Embed Size (px)
Citation preview

Variabilité morphologique, caryologique et enzymatique chez quelques populations dePhragmites australis (Cav.) Trin. ex SteudAuthor(s): Mohamed DjebrouniSource: Folia Geobotanica & Phytotaxonomica, Vol. 27, No. 1 (1992), pp. 49-59Published by: SpringerStable URL: http://www.jstor.org/stable/4181118 .
Accessed: 18/06/2014 11:15
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Springer is collaborating with JSTOR to digitize, preserve and extend access to Folia Geobotanica&Phytotaxonomica.
http://www.jstor.org
This content downloaded from 185.2.32.49 on Wed, 18 Jun 2014 11:15:59 AMAll use subject to JSTOR Terms and Conditions

MOHAMED DJEBROUNI
Laboratoire d'Ecogen6tique, Institut des Sciences de la Nature, U.S.T.H.B.B.P. 139 El Alia, 16111 Alger, Algerie
Variabilitd morphologique, caryologique et enzymatique chez quelques populations de Phragmites australis (CAV.) TRIN. ex STEUD.
KEYWORDS
Phragmites austrajis, Variability, Morphology, Karyology, Enzymes
ABSTRACT
Use of morphological, karyological and enzymatic approaches in the study of 11 populations of Phragmites australis (CAV.) TRIN. ex STEUD. originating from different geographic areas, climates and ecological conditions reveals a spatial differentiation of populations.
There is no relationship between the two levels of ploidy, 2n =48 and 2n=%9, and the habitat. In contrast, the climate and environmental conditions act to the selection of the well-adapted genotypes. So, the considerable phenotypic variability of P. australis is due to a high adaptability and an important genetic diversity.
INTRODUCTION
Le roseau commun, Phragmites australis (CAV.) TIUN. ex. STEUD., g6neralement appele P. communis TRN. (CLAYrON 1968), est une espece hautement polymorphe. Il se developpe dans des conditions environnementales diverses oiu il montre une grande adaptabilite ecologique.
Morphologiquement, le P. australis varie en hauteur, en largeur des feuilles ainsi que pour les caracteres relatifs aux inflorescences et aux epillets (CLAYrON 1967, VAN DER TOORN 1971, HAsLAm 1972, HRADECKA 1973a,b, ATER 1985, KSENOFONTOVA 1988).
11 constitue un complexe polyploide important. De nombreux auteurs ont rapporte differents niveaux de ploidie, de triploide, 2n=36 a octoploide, 2n-96 (REEZE 1957, RAICU et al. 1972, GORENFLOT 1976, GORENFLOT et al. 1972, 1974, STOLAN et al. 1974). La forme pentaplofde, 2n=60 est inconnue et les individus diploides, 2n =24, recoltes par PAZOURKOVA (1973) en Tfebo'n sont interpretes comme un accident dans la descendance d'un polyploide
Folia Geobot. Phytotax., Praha, 27: 49-59
This content downloaded from 185.2.32.49 on Wed, 18 Jun 2014 11:15:59 AMAll use subject to JSTOR Terms and Conditions

50 FOLIA GEOBOTAINICA ET PHYTOTAXONOMICA, 27, 1992
Tab. 1. Localisation et caract6ristiques climatiques et ecoloqigues des populations etudiees. Les approches effectu6es sur chaque population sont indiqu6es par M = morphologique, C = caryologique, E= enzymatique.
Localit6 Code Lat. Long. Climat Site Approche
ALGERIE
Laghouat LAGH 33049'N 2r55'E aride lit d'oued M El Mesrane MESR 34057'N 3004'E aride dunes
continentales M C E Boughzoul BOUG 35042'N 251'E semi-aride lac sal6 M C E Ghrib GHRI 36015'N 2r18'E sub-humide barrage M C E Constantine CONS 36022'N 6040'E semi-aride marais M R6ghaia REGH 36051'N 3018'E sub-humide marais M C E Cap Djinet ISSE 36053'N 3043'E sub-humide embouchure
d'oued M C E Akbou SOUM 36029'N 4032'E sub-humide lit d'oued M
FRANCE
Rochefort ROCH 45047'N 0058'W atlantique marais C E Suscinio SUSC 47031'N 2r44'W atlantique marais C E Ile-Grande ILGR 48048'N 3035'W atlantique bas de falaise
maritime C E
et non comme des formes originelles. Cependant, selon DYKYJOVA et PAZOURKOVA (1979) la diploidie pourrait etre une reponse cytogenetique a la transplantation d'un milieu saumatre a un milieu d'eau douce.
