Embed Size (px)
Citation preview

7
Chapitre 2
Revue de littérature
La compréhension des métabolismes primaire et secondaire est une étape essentielle dans
la démarche de modélisation, car la vraisemblance du modèle dépend directement des hypo-
thèses proposées pour son développement. Il est donc nécessaire de cerner dans un premier
temps les mécanismes qui permettent à une plante de vivre. La constitution biochimique, le
rôle des différents nutriments qui lui sont nécessaires et leur dynamique intracellulaire posent
les bases de la compréhension du métabolisme primaire de la cellule végétale. Le métabo-
lisme secondaire décrit les multiples voies de biosynthèse de composés non essentiels pour le
développement, la croissance et la respiration de la plante. Certains mécanismes d’activation
de ces voies métaboliques commencent à être élucidés et leur connaissance apporte à l’in-
génieur un outil supplémentaire pour augmenter la production de métabolites secondaires.
De manière plus spécifique, l’étude du métabolisme secondaire deCatharanthus roseusré-
vèle des voies métaboliques complexes et fortement régulées par la différenciation cellulaire.
Dans ce contexte, les racines transformées parAgrobacterium rhizogenesreprésentent un ou-
til de choix lorsque la différenciation cellulaire est une nécessité. La transformation génétique
parA. rhizogeness’effectue en trois étapes dont la connaissance a permis d’établir plusieurs
protocoles expérimentaux de transformation de plantes avecA. rhizogenes. Les racines trans-
formées deC. roseusfournissent ainsi un système modèle utilisé pour étudier le métabolisme
des alcaloïdes à noyau indole.

8
Les revues de littérature sur les techniques de quantification des alcaloïdes à noyau indole
et sur l’utilisation de phases extractives sur des racines transformées et des cellules en sus-
pension de plante apparaissent dans les publications traitant des sujets respectifs. Les notions
et leur références utilisées dans la structure du modèle cinétique proposé au dernier chapitre
sont présentées avec la description du modèle.
2.1 Métabolisme primaire
2.1.1 Constitution des cellules de plantes
Les cellules de plantes sont des cellules eucaryotes possédant une paroi cellulaire com-
plexe, composée de polymères de glucides. Les principaux constituants de la cellule sont les
lipides et les phospholipides à partir desquels les membranes des différents compartiments
cellulaires sont synthétisées. Les acides aminés sont les sous-unités d’assemblage des pro-
téines. Certains d’entre eux, comme le tryptophane, sont utilisé par la cellule pour synthétiser
des facteurs de croissance tel que l’acide indole-3-acétique (IAA). Le génome de la plante est
porté par les acides nucléiques, polymères de nucléotides (ADN et ARN). Les glucides sto-
ckés sous forme de saccharose ou d’amidon servent de réserves énergétiques. Les cofacteurs
enzymatiques tels que la coenzyme A, le NAD, le NADP sont essentiels pour le fonction-
nement du métabolisme de la cellule. De nombreux intermédiaires métaboliques comme le
pyruvate et l’acétyl-coenzyme A interviennent dans de nombreuses réactions de catabolisme
et d’anabolisme (Brownleader et al., 1997). L’ensemble de ces constituants cellulaires est
constitué d’atomes de carbone (45 %), d’hydrogène (6 %), d’oxygène (45 %), d’azote (1,5
%), de potassium (1 %), de calcium (0,5 %), de magnésium (0,2 %), de phosphate (0,2 %),
de soufre (0,1 %) et de silicium (0,1 %) (Epstein, 1972). D’autres éléments tels que manga-
nèse, le zinc, le fer, le nickel, le bore, le cuivre et le molybdène entrent dans la composition
de certaines espèces chimiques, mais leur concentration dans la matière sèche végétale est
extrêmement faible (de l’ordre de quelques ppm) (Taiz et Zeiger, 1998). Ces éléments sont
appelés micronutriments. D’autres ions comme le chlorure, le nitrate sont présents dans la

9
cellule, mais ne sont pas incorporés dans des composés cellulaires. Ainsi, de nombreux nu-
triments sont nécessaires pour synthétiser les métabolites cellulaires.
2.1.2 Nutriments utilisés par la plante
Macronutriments. Les différents constituants de plante sont apporté par des minéraux qui
assurent la fonction de nutriments pour la plante. Le rôle essentiel du potassium est de par-
ticiper au gradient électrochimique de part et d’autre de la membrane plasmique grâce à sa
concentration cytoplasmique intracellulaire élevé qui est de l’ordre de 100 mM (Clarkson et
Hanson, 1980). Le chlorure assure l’électroneutralité du cytosol (Clarkson et Hanson, 1980).
Il intervient également dans certaines réaction photosynthétiques (Haehnel, 1984). Il jouerait
également un rôle dans la division cellulaire des feuilles et des racines (Terry, 1977).
Le calcium est un messager secondaire pour de nombreuses mécanismes de réponse aux
signaux environnementaux et hormonaux (Hepler et Wayne, 1985). Il peut être séquestré par
la calmoduline, une protéine cytoplasmique. Le complexe calcium-calmoduline est impli-
qué dans la régulation de nombreuses voies métaboliques associés à la réponse à un signal
(Sanders et al., 1999). Le magnésium est aussi un messager secondaire pour la régulation de
l’expression de gènes (Prescott, 1993) et joue un rôle dans l’activation des enzymes impli-
quées dans la respiration, la photosynthèse et la synthèse d’ADN et d’ARN (Taiz et Zeiger,
1998).
Le soufre est utilisé pour les réactions d’oxydoréduction (Bouma, 1975), et est intégré
dans méthionine et la cystéine, 2 acides aminés essentiels pour la formation de la structure
tertiaire des protéines. Enfin, il entre dans la composition de plusieurs cofacteurs comme la
coenzyme A (Michal, 1999).
Enfin, les squelettes carbonés dont la plante a besoin pour vivre et croître proviennent de
la fixation du CO2 en trioses puis en hexoses par les chloroplastes des parties aériennes. Les
hexoses (glucose et fructose) sont condensés en saccharose par la saccharose synthétase. Le

10
saccharose est transporté dans les parties souterraines de la plante. Ces dernières ne possédant
pas de chloroplaste, il joue le rôle de source de carbone (Taiz et Zeiger, 1998).
Trois types de nutriments ont été plus particulièrement étudiés au laboratoire, à cause de
leur importance physiologique: le phosphate, les sources d’azote (nitrate et l’ammonium), et
le saccharose.
Phosphate. Le phosphate est utilisé par la cellule sous forme d’orthophosphate ou phos-
phate inorganique (Pi), de pyrophosphate ou de polyphosphates1. Le Pi possède 3 pKa : 2.16,
7.21, et 12.32. Dans les conditions physiologiques des cellules de plantes (pH entre 5 et 7
selon les compartiments cellulaires), H2PO−4 et HPO2−4 sont les formes prédominantes. La
cellule ne peut absorber et stocker que la forme monovalente H2PO−4 (Bieleski, 1973).
Le phosphate entre dans la composition des phospholipides, des nucléotides, des acides
nucléiques, et de nombreux intermédiaires de réaction phosphatés. Outre ce rôle structural,
le phosphate contenu dans les nucléotides triphosphatés tel que l’ATP et le GTP fourni à la
cellule des molécules ayant deux liaisons phosphatées extrêmement riches en énergie. Ces
molécules servent de navettes énergétiques. Par exemple, l’hydrolyse d’ATP en ADP libère
une enthalpie libre standard de 33 kJ/mol (Taiz et Zeiger, 1998). Inversement l’énergie libérée
par la dégradation de molécules complexes est utilisée pour la regénération d’ATP. De plus,
le Pi régule l’activité des protéines par leur phosphorylation et leur déphosphorylation. Selon
son état de phosphorylation, une protéine est soit active, soit inhibée.
Le pyrophosphate est en général produit lors des réactions de biosynthèse faisant interve-
nir des composés phosphatés. Il est rarement utilisé, sauf en cas de limitation en phosphate ou
son recyclage en 2 molécules de Pi libére de l’énergie. Dans ce cas, l’hydrolyse du pyrophos-
phate remplace l’hydrolyse d’ATP pour certaines réactions clef de la glycolyse et le pompage
de protons vers la vacuole (Plaxton, 1998).
1. utilisé par les plantes inférieures.

11
Nitrate. L’azote est extrêmement important pour la plante, car les acides aminés contiennent
tous au moins un atome d’azote. Ils servent dans la production de protéines, de nucléotides,
d’acides nucléiques de facteurs de croissance et d’autres métabolites primaires (Wink, 1997).
La plante est capable de se servir du nitrate (NO−3 ), de l’ammonium (NH+4 ) et du diazote (N2)
comme source d’azote. L’azote ne joue pas de rôle énergétique comme le phosphate, si ce
n’est qu’il entre dans la composition des nucléotides triphosphatés.
L’incorporation de l’azote a lieu par l’incorporation d’un ion ammonium dans le 2-oxo-
glutarate, un intermédiaire du cycle de l’acide citrique. Cette réaction produit le glutamate,
acide aminé utilisé pour les réactions de transamination menant aux autres acides aminés
(Michal, 1999). L’incorporation de l’ammonium peut aussi se produire lors de la synthèse
de la glutamine (qui possède 2 atomes d’azote) à partir du glutamate. Cela se produit de
manière plus spécifique au niveau du cycle de la glutamine synthétase/glutamate synthétase
(Ivashikina et Sokolov, 1997). Pour utiliser le nitrate, la cellule doit d’abord le réduire en
nitrite à l’aide de la nitrate réductase. Le nitrite est ensuite réduit en ammoniaque par la
nitrite réductase. L’ammonium est en équilibre avec sa base faible, l’ammoniaque. Le pKa
du couple est de 9,25. Ceci implique que dans le cytosol, la forme basique du couple ne
représente qu’environ 1 % de l’ammonium.
Saccharose et hexoses.Le saccharose est un dimère constitué d’une molécule de glucose
et d’une molécule de fructose. C’est le glucide qu’on retrouve dans la sève des plantes supé-
rieures et qui sert de molécule énergétique pour les plantes. La plante ne peut pas se servir
du saccharose directement, elle doit l’hydrolyser en glucose + fructose par l’action des inver-
tases (Sturm, 1999) ou par l’action de la saccharose synthétase (Huber et Huber, 1996). Dans
la cellule, le glucose et le fructose sont rapidement phosphorylés en glucose-6-phosphate et en
fructose-6-phosphate pour rentrer dans la glycolyse et la voie des pentoses phosphates (voir
la section 2.1.5). Ainsi, le métabolisme du saccharose est intimement lié à la disponibilité du
Pi dans le cytosol. Le saccharose a également un rôle de régulation de voies métaboliques et
de signalisation au sein de la plante, notamment au niveau du développement (Gibson , 2000;
Smeekens , 2000; Williams et al., 2000; Lalonde et al., 1999). Cette signalisation passe par

12
le clivage du saccharose en glucose et en fructose. Les hexokinases, enzymes activées par
les hexoses, déclenchent ensuite une cascade d’événements en phosphorylant des protéines
spécifiques (Farrar et al., 2000).
2.1.3 Bassins de stockage des nutriments
La cellule de plante contient plusieurs compartiments cellulaires comme le cytoplasme, la
vacuole, les chloroplastes les amyloplastes, les mitochondries. Ces différents compartiments
ont un rôle qui leur est propre dans la gestion des différents nutriments.
Phosphate. Les plantes sont capables de stocker le phosphate dans la vacuole. Ce méca-
nisme est essentiel à leur survie dans un environnement où en général, le phosphate n’est pas
toujours disponible dans le sol ou à des concentrations très faibles, souvent inférieures à 10
µM (Schachtman et al., 1998). Elle peuvent utiliser leur réserves pour continuer à croître si le
milieu externe ne contient plus de phosphate. Pour que le métabolisme de la plante fonctionne
correctement, la concentration cytoplasmique en Pi reste stable en conditions non limitantes.
Elle varie selon les espèces entre 5 et 30 mM (Plaxton, 1998; Mimura, 1999). En revanche,
la concentration du Pi dans la vacuole dépend des besoins de la cellule et de la disponibi-
lité du Pi extracellulaire. Elle peut monter jusqu’à 120 mM dans l’orge cultivé sur un milieu
contenant 40 mM de Pi (Mimura et al., 1990).
Nitrate et ammonium. Miller et Smith (1996) ont montré que la concentration cytoplas-
mique en nitrate est stabilisé autour de 10 à 35 mM, sauf en cas de limitation en nitrate. Les
plantes peuvent aussi stocker le nitrate dans la vacuole. Ces réservent peuvent monter jusqu’à
100 mM (Crawford et Glass, 1998). Tout comme le Pi, les limites de stockage du nitrate dans
la vacuole dépendent des espèces.
L’ammonium présent dans le cytosol est en général incorporé par la glutamate synthé-
tase qui a unKm de l’ordre de 10 a 20µM. La concentration cytoplasmique de l’ammonium
mesurée par RMN est du même ordre de grandeur. Les estimations de la concentration va-

