Embed Size (px)
Citation preview

Génétique de la susceptibilité au paludisme: approches épidémiologiques sur le terrain
Atelier paludisme 2004
David Modiano
Département de Sciences de Santé PubliqueUniversité de Rome “La Sapienza” Italie
World Health Organization Collaborating Centrefor Malaria Epidemiology and Control
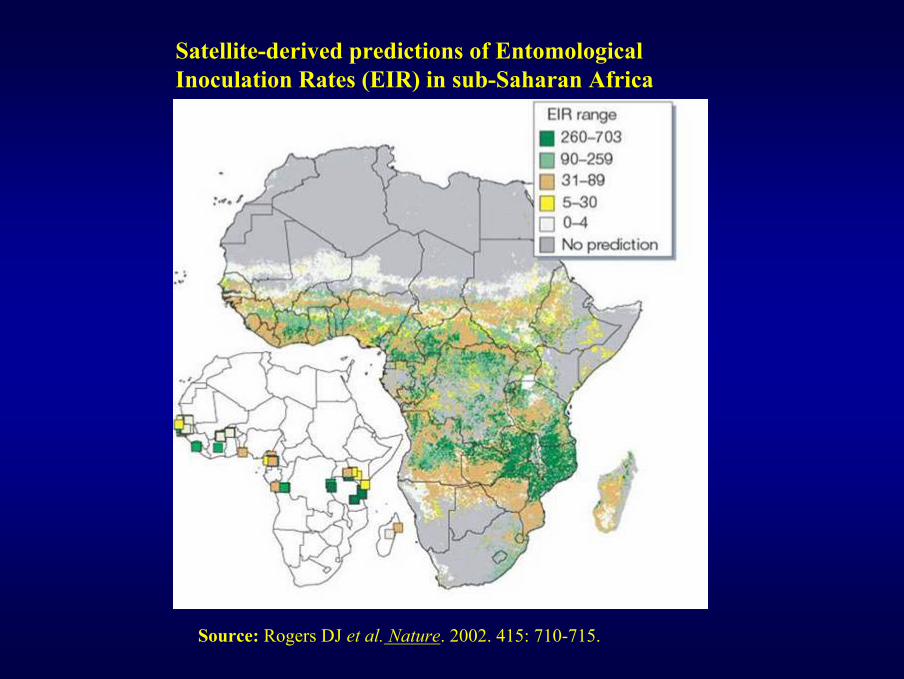
Source: Rogers DJ et al. Nature. 2002. 415: 710-715.
Satellite-derived predictions of Entomological Inoculation Rates (EIR) in sub-Saharan Africa
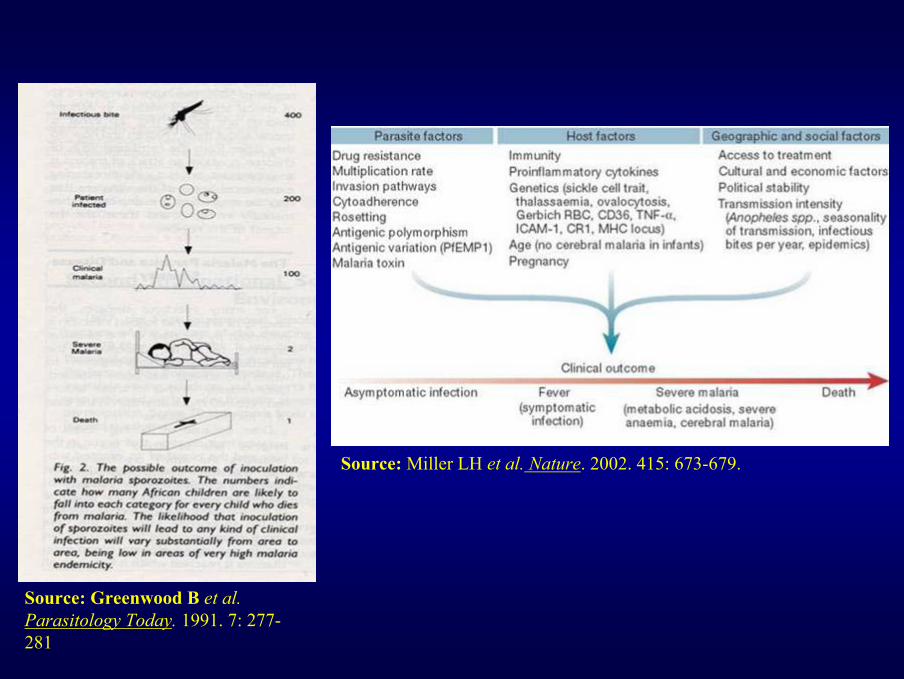
Source: Miller LH et al. Nature. 2002. 415: 673-679.
