris cellulaires (neutrophiles apoptotiques). Il y a eu contribution du pathogène et des éléments dérivés de l’hôte dans la pathologie, à l’infection, inflammation dans ce SNC. Le challenge du système immunitaire est de reconnaître à la fois les composés microbiens : PAMPs (Pathogen Associated Molecular Patterns) et les facteurs dérivés de l’hôte, les débris appelés ici DAMPs (Damage/Danger Associated Molecular Patterns) ou alarmins (reconnaissance par les PRR) pour résoudre cette infection associée à une accumulation des débris de l’hôte.
Un alarmins est un DAMPs, cela peut être de l’ADN, une protéine HMGB1, l’objectif étant d’alerter le système immunitaire pour éliminer ces débris avant que le système immunitaire adaptatif reconnaisse ces éléments dérivés de l’hôte et se mette à générer des auto Ac. Rôle déterminant du système immunitaire inné pour activer le système immunitaire adaptatif contre les microbes et pour protéger contre l’initiation de l’auto immunité.
D’autres exemples où votre système immunitaire va jouer un rôle déterminant dans l’élimination de ces facteurs dérivés de l’hôte :
Innate immune response in adversity
-L'aspergillus (vu précédemment)
-Dans l'ischémie cérébrale (stroke), il y a une très forte destruction du tissu cérébral. Il faut que le système immunitaire les reconnaisse et les éliminent avant qu’elles deviennent elles mêmes pathogènes. Les neurones meurent par apoptose (noyau en brun, technique Tunnel). Le SI doit reconnaître ces cellules apoptotiques et les éliminer. Forte accumulation de myéline dans le cerveau de ce patient (débris de myéline représentés en bleu) que le SI doit aussi éliminer. S’il n’y a pas de reconnaissance et d’élimination par le SI, le cerveau va connaître un mécanisme d’exacerbation de l’inflammation et va s’auto détruire.
-Dans le cas du cancer du cerveau, le SI peut reconnaître et éliminer les cellules cancéreuses. Le glioblastome multiforme (GBM) est le cancer du cerveau le plus dangereux, foudroyant avec un très mauvais pronostic car ce sont des cellules extrêmement proliférantes dans le SNC. A ce stade (grade IV), l'exérèse est nécessaire. La survie est possible que si l'exérèse est parfaitement réalisée. Il faut connaitre la différence entre la protéine prion C et SC.
-La protéine amyloïde est une protéine anormale qui s'accumule chez les patients atteints d’Alzheimer. Rôle du SI dans l’élimination d’autres facteurs dérivés de l’hôte (débris cellulaires) : plaque amyloïde et les

« neurofibrillary tangles » (neurones mourant par apoptose et nécrose caractérisant la maladie d’Alzheimer).
Aujourd’hui on sait que le SI dans le cerveau au travers des microglies, neurones, astrocytes va reconnaître ces éléments comme étant des dérivés dangereux donc des DAMPs = des alarmins et va essayer de les éliminer. Un des concepts de la maladie d’Alzheimer est qu’avec l’âge, il y a une incapacité à éliminer ces plaques amyloïdes et neurones mourant par apoptose. Cela pouvant conduire à une maladie d’Alzheimer beaucoup plus précoce.
-Dernier exemple donné de DAMPS = alarmins : protéine prion. Ne pas oublier que le prion est une protéine normale exprimée par nos cellules et jouant un rôle dans la capacité à capter les et à éliminer les ???. Cependant, cette protéine peut être transformée en une forme peu soluble s’appelant protéine fibrillaire prion que l’on voit dans la pathologie scrapie ou tremblante du mouton. Pour la petite histoire (le professeur ayant vécu en Grande-Bretagne pendant très longtemps) : dans les années 90 à 98, il y a eu plus de 100 000 carcasses de vache folle qui étaient infectées et sont allées dans la chaîne alimentaire. Cela est probablement responsable de la nouvelle variante de prion qu’on a vu chez les enfants entre 10 et 15 ans due aux hormones de croissances. Tout ça pour dire que prion est un agent infectieux et pathogène bien qu’il fasse partie du soi et là fort heureusement le SI est capable de le reconnaître particulièrement la forme Sc et non C normale et contribue à son élimination sur le même principe que nous avons vu pour les microbes classiques (virus, bactéries). La protéine amyloïde ou la protéine prion va exprimer des motifs reconnus par des senseurs, récepteurs exprimés par les cellules type macrophage, astrocyte, neurone (ce que l’on veut), ceci contribuant à leur élimination.
A retenir: le SI ne reconnaît pas uniquement que des microbes mais également des débris cellulaires dont la cellule cancéreuse.
C. Les mécanismes de reconnaissance :
1. Introduction :
How do your cells define "self" versus "danger-sef" and "non-self"?
(In the absence of acquired knowlegde!!!)
Question : si nous avions à reconnaître les virus, bactéries, parasites, protéine amyloïde, prions comme étant pathogènes, comment ferions-nous ? Ne pas oublier que les cellules n’ont pas de yeux, pas de cerveau et ne peuvent pas savoir ce qui est pathogène et ce qui ne l’est pas. Nous avons une information que nos cellules n’ont pas permettant la distinction entre la pathogénicité et non pathogénicité. Des mécanismes vont permettre de reconnaître un élément pathogène lorsqu’on a aucune instruction pour le faire. La réponse est donnée par l’enfant, le bébé : il ne sait pas lorsqu’il est né ce qui est bien, ce qui est bon ou pas pour lui. Il va alors goûter : un enfant vous lui donnez n’importe quoi, il va le mettre dans sa bouche.

C’est exactement ce qu’il se passe pour les cellules. Elles ne savent pas ce qui est bon de ce qui ne l’est pas. En l’occurrence bon : je vais manger, phagocyter, éliminer. Les cellules vont donc goûter, venir au contact de quelque chose les entourant et elles vont questionner : est-ce que tu es bon ?
- si c’est le cas, je te phagocyte, - sinon (amer et pas bon) : je ne fais pas de phagocytose.
L’apprentissage du SI peut donc être vu comme celui d’un enfant : si on lui donne un citron, il ne le mangera pas car il sait que c’est acide et donc désagréable pour lui. C’est la même chose pour notre SI : reconnu par les PRR, nos macrophages ont des sensors (PRR) ou dégustateurs qui vont détecter les sucres. Si ceux-ci sont plaisants, les macrophages les phagocytent. On verra que le complément, au travers de lectines telles que Mannan Binding Lectin (MBL), est capable de détecter des pathogènes exprimant des sucres particuliers (carbohydrates) et qui détermineront s’il y aura phagocytose ou non. (Ces 2 paragraphes ci-dessus sont tirés de la ronéo de l'année précédente. L'explication est plus claire que celle donnée en cours)
Entre quelque chose du soi et quelque chose du non soi, en passant par tous les scénarios possibles (plaques amyloïdes, protéines prions ou bactéries), la tâche du SI est extrêmement difficile puisqu’il devra d’un côté préserver et de l’autre reconnaître et s’engager dans l’élimination de ce pathogène. Le tout sans aucune connaissance de ce qui est pathogène et de ce qui ne l’est pas.
2. Le rôle du PRR (TLR4) :
All cells express innate immune receptor (Pattern Recognition Receptors, PRRs) sense and respond to "pathogens"
Quelqu’un qui a joué un rôle clé dans l’identification des mécanismes de reconnaissance et de ‘taste’ (de goût) des éléments pathogènes en général : Charles Janeway. Il a permis de mettre en place ce concept que peu importe si on a des cellules apoptotiques, microbes ou des protéines pathogéniques (Alzheimer, protéine amyloïde, fibrille), nous aurons des senseurs : des Pattern Recognition Receptors = PRR pour détecter des motifs exprimés par tous ces composés. L’engagement de ces récepteurs va induire la phagocytose et la lyse de l’élément pathogène.
⇨ Charles Janeway, le «pape de l'immunité inné» a introduit le concept du PRR capable à la fois de reconnaître des pathogènes, des cellules apoptotiques et des protéines pathogènes (prion, amyloïde).
Description schéma : le ligand indiqué comme étant un motif de reconnaissance s’appelle PAMPs ou DAMPs, le récepteur s’appelle PRR (multiples dans une liste restreinte) et est capable de reconnaître la grande complexité de tous ces éléments pathogènes (microbes, débris tissulaires).

$
Il a fallu ensuite déterminer quels étaient ces PRR, PAMPs et DAMPs. Le concept étant donné, il fallait à cette époque (et ce n’est pas si vieux que ça) se poser la question : quelle est la nature de ces PRR, PAMPs et DAMPs ? Un élément important déjà donné en rapport avec l’analogie de l’enfant : le goût. Un des goûts que nous avons et que le SI utilise aussi c’est le goût pour le sucré et le gras, il détecte des sucres (hydrate de carbone) ou des lipides (LPS ou lipopolysaccharide). On se focalisera sur l’exemple des LPS : comme son l’indique c’est quelque chose de gras et c’est aussi un saccharide donc sucré. Il est exprimé par les bactéries. La molécule du PRR capable de reconnaître un LPS est le TLR4. Le Toll Like Receptor 4 (TLR4) est aujourd’hui l’exemple le plus connu de PRR. Il est le récepteur du LPS.
Nos cellules adorent le sucre et comme l’ARN, ADN et le peptidoglycane sont des sucres, il va y avoir reconnaissance des virus (car contenant une longue chaîne de sucre : ribose pour les virus à ARN et désoxyribose pour les virus à ADN) et des bactéries par notre SI (reconnues par leur peptidoglycane). Les cellules goutant ces longues chaînes de sucres vont avoir un très fort signal et cela augmentera leur appétit. Elles vont se mettre à phagocyter ces virus et bactéries.
Ces PRR agissent comme un système de surveillance qui donne l'alerte en contact avec un élément pathogène exprimé en intracellulaire au niveau du cytosol pour détecter les virus ou en extracellulaire à la membrane pour détecter les bactéries.
Dans les années 90-95, s’est posée la question de la nature des PAMPs, DAMPs et PRR sachant que l’hypothèse proposée ici était : le pathogène exprime le ligand reconnu par un récepteur et ce sont les interactions entre ces deux éléments qui faciliteront la reconnaissance de pathogène : si la cellule nécrose alors l’ADN et ARN sont exposés en extra cellulaire et sont donc phagocytés.
Hypothesis: Defect in a gene coding for a PRR of innate immune system will increase susceptibility to infection