13
cuolaire de l’ammonium dans les racines de maïs indiquent qu’elle est 2 à 5 fois supérieure à
la concentration cytoplasmique (Howitt et Udvardi , 2000).
Glucides. Les cellules de plantes peuvent stocker les hexoses sous forme d’amidon, poly-
mère de glucose, dans les plastides. La biosynthèse de l’amidon passe par la conversion du
glucose-6-phosphate en glucose-1-phosphate par la phosphoglucose isomérase. Le glucose-
1-phosphate est condensé avec une molécule d’ATP pour donner l’ADP-glucose et une molé-
cule de pyrophosphate. L’amidon synthétase polymérise l’ADP-glucose en amidon (Avigad
et Dey, 1997). La dégradation d’amidon a lieu lorsque la cellule est limité en sources de glu-
cides. L’amidon phosphorylase permet de dépolymériser l’amidon en glucose-6-phosphate en
consommant le Pi nécessaire. Lorsque la cellule est en limitation de phosphate et donc d’ATP,
son elle peut libérer du pyrophosphate en synthétisant de l’amidon. Le pyrophosphate peut
remplacer l’ATP dans certaines voies métaboliques (voir la section 2.1.5). Le saccharose peut
également être stocké dans la vacuole pour une utilisation ultérieure (Avigad et Dey, 1997).
2.1.4 Transport des nutriments dans les compartiments cellulaires
Les nutriments sont transportés dans la cellule par plusieurs systèmes. Le système le plus
simple est la diffusion passive, gouvernée par la loi de Fick. Les molécules diffusent libre-
ment à travers la membrane plasmique selon le gradient électrochimique de part et d’autre de
la membrane plasmique. Certaines molécules passent difficilement la membrane plasmique,
notamment les espèces chimiques très hydrophiles. La cellule dispose pour cela de canaux io-
niques qui permettent le passage de ces espèces à travers la membrane plasmique. Les espèces
suivent toujours le gradient électrochimique, mais la vitesse de leur transport est gouverné par
une loi de type Monod:
V = VmS
S+Km
où Vm est la vitesse maximale enzymatique etKm l’affinité de l’enzyme pour le substrat.
On parle de diffusion facilitée.

14
Enfin, beaucoup de nutriments doivent être absorbé par la plante dans le sens contraire du
gradient électrochimique. La plante a développé des systèmes de transport actif dans lesquels
un apport d’énergie est nécessaire pour transporter une espèce contre son gradient électro-
chimique. Cette énergie est fournie par l’hydrolyse d’ATP ou de pyrophosphate ou par l’uti-
lisation de cotransporteurs qui utilisent le gradient électrochimique d’une deuxième espèce
chimique comme source d’énergie. Ainsi, la cellule possède des ATPases qui transporte des
protons contre leur gradient électrochimique. Les H+-ATPase expulsent 1 proton par ATP
consommé vers le milieu extérieur tandis que les V-ATPase transportent 2 a 3 protons par
ATP dans la vacuole. Les V-PPases transportent un proton dans la vacuole par pyrophosphate
hydrolysé et se situent sur la membrane du tonoplaste (Sze et al., 1999). Les nutriments qui
doivent passer du milieu extérieur vers le cytosol (phosphate, nitrate, saccharose, hexoses)
utilisent des cotransporteurs en symport avec l’ion H+. Ceux qui doivent être expulsés du
cytosol (ex: sodium), utilisent des cotransporteur en antiport avec l’ion H+. Certains cotrans-
porteurs utilisent d’autres cations à la place du proton. Un cotransporteur Na+/Pi a été mis en
évidence dans des cellules deChara(Reid et al., 1998).
Phosphate. Le phosphate inorganique est transporté du milieu extérieur dans le cytosol par
deux H+-cotransporteurs différents: un transporteur à haute affinité (Km ≈ 5µM) et un trans-
porteur à basse affinité (Km ≈ 1−10mM) (Raghothama, 1999). Mimura (1999) a rassemblé
les Vm des transporteurs à haute et basse affinité de plusieurs espèces dontC. roseusselon
différentes conditions de cultures. Les valeurs varient énormément selon la concentration en
phosphate extracellulaire: 10-330 nmol/min/g FW pour le transporteur à haute affinité et 4-
110 nmol/min/g FW pour le transporteur à basse affinité. La stœchiométrie de transport du
phosphate est évaluée à 2-4 protons par Pi transporté (Raghothama, 1999).
Des canaux ioniques régulés par le pH et le potentiel membranaires sont responsable
de l’efflux de phosphate vers le milieu extérieur (Raghothama, 1999). Bieleski (1973) sug-
gère que l’efflux de Pi hors de la cellule sert à réguler l’homéostasie du phosphate et que
l’augmentation de l’absorption du phosphate est due la diminution de l’efflux plutôt qu’a
l’augmentation de l’influx.

15
Tableau 2.1 –Paramètres cinétiques des trois systèmes de transport de nitrate.
Transporteur Km Vm
CHATS 6-20µM 0,3-0,82µmol/g FW/h
IHATS 20-100µM 3-8 µmol/g FW/h
LATS > 50 mM ne sature pas
Le transport du Pi dans la vacuole ferait appel à deux systèmes de transport, un actif
dépendant de l’ATP, l’autre passif (Raghothama, 1999). D’un point de vue thermodynamique,
ces deux systèmes sont nécessaires car le Pi existe sous sa forme monovalente et divalente
selon pH du compartiment cellulaire (Mimura, 1999). De plus, l’énergétique du transport du
phosphate dans la vacuole est différente selon la concentration vacuolaire en phosphate qui
varie énormément selon la disponibilité du phosphate extracellulaire. Le statut en phosphate
des cellules serait le régulateur des deux systèmes. L’efflux de Pi hors de la vacuole serait de
type diffusion facilité avec unVm assez faible (Mimura, 1999).
Nitrate et ammonium. Le nitrate est transporté du milieu externe vers le cytosol des cel-
lules de racines par 3 systèmes de transport: un à basse affinité (LATS) et deux à haute affinité
(CHATS et IHATS). Le LATS a une cinétique linéaire et ne sature pas pour des concentrations
supérieures à 50 mM de nitrate extracellulaire. Les deux systèmes à haute affinité ont une ci-
nétique de type Monod. Le premier (CHATS) est exprimé de manière constitutive tandis que
le second est induit par la présence de nitrate dans le milieux extracellulaire. Le tableau 2.1
résume les données cinétiques de ces transporteurs (Crawford et Glass, 1998). Certains trans-
porteurs pourraient se comporter comme un transporteur à double affinité, ce qui complique
quelque peu la compréhension du système de transport de nitrate dans le cytosol (Williams et
al., 2000). La stœchiométrie de transport du nitrate est de 2 protons pour 1 nitrate (Crawford
et Glass, 1998). La régulation de l’expression des transporteurs de nitrate est dépendante du
nitrate extracellulaire. IHATS est fortement induit par une hausse du nitrate extracellulaire,
alors que CHATS et LATS sont exprimés de manière constitutive (Forde , 2000).

16
L’efflux de nitrate hors du cytosol est possible grâce à un canal ionique dont leKm est
de l’ordre de 5 mM, valeur proche de la concentration cytoplasmique du nitrate (Williams et
Miller , 2001).
Du point de vue thermodynamique, le transport du nitrate dans la vacuole nécessite un
apport d’énergie si le nitrate s’accumule à des niveaux supérieurs à 12 mM dans la vacuole.
Le transport actif est réalisé par un antiport de proton (Miller et Smith, 1992). L’efflux de
nitrate vacuolaire est pris en charge par des canaux ioniques de type CLC (canaux de l’ion
chlorure) (Williams et Miller , 2001).
Le fait que l’ammonium soit un acide faible implique que son transport dans les diffé-
rents compartiments cellulaires dépend non seulement du potentiel électrochimique∆Ψ de la
membrane à passer, mais aussi de la différence de pH. Des systèmes à haute et à basse affinité
sont responsable de l’assimilation de l’ammonium dans le cytosol. LeKm du système à haute
affinité est de l’ordre de 100µM. Son expression augmente lorsque l’ammonium extracel-
lulaire n’est plus disponible. Cette régulation a lieu au niveau protéique, par modification
allostérique et au niveau de la transcription. Le système à basse affinité se présente comme
un système non saturable, à cinétique linéaire, et est exprimé constitutivement (Howitt et Ud-
vardi , 2000). L’efflux de l’ammonium cytosolique est probablement régulé par des canaux
cationiques.
Le transport de l’ammonium dans la vacuole se fait par le biais de la diffusion passive
de l’ammoniaque. La haute concentration de l’ammonium dans la vacuole résulte du pH
vacuolaire plus faible de 1 unité environ par rapport au pH cytosolique. L’efflux d’ammonium
de la vacuole vers le cytosol fonctionne sur le même mécanisme qui reste peu documenté
(Howitt et Udvardi , 2000).
Glucides. Dans la nature, les parties aériennes des plantes excrètent du saccharose dans le
phloème pour nourrir les parties souterraines de la plante. Les cellules des racines assimilent
le saccharose par des transporteurs spécifiques. Le transporteur le plus répandu, SUT1 fonc-

17
tionne en mode symport avec un proton par saccharose transporté (Lalonde et al., 1999). Le
Km de ces transporteur est de l’ordre de 1 mM (Lemoine , 2000).
Le saccharose est stocké dans la vacuole. Aucun transporteur de saccharose vacuolaire
n’a été caractérisé, mais le transport du saccharose du cytosol vers la vacuole semble être
réalisé par un antiport à proton, tandis que sa libération dans le cytosol serait un phénomène
passif (Lalonde et al., 1999).
La cellule clive le saccharose en glucose + fructose par l’action des invertases que l’on
retrouve dans le cytoplasme, l’apoplaste et la vacuole (Avigad et Dey, 1997). Dans les tissus
souterrains de la plante, les invertases apoplastiques sont souvent exprimées. Ces cellules syn-
thétisent des transporteurs de monosaccharides (glucose, fructose) pour assimiler les hexoses
produits par les invertases apoplastiques (Lalonde et al., 1999). Des études cinétiques sur la
consommation de saccharose, de glucose et de fructose par des racines transformées de ca-
rotte montrent que ces cellules utilisent exclusivement ce mécanisme, ce qui suggère qu’elles
n’expriment pas de transporteur de saccharose (Jolicoeur, 1998). Les transporteurs d’hexoses
fonctionnent en mode symport de proton (Tanner et Caspari, 1996).
2.1.5 Métabolisme primaire des cellules de plante non photosynthétiques
Le métabolisme primaire des cellules de plantes possède quelques particularités par rap-
port à celui des cellules animales, des levures ou des bactéries. Pour simplifier l’étude, on se
restreint au métabolisme des tissus non photosynthétiques.
Navettes d’oxydo-réduction. Beaucoup de réactions font intervenir un transfert d’atome
d’hydrogène par oxydo-réduction. La cellule dispose de certaines molécules dont le rôle
est précisément de réduire d’autres espèces chimiques. Ces navettes d’oxydo-réduction sont
le NAD+/NADH, NADP+/NADPH et le FAD/FADH2. Les différentes navettes d’oxydo-
réduction participent à certains types de réactions. Le NAD+/NADH est utilisé dans toutes les
sortes de réactions, le NADP+/NADPH est utilisé dans beaucoup de réactions de biosynthèse
et le FAD/FADH2 est utilisé dans les réactions du cycle de l’acide citrique et de la respiration.