En plus des series euploides, une serie aneuplofde est centree autour de 2n = 48 (PAZOURKOVA 1973, GORENFLOT et al. 1972, LENOIR et al. 1975).
Les approches biochimiques menees par BAHRMAN et GORENFLOT (1983) et GORENFLOT et al. (1982, 1984) ne montrent aucune relation entre la composition flavonique et le nombre chromosomique, alors que la distribution des proteinogrammes revele deux groupes de populations a chaque niveau de ploidie, 2n =48 et 2n = 96.
A notre connaissance aucune donnee n'est actuellement disponible concernant la variabilite enzymatique chez le P. australis. De plus, en Algerie, peu d'etudes cytogenetiques ont etd menees sur cette espece oii seulement une population tetraploide et deux populations octoploYdes ont ete decrites (REEZE 1957, GORENFLOT et al. 1974, 1984).
Ces dernieres remarques ont induit cette etude prospective sur la variabilite morphologique, caryologique et enzymatique de quelques populations de P. australis recoltees en Algerie du nord et dans l'ouest de la France. Les denombrements chromosomiques, les variations des caracteres morphologiques et les isomorphes de plusieurs systemes enzymatiques sont analyses dans le but d'apprehender les differences phenotypiques et genetiques entre des populations isolees geographiquement.
MATERIEL ET METHODES
Les onze populations etudiees se developpent dans des conditions climatiques et ecologiques differentes dans le nord de l'Algerie et dans l'ouest de la France. Les trois
This content downloaded from 185.2.32.49 on Wed, 18 Jun 2014 11:15:59 AMAll use subject to JSTOR Terms and Conditions

DJEBRoUNI: VARIABILITY IN PHPAGMITES A USTRALIS 51
Tab. 2. Caracteres biometriques retenus pour 1'etude morphologique
A Hauteur totale des pousses B Hauteur de la panicule C Longueur du rachis D Distance separant le ler et le 6eme noeud en partant du niveau du sol E Nombre de nocuds du chaume F Diametre A la base du rachis G Diam6tre A la base du chaume
approches, morphologique, caryologique et enzymatique ont ete effectuees sur cinq populations. Les autres populations ont ete traitees soit sur le plan morphologique, soit sur les plans caryologique et enzymatique (Tabl. 1). Dans chaque population dix pousses fertiles et trois sections de rhizomes sont recoltees.
Les pousses fertiles font l'objet de comptages et de mesures biometriques destinees aux analyses multidimensionnelles. Les caracteres retenus pour l'analyse de la variabilite morphologique sont consignes dans le tableau 2.
Les sections de rhizome sont cultives sous serre, pendant cinq mois, dans des conditions identiques. Cette periode permet d'6liminer un eventuel choc dfu a la transplantation et la formation de nouvelles emergences.
Les denombrements chromosomiques sont r6alises sur l'extremite des racines pretraitees pendant 16 heures a 4 ?C dans une solution saturee d'ca -bromonaphtalene. L'hydrolyse, effectuee dans une solution d' HCl 1N a 60 ?C pendant 12 a 15 minutes, est suivie d'une coloration des chromosomes au reactif de Schiff. L'etalement des chromosomes est ameliore par l'addition d'une solution de pectinase a 5% avant l'ecrasement des racines dans de l'acide acetique a 45%.
Pour minimiser les variations intrapopulationnelles dans l'6tude de la variabilite enzymatique, trois emergences sont prelevees dans les trois rhizomes de chaque population. Elles sont melangees puis broyees, a 4 OC, au polytron dans un tampon 0.125 M Tris-HCl pH 6.8 contenant 2% de saccharose. Le tampon est utilise dans la proportion 3/1 (V/Poids). Du PVP (polyvinylpolypyrrrolidone) est ajoute au broyat a raison de 50% du poids de materiel utilise. Le surnageant contenant les proteines solubles est preleve apres centrifugation a 12.000 g pendant 30 minutes a 4 OC.