Rôle déterminant du français Jule Hoffman de Strasbourg ayant beaucoup œuvré dans l’identification des mécanismes de reconnaissance et de signalisation permettant de contrôler l'infection. Il fut le 1er à répondre à cette question. Son hypothèse était d’identifier le PAMP et le récepteur capable de détecter des éléments pathogènes et surtout prouver en quoi l’absence d’un gène exprimé par un organisme l’empêcherait de lutter contre un agent infectieux. Il s’est alors posé la question suivante : existe-t-il une mutation dans un gène qui sensibiliserait un organisme à des maladies infectieuses ? Et auquel cas, il y aurait de fortes chances que ce gène code pour un PRR. Dans ce temps, nous n’avions pas tous les outils disponibles aujourd’hui (séquençage du génome pas encore réalisé, en tout cas pas toutes les techniques de biologie cellulaire et moléculaire n’étaient disponibles).
Chose inhabituelle, il n'a pas travaillé avec des souris ni chez l'homme mais sur des drosophiles car la question de l’II qu’on l’a pose sur un ver de terre, une étoile de mer, un homme c’est la même question : comment ces organismes arrivent à lutter contre des agents infectieux ? Un ver de terre baigne dans un milieu enrichis de pathogènes, il faut qu’il lutte également (infections bactériennes). La drosophile c’est la même chose, elle a tendance à aller là où nous n’irions pas, elle va donc subir une forte exposition à des éléments infectieux (bactériens, fongiques).
L’objectif d’Hoffman était d’identifier des mutants de la drosophile qui ne seraient plus capables de lutter contre un agent infectieux. C’est le cas sur cette image prise au microscope de la drosophile (ci-dessous) : elle est morte recouverte d’énormément de champignons. C’est un mutant très facile a générer :
- Le temps de reproduction et la durée d'exposition nécessaire pour induire une mutation sont beaucoup plus courts chez la drosophile que chez la souris.
- Une drosophile est exposée à un produit chimique (gaz, insecticides, pesticides, tout ce que vous voulez) ou aux UV. Comme une drosophile ne met que quelques jours pour se multiplier : sa descendance au bout d'une semaine aura cette mutation.
⇨ Pour cette étude, la drosophile est un modèle de choix. = autre technique possible en rendant les souris efficientes mais cette technique était peu au point à l’époque.
Grand patron de l’immunologie française, il obtient son prix Nobel en 2011 en même temps que Bruce Beutler et Ralph Steinman. Les travaux de Ralph Steinman concernent l'IA et l'identification des cellules dendritiques tandis que ceux de J. Hoffman et de B. Beutler se portent plutôt sur l’II. Ainsi qu’un bel article séminal dans la revue Cell (unes des meilleures revues scientifiques au monde) avec un fort facteur d’impact, publié en 1996 dans lequel est reporté son expérience. NB: Les articles scientifiques sont publiés dans des journaux scientifiques dont la réputation est représentée par le facteur d'impact. Impact de Cell: 40 Impact du Journal de l'île: de l'ordre de 0,00qqchose Les meilleurs journaux sont Cell, Nature, Nature Medicine avec un très fort facteur d'impact.
Rappel de la problématique soulevée : si un gène codant pour un PRR est muté, est-ce que le SI perdrait sa capacité à lutter contre l'infection? L'expérience consiste à induire des mutations chez des drosophiles et ensuite à les exposer soit à des bactéries soit des champignons. On observe alors le taux de survie des drosophiles. Sur le graphique, on voit qu’il a pu identifier le % de survie des drosophiles selon qu’elles sont exposées à aspergillus (un champignon) ou e.coli (une bactérie). Les drosophiles OR sont résistantes et si elles

présentent une mutation entrainant un défaut d’expression du gène Tl- ou imd ou les 2, il y avait alors une augmentation de la susceptibilité à l’infection avec une chute du taux de survie très rapidement.

The role of the Toll and imd pathways in the immunity in D melanogaster
Sur la courbe de survie, la flèche jaune indique un mutant marqué Tl-. Ce clone de drosophile avait une mutation dans un gène appelé Toll. Il s'avère que le taux de survie des drosophiles diminue fortement lorsque leur gène Toll est muté. Toll est donc impliqué dans la lutte contre des agents infectieux de type champignon. Ce qui est vrai également pour la bactérie E. Coli. Aujourd’hui Tl code pour TLR4 de drosophile. Donc Tl est le récepteur chez la drosophile Toll permettant la reconnaissance de bactéries et champignons.
Très rapidement, il y a eu un article dans Nature (summum des journaux scientifiques) qui a été réalisé par Charles Janeway (celui ayant émis l’hypothèse initiale) et son co doctorant Ruslan Medzhitov.
Ils ont identifié l’homologue humain de Toll de la drosophile, appelé Toll-like récepteur = celui qui ressemble au gène Toll de la drosophile, simplement par séquençage de la drosophile et par homologie de séquences. Aujourd’hui il y a 11 à 12 TLR impliqués dans la reconnaissance de pathogènes. Ces études montrent que les TLR sont impliqués en tant que PRR dans l'élimination des pathogènes.
3. Le rôle du LPS : ( grâce à la fabrication de souris pas capable de répondre au LPS tué à une mutation).
Si on compare ce qu’il se passe chez la drosophile et chez l’homme, ces études ont permis d’identifier ces 2 récepteurs : Toll dans le modèle de la drosophile et TLR dans le modèle humain. Cependant, le ligand n'avait toujours pas été identifié : quel est le ligand (exprimé par le pathogène) capable de se fixer sur Toll et d'induire les activités anti- infectieuses ? On sait aujourd’hui que ces récepteurs vont détecter des composés issus des éléments pathogènes (de type bactérien et champignons). La fixation de ses ligands sur Toll va engager des voies d’activation qui vont faire intervenir quelque chose de très important revu systématiquement en sciences : les facteurs de transcription, dont un important ici : Nf-κB. Ce FT va permettre l’activation de gènes, la production de protéines pouvant être des peptides antimicrobiens luttant contre l’infection.
On voit sur ce schéma l’adéquation entre l’identification d’un pathogène et en quoi nos cellules vont être capables de lutter contre ce pathogène car il y aura la reconnaissance, l’activation des cellules immunitaires et la production de substances antimicrobiennes capables de se fixer sur la membrane

d’une bactérie et de former des pores à la membrane de la bactérie et de lyser la bactérie.
First data arguing that LPS is à ligand of TLR (4)
$
Sur ce schéma indiqué par la flèche bleue et à l’époque (en 1997), on n’avait toujours pas identifier quel pouvait être le ligand chez l’homme de TLR4. Un des ligands possibles suggéré et mis sur la scène : le LPS (pour les raisons précédentes : possède des sucres et du gras). Ce LPS pourrait être un très bon ligand pour l’activation de TLR4 au niveau du système immunitaire. C’est un constituant majeur des bactéries gram négatives telles que e.coli (utilisée par Hoffman dans son expérience). Encore fallait-il le prouver.
Un des premiers articles ayant prouvé que le LPS était probablement un ligand de TLR4 chez l’homme, souris (mammifères donc) a été publié dans Science dont l’auteur n’est autre que Bruce Beutler. Il a reçu un prix Nobel en même temps qu’Hoffman (cf ci-dessus). Une expérience est menée sur des souris dont le gène TLR4 est muté (appelées C3H/Hej). On observe que ces souris n'étaient pas capables d'induire une réponse anti-infectieuse lorsqu'on leur administre le LPS. Ce qui laissait subodorer que le LPS serait un ligand de TLR4.
Le vrai article ayant confirmé cela vient d’Akira :

Shizuo Akira (une figure emblématique de l’II, à solliciter si besoin d’un stage sur ce thème) et son groupe de recherche japonais, auteurs de 800 publications sur l'immunologie innée mènent des expériences 24h/24. Des souris mutantes (TLR4-/-) ont été exposés à du LPS. Au contact du LPS, les souris Wild type ont une très bonne réponse de ses cellules avec une grande production de cytokines de type TNFα. En revanche, chez les souris knock out (rond blanc), la production de TNFα induite est fortement diminuée (réponse réduite). Cette expérience a permis d'identifier le ligand du TLR4 le LPS. Une bactérie qui exprime du LPS sera reconnu par le TLR4 exprimés par les cellules immunitaires. Ce qui va induire la production des cytokines et l'activation d'autres cellules immunitaires qui vont phagocyter la bactérie.
LPS (Lipopolysaccharide) component of Gram- bacteria
Le LPS est exprimé en grande majorité par les bactéries GRAM-. Ceci explique le principe de la coloration. Le LPS empêche le colorant de pénétrer la bactérie et lorsqu'on ajoute la -mot inaudible- elle devient rose au lieu d'être violette. (Vu avec Jaffar Bandjee). Le SI reconnaît la bactérie GRAM- par le LPS. Les GRAM+ seront reconnus par les lipoprotéines, l'acide lipotéichoïque et les peptidoglycanes (rouge) fortement exprimés.
4. Les différents PRR :
Several PRRs (many are TLR) are involved against every single pathogen (PAMPs)
Rôle des différents TLR (il y en a 13, arrêt à 11 dans ce tableau), à retenir : TLR4 : reconnaissance LPS bactérien. On retrouve aussi les TLR dans la reconnaissance des virus, champignons, parasites. Même en ayant un jeu de 11 voire 13 TLR, on voit que cela est suffisant pour reconnaître les 4 grandes familles d’éléments pathogènes. Ces même TLR jouent exactement le même rôle dans la reconnaissance de la protéine amyloïde, prion, HMGB1 … Même en ayant si peu de TLR (que 13) : c’est suffisant pour reconnaître toute la complexité des pathogènes car la reconnaissance repose uniquement sur des motifs, des lipides, des saccharides qui ne sont normalement pas exprimés par nos cellules.