18
La cellule consomme les formes réduites de ces navettes et une grande partie de son métabo-
lisme est utilisé pour les régénérer, à partir de molécules organiques, telle que les hexoses, ou
par la photosynthèse. Ces formes réduites servent aussi à produire de l’ATP à partir de l’ADP.
Glycolyse. La régénération de l’ATP à partir d’ADP est réalisée en plusieurs étapes. La pre-
mière étape consiste en la transformation du saccharose en pyruvate. Cette étape est réalisée
par la glycolyse et la voie des pentoses phosphates, deux voies métaboliques qui partagent
quelques intermédiaires de réaction et qui fonctionnent en parallèle dans le cytosol.
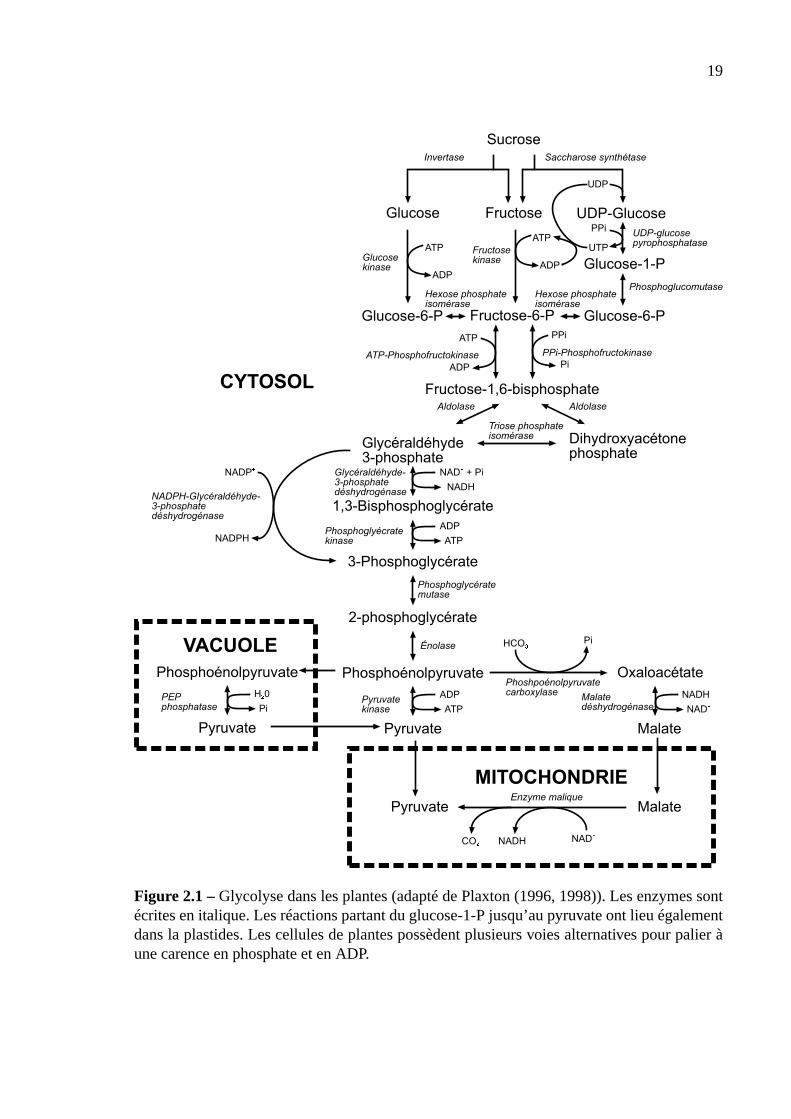
Le schéma général de la glycolyse est donné sur la figure 2.1. Bien que cela n’appa-
raisse pas sur la figure, il est important de noter que toutes les réactions à partir du glucose-
1-phosphate jusqu’au pyruvate se produisent également en parallèle dans les plastides. Le
glucose-1-phosphate provient dans ce cas de la dégradation de l’amidon. La glycolyse, comme
de nombreuses réactions, nécessite l’hydrolyse d’ATP en ADP. Cependant, en cas de carence
d’ATP et de Pi, la plupart des réactions de la glycolyse continuent d’avoir lieu grâce à l’uti-
lisation d’enzymes non dépendantes en ATP (Plaxton, 1998). En effet, plusieurs réactions de
la glycolyse sont catalysées par l’hydrolyse d’ATP ou le pyrophosphate ou le NADPHvia
des enzymes différentes. Par exemple, le mécanisme de navette de la saccharose synthétase
permet notamment de transformer une molécule de saccharose en 1 une molécule de fructose-
6-phosphate et une molécule de glucose-6-phosphate en utilisant l’énergie et les deux groupes
phosphate d’un pyrophosphate. L’hydrolyse classique du saccharose est normalement menée
par l’invertase et donne du glucose et du fructose qui doivent être phosphorylés en utilisant 2
ATP. La glycolyse produit des molécules de NADH et de NADPH en cas de limitation en Pi.
Globalement, la conversion d’une molécule de glucose en deux molécules de pyruvate
produit 2 ATP, et 2 NADH quand la cellule n’est pas en limitation de phosphate. Les intermé-
diaires de la glycolyse sont aussi des précurseurs de nombreuses voies métaboliques, comme
la synthèse de la sérine.
La glycolyse est contrôlé par la présence des intermédiaires du cycle de l’acide citrique
(voir plus loin), tel que le malate. Une accumulation de ces intermédiaires inhibe la pyruvate
19
��
�
���
�
�
�
Figure 2.1 –Glycolyse dans les plantes (adapté de Plaxton (1996, 1998)). Les enzymes sontécrites en italique. Les réactions partant du glucose-1-P jusqu’au pyruvate ont lieu égalementdans la plastides. Les cellules de plantes possèdent plusieurs voies alternatives pour palier àune carence en phosphate et en ADP.

20
�
�
�
Figure 2.2 –Voie des pentoses phosphates dans les plantes (adapté de Taiz et Zeiger (1998)).Les enzymes sont écrites en italique. Les intermédiaires réactionnels communs à la glycolysesont notés en caractères gras.
kinase, ce qui entraîne l’accumulation de phosphoénolpyruvate qui inhibe alors la conversion
du fructose-6-phosphate en fructose-1,6-bisphosphate (Taiz et Zeiger, 1998).
Voies des pentoses phosphates.La voie des pentoses phosphates est principalement res-
ponsable de la synthèse de NADPH à partir de NADP+. Elle démarre avec l’oxydation du
glucose-6-phosphate pour produire du ribulose-5-P (figure 2.2). C’est la seule voie de dégra-
dation des hexoses qui permet la réduction du NADP+. Elle produit également l’érythrose-
4-phosphate qui est utilisé dans la voie de l’acide shikimique, le ribulose-5-phosphate pour
la synthèse de l’isopentényl pyrophosphate (IPP) et des composés de la paroi cellulaire et

21
le ribose-5-phosphate, précurseur des nucléotides (Taiz et Zeiger, 1998; Bramley, 1997). Son
importance dans l’anabolisme est donc primordiale. Si ses intermédiaires réactionnels ne sont
pas utilisé dans des voies de biosynthèse, le bilan net est le suivant:
3 GLUCOSE-6-P+ 6 NADP+ −→2 FRUCTOSE-6-P+ GLYCÉRALDÉHYDE-3-P + 6 NADPH
La voie des pentoses phosphate est contrôlé par le ratio NADPH/NADP+qui influence
l’activité de la glucose-6-phosphate déshydrogénase. Plus le ratio est haut, plus l’inhibition
est importante (Taiz et Zeiger, 1998). Ces réactions ont lieu dans le cytosol.
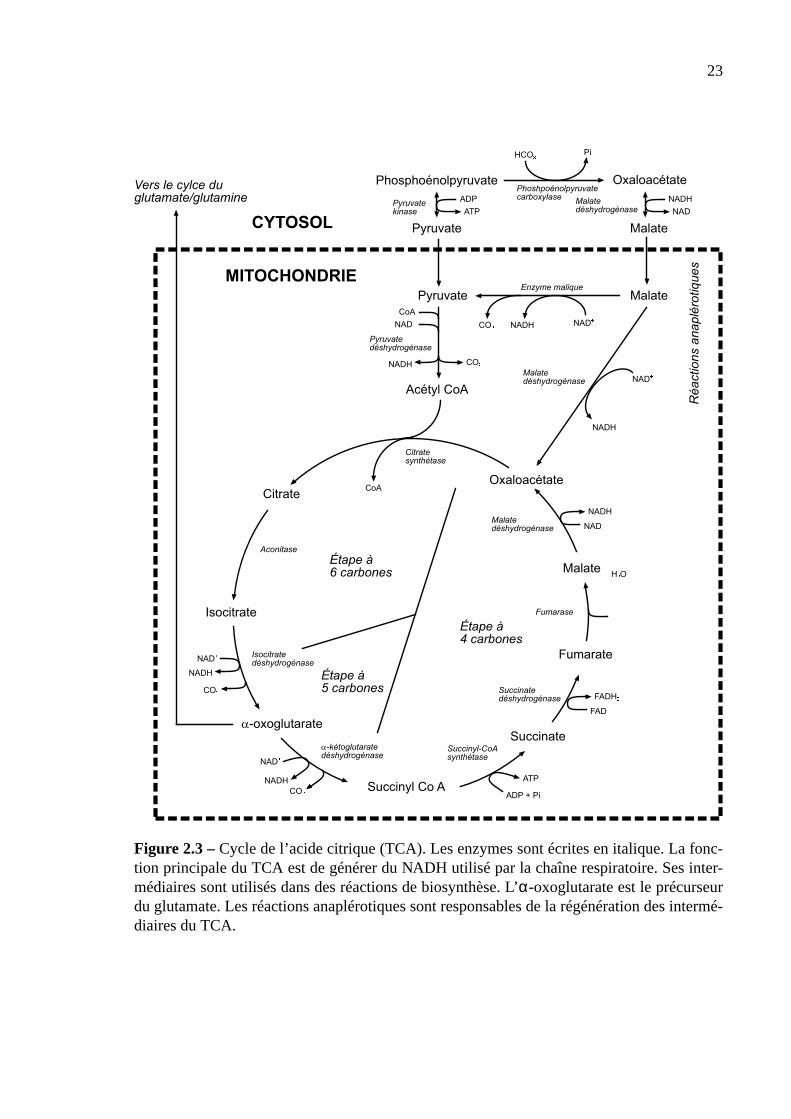
Cycle de l’acide citrique. Le cycle de l’acide citrique ou cycle des acides tricarboxyliques
(TCA) est responsable de la dégradation du pyruvate (figure 2.3) et a lieu dans la mitochon-
drie. Le pyruvate est converti en acéyl-coenzyme A. Ce dernier est condensé avec l’oxaloa-
cétate (quatre atomes de carbone) pour donner le citrate (six atomes de carbone). Le citrate
est ensuite dégradé en oxaloacétate et une autre molécule de d’acétyl-coenzyme A peut venir
compléter un nouveau cycle. Au bilan, la dégradation d’une molécule de pyruvate produit
quatre molécules de NADH, une molécule de FADH2 et la régénération d’un ATP. Trois mo-
lécules de CO2 sont également produites.
Les intermédiaires du TCA servent également à de nombreuses biosynthèses. L’oxoglu-
tarate est le précurseur du glutamate et de la glutamine. L’oxaloacétate est le précurseur de
l’aspartate. L’acétyl-coenzyme A est utilisé pour la synthèse des lipides et dans la voie de
l’acide mévalonique. Contrairement à la glycolyse ou à la voie des pentoses phosphates, le
TCA nécessite la présence d’intermédiaires pour fonctionner. Si les réactions de biosynthèse
venaient épuiser tous les intermédiaires du TCA, il n’y aurait plus d’oxaloacétate et le cycle
finirait par s’arrêter. Pour pallier à ce danger, la cellule dispose des réactions anaplérotiques
qui produisent du malate et de l’oxaloacétate à partir du pyruvate (Brownleader et al., 1997;
Taiz et Zeiger, 1998). Le TCA est controlé par la concentration en NADH, qui à forte concen-