La separation des isozymes est realis6e par electrophorese verticale sur gel de polyacrylamide (30:1.5) en utilisant un svsteme discontinu. Le gel de concentration 'a 5% d'acrylamide et 0.125 M Tris-HCl pH 6.8 est le meme pour tous les systemes enzymatiques etudies. La concentration en acrylamide du gel de s6paration a 0.375 M Tris-HCl pH 8.8 est de 7.5% pour les amylases (E.C.3.2.1.1.) et les glutamate oxaloacetate transaminases ou GOT (E.C.2.6.1.1.), et de 10% pour les malate deshydrogenases ou MDH (E.C.1.1.1.37.), les est6rases non sp6cifiques (E.C.3.1...), les phosphatases acides ou ACP (E.C.3.1.3.2.) et les superoxyde dismutases ou SOD (E.C.1.15.1.1.). Apr&s 6lectrophorese les systemes enzymatiques sont reveles d'apres SOLTIs et al..(1983) et PASTEUR et al. (1987).
Pour chaque zymogramme rev616, la pr6sence ou l'absence de chaque bande est not6e 1 ou 0. Les donnees ainsi obtenues pour toutes les populations et l'ensemble des systemes enzymatiques sont soumises aux analyses multidimensionnelles.
Les donnees biometriques sont trait6es par l'analyse en composantes principales (A.C.P.) et les doniees enzymatiques par l'analyse factorielle des correspondances (A.F.C.) et la
This content downloaded from 185.2.32.49 on Wed, 18 Jun 2014 11:15:59 AMAll use subject to JSTOR Terms and Conditions

52 FOLIA GEOBOTANICA ET PHYTIOTAXONOMICA, 27, 1992
Tab. 3. Matrice des correlations entre les caracteres morphologiques
A B C D E F G
A 1.000 B 0393 1.000 C -0.085 0.387 1.000 D 0.833 0.267 -0.093 1.000 E 0.781 0329 -0.149 0.491 1.000 F 0.206 0.155 0.026 0.186 -0.183 1.000 G 0.037 0.103 -0.005 0.156 -0.035 0.141 1.000
Tab. 4. Nombres chromosomiques des populations 6tudi6s
El Mesrane 2n=96 Boughzoul 2n=48 Ghrib 2n=48 Reghaia 2n=48 Cap Djinet 2n = 48 Rochefort 2n = 48 Suscinio 2n=48 Ile-Grande 2n=48
classification ascendante hi6rarchique (C.A.H.). Pour cette derniere nous avons calcule la distance du khi-deu.x et la minimisation du moment centre deux des classes comme critere d'agregation.
Les traitements informatiques ont ete realises au C.I.C.B. (Centre interuniversitaire de calcul de Bretagne) en utilisant les programmes ANCOMP, ANCORR et CAHTRA de la collection A.D.D.A.D. (association pour la diffusion et le d&veloppement de I'analyse des donnees).
RESULTATS ET DISCUSSIONS
VARIABIL1TE MORPHOLOGIQUE
La matrice des correlations 6tablie a partir des donnees biom6triques (Tabl. 3) revele que les descripteurs de la vigueur des pousses sont fortement lies. Ainsi la hauteur (A), ia distance entre le premier et le sixieme noeud (D) et le nombre de noeuds du chaume (E) montrent des corr6lations elevees. De meme, mais a un moindre degre, la hauteur de la panicule (B) varie dans le mime sens que la hauteur totale (A), la longueur du rachis (C) et le nombre de noeuds du chaume (E).