Si on prend un pathogène, il est important de ne pas avoir qu’un PRR capable de le reconnaître. Si il y a une mutation dans ce gène codant pour ce PRR, il n’y aura pas de survie et le sujet sera souvent atteint d’infections.
Même si on a une mutation dans TLR4 : ex : gram negative : il reste TLR9,2 et 5 qui vont participer à la reconnaissance et élimination de cette bactérie.
Les autres récepteurs impliqués dans la reconnaissance des pathogènes qu’il faut peut-être mentionner : RIG1 et MDA5, NALP3, NOD 1 et 2. Il y a à peu près une trentaine, quarantaine de PRR aujourd’hui en plus des TLR capables de reconnaître les éléments pathogènes (et aussi le complément sur lequel on y revient plus bas).
Ces PRR doivent être présents à la membrane des cellules et à l’intérieur au niveau du RE ou du phagolysozome car jouant un rôle important dans la détection intracellulaire (PRR cytosoliques) des virus.
Le peptidoglycane (PGN) des bactéries GRAM+ est reconnu par le TLR2. Les virus à ADN sont reconnus par TLR9. Les virus à ARN sont reconnus par TLR3 et TLR7.
Une grande famille de TLR est donc impliqué dans la reconnaissance des pathogènes: -de TLR 1 à 11 chez l'homme -de TLR 1 à 13 chez la souris
Ces PRR de type TLR ont la capacité de reconnaître sélectivement différents éléments pathogènes. Dans la grande majorité des cas, les TLR sont exprimés à la membrane pour détecter des bactéries extracellulaires. D'autres PRR cytosoliques, sont exprimés à l’intérieur des cellules pour détecter les virus et les bactéries intracellulaires. Ex: RIG-1 et MDA5 Mycobacterium tuberculosis est un exemple de bactérie intracellulaire qui se cache dans les macrophages et rendre le patient totalement immunodéprimé. Connaitre un peu bactérie gram - sur le schéma.
Co-engagement of many PRRs will boost innate
immune response to clear pathogens Représentation
schématique du tableau précédent
CONNAITRE PAR COEUR
mais surtout que LPS = TLR4 ADN = TLR9 ARN = TLR7
D. exemple de PRR: Les Toll Like Receptor (TLR) : A RETENIR
TLR signaling pathways in macrophages will alert surrounding cells (role of cytokin
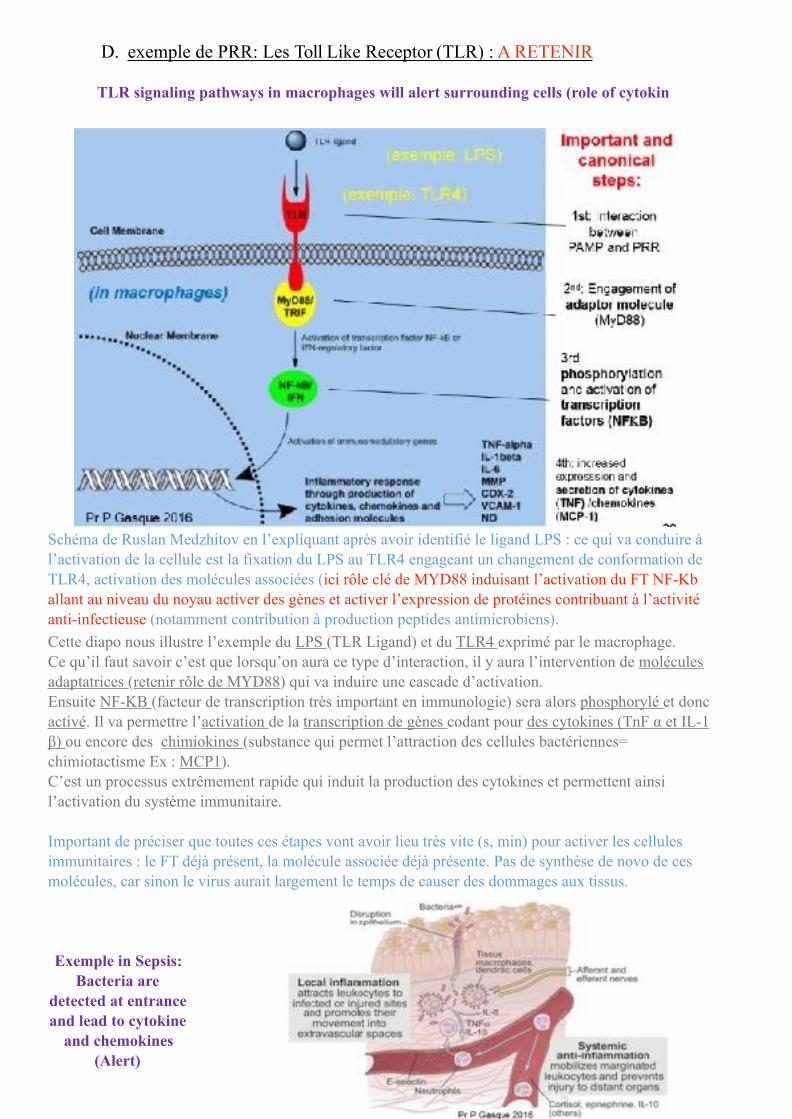
Schéma de Ruslan Medzhitov en l’expliquant après avoir identifié le ligand LPS : ce qui va conduire à l’activation de la cellule est la fixation du LPS au TLR4 engageant un changement de conformation de TLR4, activation des molécules associées (ici rôle clé de MYD88 induisant l’activation du FT NF-Κb allant au niveau du noyau activer des gènes et activer l’expression de protéines contribuant à l’activité anti-infectieuse (notamment contribution à production peptides antimicrobiens). Cette diapo nous illustre l’exemple du LPS (TLR Ligand) et du TLR4 exprimé par le macrophage. Ce qu’il faut savoir c’est que lorsqu’on aura ce type d’interaction, il y aura l’intervention de molécules adaptatrices (retenir rôle de MYD88) qui va induire une cascade d’activation. Ensuite NF-KB (facteur de transcription très important en immunologie) sera alors phosphorylé et donc activé. Il va permettre l’activation de la transcription de gènes codant pour des cytokines (TnF α et IL-1 β) ou encore des chimiokines (substance qui permet l’attraction des cellules bactériennes= chimiotactisme Ex : MCP1). C’est un processus extrêmement rapide qui induit la production des cytokines et permettent ainsi l’activation du système immunitaire.
Important de préciser que toutes ces étapes vont avoir lieu très vite (s, min) pour activer les cellules immunitaires : le FT déjà présent, la molécule associée déjà présente. Pas de synthèse de novo de ces molécules, car sinon le virus aurait largement le temps de causer des dommages aux tissus.
Exemple in Sepsis: Bacteria are
detected at entrance and lead to cytokine
and chemokines (Alert)

-On a sur cette diapo l’exemple d’un sepsis (infection systémique par une bactérie). On aura une très forte concentration de bactérie dans le sang. Le patient peut très rapidement décéder mais il décédera que si son système immunitaire n’est pas efficace. Ex: VIH, immune déficience de manière globale.
-Pour résumer le mécanisme d’action, une bactérie pénètre à l’intérieur d’une muqueuse fragilisée au niveau de la peau. Dans la majorité des cas il n’y aura pas de sepsis. Il y a un mécanisme de barrière physique de la peau qui va nous protéger de ces infections, mais si elle a subi des dommages et qu’elle est abrasée, il y aura pénétration de la bactérie. Les cellules au niveau de l’épiderme : kératinocytes, mélanocytes, cellules dendritiques, langerhans vont reconnaître ce constituant bactérien car ont tous TLR4. Elles vont s’activer car ayant reconnu le LPS de la bactérie et vont se mettre à produire des facteurs antimicrobiens, des cytokines et chemokines (ou chemoattractant) qui vont permettre d’attirer les macrophages ou neutrophiles qui sont dans la circulation sanguine. L’infiltration des macrophages et neutrophiles ne se fera qu’à l’endroit où il y a l’infection bactérienne car si cette infiltration ne se faisait pas à cet endroit mais ce faisait de manière sporadique, vous diluez les effets des cellules immunitaires dans la lutte contre cet agent infectieux. Ces cellules vont alors infiltrer des tissus sains n’ayant jamais rien demandé et là il faut que le SI soit rapidement mobilisé sur le site infectieux et nulle part ailleurs. Les LPS de ces bactéries vont activer les macrophages. Ces macrophages vont produire beaucoup de cytokines (TnF α et IL-1 β) et des chimioquines (IL-8). Ces substances vont diffuser au niveau des vaisseaux. Ces vaisseaux sont des sources très riches en cellules immunitaires (macrophages, neutrophiles, monocytes) qui vont alors percevoir le signal d’infection et infiltrer le site d’infection pour aller lutter contre l’infection bactérienne. - Si votre patient est immuno déprimé ou si la charge bactérienne est trop importante, il ne pourra pas lutter. Il y aura deux complications : les bactéries libèrent beaucoup de LPS et le système immunitaire qui détruit nos propres cellules. En général le patient est en choc septique, il est très vite en réanimation car il aura « un multi organ failure » (tous les organes principaux vont lâcher) il faut rapidement le maintenir en lui donnant des antibiotiques. Les antibiotiques tuant les bactéries vont libérer encore plus de LPS.
RESUME 1: - Toutes nos cellules sont capables de détecter une infection (toutes sans exception : même les
neurones, spermatozoïdes, ovocytes, cellules rénales). - Toutes nos cellules sont capables d’exprimer des sensors (PRRs) pour détecter les
pathogènes microbiens tels que les PAMPs. - Les PRRs peuvent être extracellulaire (pour détecter les bactéries, les parasites) ou
intracellulaire (pour les virus, bactéries intracellulaires). - Les PRRs sans exception sont capables également de détecter des facteurs dérivés de l’hôte
(débris cellulaires appelés DAMPs ou alarmins). - Les PRRs vont engager des voies de signalisation. - Ces voies de signalisation vont permettre de libérer des cytokines et des chimiokines pour
alerter le système immunitaire. - Les cytokines n’ont aucun effet sur les virus.
II Virus
Dans cette partie, on verra l’historique des mécanismes qui ont été mis en exergue concernant l’élimination plus particulièrement des virus, car comme vous le savez un virus ne circule pas à l’extérieur comme une