Source: Greenwood B et al.Parasitology Today. 1991. 7: 277-281

Genes studied for association with malaria IABO Lell 99 The role of red blood cell polymorphisms in resistance and susceptibility to malariaCD36 Aitman 00 Malaria susceptibility and CD36 mutationPain 01 A non-sense mutation in Cd36 gene is associated with protection from severe malaria
CR1 Rowe 97 P. falciparum rosetting mediated by a parasite-variant erythrocyte membrane protein and complement-receptor 1Bellamy 98 Absence of an association between intercellular adhesion molecule 1, complement receptor 1 and interleukin 1 receptor antagonist gene
polymorphisms and severe malaria in a West African populationFCGR2A Omi 02 Fcgamma receptor IIA and IIIB polymorphisms are associated with susceptibility to cerebral malariaFY Miller 76 The resistance factor to Plasmodium vivax in blacks. The Duffy-blood- group genotype, FyFy
Chitnis 94 Identification of the erythrocyte binding domains of Plasmodium vivax and Plasmodium knowlesi proteins involved in erythrocyte invasionTournamille 95 Disruption of a GATA motif in the Duffy gene promoter abolishes erythroid gene expression in Duffy-negative individuals
G6PD Gilles 67 Glucose-6-phosphate-dehydrogenase deficiency, sickling, and malaria in African children in South Western NigeriaRuwende 95 Natural selection of hemi- and heterozygotes for G6PD deficiency in Africa by resistance to severe malariaRuwende 98 Glucose-6-phosphate dehydrogenase deficiency and malaria
GYPC Patel 01 The association of the glycophorin C exon 3 deletion with ovalocytosis and malaria susceptibility in the Wosera, Papua New GuineaHBA1 Allen 93 A prospective study of the influence of alpha thalassaemia on morbidity from malaria and immune responses to defined Plasmodiumfalciparum antigens in Gambian children
Williams 96 High incidence of malaria in alpha-thalassaemic childrenAllen 97 alpha+-Thalassemia protects children against disease caused by other infections as well as malaria
HBB Neel 49 (Science: 110, 64-6) The inheritance of sickle cell anemiaPauling 49 (Science: 110, 543-8) Sickle cell anemia, a molecular diseaseGilles 67 Glucose-6-phosphate-dehydrogenase deficiency, sickling, and malaria in African children in South Western NigeriaHill 88 Beta thalassemia in Melanesia: association with malaria and characterization of a common variant (IVS-1 nt 5 G----C)Hill 91 Common west African HLA antigens are associated with protection from severe malariaAllen 92 Morbidity from malaria and immune responses to defined Plasmodium falciparum antigens in children with sickle cell trait in The GambiaStirnadel 99 Malaria infection and morbidity in infants in relation to genetic polymorphisms in TanzaniaAgarwal 00 HBC associated with protection from severe malaria in the dogon of mali, a westafrican population with a low prevalence of hemoglobin SModiano 01 Haemoglobin C protects against clinical Plasmodium falciparum malariaModiano 01 The lower susceptibility to Plasmodium falciparum malaria of Fulani of Burkina Faso (west Africa) is associated with low frequencies of
classic malaria-resistance genesHLA-B Hill 91 Common west African HLA antigens are associated with protection from severe malaria
Hill 92 (Parasitology Today: 8, 57) HLA and malaria: on the mechanisms of dominant protective associations [letter].Hill 92 Molecular analysis of the association of HLA-B53 and resistance to severe malariaBennett 93 Human leucocyte antigen (HLA) and malaria morbidity in a Gambian communityGilbert 98 Association of malaria parasite population structure, HLA, and immunological antagonismModiano 01 HLA class I in three West African ethnic groups: genetic distances from sub-Saharan and Caucasoid populations
SOURCE: http://www.gmap.net/topics/malaria.htm

Genes studied for association with malaria IIHLA-DRB1 Hill 91 Common west African HLA antigens are associated with protection from severe malariaHP Elagib 98 Association of the haptoglobin phenotype (1-1) with falciparum malaria in Sudan
Quaye 00 Haptoglobin 1-1 is associated with susceptibility to severe Plasmodium falciparum malariaHunt 01 Haptoglobin and malariaAucan 02 Haptoglobin genotypes are not associated with resistance to severe malaria in The Gambia
ICAM1 Fernandez-Reyes 97 A high frequency African coding polymorphism in the N-terminal domain of ICAM-1 predisposing to cerebral malaria in Kenya
Bellamy 98 Absence of an association between intercellular adhesion molecule 1, complement receptor 1 and interleukin 1 receptor antagonist gene polymorphisms and severe malaria in a West African population
Kun 99 Association of the ICAM-1Kilifi mutation with protection against severe malaria in Lambarene, GabonIFNGR1 Koch 02 IFNGR1 Gene Promoter Polymorphisms and Susceptibility to Cerebral MalariaIL12B Morahan 02 A promoter polymorphism in the gene encoding interleukin-12 p40 (IL12B) is associated with mortality from cerebral malaria and with reduced nitric oxide productionIL4 Luoni 01 Antimalarial antibody levels and IL4 polymorphism in the Fulani of West AfricaMBL2 Bellamy 98 Mannose binding protein deficiency is not associated with malaria, hepatitis B carriage nor tuberculosis in Africans
Luty 98 Mannose-binding lectin plasma levels and gene polymorphisms in Plasmodium falciparum malariaNOS2A Burgner 98 Inducible nitric oxide synthase polymorphism and fatal cerebral malaria