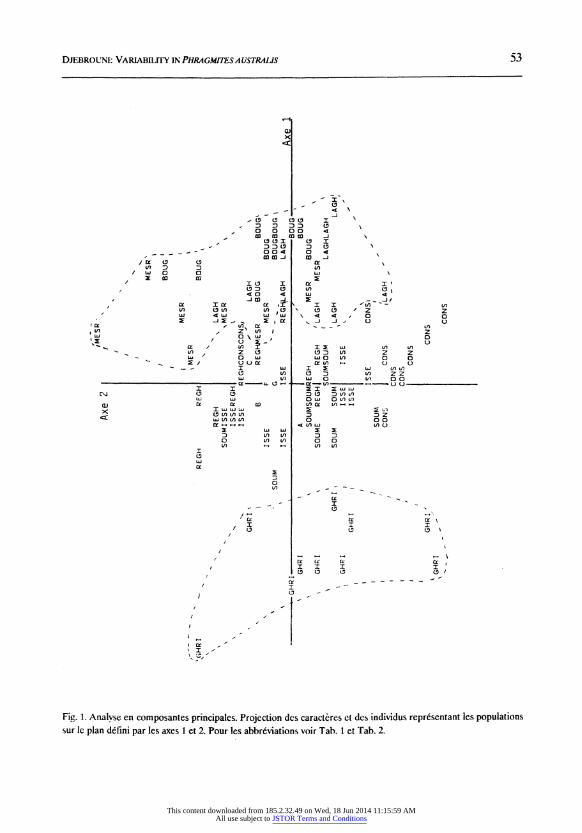
La projection des caracteres et des individus dans le systeme d'axes 1-2 de l'A.C.P. (Fig. 1) permet d'isoler les populations selon un gradient climatique et 6cologique. Les populations provenant d'un climat aride ou semi-aride, ou se developpant dans des milieux sales
This content downloaded from 185.2.32.49 on Wed, 18 Jun 2014 11:15:59 AMAll use subject to JSTOR Terms and Conditions
DJEBROUNI: VARIABILrrY IN PHRAGMITESAUSTR4ALS 53
x
_ - C )
-0 C)X 0 I - D 'n: 0
' 0 0 D 4
/~~~~~~~~~~C co Ca co z
} 0 01 0 I
8 ~ ~~~~~~~ ~ ~~~ I O I ) tI
- ~~~~~00 4 0 4 a to CO -
/CC 0 0 CC / L~~~~J 0 0 LI~ 0 c
a I ) I ,- C) 0 LI)0
/ l,s,@ < S ~ .~ W .4
U j .4,< / x I ' a I I )V U)
'>: ~ ~ U ,D') o ~ 0 0 ,1 Z
_ i~ ~ ~ ~~~tn Zn LA Zo z /
WL 4 W Wi 1? 0 0
_F ~ ~ .~ zLiL .- O4 U C) 1 _ I / t ) , x I a )
_ c _ _) III U) U)u i U), Z 0 n 0 U) Z Z
0 u 1 W O U) C O
I I W I I LflU iL U) 0 O n u l
as 0K tr t D 0 ' 0) 0
I I II ;IW :CL
IWWLu I~~~~~C L u L
) n u ) D Z o U) U) 0 u7 O O
(X J. 0 - - - (n - - L.) o
C\J O D 1 L5 un D OL LJ 0n w O )E
I) _ tn n LL ~ ~ ~ n L
r~~~~ 0
_ - - I
-- C) II
/ I
S :2: :I I I C) C) C) C)
/ CL.
CL I ' I
Fig. 1. Analyse en composantes principales. Projection des caracteres ct des individus representant les populations
sur le plan defini par les axes I et 2. Pour les abbreviations voir Tab. 1 et Tab. 2.
This content downloaded from 185.2.32.49 on Wed, 18 Jun 2014 11:15:59 AMAll use subject to JSTOR Terms and Conditions

54 FOLIA GEOBOTANICA ET PHYrOTAXONOMICA, 27, 1992
gS~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~: C1 tV -r - (D. cc t O L/ Ln tz ) cyQ / ) CD tU0t
en O cc L O D J O U~~~~~~ Li V) ~~~~~~~~~~~~~~~~~~~ ~ ~ ~ ~ ~ ~ i (3 ~~~~~~~~~~~~x L
LI 0r _ : o cr v ) =m~c nc
ESTIERASES S.O.D.
AMYLASES G.O.T.