bactérie, un virus est nécessairement à l’intérieur de nos propres cellules. C’est pour ça que les mécanismes de reconnaissance et d’élimination d’un virus sont des mécanismes très particuliers, parce qu’il faut que notre système immunitaire (=SI) soit capable de voir la cellule infectée car il ne verra pas le virus.
Dans cette partie, contrairement aux mécanismes de reconnaissance des bactéries avec les LPS, ici le point focale, c’est que le SI va reconnaître toujours un sucre mais qui va être ici soit de l’ARN sous forme viral soit de l’ADN viral. Si on prend l’exemple du virus du chikungunya, il y aura énormément d’ARN, il y aura peu de protéines virales, c’est pour ça que notre SI va s’intéresser à reconnaître ce composé (=ARN) plutôt que les protéines.
Model to study the innate immune response against viruses (e.g. Influenzae)
" -Comment éliminer un virus ?
La détection d’interférons = interfère avec la réplication d’un virus.
-Jean Lindenmann, prix Nobel, a déposé un modèle en 1957 qui a permis de répondre à cette question. -Pour son expérience il a utilisé un incubateur naturel : un œuf qui contient l’embryon dans lequel il a pu injecter des virus (Remarque : La protein majeure du blanc d’oeuf est le lysozyme,
qui a pour function de protéger l’embryon qui est a.n. du jaune contre les infections.). -Il a voulu identifier les substances qui étaient libérées par l’embryon pour essayer de se protéger de l’infection virale. Il savait que le lysozyme protégeait l’embryon contre les infections bactériennes et fongiques mais pas contre les infections virales.
Very simple experiment to identify a molecule called interferon (alpha/beta)

L’expérience est illustrée ci-contre : il a injecté dans un premier œuf (en bleu) le virus influenza (virus de la grippe) inactivé. -Il a considéré que ce virus serait capable d’induire une réponse permettant de protéger un autre embryon. -Suite à cette expérience il a pris l’extrait de l’embryon exposé au virus la première fois et il l’a mis sur un autre œuf dans lequel il a injecté un virus vivant (capable de tuer l’embryon). -On remarque dans ce deuxième œuf que l’embryon est protégé. -Donc il a bien identifié un facteur qui est libéré dans la 1ere expérience et qui est capable de protéger le 2eme embryon. - Cette substance s’appelle l’interféron. -Il a montré que ce facteur est soluble, sensible à la trypsine et labile au contact de la chaleur. -L’interféron est donc une protéine (trypsine=protéase ⇒ sensible à la trypsine, donc c’est une protéine).
IFN-type I (alpha and beta) mode of action
$ -Sur ce schéma on voit que la 1ere cellule est infectée par un virus, cette cellule va répondre en produisant ce facteur interféron.
-Cet interféron va par la suite induire la production de protéines antivirals(=ISG : Interferon Stimulates Genes) en se fixant sur une autre cellule. Ce qui fait que le virus ne pourra pas infecter l’autre cellule.
IFN type I drives an anti-viral state in neighboring cells
-Retenir la 3eme étape sur cette diapo : quand l’interféron se fixe sur son récepteur, il va induire la production d’ISG (interferon stimulated G).
ISG (IRG) and antiviral effects
Illustration de toutes les ISG. -Retenir la première ISG (encadré rouge) qui code pour une RNASE (OAS, RNASEL). -L’interféron va induire la production de ces ISG. -Ces ISG vont empêcher les virus de se
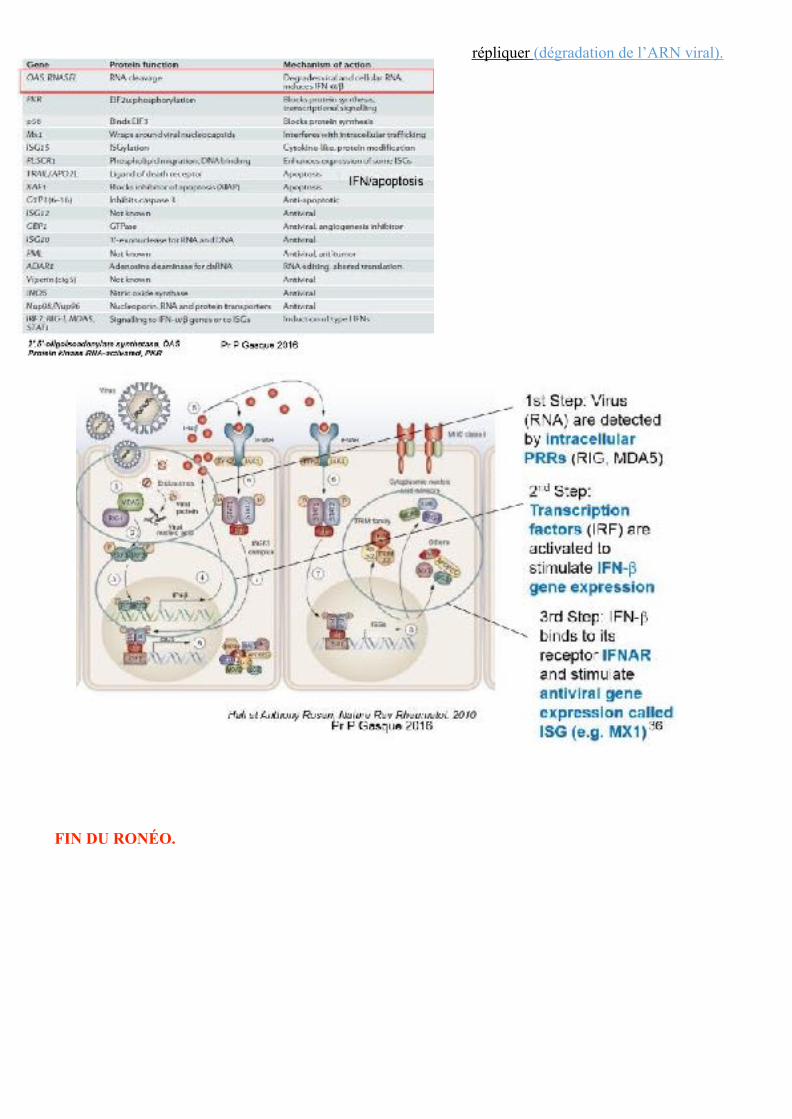
répliquer (dégradation de l’ARN viral).
FIN DU RONÉO.
RESUME 2 : −Les virus sont des pathogènes intracellulaires −Les PRRs comme TLR3/7 et RIG-I, ou encore MDA5 reconnaissent les virus −Les voies de signalisations sont activées pour induire l'expression d'interférons de type I (alpha et beta) −Les interférons de type I se fixent à leurs récepteurs et engagent des voies de signalisations plus poussées −Les ISG produits contribuent à limiter l'infection. −Mais qu’en est –il des bactéries?
III .Bactéries
-Contrairement aux virus, les bactéries vont être éliminées par phagocytose et par lyse Engagement of one or many PRRs will induce lysis/phagocytosis of pathogens?

A. Phagocytose
-Les bactéries vont être détectées par les macrophages, puis phagocytées. A l'instar des virus, nous aurons un ligand, un pathogène et un récepteur. Mais cette fois, il y aura induction de la lyse et de la phagocytose. Le macrophage n’est pas capable de reconnaître un virus directement puisque les virus sont à l’intérieur des cellules. Il va reconnaître la cellule infectée, phagocyter cette cellule infectée. C’est important d’avoir une phagocytose d’une cellule infectée par un virus parce que si on a une lyse d’une cellule infectée par un virus, on répandra encore plus ce virus. En le phagocytant, on tuera les virus dans la cellule infectée.
Ilya Ilyich Mechnikov (Élie Metchnikoff) (15 May 1845 – 15 July 1916) He received the Nobel Prize in Medicine in 1908, shared with Paul Ehrlich, for his work on phagocytosis (starfish) at the root of our current understanding of innate immunity. He is also credited by some sources with coining the term gerontology in 1903, for the emerging study of aging and longevity.
Elie Metchnikoff a travaillé avec Pasteur sur la compréhension des mécanismes de phagocytose. Il a identifié dans les étoiles de mer de grandes cellules grises (image du milieu) avec à l’intérieur des bactéries (en rouge). C’est en fonction du temps d’incubation de ces cellules avec la bactérie qu’il y avait la formation de plus grand pool de ces bactéries intracellulaires et surtout la formation de ces larges vésicules denses rouges (=vésicules de phagocytose). La bactérie va être très rapidement phagocyté, ensuite elles vont être dirigées vers ces vésicules (qu’on appelle des phagolysosomes) pour être détruite. Il a donc décrit la capacité des cellules à ingérer des pathogènes. Il est considéré comme le père de l’immunologie.
Depuis il y a eu énormément d’études où on a utilisé cette fois-ci des cellules humaines sur le même principe.
Pathogens and host-derived DAMPs recognized by MØ