Kun 98 Polymorphism in promoter region of inducible nitric oxide synthase gene and protection against malariaLevesque 99 Nitric oxide synthase type 2 promoter polymorphisms, nitric oxide production, and disease severity in Tanzanian children with malariaKun 01 Nitric oxide synthase 2(Lambarene) (G-954C), increased nitric oxide production, and protection against malariaHobbs 02 A new NOS2 promoter polymorphism associated with increased nitric oxide production and protection from severe malaria in Tanzanian and
Kenyan childrenSLC4A1 Foo 92 Ovalocytosis protects against severe malaria parasitemia in the Malayan aborigines
Genton 95 Ovalocytosis and cerebral malariaAllen 99 Prevention of cerebral malaria in children in Papua New Guinea by southeast Asian ovalocytosis band 3
TNF McGuire 94 Variation in the TNF-alpha promoter region associated with susceptibility to cerebral malariaKnight 99 A polymorphism that affects OCT-1 binding to the TNF promoter region is associated with severe malariaMcGuire 99 Severe malarial anemia and cerebral malaria are associated with different tumor necrosis factor promoter allelesStirnadel 99 Malaria infection and morbidity in infants in relation to genetic polymorphisms in TanzaniaWattavidanage 99 TNFalpha*2 marks high risk of severe disease during Plasmodium falciparum malaria and other infections in Sri LankansMeyer 02 TNFalpha-308A associated with shorter intervals of Plasmodium falciparum reinfectionsKnight 03 In vivo characterization of regulatory polymorphisms by allele-specific quantification of RNA polymerase loading
SOURCE: http://www.gmap.net/topics/malaria.htm
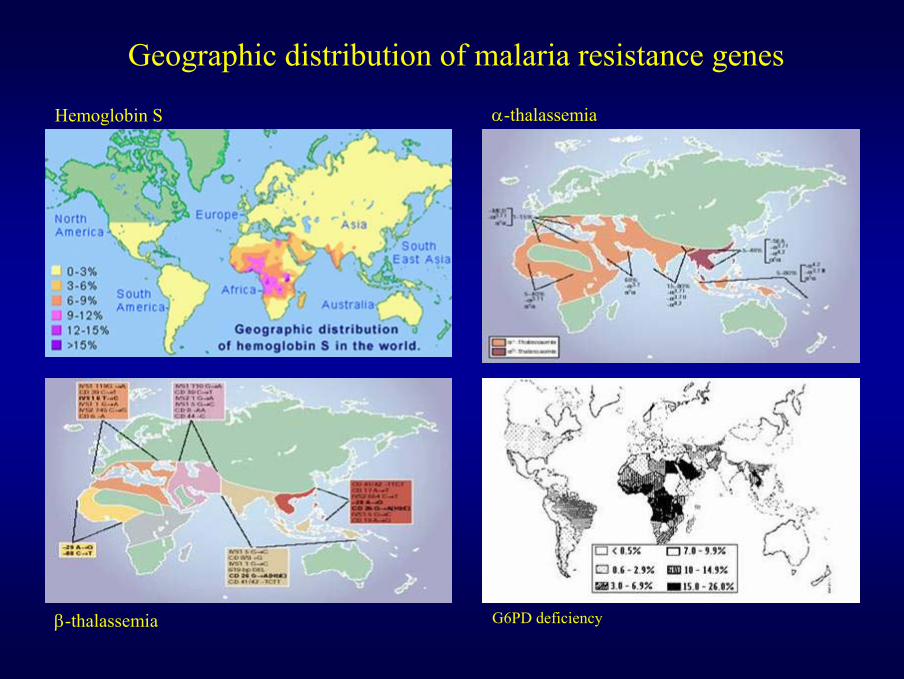
α-thalassemia
β-thalassemia G6PD deficiency
Hemoglobin S
Geographic distribution of malaria resistance genes
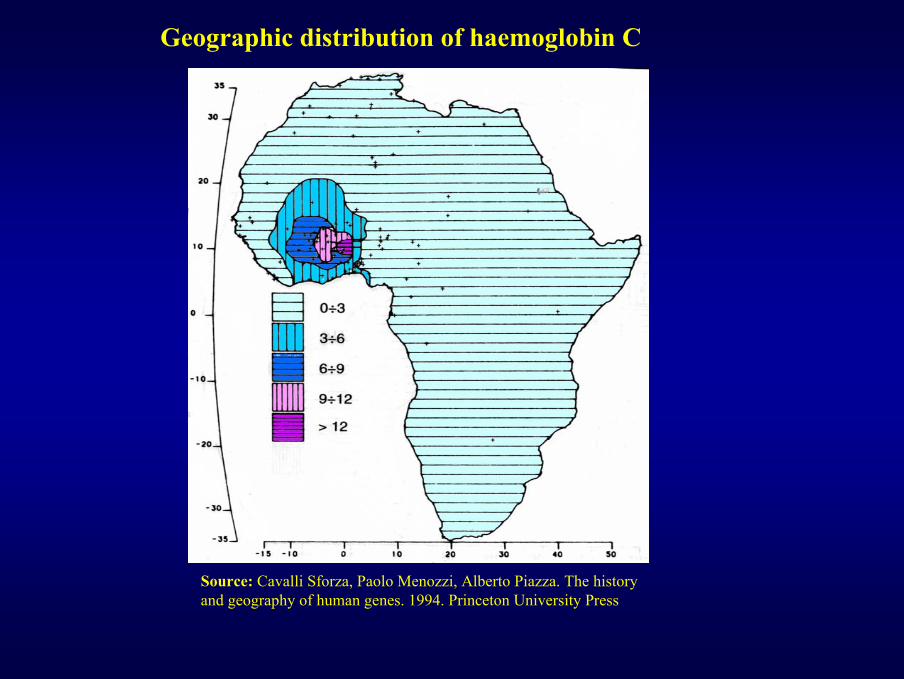
Geographic distribution of haemoglobin C
Source: Cavalli Sforza, Paolo Menozzi, Alberto Piazza. The history and geography of human genes. 1994. Princeton University Press

Host Factors – GeneticsTwo complementary populationalapproaches/models:
- intra-ethnic
- inter-ethnic

Intra-ethnic approach I
Comparison of genotype and allele frequencies of candidate resistance/susceptibility genes
between the general healthy population and hospitalized malaria patients (severe malaria, non complicated malaria)
vs

Intra-ethnic approach IIComparison of genotype and allele frequencies of candidate resistance/susceptibility genes among groups belonging to the same genetic background but living in endemic and non endemic areas
The inverse relationship between altitude, Thal heterozygotes (left) and GD-allele (right) distribution in Sardinia, Italy. Shaded areas > 500 m a.s.l.
Source: Modiano G. et al, 1986. Atti Accademia Nazionale dei Lincei, Serie VIII – Volume 18, Sezione III, Fascicolo 4

Intra-ethnic approach IIIFamily studies: comparison of genotype and allele frequencies of candidate resistance/susceptibility genes among families showing different capacity to control parasite density
Abel 92 Segregation analysis detects a major gene controlling blood infection levels in human malariaGarcia 98 Genetic control of blood infection levels in human malaria: evidence for a complex genetic modelRihet 98 Human malaria: segregation analysis of blood infection levels in a suburban area and a rural area in Burkina FasoStirnadel 99 Heritability and segregation analysis of immune responses to specific malaria antigens in Papua New Guinea
Chromosomal regions linked to malaria
Human chromosome 5Garcia 98 Linkage analysis of blood Plasmodium falciparum levels: interest of the 5q31-q33 chromosome regionRihet 98 Malaria in humans: Plasmodium falciparum blood infection levels are linked to chromosome 5q31-q33
Human chromosome 6Jepson 97 Genetic linkage of mild malaria to the major histocompatibility complex in Gambian children: study of affected sibling pairsJepson 97 Quantification of the relative contribution of major histocompatibility complex (MHC) and non-MHC genes to human immune responses to foreign antigensFlori 03 Linkage of mild malaria to the major histocompatibility complex in families living in Burkina Faso

Inter-ethnic approach
Comparison of genotype and allele frequencies of candidate resistance/susceptibility genes
among ethnic groups with different genetic background and different susceptibility to malaria living in the same epidemiological context, i.e. exposed to the same transmission levels and to the same parasite strains
vs
Fulani Mossi-Rimaibé

CONDITIONS NEEDED FOR INTER-ETHNIC APPROACH
Fulani
vs
Mossi -Rimaibé
ETHNIC GROUPS must show:
- similar exposure to malaria (higher the level, lower the risk of confounders)
- similar access to anti-malaria drugs
- similar use of control methods (ex. ITN)
- different genetic background
- different susceptibility (parasite rates, parasite densities, incidence and of clinical malaria)

Intra-ethnic approach

Sample N Relative and (absolute) genotype frequencies Relative and (absolute) allele frequencies AA AC AS CC SC SS βA βC βS Healthy subjects 3513 0.6641
(2333) 0.2172 (763)
0.0954 (335)
0.0165 (58)
0.0065 (23)
0.0003 (1)
0.8204 (5764)
0.1284 (902)
0.0512 (360)
Severe malaria 359 0.8078 (290)
0.1755 (63)
0.0111 (4)
0.0028 (1)
0.0028 (1)
0 (0)
0.9011 (647)
0.0919 (66)
0.0070 (5)
Non-complicated malaria 476 0.8004 (381)
0.1555 (74)
0.0399 (19)
0 (0)
0.0042 (2)
0 (0)
0.8981 (855)
0.0798 (76)
0.0221 (21)
Malaria patients (total) 835 0.8036 (671)
0.1641 (137)
0.0275 (23)
0.0012 (1)
0.0036 (3)
0 (0)
0.8994 (1502)
0.0850 (142)
0.0156 (26)
Comparisons Odds ratio (95% confidence interval) and P values * Healthy subjects vs. Malaria patients 2.07 Ω
(1.71-2.50)<<0.001
0.71 ψ (0.58-0.87)
0.0008
0.27 φ (0.17-0.42)
<<0.001
0.07 δ (0.00-0.48)
0.0011
n.s.