A. .P. M.D. H
Fi_ .Zyorame,de ixsstme nzmtiue tuis - - _l- - -
- - - - -_
---~~~~~~~ - - _ -
Fig.~~~~~~~~ ~ ~~~ 2. Z_igamsdssxss~e nyatqe td u utppltos
This content downloaded from 185.2.32.49 on Wed, 18 Jun 2014 11:15:59 AMAll use subject to JSTOR Terms and Conditions

DJEBROUNI: VARIABILrrY IN PHRAGMITES AUSTRALIS 55
Axe 2
, ILGR
- P REGH' .. t - -. _ _ R BOUG
ROCH G,-HRI'. Axe 1 \ x 5UR Isuc 1SSEI
MESR/
Fig. 3. Analyse factorielle des correspondances. Projection des populations en fonction des doonns enzymatiques sur le systdme d'axes (1-2).
(Laghouat, Mesrane et Boughzoul) sont projet6es dans la partie positive de l'axe 1; elles s'opposent ~ celles de Ghrib et de Soummam qui sont r6colt6s dans un climat sub-humide. Les autres populations occupent une position interm6diaire dans le diagramme 1-22 de l'A.C.P., cette position se retrouve aussi sur les plans g6ographique, climatique et 6cologique.
Ces r6sultats sont en accord avec ceux de&j"a obtenus par VAN DER TooRN (1971), HRADECKA, (1973a,b) et KSENOFONTOVA (1988) qui ont montr6 que les caract&res de vigueur sont li6s aux conditions climatiques et trophiques. Par ailleurs, les caract6res v6g6tatifs, ceux des inflorescences et des e6pillets pr6sentent de fortes variations d'une population a une autre et d'une ann6e sur l'autre pour une m8me population. Ceci sugg6re une forte variabilit6 ph6notypique de 1'ensemble des caract~res morphologiques. Cependant, les &tudes men6es r6v6lent une diff6renciation des populations sur le plan morphologique; HAsLANM (1973) a montr6 que ces caract&res varient peu apr&s transplantation ou apres culture 'a partir de graines, ce qui leur conf&re une base g6n6tique.
This content downloaded from 185.2.32.49 on Wed, 18 Jun 2014 11:15:59 AMAll use subject to JSTOR Terms and Conditions

56 FOLIA GEOBOTA.NICA ET PHYTOTAXONOMICA, 27, 1992
VARIABILITE CARYOLOGIQUE
Sur les huit populations etudiees, sept sont tetraploides, avec 2n=48 et une seule est octoploide, avec 2n = 96 (Tabl. 4). La forme octoploide est originaire de Mesrane, situee dans les hautes plaines steppiques en Alg6rie. Les populations tetraploYdes se developpent dans des zones geographiques et des conditions environnementales extremement variables, du lac sale de Boughzoul et du barrage de Ghrib en Algerie aux marais d'eau douce de Rochefort et de Suscinio en France.
11 apparalit que 2n =48 est le nombre chromosomique le plus courant dans les populations etudiees et qu'il n'y aurait aucune relation entre les conditions 6cologiques, la distribution geographique et le degre de ploidie.
Ces resultats confirment ceux deja obtenus par REEZE (1957) et par GORENFLOT et al. (1972, 1974) pour des populations de France et du reste de l'Europe.
D'apres RAIcU et al. (1972) et STOTAN et al. (1974) le nombre chromosomique de base chez le P. australis est x = 12. Des analyses comparatives de caryotypes et de meioses polliniques ont montre que 2n = 8x =96 est probablement un autoallooctoploide, alors que l'origine des tetraploides, 2n = 4x =48, est incertaine; ils pourraient resulter d'une allotetraploidie.
Par ailleurs, LENOIR et al. (1975) etudiant les meioses chez les tetraploides et les octoploides concluent que x = 6 serait le nombre chromosomique de base, avec une polyploidisation deja ancienne suivie d'une diploidisation poussee; le mode de polyplofdisation restant A elucider.
POLYMORPHISME ENZYMATIQUE
La genetique formelle du P. australis n'est pas encore connue, aussi le nombre de locus codant pour chaque systeme enzymatique ne peut etre affirme. Cependant, les zymogrammes obtenus pour les six systemes enzymatiques analyses presentent une forte variabilite entre les huit populations etudiees (Fig. 2).