"
MØ: Sensing and responding to all kind of danger!!!
On s’est rendu compte que cette fonction n’était pas uniquement contre les bactéries, mais aussi contre les débris de l’hôte. Image de gauche : Capacité des macrophages à reconnaître des gouttelettes lipidiques et de les éliminer. Si on ne les élimine pas, on peut aboutir à l’athérosclérose (accumulation de lipides dans les artères).
Image de droite : La goutte est-elle due à l’accumulation d’acide urique ? Non, ce n’est pas l’acide urique qui est à l’origine de la maladie. Le macrophage est minuscule par rapport au cristal d’acide et il tente de le ramener vers lui pour le phagocyter. Cependant il n’y arrive pas et va se mettre à libérer énormément de composés toxiques par hyper activation.
-Les cellules qui vont être impliquées dans la phagocytose jouent un rôle extrêmement important notamment pour tout ce qui est pathogène extracellulaire Rappel : quand il dit pathogène ce n’est pas uniquement microbes mais aussi les lipides, l’acide urique…
Phagocytosis of pathogens by macrophages
Ce rôle de phagocytose est d’autant plus important qu’il va permettre d’initier la réponse immunitaire adaptative. Si on a pas de phagocytose de pathogènes, on n’aura pas d’initiation de la réponse immunitaire adaptative.
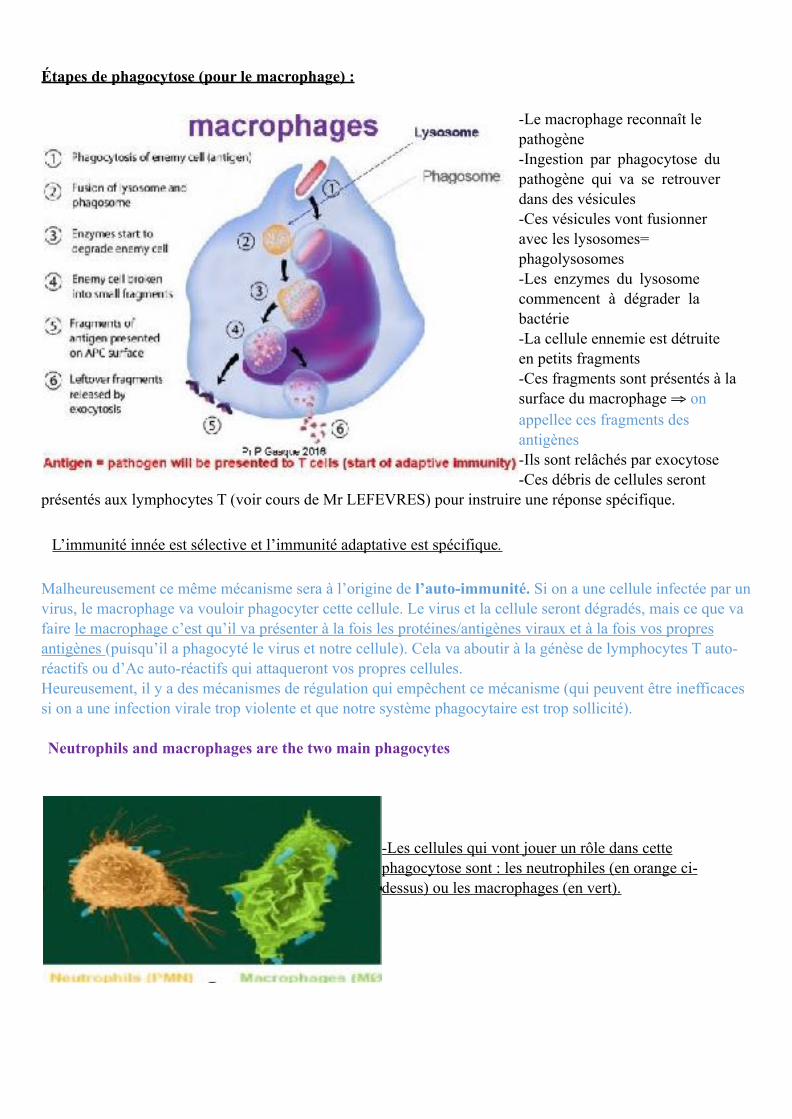
Étapes de phagocytose (pour le macrophage) :
-Le macrophage reconnaît le pathogène -Ingestion par phagocytose du pathogène qui va se retrouver dans des vésicules -Ces vésicules vont fusionner avec les lysosomes= phagolysosomes -Les enzymes du lysosome commencent à dégrader la bactérie -La cellule ennemie est détruite en petits fragments -Ces fragments sont présentés à la surface du macrophage ⇒ on appellee ces fragments des antigènes -Ils sont relâchés par exocytose -Ces débris de cellules seront
présentés aux lymphocytes T (voir cours de Mr LEFEVRES) pour instruire une réponse spécifique.
L’immunité innée est sélective et l’immunité adaptative est spécifique.
Malheureusement ce même mécanisme sera à l’origine de l’auto-immunité. Si on a une cellule infectée par un virus, le macrophage va vouloir phagocyter cette cellule. Le virus et la cellule seront dégradés, mais ce que va faire le macrophage c’est qu’il va présenter à la fois les protéines/antigènes viraux et à la fois vos propres antigènes (puisqu’il a phagocyté le virus et notre cellule). Cela va aboutir à la génèse de lymphocytes T auto-réactifs ou d’Ac auto-réactifs qui attaqueront vos propres cellules. Heureusement, il y a des mécanismes de régulation qui empêchent ce mécanisme (qui peuvent être inefficaces si on a une infection virale trop violente et que notre système phagocytaire est trop sollicité).
Neutrophils and macrophages are the two main phagocytes
-Les cellules qui vont jouer un rôle dans cette phagocytose sont : les neutrophiles (en orange ci- dessus) ou les macrophages (en vert).

Neutrophils phagocytose and kill bacteria (role of lysozyme)
-Sur la diapo ci-dessus on observe la bactérie(en jaune), le neutrophile(en bleu) et son noyau (en mauve). -Ce qu’on voit en zoomant c’est que cette bactérie est à l’intérieur d’une vacuole, contenu dans le neutrophile. -Ces neutrophiles vont libérer de nombreuses substances pour le lysozyme et de l’eau oxygénée. La bactérie pourra donc être éliminée. - Ils contiennent d’autres molécules telles que les défensines, qui sont des molécules antimicrobiens, qui vont se fixer à la bactérie et former des pores à la membrane de la bactérie et ainsi lyser la bactérie.
RESUME 3 : -Les macrophages et neutrophiles sont des phagocytes -Ils expriment des PRRs capable de reconnaître le pathogène et d’induire la phagocytose -Ils vont répondre également à des chimioquines pour aller au site infectieux -Ils vont ingérer le pathogène (bactérie, parasite) -Ils vont digérer le pathogène -Les pathogènes vont être présentés aux lymphocytes T pour instruire l’immunité adaptative
B. Facteurs du complément
1. Introduction, historique, généralités
-Les macrophages et neutrophiles ont besoin de facteurs du complément pour phagocyter et tuer les pathogènes.
-Le complément est un élément clé de l’immunité innée. Si l’on a une déficience en complément, il y a de forte chance qu’on ne puisse pas survivre. On peut le détecter très tôt en observant chez un patient qui a des maladies infectieuses à répétition, on peut donc essayer de compenser cette déficience en donnant du sang. (Car les compléments se trouvent dans le sang)
Macrophages and neutrophils will require complement to phagocytose and kill pathogens
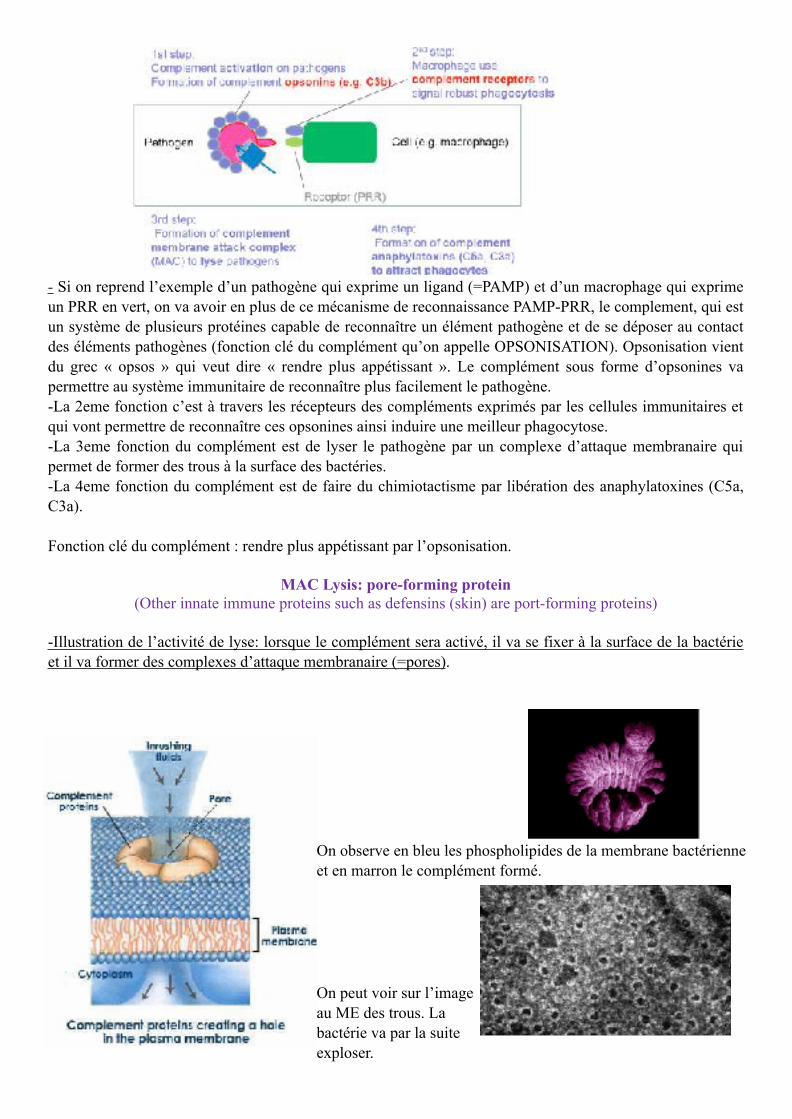
- Si on reprend l’exemple d’un pathogène qui exprime un ligand (=PAMP) et d’un macrophage qui exprime un PRR en vert, on va avoir en plus de ce mécanisme de reconnaissance PAMP-PRR, le complement, qui est un système de plusieurs protéines capable de reconnaître un élément pathogène et de se déposer au contact des éléments pathogènes (fonction clé du complément qu’on appelle OPSONISATION). Opsonisation vient du grec « opsos » qui veut dire « rendre plus appétissant ». Le complément sous forme d’opsonines va permettre au système immunitaire de reconnaître plus facilement le pathogène. -La 2eme fonction c’est à travers les récepteurs des compléments exprimés par les cellules immunitaires et qui vont permettre de reconnaître ces opsonines ainsi induire une meilleur phagocytose. -La 3eme fonction du complément est de lyser le pathogène par un complexe d’attaque membranaire qui permet de former des trous à la surface des bactéries. -La 4eme fonction du complément est de faire du chimiotactisme par libération des anaphylatoxines (C5a, C3a).
Fonction clé du complément : rendre plus appétissant par l’opsonisation.
MAC Lysis: pore-forming protein (Other innate immune proteins such as defensins (skin) are port-forming proteins)
-Illustration de l’activité de lyse: lorsque le complément sera activé, il va se fixer à la surface de la bactérie et il va former des complexes d’attaque membranaire (=pores).
On observe en bleu les phospholipides de la membrane bactérienne et en marron le complément formé.
On peut voir sur l’image au ME des trous. La bactérie va par la suite exploser.