n.s.
<<0.001
<<0.001
<<0.001
Severe malaria vs. non-severe malaria n.s. n.s 0.27 ϕ (0.08-0.86)
0.021
n.s. n.s. n.s. n.s. n.s. 0.023
vs
INTRA-ETHNIC APPROACH
β globin genotype and allele frequencies in healthy subjects and in malaria patients from Burkina Faso, West Africa (Source: Modiano D, et al., Nature, 2001: 414: 305-308)
Healthy subjects Severe malaria patients
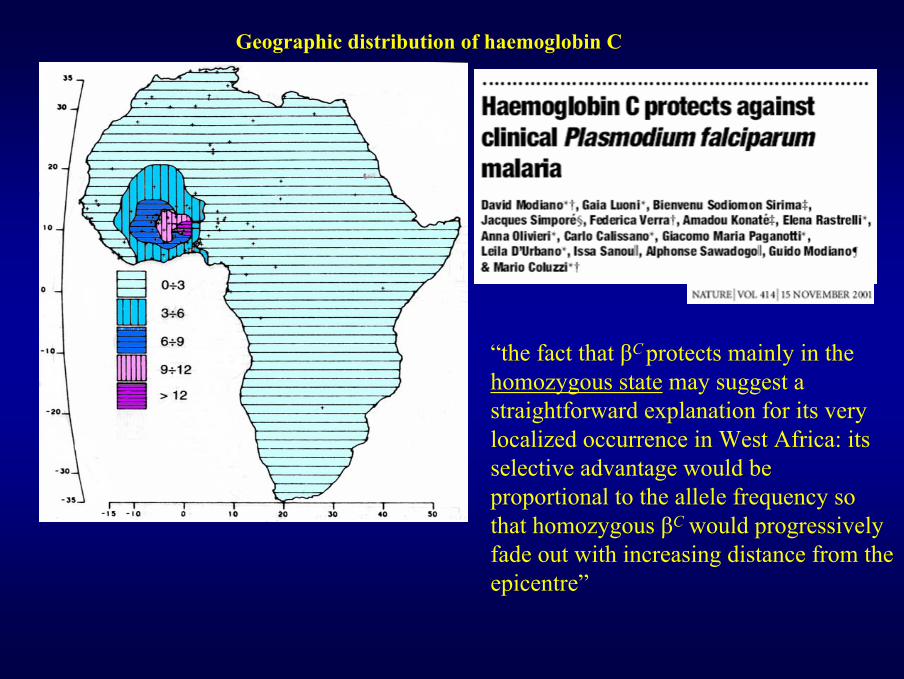
Geographic distribution of haemoglobin C
“the fact that βC protects mainly in the homozygous state may suggest a straightforward explanation for its very localized occurrence in West Africa: its selective advantage would be proportional to the allele frequency so that homozygous βC would progressively fade out with increasing distance from the epicentre”

Inter-ethnic approach

M MM
R
F
FF
F
F
FF
FF
FF
F
R
RR
R
RRR
MM
M MM M M
MR
RR R
R
F
Barkoundouba
Barkoumbilen
MALI NIGER
CÔTE
GHANABENIN
100
Km
D'IVOIRE
BURKINAFASOOuagadougou
BoboDioulasso
Study area
Km
0 0.5 1
F = FulaniM = MossiR = Rimaibé
Kaya
Ouagadougou
INTER-ETHNIC APPROACH
Fulani
vs
Mossi -Rimaibé
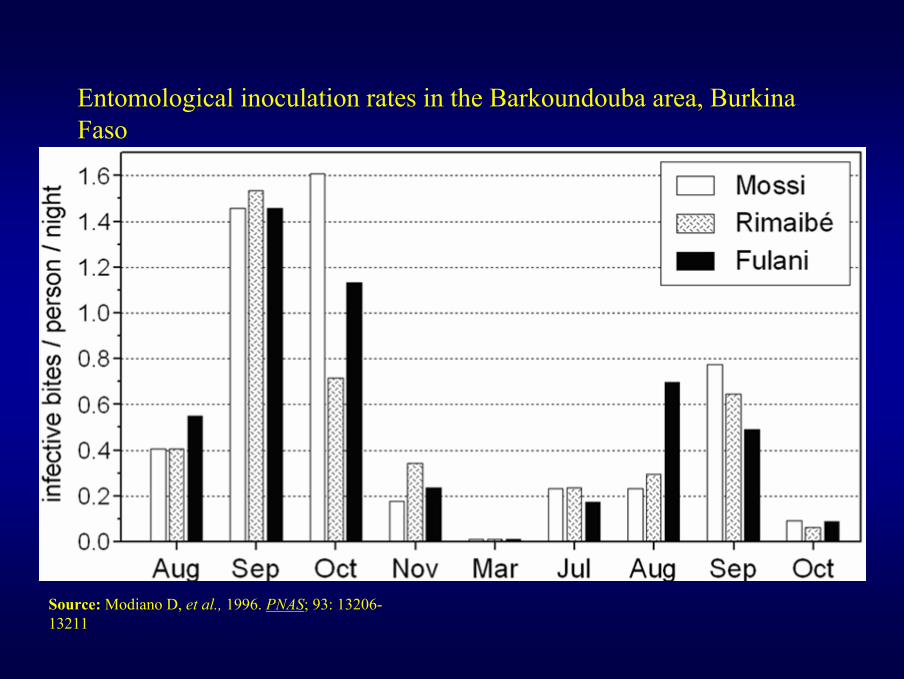