Les esterases montrent dix bandes dont aucune n'est commune a toutes les populations. Ces bandes se repartissent en deux groupes distincts qui peuvent etre assimiles a deux locus, le locus rapide est.-1 et le locus lent est.-2. Les populations de Rochefort et de Suscinio sont les seules A presenter les memes profils alors que celles de Mesrane, Boughzoul et Ghrib ont chacune un electromorphe qui les differencie des sept autres populations.
Sur les sept bandes des amylases, une seule est commune a toutes les populations; elle correspond a l'lectromorphe le plus lent du locus amyl.-1. Le locus amyl.-2 presente une bande qui n'est absente que dans la population de Rochefort et une autre dans la population d'Isser.
Les phosphatases acides (ACP) presentent huit bandes dont deux sont communes A toutes les populations. Les echantillons de Boughzoul, Isser et Suscinio ont des profils identiques pour cette enzyme.
Les superoxyde dismutases (SOD) montrent un grand nombre de bandes qui pourraient correspondre A trois locus, SOD.-1 avec trois bandes, SOD.-2 avec deux bandes et SOD.-3 avec trois bandes. Sur les huit electromorphes, cinq sont communs a toutes les populations.
Les glutamate oxaloacetate transaminases (GOT) revelent deux groupes de bandes A mobilite electrophoretique differente. Le locus rapide GOT.-1 a deux electromorphes et le locus lent GOT.-2 en presente quatre. Seules deux bandes sont communes a toutes les populations.
This content downloaded from 185.2.32.49 on Wed, 18 Jun 2014 11:15:59 AMAll use subject to JSTOR Terms and Conditions

DJEBROuNI: VARIABILITY IN PHRRAGMITES AUSTRALIS 57
1 0.8 0.6 04 0.2 0 GHRI I i I I - I t
REGH
susc ROCH
+ ILGR
MESR
ISSE
BOUG
Fig. 4. Classification ascendante hidrarchique des populations a partir des donnees enzymatiques.
Les malate deshydrogenases presentent cinq bandes. Les echantillons de Boughzoul, Isser, Ghrib et Rochefort ont les memes profils pour cette ernzyme.
La projection des populations en fonction des variables (electromorphes) sur le plan defini par les axes 1 et 2 de 1'A.F.C. (Fig. 3) permet de repartir les populations en deux ensembles. Les populations originaires d'un habitat dulsaquicole (Rochefort, Suscinio, Ghrib et Reghaia) s'opposent a celles recoltees dans un milieu saumatre (Isser, Boughzoul, Ile-Grande) ou aride (Mesrane).
La C.AH. des populations (Fig. 4) confirme ces resultats et les precise. Ainsi, pour le meme habitat d'eau douce les populations d'Algerie (Reghaia, Ghrib) s'isolent nettement des populations de France (Suscinio, Rochefort).
CONCLUSIONS
Les populations de P. australis etudiees montrent une grande variabilite interpopulationnelle, ce qui permet de les distinguer les unes des autres notamment sur les plans morphologiques et enzymatiques. Cette forte variabilite phenotypique est due 'a un grand pouvoir d'accommodation aux milieux les plus varies.
La variabilite des caracteres morphologiques dans chaque population reviendrait principalement a la plasticite phenotypique des individus. En effet, la nature clonale du P.australis implique que le nombre de pousses par population ne reflete pas le nombre effectif d'individus, chaque population serait dominee par peu de genotypes.
La complexite des zymogrammes obtenus suggere que le P. australis est une espece hautement heterozygote. L'isolement geographique et ecologique des populations se traduit par une selection des genotypes les mieux adaptes aux conditions environnementales locales.
La diversite genetique du P. australis est accentuee par la possibilite de reproduction sexuee malgre une importante letalite du pollen due a une forte proportion de meioses irregulieres et a des phenomenes d'autoincompatibilite (GUSTAFFSON et SIMAK 1963, CURRAN 1969, LENOIR et al. 1975).