Complement : Three main effector functions
-Ce schéma illustre les 3 fonctions que l’on doit retenir du système de compléments: à savoir le complément joue un rôle clé dans la phagocytose, dans la lyse et dans le chimiotactisme.
HISTORIQUE DU COMPLEMENT
Belgian immunologist (1870–1961) Bordet was born in Soignies, Belgium, and graduated in medicine from Brussels University in 1892. In 1894
joined the Pasteur Institute, Paris, where he worked under the bacteriologist Elie Metchnikoff. In collaboration with Octave Gengou, Bordet discovered that in an immunized animal the antibodies produced by the immune response work in conjunction with another component of blood (which Bordet termed ‘alexin’ but which is now called ‘complement’) to destroy foreign cells that invade the body. This component, Bordet found, was present in both immunized and nonimmunized animals and was destroyed by heating to over 55°C. In 1901 Bordet left Paris to found and direct the Pasteur Institute in Brussels and in 1907 he was appointed professor of pathology and bacteriology at Brussels University. In 1906 Bordet isolated the bacteria responsible for cough, which is named after him: Bordetella (Haemophilus) pertussis. For his discovery of complement and other contributions to medicine, he was awarded the 1919 Nobel Prize in physiology or medicine.
-Bordet a été le premier à caractériser le complement. On le connait surtout pour ses travaux en microbiologie : il a identifié la bactérie Bordetella (⇒ Prix Nobel). En incubant le sérum d’animaux immunisés avec des bactéries, il se rendit compte qu’il y avait dans ce sérum autre chose que des Ac, qui étaient capable de favoriser la lyse des bactéries. Il a appelé cette substance « complément » car c’était une substance capable de complémenter l’action des Ac.
Aujourd’hui on sait que le système du complément est composé de 30 substances.
Qu’est-ce que le Complément?
• Système ancestral de l’immunité innée (apparu avant les anticorps) Remarque : Il y a très longtemps de cela, un virus a infecté nos ancêtres et nous a donné la possibilité de générer plusieurs types d’anticorps grâce à deux gènes REC 1et 2 (recombinases)
• Produit par le foie et autres tissus (adipeux⇒ role important dans la lutte contre agents infectieux, toute espèce confondue)
• Reconnaît des éléments du non-soi et débris cellulaires • Implique des protéines C3, C4, C5 thioesther capables de se fixer à des éléments du non-soi La protéine C3 est l’élément central des compléments car c’est elle qui va s’opsoniser et se déposer à la surface des pathogènes avec l’aide des autres protéines.
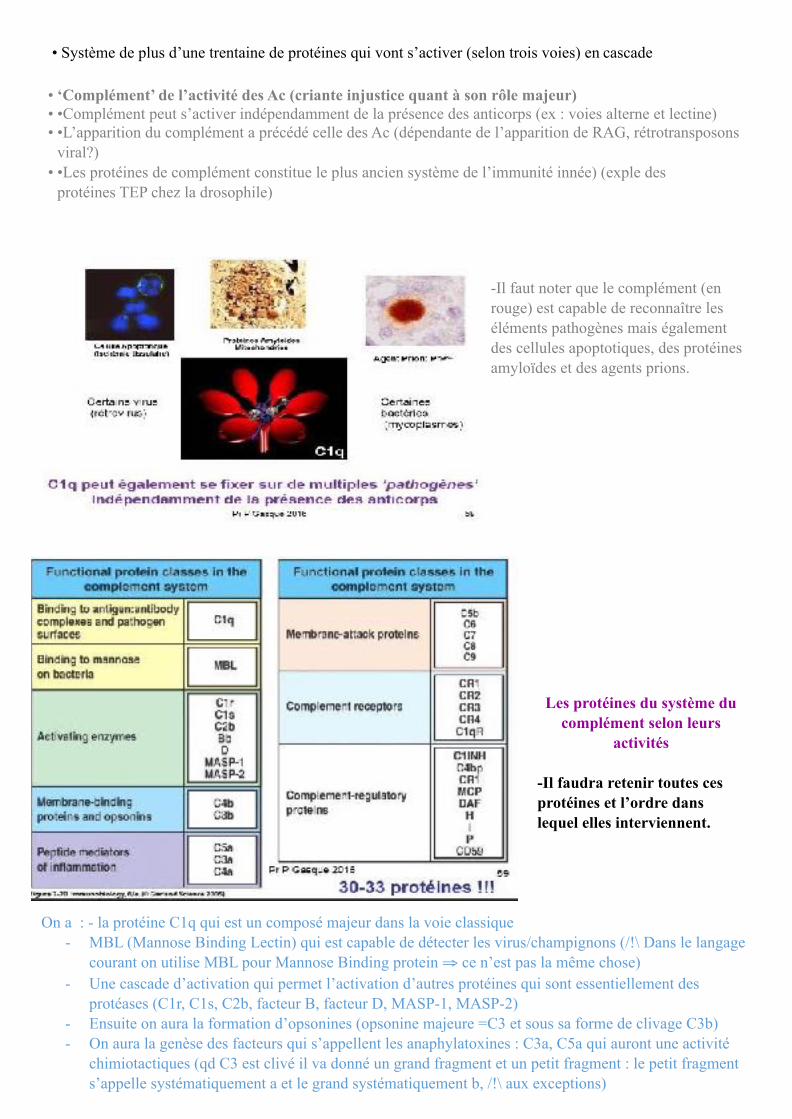
• Système de plus d’une trentaine de protéines qui vont s’activer (selon trois voies) en cascade
• ‘Complément’ de l’activité des Ac (criante injustice quant à son rôle majeur) • •Complément peut s’activer indépendamment de la présence des anticorps (ex : voies alterne et lectine) • •L’apparition du complément a précédé celle des Ac (dépendante de l’apparition de RAG, rétrotransposons
viral?) • •Les protéines de complément constitue le plus ancien système de l’immunité innée) (exple des
protéines TEP chez la drosophile)
-Il faut noter que le complément (en rouge) est capable de reconnaître les éléments pathogènes mais également des cellules apoptotiques, des protéines amyloïdes et des agents prions.
Les protéines du système du complément selon leurs
activités
-Il faudra retenir toutes ces protéines et l’ordre dans lequel elles interviennent.
On a : - la protéine C1q qui est un composé majeur dans la voie classique - MBL (Mannose Binding Lectin) qui est capable de détecter les virus/champignons (/!\ Dans le langage
courant on utilise MBL pour Mannose Binding protein ⇒ ce n’est pas la même chose) - Une cascade d’activation qui permet l’activation d’autres protéines qui sont essentiellement des
protéases (C1r, C1s, C2b, facteur B, facteur D, MASP-1, MASP-2) - Ensuite on aura la formation d’opsonines (opsonine majeure =C3 et sous sa forme de clivage C3b) - On aura la genèse des facteurs qui s’appellent les anaphylatoxines : C3a, C5a qui auront une activité
chimiotactiques (qd C3 est clivé il va donné un grand fragment et un petit fragment : le petit fragment s’appelle systématiquement a et le grand systématiquement b, /!\ aux exceptions)
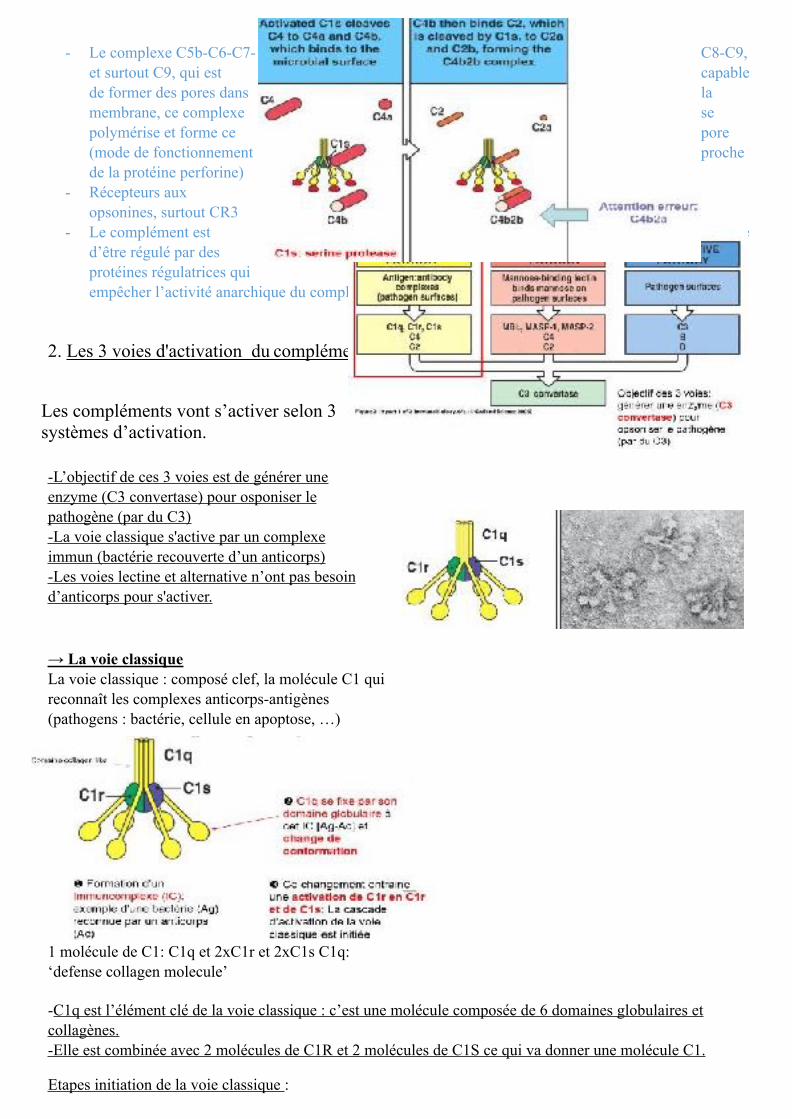
- Le complexe C5b-C6-C7- C8-C9, et surtout C9, qui est capable de former des pores dans la membrane, ce complexe se polymérise et forme ce pore (mode de fonctionnement proche de la protéine perforine)
- Récepteurs aux opsonines, surtout CR3
- Le complément est capable d’être régulé par des protéines régulatrices qui vont empêcher l’activité anarchique du complément = CD59
2. Les 3 voies d'activation du complément
Les compléments vont s’activer selon 3 systèmes d’activation.
-L’objectif de ces 3 voies est de générer une enzyme (C3 convertase) pour osponiser le pathogène (par du C3) -La voie classique s'active par un complexe immun (bactérie recouverte d’un anticorps) -Les voies lectine et alternative n’ont pas besoin d’anticorps pour s'activer.
→ La voie classique La voie classique : composé clef, la molécule C1 qui reconnaît les complexes anticorps-antigènes (pathogens : bactérie, cellule en apoptose, …)
1 molécule de C1: C1q et 2xC1r et 2xC1s C1q: ‘defense collagen molecule’
-C1q est l’élément clé de la voie classique : c’est une molécule composée de 6 domaines globulaires et collagènes. -Elle est combinée avec 2 molécules de C1R et 2 molécules de C1S ce qui va donner une molécule C1.
Etapes initiation de la voie classique :