Entomological inoculation rates in the Barkoundouba area, Burkina Faso
Source: Modiano D, et al., 1996. PNAS; 93: 13206-13211
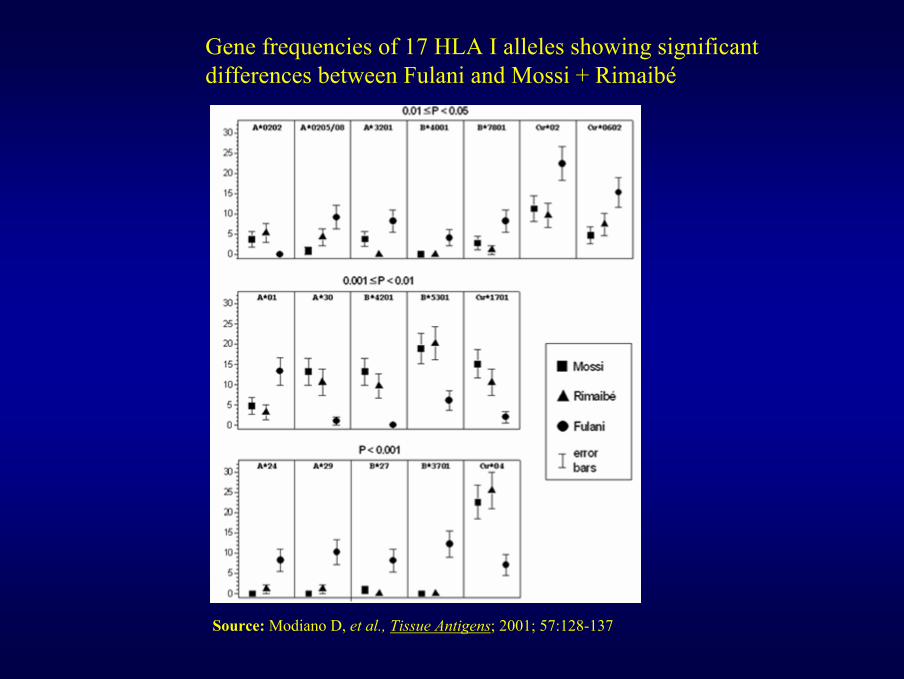
Gene frequencies of 17 HLA I alleles showing significant differences between Fulani and Mossi + Rimaibé
Source: Modiano D, et al., Tissue Antigens; 2001; 57:128-137
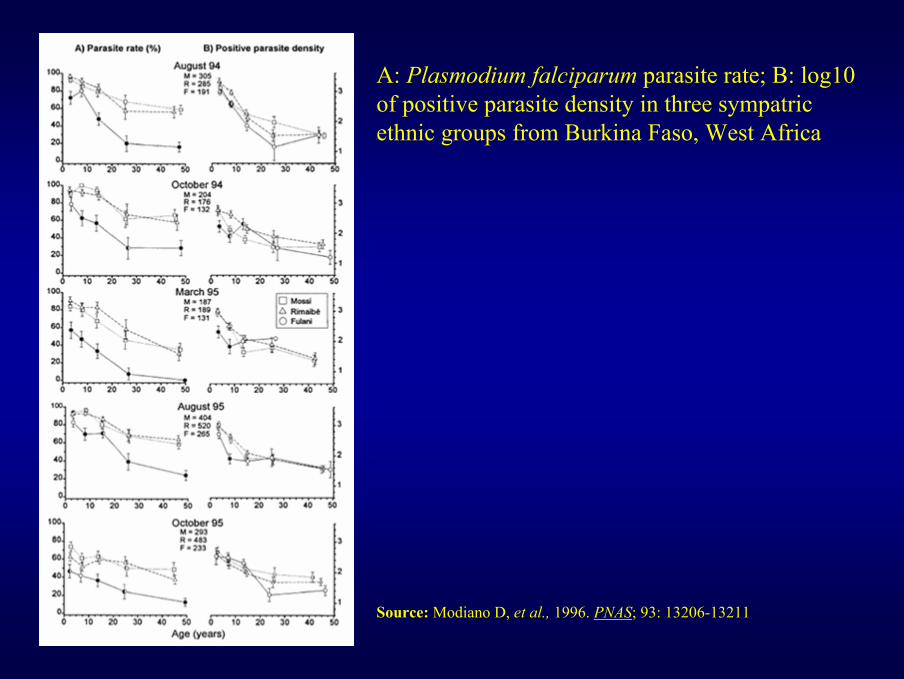
A: Plasmodium falciparum parasite rate; B: log10 of positive parasite density in three sympatric ethnic groups from Burkina Faso, West Africa
Source: Modiano D, et al., 1996. PNAS; 93: 13206-13211
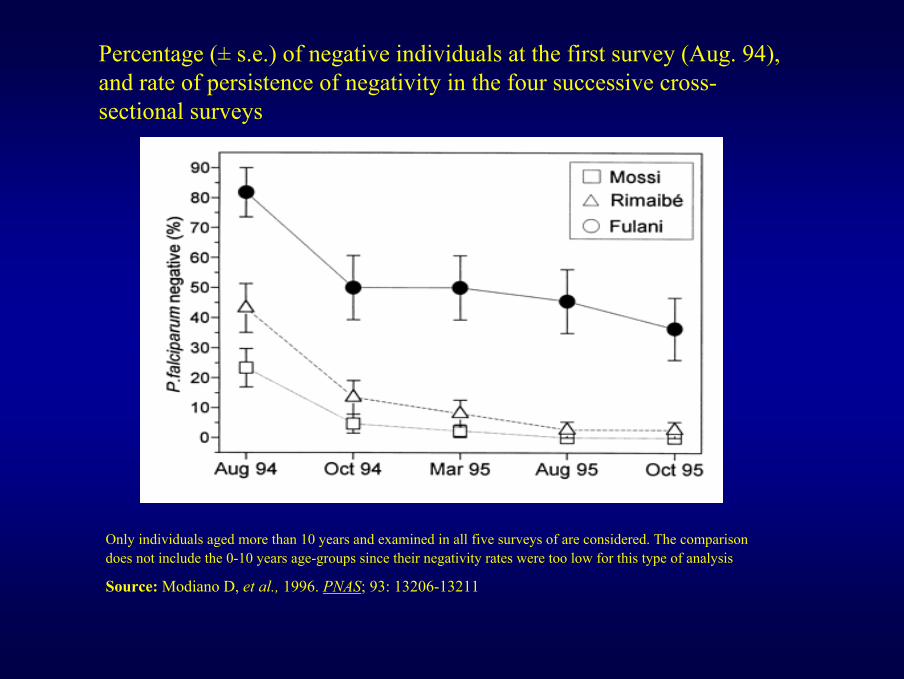
Percentage (± s.e.) of negative individuals at the first survey (Aug. 94), and rate of persistence of negativity in the four successive cross-sectional surveys
Only individuals aged more than 10 years and examined in all five surveys of are considered. The comparison does not include the 0-10 years age-groups since their negativity rates were too low for this type of analysis
Source: Modiano D, et al., 1996. PNAS; 93: 13206-13211
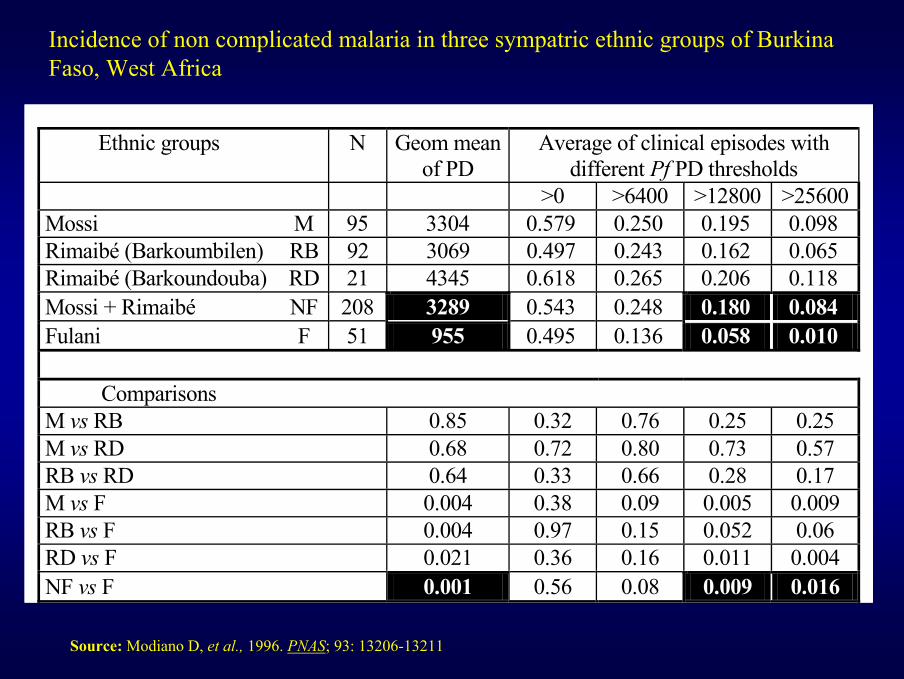
Ethnic groups N Geom mean of PD