This content downloaded from 185.2.32.49 on Wed, 18 Jun 2014 11:15:59 AMAll use subject to JSTOR Terms and Conditions

58 FOLIA GEOBOTANICA Er PHYTOTAXONOMICA, 27, 1992
Ainsi, la forte variabilite sur les plans morphologique et caryologique est confirmee par une importante diversit6 gen6tique permettant au P. australis de s'adapter aux variations spatiales et temporelles de l'environnement.
RESUME
Onze populations de Phragmnites australis (CAV.) TRIN. ex STEUD. provenant de zones geographiques, de climats et de conditions ecologiques diff6rentes sont analysees sur les plans morphologique, caryologique et enzymatiques.
Les deux niveaux de ploidie determines, 2n = 48 et 2n = 96, ne sont pas en relation directe avec I'habitat. Les descripteurs de la vigueur des pousses sont correles positivement et seraient lies aux conditions
trophiques. L'analyse en composantes principales effectuee a partir des donn6s biometriques isole les populations selon un gradient ecologique et climatique.
Les zymogrammes obtenus pour six systemes enzymatiques revelent une grande variabilite interpopulationnelle. Leur etude par l'analyse factorielle des correspondances et par la classification ascen-dante hierarchique montre une differenciation ecologique et gdographique des populations. Ces resultats suggerent que la grande adaptabilit6 du P. australis est due a une forte h6t6rozygotie et a une importante diversite genetique.
SUMMARY
Eleven populations of Phragmites australis (CAV.) TRIN. ex STEUD. originating from different geographic areas, climatic and ecological conditions were studied through morphological, caryological and enzymatic approaches.
The two ploidy levels found, 2n = 48 and 2n = 96, were not directly related to the habitat. Characters describing the vigour of the shoots were highly correlated; they seems to be controlled by trophic
conditions. Principal conponent analysis done with biometric data separate populations along ecological and climatic
gradient. Banding pattern of the zymograms obtained from the six enzymatic systems revealed a high interpopulation variability. Their analysis using factorial correspondences analysis and hierarchical ascendent classification showed ecological and geographic differentiation of the populations. These results suggest that the considerable adaptability of P. communis is due to a high heterozygosity and an important genetic diversity.
LITERATURE CITED
ATER M. (1985): La variation pollinique du Phragrnites australis (CAV.) TRIN. ex STEUD. - Bull. Soc. Bot. Fr., 21: 111-117.
BAHRMAN N. et R. GORENFLOT (1983): Apports des prot6ines foliaires solubles dans l'interpretation du complexe polyploide du Phragmites australis (CAV.) TRIN. ex STEUD. - Rev. Gen. Bot., 90: 177-184.
CLAYTON W.D. (1967): Studies in the Gramineae: XIV - Kew. Bull., 21: 111-117. CLAYTON W.D. (1968): The correct name of the common reed. - Taxon, 17: 168-169. CLRRAN P.L. (1969): Fertility of Phragmites communis TRIN. - Irish Naturalist's J., 16: 242. DYKYJOVA D. et PAzouRXovA Z. (1979): A diploid form of Phragnites communis, as a possible result of
cytogenetical response to ecological stress. - Folia Geobot. Phytotax., 14: 113-120. GORENFLOT R. (1976): Le complexe polyploide du Phragmites australis (CAV.) TRIN. ex STEUD. (= P. comnzunis
TRIN.). - Bull. Soc. Bot. Fr., 123: 261-271. GORENFLOT R., CARTIER D. et LENOIR A. (1974): La polypldidie du P. australis (CAV.) TRIN. ex SEUD. dans le
bassin mediterraneen. In: "La flore du bassin mediterraneen. Essai de syst6matique synthetique" (colloque du
This content downloaded from 185.2.32.49 on Wed, 18 Jun 2014 11:15:59 AMAll use subject to JSTOR Terms and Conditions

DJEBRouNi: VARiABiLrTY IN PHRAGAMTESAUJSTRALIS 59
CNRS N0 235): 165-173. GORENFLOT R., HUBAC J.M., et JAY M. (1984): Le complexe polyploide du Phragmites australis (CAV.) TRIN. ex
STEUD. dans la region medit6rranmenne. - Webbia, 38: 715-721. GORENFLOT R., HuBAc J.M., JAY M. et LALANDE P. (1982): Geographic distribution, polyploidy and patterns of
flavonoids in Phragmites australis (CAV.) TRIN. ex STEUD. - Proceedings of NATO Advanced Study Institute. Ser. G. (Ecological Sciences), 1 474-478.