22
tration inhibe plusieurs enzymes du cycle.
Le TCA des plantes se distingue de celui des cellules de mammifères par la présence de
l’enzyme malique qui permet de produire du pyruvate à partir de malate. Ce mécanisme est
mis en place lors d’une limitation en phosphate qui entraîne généralement une limitation en
ADP. Dans une telle situation, la pyruvate kinase est limitée par le manque d’ADP. Le malate
peut aussi être excrété dans le milieu extérieur pour solubiliser le Pi minéral (Plaxton, 1996).
Respiration. La chaîne respiratoire est responsable de l’oxydation du NADH et du FADH2
en utilisant comme accepteur d’électrons le dioxygène. Ce transfert d’électrons a lieu dans la
membrane interne de la mitochondrie et est accompagné du transfert de protons de la matrice
vers l’espace intermembranaire de la mitochondrie. Ceci crée un gradient de protons qui est
utilisé par les F0F1ATPases pour produire de l’ATP à partir de l’ADP. Chaque NADPH libère
deux électrons. Le passage de ces deux électrons dans les complexes respiratoires exporte 10
protons dans l’espace intermembranaire. Le FADH2 est oxydé comme le NADH mais son
potentiel d’oxydo-réduction plus élevé (-45 mV contre -320 mV) ne lui permet pas de parti-
ciper à toute la chaîne respiratoire. Son oxydation ne permet le transport que de six protons.
Chaque F0F1ATPase nécessite trois protons pour la synthèse d’un ATP. Un proton est égale-
ment nécessaire pour le transport d’un ADP + Pi dans la mitochondrie et d’un ATP vers le
cytosol. Ainsi, 2,5 ATP sont théoriquement synthétisés par molécule de NADPH oxydée dans
la chaîne respiratoire (Brownleader et al., 1997). La chaîne respiratoire a donc deux fonctions:
générer de l’ATP en grandes quantités et oxyder le NADH et le FADH2. Cependant, il faut
noter que l’oxydation du NADH est réalisé par beaucoup d’autres réactions biochimiques.
La chaîne respiratoire est contrôlé par le ratio ATP/ADP et la concentration en ADP et
Pi. Un manque d’ADP ou de Pi a pour effet d’inhiber la chaîne respiratoire. Ceci induit une
accumulation de NADH qui peut à son tout inhiber le TCA. L’inhibition du TCA mène à
l’accumulation de malate qui à son tour inhibe la pyruvate kinase. Le phosphoénolpyruvate
s’accumule alors, provoquant l’inhibition de la conversion du fructose-6-P en fructose-1,6-
bisphospate. Ainsi, une baisse de la concentration en ADP peut mener au blocage de l’en-
23
���
�
�
�
�
�
�
�
�
�
�
Figure 2.3 –Cycle de l’acide citrique (TCA). Les enzymes sont écrites en italique. La fonc-tion principale du TCA est de générer du NADH utilisé par la chaîne respiratoire. Ses inter-médiaires sont utilisés dans des réactions de biosynthèse. L’α-oxoglutarate est le précurseurdu glutamate. Les réactions anaplérotiques sont responsables de la régénération des intermé-diaires du TCA.

24
semble du processus de la respiration. La voie des pentoses phosphates dans ce cas ne pourra
pas lever l’inhibition, car elle accumulera du fructose-6-P (Taiz et Zeiger, 1998).
Synthèse du tryptophane. La synthèse du tryptophane mérite d’être soulignée, car le tryp-
tophane est non seulement utilisé pour le métabolisme primaire, mais aussi pour le métabo-
lisme secondaire. Elle est réaliséevia la voie de l’acide shikimique. La réaction globale peut
s’écrire:
2 Phosphoénolpyruvate+ Érythrose-4-P+ ATP + NADH + Glutamate
+ 5-P-Ribosyl-PP+ Sérine−→Tryptophane+ 2 Pi + PPi + Glycéraldéhyde-3-P+
2 Pyruvate+ NAD+ + ADP
On peut scinder cette équation en deux parties. La voie de l’acide shikimique mène au
chorismate:
2 Phosphoénolpyruvate+ Érythrose-4-P+ ATP + NADH −→Chorismate+ 2 Pi + NAD+ + ADP
Le chorismate est ensuite converti en anthranilate puis en indole et enfin en tryptophane:
Chorismate+ Glutamate+ 5-P-Ribosyl-PP+ Sérine−→Tryptophane+ PPi + Glycéraldéhyde-3-P+ 2 Pyruvate
Il est important de noter que la voie de l’acide shikimique utilise des précurseurs prove-
nant de la glycolyse (phosphoénolpyruvate) et de la voie des pentoses phosphates (Érythrose-

25
4-P et 5-P-ribosyl-PP qui vient du Ribose-5-P) (Michal, 1999). Le chorismate est aussi le
précurseur de la tyrosine et de la phénylalanine. Cette dernière est elle même précurseur de
nombreux composé phénoliques. Aussi, le chorismate est un intermédiaire réactionnel impli-
qué dans le métabolisme primaire et secondaire.
2.2 Métabolisme secondaire
2.2.1 Classification des métabolites secondaires
Classes de métabolites secondaires.Les plantes sont capables de produire de très nom-
breux métabolites secondaires qui sont classés selon leur structure chimique. A l’inverse des
métabolites primaires, les métabolites secondaires ne sont pas synthétisés de manière uni-
forme dans le règne végétal. Un métabolite secondaire particulier est souvent spécifique à
quelques espèces. Trois grandes catégories de métabolites secondaires ont été définies : les
composés phénoliques, les isoprénoides et les composés azotés. Le tableau 2.2 (adapté de
Bramley (1997); Strack (1997); Wink (1997)) présente quelques exemples de métabolites
secondaires appartenant à ces trois familles.
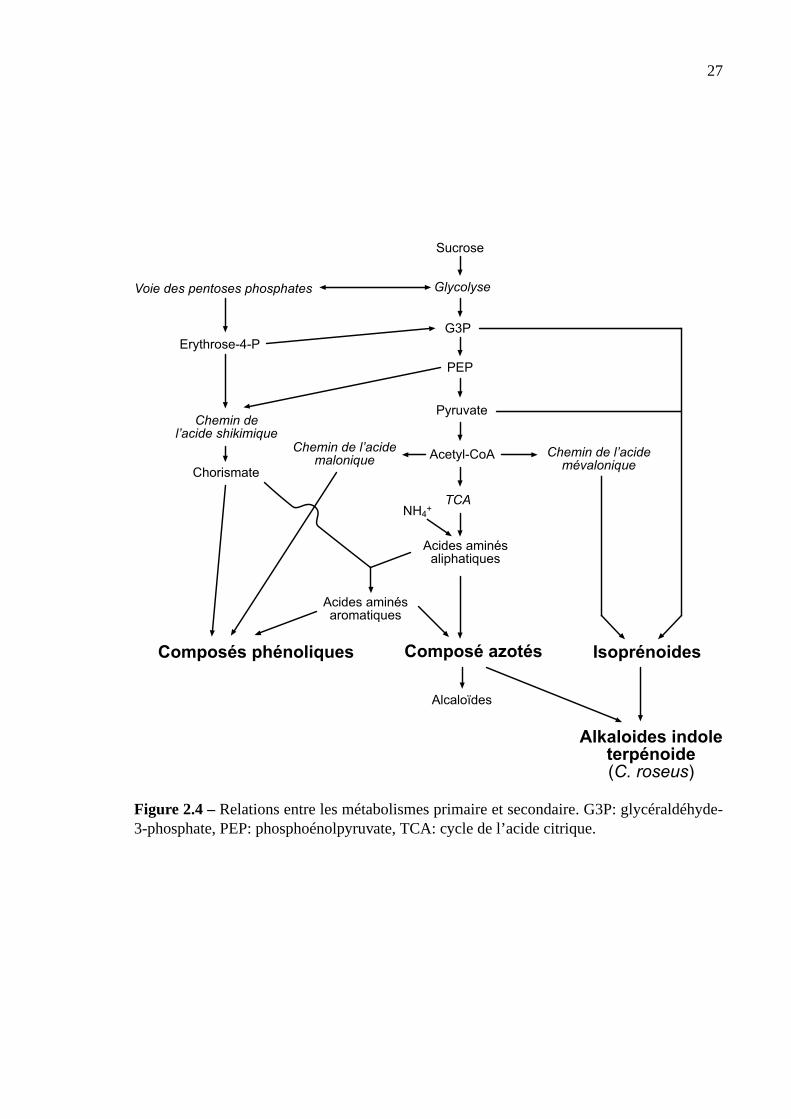
Voies de biosynthèse. Le métabolisme secondaire des plantes est lié au métabolisme pri-
maire par cinq voies métaboliques principales: la voie de l’acide shikimique, de l’acide ma-
lonique, de l’acide mevalonique, des acides aminés (Taiz et Zeiger, 1998) et du G3P via la
voie des pentoses phosphates (Contin et al., 1998). La figure 2.4 donne un aperçu des interac-
tions entre métabolismes primaire et secondaire. Les précurseurs principaux de la plupart des
métabolites secondaires sont l’acétyl-CoenzymeA, l’erythrose-4-phosphate, le phosphoénol-
pyruvate, les acides aminés, le pyruvate et le 3-phosphoglycérate.
2.2.2 Caractéristiques des alcaloïdes
Les métabolites secondaires synthétisés parC. roseusfont partie de la famille des alca-
loïdes. Les alcaloïdes sont une sous famille de composés azotés qui désignent l’ensemble

26
Tableau 2.2 –Familles de métabolites secondaires.
Famille Sous-famille Exemple
Composésphénoliques
phenols simples Arbutin dans les Rosaceae et Ericaceaehydroxybenzoates 4-Hydroxybenzoate dans plusieurs famillesAcetophenones Picein dansPicea abiesPhenyl acetates 4-Hydroxyphenylacetate dansTaxacum officinaleHydroxycinnamates Chlorogenate dans les SolanaceaePhenylpropenes Eugenol dans les huiles essentielles de plusieurs
espècesCoumarines Cichoriine dansCihorium intybusChromones Eugenine dansEugenia aromaticaNaphthoquinones Juglone dans les JuglandaceaeXanthones Mangiferine dansMangifera indicaStilbenes Resveratrol dansEucalytpusAnthraquinones Emodine dansRheum palmatumFlavonoides Rutine et anthocyanines dans plusieurs famillesLignanes Pinoresinol dansPiceaetPinusBiflavonoides Amentoflavone dans les gymnospermesCatechol melanines polymères de Naphtalène dansDaldinia
concentricaTannins hydrolisables tannin chinois dansRhus semialataLignines Lignines guaiacyl dans les gymnospermesTannins condensés Tannins dansQuercus robur
Isoprenoides
Monoterpenoides geraniol, menthol, iridoides (loganine,secologanine dansCatharanthus roseus)
Sesquiterpenoides Acide abscisique dans la plupart des plantes,lactones sesquiterpenoides
Diterpenoides Gibberellines dans toutes les plantesTriterpenoides Phytosterols, saponines, cardenolides dans la
plupart des plantesCarotenoides β-carotène dansDaucus carota, prolycopène
dansEscultentum, persicaxanthine danscitrusPolyterpenoides Caoutchouc dansHevea brasilienesis
Composés azotés
Acides aminés nonprotéiques
acide L-azetidine-2-carboxylique dans lesLiliaceae
Amines amines aliphatiques, putrescine, spermidine,tryptamine dans la plupart des plantes,cadaverines dans les Leguminosae
Glycosides cyanogéniques Linamarine, lotaustraline, lucumine dansplusieures familles
Glycosinolates Glucobrassicine, sinalbine, sinigrine dans lesdicotylédons
Alcaloïdes Cocaïne dans les Solanaceae, nicotine dans lesdifférentesNicotiana, ajmalicine, vincristine dansCatharanthus roseus, sanguinarine et berberinedans les Papaveraceae
27
Figure 2.4 –Relations entre les métabolismes primaire et secondaire. G3P: glycéraldéhyde-3-phosphate, PEP: phosphoénolpyruvate, TCA: cycle de l’acide citrique.