Average of clinical episodes with different Pf PD thresholds
>0 >6400 >12800 >25600Mossi M 95 3304 0.579 0.250 0.195 0.098 Rimaibé (Barkoumbilen) RB 92 3069 0.497 0.243 0.162 0.065 Rimaibé (Barkoundouba) RD 21 4345 0.618 0.265 0.206 0.118 Mossi + Rimaibé NF 208 3289 0.543 0.248 0.180 0.084 Fulani F 51 955 0.495 0.136 0.058 0.010
Comparisons
M vs RB 0.85 0.32 0.76 0.25 0.25 M vs RD 0.68 0.72 0.80 0.73 0.57 RB vs RD 0.64 0.33 0.66 0.28 0.17 M vs F 0.004 0.38 0.09 0.005 0.009 RB vs F 0.004 0.97 0.15 0.052 0.06 RD vs F 0.021 0.36 0.16 0.011 0.004 NF vs F 0.001 0.56 0.08 0.009 0.016
Incidence of non complicated malaria in three sympatric ethnic groups of Burkina Faso, West Africa
Source: Modiano D, et al., 1996. PNAS; 93: 13206-13211
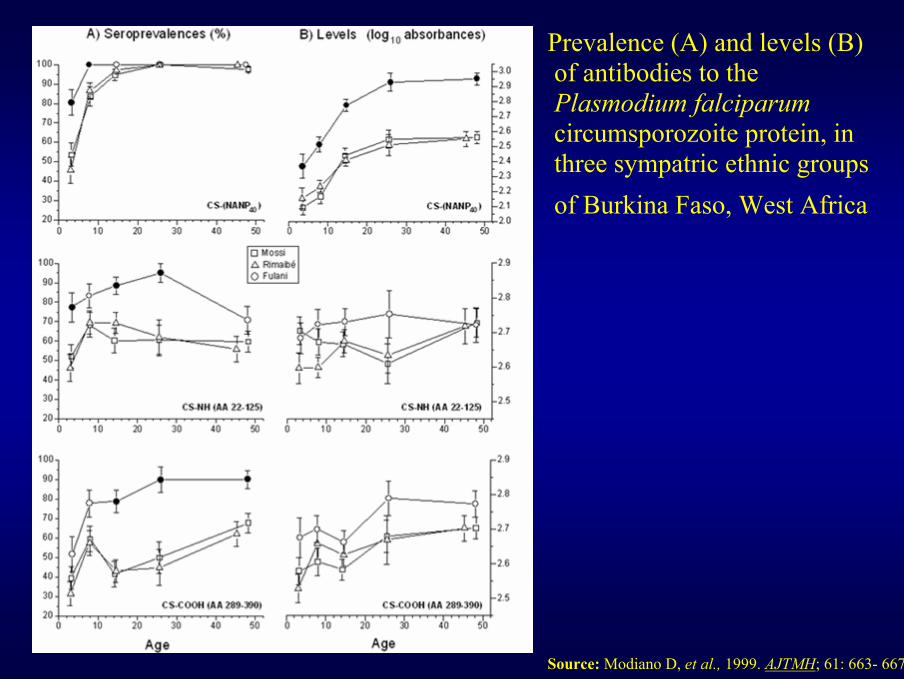
Prevalence (A) and levels (B)of antibodies to thePlasmodium falciparumcircumsporozoite protein, inthree sympatric ethnic groupsof Burkina Faso, West Africa
Source: Modiano D, et al., 1999. AJTMH; 61: 663- 667
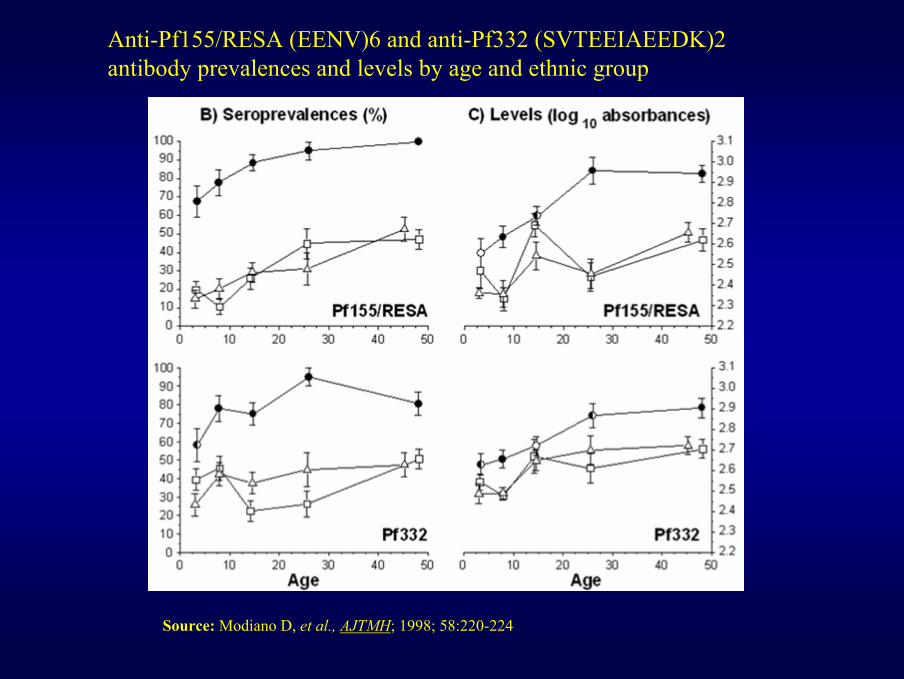
Anti-Pf155/RESA (EENV)6 and anti-Pf332 (SVTEEIAEEDK)2 antibody prevalences and levels by age and ethnic group