1-Formation d’un Immuncomplexe (IC) : exemple d’une bactérie (Ag) reconnue par un anticorps (Ac) 2-C1q se fixe par son domaine globulaire à cet IC [Ag-Ac] et change de conformation (il faut qu’au moins 2 domaines globulaires soient engagés) 3-Ce changement entraîne une activation de C1r en C1r (activé) et de C1s (activé): La cascade d’activation de la voie classique est initiée (sur le schéma : qd on veut indiquer qu’une molécule est active, on lui met une barre au dessus) C1r et C1s sont couplées à la molécule de c1q. Ce couplage formant la molécule C1. C1 est en fait constituée d’une molécule de c1q, de deux molécules de c1r et de deux molécules de c1s. Cls est une serine protéase.
Nous avons ici les molécules associées au pathogène en rouge, c1q qui s’est fixé au contact de l’Immuncomplexe, les molécules c1r et c1s qui s’activent et en s’activant la molécule de c1s clive c4 en c4a et c4b, c4b se fixant au contact du pathogène. C1s clive ensuite c2 en c2a et c2b.
-C1s clive C4 en C4a (qui diffuse) et C4b (qui se fixe au contact du pathogène par sa fonction thioesther) -Ensuite C1s clive C2 en C2a (qui se fixe au contact du pathogène) et C2b (qui diffuse) -Formation de C4b2a= C3 convertase Le prof nous prévient ici d’une erreur récurrente dans les livres : ce n’est pas c4b2b mais c4b2a (et il risque de nous poser la question) ! De même il s’agit du c2b et non du c2a qui va diffuser. Le complexe c4b2a généré est en fait la c3 convertase classique puisqu’elle aura été initiée par le composé c1q de la voie classique (c4b2a= C3
convertase classique). Question/réponse : c’est c1s qui clive à la fois c4 et c2. C1s est une sérine protéase qui clivera donc une sérine.
ATTENTION erreur sur le schéma : C4b2a et non C4b2b qui est la C3 convertase classique.
-C4b2a (C3 convertase classique) va cliver C3 en C3a et C3b (=opsonines). -C4b2a3b (C5 convertase) va convertir C5 par la suite.
Avec une seule molécule C1q, on aboutit à la formation de 1000 molécules de C3.
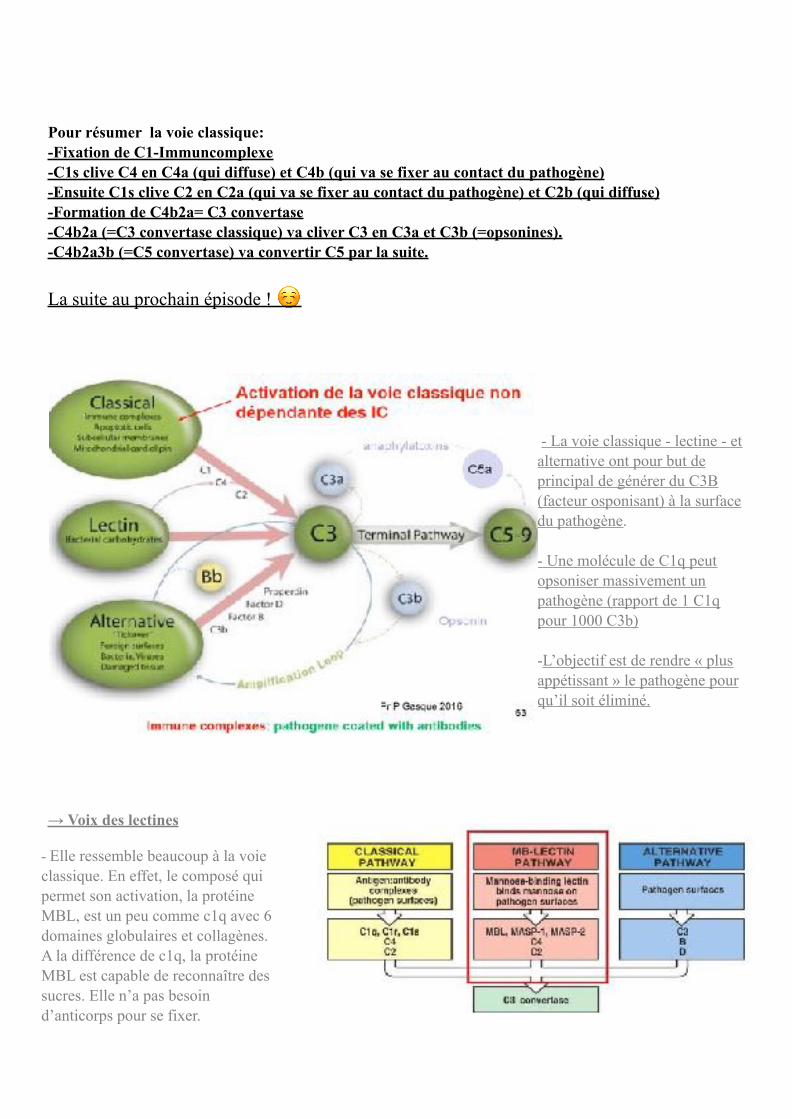
Pour résumer la voie classique: -Fixation de C1-Immuncomplexe -C1s clive C4 en C4a (qui diffuse) et C4b (qui va se fixer au contact du pathogène) -Ensuite C1s clive C2 en C2a (qui va se fixer au contact du pathogène) et C2b (qui diffuse) -Formation de C4b2a= C3 convertase -C4b2a (=C3 convertase classique) va cliver C3 en C3a et C3b (=opsonines). -C4b2a3b (=C5 convertase) va convertir C5 par la suite.
La suite au prochain épisode ! ☺
- La voie classique - lectine - et alternative ont pour but de principal de générer du C3B (facteur osponisant) à la surface du pathogène.
- Une molécule de C1q peut opsoniser massivement un pathogène (rapport de 1 C1q pour 1000 C3b)
-L’objectif est de rendre « plus appétissant » le pathogène pour qu’il soit éliminé.
→ Voix des lectines
- Elle ressemble beaucoup à la voie classique. En effet, le composé qui permet son activation, la protéine MBL, est un peu comme c1q avec 6 domaines globulaires et collagènes. A la différence de c1q, la protéine MBL est capable de reconnaître des sucres. Elle n’a pas besoin d’anticorps pour se fixer.

-Les enzymes reconnaissent des sucres à la surface du pathogène. Ces sucres sont de type mannose et permettent, lorsqu’ils sont exposés sur les pathogènes, la fixation de MBL. S’il y en a peu, elle ne se fixera pas mais s’il y en a beaucoup, elle se fixe et s’active.

Dans la figure remplacer C1s par MASP2 Attention erreur C4b2a : C3 convertase
-C1s et c1r sont des sérines protéases, mais ici on ne les appelle pas des c1r ou s mais des « MBL associated serine protease». On aura donc MASP1 homologue de c1r et MASP2 homologue de c1s. La fonction étant la même : ce sont des sérines protéases qui vont activer c4 en c4a et c4b, c2 en c2a et c2b et ainsi le complexe c4b2a généré est le même que celui de la voie classique et permettra l’activation et le clivage de c3 en c3a et c3b et à chaque fois pour une molécule de c4b2a, on aura 1000 molécules de c3b.
Rendez-vous au prochain cours de ce cher professeur pour découvrir la suite !