28
Figure 2.5 –Structures de la norclaurine et de la strictosidine.
des composés azotés produits par les plantes qui ne sont pas classés dans les sous familles
suivantes: peptides, acides aminés non protéiques, amines, glycosides cyanogéniques, glu-
cosinolates, cofacteurs, phytohormones ou métabolites primaires (comme les purines et les
pyrimidines qui sont des précurseurs des acides nucléiques). En général, ces composés pos-
sèdent au moins un atome d’azote hétérocylique. Actuellement, la structure chimique d’en-
viron 16000 alcaloïdes est connue. Environ 20 % des espèces de plantes produisent des al-
caloïdes (Memelink et al., 2001). Les alcaloïdes ont en général une activité biologique et
à ce titre, ils entrent dans la composition de nombreux médicaments comme principe actif.
Les plantes les utilisent pour la plupart d’entre eux dans leur système de défense contre les
herbivores et les pathogènes (Caporale, 1995; Wink, 1999), car ces composés sont toxiques.
Biosynthèse. La plupart des alcaloïdes sont dérivés d’acide aminés tels que le tryptophane,
l’ornithine, la lysine, l’asparate, l’anthranilate, la phénylalanine et la tyrosine. Ces acides ami-
nés sont décarboxylés en amines qui sont couplées à d’autres squelettes carbonés. La stric-
tosidine et la norcoclaurine sont deux composés centraux source de la moitié des alcaloïdes
connus (figure 2.5).
2.2.3 Métabolisme secondaire deCatharanthus roseus
C. roseusproduit essentiellement des alcaloïdes indole terpénoïques (TIA) comprenant
environ 3000 composés qui incluent la vincristine (anti-cancéreux), la quinine (utilisée contre
la palludisme) et la strychnine (pesticide contre les rats). La structure de base des TIA com-
prend un noyau de type indole provenant de la tryptamine et une composante terpénoïque

29
provenant de la sécologanine.
Biosynthèse de la tryptamine. La voie de l’acide shikimique est responsable de la forma-
tion de l’acide shikimique à partir d’une molécule de PEP (glycolyse) et d’une molécule de
E4P (voie des pentoses phosphates) (Herrmann et Weaver, 1999). Les différentes réactions
utilisent le pouvoir réducteur d’une molécule de NADPH pour donner une molécule d’acide
shikimique. Ce dernier se combine avec une molécule de PEP pour donner l’acide choris-
mique (CHO). Le CHO est aminé en anthranilate par une réaction de transamination par la
glutamine (Taiz et Zeiger, 1998). Finalement, l’anthranilate est transformé en indole par plu-
sieurs réactions enzymatiques qui utilisent une molécule de 5-P-ribosyl-pyrophosphate (voie
des pentoses phosphates) et donnent une molécule de pyrophosphate et une molécule de G3P.
L’indole est convertit en tryptophane par l’incorporation d’une molécule de sérine (Michal,
1999). La tryptamine est obtenue par décarobxylation du tryptophane par la tryptophane de-
carboxylase (TDC) (Noé et al., 1984). ChezC. roseus, la TDC est régulée par les facteurs
de développement et par les mécanismes de défense de la plante (Menke et al., 1999). L’ex-
pression de la TDC est aussi induite par les UV (Ouwerkerk et al., 1999). Sa surexpression
dans des cellules en suspension deC. roseusest toxique et non nécessaire pour augmenter la
production de TIA (Canel et al., 1998).
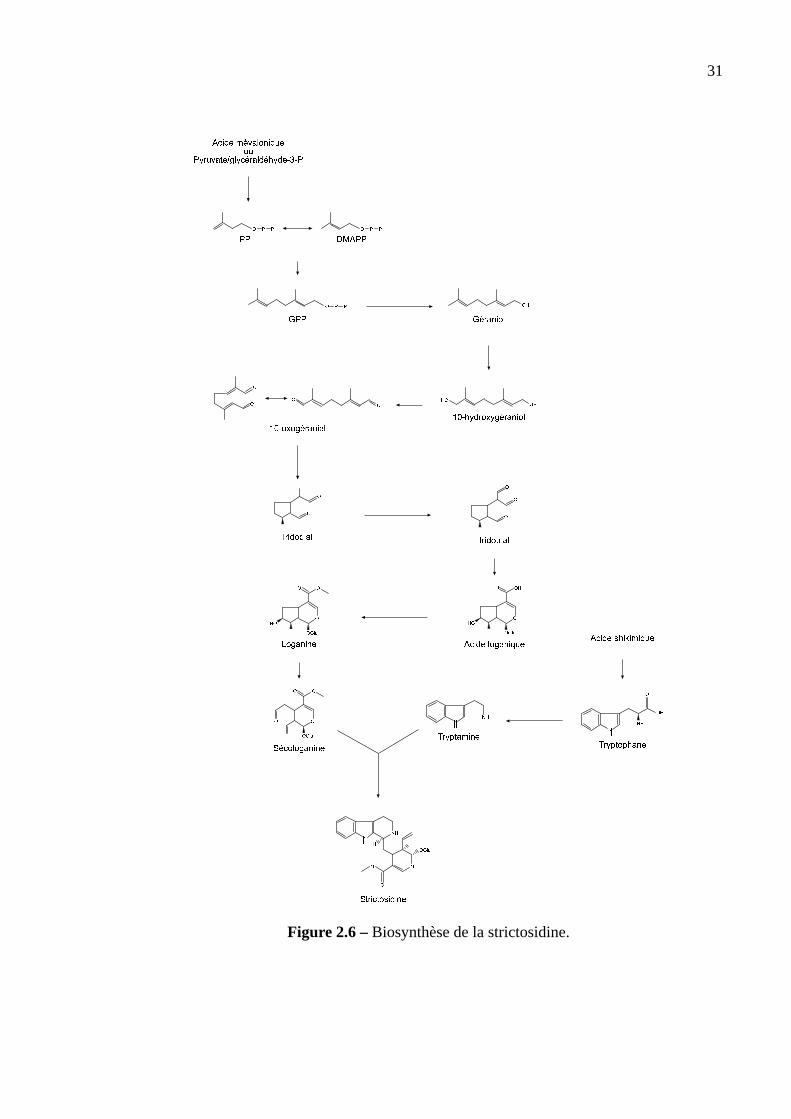
Biosynthèse de la sécologanine.La sécologanine est synthétisée via l’isopentényl pyro-
phosphate (IPP), précurseur de tous les isoprénoides. Deux voies de biosynthèse sont connues.
La première utilise la voie de l’acide mévalonique, synthétisé à partir de 3 molécules d’acétyl-
Coenzyme A, via plusieurs réactions enzymatiques qui consomment 2 NADPH. La conver-
sion de l’acide mévalonique en IPP consomme 3 molécules d’ATP et fixe 2 Pi (Taiz et Zeiger,
1998). Le bilan s’écrit:
3 Acétyl-coenzyme A+ 2 NADPH + 3 ATP −→IPP + 3 ADP + Pi + 2 NADP+

30
La seconde voie a été proposée récemment grâce à un marquage glucose au14C et un
suivi par RMN. Elle utilise du glycéraldéhyde-3-phosphate (G3P) provenant de la voie des
pentoses phosphates et du pyruvate (Contin et al., 1998). Les réactions intermédiaires de cette
seconde voie ne sont pas encore bien connues.
L’IPP est ensuite convertit en sécologanine par plusieurs réactions. Certaines d’entre elles
font intervenir des enzymes qui ont été caractérisées. L’IPP est transformé en géraniol, lui
même oxydé en 10-hydroxygéraniol via l’action d’une monooxygénase P450 dépendante du
NADPH qui semble être l’étape limitante de la synthèse de la sécologanine (Meehan et Coc-
sia, 1973). Cinq réactions transforment le 10-hydroxygéraniol en loganine (Sampaio-Santos
et Kaplan , 2001). Un cytochrome P450 catalyse la rupture oxydative de l’anneau cyclopen-
tane de la loganine pour donner la sécologanine (Yamamoto et al., 2000). La figure 2.6 résume
ces étapes.
Biosynthèse du précurseur des TIA. La strictosidine synthétase (STR) est responsable
de la condensation de la tryptamine avec la sécologanine en strictosidine. Cette enzyme est
codée par un seul gène dont la séquence a été déterminée (McKnight et al., 1990). Comme
la TDC, elle est fortement régulée par le développement de la plante et par l’induction des
mécanismes de défense (Menke et al., 1999). La surexpression de STR dans des cellules de
C. roseusa permis d’augmenter un peu la production d’alcaloïdes (Canel et al., 1998). La
strictosidine possède une groupe glucoside qui est éliminé par l’action de la strictosidineβ-
D-glucosidase (SGD) (Luijendijk et al., 1998). La strictosidine déglycosylée est convertie en
plusieurs étapes en 4,21-déhydrogeissoschizine, précurseur commun de tous les TIA.
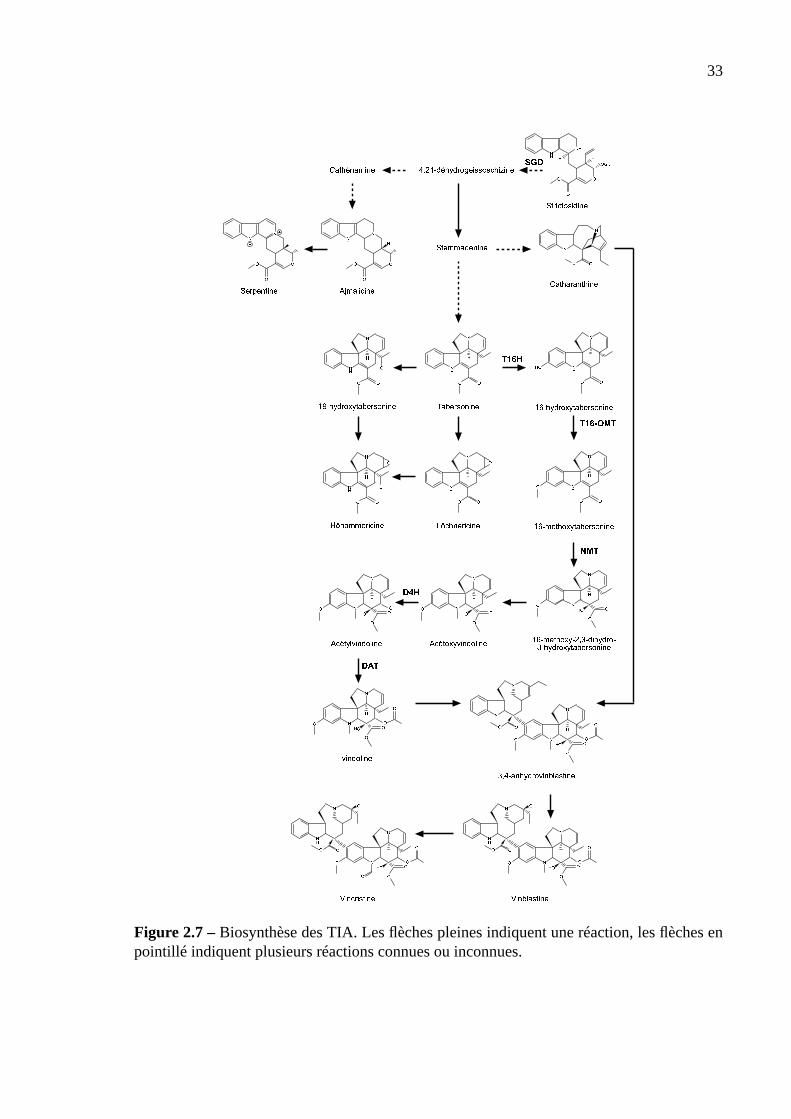
Biosynthèse des TIA. Bien qu’une centaine de TIA aient été identifiés, peu d’enzymes
impliquées dans leur biosynthèse ont été identifiées (Meijer et al., 1993).C. roseusproduit
trois types de TIA: corynanthe, iboga et pluméra, dont les représentants typiques sont respec-
tivement l’ajmalicine, la catharanthine et la vindoline. La 4,21-déhydrogeissoschizine peut
donner la cathénamine qui donne ensuite l’ajmalicine oxydée en serpentine. Ces deux der-
niers composés sont utilisés comme hypertenseurs. La biosynthèse de l’ajmalicine à partir de
31
����� ��������� ����� ��������
������������� ��! "�# ��$�%�����# &�%�'���&���(�)�(*�
+�,�- .�/0�1�- 2�- 3- 4�5�/
687:9�; <�7 =�>�; ?�; @�A
B
CEDEF G
HJIKL MN
O
P
QSR�T�U�V W�XY Z�[
\J]_^`a
bSc�d�e�f:g�e�h�i�j�k
l*m_nop
qsr
t
uwv�x�y�z y�{�|�}�~ }��
������ �
��
�
����� ���� ��������� �����
�����E�E� �
���
¡�¢�£�¤�¥�¦ ¥�§
¨©«ª¬EE® ¯
°±
² ³�´ µ�¶�·:³�´ ¸�¹
º
»
¼
½ ¾�¿ À�Á�À�¿ Â�Ã
Ä
Å
ÆÈÇ�É�Ê�Ë�Ê�Ì�Í�ÎÐÏ�Ñ�Ò Ï�Ó
ÔÕÖ
×ØÈÙ�Ú�Û�Ü�Ý�Þ�ß�à�Ü�á�â�Þ�ã�ä�å ß�æ
çsèé«ê
ëíì�î�ï�ð�ñ ò�óô�õ
öø÷�÷ùûúýü
þ ÿ���������
� ���
����
Figure 2.6 –Biosynthèse de la strictosidine.