GORENFLOT R., RAICU P., CARTIER D., CIOBANU I., STOIAN V. et STAICU S. (1972): Le complexe polyploide du
Phragmites communis TRIN. - C.R. Acad. Sci., Paris, 274: 1501-1504. GORENFLOT R. et SA-NEI-CHARIAT PANAHI M.(1979): Le complexe polyploide du Phragmites australis (CAV.)
TRIN. ex STEUD. (= P.communis TRIN.) en Iran. - Rev. Cyt. Biol. Veget., 2:67-81. GUSTAFSSON A. et SIMAK M. (1963): X-Ray photography and seed sterility in Phragmites communis TRIN. -
Hereditas, 49: 442-450. HASLAM S.M. (1972): Biological flora of the British isles. Phrag,mites communis TRIN. - J. Ecol., 60: 585-610. HASLAM S.M. (1973): Some aspects of the life history and autoecology of Phragmites communis TRIN. A review. -
Pol. Arch. Hydrobiol., 20: 79-100. HRADECKA D. (1973a): Common reed (Phragmites communis TRIN.) in south Bohemia, south Moravia and south
Slovakia. Morphology of the inflorescences and flower wraps. - In: "Ecosystem study on Wetland Biome in Czechoslovakia. Czechosl. IBP/PT-PP Report No. 3, Tfeboni: 47-53.
HRADECKA D. (1973b): Morphological characteristics of two types of common reed (Phraginites communis TRIN.) from the Opatovick4 fishpond. - In: "Ecosystem study on Wetland Biome in Czechoslovakia". Czechosl. IBP/PT-PP Report No.3, Tfebof: 55-58.
KSENOFONTOVA T. (1988): Morphology, production and mineral contents in Phragmites australis in different waterbodies of the Estonian SSR. - Folia Geobot. Phytotax., 23: 17-44.
LENOIR A., STOLAN V., CARTIER D., GORENFLOT R. et RAICU P. (1975): Polyploidie et m6foses polliniques du Phragmites australis (CAV.) TRIN. ex STEUD. - C. R. Acad. Sci. Paris, 280: 621-624.
PASEDUR N., PASTEUR G., BONHOMME F., CATALAN J. et BR1rION-DAvIDIAN GJ. (1987): Manuel technique de genetique par electrophorese des prot6ines. - Lavoisier Tech. & Doc., Paris, 217 pp.
PAZOURKOVA Z. (1973): Caryology of some forms of Phragmites communis TRIN. - In: "Ecosystem study on Wetland Biome in Czechoslovakia". - Czechosl. IBP/PT-PP Report No. 3, Tfeboii: 59-62.
RAIcu P., STAIcu S., STOIAN V. et ROMAN T. (1972): The Phragmnites communis TRIN. chromosome complement in the Danube delta. - Hydrobiologia, 39: 83-89.
REEZE G. (1957): Uber die Polyploidiespektren in der nordsaharischen Wustenpflanzen. - Flora 144: 598-634. SOLTIs D.E., HAUFLER C.H., DARROW D.C. et GASTONY GJ. (1983): Starch gel electrophoresis of ferns: a
compilation of grinding buffers, gel and electrode buffers, and staining schedules. - Am. Fern J., 73: 9-27. STOIAN V., LENOIR A., RAIcu P. et GORENFLOT R. (1974): La meiose et la taille polliniques d'individus
octoploides du Phragmites australis (CAV.) TRIN. ex STEUD. - C. R. Acad. Sci., Paris. 278: 457-459. VAN DER TooRN J. (1971): Investigations on the ecological differentiation of Phragmites communis TRIN. in the
Netherlands.- Hydrobiologia, 12: 97-106.
ReNu 21 Aoat 1989, accepte 4 Juin 1990
This content downloaded from 185.2.32.49 on Wed, 18 Jun 2014 11:15:59 AMAll use subject to JSTOR Terms and Conditions





