Annales :
2013-2014
1. Concernant l’immunité innée : A. Elle fait appel à des récepteurs invariants. B. Son apparition est concomitante à celle de l’immunité adaptative. C. Elle est capable de faire face à la diversité des microbes. D. Elle permet d’éliminer les cellules apoptotiques. E. Elle fait intervenir uniquement des récepteurs exprimés à la membrane.
31. Concernant le rôle des opsonines du complément dans l’activation leucocytaire? A. Elles rendent plus appétissant les pathogènes pour l’élimination par les macrophages. B. Elles permettent le recrutement des macrophages au site infectieux. C. Elles permettent de favoriser la phagocytose du pathogène. D. Elles permettent le clivage de la molécule C4 du complément. E. Elles contribuent à l’inhibition de la voie terminale du complément.
32. Remettre dans l’ordre chronologique la cascade d’activation de la voie classique du complément:
1. Le facteur C4 est clivé en C4a et C4b. 2. Le facteur C1 se lie à un agent pathogène. 3. C1r activé clive C1s. 4. Le facteur C2 est clivé en C2a et C2b pour former la C3 convertase. A. 2431 B. 3214 C. 2314 D. 3124 E. 4213
33. Concernant le rôle de la protéine « mannan binding lectin » au contact de Candida albicans :
A. Elle reconnaît les complexes antigènes-anticorps. B. Elle s’associe aux protéines C1r et C1s. C. Elle se fixe aux composés riches en mannose du pathogène. D. Elle induit la formation de la C4b2a, la C5 convertase. E. Elle permet l’activation de sérines protéases.
34. Concernant le rôle des anaphylatoxines du complément dans l’initiation de la réponse inflammatoire :
A. Elles stimulent le chimiotactisme des plaquettes. B. Elles activent et induisent le chimiotactisme des neutrophiles. C. Elles se fixent sur des récepteurs de type RCPG. D. Elles clivent les composés C3 et C5. E. Elles se fixent aux membranes des pathogènes.
2014-2015
1. L’immunité innée est un mécanisme de défense : A. spécifique des antigènes. B. qui empêche les réinfections. C. qui peut être impliqué dans l’élimination des tumeurs. D. qui fait appel à des agents chimiques non spécifiques.

E. Aucune des propositions ci-dessus.
2. Les PAMPs (motifs moléculaires associés aux pathogènes) sont : A. produits par l'organisme-hôte en réponse à une infection par des microbes. B. exprimés à la surface des cellules apoptotiques. C. communs à de grandes classes d'agents infectieux. D. des motifs qui mutent fréquemment pour permettre l’échappement aux systèmes de
reconnaissance de l’immunité innée. E. Aucune des propositions ci-dessus.
3. Les substances chimiques de l’immunité non spécifique comprennent: A. les interférons B. le lysozyme C. la cryptidine D. le facteur du complément C3b E. Aucune des propositions ci-dessus.
4. Les cellules phagocytaires comprennent: A. Les mastocytes B. Les basophiles C. Les lymphocytes NK D. Les éosinophiles E. Aucune des propositions ci-dessus.
6. Un macrophage : A. est une cellule monocytaire circulante. B. est un phagocyte. C. possède des lysosomes plus nombreux et plus complexes que les monocytes. D. secrète des médiateurs solubles agissant sur leucocytes. E. Aucune des propositions ci-dessus.
5. Remettez dans l’ordre chronologique, la séquence de la phagocytose : 1. Le phagosome fusionne avec des lysosomes. 2. Les bactéries se lient aux récepteurs des phagocytes. 3. Les produits de digestion sont libérés des cellules. 4. Les bactéries sont ingérées formant un phagosome. 5. Les enzymes lysosomales digèrent le matériel ingéré. A. 12345 B. 24153 C. 53142 D. 21453 E. 51243
7. Concernant les interactions entre les cellules endothéliales et les phagocytes : A. Les intégrines permettent des interactions fortes. B. La L-selectine est exprimée par les cellules endothéliales. C. Les sélectines interviennent dans les étapes de roulement (rolling). D. PECAM est exprimé par les cellules endothéliales et la plupart des leucocytes activés. E. Aucune des propositions ci-dessus.
8. Les récepteurs de reconnaissance de motifs moléculaires (PRRs) : A. reconnaissent des motifs moléculaires associés aux pathogènes (PAMPs). B. reconnaissent des motifs moléculaires associés à des signaux de danger (DAMPs).

C. sont présents exclusivement à la surface des cellules phagocytaires. D. peuvent être sécrétées. E. Aucune des propositions ci-dessus.
9. La lectine liant le mannose (MBL) : A. est un PRR endocytique. B. appartient à la famille des lectines. C. est synthétisée par les hépatocytes. D. joue un rôle important dans la reconnaissance des déterminants protéiques. E. Aucune des propositions ci-dessus.
10. La reconnaissance du LPS par le TLR4 : A. fait intervenir l’hétérodimère TLR4/TLR7. B. requiert le facteur MD-2. C. fait intervenir la molécule CD14 chez les monocytes. D. active la phagocytose. E. Aucune des propositions ci-dessus.
11. La signalisation intracellulaire des TLRs : A. nécessite des protéines adaptatrices qui interagissent avec les domaines TIR. B. L’adaptateur le plus commun des TLRs est MyD88. C. fait intervenir des protéines kinases. D. permet d’activer la voie des MAP kinases. E. Aucune des propositions ci-dessus.
12. Les fonctions immunes des TLRs sont, entre autre : A. d’induire une réponse immunitaire innée immédiate. B. d’activer les cellules dendritiques. C. d’induire la production de cytokines anti-inflammatoires. D. d’induire la présentation antigénique. E. Aucune des propositions ci-dessus.

13. Concernant l’activité bactéricide intracellulaire des phagocytes : A. Elle conduit toujours à la destruction de l’agent pathogène phagocyté. B. Elle fait intervenir des dérivés toxiques oxygénés et azotés. C. Elle s’effectue dans les lysosomes. D. Elle ne présente aucun danger pour les constituants de l’hôte. E. Aucune des propositions ci-dessus.
26. Qu’est-ce que le complément ? A. C’est un système de plusieurs protéines solubles qui s’activent en cascade. B. C’est un système ancestral de l’immunité adaptative. C. C’est un système qui reconnait spécifiquement des éléments du soi. D. C’est un système qui permet la lyse d’agents pathogènes. E. Aucune des propositions ci-dessus.
27. Remettez dans l’ordre chronologique la cascade d’activation de la voie classique du complément: 1. Le facteur C4 est clivé en C4a et C4b par C1s activé 2. Le facteur C1 se lie à un agent pathogène 3. C1r activé clive C1s 4. Le facteur C2 est clivé en C2a et C2b A. 2431 B. 3214 C. 2314 D. 3124 E. Aucune des propositions ci-dessus
29. Concernant les C3 convertases : A. La C3 convertase classique est la C4b2a. B. La C3 convertase alterne est la C3bBb. C. La voie lectine du complément conduit à la formation de la C3 convertase classique. D. La voie lectine et alterne du complément conduisent à la formation de la C3 convertase alterne. E. Aucune des propositions ci-dessus
2015-2016
55. Concernant le rôle des opsonines du complément dans l’activation leucocytaire : A. Il rend plus appétissant les pathogènes pour l’élimination par les macrophages B. Il permet le recrutement des macrophages au site infectieux C. Il permet de favoriser la phagocytose du pathogène. D. Il permet le clivage de la molécule C4 du complément. E. Il contribue à l’inhibition de la voie terminale du complément.
57. Concernant le rôle de l’anaphylotoxine C5a du complément dans l’initiation de la réponse inflammatoire : A. Elle stimule le chimiotactisme des plaquettes. B. Elle active et induit le chimiotactisme des neutrophiles. C. Elle se fixe sur un récepteur de type RCPG. D. Elle clive les composés C3 et C5. E. Elle se fixe à la membrane des neutrophiles
58. CD59 A. - est une petite protéine soluble qui se fixe à la molécule C5. B. – est couplée via un groupement GPI et bloque la lyse dépendante du complément. C. – est ancrée à la membrane et inhibe la polymérisation de C9.

D. – bloque le complexe d’attaque membranaire. E. – induit la libération d’histamine par les mastocytes.
2016-2017:
29. Concernant l’immunité innée: A. C’est un mécanisme de défense inactif contre les infections virales. B. Son fonctionnement repose essentiellement sur des facteurs moléculaires. C. Son activation peut contribuer au bon développement de la mémoire immunitaire. D. Le système du complément n’en fait pas parti. E. Aucune des propositions n’est exacte.
30. Concernant les PAMPs: A. Ce sont des motifs moléculaires associés aux pathogènes. B. Ce sont des motifs moléculaires associés à l’ironie hote. C. Les PRRs en font partie. D. Les acides lipothéchoïques en font partie. E. Aucune des propositions n’est exacte.
31. Concernant les LPS: A. C’est un PAMP exprimé par la majeure partie des bactéries gras négatives. B. Il est reconnu par un PRR de signalisation. C. Sa reconnaissance chez le macrophage fait intervenir le CD14. D. Sa présence dans le sang peut entrainer un choc anaphylactique. E. Aucune des propositions n’est exacte.
37. Concernant les TLRs: A. Ce sont des protéines à 7 domaines transmembranaire couplés aux protéines G. B. Le domaine TIR des TLR présente un site de fixation pour des molécules adaptatrices telles que MyD88. C. Le TLR4 est impliqué dans la reconnaissance des ARN double brin. D. Le TLR5 intervient dans la reconnaissance des bactéries flagellées. E. Aucune des propositions n’est exacte.