32
la strictosidine implique l’action d’au moins 10 enzymes dont une a été clonée (Dogru et al.,
2000). La 4,21-déhydrogeissoschizine conduit également à la stemmadénine qui est convertie
soit en tabersonine, soit en catharanthine (Meijer et al., 1993).
Aucune enzyme menant à la synthèse de la catharanthine n’est actuellement connue. En
revanche, la biosynthèse de la vindoline à partir de la tabersonine a été très bien caracté-
risée. Cette synthèse a lieu en 6 étapes. La première étape est consiste en une hydroxyla-
tion par la tabersonine 16-hydroxylase (T16H), une monooxygénase de type P450 et dé-
pendante en NADPH (St-Pierre et DeLuca, 1995). Les étapes suivantes consistent en une
16-O-méthylation par une O-méthyltransférase (OMT) (Fahn et al., 1985), une hydratation
de la double liaison 2-3 par une enzyme encore inconnue puis une N-méthylation de l’atome
d’azote de l’anneau indole par une N-méthyltransférase (NMT) (DeLuca et al., 1987; Dethier
et DeLuca, 1993) pour former l’acétoxyvindoline. L’avant dernière étape est effectuée par un
dioxygénase (D4H) utilisant comme substrat supplémentaire une molécule de 2-oxoglutarate
(DeCarolis et DeLuca, 1993). D4H existerait sous une forme inactive et une forme active en
présence de lumière (Vazquez-Flota et al., 2000). La formation de la vindoline est catalysée
par la déacétylvindoline 4-O-acétyltransférase (DAT) (DeLuca et al., 1985; Fahn et al., 1985),
qui utilise une acétyl-coenzyme A comme source du groupement acétyl. DAT est inhibée for-
tement par la tabersonine et l’acétyl-coenzyme A (K i de l’ordre de 50µM) et faiblement par
la tryptamine, la secologanine et la vindoline (K i de l’ordre de 500µM) (Power et al., 1990).
T16H, NMT, D4H et DAT sont très fortement contrôlés par le développement de la plante
et par la lumière (DeLuca et al., 1986, 1988; St-Pierre et DeLuca, 1995). Ces enzymes se
retrouvent dans les parties aériennes de la plante uniquement (St-Pierre et al., 1999), ce qui
expliquerait pourquoi la biosynthèse de la vindoline n’est que très rarement observée dans les
racines transformées ou les cellules deC. roseus(Parr et al., 1988).
Dans les racines transformées et les cellules en suspension deC. roseus, la vindoline n’est
pas synthétisée (VanderHeijden et al., 1989; Moreno et al., 1995). La compartimentation de
D4H et de DAT dans les parties aériennes de la plante pourraient expliquer cette inhibition
(St-Pierre et al., 1999). Cependant, la tabersonine, précurseur de la vindoline, n’est pas pour
33
������������� �� ��
�
�
���
�
�
� �
����� ���! #" $�%���& '�(�)�* +!,#-�. -�)
/
0
123
45 6
7�8�9�:�; <�=�>�? >�:
@
A
BCD
E
F�G�H�I�J�K�L M H�M J�K
N
O
PQR
S
T
U�V�W X�Y�Z�[ \�]�Y�^ _�`�a�b c�d�e�f e�a
g
h
i jk
lm n
o�p�q rs�t u�v�w�x�t y�z�{�| }!~#��� ��{
�
�
���
��
���!��� ����� ��� ������� � ���
�
��
� ��
� �
���� �¡ ¢�£ ¤�¥ ¦�§�¨�£ ¥ ¦�©
ª
«¬
®¯
° ± ² ³
´¶µ�·¹¸
º¼»�½¿¾ ÀÂÁú
Ä�Å Æ�Ç�È�É Å Æ�Ê
Ë
ÌÍ
Î ÏÐ
Ñ Ò Ó
Ô
ÕÖØ×
Ù¶Ú�Û
Ü�ݼÞ
ß�à á�â ã�ä�å�æ!ç#è é�ê�ë ä�ì�í ã�î�ï ë ä�ð
ñ
òó
ô õö
÷ ø ù
ú
û
üýþÿ
��� ����� ��� � �
�
� �� �
�� � ��
�
�� ��
� �
�! " # %$ �&" #�' &(
)*
+,
-�. /01 . 2�3 . /�4
5
6 78 9
:; < =>
?
@A BC
D E
F
G�HI JLK�M N�OPQ�I RTS U�I V%W NQXVZY O�I[\ ]�^�_%` ab^Xc d�ef%g h�i%j�k jf
l
mno
p qrs t
uwv xLy%z { |{ }~�
�� � �
���w��� ����� � ��
�
�� �
��
��
�!�� ��%������ ���
�Z ¡�¢�¢L£�¤�¥%¦�§ ¦¡
¨Z© ª « ¬ © �®« ¯Z« °�±²³ ´ µ ¶
· ¸¹º »¼½¾
¿TÀ ÁÃÂ Ä Å%Æ�ÇÈXÅZÉ Ê�Ë�Ì�Í Î�Î�Ê�ÎÏ�Ç�Í ÐÍ Ñ�Ì Ò!ÓÕÔ
Figure 2.7 –Biosynthèse des TIA. Les flèches pleines indiquent une réaction, les flèches enpointillé indiquent plusieurs réactions connues ou inconnues.

34
autant accumulé. Shanks et al. (1997) ont mis en évidence la production de löchnericine et
de hörhammericine dans des racines transformées deC. roseus. Ces composés semblent être
des dérivés de la tabersonine, bien qu’aucune enzyme responsable de leur formation n’ait été
identifiée.
La catharanthine et la vindoline sont condensées par une peroxydase non spécifique pour
produire la vinblastine puis la vincristine (Sottomayor et al., 1998). L’ensemble du métabo-
lisme des TIA est résumé sur la figure 2.7.
Compartimentation des alcaloïdes. La biosynthèse des TIA est effectuée dans au moins
trois compartiments cellulaires : les plastides pour la production du tryptophane et de gé-
raniol, le cytosol pour la décarboxylation du tryptophane en tryptamine et la vacuole pour
la condensation de la tryptamine avec la secologanine. La plupart des alcaloïdes qui s’ac-
cumulent dans la cellule sont stockés dans la vacuole, mais sont parfois synthétisées dans
d’autres compartiments. La 4,21-déhydrogeissoschzine, la catharanthine, l’ajmalicine et la
vindoline sont synthétisées dans le cytosol. Les alcaloïdes bisindole comme la vinblastine et
la vincristine sont synthétisés dans la vacuole (Verpoorte et al., 1999).
2.2.4 Activation du métabolisme des alcaloïdes
Activation par élicitation. L’activation du métabolisme secondaire de la plante est consi-
déré comme étant une réponse à un stress. La production en métabolites secondaires est sou-
vent augmentée lorsque les cellules de plante ou les racines transformées sont soumises à des
produits chimiques qui miment l’attaque d’un pathogène, comme des extraits de levure, des
extraits de champignons (comme la chitine) ou de la pectinase (Lamb et al., 1989; Darvill
et Albersheim, 1984). L’élicitation désigne l’action de mimer une telle attaque sur une cel-
lule de plante. L’acide jasmonique est une molécule synthétisée par les cellules de plantes en
réponse à ce genre de stress (Doares et al., 1995). L’accumulation simultanée d’acide jasmo-
nique endogène et de métabolites secondaires suite à une élicitation a été montré dans des
cellules dePapaver somniferum(Mueller et al., 1993) et de riz (Nojiri et al., 1996). Ainsi,

35
�������
������
������
Figure 2.8 –Voie de signalisation par l’acide jasmonique. Un signal extérieur est transforméen signal chimique (1), un récepteur membranaire reçoit le signal (2) et active une lipase (3)qui libère de l’acide linolénique (4 et 5). Ce dernier est convertit en acide jasmonique par cinqréactions enzymatiques (6). L’acide jasmonique peut induire différents types de réponses (8)dont l’activation la transcription detrp et str dansC. roseus. Adapté de Creelman et Mullet(1997).
l’acide jasmonique est un messager secondaire dans la cascade de signalisation de la réponse
de la plante à un stress. La figure 2.8 présente le mécanisme de synthèse d’acide jasmonique
suite à un signal externe. Dans ce mécanisme, l’acide linolénique présent dans la membrane
plasmique est libéré dans le cytoplasme suite à l’activation d’un récepteur transmembranaire
par un signal externe. L’acide linolénique est convertit en acide jasmonique (Farmer et Ryan,
1992) et active la transcription de certains gènes par des mécanismes inconnus.
Menke et al. (1999) ont montré que l’élicitation de cellules deCatharanthus roseusmène
à la biosynthèse de l’acide jasmonique et à l’augmentation de la transcription des gènesstr et
tdc, responsables de la biosynthèse de la strictosidine et de la tryptamine. L’acide linolénique

36
et l’acide jasmonique induisent aussi la transcription destr et tdc lorsqu’ils sont ajoutés dans
le milieu de culture. Ces effets disparaissent lors de l’ajout d’un inhibiteur de protéine kinase.
De plus, le même inhibiteur empêche la biosynthèse d’acide jasmonique après élicitation.
Ainsi, l’action de l’acide jasmonique sur le métabolisme secondaire est clairement démontré
et fait intervenir des protéines kinases au niveau de la biosynthèse de l’acide jasmonique et
au niveau de son action régulatrice sur les gènesstr et tdc.
L’ajout d’acide jasmonique sur les racines transformées deC. roseusà différentes doses a
été étudié (Rijhwani et Shanks, 1998; Bhadra et Shanks, 1997). Des concentrations comprises
entre 10 et 50 mg/l d’acide jasmonique dans le milieu de culture on un effet sur la production
de serpentine, d’ajmalicine et de tabersonine. Dans certains cas, la production de tabersonine
est diminuée par l’élicitation à l’acide jasmonique.
Effet du statut en Pi de la cellule. La production de métabolites secondaires est également
fonction du statut nutritionnel de la plante. Une des fonctions du métabolisme secondaire est
la défense de la plante par la biosynthèse de composés toxiques pour l’agent pathogène ou
le prédateur. Néanmoins, ces mécanismes de défense sont moins importants si la plante n’est
pas limitée en nutriments, car elle peut remplacer les organes qui sont attaqués. Dans le cas
contraire, il est important pour la plante de conserver ses organes existants (Harbone, 1997).
D’autres auteurs proposent que l’arrêt de la croissance du à une limitation en phosphate par
exemple rend disponible des nutriments pour le métabolisme secondaire. Ce type de com-
portement est observé dans les cultures en suspension de cellules d’Escholtzia californica
dans le laboratoire BIOPRO. Ces cellules ne produisent pas de sanguinarine lorsque elle ne
sont pas limitées en phosphate. De plus, des racines transformées deHyoscyamus muticus
limitées en phosphate produisent beaucoup plus de solavetivone, cet effet étant renforcé par
une élicitation par des extraits de champignons (Dunlop et Curtis, 1991). Ainsi, les effets
d’une limitation en phosphate et de l’élicitation permettent d’établir une optimisation de la
production de métabolites secondaires. L’effet d’une limitation en nitrate sur la production
de métabolites secondaires devrait dépendre de leur nature. L’azote entre dans la composi-
tion des alcaloïdes. Par conséquent on s’attend à ce que la production d’alcaloïdes ne soit