Source: Modiano D, et al., AJTMH; 1998; 58:220-224
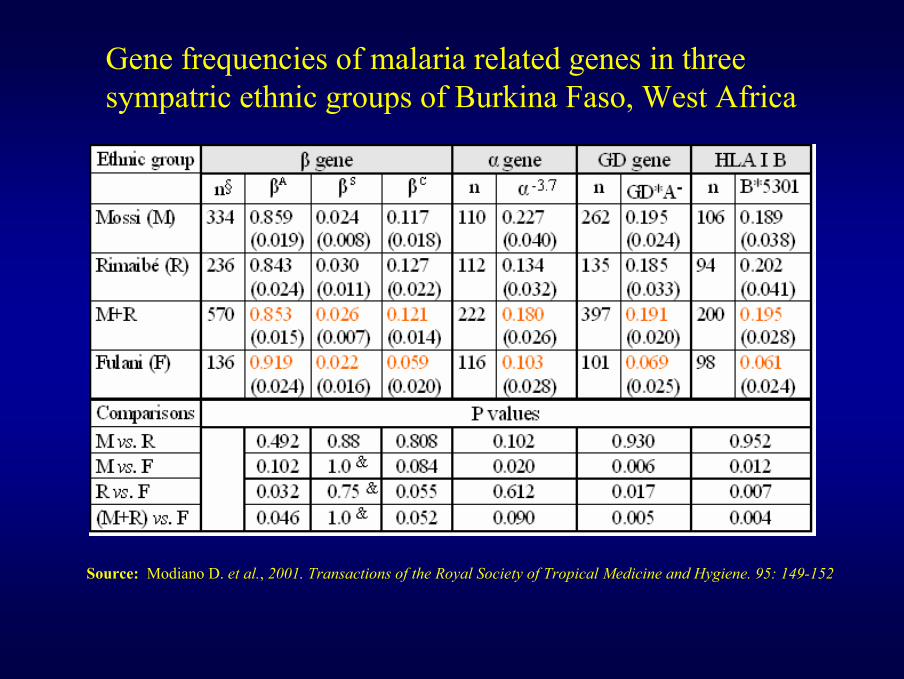
Source: Modiano D. et al., 2001. Transactions of the Royal Society of Tropical Medicine and Hygiene. 95: 149-152
Gene frequencies of malaria related genes in three sympatric ethnic groups of Burkina Faso, West Africa
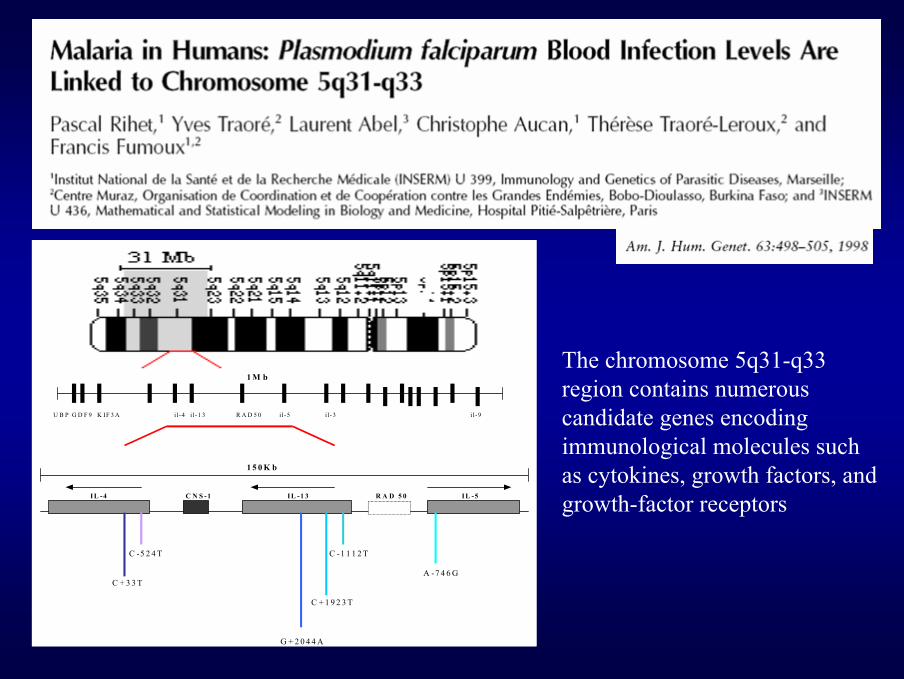
1 M b
U B P G D F 9 K IF 3 A i l-4 i l-1 3 R A D 5 0 i l -5 i l-3 i l-9
1 5 0 K b IL -4 C N S -1 IL -1 3 R A D 5 0 IL -5 C -5 2 4 T C -1 1 1 2 T
A -7 4 6 G C + 3 3 T
C + 1 9 2 3 T G + 2 0 4 4 A
The chromosome 5q31-q33 region contains numerous candidate genes encoding immunological molecules such as cytokines, growth factors, and growth-factor receptors
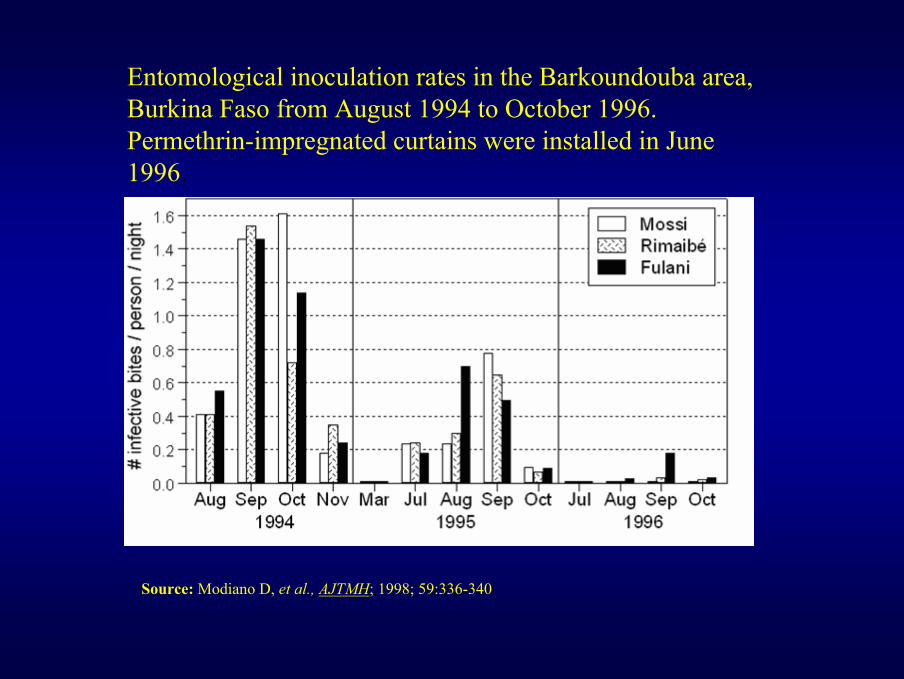
Entomological inoculation rates in the Barkoundouba area, Burkina Faso from August 1994 to October 1996. Permethrin-impregnated curtains were installed in June 1996.
Source: Modiano D, et al., AJTMH; 1998; 59:336-340
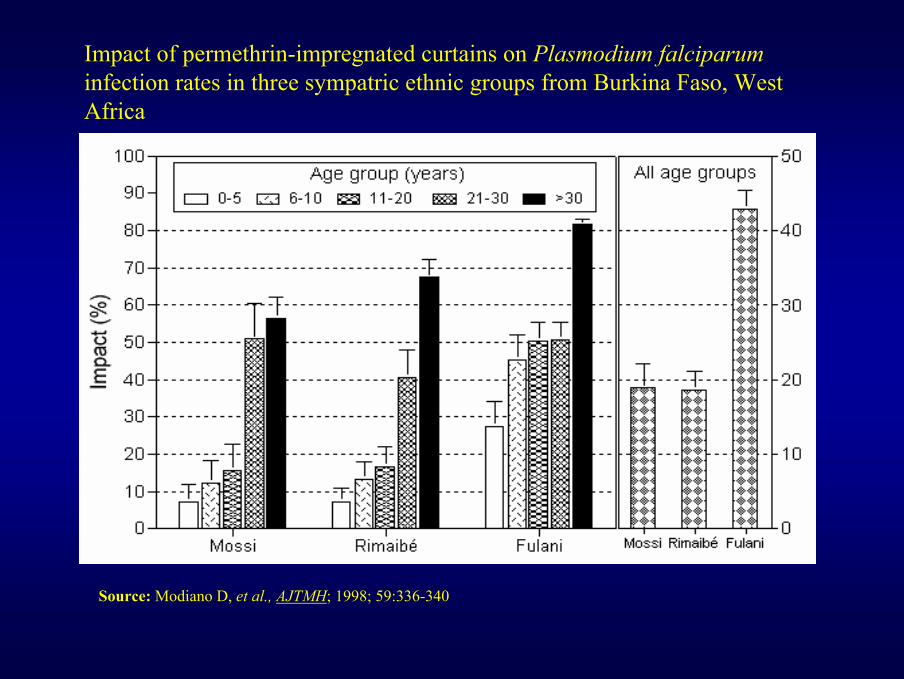
Impact of permethrin-impregnated curtains on Plasmodium falciparuminfection rates in three sympatric ethnic groups from Burkina Faso, West Africa