37
pas augmentée aussi significativement par une limitation en nitrate que par une limitation en
phosphate. La production de métabolites ne contentant pas d’atome d’azote devrait répondre
différement à une limitation en nitrate.
2.3 Utilisation des racines transformées parAgrobacterium
rhizogenes
Agrobacteriumest un genre de bactéries gram-négatives vivant dans le sol. Les deux es-
pèces les plus connues sontA. tumefacienset Agrobacterium rhizogenesqui sont des patho-
gènes pour de nombreuse plantes dicotélydones et quelques espèces monocotylédones. L’in-
fection a lieu lorsque la bactérie a accès sur une zone blessée de la plante. La maladie en résul-
tant est un sorte de tumeur dont les caractéristiques dépendent de l’espèce d’Agrobacterium.
Ainsi, les racines transformées sont en fait une tumeur cancéreuse particulière dues à une
infection parA. rhizogenes. La caractéristique commune de ces tumeurs est qu’elles sont ca-
pables de biosynthétiser des dérivés de sucres et d’acides aminés appelés opines et qui sont
métabolisées exclusivement par la bactérie.Agrobacteriumest donc avant tout un parasite qui
se sert de la machinerie métabolique de la plante pour synthétiser le substrat qui lui sert de
source de carbone et d’azote. Il existe différents types d’opines, qui sont utilisées par diffé-
rentes lignées d’Agrobacterium.
2.3.1 Aspects historiques
La maladie decrown gall. C’est au début du siècle que le pathogène de la maladie appe-
léecrown galla été identifié comme étantA. tumefaciens(Smith et Townsend, 1907). Cette
maladie se caractérise par l’apparition de tumeurs non différenciées au niveau de la blessure
de la plante. Ce n’est que dans les années 70 que l’origine de la virulence de la bactérie est
identifiée, bien qu’en 1941, White et Braun aient déjà montré que le le tissu une fois infecté
pouvait garde les propriétés de la tumeur même en l’absence d’A. tumefaciens. En 1974, la
découverte d’un plasmide de 200-800 kb porté par les souches virulentes deA. tumefaciens

38
(Zaenen et al., 1974) permit de nombreuses expérimentations qui montrèrent que i), l’élimina-
tion du plasmide dans une souche virulente la rend non-virulente (VanLarebeke et al., 1974)
et ii), le transfert du plasmide sur une souche non-virulente la rend virulente et le plasmide
n’est pas intégré au sein de l’ADN bactérien (Watson et al., 1975). Les résultats de Braun et
White furent partiellement expliqués lorsqu’il fut montré parSouthern blotque les cultures
de tumeurs de lacrown gallen l’absence d’A. tumefaciensavaient acquis de manière stable
une portion du plasmide (Chilton et al., 1977). Le plasmide a été nommé plasmideTumor
inducing(Ti). Deux zones distinctes le composent. La portion qui est transférée dans l’ADN
de la plante hôte est appelé T-DNA, la portion qui code pour les facteurs nécessaires à ce
transfert est appelée la région de virulence (vir). Les distinctions principales entre les diffé-
rents T-DNA proviennent du type d’opines dont ils codent les enzymes responsables de leur
synthèse. Les tumeurs proviennent de la synthèse d’auxines et de cytokinines par certainsloci
du T-DNA exprimés dans la plante. Un plasmide Ti peut contenir un ou plusieurs T-DNA, qui
sont alors séparés par des séquences conservées de 25 base.
La maladie des racines chevelues.Les symptômes de cette maladie sont l’apparition d’une
tumeur sous forme de racines organisées, beaucoup plus ramifiées que les racines normales.
Le pathogène responsable de cette maladie estA. rhizogenes. La biologie de l’infection d’A.
rhizogenesest très similaire à celle d’A. tumefaciens: les souches virulentes contiennent un
plasmide appeléRoot-inducing(Ri) dont une partie est intégrée à l’ADN de la plante infectée.
Les racines transformées sont capables de synthétiser un type d’opines qui sont métabolisées
par le parasite. La classification des plasmides Ri est établie comme celle des plasmides Ti,
selon le type d’opines synthétisées. Les lignées d’A. rhizogenesétudiées le plus ancienne-
ment sont les lignées A4 et 8196. La lignée 8196 contient un seul T-DNA, qui n’a aucune
homologie avec le T-DNA d’A. tumefaciens. La lignée A4 contient deux T-DNA, nommés
Ri-TR-DNA et Ri-TL-DNA (White et al., 1985). Le Ri-TR-DNA est très proche du T-DNA
d’A. tumefaciens. Le Ri-TL-DNA a un quelques homologies avec Ri-T-DNA de la lignée
8196 d’A. rhizogenes. Ri-TL-DNA pourait être suffisant pour induire la maladie deshairy
roots: on a observé l’absence de Ri-TR-DNA dans des plantes transformées avec la lignée

39
A4 d’A. rhizogenes(Jouanin et al., 1987). Le Ri-TR-DNA, qui est responsable de la pro-
duction d’auxines et de cytokinines, stimulerait l’action du Ri-TL-DNA, ce qui suggère des
modes d’infection entraînant les symptômes de la maladie complètement différents entreA.
tumefaciensetA. rhizogenes. Le plasmide pRiA4 est devenu le plasmide de référence pour la
nomenclature des différentsloci présents sur Ri-TL-DNA selon leur fonctions.
Le Ri-TR-DNA porte 4loci responsables de la morphologie des racines transformées :
rolA, rolB, rolC et rolD (White et al., 1985).rolA semble être lié au métabolisme de l’acide
gibbérellique impliqué dans le développement de la plante. Son expression est cruciale pour
l’établissement du phénotype "racines transformées" (Meyer et al., 2000). Selon le modèle
actuel, l’expression derolB produit une protéine membranaire spécialisée dans le reconnais-
sance d’auxines qui modifie la réponse de la plante aux auxines. Ceci se traduit par un tissu
racinaire plus dense que la normale. CommerolA, rolC est impliqué dans le métabolisme de
l’acide gibbérellique. Un modèle récent propose querolC serait impliqué dans le métabolisme
et le transport des sucres (Nilson et al., 1997).rolD n’est pas essentiel pour induire le phéno-
type des racines transformées, mais son absence retarde l’apparition des racines transformées
(Meyer et al., 2000).
2.3.2 Mécanismes cellulaires de transformation
A. tumefaciensa été beaucoup plus étudié queA. rhizogenes. Aussi, les mécanismes cel-
lulaire de transformation sont généralement décrit chezA. tumefaciens. Néanmoins, ces mé-
canismes restent semblables à ceux d’A. rhizogenes. On peut dissocier le mécanisme de la
transformation en trois étapes : l’adhésion bactérie-plante, l’activation des gènes de virulence
vir et l’exportation de plasmide T-DNA de la cellule hôte.
Adhésion bactérie-plante. L’adhésion de la bactérie à la plante est une nécessité pour que
l’infection ait lieu. Les mutants d’A. tumefaciensincapables de s’attacher aux cellules de
plantes sont toujours avirulents. D’autre part, les souches d’A. tumefaciensavirulentes mais
capables de s’attacher aux cellules de plantes peuvent inhiber la formation des tumeurs lors-
qu’elles sont incubées avec une forme virulente d’A. tumefacienslors de l’infection d’une

40
plante (Lippincott et Lippincott, 1969; Tanimoto et al., 1979).
Mécanisme bactérien de l’adhésion. Les loci connus responsables de la synthèse des
enzymes et protéines impliquées dans l’adhésion d’A. tumefacienssur les cellules de plantes
ne sont pas localisés sur le Ti-DNA, mais sur l’ADN chromosomal d’A. tumefaciens. Le
Ti-DNA ne contiendrait donc aucun gène permettant l’adhésion bactérie plante. Aussi, l’in-
corporation du Ti-DNA dans un micro-organisme incapable de s’attacher sur les cellules de
plantes résulte en un micro-organisme incapable d’initier des tumeurs engendrées par le Ti-
DNA. Un système tel queE. coli, qui ne peut pas s’ancrer sur les cellules de plantes, contenant
le Ti-DNA ou le Ri-DNA est donc incapable de transformer une plante.
Huit loci du chromosome bactérien sont principalement impliqués dans l’adhésion de la
bactérie sur la plante. Ce sont :chvA, chvB(Douglas et al., 1985),chvD, exoC(Cangelosi et
al., 1987),cel (Matthysse, 1983),att (Mathysse, 1987),miaA et ros. Les deuxloci chvAet
chvBainsi queexoCsont impliqués dans la synthèse et l’excrétion d’unβ-1,2-glucane hors
de la bactérie, un polysaccharide normalement périplasmique, dont le rôle dans l’adhésion
bactérie-plante, bien qu’indispensable, est encore mal compris. Lelocus celest responsable
de la synthèse d’un polysaccharide sous forme de fibriles, dont le rôle est d’améliorer l’adhe-
sion bactérie-plante une fois qu’elle a été initiée (Mathysse et Gurlitz, 1982). Le rôle dulocus
att n’est pas encore clair, bien que son expression soit impérative pour l’adhésion bactérie-
plante. Lesloci attB, attD et attRont été étudiés avec plus d’attention. Il est suggéré que la
synthèse de la cellulose serait impliquée dans la capacité pathogène d’A. tumefaciens(Ma-
thysse et McMahan, 1998). Les rôles demiaA, chvDet rossont mal compris.
Sites d’adhésion des cellules végétales.A. tumefaciensa pour cible les parois cellu-
laires des cellules végétales contenant de la pectine. Ceci est confirmé par le fait que i)A.
tumefaciensco-incubée avec des parois cellulaires de cellules végétales ne provoque pas la
formation de tumeursin vivo (Lippincott et Lippincott, 1977), ii)A. tumefaciensincubée
in vitro en présence de paroi cellulaires végétales enrichies en pectine sur des cellules en
suspension de tomate perd ses propriétés d’adhésion (Neff et Binns, 1985) et iii) la pectine

41
ajoutée seule n’inhibe pas l’adhésion d’A. tumefacienssur des cellules en suspension de ca-
rotte (Mathysse et Gurlitz, 1982). L’importance de la paroi cellulaire végétale a été montrée
par le fait que la régénération de la paroi cellulaire de protoplastes de tabac est nécéssaire
pour l’adhésion d’A. tumefacienset la transformation des protoplastes (Krens et al., 1985).
Activation des gènesvir. Les gènes qui codent pour les protéines impliquées dans la pré-
paration du plasmide T-DNA et dans son transport sont portés par le Ti-DNA et le Ri-DNA.
25 opérons ont été identifiés. Les plus étudiées sont :virA, virB, virC, virD, virE, virG, virH,
virJ, virK virL et virM. Cinq loci sont essentiels pour le pouvoir pathogène:virA, virB, virD
et virG. virC et virE augmentent ce pouvoir pathogène. Ils sont localisés dans la portionvir
(35 kb) du Ti-DNA et Ri-DNA. ExceptévirA et virG, les opéronsvir sont polycistroniques.
A gauche devir se trouve un autre opéronpinF dont la présence n’est pas requise pour en-
gendrer un tumeur, mais dont la régulation dépend des gènesvir. De même,virH, virK, virL
et virM ne sont pas indispensables pour induire la formation de tumeurin vitro (Kalogeraki
et Winans, 1998). Des gènes chromosomiques sont également impliqués dans le transfert du
T-DNA : chvEetacvF.
Les gènesvir sont activés par trois types de signaux chimiques que la plante relâche en
cas de blessure : i) composés phénoliques dérivés du syringol (le plus actif étant l’acétosyrin-
gone), ii) sucres tels que glucose et acide glucorinique et iii) pH acide.
Voie de signalisation par les protéines Vir. L’activation des gènesvir est un phéno-
mène indépendant de l’adhésion d’Agrobacteriumsur une cellule végétale. VirA, qui aurait
deux segments transmembranaires, est exprimé à la surface de la bactérie (Leroux et al.,
1987). VirA est une protéine kinase qui s’autophosphoryle lorsqu’elle est activée par un com-
posé phénolique présent en concentration élevée. Cette activation se fait par une interaction
directe entre VirA et le composé phénolique inducteur (Lee et al., 1995, 1996). Une fois
phosphorylée, elle active la protéine cytoplasmique VirG par phosphorylation. Cette dernière
déclenche l’expression des autres gènesvir. La spécificité sur les composés phénoliques de
VirA est très étroite et dépend du plasmide Ti (Lee et al., 1996). Une étude par mutagénèse

42
dirigée a donné deux mutants de VirG actifs même en absence d’acétosyringone (Scheeren-
Groot et al., 1994).
Voie de signalisation par des protéines chromosomiques.Le deuxième mécanisme
d’activation des gènesvir est pris en charge par ChvE qui est une protéine périplasmique
chromosomique pouvant lier des sucres. Elle est homologue auxgalactose-binding protein
deE. coli (Doty et al., 1993). Lorsque ChvE est activée par son ligand, elle augmente la sen-
sibilité de VirA aux composés phénoliques, permettant une activation de VirA en présence de
faibles concentrations de ces composés. Lorsque ChvE est hautement exprimée dansA. tume-
faciens, la sélectivité de VirA s’étend à d’autre composés phénoliques n’ayant normalement
qu’un faible pouvoir inducteur des gènesvir (Peng et al., 1998). ChvE aurait des caractéris-
tiques différentes selon la lignée d’A. tumefaciens, permettant une association optimale avec
VirA de la même lignée (Bélanger et al., 1997). ChvE interagit avec la partie périplasmique
de VirA, au niveau de sites de liaison de ChvE sur VirA (Banta et al., 1994). Le tandem
ChvE/VirA est donc sensible à deux types de signaux chimiques : hautes concentrations en
composés phénoliques spécifiques d’une part et faibles concentrations en composés phéno-
liques moins spécifiques en présences de sucres d’autre part.
Activation de VirG. L’acidité du milieu externe est le troisième élément intervenant
dans l’activation des gènesvir. Elle active l’expression devirG, placé sous le contrôle de
deux promoteurs, dont l’un est sensible à une acidification du milieu externe,via une protéine
chromosomique ChvI, dont l’action n’est pas tout à fait confirmée (Mantis et Winans, 1993;
Charles et , 1993). Néanmoins, aucune protéine du plasmide Ti ne semble être impliquée dans
l’activation de ce promoteur (Mantis et Winans, 1992). Le deuxième promoteur est sensible
à VirG phosphorylée et à une carence en phosphate.
Exportation du plasmide T-DNA dans l’ADN de la cellule végétale cible.
Préparation du plasmide T-DNA. Le plasmide T-DNA est initialement intégré dans le
Ri-DNA ou le Ti-DNA. C’est sous forme de simple brin qu’il est exporté dans le noyau de la

43
cellule végétale cible. Les protéines VirD1 et VirD2 sont des endonucléases spécifiques qui
reconnaissent les bords du T-DNA ainsi que le brin qui sera exporté. Elles coupent le brin
selon les séquences spécifiques bordant le T-DNA. VirD2 s’attache de manière covalente sur
l’extrémité 5’ du simple brin du T-DNA.
Transport du plasmide T-DNA. Le brin de T-DNA est recouvert par plusieurs copies
(environ 600) de la protéine VirE2, ce qui permettrait la protection du T-DNA contre les
nucléases de la cellule végétale cible (Howard et Citovsky, 1990). Les protéines VirE1 stabi-
lisent les protéines VirE2 (Citovsky et al., 1992; Sundberg et al., 1996). L’ensemble VirD2,
VirE2 et T-DNA simple brin constitue le « compexe T ». Des résultats récents suggèrent que
les protéines VirE2 sont exportées séparément du T-DNA vers la cellule végétale (Lee et al.,
1999). On suppose que les protéines VirE2 et VirD2 sont aussi responsables de l’adressage
du T-DNA vers le noyau, à travers les pores nucléaires. Les protéines virB (11 protéines pro-
duites par l’opéronvirB) ainsi que VirD4 serviraient de canal entre la bactérie et la cellule
végétale cible. De nombreuses études comparent le système de transfert du T-DNA d’A. tu-
mefaciensavec le système de conjugaison du plasmide F deE. coli, régulé par les gènestra
qui permettent l’établissement d’un tunnel entre la cellule hôte possédant le plasmide et une
cellule cible dépourvue du plasmide. Ce « tunnel » est formé par unpilus par lequel transite
le plasmide. Dans le cas d’A. tumefaciens, la protéine VirB2 serait la composante essentielle
dupilusqu’A. tumefaciensforme pour se connecter sur la cellule végétale cible. La structure
prédite de VirB2 est la même que TraA, impliquée dans la conjugaison du plasmide F deE.
coli. virB2, virB3 etvirB9 sont impliquées dans la synthèse dupilusde conjugaison (Jones et
al., 1996). L’export de VirB2 nécessiterait l’action d’autres protéines Vir (Lai et Kado, 1998).
VirB1, située sur la membrane interne d’A. tumefacienset ayant une structure N-terminale
similaire aux transglycosidases bactériennes, serait responsable de la lyse local de la paroi
cellulaire de la bactérie, permettant la formation dupilus (Baron et al., 1997).virB1 est aussi
responsable de la synthèse d’unpropilin, impliqué dans le contact initial avec la cellule cible,
par l’assemblage de protéines VirB1, sécrétée dans le milieu de culture (Baron et al., 1997;
Chumakov et Kurbanova, 1998). La protéine chromosomique AcvB jourait un rôle essentiel

44
dans le transfert du plasmide, mais ce rôle pourrait être remplacé par l’expression devirJ,
regulée par l’acetosyringone (Pan et al., 1995). VirB8 est située sur la membrane interne d’A.
tumefaciens(Thorstenson et Zambryski, 1994).
Intégration du plasmide T-DNA. Le plasmide est exporté dans la cellule végétale sous
forme de simple brin. Il n’est pas encore clair si ce simple brin est complété en double brin
puis intégré au sein du génome de la plante ou le simple brin est d’abord intégré dans le
génome puis le brin complémentaire est synthétisé avec les mécanismes de réparation de
l’ADN de la plante. De nombreux résultats suggèrent que les protéines VirD2 participent dans
l’intégration du T-DNA dans le génome de la plante (Mysore et al., 1998; Narasimhulu et al.,
1996). VirE2 semble également y jouer un rôle indirect, car les mutations sur VirE2 donne
des souches d’Agrobacteriumdont la virulence est très atténuée (Dombek et Ream, 1997;
Stachel et Nester, 1986). Il est maintenant reconnu que le T-DNA s’intègre dans le génome
de la cellule végétale par une recombinaison illégitime (Gheysen et al., 1991; Matsumoto
et al., 1990; Ohba et al., 1995). De plus, cette intégration a lieu préférentiellement dans les
parties du génome qui sont transcrites régulièrement (Herman et al., 1990; Koncz et al., 1989,
1992).
2.3.3 Protocoles de transformation parAgrobacterium rhizogenes
L’obtention de racines transformées passe par plusieurs étapes : culture et activation de la
bactérie, infection et co-culture de la plante, isolation des racines transformées et confirmation
génétique de la transformation.
Culture d’ A. tumefacienset A. rhizogenes.
Préparation de la bactérie. Agrobacteriumest cultivé en routine sur un milieu solide
à 26◦C. Pour infecter une plante, on utiliseAgrobacteriumcultivée en milieu liquide. Le
transfert d’Agrobacteriumen milieu liquide a lieu entre 1 et 3 jours avant le jour de l’infection,
sur agitateur orbital à 80 rpm. Les concentrations utilisées sont en général entre108 et 1010

45
bactéries par ml (Hamill et al., 1987) ou une absorbance de A600=0,9 à A600=1. Le milieu
utilisé peut être du milieu Nutrient Broth. Parfois, les bactéries sont centrifugées à 5000g
pendant 10 min, puis resuspendues dans un milieu propre aux cellules végétales, tels que le
milieu B5 (Gamborg et al., 1968).
Sur-activation de la virulence. Pour activer les gènesvir, il est possible d’utiliser d’uti-
liser un milieu ayant contenu pendant un certain temps des feuilles blessées de l’espèce à in-
fecter qui auront libéré les composés phénoliques responsables de l’activation des gènesvir.
Plusieurs protocoles sont utilisés, mais le trait commun consiste en la culture de morceaux de
feuilles dans un milieu nutritif liquide pendant quelques jours. La bactérie est ensuite cultivée
dans ce milieu au lieu d’un milieu de culture de bactéries (Sunilkumar et al., 1999). On peut
également utiliser l’acétosyringone pour activerAgrobacterium. Il est ajouté dans la suspen-
sion bactérienne dans les concentrations de l’ordre duµM: 60 µM avecA. tumefaciensA348
sur des feuilles de tabac (Sunilkumar et al., 1999), 200µM avecA. tumefaciensC58 surAr-
temisia annua(Ghosh et al., 1997) et 10µM avecA. rhizogenessurC. roseus(Hamill et al.,
1987).
Infection et co-culture. Le matériel végétal utilisé provient soit de plantes non stériles, au-
quel cas il est stérilisé, soit de plantes stériles âgées de 15 à 30 jours. La période de co-culture
se fait en général dans l’obscurité. En cas d’infection parA. rhizogenes, selon l’espèce infec-
tée, les racines apparaissent directement au niveau des blessures ou il y a d’abord formation
d’un cal puis les racines transformées émergent de ce cal. Plusieurs protocoles d’infection ont
été décrits.
Infection par immersion. Le matériel végétal (en général des morceaux de feuilles)
est plongé dans la suspension bactérienne pendant une période variant de quelques minutes
(Santos et al., 1998) à deux jours (Garnier et al., 1996). On peut envisager l’inoculation sur
un agitateur orbital à une vitesse de 30 rpm (Godo et al., 1997). Une étape de rinçage dans
du milieu de culture neuf peut avoir lieu. Le matériel végétal est ensuite séché sur du papier

46
filtre stérile, puis déposé sur un milieu neuf solide pour co-culture pendant deux (Zhan et al.,
1997) à cinq jours (Hoshino et al., 1998).
Infection par dépôt. Le matériel végétal (en général des tiges) est blessé avec une se-
ringue contenant la suspension bactérienne (Hamill et al., 1987). Quelquesµl de suspension
bactérienne sont injectés sur la blessure. On peut aussi couper la tige, appliquer la suspension
bactérienne sur la partie basale de la tige, et enfoncer la partie apicale dans du milieu MS soli-
difié (Pavlova et al., 1998). La durée de la co-culture varie entre un jour et jusqu’à apparition
des racines (Tepfer, 1984), bien que des durées de un à trois jours soient les plus répandues. Il
est souvent nécessaire pendant la co-culture de ne pas mettre en contact les parties infectées
de la plante avec le milieu de culture pour éviter la croissance d’Agrobacterium.
Récupération des racines transformées La première étape consiste à éliminerA. rhizo-
genesen utilisant un antibiotique, puis à cultiver les racines qui émergent des sites d’infection
sur un milieux de culture solidifié.
Utilisation des antibiotiques. Habituellement, on utilise la carbenicilline à une dose de
500 mg/l pour éliminerAgrobacterium. Il semblerait qu’une telle concentration ne soit pas
suffisante et que pour avoir une destruction totale d’Agrobacterium, il faille utiliser 1 g/l de
carbenicilline. Néanmoins, la carbenicilline se révèle être calogène, ce qui n’est pas désiré
lors de l’établissement de racines transformées (Nauerby et al., 1997). La timentine à une
concentration de 150 mg/l pourrait, selon les mêmes auteurs, inhiber totalement la croissance
d’Agrobacterium, en supprimant les effets calogènes de la carbenicilline. On peut utiliser la
céfotaxime à une concentration de 500 mg/l, mais elle provoquerait des effets secondaires sur
la plante tels qu’une croissance ralentie (Nauerby et al., 1997).
Lorsque la lignée utilisée possède un gène de résistance à la kanamycine sur le T-DNA,
on utilise cette dernière à une concentration de 100 mg/L à 300 mg/L. La kanamycine tue les
bactéries non résistantes, ainsi que les cellules végétales qui n’ont pas le T-DNA. Ceci permet
de faire une sélection des cellules végétales transformées. Cependant, la kanamycine à 300

47
mg/L peut inhiber la croissance deCatharanthus roseus(Garnier et al., 1996).
Culture du matériel végétal transformé. En général, le fait de couper des racines
transformées émergeant du site d’infection suffit à en faire des cultures stables. Il arrive
qu’il faille tamponner le pH du milieu, utiliser des milieux avec des concentrations dimi-
nuées de moitié en sels majeurs, ou ajouter des facteurs de croissance ou des phytohormones.
Néanmoins, ces techniques causent souvent l’apparition de cals indésirables (références dans
Rhodes et al. (1987)).
Confirmation de la transformation. L’ancienne méthode pour confirmer la transformation
d’un tissu végétal parAgrobacteriumest le dosage des opines par électrophorèse sur papier et
coloration au nitrate d’argent (Tepfer et al., 1981; Bécard et Fortin, 1988). Néanmoins, l’ex-
pression des gènes codant pour la synthèse des opines n’est pas stable dans le temps (Tepfer,
1984; Kamada et al., 1986). Deux méthodes plus précises sont utilisées : une détection du
T-DNA par hybridisation parSouthern blot(Rhodes et al., 1987), ou une amplification du
T-DNA du tissu par PCR (Pinol et al., 1999).





















![[tel-00334974, v1] Biodégradation du 2-éthylhexyl nitrate](https://img.pdfslide.fr/doc/110x75/62ae5b320702301cb711ca30/tel-00334974-v1-biodgradation-du-2-thylhexyl-nitrate-.jpg)