Source: Modiano D, et al., AJTMH; 1998; 59:336-340

Dipartimento di Scienze di Sanità Pubblica, Sezione di Parassitologia, Università “La Sapienza” Rome, ItalyD. Modiano, G. Luoni, F. Verra, G.Paganotti, V. Petrarca, M. Coluzzi
Dipartimento di Biologia, Università “Tor Vergata” Rome, Italy;G. Modiano, F. Pompei, G. Bancone, BM Ciminelli
Associazione Nazionale Lotta contro le Microcitemie, Rome, Italy; I. Bianco, P. Grisanti, E. Foglietta
Tissue Antigen Lab., Imperial Cancer Research Found,London, UK;J. Bodmer
Department of Immunology Stockholm University, Sweden; P Perlmann, H Perlmann, M Troye Blomberg
Institute of Cell, Animal and Population Biology, Edinburgh, UK;D Walliker, H Babiker
Wellcome Trust Center for Human Genetics, Oxford, UK; D. Kwiatkowski
Centre National de Récherche et Formation sur le Paludisme, Ministère de la Santé, Burkina Faso;
BS Sirima, I Nebié, D Diallo, A Konaté, J Simporé
Hopital Yalgado Ouedraogo, Ouagadougou, Burkina FasoA. Sawadogo, I Sanou
Ecole de Medecine et Pharmacie, Université de Bamako, MaliO Doumbo, A Dolo